Vias metabolicas
30
1 METABOLISMO OXIDATIVO Utilización de la energía almacenada en los nutrientes para la generación de ATP Entonces, ¿para qué comemos? La ingesta de alimentos constituye una necesidad fisiológica primordial, porque cuando comemos ingerimos fuentes de energía. ¿Qué significa esto? Los alimentos son combustibles, y al quemarlos (u oxidarlos, químicamente hablando) obtenemos energía, que conservada en forma de ATP, se utiliza para suplir las necesidades energéticas del organismo. En todas las células ocurren sistemas complejos de reacciones químicas que están regulados en forma compleja y producen y utilizan energía. Todos los procesos que ocurren en nuestras células son procesos de transformación de energía. Los combustibles metabólicos (glúcidos, lípidos y proteínas) constituyen una mezcla compleja de macromoléculas que contiene una gran cantidad de enlaces C-C y C-H, que al romperse permiten obtener otras formas de energía. El gradiente electroquímico de energía potencial que permite crear un medio intracelular de composición diferente del externo, o el movimiento organizado de grupos de células, son algunos ejemplos de utilización de energía. Los procesos de transformación involucrados en la utilización de la energía del enlace químico de los combustibles pueden dividirse en tres fases: 1) obtención de energía a partir de la oxidación de combustibles, 2) conversión de esa energía en la forma biológicamente útil que se encuentra en las uniones de alta energía del ATP y 3) utilización de la energía de las uniones del ATP. Las primeras dos fases son parte del proceso de respiración celular. Bioquímicamente, al hablar de respiración celular, nos referimos a los procesos celulares que al oxidar los combustibles a CO 2 , consumen oxígeno y generan ATP. La respiración celular es un conjunto complejo de procesos que se dividen en 3 fases (ver Figura). En la Fase 1 de la respiración celular, se produce la oxidación enzimática de los combustibles metabólicos con transferencia de electrones a las coenzimas aceptoras NAD + y FAD que se reducen a NADH y FADH 2 , respectivamente. Además, en esta Fase de la respiración celular, la mayoría de los combustibles metabólicos (glúcidos, ácidos grasos y muchos aminoácidos) convergen en la generación del grupo acetilo (de 2 C) activado del acetilCoA. En la Fase 2 de la respiración celular, la oxidación completa del grupo acetilo del acetilCoA a CO 2 (el compuesto más estable del carbono) ocurre en el ciclo de los ácidos tricarboxílicos (o ciclo de Krebs, o ciclo del ácido cítrico) y también aquí, la energía se almacena principalmente como NADH y FADH 2 . La Fase 2 de la respiración celular ocurre exclusivamente en las mitocondrias.
-
Upload
roger-casas -
Category
Education
-
view
696 -
download
3
Transcript of Vias metabolicas

1
METABOLISMO OXIDATIVO
Utilización de la energía almacenada en los nutrientes para la generación de ATP
Entonces, ¿para qué comemos? La ingesta de alimentos constituye una necesidad fisiológica
primordial, porque cuando comemos ingerimos fuentes de energía. ¿Qué significa esto? Los
alimentos son combustibles, y al quemarlos (u oxidarlos, químicamente hablando) obtenemos
energía, que conservada en forma de ATP, se utiliza para suplir las necesidades energéticas del
organismo.
En todas las células ocurren sistemas complejos de reacciones químicas que están regulados en
forma compleja y producen y utilizan energía. Todos los procesos que ocurren en nuestras
células son procesos de transformación de energía. Los combustibles metabólicos (glúcidos,
lípidos y proteínas) constituyen una mezcla compleja de macromoléculas que contiene una gran
cantidad de enlaces C-C y C-H, que al romperse permiten obtener otras formas de energía. El
gradiente electroquímico de energía potencial que permite crear un medio intracelular de
composición diferente del externo, o el movimiento organizado de grupos de células, son algunos
ejemplos de utilización de energía.
Los procesos de transformación involucrados en la utilización de la energía del enlace químico
de los combustibles pueden dividirse en tres fases: 1) obtención de energía a partir de la
oxidación de combustibles, 2) conversión de esa energía en la forma biológicamente útil que se
encuentra en las uniones de alta energía del ATP y 3) utilización de la energía de las uniones del
ATP.
Las primeras dos fases son parte del proceso de respiración celular. Bioquímicamente, al hablar
de respiración celular, nos referimos a los procesos celulares que al oxidar los combustibles a
CO2, consumen oxígeno y generan ATP.
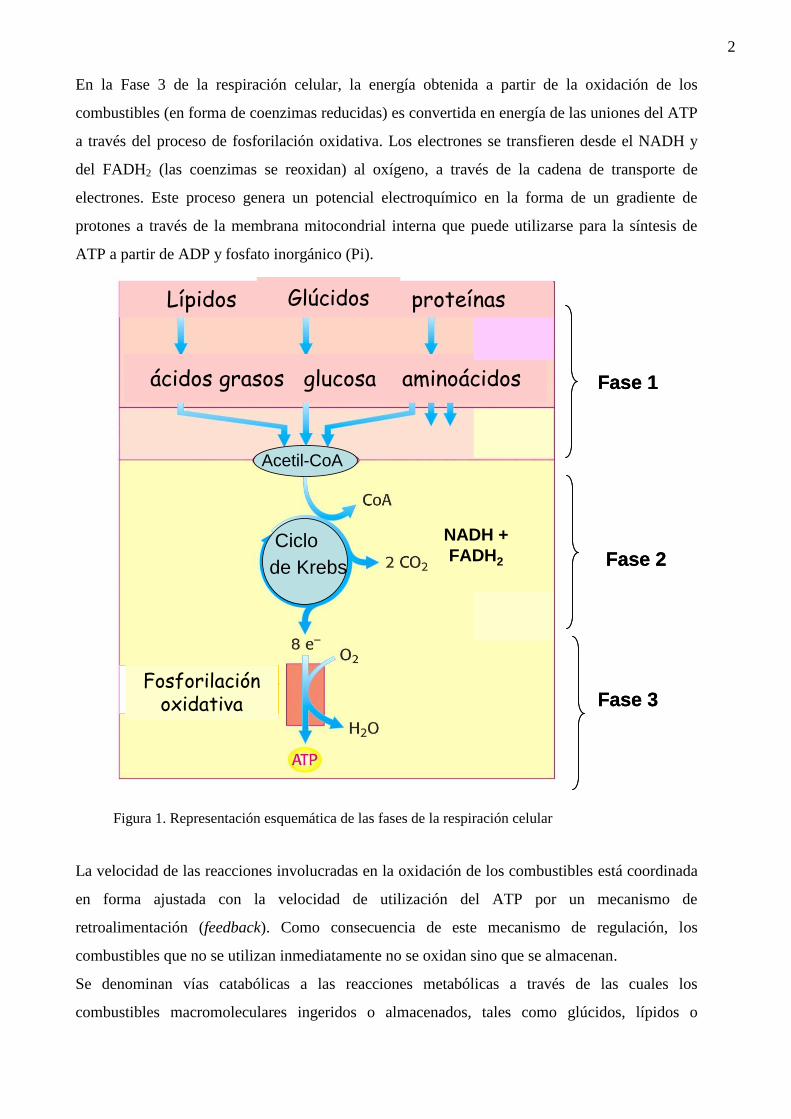
La respiración celular es un conjunto complejo de procesos que se dividen en 3 fases (ver
Figura). En la Fase 1 de la respiración celular, se produce la oxidación enzimática de los
combustibles metabólicos con transferencia de electrones a las coenzimas aceptoras NAD+ y
FAD que se reducen a NADH y FADH2, respectivamente. Además, en esta Fase de la respiración
celular, la mayoría de los combustibles metabólicos (glúcidos, ácidos grasos y muchos
aminoácidos) convergen en la generación del grupo acetilo (de 2 C) activado del acetilCoA. En
la Fase 2 de la respiración celular, la oxidación completa del grupo acetilo del acetilCoA a CO2
(el compuesto más estable del carbono) ocurre en el ciclo de los ácidos tricarboxílicos (o ciclo de
Krebs, o ciclo del ácido cítrico) y también aquí, la energía se almacena principalmente como
NADH y FADH2. La Fase 2 de la respiración celular ocurre exclusivamente en las mitocondrias.
2
En la Fase 3 de la respiración celular, la energía obtenida a partir de la oxidación de los
combustibles (en forma de coenzimas reducidas) es convertida en energía de las uniones del ATP
a través del proceso de fosforilación oxidativa. Los electrones se transfieren desde el NADH y
del FADH2 (las coenzimas se reoxidan) al oxígeno, a través de la cadena de transporte de
electrones. Este proceso genera un potencial electroquímico en la forma de un gradiente de
protones a través de la membrana mitocondrial interna que puede utilizarse para la síntesis de
ATP a partir de ADP y fosfato inorgánico (Pi).
Figura 1. Representación esquemática de las fases de la respiración celular
La velocidad de las reacciones involucradas en la oxidación de los combustibles está coordinada
en forma ajustada con la velocidad de utilización del ATP por un mecanismo de
retroalimentación (feedback). Como consecuencia de este mecanismo de regulación, los
combustibles que no se utilizan inmediatamente no se oxidan sino que se almacenan.
Se denominan vías catabólicas a las reacciones metabólicas a través de las cuales los
combustibles macromoleculares ingeridos o almacenados, tales como glúcidos, lípidos o
ácidos grasos glucosa aminoácidos
Fase 2
Fase 1
Fase 3
Lípidos polisacáridos proteínas
NADH +
FADH2
Fosforilación oxidativa
Acetil-CoA
Ciclo
de Krebs
ácidos grasos glucosa aminoácidos
Fase 2
Fase 1
Fase 3
Lípidos polisacáridos proteínas
NADH +
FADH2
Fosforilación oxidativa
Acetil-CoA
Ciclo
de Krebs
Glúcidos

3
proteínas son degradados secuencialmente para obtener energía. Las reacciones catabólicas
resultan generalmente en la conversión de moléculas grandes y complejas en moléculas pequeñas
(y finalmente a CO2 y H2O), en la producción de energía almacenable o conservable, y en general
en el consumo de oxígeno (es decir, generalmente son reacciones de oxidación). De hecho, la
única vía que puede generar ATP sin oxígeno es la glucólisis anaeróbica. Por el contrario, las
vías metabólicas involucradas en la biosíntesis de macromoléculas se denominan vías anabólicas
y generalmente resultan en la síntesis de moléculas grandes y complejas a partir de precursores
pequeños. A diferencia de las reacciones catabólicas, los procesos anabólicos son reductivos
(reacciones de reducción) y consumen energía.
Los ácidos grasos son el principal combustible metabólico del organismo. Luego de una
comida rica en glúcidos (por ejemplo pastas, pan, azúcar, o papas), su exceso se almacena como
las unidades glucosilo del glucógeno. Sin embargo, esta forma de almacenamiento es
relativamente limitada, de manera que el exceso de glúcidos por encima de la capacidad de ser
almacenados como glucógeno, se convierte en triacilglicéridos (“depósitos lipídicos”) que se
almacenan en el tejido adiposo. Obviamente, también el exceso de lípidos de la dieta se almacena
de esta forma.
Entre las comidas, se liberan ácidos grasos de estos depósitos, circulan por la sangre unidos a
albúmina y se oxidan a acetilCoA en el músculo, hígado y muchos otros tejidos por la vía de la
-oxidación. Uno de los principales destinos metabólicos del acetilCoA es el tema de esta clase.
En el hígado, sin embargo, el acetilCoA también puede convertirse en cuerpos cetónicos que se
oxidan en músculo y otros tejidos y se convierten en el combustible principal del cerebro en un
ayuno prolongado. Las vías de oxidación de ácidos grasos y de cuerpos cetónicos utilizan NAD+
y FAD como aceptores de electrones.
Todas las células requieren ATP en forma continua. Sin embargo, el aporte de combustibles para
obtener energía no es un proceso continuo. La disponibilidad constante de combustibles a pesar
de la discontinuidad de las ingestas alimenticias y de la variación en la velocidad de su
utilización, se denomina homeostasis metabólica y se logra a través de la regulación hormonal
de las vías de almacenamiento y de movilización de combustibles, principalmente mediada por
las hormonas metabólicas, esencialmente la insulina y las hormonas contrarregulatorias de la
insulina, el glucagon, la adrenalina y el cortisol.
La glucosa tiene un rol central en la homeostasis metabólica debido a que el cerebro y algunos
otros tejidos requieren glucosa para una gran parte de sus necesidades energéticas. Los niveles
sanguíneos de glucosa en personas sanas se mantienen en alrededor de 80 mg%. Niveles

4
inadecuados de insulina o resistencia de los tejidos a los efectos de la insulina ocasionan la
hiperglucemia característica de la Diabetes Mellitus.
El ciclo de Krebs constituye la etapa intermedia del metabolismo. Además de ser la vía final de
oxidación de los combustibles, también provee de intermediarios para procesos anabólicos. Este
rol central en el metabolismo hace de la comprensión de su funcionamiento y regulación, un
tema clave para el estudio del metabolismo normal y de su alteración en diferentes patologías.
PIRUVATO: ¿DE DONDE PROVIENE Y CUÁL ES SU DESTINO METABOLICO?
Antes de empezar a hablar en detalle del ciclo de Krebs (ciclo de los ácidos tricarboxílicos o
CAT) vamos a analizar primero una de las reacciones que genera acetilCoA. Nos referimos a la
reacción de descarboxilación oxidativa del piruvato catalizada por el complejo de la piruvato
deshidrogenasa. Esta reacción NO FORMA PARTE DEL CICLO DE KREBS pero la mayor
parte de los libros de texto la incluyen en esta unidad, posiblemente porque no forma parte de
ninguna otra vía metabólica. Esta reacción también ocurre en las mitocondrias y el complejo
enzimático que la cataliza es similar a uno que interviene en una de las reacciones del ciclo. Sin
embargo, no confundirse, pues la importancia de esta reacción como fuente de acetilCoA
depende del tejido y además NO ES LA UNICA FUENTE DEL ACETILCoA para el CAT.
El piruvato es el producto final de la glucólisis aeróbica que ocurre en el citosol y también de la
degradación de algunos aminoácidos como alanina, serina y cisteína.
El destino del piruvato depende del tejido en el que se produce y de su estado metabólico.
Además de ser sustrato del complejo de la piruvato deshidrogenasa puede seguir otras vías
metabólicas como por ejemplo: transaminación a alanina, carboxilación para generar
oxaloacetato (sustrato gluconeogenético) y reducción a lactato (glucólisis anaeróbica)
PIRUVATO DESHIDROGENASA

5
Como ya hemos dicho, el piruvato se transforma en acetilCoA por descarboxilación oxidativa, en
una reacción catalizada por un complejo multienzimático denominado piruvato
deshidrogenasa, que cataliza la siguiente reacción:
Piruvato + NAD+ + CoASH acetil CoA + CO2 + NADH + H
+ Gº’= -8 kcal/mol
Esta enzima se localiza exclusivamente en mitocondrias, en altas concentraciones en tejidos
como el músculo cardíaco y el riñón. Dado el alto valor negativo del Gº’ de la reacción, en
condiciones fisiológicas la reacción es esencialmente irreversible, y este hecho es la principal
razón por la cuál la conversión neta de ácidos grasos a carbohidratos no puede ocurrir en
nuestro organismo.
El peso molecular del complejo de riñón, corazón o hígado varía entre 7 y 8.5 x 106 y en
mamíferos consta de tres tipos diferentes de subunidades catalíticas:
Número de
subunidades
tipo peso
molecular
estructura
20 ó 30 Piruvato deshidrogenasa (E1) 154.000 2- 2
tetrámero
60 Dihidrolipoil transacetilasa (E2) 52.000 idénticas
6 Dihidrolipoil deshidrogenasa
(E3)
110.000 2 Dímero
El complejo de la piruvato deshidrogenasa está compuesto por tres actividades enzimáticas,
denominadas: piruvato deshidrogenasa propiamente dicha (E1), dihidrolipoil transacetilasa (E2)
y dihidrolipoil deshidrogenasa (E3). Cinco coenzimas diferentes o grupos prostéticos participan
de la reacción de la piruvato deshidrogenasa: pirofosfato de tiamina, ácido lipoico, coenzima A,
FAD y NAD+
(ver capítulo de reacciones rédox y cadena de transporte de electrones). La E1 tiene
covalentemente unido pirofosfato de tiamina, la E2 tiene covalentemente unido ácido lipoico y la
E3 tiene covalentemente unido FAD.
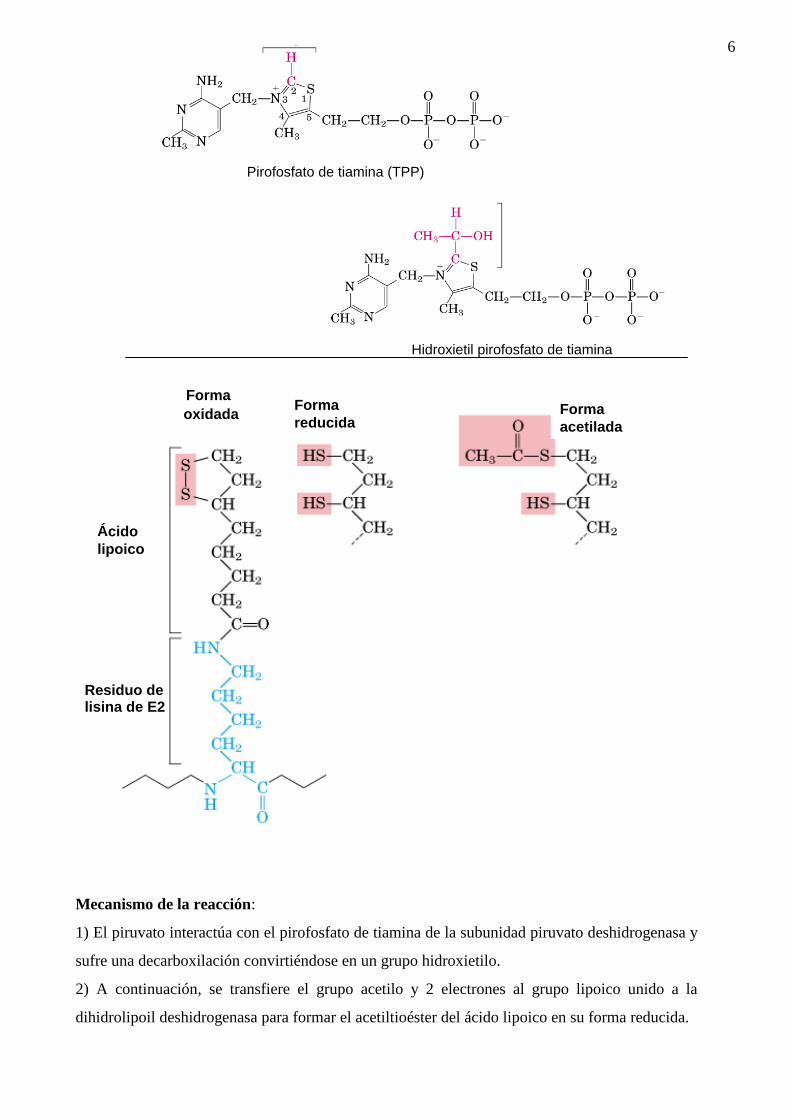
En la siguiente figura se indican las estructuras del pirofosfato la reacción catalizada por el
complejo de la piruvato deshidrogenasa.
6
Mecanismo de la reacción:
1) El piruvato interactúa con el pirofosfato de tiamina de la subunidad piruvato deshidrogenasa y
sufre una decarboxilación convirtiéndose en un grupo hidroxietilo.
2) A continuación, se transfiere el grupo acetilo y 2 electrones al grupo lipoico unido a la
dihidrolipoil deshidrogenasa para formar el acetiltioéster del ácido lipoico en su forma reducida.
Forma acetilada
Forma reducida
Forma oxidada
Á cido lipoico
Residuo de lisina sw
Forma acetilada
Forma acetilada
Forma reducida
Forma
oxidada
Á cido lipoico
Residuo de lisina de E2
E2
sw
Forma acetilada
Pirofosfato de tiamina (TPP)
Hidroxietil pirofosfato de tiamina
Pirofosfato de tiamina (TPP)
Hidroxietil pirofosfato de tiamina
7
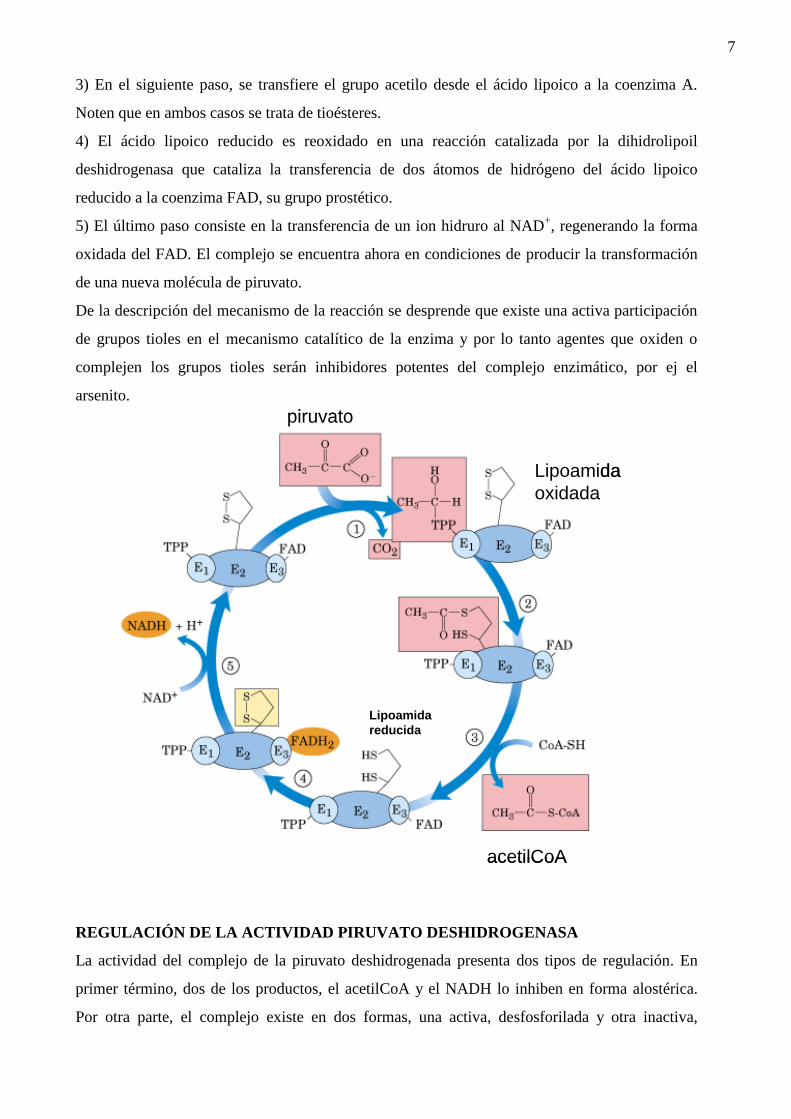
3) En el siguiente paso, se transfiere el grupo acetilo desde el ácido lipoico a la coenzima A.
Noten que en ambos casos se trata de tioésteres.
4) El ácido lipoico reducido es reoxidado en una reacción catalizada por la dihidrolipoil
deshidrogenasa que cataliza la transferencia de dos átomos de hidrógeno del ácido lipoico
reducido a la coenzima FAD, su grupo prostético.
5) El último paso consiste en la transferencia de un ion hidruro al NAD+, regenerando la forma
oxidada del FAD. El complejo se encuentra ahora en condiciones de producir la transformación
de una nueva molécula de piruvato.
De la descripción del mecanismo de la reacción se desprende que existe una activa participación
de grupos tioles en el mecanismo catalítico de la enzima y por lo tanto agentes que oxiden o
complejen los grupos tioles serán inhibidores potentes del complejo enzimático, por ej el
arsenito.
REGULACIÓN DE LA ACTIVIDAD PIRUVATO DESHIDROGENASA
La actividad del complejo de la piruvato deshidrogenada presenta dos tipos de regulación. En
primer término, dos de los productos, el acetilCoA y el NADH lo inhiben en forma alostérica.
Por otra parte, el complejo existe en dos formas, una activa, desfosforilada y otra inactiva,
piruvato
Lipoamida
oxidada
acetilCoA
Lipoamida
reducida
piruvato
Lipoamida
oxidada
acetilCoA
Lipoamida
reducida

8
fosforilada. La inactivación del complejo es catalizada por una proteína quinasa dependiente de
ATP-Mg2+
que está fuertemente unida al complejo. La reactivación del complejo está catalizada
por una fosfoproteína fosfatasa, que desfosforila al complejo en forma dependiente de Mg2+
y de
Ca2+
. Tres residuos diferentes en la subunidad de la piruvato deshidrogenasa se fosforilan por
la proteína quinasa, pero la fosforilación de sólo uno de ellos está relacionada con la actividad
del complejo. La regulación diferencial de la quinasa y de la fosfatasa es la clave de la regulación
de la actividad del complejo.
La formación de acetilCoA está coordinada con la velocidad de utilización del ATP. Esto es
consecuencia del efecto que tienen en su actividad las variaciones en las concentraciones de
ADP, piruvato, CoASH, NAD+,
acetilCoA y del Ca2+
intramitocondrial. Todos estos compuestos
regulan la conversión de la enzima a la forma inactiva (fosforilada). La quinasa que cataliza su
fosforilación es inhibida por ADP, cuyos niveles aumentan cuando aumenta el consumo de ATP.
Esta inhibición mantiene a la piruvato deshidrogenasa en la forma activa, no fosforilada. En
algunos tejidos, el aumento del Ca2+
intramitocondrial que estimula la fosfatasa, contribuye al
incremento de la forma defosforilada y por lo tanto, activa. El acetilCoA y el NADH, productos
de la reacción, inhiben a la forma desfosforilada (activa) de la enzima, y también activan a la
quinasa, llevando a un desplazamiento del complejo a su forma inactiva. Además, la CoASH y el
NAD+ inhiben a la quinasa. Por lo tanto, cuando se produce un aumento en las relaciones
NADH/NAD+ o acetilCoA/CoA (como por ejemplo durante la -oxidación de ácidos grasos,
situación en la que se produce un gran aporte de sustratos al CAT), la actividad del complejo
enzimático disminuye marcadamente. Además, el piruvato es un poderoso inhibidor de la
quinasa, y por lo tanto, en presencia de altas concentraciones de piruvato, la quinasa estará
inhibida y el complejo tendrá su máxima actividad. Finalmente, se ha demostrado que la
administración de insulina puede activar a la piruvato deshidrogenasa del tejido adiposo y que las
catecolaminas (como la adrenalina) pueden activar la del tejido cardíaco. Los mecanismos aún no
están aclarados completamente, pero podría estar involucrada una alteración en la distribución
intracelular de calcio, de modo de afectar a la fosfoproteína fosfatasa. Estos efectos hormonales
no están mediados directamente por alteraciones en los niveles de AMPc dado que tanto la
quinasa como la fosfatasa del complejo son independientes de AMPc.

9
CoASH
Piruvato
NAD+CO2
Acetil CoA
NADH + H+
PDH inactiva
PDH activa
Pi
ATP
ADPquinasa fosfatasaMg2+ Ca2+ Mg2+
(-)
(-)
(-)
(-)
(+)
Pi
(-)
En conclusión, la actividad del complejo enzimático aumenta cuando se requiere un aporte de
acetilCoA al CAT o cuando aumentan sus sustratos, aunque en ese último caso el destino final
del acetilCoA no será únicamente el ciclo.
AcetilCoA
Como ya dijimos, la mayor parte de las principales vías catabólicas generan la unidad de dos
carbonos del acetilCoA. El catabolismo de los glúcidos almacenados o ingeridos, a través de la
vía glucolítica, el de los ácidos grasos de cadena larga provenientes de la lipólisis de los
triacilglicéridos por la secuencia de -oxidación, la oxidación de cuerpos cetónicos y de etanol o
el catabolismo de ciertos aminoácidos, producto de proteólisis, luego de su transaminación o
desaminación y posterior oxidación proveen precursores para la formación de acetilCoA.
La coenzima A, cuya estructura se muestra en la Figura, se compone de beta-mercaptoetilamina,
la vitamina ácido pantoténico y un nucleótido de adenina, adenosina 3’-fosfato 5’-disfosfato. Su
grupo tiol se encuentra reducido y está involucrada en muchas reacciones de transferencia de
grupos acilo en las que la coenzima A alternativamente funciona como el aceptor y luego el
dador de la función acilo.

10
S C CH3
O
CH2
CH2
NH
C O
CH2
CH2
NH
OC
C
C
CH2
HO H
CH3H3C
CH2O
NH2
N
N
P O P O
O O
O O- -
-O P O
O
O-
1'
2'3'
4'
5'
3' fosfoadenosina difosfato
D-ribosa 3' fosfato
adenina
grupo acetilo
beta-mercaptoetilamina
ácido pantoténico
ACETIL COENZIMA A
N
N
En muchas vías metabólicas, todos los intermediarios son compuestos donde participa la
coenzima A, como por ejemplo, en la -oxidación de ácidos grasos y en la degradación de
aminoácidos ramificados. Como muchos otros nucleótidos, los derivados de la CoASH no se
transportan libremente a través de las membranas celulares, sino que lo hacen a través de
transportadores o mecanismos específicos. También es importante señalar que el enlace tioéster
del acetilCoA es un enlace rico en energía, y por lo tanto estos compuestos pueden servir como
dadores eficientes de grupos acilo en reacciones de transferencia de acilo. En la síntesis de
acetilCoA catalizada por la enzima acetato tioquinasa debe utilizarse una molécula de ATP, pero
la reacción se produce con gasto de 2 enlaces de alta energía:
acetato + CoASH + ATP acetilCoA + AMP + PPi
El destino del acetilCoA generado en la mitocondria puede ser:
a) oxidación completa del grupo acetilo en el CAT con generación de energía,
b) conversión del exceso de acetilCoA a cuerpos cetónicos: acetoacetato y -hidroxibutirato (en
hígado)
c) transferencia de acetilos al citosol con la subsecuente biosíntesis de moléculas complejas
como esteroles y ácidos grasos de cadena larga.
CICLO DE LOS ÁCIDOS TRICARBOXÍLICOS

11
Como ya mencionamos, en la mayoría de las vías de oxidación de combustibles metabólicos se
terminan formando los dos carbonos activados del acetilCoA. El destino principal del acetilCoA
es su oxidación completa a CO2 en una serie de reacciones oxidativas cíclicas, denominadas ciclo
de los ácidos tricarboxílicos (CAT). También se lo conoce como ciclo del ácido cítrico o ciclo de
Krebs, en homenaje al investigador que postuló las características esenciales del ciclo en el año
1937. La localización de las enzimas del ciclo es la mitocondria, lo que coincide con la
localización del complejo de la piruvato deshidrogenasa y las enzimas de la -oxidación, las dos
fuentes principales de acetilCoA. Una de las funciones principales del ciclo es la generación de
equivalentes de reducción, que se utilizan para generar energía (ATP), en la secuencia de
transporte de electrones y fosforilación oxidativa, procesos que también ocurren en las
mitocondrias. La oxidación ocurre en cuatro reacciones que transfieren electrones a las
coenzimas aceptoras NAD+ o FAD. En otras reacciones del ciclo se rearreglan los electrones para
facilitar esta transferencia. Inicialmente la porción acetilo del acetilCoA se combina con el
intermediario de cuatro carbonos oxaloacetato para formar citrato (6 carbonos). Un rearreglo
subsecuente de enlaces en el citrato es seguido por dos decarboxilaciones oxidativas. Estas
reacciones transfieren los electrones al NAD+ y liberan 2 carbonos como CO2. En el siguiente
paso, se genera un enlace de alta energía del GTP por fosforilación a nivel de sustrato. En la
porción remanente del ciclo hay dos reacciones de transferencia electrónica adicionales y se
regenera el oxaloacetato. El proceso completo ocurre con la conservación de la mayor parte de la
energía de los enlaces químicos del grupo acetilo como 3 NADH, 1 FADH2 y 1 GTP.
Aproximadamente 2/3 del NADH y del FADH2 generados en todas las vías de oxidación de
combustibles provienen del ciclo. La velocidad del ciclo está fuertemente coordinada a la
velocidad de utilización de ATP a través de la regulación de enzimas individuales por
compuestos relacionados con el ATP, por el estado de reducción del NAD+ y por la
concentración mitocondrial de Ca2+
. Otra de las características del CAT es la existencia de
“puntos de fuga”, es decir reacciones en las cuales algunos de los intermediarios del ciclo pueden
“salir” del ciclo y actuar como sustratos de otras reacciones ajenas a éste.
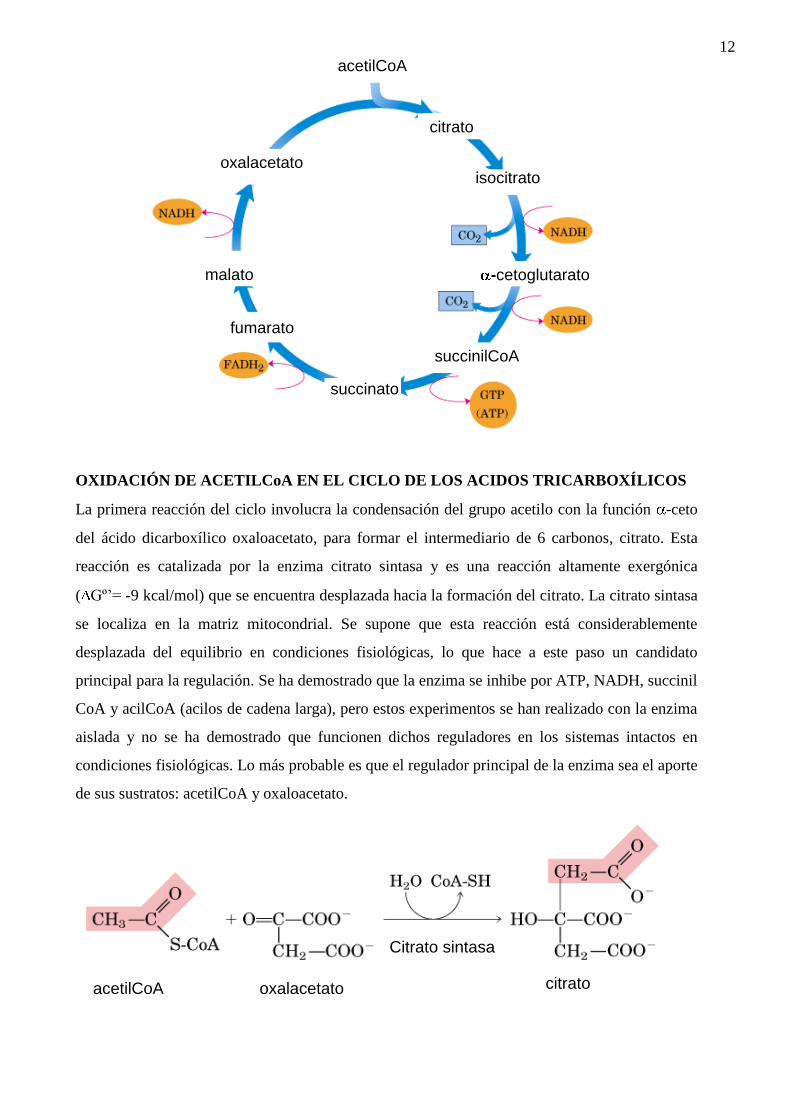
12
OXIDACIÓN DE ACETILCoA EN EL CICLO DE LOS ACIDOS TRICARBOXÍLICOS
La primera reacción del ciclo involucra la condensación del grupo acetilo con la función -ceto
del ácido dicarboxílico oxaloacetato, para formar el intermediario de 6 carbonos, citrato. Esta
reacción es catalizada por la enzima citrato sintasa y es una reacción altamente exergónica
( Gº’= -9 kcal/mol) que se encuentra desplazada hacia la formación del citrato. La citrato sintasa
se localiza en la matriz mitocondrial. Se supone que esta reacción está considerablemente
desplazada del equilibrio en condiciones fisiológicas, lo que hace a este paso un candidato
principal para la regulación. Se ha demostrado que la enzima se inhibe por ATP, NADH, succinil
CoA y acilCoA (acilos de cadena larga), pero estos experimentos se han realizado con la enzima
aislada y no se ha demostrado que funcionen dichos reguladores en los sistemas intactos en
condiciones fisiológicas. Lo más probable es que el regulador principal de la enzima sea el aporte
de sus sustratos: acetilCoA y oxaloacetato.
acetilCoA
citrato
isocitrato
-cetoglutarato
succinilCoA
succinato
fumarato
malato
oxalacetato
acetilCoA
citrato
isocitrato
-cetoglutarato
succinilCoA
succinato
fumarato
malato
oxalacetato
acetilCoA citrato
Citrato sintasa
oxalacetatoacetilCoA citrato
Citrato sintasa
oxalacetato

13
El citrato, que es el primer producto del ciclo, es también el primer “punto de fuga” dado que
además de intermediario en el ciclo, puede funcionar como efector regulatorio o como fuente de
carbonos en otras vías metabólicas que ocurren en el citosol:
CITRATO
Fuente de equivalentes Fuente de carbonos para la
de reducción para procesos biosíntesis citosólica de ácidos grasos y esteroles
de síntesis citosólicos
Regulador de actividad enzimática:
(-) de la fosfofructoquinasa I
(+) de la acetilCoA carboxilasa
En el siguiente paso del ciclo, catalizado por la aconitasa, se forma isocitrato por la transferencia
de un grupo hidroxilo a un carbono adyacente que posee un enlace C-H.
Esta reacción involucra la generación de un intermediario unido a la enzima, el cis-aconitato. En
el equilibrio existe un 90% de citrato, 3% de cis-aconitato y un 7% de isocitrato. Por lo tanto, en
el equilibrio, predomina el citrato. Aunque la reacción no requiere cofactores, requiere hierro
(Fe2+
) y existe evidencia de que éste estaría involucrado en un centro Fe-S, componente esencial
de la actividad hidratasa de la aconitasa.
La isocitrato deshidrogenasa cataliza la primera reacción de deshidrogenación del ciclo. El
isocitrato se convierte en -cetoglutarato en una reacción de descarboxilación oxidativa. En este
paso del ciclo se produce el primero (de dos) CO2 y se genera el primero (de tres) NADH + H+.
citrato Cis-aconitato isocitrato
aconitasaaconitasa
citrato Cis-aconitato isocitrato
aconitasaaconitasa

14
La enzima requiere NAD+ como aceptor oxidado para los equivalentes de reducción. Si bien
existe otra actividad de isocitrato deshidrogenasa en el citosol que requiere NADP+ como
coenzima oxidada, no es ésta la que participa del ciclo y su función en el citosol sería la de
proveer equivalentes de reducción para procesos citosólicos reductivos. El equilibrio de la
reacción está muy desplazado hacia la formación de -cetoglutarato con un Gº'= -5 kcal/mol.
La enzima tiene un peso molecular de 380.000 y está constituida por 8 subunidades idénticas. Se
requiere un metal divalente (Mn2+
o Mg2+
) para la descarboxilación de la posición beta del
isocitrato. La enzima se activa por ADP y en algunos casos por AMP y se inhibe por ATP y
NADH. Por lo tanto, en condiciones de alta energía (alto ATP/ADP + Pi y alto NADH/NAD+) se
inhibe. En situaciones de baja energía, la actividad de la enzima se estimula, lo que permite
acelerar la generación de energía por el ciclo. El -cetoglutarato es otro “punto de fuga” del
CAT, porque puede ser sustrato de una reacción que intersecta el ciclo, en la cual el -
cetoglutarato se condensa con amoníaco (o amonio) en presencia de NADH o NADPH y se
sintetiza glutamato y NAD+ o NADP
+. Esta reacción ocurre en las mitocondrias, y es catalizada
por la enzima glutamato-deshidrogenasa, una de las enzimas más importantes en el metabolismo
de aminoácidos
GLUTAMATO DESHIDROGENASA
NH4+ + H
CH2
C
COO-
2
COO-
C O
NAD(P)H NAD(P)+
H
CH2
C
COO-
2
C NH3
+
-COO
ALFA-CETOGLUTARATO GLUTAMATO
isocitrato-cetoglutarato
Isocitrato-deshidrogenasa
isocitrato-cetoglutarato
Isocitrato-deshidrogenasa

15
La conversión de -cetoglutarato a succinil-CoA es catalizada por el complejo multienzimático
de la -ceto-glutarato deshidrogenasa. Este complejo está compuesto por 3 actividades
enzimáticas diferentes: alfa-cetoglutarato deshidrogenasa, dihidrolipoil transuccinilasa y
dihidrolipoil deshidrogenasa. Su peso molecular es de varios millones de daltons y pertenece a
una familia de complejos similares que descarboxilan oxidativamente al -cetoglutarato, al
piruvato, al -cetobutirato y a los -cetoácidos de los aminoácidos de cadena ramificada (valina,
leucina e isoleucina).
Este complejo es casi idéntico al de la piruvato deshidrogenasa en cuanto al tipo de reacción que
cataliza y a algunas de sus características estructurales. También en este caso, el pirofosfato de
tiamina, el ácido lipoico, la CoASH, el FAD y el NAD+ participan del mecanismo catalítico. El
equilibrio está desplazado hacia la formación de succinil-CoA con un Gº’=-8 kcal/mol. En esta
reacción se genera la segunda molécula de CO2 y el segundo equivalente de reducción del ciclo
(NADH + H+). El otro producto, el succinil-CoA, es un ejemplo de tioéster rico en energía
similar al acetilCoA. A diferencia del complejo de la piruvato deshidrogenasa, la actividad de
este complejo no está regulada por fosforilación. Los nucleósidos trifosfato, ATP y GTP, NADH
y succinil-CoA inhiben el complejo, mientras que el aumento en la concentración de Ca2+
lo
activa. La energía del enlace tioéster del succinil-CoA se conserva en una reacción de
fosforilación a nivel de sustrato en el siguiente paso del ciclo. Una reacción de fosforilación a
nivel de sustrato es la formación de un nuevo enlace de fosfato de alta energía en una reacción en
la que no participa directamente el oxígeno. La reacción catalizada por la succinil-CoA sintetasa
o succinato tioquinasa convierte el succinilCoA a succinato y en tejidos de mamíferos resulta en
la fosforilación del GDP para dar GTP. La reacción es libremente reversible con un Gº’= -0.7
kcal/mol. Dada la presencia de la nucleósido difosfato quinasa, el grupo fosfato del GTP puede
generar ATP por transferencia al ADP.
-cetoglutarato Succinil-CoA
-cetoglutarato deshidrogenasa
-cetoglutarato Succinil-CoA
-cetoglutarato deshidrogenasa

16
NDQ (Mg2+
)
ADP + GTP <===>ATP + GDP
El succinilCoA es otro punto de ramificación (de fuga) metabólica, pues los intermediarios
pueden entrar o salir del ciclo a partir de este intermediario. El succinil-CoA puede formarse a
partir de -cetoglutarato o a partir de metilmalonil-CoA en los pasos finales de la degradación de
ácidos grasos de número impar de carbonos o de aminoácidos ramificados como valina e
isoleucina. Su destino metabólico incluye su conversión a succinato por la reacción de la
succinil-CoA sintetasa y su condensación con glicina para formar el -aminolevulinato en la
reacción catalizada por la -aminolevulinato sintetasa, que es la reacción inicial de la síntesis de
porfirinas.
Propionil CoA-cetoglutarato
Metilmalonil CoA Ciclo de Krebs
Metabolismo de ácidos
grasos de nº impar de C y
de aminoácidos ramificados
Succinil CoA
Ciclo de Krebs Acetoacetato
Utilización de cuerpos Biosíntesis de Porfirinascetónicos
AcetoacetilCoA
Succinato -aminolevulinato
Propionil CoA-cetoglutarato
Metilmalonil CoA Ciclo de Krebs
Metabolismo de ácidos
grasos de nº impar de C y
de aminoácidos ramificados
Succinil CoA
Ciclo de Krebs Acetoacetato
Utilización de cuerpos Biosíntesis de Porfirinascetónicos
AcetoacetilCoA
Succinato -aminolevulinato
SuccinilCoA
SuccinilCoA sintetasa
Succinato
o
SuccinilCoA
Succinil-CoA sintetasa o Succinato tioquinasa
Succinato
o

17
El succinato se oxida a fumarato en la reacción catalizada por la succinato
deshidrogenasa. Se transfiere un electrón de cada grupo metileno adyacente del succinato a un
FAD unido a la succinato deshidrogenasa, formándose entonces el doble enlace del fumarato.
Del FAD reducido unido a la enzima, los electrones pasan a la cadena de transporte. La enzima
está fuertemente unida a la membrana mitocondrial interna y está compuesta por dos
subunidades. La mayor de las subunidades contiene el sitio de unión del sustrato, el FAD
covalentemente unido a una lisina, varios átomos de hierro no-hémico y de azufre lábiles,
mientras que la menor contiene hierros no-hémicos y azufres lábiles. Esta enzima es un ejemplo
típico de una proteína hierro-azufre en la cual el hierro no-hémico sufre un cambio de valencia
durante la remoción de los electrones y de los protones del succinato y la subsecuente
transferencia de estos equivalentes de reducción por el FAD covalentemente unido a la cadena de
transporte de electrones a nivel de la coenzima Q-citocromo b. La succinato deshidrogenasa es
inhibida fuertemente por malonato y oxaloacetato y es activada por ATP y succinato. El
malonato es un inhibidor competitivo debido a la similitud estructural con el succinato.
El fumarato se hidrata para formar L-malato en el siguiente paso del ciclo, reacción catalizada
por la enzima fumarasa. Un grupo OH- y un protón del agua se agregan al doble enlace del
fumarato transformándolo en malato. La fumarasa es un tetrámero de PM 200.000 y es
estereoespecífica para la forma trans del sustrato (la forma cis, el maleato o ácido maleico, no es
su sustrato) y el producto de la reacción sólo es L-malato (no su isómero, el D-malato). La
reacción es libremente reversible en condiciones fisiológicas.
Succinato
Succinato deshidrogenasa
Fumarato Succinato
Succinato deshidrogenasa
Fumarato

18
La última reacción del ciclo es catalizada por la malato deshidrogenasa, en la que se remueve el
último (de tres) de los equivalentes de reducción como NADH + H+. El grupo alcohol del malato
se oxida a grupo ceto cediendo sus electrones a la coezima.
En el equilibrio predomina el malato, dado que para esta reacción el Gº’= + 7.0 kcal/mol. La
reacción es por lo tanto endergónica considerada en la dirección de formación de oxaloacetato.
Sin embargo, la reacción catalizada por la citrato sintasa y las otras del ciclo permiten la
formación del oxaloacetato pues mantienen sus niveles muy bajos. Además, el NADH producido
se oxida rápidamente en la cadena de transporte de electrones. El malato constituye otro punto de
ramificación, pues puede ser transportado al citosol, donde genera oxaloacetato en la reacción
catalizada por la enzima malato deshidrogenasa citosólica. Éste, a su vez, es un sustrato
gluconeogénico.
Es importante destacar que esta última secuencia de reacciones: oxidación por formación de un
doble enlace, adición de agua al doble enlace y oxidación del alcohol resultante a cetona, se
Fumarato (trans) L-malato
fumarasa
Fumarato (trans) L-malato
fumarasa
L-Malato
Malato deshidrogenasa
OxalacetatoL-Malato
Malato deshidrogenasa
Oxalacetato

19
encuentra en muchas vías metabólicas tales como la oxidación de los ácidos grasos y de los
aminoácidos ramificados.
¿Qué transformación neta ocurre entonces en el ciclo? En primer lugar, el grupo acetilo de dos
carbonos del acetilCoA se oxida generando 2 moléculas de CO2. Una función del ciclo es
conservar la energía de esta oxidación en forma de coenzimas transportadoras de electrones en
estado reducido. El balance global del ciclo muestra que el grupo acetilo funciona como fuente
de carbonos para el CO2 y como fuente de electrones para la transferencia a las coenzimas.
Acetil CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O 2 CO2 + CoASH + 3 NADH + 3 H
+ +
FADH2 + GTP
El acetilCoA es el combustible del ciclo, su oxidación no involucra O2 directamente, y se lleva a
cabo removiendo electrones como parte del hidrógeno o del ión hidruro y agregando H2O. Los 4
oxígenos de los 2 CO2 provienen del grupo carbonilo del acetilCoA, 2 H2O y la adición de un
PO43-
al GDP. Las diferentes reacciones del ciclo involucran el rearreglo de los carbonos y los
electrones donados por el grupo acetilo de modo que los mismos carbonos y electrones no entran
y salen del ciclo en una sola vuelta.

20
En el esquema se muestran marcados (en rojo) los C que provienen del AcetilCoA y como se
observa, en la primera vuelta del ciclo, estos C no abandonan el ciclo como CO2 sino que quedan
en el oxalacetato.
El rendimiento completo en compuestos que contienen energía es de 3 NADH, 1 FADH2 y 1
GTP. Las reacciones rédox son catalizadas por 4 deshidrogenasas: isocitrato deshidrogenasa,
alfa-cetoglutarato deshidrogenasa, succinato deshidrogenasa y malato deshidrogenasa. El enlace
de alta energía del GTP se origina en una reacción de fosforilación a nivel de sustrato catalizada
por la succinato tioquinasa.
Coenzimas
Diferencias entre NAD+
y FAD
En la oxidación del succinato a fumarato y en la oxidación del lipoato reducido a lipoato
disulfuro, el FAD es el aceptor de electrones. En las otras oxidaciones esa función la cumple el
NAD+.
¿Por qué? Estas coenzimas tienen diferentes propiedades y realizan funciones diferentes.
El FAD puede aceptar electrones individuales (H.) y formar un intermediario semirreducido con
un electrón. Por lo tanto, participa en reacciones donde se transfieren electrones individuales
desde dos átomos diferentes tales como dos C-H adyacentes en el succinato o los dos grupos SH
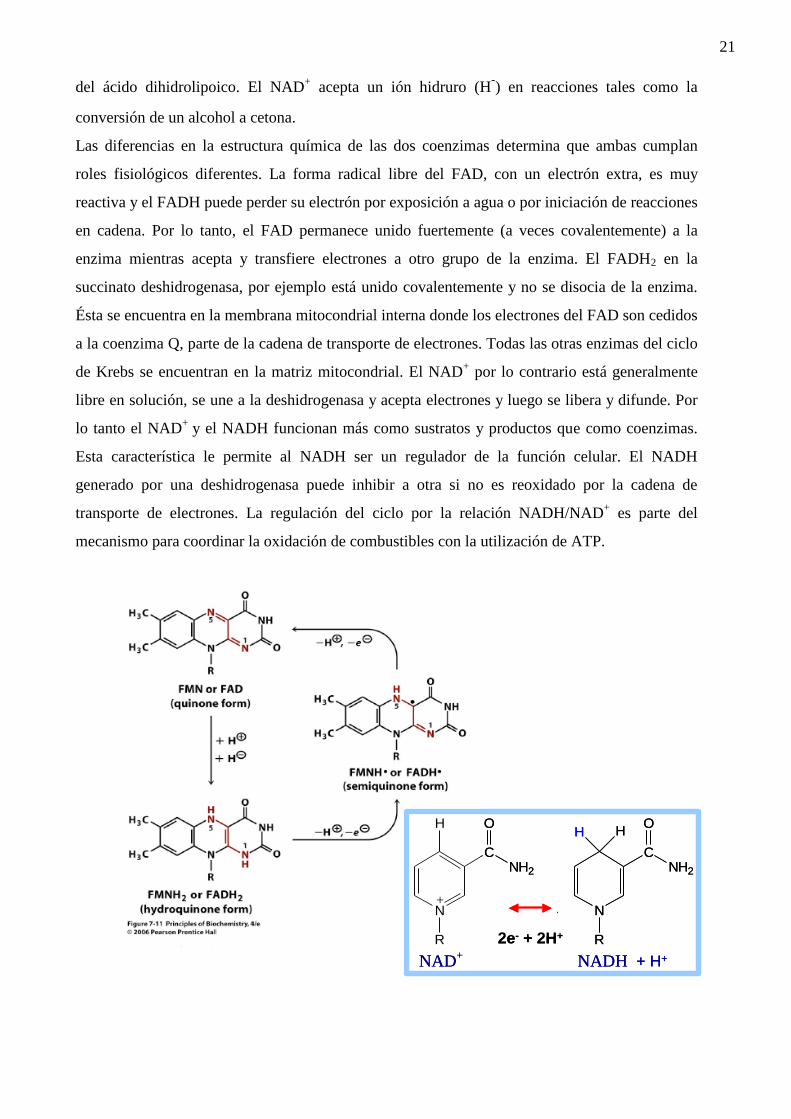
21
del ácido dihidrolipoico. El NAD+ acepta un ión hidruro (H
-) en reacciones tales como la
conversión de un alcohol a cetona.
Las diferencias en la estructura química de las dos coenzimas determina que ambas cumplan
roles fisiológicos diferentes. La forma radical libre del FAD, con un electrón extra, es muy
reactiva y el FADH puede perder su electrón por exposición a agua o por iniciación de reacciones
en cadena. Por lo tanto, el FAD permanece unido fuertemente (a veces covalentemente) a la
enzima mientras acepta y transfiere electrones a otro grupo de la enzima. El FADH2 en la
succinato deshidrogenasa, por ejemplo está unido covalentemente y no se disocia de la enzima.
Ésta se encuentra en la membrana mitocondrial interna donde los electrones del FAD son cedidos
a la coenzima Q, parte de la cadena de transporte de electrones. Todas las otras enzimas del ciclo
de Krebs se encuentran en la matriz mitocondrial. El NAD+ por lo contrario está generalmente
libre en solución, se une a la deshidrogenasa y acepta electrones y luego se libera y difunde. Por
lo tanto el NAD+
y el NADH funcionan más como sustratos y productos que como coenzimas.
Esta característica le permite al NADH ser un regulador de la función celular. El NADH
generado por una deshidrogenasa puede inhibir a otra si no es reoxidado por la cadena de
transporte de electrones. La regulación del ciclo por la relación NADH/NAD+ es parte del
mecanismo para coordinar la oxidación de combustibles con la utilización de ATP.
N
R
H
CNH2
O
N
R
CNH2
OH H
+
2e + H+
NAD+ NADH
2e- + 2H+
+ H+
N
R
H
CNH2
O
N
R
CNH2
OH H
+
2e + H+
NAD+ NADH
2e- + 2H+
+ H+

22
Coenzima A
La CoASH, la coenzima de acilación, participa en reacciones en las que se forma un enlace
tioéster con un grupo acilo (acetilCoA, succinil-CoA). El carbono carbonílico, el carbono alfa y
el carbono beta del grupo acilo se activan en diferentes tipos de reacciones por la formación del
enlace tioéster con el grupo sulfhidrilo de la coenzima A. La hidrólisis de este enlace tiene un
gran valor negativo del Gº’ (aprox. -13 kcal/mol). El enlace tioéster es mucho menos estable
que un enlace éster con oxígeno, pues el azufre no comparte sus electrones en estados de
resonancia. La energía liberada por la ruptura del tioéster cumple dos funciones en el CAT:
provee la energía para la generación del GTP y hace que la entrada del acetilCoA al ciclo sea más
favorable termodinámicamente.
En la condensación de la porción acetilo con el oxaloacetato para formar citrato, la ruptura del
enlace tioéster del acetilCoA ocurre con un Gº’= -7.7 kcal/mol. Cuando este valor negativo se
agrega al gran valor positivo para la reacción de la malato deshidrogenasa, se obtiene un valor
neto cercano a 0 para la conversión de malato a citrato. Por lo tanto la entrada del acetilCoa al
ciclo se facilita por la ruptura del enlace tioéster.
Dado que el enlace tioéster del acetilCoA tiene un Gº’ de hidrólisis de -13 kcal, su formación
requiere energía, como ya se mencionó. Cuando se forma el acetilCoA por la oxidación de la
glucosa, la energía es aportada por la descarboxilación oxidativa del piruvato en la reacción
catalizada por el complejo de la piruvato deshidrogenasa. Cuando el acetilCoA proviene de las
vías de degradación de ácidos grasos, cuerpos cetónicos o aminoácidos, la energía proviene de
reacciones de oxidación y ruptura de enlaces C-C. El acetato también puede convertirse
directamente en acetilCoA utilizando la energía aportada por el ATP, en una reacción catalizada
por la acetato tioquinasa, que ya se describió.
BALANCE ENERGETICO DEL CICLO DE KREBS
Las reacciones del ciclo son extremadamente eficientes en la conversión de la energía de las
uniones químicas del acetato. La energía total contenida en el acetato, liberada como calor en la
combustión completa de un mol de acetato a CO2 en un calorímetro, es de 243 kcal/mol. Para
activar el acetato a acetilCoA se requiere el equivalente a dos enlaces de alta energía o alrededor
de 15 kcal/mol, de modo que sólo se producen 228 kcal/mol de la oxidación del acetato. Los
productos del ciclo almacenan alrededor de 206 kcal/mol, por lo tanto las reacciones del ciclo
son capaces de conservar alrededor del 90% de la energía accesible de la oxidación del
acetil CoA.

23
Tres reacciones del ciclo tienen valores muy negativos de Gº’: son las catalizadas por la citrato
sintasa, la isocitrato deshidrogenasa y la -cetoglutarato deshidrogenasa. Estas reacciones son
irreversibles en condiciones fisiológicas debido a sus altos valores (negativos) de Gº’ y porque
las enzimas involucradas catalizan las reacciones inversas muy lentamente. Estas reacciones
realizan la contribución principal al elevado Gº’ negativo del ciclo.
Enzima Gº’ (kcal/mol)
Citrato Sintasa -7.7
Aconitasa + 1.5
Isocitrato Deshidrogenasa -5.3
-Cetoglutarato
Deshidrogenasa
-8.0
Succinato Tioquinasa -0.7
Succinato Deshidrogenasa 0
Fumarasa 0
Malato Deshidrogenasa + 7.1
Como ya hemos mencionado, durante el proceso de oxidación completa del grupo acetilo se
producen 4 equivalentes de reducción, que se usan subsecuentemente para generar energía. Como
se discutirá más adelante, la oxidación de cada mol de NADH + H+ resulta en la formación de
2,5 moles de ATP mientras que la de FADH2 en 1,5. También se forma un enlace de alta energía
en la reacción de la succinil-CoA sintetasa. Por lo tanto la ganancia neta para la oxidación de
cada mol de acetilCoA en el CAT es de 10 moles de ATP.
REACCIONES ANAPLEROTICAS
Teniendo en cuenta los ya mencionados “puntos de fuga” del ciclo, para que éste siga
funcionando los tejidos aportan intermediarios para compensar su salida hacia otras vías como la
gluconeogénesis o la síntesis de ácidos grasos. En cada tejido, los puntos de fuga intersectan al
ciclo y remueven intermediarios como el citrato, el -cetoglutarato, el succinil-CoA y el malato.
En el tejido nervioso, el -cetoglutarato se convierte en glutamato y luego en GABA. El succinil-
CoA se utiliza para la síntesis de hemo en el hígado. Los esqueletos carbonados de algunos
aminoácidos también derivan de intermediarios del ciclo. El oxaloacetato que es imprescindible
para la oxidación del acetilCoA, siempre se regenera: en cada ciclo, 2 carbonos se incorporan en
la forma del grupo acetilo, 2 se eliminan como CO2, y la unidad de 4 carbonos está presente en el

24
succinato, fumarato, malato y oxaloacetato. Sin embargo, si un intermediario entra en otra vía, el
nivel de intermediarios debe restaurarse para que el ciclo (y con ello la obtención de energía) no
se detenga.
Las reacciones que aportan intermediarios al CAT se denominan reacciones anapleróticas o de
relleno. Una de las principales reacciones anapleróticas es la conversión de piruvato y CO2 a
oxaloacetato, catalizada por la piruvato carboxilasa. Esta enzima contiene biotina, que en
presencia de ATP y Mg2+
forma un intermediario covalente con el CO2 que se agrega luego como
grupo carboxilo al piruvato, para formar oxaloacetato. Las carboxilasas de otras vías metabólicas
también requieren biotina y su mecanismo de acción es similar. El acetilCoA es un efector
alostérico positivo de la piruvato carboxilasa.
PIRUVATO CARBOXILASA
COO- COO
-
ATP + HCO3- + C=O C=O + ADP + Pi
CH3 CH2
COO-
PIRUVATO OXALOACETATO
La piruvato carboxilasa se encuentra en altas concentraciones en el hígado y el tejido nervioso ya
que estos tejidos tienen un constante eflujo de intermediarios. Como la mayoría de las reacciones
anapleróticas, esta reacción forma parte de una vía que intersecta al ciclo. En el hígado, es parte
de la vía gluconeogénica que convierte alanina y lactato en glucosa.
A partir de ácidos grasos con número impar de C se obtiene propionilCoA (ver reacciones de la
-oxidación). Este intermediario puede convertirse en succinilCoA en una reacción anaplerótica

25
Los aminoácidos son otra fuente de intermediarios para el CAT. La vía que convierte isoleucina,
valina, metionina y otros compuestos en succinil-CoA es una ruta anaplerótica importante. La
mayor parte de los tejidos, incluidos cerebro, músculo esquelético, y corazón degradan isoleucina
y valina a succinil-CoA por esta ruta. En el hígado, el ciclo es parte de la vía que convierte valina
e isoleucina en glucosa, en el músculo esquelético es parte de la vía que los convierten en
glutamina.
La reacción catalizada por la glutamato deshidrogenasa, que ya hemos mencionado, cataliza la
conversión reversible de -cetoglutarato en glutamato y también es un buen ejemplo de reacción
anaplerótica. En cambio la reacción catalizada por la glutamato-oxaloacetato transaminasa no es
una reacción anaplerótica. ¿Por qué?
Otra reacción anaplerótica es la catalizada por la enzima málica (diferente de la enzima málico
deshidrogenasa) que convierte el piruvato en malato, consume NADPH y requiere del aporte de
1C como CO2.
PropionilCoAPropionil-CoA
D-Metilmalonil-CoA
L-Metilmalonil-CoA Succinil-CoA
PropionilCoAPropionil-CoA
D-Metilmalonil-CoA
L-Metilmalonil-CoA Succinil-CoA

26
ENZIMA MALICA
COO-
N ADPH + H
+ NADP
+ COO
-
C=O + CO2 HO C H
CH3 COO-
REGULACION DEL CICLO DE LOS ACIDOS TRICARBOXILICOS
La oxidación del acetilCoA y la conservación de la energía en las coenzimas reducidas para la
generación de ATP son esenciales en todos los tejidos que contienen mitocondrias. La velocidad
de utilización de ATP es, por lo tanto, la fuerza fisiológica primaria impulsora del ciclo. Existen
dos señales principales de la velocidad de utilización de ATP: a) el estado de fosforilación de los
nucleótidos de adenina, que se refleja en los niveles de ATP y ADP, y b) el estado de reducción
del NAD+, reflejado en la relación NADH/NAD
+. Dentro de la célula y aún dentro de la
mitocondria, la suma de la concentración de nucleótidos de adenina (AMP, ADP y ATP) y la de
NAD+ (NAD
+ y NADH) son relativamente constantes. Sin embargo, la velocidad de
interconversión de los nucleótidos de adenina varía considerablemente como lo hace la velocidad
de oxidación y reducción de las coenzimas. Por lo tanto, un aumento en la utilización de ATP
resultaría en una pequeña disminución en sus niveles y un aumento en la concentración de ADP.
Análogamente, un aumento en la oxidación de NADH por la cadena de transporte provocaría un
aumento en el contenido de NAD+
y una disminución en el NADH.
Dado que las deshidrogenasas principales son dependientes del aporte continuo de NAD+
y de
FAD, sus actividades son estrictamente controladas por la cadena respiratoria que es responsable
de su oxidación y que está obligatoriamente acoplada a la generación de ATP. De este modo la
actividad del ciclo es muy dependiente del control respiratorio, que a su vez está afectado por el
aporte de ADP + Pi y de oxígeno. Un agente inhibitorio o una condición metabólica que
interrumpa el aporte de oxígeno, el aporte continuo de ADP o la fuente de equivalentes de
reducción, anularán la actividad del ciclo. Este tipo de control se conoce como “control grueso”.
Hay también un número de interacciones mediadas por efectores entre intermediarios,
nucleótidos y las enzimas del ciclo que pueden ejercer un control fino de su actividad.
Los conceptos generales con respecto a los mecanismos de regulación que se aplican a todas las
vías metabólicas pueden ser aplicados también en este caso. La regulación de la velocidad de la
vía se da sobre las enzimas que catalizan los pasos limitantes de la velocidad, o sea los más

27
lentos. Se puede hacer una analogía considerando la velocidad promedio en una autopista cuando
en alguna zona hay un bloqueo que sólo deja libre un carril. En ese caso, el tránsito en esa zona
se hace más lento y esta disminución de la velocidad se transmite hacia el principio de la
autopista. Suponiendo que el lugar del bloqueo constituya la etapa limitante de la velocidad, una
modificación en el mismo, por ejemplo, la apertura de un nuevo carril para el tránsito, aumentará
significativamente la velocidad global en la autopista. En forma semejante, la velocidad de toda
una vía metabólica está determinada por la de la reacción más lenta y por lo tanto la regulación
de la actividad de la vía ocurre principalmente a ese nivel.
Otro de los conceptos generales que también se aplica aquí, es que las vías que deben mantener
un nivel constante de un producto final, como el ATP se regulan por un feedback de ese producto
o un metabolito relacionado. En este caso, ATP, ADP, NADH y NAD+ participan en la
regulación por feedback (o retroalimentación).
Dos de los principales sitios de regulación del ciclo son la isocitrato deshidrogenasa y la -
cetoglutarato deshidrogenasa. La capacidad de ambas enzimas es relativamente baja en
condiciones fisiológicas y por lo tanto constituyen los pasos limitantes. La isocitrato
deshidrogenasa es una enzima oligomérica (formada por varias subunidades) activada
alostéricamente por ADP e inhibida por NADH. En ausencia de ADP exhibe cooperatividad
positiva, cuando el isocitrato se une a la primera subunidad, las otras subunidades se convierten a
una conformación activa. En presencia de ADP cambia la conformación de todas las subunidades
de modo que el isocitrato se une más rapidamente y el Km aparente cambia a un valor mucho
menor. Por lo tanto, a la concentración de isocitrato encontrada en la matriz mitocondrial, un
pequeño cambio en la concentración de ADP puede producir un gran cambio de flujo. Pequeños
cambios en la concentración del producto NADH y del cosustrato NAD+ también afectan la
velocidad de la enzima más de lo que lo harían en una enzima no alostérica.
La -cetoglutarato deshidrogenasa, aunque no es alostérica, es inhibida por los productos NADH
y succinilCoA. Por lo tanto, ambas enzimas responden directamente a cambios en el estado de
fosforilación o de reducción. Ambas enzimas también se activan por aumento en la
concentración de Ca2+
mitocondrial. En el músculo esquelético en contracción y posiblemente en
otros tejidos musculares, la liberación de Ca2+
del retículo sarcoplásmico y el aumento en la
concentración mitocondrial de Ca2+
durante la contracción puede proveer una activación
adicional de estas enzimas cuando el ATP se está hidrolizando rápidamente.
Ninguna de estas enzimas cataliza la primera reacción del ciclo, y por lo tanto no afectan
directamente la entrada del acetilCoA. La citrato sintasa, es una enzima simple en tejidos de
mamíferos y no se regula alostéricamente. Su velocidad se controla principalmente por la

28
concentración de citrato, el producto inhibidor y por la de oxaloacetato, su sustrato. Cuando la
isocitrato deshidrogenasa se activa, la concentración de citrato disminuye, liberando la inhibición
por producto y aumentando su velocidad. Dado que el equilibrio malato-oxaloacetato favorece al
malato la concentración de oxaloacetato es muy baja dentro de la mitocondria, por debajo del
Km aparente para la citrato sintasa. Cuando la relación NADH/NAD+ disminuye, la relación de
oxaloacetato/malato aumenta. El aumento en los niveles de oxaloacetato aumenta la velocidad de
la reacción de la citrato sintasa. En el hígado el cociente de las formas reducida a oxidada de esta
coenzima determina si el acetilCoA entra al ciclo o se utiliza para la síntesis de cuerpos
cetónicos.
Además de lo ya mencionado, otros factores están involucrados en la regulación de la actividad
del CAT. Por ejemplo, el aporte de sustratos como las unidades de acetilo que provienen del
piruvato o de ácidos grasos, es un factor crucial para determinar la velocidad del ciclo. Por lo
tanto, la regulación de la piruvato deshidrogenasa tendrá un efecto importante en la velocidad del
ciclo. De modo similar, cualquier mecanismo que regule los procesos de transporte y -
oxidación de ácidos grasos será un determinante efectivo de la actividad del ciclo.
En conclusión la regulación de la actividad del ciclo le permite a la célula adaptarse para afrontar
diferentes situaciones metabólicas, es decir, en aquellas donde se requiera una gran actividad
para producir la oxidación completa de moléculas combustibles las enzimas funcionarán con su
máxima capacidad y en aquellas donde el requerimiento sea menor, también disminuirá la
oxidación del acetilCoA que se desviará hacia la formación de triacilglicéridos que se
depositarán principalmente en el tejido adiposo. De este modo, el organismo logra mantener la
homeostasis calórica a pesar de las variaciones en el aporte de combustibles y en su utilización.
A modo de resumen,

29
Sistemas de lanzaderas o mecanismos de oxidación del NADH citosólico
Como ya hemos dicho, tanto las enzimas del ciclo de los ácidos tricarboxílicos como la cadena
de transporte de electrones se localizan en la mitocondria. También son mitocondriales las
enzimas de la -oxidación y de otras vías oxidativas. Ahora bien, en estas vías, cuando se oxidan
los sustratos, se reducen las coenzimas (NAD+, FAD). La cantidad de coenzimas dentro de la
mitocondria y en general dentro de la célula es muy baja, por lo tanto para que la vía metabólica
pueda continuar funcionando es necesario que se produzca su reoxidación. Las coenzimas se
reoxidan cediendo sus electrones a la cadena de transporte de electrones. Para las vías
metabólicas intramitocondriales, esto es bastante sencillo porque las coenzimas se reducen y
luego se reoxidan en la misma organela. Pero, ¿qué ocurre con las coenzimas reducidas en el
citosol?
Un mecanismo de reoxidación del NADH citosólico consiste en el transporte de los electrones a
la mitocondria por las lanzaderas del glicerol 3-fosfato o del malato-aspartato y finalmente al
oxígeno a través de la cadena de transporte de electrones.
Los sistemas de lanzaderas son una serie de reacciones que transfieren equivalentes de reducción
desde el NADH localizado en el citosol a la cadena de transporte de electrones. La membrana
mitocondrial interna es impermeable al NADH, no existen proteínas transportadoras que lo
pueda translocar. Los equivalentes de reducción (en forma de ión hidruro) se transfieren por una
reacción rédox citosólica a un compuesto que tiene un transportador en la membrana
mitocondrial interna. En ese paso inicial se regenera el NAD+ en el citosol. Los electrones se
transfieren a la cadena donde se generan aproximadamente 1,5 moles de ATP por mol de NADH
transportado por la lanzadera del glicerol 3-fosfato y 2,5 moles de ATP si la lanzadera es la del
malato-aspartato.
Lanzadera del glicerol 3-fosfato
Es la lanzadera predominante en el músculo esquelético y en el cerebro. En el citosol, los
electrones se transfieren del NADH a la dihidroxiacetona fosfato formando el glicerol 3-fosfato
(catalizada por la glicerol 3-fosfato deshidrogenasa citosólica). Este compuesto se transporta a la
membrana mitocondrial interna donde cede sus electrones a la glicerol fosfato deshidrogenasa
que contiene FAD, la que a su vez los cede a la coenzima Q. Esta vía, como otras que ceden sus
electrones a la coenzima Q, genera aproximadamente 1,5 ATP por la transferencia de estos
electrones al oxígeno. La dihidroxiacetona fosfato vuelve al citosol.

30
Lanzadera del malato-aspartato
Es una lanzadera más compleja que la anterior pero funciona en forma similar. Se regenera el
NAD+ en el citosol por la reducción del oxaloacetato a malato catalizada por la enzima malato
deshidrogenasa citosólica. El malato es translocado por una proteína transportadora (transporta
malato y -cetoglutarato) a la matriz mitocondrial donde se regenera el NADH por la malato
deshidrogenasa mitocondrial. La reoxidación del NADH por esta vía genera 2,5 moles de ATP
por mol de NADH reoxidado. El oxaloacetato, que no puede atravesar la membrana mitocondrial
interna es transaminado a aspartato en la mitocondria. El aspartato se trasloca (transportador de
glutamato-aspartato) hacia el citosol donde es transaminado nuevamente a oxaloacetato. Esta
lanzadera es la más activa y funciona en el hígado y en el corazón. Los tejidos con mitocondrias
poseen ambos sistemas de lanzaderas.