Sistema de complemento
45
SISTEMA DEL COMPLEMENTO DR. JORGE CHAVEZ MEZONES MEDICO INFECTOLOGO
description
Inmunología General de Medicina
Transcript of Sistema de complemento

SISTEMA DEL COMPLEMENTO
DR. JORGE CHAVEZ MEZONESMEDICO INFECTOLOGO

El complemento fue descubierto por Jules Bordet del Instituto Pasteur en París en 1890, cuando demostró que el antisuero de oveja contra la bacteria Vibrio cholerae lisaba la bacteria y el calentamiento del antisuero destruía su actividad bacteriolítica.
Bordet consideró que la actividad bacteriolítica requiere 2 sustancias diferentes: primero, anticuerpos antibacterianos específicos que sobreviven al proceso de calentamiento, y un segundo componente sensible al calor que causa la actividad lítica.
Diseñó una prueba simple para la actividad lítica: la lisis (que se detecta con facilidad) de glóbulos rojos recubiertos con anticuerpo, la denominada hemólisis.

JULES BORDET (1870 – 1961)
PREMIO NÓBEL DE FISIOLOGÍA (MEDICINA) 1919

Paul Ehrlich, en Berlín, llevó a cabo de manera independiente experimentos similares e ideó el término complemento y lo definió como “la actividad del suero sanguíneo que completa la acción antibacteriana del anticuerpo”.
Se demostró además que los resultados de la activación del complemento van más allá de la lisis celular mediada por anticuerpo, y que el complemento tiene un papel clave tanto en la inmunidad innata como en la adaptativa.
Posición en el sistema inmunitario innato: una función importante de este sistema es el reconocimiento y la destrucción de patógenos con base en el reconocimiento de patrones moleculares relacionados con patógeno, o PAMP, más que en la especificidad de anticuerpo.

La activación de la cascada del complemento puede ser iniciada por varias proteínas que circulan en el suero normal.
Estas moléculas, denominadas proteínas de fase aguda, poseen capacidad de reconocimiento de patrones y experimentan cambios de concentración durante la inflamación.
De este modo, todas las diversas acciones del sistema del complemento pueden ser desencadenadas no sólo por anticuerpos, como lo demostró originalmente Bordet, sino también por componentes de la inmunidad innata.

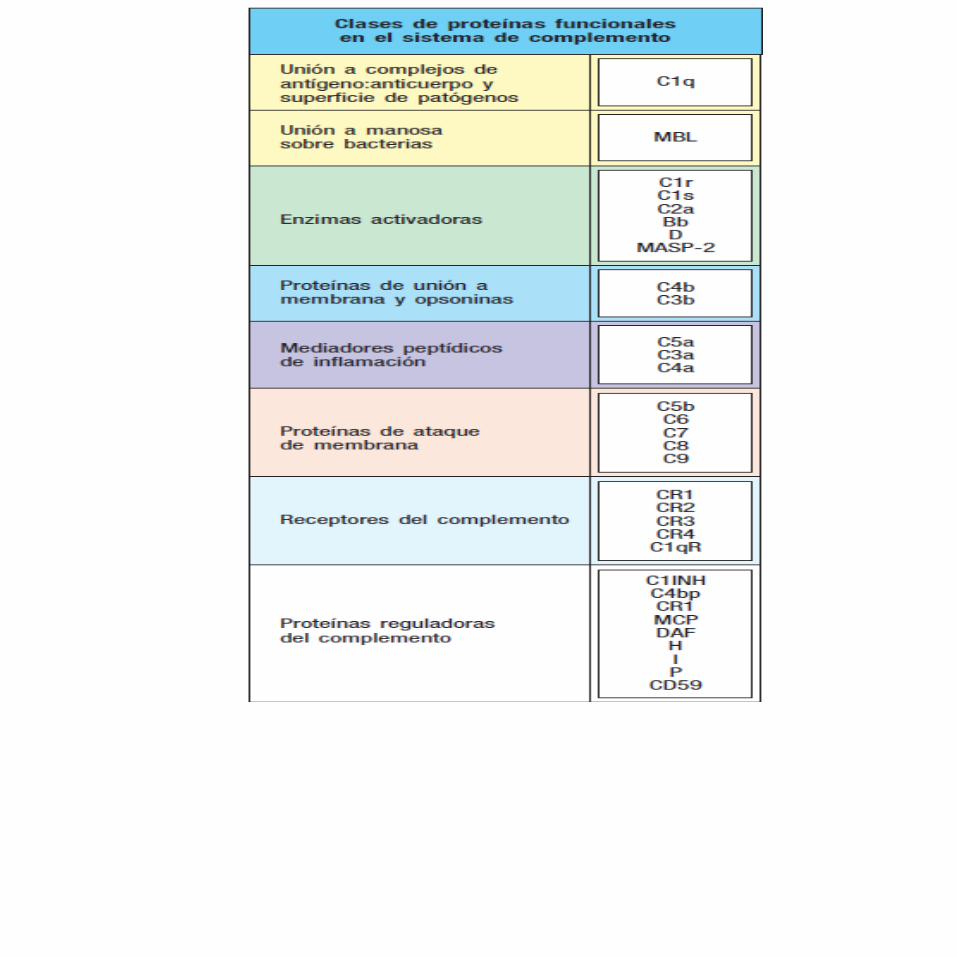
FUNCIONES DEL COMPLEMENTOEl sistema de complemento consta de más de 30 proteínas solubles y unidas a células.
Una característica del sistema es que varias proteínas del complemento son proteasas que sólo quedan activadas luego de división, generalmente por otra proteasa específica.
En su forma inactiva, esas enzimas se llaman proenzimas ozimógenos.
Los zimógenos precursores del sistema de complemento están ampliamente distribuidos en los líquidos y tejidos del cuerpo.

Después de la activación inicial, los diferentes componentes del complemento interactúan en una cascada muy regulada para llevar a cabo varias funciones básicas:
Lisis de células, bacterias y virus.Opsonización, que promueve la fagocitosis de Ag particulados.Unión a receptores de complemento específicos en células del sistema inmunitario, que desencadena funciones efectoras, inflamación y secreción de moléculas inmunorreguladoras.Depuración inmunitaria (inmunodepuración), que elimina complejos inmunitarios de la circulación y los deposita en el bazo y el hígado.
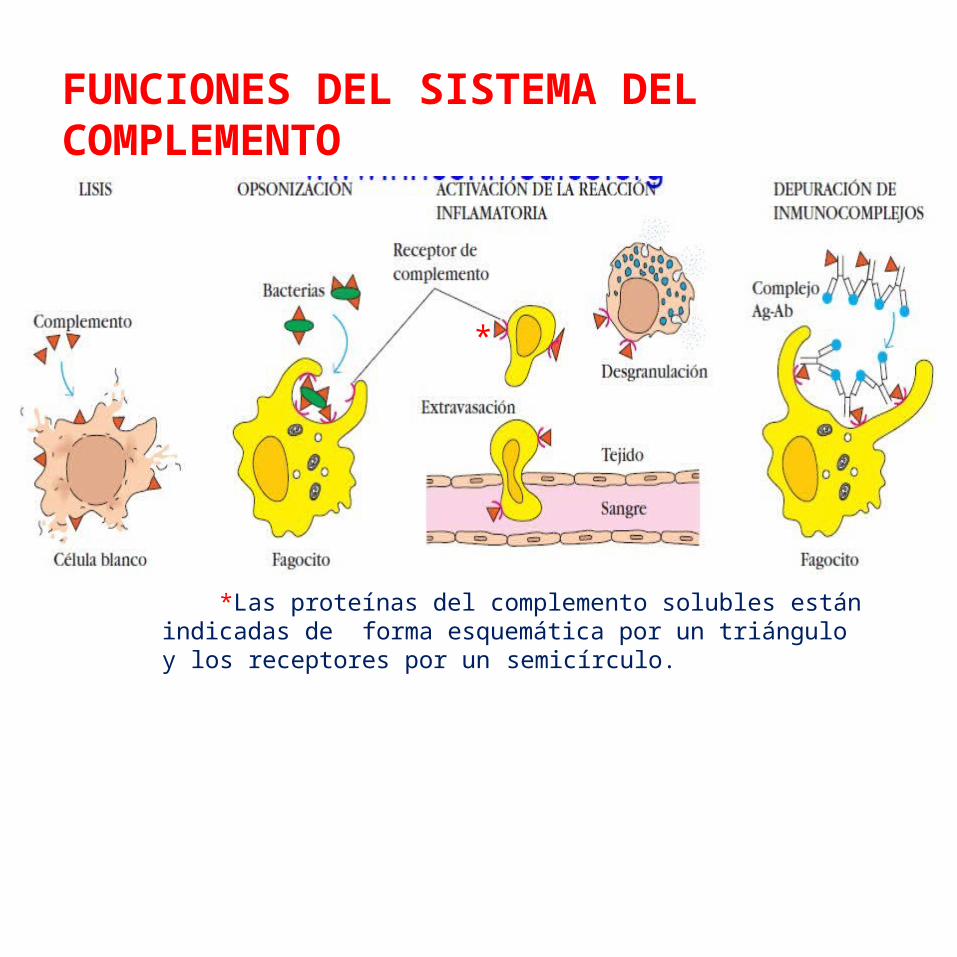
FUNCIONES DEL SISTEMA DEL COMPLEMENTO
*Las proteínas del complemento solubles están indicadas de forma esquemática por un triángulo y los receptores por un semicírculo.
*

COMPONENTES DEL COMPLEMENTO
Las proteínas y glucoproteínas solubles que componen elsistema del hepatocitos, importancia
aunque también producen cantidadesmonocitos sanguíneos, macrófagos
tisulares
complemento se sintetizan sobre todo en losde
ycélulas epiteliales de los aparatos digestivo y genitourinario.
Casi todos circulan en el suero en formas funcionales inactivascomo proenzimas, o zimógenos.
Los componentes del complemento se designan con numerales (C1-C9), letras (p. ej., factor D) o nombres (factor de restricción homólogo).

Los fragmentos peptídicos que se forman por activación de un componente se indican con letras pequeñas. En la mayor parte de los casos, los fragmentos más pequeños que resultan de la escisión de un componente se designan “a”, y el fragmento más grande se designa “b” (p. ej., C3a, C3b).C2 es una excepción: C2a es el fragmento de escisión más grande).
Los fragmentos más grandes se unen al blanco cerca del sitio de activación, y los más pequeños se difunden desde el sitio y pueden precipitar reacciones inflamatorias localizadas por unión a receptores específicos.
Los fragmentos del complemento interactúan entre sí para formar complejos funcionales. Los complejos que tienen actividad enzimática se designan con una línea sobre el número o signo (p. ej., C4b2a, C3bBb).

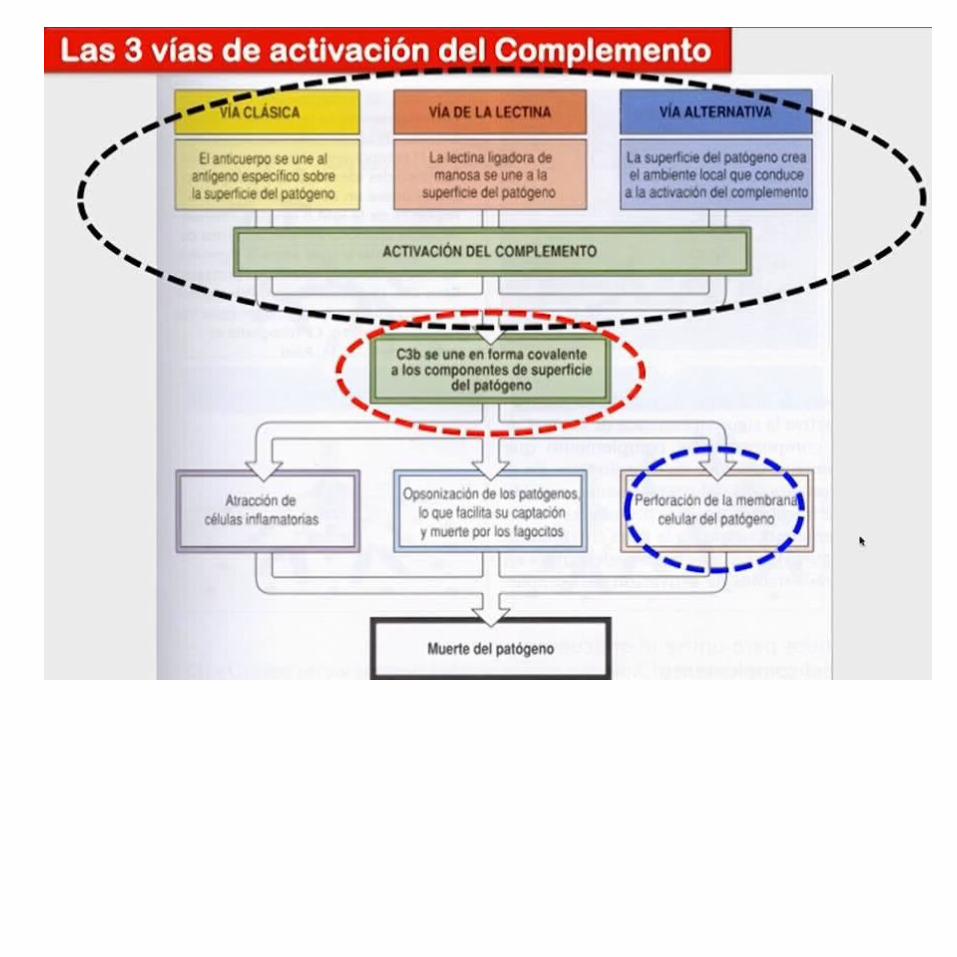
ACTIVACIÓN DEL COMPLEMENTOSitio clave para la activación de la vía del complemento: la superficie depatógenos, y hay 3 vías mediante las cuales puede activarse.Estas vías dependen de diferentes moléculas para su inicio, pero convergen para generar el mismo grupo de proteínas del complemento efectoras.También hay 3 modos en los cuales el sistema de complemento protege contra Infección:1. Genera gran número de proteínas del complemento activadas que se
unen de manera covalente a patógenos, y los opsoniza para fagocitosis por fagocitos que portan receptores para complemento.
2. Los fragmentos pequeños de algunas proteínas del complemento actúan como quimioatrayentes para reclutar más fagocitos hacia el sitio de activación del complemento, y para activar también estos fagocitos.
3. Los componentes finales en la vía del complemento dañan ciertas bacterias al crear poros en la membrana bacteriana.
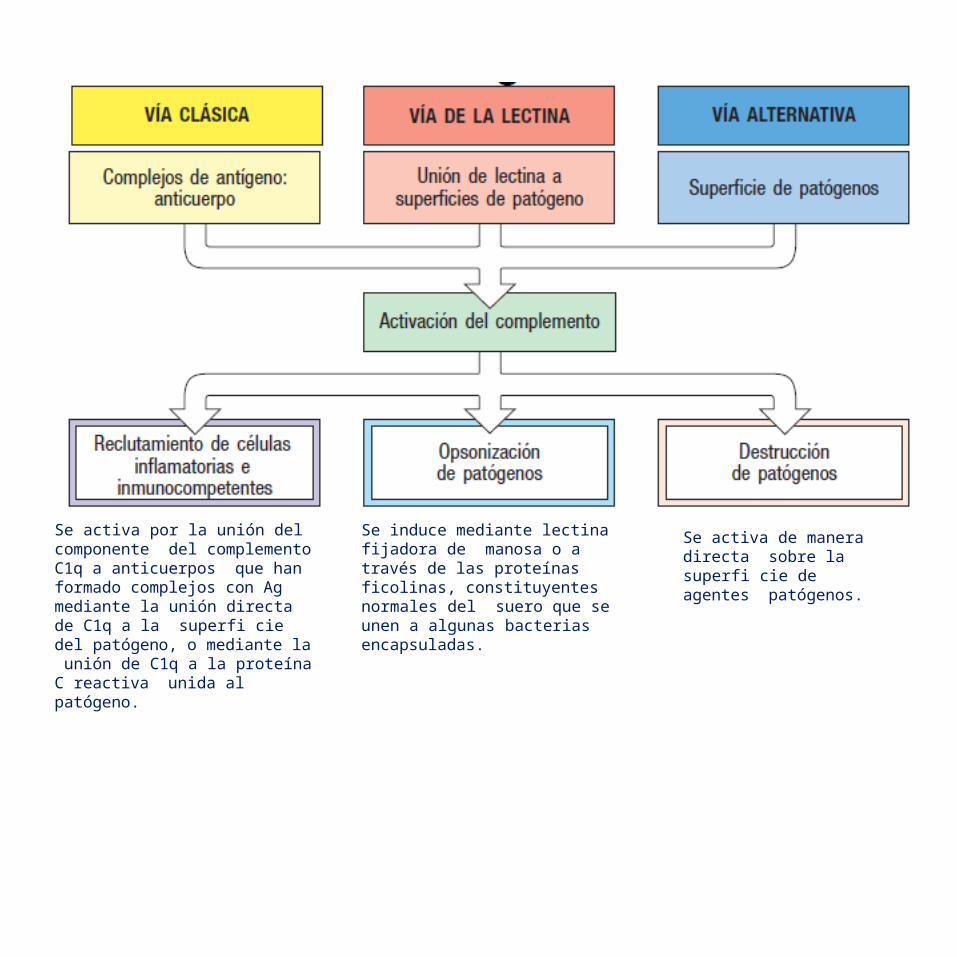
Se activa por la unión del componente del complemento C1q a anticuerpos que han formado complejos con Ag mediante la unión directa de C1q a la superfi cie del patógeno, o mediante la unión de C1q a la proteína C reactiva unida al patógeno.
Se induce mediante lectina fijadora de manosa o a través de las proteínas ficolinas, constituyentes normales del suero que se unen a algunas bacterias encapsuladas.
Se activa de manera directa sobre la superfi cie de agentes patógenos.

El complemento no sólo se activa por microorganismos infecciosos. Las células moribundas, como las que están en sitios de lesión isquémica (lesión de tejidos causada por falta de oxígeno) pueden desencadenar la activación del complemento.
Dado que las partículas cubiertas con complemento soncaptadas con mayoreficiencia complemento tiene
importancia
porfagocitos, el en
laeliminación
eficiente de células muertas, dañadas y apoptóticas y, al hacerlo, protege contra la aparición de autoinmunidad, el ataque de los antígenos propios del cuerpo por el sistema inmunitario.

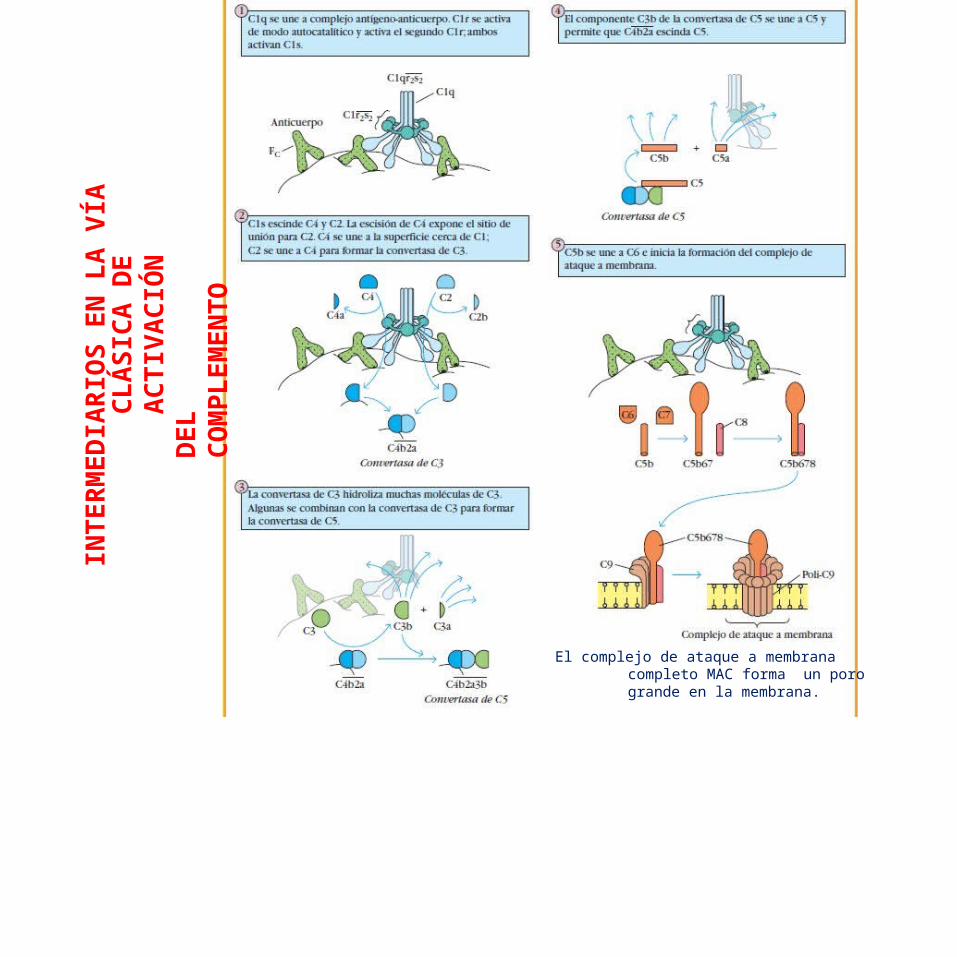
En etapas tempranas en las 3 vías de activación del complemento una serie de reacciones de división culmina en la formación de una enzima activa llamada convertasa de C3, que divide el componente del complemento C3 en C3b y C3a.
La producción de la convertasa de C3 es el punto en el cual convergen las 3 vías y se generan las principales funciones efectoras del complemento.
C3b se une de modo covalente a la membrana de la célula bacteriana y opsoniza a las bacterias, lo que les permite a los fagocitos internalizarlas.

C3a es un mediador peptídico de inflamación local.
C5a y C5b se generan mediante la división de C5b poruna convertasa de C5.
C5b desencadena los eventos tardíos en los cuales los componentes terminales del complemento se ensamblan para formar un complejo de ataque de membrana MAC capaz de dañar la membrana de ciertos patógenos.

INTE
RMED
IARI
OS
EN L
A VÍ
A CL
ÁSIC
A D
EAC
TIVA
CIÓ
N
DEL
CO
MPL
EMEN
TO
El complejo de ataque a membrana completo MAC forma un poro grande en la membrana.


VÍA ALTERNATIVA DEL COMPLEMENTOEl nombre de esta vía se debe a que se descubrió como una segunda vía, para la activación del complemento.
Tiene lugar en muchas superficies microbianas en ausencia de anticuerpo específico, y da pie a la generación de una convertasa C3 distinta designada C3bBb.
El inicio de la vía alternativa no depende de una proteína de unión a patógeno; se inicia por medio de la hidrólisis espontánea de C3.
El complemento activado por la vía alternativa ataca patógenos mientras preserva las células hospedadoras, que están protegidas por proteínas reguladoras del complemento.

La vía alternativa de activación del complemento amplifica la vía clásica o la vía de la lectina al formar una convertasa de C3 alternativa y depositar más moléculas de C3b sobre el patógeno.
Genera productos activos similares a los de la vía clásica, pero lo hace sin necesidad de complejos de antígeno y anticuerpo para iniciarse.
Esta vía mayor de activación del complemento incluye 4 proteínas séricas: C3, factor B, factor D y properdina.
Debido a que no se requiere anticuerpo, la vía alterna es un componente del sistema inmunitario innato.

INICIADORES DE LA VÍA ALTERNA DE ACTIVACIÓN DEL COMPLEMENTO


RELACIÓN ENTRE LOS FACTORES DE LAS VÍAS DE ACTIVACIÓN DEL COMPLEMENTO ALTERNATIVA, DE LA LECTINA, Y CLÁSICA.


LA VÍA DE LECTINA
como blancosLas lectinas son proteínas quereconocen carbohidratos específicos y se unen a ellos.Debido a que la lectina que activa el complemento se une a residuos manosa, algunos autores llaman a esta vía la vía de MBL o vía de lectina de unión a manosa.
Al igual que en el caso de la vía alterna, la activación de la vía de lectina no depende de anticuerpos.
Sin embargo, el mecanismo es más similar al de la vía clásica que al de la vía alterna, porque después de iniciarse prosigue a través de la acción de C4 y C2 para producir proteínas activadas del sistema del complemento.

La vía de lectina se activa cuando la lectina de unión a manosa (MBL, del inglés mannose-binding lectin) se une a residuos manosa en glucoproteínas o carbohidratos sobre la superficie de microorganismos, incluidas ciertas cepas de Salmonella, Listeria y Neisseria, además de los hongos Cryptococcus neoformans y Candida albicans y algunos virus, como VIH-1 y virus sincicial respiratorio.
Las células humanas normalmente tienen residuos de ácido siálico que cubren los grupos azúcar reconocidos por MBL, y no son blancos para la unión.
La MBL, miembro de la familia de las colectinas, es una proteína de fase aguda y su concentración aumenta durante las reacciones inflamatorias.

Resumen de las tres vías de activación del complemento.Se muestra el inicio ya sea por la vía clásica, a partir de la unión de C1 a un complejo antígeno-anticuerpo, o por las vías alterna o de lectina, que son independientes de anticuerpo. Todas las vías convierten C3 en su forma activa. Con la formación de C5b, las tres vías convergen para generar el complejo de ataque a membrana.

REGULACIÓN DEL SISTEMA DEL COMPLEMENTODebido a que muchos elementos del sistema del complemento son capaces de atacar tanto células del huésped como células extrañas y microorganismos, existen por evolución mecanismos reguladores complejos que restringen la actividad del complemento a los blancos designados.
La regulación activa de la actividad del complemento está a cargo de una serie de proteínas reguladoras que desactivan diversos componentes del complemento y contienen secuencias repetitivas de aminoácidos de unos 60 residuos, llamadas repeticiones de consenso cortas (SCR, del inglés short consensus repeats).
En el ser humano, todas estas proteínas se codifican en un mismo sitio en el cromosoma 1, que se conoce como grupo génico de reguladores de la activación del complemento (RCA, del inglés regulators of complement activation).

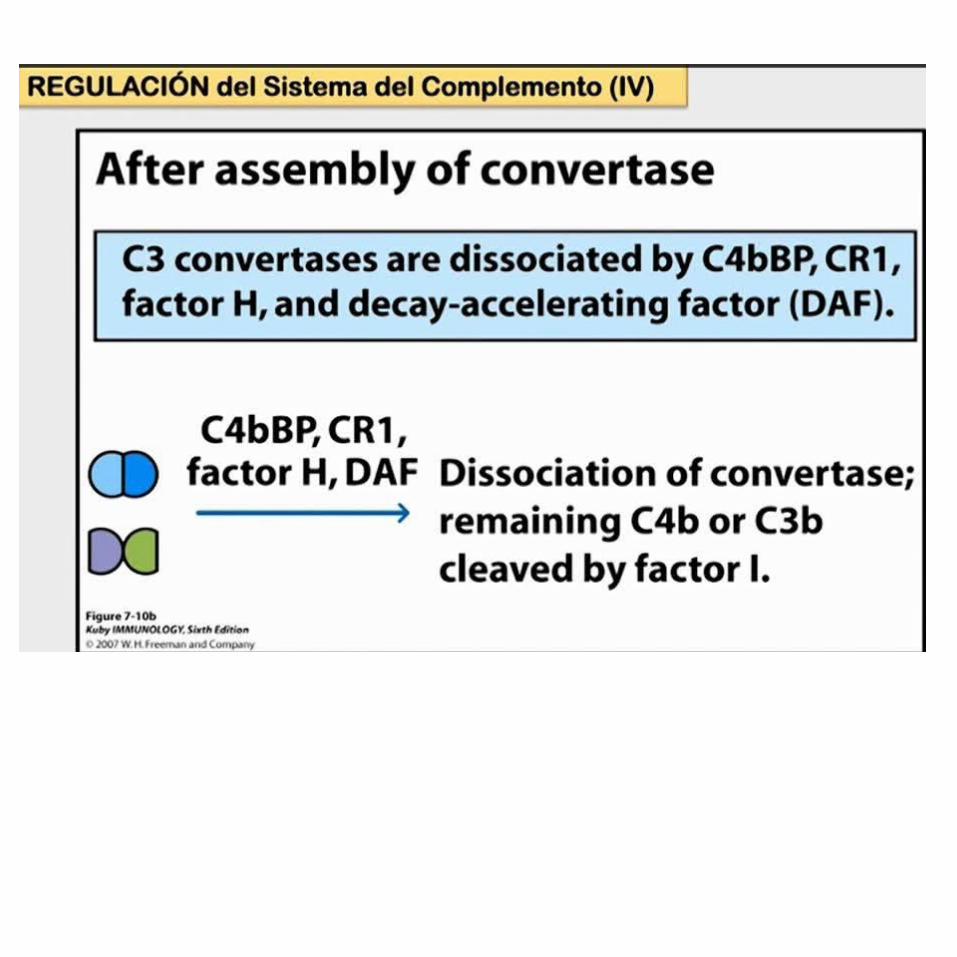


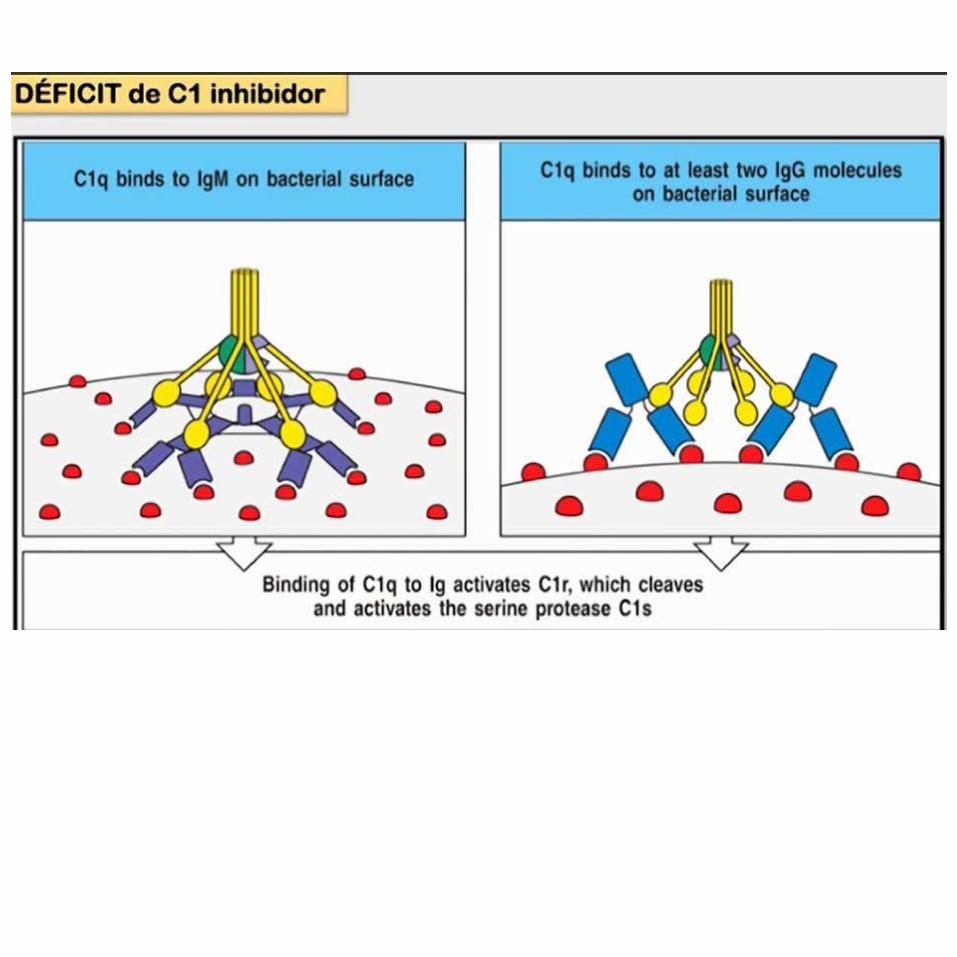




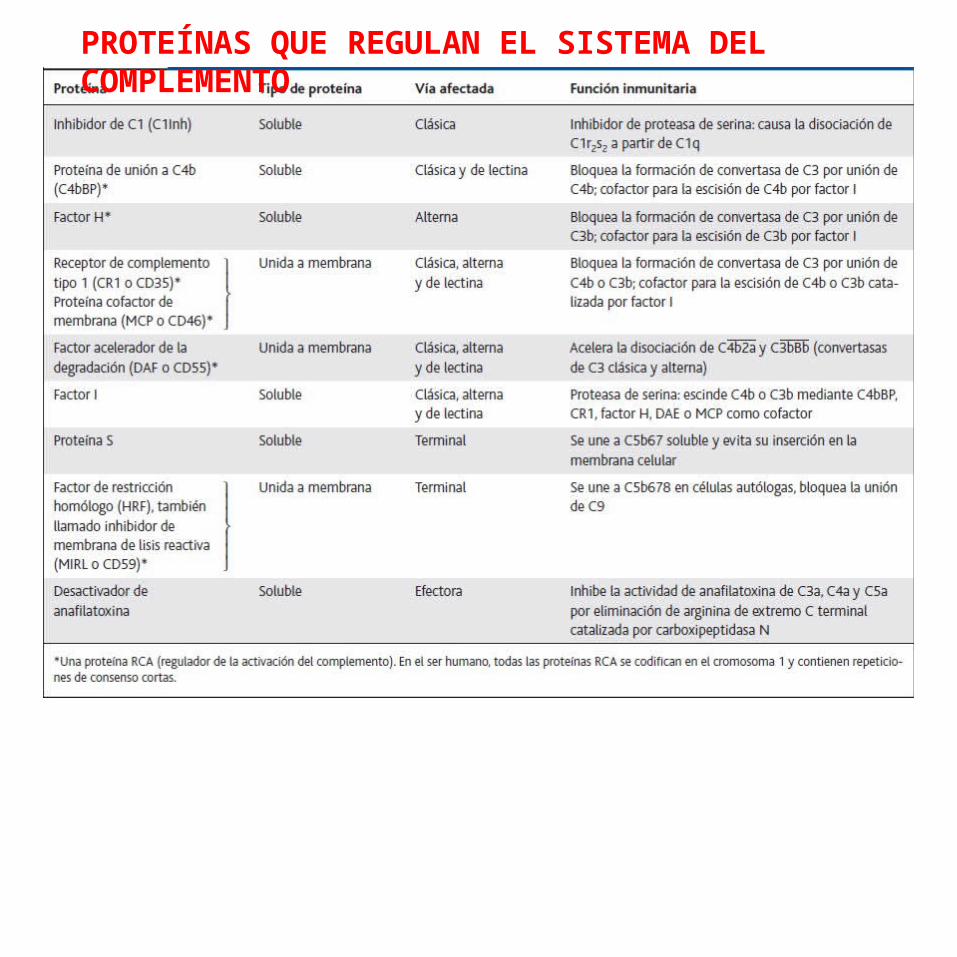
PROTEÍNAS QUE REGULAN EL SISTEMA DEL COMPLEMENTO

CONSECUENCIAS BIOLÓGICAS DE LA ACTIVACIÓN DEL COMPLEMENTO
EFECTOS BIOLÓGICOS MEDIADOS POR PRODUCTOS DEL COMPLEMENTO

Por lo general, el sistema del complemento es muy eficaz para lisar bacterias gramnegativas. Sin embargo, algunas de estas últimas y la mayor parte de las bacterias grampositivas tienen mecanismos para evitar el daño mediado por Complemento.
Por ejemplo, algunas bacterias gramnegativas pueden desarrollar resistencia a lisis mediada por complemento que se correlaciona con la virulencia del microorganismo.
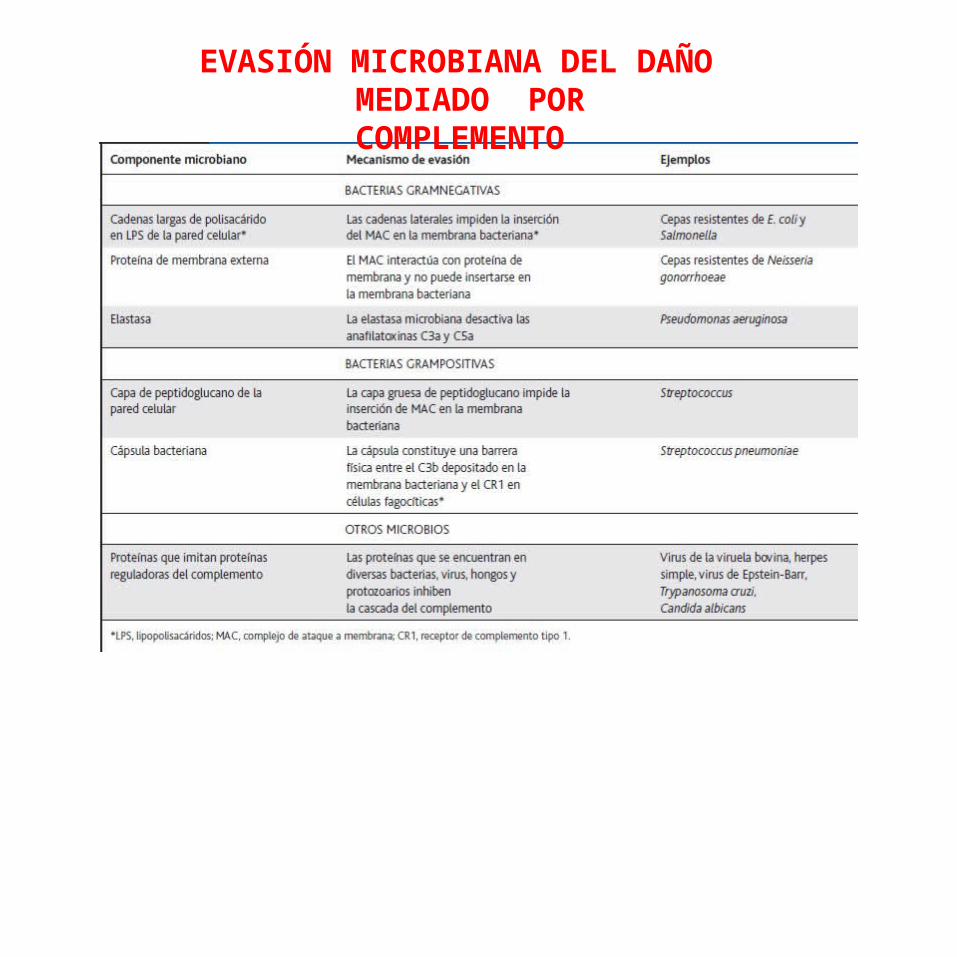
EVASIÓN MICROBIANA DEL DAÑO MEDIADO POR COMPLEMENTO

1. El sistema del complemento comprende un grupo de proteínas séricas, muchas de las cuales existen en formas inactivas.
2. La activación del complemento ocurre por las vías clásica, alterna o de lectina, cada una de las cuales se inicia de manera diferente.
3. Las 3 vías convergen en una secuencia clásica de fenómenos que conducen a la generación de un complejo molecular que causa lisis celular.
4. La vía clásica se inicia por la unión de anticuerpo a una célula blanco; las reacciones de IgM y ciertas subclases de IgG activan esta vía.
5. La activación de las vías alterna y de lectina es independiente de anticuerpo. Estas vías se inician por la reacción de proteínas del complemento con moléculas superficiales de los microorganismos.
RESUMEN

6. Además de su papel fundamental en la lisis celular, el sistema del complemento media la opsonización de bacterias, la activación de la inflamación y la depuración de inmunocomplejos.
7. Las interacciones de proteínas del complemento y fragmentos proteínicos con receptores en las células del sistema inmunitario controlan las reacciones inmunitarias innata y adquirida.
8. Debido a su capacidad de dañar al organismo hospedador, el sistema del complemento requiere mecanismos reguladores pasivos y activos complejos.
9. Las consecuencias clínicas de las deficiencias hereditarias del complemento van desde mayor susceptibilidad a infecciones hasta daño de tejidos causado por inmunocomplejos.