
Xenética molecular
46
XENÉTICA MOLECULAR Profesor: Adán Gonçalves
-
Upload
adan-goncalves-conselleria-de-educacion-xunta-de-galicia -
Category
Education
-
view
17 -
download
2
Transcript of Xenética molecular

XENÉTICA MOLECULAR
Profesor: Adán Gonçalves

1. O ADN, PORTADOR DA INFORMACIÓN XENÉTICA
As primeiras análises químicas revelaron que os cromosomas
(portadores da información xenética como reflectía a teoría
cromosómica) están constituídos por ADN, un ácido nucleico, e por
proteínas a partes máis ou menos iguais. Esto xa se sabía a principios
do século XX.
Nun principio os científicos dubidaban entre estes dous candidatos
(proteínas e ácidos nucleicos) como portadores desta información.
Incluso as proteínas tiñan máis apoios, pois había un coñecemento
maior da natureza química das proteínas (se sabía xa que eran longas
cadeas de aa cuxa orde variaba) que dos ácidos nucleicos que se
pensaba que eran un tetranucleótido cíclico.
O experimento de Frederick Griffith en 1928, sen sabelo nin el mesmo,
supuxo o primeiro paso para resolver este dilema…

O PRIMEIRO PASO: O EXPERIMENTO DE GRIFFITH (1928-29)
Griffith facía experimentos co pneumococo (unha
bacteria que causa pneumonía). A inoculación
desta bacteria en ratos pode causar a morte en 24
h debido a unha cápsula que posúen por fóra da
parede.
Hai 2 cepas desta bacteria: cepa S (con cápsula e
mortal) e cepa R (sen cápsula e non virulenta).
Cos seus experimentos Griffith comprobou que
unha mestura de cepa S fervidas (mortas) e cepa
R vivas provocaba a morte nos ratos. É dicir as
bacterias R volvíanse virulentas só coa presencia
de S mortas; TRANSFORMÁBANSE.
Debido a que Griffith non sabía cal era a molécula
responsable denominouna “PRINCIPIO
TRANSFORMANTE”.

O SEGUNDO PASO: OS EXPERIMENTOS DE AVERY, McCLEOD E
McCARTHY (1944)
En 1944, estos investigadores demostraron mediante varias experiencias
que o “principio transformante” de Griffith era o ADN, e é esta molécula
a que se transfire dende as bacterias S mortas (virulentas) ás R e que
convirte a estas últimas en virulentas. Só os extractos inoculados que
mantiñan o ADN intacto permitían a transformación. Por tanto, é o ADN
o portador da información xenética. Posteriores experimentos
demostraron que o ADN é o material xenético en todos os seres vivos.
F. Griffith O. Avery McCleod McCarthy

EXPERIMENTO DE HERSEY Y CHASE

2. A DOBRE HÉLICE
A publicación do modelo de Watson e Crick en 1953 (que xa explicamos en
temas anteriores) e a súa difusión rematou co escepticismo acerca do papel
do ADN como portador da información xenética e sentou as bases para o
nacemento dunha nova disciplina, a Bioloxía Molecular que supuxo unha
revolución sen precedentes no eido das ciencias biolóxicas e que debido ao
seu grande desenvolvemento nas décadas seguintes trouxo consigo unha
contribución esencial á comprensión do funcionamento molecular dos
sistemas vivintes.
Sen esquecer que na actualidade é unha das ramas da Bioloxía cun futuro
máis prometedor como veremos.

3. O CONCEPTO DE XENE
Cada ser vivo presenta unha características anatómicas, fisiolóxicas e
comportamentais que o fan único. Cada unha desas características ou
trazos distintivos denomínase, en Xenética, carácter.
Estos caracteres veñen determinados pola herdanza (os xenes), pero
tamén polo ambiente que rodea ao individuo. Así unha persoa pode ter
recibido dos seus proxenitores unha tendencia clara ao sobrepeso, pero a
súa dieta diaria (o ambiente) terá unha influenza definitiva na expresión
desta condición.
Dito isto, que é un xene?
Dende un punto de vista xenético, un xene é un fragmento de ADN que
porta a información xenética para un carácter.

Os experimentos de Beadle e Tatum
Estes experimentos realizados no fungo Neurospora crassa foron
esenciais para determinar as funcións dos xenes. A partir deles
enunciaron en 1948 a teoría un xene - un enzima pola que recibirían
o premio Nobel en 1958. Da idea xene-carácter a xene-substancia.
Posteriormente comprobouse que esta idea podía extenderse a
todas as proteínas e reformulouse como teoría un xene – unha
cadea polipeptídica.
Dende un punto de vista molecular, un xene é un fragmento de
ADN que leva información para a síntese ou a regulación da síntese
de alomenos unha proteína, necesaria para que se exprese un
determinado carácter nun individuo. É a unidade básica da
transcrición.
3. O CONCEPTO DE XENE

4. A REPLICACIÓN É SEMICONSERVATIVA
Watson e Crick xa esbozaran no mesmo ano en que publicaron o modelo
da dobre hélice o mecanismo polo cal o ADN faría unha copia de si mesmo
para poder repartir equitativamente o material xenético entre as células
fillas cando unha célula vai a dividirse. Eles afirmaban que a replicación
era semiconservativa mediante a apertura das cadeas para que serviran
como molde, pero se plantexaron tamén outras posibilidades.
Hipótese conservativa: O ADN ábrese e cópiase orixinando un ADN fillo
formado por dúas cadeas totalmente novas. O resultado son dúas
moléculas unha a que servíu de patrón e outra de síntese “de novo”.
Hipótese semiconservativa: O ADN ábrese e cada cadea serve de patrón
para copiar unha febra nova orixinanado polo tanto dúas moléculas
híbridas (cada unha cunha cadea patrón e cunha cadea filla).
Hipótese dispersiva: O ADN ábrese e o copiado alterna nas febras
fragmentos novos e antigos.


O EXPERIMENTO DE MESELSON Y STAHL (1957)
Estes dos científicos demostraon nun elegante experimento que Watson e
Crick tiñan razón, a replicación é semiconservativa.
Para elo pasaron bacterias de E. Coli cultivadas nun medio con 15N a un
medio con 14N durante media hora, o tempo preciso para un ciclo de
replicación. Despois extraeron o ADN e centrifugárono. Mediante
ultravioleta observouse que o ADN obtido ocupaba unha posición
intermedia entre o ADN con 15N e o ADN con 14N. Esto permitía descartar
xa a hipótese conservativa.
Posteriormente deixaron pasar dous ciclos de replicación e repetiron o
procedemento. O resultado foi ADN de dúas densidades, un que coincidía
co ADN con 14N e outro que era intermedio entre o 14N e o 15N.
Para asegurarse repetiron o experimento permitindo ata 3 replicacións e
observaron que o ADN híbrido (14N-15N) diminuía o que descartaba
definitivamente a hipótese dispersiva e demostraba a semiconservativa.


Se deixaban un maior número de ciclos de replicación a proporción de
ADN híbrido era cada vez máis pequena.
Ademais como procedemento definitivo aillaron o ADN e separaron as
dúas febras comprobando que unha era lixeira e outra pesada.

5. A REPLICACIÓN
Tras comprobar que efectivamente a replicación é semiconservativa houbo
numerosos esforzos por tratar de dilucidar como sucedía exactamente o
proceso:
Arthur Kornberg illou en 1956 unha ADN polimerasa capaz de sintetizar
ADN.
John Cairn e outros nos anos 60 demostraron que a replicación é
ordenada e secuencial e que se forma unha “horquilla de replicación”.
Okazaki , en 1968, descubriu os fragmentos que levan o seu nome e
demostrou a síntese retardada.
O proceso é moi similar en procariotas e eucariotas e está moi ben
estudado en E. Coli. Comentaremos agora como é proceso nesta bacteria e
salientaremos as diferenzas máis importantes que se dan do proceso en
eucariotas.
Lembremos que se produce no período S do ciclo celular.

5. A REPLICACIÓN
A replicación ou duplicación do ADN é un proceso complexo no que
participan moitas enzimas entre as que destacan as ADN-polimerasas que se
encargan de sintetizar as novas cadeas a partir das cadeas patrón. En
procariotas hai 3 ADN-polimerasas e en eucariotas coñecemos 5.
Para que se poida levar a cabo a síntese das novas cadeas é preciso que a
dobre hélice se desenrrole para permitir o acceso das polimerasas ás cadeas
que van a servir de molde.
Fase de Inicio: en procariotas comeza nun só punto denominado ORI C
(Monofocal), mentres que en eucariotas sucede simultaneamente en varios
puntos chamados replicones (Multifocal) grazas a existencia dunha señal de
incio (rica en A e T). Na separación da dobre hélice participan diversas
enzimas: helicasas (rompen as pontes de H entre as bases), topoisomerasas
(impiden as torsións), as proteínas SSB (estabilizan a estrutura)... A
actuación destas enzimas forma a horquilla de replicación que permite ás
polimerasas acceder ás cadeas patrón.

5. A REPLICACIÓN
Fase de Elongación: para que a polimerasa sexa capaz de iniciar o proceso
para ir sintetizando nucleótidos (nt) complementarios da cadea que serve
como molde (parental ou patrón) precisa de fragmentos curtos de ARN
(primer ou cebador) que son sintetizados por unha enzima chamada
primasa.
A ADN-polimerasa III engade desoxirribonucleótidos complementarios da
cadea parental a partir do cebador, pero só pode engadilos a partir do
extremo 3´-OH polo que a síntese da nova cadea prodúcese sempre en
dirección 5´-3´. Como consecuencia deste feito fórmase unha cadea de xeito
continuo (febra condutora ou continua) e a que se está a sintetizar enfronte
faino de modo discontinuo (febra retardada) en fragmentos (fragmentos de
Okazaki). Esto é así porque a replicación é bidireccional o que provoca que a
horquilla de replicación se mova nas dúas direccións, pero por contra as
polimerasas só actúan, como dixemos, nunha dirección tendo que esperar
nunha das cadeas a que a horquilla se abra para poder sintetizar a cadea.

©César Benito Jiménez
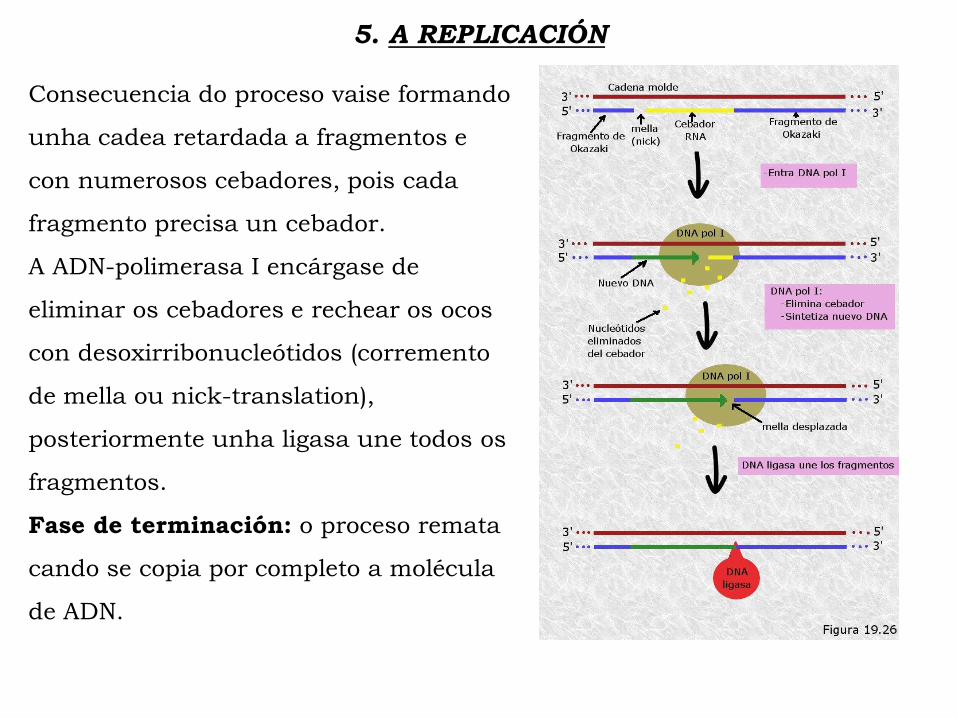
5. A REPLICACIÓN
Consecuencia do proceso vaise formando
unha cadea retardada a fragmentos e
con numerosos cebadores, pois cada
fragmento precisa un cebador.
A ADN-polimerasa I encárgase de
eliminar os cebadores e rechear os ocos
con desoxirribonucleótidos (corremento
de mella ou nick-translation),
posteriormente unha ligasa une todos os
fragmentos.
Fase de terminación: o proceso remata
cando se copia por completo a molécula
de ADN.

Diferenzas na replicación en eucariotas
É multifocal e comeza nos replicóns.
Hai 5 tipos de ADN-polimerasas
Ao completar cada ciclo de replicación prodúcese o acurtamento dos
telómeros (extremos dos cromosomas) que está relacionado co
envellecemento celular.

6. A EXPRESIÓN DA INFORMACIÓN XENÉTICA
Unha vez que Badle e Tatum en 1948 estableceran o paralelismo entre
xene e enzimas e tras a proposta da dobre hélice por Watson e Crick en
1953, o propio Crick formula a “hipótese da colinealidad” na que establece
a correspondencia entre a secuencia de nucleótidos e a secuencia de
aminoácidos da enzima que codifica o xene.
No paso da secuencia de nucleótidos á secuencia de aminoácidos
diferéncianse dous procesos:
Transcrición
Tradución
Este fluxo de información entre nucleótidos e aminoácidos foi proposto por
Crick como o dogma central da bioloxía molecular, que como veremos un
pouco máis adiante na actualidade engloba algúns procesos máis.

6. A EXPRESIÓN DA INFORMACIÓN XENÉTICA
A información xenética está contida no ADN, na secuencia de nucleótidos.
Esta información exprésase en último termo en forma de proteínas. Para
que esto suceda, ocorren dous procesos: TRANSCRICIÓN (copiado de ADN
a ARN) e TRADUCIÓN (de ARN a proteínas).
O ADN non pode saír do núcleo nos eucariotas para dirixirse ós
ribosomas, orgánulos onde se produce a síntese de proteínas. Por iso hai
que “copiar” (TRANSCRIBIR) o fragmento de ADN que interese á célula
nese momento. A molécula que leva a información do ADN do núcleo cara
ós ribosomas, é o ARNm constituído polas bases complementarias dunha
das dúas cadeas de ADN que lle serviu de molde. Este ARN leva como
pentosa a ribosa e como base complementaria da adenina o uracilo.

6. A EXPRESIÓN DA INFORMACIÓN XENÉTICA
Pero ademais disto sabemos que hai virus, retrovirus, que posúen ARN
que poden empregar como molde para a síntese de ADN grazas a que
posúen unha enzima denominada transcriptasa inversa ou
retrotranscriptasa.
E outros virus de ARN posúen outra enzima, a replicasa, que lles
permite facer copias do seu ARN.

7. TRANSCRICIÓN
De xeito xeral a transcrición consiste non só na síntese de ARNm, senón de
tódolos tipos de ARN tomando como patrón unha febra de ADN. O proceso
en liñas xerais é moi similar en todos os casos. Para que se copie a
información contida no ADN a ARN son necesarias unha enzimas
chamadas ARN-polimerasas. Nos procariotas hai só unha, nos ecucariotas
hai tres. O proceso pode resumirse nas seguintes etapas:
Iniciación
Elongación
Terminación
Maduración
En eucariotas este proceso sucede no núcleo.

7. TRANSCRICIÓN
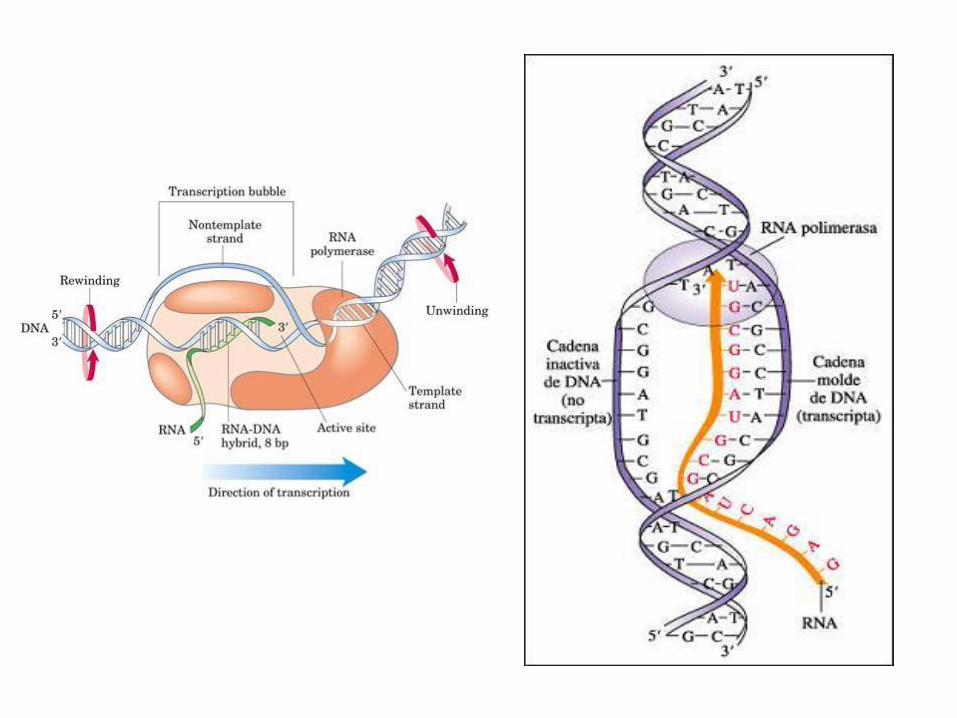
Iniciación: a ARN-polimerasa únese a un sitio específico do ADN, o
promotor. O promotor ten unha secuencias consenso que recoñece a
polimerasa. Estas secuencias varían entre procariotas e eucariotas.
Elongación: o ADN nesa rexión desenrrólase e queda o descuberto a
febra que actuará como patrón e a ARN-polimerasa ira colocando fronte
ela ribonucleótidos (lembremos que sintetiza ARN) complementarios;
como se está a formar ARN fronte a A poñerá U. A dirección de síntese da
cadea é 5´-3´.
Terminación: a síntese de ARN finaliza cando a polimerasa chega a unha
rexión do xene denominada “sinal de terminación”. En procariotas
habitualmente é unha rexión palindrómica de GC, en eucariotas é unha
sinal de poliadenilización.

En procariotas, para a síntese de ARNm o proceso remata aquí, pois éste
é ARN que xa pode ser traducido a proteínas. De feito, é habitual que
mentres se está a transcribir a cadea, polo extremo contario se esté
traducindo (transcrición e tradución acoplada). Tamén é frecuente a
formación de polisomas.
Maduración: en eucariotas en todos os ARN e en procariotas no caso
do ARNt e ARNr hai un último proceso preparativo antes de que os
ARN desempeñen a súa función denominado maduración.
En procariotas o ARNt e o ARNr sofren corte e empalme e despois
plegamento para dar lugar a forma tridimensional característica.
En eucariotas o proceso é máis complexo e hai unha serie de
modificacións post-transcricionais que afectan a todos os tipos de ARN.
7. TRANSCRICIÓN

7. TRANSCRICIÓN
Modificacións post-transcricionais en el ARNm de eucariotas:
Formación da CAP: no extremo 5´ engádese a 5-metil-guanosina-
trifosfato denominada cap ou caperuza que protexe da degradación e
facilitará a formación do complexo de inicio do próximo proceso de
tradución.
Formación da Poli-A: no extremo 3´engádese unha cadea de A que
tamén protexen contra a degradación da molécula e ademais facilitan a
saída do ARNm do núcleo.
Splicing ou corte e empalme: en eucariotas tras a fase de terminación o
ARNm é en realidade un transcrito primario ou ARNhn que non pode ser
traducido todavía. Nesta molécula hai rexións con información para
síntese peptídica (exones) e outras sen esta información (intróns). Estes
intróns deben ser eliminados mediante o mecanismo de “corte e empalme”
(splicing) que ten lugar grazas a enzima ribonucleoproteína pequena
nuclear e ao ARNpn. O empalme sucede grazas a ligasas.
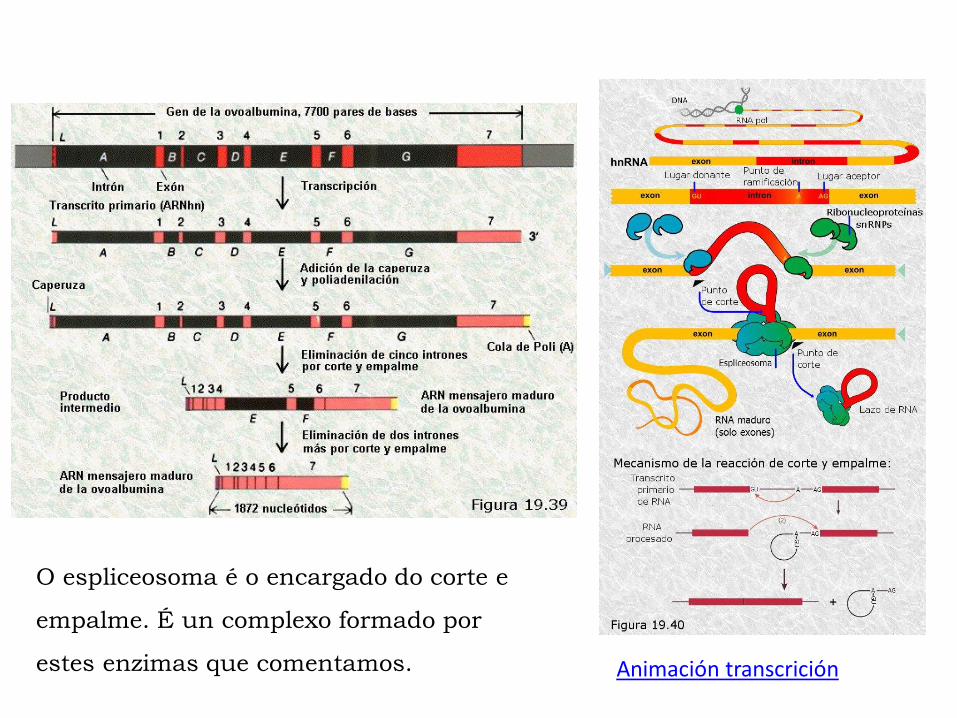
O espliceosoma é o encargado do corte e
empalme. É un complexo formado por
estes enzimas que comentamos. Animación transcrición
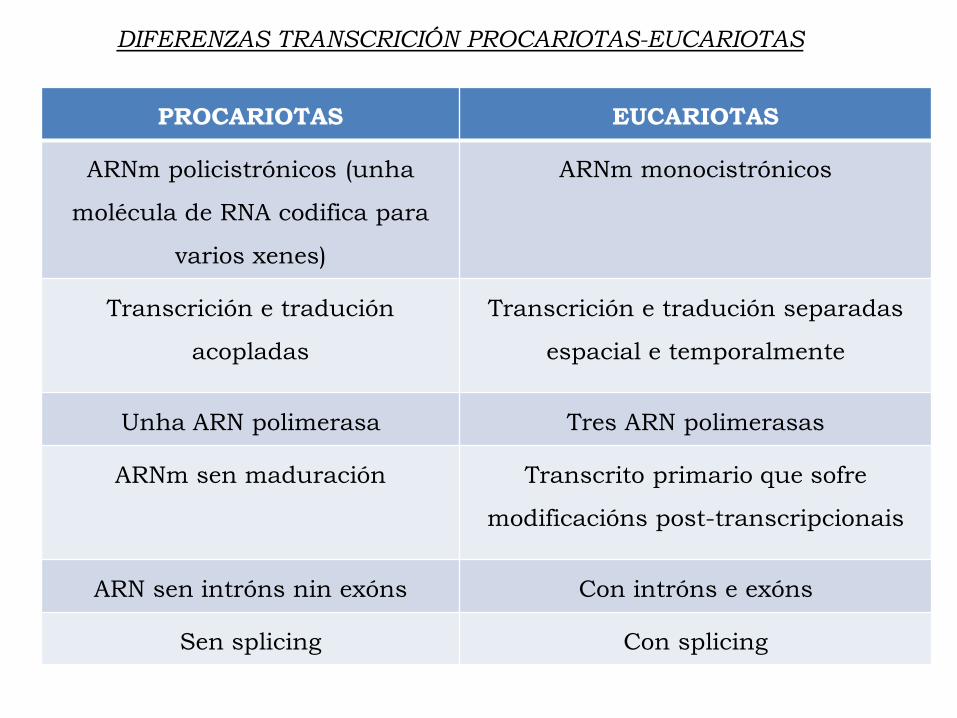
PROCARIOTAS EUCARIOTAS
ARNm policistrónicos (unha
molécula de RNA codifica para
varios xenes)
ARNm monocistrónicos
Transcrición e tradución
acopladas
Transcrición e tradución separadas
espacial e temporalmente
Unha ARN polimerasa Tres ARN polimerasas
ARNm sen maduración Transcrito primario que sofre
modificacións post-transcripcionais
ARN sen intróns nin exóns Con intróns e exóns
Sen splicing Con splicing
DIFERENZAS TRANSCRICIÓN PROCARIOTAS-EUCARIOTAS

8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
O CÓDIGO XENÉTICO
As proteínas sintetízanse cunha secuencia de aa determinada pola tradución da
información contida no ARNm. No ARNm os aa veñen especificados por unidades de
información de tres nucleótidos, os tripletes ou codóns. A correspondencia entre estes
codóns e os aa proteicos denomínase código xenético.
Como se descubríu o código xenético?
Grazas sobre todo a dous hitos da bioloxía molecular:
En 1955 Severo Ochoa e Marianne Grunberg-Manago aillan a enzima
polinucleótido fosforilasa que é capaz de sintetizar sen molde ARNm. A partir dun
medio con soamente UDP obteñen un poli-U (UUUUUU...)
En 1961 M.W. Nirenberg dispuxo 20 tubos cos 20 aa proteicos marcados con 14C.
En todos engadíu poli-U e só no tubo da fenilalanina apareceu un polipéptido
marcado. Polo tanto este aa está codificado por UUU. Posteriormente fixo o mesmo
con poli-C (prolina), con poli-A (lisina) e poli-G (sen resultado).
Finalmente e a partir de aquí outros experimentos conseguiron dilucidar o código
xenético ao completo.
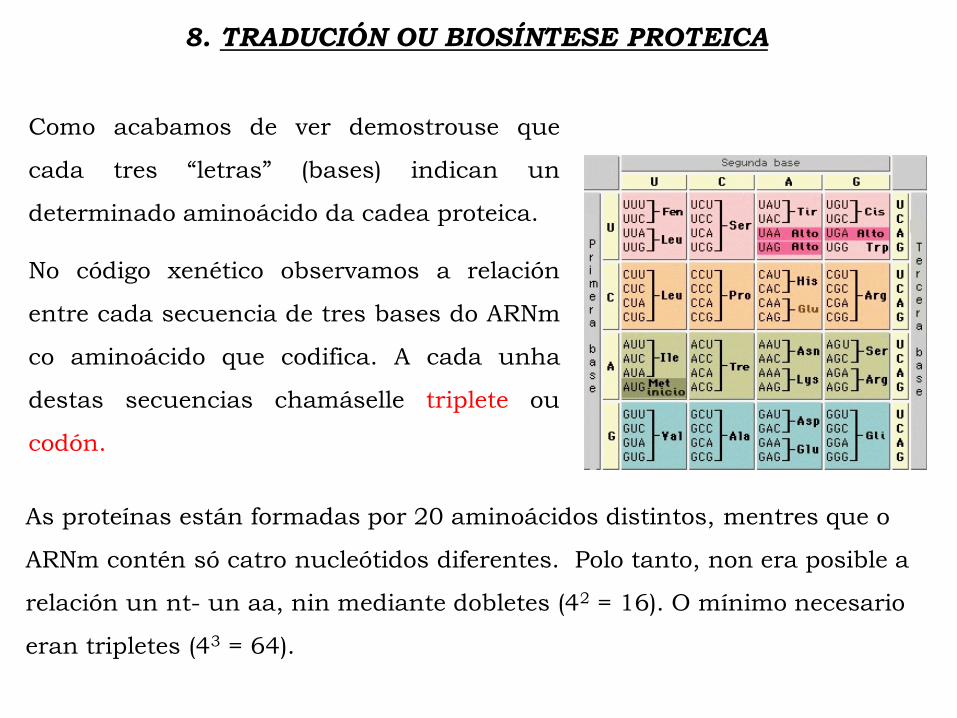
Como acabamos de ver demostrouse que
cada tres “letras” (bases) indican un
determinado aminoácido da cadea proteica.
No código xenético observamos a relación
entre cada secuencia de tres bases do ARNm
co aminoácido que codifica. A cada unha
destas secuencias chamáselle triplete ou
codón.
8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
As proteínas están formadas por 20 aminoácidos distintos, mentres que o
ARNm contén só catro nucleótidos diferentes. Polo tanto, non era posible a
relación un nt- un aa, nin mediante dobletes (42 = 16). O mínimo necesario
eran tripletes (43 = 64).
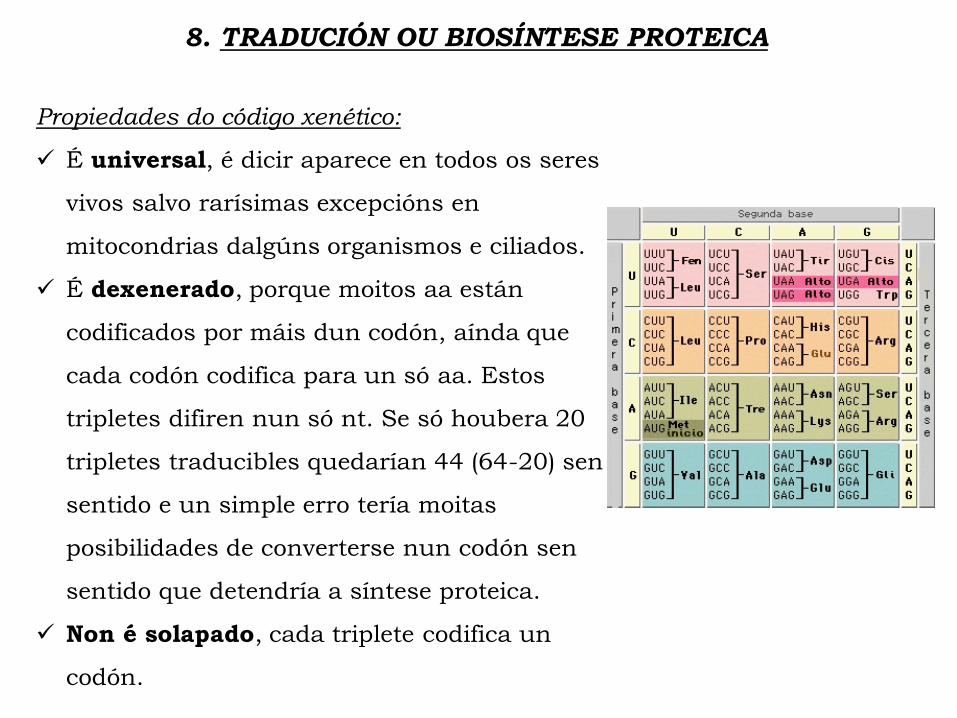
8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
Propiedades do código xenético:
É universal, é dicir aparece en todos os seres
vivos salvo rarísimas excepcións en
mitocondrias dalgúns organismos e ciliados.
É dexenerado, porque moitos aa están
codificados por máis dun codón, aínda que
cada codón codifica para un só aa. Estos
tripletes difiren nun só nt. Se só houbera 20
tripletes traducibles quedarían 44 (64-20) sen
sentido e un simple erro tería moitas
posibilidades de converterse nun codón sen
sentido que detendría a síntese proteica.
Non é solapado, cada triplete codifica un
codón.

No código xenético o codón de inicio da tradución sempre é AUG que
codifica para a metionina (Met) en eucariotas e formilmetionina en
procariotas, hai sen embargo varios codóns de paro.

8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
O proceso de tradución pódese dividir en varias fases:
Activación: cada un dos 20 aa proteicos se une no hialoplasma ao seu
ARNt específico. Este proceso, como xa comentamos, é mediado pola
aminoacil-sintetasa a expensas do ATP que se convirte en AMP+ PPi.
Cada ARNt só pode transportar un aa específico, pero un aa dado pode
ser transportado por varios ARNt.
Cada ARNt ten unha
secuencia de tres
nucleótidos (anticodón) que
lle permitirá a unión ao
ARNm no ribosoma
específicamente.

8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
Inicio: o ARNm únese polo seu extermo 5´ (onde está CAP) á subunidade
menor do ribosoma, despois únese o primeiro ARNt co seu aa específico
que ven codificado polo primeiro codón do ARNm. A unión no ribosoma
do ARNt e o ARNm prodúcese por complementariedade entre o codón do
ARNm e o anticodón do ARNt. Este codón de inicio é sempre AUG
(Metionina en eucariotas; formilmetionina en procariotas), polo tanto o
ARNt ten como anticodón UAC.
Na subunidade menor hai dous centros de unión, o centro P (centro
peptidilo) e o centro A (centro aminoacilo). O primeiro ARNt (que porta a
Met ou a formilMet) únese ao centro P, depois únese a subunidade maior
e queda constituído o complexo de inicio.

INICIO
Debuxos realizados por Milagros Nespereira

8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
Elongación: o seguinte aa en entrar no ribosoma ven dado polo codón do
centro A. O ARNt específico para o aa codificado por ese codón únese,
polo tanto ao centro A (neste momento temos 2 ARNt no ribosoma).
Posteriormente rómpese o enlace entre o primeiro ARNt e a Met que pasa
a unirse ao segundo ARNt que entrou no ribosoma (catalizado por
aminoacil-transferasas). O primeiro ARNt sae do ribosoma e éste móvese
un codón quedando agora o 2º ARNt que porta neste momento 2 aa no
centro P e deixando libre o centro A ao que se unirá o seguinte aa
codificado polo correspondente na cadea de ARNm e exposto no centro A.
A medida que o ribosoma avanza ao longo da cadea de ARNm en
dirección 5´-3´vaise formando a cadea polipeptídica en dirección N-
terminal a C-terminal.

ELONGACIÓN
Debuxos realizados por Milagros Nespereira

8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
Terminación: o proceso repítese ata que o ribosoma chega a un codón que
non codifica para ningún aa, un codón de terminación. Neste momento,
desensámblase o ribosoma e a cadea peptídica libérase.
Debuxos realizados por Milagros Nespereira

8. TRADUCIÓN OU BIOSÍNTESE PROTEICA
Como paso final a cadea peptídica adopta a súa forma tridimensional
biolóxicamente activa (pregamento).
9. REGULACIÓN DA EXPRESIÓN XÉNICA
A regulación a cal se sintetiza un produto xénico pódese regular en calquera
parte da ruta: a nivel transcricional, de procesamento ou estabilidade do
RNA, da tradución ou das modificacións postranscricionais.
En xeral, a expresión da maior parte do xenes foise optimizando ao longo da
evolución e as proteínas máis empregadas posúen promotores máis fortes.
Denomínanase xenes regulados a aqueles cuxa expresión varía coas
condicións da célula (concentración de metabolitos, pH, Tª...). A maioría dos
xenes son regulados por proteínas reguladoras que se unen ao DNA en
zonas cercanas ao promotor. Éstas poden ser represores ou activadores.

9. REGULACIÓN DA EXPRESIÓN XÉNICA
Represión: xene regulado negativamente por un represor que inhibe a
transcrición.
Activación: xene regulado positivamente por un activador que permite a
súa transcrición.
Represores e activadores son enzimas alostéricos.
Hai 3 mecanismos xerais de regulación da transcrición:
Activación: o activador e o seu ligando únense ao xene e estimulan a
transcrición, sen activador a transcrición é moi reducida.
Indución: un represor inhibe a transcrición. Cando o inductor (ligando)
únese ao represor diminúe a afinidade para unión ao xene e prodúcese a
transcrición.
Correpresión: o represor non se une ao ADN cando non está o
correpresor (ligando). Se hai correpresor, o represor se une ao xene e o
inhibe.

9. REGULACIÓN DA EXPRESIÓN XÉNICA
De xeito xeral, a síntese de ARNm en procariotas depende do substrato
dispoñible, mentras que en eucariotas adoita depender dos niveis
hormonais do medio interno.
9.1. O Operón
Este sistema foi proposto por Jacob e Monod como modelo de regulación
da expresión xénica en procariotas en base aos seus descubrimentos no
funcionamento do Operon Lac de E. Coli.
Un único promotor controla a expresión de varios xenes estruturales
(xenes policistrónicos) e este promotor está regulado por represores.
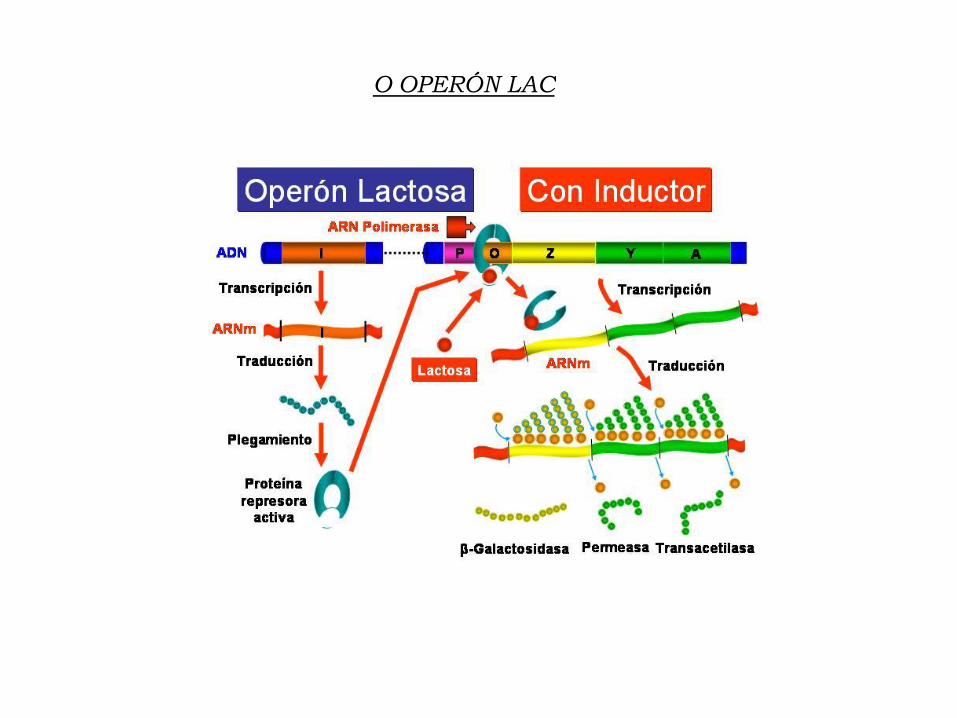
O OPERÓN LAC

9. REGULACIÓN DA EXPRESIÓN XÉNICA
9.2. Control da expresión xénica en eucariotas
Nos pluricelulares todas as células teñen tamén o mesmo ADN, pero non en
todas se expresa o mesmo, como xa sabemos, e ésta é a base da
diferenciación celular. Tamén sabemos que o nivel de condensación do ADN
está relacionado coa súa actividade transcricional e isto está regulado nas
células:
A acetilación das histonas por parte de enzimas favorece a transcrición.
A metilación do ADN impide a expresión.
Ademais a nivel de membrana hai distintos receptores hormonais que
permite a cada célula diana respostar a estos mensaxeiros químicos. A
regulación difire segundo o tipo de hormonas:
Hormonas lipídicas: atravesan a membrana, forman complexos hormona-
receptor intracelulares que poden unirse a xenes específicos.
Hormonas proteicas: mediante o sistema da adenilato-ciclasa que xa
vimos.

GRAZAS POR ATENDERME

WEBGRAFÍA
http://biologiacampmorvedre.blogspot.com.es/2013/02/bloque-iii_7399.html
http://www.bionova.org.es/biocast/tema19.htm
http://3.bp.blogspot.com/-
1DYI9JlJUK0/T1_42CIw1fI/AAAAAAAAABw/QnvhnVw_oL4/s1600/TTTTTT.jpg
https://commons.wikimedia.org/wiki/File:ADN_animation.gif
http://pendientedemigracion.ucm.es/info/genetica/grupod/Replicacion/Replicacion.
htm#Inicio
https://pendientedemigracion.ucm.es/info/genetica/grupod/Transcripcion/Transcripc
ion.htm
http://planetas.unipe.edu.ar/cienciayt/?cat=37


















