
RESUMEN - digeset.ucol.mxdigeset.ucol.mx/tesis_posgrado/Pdf/JOSE_ENCARNACION_DEL_RIO... · es un...
49
RESUMEN: Se investigaron los efectos del entrenamiento de resistencia y de velocidad sobre las propiedades mecánicas pasivas de los músculos rápidos y lentos. Ratas macho Wistar (n= 32) de 8 semanas de edad fueron divididas en tres grupos; Control (C), entrenados para desarrollar Resistencia (R) y entrenados para desarrollar velocidad (V). Los animales entrenaron durante 10 semanas en una banda sinfín motorizada. Al finalizar el programa de entrenamiento, los músculos Soleo y Plantaris fueron sometidos a dos protocolos de entrenamiento; en el primero, aplicando un estimulo supramáximo se provoco un tétanos y sobre la meseta del tétanos el músculo se deformo 0.33, 1 y 6 mm y se midió al amplitud del tétanos antes y después de la deformación. En el segundo protocolo, los músculos fueron deformados cíclicamente a 0.33 mm s -1 con incrementos de 1 mm hasta la ruptura. Las curvas esfuerzo deformación fueron construidas usando los valores de esfuerzo y deformación alcanzado en cada ciclo. Las curvas fueron ajustadas por medio de una ecuación exponencial sigmoidea. Los parámetros de ajuste para músculos entrenados y no entrenados muestran diferencias estadísticamente significativas. Los resultados descritos sugieren que el entrenamiento modifica la plasticidad de los elementos responsables de la tensión pasiva.
Transcript of RESUMEN - digeset.ucol.mxdigeset.ucol.mx/tesis_posgrado/Pdf/JOSE_ENCARNACION_DEL_RIO... · es un...

RESUMEN: Se investigaron los efectos del entrenamiento de resistencia y de velocidad sobre las
propiedades mecánicas pasivas de los músculos rápidos y lentos. Ratas macho Wistar
(n= 32) de 8 semanas de edad fueron divididas en tres grupos; Control (C), entrenados
para desarrollar Resistencia (R) y entrenados para desarrollar velocidad (V). Los
animales entrenaron durante 10 semanas en una banda sinfín motorizada.
Al finalizar el programa de entrenamiento, los músculos Soleo y Plantaris fueron
sometidos a dos protocolos de entrenamiento; en el primero, aplicando un estimulo
supramáximo se provoco un tétanos y sobre la meseta del tétanos el músculo se
deformo 0.33, 1 y 6 mm y se midió al amplitud del tétanos antes y después de la
deformación.
En el segundo protocolo, los músculos fueron deformados cíclicamente a 0.33 mm s-1
con incrementos de 1 mm hasta la ruptura. Las curvas esfuerzo deformación fueron
construidas usando los valores de esfuerzo y deformación alcanzado en cada ciclo. Las
curvas fueron ajustadas por medio de una ecuación exponencial sigmoidea.
Los parámetros de ajuste para músculos entrenados y no entrenados muestran
diferencias estadísticamente significativas.
Los resultados descritos sugieren que el entrenamiento modifica la plasticidad de los
elementos responsables de la tensión pasiva.

i
ABSTRACT: We investigated the effects of endurance and sprint training on the passive
mechanical properties of fast-twitch (FT) and slow-twitch (ST) skeletal muscles.
Eigth-week-old male Wistar rats (n= 32) were divided into three groups; control
(C), sprint-trained (S) and endurance-trained. The trained animals exercised for
10 weeks on a treadmill. Under anaesthesia, After the training the Plantaris and
Soleo muscles were submitted in two protocols of investigation; in the first one,
stimulus supramaximum were applied across the motor nerve to provoke
tetanus. On the plateau of this one deformations of 0.33, 1 and 6 mm were
applied and the force measured up, earlier and after the deformation, The
second protocol the muscles were deformed cyclically in vivo at 0.33 mm s-1
with length increments of 1 mm in successive cycles until rupture. The rupture
of muscle occurs at belly. Stress-strain relation were constructed using the
maximum stress and maximum strain in each cycle. The data were fitted to an
S-shaped curve. The curve-fitting parameters for trained and untrained muscles
showed significant statistical differences. Stress-strain at rupture and maximum
deformation energy were statistically greater for trained ST musclers (both
groups) than the controls.. The above results suggest the plasticity of passive
structures caused by activity demands.

i
ii

INDICE GENERAL Pag. RESUMEN i ABSTRACT ii INTRODUCCION 1 Músculos Esqueleticos 4 Constitución de las Miofibrillas y sus componentes 5 Ultraestructura del filamento delgado 5 Ultraestructura del filamento grueso 6 Ultraestructura del filamento conector 6 Filamentos intermedios 8 Otras estructuras sarcómericas 9 Tejido conectivo 9 Tejido elástico 11 MARCO TEORICO 12 Componentes de la fuerza muscular 13 HIPOTESIS 17 OBJETIVOS 19 MATERIALES Y METODOS 21 Animales de experimentación 22 Programa de entrenamiento 22 Cirugía 24 Protocolo experimental 1 25. Protocolo experimental 2 27 Análisis mecánico 27 Método estadístico 28 RESULTADOS 29 Efectos del entrenamiento sobre el músculo Soleo 31 Primer protocolo 31 Segundo protocolo 31 Efectos del entrenamiento sobre el músculo Plantaris 35 Primer protocolo 35 Segundo protocolo 36 DISCUSION 40 CONCLUSIONES 44 BIBLIOGRAFIA 46 APENDICES 51 Principios del entrenamiento deportivos 52 Trabajos publicados 54

I N T R O D U C C I O N

Toda actividad en los seres vivos requiere de movimiento, si caminamos o
corremos, si lanzamos algo o lo atrapamos, si miramos o escuchamos o aun, si
solo estamos parados requerimos de la contracción de los músculos
esqueléticos (MacComas, 1996)
El funcionamiento de los músculos esqueléticos se basa en mecanismos
complejos, así la enorme diversidad de funciones de los músculos depende de
su tamaño, forma y composición. Los músculos esqueléticos de mamífero están
formados por dos diferentes tipos de fibras musculares que por su aspecto las
clasificaron en fibras rojas y fibras blancas. Más tarde explorando sus
propiedades mecánicas e histoquímicas, se agruparon en fibras de Sacudida
Rápida (FT) y fibras de sacudida lenta (ST).
Los músculos esqueléticos se han clasificado en rápidos y lentos según el
tiempo de contracción que ejecutan la mayoría de sus fibras musculares, sin
embargo, los estudios realizados demuestran que, los músculos ademas de
realizar tensión activa (contracción), responden a los cambios de longitud
desarrollando tensión pasiva. En condiciones fisiológicas, la tensión pasiva, se
origina a partir del tono muscular y de las propiedades elásticas de las proteínas
estructurales y es modulada por estímulos externos.
Las fibras musculares de sacudida lenta (ST) resisten la fatiga y pueden
mantener tensión por largos periodos de tiempo, muestran una menor actividad
de la miosina ATPasa y están adaptadas para la producción aeróbica de
energía. Por lo tanto, los músculos formados por una mayor proporción de fibras
de sacudida lenta desarrollan una fuerza sostenida, lo que les permite mantener
la postura pero también realizan contracciones lentas y graduadas para realizar
movimientos finos, como el control del movimiento muscular esta dirigido por el
Sistema Nervioso Central (SNC) a través de las motoneuronas, cada
motoneurona inerva a cierto número de fibras musculares de un tipo específico
formando las Unidades motoras. Las unidades motoras de las fibras ST están

inervadas por moto-neuronas que disparan impulsos nerviosos a baja frecuencia
y con baja velocidad de conducción.
Por otro lado, Los músculos que contienen mayor proporción de fibras de
sacudida rápida (FT) desarrollan una fuerte y rápida tensión; las unidades
motoras, que conforman estos músculos, están inervadas por motoneuronas
que conducen sus impulsos a altas velocidades pero que mantienen su actividad
por cortos periodos de tiempo. Estos músculos están adaptados para realizar
movimientos rápidos.
La proporción de los diferentes tipos de fibras en cada músculo esta
determinada genéticamente, así, por ejemplo el músculo Soleo de Ratas Wistar
es un músculo lento, porque esta compuesto por 85% de fibras ST, mientras que
el Plantaris es un músculo rápido porque esta compuesto por un 85% de fibras
FT. (Bigard y cols, 1991; Saubert y cols, 1973).
Los componentes contráctiles de las fibras musculares, actina y miosina,
son proteínas intracelulares dispuestas en un arreglo ordenado y que junto a los
elementos mecánicos pasivos, permiten que el músculo, desde el punto de vista
de la ingeniería, se considere una maquina diseñada para realizar movimientos.
Estímulos externos tales como: inmovilización de un miembro (Guezennec
y col.; 1990), estimulación crónica (Jarvis y cols; 1996) y actividad física
(Almeida-Silveira y cols, 1994; Bigard y cols, 1991; Goubel y cols, 1987;
Kovanen 1984; Takekura y Yoshioka, 1990; Woo y cols, 1980; Pousson, 1991;
Woo y cols, 1981). Inducen alteraciones morfométricas, histoquímicas y
contráctiles en los músculos
Woo y cols (1981) mostraron que el ejercicio prolongado incrementa la
fuerza máxima y la pendiente de la curva esfuerzo-deformación del músculo
extensor del tendón del cerdo; Kovanen y cols. (1984), reportan que el
entrenamiento de resistencia incrementa el modulo tangente y la máxima
energía de deformación del músculo Soleo de rata e incrementa la carga

máxima, la deformación máxima y la máxima energía de deformación del
músculo rectus femoris.
Se obtienen efectos diferentes dependiendo del tipo de entrenamiento que
se realice: por ejemplo, el entrenamiento de resistencia incrementa la rigidez del
tendón (Woo y cols, 1980) y la rigidez muscular (Goubel y Marini, 1987) del
músculo Soleo; mientras que el entrenamiento de velocidad disminuye la rigidez
muscular (Pousson y cols., 1991).
Almeida-Silveira y col. (1994), estudiaron los efecto de ciclos de
acortamiento-estiramiento un tipo de entrenamiento de velocidad en el músculo
Soleo de rata y encontraron que este tipo de estimulo incrementa la rigidez de
los componentes elásticos en serie del músculo.
Si una fuerza externa es aplicada al músculo, este sufre algunas
modificaciones en sus dimensiones. Bajo ciertos limites, si la fuerza externa
deformante es retirada antes de llegar al limite elástico, el músculo regresa a su
forma original.
Así, las propiedades mecánicas pasivas del músculo se relacionan con el
desarrollo de la tensión pasiva y con la capacidad del músculo de tolerar el
esfuerzo mecánico. Las curvas Esfuerzo-deformación una herramienta de la
ingeniería de materiales, que aplicada al tejido muscular, nos permite estudiar
las propiedades mecánicas pasivas de los músculos (Nubar, 1962).
Músculos esqueléticos:
Los músculos esqueléticos varían en forma y tamaño, sin embargo si un
músculo es cortado transversalmente se pueden observar diferentes
compartimentos llamados fascículos. Cada fascículo esta formado por un
numero aproximado de 100 o 150 células musculares individuales llamadas
fibras musculares, cuyo diámetro varia de entre 10 a 100 ìm y con una longitud
de 1 a 40 mm. Cuando las fibras musculares son observadas a través del
microscopio, muestran una secuencia de bandas o estriaciones. Este patrón de

organización en bandas, refleja la organización estructural de las miofibrilla. Así,
para entender como el músculo se contrae, se relaja o se estira, debemos
conocer la estructura de las miofibrillas.
Constitución de las miofibrillas y sus componentes:
Utilizando microscopio electrónico (figura 1), se observa que cada fibra
muscular esta formada por un gran numero de unidades mas pequeñas
ordenadas en sentido longitudinal a la fibra muscular llamadas miofibrillas,. Cada
miofibrilla, tiene un diámetro de entre 1 a 2 ìm y esta constituida por un patrón
repetido de bandas que constituyen los sarcómeros. Los sarcómeros están
delimitados en cada extremo por unas bandas densas llamadas línea Z (tambien
conocidas como disco Z o bandas Z) y representan la unidad funcional del
músculo. Entre cada linea Z, dentro de los sarcómeros, se encuentran
estructuras aun mas pequeñas, llamados filamentos o miofilamentos.
Originalmente, se pensaba que solo habia dos clases de filamentos; los
filamentos gruesos formados por la proteína Miosina y los filamentos delgados
formados principalmente por la Actina. Sin embargo entre 1970 y 1980 fue
descrito un tercer tipo de filamento llamado, filamento conector.
Ultraestructura del filamento delgado:
El filamento delgado, llamado de Actina tiene un diámetro de
aproximadamente 5 o 6 nm y una longitud de 1 ìm. A baja resolución este
filamento se asemeja a dos collares de perlas enrollados entre si.
Sin embargo, la actina no es el único componente del filamento delgado,
la troponina, tropomiosina y nebulina, forman parte del mismo.

Ultraestructura del filamento grueso:
El filamento grueso o de Miosina, tienen un diámetro de aproximadamente
10 a 15 nm y una longitud de 1.5 um. Este filamento posee numerosas
proyecciones laterales, que se extienden hacia los filamentos de Actina. Estas
proyecciones son llamadas colectivamente “Puentes cruzados” y son los sitios
de unión entre los filamentos de Actina y de Miosina.
Ultraestructura del filamento conector:
El modelo contemporáneo de dos filamentos, propuesto por H. E. Huxley y
Hanson (1954) y por A. F. Huxley y Niedergerke (1954), sugiere que el músculo
se acorta debido a la unión y deslizamiento de los filamentos gruesos y
delgados. Sin embargo, si observamos una ilustración clásica de la organización
del sarcómero de aquel modelo (Figura 1), surgen dos dudas: (1) ¿Que hace
que el filamento grueso se mantenga centrado en el sarcómero? y (2) ¿que
mantiene unido al sarcómero?.
La respuesta es; la presencia de un filamento conector.
Wang (1985), comenzó un análisis bioquímico detallado de la proteína
conectina. Las investigaciones de Wang indicaron que la Conectina esta
compuesta de dos proteínas: titina y Nebulina.
La titina es la proteína mas grandes descrita hasta este momento, con una
masa molecular de 2.5 a 3.0 daltones (Maruyama y cols, 1984) y constituye
cerca del 10% de la masa miofibrilar. Esta proteína se extiende desde la línea Z,
en el extremo del sarcómero hasta unirse firmemente al filamento grueso.
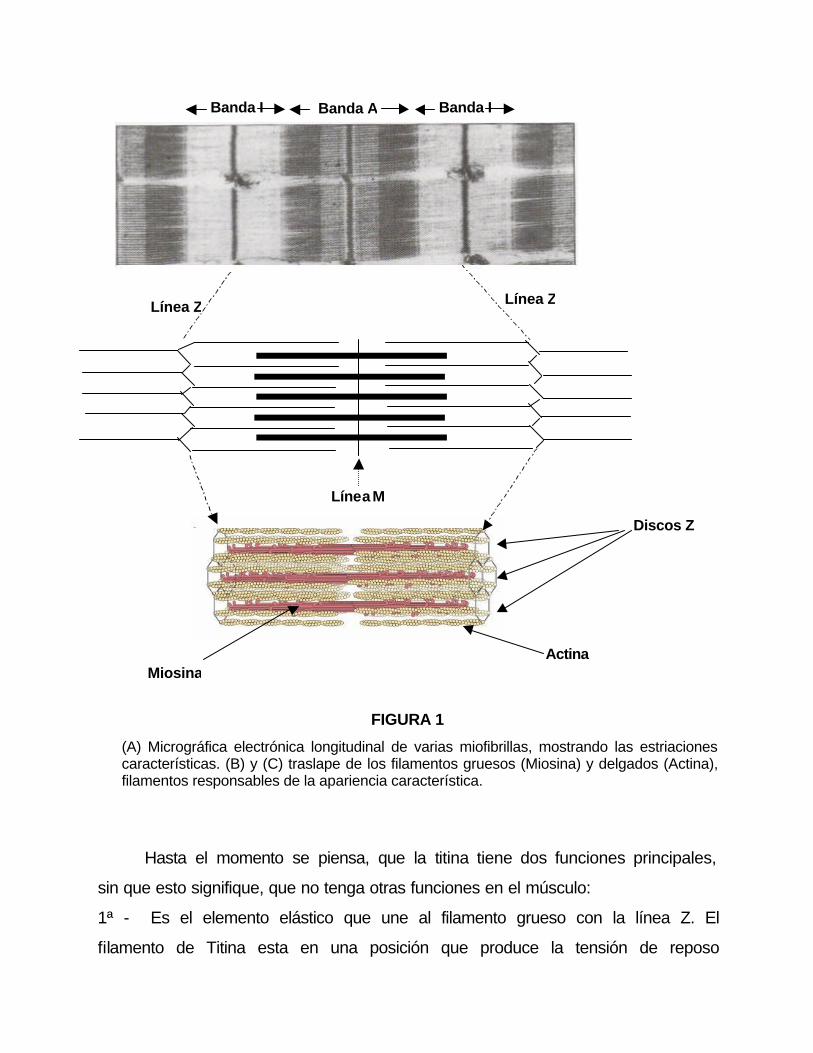
FIGURA 1
(A) Micrográfica electrónica longitudinal de varias miofibrillas, mostrando las estriaciones características. (B) y (C) traslape de los filamentos gruesos (Miosina) y delgados (Actina), filamentos responsables de la apariencia característica.
Hasta el momento se piensa, que la titina tiene dos funciones principales,
sin que esto signifique, que no tenga otras funciones en el músculo:
1ª - Es el elemento elástico que une al filamento grueso con la línea Z. El
filamento de Titina esta en una posición que produce la tensión de reposo
Banda I Banda A Banda I
Línea M
Discos Z
Línea ZLínea Z
MiosinaActina

(tensión que esta presente cuando la fibra muscular esta en una longitud
fisiológica y que incrementa cuando una fibra relajada es estirada),
proporcionando la fuerza que tiende a centrar al filamento grueso dentro del
sarcómero. Además, estos filamentos elásticos podrían transmitir la fuerza activa
hacia los discos Z, fuerza que de otra manera no podría alcanzar nunca los extremos
del sarcómero
2º - Se ha especulado que la Titina, juega un papel importante en la
morfogénesis de la miofibrilla (Pollack, 1990).
Filamentos intermedios
Las proteínas Desmina, Vimectina y Sinemina forma parte de una
compleja red de proteínas que envuelven a las miofibrillas a nivel de los discos Z
y los mantienen juntos y alineados tranversalmente. Este arreglo proteico que
conecta los discos Z de una miofibrilla con los discos Z vecinos, se une tambien
a proteínas del citoesqueleto (Actina, Distrofina y Espectrina), constituyendo una
superestructura que se repite periódicamente a lo largo del interior de la fibra
muscular y que se denomina “Costámero”. (figura 2). Desde la parte interna de
la membrana plasmática, la Actina del citoesqueleto se liga con dímeros de
Distrofina y vinculina, los cuales a su vez se ensamblan con otras proteínas
transmembranales. Estas proteínas unen al plasmalema con la matriz
extracelular, anclando a los costámeros a la matriz extracelular. Debido a esta
organización proteíca, las fibras musculares se encuentran alineadas
transversalmente desde el sarcomero, las miofibrillas, las fibras musculares y el
músculo completo, confiriéndole a los músculos esqueléticos un acople
mecánico para la contracción (figura 1).

Otras estructuras sarcomericas:
Las estructuras miofibrilares de los sarcómeros, deben de mantenerse
juntos no solo en el sentido longitudinal, ya que tambien necesitan de soporte
en el sentido transversal, por ejemplo, deben proteger al músculo cuando este
es aplastado.
Las fuentes de resistencia transversal que mantiene la integridad del
sarcómero son tres estructuras como puentes denominados: puentes M,
puentes A y puentes I (figura 3).
Tejido conectivo:
El tejido conectivo esta formado por una amplia variedad de células
especializadas, cuyas funciones son: protección de órganos huecos,
almacenamiento de energía, unión, conexión y en general soporte y reparación.
FIGURA 2 Esquema de la fijación de las líneas Z por medio de los filamentos intermedios, los últimos se unen tambien a elementos de citoésqueleto por debajo del plasmalema. Tomado de McComas, A. J. Skeletal muscle, Human Kinetics, 1996. p. 15
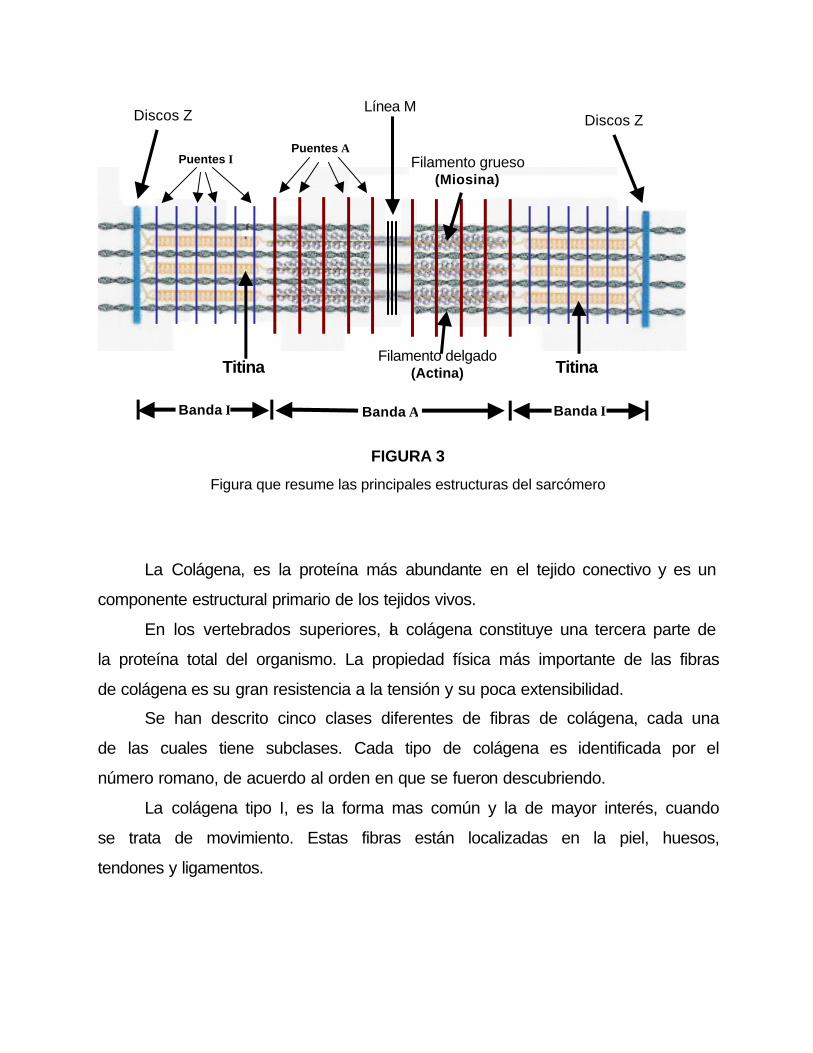
FIGURA 3
Figura que resume las principales estructuras del sarcómero
La Colágena, es la proteína más abundante en el tejido conectivo y es un
componente estructural primario de los tejidos vivos.
En los vertebrados superiores, la colágena constituye una tercera parte de
la proteína total del organismo. La propiedad física más importante de las fibras
de colágena es su gran resistencia a la tensión y su poca extensibilidad.
Se han descrito cinco clases diferentes de fibras de colágena, cada una
de las cuales tiene subclases. Cada tipo de colágena es identificada por el
número romano, de acuerdo al orden en que se fueron descubriendo.
La colágena tipo I, es la forma mas común y la de mayor interés, cuando
se trata de movimiento. Estas fibras están localizadas en la piel, huesos,
tendones y ligamentos.
Discos Z Discos Z Línea M
Filamento grueso (Miosina)
Filamento delgado (Actina) Titina Titina
Banda I Banda A Banda I
Puentes IPuentes A

Tejido elástico:
El tejido elástico, es tambien un componente estructural en los tejidos
vivos. En el sarcolema de la fibra muscular (tejido conectivo que rodea el
sarcómero), se encuentran grandes cantidades de tejido elástico, que determina
en gran medida el rango de extensibilidad de las fibras musculares. El
componente principal del tejido elástico es la Elastina, que es un estructura
compleja con propiedades elásticas, como su nombre lo indica, y que se deben
a la composición bioquímica y el arreglo físico de sus moléculas.

MARCO TEORICO
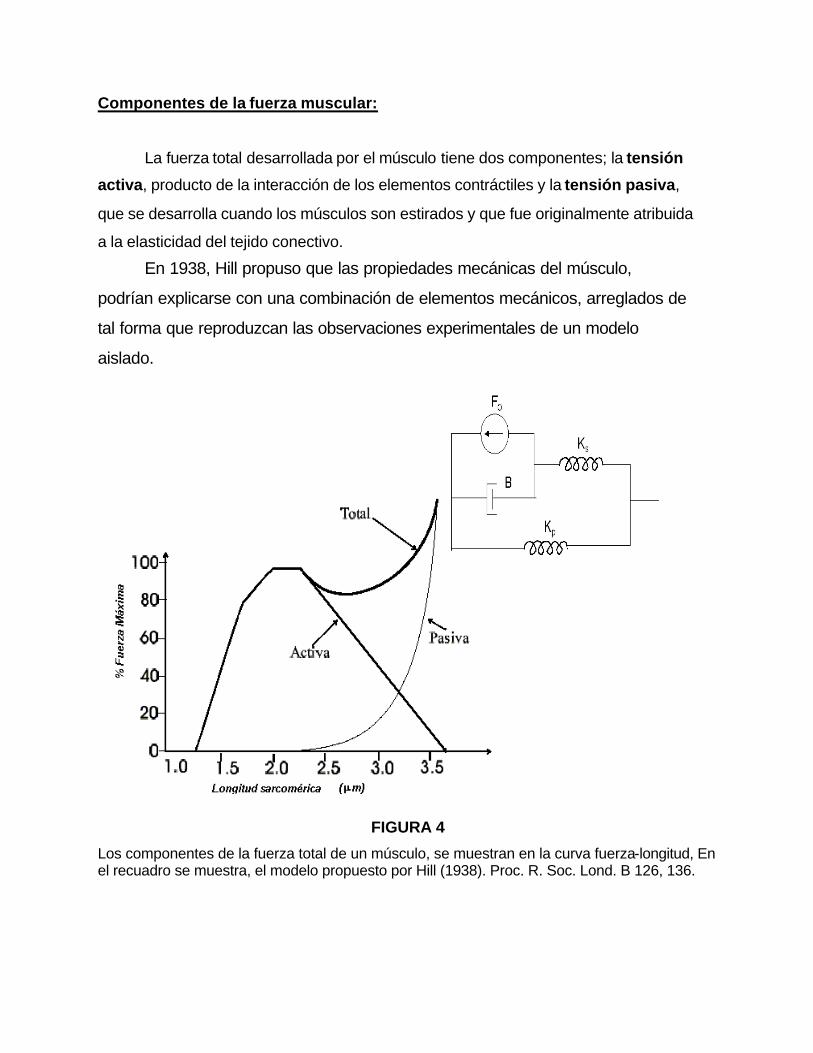
Componentes de la fuerza muscular:
La fuerza total desarrollada por el músculo tiene dos componentes; la tensión
activa, producto de la interacción de los elementos contráctiles y la tensión pasiva,
que se desarrolla cuando los músculos son estirados y que fue originalmente atribuida
a la elasticidad del tejido conectivo.
En 1938, Hill propuso que las propiedades mecánicas del músculo,
podrían explicarse con una combinación de elementos mecánicos, arreglados de
tal forma que reproduzcan las observaciones experimentales de un modelo
aislado.
FIGURA 4
Los componentes de la fuerza total de un músculo, se muestran en la curva fuerza-longitud, En el recuadro se muestra, el modelo propuesto por Hill (1938). Proc. R. Soc. Lond. B 126, 136.

Cada uno de los componentes de la curva fuerza-longitud (figura 4),
pueden ser representados por los elementos mostrados en el modelo del
recuadro superior derecho. Según este modelo, la tensión activa es
representada por un motor (Fo), mientras que la tensión pasiva, depende de la
interacción de los siguientes elementos
-El elemento viscoso (B), que lo producen la interacción de las proteínas
con el citoplasma de la fibra muscular.
-El elemento elástico en serie (Ks), que se atribuye a los puentes cruzados.
-El elemento elástico en paralelo (Kp), que es responsabilidad de dos
estructuras; por un lado, la amplia red de filamentos intermedios, que interconectan
a los sarcómeros a nivel de los discos Z, con el citoesqueleto asociado al sarcolema,
estructuras denominadas costámeros (figura 2), que son potencialmente productora de
tensión pasiva en las células musculares. Por otro lado, el estiramiento de los
filamentos conectores de titina, contribuye en gran parte al desarrollo de tensión
pasiva, (Horowits, y col., 1986; Horowits y Podolsky, 1987; Funatsu, y col.., 1990;
Horowits, 1992). La titina cumple con el papel principal de generar tensión pasiva
y transmitir tensión activa, ademas, contribuye a la estabilización de los
filamentos gruesos en el centro de la sarcómera
Según el modelo de Wang (1993), la curva de pasiva–longitud
sarcomérica puede ser dividida en cuatro zonas (figura 5);
Zona I, donde no se genera tensión significativa, como resultado del
enderezamiento de los filamentos laxos de titina sin un cambio neto en la
longitud del contorno.
Zona II, donde la tensión aumenta de manera exponencial hasta alcanzar un
máximo, en esta zona el filamento conector de titina extiende su longitud
linealmente, generando fuerza.

Figura 5 (A).- Interpretación del comportamiento estructural del sarcómero durante el estiramiento. (B).- Descripción de las diferentes Zonas descritas en A. (C).- Interpretación de la participación de los filamentos intermedios durante el estiramiento, antes y después de la extracción de los miofilamentos con yoduro de potasio.
Zona III, inicia donde la tensión alcanzó. Esta reducción puede ser causada por
un incremento neto de la longitud del segmento extensible de la titina.
Zona IV, se presenta una segunda elevación de la tensión, que refleja la
contribución adicional de la red de filamentos intermedios que rodean cada
miofibrilla.
. En animales superiores, la producción de tensión de reposo durante el
alargamiento pasivo podría limitar el grado de movimiento producido por el
músculo opuesto durante el acortamiento activo. Ademas, la producción de
tensión de reposo podría ayudar a proteger a los músculos del daño debido a la
sobreextensión.

Muchas investigaciones se han centrado en estudiar los efectos del
entrenamiento sobre la tensión activa de los músculos esqueléticos, sin
embargo, este trabajo considera que los elementos responsables de la tensión
pasiva, participan desde el inicio de la curva de tensión (figura 4), cuando la
longitud de los sarcómeros no es muy grande y que el entrenamiento puede
modificar estos elementos.

H I P O T E S I S

Las propiedades mecánicas pasivas de los músculos esqueléticos son moduladas por el entrenamiento físico y están relacionadas con su habilidad para tolerar el esfuerzo mecánico.

O B J E T I V O S

Estudiar los efectos del entrenamiento de resistencia sobre las
propiedades mecánicas pasivas de músculos completos rápidos (Plantaris) en
ratas macho Wistar, utilizando las curvas esfuerzo-deformación y medir la
persistencia de desarrollo activo de tensión, posterior a los ciclos de
deformación progresivos.
Estudiar los efectos del entrenamiento de velocidad sobre las propiedades
mecánicas pasivas de músculos completos rápidos (Plantaris) en ratas macho
Wistar, utilizando las curvas esfuerzo-deformación medir la persistencia de
desarrollo activo de tensión posterior a los ciclos de deformación progresivos.
Estudiar los efectos del entrenamiento de resistencia sobre las
propiedades mecánicas pasivas de músculos completos lentos (Soleo) en ratas
macho Wistar, utilizando las curvas esfuerzo-deformación y medir la persistencia
de desarrollo activo de tensión posterior a los ciclos de deformación progresivos.
Estudiar los efectos del entrenamiento de velocidad sobre las propiedades
mecánicas pasivas de músculos completos lentos (Soleo) en ratas macho
Wistar, utilizando las curvas esfuerzo-deformación y medir la persistencia de
desarrollo activo de tensión posterior a los ciclos de deformación progresivos.

MATERIALES Y METODOS

Animales de experimentación:
Se utilizaron para el estudio Ratas macho Wistar, de 8 semanas de edad,
que fueron separados en tres grupos: un grupo de animales no entrenados o
Control (C, n = 12; 232 ± 38 g. de peso corporal) y dos grupos de animales
entrenados para desarrollar Resistencia (R, n = 11, 256 ± 22 g. de peso
corporal) y Velocidad (V, n = 9, 258 ± 26g. de peso corporal), no se observo
diferencia estadísticamente significativa en el peso corporal de los diferentes
grupos experimentales. Todos los animales de experimentación fueron alojados,
en jaulas individuales de acrílico bajo condiciones ambientales naturales
(Temperatura 26 ± 4 °C, 60-70% de humedad y ciclo natural de luz / oscuridad).
La comida y el agua fueron proporcionadas ad libitum. La dieta de los animales
contenía; 24% de proteína, 8% de grasas, 4.5% de fibra, 39.3% de
carbohidratos, 12% de agua y 12.2% de minerales y vitaminas.
Programa de entrenamiento:
Las ratas fueron entrenadas durante 10 semanas en una banda sinfín
motorizada (Modular Enclosed Treadmill; Columbus Instrument, Columbus, OH,
USA). En la parte posterior de la banda se utilizó un dispositivo eléctrico para
aplicar estímulos de 0.35 a 1.5 mA.
Las primeras 4 semanas, los animales realizaron una sesión diaria de
entrenamiento, con duración de 15-20 minutos a una velocidad de 16 m min-1
(65% del VO2max.). Al finalizar la séptima semana, las ratas del grupo R,
corrieron una hora diaria a 20 m min-1,(80% del VO2max) 5 días por semana.
Mientras que las ratas de grupo V tres veces por semana, realizaron series
crecientes de carreras de 20 segundos de duración a una velocidad de 60 m
min-1 (95% del VO2max) intercaladas por periodos de reposo de 5 minutos. Los
2 días restantes realizaron sesiones de recuperación con 15 a 20 minutos de
duración, a una velocidad de 16 m min-1.
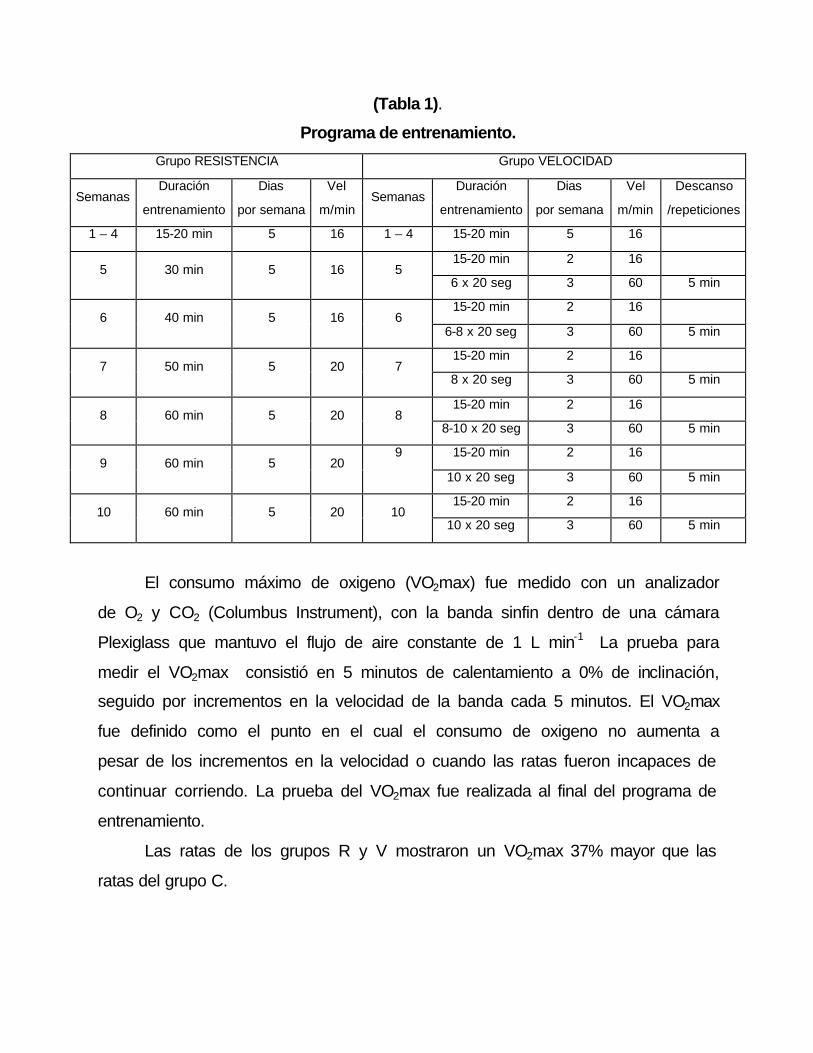
(Tabla 1).
Programa de entrenamiento.
Grupo RESISTENCIA Grupo VELOCIDAD
Duración Dias Vel Duración Dias Vel Descanso Semanas
entrenamiento por semana m/min Semanas
entrenamiento por semana m/min /repeticiones
1 – 4 15-20 min 5 16 1 – 4 15-20 min 5 16
15-20 min 2 16 5 30 min 5 16 5
6 x 20 seg 3 60 5 min
15-20 min 2 16 6 40 min 5 16 6
6-8 x 20 seg 3 60 5 min
15-20 min 2 16 7 50 min 5 20 7
8 x 20 seg 3 60 5 min
15-20 min 2 16 8 60 min 5 20 8
8-10 x 20 seg 3 60 5 min
9 15-20 min 2 16 9 60 min 5 20
10 x 20 seg 3 60 5 min
15-20 min 2 16 10 60 min 5 20 10
10 x 20 seg 3 60 5 min
El consumo máximo de oxigeno (VO2max) fue medido con un analizador
de O2 y CO2 (Columbus Instrument), con la banda sinfin dentro de una cámara
Plexiglass que mantuvo el flujo de aire constante de 1 L min-1 La prueba para
medir el VO2max consistió en 5 minutos de calentamiento a 0% de inclinación,
seguido por incrementos en la velocidad de la banda cada 5 minutos. El VO2max
fue definido como el punto en el cual el consumo de oxigeno no aumenta a
pesar de los incrementos en la velocidad o cuando las ratas fueron incapaces de
continuar corriendo. La prueba del VO2max fue realizada al final del programa de
entrenamiento.
Las ratas de los grupos R y V mostraron un VO2max 37% mayor que las
ratas del grupo C.

Cirugía:
Después de 10 semanas de entrenamiento, las ratas fueron anestesiadas
por la vía intraperitoneal con pentobarbital sodico (Anestesal; 4.0-6.0 mg x
100mg de PC). Los músculo Soleo y Plantaris del miembro posterior derecho,
fueron liberados del tejido circundante, dejando intactos la inserción ósea y el
riego sanguíneo. En nervio motor fue cortado lo mas retirado posible del
músculo. Durante la cirugía, el tejido se mantuvo húmedo por la aplicación
constante de solución salina.
Se colocaron marcas en las uniones músculo-tendón, para facilitar la
medición del músculo. Posteriormente utilizando un taladro de dentista se
perforó el fémur para fijar el animal al dispositivo experimental (figura 5), que
consiste en una placa de acrílico montada en una base que permite alinear el
músculo perpendicularmente a un transductor de fuerza (FT10, Grass Co.,
Quincy, MA, USA), el Transductor estaba conectado a un Polígrafo Grass y
montado en un micromanipulador que a su vez era movido por una motor de
pasos controlado por medio de una computadora. Dos postes fijos en la placa de
acrílico servían para fijar la extremidad del animal, pasando una barra de acero
a través del orificio previamente realizado en el fémur. La inserción distal del
músculo se unió al transductor por medio de un gancho y una cadena de acero.
El nervio motor correspondiente fue colocado sobre un electrodo de
estimulación que estaba conectado a un estimulador (S88, Grass).
Con el fin de obtener el rango fisiológico de deformación, se medio la
longitud muscular aplicando hiper-extensión del talón y tétanos a cada músculo
antes de realizar la perforación en el fémur y el la inserción distal.
Previo a los protocolos experimentales que se describen a continuación,
se aplicaron estímulos supramáximos a través del nervio motor a diferentes
longitudes del músculo hasta alcanzar la longitud optima (L0), definida por la
máxima amplitud de la sacudida muscular en respuesta a un estimulo
supramaximal.

FIGURA 5
El dispositivo experimental consistió en una placa de acrílico (P) montada sobre una base que permite que el músculo sea alineado perpendicularmente a un transductor de fuerza FT-10 (T); (Grass, Co.). El transductor (T) estaba montado sobre un micromanipulador movido por un motor de pasos (M) por medio de una computadora y conectado a un Polígrafo Grass. La placa de acrílico (P), Tenia dos postes de metal donde se fijo una barra que atravesaba el fémur. La inserción distal del músculo se unió al transductor (T) por medio de un gancho y una cadena metálica. El nervio motor correspondiente fue colocado sobre los electrodos de estimulación (S) que a su vez estaban conectados a un estimulador (S88, Grass, Co.).
Durante los experimentos la temperatura corporal de la rata fue mantenida
a 37 ± 0.5 °C. La temperatura ambiental fue de aproximadamente 24 °C.
Protocolo experimental 1:
Con el fin de valorar la tolerancia a la deformación en el estado activo
(tétanos), se estimulo el nervio motor, para provocar tétanos con una duración
de 6 s, Poco antes de la mitad de la meseta de cada tétano se efectuaban
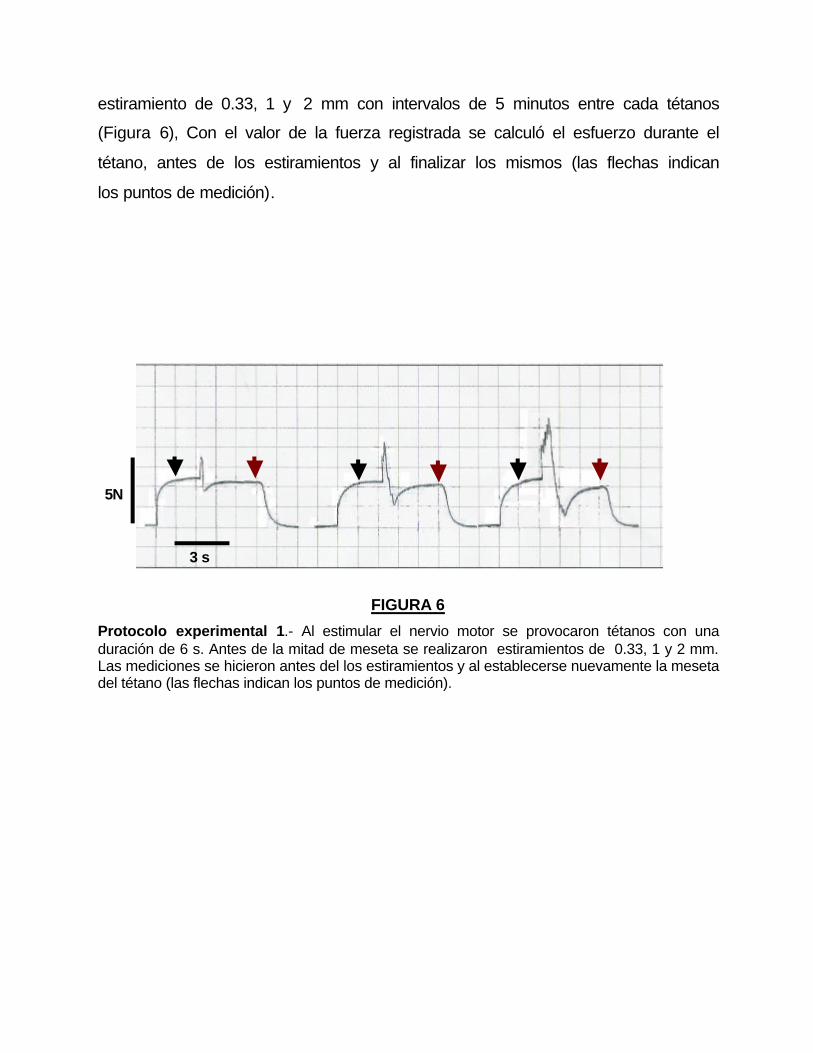
estiramiento de 0.33, 1 y 2 mm con intervalos de 5 minutos entre cada tétanos
(Figura 6), Con el valor de la fuerza registrada se calculó el esfuerzo durante el
tétano, antes de los estiramientos y al finalizar los mismos (las flechas indican
los puntos de medición).
FIGURA 6
Protocolo experimental 1.- Al estimular el nervio motor se provocaron tétanos con una duración de 6 s. Antes de la mitad de meseta se realizaron estiramientos de 0.33, 1 y 2 mm. Las mediciones se hicieron antes del los estiramientos y al establecerse nuevamente la meseta del tétano (las flechas indican los puntos de medición).
5N
3 s

Protocolo experimental 2:
Para determinar la persistencia de la tensión activa después del
estiramiento, se aplicaron cada 5 minutos ciclos de deformación con una
velocidad de
0.33 mm s-1, con incrementos de 1 mm a partir de L0 hasta llegar a la ruptura del
músculo, después de cada ciclo se registraba una sacudida y un tétano. (Figura
7). Al final de los experimentos las ratas recibieron una sobredosis de
pentobarbital sódico.
FIGURA 7
Protocolo experimental 2: La figura muestra que posterior a los ciclos de deformación (δδ ) eran provocados por estimulación del nervio motor, una sacudida (s) y un tétanos (t). Con los valores de tensión en cada ciclo de deformación se construyeron las curvas σσ -δδ , mientras que, de la tensión registrada en las sacudidas (s) y tétanos (t) se obtuvieron las figuras 6 y 9.
Análisis mecánico:
La tensión máxima en cada deformación (Fuerza) fue usada para construir
las curvas Esfuerzo-deformación (curvas σ-δ).
El esfuerzo (σ en Nm-2) fue calculado a partir de la formula:
1.- σ = Fuerza / ACT
1N3
6
9
5N
s t

El área de corte transversal (ACT) fue calculado por medio de la siguiente
ecuación (Méndez y Keys, 1990):
2.- ACT = P / Lf * 1.056
Donde P es el peso del músculo en gramos; Lf es la longitud en centímetros, al
final de cada estiramiento; y 1.056 g cm-1 es la densidad del músculo.
La deformación del músculo en cada ciclo fue obtenida a partir de:
3.- δδ = L f - L 0
L0
Las curvas esfuerzo-deformación fueron ajustadas con una ecuación
exponencial sigmoidea.
4.- σ = σmax / (1 ± exp(-k (δ - δ50)))
Donde: δ50 es el valor de la deformación al cual la curva de ajuste
alcanza el 50% del esfuerzo máximo (σmax). El esfuerzo máximo es el mejor
ajuste de la curva en la ruptura del músculo y ê es una constante que representa
la velocidad de crecimiento de la curva. Estos parámetros fueron utilizados para
comparar las curvas σ-δ de los animales entrenados y no entrenados.
Los valores del esfuerzo (σU) y la deformación (δU) de ruptura son los
valores del esfuerzo y la deformación al momento de rompimiento del músculo,
fueron obtenidos de las curvas σ-δ y promediadas en cada grupo.
Con la ayuda de un digitalizador la Energía Máxima de Deformación (EMD) fue
medida como el área bajo la curva σ-δ.
Método estadístico:
Se calcularon para la estadística descriptiva, las medias de los valores y el error
estándar, utilizando análisis de varianza (ANOVA), con un test de Scheffe post hoc
para comparar los valores de los grupos C, R y V.

R E S U L T A D O S .
Después de 10 semanas de entrenamiento el peso corporal de los
animales de los grupos Resistencia (n = 11, 386 ± 37 g.) y Velocidad (n = 9, 355
± 46 g) no fueron significativamente diferentes con el grupo Control (n = 12; 381
± 52).
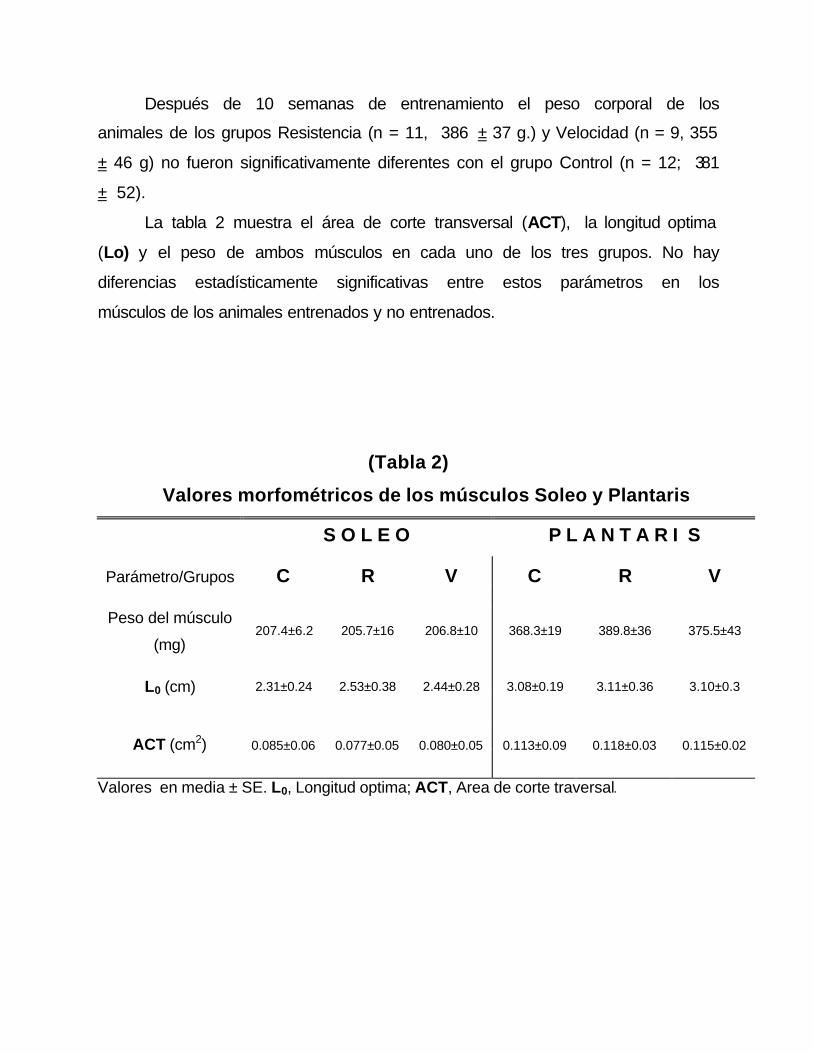
La tabla 2 muestra el área de corte transversal (ACT), la longitud optima
(Lo) y el peso de ambos músculos en cada uno de los tres grupos. No hay
diferencias estadísticamente significativas entre estos parámetros en los
músculos de los animales entrenados y no entrenados.
(Tabla 2)
Valores morfométricos de los músculos Soleo y Plantaris
S O L E O P L A N T A R I S
Parámetro/Grupos C R V C R V
Peso del músculo
(mg) 207.4±6.2 205.7±16 206.8±10 368.3±19 389.8±36 375.5±43
L0 (cm) 2.31±0.24 2.53±0.38 2.44±0.28 3.08±0.19 3.11±0.36 3.10±0.3
ACT (cm2) 0.085±0.06 0.077±0.05 0.080±0.05 0.113±0.09 0.118±0.03 0.115±0.02
Valores en media ± SE. L0, Longitud optima; ACT, Area de corte traversal.

Efectos del entrenamiento en los músculos Soleo:
Primer protocolo:
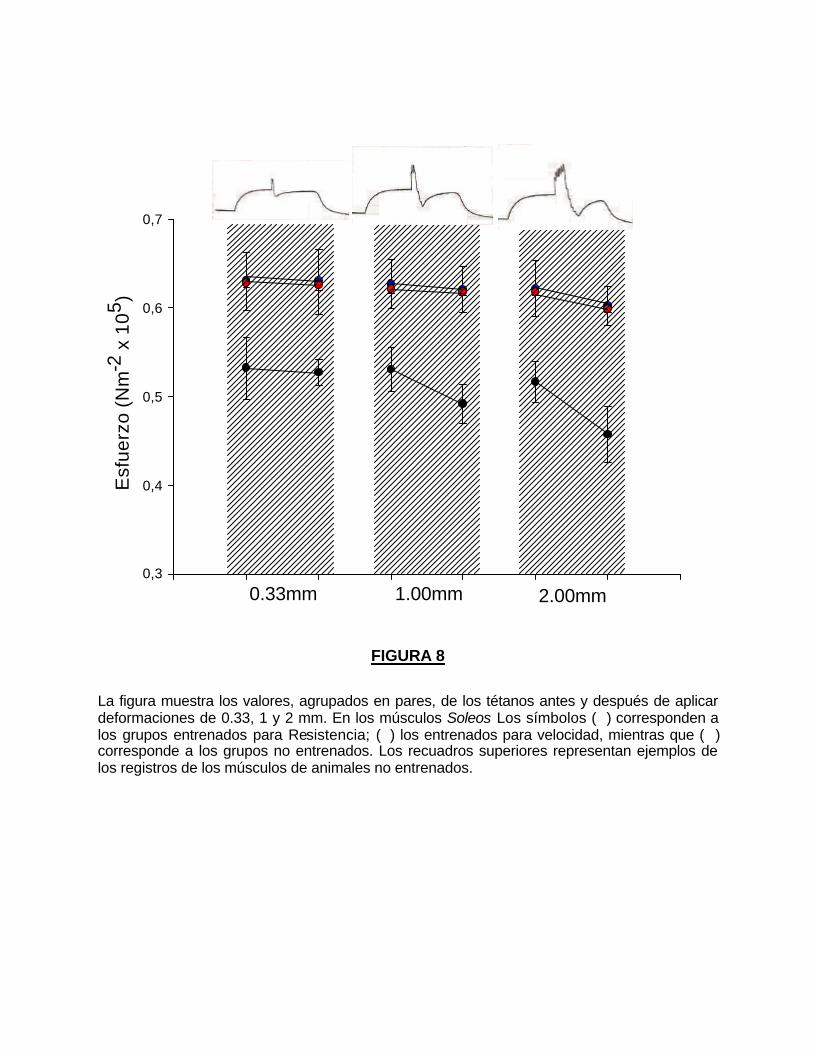
La figura 8 muestra los resultados de la aplicación de estiramientos de
0.33, 1 y 2 mm. a los músculos Soleos durante un tétanos de 6 seg. de duración
a una frecuencia de estimulación de 120 pps
Los tétanos de los músculos de los grupos entrenados, son de mayor
amplitud y alcanzan un valor de tensión más cercano al inicial, después de los
estiramientos con respecto a los grupos no entrenados.
El valor del tétanos después de la deformación de 0.33 mm, es 0.05%
menor en el grupo R y 0.33% menor en el grupo V con respecto al valor del
tétanos antes de la deformación, mientras que en el grupo C es 0.88% menor.
Al aplicar estiramientos de 1 y 2 mm sobre el tétano las diferencias en la
amplitud antes y después de la deformación fueron significativamente mas
notables en los animales no entrenados (en deformaciones de 1 mm 7.27%
menor y en 2 mm 11.45% menor ) con relación a los valores de los grupos
entrenados (R, 0.98% y 3.11%; V, 0.56% y 2.98% respectivamente).
Segundo protocolo:
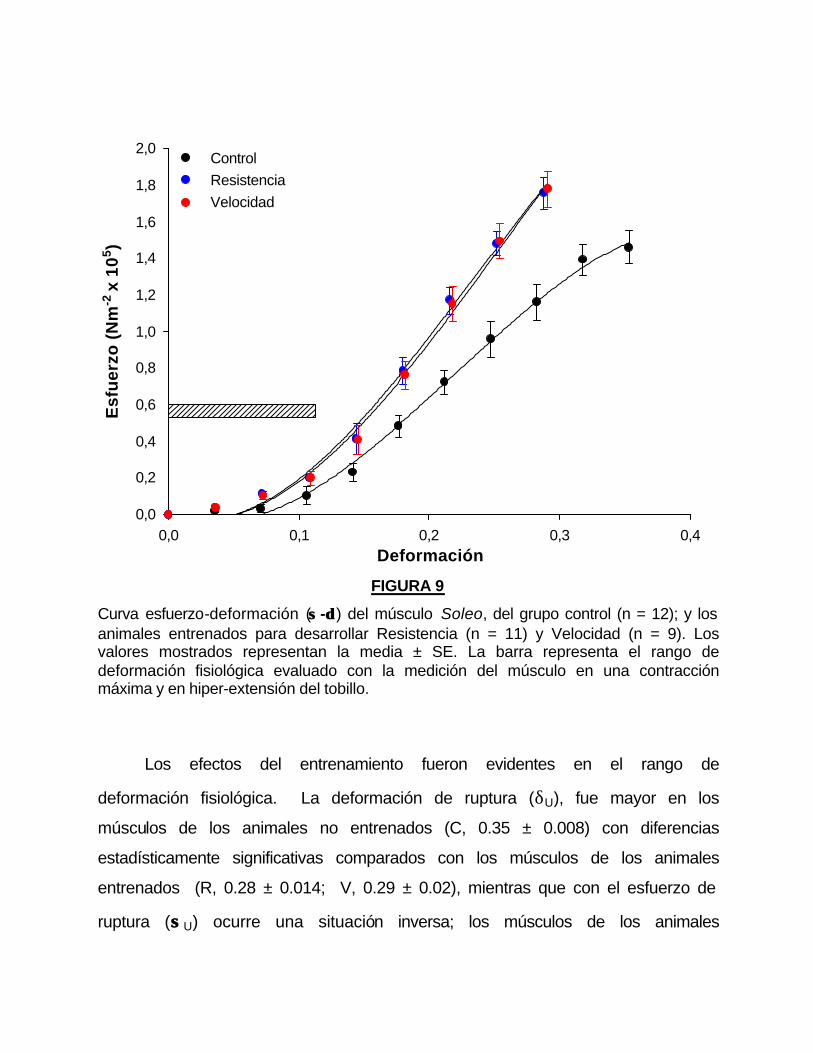
Las curvas esfuerzo-deformación (σσ -δδ ) de músculos Soleo (figura 9),
mostraron un comportamiento similar en los animales entrenados y no
entrenados.
Los parámetros de ajuste de la ecuación exponencial (4) se muestran en
la tabla 3. Los valores de k fueron similares en los grupos entrenados con
diferencias significativas respecto a los no entrenados. El valor de δ50 fue menor
en los grupos entrenados respecto al control y la diferencia fue estadísticamente
significativa.
E
sfu
erz
o (
Nm
-2 x
10
5)
0,3
0,4
0,5
0,6
0,7
0.33mm 1.00mm 2.00mm
FIGURA 8
La figura muestra los valores, agrupados en pares, de los tétanos antes y después de aplicar deformaciones de 0.33, 1 y 2 mm. En los músculos Soleos Los símbolos (�) corresponden a los grupos entrenados para Resistencia; (�) los entrenados para velocidad, mientras que (�) corresponde a los grupos no entrenados. Los recuadros superiores representan ejemplos de los registros de los músculos de animales no entrenados.
Deformación0,0 0,1 0,2 0,3 0,4
Esf
uer
zo (
Nm
-2 x
10
5)
0,2
0,4
0,6
0,8
1,2
1,4
1,6
1,8
0,0
1,0
2,0Control
Resistencia
Velocidad
FIGURA 9
Curva esfuerzo-deformación (σσ -δδ ) del músculo Soleo, del grupo control (n = 12); y los animales entrenados para desarrollar Resistencia (n = 11) y Velocidad (n = 9). Los valores mostrados representan la media ± SE. La barra representa el rango de deformación fisiológica evaluado con la medición del músculo en una contracción máxima y en hiper-extensión del tobillo.
Los efectos del entrenamiento fueron evidentes en el rango de
deformación fisiológica. La deformación de ruptura (δU), fue mayor en los
músculos de los animales no entrenados (C, 0.35 ± 0.008) con diferencias
estadísticamente significativas comparados con los músculos de los animales
entrenados (R, 0.28 ± 0.014; V, 0.29 ± 0.02), mientras que con el esfuerzo de
ruptura (σσU) ocurre una situación inversa; los músculos de los animales

entrenados mostraron un valor mayor (R, 1.75 ± 0.1; V, 1.78 ± 0.06) con
respecto a los músculos de los animales no entrenados (C, 1.47 ± 0.14).
La Energía Máxima de Deformación (EMD), fue mayor en los grupos
entrenados (R y V), que en los no entrenados (C), con diferencias
estadísticamente significativas (tabla 4).
La figura 10 muestra las graficas construida con los valores del esfuerzo
ocasionado por las sacudidas y los tétanos provocados después de cada
deformación. Los Esfuerzos de las sacudidas y los tétanos en los grupos
entrenados fueron mayores al iniciar el protocolo y después de las primeras
deformaciones, con respecto a los músculos no entrenados, sin embargo los
valores fueron disminuyendo más notablemente en estos grupos a
deformaciones mayores. Ambos parámetros (tétanos y sacudidas) mostraron
una mayor persistencia en los animales no entrenados conforme aumenta
amplitud de las deformaciones.
Deformación0,0 0,1 0,2 0,3 0,4
Esfu
erzo
(Nm
-2 x
10
5)
0,00
0,02
0,04
0,06
0,08
0,10
0,12
Deformación0,0 0,1 0,2 0,3 0,4
Esfu
erzo
(Nm
-2 x
10
5 )
0
1
A B
FIGURA 10
La figura muestra los valores del esfuerzo calculados a partir de la amplitud de las sacudidas (A) y los tétanos (B) del músculo Soleo, provocados por la estimulación del nervio motor después de cada deformación. Antes de iniciar el protocolo y durante las primeras deformaciones los valores del esfuerzo de la sacudida y los tétanos, son mayores en los grupos entrenados para desarrollar resistencia (�) y para desarrollar velocidad (�). Sin embargo, en los grupos no entrenado (�), estos parámetros resisten mayores deformaciones.
Efectos del entrenamiento en los músculo Plantaris:
Primer protocolo:
La figura 11 muestra los resultados de la aplicación de estiramientos de
0.33, 1 y 2 mm. a los músculos Plantaris mientras se encontraba tetanizado por
la estimulación del nervio motor durante 6 seg. a una frecuencia de 200 pps.
La diferencia entre el valor del tétanos antes y después de las
deformaciones en el grupo de animales no entrenados fueron de 1.8, 3.8 y 4.5%
menores. Mientras que en los grupos entrenados fueron de 0.62, 1.1 y 2.5%
menores para el grupo R y 0.79, 1.5 y 2.5 para el grupo V.
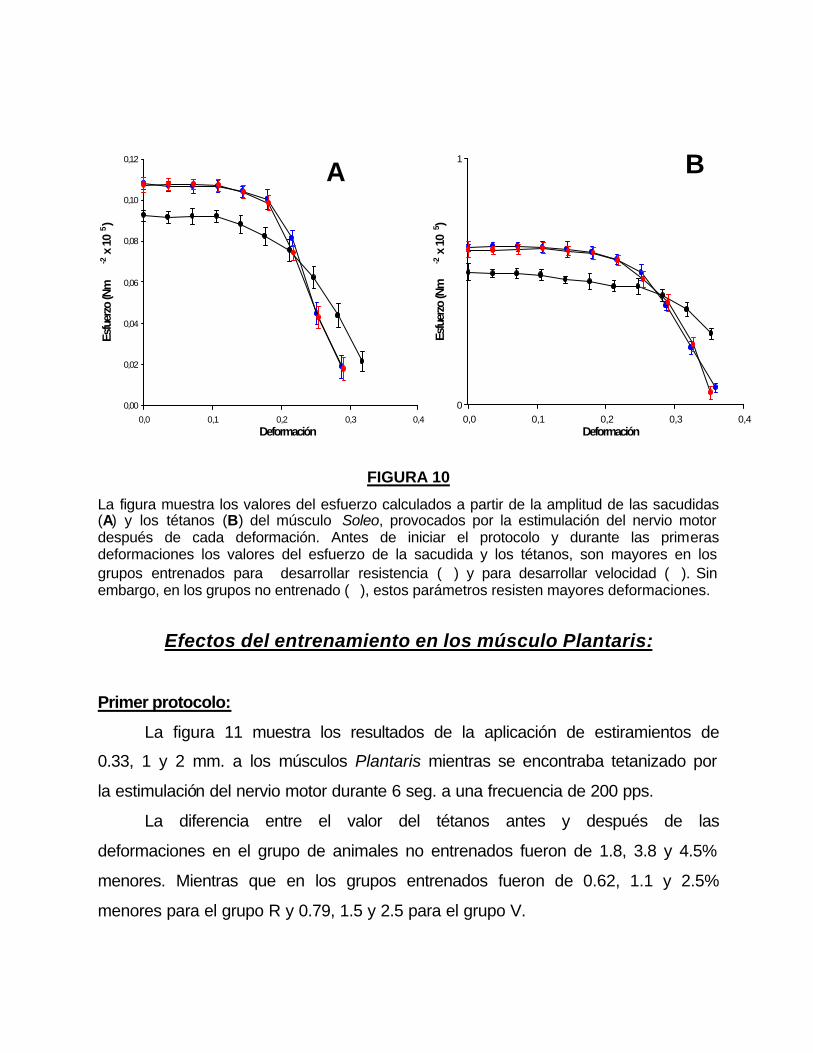
Segundo protocolo:
Las curvas esfuerzo-deformación (σσ -δδ ), obtenidas de los músculos
Plantaris de los grupos entrenados y no entrenados, mostradas en la figura 11,
presentan un patrón similar a las que construyeron con los músculos Soleos de
los diferentes grupos estudiados. Los músculos Plantaris toleran más ciclos de
deformación que los músculos Soleos.
El valor de σmax obtenido con la ecuación 4 fue significativamente mayor
en los grupos entrenados que en los controles (tabla 3).
Esf
uerz
o (N
m-2
x 1
05 )
0,70
0,75
0,80
0,85
0,90
0,95
1,00
0.33mm 1.00mm 2.00mm
FIGURA 10
Valores de los tétanos antes y después de la aplicación de ciclos de deformación de 0.33, 1 y 2 mm en los músculos Plantaris de animales entrenados para Resistencia (�), para Velocidad (�) y los animales no entrenados (�). Los recuadros superiores muestran un ejemplo de los registros obtenidos en músculos de animales del grupo de Resistencia.
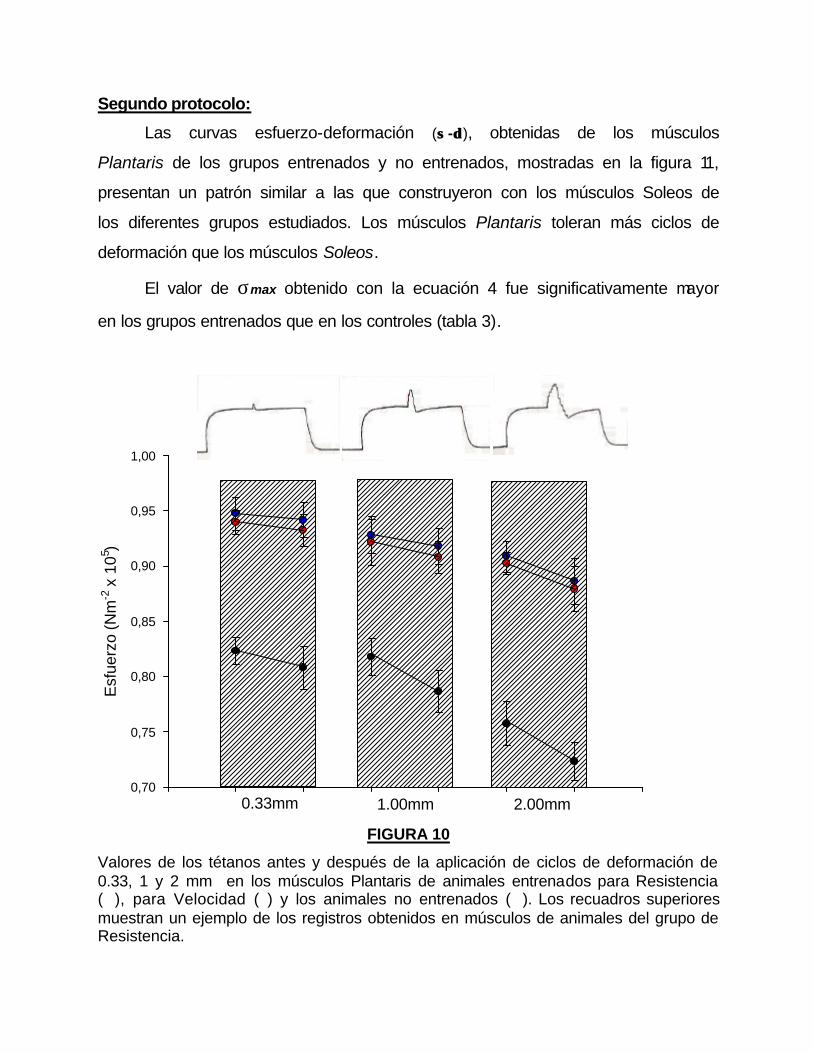
Los valores de k no muestran diferencias significativas entre los músculos
de los animales entrenados y los no entrenados, mientras que δδ 50 es mayor en el
grupo no entrenado.
La δU de los músculos del grupo Control fue de 0.37±0.07, mientras que
para los grupos de Resistencia y Velocidad, fueron de 0.35±0.001 y 0.34±0.01
respectivamente, estos valores no muestran diferencias significativas.
Los valores de σU para los músculos de los animales entrenados (R,
2.29±0.02; V, 2.21±0.01) fueron significativamente mayores con respecto a los
no entrenados (C, 1.78±0.02).
Deformación
0,0 0,1 0,2 0,3 0,4
Esf
uer
zo (
Nm
-2 x
105
)
0
1
2
3Control
ResistenciaVelocidad
FIGURA 11
Curva esfuerzo-deformación (σσ -δδ ) del músculo Plantaris, del grupo control (n = 12); y de los animales entrenados para desarrollar Resistencia (n = 11) y Velocidad (n = 9). Las líneas que unen los valores fueron obtenidos con la ecuación 4. Los valores mostrados representan la media ± SE.
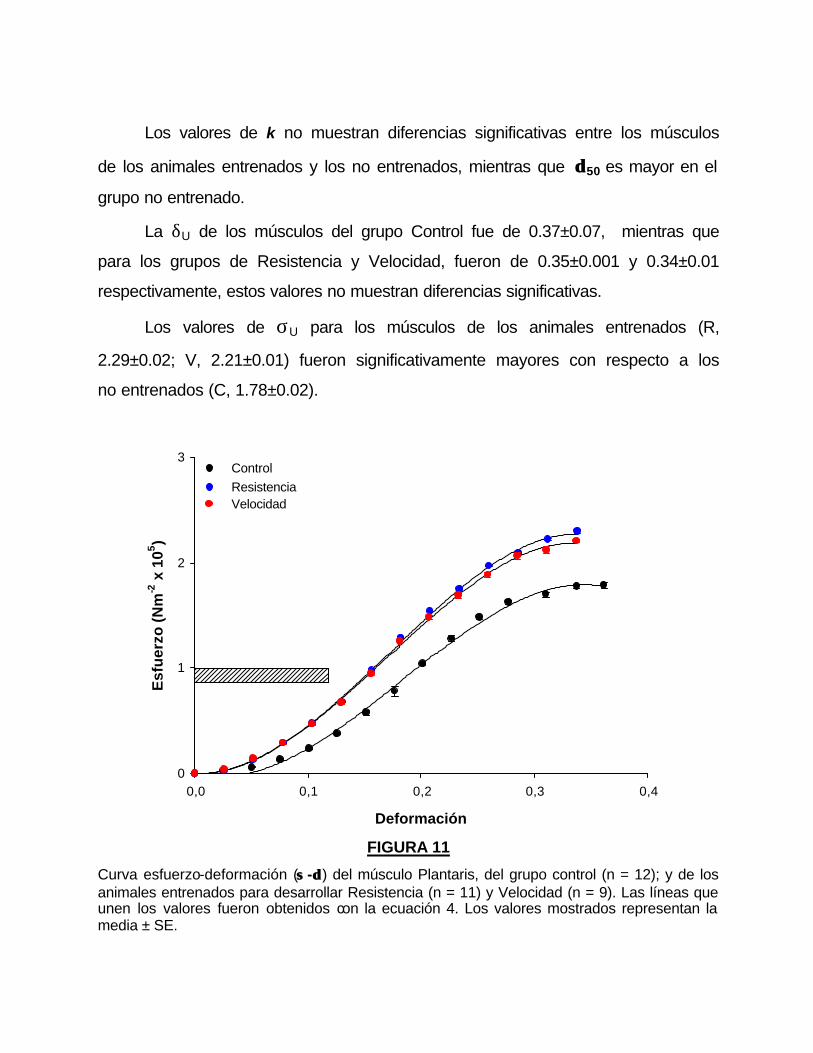
De igual forma la EMD mostró diferencias significativas entre los grupos
entrenados para resistencia y velocidad comparados con los animales del grupo
no entrenado (tabla 4).
La figura 12 muestra los valores del esfuerzo de las sacudidas (A) y los
tétanos (B) provocados después de cada deformación. Los músculos Plantaris
de los grupos entrenados (R y V) mostraron un valores de esfuerzo mayores en
los tétanos y las sacudidas y toleraron, desde el inicio del protocolo hasta el
rompimiento del músculo, mas ciclos de deformación que los músculos del
grupo control.
Deformación0,0 0,1 0,2 0,3 0,4
Esf
uerz
o (N
m-2
x 1
05 )
0,00
0,05
0,10
0,15
0,20
0,25
A
Deformación0,0 0,1 0,2 0,3 0,4
Esf
uerz
o (N
m-2
x 1
05 )
0
1 B
FIGURA 12 La figura muestra los valores del esfuerzo calculados a partir de la amplitud de las sacudidas (A) y los tétanos (B) provocados por la estimulación del nervio motor después de cada deformación en el músculo Plantaris. Estos parámetros persisten a deformaciones mayores, en los músculos de animales entrenados para Resistencia (�) y Velocidad (�) al compararlos con el grupo Control (�).
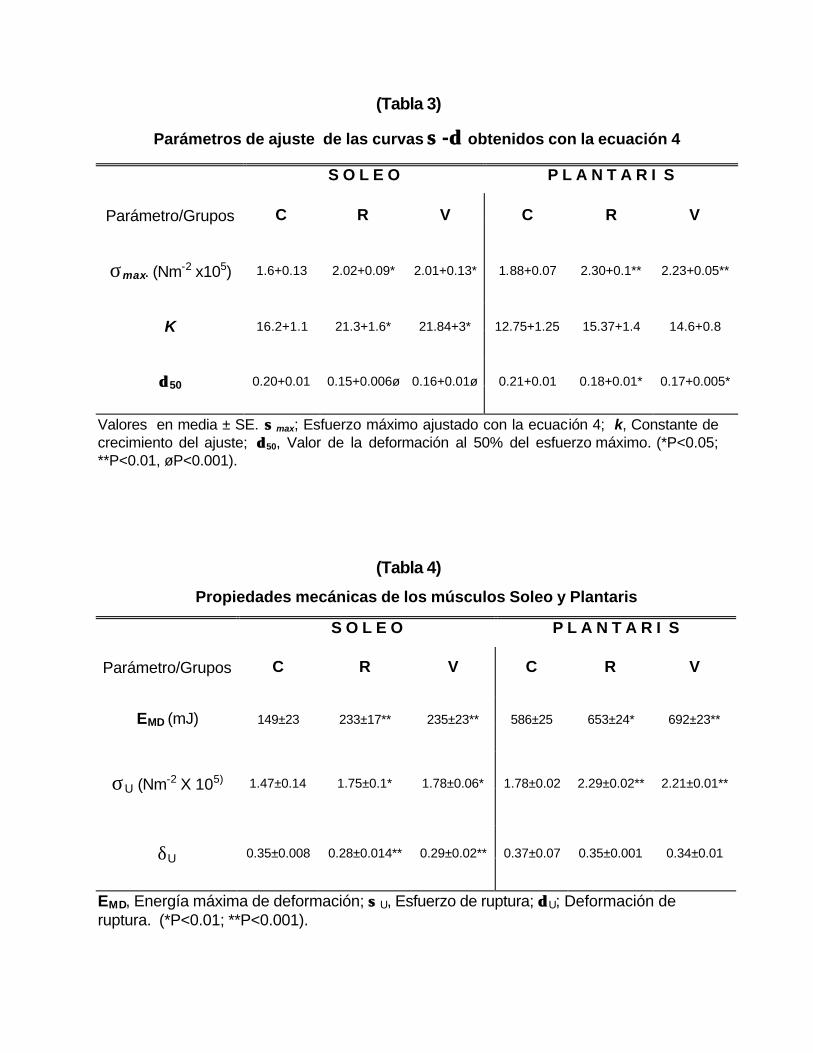
(Tabla 3)
Parámetros de ajuste de las curvas σσ -δδ obtenidos con la ecuación 4
S O L E O P L A N T A R I S
Parámetro/Grupos C R V C R V
σmax. (Nm-2 x105) 1.6+0.13 2.02+0.09* 2.01+0.13* 1.88+0.07 2.30+0.1** 2.23+0.05**
K 16.2+1.1 21.3+1.6* 21.84+3* 12.75+1.25 15.37+1.4 14.6+0.8
δδ 50 0.20+0.01 0.15+0.006ø 0.16+0.01ø 0.21+0.01 0.18+0.01* 0.17+0.005*
Valores en media ± SE. σσ max; Esfuerzo máximo ajustado con la ecuación 4; k, Constante de crecimiento del ajuste; δδ 50, Valor de la deformación al 50% del esfuerzo máximo. (*P<0.05; **P<0.01, øP<0.001).
(Tabla 4)
Propiedades mecánicas de los músculos Soleo y Plantaris
S O L E O P L A N T A R I S
Parámetro/Grupos C R V C R V
EMD (mJ) 149±23 233±17** 235±23** 586±25 653±24* 692±23**
σU (Nm-2 X 105) 1.47±0.14 1.75±0.1* 1.78±0.06* 1.78±0.02 2.29±0.02** 2.21±0.01**
δU 0.35±0.008 0.28±0.014** 0.29±0.02** 0.37±0.07 0.35±0.001 0.34±0.01
EMD, Energía máxima de deformación; σσ U, Esfuerzo de ruptura; δδ U; Deformación de ruptura. (*P<0.01; **P<0.001).

D I S C U S I O N.

Para probar los efectos del ejercicio, en las propiedades mecánicas
pasivas, se escogieron dos tipos básicos de entrenamiento: el de resistencia
(Power y col. 1992, Deschenes y col. 1995) y el de velocidad (Saubert y col.
1973, Guezennec y col. 1990).
Los Resultados obtenidos en este estudio con Ratas Wistar, confirman
que los músculos lentos (Soleo) y rápidos (Plantaris) exhiben un
comportamiento visco-elástico no lineal (Nubar1962, Halpern y Moss 1976,
Kovanen y col. 1984, Magid y Law 1985). Las diferencias en los valores de
ajuste resultantes de la ecuación 4 entre los músculos estudiados, podrían estar
relacionados con la cantidad y organización de los componentes elásticos de los
mismos (Kovanen y col. 1984, Takekura y Yoshioka 1990, Bigart y col 1991).
Los entrenamientos de resistencia y de velocidad utilizados en este
estudio incrementan la EMD y el σU en ambos músculos pero disminuyen la δU.
Los estudios de Kovanen y cols (1984), fueron realizados in vitro, a la
temperatura ambiente. Otros estudios sugieren que la temperatura podría
afectar las propiedades elásticas de las fibras musculares (Maruyama y col.,
1977, Mutungi y Ranatunga, 1996a). En nuestros experimentos la temperatura y
el aporte sanguíneo, se mantuvieron lo más cercano a las condiciones
fisiológicas. Esto podría explicar las diferencias entre nuestros datos y los
reportados previamente (Kovanen y col., 1984).
Nuestros resultados muestran que el Plantaris, un músculo rápido,
desarrolla mayor esfuerzo a una deformación dada comparado con el Soleo,
considerado como un músculo lento y esto podria deberse a las diferencias en la
composición de Titina entre ambos músculos.
El parámetro σU esta relacionado con el punto de ruptura del músculo y
es análogo al esfuerzo máximo (σmax) propuesto por Kovanen y col. (1984). En
nuestros resultados es del mismo orden de magnitud (1 x 105 Nm-2) al reportado
por Wang y col. (1993) en el músculo Psoas de conejo. Los cambios en σσU, δU,

δ50, k, indican que los músculos de los animales entrenados aumenta la
capacidad para almacenar energía elástica, comparado con los animales no
entrenados.
Los valores de σσmax, δU y k tienden a ser más grandes en los músculos de
los animales entrenados para desarrollar resistencia y velocidad, que en los
músculos a de los animales no entrenados. El mayor valor de k, mostrado por
los músculos Soleos y sus valores menores de δU, y δ50, confirma que este
músculo lento tiene menos rigidez que el Plantaris.
El hallazgo de que la EMD, fuera mayor en los músculos Plantaris con
respecto a los soleos, demuestra que este músculo rápido, es mas eficiente para
recuperar la energía después de una contracción activa. Adicionalmente los
resultados de este estudio demostraron que el entrenamiento mejora la EMD en
ambos músculos.
Trabajos previos han propuesto que el entrenamiento incrementa la
eficiencia de la contracción y podría proteger a los músculos contra lesiones. La
mayor amplitud de los tétanos y las sacudidas desarrolladas por los músculos
Soleo y Plantaris de animales entrenados para desarrollar resistencia ( R ) y
velocidad ( V ) mostrados en las figuras 6 y 9 y los resultados de las curvas
esfuerzo-deformación (σσ -δδ ) de las figuras 5 y 8, demuestran una mayor eficiencia
de los músculos entrenados con respecto a los controles.
En un músculo relajado, la tensión pasiva se debe, probablemente a la
activación espontánea de algunos puentes cruzados y a la participación de los
filamentos conectores compuestos principalmente por moléculas de Titina.
(Magid y Law, 1985).
En el estado activo alcanzado en el tétanos completo, se estima que,
alrededor del 70% de los puentes cruzados están activos generando tensión, la
cual se suma a que generan los filamentos de Titina y su conexión con los
discos Z.

La recuperación de la tensión del tétanos después de la aplicación de
estiramientos mostrada en las figuras 6 y 9 es mas eficiente en los animales
entrenados, los cual podría relacionarse con una mayor capacidad de activación
de los puentes cruzados.
En el presente trabajo, medimos el pico de tensión en los ciclos de
deformación, que depende de los componentes musculares visco-elásticos. La
velocidad de los ciclos de deformación fue constante (0.33mm s-1) en todos los
experimentos realizados. El coeficiente de viscosidad se calcula dividiendo el
esfuerzo entre la velocidad de estiramiento y en nuestro caso fue del orden de
108 Nsm-3, similar a lo reportado previamente en fibras musculares aisladas
(Bagni y col., 1992; Mutungi y Ranatunga, 1996b).
A pesar que la literatura reporta que el ejercicio produce hipertrofia en las
fibras musculares, no se encontraron diferencias en los valores morfométricos
en los músculos Soleos de los animales entrenados y no entrenados, igual que
ocurre con los músculos Plantaris (tabla 2). Por lo cual, podremos asumir que
los dos tipos de entrenamiento utilizados en el estudio, tienen un efecto sobre
las propiedades mecánicas de los músculo rápidos y lentos de rata, al efectuar
cambios en las cantidades relativas de los componentes musculares y/o por un
re-arreglo estructural de las proteínas miofibrilares y no a cambios por hipertrofia
o atrofia.
Parte de los nuestros resultados experimentales fueron publicados en las
revistas: General. Physiology & Biophys. Journal (1999) y Acta Physiological
Scandinavian (2001). Ambos trabajos se anexan en el apendice

C O N C L U S I O N E S

En los músculos rápidos y lentos de los animales entrenados la recuperación de
la tensión tetánica después de ser sometidos a una deformación es mayor que en
los animales No entrenados.
En el rango fisiológico, la tensión de tétanos y la sacudida, es significativamente
mayor en los animales entrenados.
La persistencia del tétanos y la sacudida, después de la deformación, es mayor en
los Soleos de los animales No entrenados.
La persistencia del tétanos y la sacudida, fue mayor en los Plantaris de los
animales entrenados.
Los entrenamientos de resistencia y de velocidad utilizados en este estudio
incrementan la EMD y σσ U en ambos músculos, lo cual puede interpretarse como
indicativo de que los músculos entrenados desarrollan una mayor rigidez.
Los cambios en σσ U, δδ U, δ50 y ê, indican que los músculos de los animales
entrenados muestran una mayor capacidad para almacenar energía elástica
comparados con los músculos de los animales No entrenados.
El hallazgo de que la EMD fue mayor en los Plantaris, en comparación con el
Soleo, demuestra que es mas eficiente para recuperar la energía después de una
contracción activa, adicionalmente, los resultados de este estudio demostraron que
los entrenamientos utilizados mejoraron la EMD en ambos músculos.
Los músculos lentos y rápidos exhiben un comportamiento visco-elástico no
lineal que puede explicarse con los modelos clásicos usados para describir el
comportamiento de los polímeros (Muñiz y col, 1999).
Finalmente como no se observaron diferencia en los parámetros morfométricos
de los músculos de animales entrenados y no entrenados, podemos asumir que el
entrenamiento modifica la plasticidad de los elementos responsables de las
propiedades mecánicas pasivas de los músculos por cambios en la cantidad relativa de
los componentes musculares y/o una reorganización estructural de las proteínas
miofibrilares (Muñiz y col, 2001).


















