I UNIDAD INMUNOLOGÍA GENERAL,...
28
UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA FACULTAD : CIENCIAS DE LA SALUD ESCUELA : FARMACIA Y BIOQUÍMICA ASIGNATURA : INMUNOLOGÍA DOCENTES : Mblgo. JOSÉ LUIS GUTIERREZ APONTE (TITULAR) Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO (TUTOR) I UNIDAD INMUNOLOGÍA GENERAL, INMUNOQUÍMICA E INMUNOBIOLOGÍA TEMA 03: MEDIADORES SOLUBLES DE LA INMUNIDAD CONTENIDOS: MEDIADORES SOLUBLES DE LA INMUNIDAD: CARACTERÍSTICAS GENERALES. LAS CITOCINAS: CARACTERÍSTICAS Y PROPIEDADES. ACTIVIDAD BIOLÓGICA DE LAS CITOCINAS EN LA INMUNIDAD INNATA Y ADAPTATIVA, RECEPTORES DE LAS CITOQUINAS Y SEÑALIZACIÓN. PROTEÍNAS DEL SISTEMA DEL COMPLEMENTO: CARACTERÍSTICAS GENERALES Y FUNCIÓN. VÍAS DE ACTIVACIÓN DEL SISTEMA DEL COMPLEMENTO (VÍA ALTERNATIVA Y VÍA CLÁSICA): MECANISMO DE ACCIÓN. LOS ANTICUERPOS: CARACTERÍSTICAS GENERALES, ESTRUCTURA Y RECONOCIMIENTO DEL ANTÍGENO. CITOQUINAS Las citoquinas o interleuquinas son proteínas de bajo peso molecular esenciales para comunicación intercelular, son producidas por varios tipos celulares, principalmente por el Sistema Inmune (SI). Estos mediadores solubles controlan muchas funciones fisiológicas críticas tales como: diferenciación y maduración celular, inflamación y respuesta inmune local y sistémica, reparación tisular, hematopoyesis, apoptosis y muchos otros procesos biológicos1, 3,4. Este escrito describe la biología básica de las citoquinas y su papel central en la regulación de la respuesta inmune en salud y enfermedad. Mblgo. JOSÉ LUIS GUTIERREZ APONTE 1 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO
-
Upload
truongtuong -
Category
Documents
-
view
212 -
download
0
Transcript of I UNIDAD INMUNOLOGÍA GENERAL,...

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
FACULTAD : CIENCIAS DE LA SALUDESCUELA : FARMACIA Y BIOQUÍMICAASIGNATURA : INMUNOLOGÍADOCENTES :Mblgo. JOSÉ LUIS GUTIERREZ APONTE (TITULAR)Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO (TUTOR)
I UNIDAD
INMUNOLOGÍA GENERAL, INMUNOQUÍMICA E INMUNOBIOLOGÍA
TEMA 03: MEDIADORES SOLUBLES DE LA INMUNIDAD
CONTENIDOS:
MEDIADORES SOLUBLES DE LA INMUNIDAD: CARACTERÍSTICAS
GENERALES.
LAS CITOCINAS: CARACTERÍSTICAS Y PROPIEDADES.
ACTIVIDAD BIOLÓGICA DE LAS CITOCINAS EN LA INMUNIDAD INNATA Y
ADAPTATIVA, RECEPTORES DE LAS CITOQUINAS Y SEÑALIZACIÓN.
PROTEÍNAS DEL SISTEMA DEL COMPLEMENTO: CARACTERÍSTICAS
GENERALES Y FUNCIÓN.
VÍAS DE ACTIVACIÓN DEL SISTEMA DEL COMPLEMENTO (VÍA
ALTERNATIVA Y VÍA CLÁSICA): MECANISMO DE ACCIÓN.
LOS ANTICUERPOS: CARACTERÍSTICAS GENERALES, ESTRUCTURA Y
RECONOCIMIENTO DEL ANTÍGENO.
CITOQUINAS
Las citoquinas o interleuquinas son proteínas de bajo peso molecular esenciales para
comunicación intercelular, son producidas por varios tipos celulares, principalmente por el
Sistema Inmune (SI). Estos mediadores solubles controlan muchas funciones fisiológicas
críticas tales como: diferenciación y maduración celular, inflamación y respuesta inmune
local y sistémica, reparación tisular, hematopoyesis, apoptosis y muchos otros procesos
biológicos1, 3,4. Este escrito describe la biología básica de las citoquinas y su papel central
en la regulación de la respuesta inmune en salud y enfermedad.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 1 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
Las citoquinas son proteínas que regulan la función de las células que las producen u
otros tipos celulares. Son los agentes responsables de la comunicación intercelular, inducen
la activación de receptores específicos de membrana, funciones de proliferación y
diferenciación celular, quimiotaxis, crecimiento y modulación de la secreción de
inmunoglobulinas. Son producidas, fundamentalmente, por los linfocitos y los macrófagos
activados, aunque también pueden ser producidas por leucocitos polinucleares, células
endoteliales, epiteliales y del tejido conjuntivo. Según la célula que las produzca se
denominan linfocinas (linfocito), monocinas (monocitos) o interleucinas (células
hematopoyéticas). Su acción fundamental es en la regulación del mecanismo de la
inflamación. Hay citocinas proinflamatorias y otras antiinflamatorias.
PROPIEDADES GENERALES DE LAS CITOCINAS
Las citocinas son polipéptidos producidas en respuesta a microorganismos y otros
antígenos que median y regulan las reacciones inmunitarias e inflamatorias:
Las citoquinas son un grupo de proteínas secretadas de bajo peso molecular (por lo
general menos de 30 kDa), producidas durante las respuestas inmunes natural y específica.
Se unen a receptores específicos de la membrana de las células donde van a ejercer su
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 2 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
función, iniciando una cascada de transducción intracelular de señal que altera el patrón de
expresión génica, de modo que esas células diana producen una determinada respuesta
biológica.
Las citoquinas son producidas por múltiples tipos celulares, principalmente del
sistema inmune. Dentro del sistema inmune natural, los macrófagos son de las
células más productoras de citoquinas, mientras que en el sistema específico lo son
las células T colaboradoras.
La producción de las citoquinas suele ser breve (transitoria), limitada al lapso de
tiempo que dura el estímulo (es decir, el agente extraño). En muchos casos ello se
debe a que los correspondientes ARNm tienen una corta vida media, que a su vez
depende de que las zonas 3’ no traducibles son ricas en A y U.
Considerando las diversas citoquinas, éstas pueden exhibir una o varias de las
siguientes cualidades:
Pleiotropía (múltiples efectos al actuar sobre diferentes células).
Redundancia (varias citoquinas pueden ejercer el mismo efecto).
Sinergismo (dos o más citoquinas producen un efecto que se potencia
mutuamente).
Por ejemplo, la acción conjunta de IL4 e IL5 induce en células B el cambio
de clase para que produzcan IgE.
antagonismo (inhibición o bloqueo mutuo de sus efectos).
Por ejemplo, el IFNg bloquea el cambio de clase promovido por IL4.
Las citoquinas ejercen su acción al unirse a receptores específicos para cada
citoquina en la superficie de la célula en la que ejercen el efecto. La afinidad de cada
receptor hacia su citoquina correspondiente suele ser bastante alta, del orden de lo
femtomolar (1015 M) a lo picomolar (1012 M).
Utilizando la analogía de lo que ocurre con las hormonas del sistema endocrino, las
acción de las citoquinas se puede clasificar en:
de tipo autocrino
de tipo paracrino
(en pocas ocasiones) de tipo endocrino.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 3 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
Las citoquinas "controlan" el sistema inmune de varias maneras, que podemos
agrupar de la siguiente manera:
Regulando (activando o inhibiendo) la activación, proliferación y diferenciación de
varios tipos de células;
Regulando la secreción de anticuerpos y de otras citoquinas.
ESTRUCTURA Y FUNCIÓN:
Las citoquinas son proteínas o glicoproteínas de menos de 30 kDa. Muchas de ellas
pertenecen a la llamada familia de las hematopoyéticas, y tienen estructuras terciarias
parecidas: una configuración a base de un conjunto de cuatro hélices a , con poca estructura
en lámina b.
Generalmente actúan como mensajeros intercelulares que suelen intervenir en la
maduración y amplificación de la respuesta inmune, provocando múltiples actividades
biológicas una vez que se unen a los receptores específicos de las células diana adecuadas.
Aunque existen muchos tipos de células productoras citoquinas (ya hemos ido viendo unas
cuantas en los temas anteriores), los más importantes son los linfocitos TH y los
macrófagos, ya que sus citoquinas son esenciales para que se produzca la respuesta
inmune una vez que se activan las células T y B por el contacto con las correspondientes
células presentadoras de antígeno.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 4 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
Principales tipos de respuesta mediatizados por la acción de las citoquinas:
1. activación de los mecanismos de inmunidad natural:
a. Activación de los macrófagos y otros fagocitos
b. Activación de las células NK
c. Activación de los eosinófilos
d. Inducción de las proteínas de fase aguda en el hígado (ver tema sobre el
complemento).
2. Activación y proliferación de células B, hasta su diferenciación a células plasmáticas
secretoras de anticuerpos.
3. Intervención en la respuesta celular específica.
4. Intervención en la reacción de inflamación, tanto aguda como crónica.
5. Control de los procesos hematopoyéticos de la médula ósea.
6. Inducción de la curación de las heridas.
Hay una aparente paradoja de las citoquinas que debemos explicar: ¿Por qué las
citoquinas, que son inespecíficas respecto del antígeno, pueden ejercer acciones de modo
específico? Veamos varios mecanismos:
Regulación muy fina de los receptores de cada citoquina: los receptores celulares
indispensables para que una citoquina ejerza su papel sólo se expresan en tipos
celulares concretos una vez que éstos han interaccionado con el antígeno
(pensemos por ejemplo en los linfocitos cebados con antígeno).
Requerimientos de contactos estrechos célula a célula: la citoquina sólo alcanza
concentraciones adecuadas para actuar en el estrecho espacio que queda entre dos
células interactuantes; recordar por ejemplo las "bolsas" que se forman en el
conjugado TH:B, donde se alcanzan mejor esos niveles de citoquinas.
Corta vida media de las citoquinas en sangre y fluidos, lo que asegura que sólo van a
actuar en un estrecho margen de tiempo, en las cercanías de la zona donde se
produjeron.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 5 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
CATEGORIAS FUNCIONALES:
Citoquinas en Inflamación:
Las principales citoquinas que actúan en la respuesta inespécifica o inflamación son:
Interleukina 1 (IL1), Factor de Necrosis Tumoral Alfa (TNF ), Interleukina 8 (IL8 yα
chemoquinas), Interleukina 12 (IL12), Interleukina 16 (IL16) e Interferones10,11,12, todas
ellas son proinflamatorias. IL6 e IL12, además, actúan en la inmunidad especifica, IL6 es
un factor autocrino de linfocitos B7 mientras que IL12 estimula la Inmunidad celular
citotóxica.
Citoquinas en Inmunidad Celular
Durante la inflamación los macrófagos y otras células presentan los antígenos a los
linfocitos T ayudadores (Th o CD4+), los cuales son muy importantes (si no los principales)
moduladores intrínsecos del sistema inmune regulando las dos vías principales de defensa
específica: Celular Vs Humoral, a través de la secreción de citoquinas.
En este instante es relevante mencionar que el pérfil o “set” de citoquinas secretadas
por los linfocitos Th polariza la respuesta inmune hacia una predominantemente citotóxica o
celular o hacia el otro extremo predominantemente humoral, esas respuestas son
antagónicas o excluyentes entre sí, creando una especie de regulación cruzada muy
articular; porque las citoquinas que favorecen la inmunidad humoral inhiben las acciones de
las citoquinas que ayudan a la inmunidad celular y viceversa. Los linfocitos Th que inducen
respuesta inmune celular se denominan Th1 mientras que aquellos que favorecen las
respuestas humorales son Th2.
Dos son las principales citoquinas de Inmunidad Celular o Th1: Interferón gamma
(IFN ) o tipo 2, llamado también interferón Inmune porque solo es producido por célulasγ
inmunológicas activadas; la otra citoquina es Interleukina 2 o Factor de Crecimiento de
Células T (IL2 o TCGF). IFN es el principal activador de macrófagos y células citotóxicasγ
T y NK. Interesantemente IFN tiene acción en la Inmunidad Humoral, induciendo laγ
producción de IgG211,12. IL2 descubierta en 1977 por Robert Gallo (codescubridor del
VIH) es el factor autocrino de crecimiento de las células T, esencial para mantener viables
los cultivos de linfocitos T, también genera células citotóxicas especialmente NK y
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 6 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
macrófagos antitumorales).
Citoquinas de Inmunidad Humoral :
La Inmunidad humoral se caracteriza por la secreción de anticuerpos por los
linfocitos B o células plasmáticas, las cuales son moduladas por las siguientes citoquinas:
Interleuquina 4 o factor estimulante de células B (IL4 o BCSF), Interleuquina 5 (IL5)23,
Interleukina 6 (IL6), Interleukina 10 (IL10) e interleukina 13 (IL13). Estas linfoquinas son
secretadas por linfocitos del tipo Th2, linfocitos B, mastocitos, eosinófilos y algunas por
macrófagos (IL6, IL13).
IL4 es la citoquina mejor caracterizada en la regulación de la respuesta inmune
humoral; en pocas cantidades induce secreción de las subclases de Inmunoglobulina G:
IgG1, IgG3 e IgG4; mientras que en excesiva cantidad induce la producción de IgE. Esta
citoquina antagoniza las acciones biológicas de IFN , tales como la activación de Mf y elγ
desarrollo de células citotóxicas21, 25 así inhibe las células Th1.
IL5 es la citoquina con rango de acción más reducido al inducir la generación de
Inmunoglobulina A (IgA) y Eosinófilos. IL6 es la mejor estudiada de una familia de
citoquinas hematopoiéticas (los otros miembros son de muy reciente descubrimiento:
Interleukina, Factor inhibitorio de leucemias (LIF), Oncostatin M (OSM), Factor neurotrófico
ciliar (CNTF)31 y cardiotrofina. IL6 es una citoquina pleotrófica, en inflamación es la más
potente inductora de hepatocitos para la síntesis de reactantes de fase aguda; potencia los
efectos de IL1 y TNF aunque no posee la toxicidad de estas y en la inmunidad humoral
tiene efectos similares a IL11 promoviendo la diferenciación, proliferación de linfocitos B y la
síntesis de inmunoglobulinas. Adicionalmente, IL6 es el factor autocrino de crecimiento de
células tumorales B malignas y benignas (Mieloma múltiple, Mixoma cardiaco), también esta
elevada en Lupus Eritematoso Sistémico (autoinmunidad).
Citoquinas en Homeóstasis:
Las citoquinas actúan en grupos formando secuencias, o cadenas interactivas en
procesos tisulares no inmunológicos como, hemopoiésis, remodelación ósea y en sitios
diversos tales como el desarrollo embrionario fetal37. Las células progenitoras
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 7 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
hematológicas dependen esencialmente del microambiente de la médula ósea finamente
regulado por citoquinas secretadas principalmente por (células estromales) para controlar su
diferenciación y proliferación hacia células sanguíneas maduras, aunque es difícil
clasificarlas por su sobre posición funcional, se distinguen tres categorías de citoquinas: A
Aquellas que actúan en las células primordiales multipotentes (multilineales como
Interleukina 3 (IL3) y el factor estimulante de monocitos y granulocitos (GMCSF). B Las
que actúan en líneas celulares ya definidas o comprometidas hacia diferenciación
(Restringidas o especificas de líneas definidas tales como Eritropoyetina (EPO), eritrocitos,
TPO (megacariocitos) GCSF (granulocitos), MCSF (monocitos), IL2 (linfocitos), IL5
(origina eosinófilos)35. 3 Las que tienen poco efecto por si solas pero que inhiben o hacen
sinergia funcional de otras citoquinas (¨stem cell factor¨(SCF), IL6, IL1).
RECEPTORES:
Hay diversos tipos de receptores de membrana para citoquinas, pero se pueden
agrupar en cinco familias:
Familia de receptores de citoquinas de la superfamilia de las inmunoglobulinas, que
poseen varios dominios extracelulares de tipo Ig. Como ejemplo, el receptor
específico para la IL1.
Familia de clase I de receptores de citoquinas (=familia de receptores de
hematopoyéticas).
Familia de clase II de receptores de citoquinas (=familia de receptores de
interferones).
Ejemplos de ligandos son los interferones no inmunes (IFNa y b) y el IFNg.
Familia de receptores de TNF: sus miembros se caracterizan por un dominio
extracelular rico en cisteínas.
Ejemplos de ligandos: TNFa, TNFb, CD40.
Familia de receptores de quimioquinas: son proteínas integrales de membrana, con 7
hélices a inmersas en la bicapa lipídica. Interaccionan, por el lado que da al
citoplasma con proteínas de señalización triméricas que unen GTP. Ejemplos de
quimioquinas que se unen a miembros de esta familia: IL8.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 8 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
La mayor parte de los receptores de citoquinas del sistema inmune pertenecen a la
familia de clase I (de receptores de hematopoyéticas). Todos sus miembros tienen en
común poseer una proteína anclada a membrana, con un dominio extracelular en el que hay
al menos un motivo característico llamado CCCC (cuatro cisteínas cercanas en posiciones
equivalentes) y el llamado motivo WSXWS (TrpSerXTrpSer). (Adicionalmente, algunos
miembros poseen dominios de tipo Ig y/o dominios de tipo fibronectina). Tras su porción
transmembrana se encuentra una larga cola citoplásmica con ciertas tirosinas susceptibles
de fosforilación.
La mayor parte de los receptores de clase I poseen dos proteínas de membrana:
Cadena a, que es la subunidad específica de la citoquina, sin capacidad de enviar
señales al citoplasma;
Cadena b, una subunidad transductora de señal, que a menudo no es específica de
citoquina, sino que es compartida por receptores de otras citoquinas. Esta cadena b
es del tipo que hemos descrito más arriba (motivos CCCC y WSXWS).
La subunidad transductora de señal se necesita para formar el receptor de alta
afinidad, y para transducir la señal al interior. Ello se logra porque tras la unión, se fosforilan
ciertas tirosinas de la larga cola citoplásmica de la cadena transductora de señal.
En los esquemas comparativos de distintos receptores se puede apreciar que
algunos de ellos poseen el mismo tipo de subunidad b. Este hecho permite explicar dos
cualidades a las que ya hemos aludido: la redundancia y el antagonismo.
Por ejemplo: consideremos los receptores de IL3, IL5 y GMCSF, que como se ve,
comparten el mismo tipo (llamado KH97) de cadena b.
Redundancia: por separado, las tres citoquinas citadas, al tener sendos receptores que
tienen el mismo tipo de cadena b , provocan los mismos efectos biológicos: proliferación de
eosinófilos y desgranulación de basófilos.
Antagonismo: las tres citoquinas compiten entre sí por la unión de un número limitado de
cadenas b con las a específicas de cada receptor.
La subfamilia de receptores a la que pertenece el receptor de IL2 (el llamado IL2R) consta
de tres subunidades:
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 9 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
✔ Cadena a específica de cada citoquina;
✔ Dos subunidades (cadena b y cadena común gC) transductoras de señal.
Esta versión trimérica de IL2R es la que une IL2 con mayor afinidad, pero en
realidad, el receptor para IL2 no siempre aparece en esta forma, sino que existe una forma
monomérica (IL2Ra) de baja afinidad, sin cadenas transductoras, y por lo tanto incapaz de
reenviar la señal, y una forma de afinidad intermedia (IL2Rbg).
Citocina AcciónLugar de síntesis
InductorAcciones más importantes
IL1 ProinflamatoriaCélulas mononucleares
Microbiana o activación cascada inflamatoria (CI)
Pirógeno
IL2 Antiinflamatorialinfocitos Th colaboradores
Sustancias microbianas o activación de CI
factores de crecimiento de células T induciendo a la proliferación de todos los tipos de subpoblaciones linfocitarias.Estimula síntesis de interferón liberación de IL1, TNF y betaα
IL3
IL4 AntiinflamatorioLinfocitos Th, mastocitos y basófilos
DesconocidoBloquea síntesis de citocinas, inhibe la síntesis de NO
IL6 Proinflamatoriaantiinflamatoria
Monocitos, macrófagos, célula endotelial y fibroblastos
Il1 y endotoxinas
Pirógeno, síntesis de inmunoglobulinas. Activación de la síntesis de de proteínas de fase aguda
IL8 Proinflamatoria
Monocitos, macrófagos, célula endoltelias y fibroblastos
IL1,TNFalfa y endotoxinas
Factor quimiotáctico y activador de neutrófilos
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 10 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
ANTICUERPOS (ACs)
“INMUNOGLOBULINAS (Ig)”
Es una proteína, producida por las células y capaces de reaccionar con un Ag. El
organismo produce un Ac distinto para cada antígeno por lo cual la reacción es distinto para
cada antígenoanticuerpo es específica.
Hay cinco clases de inmunoglobulinas concentrantes IgG, IgA, IgM, IgD IgE, estas
clases se diferencian entre si, en cuanto a su tamaño, carga, composición de aminoácidos y
contenidos de hidratos de carbono.
Son moléculas bifuncionales porque es capaz de unirse al antígeno mientras que
otra región ejerce las denominadas fuerzas efectoras.
ESTRUCTURA DE LAS INMUNOGLOBULINAS:
Las inmunoglobulinas son glicoproteínas que, según ya indicó Porter en 1959, están
formadas por cadenas polipeptídicas agrupadas, dependiendo del tipo de inmunoglobulina,
en una o varias unidades estructurales básicas.
Unidad estructural básica:
Cada unidad está compuesta por cuatro cadenas polipeptídicas unidas entre sí por
puentes disulfuro y otras uniones de tipo no covalente (figura 03). Para su estudio se han
empleado diferentes procedimientos. Por ejemplo, tras la rotura de los puentes disulfuro por
sustancias de carácter reductor, como el mercaptoetanol, se individualizan las cuatro
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 11 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
cadenas polipeptídicas y éstos atendiendo a su tamaño, son de dos tipos: de bajo peso
molecular (aproximadadamente 22 KD) y de alto peso molecular (5070 KD, dependiendo
del tipo de Ig). Los polipéptidos de bajo peso molecular reciben el nombre de cadenas
ligeras o cadenas L (Light) y las de alto peso molecular, cadenas pesadas o cadenas H
(Heavy).
Figura 03
Dos cadenas ligeras y dos cadenas pesadas se agrupan de tal manera que existe
una proximidad espacial entre los cuatro extremos amínicos de las cadenas ligeras y
pesadas por una parte, y entre los dos extremos carboxílicos de las cadenas pesadas por
otra.
Figura 04
Esta estructura
básica de las
inmunoglobulinas puede
ser fraccionada
mediante la utilización
de enzimas (papaína,
pepsina, etc.), como fue
efectuado por Porter en
1959, obteniéndose
diferentes tipos de
fragmentos (figura 04).
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 12 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
El tratamiento con papaína produce la ruptura específica de las cadenas H, en el espacio
comprendido entre el puente disulfuro que las une entre sí y los que las unen a las cadenas
ligeras. Se obtienen tres fragmentos: uno denominado Fc, que determina la actividad
biológica, contiene el alotipo y determina la clase y subclase de cadena pesada y dos
denominados cada uno de ellos Fab, que contienen el idiotipo y es por donde la molécula se
une al antígeno.
Cadenas Ligeras:
Hay dos tipos de cadenas ligeras, estructuralmente diferentes, que se conocen como
cadenas ligeras tipo kappa (k) y cadenas ligeras tipo lambda (l). La familia de genes que
codifica para la cadena ligera k se localiza en el cromosoma 2 y los loci de los genes
homólogos que codifican para la cadena l, en el cromosoma 22. En cada molécula de
inmunoglobulina las dos cadenas ligeras son del mismo tipo, k o bien l, pero nunca existe
una de cada tipo en la misma inmunoglobulina.
Las cadenas ligeras están formadas por unos 200 aminoácidos con la particularidad
de que existen dos puentes disulfuro que unen grupos de unos cincuenta aminoácidos.
Concretamente la IgG1 posee 214 aminoácidos y su estructura secundaria y terciaria están
determinadas por dos puentes disulfuro intracatenarios que unen los aminoácidos 23 con el
88 y 134 con el 193 (figura 06). A su vez, estas cadenas ligeras tienen otro puente disulfuro
intercatenario, por el cual cada una de ellas se une a una cadena pesada para constituir la
unidad básica de las inmunoglobulinas. Este puente se encuentra en el último aminoácido
(214) de la parte carboxílica para el tipo k y en el penúltimo para el tipo l.
Figura 05: Las estructuras de color verde
vienen hacer la cadenas ligeras.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 13 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
Figura 06
Cadenas pesadas:
Estas cadenas poseen unos cuatrocientos aminoácidos estableciéndose entre
algunos de ellos puentes disulfuro (intracatenarios) que asocian unos 60 aminoácidos y que
condicionan la estructura secundaria del polipéptido. Por ejemplo, las cadenas pesadas de
la IgG1 poseen 440 aminoácidos y los puentes disulfuro unen el aminoácido 22 con el 96, el
144 con el 200, el 261 con el 321 y el 367 con el 425.
Estas dos cadenas pesadas están unidas la una a la otra por puentes disulfuro
intercatenarios, ya indicados anteriormente, y que pueden ser de uno a cinco dependiendo
del tipo de inmunoglobulina.
En estas cadenas pesadas, y a nivel de los puentes disulfuro intercatenarios, hay
una zona de unos 15 aminoácidos, de gran flexibilidad debido a su estructura y constituye lo
que se denomina zona bisagra por donde se deforma la molécula de inmunoglobulina
cuando se produce la unión con el antígeno,
facilitándose así su acoplamiento con éste. Los loci de
los genes que codifican para la cadena pesada se
localizan en el brazo largo del cromosoma 14.
Figura 076: Estructuras de color rojo vendrían hacer
las cadenas pesadas
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 14 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
Parte variable y constante de las cadenas ligeras y pesadas:
Estructuralmente, las cadenas ligeras poseen dos partes: una corresponde al
extremo carboxílico que diferencia las cadenas ligeras en dos tipos k y l, y constituye la
parte constante de las cadenas ligeras (CL). La otra corresponde al extremo amínico, que es
muy variable y constituye la parte variable de las cadenas ligeras (VL) y corresponde a la
zona de interacción con el antígeno. Las partes constante y variable son prácticamente de
igual tamaño en las cadenas ligeras.
También las cadenas pesadas poseen una parte variable y otra constante.
Aproximadamente el tercio del extremo amínico de estas cadenas se caracteriza por ser
estructuralmente muy variable, por lo que se conoce como parte variable de las cadenas
pesadas (VH). La estructura de este fragmento, al igual que en las cadenas ligeras, depende
del tipo de antígeno que reconoce, dado que este extremo también participa en la unión de
la inmunoglobulina con el antígeno. Por el contrario, aproximadamente los dos tercios del
extremo carboxílico de todas las cadenas pesadas de un mismo tipo de inmunoglobulinas
poseen una estructura idéntica. De ahí que esta parte de las cadenas pesadas se conozca
como parte constante de las cadenas pesadas (CH).
Esta parte constante es diferente según la clase de inmunoglobulina que
consideremos, determinando la existencia de cinco tipos de cadenas pesadas: g, a, m, d y e
que definen a su vez las cinco clases de inmunoglobulinas: IgG, IgA, IgM, IgD e IgE
respectivamente (figura 08)
Figura 08
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 15 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
.
Figura 09: Parte variable y constante
de las cadenas ligeras y pesadas.
Características de los distintos tipos de inmunoglobulinas:
Debido a esta distinta estructura, las cadenas pesadas van a presentar distintas
propiedades biológicas, tales como la capacidad de unirse entre sí, fijar complemento, fijar la
pieza de secreción y unirse a macrófagos, neutrófilos y células NK.
Propiedades biológicas de las subclases de IgG
Características IgG1 Igg2 Igg3 Igg4
Fijación de complemento ++++ ++ ++++ +/
Anafilaxis cutánea + +
Unión a receptores Fc de
polimofornuclares
monocitos
+
+
+
+
+
+
Transferencia placentaria + + + +
Isotipos:
Si inmunizamos un animal de una especie con inmunoglobulinas procedentes de una
especie distinta, la mayoría de los anticuerpos generados (antisuero heterólogo) irán
dirigidos contra la región constante de la inmunoglobulina que hayamos inyectado,
permitiendo definir lo que llamamos el isotipo de una inmunoglobulina determinada. Los
genes que codifican para las distintas variantes isotipicas están presentes en todos los
individuos sanos, es decir, todos los individuos sanos poseen los genes g1, g2, g3, g4, m,
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 16 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
a1, a2, d, e, k y l; que codifican respectivamente para las regiones constantes G1, G2, G3,
G4, M, A1, A2, D y E de las cadenas pesadas y para las regiones kappa y lambda de las
cadenas ligeras. Existen cinco isotipos de cadena pesada (M, G, A, D y E) y dos de cadena
ligera (k y l). Así diremos que el isotipo de una determinada inmunoglobulina es G1 o que
esa inmunoglobulina es de la clase G y subclase 1, que a su vez puede tener unas cadenas
ligeras del isotipo kappa o lambda.
Dominios moleculares en las cadenas ligeras y pesadas:
Tanto las cadenas pesadas como las ligeras poseen grupos de aminoácidos unidos
por puentes disulfuro intracatenarios. Estos segmentos repetidos en las cadenas L y H se
conocen como dominios. Los dominios de la parte constante de las cadenas pesadas
presentan una gran homología secuencial no sólo entre ellos, sino también con la región
constante de la cadena L. De forma similar, los únicos segmentos variables en las cadenas
L y H presentan cierta homología entre ellos y en menor grado a los de la región constante.
Figura 10
La cadena L tiene dos dominios, uno corresponde a la región variable (VL) y otro a la
constante (CL). La cadena H tiene un dominio en la región variable (VH) y tres o cuatro en la
constante dependiendo de la clase de inmunoglobulina que consideremos (tres en la IgG,
IgA e IgD y cuatro en las IgM e IgE). Estos dominios de la región C se denominan CH1,
CH2, CH3 y CH4 cuando aparece este último.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 17 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO
UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
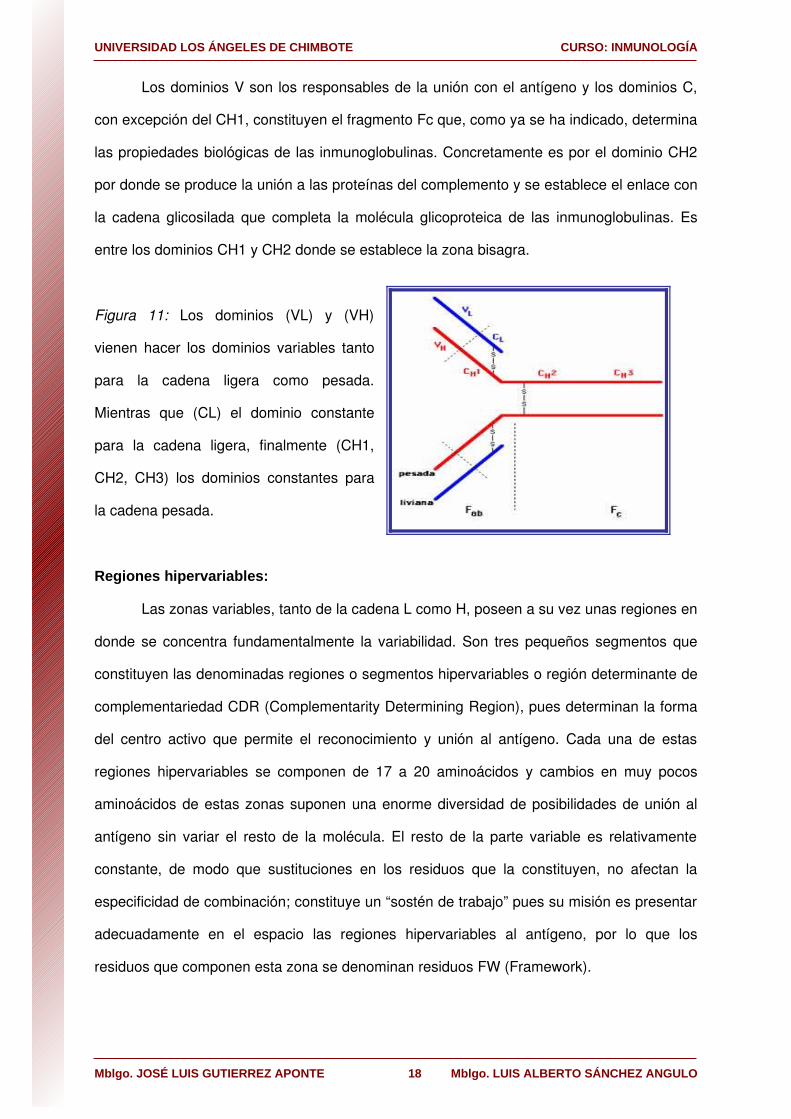
Los dominios V son los responsables de la unión con el antígeno y los dominios C,
con excepción del CH1, constituyen el fragmento Fc que, como ya se ha indicado, determina
las propiedades biológicas de las inmunoglobulinas. Concretamente es por el dominio CH2
por donde se produce la unión a las proteínas del complemento y se establece el enlace con
la cadena glicosilada que completa la molécula glicoproteica de las inmunoglobulinas. Es
entre los dominios CH1 y CH2 donde se establece la zona bisagra.
Figura 11: Los dominios (VL) y (VH)
vienen hacer los dominios variables tanto
para la cadena ligera como pesada.
Mientras que (CL) el dominio constante
para la cadena ligera, finalmente (CH1,
CH2, CH3) los dominios constantes para
la cadena pesada.
Regiones hipervariables:
Las zonas variables, tanto de la cadena L como H, poseen a su vez unas regiones en
donde se concentra fundamentalmente la variabilidad. Son tres pequeños segmentos que
constituyen las denominadas regiones o segmentos hipervariables o región determinante de
complementariedad CDR (Complementarity Determining Region), pues determinan la forma
del centro activo que permite el reconocimiento y unión al antígeno. Cada una de estas
regiones hipervariables se componen de 17 a 20 aminoácidos y cambios en muy pocos
aminoácidos de estas zonas suponen una enorme diversidad de posibilidades de unión al
antígeno sin variar el resto de la molécula. El resto de la parte variable es relativamente
constante, de modo que sustituciones en los residuos que la constituyen, no afectan la
especificidad de combinación; constituye un “sostén de trabajo” pues su misión es presentar
adecuadamente en el espacio las regiones hipervariables al antígeno, por lo que los
residuos que componen esta zona se denominan residuos FW (Framework).
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 18 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO
UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
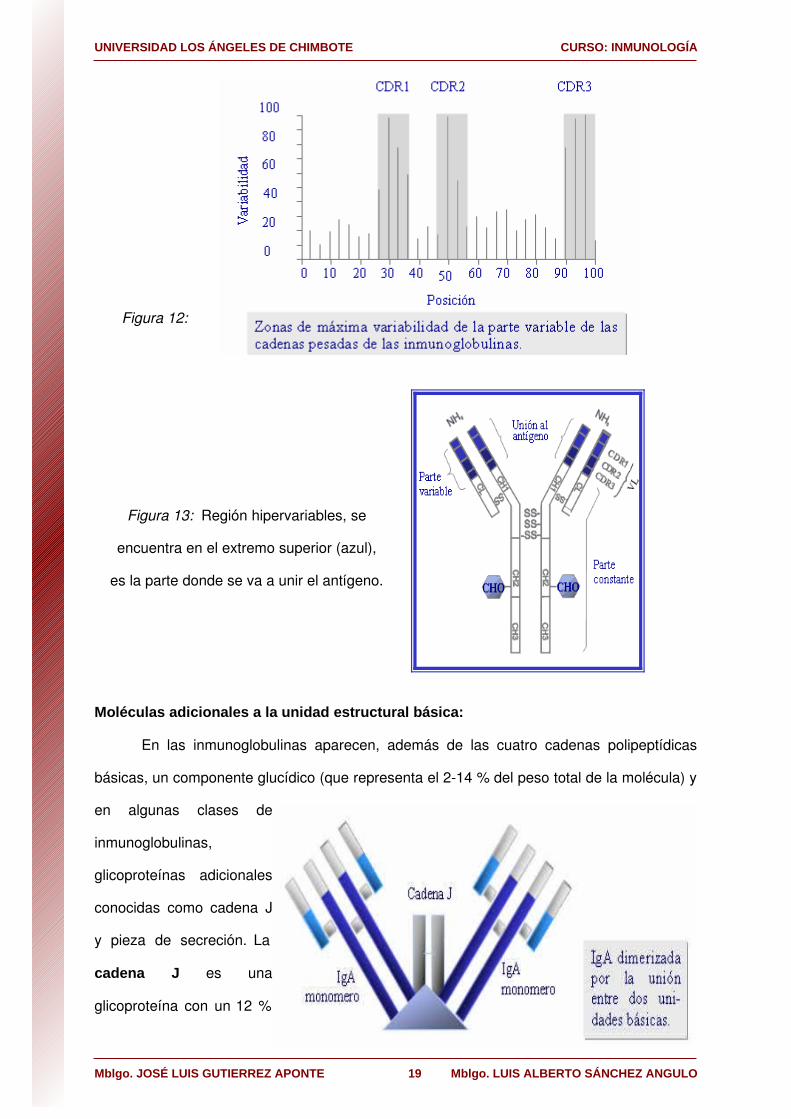
Figura 12:
Figura 13: Región hipervariables, se
encuentra en el extremo superior (azul),
es la parte donde se va a unir el antígeno.
Moléculas adicionales a la unidad estructural básica:
En las inmunoglobulinas aparecen, además de las cuatro cadenas polipeptídicas
básicas, un componente glucídico (que representa el 214 % del peso total de la molécula) y
en algunas clases de
inmunoglobulinas,
glicoproteínas adicionales
conocidas como cadena J
y pieza de secreción. La
cadena J es una
glicoproteína con un 12 %
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 19 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
de azúcares y un peso molecular de 15 kD que une, mediante puentes disulfuro, extremos
Fc en la IgA e IgM. La pieza de secreción es una glicoproteína de 58 kD de peso molecular
que sintetizan las células epiteliales de las mucosas y glándulas exocrinas.
TIPOS Y FUNCION QUE CUMPLEN:
➢ Todas las Ig son glicoproteínas pero su concentraciones de hidratos de carbono
oscila entre el 23% de las IgG y el 1214% de las IgM. IgD e IgE.
IgG: es la mas abundante en el suero humano normal, en donde constituye el 7075% de
las Ig totales. Los Anticuerpos de clase IgG ,se encuentran distribuidos en los
compartimientos intra y extra vascular:
● Son los únicos que presentan actividad frente a las toxinas.
● Predominante y confieren las respuestas inmunitarias secundarias.
● Son procedentes de la inmunidad al recién nacido durante los primeros meses de
vida.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 20 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO
IgG1
UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
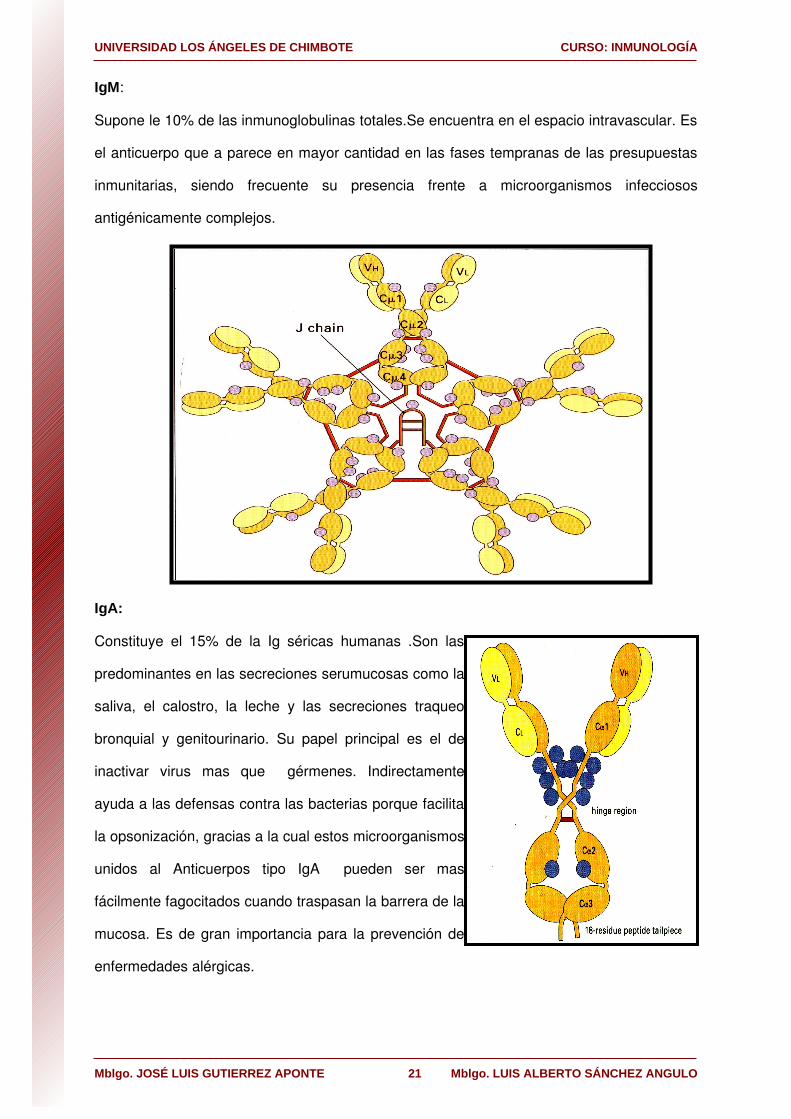
IgM:
Supone le 10% de las inmunoglobulinas totales.Se encuentra en el espacio intravascular. Es
el anticuerpo que a parece en mayor cantidad en las fases tempranas de las presupuestas
inmunitarias, siendo frecuente su presencia frente a microorganismos infecciosos
antigénicamente complejos.
IgA:
Constituye el 15% de la Ig séricas humanas .Son las
predominantes en las secreciones serumucosas como la
saliva, el calostro, la leche y las secreciones traqueo
bronquial y genitourinario. Su papel principal es el de
inactivar virus mas que gérmenes. Indirectamente
ayuda a las defensas contra las bacterias porque facilita
la opsonización, gracias a la cual estos microorganismos
unidos al Anticuerpos tipo IgA pueden ser mas
fácilmente fagocitados cuando traspasan la barrera de la
mucosa. Es de gran importancia para la prevención de
enfermedades alérgicas.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 21 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
IgD:
Supone el1% de las Ig plasmáticas totales, perro
abundan en las membranas de las células B, No
se sabe bien cual es la función biológica exacta
de la IgD pero puede desempeñar un papel muy
importante de los linfocitos inducidos por el
antígeno.
IgE:
Se encuentra en la superficie de los basófilos y los mastocitos de todos los individuos.
También sensibiliza a la célula como
algunas mucosas, como la conjuntiva, la
mucosa braquial y nasal. Cumple una
función importante como el control de las
enfermedades parasitarias. En los países
desarrollados se suele asociar con las
enfermedades alérgicas como el asma o la
fiebre del heno.
.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 22 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
SISTEMA DEL COMPLEMENTO
El sistema de complemento forma parte del sistema inmunitario, y a lo largo de la
evolución ha desarrollado mecanismos para distinguir lo propio de lo ajeno.
LA NOMENCLATURA DEL SISTEMA DE COMPLEMENTO ES COMPLEJA:
En la nomenclatura se asigna un numero a cada de las proteínas de la vía clásica y
del complejo de ataque a la membrana. Las proteínas del complemento se agrupan en
superfamilias, esta familia de proteínas son:
Factor H
Proteína fijadora
Factor acelerador de la degradación
Cofactor proteico de la membrana
Receptores de complemento de tipo I
LAS PROTEINAS DEL COMPLEMETO PRESENTAN UNA ESTRUCTURA EN MOSAICO:
Muchas proteínas del sistema del complemento están formadas por un mosaico de
exones procedentes de otras familias. Las 2 vías principales de activación de complemento
reflejan las respuestas inmunitarias innatas y adaptativas.
Las consecuencias de la activación del complemento son:
Opsonización
Activación de los leucocitos
Lisis de las células dianas
LA VÍA CLÁSICA:
Es el mecanismo fundamental de la activación del complemento mediada por
anticuerpos.
La activación se inicia tras la unión de C1 a un inmunocomplejo
Receptor del complemento.
Muchos de los fragmentos de las proteínas del complemento k se generan durante
el proceso de activación, se unen a receptores específicos situados sobre la
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 23 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
superficie de las células del sistema inmunitario. este es un mecanismo importante
de mediación de lo efectos fisiológicos del complemento, entre los k se encuentran la
ingestión de partículas opsonizadas y la activación de las células poseedoras de los
receptores.
Tipos de receptores de fragmentos de C3 unidos covalentemente
Se conocen cuatro receptores distintos de estos fragmentos opsonizantes , que se
denominan receptores del complemento de tipo 1 a 4 .
• CR1
• CR2
• CR3
• CR4
CR1
Denominada receptor de adherencia inmunitario, receptor del C3b /C4b.
Desempeñando 4 funciones biológicas
CR2 Receptor relacionado con la activacion de los linfocitos B.
En células B el CR2 actúa como receptor accesorio de anticuerpos en la activación
de las respuestas inmunitarias especificas.
La unión de iC3b o C3dg a CR2 reduce el umbral necesario para que la ocupación
de los receptores específicos de antígeno de células B, los anticuerpos de
membrana , desencadene la activación celular.
VIA ALTERNATIVA:
Es una de las vías de activación del complemento que se origina por el
reconocimiento directo de ciertas estructuras de las superficies de los microorganismos por
una reacción que se produce en el plasma mediante la activación al ralentí de C3 que
origina el pequeño depósito de pequeñas cantidades de moléculas de C3 sobre la superficie
propia y ajena.
CARACTERISTICAS:
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 24 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
Para que se active esta vía es necesaria la presencia de superficies protegidas.
Se da el catabolismo de las moléculas de C3 depositadas sobre la superficie del
huésped por medio de las moléculas reguladoras que interrumpen el proceso de
activación del complemento.
El C3 depositado por la activación al ralentí en superficies extrañas que carecen de
moléculas reguladoras activa la vía alternativa del complemento por tener mayor
afinidad por el factor B que se encuentra en ellas
A comparación de la vía clásica no necesita de anticuerpos para activarse ya se
activa por el reconocimiento directo de ciertas estructuras de la superficie de los
microorganismos y por ello forma parte de la inmunidad innata.
Fue descubierta posteriormente a la vía clásica pero filogenéticamente es mas
antigua.
Se pone en manifiesto la protección de lo propio y la lisis de lo ajeno mediante los
factores de la regulación. Estas proteínas de membrana se detectan
fundamentalmente en eritrocitos y células endoteliales que son las células que se
encuentran en mayor peligro de autolisis por el complemento.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 25 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
MEDIO DE ACTIVACION:
➢ Se inicia con la activación al ralentí de C3 depositado en la superficie extrañas
(microorganismo) que carecen de moléculas reguladoras desencadenando la
síntesis de C3b (los pasos iníciales).
➢ C3b inicia los pasos finales en la activación del complemento que culmina en la
producción de péptidos que estimulan la inflamación C5a (fenómeno básico en la
defensa contra la infección), y C9 polimerizado, que forma el complejo de ataque a la
membrana.
Se distinguen 02 fases:
Fase de reposo: esta se va a dar cuando la molécula C3iBb al actuar en fase líquida la
mayor parte de C3b* generado es hidrolizados e inactivado.
Fase de amplificación: se va dar cuando la molécula se encuentra situado en las
proximidades de una superficie perteneciente a una estructura ajena, como la membrana de
una célula bacteriana, se une covalentemente a la misma e inicia el circuito de amplificación
de la vía alternativa.
La regulación del mecanismo de amplificación es importante en el huésped, ya que
se trata de mecanismo de retroalimentación positiva que, de no ser controlado, no se
interrumpiría hasta que se agote todo el C3.
En resumen
La regulación del destino de C3b unido a superficies es el factor crítico que permite al
sistema del complemento distinguir entre lo propio y lo ajeno. El C3b unido puede correr dos
tipos de suertes diferentes: amplificación e inhibición
Amplificación: C3b actúa como punto de unión para el factor B, da lugar a una convertasa
e induce el depósito de C3b sobre la misma superficie.
Inhibición: C3b es catabolizado por el factor I con la intervención de uno de estos tres
cofactores: el factor H (plasmático), CR1 o MCP (membrana).
La clave para que se produzca esta rápida amplificación de C3b radica en la
formación de una C3 CONVERTASA de SUPERFICIE.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 26 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
La naturaleza de la superficie a la que se encuentra unido C3b es el factor que
determina cuál de estas dos vías se activa.
Acción facilitadora de la fagocitosis (opsonización): los macrófagos y polimorfo
nucleares neutrófilos presentan en sus membranas receptores capaces de unir la molécula
C3b y sus derivados resultantes de la activación del complemento. De esta forma, si el C3b
está fijado sobre la superficie de un germen, los fagocíticos pueden conectar con éste
mediante los receptores para C3b, facilitándose así el fenómeno la fagocitosis.
Aclaramiento de inmunocomplejos: Existe un mecanismo fisiológico de aclaramiento de
IMC. Los eritrocitos, las células más abundantes de la sangre, presentan CR1 en su
membrana y mediante este receptor captan IMC circulantes a través del factor C3b. Cuando
los hematíes atraviesan el hígado o el bazo, los macrófagos de estos órganos, mediante sus
receptores unen los IMC a través de C3b y lo fagocitan.
Acción reguladora de la respuesta inmune: el factor C3b tiene importantes funciones
reguladoras de la respuesta inmune. Así, el C3b o sus derivados, unido a membranas facilita
la cooperación entre las células inmunes y actúa en la estimulación de las células T y B
probablemente debido a su capacidad de promover la adhesión célulacélula.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 27 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO

UNIVERSIDAD LOS ÁNGELES DE CHIMBOTE CURSO: INMUNOLOGÍA
LA FASE FINAL O VIA LITICA
La fase final de la cascada del complemento es la formación del complejo de ataque
a la membrana (CAM) las enzimas convertasas de C5, formadas ya sea en la vía clásica o
alternativa, escinde a C5 para dar lugar a C5a que es una potente anafilotoxina que pasa al
medio fluido y el fragmento C5b de mayor peso molecular se une a C6 y C7 y da lugar a
C5b67, de naturaleza hidrófoba que contiene un punto de unión que le permite fijarse a las
membranas plasmáticas próximas a las zonas de reacción. C8 se une a este complejo y
penetra en la membrana, seguido de inserción y polimerización de una serie de moléculas
C9 con lo que queda formado un “enchufe” lítico o molécula formadora de poros. El principal
inductor de la misma es el componente C 9 polimerizado. Este complejo se le puede
representar como C5b9y los complejos previos se pueden abreviar de la misma forma por
ejemplo, C5b8.
Mblgo. JOSÉ LUIS GUTIERREZ APONTE 28 Mblgo. LUIS ALBERTO SÁNCHEZ ANGULO