FISIOLOGIA NEUROMUSCULAR 2011
38
-
Upload
luis-alberto-isea-m -
Category
Documents
-
view
2.595 -
download
0
description
Fisiología de los tejidos excitables, realizada por el Dr. Luis Alberto Isea M.
Transcript of FISIOLOGIA NEUROMUSCULAR 2011

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 1/38

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 2/38
POTENCIAL DE MEMBRANA
La membrana celular es una bicapa lipidica selectivamente permeable a diferentes moléculas, hecho, es prácticamente impermeable a los iones, comportándose como una especie de aislanentre 2 soluciones llenas de iones (cargas eléctricas), el liquido intracelular y el liquido extracelulComo sabemos existe una diferencia en la distribución ionica entre el exterior y el interior celuexistiendo una gran cantidad de Na+ y Cl- en el exterior de la celula y una gran cantidad de K+ y
(proteínas, AA) en el interior celular, en base a lo establecido según el equilibrio de Gibbs-donn(el cual determina dicha distribución de cargas a ambos lados de la membrana). Al ser iones moléculas cargadas eléctricamente (bien sea positiva o negativamente), hay una distribucdiferencial de cargas eléctricas dentro y fuera de la célula, es decir, existe un potencial eléct
diferente a ambos lados de la membrana, cuando decimos potencial eléctrico hablamos de energpotencial eléctrica (sencillamente energía eléctrica generada por un acumulo de cargas eléctricque son en este caso los iones).
Esta separación de cargas se mantiene porque la doble capa lipídica de la membrana bloquealibre difusión de los iones. Por lo que esta separación de cargas da origen a una diferencia
potencial eléctrico, o voltaje, a través de la membrana (es decir, debido a los diferentes potenciaeléctricos a ambos lados de la membrana se genera una diferencia de potencial, o diferencia voltaje), denominado POTENCIAL DE MEMBRANA (Vm)
El potencial de membrana se define como: El potencial eléctrico en la parte interna de la cél
menos el potencial en la parte externa. Es decir, Vm = Vic - Vec
Sin embargo, por convención, el potencial fuera de la célula se establece en 0. Por lo cualpotencial de membrana va a ser igual al potencial en la parte interna de la celula. Vm = Vic
Si aplicamos la Ley de Ohm para determinar la diferencia de potencial, I = ∆P/R, podríamos despe
la formula, y concluir que la diferencia de potencial (∆P), es igual a la corriente eléctrica que fluye multiplicada por la resistencia de la membrana (R), siendo esta otra forma de interpretar el potencde membrana.
El potencial de membrana depende entonces de la diferente distribución ionica a través demembrana plasmática, lo cual a su vez, esta regulado por el trafico o movimiento de iones a travde la misma. Debemos acotar que toda celula excitable tiene la capacidad de responder activamea un estímulo mediante la modificación de su potencial de membrana, por lo que debemos defdos estados celulares: reposo y excitable. Ambos presentaran diferente potencial de membrana, lo que el movimiento de iones a través de la membrana es distinto en ambos estados.
El movimiento de los iones (responsable del potencial de membrana), esta mediado por canaiónicos, una clase de proteínas integrales de membrana que se encuentra en todas las células organismo, y que permiten el pasaje de ciertos iones a través de la membrana. Como todsabemos la membrana celular es una bicapa lipidica selectivamente permeable, por lo cual no todlas moléculas pueden atravesarla libremente, de hecho, la membrana es impermeable a los ion(ya que son moléculas con carga eléctrica), por lo cual para el tránsito de iones en la célula necesario la presencia de estos canales iónicos. Los canales iónicos son altamente selectivospermiten solamente el paso de iones con características específicas. Esta selectividad se basa tanen el tamaño del ion como en la carga del mismo. Por ejemplo, canales que presenten ca

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 3/38
negativa en su interior (debido a la presencia de residuos de aminoácidos con carga negativa, dende la estructura del canal) suelen permitir el paso de cationes (iones con carga positiva), peexcluyen el paso de aniones (iones con carga negativa), y viceversa. Igualmente, un canal qpermita el paso de cationes podría permitir el paso solo de Na+ y excluir el paso de K+, a pesar que sean ambos iones positivos, debido a que el Na+ es en la practica un ion de mayor tamaño qel K+ y no podría atravesar el canal.
Los canales iónicos constituyen un mecanismo de transporte pasivo, al no requerir un gasto extra energía metabolica para permitir la movilización del ión, de tal manera, que este se desplaza en baa su gradiente electroquímico. Siendo subclasificados como difusión facilitada, al estar etransporte mediado por un canal proteico. Poseen 3 propiedades importantes:
a) Conducen iones a velocidades extremadamente altasb) Reconocen y seleccionan iones específicosc) Presentan una cinetica lineal, no saturable
Se pueden distinguir 2 tipos de canales ionicos por sus diferentes funciones en la señalizacneuronal y su diferente mecánica de activación:
- En reposo: Son canales iónicos que se encuentran activos (abiertos) cuando la celula estaestado de reposo, es decir, en ausencia de estimulación o excitación. Estos canales reposo contribuyen significativamente al potencial de reposo de la celula.
- Regulados: Los cuales se abren y cierran en respuesta a diferentes estimulos.Existen canales voltaje-dependientes (o regulados por voltaje), los cuales responden avariación del potencial de membrana; canales ligando-dependientes los cuales respondenmensajeros químicos (hormonas, neurotransmisores, segundos mensajeros), y los regulad
mecánicamente los cuales son sensibles a la presión o estiramiento de la membrana.
El potencial de membrana de una celula en reposo recibe el nombre de potencial de membranareposo, y suele ser de alrededor de -60 a -70 mV, debido al gran acumulo de cargas eléctricnegativas en el interior celular.
Para lograr entender como la diferencia de distribución de los iones genera este potencial de repocomenzaremos con un ejemplo de permeabilidad exclusiva de membrana, como es el caso de células gliales, las cuales son exclusivamente permeables a un tipo de ion en reposo (potasio), eexplicación nos detallara igualmente lo que es el potencial de equilibrio de un ion.
La celula glial tiene un potencial de reposo de -75mV, en ella la inmensa mayoría de los canales
reposo no son permeables mas que a K+. Como resultado de ello, la membrana de la celula glialpermeable casi de forma exclusiva a los iones de K+. Al igual que el resto de las células, la glia tieuna elevada concentración de K+ y A- en el interior celular y una gran concentración de Na+ y Cl- la externa. Como el K+ esta presente en una gran concentración en el interior de la celula se geneun gradiente químico que lo impulsa a salir al exterior, como resultado de esto, progresivamenteinterior celular se va haciendo mas negativo (ya que están saliendo cargas positivas, el K+, y se equedando los A-, que son proteínas, que no pueden salir a través de la membrana), esto lleva a qa medida que el K+ sale (y el interior se va haciendo cada vez mas negativo), la misma difusión autolimita, llega un punto en que el interior es tan negativo, que comienza a atraer por gradieeléctrico al potasio, contrarrestando su salida por gradiente quimico. Asi podemos ver que el traf

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 4/38
de iones esta sujeto a 2 fuerzas: una fuerza de arrastre química que depende del gradiente concentración a través de la membrana (que, en este caso, impulsa al K+ a salir); y una fuerza
arrastre eléctrica que depende de la diferencia de potencial a través de la membrana (que, en ecaso, al salir progresivamente el K+, y volviéndose el interior celular mas negativo, atrae K+ medio externo).
En resumen, una vez que la difusión de K+ ha alcanzado cierto punto, se desarrolla un potencia
través de la membrana con el cual la fuerza eléctrica que arrastra K+ al interior de la celula esta perfecto equilibrio con la fuerza química que arrastra al K+ fuera de la celula. Es decir, el movimiehacia fuera de K+ (arrastrado por su gradiente de concentración químico) es igual al movimiehacia dentro de K+ (arrastrado por la diferencia de potencial eléctrico de la membrana). Espotencial de membrana al cual ocurre dicho fenómeno, recibe el nombre de potencial de equilib
del potasio. En una celula permeable tan solo a los iones de K+, el Ek (potencial de equilibrio potasio) es igual al potencial de membrana en reposo, que es de -75 mV.
Entonces podemos definir al potencial de equilibrio de un ion como el potencial de membrana al c
no hay un flujo neto del ion a través de la membrana , o bien como el potencial de membrana q
contrarresta la tendencia del ion a seguir su gradiente de concentración químico. Ver figura 1.En el potencial de equilibrio se alcanza un equilibrio electroquímico en donde las fuerzas químicaeléctricas que actúan sobre un ion son iguales y opuestas y no ocurre difusión neta del mismo.
Figura 1. Difusion del Na+ a traves de una membrana exclusivamente permeable al mismo, que demuestra quedifusión del mismo se mantendra hasta que las cargas negativas en el compartimiento 1, tiendan a arrastrar al interiomisma cantidad de ion que tiende a salir, y se establezca el potencial de equilibrio.
El potencial de equilibrio para cualquier ion puede calcularse mediante una ecuación derivada 1888 de los principios básicos de la termodinámica por el fisicoquímico alemán Walter Nernst.ecuación de Nernst puede utilizarse para calcular el potencial de equilibrio de cualquier ion qse encuentra a ambos lados de una membrana permeable para el.
Una vez descrito como los gradientes ionicos generan el potencial de reposo glial, y como este igual al potencial de equilibrio del potasio, (por tener una permeabilidad exclusiva al mismAnalizaremos el caso de las neuronas y el resto de las células, en donde esto no ocurre igual, ya qlas neuronas son permeables a 3 tipos de iones en reposo, el Na+, el K+ y el Cl-.
Para entender como se relacionan estos iones, vamos a pensar rápidamente en nuestro ejemanterior de las células gliales, imagínense que a estas células (exclusivamente permeables al pota

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 5/38
en reposo), le ponemos ahora unos canales ionicos abiertos en reposo para el Na+, que va a pasapues que el Na+ va a tender a entrar al interior de la celula, siguiendo su gradiente electroquími(eléctrico por la carga negativa en el interior de la celula, derivada de la salida de potasio en reposy el acumulo de las proteínas intracelulares con carga negativa, y químico por la miniconcentración del mismo en el medio intracelular). Este influjo de cargas positivas (Na+) despolarla celula (es decir, lleva su potencial de membrana a valores mas positivos, ya que estoy metiencargas positivas al interior celular, disminuyendo la diferencia de cargas entre el interior y el exter
celular), pero solo ligeramente por encima del potencial de equilibrio del K+. El nuevo potencial membrana no se aleja mucho del potencial inicial ya que la “conductancia” ó permeabilidad demembrana en reposo al Na+ es muy poca, es decir, solamente hay pocos canales de Na+ abieren reposo, por lo cual no logra entrar tantos iones como para despolarizar significativamentemembrana.
En el ejemplo glial estábamos en un potencial de equilibrio, ya que había un equilibrio entrecantidad de K+ que salía y que entraba por la membrana, pero ahora ya que se despolariza un pola membrana por la entrada de Na+, esto reduce la fuerza eléctrica negativa que arrastraba al K+interior de la celula, por lo tanto se establece un flujo neto de K+ al exterior de la celula (siguiendo
gradiente electroquímico), este flujo neto tiende a contrarrestar el flujo de entrada del Na+. Porque, el potencial de membrana alcanza un nuevo potencial de reposo, un nuevo punto de equilibcon el cual el movimiento hacia fuera de K+ neto equilibra exactamente el movimiento hacia dende Na+. Este punto de equilibrio suele ser de -60mV a -70 mV, en el caso de la neurona.
Para comprender como se determina este punto de equilibrio, téngase en cuenta que la magnitud flujo de un ion a través de una membrana celular es el producto de su fuerza de arraselectroquímica (la suma de las fuerzas de arrastre químicas y eléctrica) por la conductanciapermeabilidad de la membrana al ion.
Una célula tiene relativamente pocos canales de reposo abiertos para el Na+, de forma que, estado de reposo, la conductancia para el Na+ es muy baja. Por lo que, pese a las grandes fuerzquímicas y eléctricas que arrastran al Na+ al interior de la celula, la entrada de este es escasa. contraste con ello, como hay muchos canales de reposo de K+, la conductancia de membrana pael K+ es relativamente grande. Como resultado de ello, la pequeña fuerza neta hacia fuera que actsobre el K+ con el potencial de membrana en reposo es suficiente para producir un flujo de K+ hafuera igual al de Na+ hacia dentro.
Ahora, si el potencial de membrana en reposo depende del gradiente electroquímico para el sodipara el potasio, debemos suponer, que estos gradientes no pueden mantenerse de mane
indefinida. La salida constante de potasio estaría limitada al agotarse el potasio intracelulalcanzando unas concentraciones similares a las del medio extracelular, terminando la difusión, y a versa para el sodio. La disipación de los gradientes ionicos se evita gracias a la actividad debomba Na+-K+, que mueve estos iones en contra de sus gradientes electroquímicos, extrayenNa+ de la celula e introduciendo en ella K+.
La bomba juega 2 roles importantes en el mantenimiento del potencial de membrana en reposo. primer lugar, tiene un pequeña contribución directa electrogenica, ya que saca 3 cargas positivas Na+ e introduce 2 cargas positivas de K+ al interior celular (dejando el interior celular mas negativEn segundo lugar, la contribución indirecta mas importante es mantener los gradientes ionicos,

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 6/38
que saca fuera de la celula el Na+ que esta entrando y permite que se mantenga el gradienquímico, e igualmente introduce el K+ que esta saliendo, para que su gradiente se mantenga.
En relación con el Cl-, vamos a ignorar su contribución actualmente, y solamente acotaremos, qen reposo no hay un flujo neto del ion, ya que en el potencial de membrana en reposo igualapotencial de equilibrio del ion. Igualmente, en relación con el calculo del potencial de membrana, que esta, en el caso neuronal, determinado por dos o mas clases de iones, la influencia de cada
no esta solo determinado por la concentración del ion, sino también por la facilidad con la que el cruza la membrana, la ecuación que permite relacionar todas estas variables en relación con el NK+ y Cl-, se conoce como ecuación de Goldman (cálculo del potencial de membrana)
Una forma de representar las variaciones del potencial de membrana celular, bien estando la celen reposo o en actividad, es el presente grafico:
Figura 2. Variación del potencial de membrana celular. El presente grafico, no es mas que una demostración de cópuede variar el potencial celular, en donde vemos un primer punto (E K+), el cual representa el potencial de equilibrio potasio (mas negativo que el potencial de membrana en reposo, -90 mV), luego evidenciamos el potencial de repocelular (que como vemos, se acerca al potencial de equilibrio del K+, es decir, en reposo, hay una mayor permeabilide la membrana al K+, sin embargo, no es completa, ya que el potencial no se iguala al potencial de equilibrio del ioluego se evidencia un ascenso posible (en el caso del potencial de accion), hasta incluso el potencial de equilibrio sodio. Si por ejemplo, la linea que dice potencial de reposo -70, estuviese arriba, después de 0, diciendo potencial
accion +30, esto indicaria que en ese momento la celula presenta una mayor permeabilidad al Na+, y por endepotencial se acerca al potencial de equilibrio del sodio.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 7/38
POTENCIAL DE ACCION.
La señalización y comunicación celular depende de variaciones rápidas de la diferencia de potenceléctrico a través de las membranas celulares neuronales en respuesta a un estimulo determinaEstas “señales” eléctricas (potenciales de receptor, potenciales sinápticos) se producen, todas ellpor cambios temporales del flujo de corriente de entrada y salida de la celula que arrastranpotencial eléctrico a través de la membrana celular y lo alejan de su valor en reposo, co
consecuencia de la regulación en la actividad de los canales iónicos de la membrana celular. Y sestas señales (estas variaciones del potencial de membrana) las que permiten el transporte deinformación celular, expresada como una señal electrica, de una forma rápida y segura a largdistancias. Esta señal electrica se denomina potencial de acción. Estos son señales eléctricautoregenerativas cuya amplitud no disminuye a medida que se propaga por el axón, y que surgeraíz del cambio de la conductancia y flujo iónico de la membrana celular.
Desde estudios anteriores de Cole y Curtis en el axón gigante del calamar se sabía que el potencde acción se originaba por cambios de la conductancia iónica a través de la membrana, la caumentaba significativamente en relación al estado de reposo, sin embargo, una cuestión importa
ahora era que iones eran los responsables del potencial?.Los estudios de Hodgkin y Katz hallaron que los principales iones responsables eran el Na+ y el Kya que observaron que la amplitud del potencial disminuía cuando se reducía la concentracexterna de Na+, lo que indicaba que el flujo interno de Na+ era el responsable de la fase elevación del potencial de acción. Igualmente sus datos sugerían que la fase de caída del potencde acción estaba causada por un aumento posterior de la permeabilidad para el K+.
Entonces son las variaciones en las corrientes iónicas de Na+ y de K+ las responsables degeneración del potencial de acción. Las corrientes de Na+ y de K+, al igual que el resto de iondependen de 2 factores: la conductancia para cada ion y la fuerza de arrastre electroquímica qactúa sobre el ion. Tomando en cuenta que la fuerza de arrastre electroquímica se mantiene mamenos estándar para cada ion debido a la actividad de la bomba Na+-K+ ATPasa, la variación decorriente va a depender de cambios en la conductancia de la membrana para el ion, lo cual a su vdepende de modificaciones en la actividad de los canales ionicos correspondientes a cada uno. habíamos descrito que existían varios tipos de canales iónicos, en reposo, y regulados (voltadependientes, ligando-dependientes, mecánico-dependientes), cuando la célula se encuentra reposo, los canales iónicos en reposo para K+ y Na+ se encuentran abiertos (en mucha maproporción los de K+ que los de Na+) y esto generaba el potencial de membrana en reposo; embargo durante el potencial de acción van a activarse unos canales iónicos de Na+ y de K+ volta
dependientes, los cuales se abren en respuesta a la despolarización del potencial de membranestos canales sin embargo difieren un poco en cuanto a su velocidad de apertura y a su respuestauna despolarización prolongada.
Todas las neuronas establecen multiples sinapsis con otras celulas, por lo que cada soma realmenrecibe multiples señales provenientes de diversas celulas, unas de ellas excitator(despolarizantes, Potenciales Postsinapticos Excitatorios) y otras de ellas inhibitor(hiperpolarizantes, Potenciales Postsinapticos Inhibitorios), por lo que la celula debe sopesar señales excitatorias y contrarrestarlas con las inhibitorias, y si al final las señales excitatorias smayores que las inhibitorias, esta excitación (despolarización), puede llegar a un nivel que permita

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 8/38
apertura de los canales de sodio voltaje dependientes y se de inicio a la secuencia del potencial acción. Esto se conoce como sumacion espacial y temporal de los potenciales; espacial, porque realiza recibiendo señales desde diferentes puntos de la celula, y temporal, porque se realizamismo tiempo en el cono axonico, siendo este el sitio de la neurona donde se decidira el disparono del potencial de accion.
Si las señales excitatorias predominan en la celula, y se alcanza una despolarización umbral,
procederá a la activación y apertura de los canales de sodio voltaje dependientes, permitiendoentrada masiva de sodio al interior celular, desplazando el potencial de membrana hacia valopositivos, cercanos al potencial de equilibrio del Na+ (cerca de +50 mV), y esto constituye la fase disparo (overshoot) del potencial de acción
De tal manera, que los canales de Na+ voltaje-dependientes son los responsables de la fase disparo del potencial de acción, presentándose su apertura una vez que se hay una despolarizacde la membrana (producida por un aumento de cargas positivas en el interior celular, probablemena consecuencia de la activación previa de un receptor o canal iónico que permitió la entrada de ionpositivos). Estos canales se caracterizan por presentar una cinetica particular, estos cana
presentan tres estados diferentes, que representan 3 conformaciones diferentes de la proteína canal de Na+: reposo, activación (abierto) o inactivación. Ante el potencial de membrana en repoel canal se encuentra en reposo (cerrado), con la despolarización, el canal va del estado de repo(cerrado) al de activado (abierto). Si la despolarización es breve, los canales vuelven directamenteestado de reposo con la repolarización. Si se mantiene la despolarización los canales van del estaabierto al inactivo (cerrado, aunque en una conformación diferente al estado de reposo). Una vque el canal esta inactivado ya no puede abrirse por una nueva despolarización. La inactivacpuede ser invertida solo mediante repolarización de la membrana a su potencial de reposo originpasándolo nuevamente al estado de reposo. Este cambio requiere algún tiempo, porque los canaabandonan el estado inactivado de una forma relativamente lenta.
Esta cinetica característica del canales de Na+ se debe a que cada canal tiene 2 clases compuertas que deben abrirse de forma simultanea para que el canal conduzca los iones de NCuando la membrana esta en su potencial negativo de reposo hay una compuerta de activación qesta cerrada y que se abre rápidamente con la despolarización; la compuerta de inactivación, porparte, esta abierta durante el potencial de reposo y se cierra lentamente en respuesta a despolarización. El canal no conduce mas que durante el breve periodo de la despolarización en qambas compuertas están abiertas. La repolarización invierte ambos procesos, cerrando rápidamela compuerta de activación y abriendo mas lentamente la de inactivación. Una vez que el canal vuelto al estado de reposo, puede ser activado de nuevo mediante la despolarización. Ver Figura 3

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 9/38
Figura 3. Cinetica y conformacion estructural del canal de sodio, donde se evidencia que en reposo la compuertaactivacion esta cerrada, la cual se abre durante el disparo del potencial, para posteriormente cerrarse la compuertainactivacion durante la repolarizacion, determinando entonces, el potencial refractario absoluto celular.
Una vez que se desarrolla la fase de despolarización del potencial de acción, esta misma limita duración ya que causa la inactivación de forma gradual de los canales de Na+, por lo tanto reduceentrada de Na+, y abre, con cierto retraso, canales de K+ sensibles al voltaje. Por lo tantocorriente de Na+ hacia dentro que se origino en el disparo del potencial de acción, disminuye, y
seguida de una corriente de K+ hacia fuera que tiende a repolarizar la membrana llevandopotencial de membrana nuevamente hacia el valor de reposo, evidenciándose una fase
repolarización. Sin embargo, en la mayoría de las células nerviosas, el potencial de acción seguido de una hiperpolarizacion transitoria, conocida como potencial posterior o hiperpolarizante
cual se origina debido a que los canales de K+ que se abren durante la fase de disparo del potencde acción se cierran lentamente, incluso después de que el potencial de membrana haya vuelto avalor en reposo. Durante este tiempo la conductancia de la membrana al K+ es mayor que enreposo (por los canales de K+ abiertos en reposo, mas los canales de K+ voltaje-dependientabiertos), por lo tanto la membrana se hiperpolariza (alcanza valores negativos), por lo tanpresenta un potencial de membrana cercano al potencial de equilibrio del K+. (ver figura 4)
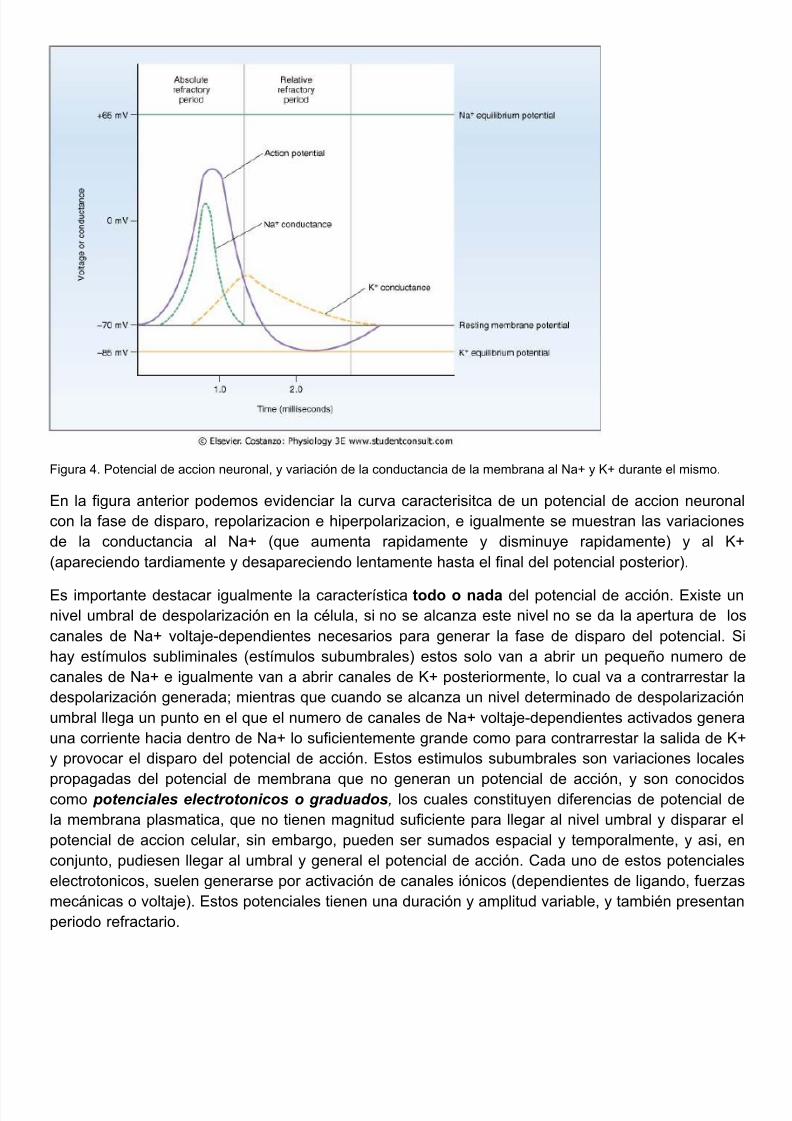
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 10/38
Figura 4. Potencial de accion neuronal, y variación de la conductancia de la membrana al Na+ y K+ durante el mismo.
En la figura anterior podemos evidenciar la curva caracterisitca de un potencial de accion neurocon la fase de disparo, repolarizacion e hiperpolarizacion, e igualmente se muestran las variacionde la conductancia al Na+ (que aumenta rapidamente y disminuye rapidamente) y al (apareciendo tardiamente y desapareciendo lentamente hasta el final del potencial posterior).
Es importante destacar igualmente la característica todo o nada del potencial de acción. Existe nivel umbral de despolarización en la célula, si no se alcanza este nivel no se da la apertura de canales de Na+ voltaje-dependientes necesarios para generar la fase de disparo del potencial.hay estímulos subliminales (estímulos subumbrales) estos solo van a abrir un pequeño numero canales de Na+ e igualmente van a abrir canales de K+ posteriormente, lo cual va a contrarrestadespolarización generada; mientras que cuando se alcanza un nivel determinado de despolarizacumbral llega un punto en el que el numero de canales de Na+ voltaje-dependientes activados geneuna corriente hacia dentro de Na+ lo suficientemente grande como para contrarrestar la salida de y provocar el disparo del potencial de acción. Estos estimulos subumbrales son variaciones loca
propagadas del potencial de membrana que no generan un potencial de acción, y son conocidcomo potenciales electrotonicos o graduados, los cuales constituyen diferencias de potencial la membrana plasmatica, que no tienen magnitud suficiente para llegar al nivel umbral y dispararpotencial de accion celular, sin embargo, pueden ser sumados espacial y temporalmente, y asi, conjunto, pudiesen llegar al umbral y general el potencial de acción. Cada uno de estos potenciaelectrotonicos, suelen generarse por activación de canales iónicos (dependientes de ligando, fuerzmecánicas o voltaje). Estos potenciales tienen una duración y amplitud variable, y también presenperiodo refractario.
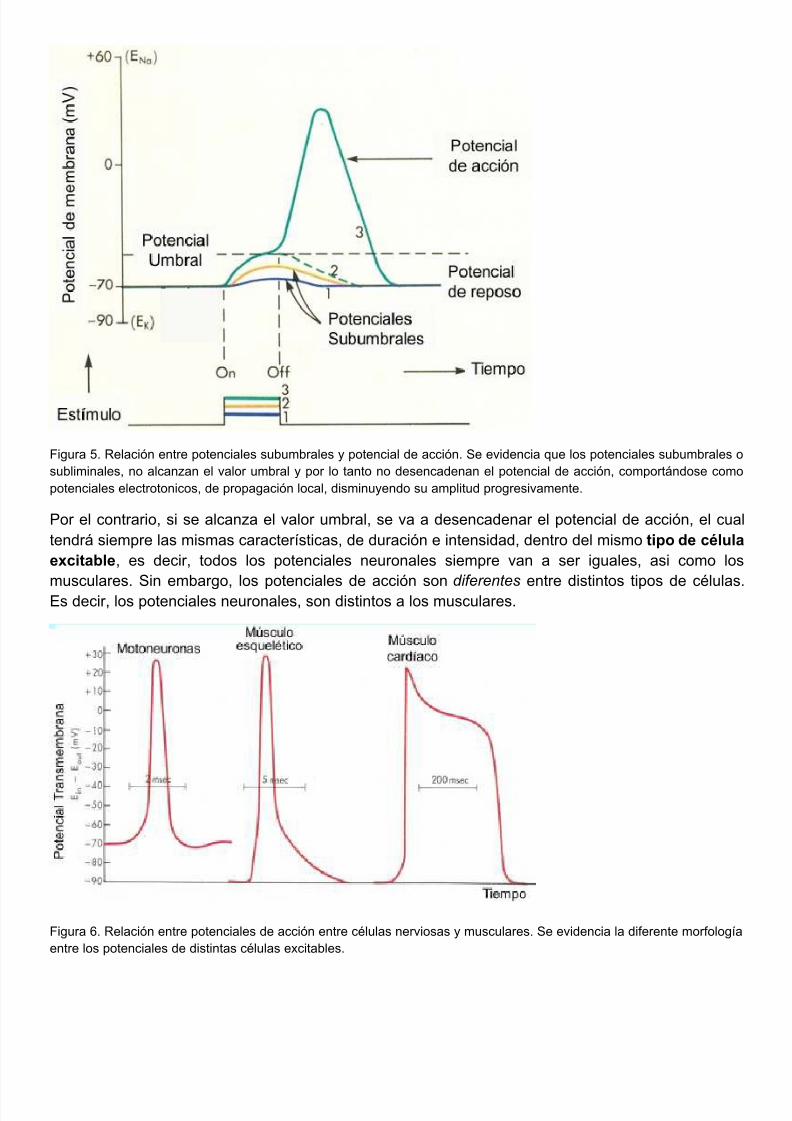
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 11/38
Figura 5. Relación entre potenciales subumbrales y potencial de acción. Se evidencia que los potenciales subumbralesubliminales, no alcanzan el valor umbral y por lo tanto no desencadenan el potencial de acción, comportándose copotenciales electrotonicos, de propagación local, disminuyendo su amplitud progresivamente.
Por el contrario, si se alcanza el valor umbral, se va a desencadenar el potencial de acción, el ctendrá siempre las mismas características, de duración e intensidad, dentro del mismo tipo de cél
excitable, es decir, todos los potenciales neuronales siempre van a ser iguales, asi como
musculares. Sin embargo, los potenciales de acción son diferentes entre distintos tipos de célulEs decir, los potenciales neuronales, son distintos a los musculares.
Figura 6. Relación entre potenciales de acción entre células nerviosas y musculares. Se evidencia la diferente morfoloentre los potenciales de distintas células excitables.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 12/38
Todo potencial de acción lleva a un breve periodo de menor excitabilidad celular, o perio
refractario, el cual puede dividirse en 2 fases. El periodo refractario absoluto, se produinmediatamente tras el disparo del potencial de acción; durante este periodo es imposible excitarcelular por más grande que sea la corriente de estimulación que se le aplique, ya que la totalidad los canales de Na+ voltaje-dependiente que generaron la fase de despolarización del potencial acción se encuentran en estado inactivo y por lo tanto no pueden ser excitados nuevamente haque vuelvan a su estado de reposo. Esta fase va seguida del periodo refractario relativo, durante
cual es posible desencadenar un potencial de acción, pero solo aplicando estímulos mayores que requeridos normalmente para alcanzar el umbral, ya que ya hay algunos canales de Na+ voltadependientes activos listos pero debido a que se encuentran activados los canales de K+ voltadependientes y el potencial de membrana esta hiperpolarizandose es necesario un mayor estimpara llegar al nivel umbral. (Ver figura 7).
En relación con la propagación del potencial de acción a lo largo del axón, ésta siguecaracterísticas importantes:
- Ocurre por la propagación de corrientes locales desde regiones activas hacia region
adyacentes inactivas.Inicialmente un segmento del axón es despolarizado al nivel umbral y dispara un potencial acción (región activa). Como resultado de la corriente interna de Na+ en el pico del potencde acción, la polaridad del potencial de membrana se revierte y el interior celular se vuepositivo, mientras que la región adyacente del axón se mantiene inactiva aun, con su intecelular negativo. Luego las cargas positivas en el interior de la región activa viajan hacia cargas negativas del interior de la región adyacente negativa. Este flujo causa que la regadyacente se despolarice al umbral, por lo que dispara un potencial de acción, nuevamentepolaridad de la membrana se revierte y el interior celular se vuelve positivo. Para e
momento, la región activa original ya ha sido repolarizada nuevamente a su potencial membrana en reposo y ha vuelto a su polaridad interna negativa, entrando en periorefractario, lo que impide que las cargas eléctricas pueden regresarse hacia esta regiimplicando la propagación unidireccional, ortodromica (del soma al terminal presinapticdel potencial de acción.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 13/38
Figura 7. Propagacion por corrientes locales del potencial de accion, demostrando que la propagacion ocurre despolarizacion del segmento adyacente a donde se disparo el potencial.
- Se propaga saltatoriamente.
La mayoría de los axones neuronales se encuentran revestidos por un aislante lipiddenominado mielina, este aislante incrementa la resistencia de la membrana y permite que cargas eléctricas viajen por el interior celular y no tiendan a escaparse por la membra
(recordemos que la corriente eléctrica sigue la ruta de menor resistencia, por lo queaumentamos la resistencia de la membrana, la corriente prefiere recorrer el interior celular cmenor resistencia al flujo), esto permite que la magnitud del potencial se mantenga constanSin embargo, si todo el axón estuviese revestido y no pudiese haber ninguna alteración potencial a lo largo del axón, la corriente tendería a disminuir por lo que se hace necesario“redisparo” del potencial cada cierto tramo de axón, por lo tanto, cada cierto intervalo enaxón se presentan partes desmielinizadas en donde los canales iónicos pueden generarcorriente iónica interna necesaria para propulsar el potencial de acción; estos lugadesmielinizados en el axón se conocen como nodos de Ranvier , y debido a esta forma conducción del potencial como saltando de un nodo a otro, esperando ser generanuevamente, se dice que los potenciales de acción se propagan saltatoriamente. Existen
factores que determinan la velocidad de conducción del potencial de acción: el diámede la fibra nerviosa y la mielinizacion del axón. A mayor diámetro, y mas cantidad de mielimayor velocidad. Las fibras nerviosas se pueden clasificar en base a estos parámetros, deslas fibras A alfa y beta (gruesas y mielinicas, de transmisión rápida), a las fibras A delta (fincon poca mielina) y C (amielinicas) de transmisión lenta. En las fibras amielinicas, no exitransmisión saltatoria, sino continua, ameritando un mayor gasto de energía, al tener qredisparar el potencial de acción a lo largo de cada tramo de la membrana plasmática.
ASPECTOS GENERALES DE LA TRANSMISION SINAPTICA.
Ya hemos visto la capacidad que tienen las células excitables para generar y transmitir una seeléctrica, ahora estudiaremos los mecanismos por medio de los cuales se transmiten estas señaentre estas células.
La región en donde se establece la comunicación entre 2 celulas excitables, bien sea neurona cneurona, musculo con musculo, con neurona con musculo, se denomina sinapsis.
Toda sinapsis, esta constituida por 3 elementos, una celula presinaptica, a partir de la cual se origla señal; una celula postsinaptica, que recibe dicha señal, y modifica asi, su potencial de membran
y un espacio variable entre ambas celulas, denominado hendidura sináptica.Una neurona media es capaz de formar 1000 conexiones sinapticas, y de recibir unas 10000 o has100000 (como las celulas de Purkinje del Cerebelo). Aunque todas conexiones son de un tespecializado, todas las neuronas usan 1 de los 2 tipos de transmisión sinaptica.
Existen sinapsis electricas y quimicas
El termino Sinapsis fue acuñado a principios del siglo XIX por Charles Sherrington, que lo deficomo el punto en el cual se comunican 2 neuronas, aunque ya antes esta sinapsis había sdescrita por Ramon y Cajal. Al principio se pensaba que en la sinapsis solo existían mecanismos

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 14/38
transmisión eléctrica, pero a mediados de los años veinte Otto Lewis demostró que la Acetil Col(Ach, un mensajero quimico) interviene en la transmisión de señales desde el nervio vago al coraz
Aquí comenzaron una serie de interrogantes, como puede una señal química acoplarse para produuna señal eléctrica en otra celula?...
Surgieron 2 escuelas de pensamiento, una fisiológica y una farmacológica, la escuela fisiologica (cJohn Eccles a la cabeza) defendía la teoría de que en la sinapsis solo intervenían mecanismos transmisión electricos (la señal se transmitia directamente de una celula a otra); mientras queescuela farmacología defendia que el potencial de acción inducía a la liberación de un mediaquimico en la celula presinaptica, el cual inducía a la generación de una señal electrica en la celpost-sináptica.
Con el tiempo se demostro que ambos mecanismos existen en los organismos vivos, con predominio de las sinapsis quimicas sobre las electricas.
En las sinapsis electricas, las células se encuentran unidas por medio de uniones intercelula
comunicantes (gap junction), las cuales les confieren una continuidad citoplasmica que permite
transmisión directa, casi instantanea de la señal electrica generada en la celula presinaptica.
En las sinapsis quimicas, el potencial de accion generado en la celula presinaptica induceliberación de mensajeros quimicos (neurotransmisores), los cuales inducen la generacion de potencial postsinaptico en la celula efectora.
Figura 8. Flujo de cargas electricas en las sinapsis electrica (A), evidenciando propagacion de las cargas de mandirecta de uina celula a otra, y en una sinapsis quimica (B), en donde, al no haber, conexión mecanica entre las celuel flujo electrico no es continuo entre las celulas
Las Sinapsis electricas generan una transmisión casi instantanea de la señal.
La corriente que despolariza la celula postsinaptica es generada por los canales ionicos voltadependientes de la celula presinaptica, los cuales no solo tienen que generar una despolarizacionsuficientemente grande como para producir un potencial de accion, sino que tienen que generar ucorriente ionica grande como para despolarizar hasta el umbral a la celula postsinaptica.
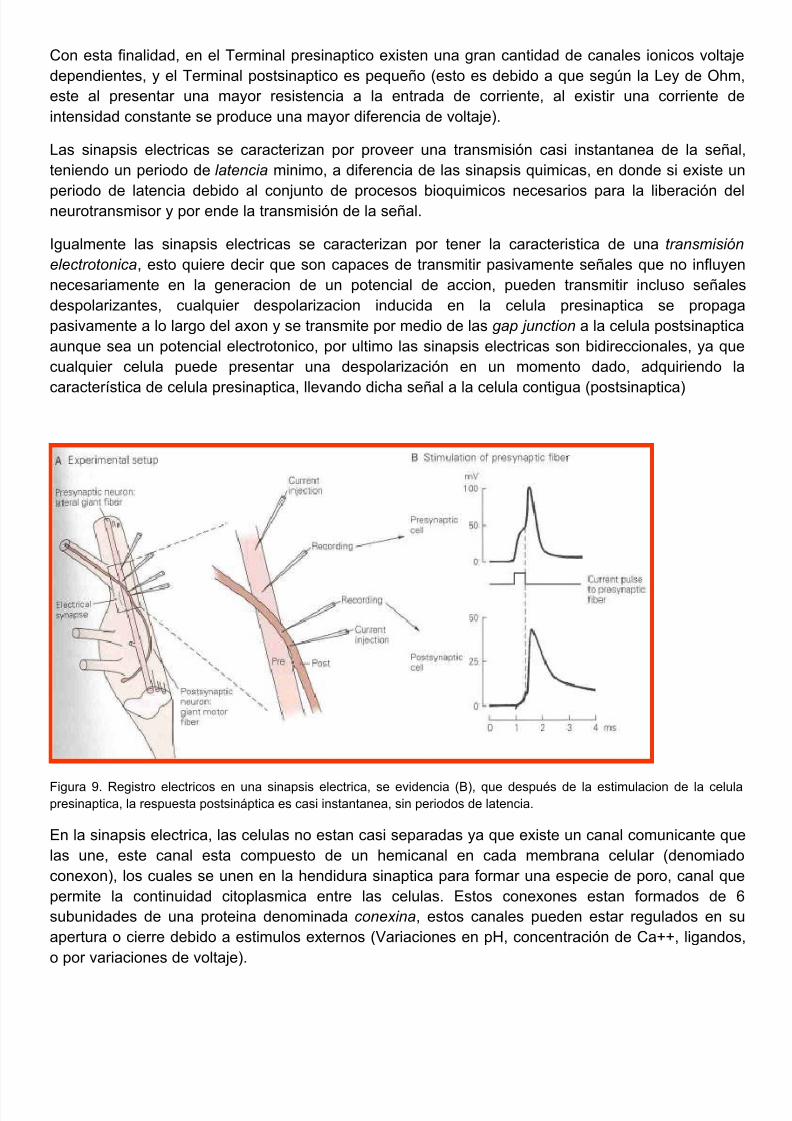
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 15/38
Con esta finalidad, en el Terminal presinaptico existen una gran cantidad de canales ionicos voltdependientes, y el Terminal postsinaptico es pequeño (esto es debido a que según la Ley de Oheste al presentar una mayor resistencia a la entrada de corriente, al existir una corriente intensidad constante se produce una mayor diferencia de voltaje).
Las sinapsis electricas se caracterizan por proveer una transmisión casi instantanea de la señteniendo un periodo de latencia minimo, a diferencia de las sinapsis quimicas, en donde si existe
periodo de latencia debido al conjunto de procesos bioquimicos necesarios para la liberación neurotransmisor y por ende la transmisión de la señal.
Igualmente las sinapsis electricas se caracterizan por tener la caracteristica de una transmis
electrotonica, esto quiere decir que son capaces de transmitir pasivamente señales que no influynecesariamente en la generacion de un potencial de accion, pueden transmitir incluso señadespolarizantes, cualquier despolarizacion inducida en la celula presinaptica se propapasivamente a lo largo del axon y se transmite por medio de las gap junction a la celula postsinaptaunque sea un potencial electrotonico, por ultimo las sinapsis electricas son bidireccionales, ya qcualquier celula puede presentar una despolarización en un momento dado, adquiriendo
característica de celula presinaptica, llevando dicha señal a la celula contigua (postsinaptica)
Figura 9. Registro electricos en una sinapsis electrica, se evidencia (B), que después de la estimulacion de la cepresinaptica, la respuesta postsináptica es casi instantanea, sin periodos de latencia.
En la sinapsis electrica, las celulas no estan casi separadas ya que existe un canal comunicante qlas une, este canal esta compuesto de un hemicanal en cada membrana celular (denomiaconexon), los cuales se unen en la hendidura sinaptica para formar una especie de poro, canal qpermite la continuidad citoplasmica entre las celulas. Estos conexones estan formados desubunidades de una proteina denominada conexina, estos canales pueden estar regulados en apertura o cierre debido a estimulos externos (Variaciones en pH, concentración de Ca++, ligando por variaciones de voltaje).

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 16/38
La sinapsis electrica nos permite una gran velocidad en la transmisión de las señales, lo cual es en respuestas de huida, etc.., igualmente conecta a las celulas permitiendo que se comporten de umanera sincronica, es decir que todas se comporten siguiendo el concepto todo o nada, permite qformen un sincitio, y que al generarse un potencial de accion en una de las celulas, todas puedresponder a ese potencial de accion de una manera rapida y en conjunto.
La sinapsis quimica permite una amplificación de la señal.
En la sinapsis quimica, las celulas si se encuentran separadas por una hendidura sinápt
significativa, cuando se genera un potencial de accion en la celula presinaptica este se transmhasta el Terminal presinaptico donde se estimula la apertura de canales de Ca++ voltdependientes, los cuales generan una corriente ionica de ca++ hacia el interior celular, este Capermite la fusion de las vesículas que contienen los neurotransmisores (mediadores químicos) conmembrana plasmatica y que sea volcado su contenido hacia la hendidura sinaptica mediaexocitosis. El aumento en las concentraciones de calcio en el terminal, su unión con la calmoduliactiva una quinasa dependiente de calcio y calmodulina que media la fosforilación de las proteínsinapsina y sinaptobrevina que favorecen esta exocitosis. Estos neurotransmisores interactu
después con receptores ubicados en la Celula Post Sinaptica y estos generan un cambio enpotencial de membrana local, llamado potencial Post Sinaptico (que puede ser excitatorio “PPSEinhibitorio “PPSI”)
En este mecanismo de transmisión sinaptica existe un periodo de latencia (retardo sinaptico) debal conjunto de procesos necesarios para la transmisión de la señal (la liberación del NT), embargo este permite una amplificación importante de la señal. Son necesarias pocas moléculas NT para activar a un receptor de la celula Post Sinaptica, y durante cada potencial de acción sliberadas miles de moléculas de NT, lo cual permite la activacion de numerosos receptores, cuales son capaces de despolarizar la membrana de la celula Post Sinaptica hasta el umbral pa
generar un Potencial de Accion, con lo cual un pequeño Terminal presinaptico es capaz despolarizar toda una gran celula post sinaptica.
Figura 10. Registro electrico en la celula presinaptica y postsináptica en una sinapsis quimica, se evidencia el retrasinaptico (A), tiempo presente entre la estimulacion presinaptica y la respuesta postsinaptica
El efecto del potencial sinaptico, ya sea inhibitorio o excitatorio, depende tanto del tipo neurotransmisor que sea liberado, asi como de los tipos de receptores sobre los que actue en

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 17/38
celula postsinaptica, ya que un mismo neurotransmisor puede generar respuestas excitatoriasInhibitorias dependiendo de con cual receptor de membrana se una.
Sinapsis Electrica Sinapsis Química
La hendidura sináptica es muy estrecha Presenta una hendidura sinapticaamplia
Las celulas se encuentran acopladaselectromecánicamente entre si, graciasa las gap junctions
Las celulas se encuentran separadasuna de otra, sin contacto fisico
La despolarizacion de la celula post-sinaptica, es inducida por la corrienteionica de la celula presinaptica
La despolarizacion o hiperpolarizacionpost-sinaptica es mediada por laliberacion de neurotransmisores desdeel terminal presinaptico
Es bidireccional, siempre excitatoria Es unidireccional, puede ser excitatoria
o inhibitoria dependiendo delneurotransmisor
No existe retardo sinaptico Hay retardo sinaptico
Figura 11. Comparacion entre las caracteristicas de una sinapsis electrica, y una sinapsis quimica
PROPAGACION DE SEÑALES EN LA SINAPSIS NERVIO-MUSCULO
La sinapsis neuromuscular es un lugar ideal para estudiar la transmisión de señales debido a que
relativamente sencilla y accesible para su experimentación. Toda celula de musculo estriado encuentra inervada por ramificaciones nerviosas, las cuales en el musculo esquelético, recibennombre de motoneuronas.
El axon de la motoneurona inerva al músculo en una region especializada de la membrana muscudenominada placa Terminal , aquí el axon pierde su vaina mielinica y se ramifica en un conjunto prolongaciones, cada una de las cuales termina en un boton sinaptico, estos son los que contientoda la maquinaria que interviene en la liberación del NT, aquí se encuentran una gran cantidad vesiculas sinápticas, y que se agrupan en una region del boton sinaptico denominada zona acticada una de estas zonas activas del Terminal sinaptico se encuentran colocadas sobre u
invaginacion de la membrana muscular denominada pliegues de union en donde se encuentran receptores para el neurotransmisor y una gran cantidad de canales de sodio voltaje dependientes.
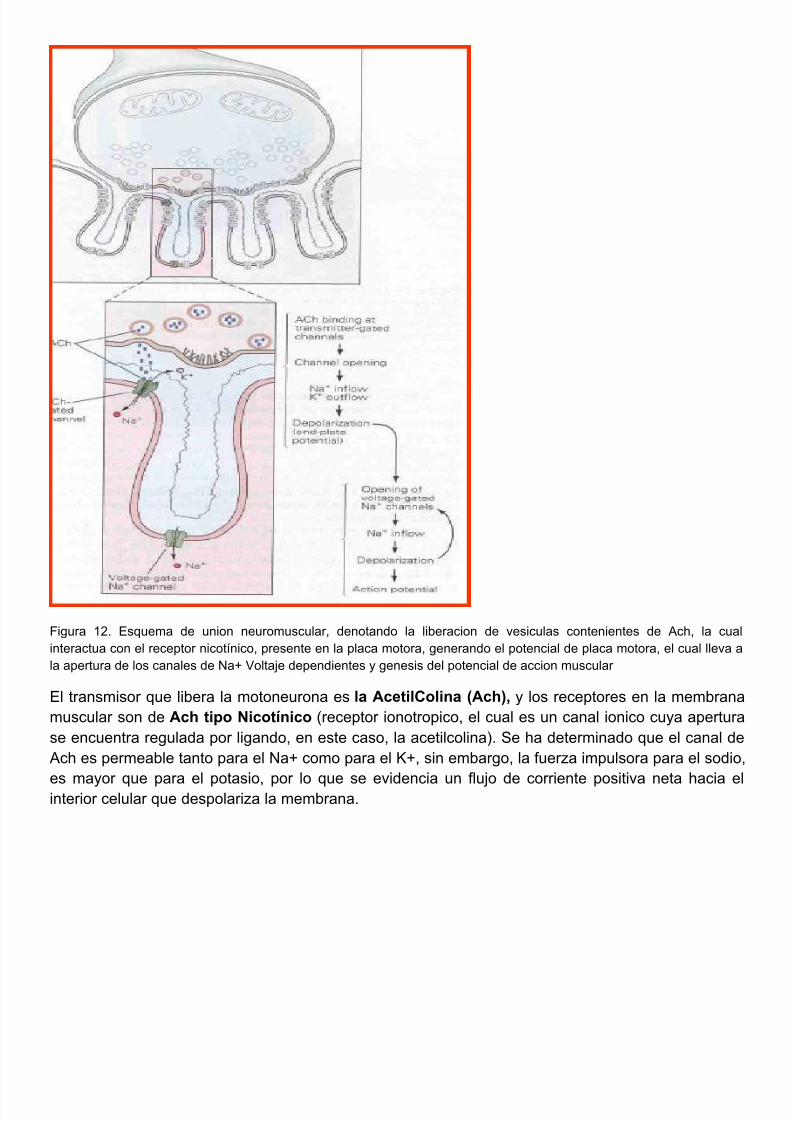
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 18/38
Figura 12. Esquema de union neuromuscular, denotando la liberacion de vesiculas contenientes de Ach, la cinteractua con el receptor nicotínico, presente en la placa motora, generando el potencial de placa motora, el cual llevla apertura de los canales de Na+ Voltaje dependientes y genesis del potencial de accion muscular
El transmisor que libera la motoneurona es la AcetilColina (Ach), y los receptores en la membramuscular son de Ach tipo Nicotínico (receptor ionotropico, el cual es un canal ionico cuya apertse encuentra regulada por ligando, en este caso, la acetilcolina). Se ha determinado que el canal Ach es permeable tanto para el Na+ como para el K+, sin embargo, la fuerza impulsora para el sod
es mayor que para el potasio, por lo que se evidencia un flujo de corriente positiva neta haciainterior celular que despolariza la membrana.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 19/38
Figura 13. Conformacion del canal nicotínico de Ach
Las membranas pre y póstsinapticas estan separadas por un espacio de alrededor 100nm conoccomo hendidura sinaptica, en este espacio se encuentra, hacia el sarcolema, una membrana baconstituida por colageno y diversas proteinas, en esta membrana basal podemos encontrar uenzima denominada acetilcolinesterasa , la cual se encarga de degradar rapidamente la AchAcetato y Colina, y asi, finalizar la señal nerviosa.
La motoneurona excita al músculo mediante la apertura de canales ionicos en la placa Terminal .
Tras la liberación de Ach por el Terminal presinaptico, la membrana de la placa Terminal
despolariza rápidamente (por la entrada masiva de sodio), este Potencial Post Sinaptico Excitatorecibe el nombre de Potencial de Placa Terminal, o Placa motora.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 20/38
Figura 14. A. Comparacion entre el potencial de placa motora, y el potencial de accion muscular, denotando que siemel potencial de placa motora, es de intensidad suficiente como para superar el umbral y provocar el potencial de accmuscular. B. Como se modifica el Potencial de placa motora, ante la presencia de curare (d-tubo curarina), la cbloquea competitivamente el receptor nicotínico, disminuyendo la amplitud del PPM.
Este PPSE se produce gracias a un flujo neto de corriente ionica positiva hacia el interior demembrana gracias a la apertura de canales ionicos dependientes de Ach como ligando. El PPSEincrementa rapidamente gracias a la brusca liberación de Ach a la hendidura sinaptica, sin embarg
no todo este Ach interactua con los receptores, la mayoria es rapidamente catabolizado gracias aAcetilcolinesterasa y el resto difunde fuera de la hendidura sinaptica; la disminución del PPSE ocumas lentamente debido a que para esta es necesario el cierre aleatorio de los canales dependiende ligando, este proceso tiende a tardar unos milisegundos.
Existen 4 factores determinantes en el flujo de corriente para el PPSE en la Placa Terminal: 1)numero total de canales de Ach, 2) La probabilidad de que un canal este abierto (que viene relacion con la cantidad de NT liberado), 3) La conductancia de cada canal abierto, 4) La fuerza arrastre que actua sobre los iones.
Este potencial suele tener una amplitud bastante grande (de aprox. 70 mV), lo cual es suficientemente grande como para abrir todos los canales de Na++ voltaje dependientes en la fibmuscular, con lo que este Potencial de Placa Terminal se convierte ahora en un Potencial de Accque se propaga activamente a lo largo de la fibra muscular, a diferencia de las neuronas, en donun PPSE suele tener solo una amplitud de 1mV con lo cual es necesario la convergencia de varPPSE para la generacion de un Potencial de Accion. Este potencial de acción muscular seráresponsable de desencadenar la contracción muscular, mediante el denominado, acoplamieexcitación – contracción.
MUSCULATURA ESTRIADA Y LISA.
El movimiento en los seres vivos es ejecutado por organos llamados músculos. Existen 2 tipos musculos: El músculo estriado y el músculo liso. El músculo estriado a su vez puede dividirse músculo esqueletico y músculo cardiaco.
El músculo esqueletico es el que forma la musculatura somatica, tiene estriaciones transversaevidentes, no es capaz de contraerse sin una estimulacion nerviosa, y sus fibras se comporcomo unidades anatomicas y funcionales independientes.
El músculo cardiaco tiene igualmente estriaciones transversales, pero sus fibras se comportan coun sincitio, por la presencia de sinapsis eléctricas entre sus celulas, y gracias a la presencia
celulas marcapasos en el miocardio, se contrae de forma ritmica sin estimulacion nerviosa.
El músculo liso carece de estriaciones transversales. El musculo liso unitario que forma las viscehuecas se comporta como un sincicio y contiene marcapasos para contracciones ritmicas.
Al igual que las celulas nerviosas, las celulas musculares pueden excitarse en respuesta a estimuquimicos, electricos o mecanicos, y generar un potencial de accion que se propague a lo largo defibra muscular. La diferencia basica entre ambas celulas radica en que el potencial de accion enfibra muscular desencadena un mecanismo contráctil, el cual esta mediado por la interaccion proteinas contráctiles, como lo son la actina y la miosina.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 21/38
El músculo esqueletico es un órgano que puede equipararse con un motor flexible y elastico que une a palancas rigidas (los huesos), y que al contraerse imprime un giro sobre esta palanca a travde las articulaciones, que actuan como puntos de apoyo.
Este conjunto de elementos, los músculos, los huesos y las articulaciones forman el aparalocomotor del organismo, que esta bajo control de los sistemas nervioso y endocrino.
El músculo esquelético esta formado por un conjunto de fibras musculares, cada una de estas fibse corresponde con una célula muscular, cilíndrica, alargada y rodeada de una membrana celular sarcolema), este sarcolema se invagina de trecho en trecho en la célula para formar los túbulos T.
El citoplasma de las células musculares contiene una gran cantidad de enzimas, lípidos, partícude glucogeno, mitocondrias y miofibrillas. Las miofibrillas constituyen el mecanismo contráctil músculo, están formadas por un conjunto de filamentos, que pueden separarse en: Filamendelgados, Filamentos gruesos y Filamentos conectores.
Las estriaciones características del músculo esquelético se deben a los diferentes índices refracción que presentan las diferentes partes de la fibra muscular. En la miofibrilla, se evidencia u
banda I (isotrópica) que se corresponde con la presencia de los filamentos finos, las bandas I acoplan entre sí en la denominada línea Z; se evidencia tambien una banda A (anisotropica) oscuque se corresponde con la presencia de filamentos gruesos, en la banda A se encuentra una regcentral denominada zona H, donde se encuentran exclusivamente los filamentos gruesos, enmedio de esta zona H se encuentra la linea M, que sirve de anclaje a los filamentos de miosina.area entre 2 lineas Z se denomina Sarcomera, y es la unidad funcional del músculo estriado, es aqdonde se interdigitan e interaccionan los filamentos finos y gruesos que permiten el mecanismo contracción muscular.
Figura 15. Conformacion del sarcomero muscular
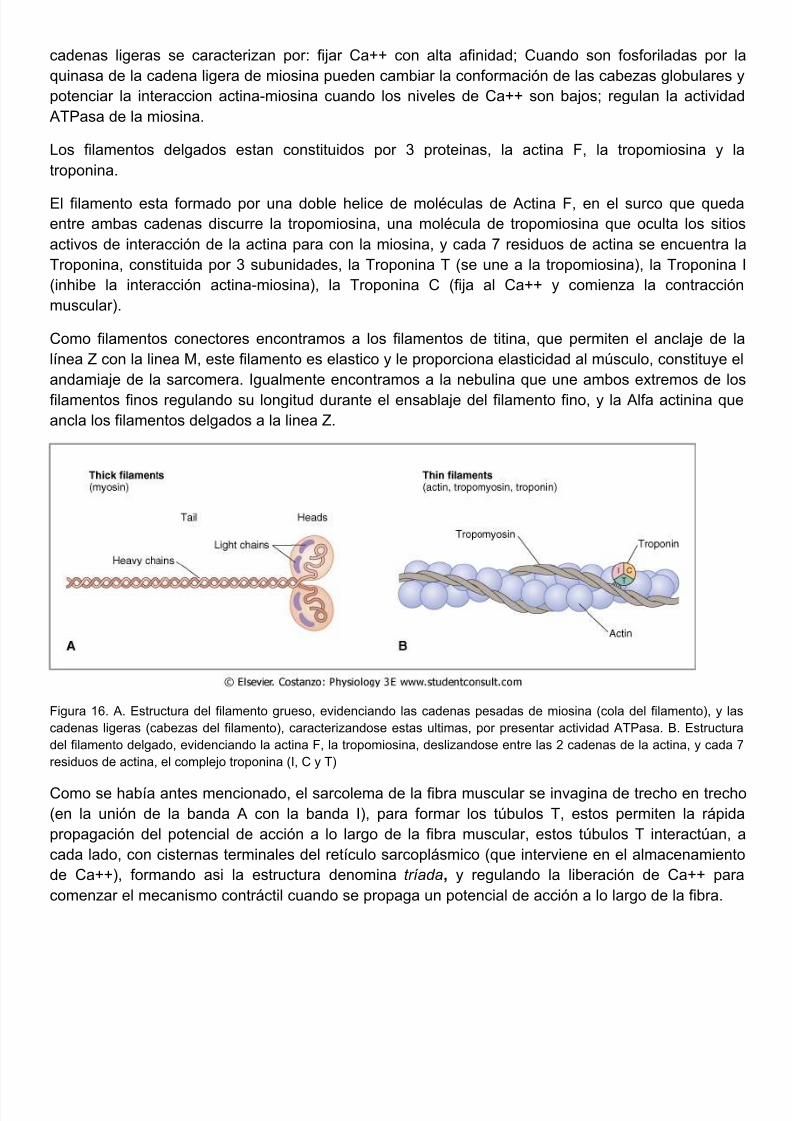
Los filamentos gruesos estan constituidos por moléculas de Miosina II, esta molécula esta constitupor 2 cabezas globulares (cadenas pesadas) con alta afinidad para la actina y actividad ATPasauna cola, constituida por 2 cadenas ligeras, una cadena reguladora y una cadena esencial, es
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 22/38
cadenas ligeras se caracterizan por: fijar Ca++ con alta afinidad; Cuando son fosforiladas porquinasa de la cadena ligera de miosina pueden cambiar la conformación de las cabezas globularepotenciar la interaccion actina-miosina cuando los niveles de Ca++ son bajos; regulan la actividATPasa de la miosina.
Los filamentos delgados estan constituidos por 3 proteinas, la actina F, la tropomiosina ytroponina.
El filamento esta formado por una doble helice de moléculas de Actina F, en el surco que queentre ambas cadenas discurre la tropomiosina, una molécula de tropomiosina que oculta los sitactivos de interacción de la actina para con la miosina, y cada 7 residuos de actina se encuentraTroponina, constituida por 3 subunidades, la Troponina T (se une a la tropomiosina), la Troponin(inhibe la interacción actina-miosina), la Troponina C (fija al Ca++ y comienza la contraccmuscular).
Como filamentos conectores encontramos a los filamentos de titina, que permiten el anclaje delínea Z con la linea M, este filamento es elastico y le proporciona elasticidad al músculo, constituyeandamiaje de la sarcomera. Igualmente encontramos a la nebulina que une ambos extremos de filamentos finos regulando su longitud durante el ensablaje del filamento fino, y la Alfa actinina qancla los filamentos delgados a la linea Z.
Figura 16. A. Estructura del filamento grueso, evidenciando las cadenas pesadas de miosina (cola del filamento), y cadenas ligeras (cabezas del filamento), caracterizandose estas ultimas, por presentar actividad ATPasa. B. Estructdel filamento delgado, evidenciando la actina F, la tropomiosina, deslizandose entre las 2 cadenas de la actina, y cadresiduos de actina, el complejo troponina (I, C y T)
Como se había antes mencionado, el sarcolema de la fibra muscular se invagina de trecho en trec(en la unión de la banda A con la banda I), para formar los túbulos T, estos permiten la ráppropagación del potencial de acción a lo largo de la fibra muscular, estos túbulos T interactúancada lado, con cisternas terminales del retículo sarcoplásmico (que interviene en el almacenamiende Ca++), formando asi la estructura denomina tríada, y regulando la liberación de Ca++ pcomenzar el mecanismo contráctil cuando se propaga un potencial de acción a lo largo de la fibra.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 23/38
Figura 17. Triada del musculo esqueletico, se denota la invaginacion del sarcolema, constituyendo los túbulosasociados a ambos lados con las cisteras del reticulo sarcoplasmico, sitio este, en donde se desarrolla el acoplamieexcitación-contraccion.
Todo el proceso de la contracción muscular suele desarrollarse en respuesta a la excitación nervio
de la placa motora, y la generación del potencial de acción muscular.El proceso por el cual la despolarización del músculo inicia la contracción se llama acoplamie
excitación-contracción.
El potencial de acción se transmite a lo largo de toda la fibra por el sistema T; se inicia la liberacde Ca++ de las cisternas terminales, este se une con la troponina C y comienza el proceso contracción, por lo que este acoplamiento implica una interaccion entre las triadas (tubulo T ycisternas del reticulo sarcoplasmico) y los miofilamentos.
La despolarización de la membrana del túbulo T activa al retículo sarcoplásmico mediante
receptores para dihidropirina, estos actúan como canales iónicos de Ca++ voltaje dependientesel músculo cardiaco, mientras que en el músculo esqueletico, actúan mas como sensores
voltaje. Cuando se propaga el potencial de acción a lo largo de la fibra, se activan estos receptoreeste actúa como un “disparador” para la liberación de Ca++ del RS. Este receptor cuando se actiinduce a la activación de los receptores de Rianodina, (ya que esta acoplado electromecánicamecon estos receptores) ubicados en la membrana del RS, y así se permite la liberación de Ca++ hael citosol celular. El aumento de las concentraciones de calcio intracelular es el desencadenante la contracción muscular, permitiendo la interacción acto miosina. Mientras mas calcio haya enfibra, mas interacción de miofilamentos habrá, y mayor tensión se generara.
La contracción consiste en el acortamiento de la longitud del músculo, esto se debe a udisminución en la longitud sarcomerica. Se han expuesto diversas teorías para explicar efenómeno, entre las cuales, la teoría de los puentes cruzados es la mas importante.
Teoría de los filamentos deslizantes (Mecanismo de los Puentes cruzados).
Durante la contracción o el estiramiento muscular la banda A del sarcomero no modifica su longita diferencia de la banda I que aumenta su longitud (en el estiramiento) o la disminuye (en contracción), este fenómeno se explica mediante movimientos deslizantes de los filamendelgados sobre los filamentos gruesos.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 24/38
Figura 18. Acortamiento de las bandas I durante la contracción muscular, se evidencia que la banda A permanconstante, mientras lo que se evidencia es un acortamiento de las bandas I (constituidas por filamentos finos), sieneste una evidencia de que en la contracción muscular lo que realmente hay es un deslizamiento del filamento fino soel filamento grueso.
Una vez establecido este movimiento deslizante, es importante establecer cual es el mecanismo qimpulsa este movimiento, el cual depende de la interacción de los puentes cruzados que proyectan desde el filamento grueso hacia el filamento delgado.
La teoria de los puentes cruzados implica la union intermitente de la actina de los filamendelgados con los puentes cruzados de miosina de los filamentos gruesos.
En condiciones estrucutrales normales del sarcomero, la interaccion actina-miosina se encueninhibida por la presencia de la troponina I, al liberarse Ca++ al medio intracelular este se une atroponina C, esto trae como consecuencia un cambio conformacional que inactiva a la troponina la tropomiosina se desplaza lateralmente, permitiendo la exposición de los sitios activos de la actpara con la miosina.
Inicialmente el filamento de miosina se encuentra unido a la actina, la cabeza de la miosina encuentra en una orientación de 45 grados (después del ultimo ciclo de puentes cruzados), se uneATP lo que causa una disminución de la afinidad de la actina por la miosina y permite la disociacde los filamentos asi como la reorientación de la cabeza de miosina a su estado de reposo en grados. Luego el ATP es hidrolizado rápidamente gracias a la actividad ATPasa de las cabezas miosina para formar el complejo Miosina-ADP Pi, este complejo se caracteriza por tener una elevaenergía libre y una alta afinidad para con la actina, después de la unión con la actina, se liberaADP y Pi, esto trae como consecuencia un cambio conformacional en la cabeza de miosina, la crota para pasar de una orientación de 90 grados a una de 45 grados con respecto al filamedelgado, este cambio conformacional origina una fuerza que impulsa al filamento delgado hacia

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 25/38
filamento grueso, esta fuerza se transmite al citoesqueleto de la célula gracias a la proteina distrof
y produce el acortamiento general de la misma; Luego a este complejo actina-miosina se le une umolécula de ATP, el complejo resultante (miosina-ATP) tiene una baja afinidad por la actina y etrae como consecuencia la disociación del complejo, posteriormente este ATP es hidrolizanuevamente y comienza el ciclo. Ver figura 19.
Figura 19. Ciclo de puentes cruzados.
Este ciclo continua realizándose hasta que se acabe el ATP o los mecanismos reguladores entren acción, disminuyendo las concentraciones de Ca++ intracelular, lo cual retrae a los sitios activos la actina para con la miosina y finaliza la contracción muscular.
Una vez, termina la señal nerviosa, la concentración de calcio en el citoplasma disminuye p
diversos mecanismos: 1) el retículo sarcoplásmico empieza a reacumularlo mediante el transpoactivo mediado por ATP (por la proteina transportadora SERCA) hacia las porciones longitudinadel retículo, y de aquí difunden hacia las cisternas terminales, donde es capturado por la protecalsecuestrina y almacenado hasta que es liberado por el siguiente potencial de acción. 2) El cales exportado hacia el exterior celular, por una bomba de calcio de la membrana plasmática, llamaCa++-ATPasa de la membrana o PMCA, 3) puede salir al exterior celular en un intercambio por Nagracias a la actividad del antiporter Na+/Ca++ de la membrana. Una vez que la concentración Ca++ fuera del retículo se redujo lo suficiente, cesa la interacción química entre la miosina yactina, y el músculo se relaja. Nótese que el ATP proporciona la energía tanto para la contracccomo para la relajación. El deslizamiento de regreso de los filamentos delgados a su posición
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 26/38
relajación es dependiente de la energía elástica almacenada en las estructuras sarcoméricasextrasarcoméricas.
Cuando las fibras musculares agotan por completo el ATP y la fosforilcreatina desarrollan un estade rigidez llamado rigor . Cuando esto sucede después de la muerte, la situación se denomina ri
mortis. En el rigor casi todas las cabezas de miosina se unen con la actina, pero de manera anormfija y resistente.
Si los mecanismos de recaptación de Ca++ se encuentran inhibidos, o bien se estimula la apertudirecta de los receptores de rianodina, en ausencia de estimulación nerviosa (como la producida pla cafeína), se puede producir una contracción sostenida, conocida como contractura.
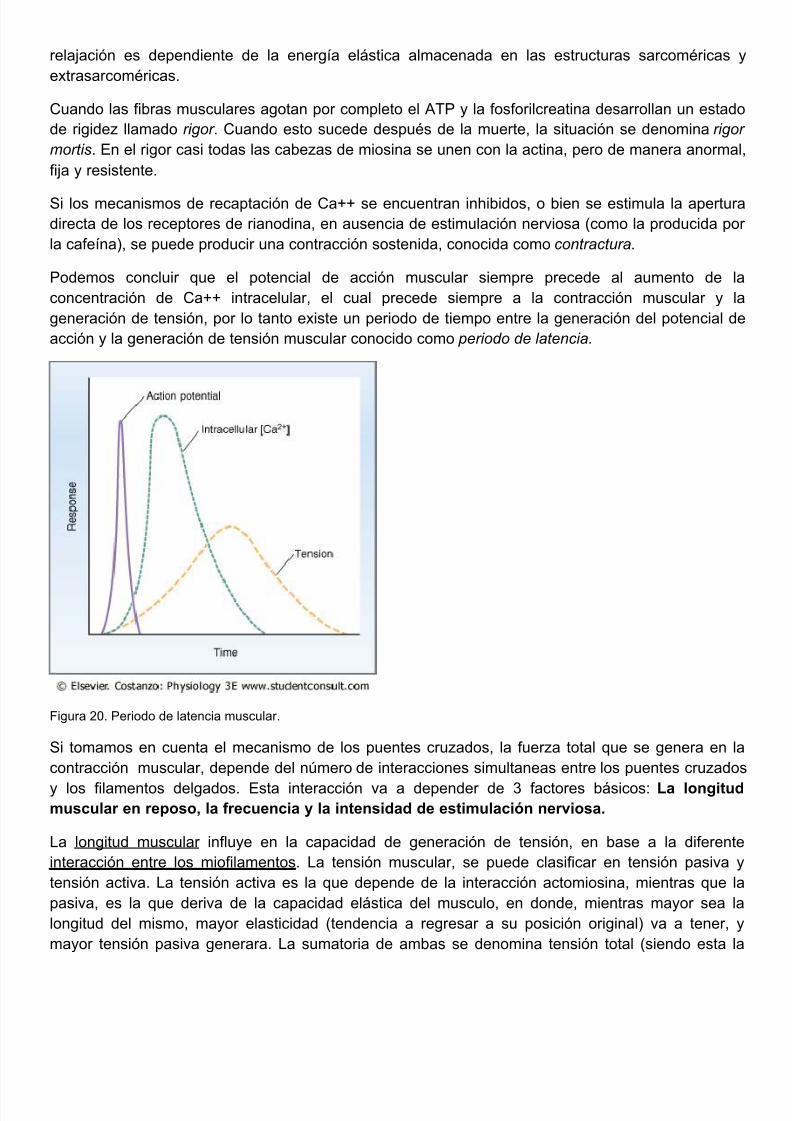
Podemos concluir que el potencial de acción muscular siempre precede al aumento de concentración de Ca++ intracelular, el cual precede siempre a la contracción muscular y generación de tensión, por lo tanto existe un periodo de tiempo entre la generación del potencial acción y la generación de tensión muscular conocido como periodo de latencia.
Figura 20. Periodo de latencia muscular.
Si tomamos en cuenta el mecanismo de los puentes cruzados, la fuerza total que se genera encontracción muscular, depende del número de interacciones simultaneas entre los puentes cruzad
y los filamentos delgados. Esta interacción va a depender de 3 factores básicos: La longit
muscular en reposo, la frecuencia y la intensidad de estimulación nerviosa.
La longitud muscular influye en la capacidad de generación de tensión, en base a la difereinteracción entre los miofilamentos. La tensión muscular, se puede clasificar en tensión pasivatensión activa. La tensión activa es la que depende de la interacción actomiosina, mientras quepasiva, es la que deriva de la capacidad elástica del musculo, en donde, mientras mayor sealongitud del mismo, mayor elasticidad (tendencia a regresar a su posición original) va a tenermayor tensión pasiva generara. La sumatoria de ambas se denomina tensión total (siendo esta
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 27/38
única posible de registrar con un transductor de tensión). Si se modifica la longitud del músculo, sea por estiramiento o acortamiento, este numero de interacciones se ve modificado, si sobreestira el músculo, la ínter digitación entre los filamentos va a disminuir progresivamente hadesaparecer por completo (al igual que la tensión activa generada, sin embargo va a aumentartensión pasiva generada ya que aumenta la resistencia elástica del musculo que tiende a volverlosu estado normal), si se acorta la longitud del músculo, los filamentos van a distorsionarinteracción en los puentes cruzados; a medida que modificamos la longitud del musculo y
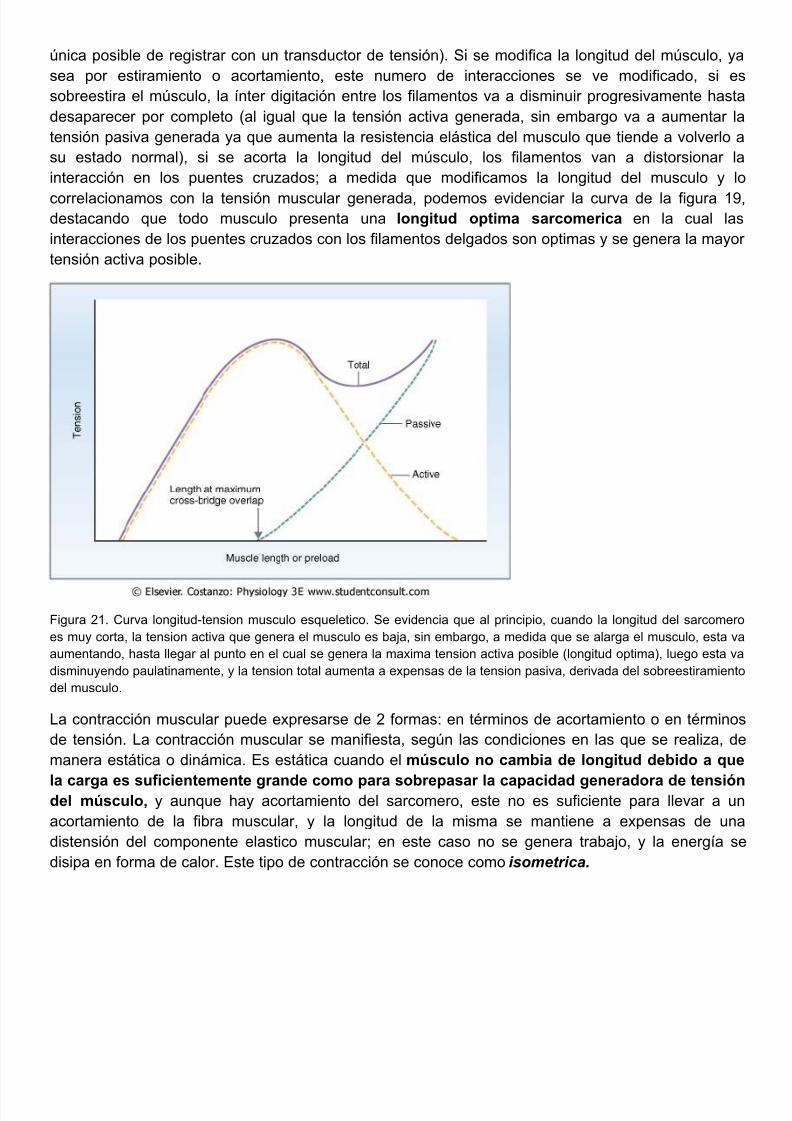
correlacionamos con la tensión muscular generada, podemos evidenciar la curva de la figura destacando que todo musculo presenta una longitud optima sarcomerica en la cual interacciones de los puentes cruzados con los filamentos delgados son optimas y se genera la maytensión activa posible.
Figura 21. Curva longitud-tension musculo esqueletico. Se evidencia que al principio, cuando la longitud del sarcomes muy corta, la tension activa que genera el musculo es baja, sin embargo, a medida que se alarga el musculo, estaaumentando, hasta llegar al punto en el cual se genera la maxima tension activa posible (longitud optima), luego estadisminuyendo paulatinamente, y la tension total aumenta a expensas de la tension pasiva, derivada del sobreestiramiedel musculo.
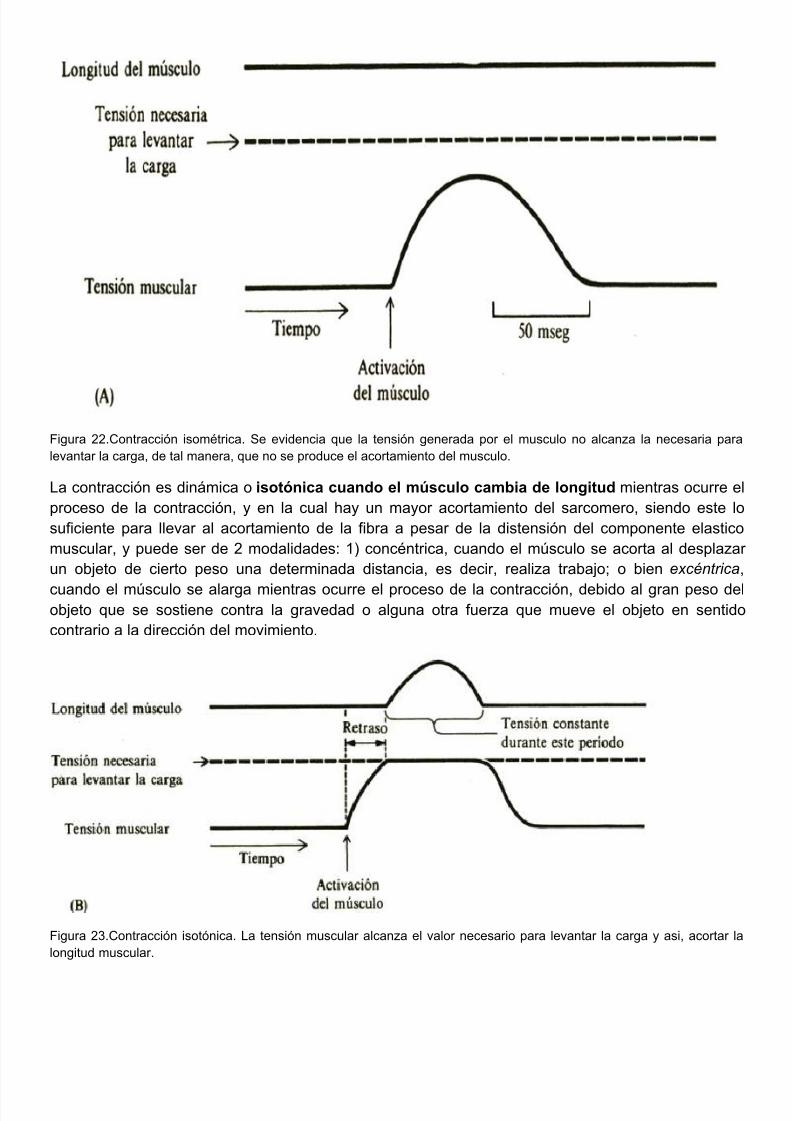
La contracción muscular puede expresarse de 2 formas: en términos de acortamiento o en términde tensión. La contracción muscular se manifiesta, según las condiciones en las que se realiza, manera estática o dinámica. Es estática cuando el músculo no cambia de longitud debido a q
la carga es suficientemente grande como para sobrepasar la capacidad generadora de tensidel músculo, y aunque hay acortamiento del sarcomero, este no es suficiente para llevar a acortamiento de la fibra muscular, y la longitud de la misma se mantiene a expensas de udistensión del componente elastico muscular; en este caso no se genera trabajo, y la energía disipa en forma de calor. Este tipo de contracción se conoce como isometrica.
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 28/38
Figura 22.Contracción isométrica. Se evidencia que la tensión generada por el musculo no alcanza la necesaria plevantar la carga, de tal manera, que no se produce el acortamiento del musculo.
La contracción es dinámica o isotónica cuando el músculo cambia de longitud mientras ocurreproceso de la contracción, y en la cual hay un mayor acortamiento del sarcomero, siendo estesuficiente para llevar al acortamiento de la fibra a pesar de la distensión del componente elastmuscular, y puede ser de 2 modalidades: 1) concéntrica, cuando el músculo se acorta al desplaun objeto de cierto peso una determinada distancia, es decir, realiza trabajo; o bien excéntri
cuando el músculo se alarga mientras ocurre el proceso de la contracción, debido al gran peso objeto que se sostiene contra la gravedad o alguna otra fuerza que mueve el objeto en sentcontrario a la dirección del movimiento.
Figura 23.Contracción isotónica. La tensión muscular alcanza el valor necesario para levantar la carga y asi, acortalongitud muscular.
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 29/38
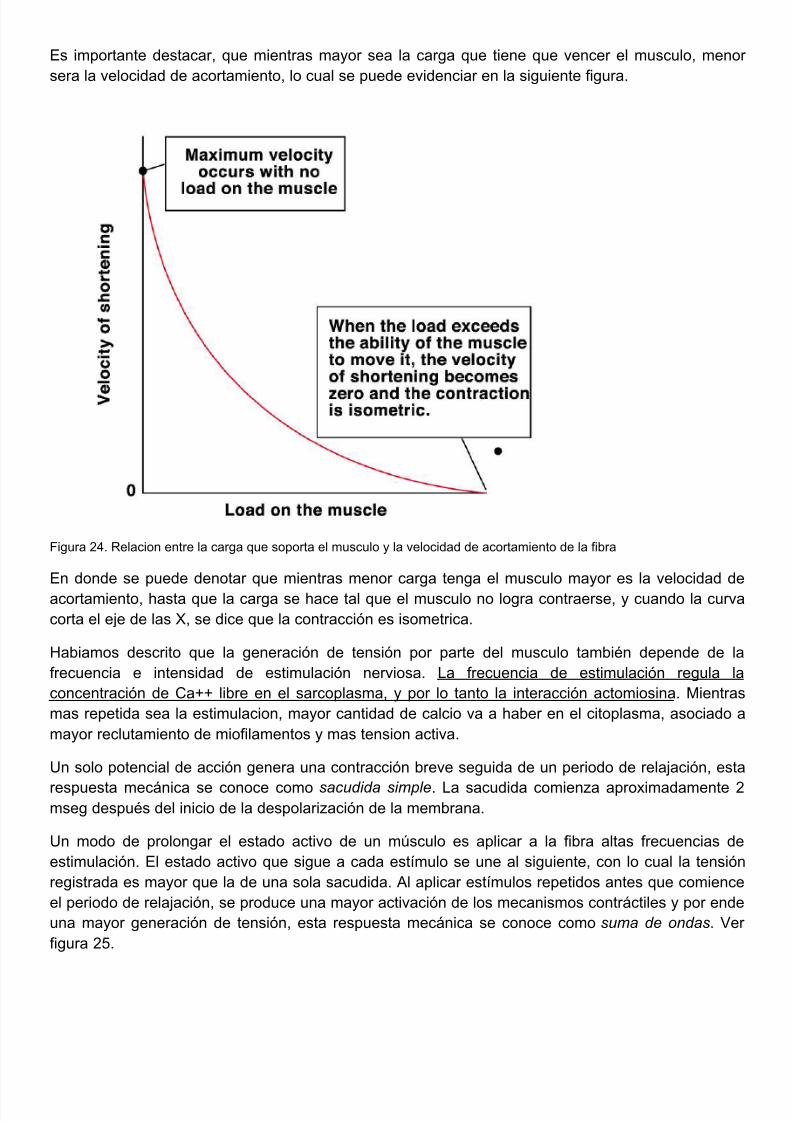
Es importante destacar, que mientras mayor sea la carga que tiene que vencer el musculo, mensera la velocidad de acortamiento, lo cual se puede evidenciar en la siguiente figura.
Figura 24. Relacion entre la carga que soporta el musculo y la velocidad de acortamiento de la fibra
En donde se puede denotar que mientras menor carga tenga el musculo mayor es la velocidad acortamiento, hasta que la carga se hace tal que el musculo no logra contraerse, y cuando la cucorta el eje de las X, se dice que la contracción es isometrica.
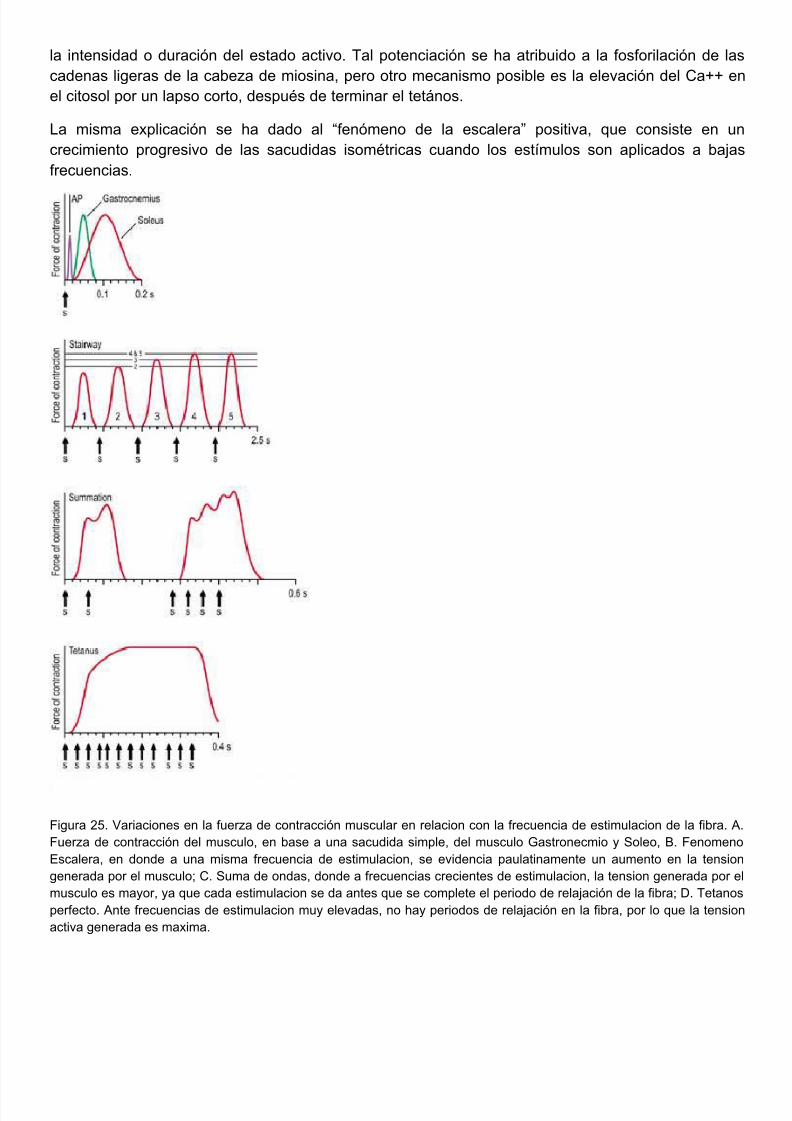
Habiamos descrito que la generación de tensión por parte del musculo también depende defrecuencia e intensidad de estimulación nerviosa. La frecuencia de estimulación regula concentración de Ca++ libre en el sarcoplasma, y por lo tanto la interacción actomiosina. Mientrmas repetida sea la estimulacion, mayor cantidad de calcio va a haber en el citoplasma, asociadomayor reclutamiento de miofilamentos y mas tension activa.
Un solo potencial de acción genera una contracción breve seguida de un periodo de relajación, e
respuesta mecánica se conoce como sacudida simple. La sacudida comienza aproximadamentemseg después del inicio de la despolarización de la membrana.
Un modo de prolongar el estado activo de un músculo es aplicar a la fibra altas frecuencias estimulación. El estado activo que sigue a cada estímulo se une al siguiente, con lo cual la tensregistrada es mayor que la de una sola sacudida. Al aplicar estímulos repetidos antes que comienel periodo de relajación, se produce una mayor activación de los mecanismos contráctiles y por enuna mayor generación de tensión, esta respuesta mecánica se conoce como suma de ondas. Vfigura 25.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 30/38
Si la frecuencia de estimulación es lo suficientemente alta, los músculos alcanzan la tensión máxiposible, conocida como tensión tetánica completa, que corresponde al máximo valor del estaactivo.
La tension generada durante el tetanos es mayor a la generada en la sacudida simple, debido a qcon la estimulacion rapida y repetida de las fibras musculares antes de que aparezca la relajacise va realizando una suma de ondas sucesiva que permite una activacion prologada de todos
mecanismos contráctiles de la fibra, esto debido a un aumento en la concentración de Caintracelular, igualmente debido a esta alta frecuencia en los estimulos no existe un periodo evidende relajación que permita la disminución de esta concentración (ya sea por receptación del mismoreticulo o difusión hacia el exterior celular), esta elevación prolongada de los niveles de Caintracelular permite una mayor interaccion entre los filamentos delgados y los puentes cruzados miosina), que se evidencia en un aumento en la generación de tensión por parte del músculo.
Existen 2 tipos de tetanos: perfecto e imperfecto, el tetanos perfecto se consigue cuando debido aalta frecuencia de los estimulos, las respuestas unitarias de contracción se fusionan en una socontracción, sin que hayan periodos de relajación evidentes, con lo cual la concentración de cal
intracelular es maxima y se encuentran interactuando la mayor cantidad de puentes cruzadgenerando la maxima tension activa posible ; en el tetanos imperfecto, hay una ligera disminuciónla frecuencia de los estimulos, y por ende existen periodos de relajación incompleta entre estimulos repetidos, en este tetanos debido a este corto periodo de relajación hay una disminucrelativa de los niveles de Ca++ intracelular que traen como consecuencia algunos desacoplamienentre los puentes cruzados y los filamentos delgados en la fibra muscular, disminuyendo ligeramela tension activa generada. Ver figura 25.
Durante la actividad muscular sostenida (tetanos completo), el músculo presenta paulatinamente udisminución temporal en la capacidad maxima para generar fuerza (tension), esto se conoce co
fatiga muscular .
El estado de fatiga de un músculo actua como un mecanismo protector que inhibe el suicidio de celulas musculares por extinguir sus reservas de ATP, ante una actividad sostenida.
Con la actividad intermitente repetida, la disminución inicial en la producción de fuerza es debidoefectos sobre el aparato contráctil, y la posterior disminución de fuerza es debido a la disminución la liberación de Ca++ del RS, ambos causados por los cambios metabolicos dentro del músculo.
Esta disminución de generacion de fuerza se ha determinado que puede ocurrir bien pordisminución progresiva de las reservas de glucogeno y síntesis de ATP (necesario para
contracción muscular, y para la recaptacion de Ca++ al reticulo, por lo que la relajación se retrasasi como por un aumento en la concentración de protones intracelular (liberados en la rglicolitica), que trae como consecuencia una disminución del pH intracelular; y a una elevación enconcentración de fosfato inorganico (Pi), por el desdoblamiento del ATP por la miosina.
El pH acido y las altas concentraciones de Pi por ejemplo, afectan la producción de fuerza yrecaptura de Ca++ por el reticulo.
Después de un tetános, la amplitud de la sacudida muscular simple es mayor. Este comportamieha sido llamado “potenciación post tetánica” y probablemente se debe a una alteración temporal
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 31/38
la intensidad o duración del estado activo. Tal potenciación se ha atribuido a la fosforilación de cadenas ligeras de la cabeza de miosina, pero otro mecanismo posible es la elevación del Ca++ el citosol por un lapso corto, después de terminar el tetános.
La misma explicación se ha dado al “fenómeno de la escalera” positiva, que consiste en crecimiento progresivo de las sacudidas isométricas cuando los estímulos son aplicados a bafrecuencias.
Figura 25. Variaciones en la fuerza de contracción muscular en relacion con la frecuencia de estimulacion de la fibraFuerza de contracción del musculo, en base a una sacudida simple, del musculo Gastronecmio y Soleo, B. FenomEscalera, en donde a una misma frecuencia de estimulacion, se evidencia paulatinamente un aumento en la tensgenerada por el musculo; C. Suma de ondas, donde a frecuencias crecientes de estimulacion, la tension generada pomusculo es mayor, ya que cada estimulacion se da antes que se complete el periodo de relajación de la fibra; D. Tetaperfecto. Ante frecuencias de estimulacion muy elevadas, no hay periodos de relajación en la fibra, por lo que la tensactiva generada es maxima.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 32/38
La contracción muscular requiere energía y el músculo se compara con “una máquina para conveenergía química en trabajo mecánico”. La fuente inmediata de esta energía es el ATP, y éste forma por el metabolismo de carbohidratos y lípidos.
El músculo tiene 3 vías para regenerar ATP.
El ATP se sintetiza de nuevo a partir del ADP por la adición de un grupo fosfato. Parte de la energpara esta reacción endotérmica proviene de la degradación de glucosa hasta CO2 y agua, peromúsculo también cuenta con otro compuesto de fosfato de alta energía que puede aportar eenergía por periodos cortos. Este compuesto es fosforilcreatina, que se hidroliza hasta grupcreatina y fosfato con la liberación de una cantidad considerable de energía. Durante el ejerciceste compuesto se hidroliza en la unión entre las cabezas de miosina y la actina, con lo que se forATP a partir de ADP y esto permite que continúe la contracción, siendo esta la forma mas rapida
obtener ATP por parte del musculo, la que se desarrolla inmediatamente al inicio de la contraccmuscular.
Como sabemos, el principal combustible muscular son los carbohidratos, el músculo almaceglucosa en forma de glicógeno, este sirve como una fuente de reserva de glucosa en el múscuDurante el ejercicio este glucogeno es degradado y la glucosa entra en una vía metabólica conoccomo glicólisis, la cual da como resultado final piruvato y 2 ATP, en condiciones aeróbicas epiruvato entra al ciclo de Krebs y se produce ATP en la fosforilación oxidativa. Durante el ejerciextenuante, cuando la síntesis aeróbica de energía es insuficiente para cubrir la demanmetabólica, este piruvato puede reducirse hasta lactato sin que tenga que entrar en el ciclo de Kreeste proceso se denomina glicólisis anaeróbica y conlleva la producción neta de mucho menenlaces fosfato de alta energía, pero no requiere la presencia de O2. El uso de la vía anaeróbicaautolimita porque, a pesar de la rápida difusión del lactato a la corriente sanguínea, se produce uacumulación muscular de lactato tal, que finalmente rebasa la capacidad de los amortiguado
titulares y causa un descenso en el pH que inhibe las enzimas. No obstante, por periodos cortos,presencia de una vía anaeróbica para la degradación de la glucosa permite un esfuerzo muscumucho mayor al que sería posible sin ella. Sin embargo, la via de la fosforilacion oxidativa, aunqes lenta, es la via de mayor rendimiento metabolico, al proporcionar 36 moleculas de ATP, por lo qes la ruta metabolica de eleccion de las fibras musculares lentas.
Las células músculo esqueléticas están especializadas en 2 tipos principales. Esta especializacpermite elevadas velocidades de contracción o contracciones de larga duración. Las dos clases células se diferencias en que unas se expresa el gen para una isoenzima de la miosina lenta y otras el de la miosina rápida (es decir, tiene una actividad ATPasa alta).
Las fibras lentas (tipo I), caracterizadas por velocidades moderadamente bajas, consumen ATPritmos moderados, reciben un buen aporte sanguíneo y suelen llamarte fibras rojas. Se caracterizpor ser oxidativas, al no requerir de una gran cantidad de ATP, este puede ser obtenido a partir defosforilación oxidativa. Si el aporte sanguíneo es apropiado, las fibras lentas proporcionan una grresistencia.
Las fibras rápidas (tipo II), tienen un alto consumo de ATP, que solo puede conseguirse medianteactivación de la ruta glicolitica. Las fibras rápidas se fatigan enseguida con el agotamiento glucógeno.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 33/38
Figura 26. Clasificación de las fibras musculares en las unidades motoras, de acuerdo a sus propiedades fisiológicas
Ya que los axones de las motoneuronas espinales que inervan a los músculos esqueléticos ramifican para llegar a varias fibras musculares, la cantidad mas pequeña de músculo que se puecontraer en respuesta a un estímulo de una sola motoneurona no es una fibra muscular, sino todlas fibras inervadas por esa neurona. Cada motoneurona y las fibras musculares a las que inervadenominan unidades motoras. Cada motoneurona inerva solo un tipo de fibra muscular, por lo qtodas las fibras musculares de una unidad motora son del mismo tipo, también es importadestacar, que mientras menos fibras musculares tenga una unidad motora determinada, mafineza y regulación de la contracción va a existir, ejemplo: los músculos oculares presentan solofibras por cada unidad motora. Con base en el tipo de músculo que inervan y, por tanto, con base
la duración de la sacudida, las unidades motoras se dividen en rápidas y lentas.
Describimos anteriormente que la intensidad de estimulación nerviosa también era capaz regulación la tensión muscular, elemento que se relaciona con el reclutamiento paulatino
unidades motoras. Dentro de cada nervio motor, cada axón presenta un nivel umbral distinto, poque a bajas intensidades de estimulación se activan unos axones y por lo tanto algunas unidadmotoras, mientras que progresivamente, a mayor intensidad de estimulación, se reclutan maxones, y mas unidades motoras, aumentando así, la capacidad de generar tensión muscular. Ereclutamiento paulatino de unidades motoras se hace en base a los requerimientos fisiológicos generar tensión, por ejemplo, cuando estamos caminando, no es necesario activar múltip
unidades motoras rápidas resistentes a la fatiga, sino mas bien con unidades motoras lenpodemos llevar a cabo esta tarea, y si paulatinamente empezamos a correr, pues reclutaremgradualmente unidades motoras rápidas y responderemos generando mas tensión, esta capacid
que tiene el organismo de reclutar unidades motoras paulatinamente, en base a l
requerimientos fisiológicos se denomina Respuesta Graduada.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 34/38
Figura 27. Respuesta graduada muscular, se evidencia el reclutamiento paulatino de unidades motoras lentas, rápifatigables, y rápidas resistentes a la fatiga, en relación con la frecuencia de estimulación nerviosa y las necesidadfisiológicas acordes.
Músculo Liso
Los órganos huecos del organismo, excepto el corazón, para cumplir su función requieren ejecuuna contracción lenta y sostenida, proporcionada por el músculo liso de sus paredes.
El músculo liso esta compuesto por células uninucleares, delgadas y fusiformes. Su núcleo y organelos citoplásmicos se ubican en el centro de la fibra.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 35/38
Desde el punto de vista anatómico, el músculo liso difiere del músculo esquelético y cardiaco, en qno presenta estriaciones transversales.
Si presenta filamentos de actina y miosina II y estos se deslizan para producir la contracción, pero están dispuestos de forma regular, por lo cual no se observan las estriaciones.
En vez de discos Z se observan cuerpos densos, los cuales se encuentran anclados al sarcolema los filamentos delgados por la alfa actinina.
El músculo liso contiene tropomiosina, pero no se ha evidenciado la troponina. Las isoformas deactina y la miosina del músculo liso difieren a la del músculo esquelético.
La fibra lisa no contiene un sistema de túbulos T ni de cisternas terminales, pues su retícsarcoplásmico esta pobremente desarrollado, pero sí contiene numerosas cisternas membranosacavéolas. Al no presentar un retículo desarrollado, no contiene altos niveles de calsecuestrina.
En general, el músculo liso contiene pocas mitocondrias, por lo cual depende fundamentalmente la glicólisis para la generación de energía.
Además de su capacidad contráctil, el musculo tiene la capacidad de sintetizar colágeno tipo elastina y proteoglicanos.
Existen varios tipos de estímulos que promueven la contracción; los más importantes son: señanerviosas, estimulación hormonal y distensión del tejido. La musculatura lisa esta inervada nervios autónomos de los sistemas simpático y parasimpático, los cuales no entran en contadirecto con ellas, por lo tanto no existen uniones neuromusculares. Los axones terminales tienmúltiples varicosidades muy próximas a la superficie de la célula muscular, las cuales secresustancias neurotransmisoras (norepinefrina y acetilcolina principalmente).
El potencial de membrana de las células musculares lisas oscila entre -60 y -70mV y contienen mcanales de calcio que la musculatura esquelética, aunque mas pocos canales de sodio. El flujo Ca++ es el principal responsable de la generación de los potenciales de acción. Se distinguen 2 tipde musculatura lisa:
Unitaria o visceral: Las células se encuentran unidas por uniones estrechas (tight junction) y unioncomunicantes (gap junction) que permiten el acoplamiento mecánico y eléctrico entre las célulcomportándose como un sincicio.
Debido a la presencia de estas uniones, la estimulación nerviosa es menor, ya que esta se propa
a lo largo de las celulas.El músculo liso visceral se caracteriza por la inestabilidad de su potencial de membrana, este pueexperimentar oscilaciones espontaneas denominadas ondas lentas, las cuales pueden alcanzar valor umbral y desencadenar potenciales de acción espontaneo; y porque presenta contraccioncontinuas e irregulares independientes de la inervación, dicho estado mantenido de contraccparcial es conocido como tono.

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 36/38
Figura 28.Contracción tónica. Se evidencia como se mantiene la fuerza tensil generada por el musculo a lo largo
tiempo
Multiunitario: Cada célula muscular se comporta como una unidad individual, se asemeja bastantemúsculo esquelético en este sentido. Presentan pocas uniones comunicantes y estrechas por lo cexiste una mayor densidad de estimulación nerviosa.
El Ca++ participa en el inicio de la contracción del músculo liso, tal como sucede en el múscesquelético. No obstante, el músculo liso visceral casi siempre tiene un retículo sarcoplásmico podesarrollado y el aumento en la concentración intracelular de Ca++ , que inicia la contracción, debe, sobre todo, a la entrada de este ion a partir del liquido extracelular por la apertura de canade Ca++ voltaje o ligando dependientes.
En presencia de una sustancia agonista (noradrenalina, angiotensina, endotelina) se activdeterminados receptores de membranas que comienzan una cascada de transducción de señamediada por la acción de la fosfolipasa C que lleva a la Protein Kinasa C que va a fosforilar a canales de calcio tipo L, activándolos. Estos canales de calcio tipo L, también se abren en respuea la despolarización de membrana en respuesta al estiramiento de la fibra muscular.
En el músculo liso, la regulación de la contracción esta asociada a la miosina y no a la actina, debia la ausencia de troponina. Como consecuencia de el aumento en la concentración intracelular Ca++ este se une a la calmodulina, formando el complejo Ca++- Calmodulina, este compl
promueve: la exposición de los sitios activos de la actina, mediante el desplazamiento lateral detropomiosina; y la activación de la cinasa de la cadena ligera de miosina dependiente
calmodulina, esta proteina fosforila a la molécula de miosina, estimulando su actividad ATPanecesaria para el inicio de la contracción.
En algunas fibras musculares lisas la fosforilación es mantenida a un bajo nivel en ausencia de estímulo externo o de una activación mecánica; esto proporciona el tono muscular, cuya intensidpuede ser variable y depende de una interaccion diferente entre la miosina y la actina llamada lat
bridges para diferenciarla de los cross-bridges, estos puentes no se disocian o se disoclentamente y mantienen el nivel tónico de tensión con minimo gasto de ATP.
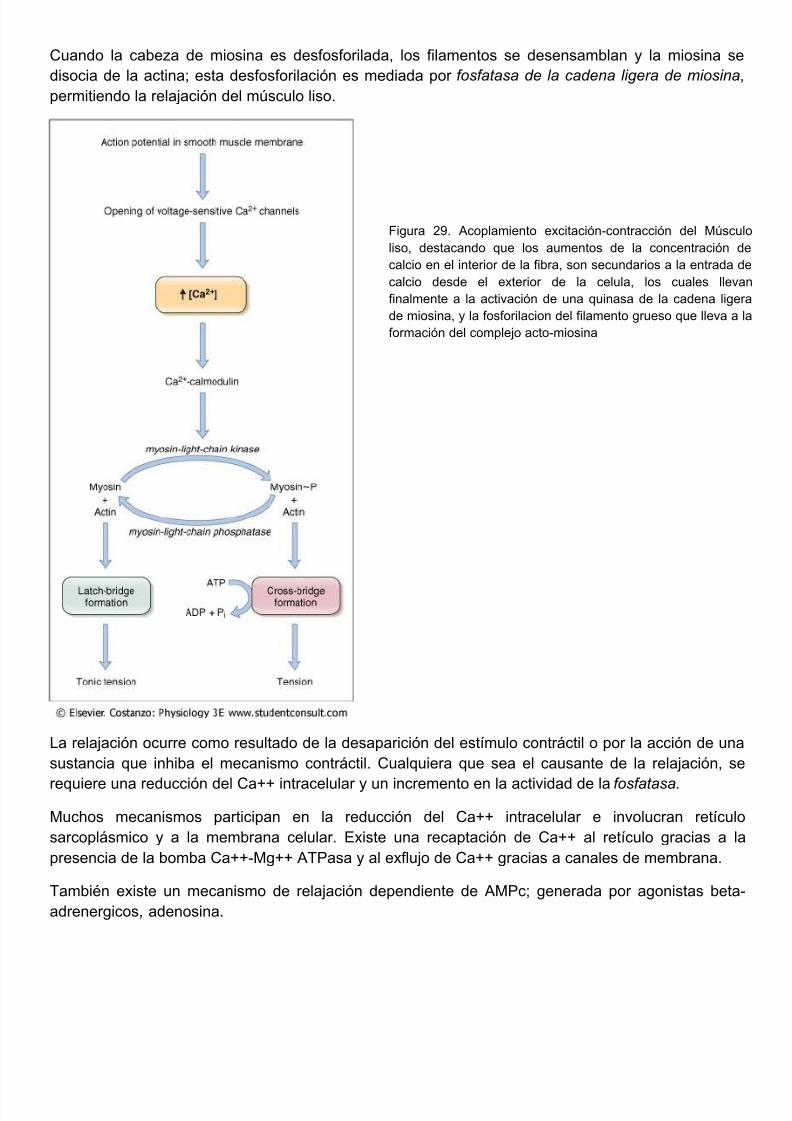
5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 37/38
Cuando la cabeza de miosina es desfosforilada, los filamentos se desensamblan y la miosina disocia de la actina; esta desfosforilación es mediada por fosfatasa de la cadena ligera de miosi
permitiendo la relajación del músculo liso.
La relajación ocurre como resultado de la desaparición del estímulo contráctil o por la acción de usustancia que inhiba el mecanismo contráctil. Cualquiera que sea el causante de la relajación,
requiere una reducción del Ca++ intracelular y un incremento en la actividad de la fosfatasa.
Muchos mecanismos participan en la reducción del Ca++ intracelular e involucran retícsarcoplásmico y a la membrana celular. Existe una recaptación de Ca++ al retículo gracias apresencia de la bomba Ca++-Mg++ ATPasa y al exflujo de Ca++ gracias a canales de membrana.
También existe un mecanismo de relajación dependiente de AMPc; generada por agonistas beadrenergicos, adenosina.
Figura 29. Acoplamiento excitación-contracción del Músliso, destacando que los aumentos de la concentraciócalcio en el interior de la fibra, son secundarios a la entradcalcio desde el exterior de la celula, los cuales lfinalmente a la activación de una quinasa de la cadena lde miosina, y la fosforilacion del filamento grueso que llevaformación del complejo acto-miosina

5/10/2018 FISIOLOGIA NEUROMUSCULAR 2011 - slidepdf.com
http://slidepdf.com/reader/full/fisiologia-neuromuscular-2011 38/38
A diferencia del músculo liso visceral, el multiunitario no presenta funcionamiento sincitial y contracciones se diseminan por todo el músculo. Por ello, las contracciones de este tipo de múscliso son mas delicadas, diestras y localizadas que las del músculo visceral.
La respuesta contráctil de este músculo suele ser mas intensa e irregular que la del múscesquelético.
Bibliografía:
Koester J, Siegelbaum S. Potencial de Membrana. En: Kandel E, Schwartz J, Jessell T. Principios Neurociencia. 4ta Edicion. Madrid: Editorial Mc Graw Hill;2000. p 125-139
Koester J, Siegelbaum S. Propagación de las señales: El potencial de acción. En: Kandel E, SchwartzJessell T. Principios de Neurociencia. 4ta Edicion. Madrid: Editorial Mc Graw Hill;2000. p 150-169
Kandel E, Siegelbaum S. Propagacion de señales en la sinapsis nervio-musculo: transmisión activadirectamente. En: Kandel E, Schwartz J, Jessell T. Principios de Neurociencia. 4ta Edicion. Madrid: EditoMc Graw Hill;2000. p 187-205
Constanzo L. Cellular Physiology. In: Constanzo L. Physiology. 3rd Edition. China: Editorial Elsevier;2006. p44.
Jesús Tresguerres. Fisiología Humana. Capítulo 2. Fisiología del Músculo. Editorial McGraw Hill. 3era Edici2005
William Ganong. Fisiología Médica. Capítulo 3. Tejido Excitable: Músculo. Editorial El Manual Moderno. 20Edición. 2006
Página web http://www.uam.es/personal_pdi/medicina/algvilla/musculo/3c.html
Imágenes tomadas de: Linda S. Constanzo. Physiology. Capitulo 1. Cellular Physiology. Editorial Elsev
3era Edicion. 2006.
Portal web http://163.178.103.176/fisiologia . Universidad de Costa Rica. Facultad de Medicina, Escuela Medicina, Departamento de Fisiologia.
This work is licensed under the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 UnporLicense. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/ or send a leto Creative Commons, 444 Castro Street, Suite 900, Mountain View, California, 94041, USA.
Fisiologia Neuromuscular by Luis Alberto Isea M is licensed under a Creative Commons Reconocimien
NoComercial-CompartirIgual 3.0 Unported License.
LAIM / 20