
Claudio Díaz Carvallo -...
55
1. INTRODUCCIÓN El arándano o blueberry (Vaccinium corymbosum), es un frutal considerado menor en nuestro país y es originario de Norteamérica. Fue introducido en Chile en la década de los ochenta, desde allí ha mostrado un importante aumento en superficie plantada. En el año 1995, Chile contaba con 1.000 ha plantadas, llegando a ser el tercero en superficie a nivel mundial (BUZETA, 1997). En el año 2005 se estima que la superficie plantada llega a las 3.500 ha (VIAL, 2005). Sin duda este fuerte desarrollo se debe a los interesantes precios alcanzados por el arándano fresco en los países del hemisferio norte, donde el principal importador de la producción chilena es EEUU, entre los meses de noviembre y mayo (BUZETA, 1997) y en Europa y países del lejano oriente como China y Japón, que han aumentado el consumo de este fruto en los últimos años (ALLENDE, 2005). En arándano, al igual que en la gran mayoría de los cultivos, una de las principales ventajas para la exportación, es la ubicación geográfica con respecto a los mercados de destino, lo que permite el abastecimiento en contra estación. Con respecto a lo anterior, en este cultivo se da una situación muy particular, ya que es un frutal muy arraigado a la tradición norteamericana, lo que genera una demanda y precios muy particulares antes del 20 de diciembre (PRITTS y HANCOCK, 1992). Esta condición ha generado en Chile tres zonas de cultivos: zona de producción temprana ubicada entre la RM y IV región; zona de producción de media estación, comprendida entre la VI y VIII región, y zona tardía, ubicada entre la IX y X región (VIAL, 2005).
Transcript of Claudio Díaz Carvallo -...

1. INTRODUCCIÓN
El arándano o blueberry (Vaccinium corymbosum), es un frutal considerado menor
en nuestro país y es originario de Norteamérica. Fue introducido en Chile en la
década de los ochenta, desde allí ha mostrado un importante aumento en superficie
plantada. En el año 1995, Chile contaba con 1.000 ha plantadas, llegando a ser el
tercero en superficie a nivel mundial (BUZETA, 1997). En el año 2005 se estima
que la superficie plantada llega a las 3.500 ha (VIAL, 2005).
Sin duda este fuerte desarrollo se debe a los interesantes precios alcanzados por el
arándano fresco en los países del hemisferio norte, donde el principal importador de
la producción chilena es EEUU, entre los meses de noviembre y mayo (BUZETA,
1997) y en Europa y países del lejano oriente como China y Japón, que han
aumentado el consumo de este fruto en los últimos años (ALLENDE, 2005).
En arándano, al igual que en la gran mayoría de los cultivos, una de las principales
ventajas para la exportación, es la ubicación geográfica con respecto a los
mercados de destino, lo que permite el abastecimiento en contra estación. Con
respecto a lo anterior, en este cultivo se da una situación muy particular, ya que es
un frutal muy arraigado a la tradición norteamericana, lo que genera una demanda y
precios muy particulares antes del 20 de diciembre (PRITTS y HANCOCK, 1992).
Esta condición ha generado en Chile tres zonas de cultivos: zona de producción
temprana ubicada entre la RM y IV región; zona de producción de media estación,
comprendida entre la VI y VIII región, y zona tardía, ubicada entre la IX y X región
(VIAL, 2005).

2
Debido a la demanda temprana de arándanos en EEUU, y los excelentes precios
originados en esta temporada, la zona de producción precoz se ha desarrollado
rápidamente. En el año 2000 existían 250 ha entre la RM y la IV región, mientras
que el 2005 se estima que existe una superficie de 470 ha (VIAL, 2005).
La zona de producción temprana difiere considerablemente al hábitat natural del
arándano, por lo tanto, la colonización de este sitio de producción se ha logrado con
variedades de bajo requerimiento de frío, además de los manejos agronómicos,
como la incorporación de materia orgánica al suelo y la modificación del pH entre
otros (SALAS, 1997).
Para lograr el máximo potencial productivo de este frutal, se deben generar
tecnologías productivas que proporcionen ventajas competitivas, principalmente en
esta zona de producción, donde el futuro panorama comercial augura una fuerte
competencia con países como Argentina y Australia (ALLENDE, 2005;
RODRIGUEZ, 2005).
Dentro de las etapas de producción del arándano, la propagación adquiere
relevancia y demanda desarrollo, debido a la fuerte necesidad por obtener plantas
de calidad, para los nuevos proyectos.
En este escenario, el uso del gas carbónico podría convertirse en una alternativa
interesante en la etapa de propagación.
El dióxido de carbono o CO2, asimilado en los órganos fotosintéticos junto con el
hidrógeno procedente del agua y los elementos minerales absorbidos por las raíces,

3
son la base estructural del crecimiento de los vegetales, en consecuencia, de la
producción primaria de los ecosistemas y de la biosfera (MEDRANO Y FLEXAS,
2000).
La concentración atmosférica de CO2, que está disponible para las plantas, se
encuentra en un nivel menor al óptimo para la absorción de éste mismo, esta
situación se acentúa mucho más en cultivos bajo invernadero (FRANCO et al.,
1991).
La técnica de la fertilización carbónica, consiste en aumentar la concentración de
CO2 por medio aéreo o radicular (fertirrigación carbónica o enriquecimiento
carbónico vía riego), de esta forma se busca elevar la tasa metabólica cuando las
condiciones de luz, temperatura y nutrición se encuentran en forma óptima, y la
concentración de CO2 se transforma en el elemento limitante.
Considerando los antecedentes del cultivo del arándano y esta técnica no probada
en nuestro país, esta investigación evaluó el efecto en la etapa de producción de
plantas en vivero, del aumento de la concentración de CO2, vía disolución de este
gas en el agua de riego. Los objetivos planteados en el presente estudio son:
1.1 Objetivo general
Implementar el uso de la fertirrigación carbónica, vía riego en la producción de
plantas de arándano en vivero, para favorecer su crecimiento.

4
1.2 Objetivos específicos
• Evaluar la factibilidad técnica del aumento de la concentración de CO2 en el
ambiente radical, a través del sistema de riego.
• Evaluar el efecto del incremento de la concentración de CO2, vía riego, sobre
el crecimiento de plantas de arándano en vivero.
• Diferenciar el efecto que tiene el aumento de la concentración de CO2,
versus el efecto por la acidificación del agua de riego, producido por la
incorporación de CO2 sobre el crecimiento vegetativo y la fotosíntesis.

5
1. REVISIÓN BIBLIOGRÁFICA
2.1. Descripción de la especie
El arándano (Vaccinium corymbosum), es un frutal menor nativo de Norteamérica,
pertenece a la familia Ericáceas, subfamilia Vacciniaceae, subgénero Cynacoccus y
es considerado dentro del grupo de los berries (PRITTS y HANCOCK, 1992).
El arándano azul de mata alta o arándano alto, ha ganado considerable interés en
EE.UU, también en partes de Europa, además del centro sur de Chile; lugares de
frío invernal y suelo ácido (GIL, 2000).
Fue introducido en Chile a principios de la década de los ochenta. Existen tres tipos:
el arándano alto; Vaccinium corymbosum, el arándano ojo de conejo; Vaccinium
ashei y el arándano bajo; Vaccinium angustifolium (BUZETA, 1997).
El arándano alto, primer cultivar introducido, proviene de una selección del
V.corymbosum y V. austral. Es una planta tetraploide, que podría alcanzar una
altura de hasta 2,5 m. En la actualidad ésta es la variedad más utilizada, ya que es
la que logra producir la mejor calidad en cuanto a tamaño y sabor. En este grupo
encontramos una gran lista de variedades, dentro de ellas O´Neal (BUZETA, 1997).
GOUGH (1994) caracteriza a la variedad O´Neal proveniente de la cruza de
´Wolcott¨ X Fla. 64-15. 1987. 9ª-7b, como un arbusto vigoroso, semierecto, con fruta
de gran tamaño, medianamente azul, de excelente sabor y muy firme, que requiere
entre 400 a 500 horas frío para superar el reposo invernal.

6
El arándano ojo de conejo, es una especie hexaploide que se origina más al sur del
continente norteamericano y alcanza alturas de hasta cuatro metros. Tiene la
cualidad de comportarse bien en suelos de pH más altos, pero en cuanto a calidad
de fruta, no supera al arándano alto (BUZETA, 1997).
El arándano bajo en EEUU es una especie silvestre, alcanza alturas de hasta un
metro y tiene una gran habilidad para generar brotes vegetativos de sus raíces
rizotomosas, también tiene una alta importancia en la base genética para la
selección de clones mejorados de arándano alto (BUZETA, 1997).
Los principales países productores son: EEUU, Canadá, Chile, Argentina, México,
Polonia, Nueva Zelanda y Australia (VIAL, 2005). Mientras que los principales
consumidores son: EEUU con un consumo del 83%, Europa con un 14% y Oriente
con un 3% (ALLENDE, 2005).
2.1.1. Botánica
El arándano es un arbusto: perenne, longevo, de hoja caduca, de madera leñosa.
Puede llegar a alcanzar en su madurez alturas de hasta tres metros (BUZETA,
1997).
Los arándanos más pequeños, menores a un metro, tienen la habilidad de formar
colonias extensas, debido a que las raíces rizotomosas emiten brotes vegetativos
(arándano bajo). Las especies que alcanzan alturas mayores a 1,5 m, por el
contrario, no tienen rizomas, pero la raíz tiene la capacidad de emitir brotes
adventicios, por lo que generalmente están desprovistas de un tronco único y más

7
bien forman coronas de brotes múltiples. El arándano alto y el ojo de conejo
pertenecen a este grupo (MUÑOZ, 1988).
Está provisto de un sistema radical superficial, de raíces finas, fibrosas y de poca
extensión. Muy importante es la desventaja de no contar con pelos radicales, por lo
tanto, las raíces más jóvenes son las encargadas de la absorción. Esta situación
genera una capacidad de absorción mucho menor comparado con otras especies
(BUZETA, 2000).
El sistema radical del arándano, aunque requiere de una humedad constante, es
muy sensible a terrenos con pobres drenajes y en condición de saturación podrían
morir en dos días (VALENZUELA, 1988).
SOTO (1993) al probar sustratos, ratifica que el crecimiento óptimo de las raíces se
genera en medios con una adecuada porosidad que se mantengan bien oxigenadas
y constantemente húmedas. Por lo tanto, manejos que favorezcan esta condición
como: el uso de materia orgánica y mulch, resultan ser muy positivos para aumentar
la distribución radicular y favorecer el crecimiento de las plantas (VALENZUELA,
1988). Por otro lado, según RIVEROS (1996) la tecnificación del riego tiene un
marcado efecto positivo en la producción de frutos y en el largo de los brotes. Estos
resultados según SIMPFENDORFER (1995) se obtienen con un 100% de reposición
de la evapotranspiración de la bandeja, para un riego por goteo; y un 75% de
reposición de la evapotranspiración de la bandeja, para riego con microjet.
Las raíces, encargadas de la absorción, son finas y fibrosas. Tienen un diámetro de
50 µm (GOUGH, 1994). Las células de la epidermis de éstas, se encuentran
asociadas bajo condiciones naturales a hongos simbiontes, pertenecientes a las

8
micorrizas ericoides, las cuales intervienen en el metabolismo del N y el P
(BUZETA, 1997).
Las micorrizas son hongos simbióticos, que entregan a las plantas hospedantes
ventajas nutricionales, hídricas y/o sanitarias. Las micorrizas ericoides le confieren
al arándano importantes ventajas en cuanto al aprovechamiento del suelo (VEGA y
MUÑOZ, 1994). El hongo más frecuentemente asociado al arándano cultivado es
Hymenoscyphus ericae o Pezizella ericae (MUÑOZ, 1988).
Las hojas son simples, se distribuyen en forma alterna en la ramilla, varían entre
uno a ocho cm en el largo y la forma puede ir de ovada a lanceolada. Tienen color
verde pálido y en otoño desarrollan una pigmentación rojiza. Hay estomas
solamente en el envés de las hojas encontrándose en densidades de 300 por mm
cuadrado (BUZETA, 1997).
Anatómicamente, las hojas tienen una epidermis compuesta de una capa de células
de empalizada y un parénquima esponjoso con abundantes espacios aéreos
(MUÑOZ, 1988).
La flor del arándano está compuesta por un ovario unido al cáliz; tiene entre cuatro a
cinco celdas con uno o más óvulos en cada lóculo; el pistilo consiste en un tubo
filiforme que termina en un estigma pequeño no modificado. La flor tiene entre ocho
a 10 estambres insertos en la base de la corola. Florece generalmente en racimos
axilares, pero también se pueden dar en forma terminal (BUZETA, 1997).

9
Las flores son perfectas y epígenas, están en racimos que emergen de yemas
laterales simples, se diferencian en verano al mismo tiempo que se agrandan en
dirección basipétala (GOUGH y SHUTAK, 1978).
La diferenciación se manifiesta por un abultamiento notorio de las yemas, las que se
recubren de escamas color café, fácilmente distinguibles de las yemas axilares
vegetativas (GOUGH, 1994).
En otoño ya son distinguibles las partes florales. La diferenciación del polen y óvulos
ocurre al reiniciarse el crecimiento en primavera, lo que toma entre ocho a nueve
meses (GOUGH y SHUTAK, 1978).
La floración ocurre sobre yemas que se diferencian al inicio del otoño, generalmente
cuando se detiene el crecimiento vegetativo, probablemente en respuesta al
fotoperíodo (MUÑOZ, 1988). Normalmente, se forma una inflorescencia por nudo,
pero en brotes medianamente gruesos pueden formarse dos. El número de nudos
florales en un brote, como el número de flores por inflorescencia, son características
de la variedad (GOUGH y SHUTAK, 1978).
El día largo de 16 horas, promueve el crecimiento vegetativo, pero evita el desarrollo
de yemas florales, el cual es máximo bajo un fotoperíodo de ocho horas por seis
semanas, tanto en el arándano alto, el bajo y el ojo de conejo (HALL et al., 1963).
Considerando que la inducción y diferenciación ocurre sobre crecimientos
vegetativos, según BAÑADOS (2005), la renovación de madera por medio de la

10
poda, tiene un efecto positivo en los calibres y la calidad de la fruta, por lo tanto,
debería ser considerada dentro de los manejos de un huerto comercial.
El fruto corresponde a una baya casi esférica que varia en tamaño desde 0,7 a 1,5
cm de diámetro. Dependiendo de la variedad, su color va desde azul claro hasta un
negro intenso, posee secreciones cerosas que le dan una terminación atractiva. El
fruto puede poseer hasta 100 semillas pequeñas ubicadas al interior del endocarpio.
Una característica del fruto, es su cicatriz, que comercialmente se busca que sea
pequeña y seca, además de un fruto firme, esto relacionado con el groso de la
epidermis (MUÑOZ, 1988 y BUZETA, 1997).
Es un fruto que presenta un bajo nivel de calorías y un alto número de compuestos
beneficiosos para la salud humana, como anticancerígenos y antioxidantes que
previenen variadas enfermedades, es por esto, que se ha convertido en un
componente importante de un dieta sana (PRITTS y HANCOCK, 1992 y GOUGH,
1994).
2.1.2 Requisitos edafoclimáticos
La mayoría de las especies de esta familia, Ericáceas, a la cual pertenecen también
el rododendro y las azaleas, tienen requerimientos muy especiales, pues prefieren
los suelos ácidos con valores de pH entre 4.0 a 5.0 para su óptimo desarrollo,
además de suelos livianos con abundante macroporosidad y materia orgánica (entre
1 a 5%). Bajo estas condiciones es donde se obtienen los mejores resultados
(VALENZUELA, 1988 y BUZETA, 1997).

11
Según FERREYRA et al. (2001) la zona central de nuestro país tiene un gran
potencial para este cultivo, pero no cuenta con las características adecuadas de pH
y porosidad en el suelo y el agua de riego, por lo general, tiene características
químicas que superan los limites recomendados, por lo tanto, el uso de aserrín,
acidificación del suelo y agua resulta una técnica beneficiosa para esta especie.
Una forma de bajar el pH es por medio del ácido sulfúrico, éste puede además
disminuir las pérdidas de amoniaco en la fertirrigación y mejorar suelos con alto
contenido de boro. Por otra parte, el ácido sulfúrico también ha demostrado su
efectividad en el aumento de la disponibilidad de fósforo, manganeso, zinc y/o hierro
en suelos calcáreos (FERREYRA, 1998).
Los suelos con altos contenidos en fósforo o calcio no son buenos para esta
especie, así como tampoco lo son los suelos en que se ha quemado leña, ya que
las cenizas elevan el pH de éste, así como los suelos calcáreos donde se provocan
severas deficiencia de fósforo (VALENZUELA, 1988).
Según BUZETA (1997) hay períodos del cultivo donde el agua debe ser muy bien
manejada, éstos corresponden a dos semanas después de la caída de pétalos, las
dos semanas previas a la cosecha y las dos a tres semanas posteriores a ésta. Al
igual que la mayoría de las especies frutales, la cuaja y crecimiento de la baya son
también períodos donde el agua es fundamental. En el ensayo realizado por
RIVEROS (1996) se determinó que para una plantación de tercer año el requisito de
agua por temporada corresponde a 4040 m3/ha para un sistema de goteo.
El clima para el arándano puede variar de acuerdo a la zona de producción, por
ende a la variedad cultivada, pero en marcos generales tiene un requerimiento de

12
frío invernal entre 650 a 850 horas frío bajo 7,2º C, para asegurar una floración
pareja y abundante en primavera. Hay programas que han obtenido variedades con
menores requerimientos que éstos como O´Neal, Mysti, Star, Jewel entre otras.
Requiere un período de crecimiento mínimo de 160 días. Las flores presentan daños
en primavera con temperaturas inferiores a -1º C; temperaturas sobre los 30º C en
las hojas detiene el crecimiento vegetativo y producen deshidratación de la fruta
(VALENZUELA, 1988 y BUZETA, 1997).
El viento también es un factor importante dentro del cultivo, porque puede provocar
importantes daños por destrucción del follaje, russet en la fruta, y además dificulta el
trabajo de polinización de las abejas (VALENZUELA, 1988).
2.1.3. Propagación
El arándano alto se propaga directamente por estacas duras de diámetro mayor a
cinco mm. También se propagan estacas tiernas con hojas y por micropropagación
(MUÑOZ, 1988).
La micropropagación o propagación in vitro es una técnica que en la práctica resulta
muy satisfactoria, ya que permite la obtención de plantas homogéneas. Este método
consiste en la proliferación de una estaquilla a partir del tejido extraído desde una
yema axilar y cultivada en un medio nutritivo, compuesto de macro y
microelementos, además de reguladores de crecimiento. Luego, estos brotes
obtenidos in vitro, son sometidos a enraizamiento, para luego ser llevados a bolsas
plásticas y terminar su etapa de vivero, en un período de uno a dos años (MUÑOZ,
1988 y BUZETA, 1997).

13
En cuanto a la fertilización de la plantas, debe comenzar una vez ya enraizadas,
sobre todo cuando se usan medios inertes, como la turba (BUZETA, 1997).
El arándano al ser una especie proveniente de zonas arenosas o pantanosas,
donde los niveles de nutrientes son muy bajos, requiere una baja cantidad de
fertilizantes (VIDAL et al., 1999).
Por lo mismo, la fertilización, durante el período de propagación de las plantas, debe
ser muy cuidadosa, ya que ante cualquier exceso éstas se intoxican y mueren en
gran proporción (BUZETA, 1997). Por otro lado, los fertilizantes foliares presentan
un gran potencial para fertilizar esta especie (GOUGH, 1997). Si ésto no es posible,
al menos se debe agregar nitrógeno en forma amoniacal y en cantidades no
superiores a 2,5 g/m2 de N total, aplicado vía riego y lavando posteriormente el
follaje para evitar quemaduras. Estas dosis pueden aplicarse hasta dos veces
durante la temporada, dependiendo de la densidad de las plantas (BUZETA, 1997).
Las fuentes de nitrógeno más recomendadas para arándanos, son las amoniacales,
ratificado en los estudios de MERHAUT y DARNELL (1995), donde el amonio tuvo
una mayor acumulación en los brotes que los nitratos, demostrando ser una mejor
fuente de nitrógeno para esta especie.
2.1.5 Fenología
La fenología de la variedad O´Neal fue estudiada por SALAS (1997) en la zona
central, determinando que la brotación ocurría a finales de julio, la floración a
comienzos de agosto con la plena flor la segunda semana de este mes, la

14
fructificación la tercera semana de agosto y la maduración, y cosecha entre finales
de octubre y diciembre.
2.2 Fertilización carbónica
2.2.1 Fundamentos del enriquecimiento carbónico
El gas carbónico CO2 es un componente básico para los ciclos de la vida, es el
material de base para la fotosíntesis. El CO2 utilizado en la fotosíntesis más el agua
y la energía lumínica producirán los carbohidratos y oxigeno. Este gas es captado
por los estomas, luego es disuelto en el agua de las cámaras estomáticas para
finalmente llegar a los cloroplastos, donde luego de numerosas reacciones el
resultado final es la fotosíntesis (LORENZO et al., 1997).
Una parte de este CO2 es absorbido por la raíz, aunque en menor proporción,
aproximadamente el 0,1 % del total fijado para el proceso fotosintético, aunque para
especies como la papa los valores ascienden a un 18% del total de CO2 fijado
(ARTECA et al., 1979).
La importancia del CO2 en las plantas radica en que del total de la materia seca que
la componen, un 40% corresponde a carbonos (LORENZO et al., 1997).
La concentración normal de CO2 en el aire está en el orden de 340 ppm. Esta
concentración se encuentra en un rango más bajo al requerido por las plantas, para
obtener el máximo en la actividad fotosintética (HICKLENTON, 1988). Según
FRANCO et al. (1991) esto ocurre, ya que existe una competencia entre el CO2 y el

15
O2 atmosféricos para ser fijados por la enzima ribulosa-difosfato carboxilasa, ya que
la concentración normal de O2 en el aire (21%) inhibe la absorción de CO2 y se
incrementa el proceso de fotorespiración, lo que supone una pérdida neta de CO2
especialmente a elevada temperatura.
Según FRANCO et al. (1991) este efecto en un invernadero es mucho más
acentuado, porque el consumo mismo dentro de él hace que la concentración del
CO2 llegue en un momento a ser tan baja que incluso pueda ser limitante para la
actividad fotosintética, llegando a detenerse, PENNINGSFELD (1983) afirma que
esta situación produce una disminución de los rendimientos, y a la vez, genera una
ineficiencia en el sistema, ésto principalmente en plantas con mucho follaje y de
rápido crecimiento, sobre todo, en los períodos en que se encuentran los otros
componentes de la fotosíntesis en su máxima expresión como luz, temperatura,
agua y nutrición. Con respecto a ésto GAASTRA (1959) señala que la respuesta es
dependiente de la temperatura y de la luminosidad; a bajo nivel de radiación (10%
del total) ocurre saturación con la concentración atmosférica normal de CO2 igual a
300 ppm, mientras que con plena radiación la saturación puede llegar a ocurrir con
una concentración cuatro veces mayor igual a 1200 ppm.
2.2.2 Descripción de la técnica de enriquecimiento carbónico
El enriquecimiento carbónico vía riego consiste en el uso de agua carbonatada para
el riego. Ésta se consigue mediante la inyección a presión del CO2 en estado
gaseoso a la matriz de riego, al disolverse en el agua produce ácido carbónico, el
que reduce el pH del agua, originando diversos bicarbonatos al reaccionar con
carbonatos y otras sales presentes. Luego de esto el agua puede ser fertilizada
normalmente (AGUILERA y CHECA, 2000).

16
2.2.3 Efectos del enriquecimiento carbónico
AGUILERA y CHECA (2000) describen en su trabajo en tomate, los distintos efectos
de la aplicación de gas carbónico, vía aérea y vía riego, conociendo muy bien el
efecto sobre la fotosíntesis de las aplicaciones de CO2 al ambiente aéreo, no así del
efecto real que tiene la aplicación del CO2 en el agua de riego.
De esta interrogante han surgido tres hipótesis: la primera planteada por D´ANDRIA
et al. (1991), en la cúal se explica el efecto positivo en distintas variables medidas
en tomates, gracias a una mejora en las características de la solución nutritiva del
suelo, al regar con un agua ligeramente más ácida, esto es producto de la formación
de ácido carbónico en el agua de riego. De esta manera al bajar levemente el pH del
agua de riego, se estaría favoreciendo la absorción de determinados nutrientes que
en condiciones normales son más difíciles de absorber como: fierro, manganeso,
cobre y zinc (MILLAR et al., 1975). Según BAKER (1988) también, el efecto leve en
la acidificación favorecería la actividad de ciertos microorganismos del suelo. De
esta forma la primera hipótesis hace referencia a un efecto producto del pH del agua
y no del aumento de la concentración de gas carbónico.
Una segunda hipótesis es la planteada por GOVINDARAJAN y POOVAIAH (1982)
que explican el efecto positivo del CO2, por medio del equilibrio competitivo que
ocurre en el suelo entre el CO2 y el etileno, así éste desplazaría al etileno y
favorecería la extensión de la raíz, mejorando ,por lo tanto, el área de absorción,
explicación al mayor desarrollo radicular observado.
Finalmente la tercera hipótesis: planteada por AGUILERA y CHECA (2000), es la
absorción adicional de CO2 por medio de las raíces, aumentando de esta manera la

17
fotosíntesis de las plantas, además de mantenerla en períodos de estrés cuando las
hojas no pueden absorber el CO2 por tener los estomas cerrados.
2.2.4 Resultados del enriquecimiento carbónico
Existe un amplio número de experiencia en el uso de CO2, a continuación se
presentarán algunas de éstas, distinguiendo el uso de CO2 al ambiente aéreo del
disuelto en el agua de riego.
En rododendro, especie que pertenece a la misma familia que el arándano, se
obtuvo entre un 10 a un 20% de aumento en el peso y un 20 a 30% de aumento en
el número de brotes laterales, producto de un aumento entre 800 a 900 ppm de CO2
en la composición atmosférica (FRANCO et al., 1991).
FRENCH (1989), observó en rododendro que el uso de CO2 afectaba negativamente
el período de enraizamiento de estacas durante el otoño, mientras que en primavera
algunas variedades como Pink Bountiful y Vulcan respondían positivamente en el
enraizamiento al aumentar la concentración de CO2 en el ambiente. Finalmente
FRENCH (1990) propuso que al aumentar la concentración de CO2 en el ambiente
aumentaba también la concentración de fructosa en el tejido interno del tallo,
situación que se plantea como un posible mecanismo de inhibición al enraizamiento.
Sin embargo, HARTMANN y KESTER (1995) recomiendan el aumento de la
concentración de CO2 en el ambiente, ya que junto con la iluminación se mantiene
activa la fotosíntesis y por lo tanto, una tasa alta de formación de carbohidratos.

18
En Rosa sp se obtuvieron aumentos en el peso; entre un 20 a un 30%, entre un 10 y
20% en la longitud del tallo, entre un 20 a 30% en el número de flores y entre un 10
a 20% en la precocidad, al aumentar la concentración de CO2 a 900 ppm (FRANCO
et al., 1991).
BIEL y SAVE (2004) en su ensayo sobre plantas de vivero de Pinus nigra,
obtuvieron un aumento en el total de la biomasa al aumentar la concentración de
CO2 a 550 y 750 ppm.
Según BOUTHERIN (1994) para niveles de enriquecimiento atmosférico entre 1000-
2000 ppm de CO2, en función de la especie y de la estación, las ganancias
obtenidas son de un 25-30% en el aumento del número de esquejes producidos,
para especies en general, además de una tasa de mantenimiento más importante y
una duración de enraizamiento más corta.
En los ensayos de enriquecimiento carbónico vía riego, AGUILERA y CHECA (2000)
obtuvieron en su ensayo de fertirrigación carbónica en tomate: un aumento del
27,4% en la cosecha acumulada con una dosis de 0,2 g de CO2/l y un aumento de
15,6% en la cosecha acumulada con una dosis de 0,35 g de CO2/l.
Según FONSECA (2001) en almácigos de tomates, la aplicación de agua
carbonatada influyó positivamente en la tasa de crecimiento de esta especie,
aumentando parámetros como el área foliar y la materia seca de distintos órganos.
D´ALBUQUERQUE (2003) en el cultivo del melón observó un aumento del 18% en
la fructificación y un 17% en la floración, al aplicar agua carbonatada, mientras que

19
en otros ensayos la inyección CO2 en el agua aumentó en un 11% el rendimiento,
20% en el rendimiento comercial y 23% en el aumento de frutos comerciales para el
cultivo del melón (KANO, 2001).
ARAÚJO (2002) en el cultivo de la calabaza, observó un aumento en el rendimiento
(número de frutos) de un 65% al aplicar agua carbonatada.

20
2. MATERIALES Y MÉTODOS
La presente investigación se desarrolló entre marzo del año 2005 y marzo del año
2006, en las instalaciones del vivero de la Estación Experimental “La Palma” de la
Facultad de Agronomía; Pontificia Universidad Católica de Valparaíso, ubicada en
La Palma s/n, Quillota, Provincia de Quillota, V región, Chile.
3.1 Instalación del equipo de inyección de CO2
Como primera parte de este ensayo, se debió instalar el equipo de inyección de CO2
al agua de riego, para lograr esto se modificó el sistema de riego del invernadero,
dónde se montó el ensayo, con esta adaptación se dejó una salida para el agua de
riego sin ninguna modificación y una para el acople del equipo de inyección de CO2
(Figura 1).
Luego se efectuó la instalación del equipo de inyección de CO2 al riego. Éste se
encontraba compuesto por: un inyector de gas, el cual entregaba el CO2 en
pequeñas burbujas a la corriente de agua de la cañería, un regulador de presión que
permite mantenerla estándar durante todo el ensayo, un caudalimetro de gas que
permitió dosificar la cantidad de CO2, un cilindro de CO2 como fuente de este gas,
fitting, además de 10 metros de cañería de PVC, que conformaba el sistema de
disolución del gas en el agua (Figura 2).
A continuación del equipo de inyección se encontraba todo el juego de matrices
portalaterales y laterales del sistema de riego (Figura 3). Para finalmente regar cada
planta a través de un gotero de 4 l/hr, más un microtubulo (Figura 4).

21
Figura 1. Modificación de la matriz de riego del vivero donde se realizó la
investigación. La modificación consistió en la adaptación de la matriz para permitir dos vías de riego, una para el agua y otra para el acople del sistema de inyección de CO2 al agua.
Salida agua de riego normal
Salida para acople del equipo de inyección de CO2

22
Figura 2. Equipo para la inyección de CO2 en el agua y sus partes principales. El
equipo de inyección de CO2 se encontraba compuesto de: (a) cilindro de CO2; (b) regulador de presión del cilindro; (c) caudalimetro de gas; (d) inyector de CO2; (e) set de cañerías dispuestos como sistema para la dilución del CO2 en el agua.
(d) Inyector de CO2 (a) Cilindro de CO2
(b) Regulador de presión
(c) Caudalimetro
(e) Sistema de dilución de CO2

23
Figura 3. Matriz de cañerías para la distribución del agua de riego con y sin CO2.
Sistema compuesto por la matriz principal, lateral y portalateral de riego, independiente para el agua con y sin CO2.
Matriz, portalateral y lateral de agua normal
Matriz, portaleral y lateral de agua con CO2

24
Figura 4. Sistema de distribución de agua para cada planta. Sistema compuesto de
un gotero de cuatro l/hr mas un microtubulo fijado al contenedor de cada planta de arándano.
Gotero individual de 4 l/hr
Microtubulo

25
3.2 Metodología y descripción de los ensayos
Para evaluar el efecto del aumento de la concentración de gas carbónico en el
arándano, se utilizaron plantas de la variedad O´Neal de 18 meses de edad, todas
éstas obtenidas por medio de propagación in vitro.
Estas plantas fueron sometidas a un aumento de la concentración carbónica, por
medio de la inyección y dilución de este gas en el agua de riego, el cual fue aplicado
como tal y como aplicación foliar, basada en una aspersión diaria al follaje, entre las
12:00 y 13:00 horas, período del día en que por su alta luminosidad se favorecería
la fotosíntesis (HICKLENTON, 1988). La concentración de CO2 en el agua fue
cuantificada por medio del caudalímetro de gas, siendo ésta de 300 ± 50 mg de CO2
por litro de agua (300 ±50 ppm) y se mantuvo durante el experimento (Anexo 1).
La frecuencia de riego fue la misma para todas las plantas y correspondió a dos
riegos por semana para los meses de septiembre, octubre y noviembre, y de tres
veces por semana para diciembre y enero, éste fue determinado de forma práctica a
través de la observación de la humedad del sustrato, cada riego tuvo una duración
de cuatro minutos. El volumen de agua aplicado fue de 0,28 litros por riego, por lo
tanto al final del período del ensayo se aplicaron 13.44 litros de agua por planta, por
lo que las plantas sometidas al enriquecimiento carbónico vía riego al final del
ensayo tuvieron una dosis de 4,03 g de CO2 por planta.
Para las plantas sometidas a la aspersión al follaje de agua enriquecida con CO2, el
volumen de agua aplicada fue de 0,01 litros de agua por planta, lo que al final del
ensayo correspondió a una dosis de 1,5 litros por planta, por lo tanto las plantas
sometidas al enriquecimiento carbónico de aspersión foliar al final del ensayo
tuvieron un consumo de 0,45 g de CO2 por planta.

26
Las plantas estuvieron sometidas bajo estas condiciones entre el 2 de septiembre
del año 2005 y el 20 de enero del 2006 y constituyeron los siguientes tratamientos:
T 0 Testigo, plantas regadas con agua.
T 1 Plantas regadas con agua con CO2 incorporado.
T 2 Plantas regadas con agua mas aplicación foliar de agua con CO2
Incorporado.
T 3 Plantas regadas con agua con CO2 incorporado mas aplicación foliar de
agua con CO2 incorporado.
La fertilización fue la misma para todos los tratamientos y estuvo basada en la
aplicación de un gramo de urea por planta semana por medio (BUZETA, 1997).
Complementariamente a esto se realizó un análisis constante del pH, tanto en el
sustrato, como en el agua de riego. Para el primero, se introdujo el electrodo del pH-
metro en los contenedores, en la parte alta y baja, y para el agua se tomó una
muestra en un frasco directamente desde el microtúbulo de distintas plantas para
luego introducir el electrodo. La lectura del pH fue entregada directamente por
medio de un pH-metro portátil digital marca Veto, modelo Exstill (Anexo 2).
3.3 Variables evaluadas
Las variables evaluadas en este ensayo corresponden a parámetros que tienen
relación directa con la calidad de una planta terminada de arándanos, por lo tanto,
cualquier efecto positivo sobre una de ellas significa la obtención de una planta de
mejor calidad.

27
3.3.1 Desarrollo vegetativo
Los parámetros de crecimiento que fueron evaluados son: el diámetro de la ramilla a
tres cm de altura; la longitud del brote apical y subapical de la ramilla; el número de
entrenudos del brote apical y subapical; el número de anticipados de la ramilla; el
número de brotes nuevos emitidos desde la corona; la longitud de los brotes nuevos
emitidos de la corona; el peso fresco, y seco de raíz.
Las mediciones comenzaron el 2 de septiembre, siendo esta fecha la primera
medición, además ésta correspondió al nivel inicial para todos los parámetros
medidos, ya que los datos analizados fueron las tasas de crecimientos y producción,
excepto para el número de ejes nuevos emitidos desde la corona, su longitud y el
peso de raiz, que fueron medidos tiempo después de comenzar la aplicación de los
tratamientos. Las mediciones finalizaron el 20 de enero, realizándose así un total de
ocho mediciones (Anexo 3).
Para medir las longitudes y diámetros se utilizó un piedemetro digital marca
Mitutoyo, modelo CD-G”65, 0,01.
Para obtener el peso fresco de la raíz se tomaron las 12 repeticiones de cada
tratamiento, se sacaron del contenedor y bajo un chorro de agua se desprendió
cuidadosamente todo el sustrato, luego se cortaron sobre la corona y luego de
cuatro horas, para eliminar el exceso de agua, se pesaron en una balanza digital
marca PRECISA, modelo 1620c. Luego estas raíces fueron deshidratadas durante
48 horas a 70°C en una estufa marca HERAEUS modelo UT12, y posteriormente
pesadas para obtener el peso seco. Para el resto de las variables se utilizó el
conteo manual. Las mediciones se realizaron sobre plantas y ramillas marcadas al
comienzo del ensayo (Figura 5).

28
Figura 5. Ramilla marcada y estructuras de arándano sobre las cuales se realizaron
las evaluaciones de crecimiento vegetativo.
Brote apical y entrenudos
Brote subapical y entrenudos
Anticipados
Diámetro de la ramilla
Ejes nuevos emitidos de la corona y su longitud.

29
3.3.2 Eficiencia fotosintética
El estado fotosintético de las plantas sometidas a los diferentes tratamientos se
evaluó por medio de la eficiencia del fotosistema II. Tras un período de oscuridad
que se logró cubriendo una hoja marcada con una lámina de papel aluminio por 30
minutos, obteniéndose de esta manera la fluorescencia máxima (Fm) y la
fluorescencia instantánea o mínima (F0), la diferencia de éstas entrega la
fluorescencia variable (Fv) que es igual a Fm-F0, mientras que la eficiencia
fotosintética o yield se obtiene de la relación de Fv/Fm (AGUILERA y CHECA, 2000).
Para medir la eficiencia fotosintética o yield fotosintético se utilizó un fluorímetro
marca JUNIOR PAM, el cual entrega una lectura inmediata. Este valor se basa en la
reemisión de fotones o fluorescencia. La medición se realizó sobre hojas marcadas
y adaptadas a la oscuridad por un período mínimo de 30 minutos, cubriéndolas con
papel aluminio.
Se realizaron seis mediciones en total entre el 12 de septiembre y el 19 de
diciembre (Anexo 3).
3.4 Diseño experimental
La unidad experimental correspondió a cada planta o la ramilla de ésta, ubicada en
un contenedor de siete litros, en un sustrato compuesto por turba con tierra de
hojas, aciculas de pino y perlita. Se tomaron 12 repeticiones para cada tratamiento.
El diseño del montaje del experimento, fue realizado de manera tal que se
bloquearan algunas condiciones ambientales que diferían en el interior del vivero,
donde se realizó el ensayo, el factor principal para realizar el bloqueo fue la luz
(Figura 6).

30
Para determinar el efecto que tiene sobre el crecimiento y sobre la fotosíntesis el
aumento de la concentración de CO2, se utilizó un BCA (Bloques completamente al
Azar) como método estadístico de análisis de datos (Anexo 4) y como herramienta
de análisis de varianzas el Test de Tukey con un 95% de confianza, para cada una
de las variables estudiadas, estos datos fueron obtenidos utilizando el software de
análisis estadístico MINITAB, versión 13.32, 2002.
El ensayo se montó con seis bloques que contenían las cuatro repeticiones,
dispuestas al azar (Figura 7).

31
Figura 6. Disposición de la luz en el vivero. Debido a la distribución irregular de la
luz en el vivero esta variable fue bloqueada por medio del diseño estadístico.

32
Figura 7. Disposición de las plantas de arándanos en el vivero. Ensayo dispuesto en
bloques completamente al azar.

33
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1 Diámetro de la ramilla
A continuación se presentan los resultados obtenidos de la variable diámetro de
ramilla (Cuadro 1).
CUADRO 1. Diámetro de ramilla en plantas de arándano sometidas a los diferentes tratamientos de riego y aplicación foliar.
Tratamiento Diferencia en diámetro de
ramilla promedio (mm) T0: Testigo, plantas regadas con agua 0,462 a T1: Plantas regadas con agua con CO2 incorporado 0,480 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 0,430 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 0,451 a
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
El aumento de la concentración de CO2 por medio del riego o de la aplicación foliar,
no presenta efecto sobre el diámetro final de la ramilla, en arándano, este resultado
difiere de los obtenidos por BARON y GORSKI (1986) para Solanum melongena,
donde la variable diámetro del vástago presentó un aumento al ser enriquecido el
sustrato con CO2, así como la variable numero brotes, estos resultados si se
comparan con los obtenidos por GLENN y WELKER (1997), difieren, ya que este
para la especie peral no observó diferencia en la variable diámetro del tronco, pero

34
si para otros parámetros como el desarrollo radicular, estos resultados parecieran
ser un factor común dentro de este tipo de ensayos, lo que hace suponer que el
efecto del aumento de la concentración de CO2 tiene un efecto variado y particular
para las distintas especies y variedades.
4.2 Longitud de brotes
4.2.1 Tasa de crecimiento de brotes de la ramilla
A continuación se presentan los resultados de la tasa de crecimiento de la variable
longitud de brotes apical y subapical (Cuadro 2 y 3).
CUADRO 2. Tasa de crecimiento de la variable longitud del brote apical de arándano para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Tasa de crecimiento
promedio (mm) T0: Testigo, plantas regadas con agua 6,120 a T1: Plantas regadas con agua con CO2 incorporado 5,494 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 5,557 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 5,229 a
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).

35
CUADRO 3. Tasa de crecimiento de la variable longitud del brote subapical de arándano para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Tasa de crecimiento
promedio (mm) T0: Testigo, plantas regadas con agua 6,572 a T1: Plantas regadas con agua con CO2 incorporado 4,311 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 6,715 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 6,189 a
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
4.2.2 Longitud del brote emitido por la corona
A continuación se presentan los resultados de la variable longitud del brote emitido
de la corona (Cuadro 4).

36
CUADRO 4. Longitud del brote emitido por la corona para plantas de arándanos para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Longitud de brotes
promedio (mm) T0: Testigo, plantas regadas con agua 7,033 a T1: Plantas regadas con agua con CO2 incorporado 8,017 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 5,700 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 8,058 a Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
La tasa de crecimiento de la longitud del brote apícal y subapical, así como la
longitud de brotes emitidos por la corona no presentaron ningún efecto al ser
sometidas a los diferentes tratamientos de enriquecimiento carbónico, este resultado
demuestra que la especie arándano variedad O´Neal no presenta respuesta frente a
este tipo de aplicaciones, similar a los resultados expuestos por FRANCO et al.
(1991) en donde el aumento de la concentración de CO2 en distintas plantas
ornaméntales tuvo efectos variados sobre la longitud de los brotes.
4.3 Producción de nudos
A continuación se presentan los resultados de la variable producción de nudos en
brotes apical y subapical (Cuadro 5 y 6).

37
CUADRO 5. Producción de nudos en el brote apical en plantas de arándano para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Número de nudos
promedio T0: Testigo, plantas regadas con agua 3,580 a T1: Plantas regadas con agua con CO2 incorporado 4,670 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 3,830 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 3,830 a Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
CUADRO 6. Producción de nudos en el brote subapical en plantas de arándano para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Número de nudos
promedio T0: Testigo, plantas regadas con agua 2,667 a T1: Plantas regadas con agua con CO2 incorporado 1,833 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 2,500 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 1,917 a Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
La producción de nudos en los brotes apical y subapical de arándanos no
presentaron diferencias en ninguno de los tratamientos, este resultado por una parte

38
parece estar determinado por la forma de aplicación del CO2, ya que según
TISSERAT y VAUGHN (2003), quienes en sus resultados en enriquecimiento
carbónico en el ambiente aéreo para la especie Pinus taeda, obtuvieron un aumento
considerable en la variable número de nudos, esto permite suponer que para esta
variable la mejor forma de aplicación sería al ambiente aéreo, pero en este caso los
tratamientos que tenían una aplicación de CO2 vía foliar tampoco presentaron
diferencias, por lo tanto, es presumible que el tiempo y la concentración no fueron
las adecuadas para obtener un aumento del número de ellos.
Sin embargo, no es posible justificar este resultado solo por medio de la forma,
concentración y tiempo de aplicación del CO2, ya que para AGUILERA y CHECA
(2000), en su ensayo de enriquecimiento carbónico vía riego en tomates la variable
número de nudos si presentó un aumento, por lo tanto, este efecto viene dado mas
bien por un factor particular de esta especie y variedad.
4.4 Producción de brotes
4.4.1 Producción de brotes anticipados en la ramilla
A continuación se presentan los resultados obtenidos de la variable producción de
brotes anticipados en la ramilla (Cuadro 7).

39
CUADRO 7. Numero de brotes anticipados en ramilla de plantas de arándano para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Número de brotes
anticipados promedio
T0: Testigo, plantas regadas con agua 0,583 a T1: Plantas regadas con agua con CO2 incorporado 0,500 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 Incorporado 0,833 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 1,083 a Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
4.4.2 Producción de brotes nuevos desde la corona
A continuación se presentan dos tablas con los resultados del análisis; la primera
corresponde a los valores de este parámetro hasta el 7 de noviembre del 2005 y; la
segunda, a una medición realizada el 20 de enero del 2006, la cual se ejecutó con el
objetivo de evaluar la nueva emisión de brotes que tenía lugar en ese período
(Cuadro 8 y 9).

40
CUADRO 8. Número de brotes emitidos desde la corona en 90 días en plantas de arándano para los diferentes tratamientos de riego y aplicación foliar.
Tratamiento
Número de brotes
promedio T0: Testigo, plantas regadas con agua 1,014 a T1: Plantas regadas con agua con CO2 incorporado 0,875 a T2: Plantas regadas con agua normal mas aplicación foliar de agua con CO2 incorporado 0,653 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 0,958 a Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
Para la segunda medición del número de brotes emitidos por la corona (Cuadro 9),
se fusionaron los tratamientos regados con agua sin CO2 (T0 y T2) quedando
finalmente como T0, y los tratamientos regados con agua con CO2 incorporado (T1 y
T3) quedando finalmente como T1. Esta fusión de tratamientos se pudo realizar ya
que las aplicaciones foliares se habían suspendido un mes antes de este segundo
“flush” y estadísticamente no eran diferentes a los otros tratamientos como lo
muestra el cuadro anterior. Por esta razón el cuadro siguiente solo contiene los
tratamientos de riego de agua sin CO2 (T1) y con CO2 incorporado (T3).

41
CUADRO 9: Número de brotes emitidos desde la corona, para el segundo flush, (mes de enero del 2006) en plantas de arándano para los diferentes tratamientos de riego.
Tratamiento
Número de brotes promedio T0: Testigo, plantas regadas con agua 2,000 a T1: Plantas regadas con agua con CO2 incorporado 2,833 b
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
El enriquecimiento carbónico vía riego al sustrato, al follaje o a ambos lugares, no
presenta resultados positivos en el número de brotaciones anticipadas de la ramilla
de arándanos, tampoco en el numero de brotes nuevos emitidos desde la corona
medidos hasta el 7 de noviembre del año 2005, los resultados analizados hasta esta
fecha son contrarios a los de BARON y GORSKI (1986), donde el aumento de la
concentración de CO2 en el suelo, con carbono marcado (C14) evidenció su
trasloque a las nuevas brotaciones en la especie Solanum melongena, proponiendo
a las brotaciones como fuertes sink de energía. Sin embargo, el aumento de la
concentración de CO2 en el sustrato si presentó un efecto positivo en el número de
brotes nuevos emitidos por la corona en el segundo “flush” de crecimiento, medido
el 20 de enero del año 2006, este efecto positivo hace presumir que los resultados
de BARON y GORSKI (1986), son repetibles para la especie arándano y que el
aumento de la concentración de CO2 en el suelo mejora su absorción.
Al analizar que el aumento del número de nuevos brotes emitidos por la corona no
es significativo desde el comienzo del ensayo hasta el 7 de noviembre del 2005, y si
lo es para la medición del 20 de enero del 2006, hace suponer que existe un efecto
acumulativo del CO2 en los tejidos como lo describe SCHAFFER, WHILEY y
SEARLE (1999), y que los efectos pudiesen ser cuantificables a mayor plazo de

42
experimentación o que estos resultados están condicionados por un efecto en el
sustrato, donde al aumentar la concentración de CO2 aumenta también la actividad
biótica como lo demostró O´NEILL (1994), y que este efecto genera una mejora en
la asimilación de carbonos y nutrientes para las plantas de arándanos como lo
describe MAUNEY y HENDRIX (1988) o mas bien, ocurre un efecto acumulativo que
conlleva a la formación de ácidos orgánicos en el sustrato, estos son absorbidos por
las raíces e ingresados directamente al ciclo de Krebs con un bajo costo de energía
como lo menciona COOKER y SCHUBERT (1981) de esta manera asistiendo y
mejorando esta nueva brotación.
4.5 Peso fresco y seco de la raíz
A continuación se presentan los resultados para las variables peso fresco y seco de
la raíz (Cuadro 10 y 11)
CUADRO 10. Peso fresco de la raíz de plantas de arándanos para los distintos tratamientos de riego y aplicación foliar.
Tratamiento
Peso fresco promedio (g)
T0: Testigo, plantas regadas con agua 12,573 a T1: Plantas regadas con agua con CO2 incorporado 17,172 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 incorporado 14,086 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 17,022 a
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).

43
CUADRO 11. Peso seco de la raíz de plantas de arándanos para los distintos tratamientos de riego y aplicación foliar.
Tratamiento
Peso seco
promedio (g) T0: Testigo, plantas regadas con agua 3,614 a T1: Plantas regadas con agua con CO2 incorporado 4,323 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 incorporado 3,986 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 4,341 a
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
El uso de CO2 tiene un efecto positivo en el crecimiento y peso de la raíz en la
mayoría de las especies estudiadas, así lo demostró DAVIS y POTTER (1983) en su
trabajo en especies ornamentales, sin embargo, este efecto no es repetible para
todas las especies y está condicionado muchas veces por el ambiente, llegando a
ser incluso detrimental, como lo ocurrido en tomate y melón en el trabajo de HARTZ
y HOLT (1991), pese al gran número de precedentes positivos en variables de
crecimiento vegetativos y reproductivos, con el uso de CO2 en el suelo para estas
especies. Es así como incluso para el rododendro, especie de la misma familia de
los arándanos, el aumento de la concentración de CO2 en el ambiente tiene un
efecto dependiente; primero del material expuesto; segundo de la época de la
exposición, particularmente de la intensidad de luz (FRENCH y LIND, 1984),
tercero; de la variedad de la especie y cuarto; la concentración de CO2, encontrando
que para este arbusto el efecto del aumento en la concentración de CO2 tiene un
efecto inapreciable, negativo o mínimo en el desarrollo de la raíz para las distintas
condiciones evaluadas (FRENCH, 1989).

44
Los resultados de la variable peso de raíz para arándanos son contrarios a los
obtenidos por GLENN y WELKER (1997), en donde la aplicación de CO2 al suelo
influyó positivamente en el desarrollo de la raíz de la especie peral, sin embargo, en
este caso el efecto de la aplicación de CO2 al suelo vía riego afectó el valor de pH,
reduciéndolo desde 5.4 a 4.9, mejorando la disponibilidad de nutrientes,
generándose un efecto positivo, aumentando las variables peso de raíz, número de
raíces laterales y largo de ellas, por lo tanto, este efecto sobre el pH pudo haber
sido la razón de estos resultados, a diferencia del ensayo en arándanos donde el pH
en el suelo tuvo una variación mínima desde un pH de 6.0 para los tratamientos sin
CO2 al suelo a 5.9 para los tratamientos que se regaban con agua con CO2, por lo
tanto de existir un efecto sobre la disponibilidad de nutrientes este es mínimo.
Desde otro punto de vista, el efecto en la interferencia del etileno en el suelo al
aplicar CO2, aunque en este ensayo no fue evaluado, no puede ser descartado, ya
que éste mejora las condiciones para el desarrollo de la raíz y, por lo tanto, los
subsiguientes efectos, como el aumento del número de ejes. (OTANI y AE, 1993).
4.6 Rendimiento fotosintético
A continuación se presentan los resultados de la variable rendimiento fotosintético,
para cada una de las fechas en las que se realizó la medición, con el objetivo de
visualizar algún efecto dentro del período del ensayo. (Cuadro 12).
45
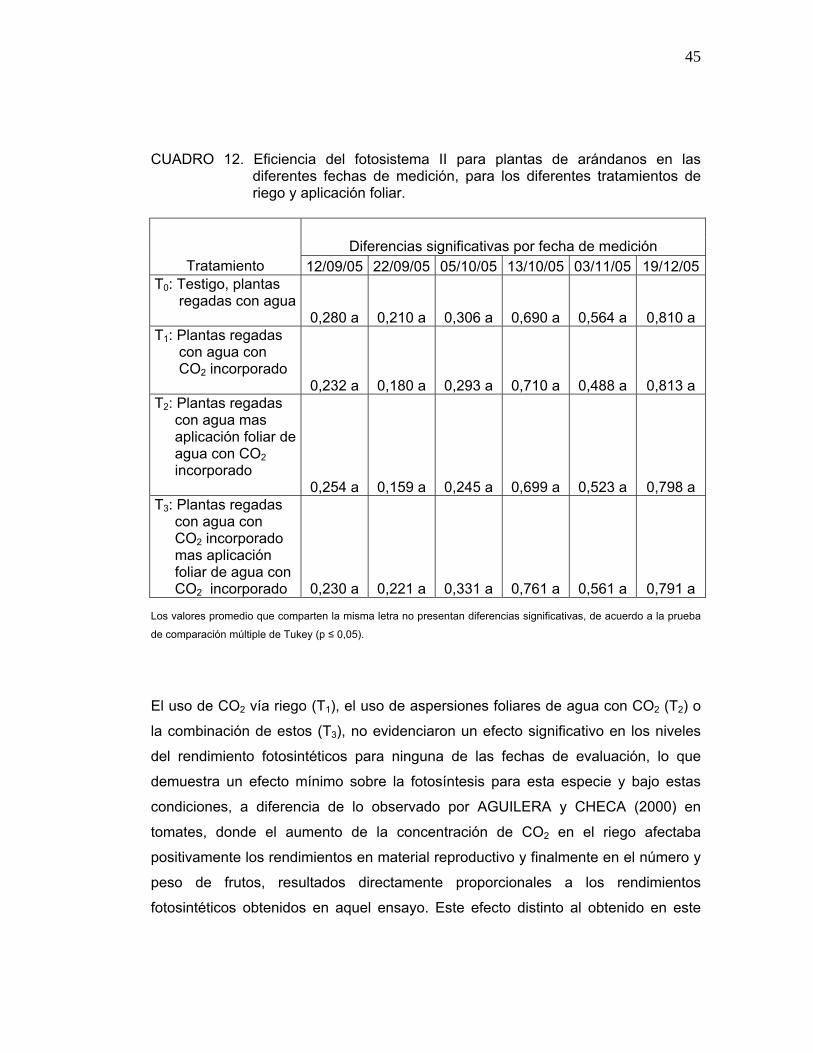
CUADRO 12. Eficiencia del fotosistema II para plantas de arándanos en las diferentes fechas de medición, para los diferentes tratamientos de riego y aplicación foliar.
Diferencias significativas por fecha de medición Tratamiento 12/09/05 22/09/05 05/10/05 13/10/05 03/11/05 19/12/05
T0: Testigo, plantas regadas con agua 0,280 a 0,210 a 0,306 a 0,690 a 0,564 a 0,810 a T1: Plantas regadas con agua con CO2 incorporado 0,232 a 0,180 a 0,293 a 0,710 a 0,488 a 0,813 a T2: Plantas regadas con agua mas aplicación foliar de agua con CO2 incorporado 0,254 a 0,159 a 0,245 a 0,699 a 0,523 a 0,798 a T3: Plantas regadas con agua con CO2 incorporado mas aplicación foliar de agua con CO2 incorporado 0,230 a 0,221 a 0,331 a 0,761 a 0,561 a 0,791 a
Los valores promedio que comparten la misma letra no presentan diferencias significativas, de acuerdo a la prueba
de comparación múltiple de Tukey (p ≤ 0,05).
El uso de CO2 vía riego (T1), el uso de aspersiones foliares de agua con CO2 (T2) o
la combinación de estos (T3), no evidenciaron un efecto significativo en los niveles
del rendimiento fotosintéticos para ninguna de las fechas de evaluación, lo que
demuestra un efecto mínimo sobre la fotosíntesis para esta especie y bajo estas
condiciones, a diferencia de lo observado por AGUILERA y CHECA (2000) en
tomates, donde el aumento de la concentración de CO2 en el riego afectaba
positivamente los rendimientos en material reproductivo y finalmente en el número y
peso de frutos, resultados directamente proporcionales a los rendimientos
fotosintéticos obtenidos en aquel ensayo. Este efecto distinto al obtenido en este

46
ensayo, no viene dado por la concentración de CO2 usada, ya que estuvo dentro del
mismo rango del ensayo de estos autores, tampoco podría ser atribuido a las
condiciones ambientales ya que ambas pruebas se realizaron durante la primavera,
por lo tanto, con condiciones de luminosidad muy similares. Bajo estos puntos
parece ser que para esta especie el aumento de la concentración carbónica bajo los
tratamientos realizados simplemente no tiene efecto en la fotosíntesis, lo que puede
ser ratificado también en los ensayos de FRENCH (1989), donde el uso de CO2
entrega resultados muy variables dentro de distintas variedades para la especie
rododendro.
Los resultados no significativos en la eficiencia fotosintética también evidencian el
nulo efecto del pH, ya que de haberse producido un cambio importante en el suelo,
producto del pH del agua de riego, se hubiesen encontrado más disponibles
nutrientes como: el Fierro, el Manganeso y el Zinc (HARTZ y HOLT, 1991), que
afectan directamente, de forma positiva la fotosíntesis, contrario a lo observado en
este ensayo, donde los valores de la eficiencia fotosintética no mostraron
diferencias.

47
5. CONCLUSIONES
1. La implementación de un sistema de enriquecimiento carbónico por medio
del sistema de riego, resultó ser una técnica de simple adaptación y uso.
2. No existió efecto del enriquecimiento carbónico vía riego y con aspersiones
foliares sobre las variables: diámetro de ramilla, longitud de brotes,
producción de nudos, producción de brotes anticipados en la ramilla y peso
seco y fresco de la raíz en plantas de vivero de arándano.
3. La variable número de ejes emitidos por la corona en el segundo “flush” de
crecimiento (mes de enero del 2006), fue mayor con el aumento de la
concentración de CO2 vía riego.
4. La acidificación del agua de riego producto del enriquecimiento carbónico no
afecto el pH del sustrato de las plantas de arándano.
5. No hubo diferencias en la eficiencia del fotosistema II, en plantas de
arándano en vivero sometidas a tratamientos de riego y aplicación foliar de
agua enriquecida con CO2.

48
6. RESUMEN
El cultivo del arándano se ha desarrollado explosivamente en superficie en los últimos años, impulsado por su alta rentabilidad. Con el desarrollo del cultivo hacia nuevas zonas de producción, se han evidenciado debilidades del sistema productivo, debido a la incipiente investigación y desarrollo que se ha realizado, factor que será vital en el futuro, donde la competencia con otros mercados y la necesidad de optar a un rubro de mayor eficiencia productiva marcaran el futuro de este cultivo. Una de las principales limitaciones en la propagación de esta especie, es el prolongado período de crecimiento en la etapa inicial, para la obtención de una planta terminada. Con respecto a esto, esta investigación evaluó el efecto del enriquecimiento carbónico vía riego y como aplicación foliar, como alternativa de aumento de la tasa metabólica para las plantas de arándano, para así adelantar su crecimiento y disminuir el período de obtención de una planta terminada. Esto se logró por medio de la inyección de CO2 en el agua de riego, para formar agua carbonatada con una concentración de 300 ± 50 ppm que posteriormente se uso para regar los tratamientos con CO2 al riego y los con aspersión foliar. El diseño estadístico utilizado correspondió a un BCA, compuesto de seis bloques con cuatro tratamientos con 12 repeticiones. El uso de agua con CO2 no tuvo efecto significativo sobre los parámetro evaluados: diámetro de la ramilla, largo del brote apical y subapical, largo de los brotes nuevos de la corona, número de nudos del brote apical y subapical, número de brotes anticipados emitidos de la ramilla, peso fresco y seco de raíz y eficiencia fotosintética, sólo se observó un efecto significativo en el número de brotes nuevos emitidos desde la corona, aspecto de especial relevancia en la capacidad de generación de brotes productivos en esta especie. La técnica de enriquecimiento carbónico resulto ser de fácil implementación y uso.

49
7. ABSTRACT
EFFECT OF CARBONIC ENRICHMENT VIA IRRIGATION ON GROWTH OF YOUNG ‘O’NEAL’ HIGH BUSH BLUEBERRY (Vaccinium corymbosum L.)
The surface area of blueberry (Vaccinium corymbosum L.) plantings has increased explosively during recent years, due to its high profitability. With the development of this crop in new production zones, weaknesses in the production systems have been observed due to the only incipient research and development that have been done. This factor will be necessary in aiming for better production efficiency to compete with new markets in the future. One of the principal limitations of blueberry propagation is the prolonged initial growth period needed to obtain new plants. The main objective of this study was to increase growth and reduce the time needed to produce finished blueberry plants by applying carbon in irrigation water and through foliar applications, thereby increasing the metabolic rate and growth of the plants. CO2 was injected into irrigation water, for carbonation at a concentration of 300 ± 50 ppm, and was subsequently used for watering and foliar application. A randomized complete block statistical design was used, composed of six blocks with four treatments and 12 replications. The use of water with added CO2 did not have any significant effect on the parameters evaluated: stem diameter, length of apical and sub-apical shoots, length of new shoots from the crown, number of nodes of apical and sub-apical shoots, number of anticipated buds on the stems, fresh and dry weight of roots, and photosynthetic efficiency. The only significant effect observed was on the number of new shoots from the crown, which is an interesting aspect in its relevance to the generation of productive shoots in this species. The CO2 water enrichment technique was easy to implement and use.

50
8. LITERATURA CITADA
AGUILERA, C.; CHECA, A. 2000. Efectos de la Fertirrigación Carbónica en Tomate. Agrícola Vergel. 19(227):747-753.
ALLENDE, J. 2005. Análisis Comercial y Visión General del Arándano en Chile.
(ASOEX) Asociación de exportadores de Chile. Berries, Arándano – Frambuesa. Santiago, 21-22 de junio del 2005. pp 1-17.
ARAÚJO, W. 2002. Aplicação de água carbonatada em abobrinha cultivada em
solo com e sem cobertura plástica. Tesis de Titulo de Mestre em Agronomia. Piracicaba. Universidad de Sao Paulo, Escola Superior de Agricultura Luiz de Queiroz. 86p.
ARTECA, R; POOVAIAH, B; SMITH, O. 1979. Changes in Carbon Fixation,
Tuberization and Growth Induced by CO2 Applications to the root zone of Potato Plants. Science. 205:1279-1280.
BAKER, R. 1989. Trichoderma spp As Plant-Growth Stimulants. Critical Reviews
in Biotechnology. 7(2):97-106. BAÑADOS, P. 2005. Fisiología del Crecimiento, Nutrición y Poda del Arándano.
(ASOEX) Asociación de exportadores de Chile. Berries, Arándano – Frambuesa. Santiago, 21-22 de junio del 2005. pp1-40.
BARON, J; GORSKI, S. 1986. Response of Eggplant to a Root Environment
Enriched with CO2. HortScience. 21(3):495-498. BIEL, C; SAVE, R. 2004. Effects af Restricted Catering and CO2 Enrichment in
the Morphology and Performance after Transplanting of Nursery-Grow Pinus nigra Seedlings. HortScience. 39(3):535-540.
BOUTHERIN, D. 1994. Multiplicación de Plantas Ornamentales. Zaragoza,
Editorial Acribia. 225p.

51
BUZETA, A. 1997. Chile: Berries para el 2000. Santiago, Fundación Chile. 133p. COOKER, R; SCHUBERT, K. 1981. Carbon Dioxide Fixation in Soybean Roots
and Nodules: 1 Characterization and Comparison with N2 Fixation and Comparison of Xylem Exudates During Early Nodule Development. Plant Physiology. 67:691-696.
DAVIS, T; POTTER, J. 1983. High CO2 Applied to Cuttings: Effects on Rooting
and Subsequent Growth in Ornamental Species. HortScience. 18(2):194-196.
D´ALBUQUERQUE JUNIOR, B. 2003. Efeito da aplicação de CO2 na água de
irrigação em diferentes fases fenológicas da cultura do melão (Cucumis melo L. var. Reticulatus) cultivado em ambiente protegido. Tesis de Titulo de Mestre em Agronomia. Piracicaba, Universidad de Sao Paulo, Escola Superior de Agricultura Luiz de Queiroz. 65p.
D´ANDRIA, R; NOVERO, R; SMITH, D; SHANAHAN, J; MORE, F. 1991.
Field-Grow Tomato Response to Carbonated Water Aplication. Agron. J. 83:911-916.
FERREYRA, R; PERALTA, J; SADZAWKA, A; MUÑOZ, A; VALENZUELA, J.
2001. Efecto de la Acidificación del Sustrato y del Agua de Riego en la Nutrición, Desarrollo y Producción del Arándano Ojo de Conejo (Vaccinium ashei reade). Agricultura Técnica. 61(4):452-458.
________ 1998. Efecto del la Aplicación de Ácido Sobre Algunas Características
Químicas de un Suelo Calcáreo. Agricultura Técnica. 58(2):163-170. FONSECA, T. 2001. Produção de mudas de hortaliças em substratos de
diferentes composições com adição de CO2 na água de irrigação. Tesis de Titulo de Mestre em Agronomia. Piracicaba, Universidad de Sao Paulo, Escola Superior de Agricultura Luiz de Queiroz. 72p.

52
FRANCO, J; BAÑON, S; GONZÁLEZ, A; FERNÁNDEZ, J. 1991. La Fertilización Carbónica en Horticultura. Agrícola Vergel. 10(117):552-561.
FRENCH, C; LIN, C. 1984. Seasonal Variations in the Effects of CO2 Mist and
Supplementary Ligthing From High Pressure Sodium Lamps on Rooting of English Holly Cutting. HortScience. 19(4):519-521.
________ 1989. Propagation and Subsequent Growth of Rhododendron Cuttings:
Varied Response to CO2 Enrichment and Supplementary Lighting. HortScience. 114(2):251-259.
________ 1990. Rooting of Rhododendron ´Anna Rose Whitney´ Cuttings as
Related to Stem Carbohydrate Concentration. HortScience. 25(4):409-411. GAASTRA, D. 1959. Photosynthesis of crop plants as influenced by light, carbon
dioxide, temperature and estomatal diffucion resístanse. Meded. Landbouwhogesschol Wageningen. 59:1-68.
GIL, G. 1997. El Potencial Productivo. 3era Edición, Santiago. Universidad
Catolica de Chile. 341p. GOUGH, R; SHUTAK, V. 1978. Anatomy and Morphology of Cultivated Highbush
Blueberry. Univer. R.I. Agr. Exp. Sta. Bull. 423p. ________ 1994. The Highbush Blueberry and Its Management. New York, Food
Product Press. 272p. GOVINDARAJAN, G; POOVAIAH, B. 1982. Effect of Root-Zone Carbon Dioxide
Enrichment on Ethylene Inhibition of Carbon Assimilation in Potato Plants. Physiologia Plantarum. 55:465-469.
GLENN, D; WELKER, W. 1997. Effects of Rhizosphere Carbon Dioxide on the
Nutrition and Growth of Peach Trees. HortScience. 32(7):1197-1199.

53
HALL, I; CRAIG, Y; AALDERS, L. 1963. The effect of Photoperiod on the Growth and Flowering of the Highbush Blueberry (Vaccinium corymbosum L.). Proc. Amer. Soc. Hort. Sci. 82:260-263.
HARTMANN, H; KESTER, D. 1995. Propagación de Plantas. Principios y
Prácticas. México, D. F., Editorial Continental. 760 p. HARTZ, T; HOLT, D. 1991. Root Zone Carbon Dioxide Enrichment in Field Does
Not Improve Tomato or Cucumber Yield. HortScience. 26(11):1423. HICKLENTON, P. 1988. CO2 Enrichment in The Greenhouse Principles and
Practice. Kentville, Timber Press, Agriculture Canada Research Station. 58p.
KANO, C. 2001. Extrações de nutrientes pelo meloeiro rendilhado cultivado em
ambiente protegido com a adição de potássio e CO2 na água de irrigação. Tesis de Titulo de Mestre em Agronomia. Piracicaba, Universidad de Sao Paulo, Escola Superior de Agricultura Luiz de Queiroz. 102p.
LORENZO, P; SANCHES-GUERRERO, M; MEDRANO, E; PERÉZ, J;
MAROTO, C. 1997. EL Enriquecimiento Carbónico en Invernadero del Sur Mediterráneo. Horticultura. 118:65-70.
MAUNEY, R; HENDRIX, D. 1988. Response of Glasshouse Grown Cotton to
Irrigation with Carbon Dioxide-Saturated Water. Crop Science. 28:835-838. MEDRANO, H; FLEXAS, J. 2000. Fijación de Dióxido de Carbono y Biosíntesis
de Fotoasimilados. In: Azcon-Bieto, J y Talon, M. Fundamentos de Fisiología Vegetal. Barcelona, McGraw – Hill. Pp 173-185.
MERHAUT, D; DARNELL, R. 1995. Ammonium and Nitrate Accumulation in
Containerized Southern Highbush Blueberry Plants. HortScience. 30(7):1378- 1381.
MILLAR, C; TURK, C; FOTH, H. 1975. Fundamentos de la Ciencia del Suelo.
Mexico D.F. Compañía Editorial Continental. 433p.

54
MUÑOZ, C. 1988. Arándano: Antecedentes Generales. Instituto de Investigación Agropecuaria. Seminario: El cultivo del Arándano. Temuco, 30 de Noviembre y 1 y 2 de Diciembre de 1988. pp 5-13.
________ 1988. Arandano: Variedades y su Propagación. Instituto de Investigación
Agropecuaria. Seminario: El cultivo del Arándano. Temuco, 30 de Noviembre y 1 y 2 de Diciembre de 1988. pp 53-65.
O´NEILL, E. 1994. Response of Soil Biota to Elevated Atmospheric Carbon
Dioxide. Plant and Soil. 165:55-65. OTANI, T; AE, N. 1993. Ethylene and Carbon Dioxide Concentration of Soil as
Influence by Rhizosphere of Crops under Field and Pot Conditions. Plant and Soil. 150:255-262.
PENNINGSFELD, F. 1983. Cultivos Hidropónicos y en Turba. Madrid, Mundi
Prensa. 343p. PRITTS, M; HANCOCK, J. 1992. Highbush Blueberry Production Guide.New
York, Northeast Regional Agricultural Engieneering Service. 200p. RIVEROS, C. 1996. Respuesta del Arándano Alto (Vaccinium corymbosum L.) de
Tercer Año al Nivel de Agua Aplicado Bajo Riego por Goteo y Microyet. Tesis Ing. Agr. Chillan, Universidad de Concepción, Facultad de Agronomía. 30p.
RODRIGUEZ, F. 2005. Situación del Arándano en Argentina. (ASOEX)
Asociación de exportadores de Chile. Berries, Arándano – Frambuesa. Santiago, 21-22 de junio del 2005. pp 1-14.
SALAS, G. 1997. Caracterización Fonológica de Tres Tipos de Arándanos del Tipo
Southern Highbush en la Zona de Curacavi. Tesis Ing. Agr. Santiago, Universidad de Chile, Facultad de Ciencias Agrarias y Forestales. 37p.
SCHAFFER, B; WHILEY, A; SEARLE, C. 1999. Atmospheric CO2 Enrichment,
Root Restriction, Photosynthesis, and Dry-matter Partitioning in Subtropical and Tropical Fruit Crops. HortScience. 34(6):1033-1047.

55
SIMPFENDORFER, C. 1995. Análisis de los Parámetros Fisiológicos, Vegetativos y Productivos en Arándano Alto (Vaccinium corymbosum L.) de Cuarto Año, Bajo Riego por Goteo y Microjet. Tesis Ing. Agr. Chillan, Universidad de Concepción, facultad de Agronomía. 75p.
SOTO, R. 1993. Efecto de las Características Físicas y Químicas de Diferentes
Mezclas de Sustratos en el Crecimiento de Arándanos en Maceta. Tesis Ing. Agr. Santiago, Universidad de Chile, Facultad de Ciencias Agrarias y Forestales. 64p.
TISSERAT, B; VAUGHN, S. 2003. Ultra-high CO2 Levels Enhance Loblolly Pine
Seedling Growth, Morphogenesis, and Secondary Metabolism. HortScience. 38(6):1083-1085.
VALENZUELA, J. 1988. Requerimientos Agroclimaticos de las Especies de
Arándano. Instituto de Investigación Agropecuaria. Seminario: El cultivo del Arándano. Temuco, 30 de Noviembre y 1 y 2 de Diciembre de 1988. pp 17-23.
VEGA, A; MUÑOS, C. 1994. Presencia de Micorrizas Ericaceas en Chile.
Agricultura Técnica. 54(3):332-339. VIAL, C. 2005. Análisis Comercial y Visión General del Arándano en Chile.
(ASOEX) Asociación de exportadores de Chile. Berries, Arándano – Frambuesa. Santiago, 21-22 de junio del 2005. pp 1-10.
VIDAL, I; AMARO, J; VENEGAS, A. 1999. Evolución Estacional de Nutrientes y
Estimación de la Extracción Anual en Arándano Ojo de Conejo (Vaccinium ashei R.). Agricultura Tecnica. 59(4):309-318.


















