13. Inmunidad celular
20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 1 de 20 file://C: \WINDOWS\ ...\C URSO DE INMUN OLOGÍ A G ENERAL 1 3_ Inmuni dad celul ar.ht 11/05/02 ÍNDICE: 13.1 INTRODUCCIÓN * 13.2 INMUNIDAD MEDIATIZADA POR CÉLULAS FAGOCÍTICAS * 13.2.1 Quimiotaxis * 13.2.2 Unión del microorganismo al fagocito * 13.2.3 Desencadenamiento de la captación * 13.2.4 Fagocitosis y mecanismos matadores * 13.2.4.1 Mecanismos dependientes de oxígeno * 13.2.4.2 Mecanismos independientes de oxígeno * 13.2.5 Activación de los macrófagos * 13.3 PAPEL DE LAS CÉLULAS T COADYUVANTES (T H ) * 13.4 CITOTOXICIDAD MEDIADA POR CÉLULAS * CURSO DE INMUNOLOGÍA GENERAL: 13. Inmunidad celular Enrique Iáñez Pareja De partamento de Microb iolo gía Universidad de Granada España
-
Upload
mario-andres-baez-allende -
Category
Documents
-
view
76 -
download
0
Transcript of 13. Inmunidad celular

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 1/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 1 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
ÍNDICE:13.1 INTRODUCCIÓN *
13.2 INMUNIDAD MEDIATIZADA POR CÉLULAS FAGOCÍTICAS *
13.2.1 Quimiotaxis *
13.2.2 Unión del m icroorganismo al fagocito *
13.2.3 Desencadenamiento de la captación *
13.2.4 Fagocitosis y m ecanismos m atadores *
13.2.4.1 Mecanismos dependientes de oxígeno *
13.2.4.2 Mecanismos independientes de oxígeno *
13.2.5 Activación de los macrófagos *
13.3 PAPEL DE LAS CÉLULAS T COADYUVANTES (TH) *
13.4 CITOTOXICIDAD MEDIADA POR CÉLULAS *
CURSO DE I NM UNOLOGÍ A
GENERAL:1 3 . I n m u n i d ad ce l u lar
Enrique Iáñez Pareja
Departamento de Microbiología
Universidad de Granada
España

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 2/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 2 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
13.4.1 Citotoxicidad directa específica (mediatizada por linfocitos CTL) *
13.4.1.1 Activación y diferenciación de los linfocitos T citotóxicos *
13.4.1.2 Fase efectora: Ataque y destrucción de la célula diana *
13.4.2 Citotoxicidad directa inespecífica mediada por células NK *
13.4.3 Citotoxicidad celular dependiente de anticuerpos (ADCC) *
13.5 RESPUESTA DE HIPERSENSIBILIDAD DE TIPO RETARDADO *
13.5.1 Fases de la hipersensibilidad de tipo retardado *
13.5.1.1 Sensibilización *
13.5.1.2 Fase efectora *
13.5.2 I nteracción entr e citoquinas y células en la DTH *
13.5.3 Efectos de la hipersensibilidad retardada *
13.1 INTRODUCCIÓN Actualmente se tiende a considerar como "inmunidad mediada por células"cualquier respuesta contra microorganismos o tumores en la que los anticuerpos jueguen un papel subordinado o secundario. Pero a estas alturas ya sabemosque en Inmunología es difícil "poner puertas al campo":
En la respuesta inmune humoral específica hay que activar células al principiode dicha respuesta.En muchas respuestas celulares los anticuerpos juegan papeles esencialescomo "puentes" entre células (lo veremos con la citotoxicidad celulardependiente de anticuerpos, ADCC).Además, cualquier respuesta celular va acompañada en paralelo deproducción de anticuerpos, que a su vez pueden modificar las respuestascelulares de varias maneras: p. ej., la formación de complejos Ag-Ac provocala activación del complemento, en la que se liberan péptidos que funcionancomo factores quimiotácticos que inducen la acumulación de mayor númerode células inmunes y el aumento de la respuesta inflamatoria.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 3/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 3 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
Por otro lado, no hay que suponer que todas las respuestas celulares dependende linfocitos T: de hecho las primeras líneas defensivas contra microorganismosson inespecíficas, y descansan en la identificación de componentes microbianoscomunes por parte de células del sistema de inmunidad natural (macrófagos,granulocitos neutrófilos, etc.).
Una de las posibles clasificaciones de la inmunidad mediada por células sería lasiguiente:
1. Reacciones dependientes de fagocitos del sistema natural:
2. El papel central de las células TH en la determinación del tipo de
mecanismo efector puesto en marcha.3. La citototoxicidad directa que conduce a la lisis de la célula diana enfermaes realizada por varios tipos celulares:
13.2 INMUNIDAD MEDIATIZADA POR CÉLULAS FAGOCÍTICAS El alumno encontrará provechoso repasar las características de las célulasmieloides fagocíticas, tratadas en el tema 2. Aquí profundizaremos en estascélulas, sobre todo en lo que atañe a los mecanismos de destrucción intracelulardel patógeno una vez que lo han engullido.
La actuación de estas células la podemos dividir a efectos pedagógicos, envarias fases:
1. Quimiotaxis2. unión del microorganismo al fagocito3. desencadenamiento de la captación del microorganismo4. fagocitosis y mecanismos matadores.
fagocitosisproducción de citoquinas
dentro del sistema específico: linfocitos T citotóxicos (matadores), que sedenominan linfocitos T citolíticos (CTL) en su fase efectora;dentro del sistema natural: células agresoras naturales (NK) y macrófagoCitotoxicidad celular dependiente de anticuerpos (ADCC)Células T CD4+ que median reacciones de hipersensibilidad de tiporetardado, que en su estado efector se denominan TDTH.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 4/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 4 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
13.2.1 Quimiotaxis
La atracción de los fagocitos del sistema de inmunidad natural hacia el foco deinfección se puede deber a varios factores:
13.2.2 Unión del microorganismo al fagocito
Esta fase es importante, ya que de su éxito depende la ocurrencia de lainternalización ulterior y el desencadenamiento de los mecanismos matadores.La unión inespecífica entre bacteria y fagocito se realiza por interacción entremoléculas de superficie del microorganismo y receptores del leucocito:
Podemos hacer una tabla parecida, pero en la columna de la bacteria ahoravamos a considerar moléculas del hospedador que recubren al microorganismo(opsonización), y que a su vez sirven para que sean reconocidas por receptores
de leucocitos:
por sustancias de origen directamente microbiano. Tal es el caso de cortospéptidos derivados de los extremos amino-terminales de proteínasbacterianas (p. ej., el formil-Met-Leu-Phe es quimiotáctico para losleucocitos, que llevan receptores capaces de reconocerlo).Ciertos componentes microbianos (como la endotoxina de bacterias Gram-negativas) activan la ruta alternativa del complemento, que libera lospéptidos C3a y C5a, que son muy quimiotácticos sobre fagocitos.
mol écu l abac te r i ana
r ecep to r de lf agoc i t o
lect i n as Ol ig osacár idos d esuper f i c ie
o l i gosacár idos d esuper f i c ie
Lec t inas
ββββ - g l ucanos CR3 y LFA-1
l íp ido A( e n d o t o x i n a )
CR3 y LFA-1
comp. opson i zan tesde l hospedadorun idos a la bac ter ia
Receptor d e l fagoc i to
C3 b CR1 ( * )
i n m u n og lob u l in as FcR ( * )

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 5/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 5 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
(*) Los receptores CR1 y FcR están entrelazados entre sí por medio del citoesqueleto, lo que explica en partesu acción sinérgica
Los receptores inespecíficos de tipo lectina disminuyen en el macrófago activado (en presencia de IFN- γ ),mientras que aumentan los CR1 y FcR. Ello permite a las células Th indicar a los macrófagos que entran altejido que fabriquen más moléculas capaces de citoadherencia.
13.2.3 Desencadenamiento de la captación
La captación del microorganismo se ve facilitada si está recubierto por elcomponente C3b del complemento y/o por anticuerpos: esta opsonizaciónpermite potenciar poderosamente el mecanismo amebiano del fagocito.
13.2.4 Fagocitosis y mecanismos matadores
En el capítulo 2 ya nos referimos a la fagocitosis, por lo que no volveremos ainsistir aquí en ella, sino que nos centraremos en lo que ocurre cuando elmicroorganismo, englobado en el fagosoma, recibe la descarga de los gránulos(lisosomas) del fagocito. El contenido preformado de estos gránulos, junto consustancias producidas de novo, constituyen una batería de mecanismosencaminados a la muerte del patógeno. Estos mecanismos se pueden clasificaren dos grandes tipos: dependientes e independientes de oxígeno.
13.2.4.1 Mecanismos dependientes de oxígeno
Los podemos estudiar a su vez bajo dos categorías: los que se basan enintermediarios reactivos de oxígeno, y los intermediarios reactivos de nitrógeno.
1 . I n te r m ed i a r i os r eact i vos de ox ígeno ( ROI ) . La unión de partículas alos receptores de la membrana del fagosoma del macrófago provoca eldenominado esta l l i do r esp i r a to r i o (por activación de la ruta de la hexosamonofosfato), que produce mucho NADH.
a. Antes de que el lisosoma se fusione con el fagosoma, en éste tienelugar una reducción de oxígeno molecular (O2) catalizada por la
NADH-oxidasa de la membrana del fagosoma; el anión superóxido
resultante (·O-2) es tóxico por sí mismo, pero a su vez da lugar aotros radicales tóxicos de vida corta, como el peróxido de hidrógeno(H2O2), el radical hidroxilo (·OH) y el oxígeno singlete (O1
2). El
macrófago se protege de los ROI que puedan salir del fagosoma pormedio de una cadena de reacciones redox en que participa glutation.
b. Una vez que el lisosoma se fusiona con el fagosoma, aquél libera lamieloperoxidasa, que actúa sobre los peróxidos en presencia dehaluros (I- y Cl-), para producir compuestos halogenados
MBP C1 qR

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 6/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 6 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
(hipohaluros) muy tóxicos y de vida larga: ácido hipocloroso (ClOH), ehipoiodoso (IOH).
2 . I n t e r m e d ia r i os r e a ct i v o s d e n i t r ó g e n o ( N R I ) a. La enzima NOS (óxido nítrico-sintetasa) combina el oxígeno molecular
con el nitrógeno guanidino de la L-arginina, para generar óxido nítrico(NO), que es tóxico para bacterias y células tumorales.
Los macrófagos de ratón (pero no los humanos) necesitan para activaresta ruta a un nivel óptimo dos señales: interferón gamma (IFN-γ )para activar la enzima NOS, y factor de necrosis tumoral (TNF) (ver el apart ado 13.2.5) .
13.2.4.2 Mecanismos independientes de oxígeno
Estos mecanismos dependen de proteínas antimicrobianas preformadas yacumuladas en los gránulos:
1. Proteínas catiónicas, con actividades de tipo antibiótico:a. defensinas (en macrófagos de conejo y en PMN neutrófilos humanos).
Son péptidos catiónicos , de unos 30 aminoácidos, ricos en cys y arg.Forman canales permeables a los iones en las bicapas lipídicas de losmicrorganismos. Actúan antes de que tenga lugar la acidificación delfagolisosoma.
b. Catepsina Gc. Azurozidina
2. Lisozima, que actúa rompiendo el peptidoglucano de bacterias (sobre todode Gram-negativas).
3. Lactoferrina (producida por neutrófilos): secuestra hierro, indispensablepara las bacterias.
13.2.5 Activación de los macrófagos
Los macrófagos en reposo pueden matar, desde luego, pero sus capacidadesmicrobicidas pueden mejorar por medio de su activación. Esta activación puedeprovocarse por:
1. productos microbianos que causan
a. activación directa de monocitos y macrófagosb. activación indirecta: los macrófagos sin activar y las células NK liberancitoquinas que a su vez activan a los macrófagos.
2. Ulterior activación por citoquinas producidas por linfocitos TH, sobre todo
por IFN-γ . Provocan atracción quimiotáctica de fagocitos y/o activación delos mecanismos dependientes e independientes de oxígeno.
En años recientes se está viendo que la activación del macrófago es algo máscomplejo de lo que se creía. La idea clave que ha surgido es que el macrófago

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 7/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 7 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
despliega unas u otras de sus funciones efectoras dependiendo de lacombinación particular de estímulos químicos que reciba, a saber, citoquinas ymoléculas inflamatorias.
La activación de los macrófagos de ratón se puede lograr mediante el IFN-γ secretado por el TH, o por la endotoxina (lípido A) de las bacterias gram-
negativas. Pero a su vez, los macrófagos activados secretan TNF-α , que alactuar sobre macrófagos ya activados provocan una mayor potenciación de suscapacidades al activarse la ruta dependiente del óxido nítrico.
En humanos, el macrófago, en presencia de IFN-gamma, activa la 1-alfahidroxilasa, que convierte el 15-hidroxicolecalciferol circulante en 1,25-dihidroxicolecalciferol (calcitriol o vitamina D3). A su vez, este calcitriol activa
más al macrófago al tiempo que inhibe los linfocitos Th1. Esta respuesta enhumanos puede ser parte de la razón de que cuando falla la eliminación delparásito haya un cambio desde TH1 a TH2, con lo que se hace crónica la
respuesta celular. La producción de calcitriol en el caso de tuberculosis osarcoidiosis puede llegar a circulación periférica, originando una afección dehipercalcemia.
Llegados a este punto, quizá sea bueno llamar la atención sobre las diferentesfases de la respuesta inmune en las que participan los macrófagos:
1. En la defensa inicial frente al patógeno: el macrófago sin activar tiene sucapacidad fagocítica basal. Al mismo tiempo secreta citoquinas que ayudana otras células (p. ej., recordar la IL-1 que participa en la activación de
linfocitos B).2. En la presentación del antígeno: actúa como célula presentadora paralinfocitos TH, que a su vez se activan secretando citoquinas.
3. En la fase efectora el macrófago se activa por citoquinas (como el IFN-γ secretado por linfocitos TH), lo cual va a potenciar sus actividades
antimicrobianas, antitumorales y de secreción de citoquinas.
Las citoquinas secretadas por macrófagos en respuesta a componentesmicrobianos cumplen importantes papeles: la IL-12 y el TNF-α , en sinergia conotros mediadores, mejoran la actividad antimicrobiana inespecífica:
el TNF-α mejora la capacidad microbicida de PMN neutrófilos y de los propiosmacrófagos;el TNF-α junto con la IL-12 provocan que las células NK liberen IFN-γ , el cualaumenta la actividad de los macrófagos;

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 8/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 8 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
13.3 PAPEL DE LAS CÉLULAS T COADYUVANTES (T H )
Las células TH desempeñan papeles esenciales en la inmunidad celular:
1. determinan la especificidad de la respuesta inmune (qué antígenos y quéepitopos son reconocidos);
2. intervienen en la selección de los mecanismos efectores destinados aeliminar al patógeno;
3. ayudan a la proliferación de las células efectoras adecuadas;4. mejoran las funciones de fagocitos y otras células efectoras.
El primero de estos papeles citados ya ha sido abordado en otro capítulo: lainteracción del conjunto de receptores TCR-CD3-CD4 con los epitopos en elMHC-II de células presentadoras condiciona la especificidad de la respuesta.
Pero el sistema inmune ha de efectuar una segunda decisión esencial: quémecanismos efectores se ponen en marcha, que sean los adecuados para lanaturaleza de cada infección en particular. Dentro de la inmunidad mediada por
células, existen varios mecanismos:
1. citotoxicidad directa específica, por medio de linfocitos T citotóxicos CD8+ (CTL);
2. degranulación de eosinófilos y mastocitos una vez que han contactado conla célula a eliminar a través de anticuerpos, que sirven de "puentes"moleculares (ADCC);
3. activación de macrófagos y reacción de hipersensibilidad de tipo retardado.
Entonces, ¿cómo se determina el tipo de acción para cada tipo de situación? La
respuesta estriba en buena parte en el patrón local de citoquinas producidas alinicio de la respuesta inmune, desencadenado ante cada tipo de patógeno, así como en la concentración local de metabolitos esteroideos y de vitamina D3 en
el tejido linfoide: estos factores determinan que predomine uno de dos subtiposde células TH, lo que finalmente condicionará el desencadenamiento de uno u
otro de los mecanismos efectores.
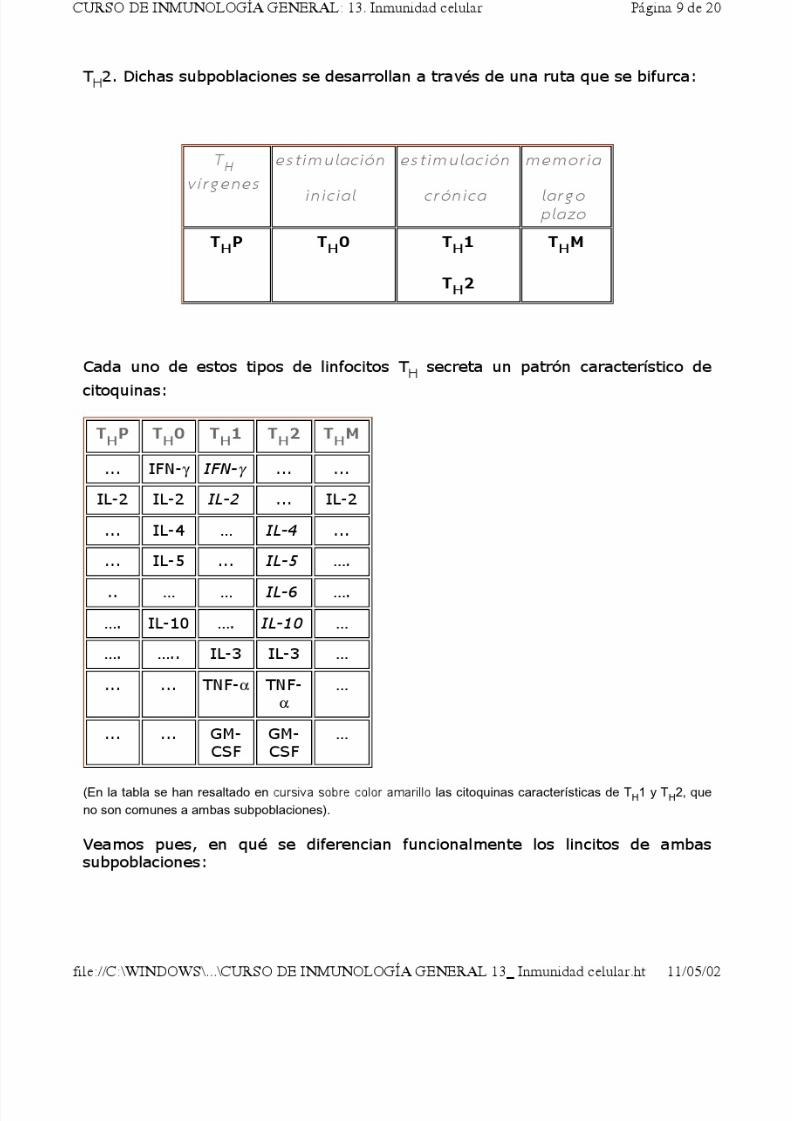
Así pues, los linfocitos TH presentan dos subpoblaciones, denominadas TH1 y
el TNF-α induce cambios en las superficies de células endoteliales y defagocitos, que pueden interaccionar de modo que los fagocitos se extravasanpara acceder al lugar de la inflamación, donde se localiza el foco de infección.
5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 9/20

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 10/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 10 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
Una vez establecidos, cada uno de estos patrones de respuesta es capaz deinhibir o suprimir al otro, debido a la acción de ciertas citoquinas de cada tipo decélula TH:
Esta diferenciación de TH1 y TH2 se descubrió en el ratón, pero existen cada vez
más pruebas de que igualmente existe en la especie humana. A los TH1 se les
suele llamar linfocitos T "inflamatorios", y a los TH2, "coadyuvantes".
Un ejemplo clínico de este papel particular de los dos tipos de células TH lo
tenemos en las dos clases extremas de lepra:
13.4 CITOTOXICIDAD MEDIADA POR CÉLULAS En este apartado vamos a tratar aquellos mecanismos efectores que dependen
de la acción de células del sistema inmune capaces de matar/lisar célulasenfermas (células diana), a las que se unen de modo directo o indirecto:
1. Citotoxicidad directa específica: es la llevada a cabo por los linfocitoscitotóxicos (TC), que están restringidos por el haplotipo propio MHC-I, y
que poseen CD8+ como moléculas correceptoras. (aunque algunas sonCD4+ y están restringididas por MHC-II)
2. Citotoxicidad directa inespecífica: las células agresoras naturales (NK)
Las células TH1 tienden a activar los macrófagos, y responden bien a
Las células TH2 tienden a incrementar la producción de mastocitos y
eosinófilos, y mejoran la producción de ciertos isotipos de inmunoglobulinas,incluyendo la IgE. Las células TH2 responden bien a antígenos presentados
por linfocitos B.
en la lepra tuberculoide se inducen sobre todo linfocitos TH1, por lo q
una buena activación de macrófagos, lo cual redunda en que se detepocas bacterias intracelulares.En cambio, en la lepra lepromatosa se induce la subpoblación TH2, q
colabora eficazmente en la respuesta específica humoral; pero losanticuerpos evidentemente no pueden acceder al interior celular, porexisten abundantes bacilos en el interior de los macrófagos; éstos, al
poder ser activados eficazmente, no pueden eliminar al parásito. Haydestrucción de tejidos.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 11/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 11 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
reconocen determinantes inespecíficos de células tumorales o infectadascon ciertos virus.
3. Citotoxicidad celular dependiente de anticuerpos: es un mecanismoindirecto por el que células matadoras como NK, eosinófilos, etc.,interaccionan con el antígeno por medio de puentes de anticuerpospreviamente unidos a receptores para Fc de la célula.
13.4.1 Citotoxicidad directa específica (mediatizada porlinfocitos CTL)
El papel más importante de este mecanismo es el de eliminar células propiasinfectadas por virus. Como veremos, los linfocitos TC en reposo, al activarse
apropiadamente, terminan diferenciándose a los llamados linfocitos T citolíticos(CTL), que son los efectores de la inmunidad celular específica.
El 90% de estos linfocitos son CD8
+
, y están restringidos por el MHC-I propio.Son capaces de reconocer a la mayoría de células nucleadas. Pero existe un10% de linfocitos TC que fenotípicamente son CD4+ y están restringidos por
MHC-II.
La actuación de estas células la podemos considerar en dos fases:
1. Activación y diferenciación de los precursores TC hasta CTL
2. Fase efectora: destrucción de la célula diana, a su vez diferenciable en:
13.4.1.1 Activación y diferenciación de los linfocitos T citotóxicos
Los linfocitos Tc vírgenes parece que se activan solamente en los órganoslinfoides secundarios, únicos entornos en los que reciben las señales necesarias.Requieren en realidad 3 señales:
1. La señal específ i ca suministrada por la interacción entre su complejoTCR-CD3-CD8 con el MHC-I de una célula presentadora profesionalinfectada (p. ej., una célula dendrítica interdigitante)
2. La señal coes t i mu l a to r i a suministrada por la misma célula (unión entreB7 y CD28).
3. La i n t e r l euc i na I L - 2.
Parece ser que el Tc expresa tras la interacción inicial una pequeña cantidad de
adhesión y formación del conjugado entre el CTL y la célula dianagolpe letaldisociación del CTLdestrucción de la célula diana.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 12/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 12 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
IL-2, pero en el caso de estos Tc vírgenes no suele ser suficiente paraactivarlos. La aportación clave de IL-2 procede de células Th1 cercanas. Dehecho, se piensa que tanto el Th como el Tc vírgenes se activansimultáneamente al unirse a la misma célula presentadora (no olvidemos quelas APCs exhiben MHC-I y MHC-II).
En cambio, las células Tc de mem or i a t i enen menos r equer i m i en tos para suactivación. De hecho, como ya sabemos, al tener moléculas de adhesión celulardiferentes a los correspondientes Tc vírgenes, pueden encontrarse el antígeno(epitopo peptídico en contexto MHC-I) en tejidos extralinfoides.
1. No requieren obligatoriamente la señal coestimulatoria.2. No requieren obligatoriamente la ayuda de Th, ya que al unirse de modo
específico a una célula propia infectada que le suministre el péptidoadecuado, provoca que el Tc produzca no sólo receptores de IL-2, sinoniveles adecuados de IL-2 capaces de auto-activarlas.
De esta manera, se provoca la pro l i f e rac ión y d i fe renc iac ión del linfocito enreposo (virgen o de memoria) hasta que se convierte en linfocito T citolítico(CTL). Este CTL se caracteriza por poseer algo más de citoplasma que el TC, y
sobre todo por poseer gránulos densos a los electrones, rodeados de unidad demembrana (llamados a veces granulosomas) y un aparato de Golgi.
13.4.1.2 Fase efectora: Ataque y destrucción de la célula diana
1) Fo r m aci ón de l con j ugado
Tras el reconocimiento específico, se produce una interacción de gran avidezentre moléculas de LFA-1 del CTL y moléculas de ICAM-1 de la célula diana.Esta unión dura unos 5-10 minutos, durante los cuales las señales de la uniónintercelular se transducen al interior del CTL por medio de cascadas de proteín-quinasas y proteín-fosfatasas, que conducen a la activación de una serie defunciones del CTL. Se dice entonces que el linfocito queda p r o g r a m a d o p a r a l al i s is.
2) Go lpe le ta l
El citoesqueleto del CTL se reorganiza, de modo que tanto el aparato de Golgicomo los granulosomas se sitúan en el polo celular que queda en contacto conla célula diana. Entonces, los gránulos se fusionan con la membranacitoplásmica, produciéndose la exocitosis de su contenido al estrecho espaciointercelular ("beso de la muerte").
3) Disoc iac ión del CTL
Antes de que se produzca la lisis de la célula diana, el CTL se separa,

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 13/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 13 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
probablemente ayudado por el hecho de que la LFA-1 vuelve al estado de bajaafinidad.
4) Dest r ucc ión de la cé lu la d iana
Parece ser que la destrucción de la célula diana puede ocurrir por varios
mecanismos, predominando uno u otro en función de la naturaleza de lasuperficie de la célula diana, y del grado de activación que haya alcanzado ellinfocito CTL.
Veamos algunas ideas actuales sobre el mecanismo de apoptosisdesencadenado por los linfocitos Tc.
Antes que nada, hay que aclarar el pape l de l a g r anz i ma B . Como ya
sabemos, se trata de una fragmentina, que induce fragmentación del ADN enmúltiplos de nucleosomas. Ahora bien, este efecto es indirecto: las fragmentinasno son DNAasas, sino proteasas. La Granzima B tiene una actividad parecida ala llamada enzima convertidora de interleucina 1-ß (ICE).
La ICE es una proteasa endógena con un papel central en la ruta endógena de laapoptosis. Dicha ruta se puede activar por una serie de estímulos externos(carencia de factores de crecimiento, ciertos virus, daño al ADN), que transformanla ICE inactiva a forma activa. Una vez activada, la ICE rompe proteolíticamente unsustrato aún desconocido, que induce las ADNasas, que fragmentarán el ADN.
Pues bien, se piensa que la granzima B secretada por el CTL (y que entra a la
célula diana a través de los canales de perforina) activa a la ICE o bien lasustituye para desencadenar el mecanismo de esta ruta apoptósica endógena.
Pero este no es el único mecanismo inductor de apoptosis por parte de los CTLs(de hecho, algunos clones de CTLs carecen de perforina y de granzimas, y sinembargo inducen muerte celular de la célula diana). Podemos encontrar otromecanismo dependiente de la interacción entre el llamado l i gando Fas del CTLy el receptor Fas de la célula diana.
Monómeros de per f o r i na contenida en los granulosomas llegan a lamembrana de la célula diana, y en presencia de Ca++ se polimerizan paraformar cilindros huecos de po l i pe r f o r i na (con unas 20 unidades), queatraviesan la bicapa lipídica. De este modo se forma un canal (con undiámetro de 5-20 nm) que es permeable a iones, y que puede provocar lalisis osmótica de la célula diana (aunque su papel principal es servir comocanal para dejar pasar a otros componentes de los gránulos).Pero no siempre el efecto de la poliperforina es la lisis. En otros casos pareceque el papel de los canales es el de permitir la entrada de otras sustanciasdel granulosoma que inducen la apop tos i s de la célula diana. De hecho seha visto que pueden entrar fragmentinas (granzimas) que inducen lafragmentación de los cromosomas en múltiplos de nuclesoma.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 14/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 14 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
La unión entre el ligando de Fas con el Fas de la célula infectada o enfermainduce en ésta una señal que dispara la ruta endógena de apoptosis, con laconsiguiente activación de la ICE y subsiguiente fragmentación del ADN.
Las vesículas del CTL pueden contener TNF- a y TNF- b ( = l i n f o tox i na ) , que junto con el IFN-g producido por TC o por otras células, desencadena efectos
citotóxicos que tardan más tiempo (>3-4 horas) que los anteriores. Sedesconoce el mecanismo, pero parece que igualmente inducen apoptosis.
¿Cómo se protege el CTL de sus propios mecanismos matadores? Por lo pronto,el condroitín sulfato (proteoglucano) del CTL se puede unir a la perforina,inactivándola. Pero puede que haya otros mecanismos: por ejemplo, la hipótesisde Peters dice que la perforina sale del granulosoma dentro de pequeñasvesículas igualmente membranosas; la membrana de estas sub-vesículascontendrían complejos TCR-CD3-CD8, que encaminarían a las pequeñasvesículas hacia la célula diana, pero no al CTL.
13.4.2 Citotoxicidad directa inespecífica mediada porcélulas NK
Recordemos que las células NK son un tipo de linfocitos granulares grandes (dehecho NK es casi sinónimo de LGL), que suponen un 5% de las células linfoidesen sangre, y que carecen de los marcadores de linfocitos T y B. Se considerancomo linfocitos "inespecíficos", pertenecientes al sistema de inmunidad natural.
El mecanismo citotóxico de las células NK es parecido al de los linfocitoscitolíticos (CTL), pero a diferencia de ellos, carecen de especificidad y dememoria, y no están restringidos por el MHC clásico.
Poseen un papel importante en los primeros días de una infección virásica, aleliminar células en las que el virus se esté multiplicando. Constituyen pues unmecanismo efector celular temprano, antes de que lleguen los linfocitos TC, yespecialmente preparado frente a ciertos virus que usan la estrategia de inducirun descenso del nivel de MHC-I
Recientemente se ha propuesto un mode l o de dos r ecep to r es para explicarcómo reconocen estas células inespecíficas a las células infectadas por virus otumorales. Se piensa que las NK reciben señales de dos receptores de modo quedistinguen entre células sanas y enfermas.
El receptor Fas (también llamado APO-1) es una proteína de membrana quepertenece a la llamada familia TNF de receptores de citoquinas.Por su lado, el ligando de Fas del CTL es una proteína de membrana queexhibe homologías de secuencia con TNF, y se une al Fas.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 15/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 15 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
Según este modelo, el receptor NKR-P1 se une a oligosacáridos deglucoproteínas de superficie de la célula enferma, procedentes deglucosilaciones anómalas. En principio, esta señal le indica a la NK que debeprepararse para matar a la célula con la que ha contactado, pero para ello debede ocurrir s i mu l t áneamen te una ausenc i a de seña l p r oceden te de o t r o
r ecep to r , e l Ly - 49 . El receptor Ly-49 de la célula NK se une a ciertos alelos de moléculas MHC-I.Cuando una célula NK se encuentra ante una célula normal, aunque reciba laseñal de NKR-P1, no la mata, ya que la interacción entre Ly-49 y las MHC-Ienvía una señal a la NK para que detenga su mecanismo matador. En cambio,muchas células enfermas (por virus o tumorales) presentan un bajo nivel deMHC-I, por lo que el mecanismo dependiente de NKR-P1 no se ve frenado.
La actividad matadora de NK se activa por IFN-a e IFN-b (liberado por muchascélulas enfermas al inicio de la infección), y por IL-12 (producido por
macrófagos). A su vez, la IL-12 y el TNF-a (también del macrófago) provocanque la NK secrete grandes cantidades de IFN-g , con lo que se mantienen a rayaciertos patógenos intracelulares hasta que se activen las células T. Como se ve,un nuevo rasgo que parece indicarnos de nuevo el carácter de línea defensivatemprana que intepretan estas células. De hecho, rara vez las células NKeliminan efectivamente el virus, pero lo mantienen a raya hasta que llegan lascélulas TC.
13.4.3 Citotoxicidad celular dependiente de anticuerpos
(ADCC)Existen varios tipos de células potencialmente citotóxicas que poseen receptorespara Fc de anticuerpos, y que pueden por lo tanto participar en la destrucciónde células diana (enfermas) o de helmintos recubiertos en ambos casos poranticuerpos. Los acontecimientos suelen tener lugar en las siguientes fases:
1. formación de inmunocomplejos (es decir, la célula enferma o el helminto serecubre de anticuerpos);
2. una célula agresora adecuada (p. ej., la NK) interacciona con la célulaenferma o el parásito a través de los anticuerpos, que se engarzan con elreceptor para Fc de la célula agresora;
3. finalmente, dicha célula agresora libera por exocitosis el contenido de susgránulos, y/o secreta productos tóxicos, que tienden a matar a la célulaenferma o parásito.
De este modo, células que son propiamente del sistema inmune natural, y por lotanto son inespecíficas, pueden llegar a destruir específicamente mediante elpuente de anticuerpos. De hecho, actúan como células efectoras finales delsistema humoral específico (es decir, pueden llegar a ser los "brazos armados" o

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 16/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 16 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
"verdugos" en una respuesta que se inició con la secreción de anticuerpos).
13.5 RESPUESTA DEHIPERSENSIBILIDAD DE TIPORETARDADO
La hipersensibilidad de tipo retardado (DTH) es una reacción inflamatorialocalizada que se induce por citoquinas secretadas por ciertas subpoblaciones delinfocitos TH (concretamente, de TH1) activados previamente por contacto conciertos tipos de antígenos, y que se caracteriza por el reclutamiento al focoinfeccioso de grandes cantidades de células inflamatorias, sobre todo demacrófagos.
Se llama retardada porque, al contrario que la inmediata, tarda 2 o 3 días en manifestarse.
La palabra hipersensibilidad alude en principio a efectos negativos sobre el propio hospedador,pero en muchos casos el efecto consiste en pequeños daños tisulares, siendo en general larespuesta beneficiosa, puesto que va encaminada a destruir a ciertos patógenos intracelulares.
13.5.1 Fases de la hipersensibilidad de tipo retardado
En la respuesta hipersensible de tipo retardado podemos distinguir una faseinicial de sensibilización en la que se produce el contacto inicial con el estímuloantigénico, y que no presenta síntomas, y una fase efectora, dependiente de uncontacto secundario con el antígeno, en la que se manifiestan tales síntomas yque es la tendente a eliminar al patógeno.
13.5.1.1 Sensibilización
La fase de sensibilización dura de una a dos semanas, tiempo durante el cual loslinfocitos TH se activan y expanden clonalmente tras el reconocimiento del
antígeno presentado por células presentadoras (APCs). Dichas célulaspresentadoras pueden ser:
Las células NK, monocitos y PMN neutrófilos poseen el receptor Fcg RIII(=CD16), de baja afinidad, que reconoce las subclases IgG1 e IgG3.Los eosinófilos posen Fcε R-I y Fcε R-II, estando especializados en destruir helmintos(p. ej., las esquistosómulas, que son las larvas de Schistosoma ).

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 17/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 17 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
Al final de esta fase se tiene pues una población expandida de linfocitos T CD4+ de memoria, que recirculan constantemente.
13.5.1.2 Fase efectora
Esta fase ocurre en tejido extralinfoide, y es la reacción de hipersensibilidad ensentido estricto, detectándose sus síntomas a las 24 horas del contactosecundario con el antígeno que provocó la sensibilización inicial, pero alcanza suapogeo a las 48-72 horas. En esta fase sale fibrinógeno al tejido, y se convierte
en fibrina, que junto con la acumulación de linfocitos T y de macrófagos haceque el tejido se hinche y se vuelva duro (esto es la base de los síntomas deinduración y edema) La podemos subdividir en tres etapas: desencadenamiento,inflamación y resolución.
Como se ve, esta respuesta tiene un claro valor adaptativo en la eliminación de
ciertos parásitos intracelulares, pero cuando el estímulo es demasiadopersistente, la DTH se vuelve destructiva para el hospedador. Este aspectoi n m u n o p a t o l ó g i c o se manifiesta en la llamada r eacc ión g r anu l om atosa: losmacrófagos, con los parásitos sin eliminar en su interior, se adhieren unos aotros, adquiriendo morfologías epitelioides; posteriormente estas célulasepitelioides se pueden fusionar entre sí, generando células gigantesmultinucleadas, que pueden llegar a desplazar al tejido normal, con lo que seforman nódu l os o t ubé r cu l os, que pueden conducir a necrosis del tejido.
células de Langerhans de la piel, que transportan el antígeno desde laepidermis hasta los ganglios linfáticos regionales, donde lo muestran a loslinfocitos TH;
macrófagos;(en humanos y otras especies) células del endotelio vascular.
En el desencadenami en to el linfocito T activado (que ahoradenominaremos como TDTH), ante el contacto secundario con el antígeno
presentado por una APC, secreta ciertas citoquinas, cuyas acciones dan pasoa la reacción de inflamación.La i n f l amac i ón se produce cuando las citoquinas secretadas reclutan y
activan a macrófagos y otras células inflamatorias hacia el sitio de infección,poniéndose en marcha una compleja serie de interacciones entre célulasinespecíficas del sistema inmunitario y citoquinas, con amplificación de larespuesta. Todo ello va encaminado a destruir al agente patógenointracelular.La reso luc ión se produce porque los macrófagos activados aumentan sucapacidad de eliminación del patógeno. Estos macrófagos son la principalcélula efectora de la hipersensibilidad retardada, pudiendo representar el90% de la población celular infiltrada.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 18/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 18 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
13.5.2 Interacción entre citoquinas y células en la DTH
En este epígrafe vamos a ampliar el aspecto molecular y celular de la respuestade hipersensibilidad retardada, resaltando cómo las distintas citoquinas actúansobre distintos tipos celulares, responsables de diversos aspectos de esta
respuesta.
aumento de la expresión de moléculas de adhesión (ICAM,ELAM, VCAM);
cambios de forma, tendentes a que las células endotelialesse separen entre sí, dejando paso para la extravasación deleucocitos;
las células endoteliales estimuladas secretan IL-8 y otrosfactores quimiotácticos que atraen a monocitos circulantes(quimiocinas).
Las consecuencias son que los PMN neutrófilos (primerosleucocitos en llegar) y los monocitos circulantes se adhierena las células endoteliales de las vénulas postcapilares, pasanpor diapédesis y entran al tejido infectado. La entrada de losmonocitos lleva consigo su diferenciación a macrófagos.
La IL-2 del linfocito TDTH funciona de modo autocrino, permitiendo la
expansión clonal, con lo que aumenta el número de estas células, que siguenproduciendo más citoquinas. Conforme avanza la respuesta, la IL-2 tambiénejerce un efecto paracrino, estimulando a otros clones de linfocitos TDTH.
La IL-3 y el GM-CSF inducen hematopoyesis del linaje granulocito-monocíticoen la médula ósea. Obviamente ello acarrea que mayores cantidades de PMNy macrófagos puedan pasar al foco de infección.El IFN-g el TNF-b de las células T, así como el TNF-a y la IL-1 de los
macrófagos actúan sobre las células endotelilales vasculares cercanas al focode infección, con los siguientes efectos:
Los macrófagos son atraídos y activados en el foco de infección debido a la
acción de IFN-g y del factor quimiotáctico y activador de los monocitos(MCAF). Otro factor secretado por TDTH, llamado MIF (inhibidor de la
migración de macrófagos) evita que el macrófago "pase de largo" el focoinfectivo, afinando su quimiotaxis.Por lo tanto, los macrófagos van llegando en grandes cantidades al focoinfeccioso, y se van activando (sobre todo por el interferón inmune),experimentando importantes cambios:

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 19/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 19 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13_ Inmunidad celular.ht 11/05/02
aumentan su tamaño
aumentan sus niveles de enzimas líticos en sus gránulos
aumentan su capacidad fagocítica (por opsonización con
IgG2a -en el ratón-)incrementan su efectividad matadora de microorganismosintracelulares
funcionan mejor como células presentadoras de antígeno,debido a que es mayor el número de moléculas MHC declase II en su membrana, porque secretan mayor cantidadde IL-1 y porque expresan más moléculas de adhesión comola B7 (coestimulatoria), ICAM-1 y LFA-3.
Este tipo de respuesta que se retroalimenta positivamente puede llegar aconvertirse en un "arma de doble filo", ya que puede degenerar en una respuestanegativa para el hospedador, una vez que se traspasa un umbral. Sin embargo, suvalor positivo se ponde de manifiesto en experimentos con ratones transgénicos
noqueados (K.O.) que tienen inactivado el gen del IFN-γ : son incapaces de matara Mycobacterium bovis , una bacteria contra la que los ratones normales sedefienden perfectamente.
13.5.3 Efectos de la hipersensibilidad retardadaEl efecto positivo de la DTH es que tiende a eliminar ciertos patógenos intracelulares y antígenos decontacto:
La DTH desempeña un papel importante en la defensa contra estos agentes, yaque el macrófago activado destruye mejor al patógeno, y el contenido de susgránulos, vertido a exterior, destruye células propias infectadas (en este últimopapel es importante la acción del TNF-b ). Obviamente, se produce cierto dañotisular, pero éste suele estar autolimitado y puede ser reparado ulteriormente.
El papel positivo de la DTH se puede comprobar "por contraste" considerando
El que los macrófagos actúen mejor ahora como APC supone que activan amás linfocitos TDTH, con lo cual se cierra el ciclo, pero amplificado.
bacterias intracelulares como Mycobacterium tuberculosis, M. leprae, Listeria monocitogenes, Brucella abortus.
Hongos intracelulares como Candida, Pneumocystis carinii, Histoplasma,Cryptococcus .Protozoos como Leishmania .Larvas de helmintos, como las de Schistosoma .
Virus.

5/14/2018 13. Inmunidad celular - slidepdf.com
http://slidepdf.com/reader/full/13-inmunidad-celular 20/20
CURSO DE INMUNOLOG A GENERAL: 13. Inmunidad celular Página 20 de 20
file://C:\WINDOWS\...\CURSO DE INMUNOLOGÍA GENERAL 13 Inmunidad celular.ht 11/05/02
aquellas enfermedades en las que precisamente este mecanismo está alterado:
por ejemplo, en el SIDA, en el que existen bajos niveles de linfocitos T CD4 +: elloacarrea que se pierda la facultad de montar una respuesta de hipersensibilidadretardada, por lo que los individuos afectados están sometidos a frecuentes yamenazadoras infecciones por Candida, Pneumocystis , Cryptococcus , o porbacterias patógenas e incluso oportunistas que no suelen causar tanta mortalidaden la población sana. De hecho, una de las maneras de seguir el curso del SIDA esevaluar periódicamente en laboratorio la capacidad de respuesta DTH con células
de los pacientes.
Sin embargo, en ciertos casos la propia respuesta defensiva degenera enpatológica: tal ocurre en la tuberculosis, en la que se pueden llegar a formarnódulos (tubérculos) en muchos tejidos (típicamente en los pulmones), quepueden terminar necrosándose (cavernas caseosas en pulmón).
En una situación de infección crónica por parásito intracelular (como en latuberculosis) las citoquinas producidas en la DTH estimulan la proliferación defibroblastos y la producción de colágeno, potenciándose los efectos por la acción
de PDGF (factor de crecimiento derivado de plaquetas) y de TGF-β del macrófagoactivado. Posteriormente emigran y proliferan células endoteliales (angiogénesis,
es decir, formación de nuevos vasos sanguíneos). Se produce pues la sustitucióndel tejido normal por tejido fibroso (cicatrización) y formación de granuloma.
Copyright © 1999 Enrique Iáñez Pareja. Prohibida la reproducción con finescomerciales.
[VOLVER AL PROGRAMA DE INMUNOLOGÍA]
[VOLVER A LA PORTADA DE ESTE SITIO WEB]
ENVIAR COMENTARIOS