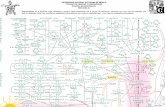
Integracion de Las Rutas Metabolicas
15
INTEGRACION DE LAS RUTAS METABOLICAS 1. GLUCOGENOGENESIS: La Glucosa se convierte en glucosa-6-fosfato mediante una reacción irreversible catalizada por la glucoquinasa o hexoquinasadependiendo del tejido en cuestión. glucosa + ATP → glucosa-6-P + ADP Glucosa-6-fosfato se convierte en glucosa-1- fosfato por la acción de la Fosfoglucomutasa, mediante la formación obligada de un compuesto intermediario, glucosa-1,6-bisfosfatasa. glucosa-6-P ←→ glucosa-1-P Glucosa-1-fosfato se convierte en UDP-glucosa por la acción de la UDP-glucosa pirofosforilasa (llamada también uridil transferasa). glucosa-1-P + UTP → UDP-glucosa + PPi Las moléculas de glucosa son acopladas en cadena por la glucógeno sintasa, este paso debe realizarse sobre un primer preexistente de glucógeno. La glucógeno sintasa actúa es decir formando alargamientos lineales de ramas preexistentes, solamente formando uniones α1- 4 permitiendo la union de glucosa a glucógeno preexistente Las ramificaciones son producidas por la enzima ramificadora del glucógeno, la cual transfiere un fragmento de 6 a 8 unidades del extremo no reductor y lo une a una glucosa por un enlace α-1,6. Esto posibilita que ambas cadenas puedan continuar alargándose mediante uniones α-1,4 de glucosas hasta poder producir nuevas ramificaciones. Es la vía generadora de glucógeno. 2. GLUCOGENOLISIS
-
Upload
lorena-reyes -
Category
Documents
-
view
188 -
download
9
Transcript of Integracion de Las Rutas Metabolicas

INTEGRACION DE LAS RUTAS METABOLICAS
1. GLUCOGENOGENESIS: La Glucosa se convierte en glucosa-6-fosfato mediante una reacción
irreversible catalizada por la glucoquinasa o hexoquinasadependiendo del tejido en cuestión.
glucosa + ATP → glucosa-6-P + ADP Glucosa-6-fosfato se convierte en glucosa-1-fosfato por la acción de
la Fosfoglucomutasa, mediante la formación obligada de un compuesto intermediario, glucosa-1,6-bisfosfatasa.
glucosa-6-P ←→ glucosa-1-P Glucosa-1-fosfato se convierte en UDP-glucosa por la acción de la UDP-
glucosa pirofosforilasa (llamada también uridil transferasa). glucosa-1-P + UTP → UDP-glucosa + PPi Las moléculas de glucosa son acopladas en cadena por la glucógeno
sintasa, este paso debe realizarse sobre un primer preexistente de glucógeno. La glucógeno sintasa actúa es decir formando alargamientos lineales de ramas preexistentes, solamente formando uniones α1-4 permitiendo la union de glucosa a glucógeno preexistente
Las ramificaciones son producidas por la enzima ramificadora del glucógeno, la cual transfiere un fragmento de 6 a 8 unidades del extremo no reductor y lo une a una glucosa por un enlace α-1,6. Esto posibilita que ambas cadenas puedan continuar alargándose mediante uniones α-1,4 de glucosas hasta poder producir nuevas ramificaciones.
Es la vía generadora de glucógeno.
2. GLUCOGENOLISIS
Es la vía por la cual se degrada glucógeno para la obtención de glucosa de una
forma rápida, esta vía se estimula por niveles bajos de glucosa, glucagon y
catecolaminas (adrenalina, noradrenalina y norepinefrina)
1. El glucogeno es degradado a glucosa 1 fosfato por la enzima glucogeno
fosforilasa que es la enzima reguladora de esta vía y la enzima
desramificante que rompe los enlaces alfa 1-4 y alfa 1-6
2. La glucosa 1 fosfato pasa a glucosa 6 fosfato por la enzima
fosfoglucomutasa
3. La glucosa 6 fosfato pasa a glucosa por la enzima glucosa 6 fosfatasa
3. GLUCOLISIS

eso se puede deducir de la formula de la glucolisis la cual es
Glucosa+2NAD(+) +2ADP+2Pi ----> 2piruvato+2NADH+2ATP+2H(+)+2H20
Los productos de esta reacción, mostrados arriba, son utilizados por la
célula en muchas funciones:
El ATP (Adenosín trifosfato) es la fuente de energía universal de la célula.
NADH y H+, otorgan la capacidad de reducir otros compuestos
pertenecientes a otras vías metabólicas, o bien para sintetizar ATP.
El piruvato es la molécula que seguirá oxidandose en el ciclo de Krebs,
como parte de la respiración aeróbica, donde dará origen a más moléculas
NADH, que podrán pasar a sintetizar ATP en la mitocondria.
1. El primer paso es HEXOQUINASA
Glucosa + ATP ----> Glucosa-6-fosfato + ADP
2do Paso: Fosfohexosa isomerasa
Glucosa-6-fosfato ---------> Fructosa-6-fosfato (este es un proceso
reversibible)
3er paso: Fosfofructoquinasa
Fructosa-6-fosfato + ATP ---> Fructosa-1,6-bifosfato + ADP
4to Paso: Aldolasa
Fructosa-1,6-bifosfato ----> Dihidroxiacetona-fosfato + Gliceraldehido-3-
fosfato (tmb reversible)
5to Paso: Triosa-fosfato-Isomerasa
Dihidroxiacetona-fosfato------> Gliceraldehido-3-fosfato (reversible)
6to Paso: Gliceraldehido-3-fosfato deshidrogenasa
7mo Paso: Fosfoglicerato quinasa
8vo Paso: Fosfoglicerato mutasa

9no Paso: Enolasa
10mo Paso: Piruvato quinasa
4. VIA PENTOSA FOSFATO
La ruta de la pentosa fosfato tiene una gran flexibilidad, de hecho, es
un módulo ideal del metabolismo, que se adapta continuamente a las
cantidades requeridas de ATP, NADPH, ribosa-5-
fosfato, piruvato o acetil-CoA, según las necesidades de la célula.
Esta ruta se ve regularizada mediante la enzima glucosa-6-fosfato
deshidrogenasa. El regulador más importante es la oferta deNADP+, el
cual actúa como activador alostérico, mientras que
el NADPH disminuye la actividad de la enzima como inhibidor
competitivo. En condiciones fisiológicas el NADPH se encuentra en
mayor proporción (70:1) respecto NADP+, si hubiese una utilización de
equivalentes de reducción conduciría rápidamente a la estimulación de
la deshidrogenasa debido al aumento de la cantidad de NADP+.
Consecuentemente, esta ruta metabólica transcurre fuertemente en
el tejido adiposo, donde hay una gran oferta de glucosa y una alta
necesidad de NADPH, requerido para la biosíntesis de ácidos grasos.
Por el contrario, en el tejido muscular, se encuentra una baja necesidad
de NADPH, por lo que se realiza la inversión de la ruta.
En el caso del tejido adiposo, se dará lugar a NADPH para las células
del tejido, pero, la formación de ribosa-5-fosfato no dará suficiente
síntesis de nucleótidos, hecho que provocará la conversión de las
pentosas en gliceraldehído-3-fosfato y fructosa-6-fosfato. Por lo
general, estas biomoléculas se incorporarán en la glucólisis, con la
ayuda de la enzima piruvato deshidrogenasa, para formar,
finalmente, acetil-CoA necesario para la síntesis de ácidos grasos. Así
pues, en la glucólisis simultáneamente se forman equivalentes de
reducción (NADPH, NADH) y también de energía (ATP). Este proceso
se detiene cuando ya hay suficiente y, además, se han cubierto las
necesidades de ATP. En este momento, los productos finales de la fase
no oxidativa de esta ruta metabólica podrán incorporarse en la
gluconeogénesis, para formar nuevamente glucosa-6-fosfato y cerrar el
ciclo.

Por último, hay otro tipo de células, las proliferantes que también se
aprovechan de la gran flexibilidad de este proceso metabólico. Éstas
necesitan una gran cantidad de ribosa-5-fosfato para poder sintetizar
ácidos nucleicos y, así, replicarse con facilidad y rapidez. De este modo,
la ruta puede invertirse, gracias a la reversibilidad de sus reacciones y, a
partir de una molécula de gliceraldehído-3-fosfato y dos de fructosa-6-
fosfato, obtendremos como producto tres moléculas de ribosa-5-
fosfato, sin formarse ningún NADPH.3
5. FERMENTACION LACTICA
En condiciones de ausencia de oxígeno (anaerobias), la fermentación
responde a la necesidad de la célula de generar la molécula deNAD+, que
ha sido consumida en el proceso energético de la glucólisis. En la glucólisis
la célula transforma y oxida la glucosa en un compuesto de tres átomos de
carbono, el ácido pirúvico, obteniendo dos moléculas de ATP; sin embargo,
en este proceso se emplean dos moléculas de NAD+ que actúan como
receptores de electrones y se reducen a NADH+H+. Para que puedan tener
lugar las reacciones de la glucólisis productoras de energía es necesario
reoxidar el NADH+H+; esto se consigue mediante la cesión de dos
electrones del NADH+H+ al ácido pirúvico, que se reduce a ácido láctico
6. FERMENTACION ALCOHOLICA

En esta fermentación a partir de la glucosa y con la participación de diferentes enzimas, se origina el ácido pirúvico, el cual es descarboxilado hasta CO2 y acetaldehído y este último reducido por la acción de la deshidrogenada hasta alcohol etílico.
7. CICLO DE KREBS
El ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo de los ácidos
tricarboxílicos) es una ruta metabólica, es decir, una sucesión de reacciones
químicas, que forma parte de la respiración celular en todas las células
aeróbicas En organismos aeróbicos, el ciclo de Krebs es parte de la vía
catabólica que realiza la oxidación de glúcidos, ácidos grasos y aminoácidos
hasta producir CO2, liberando energía en forma utilizable (poder reductor y
GTP).
Reacción 1: condensación del oxalacetato con la acetil CoA
La enzima citrato sintasa condensa a la acetil-CoA (2C) con el oxalacetato (4C)
para dar una molécula de citrato (6C). Como consecuencia de esta
condensación se libera la coenzima A (HSCoA). La reacción es fuertemente
exergónica: es irreversible.
• Reacción 2: isomerización del citrato a isocitrato

La isomerización del citrato en isocitrato ocurre por dos reacciones, que se
resumen en una.
• Reacción 3: oxidación y decarboxilación del isocitrato
El isocitrato es sustrato de la isocitrato deshidrogenasa, enzima que tiene como
cofactor un NAD, que forma parte de la cadena respiratoria. En la reacción 3 se
resumen dos reacciones a partir de las cuales el isocitrato forma α-
cetoglutarato (5C). Para lograr ese producto ocurre una decarboxilación, es
decir la liberación de una molécula de CO2, y la reducción de un NAD que
permite la formación de 3 ATP.
• Reacción 4: el α-cetoglutarato se transforma en succinil-CoA
Este paso implica la segunda decarboxilación oxidativa, catalizada por la α-
cetoglutarato deshidrogenasa, que lleva a la formación de succinil-CoA (4C). El
NAD es la coenzima de la deshidrogenasa, de manera que se formarán 3 ATP
como consecuencia de la actividad de cadena respiratoria.
• Reacción 5: la succinil-CoA rinde succinato y GTP
La succinil-CoA, es un tioéster de alta energía con un ∆G°′ de hidrólisis de -
33.5 KJ.mol-1 aproximadamente. La energía liberada por la ruptura de ese
enlace se utiliza para generar un enlace fosfoanhidro entre un fosfato y un GDP
para dar 1GTP por fosforilación a nivel de sustrato. En la reacción se libera
HSCoA.
El GTP se puede convertir en ATP según la siguiente reacción:
GTP + ADP GDP + ATP ∆G°′ = 0 KJ.mol- 1
• Reacción 6: el succinato se transforma en fumarato
El succinato es oxidado a fumarato por la succinado deshidrogenasa, enzima
que tiene como cofactor al FAD: se producen 2ATP en la cadena respiratoria.
La enzima usa FAD porque la energía asociada a la reacción no es suficiente
para reducir al NAD.
El complejo enzimático de la succinato deshidrogenasa es el único del ciclo
que está asociado a la membrana mitocondrial de eucariotas, y en la
membrana plasmática de procariotas.
• Reacción 7: el fumarato se hidrata y genera malato
La fumarasa cataliza la adición de agua, es decir la hidratación del fumarato. El
producto de la reacción es el malato.

• Reacción 8: el malato se oxida a oxalacetato
Dada la naturaleza cíclica de la vía, las reacciones en su conjunto conducen a
la regeneración del oxalacetato. La malato deshidrogenasa cataliza la
oxidación del malato a oxalacetato, con la reducción de un NAD: se forman 3
ATP en la cadena respiratoria.
7. CICLO DE LA UREA
El primer grupo amino que ingresa al ciclo proviene del amoníaco libre intramitocondrial. El amoníaco producido en las mitocondrias, se utiliza junto con el bicarbonato (producto de la respiración celular), para producir carbamoil-fosfato. Reacción dependiente de ATP y catalizada por la carbamoil-fosfato-sintetasa I. Enzima alostérica y modulada (+) por el N-acetilglutamato.
2. El carbamoil-fosfato cede su grupo carbamoilo a la ornitina, para formar citrulina y liberar Pi. Reacción catalizada por la ornitina transcarbamoilasa. La citrulina se libera al citoplasma.
3. El segundo grupo amino procedente del aspartato (producido en la mitocondria por transaminación y posteriormente exportado al citosol) se condensa con la citrulina para formar argininosuccinato. Reacción catalizada por la argininosuccinato sintetasacitoplasmática. Enzima que necesita ATP y produce como intermediario de la reacción citrulil-AMP.
4. El argininosuccinato se hidroliza por la arginino succinato liasa, para formar arginina libre y fumarato.
5. El fumarato ingresa en el ciclo de Krebs y la arginina libre se hidroliza en el citoplasma, por la arginasa citoplasmática para formar urea y ornitina.
6. La ornitina puede ser transportada a la mitocondria para iniciar otra vuelta del ciclo de la urea.
En resumen, el ciclo de la urea consta de dos reacciones mitocondriales y cuatro citoplasmáticas
8. TRANSAMINACION Y DESAMINACION DE LOS AMINOACIDOSLa transaminación es catalizada por transaminasas que transfieren el grupo amino desde el aminoácido a un cetoácido que normalmente es el -cetoglutarato. El resultado es otro cetoácido y el ácido glutámico (Glu). El objetivo de las reacciones de transaminación es precisamente obtener Glu, ya que este aminoácido puede intervenir en la siguiente etapa.
Mediante la desaminación oxidativa e Glu pierde su grupo amino, obteniéndosa amoniaco y regenerándose el -cetoglutarato. La reacción es catalizada por la

glutamato deshidrogenasa, que se encuentra en la matriz mitocondrial y precisa NAD+ como aceptor de electrones.
Por tanto, como resultado de los procesos de desaminación e obtienen aminoácidos desaminados y amoniaco.
10.CETOGENESIS
Cuando ocurre la oxidación de los ácidos grasos el acetil-CoA es excesivo lo
cual genera que esta ruta se active la cual sirve para disminuir el acetil-CoA
mediante la producción de cuerpos cetónicos que son usados para energía, la
síntesis de colesterol y otros como la acetona se desechan en la orina.
Principalmente ocurre en hígado.

11.B- OXIDACION DE ACIDOS GRASOS
se denomina así porque la oxidación ocurre en la posición del C3 o beta del
grupo acilo graso, al unirse la coenzima A con el grupo carboxílico en el C1,
dicha oxidación ocurre en las mitocondrias, y se producen en tres fases:
1) La Beta oxidación propiamente dicha.
2) Oxidación de los acetil CoA a CO2 a través del ciclo del acido cítrico.
3) Los productos generados (transportadores electrónicos reducidos NADH y
FADH2) tanto de la beta oxidación como del ciclo del acido cítrico donarán sus
electrones a la cadena respiratoria mitocondrial con acción del ADP para
generar agua.
Son 4 pasos básicos y las enzimas que participan en cada paso, a
continuación:
1) Deshidrogenación: Acil-CoA deshidrogenasa.
2) Hidratación: Enoil-CoA hidratasa.
3) Deshidrogenación: Beta hidroxiacil-CoA deshidrogenasa.
4) Acción de una tiolasa: Acil CoA acetiltransferasa (tiolasa).
Luego suponiendo para el palmitil-Coa que tiene 16 carbonos, luego de estos 4
pasos básicos se genera para su oxidación completa 7 productos de Acetil-
CoA, esto quiere decir que se repite los 4 pasos mencionados 6 veces

12. GLUCONEOGENESIS
Piruvato carboxilasa es un enzima mitocondrial, mientras que el resto de
enzimas de la gluconogénesis son citosólicos: Se debe transportar el
oxalacetato producido fuera de la mitocondria:
1. Oxalacetato es reducido a malato por una malatodeshidrogenasa
mitocondrial ligada a NADH
2. Malato es transportado al citosol por el sistema lanzadera malato-
aspartato
3. Una vez en el citosol, el malato es reoxidado a oxalacetato por una
malato deshidrogenasa citosólicaligada a NAD+ Oxalacetato es �descarboxilado y fosforilado simultaneamente por
FOSFOENOLPIRUVATO CARBOXIQUINASA (PEP
carboxiquinasa). La hidrólisis del GTP y liberación de CO2desplazan al
reacción hacia la formación de PEP.
Conversión de Fructosa-1,6-bifosfato en Fructosa-6-fosfato
Una vez formado, el Fosfoenolpiruvato es metabolizado por los enzimas �de la glicolisis pero en sentido inverso (reacciones en equilibrio).
El siguiente paso irreversible es la hidrólisis de Fructosa-1,6-bifosfato en �Fructosa- 6-fosfato y Pi:

13. FOSFORILACION OXIDATIVA

Se forman 2 NADH en la glicolisis: cada uno produce 1.5 ATP (suponiendo el
transporte de NADH por la lanzadera de glicerol fosfato).
Se forman 2 NADH en la descarboxilacion oxidativa del piruvato; cada uno
produce 2.5 de ATP.
Se forman FADH2 en el ciclo de acido cítrico; cada uno produce 1.5 de ATP.
Se forman 6 NADH en el ciclo del acido cítrico; cada uno produce 2.5 de ATP.
14. SISTEMAS LANZADERA
el NADH+H+ formado en el citosol, no puede atravesar directamente la
membrana mitocondrial interna, pero los electrones pueden incorporarse a la
cadena respiratoria mitocondrial por rutas indirectas llamadas lanzaderas de
sustrato. Se conocen dos tipos:
LANZADERA DEL GLICEROL FOSFATO. Los electrones del NADH+H+
citosólico pasan al FADH2 mitocondrial, con lo que la cadena respiratoria sólo
rendirá 2 ATP Se encuentra por ejemplo en células del cerebro y del músculo
esquelético
LANZADERA MALATO-ASPARTATO. Los electrones del NADH+H+ citosólico
pasan al NADH + H+ mitocondrial, con lo que la cadena respiratoria rendirá 3
ATP Se encuentra por ejemplo en células del corazón y del hígado.



















