
ESTUDIOS EPIDEMILÓGICOS Y TRANSMISIÓN DE AISLADOS DE...
66
1 1. INTRODUCCIÓN Los cítricos son uno de los principales cultivos en el mundo, liderando la producción mundial de frutos primarios, la que alcanzó a 103 millones de toneladas métricas el año 2003, superando con creces la producción de bananos la que llegó a 69 millones de toneladas métricas el mismo año (FAO, 2004). Este nivel de producción, sus beneficios económicos, efectos sociales y además las propiedades alimentarias y organolépticas, hacen de los cítricos uno de los cultivos mas estudiados, motivando la preocupación de numerosos investigadores en todo el mundo, ante la aparición de nuevas plagas y enfermedades que afecten a este cultivo. Entre las enfermedades importantes, la tristeza de los cítricos es la que ha causado más muertes de árboles de este género en el mundo (CAMBRA et al., 2000). En Chile la producción anual de frutos cítricos es de 268.000 toneladas métricas, con una superficie plantada de 16.100 hectáreas (FAO, 2004), sin considerar mandarinos e híbridos, los cuales según el censo agropecuario de 1997 llegan a 1267 ha (GARDIAZABAL y MAGDHAL, 1998), cifra que probablemente en la actualidad se ha duplicado. El virus de la tristeza de los cítricos (CTV), se trasmite de árboles enfermos a árboles sanos por injertos y por algunas especies de pulgones (CAMBRA et al., 1990), y es el causante de la enfermedad viral mas grave de los cítricos, conocida internacionalmente como tristeza, que empezó a manifestarse en forma epidémica en distintos países a partir de 1930, y hoy en día desde el punto de vista económico, es una de las mas destructivas e importantes que afecta a este tipo de cultivo (CAMBRA y MORENO, 2000).
Transcript of ESTUDIOS EPIDEMILÓGICOS Y TRANSMISIÓN DE AISLADOS DE...

1
1. INTRODUCCIÓN
Los cítricos son uno de los principales cultivos en el mundo, liderando la producción
mundial de frutos primarios, la que alcanzó a 103 millones de toneladas métricas el
año 2003, superando con creces la producción de bananos la que llegó a 69 millones
de toneladas métricas el mismo año (FAO, 2004).
Este nivel de producción, sus beneficios económicos, efectos sociales y además las
propiedades alimentarias y organolépticas, hacen de los cítricos uno de los cultivos
mas estudiados, motivando la preocupación de numerosos investigadores en todo el
mundo, ante la aparición de nuevas plagas y enfermedades que afecten a este cultivo.
Entre las enfermedades importantes, la tristeza de los cítricos es la que ha causado
más muertes de árboles de este género en el mundo (CAMBRA et al., 2000).
En Chile la producción anual de frutos cítricos es de 268.000 toneladas métricas, con
una superficie plantada de 16.100 hectáreas (FAO, 2004), sin considerar mandarinos
e híbridos, los cuales según el censo agropecuario de 1997 llegan a 1267 ha
(GARDIAZABAL y MAGDHAL, 1998), cifra que probablemente en la actualidad se
ha duplicado.
El virus de la tristeza de los cítricos (CTV), se trasmite de árboles enfermos a árboles
sanos por injertos y por algunas especies de pulgones (CAMBRA et al., 1990), y es el
causante de la enfermedad viral mas grave de los cítricos, conocida
internacionalmente como tristeza, que empezó a manifestarse en forma epidémica en
distintos países a partir de 1930, y hoy en día desde el punto de vista económico, es
una de las mas destructivas e importantes que afecta a este tipo de cultivo (CAMBRA
y MORENO, 2000).

2
Importantes epidemias de tristeza han destruido millones de árboles de cítricos
injertados sobre naranjo agrio (citrus aurantium (L) Osbeck). Hasta la fecha se
calcula en más de 80 millones, desde 1930, los árboles de naranjo dulce, mandarino y
pomelo injertado sobre naranjo amargo, que han muerto debido al virus de la tristeza
en todo el mundo (CAMBRA y MORENO, 2000).
En muchas regiones donde CTV es endémico (Asia, América del Sur, Australia,
África Central y del Sur), las razas muy severas pueden debilitar y destruir la
citricultura independiente del patrón usado. Además, aquellas áreas donde Toxoptera
citricida (Kirk.) no está presente aún, enfrentan el peligro de su posible introducción
(ROISTACHER y MORENO, 1991).
Aun existen en el mundo áreas productoras de cítricos en las cuales el patrón
predominante es naranjo agrio (citrus aurantium), que es extremadamente
susceptible. Por otra parte los productores y viveristas siguen importando material de
propagación no certificado, con el peligro de la introducción de razas severas que
pueden ser eficientemente transmitidas por áfidos locales y no necesariamente por T.
citricida (ROISTACHER y MORENO, 1991).
CTV ha sido detectado en árboles aislados y aún no tiene carácter epidémico en
muchos países del Mediterráneo y Próximo Oriente (Albania, Argelia, Chipre,
Egipto, Francia, Grecia, Italia, Jordania, Líbano, Libia, Marruecos, Palestina,
Portugal, Siria, Túnez y Turquía) y algunos de América Central y del Sur como
Belice, Chile, Ecuador, El Salvador, Honduras, México o Nicaragua (CAMBRA y
MORENO, 2000).
En Chile, de acuerdo a prospecciones realizadas entre las regiones I y VII, se han
encontrado muestras positivas de CTV principalmente en la zona del Oasis de Pica,
sólo en algunos huertos del resto de las regiones, y no se ha determinado la presencia

3
de Toxoptera citricida. De acuerdo con lo anterior, Chile, es un país en donde no se
han encontrado razas severas del virus en las principales zonas productoras, y
tampoco su principal vector Toxoptera citricida (CANALES et al., 2001).
Por lo tanto, es muy importante mantener un control en las zonas limítrofes más
propensas al ingreso del principal vector de CTV, y a material vegetal que pueda
contener razas del virus más severas que las descritas en Chile, además de mantener
al país libre de numerosas plagas y enfermedades presentes en el resto del mundo.
Los datos obtenidos en las prospecciones, han permitido localizar la presencia de
CTV y analizar su incidencia a lo largo de la zona citrícola del país (BESOAIN et
al., 2003a). No obstante la dinámica de dispersión, la relación existente con los áfidos
que predominan en los huertos de cítricos de Chile, y la eficiencia de transmisión de
razas de CTV presentes en el país no se conocen.
Por lo tanto, este trabajo busca determinar un patrón de distribución de la enfermedad
en el huerto, asociarlo con los áfidos portadores de CTV que visitan los predios de
cítricos de Chile, y determinar la capacidad de Aphis gossypii (Glover) de transmitir
razas del virus encontradas en Chile.
Estos estudios permitirán recopilar información acerca del carácter real de la
enfermedad en Chile, y con ello poder predecir, prevenir o controlar un posible mayor
desarrollo de esta enfermedad en el país.
De acuerdo con lo expuesto, los objetivos del presente trabajo son:
Realizar estudios espacio-temporales de la distribución de CTV en tres predios de
cítricos en la zona central de Chile.

4
Determinar mediante la metodología de brotes pegajosos, la presencia de especies
de áfidos virulíferos que visitan predios de cítricos.
Realizar transmisión de aislados mediante Aphis gossypii (Glover) a plantas
receptoras.

5
2. REVISIÓN BIBLIOGRÁFICA
2.1. Antecedentes generales del virus de la tristeza de los cítricos
La enfermedad ha recibido diversos nombres que hacen mención a las características
sintomatológicas encontradas. Los más importantes son: “graft incompatibily” y
“stunt-bush” en Sudáfrica (1904), “podredumbre de las raicillas” en Argentina
(1931), “quick decline” en California (USA) (1939), “bud-union decline” en
Australia (1941), tristeza en Brasil (1942), “hassaku dwarf “ en Japón (1948), “stem
pitting of grapefruit” en Sudáfrica (1949), “lime disease” o “lime decline” en Costa
de Oro, Ghana (1949), “seedling yellows” en Australia (1952) (BOVE y VOGEL,
1975, citados por NARANJO, 1997a).
La tristeza comenzó a expresarse producto del uso masivo de naranjo agrio como
patrón tolerante a otras enfermedades, principalmente Phytophthora spp., que
provoca la gomosis del cuello de la raíz del naranjo dulce (Citrus sinensis (L)
Osbeck) (BAR-JOSEPH et al., 1981).
En Australia, en 1860, y en Sudáfrica, a finales de 1890, se intentó injertar naranjos
dulces y mandarinos (Citrus reticulata Blanco) sobre naranjo agrio y se observó que
los árboles morían en un plazo de dos a tres años, en tanto que los limoneros (Citrus
limon (L) Burn.) sobre agrio crecían satisfactoriamente con desarrollo vegetativo y
producciones normales (WEBBER, 1943, citado por NARANJO, 1997a).
A partir de 1930 comenzaron en distintos países epidemias que causaron el
decaimiento de los cítricos, principalmente sobre naranjo amargo. Entre las más
importantes por el número de árboles afectados están las de Argentina (1930), Brasil

6
(1937), California (1939), Florida (1951), España (1957), Israel (1970) y Venezuela
(1980) (CAMBRA y MORENO, 2000).
La problemática que presentó esta enfermedad al manifestarse en forma masiva,
ocasionando la muerte de millones de árboles, llevó a la creación de la Organización
Internacional de Virólogos de Cítricos (IOCV), integrada por numerosos
investigadores de todo el mundo, cuyos trabajos e investigaciones son innumerables.
Esto denota la importancia y la trascendencia que ha tenido esta enfermedad
(ROISTACHER y MORENO, 1991).
En 1949, HUGHES y LISTER citados por NARANJO (1997a), describieron los
síntomas de “stem pitting” y aclareo de nervaduras de las hojas en plantas de lima
mexicana, coincidiendo con ellos investigadores de Brasil, los que sugirieron la
utilización de la misma como planta indicadora. A partir de esta época se hizo posible
diagnosticar cuando un árbol tiene tristeza.
Continuando con investigaciones sobre CTV, en 1979, gracias a los trabajos de
colaboración entre científicos de Israel, EEUU y España, se logró poner a punto el
ensayo Inmunoenzimático (ELISA) para la detección del virus, el cual posee una
importancia trascendental (BAR-JOSEPH et al., 1979; CAMBRA, MORENO y
NAVARRO, 1979, citados por CAMBRA et al., 1991).
2.1.1 Origen y diseminación
La enfermedad de la tristeza es originaria de Asia desde donde se extendió a las
principales zonas citrícolas de todo el mundo, mediante la introducción de material
vegetal (CAMBRA et al., 1990). Una vez instalada la enfermedad en una nueva zona,
los áfidos presentes en ella se han encargado de difundirla a más corta distancia
(CAMBRA, 1994).

7
Existen evidencias que indican que el virus pudo ser introducido desde India a
Sudáfrica en el siglo XVII por comerciantes holandeses, y desde ahí se diseminó a
otros países de África y Sudamérica. La introducción del virus a Brasil se produjo
desde Argentina y también directamente desde África del Sur, conjuntamente con
esta se introdujo T. citricida (WALLACE et al., 1956; COSTA, 1956, citados por
NARANJO, 1997a).
En España el virus de la tristeza fue introducido probablemente en la década de los
años treinta, pero fue detectado mucho más tarde, después de su manifestación en
forma epidémica en 1957 (CAMBRA et al., 1990).
CTV fue informado por primera vez en Israel (1956) en plantaciones de limonero
“Meyer” y desde 1970 comenzó a propagarse en los campos de forma natural
(RACCAH, LOEBENSTEIN y BAR-JOSEPH, 1976).
En Estados Unidos CTV fue detectado en la década de los años 50, y al igual que en
los casos de España e Israel esta enfermedad no ha provocado la muerte fulminante
de la citricultura sobre patrón agrio, si lo comparamos con los casos de Argentina y
Brasil (MORENO, 1995).
2.1.2 Distribución e importancia mundial
CTV está presente en todos los países citrícolas, si bien la incidencia es muy variable,
es prácticamente endémico en la mayor parte de Asia, sur de África, Australia y
Sudamérica; tiene elevada incidencia en las principales zonas citrícolas de Estados
Unidos, Israel y España y está dispersándose activamente en varios países de América
Central como Panamá y Costa Rica, e islas del Caribe como República Dominicana,
Jamaica o Cuba. (CAMBRA y MORENO, 2000).

8
Es importante mencionar que en la actualidad existen zonas productoras de cítricos en
el Mediterráneo, en América Central o Islas del Caribe donde el naranjo agrio es el
patrón predominante. Esta es una grave amenaza que enfrentan estos países si se
considera que CTV ya esta presente en el Caribe, España y países de América Central
(ROISTACHER y MORENO, 1991).
Se calculan unas pérdidas de 18 millones de árboles en Argentina, 10 millones en
Brasil, 3-4 millones en California, 35 millones en España, 5 millones en Florida, y 4
millones en Venezuela. Una estimación conservadora debería elevar al menos a 80
millones el número de árboles sobre patrón amargo muertos en los últimos 70 años
(CAMBRA y MORENO, 2000).
Unido ha esto se incorpora un elemento más, la imposibilidad de usar el naranjo agrio
como patrón en la mayor parte del mundo, ya que al usarlo como porta injerto de
naranjo dulce, mandarinos o pomelos, CTV es letal. Este patrón, sin lugar a dudas, es
el de mejores cualidades de adaptación a las diferentes condiciones ecológicas,
además posee extremada resistencia a Phytophthora spp., es resistente a la mayor
parte de las virosis de los cítricos y produce frutos de elevada calidad (NAVARRO,
1981, citado por NARANJO, 1997a).
2.1.3 Evolución de la enfermedad en Chile y situación actual
En Chile, el virus de la tristeza de los cítricos está presente desde hace varias décadas,
situación que fue confirmada por una prospección que se hizo en el país entre los
años 1967 y 1970, en cooperación entre la Universidad de Chile y la Universidad de
California (SÁNCHEZ, 1996). Debido a esta situación, en Chile, se prohíbe la
propagación de naranjo dulce sobre naranjo agrio, por la susceptibilidad a CTV que
confiere esta combinación (CHILE, MINIAGRI, 1968).

9
En Chile, en 1969, WEATHERS, SÁNCHEZ y PLATT indican que CTV está
presente en limoneros, cv. Meyer, en todo el país, como así mismo los áfidos vectores
del virus. Estudios preliminares revelaron que habían más de 2000 árboles de
limoneros de la variedad Meyer distribuidos a través de Chile y que cada uno de ellos
estaba afectado con el virus de la tristeza.
En ensayos realizados por SÁNCHEZ y WEATHERS (1970), el virus de la tristeza
fue encontrado en varios árboles de tangelo Sampson en un huerto en la zona de
Peumo, y también en tres limoneros Génova, dos limoneros Rugosos y dos limas
Bears en un huerto de Isla de Maipo, estos árboles fueron importados a Chile desde
Estados Unidos en 1942 y 1947 respectivamente.
Durante la temporada 1994/95, se observaron en la zona central de Chile, plantas de
limoneros injertadas sobre patrón C. macrophylla que mostraban una disminución de
crecimiento, clorosis, concavidades y gomosis en el patrón. Las plantas que
presentaban clorosis generalizada reaccionaron en forma positiva el test de ELISA
indirecto, confirmándose la presencia de partículas tipo closterovirus en muestras de
corteza de plantas afectadas (HERRERA, MADARIAGA y SANTELICES, 1995).
BESOAIN (1998) y BESOAIN et al., (1997), en un diagnóstico de enfermedades
causadas por virus y viroides en cítricos, detectan mediante serología e indexaje
biológico la presencia de aislados de tristeza en árboles de lima mexicana, limonero,
naranjo dulce, tangelos y pomelos, en huertos ubicados desde la I Región hasta la VI
Región. Este mismo estudio revela que hasta esa fecha no se ha observado
decaimiento rápido (DI) en plantas de naranjo dulce injertadas sobre naranjo agrio, ni
acanaladuras de la madera (SP) en variedades de pomelo.
Al parecer, las razas severas de CTV no se encuentran en Chile, sin embargo se
requiere un estudio más profundo, con una combinación de diferentes plantas

10
indicadoras (BESOAIN et al., 2000). No obstante, durante el año 2001, al observar
dos predios en Pica afectados por este virus se detectó la presencia de pomelos Star
Ruby injertados sobre C. macrophylla con síntomas de acordonamiento de árboles y
acanaladuras en la madera (BESOAIN et al., 2002).
Según la “Prospección de razas severas y áfidos vectores del virus de la tristeza de los
cítricos, presentes entre la I y VII Región de Chile", la incidencia de CTV estimada
por región fue de: 15,3% (I Región), 0,199% (III Región), 0,004% (IV Región),
0,079% (V Región), 0,183% (R. M.), 0,371% (VI Región), 0% (VII Región)
(BESOAIN et al., 2003a).
2.2. Agente causal y propiedades
El virus que causa los distintos síndromes de Tristeza (CTV) pertenece a los
closterovirus (virus filamentoso), con partículas flexuosas de unos 2000 nanómetros
de longitud y 10 – 12 de diámetro. Los viriones de CTV tienen dos proteínas
capsídicas de 25 y 27 kDa aproximadamente, que cubren el 95 y el 5 % de la longitud
de la partícula, respectivamente, dando un aspecto similar al de una serpiente
cascabel. Su genoma está constituido por ácido ribonucleico de cadena simple
(ssRNA) y polaridad positiva de 19226 – 19296 bases, con 12 marcos de lectura
abierta y capacidad para codificar al menos 19 proteínas. Se han descrito numerosos
aislados de CTV que difieren en sus características biológicas, serológicas, perfil de
ARNs bicatenarios, o secuencia de ARN genómico (CAMBRA y MORENO, 2000).
2.2.1 Distribución y movimiento del patógeno en la planta
Se plantea que la mayor concentración de partículas está en el pedúnculo de los frutos
y también en la corteza de los brotes jóvenes. Esta concentración por gramo de

11
corteza aumenta hasta los 6 meses tras inocular y luego disminuye (LEE et al., 1987
citado por NARANJO 1997a).
En plantas inoculadas por pulgones o injerto, las partículas se detectan a los 16 – 28
días, período análogo al necesario para recuperar inóculos. No obstante, se requieren
de 8 a 14 días para que el injerto infecte al patrón, dependiendo de las características
inherentes a la combinación injerto/patrón y de las condiciones que reúna el hábitat,
específicamente la temperatura (ROCHA-PEÑA et al., 1993; PRICE, 1966, citados
por NARANJO 1997a).
2.2.2 Rango de hospederos
BOVE y VOGEL (1980) plantean que, aunque todas las especies de cítricos son
hospederas de la enfermedad, la expresión de los síntomas se produce en forma
diferencial, lo que ha llevado a usar algunas especies como indicadoras de las
distintas razas de CTV.
También existen otras especies, fuera de los cítricos que son hospederos de CTV.
MULLER en 1974, logró transmitir CTV por medio de T. citricida, desde de naranjo
dulce a plantas de Passiflora gracilis, la cual es una planta indicadora herbácea, que
muestra síntomas de enanismo y clorosis de las hojas. Otras especies de Passiflora
también han sido utilizadas para observar síntomas específicos de CTV
(ROISTACHER y BAR-JOSEPH, 1987).
El virus también ha sido transmitido a algunas especies de géneros afines a Citrus
como: Aegle, Aeglopsis, Afraegle, Atalantia, Citropsis, Clausena, Eremocitrus
Hesperthusa, Merrillia, Microcitrus, Pamburus, Pleiospermium, Swinglea.
(CAMBRA y MORENO, 2000).

12
2.2.3 Razas del virus
CTV tiene gran diversidad de cepas, las cuales difieren en cuanto al rango de
actividad biológica, desde las razas más suaves hasta las más severas, y son causantes
de un amplio espectro de síntomas. Actualmente se han descrito una gran diversidad
de nuevos aislados de carácter severo (ROISTACHER y MORENO, 1991):
- Aislados de “stem pitting” en Sudáfrica, Australia y Japón.
- Aislado severo (B- 12) que produce “stem pitting” en naranjo dulce reportado desde
California.
- Aislado de stem pitting en naranjo dulce destruyendo la producción de naranjo
‘Navel’ en Perú y Australia, donde además afecta a las plantaciones de tangor
‘Ortanique’ (Citrus reticulata x Citrus sinensis).
- Aislado severo de tristeza afectando a naranjos dulces sobre naranjo agrio, en
Florida.
- Raza muy severa, llamada “Macapo”, capaz de inducir síntomas fuertes en naranjo
‘Valencia’ en condiciones de campo.
En presencia de cepas muy virulentas es prácticamente inviable el cultivo de naranjos
dulces y pomelos (NAVARRO, 1981, citado por NARANJO, 1997b). En Israel se
conoce un tipo destructivo de tristeza capaz de matar a los árboles injertados sobre
naranjo agrio aún antes de que el virus sea detectado por un método indicador rápido
(RACCAH, 1976).

13
MORENO et al. (1991), citados por (NARANJO 1997b) demostraron
fehacientemente que muchas entidades genéticas pueden estar presentes en un simple
aislado, además de que lograron su segregación o separación física mediante análisis
de dsRNAs. Ellos plantean que la presencia de diferentes razas del virus en una
simple planta puede estar debido a mutaciones de la raza original al pasar el tiempo, a
inoculaciones sucesivas por áfidos que cargan el virus desde otras fuentes, o más
comúnmente a ambos factores.
2.2.4 Sintomatología
Existen dos síndromes característicos de la enfermedad, que son regulados por genes
independientes:
- El primero es el llamado DI (Decline inducing) o decaimiento rápido, el cual
produce una gama de síntomas, lo más generalizado es un decaimiento lento,
acompañado de clorosis, defoliación de los árboles, asociado a pudrición de raicillas,
se observa hojas abarquilladas dentro del nervio central, disminución del vigor y
muerte progresiva de ramillas. En segundo lugar este síndrome provoca una
disminución del tamaño del árbol acompañado de frutos muy pequeños, con
maduración temprana y de mala calidad. La tercera manifestación de este síndrome
conocida como “quick decline” o colapso rápido es la más espectacular, los árboles
aparentemente sanos comienzan a mostrar un marchitamiento de las hojas similar al
producido por falta de agua y en cuestión de una o dos semanas mueren totalmente
(BESOAIN, 1998; ROCHA-PEÑA et al., 1995).
- El segundo síndrome es el denominado punteaduras del tallo (stem pitting, SP), que
produce, especialmente en variedades de pomelo y limas, acanaladura a nivel de la
madera con proyecciones puntiagudas de la corteza, las que calzan perfectamente con
las cavidades a nivel del xilema, lo que induce a un decaimiento y clorosis del follaje

14
con pérdida de productividad de las plantas afectadas (BESOAIN, 1998; ROCHA-
PEÑA et al., 1995). Estos aislados de CTV que causan SP severo predominan en la
mayoría de las áreas donde se cultiva cítricos en el mundo (GOTTWALD et al.,
1999).
El síntoma de Seedling yellows fue considerado por mucho tiempo como un tercer
síndrome, sin embargo hoy se sabe que ambos tipos de razas (DI y SP) pueden
producirlo (ROCHA-PEÑA et al., 1995).
2.2.5 Clasificación de especies y variedades por su sensibilidad
CTV manifiesta diferentes expresiones de síntomas dependiendo del cultivar y del
tipo de raza de que trate, es por ello que se han hecho diversas clasificaciones con
respecto a la susceptibilidad o tolerancia de las especies cítricas al CTV.
CARRERO (1981) y NAVARRO (en el mismo año), citados por NARANJO 1997b,
establecieron la siguiente clasificación, según el comportamiento de las diferentes
variedades ante el CTV:
- Variedades hipersensibles: naranjo agrio, limoneros y algunas variedades de
pomelos.
- Variedades muy sensibles: lima ‘Mexicana’, lima dulce de ‘Palestina’, cidro
‘Etrog’, C. hystrix, C. excelsa, C. macrophylla y citrange ‘Uvalde’.
- Variedades sensibles: Pomelos, ‘Kumkuat’, algunos tangors (‘temple’), naranjos
dulces (‘Pera’, ‘Mediterraneo’, ‘Hassaku’) y citranges (‘Savage’, ‘Morton’ y ‘Rusk’).

15
- Moderadamente tolerantes: Los restantes naranjos dulces, lima ‘Rangpur’, citrange
‘Troyer’, P. Trifoliata y limón ‘Rugoso’.
2.3. Formas de transmisión de CTV
El CTV se difunde de árboles enfermos a sanos mediante injertos y por áfidos en
forma semipersistente (CAMBRA, 1994).
No obstante, se debe señalar al hombre como el principal vector de la enfermedad a
larga distancia mediante el tráfico no controlado de material vegetal infectado
(CAMBRA et al., 1990).
GARNSEY y MULLER (1986), han realizado ensayos in vitro de transmisión
mecánica comparando distintos tipos cortes e injertos, sobre tallos y hojas de plantas
de diferentes edades.
2.3.1 Transmisión por injerto
Esta forma de transmisión es la causa de introducción de nuevas razas del virus en un
área citrícola y de la dispersión de la enfermedad a larga distancia, como
consecuencia de importaciones no controladas o clandestinas de nuevas variedades
(CAMBRA y MORENO, 2000).
CTV es fácilmente transmitido por injerto, usándose con éxito tejidos de yemas y
trozos de hojas como inóculo. Sin embargo, el porcentaje de transmisión por injerto
va a depender de: las combinaciones donante-receptor y virus-hospedero, del tipo de
tejido infectado, de la madurez del mismo, condiciones ambientales, entre otros
(ROCHA-PEÑA, 1993; GARNSEY y MULLER 1986).

16
2.3.2 Transmisión por medio de vectores
Una vez introducido en un área citrícola, a nivel local, CTV puede ser difundido a
árboles próximos por varias especies de áfidos (MORENO, 1995).
Existen muchas especies de áfidos con la capacidad de adquirir CTV, pero sólo siete
de ellas tienen capacidad para transmitir esta enfermedad. Toxoptera citricida, Aphis
gossypii, T. aurantii, A. spiraecola han sido descritos como vectores de CTV en
muchas zonas citrícolas (CAMBRA y MORENO, 2000).
El período de alimentación que requieren los áfidos para adquirir CTV puede ir de
unos segundos a 60 minutos. Igualmente el período mínimo que un pulgón virulífero
requiere para infectar a una planta mientras se alimenta de ella, es relativamente corto
(CAMBRA, 1994). Sin embargo pueden permanecer infecciosos durante 24 a 48
horas (HERMOSO DE MENDOZA, BALLESTER-OLMOS y PINA, 1984).
En el proceso de transmisión del virus, su velocidad y eficiencia tienen una marcada
influencia: la especie de áfido y su estadío, número de insectos, la raza del virus,
planta donante y receptora, temperatura, irrigación, lluvias repentinas y fertilización
(YOKOMI, 1995; HERMOSO DE MENDOZA, BALLESTER-OLMOS y PINA
1986).
2.3.2.1 Principales vectores
Los áfidos que más se destacan por su eficiencia de transmisión son el pulgón pardo
de los cítricos (T. citricida Kirk.) y el pulgón del melón (A. gossypii Glover), los
cuales difieren notablemente en su relación con el cultivo y su capacidad transmisora
de CTV (MORENO, 1995).

17
Los cítricos no son el hospedero principal en el caso de A. gossypii, por lo tanto
cuando hay otros cultivos vecinos migran y no logran formar grandes colonias en los
cítricos. Por su parte el vector de CTV más eficiente en el mundo, T. citricida, llega a
ser 25 veces más eficiente que A. gossypii, el cual viene a ser el segundo principal
vector de CTV (GOTTWALD, GARNSEY y BORBÓN, 1998).
T. citricida probablemente es originario de China, es común en el hemisferio sur y en
el este y sudeste de Asia (BOVE y VOGEL, 1975, citados por NARANJO, 1997b).
Por lo tanto el virus de la tristeza es diseminado eficientemente en gran parte del
hemisferio sur por Toxoptera citricida. Mientras que la tasa de transmisión de A.
gossypii y A. spiraecola ha sido significativamente más baja en California y Florida
(RACCAH, LOEBENSTEIN y BAR-JOSEPH, 1976).
A. gossypii, aparentemente es el principal vector en California, Florida, España e
Israel, en donde T. citricida está ausente. El rol de A. spiraecola como diseminador
de CTV a nivel de campo, aún no está completamente resuelto (GOTTWALD,
GARNSEY y BORBÓN, 1998).
En los últimos años T. citricida, se ha establecido en algunos países de América
Central y del Sur (Venezuela, Costa Rica, Panamá, Nicaragua, Colombia y Argentina),
en las islas Madeira y Reunión y en todas las islas del Caribe (excepto Bahamas y
Bermudas). Se ha demostrado que en las regiones donde se establece el pulgón pardo
comienzan a ser más comunes las razas más virulentas del virus (YOKOMI, 1995).
2.3.2.2 Eficiencia de transmisión de sus vectores
Como ya se mencionó anteriormente la eficiencia de transmisión depende de variados
factores entre ellos están la especie de áfido, la raza del virus y la especie de planta
donante y receptora.

18
HERMOSO DE MENDOZA, BALLESTER-OLMOS y PINA (1986), compararon
los porcentajes de transmisión de dos tipos de razas de CTV diferentes (un aislado
común en España y otro de reciente introducción que provoca seedling yellows), por
medio de cuatro especies de áfidos. Los resultados indicaron que A. gossypii posee
una alta capacidad para transmitir (30%) el aislado T-300, mientras que A. spiraecola
presenta una baja eficiencia de transmisión (20%) de este aislado. Con respecto al
aislado T-387 fue transmitido solo con un 10% de eficiencia por parte de A.
spiraecola, resultado mucho menor comparado con un 30% de eficiencia que
presentó A. gossypii al transmitir este aislado. Por lo tanto este estudio revela que
Aphis gossypii es para los cítricos españoles el vector más importante de tristeza.
Simultáneamente, HERMOSO DE MENDOZA et al. (1986), compararon la
transmisión de tristeza por Aphis gossypii entre varias especies comerciales de
cítricos. Obteniéndose que A. gossypii tiene mayor capacidad de transmitir CTV
sobre naranjo dulce con un 13,4% como promedio, combinando las tres especies
donantes, seguido por el mandarino con 8,5% como promedio, y al parecer el
limonero es poco apetecido por A. gossypii, con un muy bajo porcentaje de
transmisión en este caso.
YOKOMI et al. (1994), evaluando al mismo tiempo cuatro razas provenientes de
distintas partes del mundo, determinan una eficiencia de transmisión del 41 % para
Toxopteta citricida y un 13% para Aphis gossypii, con eficiencias de transmisión
individual (un áfido por planta receptora) variables según el aislado que se ocupe,
pero estableciéndose una eficiencia media de 16% y 1,4%, respectivamente.
YOKOMI y GARNSEY (1987), determinan una eficiencia de transmisión para cuatro
razas al mismo tiempo del 6,3% para Aphis spiraecola y de 17,9% para Aphís
gossypii. No obstante, se han detectado aumentos de la eficiencia de algunos vectores,
como Aphis spiraecola, Toxoptera aurantii y Aphis gossypii, siendo este último el

19
que ha experimentado los cambios más dramáticos (ROISTACHER y MORENO,
1991; HERMOSO DE MENDOZA, BALLESTER-OLMOS y PINA 1984). BAR-
JOSEPH (1978), sugiere que un cambio en la eficiencia de transmisión ocurre en
función de un cambio en la transmisibilidad del virus, y no en la del vector.
2.3.2.3 Áfidos vectores presentes en Chile
Afortunadamente, la especie de pulgón (T. citricida) más eficiente en la transmisión
del virus de la tristeza no está en Chile (SÁNCHEZ, 1996). Lo que queda
corroborado por BESOAIN et al. (2003a) en la más reciente prospección de CTV
realizada en toda la zona citrícola del país, indicando que hasta la fecha de todas las
muestras de áfidos analizadas no se ha determinado la presencia de T. citricida en
Chile.
Los áfidos que si se encuentran en Chile como vectores de CTV son A. gossypii, A.
spiraecola, T. aurantii, además, se han descrito como plaga de cítricos: Aphis
spiraecola, Aphis craccivora, Myzus persicae, Macrosiphum euphorbiae, (PRADO,
1991). Siendo A. spiraecola uno de los áfidos mas frecuentes e importantes como
plaga de los cítricos, el cual además del daño directo tiene una moderada capacidad
de transmisión de CTV (RIPA, 1999; YOKOMI y GARNSEY, 1987).
Cabe destacar que debido a la ausencia T. citricida no se ha desarrollado una
epidemia de grandes proporciones, aunque la mayoría de huertos viejos de naranjos
dulces están sobre naranjo agrio, combinación muy susceptible (SÁNCHEZ, 1996).
2.4. Caracterización y diagnóstico del virus de la tristeza
La identificación de CTV por síntomas en campo no es segura, además la ausencia de
síntomas no implica que el virus no este presente, ya que éste puede albergarse en

20
plantas tolerantes (GORRIS, MARTINEZ y CAMBRA, 2000). Además que el
decaimiento, más o menos rápido, que ocurre en árboles injertados sobre el patrón
naranjo agrio no es específico (CAMBRA et al., 1991).
Por lo tanto el diagnóstico de la tristeza por métodos de invernadero o de laboratorio
es pues imprescindible (CAMBRA et al., 1991).
2.4.1 Caracterización biológica
Consiste en estudiar los síntomas que cada aislado de tristeza produce en diferentes
hospederos, evaluando en terreno y en invernadero con plantas indicadoras infectadas
con tristeza transmitida mediante áfidos o por injerto. En ambos casos se mide
intensidad, tipo o gama de síntomas y el crecimiento de copa y raíces (BALLESTER-
OLMOS et al., 1993).
La sintomatología de tristeza y las cinco plantas indicadoras utilizadas para la
caracterización de las razas de CTV son:
• Lima mexicana: acanaladura en la madera, aclaramiento de venas y
acucharamiento de hojas.
• Naranjo dulce / naranjo agrio: necrosis del floema, enanismo y “seedling
yellows”.
• Naranjo agrio: enanismo y “seedling yellows”.
• Pomelo Duncan: acanaladura en la madera, enanismo y “seedling yellows”.

21
• Naranjo dulce Madame Vinous: acanaladuras en la madera y enanismo.
Este método de diagnóstico tiene algunos inconvenientes como: el tiempo que se
requiere para la aparición de síntomas es mucho, es costoso y además que las
diferencias entre razas no son absolutas (MORENO, 1992). Otro inconveniente es la
dificultad de su aplicación a tejidos vegétales cultivados in vitro o a áfidos virulíferos
(CAMBRA et al., 1991).
Es por ello que las investigaciones en torno a este tema se han enfocado a desarrollar
nuevos tipos de métodos de diagnóstico más confiables, certeros, rápidos, de uso
masivo y aplicable a gran número de razas de CTV.
2.4.2 Caracterización serológica
La serología ha introducido en el diagnóstico de CTV ventajas muy notables que han
sido completadas con las posibilidades que ofrece la técnica inmunoenzimática
ELISA (CAMBRA et al., 1991).
El método Inmunoimpresión – ELISA para la detección de CTV fue desarrollado
entre 1991 y 1993 en el IVIA de Mondaca (Valencia) en conjunto con el equipo del
Dr. Garnsey del USDA de Orlando (Estados Unidos) (GORRIS, MARTINEZ y
CAMBRA, 2000).
Hoy en día la tendencia es utilizar Inmunoimpresión – ELISA, la cual es más sencilla,
rápido (tres horas), sensible, económico y fiable y no necesita preparación de
extractos. Una gran ventaja que presenta este procedimiento es que las impresiones o
improntas pueden prepararse en el campo, ser almacenadas en laboratorio y
procesadas varios meses después, no tiene riesgos de contaminación y su

22
repetibilidad es muy buena (CAMBRA y MORENO, 2000). Esta técnica es utilizada
en todos los países citrícolas con los anticuerpos monoclonales españoles 3DF y
3CA5. Estos anticuerpos son los únicos que en mezcla son capaces de reconocer a
cualquier aislado de CTV sin reacción alguna con otros componentes de la planta
(GORRIS, MARTINEZ y CAMBRA, 2000).
La relativa facilidad de análisis mediante ELISA y la buena disponibilidad de
anticuerpos monoclonales, han hecho posible abordar grandes prospecciones,
estudios epidemiológicos y programas de erradicación y control de CTV (CAMBRA,
1994).
2.4.3 Caracterización por métodos bioquímicos
Prácticamente existen dos grupos de métodos bioquímicos que se utilizan con este
fin:
- El primero se refiere a las características de las proteínas de la cápside, y se basa
en la fabricación de mapas de péptidos.
- El segundo grupo se refiere a los ácidos nucleicos, bien sean los RNA de doble
cadena que se generan en las plantas infectadas o mediante el estudio del ácido
nucleico del genoma del virus usando sondas de cDNA (NARANJO, 1997b).
2.4.4 Amplificación de fragmentos de ácidos nucleicos
Esto se logra mediante la Reacción en Cadena de la Polimerasa (PCR), y es una de las
técnicas que se esta poniendo a punto en los distintos laboratorios, ya que entrega un
gran poder de detección y es un método conveniente para los estudios

23
epidemiológicos de CTV (NOLASCO et al., 1995; ACOSTA et al., 1994;
CALVERT, 1987, citados por NARANJO, 1997b).
Es así como en los últimos años se ha obtenido el ARN genómico de distintos
aislados, permitiendo el desarrollo de técnicas basadas en la detección del ARN viral,
tales como Inmunocaptura-PCR, la impresión o cachado captura- PCR o nested-PCR
en un único tubo cerrado. (CAMBRA y MORENO, 2000).
Las nuevas técnicas como el nested-PCR y hemi nested-PCR en un único tubo
cerrado, a diferencia de las anteriores, son técnicas que permiten amplificar las
secuencias de ARN viral de CTV, de plantas infectadas o áfidos virulíferos,
directamente desde el escachado realizado sobre el papel, sin necesidad de realizar
extracción de ácidos nucleicos o inmunocaptura (CAMBRA et al., 2000).
2.5. Dinámica de la distribución espacial de CTV
La velocidad de transmisión de CTV lograda por los vectores en una determinada
región citrícola no depende sólo de la eficiencia de transmisión de las especies de
áfidos presentes (ROISTACHER y MORENO, 1991). Por consiguiente entre los
factores que determinan la velocidad de dispersión de CTV, se encuentran: la
densidad de población y hábitos de vuelo de los vectores, la densidad del inóculo, las
especies de cítricos predominantes y la intensidad y duración de la brotación
(CAMBRA y MORENO, 2000).
A la vez se ha demostrado que la composición de las poblaciones de áfidos vectores
altera la evolución del patrón de distribución espacial del virus, afectando la
diseminación y transmisión de CTV (GOTTWALD, GARNSEY y GIBSON, 2000).

24
Por otro lado, se ha demostrado que la atracción de los áfidos vectores hacia su
hospedero varía según del habito de crecimiento del hospedero, turgencia de brotes,
el color de las hojas, etc. Por lo que estos parámetros, en conjunto con el número de
poblaciones de áfidos y el número de áfidos alados podrán ser incluidos en modelos
espaciales y temporales de CTV más depurados (GOTTWALD, CAMBRA y
MORENO, 1993).
2.5.1 Distribución espacial de CTV según los vectores predominantes
En zonas donde no está presente T. citricida, y Aphis gossypii es el principal vector,
se puede pasar de una incidencia de CTV relativamente baja (5%) a una incidencia
alta (95%) en 8 a 15 años, y posiblemente por su habito migratorio este áfido, infecta
a árboles distantes, obteniéndose como resultado una distribución difusa o al azar de
CTV en campo. Por el contrario, datos obtenidos desde zonas donde el vector
principal es T. citricida, indican que los mismos incrementos tienen lugar en sólo dos
a cuatro años; el hábito colonizador de esta especie de áfido hace que las nuevas
infecciones ocurran en los árboles adyacentes, lo cual hace que la enfermedad se
extienda en forma de manchas, provocando una aparición más agregada del patógeno
en la plantación (GOTTWALD et al., 1999; GOTTWALD, GARNSEY y BORBÓN,
1998; MORENO, 1995).
Si ocurre una coinfestación por ambas especies de áfidos, un gran porcentaje de los
árboles son infectados en el mismo período de tiempo, y resultado de esto puede ser
la rápida diseminación a larga distancia. Esta situación se observa frecuentemente en
la zona del caribe (GOTTWALD et al., 1999; GOTTWALD et al., 1995).
El mismo autor señala que cuando hay una dispersión extensa y difusa de CTV, es
importante considerar el efecto del viento.

25
En análisis de modelos espaciales de CTV realizados en Israel y Florida, cuando A.
gossypi es el vector predominante, y la canopia de los árboles comienza a cerrarse y a
toparse con los árboles adyacentes, se pasa de una distribución al azar a un modelo
agregado de dispersión. La razón por que ocurre esto no esta clara, pero existen
áfidos como A. spiraecola y T. aurantii que a pesar de ser menos eficientes, forman
grandes colonias en cítricos las cuales pueden contribuir a la formación de un modelo
agregado (GOTTWALD et al., 1996).
GOTTWALD et al. (1999), en investigaciones posteriores, trabajando con A. gossypii
como vector revelan que el modelo agregado de dispersión de CTV, es explicado por
infecciones provenientes desde fuera de la parcela y con lo que llaman mecanismo de
transmisión local, el cual muestra a las nuevas infecciones dependientes de
infestaciones previas dentro del huerto.
Aunque Aphis gossypii es un vector más eficiente que Aphis spiraecola, la dinámica
poblacional debe ser considerada, ya que A. spiraecola es mucho más abundante que
A. gossypii, y exhibe una actividad alada mucho mayor. No obstante, se han detectado
cambios en el balance de la fauna afídica favoreciéndose un aumento de la población
del vector más eficiente (ROISTACHER y MORENO, 1991). Esto puede ser efecto
de grandes migraciones hacia los cítricos, debido al hábito migratorio de esta especie,
lo cual se traduce en un aumento en el número de nuevas infecciones (MORENO,
1995).
El hecho de que A. gossypii sea menos abundante en cítricos que A. spiraecola y que
T. citricida (en los países en donde esta presente), se debe a que los cítricos no son el
principal hospedero para A. gossypii, el cual migra antes de formar grandes colonias
(GOTTWALD, GARNSEY y BORBÓN, 1998).

26
En el caso de A. spiraecola no se han desarrollado modelos estadísticos que
expliquen el patrón de dispersión de CTV que pueda conferir este vector, y su rol en
la diseminación de la enfermedad a nivel de campo no está resuelto (GOTTWALD,
GARNSEY y BORBÓN, 1998).
2.5.2 Modelos de distribución espacial
Los modelos estadísticos de distribución, en una comunidad biológica distinguen tres
patrones básicos de dispersión: al azar, agrupadas y uniformes. Para poder determinar
el patrón de distribución espacial de especies que forman una comunidad biológica,
sobre unidades de muestreo (hojas, frutos, árboles, etc.) los datos se deben sintetizar
en una frecuencia de distribución. Esta frecuencia de distribución consiste en el
número de unidades de muestreo con cero individuos, un individuo, dos individuos,
etc. (LUDWIG y REYNOLDS, 1988).
En el caso de los modelos al azar se asume que no hay influencia del ambiente y de
los patrones de comportamiento o de otros individuos. El modelo agregado sugiere
que los individuos se agrupan en partes más favorables para ellos, lo que puede
deberse a un comportamiento gregario, heterogeneidad del ambiente, hábito
reproductivo, intervención del hombre, etc. En cambio los patrones uniformes son
producto de interacciones negativas entre los individuos, tales como la competencia
por comida o espacio. Una vez que el patrón ha sido identificado, el investigador
puede proponer y probar hipótesis que revelen que factores pueden ser los
responsables (LUDWIG y REYNOLDS, 1988).
Al observar la relación entre la varianza del número de individuos por unidad de
muestreo y la media, se puede determinar a que de distribución espacial se ajustan
los individuos dispersos en una comunidad. Los tres tipos básicos de distribución
espacial y su relación varianza – promedio se definen a continuación:

27
Para patrones de distribución al azar: (σ² = λ)
Para patrones de distribución agrupados: (σ² > λ)
Para patrones de distribución uniforme: (σ² < λ).
Donde λ: media de los datos; σ²: desviación estándar (LUDWIG y REYNOLDS,
1988).
Otros valores que ayudan a determinar el patrón de distribución que presenta una
comunidad de individuos, son los índices de dispersión. Estos índices relacionan la
proporción varianza – promedio, sugiriendo una cierta distribución. A continuación
se definen los índices de dispersión utilizados:
- Índice de dispersión: relación o proporción existente entre la varianza y el
promedio. Este método no es tan exacto para medir el grado de agrupamiento de una
población.
- Índice de agrupamiento: variación del índice anterior, también relaciona la
proporción de la varianza y el promedio.
- Índice de morosita: este índice no se ve afectado por cambios en la densidad debido
al azar. Además de relacionar la varianza y el promedio, incluye el número total de
individuos.
En el Cuadro 1, se observan los valores de máxima uniformidad, aleatoriedad y de
máximo agrupamiento para cada uno de los índices utilizados en este estudio.

28
CUADRO 1. Propiedades de los índices de dispersión.
Valor del índice “a”
Índice
Máxima
uniformidad
Aleatoriedad Máximo
agrupamiento
Índice de dispersión 0 1 N
Índice de agrupamiento -1 0 n-1
Índice de Morosita 1-[(x-1)/(n-1)] 1 X
Fuente: LUDWIG y REYNOLDS, 1988
Donde n es el número total de individuos (árboles infectados) en la muestra y ‘x’ es el
número de cuadrículas.
2.6. Prevención y control de la enfermedad
En el caso de España, las prospecciones realizadas han permitido conocer con detalle
la distribución e incidencia real de CTV, independiente de sus síntomas, lo cual ha
sido fundamental para determinar las medidas de control que se llevan a cabo. Por lo
tanto la lucha contra el virus de la tristeza se debe concebir en forma integral, ya que
se deben incluir métodos directos (Cuarentena o impedir la entrada del virus y
erradicación) y métodos indirectos (reducir daños causados por la enfermedad con el
uso de patrones tolerantes y preinmunización) (CAMBRA, 1994; CAMBRA et al.,
1990).
2.6.1 Medidas de exclusión del inóculo
Consiste en impedir la entrada del inóculo a zonas libres del virus mediante medidas
cuarentenarias, esto también involucra evitar la entrada de razas severas en zonas

29
donde el virus esta presente pero en forma aislada o en donde predominan razas poco
agresivas. Para ello son fundamentales las estaciones de cuarentena y las inspecciones
en frontera (CAMBRA y MORENO, 2000; CAMBRA et al., 1991).
2.6.2 Medidas de erradicación
Si la enfermedad ya está presente en una zona determinada, hay que estudiar la
posibilidad de erradicación o en todo caso la realización de planes para disminuir la
cantidad de inóculo (CAMBRA et al., 1991).
Por lo tanto, la erradicación consiste en eliminar la enfermedad en áreas en donde ésta
presente el virus, procediendo al arranque de plantas infectadas. La erradicación para
ser efectiva, debe realizarse más rápidamente que la difusión natural de la enfermedad
(CAMBRA et al., 1990).
2.6.3 Protección cruzada
Cuando la tristeza (CTV) es endémica y se poseen varias razas virulentas, existe la
posibilidad de establecer sistemas de protección cruzada o preinmunización con razas
débiles o poco virulentas que impidan la manifestación de los daños de las razas mas
agresivas (CAMBRA et al., 1991). Esta no es una solución a largo plazo, sino que
considera una estrategia de control intermedia que permite alargar la vida económica
de los árboles (LEE et al., 1995).
2.6.4 Patrones tolerantes y mejoramiento genético
El método indirecto mas eficaz, de lucha contra tristeza, es la utilización de patrones
tolerantes y el control de sanidad en viveros, a partir de material libre de virus
(CAMBRA, 1994).

30
Otra posible vía para disminuir los daños causados por CTV, es la introducción de
resistencia al virus en variedades comerciales por medio de Biotecnología.
Actualmente se están desarrollando plantas transgénicas de cítricos que podrían
expresar genes naturales de resistencia o anticuerpos frente a las proteínas
estructurales de CTV (CAMBRA y MORENO, 2000).

31
3. MATERIALES Y MÉTODOS
3.1. Estudio sobre la distribución espacial del CTV:
Con el fin de estudiar la distribución espacio temporal de CTV, se realizó toma de
muestras de brotes y captura de áfidos vectores durante la primavera del año 2003 y
el otoño del 2004 en tres huertos comerciales de cítricos ubicados en las comunas de
Ovalle en la IV Región, Limache en la V Región y Paine en la Región Metropolitana.
Según el último censo agropecuario (INE, 1997) estas tres regiones comprenden el
61,3 % del total de la superficie plantada con cítricos en el país.
La elección de estos predios obedece a la presencia en éstos de árboles infectados con
CTV, según muestreos realizados por la Pontificia Universidad Católica de
Valparaíso en el proyecto Fondo – SAG VI – 15 - 0199 de: “Prospección de razas
severas y áfidos vectores del virus de la tristeza de los cítricos, presentes entre la I y
VII Región de Chile entre los años 1999 y 2002”.
Para llevar a cabo este trabajo, se seleccionaron sectores de 400 árboles por predio en
donde había árboles cuyas muestras dieron positivas a la infección de CTV según los
análisis realizados en el estudio de prospección antes mencionado.
El trabajo consistió en tomar muestras de cada sector seleccionado a las cuales se les
realizó Inmunoimpresión- ELISA para analizar la presencia de CTV, además, en los
tres predios, se capturaron pulgones por medio de la metodología de brotes pegajosos,
los cuales fueron llevados a España, donde se les realizó nested-PCR para determinar
la presencia de áfidos portadores de CTV.

32
Las muestras colectadas se analizaron mediante Inmunoimpresión-ELISA, en el
Laboratorio de Fitopatología de la Pontificia Universidad Católica de Valparaíso,
ubicado en La Palma, comuna de Quillota, V Región.
3.1.1 Muestreo y recolección del material vegetal
La metodología consistió en colectar material vegetal de los 400 árboles
seleccionados, cuatro brotes por árbol, que correspondió a tomar un brote por cada
punto cardinal. En todos los casos se tomaron brotes tiernos de la temporada y de
unos 15 a 20 cm de largo, desde la altura media del árbol, los cuales fueron
identificados con una cinta blanca que indicaba el número de la hilera y el número
correlativo del árbol dentro de la hilera. De este modo y con la confección de un
mapa del sector se pudo identificar fácilmente cuales son los árboles infectados con
CTV tras realizar Inmunoimpresión- Elisa a los brotes colectados.
Una vez marcados los brotes, estos se almacenaron en bolsas plásticas, las que se
conservaron en un contenedor refrigerado hasta la llegada al laboratorio, para su
análisis posterior.
En los tres huertos, se realizaron dos muestreos de brotes, con el fin de hacer un
seguimiento de la enfermedad a través del tiempo, el primero se realizó durante la
primavera del año 2003, y el segundo se efectuó en otoño del año 2004 como se
mencionó anteriormente.
3.1.1.1 Muestreo en la localidad de Ovalle
En este huerto que corresponde a una plantación de limoneros variedad Eureka sobre
patrón C. macrophylla, el primer muestreo se realizó el 9 de octubre del 2003. A
diferencia de los muestreos en los otros predios, en este predio se analizó además en

33
los 400 árboles un trozo de corteza de 1 cm² de la zona del PI, con la finalidad de
determinar si hay presencia de CTV en esa zona, ya que se plantea que como ambas
especies son hipersensibles (limonero y C. macrophylla), las partes del complejo viral
que se puedan multiplicar en las células del limonero serán las más débiles y por tanto
no afectan a C. macrophylla (NAVARRO, 1981).
En el segundo muestreo también se tomaron muestras de los 400 árboles, y se realizó
el 21 de junio del 2004, en esta oportunidad sólo se muestrearon brotes y no corteza.
3.1.1.2 Muestreo en la localidad de Limache
En este huerto que corresponde a una plantación de naranjo Valencia Late sobre
patrón citrange (troyer o Carrizo), se tomaron brotes desde los 400 árboles del
sector, realizándose el primer muestreo el 19 de noviembre del 2003, y el segundo el
10 de junio del 2004.
3.1.1.3 Muestreo en la localidad de Paine
Este corresponde a un huerto de naranjo variedad Thomson sobre portainjerto
troyer. El primer muestreo se realizó el día 4 de noviembre del 2003, y el segundo
muestreo se realizó el 7 de junio del 2004, y al igual que el caso anterior se
muestrearon brotes tiernos de la temporada de los 400 árboles del sector.
3.1.2 Diagnóstico
Una prueba fácil y confiable para diagnosticar y verificar la presencia de CTV en las
plantas es el test de Inmunoimpresión-ELISA, el cual utiliza técnicas
inmunoenzimáticas con anticuerpos monoclonales específicos, en este caso se

34
utilizaron los anticuerpos 3DF1 y el 3CA5, que reaccionan con epítopes determinados
de la cápside de CTV.
Esta prueba se realizó en el Laboratorio de Fitopatología de la PUCV, y el
procedimiento consistió en cortar los ápices de los brotes tiernos en forma transversal,
posteriormente, una vez hechos los cortes con un bisturí las secciones se imprimieron
con firmeza sobre una membrana de nitrocelulosa, en la cual quedaron las improntas
del material vegetal.
El siguiente paso fue bloquear los poros de la membrana y la superficie no
impresionada con una solución al 1% de albúmina de suero bovino en agua destilada
(20 ml para una membrana), la membrana se mantuvo cubierta en esta solución por
un tiempo de una hora, con una suave agitación y a temperatura ambiente, una vez
desechada la albúmina se agregaron los anticuerpos monoclonales específicos de
CTV ( para una membrana: 1 ml de agua fisiológica tamponada (AFT), más 9 ml de
agua destilada estéril, y 0,1 ml de anticuerpo) marcados con la enzima fosfatasa
alcalina. Luego de un período de incubación de dos a tres hr esta solución de
anticuerpos fue eliminada.
Posteriormente las membranas fueron lavadas con un tampón lavador (25 µl de
Tween 20 + 0,4375 g de sales minerales diluido en 50 ml de agua estéril), con el fin
de eliminar todos los anticuerpos marcados con fosfatasa alcalina que no hayan
reaccionado.
El último paso, correspondió a la aplicación de un sustrato específico (BCIP +NBT)
que reacciona con la fosfatasa alcalina, este sustrato pone de manifiesto el material
vegetal infectado después de tres a siete minutos de incubación, luego se lavaron las
membranas con agua corriente para paralizar la reacción, y se dejaron secar sobre
papel absorbente.

35
La observación de las improntas se realizó bajo lupa 20x, y basta con la presencia de
unos pocos precipitados de color violeta- purpura en la zona del floema para concluir
que se trata de material vegetal infectado.
3.1.3 Análisis espacial y temporal de la enfermedad
Una vez obtenidos los datos de los árboles infectados con CTV, se confeccionaron
mapas de cada huerto, en los cuales se marcaron las plantas infectadas.
Los mapas fueron estudiados dividiéndolos en cuadrículas de igual tamaño. No
obstante debido a que la cantidad de árboles y diseño de plantación era diferente en
un huerto, las cuadrículas no son del mismo tamaño al comparar los tres huertos entre
si.
En cada cuadrícula, se contaron los árboles con infección y luego los datos se
llevaron a una tabla, en donde se ordenaron en número de árboles infectados por
cuadrícula (Xi) y en número de cuadriculas con X árboles infectados (Ni).
Para llevar a cabo el estudio de la dispersión espacial de los árboles infectados con
CTV en cada uno de los huertos visitados, se utilizó la prueba estadística Ji-cuadrado
(X²), verificando el ajuste Poisson, ya que la estructura de los datos y las
observaciones realizadas indicaban que se trataba de una distribución al azar en los
tres predios. Además, se realizó el cálculo de los índices de dispersión para el mismo
estudio y con la misma finalidad.
Para realizar el estudio temporal de la enfermedad se realizó el calculo de Dócima
para comparar proporciones, el cual indica si hay cambios significativos en las
proporciones de árboles infectados entre el primer y segundo muestreo.

36
• Distribución de probabilidad Poisson
Para ver si los datos obtenidos se ajustan a este tipo de distribución, se realizaron los
siguientes pasos:
1. Establecer la hipótesis: La hipótesis es que el número de árboles infectados con
CTV se distribuye al azar.
2. Distribución de frecuencia, Fx: Los datos de las muestras se resumen en una
distribución de frecuencia, es decir, el número de cuadrículas con x = 0, 1, 2,...n
árboles infectados con CTV.
3. La probabilidad de Poisson, P(x): La probabilidad de encontrar x árboles
infectados en una cuadrícula, es decir, P(x), donde x = 0, 1, 2,..., n árboles
infectados, está dada por la serie Poisson:
P(x) = ( !/)ˆ xx λλ −l
Donde scuadrículadetotalNii °∑= /ˆ χηλ ; e: base del logaritmo natural; x!:
factorial de x
4. Frecuencias Poisson esperadas, Ex: Este se calculó multiplicando cada
probabilidad por el número total de cuadrículas. Esto da como resultado un total de
clases de frecuencias de individuos esperados, q = n + 1.
5. Test estadístico Ji-cuadrado para la bondad de ajuste: Esta prueba se usó para ver
cuan bien las frecuencias observadas se comparan con las frecuencias esperadas.

37
El test estadístico Ji-cuadrado se calcula de la siguiente manera:
)7(/)( 22 XEEQ iii ≈−∑= η
Este test estadístico es comparado a una tabla de probabilidades de Ji-cuadrado con q
- 2 grados de libertad. Si el test estadístico es mayor que el Ji-cuadrado tabulado a un
nivel de probabilidad dado, se concluye que es improbable que la distribución siga
una serie Poisson, por lo tanto, se rechaza la hipótesis nula (LUDWIG y
REYNOLDS, 1988).
• Índices de dispersión
Los índices de dispersión fueron utilizados para medir el grado de agrupamiento de
los árboles infectados en el huerto. Son variados los índices de dispersión que se
pueden utilizar en una comunidad ecológica con este fin, por lo cual, para este estudio
los índices utilizados fueron: Índice de dispersión, Índice de agrupamiento e Índice
de morosita.
Índice de dispersión ID = s²/ x
Índice de agrupamiento IC = (s²/ x ) – 1 = ID -1
Índice de Morosita (Id = (n/n –1) (x*/ x )
Donde: x = media de los datos; n = N° total de árboles en la muestra; x* = x + IC

38
Con los valores obtenidos en estos índices se determinó que grado de aleatoriedad o
uniformidad tienen las variables.
• Dócima para comparar proporciones
Para este análisis lo primero fue establecer la hipótesis. Ho: La proporción de árboles
infectados en el primer muestreo es igual a la proporción de árboles infectados en el
segundo muestreo.
Para determinar si la proporción de árboles infectados en el primer muestreo es igual
o significativamente diferentes a la proporción de árboles infectados se realizaron los
siguientes cálculos:
11 /ˆ nxp = ,
22 /ˆ nyp = ,
Donde: “x”es el número de árboles infectados en primer muestreo; “y” es el número
de árboles infectados en segundo muestreo; “n” corresponde al número de cuadrículas
totales.
Luego se calculó: 210 /ˆ nnyxp ++=
Donde: 00 ˆ1ˆ pQ −= ; “Z”: es el Z observado que debe ser comparado con Z tabulado,
para determinar si se rechaza o no la hipótesis nula.
)/1/1(ˆ*ˆ/ˆˆ 210021 nnQpppZ += −

39
3.1.4 Captura de áfidos
Las capturas se realizaron cada 10 días en cada uno de los tres predios en estudio,
comenzando en Limache el 15 de septiembre del 2003, en este predio se realizaron
siete capturas durante esta temporada, siendo la última el 19 de noviembre. En la
siguiente temporada en este predio se realizaron ocho capturas, desde el 19 de abril al
1 de julio del año 2004, sumando para este predio un total de 15 capturas durante las
dos temporadas.
En Paine, se realizaron seis capturas durante la primavera del 2003, y comenzaron el
14 de septiembre hasta el 4 de noviembre. Al igual que en el caso anterior las
capturas se reinician en otoño del año 2004, correspondiendo la primera al 16 de
abril y la última al 28 de junio, completando un total de 14 capturas en esta localidad.
En Ovalle, las capturas se realizaron desde el 27 de septiembre al 30 de octubre del
año 2003, luego se retomaron el 19 de abril hasta 2 de julio del 2004, haciendo un
total de 12 capturas en este predio durante las dos temporadas.
La técnica empleada para la captura de áfidos fue el método del árbol pegajoso.
3.1.4.1 Metodología del árbol pegajoso
El método empleado consistió en pulverizar cada 10 días con un adhesivo
(Soveurode aerosol) 15 brotes de entre 10 a 20 cm de longitud, de cinco árboles
elegidos al azar (tres brotes por árbol), con la finalidad de atrapar áfidos alados que
visitan, se mueven y alimentan sobre distintos árboles dentro del predio. Los brotes
pulverizados correspondieron a brotes tiernos de la parte media del árbol, y fueron
marcados con una cinta para poder recogerlos en la siguiente fecha de captura, donde
se realiza el mismo proceso en árboles diferentes.

40
Posteriormente los brotes con áfidos fueron puestos en bolsas plásticas y llevados al
laboratorio para su posterior separación e identificación.
3.1.5 Identificación de áfidos
Los brotes con áfidos adheridos, se dejaron 48 hr en un frasco con aguarras para
poder liberarlos de las hojas, luego los brotes se sacaron del frasco quedando los
pulgones en la solución de aguarras, los cuales fueron separados de otros insectos y
clasificados con claves entomológicas creadas para identificar especies de pulgones
alados. Una vez clasificados bajo lupa estereoscópica 40x, los pulgones fueron
guardados en alcohol al 70% para ser llevados a España, en donde se les realizó un
análisis de impresión-PCR.
3.1.6 Impresión o aplastado de áfidos
Los áfidos capturados durante la primavera del 2003, fueron analizados en el
Laboratorio de Inmunología del IVIA, España, en donde el 4 de marzo del 2004 se les
realizó nested-PCR para determinar la presencia de partículas virales de CTV.
En el caso de los áfidos capturados durante el otoño del 2004, estos serán analizados
con posterioridad en el IVIA, España.
3.2 Transmisión de CTV por Aphis gossypii:
3.2.1 Aislados
Los 10 aislados utilizados para este ensayo, fueron seleccionados de un total de 100
aislados recolectados en campo por el proyecto “Prospección de razas severas y
áfidos vectores del virus de la tristeza de los cítricos, presentes entre la I y VII Región

41
de Chile entre los años 1999 y 2002”. La elección de los aislados se realizó en junio
del año 2003 en España, y obedece principalmente a la severidad de las razas,
atenuadas o severas, sintomatología y lugar de recolección. A continuación se
presentan en el Cuadro 2, los 10 aislados elegidos para la transmisión de CTV.
Cuadro 2. Origen, serogrupo y severidad de los aislados de CTV chilenos utilizados
en el estudio de eficiencia de transmisión de A. gossypii
AISLADO LUGAR DE ORIGEN REACCIÓN MCA13 SEROGRUPO
231-1 Azapa (+ ) 1
240-1 Matilla (+ ) 1
281-1 Vallenar (-) 4
305-3 Limache (-) 2
369 Paine (-) 2
301-10 Pica (-) 8
496 Ovalle (-) 5
497 Ovalle (+) 3
509 Pica (+) 1
305-2 Limache (-) 7
Fuente: BESOAIN et al., 2003b
3.2.2 Inoculación a plantas donadoras
Se utilizaron 20 plantas de naranjo dulce Madame Vinous como donadoras de los
aislados (dos plantas por cada aislado de CTV), las cuales en septiembre del año 2003
fueron inoculadas mediante tres trozos de corteza con un injerto en T, cuando tenían
un diámetro de 6 mm. Estas plantas se mantuvieron en un invernadero calefaccionado
a temperaturas de entre 20 a 28 °C, luego se les realizó un rebaje a 30 cm, y además
estuvieron sujetas a un régimen continuo de macro y microelementos por medio de

42
fertirrigación, todo esto para estimular “flushes” de crecimiento y traslocación del
virus.
El 21 y 22 de abril del año 2004, se realizó toma de muestras de todos los posibles
brotes donadores para verificar la presencia de CTV mediante Inmunoimpresión, y
junto con esto se realizó una poda de rebaje para estimular el crecimiento de nuevos
brotes donantes.
El 23 de abril las muestras fueron impresas y analizadas, en el Laboratorio de
Fitopatología de la PUCV, con lo que se pudo determinar que brotes son los que
contienen el virus.
3.2.3 Plantas receptoras
Se utilizaron plantas de entre 10 a 15 cm de altura, y correspondieron a 110
naranjos dulce (Madame Vinous) y 110 Limas mexicanas, obtenidas de semilla y
micropropagadas en el Laboratorio de Propagación de la PUCV. Estas plantas de
semillas fueron germinadas en un medio de cultivo MS (Murashige Skoog), luego
fueron colocadas en un medio de multiplicación o enraizamiento, posteriormente, una
vez enraizadas (60 a 90 días desde que fueron puestas a germinar) se trasladaron a la
maternidad del Programa de Certificación de Cítricos de la PUCV, por lo tanto al ser
plantas de semillas y mantenidas en un lugar hermético existe la seguridad que son
libres de virus.
3.2.4 Identificación de Aphis gossypii
Se colectaron muestras de áfidos en la colección de cítricos de la Estación
Experimental La Palma (Quillota), las cuales fueron llevadas al Laboratorio de
Fitopatología de la PUCV, para ser observados bajo lupa, y con la ayuda de claves

43
entomológicas específicas se identificó a Aphis gossypii según características
morfológicas (CHARLIN, 1975; HERMOSO DE MENDOZA, 2004)*. Para
corroborar la identificación, se enviaron muestras de la especie clasificada al
Laboratorio de Entomología del Servicio Agrícola y Ganadero de Valparaíso, donde
fueron clasificados por el método de montaje de Hoyer’s por el entomologo Sr. Luis
Torelli, coincidiendo con la clasificación descrita anteriormente en el Laboratorio de
Fitopatología de la PUCV. Una vez identificada la especie de áfido, estos fueron
colocados por medio de un pincel sobre el sustrato para su multiplicación.
3.2.5 Confinamiento y crianza de Aphis gossypii
El sustrato que se utilizó para la crianza de Aphis gossypii fueron plantas de algodón,
por lo tanto a partir del 15 de marzo se comenzó con la germinación de 100 semillas
de algodón por semana, colocando 15 a 20 semillas por maceta, hasta completar un
total de 1000 plantas, que es la cantidad que se estimó para alimentar a 14000
pulgones aproximadamente. La importancia de tener suficiente alimento para la
multiplicación de los pulgones, radica en evitar que haya competencia por el alimento
y mantener las colonias en el tiempo, mientras se obtiene el número deseado de
pulgones, además de esta manera se evitó que se produjeran generaciones de hembras
aladas que puedan migrar hacia el exterior del invernadero.
La crianza de los áfidos, se realizó bajo condiciones controladas de temperatura (18 –
26ºC), bajo invernadero, donde se aplicó azúcar semanalmente para atraer hormigas,
las cuales favorecen la reproducción de áfidos, además las plantas fueron fertilizadas
una vez a la semana con urea en dosis de 1 g por litro, para tener brotes más turgentes
y apetecibles para los áfidos.
*HERMOSO DE MENDOZA A. Ph D. Ingeniero Agrónomo, Investigador del IVIA,
España.

44
3.2.6 Transmisión
Una vez que se seleccionaron los brotes en donde se alimentarán los pulgones, según
Inmunoimpresión- ELISA, el primer paso es la adquisición, la que se llevó a cabo el
6 de julio del año 2004, para ello se cortaron brotes y hojas de algodón con áfidos, los
cuales fueron pegados (con cinta adhesiva) al brote donante. De esta forma, los
pulgones se movilizan desde los brotes de algodón a los brotes virulíferos, donde
comenzaron su alimentación (adquisición) sobre las plantas donadoras, luego fue
necesario cubrir los brotes donadores para evitar migraciones de áfidos, para tal
efecto, se ideó un sistema, en el que se utilizaron tubos transparentes de 15 cm de
largo y 6 cm de diámetro, estos tubos fueron tapados con malla antiáfidos, dejando un
costado abierto para poder introducir el brote. Una vez puesto el tubo en el brote, se
amarró la malla antiáfidos por donde se introdujo el brote y se dejó 24 horas
adquiriendo el virus.
El segundo paso, corresponde a la transmisión propiamente tal, se realizó el 7 de julio
del 2004. Para tal efecto se cortaron los brotes, y las hojas de las plantas donantes que
contienen los pulgones con el virus adquirido, y se traspasaron a las plantas
receptoras de la misma forma como se hizo la adquisición. Los áfidos se dejaron
inoculando el virus a las plantas sanas por un período de 48 horas.
Pasado las 48 horas, se aplicó un insecticida para eliminar los áfidos. Las plantas
receptoras permanecen en un invernadero con temperaturas que oscilan entre 18 –
25º C , en donde se le realizan evaluaciones periódicas de sintomatología.

45
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Captura de áfidos
En la primera temporada de capturas, es decir, en la primavera del 2003, sólo se
encontraron áfidos en los brotes pegajosos de Limache y Paine. Este hecho no
significa que en el huerto de Ovalle, no existieran áfidos, ya que el método del árbol
pegajoso busca atrapar sólo áfidos alados y no colonias ni individuos ápteros. No
obstante, en la segunda temporada de capturas (2004), se encontraron áfidos en los
brotes pegajosos en los tres predios.
La primera captura se realizó en Ovalle, en donde los áfidos no estaban en forma
abundante, lo que provocó que en la primera temporada no se encontrara ningún
pulgón en los brotes pegajosos. Sin embargo, en la segunda temporada de capturas, se
identificó a Aphis gossypii como el áfido mas abundante, seguido muy de cerca por A.
spiraecola (Cuadro 3). YOKOMY y GARNSEY (1987), indican que A. spiraecola es
uno de los áfidos mas frecuentes e importantes como plaga de los cítricos, el cual
además del daño directo tiene una moderada capacidad de transmisión de CTV. Por
otro lado ROISTACHER y MORENO (1991), plantearon que se han detectado
cambios en la composición de la fauna afídica como consecuencia de muchos
factores, entre ellos: resistencia selectiva a determinados plaguicidas, variaciones en
la población de enemigos naturales, o cambios favorables para el aumento de otras
especies de áfidos.
En el Cuadro 3, se presentan los áfidos capturados en las dos temporadas de
muestreos, en las localidades de Ovalle, Paine y Limache.
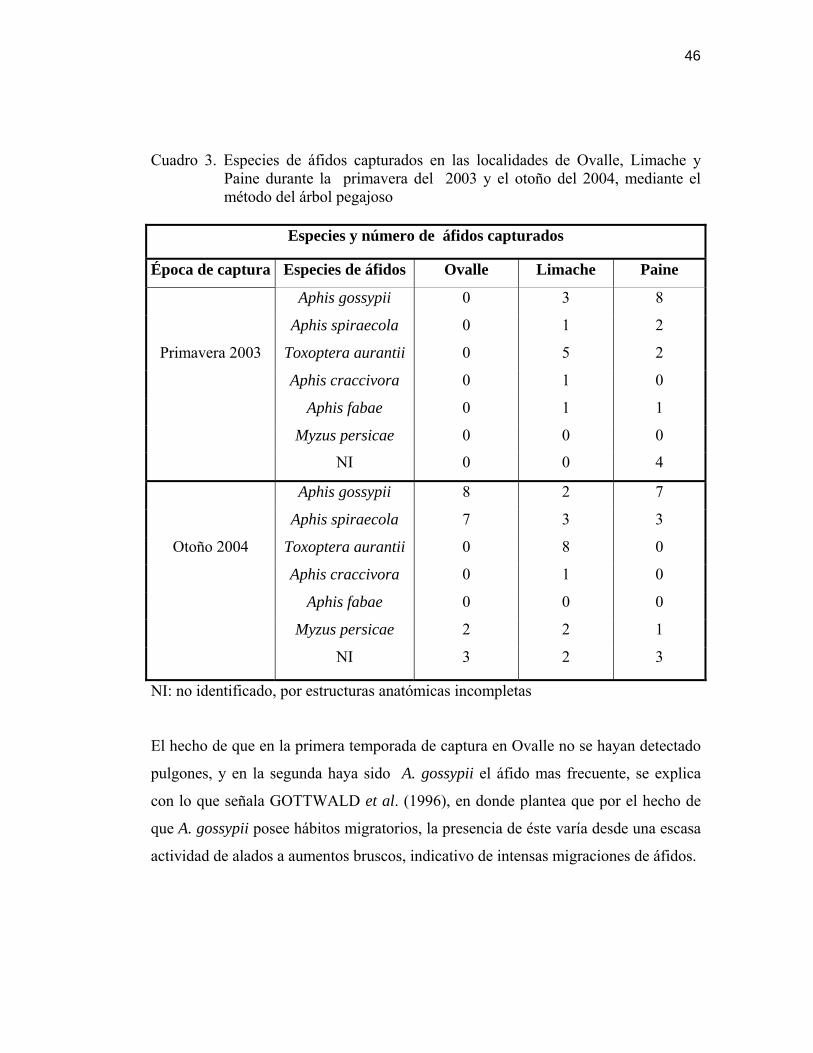
46
Cuadro 3. Especies de áfidos capturados en las localidades de Ovalle, Limache y Paine durante la primavera del 2003 y el otoño del 2004, mediante el método del árbol pegajoso
Especies y número de áfidos capturados
Época de captura Especies de áfidos Ovalle Limache Paine
Aphis gossypii 0 3 8
Aphis spiraecola 0 1 2
Toxoptera aurantii 0 5 2
Aphis craccivora 0 1 0
Aphis fabae 0 1 1
Myzus persicae 0 0 0
Primavera 2003
NI 0 0 4
Aphis gossypii 8 2 7
Aphis spiraecola 7 3 3
Toxoptera aurantii 0 8 0
Aphis craccivora 0 1 0
Aphis fabae 0 0 0
Myzus persicae 2 2 1
Otoño 2004
NI 3 2 3
NI: no identificado, por estructuras anatómicas incompletas
El hecho de que en la primera temporada de captura en Ovalle no se hayan detectado
pulgones, y en la segunda haya sido A. gossypii el áfido mas frecuente, se explica
con lo que señala GOTTWALD et al. (1996), en donde plantea que por el hecho de
que A. gossypii posee hábitos migratorios, la presencia de éste varía desde una escasa
actividad de alados a aumentos bruscos, indicativo de intensas migraciones de áfidos.

47
En el caso de Limache tampoco se observó gran cantidad de especies aladas de
áfidos, la especie capturada más frecuente en las dos temporadas de captura fue T.
aurantii, seguido por A. gossypii. Se ha visto que Toxoptera aurantii es capaz de
formar grandes colonias en cítricos, los cuales son un hospedero primario de esta
especie de pulgones (GOTTWALD et al., 1996).
En el huerto de Paine, al igual que en los otros dos predios, tampoco se observó
abundancia de áfidos, el pulgón más frecuente en las dos temporadas de captura fue
A. gossypii, en segundo lugar de frecuencia se detectó a A. spiraecola y a T. aurantii.
La baja presencia de áfidos observada en los tres predios, se debió posiblemente a los
manejos realizados en los huertos y a la presencia de numerosos enemigos naturales,
ya que durante las capturas se observó gran cantidad de controladores asociados a los
áfidos, entre los más frecuentes están: Lysiphlebus testaceipes, Adalia deficiens,
Adalia bipunctata, Scymnus bicolor y Chrysoperla, entre otros. El parasitoide L.
testaceipes tiene la característica de que utiliza varias especies de áfidos para
reproducirse. También se destacan depredadores como A. bipunctata que es el
coccinélido que se encuentra con mayor frecuencia asociado a T. aurantii y a A.
gossypii, por su parte el neuróptero Chrysoperla sp es un importante depredador que
controla a la mayoría de las especies de áfidos presentes en Chile (RIPA, 1999).
A pesar de esto, los resultados indican que A. gossypii, está presente en los tres
huertos, mostrando presencia predominante en ciertas épocas del año y bajo
condiciones favorables para su desarrollo en cítricos. Esto es una amenaza latente si
se considera que A. gossypii es el segundo vector más eficiente de CTV en el mundo.
En España CTV se difunde naturalmente por medio de T. aurantii, A. spiraecola y A.
gossypii. Este último es el más eficiente en transmitir las razas allí presente
(CAMBRA et al., 1990).

48
4.2. Impresión o aplastado de áfidos
Las especies de pulgones aplastados correspondieron a A gossypii y a A. spiraecola,
ya que estos son los posibles principales diseminadores de CTV en Chile. Los
resultados de la prueba nested-PCR realizada a los áfidos colectados en los huertos de
Limache y Paine se presentan a continuación en el Cuadro 4.
Cuadro 4: Análisis nested-PCR para detectar CTV en áfidos capturados en las
localidades de Limache y Paine durante la primavera del 2003
Lugar de
origen
Especie escachada Número individuos
escachados
Nested-PCR
positivo
Aphis gossypii 3 1 Limache
Aphis spiraecola 1 0
Aphis gossypii 8 5 Paine
Aphis spiraecola 2 0
De los áfidos capturados en el predio de Limache, se detectó la presencia de
partículas virales de CTV sólo en uno de los tres A. gossypii aplastados. En el caso de
A. spiraecola, se aplastó un individuo resultando con respuesta negativa a la
presencia de CTV. En cambio Paine fue un poco diferente, en este huerto, además de
capturar un mayor número de áfidos, éstos se encontraron en su mayoría virulíferos,
detectándose cinco individuos de A. gossypii portando el virus de un total de ocho
áfidos de esta especie aplastados. También se aplastaron dos A. spiraecola entregando
ambos respuestas negativa a la presencia del virus (Cuadro 4).
Esto indica que probablemente A. gossypii sea el principal vector de CTV en Chile, y
el que estaría transmitiendo el virus de árboles enfermos a árboles sanos,
coincidiendo la mayor proporción de árboles infectados con una mayor cantidad de
áfidos virulíferos presentes, como es el caso de Paine, el cual, al comparar los tres

49
huertos en estudio, es el que presenta mayor incidencia de la enfermedad y un mayor
número de áfidos virulíferos.
4.3. Estudio sobre la distribución espacial
4.3.1 Huerto de Ovalle
En este huerto, se encontraron sólo siete árboles infectados de un total de 400 árboles
analizados, lo que entrega una proporción de 1,75% árboles con CTV, lo cual es una
proporción baja en comparación con los otros huertos en estudio.
Además al comparar los resultados de los dos muestreos realizados en este huerto, se
observa que estos son idénticos, es decir, no hay desplazamiento del virus en el
período transcurrido entre los dos muestreos, por consiguiente, se mantiene la misma
proporción de árboles infectados.
En este caso, el mapa del huerto se dividió en 50 cuadrículas, de ocho árboles cada
uno. En el Anexo 1, se muestra el número de árboles infectados en relación al número
de cuadrículas.
Una vez realizado el análisis y con los datos obtenidos: =λ̂ 0,14 (λ: Número medio
estimado de árboles infectados por cuadrícula); 2observadoχ =0,89, se puede decir que no
se rechaza la hipótesis que la distribución de frecuencias es Poisson. Por lo tanto, la
distribución es al azar.
A continuación se presentan los valores de los índices de dispersión: Índice de
dispersión (ID) =1,1691; Índice de agrupamiento (IC)= 0,1691; Índice de Morosita
( dΙ ): 2,5758.

50
Al analizar los resultados, estos indican que los árboles se distribuyen aleatoriamente
en el huerto, lo que apoya la hipótesis de que los árboles están distribuidos al azar. En
el Anexo 2, se muestra la incidencia y distribución de los árboles infectados con CTV
en Ovalle, en las dos épocas de muestreo.
Por otra parte, los resultados de Inmunoimpresión-ELISA indicaron que en todos los
casos en donde la variedad estaba infectada lo estaba el portainjerto, coincidiendo en
todos los casos variedad y portainjerto infectados en el mismo árbol. Tal efecto puede
ser consecuencia de una contaminación del patrón previa a la plantación o por
rebrotes, ya que al emplearse C. macrophylla como patrón para limoneros, tiene las
limitantes de que se contamine en vivero antes del injerto y la tendencia a emitir
rebrotes en las raíces, las que se pueden infectar con el virus (NAVARRO, 1981). En
este predio, se descarta la posibilidad de que estos árboles hayan sido infectados por
pulgones en el huerto, ya que según el “Proyecto de prospección de razas severas en
Chile” y muestreos realizados en este predio, el virus no se ha desplazado a otros
árboles del sector, desde el año 2002 hasta el último muestreo realizado en este
estudio.
Además ensayos realizados por HERMOSO DE MENDOZA et al., (1986), indican
que cuando el limonero es el donante y a la vez receptor de CTV, hay un 0% de
transmisión del virus, siendo a la vez el limonero el peor receptor cuando el donante
es otra especie de cítrico de interés comercial.
4.3.2 Huerto Limache
En este huerto, la incidencia es mayor que en el caso anterior, encontrándose, en el
primer muestreo 15 árboles infectados de un total de 420, lo que da una proporción de
3,57% árboles infectados.

51
En el segundo muestreo, el número de árboles infectados aumentó de 15 a 17,
llegando a un 4% de los árboles infectados con CTV. Esto quiere decir que el virus se
está moviendo y está siendo transmitido por los áfidos que visitan este predio.
Para realizar el análisis de distribución del virus el huerto, se dividió en 60
cuadrículas de siete árboles cada una. En el Anexo 3, se muestra el número de
cuadrículas con el número de árboles infectados en los dos muestreos realizados.
Al realizar el análisis estadístico de los datos obtenidos en el primer muestreo se
obtienen los siguientes resultados: =λ̂ 0,25 (λ: Número medio estimado de árboles
infectados por cuadrícula); 2observadoχ =0,147, los cuales indican que no se rechaza la
hipótesis que la distribución de frecuencias es Poisson. Por lo tanto, la distribución
es al azar.
Al analizar los datos del segundo muestreo se obtienen los siguientes resultados:
=λ̂ 0,28333; 2observadoχ =0,00386, lo cual también dice que no se rechaza la hipótesis
que la distribución de frecuencias es Poisson.
Por lo tanto se mantiene la distribución de los árboles infectados al azar, y a pesar de
que aumenta la proporción de árboles infectados, esta no es estadísticamente
diferente. Los mapas con la incidencia y la distribución de la enfermedad en la
primavera del 2003 y el otoño del 2004 se presentan en Anexo 4 y 5 respectivamente.
A continuación se presentan los valores de los índices de dispersión obtenidos con
datos del segundo muestreo: Índice de dispersión (ID) = 0,9681; Índice de
agrupamiento (IC) = -0,0319; Índice de Morosita ( dΙ ) = 0,9429.

52
Estos resultados reafirman claramente la hipótesis que plantea que se trata de un
modelo de distribución al azar
4.3.3 Huerto Paine
De los tres huertos en estudio este es el que presentó la mayor proporción de árboles
infectados. En el primer muestreo de un total de 400 árboles, se encontraron 24
respuestas positivas a CTV, lo que equivale a un 6% de incidencia.
En el segundo muestreo, los árboles infectados aumentaron a 25, lo que elevó
levemente la proporción de árboles infectados a 6,25%. En este caso, este aumento en
los árboles infectados también se le atribuye a los áfidos presentes en el huerto.
En este predio, el mapa del huerto se dividió en 50 cuadrículas, de ocho árboles cada
una. El número de árboles infectados por cuadrícula en los dos muestreos, se muestra
en el Anexo 6.
Los datos obtenidos en el primer muestreo, entregaron los siguientes
resultados: =λ̂ 0,48 (λ: Número medio estimado de árboles infectados por
cuadrícula); 2observadoχ = 1,138, una vez analizados, indican que no se rechaza la
hipótesis que la distribución de frecuencias es Poisson. Por lo tanto, la distribución
de los árboles infectados es al azar.
En el segundo muestreo los resultados no fueron muy diferentes y arrojaron los
siguientes resultados: =λ̂ 0,46; 2observadoχ = 1,998, por lo tanto tampoco se rechaza la
hipótesis que la distribución de frecuencias es Poisson, manteniéndose la
distribución al azar.

53
A continuación se presentan los índices de dispersión obtenidos con datos del
segundo muestreo: Índice de dispersión (ID) = 0,7856; Índice de agrupamiento (IC)
= -0,2144; Índice de Morosita ( dΙ ) = 0,5774.
Los resultados obtenidos, indican una distribución al azar, pero en este caso, los
valores no lo expresan en forma tan marcada como fue el caso del predio anterior, lo
que podría confundirse con una distribución uniforme. Sin embargo al complementar
los datos obtenidos, se puede decir con mayor certeza que en este predio la
distribución de CTV no es significativamente diferente a una distribución al azar.
Por otro lado, al comparar las diferencias de proporciones, se puede decir que el
aumento en la proporción de los árboles infectados no es estadísticamente
significativo. Los mapas con la incidencia y distribución de la enfermedad en la
primavera del 2003 y el otoño del 2004 se presentan en Anexo 7 y 8,
respectivamente.
Por lo tanto, los resultados obtenidos indican que si bien el virus se está moviendo en
los huertos de Paine y Limache, este movimiento hacia otros árboles en el sector, no
provocó cambios en el patrón de dispersión al azar de CTV, ni tampoco hubo un
aumento significativo en la proporción de árboles infectados entre el primer y
segundo muestreo.
Este patrón de distribución al azar de CTV en los tres huertos, sugiere que las
infecciones fueron provocadas por los áfidos vectores presentes en ellos,
descartándose la posibilidad de que el virus se transmitió por injerto en vivero, ya que
de ser así el patrón de dispersión de CTV sería mas uniforme o regular,
encontrándose un mayor número de plantas infectadas. TORRES (2002), en un
estudio de prospección realizado en huertos de Pica y Matilla, al analizar la

54
distribución espacial, determina que CTV en estos huertos se distribuye con un
grado de agrupamiento o en forma regular, y señala que esto es atribuible a la
longevidad y a manejos propios del huerto, y a la alta sensibilidad a CTV de las
especies presentes como es el caso de lima mexicana. En contraste, los huertos
analizados en este estudio son relativamente nuevos, en plena producción y con una
combinación patrón/variedad tolerante a CTV. Todos estos factores estarían dando
como resultado una distribución al azar de CTV.
Por lo tanto, la diseminación de la enfermedad posiblemente está siendo provocada
por A. gossypii. En áreas como California, España, Florida donde A. gossypii es el
vector predominante, los hábitos migratorios de éste dan lugar a que las nuevas
infecciones ocurran preferentemente en árboles no adyacentes a los árboles ya
infectados, lo que hace que los árboles con CTV parezcan distribuidos al azar en los
huertos (GOTTWALD et al., 1995).
MORENO (1995), según ensayos realizados con áfidos presentes en República
Dominicana, indica que la eficiencia de transmisión de un individuo de T. citricida es
de 11%, la de A. spiraecola es de 0,3% y A. gossypii, T. aurantii y T. odinae no
parecen transmitir los aislados locales de CTV. Es de señalar, no obstante, que la
capacidad o ineficiencia de estas especies para transmitir CTV cuando el virus se
introduce en un país es sólo temporal, según se deduce de las experiencias de
California, Israel y España, y que pueden transcurrir 20 – 30 años hasta que el virus y
vector se adaptan y comienza la difusión epidémica. Por consiguiente, considerando
lo señalado anteriormente, la tasa de diseminación de CTV en estos huertos puede
aumentar. Sin embargo, este punto no se debe sobredimensionar, ya que a pesar de
que el virus está siendo diseminado, en Chile la gran mayoría de los huertos están
plantados con patrones tolerantes, y además existe un Programa de Certificación de
cítricos libres de enfermedades.

55
4.4. Transmisión de CTV por Aphis gossypii
4.4.1 Crianza de Aphis gossypii
Se logró formar colonias de A. gossypii sobre plantas de algodón, los cuales por
medio de un pincel, se fueron traspasando a otras plantas de algodón para formar
nuevas colonias y aumentar así la población sin agotar el sustrato, ya que los áfidos
tienen la característica poblacional de colonizar un hospedero mediante hembras
aladas, desarrollar la población mediante partenogénesis, hasta que comienzan a
agotar el espacio y el sustrato, momento en el cual comienza una gran formación de
individuos alados que se dirigirán a colonizar otras áreas del mismo vegetal u otro
hospedero en la cercanía, extinguiéndose la población inicial (PRADO, 1991).
La crianza se vio favorecida por la presencia de hormigas, una buena iluminación,
temperatura controlada (entre 18 – 26° C) y alta humedad relativa. Un aspecto
importante es el control de la temperatura, tanto para las crianzas como para las
transmisiones, las que en otros países en su mayoría son realizadas en invernadero
con temperaturas en el rango de 18 a 27 °C, y eventualmente con iluminación con
fotoperíodo para crianza, e iluminación las 24 horas para la transmisión (YOKOMI y
GARNSEY, 1987; HERMOSO DE MENDOZA et al., 1986).
El resultado fue el crecimiento exponencial de la población de áfidos confinados,
llegando a tener 14000 pulgones en un período de tres meses, cantidad suficiente para
poder desarrollar en forma exitosa el proceso de transmisión de CTV. El hecho de
lograr rápidamente un buen número de áfidos, se debe a que A. gossypii se reproduce
habitualmente por partenogénesis telitóquita, esto da lugar sólo a hembras, las cuales
dan origen a aproximadamente 60 descendientes y el incremento de la población es
rápido, dado que una generación puede ser completada en ocho días (RIPA, 1999).

56
4.4.2 Transmisión
En esta etapa, se desarrolló la metodología para todo el proceso de transmisión de
CTV, desde la adquisición hasta la donación a plantas receptoras por parte de Aphis
gossypii. Para ello, se ideó un sistema de tubos cerrados con malla antiáfidos por
ambos costados, los cuales resultaron prácticos y eficientes al utilizarlos en el proceso
de transmisión de CTV por áfidos (Figura 1).
Es importante destacar que si bien los resultados de la transmisión de CTV, no están
disponibles para ser expuestos en este estudio, todo el proceso de la transmisión se
realizó paso a paso, cumpliendo con todos las etapas que la metodología indicó seguir
para poder transmitir con éxito aislados de CTV. Esta metodología, se adaptó a
nuestras condiciones, según experiencias probadas en diferentes situaciones en
Estados Unidos, Israel y España principalmente. Es así como HERMOSO DE
MENDOZA, BALLESTER-OLMOS y PINA (1986), comparó los porcentajes de
transmisión de dos tipos de razas de CTV diferentes (un aislado común en España y
otro de reciente introducción que provoca amarillez de plántulas “seedling yellows”),
por medio de cuatro especies de áfidos, obteniendo diferencias en la eficiencia de
transmisión en relación a especies de áfidos y tipo de aislado.
Cabe señalar que después de un mes de realizada la transmisión las plantas no han
manifestado síntomas, esto se debe a que el virus requiere de un período de
incubación, el cual en sentido general es muy variable, pudiendo ir de tres a cuatro
semanas hasta años (CARRERO, 1981). No obstante, los síntomas en plantas
indicadoras aparecen desde los tres a seis meses, desde realizada la transmisión
(NAVARRO, 1981).
Por lo tanto, para evaluar la presencia de CTV en cada planta receptora se efectuará
Inmunoimpresión-ELISA a los seis meses de realizada la transmisión.

57
a b
c d Figura 1. Proceso de transmisión de CTV por medio de Aphis gossypi: (a) planta
donante de aislados de CTV con sus brotes marcados, (b) crianza de A. gossypii sobre plantas de algodón, (c) áfidos alimentándose de plantas donantes, (d) áfidos alimentándose de plantas receptoras.

58
5. CONCLUSIONES
El estudio epidemiológico de CTV, permitió concluir que en los tres huertos la
distribución espacial de CTV es al azar, y a pesar de que el virus aparentemente se
está dispersando en los predios de Limache y Paine, la distribución se mantuvo al
azar entre las dos épocas de muestreo. No obstante, si bien en estos dos predios
aumentó el número de árboles infectados, se puede decir que el aumento en la
proporción de los árboles infectados no es significativo. En el caso del huerto de
Ovalle no se detectó diseminación del virus.
Mediante la metodología del árbol pegajoso, se pudo capturar áfidos alados que
visitan predios de cítricos, y con ello establecer la presencia de los áfidos
predominantes y virulíferos en tres huertos de cítricos de la zona central de Chile. Los
áfidos virulíferos que se detectaron mediante nested-PCR, resultaron ser todos A.
gossypii, esto, junto con el patrón de distribución al azar de la enfermedad de los
predios en estudio, indica que posiblemente es esta especie la que está dispersando
CTV en estos huertos.
Por otro lado, se logró establecer la metodología de crianza de A. gossypii y del
proceso de transmisión de aislados de CTV por medio de este áfido, el que incluyó la
creación de un sistema de tubos específicos para transmisión del virus por medio de
áfidos.

59
6. RESUMEN
El virus de la tristeza de los cítricos (CTV), ha provocado pérdidas millonarias para el patrimonio de la citricultura mundial a lo largo de todo un siglo. Sin embargo, en Chile, no se han manifestado síntomas de carácter severo en las principales zonas citrícolas, lo cual no indica que el virus no este presente, existen antecedentes de que CTV se encuentra en el país desde 1969. Esto ha sido corroborado por estudios posteriores, como la extensa prospección realizada entre 1999 y 2002. Sin estar ajeno a lo planteado, los estudios de epidemiología y transmisión de CTV realizados en este trabajo buscan recopilar antecedentes sobre el carácter real de la enfermedad y su potencial diseminación en Chile. Por consiguiente, con los resultados obtenidos se ha podido determinar el patrón de distribución de la enfermedad y su incidencia en tres huertos de la zona central de Chile, además se ha podido determinar que en los huertos estudiados de Limache y Paine el virus se está dispersando, pero a bajas proporciones. No obstante, en estos huertos no se espera una expresión de la enfermedad, ya que su combinación “variedad/portainjerto” es tolerante a tristeza. Otra parte de este estudio epidemiológico consistió en capturar áfidos alados que visitan los predios cítricos ( por medio del método del árbol pegajoso), con el fin de determinar la presencia de los áfidos predominantes, y por medio de la prueba de nested-PCR establecer la existencia de áfidos virulíferos. Los resultados indicaron en general para los tres predios a A. gossypii y A. spiraecola como la especie más frecuente, estas especies fueron sometidas a la prueba de nested-PCR obteniéndose que sólo A. gossypii presentaba partículas virales de CTV. Por otro lado, se logró establecer la metodología de crianza de A. gossypii, y del proceso de transmisión de aislados de CTV por medio de este áfido.

60
7. ABSTRACT
Citrus Tristeza Virus (CTV) has caused millions in losses to the citrus industry worldwide over the last century. However, the major citrus-producing regions of Chile have not show severe symptoms of decay or stemp pitting, which does not necessarily indicate that the virus is absent, as there have been reports of the presence of CTV in Chile since 1969. This has been corroborated by an extensive survey done between 1999 and 2002. Considering this, the epidemiologic and CTV transmission studies done in this work attempted to compile records on the actual character of the disease and its potential spread in Chile. Consequently, the results obtained have made it possible to determine the distribution pattern of the disease as well as its incidence in three orchards in the Central Zone of Chile, additionally it was determined that for the orchards studied in Limache and Paine, the virus is spreading, although on a small scale. Nevertheless, expression of the disease is not expected in these orchards, as their variety/rootstock combination is tolerant to CTV. Another part of this epidemiologic study consisted of catching winged aphids which fly over citrus plantings (using a sticky tree), to determine the predominant aphid species visiting these orchards, and to establish the existence of virulent aphids using a nested-PCR test. Results indicated that in general A. gossypii and A. spiraecola were the most frequent visitors to the three sites. When tested using nested-PCR, only A. gossypii was found to contain CTV particles. A methodology for breeding A. gossypii was established, as well as the process for the transmission of CTV isolates by this aphid.

61
8. LITERATURA CITADA
BALLESTER- OLMOS, J. F., CARBONELL, E. A., MORENO, P., HERMOSO DE MENDOZA, A., CAMBRA, M. and NAVARRO, L. 1993. Biological diversity of citrus tristeza virus (CTV) in Spain. Plant Pathology 42 (2): 219 – 229.
BAR-JOSEPH, M., ROISTACHER, C. N., GARNSEY, M. and GUMPF, D. J. 1981.
A review of tristeza, an ongoing threat to citriculture. International Society Citriculture. Proceedings of the 6th Conference. pp. 419 – 423.
________., 1978. Cross protection incompleteness: a possible cause for natural
spread of citrus tristeza virus after a prolonged lag period in Israel. Phytopathology 68 (8): 1110 – 1111.
BESOAIN, X., RAMELLA, F., CANALES, C., VALENZUELA, M., CASTRO, M.,
GORRIS, M.T. y CAMBRA, M. 2003b. Caracterización serológica de 100 aislados de Citrus tristeza virus (CTV), colectados desde las principales regiones citrícolas de Chile. 13° Congreso Sociedad Chilena de Fitopatología. Marbella 28 – 31 de octubre del 2003 (abstract. Original no consultado).
________., SIMPSON,_M. C., RAMELLA, F., CANALES, C., VALENZUELA, M.,
CASTRO, M., TORELLI, L. y CAMBRA, M. 2003a. Incidencia del virus de la tristeza de los cítricos en Chile. 13° Congreso Sociedad Chilena de Fitopatología. Marbella 28 – 31 octubre del 2003 ( abstract. Original no consultado).
________., CANALES, C., RAMELLA, F., SIMPSON,_M. C. y CAMBRA, M.
2002. Decaimiento y acanaladuras de la madera en pomelos (Citrus paradisi) en el oasis de Pica y su asociación con el virus de la tristeza (CTV). 12° Congreso Sociedad Chilena de Fitopatología. Puerto Varas, octubre del 2002 (abstract. Original no consultado).
________., VALENZUELA, M., CASTRO, M. P., y BALLESTER-OLMOS, J.F. _________2000. Situación actual de las enfermedades tipo virus que afectan a los cítricos en Chile. Fitopatología 35(2): 98 -104. ________., 1998. Virosis en cítricos. In: Seminario “Incidencia, sintomatología y
control de virus en frutales”. PUC-HORTIFRUT. Santiago 23-25 de marzo del 1998. pp 30 – 42.

62
________., VALENZUELA, M., CASTRO, M. y BALLESTER-OLMOS, J. F. 1997. Situación actual de enfermedades tipo-virus que afectan a los cítricos en Chile. Sociedad Uruguaya de Fitopatología. IX Congreso Latinoamericano de Fitopatología, Montevideo, agosto 1997. 104 p (Abstract. Original no consultado).
BOVE, J. M. and VOGEL, R. 1980. Descriptions and illustrations of virus and
viruslike diseases of citrus. A collection of color slides. Paris. SETCO- INRA. Vol 4. 108 p.
CAMBRA, M., OLMOS, A., GORRIS, M. T., MARROQUIN, C., ESTEBAN, O.,
GARNSEY, S. M., LLAUGER, R., BATISTA, L., PEÑA, I. and HERMOSO DE MENDOZA, A. 2000. Detection of citrus tristeza virus by print capture and squash capture-PCR in plant tisúes and single aphids. Riverside, International Organization Citrus Virology. Proceedings of the 14th Conference of IOCV. pp 42 – 48.
________., y MORENO, P. 2000. Enfermedades de los cítricos. Madrid, Ediciones Mundi-Prensa. 213 p. (vol. 2). ________., 1994. El virus de la tristeza en la provincia de Castellón. Infección actual, previsiones y recomendaciones. Levante Agrícola 33(327): 105- 111. _______ ., GORRIS, M. T., SERRA, J., CAMARASA, E., PINA, J. A., SANZ,
A., VELA, C. y LLATSER, F. 1991. Diagnostico y estudio del virus de la tristeza de los cítricos utilizando técnicas inmunoenzimáticas con anticuerpos monoclonales específicos. Levante agrícola 30 (313): 176 – 189.
________., SERRA, J., MARTINEZ, J. C., VILLALVA, D. 1990. Situación de la
tristeza de los cítricos en la Comunidad Valenciana. Levante. Agrícola 29(305): 288 – 296.
CANALES, C., BESOAÍN, X., VALENZUELA, M., CASTRO, M., SIMPSON,
M.C., TORELLI, L. 2001. Prospección de aislados de CTV y sus vectores en la zona comprendida entre la primera Región y la Provincia de Melipilla de la Región Metropolitana. Sociedad Chilena de Fitopatología. 52° Congreso Agronómico Chile. Quillota 17 – 19 octubre del 2001. Puerto Varas, octubre del 2002 ( abstract. Original no consultado).

63
CARRERO, J. M. 1981. Virosis y enfermedades afines de los cítricos. 2ª Edición Arabí, Musigraf. 411p.
CHARLIN, R. 1975. Identificación de los áfidos de los citrus. Investigación Agrícola
1 (2): pp. 110 – 114. CHILE, MINISTERIO DE AGRICULTURA 1968. Decreto Ley N° 15. Modificación
Reglamento de viveros. 42p. GARDIAZABAL, F. y MAGDHAL, C. 1998. Seminario Internacional de Cítricos.
Viña del Mar, 13 – 15 mayo de 1998. 220p. GARNSEY, S. M. and MULLER, G. W. 1986. Efficiency of mechanical
transmission of citrus tristeza virus. Riverside, International Organization Citrus Virology. Proceedings of the 10th Conference of IOCV. pp. 46 - 54.
GORRIS, M. T., MARTINEZ, M. C. y CAMBRA, M. 2000. Detección simple y
fiable del virus de la tristeza de los cítricos (CTV) mediante Inmunoimpresión – ELISA. Levante Agrícola 39 (350): 69 – 74.
GOTTWALD, T. R., GARNSEY, S. M. and GIBSON, G. 2000. The effect of aphid
vector population composition on local and background components of citrus tristeza virus spread. Riverside, International Organization Citrus Virology. Proceedings of the 14th Conference of IOCV. pp. 88 – 94.
________., GIBSON, G. J., GARNSEY, S. M. and IREY, M. 1999. Examination of
the effect of aphid vector population composition on the spatial dynamics of citrus tristeza virus spread by stochastic modeling. Phytopathology 7 (89): 603 – 606.
________., GARNSEY, S. M. and BORBÓN, J. 1998. Increase and patterns of
spread of citrus tristeza virus infections in Costa Rica and the Dominican Republic in the presence of the brown citrus aphid, Toxoptera citricida. Phytopathology 7 (88): 621 – 635.
________., GARNSEY, S. M., CAMBRA, M., MORENO, P., IREY, M. and
BORBÓN, J. 1996. Differential effecs of Toxoptera citricida vs. Aphis gossypii on temporal increase and spatial patterns of spread of citrus tristeza. Riverside, International Organization Citrus Virology. Proceedings of the 13th Conference of IOCV. pp. 121 – 128.

64
________., CAMBRA, M., MORENO, P., CAMARASA, E. and PIQUER, J. 1995. Spatial and temporal analices of citrus tristeza virus in Eastern Spain. Phytopathology 1(86): 45 – 55.
________., CAMBRA, M. and MORENO, P. 1993. The use of serological assays to
monitor spatial and temporal spread of citrus tristeza virus in symptomless trees in Eastern Spain. Riverside, International Organization Citrus Virology. Proceedings of the 12th Conference of IOCV. pp. 51 – 61.
HERMOSO DE MENDOZA, A., BALLESTER-OLMOS, J. F. and PINA. J. A.
1986. Comparative aphid transmission of a common citrus tristeza virus isolates and seedling yellows isolate recently introduced into Spain. Riverside, International Organization Citrus Virology. Proceedings of the 10th Conference of IOCV. pp 68 – 70.
________., BALLESTER-OLMOS, J. F., PINA. J. A. SERRA, J. and FUERTES, C.
1986. Differences in transmisión efficiency of citrus tristeza virus by Aphis gossypii using sweet orange, mandarin or lemon trees as donor or receptor hosts plants. Riverside, International Organization Citrus Virology. Proceedings of the 10th Conference of IOCV. pp 62 – 64.
________., BALLESTER-OLMOS, J. F. and PINA. J. A. 1984. Transmisión of
citrus tristeza virus by aphids (Homoptera Aphididae) in Spain. Riverside, International Organization Citrus Virology. Proceedings of the 9th Conference of IOCV. pp 23 – 27.
HERRERA, G., MADARIAGA, M. y SANTELICES, M. 1995. Detección del virus
de la tristeza de los cítricos, en la zona central de Chile. Agricultura Técnica 55 (2) :167 – 169.
INSTITUTO NACIONAL DE ESTADÍSTICA. 1997. 6º Senso Nacional
Agropecuario. Santiago, INE. 214p. LEE, R. F., DERRICK, K. S., NIBLET, C. L. and PAPPU, H. R. 1995. When to use
mild strain cross protection (MSCP) and problems encountered. In: Proceedings 3th Int. Workshop citrus tristeza virus and brown citrus aphid in the Caribbian basin: Management Strategies. Lake Alfred, Florida: 158 – 162.
LUDWIG, J. A. and REYNOLDS, J. F. 1988. Statistical Ecology: A primer on
methods and computing. New York, Whiley. 337p.

65
MORENO, P., 1995. La tristeza y el pulgón pardo de los cítricos (Toxoptera citricidus Kirk.): Una amenaza inmediata para la citricultura en los países ribereños del caribe. Levante Agrícola 34 (333): 316– 325.
________., 1992. El virus de la tristeza. Aspectos importantes para su
caracterización. Master en Citricultura. Valencia. Universidad Politécnica de Valencia. 8 p.
________., GUERRI, J. and MUNOZ, N. 1990. Identification of spanish strains of
citrus tristeza virus by analices of double-stranded RNA. Phytopathology 80 (5): 477 – 482.
NARANJO, M. 1997a. Reseña bibliográfica sobre la enfermedad tristeza de _________los cítricos. Levante Agrícola 36 (340): 227 – 240. ________., 1997b. Reseña bibliográfica sobre la enfermedad tristeza de los _________cítricos. Levante Agrícola 36 (341): 355 -368. NAVARRO, L. 1981. Versión de la conferencia ofrecida por el Dr. Luis Navarro
Lucas sobre la enfermedad de la tristeza de los cítricos. La Habana. CIDA. 33p.
ORGANIZACIÒN DE LAS NACIONES UNIDAS PARA LA AGRICULTURA Y
LA ALIMENTACIÓN. 2004. Cuadro de superficies de cultivo a nivel mundial, (on line). http://www.fao.org.
PRADO, E. 1991. Artrópodos y sus enemigos naturales asociados a plantas ________ cultivadas en Chile. Santiago, INIA. 207p. (Bol. Técnico N° 169). RACCAH, B., LOEBENSTEIN, G. and BAR-JOSEPH, M. 1976. Transmission of
citrus tristeza virus by the melon aphid. Phytopathology (66): 1102 – 1104.
RIPA, R. y RODRIGUEZ, F. 1999. Plagas de los cítricos, sus enemigos y manejo.
Santiago, INIA. 151p. ROCHA-PEÑA, M. A., LEE, R., LASTRA, R., NIBLETT, C., OCHOA-CORONA, _________F., GARNSEY, S. and YOKOMI, R. 1995. Citrus tristeza virus its _________aphis vector Toxoptera citricida. Plant Disease. 79 (5): 437 – 445. ________., LEE, R. F. and NIBLETT, C. L. 1993. Effectiveness of diferent citrus
species as donor host for graft transmission of citrus tristeza virus. Riverside, International Organization Citrus Virology. Proceedings 12th
Conference of IOCV. pp. 84- 92.

66
ROISTACHER, C. N. and MORENO, P. 1991. Tristeza. The wordlwide threath from
destructive isolates of citrus tristeza virus. Riverside, International Organization Citrus Virology. Proceedings of the 11th Conference of IOCV. pp. 7- 19.
________., and BAR-JOSEPH. 1987. Transmission of citrus tristeza virus (CTV) by
Aphis gossypii and by graft inoculation to and from passiflora spp. Phytophylactica 19: 179 – 182.
SANCHEZ, L. 1996. La tristeza de los cítricos en Chile. In: Estereo, M y
Magunacelaya, J. eds. Avances en sanidad vegetal de frutales y vides. Santiago. Universidad de Chile. pp. 155 – 159.
________., y WEATHERS, L. 1970. La amenaza de la tristeza de los cítricos en
Chile. Agricultura Técnica 30 (2): 166 – 170. TORRES, R., 2002. Análisis de la situación de la tristeza de los cítricos en huertos
comerciales ubicados en la I y V región del país. Taller de licenciatura Ing. Agr. Quillota. Universidad Católica de Valparaíso. Facultad de agronomía. 79 p.
WEATHERS, L., SANCHEZ, L. y PLATT, R. 1969. Naturaleza y distribución de las
enfermedades virosas de cítricos en Chile. Agricultura Técnica 29 (4): 166 – 170.
YOKOMI, R. K. 1995. Why the Concern about spread of brown citrus aphids into
new citrus areas? In: Proceedings 3 er Int. Workshop citrus tristeza virus and brown citrus aphid in the Caribbian basin: Management Strategies. Lake Alfred, Florida: 27 – 31.
________., LASTRA, R., STOETZEL M. B., DAMSTEEGT, V. D., LEE, R. F.,
GARNSEY, S. M., GOTTWALD, T. R., ROCHA-PEÑA, M. A. and NIBLETT, C. L. 1994. Establishment of the brown citrus aphid (Homoptera Aphididae) in Central America and the Caribbean basin and transmission of citrus tristeza virus. Journal of Economic Entomology 87(4): 1078 – 1085.
________., and GARNSEY, S. M. 1987. Tansmission of citrus tristeza virus by Aphis
gossypi and Aphis spiraecola in Florida. Phytophylactica 19: 169 – 172.


















