
2 Puesto que el ingreso de calcio a la planta depende del...
54
1. INTRODUCCIÓN En Chile, el cultivo de tomate como cultivo forzado se realiza en invernaderos sin calefacción denominados fríos (GIACONNI y ESCAFF, 1998), por no poseer equipos de control de factores ambientales, como la temperatura, humedad e intensidad de luz. El manejo de estos factores en este tipo de invernaderos sólo se puede realizar en forma relativa a través de la construcción y orientación de éstos y de las cubiertas utilizadas, lo que los hace muy dependientes de las condiciones del medio. Los factores ambientales ejercen influencia sobre los procesos nutritivos de las plantas, y éstos afectan al desarrollo, crecimiento y producción de las mismas, principalmente porque afectan a su fisiología (GUZMÁN, 2000). En los cultivos de tomate en la modalidad de tomate primor temprano plantados en el mes de mayo, en invernaderos fríos, donde las bajas temperaturas y la alta humedad de este mes son difíciles de manejar en forma óptima, se genera un ambiente en el interior del invernadero con temperaturas levemente superiores a las externas y una humedad relativa muy alta. Cuando la humedad interior es alta, el bajo déficit de presión de vapor entre las hojas y el aire hace que la transpiración se reduzca, afectando negativamente al desarrollo de las plantas, lo que influye en el normal crecimiento de ellas (NAVAS, 2000).
Transcript of 2 Puesto que el ingreso de calcio a la planta depende del...

1. INTRODUCCIÓN
En Chile, el cultivo de tomate como cultivo forzado se realiza en invernaderos
sin calefacción denominados fríos (GIACONNI y ESCAFF, 1998), por no
poseer equipos de control de factores ambientales, como la temperatura,
humedad e intensidad de luz.
El manejo de estos factores en este tipo de invernaderos sólo se puede
realizar en forma relativa a través de la construcción y orientación de éstos y
de las cubiertas utilizadas, lo que los hace muy dependientes de las
condiciones del medio.
Los factores ambientales ejercen influencia sobre los procesos nutritivos de
las plantas, y éstos afectan al desarrollo, crecimiento y producción de las
mismas, principalmente porque afectan a su fisiología (GUZMÁN, 2000).
En los cultivos de tomate en la modalidad de tomate primor temprano
plantados en el mes de mayo, en invernaderos fríos, donde las bajas
temperaturas y la alta humedad de este mes son difíciles de manejar en
forma óptima, se genera un ambiente en el interior del invernadero con
temperaturas levemente superiores a las externas y una humedad relativa
muy alta.
Cuando la humedad interior es alta, el bajo déficit de presión de vapor entre
las hojas y el aire hace que la transpiración se reduzca, afectando
negativamente al desarrollo de las plantas, lo que influye en el normal
crecimiento de ellas (NAVAS, 2000).

2
Puesto que el ingreso de calcio a la planta depende del flujo transpiratorio, la
disminución de éste genera problemas de absorción de calcio. Este efecto
provoca una deficiencia de calcio, que se ve manifestada en una clorosis
marginal de los foliolos de las hojas en el tomate, que posteriormente se
necrosa, aumentando la susceptibilidad a enfermedades como Botrytis
cinerea (ADAMS, 1986).
Los objetivos de la presente investigación son establecer si existe relación
entre los contenidos de calcio foliar y la incidencia de clorosis apical, el grado
de asociación de dicho síntoma con el ataque de Botrytis cinerea sobre los
tejido afectados, y evaluar el efecto de la aplicación foliar de óxido de calcio
sobre la incidencia de clorosis apical y la incidencia de Botrytis cinerea.

3
2. REVISIÓN BIBLIOGRÁFICA
2.1. Clasificación botánica y descripción del tomate: El tomate (Lycopersicon esculentum Mill.) es miembro de la familia
Solanaceae, planta originaria de las planicies costeras occidentales de
Sudamérica, extendiéndose desde Ecuador hasta el norte de Chile (KINET y
PEET, 1997), concentrando la mayor diversidad en Perú.
El tomate es una planta perenne de porte arbustivo que se cultiva en forma
anual. Puede desarrollarse de forma rastrera, semi erecta o erecta y el
crecimiento es ilimitado en las variedades indeterminadas, pudiendo éstas
llegar a 10 m en un año (CHAMARRO, 1995). La ramificación es
generalmente simpodial, con lo que los ejes sucesivos se desarrollan a partir
de la yema axilar del eje precedente y la yema terminal da a lugar a la
inflorescencia (PICKEN, STEWART y KLAPWIJK, 1986).
Las hojas se disponen sobre los tallos alternadamente y son compuestas e
imparipinadas, constituidas generalmente por 7-9 foliolos lobulados o
dentados, de tamaño variable. Las hojas están recubiertas por pelos
glandulares y no glandulares que salen de la epidermis, los que le confieren
el olor característico (MAROTO, 1995). La epidermis del envés, o inferior,
contiene abundantes estomas que facilitan el intercambio gaseoso con el
exterior, mientras que éstos son escasos en la epidermis superior. Los
haces vasculares principales constan de un solo nervio primario del cual
parten en una estructura pinnada, irregular, en los nervios secundarios. Los

4
nervios primarios y secundarios presentan floema interno y externo (PICKEN,
STEWART y KLAPWIJK, 1986).
La iniciación de las hojas se produce a intervalos de 2-3 días, en función de
las condiciones ambientales. En general, la producción de hojas y primordios
foliares aumenta con la irradiación diaria y con la temperatura, siendo
constantes cuando las condiciones ambientales lo son. La velocidad de
iniciación de las hojas no se ve afectada por la irradiación diaria durante el
verano, pero sí resulta afectada en el invierno (CHAMARRO, 1995).
2.2. Nutrición con calcio:
2.2.1. Funciones fisiológicas del calcio
La mayoría de las funciones del calcio como componente estructural de
macromoléculas tienen relación con su capacidad de coordinación, mediante
la cual el calcio provee mecanismos de conexión intermoleculares estables,
pero reversibles, predominantemente en las paredes celulares y en la
membrana plasmática (MARSCHNER, 1995).
En los últimos años, el calcio ha generado mucho interés en la fisiología
vegetal y en la biología molecular, debido a su función como mensajero
secundario en la conducción de señales entre los factores medioambientales
y las respuestas de las plantas, en términos de crecimiento y de desarrollo
(FERGUSON y DROBAK, 1988).

5
2.2.1.1. Estabilización de la pared celular
El calcio es un componente integral de la pared celular y está involucrado en
la unión de las moléculas pécticas (FERGUSON y DROBAK, 1988). La unión
del calcio como un pectato en la lámina media es esencial para fortalecer las
paredes celulares y los tejidos de la planta. Esta función del calcio se refleja
claramente en la estrecha y positiva correlación entre la capacidad de
intercambio de cationes de las paredes celulares y el contenido de calcio en
los tejidos de la planta, requeridos para un óptimo crecimiento
(MARSCHNER, 1995).
La degradación de los pectatos es mediada por la poligalacturonasa, la cual
es inhibida drásticamente por las altas concentraciones de calcio. De
acuerdo con esto, en el tejido deficiente de calcio, la actividad de la
poligalacturonasa aumenta y un síntoma típico de la deficiencia de calcio es
la desintegración de las paredes celulares y el colapso de los tejidos
afectados, como los pecíolos y las partes superiores del tallo. En las hojas
de las plantas que reciben altos niveles de calcio durante el crecimiento o
que son cultivadas bajo condiciones de alta intensidad de luz, una gran
proporción del material péptico está presente en forma de pectato de calcio.
Esto hace al tejido altamente resistente a la degradación por la
poligaracturonasa. La proporción de pectato de calcio en las paredes
celulares también es importante, tanto para la susceptibilidad del tejido a las
infecciones fungosas y bacterianas, como para la maduración de la fruta
(MARSCHNER, 1995).

6
2.2.1.2. Estabilización de la membrana
La función fundamental del calcio en la estabilidad de la membrana y en la
integridad celular se refleja de varias formas. Esto se puede demostrar por el
aumento de la fuga de solutos de bajo peso molecular desde las células de
tejidos deficientes en calcio y, en las plantas con deficiencia severa de éste,
por una desintegración de las estructuras de la membrana y una pérdida de
la compartimentación de la célula (MARSCHNER, 1995).
El calcio estabiliza las membranas celulares al unir el grupo fosfato y el
carboxílico en fosfolípidos y proteínas, preferentemente en la superficie de la
membrana (MARSCHNER, 1995).
Por lo tanto, para cumplir su función en la membrana del plasma, el calcio
debe estar siempre presente en la solución externa, donde regula la
selectividad de absorción de iones y previene la pérdida desde el citoplasma
(MARSCHNER, 1995).
2.2.1.3. El calcio como mensajero secundario
La función del calcio como mensajero secundario puede influir sobre la
actividad enzimática de la célula y regular procesos tan importantes como la
elongación y división celular (POOVAIAH, 1988), además de los procesos
bioquímicos de la senescencia, en los cuales el calcio inhibe la acción de
enzimas pectolíticas (FERGUSON y DROBAK, 1988). En forma general, el
proceso comienza con la recepción del estímulo en la membrana y continúa

7
con la liberación de calcio de ciertos organelos, como la vacuola, para
aumentar la concentración citoplasmática, lo que permite su unión a una
proteína conocida como calmodulina. Esto induce un gran cambio
conformacional que posibilita al calcio para funcionar como un activador de
numerosas enzimas, liderando un retraso o una aceleración de respuestas
fisiológicas (POOVAIAH, 1988).
2.2.2. Absorción del calcio
El calcio es absorbido como ion divalente Ca++ (AZCÓN y TALÓN, 2000) a
través del sistema radicular por el flujo generado por la corriente
transpiratoria, o por difusión desde una zona de mayor concentración a una
zona de menor concentración de iones, siendo la primera responsable del
grueso de la absorción. Estos procesos ocurren principalmente en los pelos
radicales y en los ápices de raíces jóvenes, aún no suberizadas (SILVA,
1991).
La absorción de calcio es esencialmente proporcional a la absorción de agua
y ésta es afectada por la transpiración de la canopia, por la luz, por la
humedad y por la temperatura de las raíces (HO, 2000).
Las plantas pueden absorber todos los elementos nutritivos vía foliar. Sin
embargo, en la práctica, la fertilización foliar no se utiliza para aportar todos
los nutrientes, porque las absorciones son relativamente pequeñas y para
satisfacer los requerimientos de los macronutrientes se deberían efectuar
numerosas aplicaciones, las cuales serían económicamente imposibles de
realizar (TRINIDAD y AGUILAR, 1999).

8
Las plantas absorben los nutrientes en forma foliar, por tres caminos
posibles: a través de los estomas, de los ectodesmos y de la cutícula
(TRINIDAD y AGUILAR, 1999). Los estomas se pueden describir como
aberturas que se encuentran en las hojas, a través de las cuales se produce
el intercambio de oxígeno y CO2, en los procesos de respiración y
transpiración (SALISBURY y ROSS, 1994). La distribución de los estomas,
así como el tamaño y forma de éstos, varía ampliamente de una especie a
otra (SALISBURY y ROSS, 1994). Pero en general, existen tres a cuatro
veces más estomas en la cara inferior de las hojas en comparación con los
existentes en la cara superior (FLORES, 1999). Los estomas se encuentran
generalmente cerrados en la noche, durante periodos calurosos del día, ante
la presencia de viento excesivo y en un ambiente con bajo concentración de
CO2, entre otros factores (SALISBURY y ROSS, 1994).
Para un máximo ingreso por los estomas, las aplicaciones foliares deben ser
realizadas cuando los estomas se encuentran abiertos. Debido a que los
estomas se encuentran cerrados en la noche y durante el mediodía, es
recomendable realizar las aplicaciones foliares durante las primeras horas de
la mañana; asimismo, existe menos evaporación durante la mañana,
lográndose así una mejor oportunidad para una máxima absorción por parte
de las hojas. Una alta humedad relativa durante el tiempo de aplicación
favorecerá también una mayor absorción al minimizarse la evaporación
(TRINIDAD y AGUILAR, 1999).
Los ectodesmos son espacios submicroscópicos lineares en los cuales la
estructura fibrilar es laxa y bastante abierta. Éstos presentan una forma de
cavernas que se encuentran en la pared externa de la epidermis celular y en
algunas partes pueden alcanzar la superficie de la cutícula. Los ectodesmos

9
se pueden hallar en abundancia en las paredes de las células de guarda, en
las cámaras subestomáticas, en líneas sobre las paredes periclinares y
algunos, pueden estar distribuidos al azar en otros puntos de la epidermis.
Los ectodesmos se han asociado con la absorción foliar y la transpiración
periestomática (FLORES, 1999).
La cutícula cubre todos los tejidos aéreos, excepto la peridermis y las
heridas. Está compuesta por cutina, microfibrillas de polisacáridos y ceras
(FLORES, 1999). La absorción a través de la cutícula se produce porque
ésta al absorber agua, se dilata, produciéndose espacios vacíos, los cuales
permiten la difusión de las moléculas. Dado que las hojas jóvenes no tienen
una capa cuticular suficientemente desarrollada, las aplicaciones foliares de
nutrientes se verán favorecidas cuando la planta presente una mayor
proporción de follaje joven, donde se producirá una mayor absorción del
nutriente (GIL, 1995; TRINIDAD y AGUILAR; 1999).
El proceso de absorción de nutrientes por vía foliar tiene lugar en tres etapas.
En la primera, las sustancias nutritivas aplicadas a la superficie penetran la
cutícula y la pared celular por difusión libre o bien, vía intercambio iónico a
través de los ectodesmos. En una segunda etapa las sustancias son
absorbidas por la superficie de la membrana plasmática a través de los
fosfolípidos hidrofílicos. Finalmente, en la tercera, pasan al citoplasma
mediante la ocurrencia de un proceso metabólico (TRINIDAD y AGUILAR,
1999).
La velocidad de absorción foliar de los diferentes nutrientes no es igual en los iones monovalentes y en los de diámetro más pequeño como el potasio. Los
elementos secundarios y los micronutrientes se absorben en períodos que

10
van desde unas horas hasta un día. El fósforo es el único nutriente cuya
velocidad de absorción es más lenta. Este concepto es importante, porque
quiere decir que si llueve algunas horas después de la fertilización foliar, la
cantidad de nutrientes que puede lavarse es mínima (TRINIDAD y AGUILAR,
1999).
Además del tipo de nutriente, la velocidad de absorción depende de la
especie cultivada, del ion acompañante, de las condiciones ambientales
(temperatura, humedad relativa y lluvias), de la edad de la planta, de la hoja y
de las condiciones tecnológicas de la aspersión (TRINIDAD y AGUILAR,
1999).
2.3. Transporte de calcio en el interior de la planta: Una vez que ha ocurrido la absorción, las sustancias nutritivas se mueven
dentro de la planta, utilizando las siguientes vías: la corriente de transpiración
vía xilema, las paredes celulares y los espacios intercelulares (GIL, 1995).
Es bien conocido que el calcio se desplaza unidireccionalmente a larga
distancia vía xilema, y que es prácticamente inmóvil en el floema, por lo que
los factores que afectan al ritmo de transpiración afectan de forma notable la
disponibilidad de calcio en los tejidos distales de la raíz y de los órganos en
crecimiento no transpirantes (GUZMÁN, 2000).
Muy poco calcio es traslocado fuera de las hojas una vez que ha sido
asimilado por ellas. De esta forma los tejidos que están activos son

11
dependientes del transporte de calcio con el agua a través del xilema
(ADAMS, 1986).
Bajo condiciones de baja humedad, gran parte del agua es transportada
hacia las hojas viejas para mantener la tasa de transpiración. Como
resultado, una baja cantidad de calcio llega a la célula en crecimiento activo,
afectando su crecimiento. Sin embargo, durante la noche la humedad
aumenta y la tasa de transpiración decrece notablemente, permitiendo que el
agua y el calcio se muevan en dirección a los tejidos de baja transpiración.
Se sabe que el calcio es movilizado por la presión de la savia que se
desarrolla durante la noche debido a la actividad de la raíz (ADAMS, 1986).
2.4. Factores que afectan la absorción de calcio:
2.4.1 Déficit de presión de vapor
La humedad, expresada como déficit de presión de vapor se puede usar
como control de la transpiración y absorción de agua de un cultivo en
sistemas de producción en invernadero. Sin embargo, si se genera una
reducción de la transpiración en un ambiente de humedad alta (0,1 Kpa dvp),
puede reducirse la absorción y la distribución de cationes, tales como el Ca y
el K en cultivos de invernadero (ADAMS, 1991).
Ya que el movimiento del calcio se restringe casi exclusivamente al xilema,
se podría esperar que la reducción del flujo de la transpiración disminuyera la
acumulación de calcio (CHOI, CHUNG y SUH, 1997; HOLDER y
COCKSHULL, 1990). Se ha descrito que al exponer una planta a un

12
ambiente con humedad alta, se podría provocar una deficiencia leve de
calcio (HOLDER y COCKSHULL, 1990), pero MULHOLLAND et al. (2001 y
2000) y KREIJ (1996) no pudieron demostrar lo anterior, ya que no hubo una
disminución de calcio en las hojas de las plantas de tomate.
La humedad ambiental afecta el desarrollo foliar. Si la hoja es expuesta en
forma prolongada a una humedad alta, se reduce en forma irreversible su
tamaño y rendimiento; además, la humedad alta prolongada reduce
indirectamente y, en menor grado, la calidad de la fruta (MULHOLLAND et
al., 2000; ADAMS y HOLDER, 1992; HOLDER y COCKSHULL, 1990). La
deficiencia de calcio puede tener relación con la restricción de la expansión
de la hoja del tomate cultivado en un ambiente con humedad alta (HOLDER y
COCKSHULL, 1990; BAKKER, 1990).
Una alta humedad favorece el transporte de calcio hacia el interior del fruto
del tomate (ADAMS y HOLDER, 1992; BANUELOS, OFERMANN y SEIM,
1985).
2.4.2. Temperatura de la zona radicular
La temperatura de la zona radicular puede afectar la absorción y la
acumulación de nutrientes (HOOD y MILLS, 1994). La temperatura de la
zona radicular óptima para el crecimiento de la planta tiene relación con la
absorción más alta de agua y de nutrientes y con la acumulación de materia
seca en el tallo (HOOD y MILLS, 1994). Sin embargo, en un cultivo de
tomate de temporada larga, la temperatura de la zona radicular óptima para
el crecimiento, rendimiento y absorción de nutrientes puede aumentar con la

13
carga de fruta y con la temperatura del aire (BAKKER y SONNEVELD, 1988;
HURD y GRAVES, 1985).
La temperatura de la solución rizosférica influye de manera importante en la
absorción de agua y de elementos minerales, así como en el desarrollo y
crecimiento del sistema radical. La temperatura óptima de la rizósfera para
favorecer la absorción debe alcanzar cerca de 25º C. Una disminución en la
temperatura de la rizósfera produciría una disminución, tanto en el
crecimiento como en la absorción de agua, nitratos y fosfatos. Cuando la
temperatura radical se incrementa de 14 a 26 ºC (HO y ADAMS, 1989, citado
por GUZMÁN, 2000; NKANSAH y ITO, 1995) se produce un incremento en la
absorción de agua y calcio en plantas de tomate de hasta un 30%. Otros
autores han descrito fuertes disminuciones en la absorción de N, P, K, y Ca,
para temperaturas radicales inferiores a los 13ºC (GUZMÁN, 2000).
2.4.3. Incidencia de luz
La cantidad de radiación interceptada por la superficie foliar se considera el
factor principal que determina el rendimiento y la producción de biomasa, y
está estrechamente relacionada con la intercepción de la radiación
acumulada (GALLAGHER y BISCOE, 1978). Un análisis detallado de la
intercepción de radiación y producción de biomasa debería entregar una
mejor comprensión de los mecanismos de distribución de carbono dentro de
los brotes en condiciones de humedad alta (MULHOLLAND et al., 2001).
Cuando la disponibilidad de agua es abundante y la radiación incidente sobre
las hojas favorece una mayor actividad fotosintética, la demanda de CO2

14
dentro de la hoja es alta y los poros estomáticos están completamente
abiertos, disminuyendo la resistencia de la cámara estomática a la difusión
de CO2. Estas condiciones aumentan la transpiración (TAIZ y ZEIGER,
1998), incrementando la absorción de agua y calcio y la distribución de éste
hacia las partes transpirantes de la planta (ADAMS, 1991). Sin embargo,
cuando la intensidad lumínica disminuye conduce a un daño en la hoja de
tomate, asociado, probablemente, a una deficiencia de calcio (AIKMAN y
HOUTER, 1990).
2.4.4. Salinidad
Una alta salinidad reduce la absorción de agua en la planta y en el fruto
(CARVAJAL, MARTÍNEZ y CERDA, 1999; EHRET y HO, 1986), debido a que
un efecto de la salinidad es la inhibición del desarrollo de los vasos
xilemáticos, lo que aumenta así la resistencia al movimiento de agua en el
xilema y, por lo tanto, disminuye el contenido de calcio en las plantas,
particularmente en el fruto y en los tejidos jóvenes (ADAMS y HO, 1989).
2.5. Extracción de calcio en cultivo de tomate: La extracción de calcio en tomate indeterminado varía entre 255 y 390 kg ha-
1, dependiendo del rendimiento obtenido. La mayor extracción de calcio
coincide con el periodo de máximo crecimiento vegetativo (RINCON et al.,
1991). La extracción de calcio presenta cinco etapas. La primera etapa
comienza a partir del primer día de transplante hasta el día 25, donde la
planta se observa en el estado de primer racimo floral. En esta etapa, la

15
absorción de calcio es lenta, acumulándose principalmente en las hojas. La
segunda etapa comienza a partir del día 25 hasta el día 72, donde la planta
muestra ya su quinto racimo en el inicio de floración. Se produce un alza en
la absorción de calcio, manteniéndose la acumulación en las hojas. La
tercera etapa se inicia el día 72 hasta el 87, donde la planta ya muestra el
inicio de cuaja del quinto racimo y el sexto racimo en flor. Aquí se presenta
una disminución de la absorción de calcio, producto de los deshojes
realizados. La cuarta etapa comienza a partir del día 87 hasta el día 102,
donde la planta se encuentra con el quinto y sexto racimo cuajado y con el
séptimo racimo en flor. Se observa un leve aumento de la absorción de
calcio. Por último, en la quinta etapa, que parte el día 102 hasta el fin del
cultivo, disminuye la absorción de calcio en forma gradual (VALENCIA,
1996).
2.6. Deficiencia localizada de calcio en la hoja: Los síntomas de deficiencia de calcio son siempre más evidentes en tejidos
jóvenes y zonas meristemáticas de las raíces, tallos y hojas (AZCÓN y
TALÓN, 2000). Las hojas nuevas de plantas con deficiencia de calcio
presentan clorosis intervenal y necrosis marginal. En los márgenes de las
hojas nuevas en rápido crecimiento (de la 3a a la 5a hoja bajo el punto de
crecimiento), esta deficiencia se manifiesta por una clorosis y, si la humedad
persiste esta clorosis (coloración amarilla), pasa a ser café, y una vez que el
tejido está muerto puede ser colonizado rápidamente por Botrytis cinerea
(ADAMS, 1986).

16
Una forma transitoria de deficiencia de calcio es inducida por períodos largos
de alta humedad, lo que hace bajar la tasa de transpiración, y de esa forma
el calcio disminuye en las hojas y genera la deficiencia (ADAMS, 1986).
2.7. Deficiencia localizada de calcio en el fruto: El transporte del calcio desde las raíces al fruto en tomate es intrínsecamente
bajo, debido a que el calcio se mueve con la corriente transpiratoria, y el fruto
tiene tasas muy bajas de transpiración. En consecuencia, la concentración
de calcio en el fruto es menor que en cualquier otro órgano de la parte aérea.
La concentración de calcio del fruto puede ser reducida aún más si la
transpiración de la canopia es alta, porque entonces, más calcio es dirigido
lejos del fruto hacia las hojas en transpiración. Además, la distribución de
calcio en el fruto no es uniforme. El sistema xilemático, que lleva el flujo
transpiratorio, está muy escasamente desarrollado hacia el extremo distal del
fruto, y así muy poco flujo transpiratorio alcanza esta región. En
consecuencia, la concentración de calcio en el tejido distal es menor en el
fruto. El sistema xilemático en la parte distal del fruto está especialmente
poco desarrollado en los cultivares susceptibles, y su desarrollo se reduce
aún más si la solución nutritiva es muy salina. Por lo tanto, una alta salinidad
no sólo reduce la absorción de calcio de las raíces, sino que también reduce
el transporte de calcio hacia el tejido distal. Además, la concentración de
calcio en el fruto desciende sustancialmente durante el período de rápida
expansión (cerca de dos semanas después de la antesis). En consecuencia,
la concentración de calcio del tejido distal del fruto puede volverse
críticamente baja durante este período. Este es el momento en que se
induce el blossom-end rot. Cuando la concentración de calcio en el tejido

17
distal se reduce bajo un nivel crítico para la permeabilidad de la membrana
celular, se desarrolla blossom-end rot (HO, 2000).
2.8. Botrytis (Botrytis cinerea): Esta enfermedad puede atacar en distintas etapas del desarrollo de las
plantas de numerosas especies hortícolas. Su ataque puede ocurrir
preferentemente en los meses de invierno y primavera. Causa pérdidas
cuantiosas en la producción de tomates en invernadero (APABLAZA, 2000).
El agente causal de la Botrytis cinerea Pers. es un hongo que presenta una
fase saprofítica, permaneciendo en restos de cosecha en el suelo. Puede
formar esclerocios cuando el sustrato se ha empobrecido o cuando las
condiciones son desfavorables (APABLAZA, 2000).
En tomate bajo invernadero los daños más frecuentes son la necrosis de los
foliolos, los que cuelgan y se secan; el aborto floral; la necrosis en las
láminas de las hojas; la necrosis de hojas completas; la muerte de brotes y la
pudrición de frutos en cualquier estado de desarrollo. Un daño importante es
la formación de cancros en los tallos, de coloración café claro, de tamaño
variable que puede llegar a circuncidar todo el tallo y causar con ello
marchitez y muerte de la planta completa (APABLAZA, 2000).
En la producción de tomates de invernadero ésta es la enfermedad más
dañina. Su diseminación es muy rápida y eficaz, ya que el hongo aprovecha

18
todo tipo de heridas causadas por la manipulación de las plantas y por la
cosecha escalonada de frutos, para penetrar y establecerse (APABLAZA,
2000).
2.8.1. Condiciones predisponentes
Al analizar en forma global esta enfermedad se encuentra que los factores
temperatura, humedad relativa y luz inciden en forma gravitante sobre el
desarrollo de esta enfermedad (BESOAIN, 2000).
Las temperaturas cardinales en las que aparece son de 35,5 ºC máxima,
22,7 ºC óptima y de 1,6 ºC como mínima (APABLAZA, 2000). Considerando
estos antecedentes se puede inferir que esta enfermedad afectará cultivos
bajo invernadero entre los meses de otoño y primavera (BESOAIN, 2000).
El segundo factor importante de considerar es la humedad relativa,
específicamente humedades superiores al 90%, que provocan que en
algunos tejidos de las plantas exista saturación o agua libre. Este concepto
se asocia con la necesidad que posee el patógeno de germinar e ingresar
internamente a los tejidos, antes que la superficie de éstos se seque
nuevamente. Un aspecto que se encuentra ligado a la humedad relativa es,
sin duda, la temperatura. En invernaderos con una temperatura y humedad
relativa alta durante el día, se produce una condensación o agua libre sobre
los vegetales. Al descender la temperatura en la noche, y si además la
temperatura no baja a los 12 ºC, se produce la condición ideal para que

19
ocurra un ataque de pudrición gris, cuando no es factible realizar una
adecuada ventilación para impedir este efecto (BESOAIN, 2000).
Un aspecto no siempre ponderado es el efecto de la luz sobre la pudrición
gris. Este tópico debe ser analizado desde el punto de vista de la planta y del
patógeno. Al desarrollarse un cultivo bajo una condición de baja luminosidad,
la planta y los tejidos de ésta serán más vulnerables, y sobre todo, las
barreras físicas que presentan a la penetración serán delgadas y con un
menor contenido de ceras. Esto le permitirá a Botrytis cinerea una
penetración más rápida y fácil (BESOAIN, 2000).
2.8.2. Control cultural
En un invernadero frío tradicional, es la ventilación la que juega un rol
importante en la disminución y acotación del periodo de agua libre sobre los
tejidos (BESOAIN, 2000), Como alternativas de control se recomienda que
en los cultivos en invernaderos, éstos se aireen diariamente para eliminar el
exceso de humedad, siempre que haya más de 6º C en el ambiente
(APABLAZA, 2000).
Una fertilización nitrogenada excesiva, así como bajos niveles de calcio,
favorecen el desarrollo de esta enfermedad (APABLAZA, 2000), por lo tanto,
un aumento de la concentración de calcio en la solución de fertilización se
puede emplear para prevenir la aparición de esta enfermedad (CANOVAS,
1995).

20
Una práctica habitual y muy importante es la eliminación de todos los tejidos
muertos de hojas, frutos o restos florales, a modo de evitar que este
patógeno pueda esporular. Al no existir inóculo o esporas, aunque existan
condiciones favorables, no se va a desarrollar esta enfermedad (BESOAIN,
2000).

21
3. MATERIALES Y MÉTODO
3.1. Invernadero: Se utilizó una estructura de dos aguas, con lucarna, de orientación norte –
sur, elegido al azar dentro del módulo al cual pertenecía. Sus dimensiones
fueron de 7,2 m de ancho y 30 m de largo. La altura cenital de 4 m en la
parte más alta y 2,2 m en la más baja de la canaleta, con una relación de 3,1
m3 . m -2.
3.2. Material vegetal: La evaluación se efectuó en la variedad Fortaleza, siendo ésta la más
utilizada en los valles de Quillota – Limache, en cultivo bajo invernadero, con
la modalidad primor-temprano y establecidos en mayo.
3.3. Ensayo 1: Evaluación de aplicaciones foliares de óxido de calcio sobre la incidencia de desorden fisiológico de clorosis foliar en Quillota:
Este ensayo se realizó en el Fundo La Palma, perteneciente a la empresa
Inversiones Quintil S.A., ubicado en La Palma, comuna de Quillota, provincia
de Quillota, V región de Valparaíso, desde el 8 de mayo hasta el 18 de
diciembre del 2001. La plantación correspondió a la modalidad primor
temprano. El transplante se realizó el 8 de mayo. La densidad de plantación

22
que se utilizó fue de 27.500 plantas por hectárea, plantadas en una hilera a
20 cm, la cuales fueron amarradas en forma alternada permitiendo un
espacio final de 40 cm entre plantas y de 1.8 m entre hileras. Los manejos
agronómicos que se llevaron a cabo fueron los habituales del cultivo. Dentro
de estos manejos se realizó un control químico de Botrytis cinerea el día 25
de julio.
Se evaluó el efecto del óxido de calcio (Basfoliar Calcio SL, que posee un
17% de Ca) en dos dosis, asperjando la planta en el tercio superior (segunda
hoja bajo el ápice de crecimiento), con un mojamiento de 400 l. ha-1. Las
aplicaciones se efectuaron con un equipo de pulverización manual de
espalda, utilizando 40 lb .pulg2 de presión máxima y empleando boquillas de
cono hueco, con un gasto de 0.45 l. min-1.
3.3.1. Tratamientos
El ensayo comparó el efecto de dos dosis de óxido de calcio foliar y de la
aplicación de sólo agua, frente a un testigo absoluto (sin aplicación), o
tratamiento control. Los tratamientos se describen de la siguiente manera:
T1: Oxido de calcio 680 g .ha -1 (i.a.); 4 l. ha-1(p.c.)
T2: Oxido de calcio 1020 g ha -1(i.a.); 6 l. ha-1(p.c.) T3: Agua con un contenido de 10.4 mg .l de calcio
T4: Testigo
El ensayo comparó el efecto de tres aplicaciones foliares de óxido de calcio
con un intervalo de 15 días entre aplicación, comenzando el 30 de julio

23
durante la aparición del quinto racimo y terminando el 29 de agosto con la
aparición del sexto racimo.
3.3.2. Mediciones
Para la realización de las mediciones de todos los tratamientos se consideró
desde la primera hoja por sobre el quinto racimo hasta la primera hoja por
sobre el sexto racimo, que corresponde a la época donde disminuye la
absorción de calcio bajo estas condiciones de cultivo (VALENCIA, 1996).
3.3.2.1. Síntomas foliares del desorden fisiológico
Cada siete días se evaluó el número de foliolos con sintomatología de
clorosis en cada tratamiento después de las aplicaciones de calcio (Figura 1),
cuantificando el número de folíolos con la sintomatología ya descrita por hoja,
a partir de la primera hoja por sobre el quinto racimo hasta la primera hoja
por sobre el sexto racimo. Una vez terminada la evaluación se determinó el
porcentaje de incidencia del desorden fisiológico, utilizando la siguiente
fórmula:
I = (Número de foliolos afectados). 100 (Total de folíolos de la hoja)

24
FIGURA 1: Clorosis y necrosis marginal de folíolos de hoja de tomate.

25
Para determinar la intensidad del síntoma, se calculó el área foliar afectada
por foliolo, para lo cual se sacaron fotos digitales de las hojas de dos plantas
por repetición, una vez terminada la cuantificación de la sintomatología. Las
imágenes digitales se analizaron con el software Scion Image (Maryland,
Scion Corporación, www.scioncorp.com). El análisis de las imágenes
consistió en tomar la imagen desde el archivo en formato BMP o TIFF, luego
desde la barra de herramientas se pulsó “Options” y luego “Density slice”. La
imagen tomó una coloración roja en algunas partes, luego en la barra de
herramientas “LUT” se desplazó el limite inferior de la línea roja hasta el
extremo inferior, después se movió al extremo superior hasta que quedó de
color rojo sólo el área que se quería calcular; posteriormente, se pulsó
“Analyze” y luego “Set Scale”, donde apareció una ventana donde se definió
la unidad de medida y se colocó una medida conocida de la imagen. Se
pulsó “Analyze” nuevamente y luego “Measure”, y por último se pulsó
nuevamente “Analize”. Luego la opción “Show Results”, donde apareció una
ventana “Results” que contenía el área determinada en la unidad de medida
definida.
3.3.2.2. Incidencia de Botrytis cinerea
El porcentaje de incidencia de Botrytis cinerea por tratamiento fue medida en
base AGRIOS (1996), que señala la incidencia de una enfermedad que
corresponde a la proporción de plantas, hojas, tallos y frutos que muestren
cualquier tipo de síntoma. Para cuantificar la incidencia se contó el número
de folíolos que presentó la enfermedad entre la primera hoja por sobre el
quinto racimo hasta la primera hoja del sexto racimo. Una vez terminada la
evaluación se determinó el porcentaje de incidencia de Botrytis cinerea,
utilizando la siguiente fórmula:

26
I = (Número de foliolos afectados). 100 (Total de folíolos de la hoja)
3.3.2.3. Contenido foliar de calcio
Como la sintomatología de clorosis ocurre en los foliolos del tercio distal de
las hojas de tomate que han completado su desarrollo, se tomó dos muestras
de 20 gramos de cada foliolo. Se obtuvieron con un sacabocado de un
centímetro de diámetro, desde el tercio distal de los folíolos y desde el tercio
proximal de los foliolos que habían completado su desarrollo. Las muestras
se analizaron en el Laboratorio de Suelos y Análisis Foliar de la Universidad
Católica de Valparaíso y se determinó el porcentaje de calcio a través del método de calcinación, utilizando un espectrofotométrico de absorción
atómica (KALRA, 1998).
3.3.2.4. Parámetros agronómicos
Para determinar los parámetros agronómicos, se cosecharon los racimos
quinto y sexto de las plantas y se registró el peso de los frutos que poseían
un diámetro mínimo de 5 cm. El índice de cosecha utilizado fue el 40% de
coloración de los frutos; esto en base a lo expuesto por CHAMARRO (1995).
Con respecto a la distribución de calidades, la selección se realizó en tres
categorías: primera, con diámetro de 7,0 a 8,0 cm; segunda, con un
diámetro de 6,0 a 7,0 cm; y tercera con un diámetro de 5,0 a 6,0 cm,

27
excluyendo los frutos que tuvieran problemas de formación, fisiológicos o
sanitarios.
3.4. Ensayo 2: Evaluación de aplicaciones foliares de óxido de calcio sobre la incidencia de desorden fisiológico de clorosis foliar en Olmué:
Este ensayo se realizó en el Fundo El Arrayán, ubicado en la comuna de
Olmué, provincia de Quillota, V región de Valparaíso, desde el 15 de mayo
hasta el 14 de diciembre del 2002. La plantación correspondió a la
modalidad primor temprano. La plantación se realizó el 15 de mayo. La
densidad de plantación que se utilizó fue de 27.500 plantas por hectárea
plantadas en una hilera a 20 cm, la cuales fueron amarradas en forma
alternada permitiendo un espacio final de 40 cm entre plantas y de 1.8 m
entre hileras y los manejos agronómicos que se llevaron a cabo fueron los
habituales del cultivo. Dentro de estos manejos se realizo un control de
Botrytis cinerea el día 15 de agosto.
Este ensayo se desarrolló bajo las mismas condiciones que el Ensayo 1 en la
localidad de Quillota el año 2001, utilizando la misma metodología para las
mediciones. Sin embargo, el tratamiento agua presentó un cambio en la
concentración de calcio a 26.0 mg . l de calcio y las fechas de aplicación del
óxido de calcio foliar comenzaron el 3 de agosto, durante la aparición del
quinto racimo y terminando el 4 de septiembre con la aparición del sexto
racimo.

28
3.5. Diseño estadístico: Se utilizó un diseño en bloques completamente al azar, con cuatro
repeticiones. La unidad experimental correspondió a una parcela de siete
plantas . m2, que fueron medidas en su totalidad.
El modelo estadístico correspondiente es:
Yij = mij + t + β +ε
Donde:
Yij : Valor Observado en cada unidad experimental
mij : Efecto de la Media General sobre cada observación
t : Efecto del Tratamiento sobre cada observación
β : Efecto de Bloques sobre cada observación
ε : Efecto del Error Experimental Aleatorio sobre cada observación
3.6. Análisis estadístico de los datos:
3.6.1. Análisis de varianza y comparación de medias.
Se verificó si los tratamientos tuvieron un efecto utilizando el análisis de
Fischer. En aquellos casos en que el análisis de varianza indicó que al
menos un tratamiento fue distinto de los otros en cuanto a los resultados (P ≤
0.05), se realizó una comparación de medias de acuerdo al test de Tukey,

29
comparando todos los tratamientos entre sí, con una probabilidad de error de
5%. El análisis se realizó con el software Minitab (Pennsylvania, Minitab Inc.,
www.minitab.com).
3.6.2. Modelo de regresión lineal múltiple
Se realizó la regresión lineal múltiple utilizando el método de los Mejores
Subconjuntos utilizando el siguiente modelo estadístico:
Yi : b0 + b1X1 + bnXn + ε Yi : Variable dependiente b0: Punto de intersección entre la recta y el eje Y b1X1: Variable independiente o predictor1 :
bnXn: variable independiente o predictor n ε : Efecto del Error Experimental Aleatorio sobre cada observación
Se construyeron tres modelos de predicción, para los cuales se detallan a
continuación sus respectivas variables dependientes e independientes o
predictores.
Y1: Sintomatología de clorosis marginal de los foliolos
X1: Concentración de calcio del tejido foliar de la zona proximal del
foliolo.
X2: Concentración de calcio del tejido foliar de la zona distal del foliolo.
Y2: Incidencia de Botrytis cinerea

30
X1: Concentración de calcio del tejido foliar de la zona proximal del
foliolo.
X2: Concentración de calcio del tejido foliar de la zona distal del foliolo.
X3: Porcentaje de incidencia de la sintomatología de clorosis
X4: Porcentaje de área afecta por la sintomatología, obtenido mediante
análisis de imagen.
Y3: Peso de los racimos X1: Concentración de calcio del tejido proximal de los foliolos de las
hojas de tomate.
X2: Concentración de calcio del tejido distal de los foliolos de las hojas
de tomate.
El análisis se realizó con el software Minitab (Pennsylvania, Minitab Inc.,
www.minitab.com).
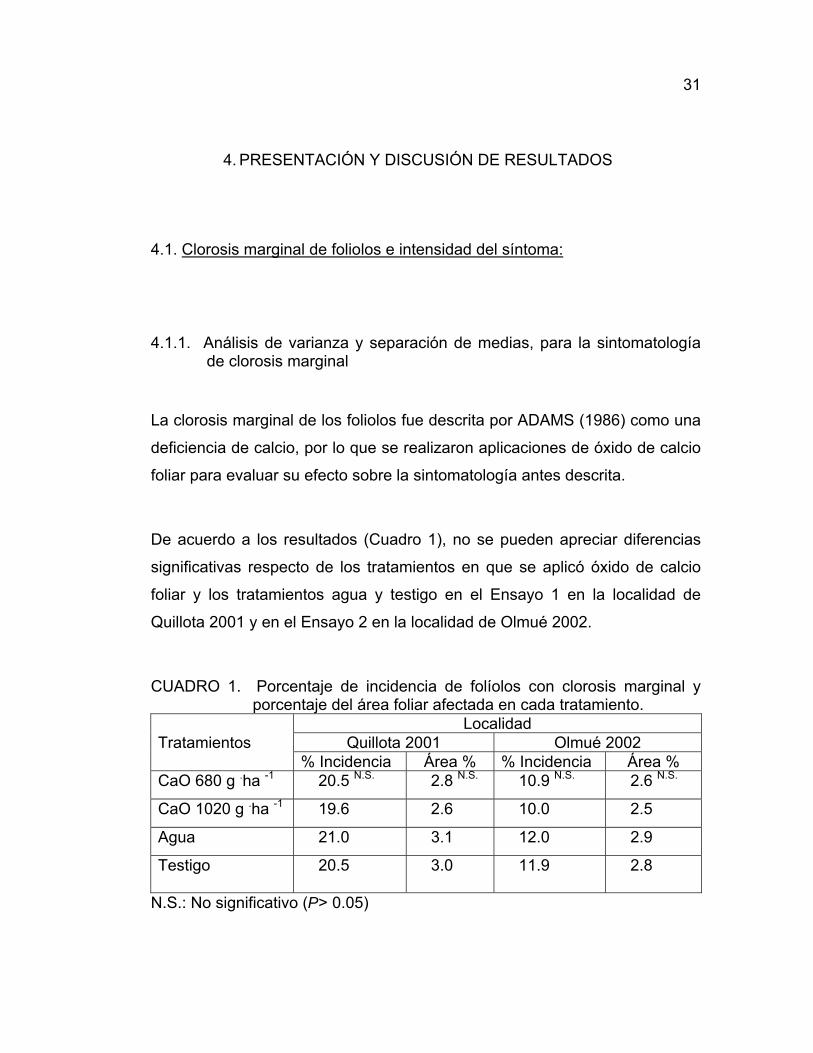
31
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Clorosis marginal de foliolos e intensidad del síntoma:
4.1.1. Análisis de varianza y separación de medias, para la sintomatología de clorosis marginal
La clorosis marginal de los foliolos fue descrita por ADAMS (1986) como una
deficiencia de calcio, por lo que se realizaron aplicaciones de óxido de calcio
foliar para evaluar su efecto sobre la sintomatología antes descrita.
De acuerdo a los resultados (Cuadro 1), no se pueden apreciar diferencias
significativas respecto de los tratamientos en que se aplicó óxido de calcio
foliar y los tratamientos agua y testigo en el Ensayo 1 en la localidad de
Quillota 2001 y en el Ensayo 2 en la localidad de Olmué 2002.
CUADRO 1. Porcentaje de incidencia de folíolos con clorosis marginal y porcentaje del área foliar afectada en cada tratamiento.
Localidad Tratamientos Quillota 2001 Olmué 2002 % Incidencia Área % % Incidencia Área % CaO 680 g .ha -1 20.5 N.S. 2.8 N.S. 10.9 N.S. 2.6 N.S.
CaO 1020 g .ha -1 19.6 2.6 10.0 2.5
Agua 21.0 3.1 12.0 2.9
Testigo 20.5 3.0 11.9 2.8
N.S.: No significativo (P> 0.05)

32
Se puede considerar, de acuerdo a los resultados, que el síntoma antes
descrito es no sólo una deficiencia de calcio localizada, sino que también
está asociado a otro nutriente como fue descrito por (MULHOLLAND et al.,
2001 y 2000), ya que éstos no pudieron relacionar el síntoma con una
deficiencia de calcio, sino que este síntoma estaría asociado a una
deficiencia localizada de potasio y calcio en el foliolo terminal en la hoja de
tomate, pero no en los otros foliolos de la hoja (ADAMS, 1994;
ADAMS,1991); además HOLDER y COCKSHULL (1990) y ADAMS (1986) y
señalan que este síntoma está asociado a una deficiencia localizada de
calcio, aunque en su estudios no midieron los contenidos de potasio en la
hoja.
4.1.2. Regresión lineal múltiple sobre la sintomatología de clorosis marginal
A continuación se presentan los modelos de regresión lineal múltiple
generados con los datos de los Ensayos 1 y 2.
Sintomatología = 80.0 - 9.25 . Calcio proximal - 2.68 . Calcio distal (1)
Sintomatología = 125 - 3.68 . Calcio proximal - 18.1 . Calcio distal (2)
Sintomatología = 34.7 - 2.72 . Calcio proximal - 3.21 . Calcio distal (3)
Sintomatología = 27.7 - 2.24 . Calcio proximal - 5.22 . Calcio distal (4)
Los modelos presentados incluyen solamente los conjuntos mínimos de
predictores que permitió obtener un coeficiente de regresión estabilizado.
La relación entre sintomatología de clorosis marginal de los foliolos y
concentración de calcio del tejido foliar de la zona proximal del foliolo y la

33
concentración de calcio del tejido foliar de la zona distal del foliolo es de un
99% (1) con los datos del testigo en el Ensayo 1 en la localidad de Quillota
2001 y de un 96% (2) en el Ensayo 2 en la localidad de Olmué 2002, pero el
porcentaje de correlación disminuye cuando se utilizan los datos de todos los
tratamientos a un 20.5% (3) en el Ensayo 1 en la localidad de Quillota 2001 y
a un 17.3% (4) en el Ensayo 2 en la localidad de Olmué 2002. Ello podría
indicar que la forma de calcio empleada no es la más adecuada para
disminuir la sintomatología.
4.2. Contenido foliar de calcio: Los análisis foliares fueron utilizados para determinar si las aplicaciones
foliares de óxido de calcio foliar aumentaron la concentración de calcio en las
hojas de tomate.
De acuerdo a los resultados (Cuadro 2), se pudo apreciar que las
concentraciones de calcio en la base de los foliolos y el contenido de calcio
distal en el Ensayo 1 de la localidad de Quillota 2001, no presentaron
diferencias significativas entre los tratamientos, lo que concuerda (Cuadro 1)
con MULHOLLAND et al. (2001 y 2000) ya que no se logró asociar el
síntoma con una deficiencia de calcio, pero discrepa con los resultados de
HOLDER y COCKSHULL (1990) y ADAMS (1986), porque éstos asociaron
una deficiencia de calcio al síntoma antes descrito.
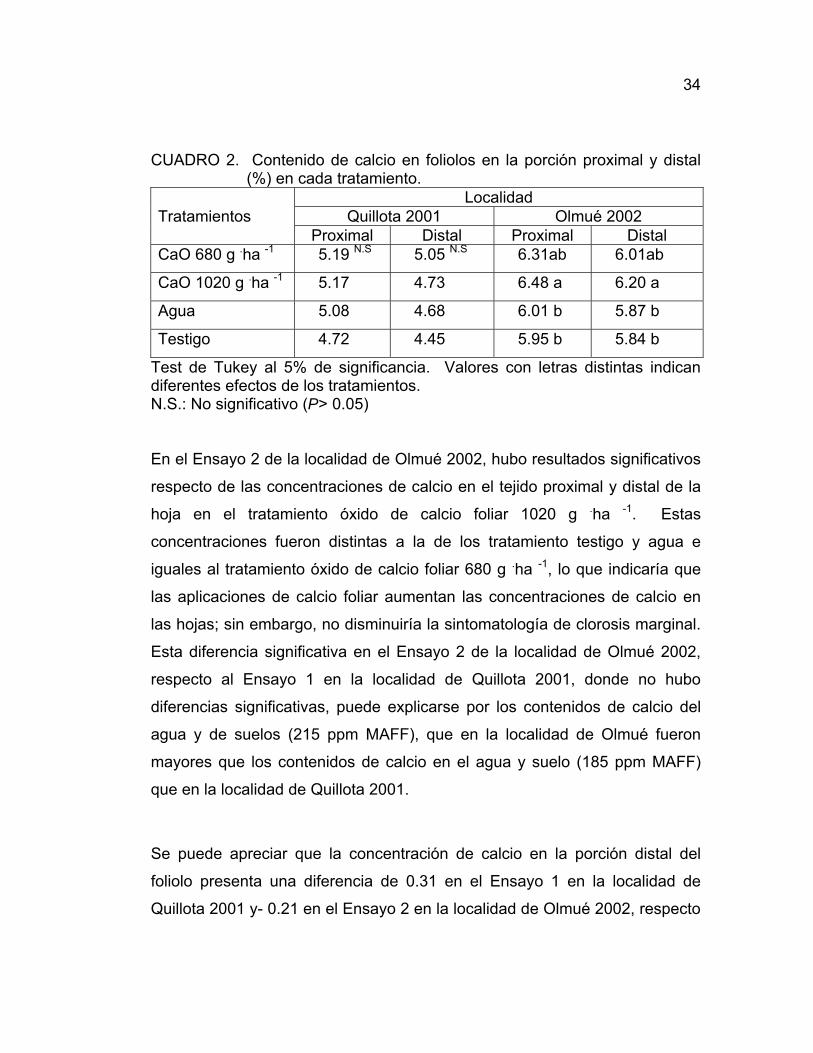
34
CUADRO 2. Contenido de calcio en foliolos en la porción proximal y distal (%) en cada tratamiento.
Localidad Tratamientos Quillota 2001 Olmué 2002 Proximal Distal Proximal Distal CaO 680 g .ha -1 5.19 N.S 5.05 N.S 6.31ab 6.01ab
CaO 1020 g .ha -1 5.17 4.73 6.48 a 6.20 a
Agua 5.08 4.68 6.01 b 5.87 b
Testigo 4.72 4.45 5.95 b 5.84 b
Test de Tukey al 5% de significancia. Valores con letras distintas indican diferentes efectos de los tratamientos. N.S.: No significativo (P> 0.05)
En el Ensayo 2 de la localidad de Olmué 2002, hubo resultados significativos
respecto de las concentraciones de calcio en el tejido proximal y distal de la
hoja en el tratamiento óxido de calcio foliar 1020 g .ha -1. Estas
concentraciones fueron distintas a la de los tratamiento testigo y agua e
iguales al tratamiento óxido de calcio foliar 680 g .ha -1, lo que indicaría que
las aplicaciones de calcio foliar aumentan las concentraciones de calcio en
las hojas; sin embargo, no disminuiría la sintomatología de clorosis marginal.
Esta diferencia significativa en el Ensayo 2 de la localidad de Olmué 2002,
respecto al Ensayo 1 en la localidad de Quillota 2001, donde no hubo
diferencias significativas, puede explicarse por los contenidos de calcio del
agua y de suelos (215 ppm MAFF), que en la localidad de Olmué fueron
mayores que los contenidos de calcio en el agua y suelo (185 ppm MAFF)
que en la localidad de Quillota 2001.
Se puede apreciar que la concentración de calcio en la porción distal del
foliolo presenta una diferencia de 0.31 en el Ensayo 1 en la localidad de
Quillota 2001 y- 0.21 en el Ensayo 2 en la localidad de Olmué 2002, respecto

35
a la concentración de calcio basal de los foliolos. Sin embargo, no se
encontró información sobre el contenido de calcio de los foliolos en la hoja de
tomate. WIEN (1997) señala que en lechuga la concentración de calcio en la
región proximal de la hoja presenta mayores concentraciones de calcio que
en la región distal de las hojas. Además, MORARD, BERNADAC y
BERTONI (1996) señalan que las concentraciones de calcio en los tejidos de
las plantas de tomate son distintas según la edad y la posición de éstas.
En los frutos de tomate la concentración de calcio se distribuye con una
mayor proporción en el tejido proximal que en el distal (HO, 2000; MINAMIDE
y HO, 1993 y BANUELOS, OFFERMAN y SEIM, 1985).
De acuerdo a los resultados de este estudio se podría establecer una
relación entre la distribución y los niveles de calcio en la hoja. Las
concentraciones de calcio son distintas en los tejidos proximales y en los
tejidos dístales en los foliolos de las hojas de tomate.
4.3. Porcentaje de incidencia de Botrytis cinerea:
4.3.1. Análisis de varianza y separación de medias sobre la incidencia de Botrytis cinerea
El porcentaje de incidencia se utilizó como sugiere AGRIOS (1996) para
determinar incidencia de Botrytis cinerea en las hojas de tomate y para
evaluar si las aplicaciones foliares de óxido de calcio disminuyen esta
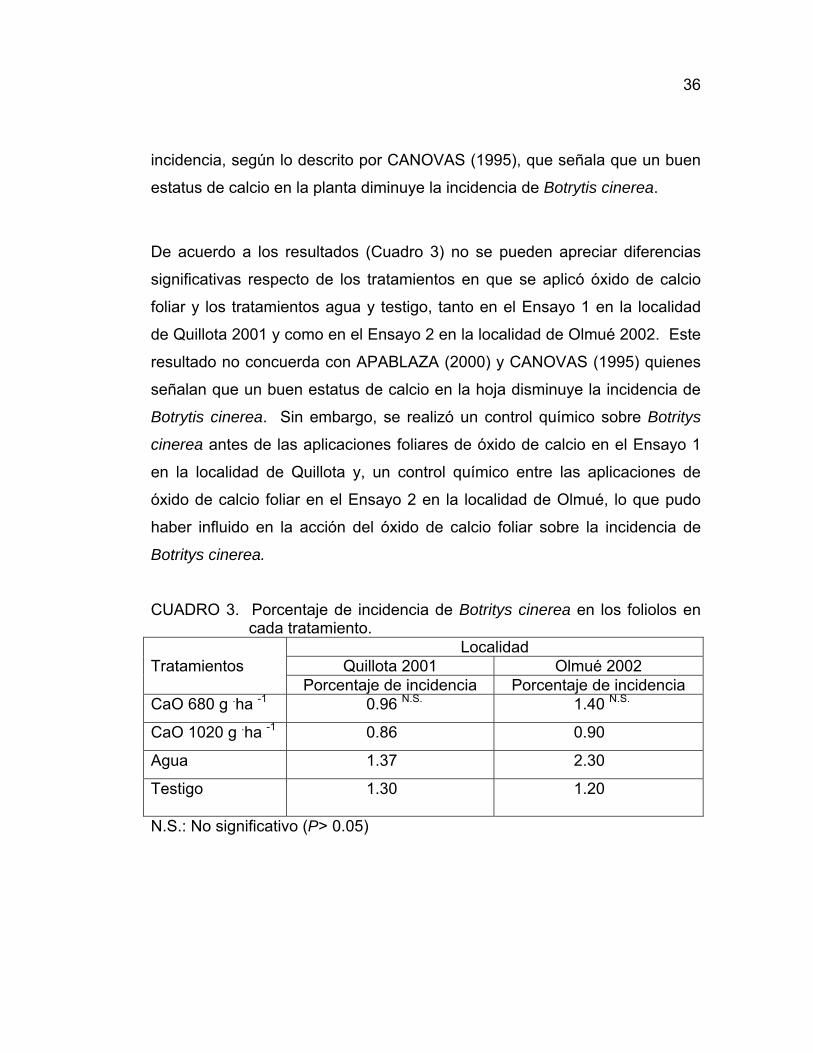
36
incidencia, según lo descrito por CANOVAS (1995), que señala que un buen
estatus de calcio en la planta diminuye la incidencia de Botrytis cinerea.
De acuerdo a los resultados (Cuadro 3) no se pueden apreciar diferencias
significativas respecto de los tratamientos en que se aplicó óxido de calcio
foliar y los tratamientos agua y testigo, tanto en el Ensayo 1 en la localidad
de Quillota 2001 y como en el Ensayo 2 en la localidad de Olmué 2002. Este
resultado no concuerda con APABLAZA (2000) y CANOVAS (1995) quienes
señalan que un buen estatus de calcio en la hoja disminuye la incidencia de
Botrytis cinerea. Sin embargo, se realizó un control químico sobre Botritys
cinerea antes de las aplicaciones foliares de óxido de calcio en el Ensayo 1
en la localidad de Quillota y, un control químico entre las aplicaciones de
óxido de calcio foliar en el Ensayo 2 en la localidad de Olmué, lo que pudo
haber influido en la acción del óxido de calcio foliar sobre la incidencia de
Botritys cinerea.
CUADRO 3. Porcentaje de incidencia de Botritys cinerea en los foliolos en cada tratamiento.
Localidad Tratamientos Quillota 2001 Olmué 2002 Porcentaje de incidencia Porcentaje de incidencia CaO 680 g .ha -1 0.96 N.S. 1.40 N.S.
CaO 1020 g .ha -1 0.86 0.90
Agua 1.37 2.30
Testigo 1.30 1.20
N.S.: No significativo (P> 0.05)

37
4.3.2. Regresión lineal múltiple sobre la incidencia de Botritys cinerea
A continuación se presentan los modelos de regresión lineal múltiple
generados con los datos de los Ensayos 1 y 2.
I. de Botritys cinerea = 2.67 – 2.68. Calcio distal + 4.21 % de área afectada(1)
I. de Botritys cinerea = 4.02 – 0.20. Calcio distal + 0.15 % de área afectada(2)
I. de Botritys cinerea = 2.51 – 3.20. Calcio distal + 4.53 % de área afectada(3)
I. de Botritys cinerea = 3.89 – 0.50. Calcio distal + 1.40 % de área afectada(4)
Los modelos presentados incluyen solamente los conjuntos mínimos de
predictores que permitió obtener un coeficiente de regresión estabilizado.
La relación entre incidencia de Botritys cinerea y la concentración de calcio
del tejido foliar de la zona distal del foliolo y el porcentaje de área afectada
con la sintomatología de clorosis marginal de foliolos es de un 92.2% (1) en
los datos del testigo Ensayo 1 en la localidad de Quillota 2001 y de un 97.9%
(2) en el Ensayo 2 en la localidad de Olmué 2002, lo que no concuerda con
los resultados obtenidos, pero la relación disminuye a un 6.2% (3) en el
Ensayo 1 en la localidad de Quillota 2001 y a un 32.9% (4) en el Ensayo 2 en
la localidad de Olmué 2002 al incluir los datos de todos los tratamientos, lo
que indicaría que la forma y/o concentración de calcio empleada no son las
más adecuadas, para la disminución de la incidencia de Botrytis cinerea, o
bien que el control químico efectuado fue eficaz en controlar esta
enfermedad.

38
4.4. Variables agronómicas:
4.4.1. Análisis de varianza y separación de medias sobre las variables agronómicas
HO (2000) señala que al aumentar los contenidos de calcio en las plantas
mejora la calidad y peso de los frutos, por lo que se evaluó si las aplicaciones
de óxido de calcio foliar presentaban relación con un aumento de la calidad y
peso de los frutos de las plantas de tomate.
El número de frutos promedio entre los dos racimos evaluados (5 y 6)
(Cuadro 4) no presentaron diferencias significativas en ninguno de los
tratamientos en el Ensayo 1 de la localidad de Quillota 2001 y en el Ensayo 2
en la localidad de Olmué 2002. Se puede establecer que las aplicaciones de
óxido de calcio foliar no afectan el número de frutos cosechados, sino que el
número de frutos cosechados depende de otros factores como la
temperatura, radiación, competencia con otros frutos y contenido endógeno
de hormonas (KINET y PEET, 1997).
Se pueden apreciar diferencias significativas respecto al peso de los racimos
en los tratamientos en los que se aplicó óxido de calcio foliar, en el Ensayo 1
de la localidad de Quillota 2001 y en el Ensayo 2 en la localidad de Olmué
2002, lo que concuerda con HO (2000) quien señala que un aumento de los
niveles de calcio en frutos de tomate en rápida expansión provoca un
aumento en la paredes y membranas celulares, mejorando la calidad y
disminuyendo los desórdenes fisiológicos asociados a deficiencia de calcio.
KOMOSA (1984) describe un aumento en el peso de los frutos de tomate,
producto de aplicaciones foliares de calcio y MOHJO et al. (1995) señalan
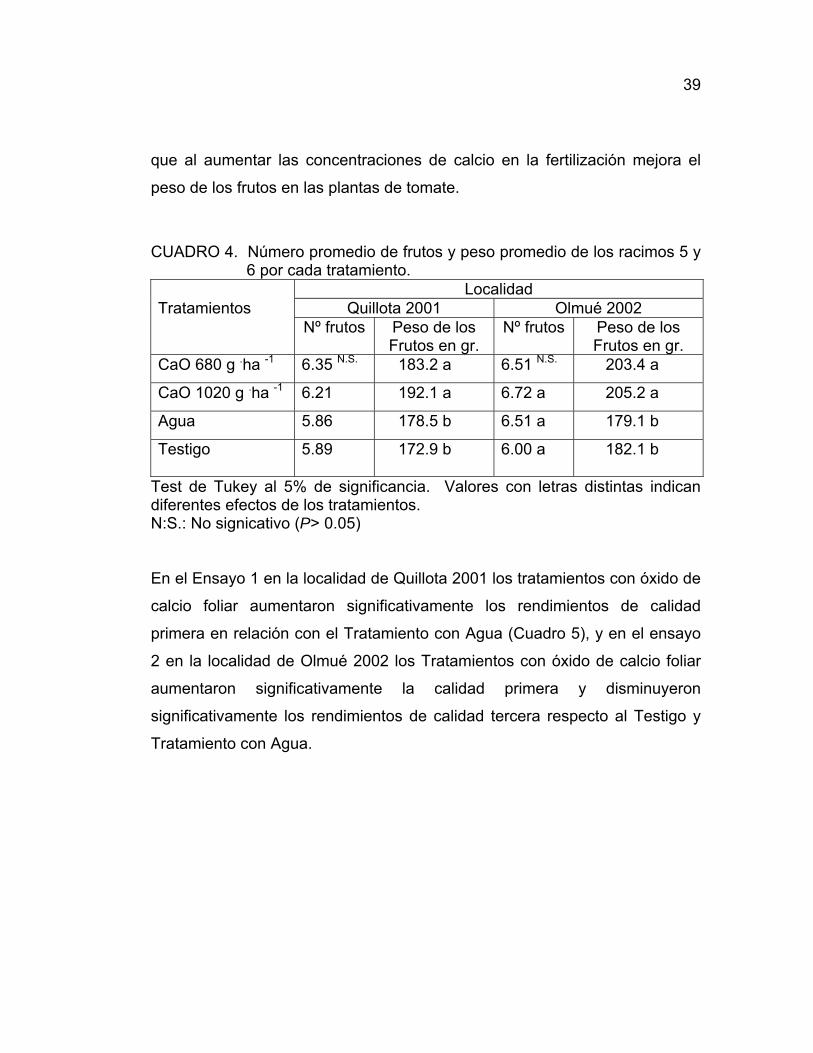
39
que al aumentar las concentraciones de calcio en la fertilización mejora el
peso de los frutos en las plantas de tomate.
CUADRO 4. Número promedio de frutos y peso promedio de los racimos 5 y 6 por cada tratamiento.
Localidad Tratamientos Quillota 2001 Olmué 2002 Nº frutos Peso de los
Frutos en gr. Nº frutos Peso de los
Frutos en gr. CaO 680 g .ha -1 6.35 N.S. 183.2 a 6.51 N.S. 203.4 a
CaO 1020 g .ha -1 6.21 192.1 a 6.72 a 205.2 a
Agua 5.86 178.5 b 6.51 a 179.1 b
Testigo 5.89 172.9 b 6.00 a 182.1 b
Test de Tukey al 5% de significancia. Valores con letras distintas indican diferentes efectos de los tratamientos. N:S.: No signicativo (P> 0.05)
En el Ensayo 1 en la localidad de Quillota 2001 los tratamientos con óxido de
calcio foliar aumentaron significativamente los rendimientos de calidad
primera en relación con el Tratamiento con Agua (Cuadro 5), y en el ensayo
2 en la localidad de Olmué 2002 los Tratamientos con óxido de calcio foliar
aumentaron significativamente la calidad primera y disminuyeron
significativamente los rendimientos de calidad tercera respecto al Testigo y
Tratamiento con Agua.

40
CUADRO 5. Distribución de calidades primera, segunda y tercera de los racimos 5 y 6 en gramos por cada tratamiento.
Localidad Tratamientos Quillota 2001 Olmué 2002 1era 2da 3era 1era 2da 3era
CaO 680 g .ha -1 468 a 522 N.S. 183 N.S. 634 a 431 N.S. 257 a
CaO 1020 g .ha -1 497 a 540 154 621 a 413 279 a
AGUA 294 b 504 222 434 b 384 347 b
TESTIGO 382 ab 484 187 440 b 360 347b
Test de Tukey al 5% de significancia. Valores con letras distintas indican diferentes efectos de los tratamientos. N.S.: No signicativo (P> 0.05)
4.4.2. Regresión lineal múltiple sobre el peso de los frutos
A continuación se presentan los modelos de regresión lineal múltiple
generados con los datos de los Ensayos 1 y 2.
Peso de los frutos = -0.06 + 0.02 . Calcio proximal + 0.03 . Calcio distal (1)
Peso de los frutos = -0.25 + 0.04 . Calcio proximal + 0.02 . Calcio distal (2)
Peso de los frutos = -0.08 + 0.01 . Calcio proximal + 0.02 . Calcio distal (3)
Peso de los frutos = 0.07 + 0.03 . Calcio proximal + 0.01 . Calcio distal (4)
Los modelos presentados incluyen solamente los conjuntos mínimos de
predictores que permitió obtener un coeficiente de regresión estabilizado.
La relación entre el peso de los frutos y la concentración de calcio del tejido
foliar de la zona proximal del foliolo y la concentración de calcio del tejido
foliar de la zona distal del foliolo es de un 97% (1) con los datos del testigo en

41
el Ensayo 1 en la localidad de Quillota 2001 y de un 98.8% (2) en el Ensayo
2 en la localidad de Olmué 2002, pero el porcentaje de correlación disminuye
cuando se utilizan los datos de todos los tratamientos a un 41.5% (3) en el
Ensayo 1 en la localidad de Quillota 2001 y a un 48.3% (1) en el Ensayo 2 en
la localidad de Olmué 2002, aunque disminuye la correlación sigue siendo
alta, por lo que esta forma de calcio en este caso sí aumenta el peso de los
racimos, lo que coincide con lo señalado por HO (2002).

42
5. CONCLUSIONES Existe relación entre la incidencia de clorosis apical y el contenido de calcio
foliar y el ataque de Botrytis cinerea, en donde una disminución de los
contenidos de calcio provoca un aumento de la incidencia de clorosis apical y
la incidencia de Botrytis cinerea.
No existe efecto de las aplicaciones de óxido de calcio sobre la clorosis y
necrosis marginal de los foliolos de las hojas en tomate. Del mismo modo,
las aplicaciones de óxido de calcio no disminuyen la incidencia de Botrytis
cinerea en las hojas de tomate.
Las aplicaciones de óxido de calcio aumentan los contenidos de calcio en los
tejidos proximal y distal del foliolo en la hoja en el Ensayo 2 en la localidad de
Olmué 2002. La concentración de calcio es mayor en el tejido proximal que
en el tejido distal de los foliolos.
Las aplicaciones de óxido de calcio aumentan el peso de los frutos. Ello se
relaciona con un aumento de la proporción de frutos de mayor tamaño
(primera), y una disminución de la cantidad de frutos de tamaño pequeño
(tercera).

43
6. RESUMEN En el presente trabajo se evaluó el efecto de la aplicación de óxido de calcio sobre la clorosis marginal de los foliolos de las hojas de tomate variedad Fortaleza, en invernadero frío, para lo cual se realizaron dos ensayos. El Ensayo 1 se desarrolló en la localidad de Quillota, entre los meses de mayo y diciembre del año 2001 y el Ensayo 2 en la localidad de Olmué entre los meses de mayo y diciembre del 2002. Cada ensayo constó de cuatro tratamientos: CaO 680 g .ha -1, CaO 1020 g .ha -1, aspersión con agua y testigo sin aspersión. Se utilizó un diseño en bloques completamente al azar con cuatro repeticiones, cuya unidad experimental correspondió a 20 plantas, a las cuales se les evaluó desde la primera hoja desde el quinto racimo hasta la primera hoja del sexto racimo, los siguientes parámetros: porcentaje de área afectada con la sintomatología de clorosis apical, incidencia de clorosis apical, contenido de calcio en los tejidos proximal y distal del foliolo, incidencia de Botrytis cinerea; y en los racimos 5 y 6 se evaluó el número de frutos, el peso y la distribución de tamaños de estos. Mediante análisis de regresión lineal múltiple se analizaron los datos del testigo, sin aplicación de calcio en ambos Ensayos, detectándose que al disminuir los contenidos de calcio foliar aumenta la aparición del síntoma de clorosis marginal de los foliolos en hojas de tomate y también aumenta la incidencia de Botrytis cinerea. Sin embargo, la aplicación de óxido de calcio no diminuyó la sintomatología de clorosis, incidencia de Botrytis cinerea y no aumentó el número de frutos, pero sí aumentó el peso de los frutos en ambos Ensayos. Sin embargo, la aplicación de óxido de calcio no aumentó el contenido de calcio foliar y la proporción de frutos de mayor tamaño en el Ensayo 1 en la localidad de Quillota 2001, pero si aumentó los contenidos de calcio foliar y la proporción de frutos de mayor tamaño en el Ensayo 2 en la localidad de Olmué 2002. Las aplicaciones de óxido de calcio aumentaron el peso de los frutos, pero debería evaluarse si éstas mejorarían el peso de todos los frutos de los racimos de la planta de tomate, ya que si el aumento de peso de los frutos fuese sobre todos los racimos, esto podría traducirse en un aumento en el rendimiento y en beneficios económicos.

44
7. LITERATURA CITADA ADAMS, P. 1994. Nutrition of greenhouse vegetables in NFT hydroponic
systems. Acta Horticulturae 361: 245-
. and HO, L. 1989. Effects of constant and fluctuating salinity on the yield, quality and status of tomatoes. Journal of Horticultural Science 64: 725-732
. and HOLDER, R. 1992. Effects of humidity, calcium and salinity on the accumulation of dry matter and calcium by the leaves and fruit of tomato (Lycopersicon esculentum). Journal of Horticultural Science 67: 137-142
. 1991. Effect of diurnal fluctuations in humidity on the accumulation of nutrients in the leaves of tomato. Journal of Horticultural Science 66: 545-550
. 1986. Mineral Nutrition. In: J Atherton and J Rudich. (eds). The Tomato Crop. London, Chapman & Hall. pp 280 – 334
AGRIOS, G. 1996. Fitopatología. 3a.ed. México, Editorial Limusa. 839 p.
AIKMAN, D. and HOUTER, G. 1990. Radiation and humidity on transpiration: Implications for calcium levels in tomato leaves. Journal of Horticultural Science 65: 245-253
APABLAZA, G. 2000. Patología de cultivos: Epidemiología y Control Holístico. Santiago, Ediciones Universidad Católica de Chile. 347 p.
AZCON-BIETO, J. y TALON, M. 2000. Fundamentos de fisiología vegetal. Barcelona, McGraw-Hill. 522 p.

45
BAKKER, J. 1990. Effects of day and night humidity on yield and fruit quality of glasshouse tomatoes (Lycopersicon esculentum Mill.) Journal of Horticultural Science 65: 323-331
. and SONNEVELD, C. 1988. Calcium deficiency of glasshouse cucumber as effected by environmental humidity and mineral nutrition. Journal of Horticultural Science 63: 241-246
BANUELOS, G., OFFERMAN, G., and SEIM, E. 1985. High relative humidity promotes Blossom-end Rot on growing tomato fruit. HortScience 20: 894-895
BESOAIN, X. 2000. Manejo de enfermedades en sistemas de producción intensiva. In: Seminario internacional: Adaptaciones Tecnológicas para la modernización de cultivos forzados. Quillota, Universidad Católica de Valparaíso, 12 y 13 de octubre 2000. s.p.
BLANCARD, D. 1990. Enfermedades del tomate. Madrid, Mundi-Prensa. 212 p.
BORKOWSKI, J. 1984. Study on the calcium uptake dynamic by tomato fruits and Blossom-end Rot. Acta Horticulturae 145: 222-229
CANOVÁS, F. 1995. Manejo del cultivo sin suelo In: F. Nuez. (ed). El cultivo del tomate. Madrid, Mundi-Prensa. pp. 227-255.
CARVAJAL, M., MARTÍNEZ, V. and CERDÁ, A. 1999. Influence of magnesium and salinity on tomato plants grown in hydroponic cultural. Journal of Plant Nutrition 22: 177-190
CHAMARRO, J. 1995. Anatomía y fisiología de la planta. In: F. Nuez. (ed). El cultivo del tomate. Madrid, Mundi-Prensa. pp. 43-91.

46
CHOI, J., CHUNG, G. and SUH, S. 1997. Effect of night humidity on the vegetative growth and the mineral composition of tomato and strawberry plants. Scientia Hoticulturae 70: 293-299
EHRET, D. and HO, L. 1986. The effects of salinity on dry matter partitioning and fruit growth in tomatoes grown in nutrient film culture. Journal of Horticultural Science 61: 361-367
FERGUSON, I. and DROBAK, B. 1988. Calcium and the regulation of plant growth and senescence. Hortscience 23: 262-266
FLORES, E. 1999. La planta estructura y función. Costa Rica, Editorial Tecnológica de Costa Rica. 337 p (Vol. I.)
GALLAGHER, J. and BISCOE, P. 1978. Radiation absorption, growth and yiel of cereals. Journal of Agricultural Science 91: 47-60
GIACONI, V and ESCAFF, M. 1998. Cultivo de las hortalizas. 13a.ed. Santiago, Editorial Universitaria. 334 p.
GIL, F. 1995. Elementos de fisiología vegetal. Madrid, Ediciones Mundi Prensa. 1047 p.
GUZMAN, M. 2000. Manejo de la nutrición en cultivos en suelo y sustrato. In: Seminario internacional: Adaptaciones Tecnológicas para la modernización de cultivos forzados. Quillota. Universidad Católica de Valparaíso, 12 y 13 de octubre 2000. s.p.
HO, L. 2000. Mejoramiento de la calidad del fruto de tomate a través del cultivo. In: Seminario internacional: Adaptaciones Tecnológicas para la modernización de cultivos forzados. Quillota. Universidad Católica de Valparaíso, 12 y 13 de octubre 2000. s.p.

47
HOOD, T. and MILLS, H. 1994. Root-zone temperature affects nutrient uptake and growth of snapdragon. Journal of Plant Nutrition 17: 279-291
HOLDER, R. and COCKSHULL, K. 1990. Effects of humidity on the growth and yield of glasshouse tomatoes. Journal Horticultural Science 65: 31-39
HURD, T. and GRAVES, C. 1985. Some effects of air and root temperatures on the yield and quality of glasshouse grown tomatoes. Journal Horticultural Science 60: 369-371
KALRA, Y. 1998. Handbook of reference methods for plant analysis: soil and plant analysis. USA, Inc CRC. 300p.
KINET, J. and PEET, M. 1997. Tomato. In: H. Wien. (ed). The physiology of vegetable crops. Wallingford, CAB International. pp 207-258
KOMOSA, A. 1984. The maximum concentration of macroelement salt solution for repeated foliar nutrition of greenhouse tomato Lycopersicon esculentum Mill. cv. Ostona. Acta Horticulturae 145: 213-221
KREIJ, C. 1996. Interactive effects of air humidity, calcium and phosphate on blossom-end rot, leaf deformation, production and nutrient contents of tomato. Journal of Plant Nutrition 19: 361-377
MOHJO, M., KUWATA, C., YOSHIKAWA, K. and ITO, T. 1995 Effects of nitrogen form, nutrient concentration and Ca concentration on the growth, yield and fruit quality in NFT tomato plants. Acta Horticulturae 396 :145-152
MORARD, P., PUJOS, A., BERNADAC, A. and BERTONI, G. 1996. Effect of temporary calcium deficiency on tomato growth and mineral nutrition. Journal of Plant Nutrition 19: 115-127

48
MAROTO, J. 1995. Horticultura herbácea especial. 4a.ed. Madrid, Ediciones Mundi-Prensa. 611p.
MARSCHNER, H. 1995. Mineral nutrition of higher plants. 2nd.ed. London, Academic Press. 889p.
MULHOLLAND, B., FUSSELL, R., EDMONDSON, J., BHASHAM, J. and MCKEE, J. 2001. Effect of vpd, K nutrition and root-zone temperature on leaf area development, accumulation of Ca and K and yield in tomato. Journal Horticultural Science 76:641-647
., FUSSELL, M., EDMONDSON, R., BURNS, I., MCKEE, J and BASHAM., J. 2000. Effect of humidity and nutrient feed K/Ca ratio on physiological responses and the accumulation of dry matter, Ca and K in tomato. Journal of Horticultural Science and Biotechnology 75: 713-722
NAVAS, M. 2000. Control ambiental en cultivos forzados. In: Seminario internacional: Adaptaciones Tecnológicas para la modernización de cultivos forzados. Quillota. Universidad Católica de Valparaíso, 12 y 13 de octubre 2000. s.p.
NKANSAH, G. and ITO, T. 1995. Comparison of mineral absorption and nutrient composition of heat-tolerant and non heat-tolerant tomato plants at different root-zone temperature. Journal of Horticultural Sciencie 70: 453-460
PICKEN, A., STEWART, K. and KLAPWIJK, D. 1986. Germination and vegetative development. In: J Atherton and J Rudich. (eds). The Tomato Crop. London, Chapman & Hall. pp 111-166
POOVAIAH, B. 1988. Molecular and cellular aspects of calcium action in plants. HortScience 23: 267-271

49
RINCON, L., SAÉZ, J., PELLICER, C. y BALSALOBRE, E. 1991. Extracción de macronutrientes en cultivo de tomate (Lycopersicon esculentum Mill.) de crecimiento indeterminado. Agrícola Vergel 4: 211-214
SALISBURRY, F y ROSS, C. 1994. Fisiología Vegetal. 4a.ed. México, Editorial Iberoamérica. 759 p.
SILVA, B. 1991. Calcio, un Problema Nutricional frecuente en numerosas especies de frutales y hortalizas. Aconex 31:17-20
TAIZ, L. and ZEIGER, E. 1998. Plant Physiology. Sunderland, Sinauer Asosciates Inc. 792 p.
TRINIDAD, A. y AGUILAR, D. 1999. Fertilización foliar, un respaldo importante en el rendimiento de los cultivos. Terra 17: 247-255
TUKEY, H., and MARCZYNSKI, S. 1984. Foliar nutrition – old ideas rediscovered. Acta Horticulturae 145: 205-212
VALENCIA, P. 1996. Acumulación de nutrientes en tomate (Licopersicon esculentum M.) de otoño c.v. FA- 144 cultivado en sustrato perlita en invernadero frío. Quillota, V Región. Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de agronomía. 152 p
WIEN, H. 1997. Lettuce. In: H. Wien. (ed). The physiology of vegetales crops. Wallingford, CAB international. pp 479-509

50
ÍNDICE DE MATERIAS 1. INTRODUCCIÓN 1 2. REVISIÓN BIBLIOGRÁFICA 3 2.1. Clasificación botánica y descripción del tomate 3 2.2. Nutrición con calcio 4 2.2.1. Funciones fisiológicas del calcio 4 2.2.1.1. Estabilización de la pared celular 5 2.2.1.2. Estabilización de la membrana 6 2.2.1.3. El calcio como mensajero secundario 6 2.2.2. Absorción del calcio 7 2.3. Transporte de calcio en el interior de la planta 10 2.4. Factores que afectan la absorción de calcio 11 2.4.1 Déficit de presión de vapor 11 2.4.2 Temperatura de la zona radicular 12 2.4.3. Incidencia de luz 13 2.4.4. Salinidad 14 2.5. Extracción de calcio en cultivo de tomate 14 2.6. Deficiencia localizada de calcio en la hoja 15 2.7. Deficiencia localizada de calcio en el fruto 16 2.8. Botrytis (Botrytis cinerea) 17 2.8.1. Condiciones predisponentes 18 2.8.2. Control cultural 19 3. MATERIALES Y MÉTODO 21 3.1. Invernadero: 21 3.2. Material vegetal: 21 3.3. Ensayo 1: Evaluación de aplicaciones foliares de óxido de calcio
sobre la incidencia de desorden fisiológico de clorosis foliar en Quillota 21
3.3.1. Tratamientos 22 3.3.2. Mediciones 23 3.3.2.1. Síntomas foliares del desorden fisiológico 23 3.3.2.2. Incidencia de Botrytis cinerea 25 3.3.2.3. Contenido de calcio foliar 26 3.3.2.4. Parámetros agronómicos 26 3.4. Ensayo 2: Evaluación de aplicaciones foliares de óxido de calcio
sobre la incidencia de desorden fisiológico de clorosis foliar en Olmué: 27
3.5. Diseño estadístico 28

51
3.6. Análisis estadístico de los datos 28 3.6.1. Análisis de varianza y comparación de media 28 3.6.2. Modelo de regresión lineal múltiple 29
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS 31 4.1. Clorosis marginal de foliolos e intensidad del síntoma 31 4.1.1. Análisis de varianza y separación de medias, para la
sintomatología de clorosis marginal 31 4.1.2. Regresión lineal múltiple sobre la sintomatología de clorosis
marginal 32 4.2. Contenido foliar de calcio 33 4.3. Porcentaje de incidencia de Botrytis cinerea 35 4.3.1. Análisis de varianza y separación de medias sobre la incidencia
de Botrytis cinerea 35 4.3.2. Regresión lineal múltiple sobre la incidencia de Botritys cinerea 37 4.4. Variables agronómicas 38 4.4.1. Análisis de varianza y separación de medias sobre las variables
agronómicas 38 4.4.2. Regresión lineal múltiple sobre el peso de los frutos 40 5. CONCLUSIONES 42 6. RESUMEN 43 7. LITERATURA CITADA 44

52
AGRADECIMIENTOS
Al profesor guía de esta investigación Sr. Javier Devia, por su constante
ayuda y dedicación, tanto en la realización de los ensayos como en la
confección del escrito.
Al Sr. Eduardo Oyanedel por su constante guía y apoyo en la confección del
escrito.
A todas las personas, compañeros y amigos, especialmente a Lorena Rojo y
Gloria Páez que de una u otra forma contribuyeron a llevar a cabo esta
investigación.

53
Al esfuerzo y dedicación que
ponen las personas humildes
para lograr cumplir con sus
sueños.

54
TALLER DE LICENCIATURA
EEffeeccttoo ddee llaa aapplliiccaacciióónn ffoolliiaarr ddee óóxxiiddoo ddee ccaallcciioo ssoobbrree llaa cclloorroossiiss
mmaarrggiinnaall eenn hhoojjaass jjóóvveenneess ddee ttoommaattee ccvv.. FFoorrttaalleezzaa bbaajjoo iinnvveerrnnaaddeerroo
ffrrííoo..
QUILLOTA 2003
UNIVERSIDAD CATÓLICA DE VALPARAÍSO
FACULTAD DE AGRONOMÍA
ÁREA DE HORTALIZAS Y FLORES


















