ESTUDIO TASA METABÓLICA BASAL EN BAHÍA MAGDALENA …biblio.uabcs.mx/tesis/te2963.pdf ·...
83
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA TESIS ANÁLISIS COMPARATIVO DE LAS ZONAS DE ALIMENTACIÓN Y DIETA DE TORTUGA VERDE (Chelonia mydas): CASO DE ESTUDIO TASA METABÓLICA BASAL EN BAHÍA MAGDALENA B.C.S. MÉXICO QUE COMO UNO DE LOS REQUISITOS PARA OBTENER EL TITULO DE BIÓLOGO MARINO PRESENTA: GABRIEL DE JESÚS PEÑA URIBE DIRECTOR: DR. RAFAEL RIOSMENA RODRÍGUEZ LA PAZ B.C.S. JUNIO 2013
Transcript of ESTUDIO TASA METABÓLICA BASAL EN BAHÍA MAGDALENA …biblio.uabcs.mx/tesis/te2963.pdf ·...

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
TESIS
ANÁLISIS COMPARATIVO DE LAS ZONAS DE ALIMENTACIÓN Y DIETA DE TORTUGA VERDE (Chelonia mydas): CASO DE
ESTUDIO TASA METABÓLICA BASAL EN BAHÍA MAGDALENA B.C.S. MÉXICO
QUE COMO UNO DE LOS REQUISITOS PARA OBTENER EL TITULO DE
BIÓLOGO MARINO
PRESENTA:
GABRIEL DE JESÚS PEÑA URIBE
DIRECTOR:
DR. RAFAEL RIOSMENA RODRÍGUEZ
LA PAZ B.C.S. JUNIO 2013

II
Dedicatoria
Especialmente dedico este trabajo a mi esposa Laura Esmeralda Montañez
González por su constante apoyo, motivación, cariño, paciencia, amor, ternura,
besos, abrazos, que me dieron las fuerzas para seguir siempre adelante y echarle
todas las ganas y sobre todo no dejarme caer en el camino para poder realizar
todas mis actividades. TE AMO PRINCESA.
Por otra parte a mis padres Ana Gabriela Uribe y Gabriel Peña, tanto como a mis
abuelitos Virginia Ramírez y José Uribe, Henna Amozorrutia y Paulino Peña; por
su apoyo en estos años que tengo viviendo aquí en la Ciudad de La Paz. A mis
tías Patricia Peña y Mónica Uribe que siempre me han brindado todo su cariño. A
mis hermanas Frida Victoria Uribe y a Gabriela Peña que siempre estuvieron
conmigo. A mis tíos Jaime Muro, Víctor Uribe y Fernando Máynez por todas las
risas brindadas y sin olvidar a mis primos queridos, José Muro, Fernando Máynez
Jr. y Paulina Máynez.
No me olvido de mis amigos, Qué sería la carrera sin estas personas Diego
Rodrigo Azcoitia Rocher (No parentesco con los chocolates), Ricardo Mirsha
Martin Mata, Daniel Alejandro Martínez, por sus temas de discusión y tonterías
que aparecían a lo largo de nuestra amistad, sobre todo por el apoyo incondicional
que me brindaron cuando más lo necesite. Abigail Aguilar y Manuel González, que
gracias a todos ellos mis días de trabajo, tanto en mi casa como en la universidad,
fueron más agradables y más relajados.

III
Agradecimientos
Agradezco principalmente a mi Director Rafael Riosmena Rodríguez y a Rocío
Marcín Medina por sus valiosas enseñanzas, que siempre me motivaron y
creyeron en mi más que yo mismo, por su valioso tiempo dedicado en la
realización de este trabajo, a pesar de las canas verdes que casi les saco, pero
mostrando siempre disponibilidad para responder mis dudas, por creer en mí,
darme siempre ánimos para seguir adelante muchas gracias Rafa y Rocío.
Alberto León G. Villacorta y Alicia G. Villacorta por ser como una familia adoptiva
aquí en esta ciudad, mostrándome cariño y apoyo en todos los sentidos de mi
vida. A los tíos Héctor Villarreal Peña y Clarisa por apoyarme en el primer año en
que me adapte a la vida en esta nueva ciudad.
A mi comité revisor, Dra. María Mónica Lara Uc y Dr. Carlos Sánchez que también
me apoyaron, siempre tratándome de llevar por un mejor camino y sobre todo una
gran disposición de ayuda. Al Dr. Gustavo Hinojosa Arango de la Escuela de
Estudios de Campo de Puerto San Carlos, por la confianza de brindarnos sus
datos de mucho tiempo de su trabajo.
Gracias a los compañeros del laboratorio de Botánica Marina, por siempre
recibirme con buenas caras y tratos, que crean o no es súper estar ahí rodeado de
ustedes.
Por último, a las personas que sin ellos no se podría encontrar nada en la
biblioteca de la uni, un agradecimiento enorme a los bibliotecarios, especialmente
a “don Deivid” que aparte de ser compañero de equipo de futbol, siempre mostro
disponibilidad de tiempo en las consultas de los libros.

IV
RESUMEN. La tortuga verde,Chelonia mydas,es un omnívoro que estáprotegido en México por la NOM-ECOL-O59 y es considerada en la IUCN como "especie en peligro". Presenta una dieta que varía ontogénicamente que va de medusas, crustáceos y pequeños teleósteoshastaplantas marinas. Se ha demostrado que la alimentación está relacionada con la abundancia de las principales especies que consume, pero también respecto a su valor nutrimental. Sin embargo, hasta ahora no se tiene un mapa detallado de las zonas de alimentación de esta especie y no se ha valorado si las principales especies que componen su alimentación. No se conoce si presentan los mismos valores calóricos, por lo cual las estrategias de conservación se han visto limitadas. A partir de observaciones sobre tallas diferentes en zonas de alimentación similares se sugiere que la dieta debe estar estrechamente vinculada con su tasa metabólica basal en donde se tienen requerimientos mínimos energéticos para que la especie logre desarrollarse. Es por ello que el objetivo de este estudio es la evaluación del estado de conocimiento sobre la alimentación de C. mydasa nivel mundial para determinar si existen diferencias en composición específica o en los volúmenes consumidos. También, como caso de estudio se determinaron los valores nutricionales de las especies que componen la dieta y la cantidad de alimento que las tortugas necesitarían consumir paraobtener, las calorías que den satisfacción a sus necesidades metabólicas primarias para la zona de Bahía Magdalena. Esto se desarrolló mediante el cálculo de su Tasa Metabólica Basal (TMB). Nuestros resultados muestran que existen 28 zonas de alimentación de C. mydasa nivel mundial, de donde se han determinado 45 estudios referentes a su dieta, de los cuales 34 han llegado a nivel género/especie en macroalgas y nueve solo se han dejado a nivel de grandes taxa. El catálogo de la dieta está compuesto principalmente porGracilaria sp, Codium sp, Ulva sp, Zostera sp, Halodule sp, Enteromorpha sp, Padina sp, Laurencia sp, Sargassum sp, Dyctiota sp, Sabellidae, Ptilosarcus undulatus, Dosidicus gigas, Antipathes galapagensis, Natantia sp y Aplysia vaccariaen donde se observa que hay plantas marinas como animales. Energéticamente se determinó que las plantas marinas aportan una gran cantidad calórica. En cuanto la TMB se encontró quepueden cubrir sus necesidades básicasalimentándose solo de plantas. Este estudio muestra que la tortuga verde tiene una dieta preferente herbívora pero en algunas zonas con poca abundancia de especies, se vuelve mono específica por lo que no se considera balanceada y puede influenciar el éxito de la especie a nivel global. Se sugiere determinar especies que son parte de su dieta continua para que se consideren en alguna categoría de protección como estrategia complementaria para la protección de esta especie.

V
Índice Página de aprobación
Dedicatoria
Agradecimientos
Resumen
Índice
Lista de Figuras
Lista de Tablas
1. Introducción
1.1. Historia natural de Chelonia mydas
1.2. Ecología trófica de Chelonia mydas
1.3. Metabolismo y Tasa Metabólica Basal de C. mydas
2. Antecedentes
2.1. Estado de Conservación de C. mydas
2.2. Ecología Trófica de C. mydas y C. mydas agassizii: Caso de
estudio Bahía Magdalena Baja California Sur 3. Justificación
4. Hipótesis
5. Objetivos Generales
5.1. Objetivos específicos
6. Material y Métodos.
6.1. Recopilación bibliográfica y extracción de los datos a nivel mundial
6.2. Biodiversidad y Valor Biológico
6.3. Área de caso de estudio
6.3.1. Bahía Magdalena Baja California Sur
6.4. Tasa Metabólica Basal e Ingesta calculada de C. mydas
7. Resultados
7.1. Recopilación bibliográfica y extracción de los datos a nivel mundial
7.2. Biodiversidad y Valor Biológico
I
II
III
IV
V
VII
VIII
1
1
3
5
7
7
8
11
12
13
13
13
13
15
16
16
19
20
20
26

VI
7.3. Caso de Estudio Bahía Magdalena: TMB e ingesta calculada de C.
mydas.
8. Discusión
8.1. Situación actual de las zonas de alimentación de C. mydas
8.2. Importancia y Conservación de las tortugas marinas en Baja
California Sur
8.3. Tasa Metabólica Basal de C. mydas agassizii en Bahía Magdalena
9. Conclusiones
10. Bibliografía
11. Apéndice
29
32
32
37
38
41
42
64

VII
Lista de Figuras
Figura 1. Complejo lagunar Bahía Magdalena B.C.S.
Figura 2. Numero de publicaciónes sobre ecología trófica de C. mydas
Figura 3. Mapa de los países con estudios de zonas de alimentación y el
número de publicaciones que se encontraron en la revisión.
Figura 4. Total de especies reportadas en los contenidos estomacales de
C. mydas en las diferentes regiones del mundo que son: Pacífico
Centroeste (Pac. CE), Pacífico Suroeste (Pac. SO), Pacífico
Noroeste (Pac. NO), Pacífico Noreste (Pac. NE), Pacífico
Central (Pac. C), Golfo de California (GC) Golfo de Tailandia
(GT), Índico Suroeste (Indico SO), Atlántico Suroeste (Atl. SO),
Atlántico Noroeste (Atl. NO), Golfo de México (GMX)
Mediterráneo (MED) y el Caribe.
Figura 5. Gráfica del ANDEVA de las especies entre regiones.
Figura 6. Dendograma del Índice de Similitud de Bray-Curtis.
Figura 7. Proporción de taxa descritos globalmente en la dieta de C.
mydas
Figura 8. Índice de Valor Biológico de los contenidos estomacales por
taxa.
Figura 9. Gráfica del ANDEVA de especies entre grandes taxa.
18
20
21
23
25
26
27
27
28

VIII
Lista de Tablas
Tabla 1. Totalde especies por taxa en la dieta de C. mydas,
recopiladas en la revisión bibliográfica.
Tabla 2. Tabla ANDEVA dos vías.
Tabla 3. Grupos homogéneos de la comprobación de hipótesis de
Fisher y Tukey entre las regiones del mundo.
Tabla 4. Grupos Homogéneos de la comprobación de hipótesis de
Fisher y Tukey entre los taxa.
Tabla 5. Indicadores de diversidad de taxa para las diferentes
regiones.
Tabla 6. Presas más representativos de la dieta de C. mydas, en
cuanto a su frecuencia en las diferentes regiones.
Tabla 7. Especies representativas para los grupos en función de la
información disponible en la red.
Tabla 8. Tendencias centrales en cuanto a requerimientos calóricos
de C. mydas agassizii por día para satisfacer su TMB y
TM.
Tabla 9. Volúmenes de ingesta de grupos.
Tabla 10. Valores porcentuales de Ingesta aproximados de grupos
por región.
22
24
25
28
29
30
30
31
32
32

1
1. Introducción 1.1. Historia natural de Chelonia mydas
El género Chelonia incluye 1 especie y 2 subespecies: la tortuga verde del
Pacífico oriental C. mydas agassizii (Boucort, 1868) de Baja California a Perú y al
oeste de las Islas Galápagos, y la tortuga verde C. mydas (Linnaeus, 1758) en el
resto del mundo (Cliffton et al., 1982). Pertenece al Phylum Chordata, Subphylum
Vertebrata, Clase Reptilia, Orden Testudines, Suborden Cryptodiria y Familia
Chelonidae. Las características morfológicas que la identifican son: cabeza
mediana y chata, un par de escamas prefrontales elongadas y cuatro escamas
postorbitales, pico córneo, filoso y aserrado en sus bordes; la presencia en el
carapacho de cinco escudos centrales, cuatro pares laterales y 12 pares
marginales; el plastrón con cuatro escudos inframarginales en cada puente, sin
poros y con una uña en el borde anterior de cada aleta. En adultos y subadultos el
color negro está distribuido en todo el cuerpo en forma de manchas o moteado
(Márquez, 1990; 2000).
El ciclo de vida de las tortugas comienza cuando eclosionan y se dirigen al
mar, pero existen una diversos factores que afectan su sobrevivencia en el
momento en que dejan sus nidos, ya sea el cansancio o los predadores como
cangrejos y gaviotas (Delgado-Trejo 2002). En el mar, se encuentran a merced de
los vientos, corrientes y mareas, hasta que llegan a tallas que les permita nadar a
su disposición (NOAA, 1998), aquí se alimentan entre mantos de algas gigantes
durante un periodo largo de tiempo que muchos definen como los “años perdidos”
(Nichols et al., 2001). En esta etapa los hábitos de alimentación son
principalmente carnívoros, alimentándose de organismos invertebrados
suspendidos en el agua (López-Mendilarahasu et al., 2005).
En mar abierto, a medida que las tortugas crecen en tamaño (35-40 cm de
largo) se desplazan a aguas costeras someras donde se reclutan con adultos y
otros juveniles (Nichols et al., 2001; Seminoff et al., 2002), donde permanecerán

2
hasta su madurez (de 9-20 años) (Seminoff et al., 2002). Es entonces, cuando
emprenden el regreso a las playas donde anidarán para depositar sus propios
huevos.Las migraciones reproductivas de esta especie pueden cubrir miles de
kilómetros en aguas tropicales y sub tropicales del Pacífico entre las zonas de
alimentación y las zonas de anidación (Alvarado y Figueroa, 1991; Bowen, 1995;
Márquez, 2000; López-Mendilarahasu et al., 2005).En estos sitios se presentan
dos aspectos clave del ciclo de vida de las tortugas, conocidos como Regreso al
Sitio de Nacimiento RSN (Natal Homing, en inglés) y Fidelidad al Sitio de
Anidación FSA (Nest Site Fidelity, en inglés) (Reece et al., 2005). RSN, se refiere
a la probabilidad de que la hembra regrese al sitio donde nació para anidar ahí.
Mientras que FSA, son las anidaciones consecutivas en un área general durante
el resto de la vida de la hembra (Reece et al., 2005).
Se estima que hay aproximadamente 150 zonas de anidación para la
tortuga verde en el mundo, con unidades demográficas independientes, no
obstante solo de 10 a 15 son lo suficientemente grandes como para implicar
alrededor de 2000 hembras anidadoras al año (Groombridge, 1982)
Las zonas de anidación más importantes en el Pacífico Este Tropical por
número de hembras que anidan se encuentran en Hawái, Michoacán ylos
Archipiélagos de Revillagigedo (México); y de Galápagos (Ecuador) (Cliffton et al.,
1982; Márquez, 2000; Chassin-Noria et al., 2004; Dutton et al., 2008; Amorocho et
al., 2012).Las tortugas que depositan sus huevos en playas de Michoacán,
México; tienen en promedio de 70 huevos depositados por cada nido (Delgado-
Trejo 2002). Los huevos generalmente eclosionan después de 6-13 semanas de
incubación, dependiendo de la temperatura (Miller, 1997). Usualmente este
periodo decrece a medida que aumenta la temperatura de incubación (Ackerman,
1997). En promedio, 86% de los huevos de cada nido eclosionan con éxito en el
estado de Michoacán (Delgado-Trejo 2002).
La tortugaC. mydas es una especie circunglobal que se encuentra en
todos los mares y océanos tropicales y subtropicales del planeta (NOAA, 1998).

3
Latortuga prieta como se conoce en Baja California Sur y en gran parte de
Latinoamérica, es una sub especieChelonia mydas agassizii, habitante común de
las aguas del Pacífico Este Tropical (Cliffton et al., 1982, Márquez, 1990; 2000).En
el continente americano se distribuyen desde las costas al sur de California hasta
Perú, aunque cuando ocurre el efecto del “Niño” y las aguas están más templadas,
se le ha observado hasta Chile(Cliffton et al., 1982; Márquez 1990; López-
Mendilarahasu et al., 2005; Dutton et al., 2008; Amorocho et al., 2012).
1.2. Ecología trófica deChelonia mydas
Al abandonar las playas de anidación las crías inician una fase pelágica,
en la cual presentan una alimentación omnívora, principalmente carnívora, que va
desde medusas, crustáceos, hasta pequeños cefalópodos y teleósteos. (Bjorndal,
1980; Mortimer, 1981; Garnett et al., 1985; Hirth, 1997; NOAA, 1998) y
frecuentemente se les encuentra asociadas a mantos de Sargassum oceánicos
(Márquez, 2002), ya que estos lugares les ofrecen alimento y resguardo (Meylan y
Meylan, 2000).
Al alcanzar una talla entre 30 y 40 cm de largo del caparazón (Musick y
Limpus, 1997),C. mydasse recluta con los adultos y otros juveniles en zonas de
alimentación costera, en la cual ocurren cambios en sus hábitos alimenticios,
iniciando una dieta principalmente herbívora, compuesta principalmente de algas y
pastos marinos (Hirth, 1997). Ocupan un nicho de alimentación único entre las
tortugas marinas debido a que es la única especie de tendencia herbívora
(Bjorndal, 1997).Se dice que éste cambio en su dieta corresponde con la madurez
en su tracto digestivo (Wood y Wood, 1981) y a sus adaptaciones morfológicas
dentro de las cuales resalta una cabeza chata y pico con bordes filosos y
aserrados que le facilitan el corte de las algas y pastos marinos (Villegas Nava,
2006), pero no se ha demostrado de manera experimental.

4
Estos migrantes de larga distancia, se mueven regularmente entre las
zonas de alimentación y anidación que por lo general son separadas por miles de
kilómetros (Meylan 1995; Pough et al.,1998). La razón para estas migraciones es
que las playas de anidación son expuestas a un alto oleaje, y que las condiciones
no son propicias para el crecimiento de macroalgas y pastos marinos de los cuales
se alimentan (Carr, 1967), aunque expertos en botánica marina, mencionan que
en zonas tropicales las algas y pastos marinos presentan bajos índices de
biomasa (Riosmena-Rodríguez, 2013). En consecuencia, algunas poblaciones, se
cree que ayunan durante varios meses durante las migraciones y en las zonas de
anidación, debido a la falta de alimentos.Estudios del comportamiento de buceo en
tortugas C. mydas, por parte de Hays y colaboradores (2002) mencionan que el
comportamiento de buceo posiblemente es empleado para maximizar la ganancia
neta de energía y reducir al mínimo la perdida de energía donde la comida puede
estar ausente.
C. mydas frecuenta lugares someros cerca de la costa como arrecifes de
coral, lagunas costeras en aguas tropicales del mundo de Norte a Sur,debido a
que son zonas de algas y pastos marinos que usan para alimentarse.En varias
partes del mundo las tortugas prietas adultas(que miden > 70 a 100 cm) pueden
alimentarse predominantemente de pastos marinos (Mortimer, 1981) o de algas
(Pritchard, 1971; Bjorndal, 1985; 1997). También se pueden alimentar de materia
animal y vegetal cuando éstos están presentes en la misma zona (NOAA, 1998).
Por otra parte la variación de la composición de su dieta puede ser una
consecuencia de la disponibilidad localen función de su abundancia, ya que se
han observado cambios en las dietas conforme se dispone de nuevo alimento
como es el caso de algas invasoras en Hawái y Australia (NOAA, 1998), la
selectividad de la tortuga y/o el tipo de hábitat (Bjorndal 1980, Garnett et al., 1985).
A Pesar de la importancia de las zonas de alimentación que son vitales
paralas tortugas marinas, es donde sufren más alteraciones causadas por las
diversas actividades humanas como pesca incidental, pesca dirigida y

5
degradación del hábitat (Gardner y Nichols, 2001; Mancini y Koch, 2009)al
momento que los quelonios utilizan estos sitios para alimentarse de macroalgas y
pastos marinos (Riosmena-Rodríguez et al., 2009). Considerando que se conocen
los disturbios que se generan en estos hábitats críticos en Baja California Sur,
aúnestamos muy lejos de lograr un buen manejo de esta especie, debido a que no
se tienen regulaciones en estas zonas y también se tendría que incorporar un
manejo para las especiespresas de C. mydasagassiziipara poderle facilitarle un
forrajeo óptimo con una gran variedad y cantidad de alimento para que
consumalos nutrientes que requiera para cumplir la teoría de MacArthur y Pianka
(1966). Hasta el momentono se han reportado diferencias en la alimentación entre
C. mydas yC. mydas agassizii, se observan algunas diferencias en sus dietas
debido a la disponibilidad de algas invasoras como alimento para la verde pero no
para la prieta (NOAA, 1998),por lo tanto la base de este estudio es conocer y
comparar dichas diferencias.
1.3. Metabolismo y Tasa Metabólica Basal de C. mydas
Se conoce como metabolismo a las transformaciones químicas que sufren
los nutrientes en los tejidos, una vez superados los procesos de digestión y
absorción correspondientes. El metabolismo incluye reacciones de tipo
degradativo, que se utilizan para obtener energía (catabolismo) y reacciones de
tipo biosintético, formando diversas biomoléculas utilizando parte de esa energía
(anabolismo) (Gil-Hernández y Sánchez de Medina, 2004).
El Metabolismo Basal (MB), es aquella fracción de gasto energético
destinada al mantenimiento de las funciones vitales del organismo. Se expresa en
calorías, producidas por el sujeto a una temperatura entre 18 y 20°C. Esta
cantidad de calor está referida a metro cuadrado por superficie del cuerpo (Vidal,
1935). Los factores que influyen en el MB son: superficie corporal (altura y peso),
edad y sexo.Hulbert y Else (2003) hablan acerca del concepto de la TMB que fue

6
desarrollado para comparar la tasa metabólica de los animales y en un principio
era importante en un contexto clínico como un medio de determinación de estado
de la tiroides de los seres humanos. También es importante la relación alométrica
entre la masa corporal y la tasa metabólica de los mamíferos,es decir que su
metabolismo basal varía con la masa corporal, con el mismo exponente alométrico
como la tasa metabólica de campo y con muchos factores fisiológicos y
bioquímicos.
Robbins (1983) en sus estudios con animales silvestres en cautiverio,
utiliza una formula muy simple para la Tasa Metabólica Basal para mamíferos, que
se representa de la siguiente manera. TMB=70Kcal*(Masa Corporal en Kg.)0.75.
Robbins (1983) también indica que para las diferentes etapas de la vida de los
animales su TMB puede aumentar, sobre todo en mamíferos en su época de
reproducción es hasta 6 veces, Mientras que para reptiles se mantiene estable. En
estudios posteriores agrega correcciones a sus estimaciones de la TMB para
organismos con diferentes metabolismos, tomando en cuenta para reptiles es el
15% de una TMB de un mamífero (Robbins, 1993).
A pesar de que las tortugas verdes juveniles realizan migraciones
estacionales (Musick y Limpus, 1997), algunos organismos permanecen en el
mismo lugar durante todo el año y experimentan fluctuaciones de las condiciones
ambientales locales. Los cambios estacionales en conjunto con factores bióticos y
abióticos tienen efectos profundos en la energética de reptiles (Bennett, 1982). Las
fluctuaciones en la temperatura ambiental afectan directamente a las tasas de las
reacciones bioquímicas y procesos fisiológicos (Hochscheid et al.,2004), y los
cambios en los patrones de actividad relacionadas a los ciclos reproductivos y en
factores ecológicos tales como la disponibilidad de alimentos también contribuyen
a las diferencias estacionales sobre el metabolismo (Christian et al., 1996). Southwood y colaboradores (2006) investigaron los cambios estacionales
en relación con la demanda energética de las tortugas verdes juveniles de una
población residente en el sur de la Gran Barrera de Coral (Australia). Usaron un

7
método de agua doblemente marcada para determinar el flujo de agua y la tasa
metabólica de campo (FMR) de C. mydas en verano (temperatura media del agua
(TW) = 25,8 °C) y el invierno (media TW = 21,4 °C) donde no obtuvieron diferencia
significativa en el metabolismo de los quelonios juveniles experimentando sólo
cambios pequeños dentro del rango de temperaturas y condiciones ambientales. En cambio, estudios que se refieran al metabolismo basal de animales son
muy escasos, los más frecuentes son con mamíferos y la mayoría de estos, se
refieren a otros tópicos relacionando la TMB con la Masa Corporal (Hayssen y
Lacy, 1985), temperatura y metabolismo enfocados a la ecología (Clarke, 2006),
pero ninguno relacionados directamente con la alimentación de reptiles y menos
con tortugas marinas.
Además en estudios de López-Castro y colaboradores (2010) encontraron
diferencias entre las tallas de las poblaciones analizadas, en las que concluyen
que las tortugas pequeñas, están asociadas a zonas más someras debido a la
protección y donde obtienen una mayor disponibilidad de alimento o de mejor
calidad, ya que estas requieren un mayor consumo debido al metabolismo más
alto que las de mayor tamaño, pero esto último no se ha evaluado profundamente
por lo tanto el presente trabajo pretende recopilar información acerca de los
estudios realizados sobre los hábitats y preferencias alimenticias de C. mydas
agassiziien el mundo y relacionarlos con su TMB para calcular los volúmenes de
ingesta que debería estar consumiendo para poder satisfacer sus necesidades
metabólicas básicas.
2. Antecedentes 2.1. Estado de Conservación de C. mydas
Desde la antigüedad, las tortugas marinas han sido usadas como
alimento, con propósitos ceremoniales, medicinales y vestimenta por pueblos
principalmente ribereños (Cliffton et al., 1982; García-Martínez y Nichols 2000;

8
Brooks, 2005). A partir de 1950 el aprovechamiento de estas tortugas era de
manera artesanal, no obstante, al disminuir drásticamente las poblaciones de
cocodrilos, la industria de las pieles se enfocó principalmente a la captura de la
tortuga golfina (Lepidochelys olivacea), ya que su piel era adecuada para esta
industria (Márquez et al., 1982a). En la década de 1960 la demanda por pieles de
tortuga tuvo un gran incremento, a tal grado, que su captura se disparó a niveles
insostenibles y sus poblaciones empezaron a disminuir de manera considerable
(SEMARNAT, 2002; Márquez et al., 1982b); en este periodo, México contribuyó
con más del 50% de la pesca mundial de tortugas marinas (Márquez y Doi, 1973;
Márquez et al., 1982a; Márquez, 1990). A mediados de 1971 y a principios de
1973, la Subsecretaría de Pesca declaró una veda para reorganizar la captura de
tortugas. No obstante, a la captura comercial, se le unió una intensa captura ilegal
y contrabando, por lo que la abundancia de las poblaciones de tortugas continuó
decayendo; hasta que en 1990 el gobierno mexicano decretó la veda total de
todas las especies de tortugas marinas (DOF, 1990).
Según la IUCN (2013) la población de tortugas continúa reduciéndose y
que de los años de 1970 al 2001 la población mundial de C. mydas disminuyo
entre 48- 67%, dejando una población aproximada de 90,000 hembras maduras.
2.2. Ecología Trófica de C. mydasy C. mydas agassizii: Caso de estudio Bahía
Magdalena Baja California Sur
Los primeros estudios sobre alimentación de C. mydas corrieron a cargo
por Bjorndal (1979) con tema en la digestión de la celulosa y producción de ácidos
grasos posteriormente en 1980 un estudio sobre la nutrición y forrajeo de esta
tortuga. Estudiosa cargo de Mortimer (1981) donde se capturaban tortugas y se
obtenían los contenidos estomacales a través de la evisceración, tomaron
importancia ya que se podía observar de una manera clara la alimentación de la
tortuga verde. Al ser las tortugas una especie en peligro, resultaba contradictorio el

9
matarlas para su conservación. No es hasta que Forbes y Limpus (1993)
desarrollan un método no letal para obtener los contenidos de la dieta de las
tortugas marinas, mediante lavados estomacales.
Después de esta aportación, diversos investigadoresen el mundo
comenzaron a utilizar este métodoy las investigaciones de ecología trófica
avanzarían de una manera más rápida y certera con el avance de nuevas
tecnologías para la identificación de las presas como los isotopos estables,
marcadores moleculares microsatélites, etc.
Diversos estudios sobre ecología trófica de tortugas C. mydas, se han
realizado alrededor del globo. De las zonas más estudiadas, se encuentran Hawái,
con estudios de la NOAA (1998)McDermid et al. (2007) y Arthur y Balazs (2008),
en donde reportaron una gran cantidad de especies de algas Rhodophyta, entre
las cuales se destacaba la presencia de un alga invasoraAcanthophora
spicifera.Esto indica que C. mydas no presenta una selección en su alimentación
de especies entre las algas rojas. También se mencionaron en menor cantidad
algas verdes, cafés y materia animal.
Otros sitios de alimentación con muchos estudios están hechos en
Australia, en el que destaca el realizado por Fuentes y colaboradores (2006), en el
que describe una gran cantidad de especies para la dieta de C. mydas, cinco
especies de pastos marinos, dos de algas verdes, cinco de algas cafés, nueve
para algas rojas y cuatro para materia animal. Lo que podría tener en común con
otros artículos es la presencia de A. spicifera, pero no se tiene información de la
especie más que el género.
En Rio de Janeiro Brasil, son recientes los estudios de ecología trófica,
como el primero elaborado por Rodríguez et al. (2013), en el que recolectaron los
contenidos estomacales de 49 tortugasC. mydas varadas, de las cuales 12
contenían desechos de origen antropogénico, que murieron asfixiadas, debido a
que mostraron bloqueos por bolsas de polietileno en la tráquea y el estómago
vacío. En el resto de las tortugas se encontró la presencia de 16 algas rojas, 13

10
algas verdes y 8 cafés. Cabe resaltar que también se reportó la presencia de A.
spicifera.
La costa occidental de la Península de Baja California es de vital
importancia para las tortugas marinas debido a que presenta surgencias que crean
un área de gran productividad primaria y secundaria (Brusca, 1980; Ortega y
Arriaga, 1991). La presencia de lagunas costeras someras donde crece una gran
cantidad de macroalgas que constituyen la dieta principal de Chelonia mydas
agassizii(López-Mendilaharsu et al., 2005; López-.Castro et al., 2007); praderas de
pasto marino, y los bosques de kelp gigante (Morgan et al., 2005), proporcionan
hábitats clave para la alimentación y el desarrollo de la tortuga prieta (Cliffton et
al., 1982; Alvarado y Figueroa, 1991; Eckert, 1993; Seminoff, 2000; Senko et al.,
2010) y además que varias especies incluidas en la NOM 059 utilizan como áreas
de alimentación, descanso y desarrollo (Cliffton et al., 1982; Márquez, 1990;
Gardner y Nichols, 2001; Riosmena-Rodríguez, 2009). Así mismo se ha
determinado que tortugas inmaduras presentan una demanda de energía mucho
mayor que las tortugas adultas, prefiriendo áreas más protegidas donde el
esfuerzo durante la búsqueda de alimento sea menor que en otras zonas (López-
Mendilaharsu, 2003).
En BCSse han reportado una notable cantidad de estudios de ecología
trófica de C. mydas agassizii, como los de Seminoff et al. (1999, 2002), López-
Mendilaharsu (2002, 2003), López-Mendilaharsu et al. (2005, 2008) en el cual se
determina que existe una selección ontogénica de los recursos alimenticios; Koch
y colaboradores (2007) mencionan agrupaciones por edad de las tortugas en
cuanto a la disponibilidad de alimento en Bahía Magdalena en el área de canales,
en los cuales al Norte se segregan individuos juveniles mientras que al sur, se
observan organismos de mayor Talla. Villegas-Nava (2006) para esta zona
cuantifica los valores energéticos que brindan las especies de macroalgas y
pastos marinos reportadas para la dieta de C. mydas agassizii en esta localidad,
mientras que Rodríguez-Barón (2010) en sus estudios con isotopos estables

11
resalta la importancia de las algas rojas en la dieta de estas tortugas y la
variabilidad de materia animal de la cual se alimentan.
No obstante en la zona de Bahía Magdalena, la tortuga prieta sufre
grandes alteraciones antropogénicas directa o indirectamente mientras esta se
encuentra en la zona (Senko et al., 2010), las cuales causan una gran mortalidad
no solo de tortuga prieta si no de todas las tortugas que se resguardan en esa
área, debido a contaminación por desechos sólidos y agrícolas, pesca incidental y
furtiva hasta colisiones con embarcaciones (Rosales-Villa, 2004; Koch, 2006).
Aún falta mucho comprender sobre la ecología trófica de la tortuga verde,
sobre todo la aportación de energía que obtiene de sus presas y si esta es lo
suficiente para satisfacer sus necesidades metabólicas de tal manera que le
permita desarrollarse adecuadamente.
3. Justificación
Las tortugas marinas presentan una disminución poblacional severa,por lo
que se encuentran en categoría de especies amenazadas y en peligro a nivel
mundial (IUCN, 2013).Esto se debe a los impactos causadospor las actividades
humanas,y debido a esto es imperante implementar medidas que permitan la
conservación y recuperación de sus poblaciones (Cliffton et al., 1982; Figueroa et
al., 1993).Las percepciones de la importancia del cuidado de los recursos
naturales en nuestro país han cambiado con el tiempo, de forma que ahora se
reconoce el cuidado y conservación de la flora y fauna como una inversión a largo
plazo (SEMARNAT, 1999).
No obstante que la ley ordena la protección total de las tortugas marinas
en México con la NOM 059 SEMARNAT 2010 y la NOM 162 SEMARNAT 2012, la
captura ilegal continua siendo de las amenazas más fuertes para las poblaciones
que se alimentan en estos campos pesqueros, debido a que la carne de las
tortugas marinas se consume todavía (especialmente en ocasiones festivas) en

12
toda la península de Baja California (Gardner y Nichols 2001; Koch et al., 2006).
Desde marzo 2006 a febrero 2008 se documentaron 1014 organismos muertos en
BC; de estos 461 (46%) se habían consumido, y el 77% eran tortugas prietas (C.
mydas) (Mancini y Koch, 2009). Todavía hay varios lugares en donde se consume
carne en el comercio local, regional, e incluso a veces en el mercado negro
internacional (Mancini y Koch, 2009).
Los esfuerzos de conservación de C. mydas están principalmente dirigidos
a proteger las zonas de anidación en la NOM-162-SEMARNAT (DOF, 2012),
mientras que no existe una Norma Oficial Mexicana que regule los efectos de la
sobreexplotación en las poblaciones y diversidad en sitios de alimentación se
encuentran aún bajo criterios científicos, ya que no hay ley que indique su cuidado
en estas zonas.
Estudios sobre el uso de hábitat de alimentación y dietas de C.
mydasagassiziien la Península de Baja California Sur destacan la importancia de
las lagunas costeras y la variabilidad de organismos de las cuales C. mydas se
alimenta(Seminoff et al., 1999; Seminoff et al., 2002; Bird et al., 2003; Brooks,
2005; Etnoyer et al., 2006; Seminoff y Jones, 2006; Koch et al., 2007; Lopez-
Castro et al., 2007; Senko et al,, 2008; Megil et al., 2009; Richardson et al., 2010;
Senko et al., 2010), pero estudios acerca de las necesidades alimentarias para
satisfacer sus necesidades metabólicas son escasos en esta zona y el mundo.
4. Hipótesis
La degradación de los hábitats están provocando una disminución de
biomasa alimento para Chelonia mydas, debido a esto la cantidad de energía
obtenida por alimento podría encontrarse muy cercana al requerimiento mínimo
para satisfacer su tasa metabólica basal.

13
5. Objetivos Generales
1. Recopilar sintetizar y analizar,información de artículos publicados en revistas
indexadas a nivel mundial, tesis, reportes técnicos publicados y no publicados
sobre hábitats de alimentación y dietas de tortuga verde (C. mydas).
2. Realizar un ejercicio de estimaciones de las cantidades de las calorías que
debería consumir para satisfacer su tasa metabólica basal con datos de
tortuga prieta (C. mydas agassizii) de Bahía Magdalena B.C.S.
5.1. Objetivos específicos
1.1 Desarrollar una recopilaciónbibliográfica de artículos publicados en revistas
indexadas, tesis y reportes técnicos sobre el uso de hábitat y ecología trófica
de C. mydas y organizarlos por regiones.
1.2 Extraer las listas de las especies encontradas en los contenidos estomacales y
lavados esofágicos de C. mydas por regiones del mundo.
1.3 Aplicar el Índice de Valor Biológico para inferir cual es la especie más
importante en la alimentación de C. mydas.
2.1 Capturar datos de informaciones nutrimentales de algas y valores calóricos,
para obtener la cantidad de calorías ingerida por C. mydas.
2.2 Calcular la tasa metabólica basal de C. mydas agassizii y compararlas para ver
si la energía consumida es la suficiente para que esta especie se desarrolle.
6. Material y Métodos 6.1. Recopilación bibliográfica y extracción de los datos a nivel mundial
Para la recopilación bibliográfica se desarrollaron búsquedas de bases de
datos en la web de Seaturtle.org y en Google Académico, artículos de revistas
indexadas, tesis, libros (impresos y electrónicos) y reportes técnicosacerca de uso

14
de hábitat de alimentación y ecología alimentaria de C. mydas en diferentes
lugares del mundo. Para la búsquedase utilizó la siguiente sistemática en criterios
de exploración: (Chelonia mydas, feeding habitat, feeding ecology, foraging
ground, diets y sus traducciones al español).Los documentoscon temas de
ecología alimentaria y proporcionaran datos de las ingestas de C. mydas, se
registraron y dividieron por regiones. Para la regionalización se dividieron los
océanos, dos partes para el Índico (Este y Oeste) y cuatro para el océano Pacífico
y Atlántico (Norte, Sur, Este y Oeste), para disminuir el número de regiones y
observar una mejor distinción en los gráficos, debido a que si usamos las regiones
ecológicas marinas ya preestablecidas el número de estas aumentaría
considerablemente.
Se realizó una base de datos de los nombres de las especies registradas
para la dieta de C. mydasy C. mydas agassizii en una tabla de presencia-
ausencia de las diferentes regiones. Debido a las diversas maneras en que los
autores de los artículos registraron las especies descritas en la dieta de C. mydas,
se optó por agruparlos en grandes Taxa (pastos marinos, Clorophyta, Rhodophyta,
Ochrophyta, Cyanophyta y materia animal).
Posteriormente se elaboró un planisferio donde se ubicaron los sitios de
alimentación estudiados y la cantidad de trabajos realizados. También se
elaboraron gráficos para facilitar el análisis de los datos registrados en la tabla.
Se realizó un ANDEVA de dos vías sin interacciones, los resultados de los
efectos por región se utilizaron para comparar estadísticamente cuales eran las
regiones más importantes por número de especies descritas. Se analizaron los
supuestos de Homogeneidad y Homocedasticidad de las varianzas para poder
realizar este análisis, los cuales fueron rechazados, por lo tanto los datos se
tuvieron que ajustar mediante una normalización para poder cumplir con estas
premisas. Por último se realizaron las pruebas de hipótesis de Tukey y Fisher para
poder observar cuales eran los grupos que son estadísticamente iguales (Grupos
Homogéneos) y saber cuál era la región más importante. A su vez se estimó el

15
grado de semejanza de los grupos reportados entre las diferentes regiones
utilizando el índice de similitud de Bray-Curtis (Primer 6), por medio de una unión
no ponderada, al igual que el ANDEVA, los datos se tuvieron que normalizar para
poder realizar esta prueba. Posteriormente se realizó un dendograma, para ver la
similitud de las regiones en una manera gráfica.
El dendograma se comparó con el ANDEVA de dos vías realizado
anteriormente, se usaron los efectos por grupos de especies para comprobar el
resultado de este índice con respecto al grupo más importante estadísticamente y
para ver cuál era la región más importante con respecto a especies reportadas.
Debido a la robustez de los datos, se tuvieron que normalizar ya a que no
cumplían con los supuestos del ANDEVA (Zar, 2010).
6.2. Biodiversidad y Valor Biológico
Para obtener cuál de los grupos de los que se alimenta C. mydas, se
realizó el Índice de Valor Biológico propuesto por Sanders (1960), que se utiliza
para ordenar la importancia de las especies el cual consiste en asignar un valor a
cada especie en función de su abundancia en cada muestra y se expresa en
puntajes. Es una herramienta que permite la selección de especies
representativas de una comunidad e integra los valores cíclicos a manera de las
especies temporales en las dominancias de las especies principales (Loya-Salinas
y Escofet, 1990). Este índice tiene como ventaja la combinación de datos de
abundancia y frecuencia de aparición de cada una de las especies. Con el
ANDEVA y sus interacciones por especie, se realizaron las pruebas de hipótesis
de Tukey y Fisher, para observar que grupos homogéneos se formaron y así
comparar lo obtenido con el IVB.
Se estimó la abundancia de los grupos a partir de las presencias
acumuladas de cada especie y la riqueza de especies de Margalefde cada
muestra para ver qué región es la que tiene una mayor cantidad de especies

16
descritas para la dieta de C. mydas. Es una medida de la cantidad de especies
presentes, que hace un margen para el número de individuos de grupos por sitio(d
= (S-1) / Log (N)). Se calculó el índice de diversidad de Shannon-Wiener (H´) que
considera de manera ponderada la porción de la abundancia de las especies, así
como la riqueza dentro de la muestra (Clarke y Gorley; 2006); el índice se expresa
de la siguiente manera:
H´=- Σ (ni/N) loge (ni/N)
Donde:
N= número total de individuos de todos los grupos en cada región
ni= número de individuos del grupo i en una región
También se estimó la Uniformidad de Pielou (J´), en el cual expresa que
tan homogéneamente se encuentran distribuidos los individuos de los diferentes
grupos en cada región. Este índice estandariza el valor de H´ en una escala de 0 a
1, considerando que la máxima diversidad se obtendría en el caso de que todas
las especies tengan la misma abundancia, mientras que si hay una mayor
dominancia de alguna especie en particular, se obtendrá un valor menor de
equitatividad (Clarke y Gorley; 2006). El índice se calcula de la siguiente manera:
J´= H´/log S
Donde:
H´= Valor de Shannon- Wiener
S=Número de grupos en la región
6.3. Área de caso deEstudio 6.3.1. Bahía Magdalena Baja California Sur
Bahía Magdalena (Fig. 1) es un sistema lagunar que se localiza en la región
central de Baja California Sur entre los 25° 20´N, 112° 15´W y 24° 15´N, 111°
30´W. Forma parte del complejo lagunar con Bahía Almejas con la que se

17
comunica a través de un canal y más de 40 esteros por lo cual es el mayor
ecosistema de humedales en la costa oeste de Baja California con más de 200 Km
de largo (Morgan, et al., 2005). Esta se divide en tres zonas hidrológicas
principales: la norte o de canales, central y la sur, también llamada Bahía Almejas.
La zona Norte se comunica con el Océano Pacífico, a través de una
angosta entrada de aproximadamente un kilómetro de ancho, llamada Boca de
Soledad. Esta da paso a un estrecho sistema de canales que alcanzan una
profundidad de 3.5 m. y rodeada por mangle, lo que lo hace de difícil acceso para
la navegación, al sur se prolonga siete kilómetros y termina al sureste, en Puerto
Adolfo López Mateos En las partes someras existen grandes áreas cubiertas de
algas y pastos marinos, con manglares en la mayoría de sus márgenes (López-
Mendilaharsu, 2002). Al sur el canal ofrece mejores condiciones para navegar con
profundidades promedio de once metros y queda separado del mar por una
estrecha barrera litoral, formado por arena muy fina. La zona central, se conecta
con el Pacífico por una boca de 38 kilómetros de profundidad aproximadamente y
5.5kilómetros de ancho. La zona Sur, también llamada Bahía Almejas, se
comunica con el Océano por una boca de 2.5 kilómetros de ancho
aproximadamente y 30 metros de profundidad que forman los extremos de Isla
Margarita e Isla Creciente.

18
Figura 1. Área de estudio complejo lagunar Bahía Magdalena B.C.S.los puntos
rojos son los sitios de muestreo
N México
Sonora
Sinaloa
Baja Californi
Baja California Su
Mar de Cortés
Océano Pacífico

19
6.4. Tasa Metabólica Basal e Ingesta calculada de C. mydas
Para calcular las cantidades de ingesta que debería estar teniendo cada
tortuga en el área de Bahía Magdalena, se realizaron despejes de la fórmula de
Robbins (1993), se seleccionaron las especies más representativas para la dieta
de C. mydasa nivel mundial mediante el criterio de frecuencia de aparición entre
las diferentes regiones en una tabla. Posteriormente se realizó otra revisión
bibliográfica en diversas fuentes como páginas de Internet, artículos publicados,
Monografías y Tesis, con los criterios de búsqueda siguientes: (contenido
energético, información nutrimental, análisis químicos, contenidos energéticos,
valores calóricos de “Especie de planta o animal” al igual que sus traducciones al
inglés) de las presasmás representativas de cada grupo en la dieta de C. mydas.
Para el cálculo de la Tasa Metabólica Basal se utilizaron colectas
realizadas entre 2000 y 2011 como parte del monitoreo en los sitios de colecta
(Fig. 1) con base en la metodología descrita en López-Mendilaharsu et al., (2005,
2008). Una vez capturado el organismo se le peso y se le midió el lago/ancho del
caparazón. Se calcularon los IMCs para cumplir con la fórmula de Robbins (1993)
debido a quenosproporcionó una base de datos con pesos y longitudes de 695
tortugasC. mydascapturadas en Bahía Magdalena Baja California Sur, que sirvió
para calcular la Tasa Metabólica Basal de esas tortugas, a su vez, se
compararon con los volúmenes de ingesta de las tortugas que se alimentan en la
Bahía Magdalena B.C.S. obtenidos de Rodríguez-Barón (2010), para saber si las
cantidades aproximadas de ingesta suficiente para satisfacer su TMB.
Los datos Fueron agrupados en una tabla de descripción estadística para
facilitar su interpretación. Para todos los análisis ecológicos se utilizó el software
Primer 6 Demo (Plymouth Routines in Multivariate Ecological Research 6.0), y
para los estadísticos el Statistica 10.
20
7. Resultados
7.1. Recopilación bibliográfica y extracción de los datos a nivel mundial
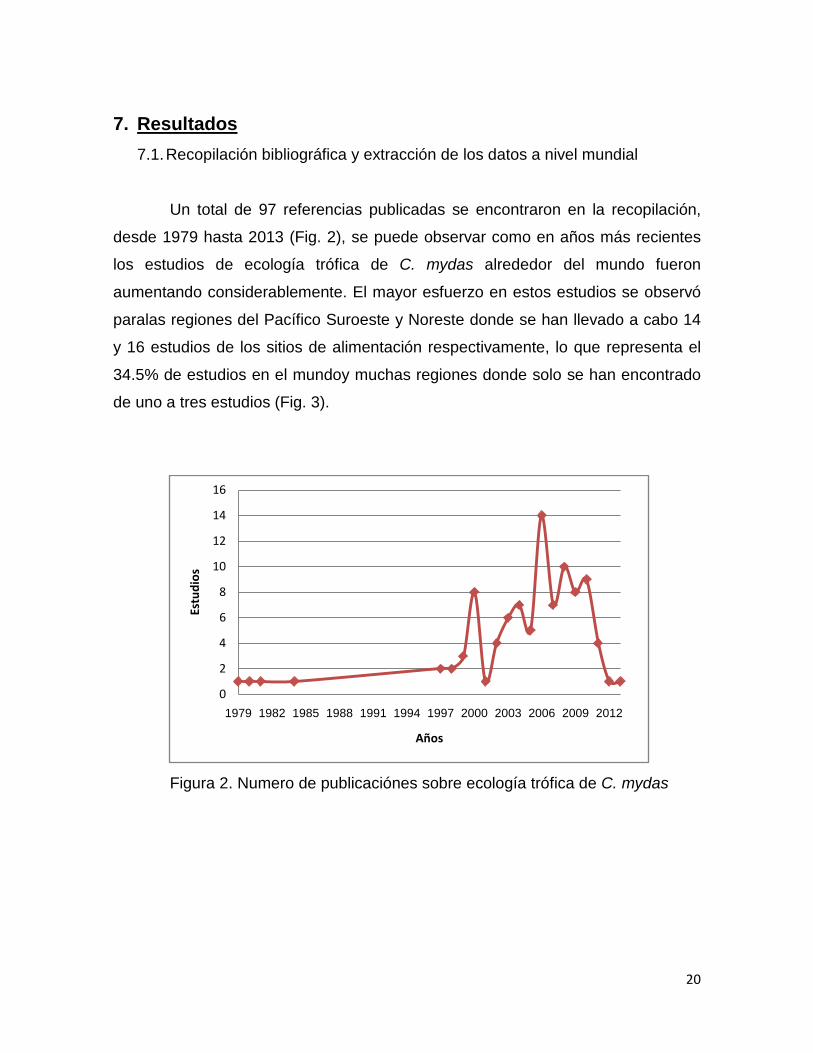
Un total de 97 referencias publicadas se encontraron en la recopilación,
desde 1979 hasta 2013 (Fig. 2), se puede observar como en años más recientes
los estudios de ecología trófica de C. mydas alrededor del mundo fueron
aumentando considerablemente. El mayor esfuerzo en estos estudios se observó
paralas regiones del Pacífico Suroeste y Noreste donde se han llevado a cabo 14
y 16 estudios de los sitios de alimentación respectivamente, lo que representa el
34.5% de estudios en el mundoy muchas regiones donde solo se han encontrado
de uno a tres estudios (Fig. 3).
Figura 2. Numero de publicaciónes sobre ecología trófica de C. mydas
0
2
4
6
8
10
12
14
16
1979 1982 1985 1988 1991 1994 1997 2000 2003 2006 2009 2012
Estu
dios
Años

21
Figura 3. Mapa de los países con estudios de zonas de alimentación y el número de publicaciones que se
encontraron en la revisión.1 – 3 ; 4 – 6 ; 7 – 9 ; 10 – 12 ; + 13 ; zonas de anidación de mayor importancia ; otros registros de anidación (Modificado de IUCN, 2003)
22
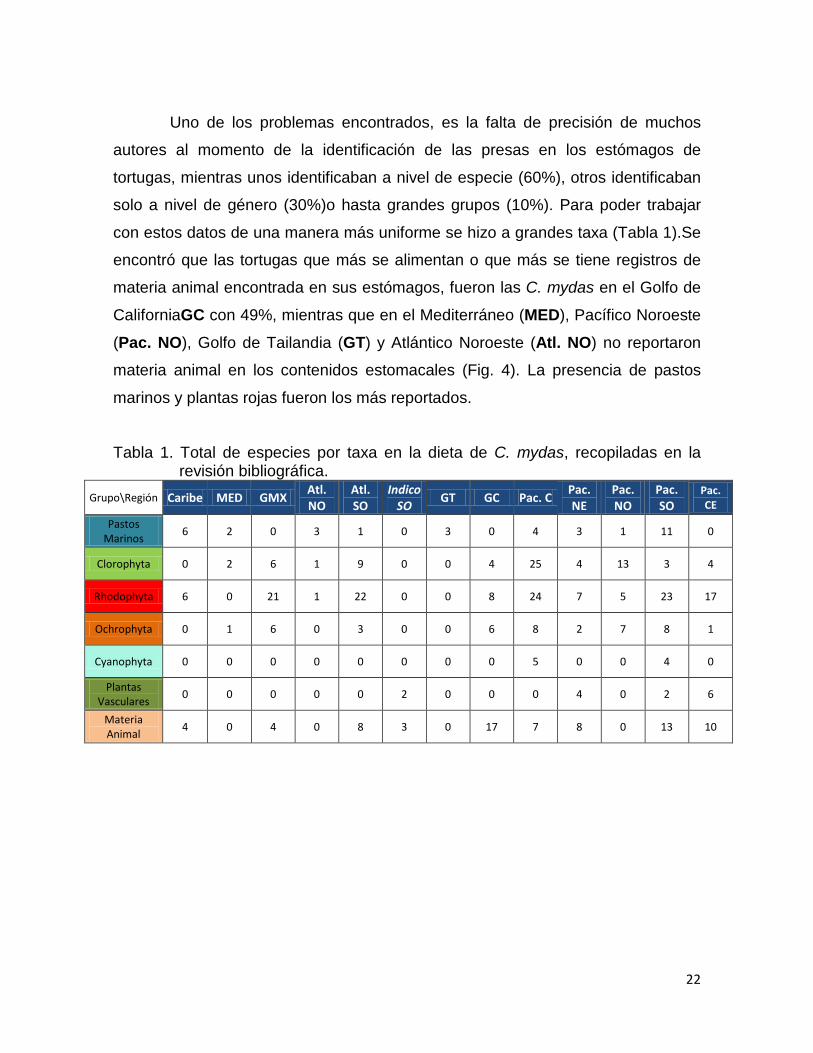
Uno de los problemas encontrados, es la falta de precisión de muchos
autores al momento de la identificación de las presas en los estómagos de
tortugas, mientras unos identificaban a nivel de especie (60%), otros identificaban
solo a nivel de género (30%)o hasta grandes grupos (10%). Para poder trabajar
con estos datos de una manera más uniforme se hizo a grandes taxa (Tabla 1).Se
encontró que las tortugas que más se alimentan o que más se tiene registros de
materia animal encontrada en sus estómagos, fueron las C. mydas en el Golfo de
CaliforniaGC con 49%, mientras que en el Mediterráneo (MED), Pacífico Noroeste
(Pac. NO), Golfo de Tailandia (GT) y Atlántico Noroeste (Atl. NO) no reportaron
materia animal en los contenidos estomacales (Fig. 4). La presencia de pastos
marinos y plantas rojas fueron los más reportados.
Tabla 1. Total de especies por taxa en la dieta de C. mydas, recopiladas en la revisión bibliográfica.
Grupo\Región Caribe MED GMX Atl. NO
Atl. SO
Indico SO
GT GC Pac. C Pac. NE
Pac. NO
Pac. SO
Pac. CE
Pastos Marinos
6 2 0 3 1 0 3 0 4 3 1 11 0
Clorophyta 0 2 6 1 9 0 0 4 25 4 13 3 4
Rhodophyta 6 0 21 1 22 0 0 8 24 7 5 23 17
Ochrophyta 0 1 6 0 3 0 0 6 8 2 7 8 1
Cyanophyta 0 0 0 0 0 0 0 0 5 0 0 4 0
Plantas Vasculares
0 0 0 0 0 2 0 0 0 4 0 2 6
Materia Animal
4 0 4 0 8 3 0 17 7 8 0 13 10

23
Figura 4. Total de especies reportadas en los contenidos estomacales de C.
mydas en las diferentes regiones que son: Pacífico Centroeste (Pac. CE), Pacífico Suroeste (Pac. SO), Pacífico Noroeste (Pac. NO), Pacífico Noreste (Pac. NE), Pacífico Central (Pac. C), Golfo de California (GC) Golfo de Tailandia (GT), Índico Suroeste (Indico SO), Atlántico Suroeste (Atl. SO), Atlántico Noroeste (Atl. NO), Golfo de México (GMX) Mediterráneo (MED) y el Caribe.
Debido a la robustez de los datos, los resultados de las pruebas a priori de
Homogeneidad de Varianzas de Cochran y Levene´s para los datos en el
Statistica 10 de la Tabla 1, resultaron ser significativos con una p<0.0001, por lo
cual se tuvieron que normalizar los datos mediante la siguiente formula
posteriormente, los datos cumplieron con los supuestos de normalidad y los
resultados del ANDEVA fueron significativos tanto como por Región como por
Grupo de especies con valores de p<0.05 mostrados en la Tabla 2. A su vez se
realizó el índice de similitud de Bray-Curtis, el cual nos muestra el grado de
similitud entre las diferentes regiones.
0 10 20 30 40 50 60 70 80
Caribe
GMX
Atl. SO
GT
Pac. C
Pac. NO
Pac. CE
No. de Especies
Regi
ones
de
Estu
dio
Pastos Marinos
Clorophyta
Rhodophyta
Ochrophyta
Cyanophita
Plantas Vasculares
Materia Animal

24
Tabla 2. Tabla ANDEVA dos vías. Gl SC CM F p
Región 12 218.6044 18.2170 3.48969 0.000439 Especies 6 216.5000 36.0833 6.91220 0.000008
Error 72 375.8571 5.2202 Total 90 810.9615
gl (grados de libertad), SC (Suma de cuadrados), CM (Cuadrado medio).
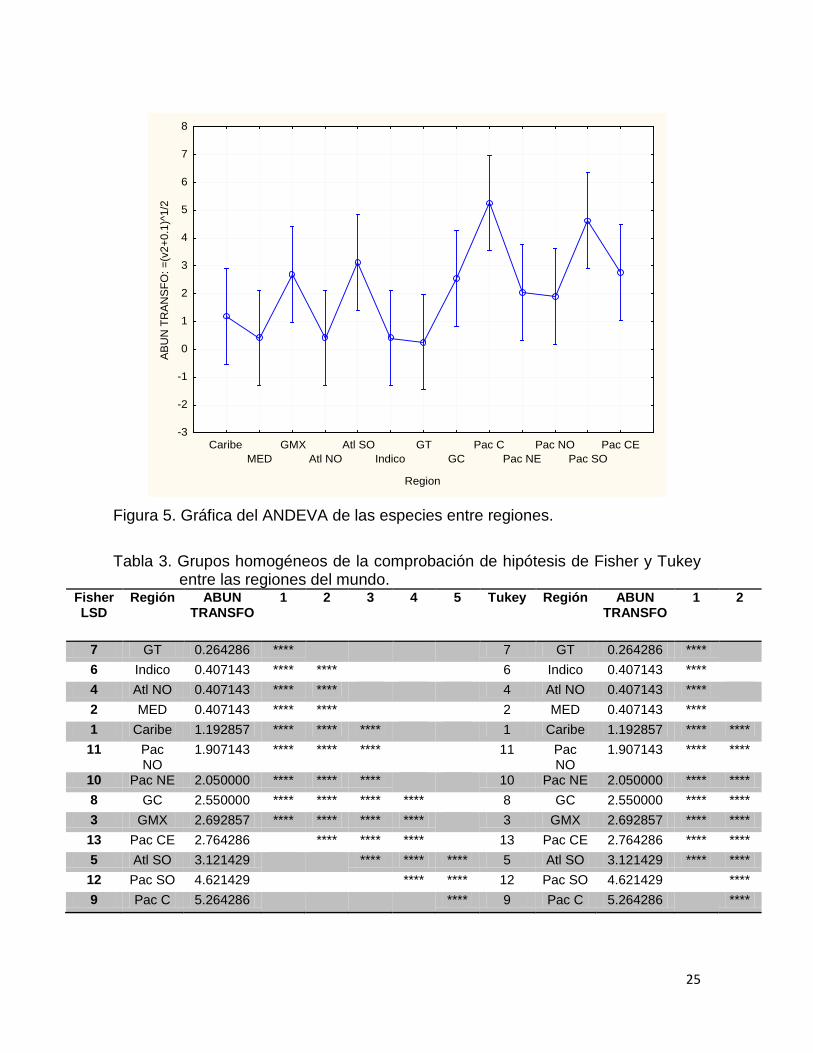
La gráfica del ANDEVA por Región (Fig. 5) con sus pruebas a posteriori de
grupos homogéneos de Tukey y de Fisher (Tabla 3) se usan para saber cuáles
son las regiones son diferentes después de que el ANDEVA resulta significativo.
Se muestran muchos traslapes entre las regiones, donde Pacífico Central,
Centroeste y Suroeste; y Atlántico Suroeste (Fig. 5) presentan los picos más altos.
La prueba de comprobación de hipótesis de Tukey, muestra solo dos grupos
homogéneos, mientras que la de Fisher presenta cinco, de los cual destacan
Pacífico Central y Suroeste; y Atlántico Suroeste como las regiones con más
especies descritas. Por otro lado el índice de similitud de Bray-Curtis, muestra el
parecido entre las regiones similar a la prueba de Tukey, sin embargo el índice de
similitud no solo se basa en comparar las cantidades de las abundancias si no la
composición de las especies y ver en qué regiones, la composición especifica
presenta mayor semejanza. Puede observarse un grupo totalmente aislado en la
parte izquierda de la figura en el cual se muestran el Mediterráneo, el Atlántico
Noroeste y el Golfo de Tailandia (Fig.6) que a su vez fueron las que tienen pocos
estudios y pocas especies registradas, por otro lado, las regiones con más
estudios y más especies descritas quedaron agrupadas, como el Golfo de
California, Pacífico Central, Noreste, Suroeste y Centroeste.
25
CaribeMED
GMXAtl NO
Atl SOIndico
GTGC
Pac CPac NE
Pac NOPac SO
Pac CE
Region
-3
-2
-1
0
1
2
3
4
5
6
7
8
ABU
N T
RAN
SFO
: =(v
2+0.
1)^1
/2
Figura 5. Gráfica del ANDEVA de las especies entre regiones.
Tabla 3. Grupos homogéneos de la comprobación de hipótesis de Fisher y Tukey
entre las regiones del mundo. Fisher LSD
Región ABUN TRANSFO
1 2 3 4 5 Tukey Región ABUN TRANSFO
1 2
7 GT 0.264286 **** 7 GT 0.264286 **** 6 Indico 0.407143 **** **** 6 Indico 0.407143 **** 4 Atl NO 0.407143 **** **** 4 Atl NO 0.407143 **** 2 MED 0.407143 **** **** 2 MED 0.407143 **** 1 Caribe 1.192857 **** **** **** 1 Caribe 1.192857 **** **** 11 Pac
NO 1.907143 **** **** **** 11 Pac
NO 1.907143 **** ****
10 Pac NE 2.050000 **** **** **** 10 Pac NE 2.050000 **** **** 8 GC 2.550000 **** **** **** **** 8 GC 2.550000 **** **** 3 GMX 2.692857 **** **** **** **** 3 GMX 2.692857 **** **** 13 Pac CE 2.764286 **** **** **** 13 Pac CE 2.764286 **** **** 5 Atl SO 3.121429 **** **** **** 5 Atl SO 3.121429 **** **** 12 Pac SO 4.621429 **** **** 12 Pac SO 4.621429 **** 9 Pac C 5.264286 **** 9 Pac C 5.264286 ****

26
Figura 6. Dendograma del Índice de Similitud de Bray-Curtis.
7.2. Biodiversidad y Valor Biológico
Los porcentajes de los contenidos estomacales encontrados por regiones
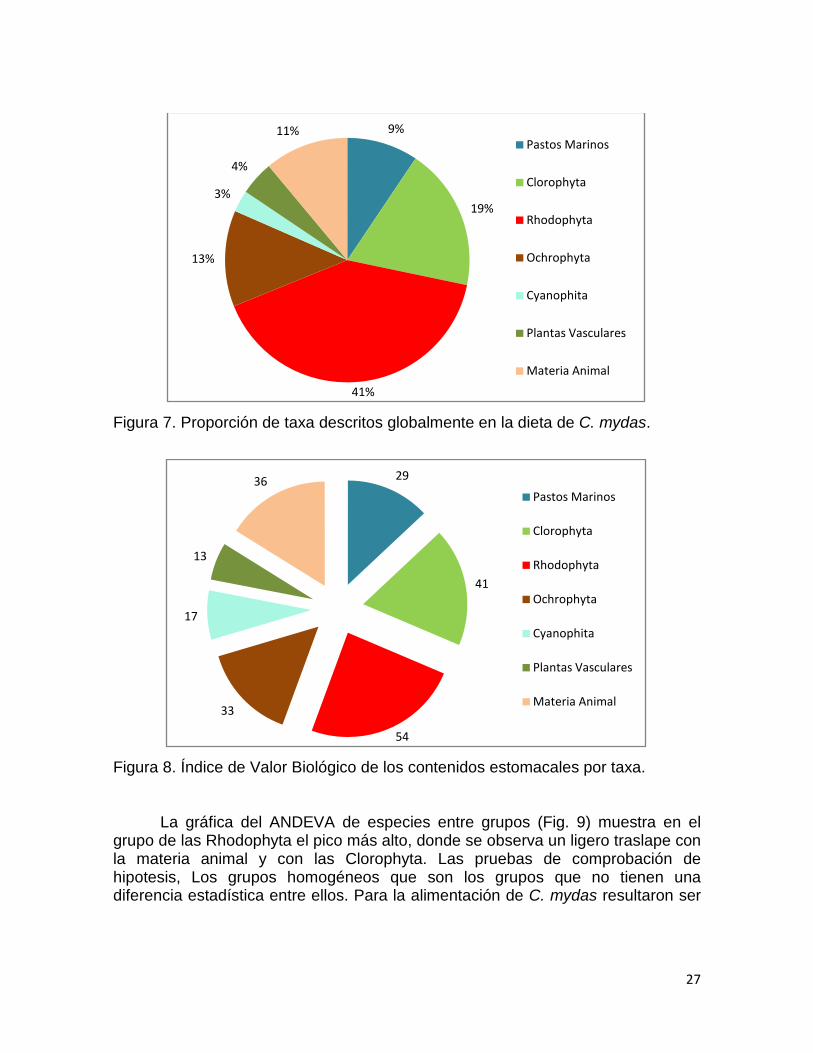
agrupadas (Fig. 7) mostró que el grupo predilecto de alimentación de C. mydasson
las algas rojas (Rhodophyta, 42%), el segundo lugar las algas verdes (Clorophyta,
19%), tercer lugar las algas cafés (Ochrophyta, 13%) y en cuarto lugar la materia
animal (11%) mientras que con el Índice de Valor Biológico (IVB, Fig. 8) las algas
rojas y verdes, se mantienen en primero y segundo lugar, mientras que la materia
animal sube a la tercera posición, en cuanto a la importancia de estas presas en la
alimentación de C. mydas.
27
Figura 7. Proporción de taxa descritos globalmente en la dieta de C. mydas.
Figura 8. Índice de Valor Biológico de los contenidos estomacales por taxa.
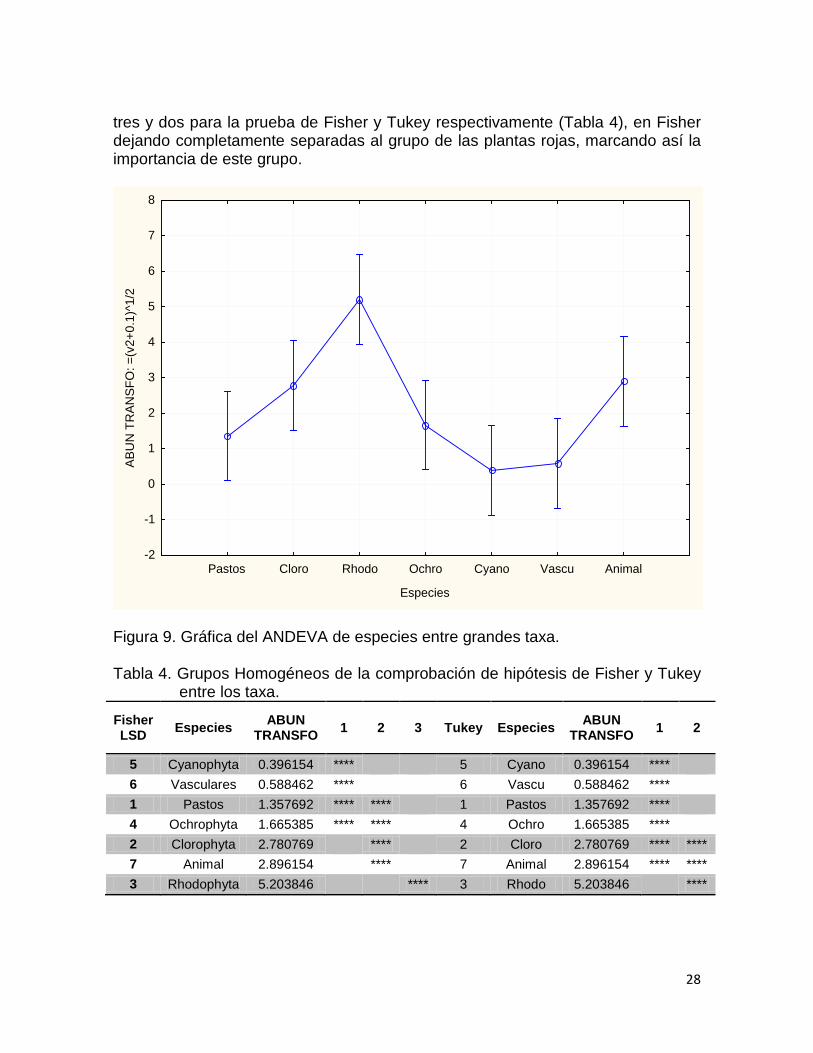
La gráfica del ANDEVA de especies entre grupos (Fig. 9) muestra en el grupo de las Rhodophyta el pico más alto, donde se observa un ligero traslape con la materia animal y con las Clorophyta. Las pruebas de comprobación de hipotesis, Los grupos homogéneos que son los grupos que no tienen una diferencia estadística entre ellos. Para la alimentación de C. mydas resultaron ser
9%
19%
41%
13%
3%
4%
11%Pastos Marinos
Clorophyta
Rhodophyta
Ochrophyta
Cyanophita
Plantas Vasculares
Materia Animal
29
41
54
33
17
13
36Pastos Marinos
Clorophyta
Rhodophyta
Ochrophyta
Cyanophita
Plantas Vasculares
Materia Animal
28
tres y dos para la prueba de Fisher y Tukey respectivamente (Tabla 4), en Fisher dejando completamente separadas al grupo de las plantas rojas, marcando así la importancia de este grupo.
Pastos Cloro Rhodo Ochro Cyano Vascu Animal
Especies
-2
-1
0
1
2
3
4
5
6
7
8
AB
UN
TR
AN
SFO
: =(v
2+0.
1)^1
/2
Figura 9. Gráfica del ANDEVA de especies entre grandes taxa. Tabla 4. Grupos Homogéneos de la comprobación de hipótesis de Fisher y Tukey
entre los taxa.
Fisher LSD Especies ABUN
TRANSFO 1 2 3 Tukey Especies ABUN TRANSFO 1 2
5 Cyanophyta 0.396154 **** 5 Cyano 0.396154 **** 6 Vasculares 0.588462 **** 6 Vascu 0.588462 **** 1 Pastos 1.357692 **** **** 1 Pastos 1.357692 **** 4 Ochrophyta 1.665385 **** **** 4 Ochro 1.665385 **** 2 Clorophyta 2.780769 **** 2 Cloro 2.780769 **** **** 7 Animal 2.896154 **** 7 Animal 2.896154 **** **** 3 Rhodophyta 5.203846 **** 3 Rhodo 5.203846 ****

29
La mayor riqueza de grupos encontrados fue en el Pacífico Suroeste (Pac. SO), con siete grupos, seguido por el Pacífico Noreste (Pac. NE) y Central (Pac. C) seis grupos reportados, esto concuerda con lo encontrado por la dde Margalef
(Tabla 5), pero su corrección por número de muestra, modificó las jerarquías. Por
otro lado la uniformidad de Pielou (J´), tiende a 1 lo que significa que los grupos
son homogéneos en cuanto a su distribución en cada región. El Índice de
Shannon-Wiener(H´) muestra las regiones con más diversidad igual a las de
riqueza, ya que son donde más diversidad de grupos se ha encontrado.
Tabla 5. Indicadores de diversidad de taxa para las diferentes regiones. Región s n d J´ H´(loge) Caribe 3 16 0.721 0.9851 1.082 MED 3 5 1.243 0.9602 1.055 GMX 4 37 0.831 0.831 1.152
Atl. NO 3 5 1.243 0.865 0.9503 Atl. SO 5 43 1.063 0.7806 1.256 Indico
SO 2 5 0.621 0.971 0.673
GC 4 35 0.844 0.8933 1.238 Pac. C 6 73 1.165 0.8609 1.543
Pac. NE 6 28 1.501 0.9423 1.688 Pac. NO 4 26 0.921 0.8239 1.142 Pac. SO 7 64 1.443 0.8629 1.679 Pac. CE 5 38 1.1 0.8297 1.335
Número de especies (s), número de individuos (n), riqueza de Margalef (d), Uniformidad de Pielou (J´), Índice de Shannon-Wiener (H´(loge)).
7.3. Caso de Estudio Bahía Magdalena: TMB e Ingesta calculada de C. mydas.
De todas las presas encontradas en la literatura, las más predominantes
fueron las que se encuentran en la Tabla 6. A estos se les priorizó la búsqueda de
los contenidos energéticos. Dadalos escases de información acerca de los
contenidos energéticos de todas las especies de la tabla 6, se tuvieron que
cambiar por los representantes con la mayor información posible, siendo los datos
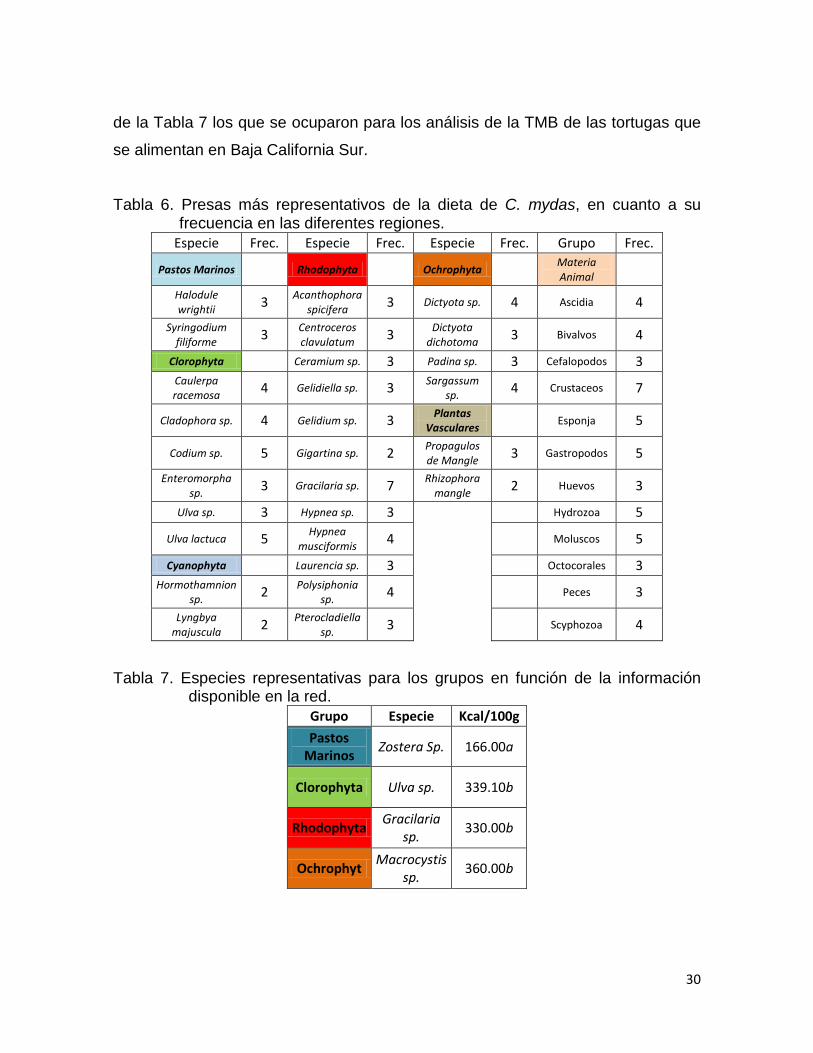
30
de la Tabla 7 los que se ocuparon para los análisis de la TMB de las tortugas que
se alimentan en Baja California Sur.
Tabla 6. Presas más representativos de la dieta de C. mydas, en cuanto a su frecuencia en las diferentes regiones.
Especie Frec. Especie Frec. Especie Frec. Grupo Frec.
Pastos Marinos
Rhodophyta
Ochrophyta
Materia Animal
Halodule wrightii 3 Acanthophora
spicifera 3 Dictyota sp. 4 Ascidia 4
Syringodium filiforme 3 Centroceros
clavulatum 3 Dictyota dichotoma 3 Bivalvos 4
Clorophyta
Ceramium sp. 3 Padina sp. 3 Cefalopodos 3 Caulerpa racemosa 4 Gelidiella sp. 3 Sargassum
sp. 4 Crustaceos 7
Cladophora sp. 4 Gelidium sp. 3 Plantas Vasculares
Esponja 5
Codium sp. 5 Gigartina sp. 2 Propagulos de Mangle 3 Gastropodos 5
Enteromorpha sp. 3 Gracilaria sp. 7 Rhizophora
mangle 2 Huevos 3
Ulva sp. 3 Hypnea sp. 3
Hydrozoa 5
Ulva lactuca 5 Hypnea musciformis 4
Moluscos 5
Cyanophyta
Laurencia sp. 3
Octocorales 3 Hormothamnion
sp. 2 Polysiphonia sp. 4
Peces 3
Lyngbya majuscula 2 Pterocladiella
sp. 3
Scyphozoa 4
Tabla 7. Especies representativas para los grupos en función de la información disponible en la red.
Grupo Especie Kcal/100g
Pastos Marinos
Zostera Sp. 166.00a
Clorophyta Ulva sp. 339.10b
Rhodophyta Gracilaria
sp. 330.00b
Ochrophyt Macrocystis
sp. 360.00b
31
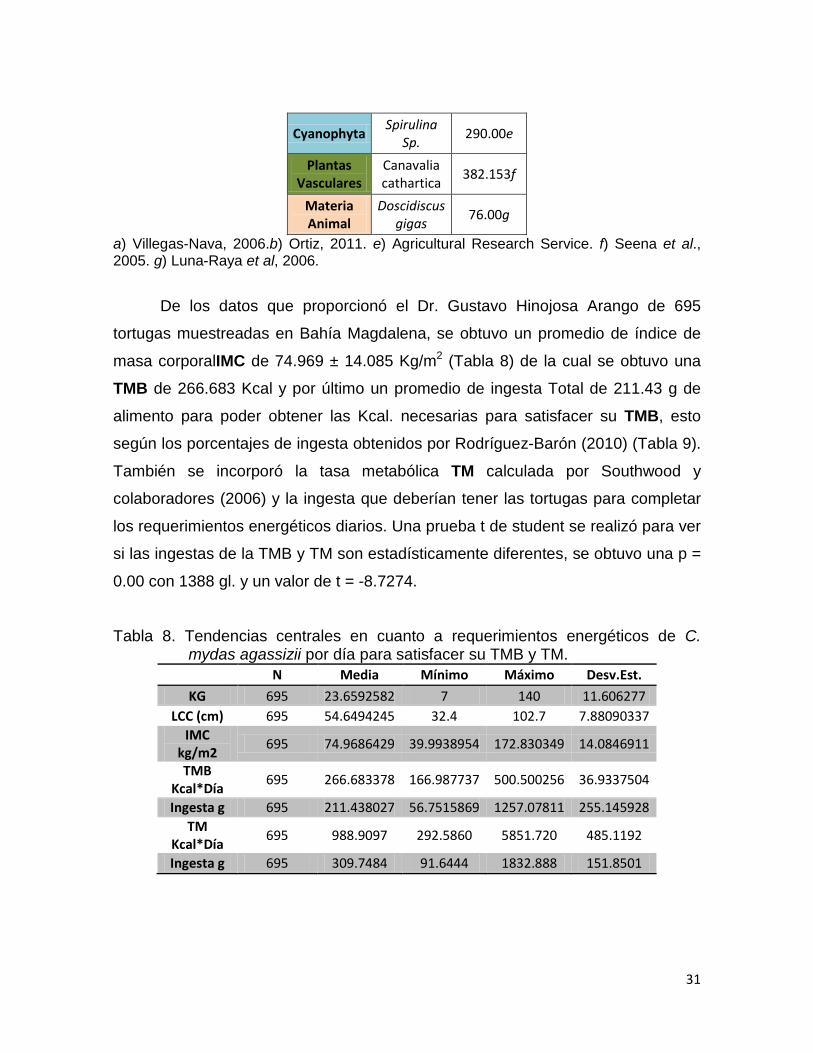
Cyanophyta Spirulina
Sp. 290.00e
Plantas Vasculares
Canavalia cathartica
382.153f
Materia Animal
Doscidiscus gigas
76.00g
a) Villegas-Nava, 2006.b) Ortiz, 2011. e) Agricultural Research Service. f) Seena et al., 2005. g) Luna-Raya et al, 2006.
De los datos que proporcionó el Dr. Gustavo Hinojosa Arango de 695
tortugas muestreadas en Bahía Magdalena, se obtuvo un promedio de índice de
masa corporalIMC de 74.969 ± 14.085 Kg/m2 (Tabla 8) de la cual se obtuvo una
TMB de 266.683 Kcal y por último un promedio de ingesta Total de 211.43 g de
alimento para poder obtener las Kcal. necesarias para satisfacer su TMB, esto
según los porcentajes de ingesta obtenidos por Rodríguez-Barón (2010) (Tabla 9).
También se incorporó la tasa metabólica TM calculada por Southwood y
colaboradores (2006) y la ingesta que deberían tener las tortugas para completar
los requerimientos energéticos diarios. Una prueba t de student se realizó para ver
si las ingestas de la TMB y TM son estadísticamente diferentes, se obtuvo una p =
0.00 con 1388 gl. y un valor de t = -8.7274.
Tabla 8. Tendencias centrales en cuanto a requerimientos energéticos de C. mydas agassizii por día para satisfacer su TMB y TM.
N Media Mínimo Máximo Desv.Est.
KG 695 23.6592582 7 140 11.606277 LCC (cm) 695 54.6494245 32.4 102.7 7.88090337
IMC kg/m2
695 74.9686429 39.9938954 172.830349 14.0846911
TMB Kcal*Día
695 266.683378 166.987737 500.500256 36.9337504
Ingesta g 695 211.438027 56.7515869 1257.07811 255.145928 TM
Kcal*Día 695 988.9097 292.5860 5851.720 485.1192
Ingesta g 695 309.7484 91.6444 1832.888 151.8501

32
Tabla 9. Volúmenes de ingesta de grupos.
Grupo % Ingesta
Rhodophyta 86.66% Clorophyta 4.70%
Pastos 5.16% Mangle 1.90% Materia Animal 2.00%
En la Tabla 10 se observan las regiones con más estudios, cada una con
los valores porcentuales de los volúmenes reportados en los distintos artículos.
Cabe mencionar que los grupos que estuvieron presentes en todos los estudios
fueron las Rhodophyta y Clorophyta, seguido por la materia animal. Los mangles
solo se reportaron en una sola región.
Tabla 10. Valores porcentuales de Ingesta aproximados de grupos por región.
Región/ Grupo Caribe GMX Atl. NO Atl. SO GC Pac. C Pac. NE Pac. SO Pac. CE
Pastos Marinos 41.47% 2.10% 74% 5.03%
Clorophyta 28.12% 8.19% 16% 43.98% 2.5% 11.60% 3% 1.60% 43.31% Rhodophyt
a 18.23% 67.15% 83% 48.83% 90.4% 78% 89% 19.23% 33.13%
Ochrophyta 12.07% 10.26% 1.70% 5.83%
Plantas Vasculares 1.45%
Materia Animal 2.16% 5.8 0.90% 4% 8.75%
8. Discusión 8.1. Situación actualde las zonas de alimentación de C. mydas
A partir de 1960 se dio la caída de las poblaciones de tortugas marinas
debido a la explotación de sus pieles (Márquez et al., 1982b) y más de 30 años
han pasado desde que se les decretó como especies amenazadas y la necesidad

33
de que sus poblaciones sean protegidas y recuperadas (Hamann et al., 2010).
Desde entonces ha sido de gran interés realizar investigaciones relacionadas con
su biología y conservación, interrelaciones con el medio físico y biológico en el que
estas se desarrollan y sobre todo de los impactos humanos y las dimensiones
asociadas a estas especies (Hamann et al., 2010).
Se encontraron97 artículos de uso de hábitat de alimentación y ecología
trófica de C. mydas, desde el año de 1979 hasta el 2013, con un promedio de 1.5
artículos al año entre 1979 y 1999, aumentando considerablemente en el 2000
hasta un promedio de 6 artículos al año. El problema es que toda vía falta
muchísima información de otros sitios donde se alimentan que han sido muy poco
estudiados (fig. 2.), y sobre todo por la dinámica en la presencia de especies
invasoras, las que afecta a su vez la composición en la dieta de C. mydas y otras
tortugas marinas.
En general, la mayoría los artículos, reportan una dieta principalmente
herbívora, donde las algas rojas y verdes son las que más preferencia tienen en la
dieta de C. mydas(NOAA, 1998; Arthur et al., 2009; Cardona et al., 2010;
Rodríguez-Barón, 2010; Rodriguez-Awabdi, 2013) debido a que presenta más
especies descritas (Tabla 1 y Fig. 3) y mayor abundancia en la mayoría de los
artículos.Rodríguez-Barón (2010) menciona la presencia y abundancia constante
de las RhodophytaGracilaria vermiculophylla y G. textorii a lo largo del año, y
obtiene un resultado que concuerda con López-Mendilaharsu y colaboradores
(2005; 2008), y explican que el género Gracilaria, fue el más representativo de las
Rhodophyta presentes en siete zonas del mundo (Tabla 6).
La importancia de las Rhodophyta se mencionan en estudios de López-
Mendilaharsu y colaboradores (2003, 2005, 2008), Villegas-Nava (2006) y
Rodríguez-Barón (2010)y se comprueba al momento de realizar el IVB que es
usado para conocer la importancia de especies de una comunidad mediante
puntajes (Sanders, 1960) (Loya-Salinas y Escofet, 1990) y acompañado de un
ANDEVA en el cual las pruebas de hipótesis de Fisher respalda la importancia de

34
las algas rojas en la dieta de la tortuga prieta y haciendo un segundo grupo
importante compuesto por pastos marinos, algas cafés, verdes y materia animal.
Esta última obtuvo su importancia de 2do lugar en el IVB (Fig. 8) debido a la gran
cantidad de especímenes identificados en los estudios de la Península de B.C.S.
(Fig. 3) por Seminoff y colaboradores (1999, 2002) y Rodríguez-Barón (2010)
sobre todo en Bahía Magdalena.
Por otro lado la presencia de algas consideradas como invasoras
reportadas en los contenidos estomacales de las tortugas son constantes temporal
y espacialmente, sobre todo en los lugares que se tienen registros de la presencia
de éstas, como es el caso del alga roja Acanthophora spicifera reportada en
diversos estudios en Hawaii, Saipan y Australia (NOAA, 1998; Russell y Balazs,
2000; Kolinskiet al., 2001; Seaborn et al., 2005; Fuentes et al., 2006; McDermind
et al., 2007; Arthur y Balazs, 2008), lo que indica que la tortuga se alimenta
dependiendo de la disponibilidad de alimento y que no es selectiva al momento de
seleccionar su presa (Hatase et al., 2005). Esto también se observa con algunos
reportes de plantas vasculares principalmente terrestres del género Ficus, lo que
significa nuevamente que C. mydas no presenta una selección específica de sus
presas.
No obstante enartículosmuy detallados en los contenidos estomacales y
en la guía de identificación de las presas de tortugas verdes en Hawái, se muestra
la presencia de algas verdeazules (Cyanophyta)(Russell y Balazs, 2000; Limpus et
al., 2004) y de substrato (arenas y pequeñas rocas), las cuales no entran en la
dieta de la tortuga de manera directa, si no como un acompañamiento del forrajeo
y que este detalle solo se observó una vez y fue en una tortuga de recién
reclutamiento (Limpus et al., 2004), sin embargo (Russell y Balazs, 2000) tiene
registradas las Cyanophyta en su guía de alimentación en Hawái, ya que al ser
devoradas, aportan nutrientes y calorías a la ingestaC. mydas, por ese motivo se
incluyó en la Tabla 7 los valores energéticos de las especies de Cyanophyta.

35
Cabe mencionar que a pesar de los estudios realizados, el ambiente se
encuentra en constante cambio, sobre todo ahora más rápidos provocados por el
hombre lo que originaque los animales también se vean obligados a modificar sus
tendencias tróficas, como es el caso de la tortuga carey (Eretmochelys imbricata),
mundialmente conocida como consumidora de esponjas, en la gran barrera de
arrecife (Australia) se demostró con lavados estomacales, un cambio a una
tendencia casi completamente herbívora (Bell, 2012) y que a su vez, las tortugas
verdes, se encuentran alimentándose de pulpos venenosos, nunca antes visto en
las costas Australianas (Townsend, et al., 2011).
Los indicadores de biodiversidad de Shannon-Weinerrevelan una alta
diversidad de especies encontradas principalmente en las regiones del Pacífico,
teniendo como la más diversa la del Noreste, esto debido a la gran aportación de
Rodríguez-Barón (2010) ya que al usar isótopos estables, logro identificar una
mayor cantidad de organismos, sobre todos los de materia animal. Por otro lado,
en las demás regiones han sido identificados con un mayor número de estudios. El
índice de uniformidad Pielou, no resultó similar al de Biodiversidad debido a la
robustez de los datos, ya que el tamaño de especies para el análisis disminuyó
considerablemente al momento de agrupar todas las especies en los siete grandes
grupos y lo que se demostró sobre cuales regiones eran las más importantes, se
confirmó con el ANDEVA y sus efectos por regiones (Tabla 3 y 4, Fig. 6),
denotando las regiones más importantes de acuerdo a la cantidad de especies
reportadas, Atlántico Suroeste, Pacífico Suroeste y Pacífico Central, que a su vez
resultaron ser de las zonas con más estudios (Fig. 2).
El índice de similitud de Bray-Curtis (Fig.6) nos permite identificar dos
grandes grupos, uno conformado por el Pacífico Central (Hawái) y el Suroeste
(Australia), ya que estos son los sitios con más especies identificadas y las cuales
comparte muchas especies incluyendo la especie invasora Acantophora
spiciferareportada en los lavados esofágicos de las tortugas, debido a que no son
selectivas en su alimento, al momento que se encuentra con una mayor

36
disponibilidad (NOAA, 1998). El segundo grupo se conforma por el Golfo de
California, Pacífico Noreste y Centro Este, los primeros, forman parte de la
península de Baja California y la región centro este comprende Colombia, Ecuador
yArchipiélago Galápagos. La diferencia entre estos dos grandes grupos es la
lejanía y barreras geográficas que presentan cada uno, debido a que Hawái y
Australia, se encuentran en lugares muy distantes separados por corrientes y
barreras geográficas naturales (Dutton et al., 2008), como para que compartan
tantas especies, inclusive las invasoras, esto se puede deber a un alto tránsito de
embarcaciones entre estas regiones. Por otro lado, el segundo grupo está formado
por regiones contiguas en una línea de costa en el continente americano, que
podría fungir como un corredor o ruta migratoria entre las zonas de anidación
tropicales y las distantes zonas de alimentación al norte del Pacífico Este Tropical
(Bolker et al., 2007).
No obstante, las colonias de anidación de C. mydas en el Pacífico Este
Tropical, se encuentran diferenciadas genéticamente (Dutton et al., 2008), por lo
que actualmente ya no es problema saber de qué colonia provienen los
organismos debido a estudios de Bolker (2007), Dutton (2008), Amorocho (2012) y
sus respectivos colaboradores en sus análisis de Stocks Mezclados, han logrado
con la región control mitocondrial, establecer la procedencia de las tortugas
presentes en los sitios de alimentación y a su vez demuestran que en una
determinada zona de forrajeo, las tortugas provienen de diferentes colonias de
anidación.
Por último se sabe que los neonatos de C. mydas en su etapa oceánica se
alimentan de materia animal, debido a algunas capturas esporádicas (NOAA,
1998), es así que existe un hueco clave en la información sobre los hábitos
alimenticios de juveniles de C. mydas en esa etapa, debido a que existen tres
grandes problemas para la obtención de datos que menciona la NOAA (1998), los
cuales son: el paradero de los neonatos en el océano es desconocido, solo se
conoce que generalmente se encuentran cerca de la superficie en los giros

37
oceánicos y a lo largo de las líneas de deriva. El segundo es la elaboración de una
técnica eficiente para obtener muestras representativas y finalmente, un medio
para determinar el origen de esas tortugas.
8.2. Importancia y Conservación de las tortugas marinas en Baja California Sur
En Baja California Sur anidan 4 especies de tortugas marinas, la tortuga
verde/prieta (Chelonia mydas), laúd (Dermochelys coriacea), carey (Eretmochelys
imbricata) y la golfina (Lepidochelys olivacea) (Arizpe et al., 2012). En la región de
los Cabos, el cuidado de los nidos y la liberación de neonatos por parte de los
hoteles, atrae cientos de turistas a esta región por lo que López-Castro y
colaboradores (2012), las mencionan como embajadoras y especies carismáticas
de la región.
Los esfuerzos de conservación y las leyes en Baja California Sur y en todo
México, son en su mayoría dirigidos al cuidado de los nidos, como en la mayor
parte del territorio mexicano (NOM-162), ya que es más fácil y menos costoso,
cuidar a las tortugas en tierra que en el mar (López-Castro et al., 2012). Debido a
esto, la conservación en su hábitat acuático recibe menos investigación. Sin
embargo si los neonatos no sobreviven el tiempo necesario para dejar
descendencia, el esfuerzo del cuidado de los nidos es insignificante (Wallace et
al., 2008). Por esto no debemos menospreciar lo que pasa en los sitios de
alimentación, si las tortugas viven o mueren, se verá reflejado en los sitios de
anidación (Bolker et al., 2007).
En Baja California Sur, debido a su gran extensión en mantos de
macroalgas y pastos marinos, recibe miles de tortugas, sobre todo C. mydas, en
sus sitios de alimentación, los cuales podrían estar albergando a tortugas
provenientes de diferentes sitios de anidación y alimentación. Si no se conservan
en estos lugares de suma importancia, se podría estar diezmando las poblaciones
de tortuga a nivel mundial.

38
Por lo que se requiere la identificación y establecimiento de sistemas de
áreas para la conservación y mediante una planeación sistemática para la
conservación, se minimicen los impactos y riesgo de extinción de las especies de
tortugas marinas y sus hábitats, a las cuales se les dé prioridad para la asignación
de recursos económicos y de investigación.
Margules y Sarkar (2009) mencionan que el concepto clave para esto, es
la complementariedad, por eso nos referimos a “Tortugas marinas y sus Hábitats”,
la cual consiste en proteger especies, comunidades y los procesos ecológicos que
se lleven a cabo.
8.3. Tasa Metabólica Basal de C, mydas agassizii en Bahía Magdalena
Los nutrientes son componentes básicos encontrados en el alimento,
útiles para el funcionamiento del organismo.Los alimentos difieren entre sí por las
cantidades y porciones del tipo de alimento y de los nutrientes que contiene
(Villegas-Nava, 2006) aparte otra serie de factores determinan la porción de los
nutrientes que son asimilados por la tortuga, como pueden ser tanto físicos como
de demanda energética. Dentro de los factores físicos interviene edad, sexo, el
potencial genético, estado fisiológico, peso corporal, fisonomía, condición corporal,
entre otros (Shimada, 2005).
La tortuga al estar en un lugar con una gran disponibilidad de alimento,
podríanelegir aquel de mejor calidad en cuanto a la cantidad de nutrientes y dejar
de lado aquellos de menor calidad, para tener así un forrajeo óptimo, el cual
menciona que el depredador va a mostrar preferencia por aquellas presas que
presenten un mayor aporte de nutrientes y un aporte de energía; al contrario que
si se encuentra en un lugar con poca diversidad y cantidad de alimento, se vuelve
menos selectivo en sus presas (MacArthur y Pianka, 1966). Estudios sobre el
metabolismo de las tortugas marinas se han reportado para las actividades de
buceo (Southwood et al., 2003; Hazel et al., 2009), los cuales mencionan que C.

39
mydas tiende a alimentarse en zonas muy someras con profundidades no mayor a
los 3 metros, que es una profundidad ideal debido a que las macroalgas tienen un
mejor crecimiento y abundancia en profundidades cercanas a 1 m (Konar et al.,
2010).
Las tortugas no presentan una variación metabólica estacional significativa
en sus zonas de alimentación, donde la temperatura del agua oscila alrededor de
los 20 y 28°C (Southwood et al., 2006), parecidas a las temperaturas en latitudes
medianas, donde zonas de surgencias permiten el aumento de biomasa y
diversidad de plantas marinas necesarias para el consumo de C. mydas.Esto se
puede observar en el mapa de las zonas de alimentación (Fig. 3), donde se
aprecia que la mayoría se encuentran en latitudes medianas y medianamente
altas lo que concuerda con Konary colaboradores (2010).
Debido a la repetida presencia de algunas especies en los contenidos
estomacales se considera que la tortuga prieta tiene algún tipo de selección
preferencial por sus presas, es decir que proponen la selectividad de la tortuga
prieta en el área de estudio pero no la corroboran con datos nutricionales o
energéticos. Estas selecciones se pueden ver a nivel de grupo de alimentación,
donde las plantas rojas son las más consumidas en casi todos los sitios de estudio
(Hilbert et al., 2005; López-Mendilaharsu et al., 2005).
Villegas-Nava (2006) presenta altos valores energéticos para las
macroalgas y pastos marinos que concuerdan con lo obtenido por Kaehler y
Kennish (1996), Castro-González ycolaboradores (1996) y Ortiz (2011), por lo que
la tortuga no requiere de una ingesta de gran volumen y su complementariedad
con la materia animal, es solo en casos de escases de alimento, sobre todo en
zonas de anidación, ya que por el alto oleaje en la mayoría de estas, las
densidades de pastos marinos y macro algas es muy baja (NOAA, 1998).
Las tendencias centrales de requerimiento energético de la TMB de C.
mydas agassizii oscilan entre 266.68 ± 36.93 kcal por día, esta cantidad depende
del IMC de la tortuga,como lo mencionan Hulber y Else (2003) la TMB tiene una

40
relación alométrica con el IMC de los individuos, que en este caso el promedio es
de 74.96 ± 14.08 Kg/m2, por lo tanto una tortuga con estas dimensiones tendría
que consumir 211.43g del cual el 87% serían macroalgas, 5% pastos marinos, 5%
Clorophyta, 2% mangle y 2% de materia animal según las cantidades de alimento
reportadas en los contenidos estomacales de los estudios de Rodríguez-Barón
(2010) para el área de Bahía Magdalena.Southwoody colaboradores(2006)
mediante el método de agua doblemente marcada (Lifson and McClintock, 1966)
calcularon la Tasa Metabólica de C. mydas juveniles en Verano e Invierno, en las
cuales no hubo diferencia significativa estacional. Con la TM de Southwood y
colaboradores (2006)se realizó la conversión pertinente y resulta 41.79
Kcal*Kg*díaque al aplicar a los datos proporcionados por el Dr. Gustavo Hinojosa
Arango, obtenemos una TM media de 988.9 Kcal*día, ahora comparable con
nuestra TMB obtenida.Incluir la TM de Southwood y colaboradores (2006), al
compararla con la TMB calculada y a pesar de tener una dispersión muy grandelas
cantidades de ingesta para satisfacer la TMB y la TMson estadísticamente
diferentes, lo que podría indicar que se realizó un buen cálculo aproximado de la
TMB.
Con los cálculos de TM y TMBdemostramos que se pueden calcular las
cantidades de alimento que requiere C. mydas para poder cubrir sus
requerimientos energéticos y en dado caso que se llegue a tener una tortuga
dañada o enferma en cautiverio, podremos suministrar el alimento necesario para
mantenerla.
Por último, la cantidad de alimento aproximada que consume C. mydas
agassizii en Bahía Magdalena, no fue posible calcularla debido a que las medias
del volumen tomado de los lavados estomacales, varían de 5 – 200 ml (Seminoff
et al., 2002) y de 1 – 80.5 ml (Lopez-Mendilaharsu et al., 2008). Amorocho y Reina
(2007) mencionan que esto sucede debido a que no se puede “cosechar” los
contenidos directos del estómago, ya que C. mydas agassizii, a diferencia de C.

41
mydas, presenta en el esófago una curva en forma de “S” antes de unirse con el
estómago y que se intenta la cosecha del estómago, podría dañar a la tortuga.
9. Conclusiones
• Las zonas de alimentación de C. mydas se encuentran regularmente entre
latitudes medianas y medianamente altas que varían entre los 25 y 60°, debido
a que aparecen las zonas de surgencias que ayudan al aumento de la
biomasa y diversidad de plantas marinas de las cuales se alimentan.
• El grupo de alimentación de C. mydas agassizii más importante, fueron las
plantas rojas (Rhodophyta) debido a que se encontró en todas las zonas y con
un mayor número de especies y cantidades, mientras que la especie
representativa de las Rhodophyta fue el género Gracilaria.
• La materia animal pese a su bajo consumo, brinda un buen aporte energético
para C. mydas por lo que podría hacer más exitosa las poblaciones que están
consumiendo, sobre todo en zonas con baja abundancia de plantas marinas.
• El cálculo de la TMB calculada en esta Investigación, resulta confiable para
realizar estimaciones de una Tasa Metabólica y por ende de consumos de
energía comparado con lo obtenido por Southwood et al., (2006).

42
10. Bibliografía
Ackerman, R. 1997. The Nest Environment and the Embryonic Development of
Sea Turtles, en The Biology of Sea Turtles.In Lutz, P.L., J.A. Musick, and
J. Wyneken (Eds.).CRC Press, 88p
Alvarado, J. y Figueroa, A. 1991. Comportamiento Reproductivo de la Tortuga
Prieta Chelonia agassizii.Ciencia y Desarrollo. 17 (98): 1-6
Agricultural Research Service. Consultado Abril 2013. National Nutrient Database
for Standard Reference. Nutrient Data Laboratory.
Amorocho, D. y Reina, R. 2007.Feeding ecology of the East Pacific green sea
turtle Chelonia mydas agassizii at Gorgona National Park,
Colombia.Endangered species research. P. 43-51.
Amorocho, D.F. y Reina, R.D. 2008. Intake passage time, digesta composition
and digestibility in East Pacific green turtles (Chelonia mydas agassizii) at
Gorgona National Park, Colombian Pacific. Journal of Experimental Marine
Biology and Ecology 360:117-124.
Amorocho, D., Abreu-Grobois, A., Dutton, P., Reina, R. 2012 Multiple Distant
Origins for Green Sea Turtles Aggregating off Gorgona Island in the
Colombian Eastern Pacific. Colombia. PlosOne V:7 Issue 2 e3148
Arizpe O., Riosmena-Rodríguez R. y Valdez N. 2012.In Los Cabos Prospectiva de
un Paraíso Turístico. Cap. 4 Flora y Fauna Marina y Costera. San Diego
State University Press. 533 pp.
Arthur, K.E., Limpus, C.J., Roelfsema, C.M.,Udy, J.W. y Shaw, G.R. 2006. A bloom
of Lyngbya majuscula in Shoalwater Bay, Queensland, Australia: An

43
important feeding ground for the green turtle (Chelonia mydas). Harm
Algae 5(3):251-265.
Arthur K. y Balazs G.H. 2008. A comparison of immature green turlte’s (Chelonia
mydas) diets among seven sites in the main Hawaiian Islands. Pacific
Science 62, 205–217.
Arthur, K.E., O’Neil, J., Limpus, C.J., Abernathy, K. y Marshall, G. 2008. Using
Animal-Borne Imaging to Assess Green Turtle (Chelonia mydas) Foraging
Ecology in Moreton Bay, Australia.Marine Technology Society Journal.
Arthur, K.E., Boyle, M. y Limpus, C. 2008. Ontogenetic changes in diet and habitat
use in green sea turtle (Chelonia mydas) life history.Marine Ecology
Progress Series. P. 303-311.
Arthur, K., McMahon, K., Limpus, C. y Dennison, W. 2009. Feeding Ecology of
Green Turtles (Chelonia mydas) from Shoalwater Bay, Australia. Marine
Turtle Newsletter 123:6-12.
Balazs, G. H. y Chaloupka, M. 2004. Spatial and temporal variability in somatic
growth of green sea turtles (Chelonia mydas) resident in the Hawaiian
Archipelago. Marine Biology, 145 5: 1043-1059.
Balazs, G. H., Rice, M., Murakawa, S. K.K. y Watson, G. 2000. Growth rates and
residency of immature green turtles at Kiholo Bay, Hawaii.
Bass, A. L., Epperly, S. P. y Braun-Mcneill, J. 2006. Green Turtle (Chelonia
mydas) Foraging and Nesting Aggregations in the Caribbean and Atlantic:
Impact of Currents and Behavior on Dispersal. Journal of Heredity.
Bell, I. 2012. Algivory in hawksbill turtles: Eretmochelys imbricata food selection
within a foraging area on the Northern Great Barrier Reef. Marine Ecology
Volume 34, Issue 1, pages 43–55.

44
Bennett, A.F. 1982. The energetics of reptilian activity.In Southwood A.L., R.D.
Reina, V.S. Jones, J.R. Speakman, and D.R. Jones. 2006. Seasonal
metabolism of juvenile green turtles (Chelonia mydas) at Heron Island,
Australia. Can. J. Zool. Vol. 84
Biasatti, D. M. 2004. Stable carbon isotopic profiles of sea turtle humeri:
implications for ecology and physiology. Palaeoecology. Volumen 206. P.
203-216.
Bird, K., Nichols, W. y Tambiah, C. P. 2003. The value of local knowledge in sea
turtle conservation: a case from Baja California, Mexico, Putting Fishers’
Knowledge to Work: Conference Proceedings, P. 178.
Bjorndal, K.A. 1979. Cellulose digestion and volatile fatty-acid production in the
green turtle Chelonia mydas.Comparative Biochemistry and Physiology A
63:127-134.
Bjorndal, K.A. 1980. Nutrition and Grazing Behavior of the Green Turtle Chelonia
mydas. Marine Biology.P147 y154.
Bjorndal, K.A. 1985 Nutritional ecology of sea turtles.Copeia. 3: 736-751.
Bjorndal K.A. 1997. Foraging ecology and nutrition of sea turtles.In Lutz P. and
Musick J. (eds) The biology of sea turtles. Boca Raton, Florida: CRC
Press, pp. 199–232.
Bjorndal K.A., Bolten, A.B. y Chaloupka, M.Y.. 2000. Green turtle somatic growth
model: evidence for density dependence. Ecol Appl 10:269–282
Bjorndal, K.A. y Bolten, A. B. 2008. Annual variation in source contributions to a
mixed stock: implications for quantifying connectivity. Molecular Ecology.
P. 2185–2193.

45
Bjorndal, K.A. 2010. Hawksbill sea turtles in seagrass pastures: success in a
peripheral habitat. Marine Biology.P135 y145.
Bolker B., Okuyama T., Bjorndal K., Bolten A. (2007) Incorporating multiple mixed
stocks in mixed stock analysis: ‘many-to-many’ analyses. Molecular
Ecology. pp 685–695.
Bowen, B. 1995.Tracking Marine Turtles with Genetic Markers.Bioscience. 45(8):
528-534.
Boyle, M.C. y Limpus, C.J. 2008. The stomach contents of post-hatchling green
and loggerhead sea turtles in the southwest Pacific: an insight into habitat
association. Marine Biology, 155 (2). pp. 233-241.
Brand, S. J., Lanyon, J. M. y Limpus, C. J. 1999. Digesta composition and
retention times in wild immature green turtles, Chelonia mydas: a
preliminary investigation. Marine and Freshwater Research, 50 2: 145-147.
Brand-Gardner, S., Limpus, C. y Lanyon, J. 1999.Diet selection by immature green
turtles, Chelonia mydas, in subtropical Moreton Bay, south-east
Queensland.Australian Journal of Zoology 47(2) 181 – 191.
Brooks, L. 2005. Abundance andcharacteristics of juvenile green turtle (Chelonia
mydas) in esterobanderitas. B.C.S. México. Tesis de Maestría.
Brusca, R. 1980. CommonIntertidal Invertebrates of the Gulf of
California.University of Arizona Press, Tucson.
Carr A. 1967.The turtle a natural history. Cassell, London. InHays G., Glen F.,
Broderick A., Godley B. y Metcalfe J. 2002. Behavioural plasticity in a large
marine herbivore: contrasting patterns of depth utilisation between two
green turtle (Chelonia mydas) populations. Marine Biology. 141: 985–990.

46
Cardona, L., Aguilar, A. y Pazos, L. 2009. Delayed ontogenic dietary shift and high
levels of omnivory in green turtles (Chelonia mydas) from the NW coast of
Africa.Marine Biology, 156: P. 1487-1495.
Cardona, L., Campos, P., Levy, Y., Demetropoulos, A. y Margaritoulis, D. 2010.
Asynchrony between dietary and nutritional shifts during the ontogeny of
green turtles (Chelonia mydas) in the Mediterranean.Journal of
Experimental Marine Biology and Ecology. P 83-89.
Carrion-Cortez, J.A, Zarate P, Seminoff JA. 2010. Feeding ecology of the green
turtle (Chelonia mydas) in the Galapagos Islands. J Mar Biol Assoc UK 90:
1005-1013.
Castro-González M.I., Pérez-Gil Romo F., Pérez-Estrella S. y Carrillo-Dominguez
S. Composición Química Del Alga VerdeUlva lactuca.Ciencias Marinas (
1996), 22(2): 205-2 13.
Chaloupka, M., Limpus, C. y Miller, F. 2004.Green turtle somatic growth dynamics
in a spatially disjunct Great Barrier Reef metapopulation. Coral Reefs,
Volume 23, Issue 3, pp 325-335.
Chassin-Noria, O., Abreu-Grobois, A., Dutton, P. y Oyama, K. 2004.Conservation
genetics of the east Pacific green turtle (Chelonia mydas) in Michoacan,
Mexico.Genetics 1891:1-12.
Christian, K., Green, B., Bedford, G., y Newgrain, K. 1996. Seasonal metabolism of
a small, arboreal monitor lizard, Varanus scalaris, Tropical Australia.J.
Zool. (Lond.), 240: 383–396.
CITES, Convention on International Trade in Endangered Species of Wild Fauna
and Flora (Convención sobre el Comercio Internacional de Especies
Amenazadas de Fauna y Flora Silvestres). 2012. http://www.cites.org

47
Clarke, K. y Gorley, R. 2006. Plymouth Routines In Multivariate Ecological
Research Primer 6. Original paintings for cover and CD by Greg Millwood,
Blue Peace Gallery, Barbican, Plymouth, UK.Primer-E ltd. 16 pp.
Cliffton, D., Cornejo, D. y Felger, R. 1982. Sea Turtles of the Pacific of Mexico.In:
Bjorndal, K. (Ed). Biology and Conservation of Sea Turtles.Smithsonian
Institution Press. Washington D.C. 199-209
Darré Castell, E., 2005. Hábitos alimentarios de juveniles de tortuga verde
(Chelonia mydas) en Cerro Verde, Rocha. Pasantía en la profundización
de Etología. Facultad de Ciencias, Universidad de la República, P. 55.
Delgado-Trejo, 2002, http://web.vims.edu/bridge/lavidaespanol.pdf?svr=www
DOF. 1990. Diario Oficial de la Federación 1990. México, D.F.
DOF. 2010. Diario Oficial de la Federación 2010. México, D.F.
DOF. 2012. Diario Oficial de la Federación 2010. México, D.F.
Domingo, A., Bugoni L, et al. (2006). The impact generated on sea turtles by
fisheries in the Southwest Atlantic Ocean. WWF Programa Marino para
Latinoamérica y el Caribe. San José, Costa Rica P. 71.
Dutton P.H., S.K. Davis. T. Guerra y D. Owens. 1996. Molecular Phylogeny for
Marine Turtles Based on Sequences of the ND4-leucine tRNA and Control
Regions of Mitochondrial DNA. Mol. Phylogenet. Evol. 5(3):511-521
Dutton, P., Balazs, G., Leroux, R., Murukawa, R., Zarate, P. y Martínez, L. 2008.
Composition of Hawaiian Green Turtle Foraging Aggregations: mtDNA
Evidence for a Distinct Regional Population. Endangered Species
Research.Vol. 5, Pág. 37-44

48
Eckert, K. 1993. The biology and population state of marine turtles inthe north
Pacific Ocean. USDepartment of Commerce, NOAATech.Memo.NMFS-
SWFSC-186.
Etnoyer, P., Canny, D., Mate, B.R., Morgan, L.E., Ortega-Ortiz, J.G. y Nichols,
W.J. 2006.Sea-surface temperature gradients across blue whale and sea
turtle foraging trajectories off the Baja California Peninsula, Mexico.Deep-
Sea Res II 53:340–358.
Figueroa, A., Alvarado, J., Hernández, F., Rodríguez, G. y Robles, J. 1993. The
Ecological recovery of sea turtles of Michoacan, México. Special attention
to the black turtle (Chelonia agassizii).Reporte Final para WWF-USFWS,
Albuquerque, NM.
Foley, A. M., Minch, K. S. 2007. Characteristics of a Green Turtle (Chelonia
mydas) Assemblage in Northwestern Florida Determined During a
Hypothermic Stunning Event. Gulf of Mexico Science 2: 131-143.
Forbes G. y Limpus C. 1993. A Non-lethal Method for Retrieving Stomach
Contents From Sea Turtles. Wild Res. 20, 339-43.
Fuentes, M.P.B., Lawler, I. R. y Gyuris, E. 2006. Dietary preferences of juvenile
green turtles (Chelonia mydas) on a tropical reef flat. Wildlife Research,33.
p. 671-678.
Garcia-Martinez, S. y Nichols, W. 2000.Sea Turtles of Bahía Magdalena, Baja
California Sur, Mexico: Demand and Supply of an Endangered Species.
10th Conf. Inst. Fish.Econ.Trade.Oregon State University, Corvallis,
Oregon. 07/2000
Gardner, S. y Nichols, W. 2001. Assessment of the Sea Turtles Mortality rates in
Bahía Magdalena Region, Baja California Sur, Mexico. Chelonia
Conservation and Biology. 4(1): 197-199.

49
Garnett ST, Pirce IR, Scott FJ. 1985. The diet of the green turtle, Chelonia mydas
(L.), in Torres Strait. Australian Wildlife Research 12: 103–112.
Gil-Hernández, G. y Sánchez de MedinaF. 2004. Funciones y Metabolismo de los
Nutrientes. http://www.biol.unlp.edu.ar/qcabiolfarmacia/LN-
fymnutrientes.pdf
Godley, B. J., Thompson, D.R., Waldron, S. y Furness, R. W. 1998. The Trophic
Status of Marine Turtles as Determinade by Stable Isotope
Analisys.Marine Ecology Progress Series.166 P. 277-284.
Godley, B.J., Lima, E. H. S.M., Akesson, S., Broderick, A. C. y otros 2003.
Movement patterns of green turtles in Brazilian coastal waters described
by satellite tracking and flipper tagging. Marine Ecology Progress Series.
P. 253:279-288.
Gómez-Gallardo, A. 2004. Uso de la Laguna San Ignacio por la ballena gris
Eschrichtius robustus, durante las temporadas invernales de 1996 y 1997.
Tesis de Maestría IPN-CICIMAR. 86 pp.
Groombridge, B. (Compiler). 1982. The IUCN Amphibia-Reptilia red data book.
Part 1.Testudines, Crocodylia, and Rhynchocephalia.IUCN, Gland,
Switzerland.426 pp.In NOAA, 1998
Hamann, M., Schäuble, C. S., Simon, T. y Evans, S. 2006. Demographic and
health parameters of green sea turtles Chelonia mydas foraging in the Gulf
of Carpentaria, Australia.Endang. Species Res. 2: P. 81–88.
Hamann, M., Godfrey, M.H., Seminoff, J.A., Arthur, K., Barata, P.C.R., Bjorndal,
K.A., Bolten, A.B., Broderick, A.C., Campbell, L.M.,Carreras, C., Casale,
P., Chaloupka, M., Chan, S.K.F, Coyne, M.S., Crowder, L.B., Diez, C.E.,
Dutton, P.H., Epperly, S.P., FitzSimmons, N.N., Formia, A., Girondot, M.,
Hays, G.C., Cheng, I.J., Kaska, Y., Lewison, R., Mortimer, J.A., Nichols,

50
W.J., Reina, R.D., Shanker, K.,Spotila, J.R., Tomás, J., Wallace, B.P.,
Work, T.M., Zbinden, J., y Godley, B.J. 2010. Global research priorities for
sea turtles: informing management and conservation in the 21st century.
Endangered Species Research, 11 (3). P. 245-269.
Hart, K. M. y Fujisaki, I. 2010. Satellite tracking reveals habitat use by juvenile
green sea turtles Chelonia mydas in the Everglades, Florida, USA.
Endangered species research.Vol. 11: P. 221–232.
Hatase, H., Sato, K., Yamaguchi, M., Kotaro, T. y Tsukamoto, K. 2006. Individual
Variation in Feeding Habitat use by adult Female Green Sea Turtles
(Chelonia mydas): Are they obligately neritic herbivores? Oecologia. 2006
Aug;149(1)52-64.
Hawkes, L. A., Broderick, A. C., Godfrey, M. H. y Godley, B. 2009. Climate change
and marine turtles. Endangered species research Vol. 7: P. 137–154.
Hays, G. C., 2000.The Implications of Variable Remigration Intervals for the
Assessment of Population Size in Marine Turtles.J Theor Biol. September
21;206(2):221-7.
Hays G., Glen F., Broderick A., Godley B. y Metcalfe J. 2002. Behavioural plasticity
in a large marine herbivore: contrasting patterns of depth utilisation
between two green turtle (Chelonia mydas) populations. Marine Biology.
141: 985–990.
Hayssen V. y Lacy R. 1984. Basal Metabolic Rates In Mammals: Taxonomic
Differences In The Allometry Of Bmr And Body Mass. Comp. Biocheni.
Physiol. Vol. 81A, No. 4, pp. 741—754, 1985.
Hazel J., Lawler I. y Hamann M. 2009. Diving at the shallow end: Green turtle
behaviour in near-shore foraging hábitat. Journal of Experimental Marine
Biology and Ecology 371 (2009) 84–92.

51
Heithaus, M. R., Frid, A. y Dill, L.M.. 2002. Shark-inflicted injury frequencies,
escape ability, and habitat use of green and loggerhead turtles. Marine
Biology February, Volume 140, Issue 2, P. 229-236.
Hilbert, S. C., S. C. Gardner, R. Riosmena-Rodriguez y W. J. Nichols. 2005. Diet
composition of East Pacific Green Turtles (Chelonia mydas) in Bahia
Magdalena, Baja California Sur, México. Proc 21th. Ann. Symp. Sea Turtle
Biology. Conservation..Philadelphia Pensylvania.
Hirth HF. 1997. Synopsis of the biological data on the green turtle, Chelonia mydas
(Linnaeus 1758). United States Fish and Wildlife Service Biological Report
97.
Hochscheid, S., Bentivegna, F., y Speakman, J.R. 2004. Longterm cold
acclimation leads to high Q10 effects on oxygen consumption of
loggerhead sea turtles Caretta caretta. Physiol. Biochem. Zool. 77: 209–
222.
Hulbert A. J. y Else P. L. 2003. Basal Metabolic Rate: History, Composition,
Regulation, and Usefulness. Physiological and Biochemical Zoology
77(6):869–876. University of Chicago. All rights reserved. 1522-
2152/2004/7706-3042.
Instituto Nacional de Ecología SEMARNAT. 1999. Programa nacional de
protección, conservación, investigación y manejo de tortugas marinas. P.
81.
Kaehler S. y Kennish R. 1996.Summer and Winter Comparisons in the Nutritional
Value of Marine Macroalgae from Hong Kong.Botanica Marina Vol. 39,
1996, pp. 11 -17.
Krebs, C.J. 1999. Ecological methodology.2nd Ed. Harper and Row.New York. 620
p.

52
Koch, V., Nichols, W., Peckham, H. y De la Toba, V. 2006. Estimates of Sea Turtle
Mortality from Poaching and By catch in Bahía Magdalena, Baja California
Sur, Mexico. Proceedings 22nd Annual Symposium on.Sea turtle Biology
and Conservation. Miami, Florida. 83 p
Koch, V., Brooks, L. y Nichols, W. 2007.Population ecology of the green/black
turtle (Chelonia mydas) in Bahı´a Magdalena, Mexico. Marine Biology
November 2007, Volume 153, Issue 1, P. 35-46.
Kolinski, S. P., Parker, D. M., Ilo, L. y Ruak. J. K. 2001. An assessment of the sea
turtles and their marine and terrestrial habitats at Saipan, Commonwealth
of the Northern Mariana Islands. Micronesica 34: P. 55–72.
Konar B, Iken K, Cruz-Motta JJ, Benedetti-Cecchi L, Knowlton A. 2010.Current
Patterns of Macroalgal Diversity and Biomass in Northern Hemisphere
Rocky Shores. PLoS ONE 5(10): e13195.
doi:10.1371/journal.pone.0013195
Kuiper-Linley.M., Johnson, C.R. y Lanyon, J.M. 2007.Effects of simulated green
turtle regrazing on seagrass abundance, growth and nutritional status in
Moreton Bay, south-east Queensland, Australia.Marine and Freshwater
Research, 58 (5). pp. 492-503. ISSN 1323-1650.
Lam,J. C. W., Tanabe, S., Chan, S. K. F., Yuen,E., Lam, M. H.W. y Lam, P. K. S.
2004. Trace element residues in tissues of green turtles (Chelonia mydas)
from South China Waters. Marine Pollution Bulletin (January 2004), 48 (1-
2), P. 174-182.
Lawler, I., Aragones, L., Berding, L., Marsh, H. y Foley, W. 2006. Near-Infrared
Reflectance Spectroscopy is a Rapid, Cost-Effective Predictor of Seagrass
Nutrients. J Chem Ecol. 2006 Jun;32(6):1353-65. Epub 2006 May 23.

53
Limpus, C. y Limpus, D. 2000. Manglares en la Dieta de Chelonia mydas en
Queensland, Australia. Noticiero de Tortugas Marinas. Ejemplar de Julio
no. 89.
Limpus, C.J., Limpus, D.J., Arthur, K.E. y Parmenter, C.J. 2005. Monitoring green
turtle population dynamics in Shoalwater Bay 2000-2004.Great Barrier
Reef Marine Park Authority.
Limpus, C., Bell, I. y Miller, J. 2009. Mixed Stocks of Green Turtles Foraging on
Clack Reef, Northern Great Barrier Reef Identified from Long Term
Tagging Studies. Marine Turtle Newsletter. P.123:3-5.
López-Castro, M., Koch, V., Mariscal-Loza, A. y Nichols, W. 2007. Long-term
monitoring of black turtles Chelonia mydas at coastal foraging areas off the
Baja California Peninsula.Endangered species research. P. 35-45.
López-Castro M., Villegas-Nava F., González-Payán M., Comer K. y Nichols W.
2012. In Los Cabos Prospectiva de un Paraíso Turístico. Cap. 4 Flora y
Fauna Marina y Costera. San Diego State University Press. 533 pp.
López-Mendilaharsu, M. 2002. Ecología alimenticia de Chelonia mydas agassizii
en Bahía Magdalena, B.C.S., México. Tesis de Maestría. CIBNOR.60p.
López-Mendilaharsu, M.; S. C. Gardner, J. A. Seminoff. 2003. Chelonia mydas
agassizii (east pacific green turtle), diet. Herpetologycal Review. 34: 139-
140.
López-Mendilaharsu, M., Gardner S., Riosmena-Rodríguez R., y Seminoff, J.
2005. Identifying critical foraging habitats of the green turtle (Chelonia
mydas) along the Pacific Coast of the Baja California Peninsula,
México.AquatConserv; 15:259–69.

54
López-Mendilaharsu, M., S.C. Gardner, R. Riosmena-Rodriguez & J.A. Seminoff.
2008. Diet selection by immature green turtles (Chelonia mydas) at
BahíaMagdalena foraging ground in the Pacific Coast of the Baja California
Peninsula,Mexico. Journal of the Marine Biological Association of the
United Kingdom.1-7.
Loya-Salinas, D. H. y A. Escofet. 1990. Aportaciones al cálculo del Índice de Valor
Biológico (Sanders, 1960). Contribution to the calculation of the Biological
Value Index (Sanders, 1960).Ciencias marinas, 16(2): 97-115.
Luna-Raya M.C., Urciaga-García J.I., Salinas-Zavala C.A., Cisneros-Mata M.A. y
Beltrán-Morales L.F. 2006. Diagnóstico del consumo del calamar gigante
en México y en Sonora. EconomEíac,o Snoocmiedíaa,d S yo cTieerdraitdo
ryio T, evrorli.t oVrI,i on,ú vmo.l .2 V2I,, 2n0ú0m6,. 52325, -2506006, 535-
560.
MacArthur, R.H. y E.R. Pianka. 1966. On optimal use of a patchy environment.
TheAmerican Naturalist. 100: 603-609.
Makowski, C., Seminoff, J. y Salmon, M. 2006. Home range and habitat use of
juvenile Atlantic green turtles (Chelonia mydas L.) on shallow reef habitats
in Palm Beach, Florida, USA. Marine Biology, 148, 1167-1179.
Makowski, C. y Fisher L. 2008. The Effect of Beach Nourishment on Juvenile
Green Turtle Distribution Along the Nearshore of Broward County, Florida.
The Open Marine Biology Journal, 2008, 2, 21-28.
Mancini, A. y Koch, V. 2009. Sea turtle consumption and black market trade in
Baja California Sur, Mexico. Endangered Species Research. México. Vol
7: 1-10.
Margules; C. R. y Sarka, S.-2009.-Planeación Sistematica de la Conservación.-
Universidad Nacional Autónoma de México, Comisión Nacional de Áreas

55
Naturales Protegidas y Comisión Nacional para el Conocimiento y Uso de
la Biodiversidad. 304 pp. México DF
Marquez, R. Y Doi, T. 1973. A Trial of Theoretical Analysis on Population of Pacific
Green Turtle Chelonia mydas carrinegra Cadwell, in Waters of Gulf of
California, Mexico. Bull. TokaiRegional Fisheries Res. Laboratory. 73, 1-22
Marquez, R., Peñaflores, C., Villanueva, A. y Díaz, J. 1982a. A model for
Diagnosis Pupulations of Olive Ridleys and Green Turtles of West Paficific
Tropical Coast.In: Bjorndal, k.a. (Ed). Biology and Conservation of Sea
Turtles.SmithsonianInstitutePress, Washington, D.C. 153-158
Marquez, R., Villanueva, A., Díaz, J. y Ríos, D. 1982b. Situación Actual y
Recomendaciones para el manejo de las Tortugas Marinas de la costa
Occidental mexicana, en Especial la Tortuga golfina Lepidochelys
olivácea. Sría. de Pesca. México. (3): 83-91
Marquez, R. 1990. FAO Species Catalogue Vol 11: Sea Turtles of the World. An
Annotated and Ilustrated Catalogue of the Sea Turtles Species Known to
Date.FAO Fisheries Synopsis. No. 125. Roma. 81 p
Márquez, R. 2000. Las Tortugas Marinas y Nuestro Tiempo. Fondo de Cultura
Económica. México. 197 p
Márquez, R. 2002. Las Tortugas Marinas y Nuestro Tiempo. La Ciencia para
Todos. México. 197pp.
McDermind, K., Stercke, B. y Balazs, G. 2007. Nutriotional Composition of Marine
Plants in the Diet of the Green Sea Turtle (Chelonia mydas) in the
Hawaiian Islands. Bulletin of Marine Science, Volume 81, Number 1, July
2007 , pp. 55-71(17).

56
Megill, W.M., Koch, V., Templeton, R. P., Mancini, A., Lewis, A., Burnham, R. y
Mayoral, R. 2009.Preliminary report on black sea turtle research activities
undertaken by the UABCS/UBath/Earthwatch team at Laguna San Ignacio,
BCS, Mexico, in June 2009.
Menafra, R. 2006. Bases para la Conservación y el Manejo de la Costa Uruguaya.
Vida Silvestre. P. 667.
Meylan A. 1995. Sea turtle migration – evidence from tag returns.In: Bjorndal KA
(eds) Biology and conservation of sea turtles,revised edition. Smithsonian
Institution Press, Washington, DC.
Meylan A.B. y P.A.Meylan. 2000. En: Técnicas de Investigación y Manejo para la
Conservación de Tortugas Marinas. Ed: K. Ecker, K. Bjorndal. A. Abreu-
Grobois y M. Donnelly. WWWF CMS SSC NOAA MTSG CMC. USA. Pag.
3-5.
Miller, J. 1997. Reproduction in sea turtles, EnThe Biology of Sea Turtles, in Lutz,
P. and Musick, J., Eds., CRC Press, 65-68
Moran, K.L. y Bjorndal, K.A. 2005. Simulated green turtle grazing affects
structureand productivity of seagrass pastures. Marine Ecology Progress
Series. 305, P. 235 e 247.
Morgan, Maxwell, L., Tara, F., Wilkinson, A., y Etnoyer, M. 2005. Áreas prioritarias
marinas para la conservación: Baja California al mar de Bering. Comisión
para la Cooperación Ambiental Conservación Marina. Biology Institute.
Montreal.
Mortimer, J.A. 1981. The feeding ecology of the West Caribbean green turtle
(Chelonia mydas) in Nicaragua. Biotropica 13: 49−58.

57
Musick, J.A., y Limpus, C.J. 1997. Habitat utilization and migration in juvenile sea
turtles.In Southwood A.L., R.D. Reina, V.S. Jones, J.R. Speakman,
and D.R. Jones. 2006. Seasonal metabolism of juvenile green turtles
(Chelonia mydas) at Heron Island, Australia. Can. J. Zool. Vol. 84
Naro-Maciel, E., Becker, J., Lima, E., Marcovaldi. M. A. y Desalle, R. 2007. Testing
Dispersal Hypotheses in Foraging Green Sea Turtles (Chelonia mydas) of
Brazil. Journal of Heredity 2007:98(1):29–39.
National Marine Fisheries Service y U.S. Fish and Wildlife Service. 1998.
Recovery Plan for U.S. Pacific Populations of the Green Turtle (Chelonia
mydas). National Marine Fisheries Service, Silver Spring, MD.
Nichols W., Brooks, L., López, M. y Seminoff, J. 2001. Record of pelagic East
Pacific green turtles associated with Macrocystis mats near Baja California
Sur, Mexico. Marine Turtle Newsletter 93: 10-11
Nichols, W., Dean, K., Pesenti, K., Koch, V. y Rangel, R. 2004.Advances on the
monitoring of the eastern pacific green turtle Chelonia mydas 2001-2004.
Ortega, A. y Arriaga, L. 1991. La reserva de la biosfera El Vizcaíno en la península
de Baja California. La Paz, México: Centro de Investigaciones Biológicas
de Baja California Sur, A.C.
Ortiz J. 2011. Composición Nutricional y Funcional de Algas Pardas Chilenas
:Macrocystis pyrifira y Durvillaea antárctica. Laboratorio de Química y
Análisis de Alimentos, Departamento de Ciencia de los Alimentos y
Tecnología Química. Universidad de Chile.
O'Toole, T. 2007. The effect of water temperature on the digestive physiology of
juvenile green turtles (Cheloniamydas) in a temperate feeding ground,
Moreton Bay, Australia PhD Thesis, School of Integrative Biology,
University of Queensland.

58
Pelletier, D., Roos, D. y Ciccione, S. 2003. Oceanic survival and movements of
wild and captive-reared immature green turtles (Cheloniamydas) in the
Indian Ocean. Aquatic Living Resources (0990-7440) (Elsevier), 2003 ,
Vol. 16 , N. 1 , P. 35-41.
Piñero D., et al,. 2008. La diversidad genética como instrumento para la
conservación y el aprovechamiento de la biodiversidad: estudios en
especies mexicanas, en Capital natural de México, vol. I :Conocimiento
actual de la biodiversidad. Conabio, México, pp. 437-494.
Pough F.H., Janis J.M., Heiser J.B. 1998.Vertebrate life.Prentice Hall, Engelwood
Cliffs, N.J.
Pritchard, P. C. 1971. Galápagos sea turtle – preliminary finding.Journal of
Herpetology. 5: 1-9.
Reece, J., Castoe, T. y Parkinson C. 2005. Historical Perspective on Population
Genetics and Conservation of Three Marine Turtles Species.Conservation
Genetics.Vol. 6.Pág.235-251.
Rice, M. R., Balazs,G.H., Hallacher, L., Dudley, W., Watson, G., Krussell, K. y
Larson, B. In Press. 2000. Diving, basking, and foraging patterns of a sub-
adult green turtle at Punalu'u, Hawaii. Proceedings of the 18th Annual
Symposium on Sea Turtle Biology and Conservation. U.S. Dep. Commer.,
NOAA Tech. Memo. NMFS-SEFSC.
Richardson, K. L., Lopez Castro, M., Gardner, S.C. y Schlenk, D.
2010.Polychlorinated Biphenyls and Biotransformation Enzymes in Three
Species of Sea Turtles from the Baja California Peninsula of
Mexico.Environmental Contamination and Toxicology.Volume 58, Issue 1,
P. 183-193.

59
Riosmena-Rodriguez, R. 2009. Praderas de Pastos Marinos en Laguna San
Ignacio, B.C.S. y Áreas Adyacentes: una Evaluación de Hábitats Críticos y
Conservación de Especies Protegidas. Programa de Investigación en
Botánica Marina, Departamento de Biología Marina, Universidad
Autónoma de Baja California Sur.
Robbins, C.T. 1983. Wildlife Feeding and Nutrition.Academic Press, Inc. 343 pp.
Robbins, C.T. 1993. Wildlife Feeding and Nutrition, Second Edition.Academic
Press, Inc. San Diego. 352 pp.
Rodríguez-Barón J. M. 2010. Afinidad Trófica a Zonas de Alimentación de la
Tortuga Verde (Chelonia mydas) en la Costa Occidental de Baja California
Sur, México. Tesis. CICIMAR-IPN
Rodriguez Awabdi, D., Siciliano, S. y Madeira Di Beneditto, A. P. 2013. First
Information abaut the Stomach Contents of Juvenile Green Turtles,
Cheloniamydas, In Rio de Janeiro, South-eastern Brazil.Marine
Biodiversity Records / Volume 6 / 2013, e5.
Rosales-Villa A.R. 2004. Dinámica de Nutrimentos en Bahía Magdalena, B.C.S.,
Russell, M. y Goettsch, C. 1992. La importancia de la diversidad Biológica en
México. México. Comisión Nacional para el Conocimiento y Uso de la
Biodiversidad.
Russell, D.J. y Balazs, G. H. 2000. Identification Manual For Dietary Vegetation of
the Hawaiian Green Turtle Cheloniamydas.
Russell, D.J., Balazs, G.H., Phillips, R.C. y Kam A.K.H. 2003. Discovery of the sea
grass Halophila decipiens (Hydrocharitaceae) in the diet of the Hawaiian
green turtle, Cheloniamydas. Pac Sci 57(4): 393-397.

60
Salmon, M., Jones,T. y Horch. K. W. 2004. Ontogeny of Diving and Feeding
Behavior in Juvenile Seaturtles: Leatherback Seaturtles (Dermochelys
coriacea L) and Green Seaturtles (Cheloniamydas L) in the Florida
Current. Journal of Herpetology, Vol. 38, No. 1, pp. 36–43.
Scott, M 1997. Feeding ecology of subadult green sea turtles in south Texas
waters. Editor Texas AM University. P. 152.
Seaborn, G. T., Moore, K. y Balazs, G. H. 2005. Depot fatty acid composition in
immature green turtles (Cheloniamydas) residing at two near-shore
foraging areas in the Hawaiian Islands. Comp Biochem Physiol B Biochem
Mol Biol. 140(2) P. 183-95.
Seena S., Sridhar K.R., Arum A.B. y Chiu-Chung Young. 2005. Effect of roasting
and pressure-cooking on nutritional and protein quality of seeds of
mangrove legume Canavaliacathartica from southwest coast of India.
Journal of Food Composition and Analysis 19 (2006) 284–293.
SEMARNAT. 2002. Programa Ambiental de México: Programa Nacional de
Protección y Conservación de tortugas Marinas (1992-1997). SEMARNAT-
INE. México. 79 pp.
Seminoff, J.A., Nichols, W.J. y Resendiz, A., Galvan, A. 1998. Diet composition of
the black sea turtle, Cheloniamydasagassizii, near Baja California, Mexico.
In: Abreu-Grobois A, Briseno R, Marquez R, Sarti L (eds) Proc 18th Int
Symp Sea Turtle Biol cons. US Department of Commerce, NOAA.
Seminoff, J. 2000. Biology of the East Pacific green turtle, Chelonia mydas
agassizii at a warm temperate foraging area in the Gulf of California,
México.

61
Seminoff, J., Reséndiz, A. Nichols, W. y Jones, T. 2002. Growth rates of wild green
turtles (Chelonia mydas) at a temperate foraging area in the Gulf of
California, Mexico. Copeia. 2002. (3):610-617.
Seminoff, J.A., Resendiz, A. y Nichols, W.J. 2002. Diet of the east Pacific green
turtle, Cheloniamydas, in the central Gulf of California, Mexico. J Herpetol
P. 36:447–453.
Seminoff, J.A., Resendiz, A. y Nichols, W.J . 2002. Home range of green turtles
Cheloniamydas at a coastal foraging area in the Gulf of California, Mexico.
Marine Ecology Progress Series P. 242: 253–265.
Seminoff, J. A., Jones, T. T., Resendiz, A., Nichols, W. J. y Chaloupka, M. Y. 2003.
Monitoring Green Turtles (Chelonia mydas) at Coastal Forraging area in
Baja California, México: Multiple Indices Describe Population Status.
Journal of the Marine Biological Association of the UK / Volume 83 / Issue
06 /December 2003, P. 1355-1362.
Seminoff, J. 2004. Chelonia mydas. En: IUCN 2011. IUCN Red List of
Threatened Species.Versión 2011.2.[En línea] Disponible en Internet
desde: http://www.iucnredlist.org/apps/redlist/details/4615/0.
Seminoff, J.A. y Jones T.T. 2006. Diel movements and activity ranges of green
turtles (Chelonia mydas) at a temperate foraging area in the Gulf of
California, Mexico. Herpetological Conservation and Biology 1(2):81-86.
Seminoff, J.A., Jones, T.T., Eguchi, T., Jones, D.R. y P.H. Dutton. 2006. Stable
isotope discrimination (δ13C and δ15N) between soft tissues of the green
sea turtle Chelonia mydas and its diet. Marine Ecology Progress Series
308: 271-278.

62
Senko, J., Kock, V. y Megill, W. 2008. Habitat use of black turtles (Chelonia
mydas) in San Ignacio Lagoon, Mexico.Other.LSI Ecosystem Science
Program.
Senko, J., Koch, V., Megill, W. M., Carthy, R. R., Templeton, R.P. y Nichols, W. J.
2010. Fine scale daily movements and habitat use of East Pacific green
turtles at a shallow coastal lagoon in Baja California Sur, Mexico. Journal
of Experimental Marine Biology and Ecology 391 2010 P.92–100.
Shimada, M. A. 2005. Nutrición animal. Trillas. México. 388pp.
Sokhannaro, H. y Sereywath, P. 2000. Seagrass meadow and green turtle in
Cambodia.
Southwood A.L., Reina R.D., Jones V.S., Speakman J.R. y Jones D.R. 2006.
Seasonal metabolism of juvenile green turtles (Chelonia mydas) at Heron
Island, Australia. Can. J. Zool. 84: 125–135 (2006) doi:10.1139/Z05-185.
Thayer, G. W., Bjorndal, K. A., Ogden, J.C., Williams, S.L. y Zieman, J.C. 1984.
Role of larger herbivores in seagrass communities. Estuaries 7:351-376.
The Pacific Sea Turtle Recovery Team ; for National Marine Fisheries Service and
Pacific Region, U.S. Fish and Wildlife Service. 1998. Recovery plan for
U.S. Pacific populations of the East Pacific green turtle (Chelonia mydas).
Townsend K., Altvater J., Thomas M., Schuyler Q. y Nette G. 2012. Death in the
octopus’ garden: fatal blue-lined octopusenvenomations of adult green sea
turtles. Mar Biol 159:689–695 DOI 10.1007/s00227-011-1846-9
Vidal A. 1935. Metabolismo Basal. Revista Médica Hondureña. p. 243.
Villegas-Nava F. E. 2006. Análisis Nutricional de Macroalgas y PastosAsociados a
la Alimentación de Tortuga PrietaChelonia mydas agassizii (Bocourt,
1968), en BahíaMagdalena, B.C.S., México.Tesis.UABCS.

63
Wallace B., Heppell S., Lewison R., Kelez S. y Crowder B. 2008. Reproductive
values of Loggerhead turtles in fisheries by catch worldwide. Journal of
Applied Ecology 45:1076-1085.
Wood, J. R. y F. E. Wood. 1891. Growth and Digestibility for the green Turtle
(Chelonia mydas) Feed Diets containing varying protein Levels.
Aquaculture.25:269-274.
Zar H.J. 2010.Biostatistical Analysis Fifth Edition.Prentice Hall Pearson Education
Inc.Upper Saddle River, New Jersey 07458.

64
11. Apéndice
Tabla con los datos de presencia-ausencia de especies obtenidas en la
revisión
Caribe MED GMX Atl. NO
Atl. SO
Indico SO
GT GC Pac.
C Pac. NE
Pac. NO
Pac. SO
Pac. CE
Pastos Marinos
1 1
1
1 1
1
Cymodocea sp.
1
Cymodocea nodosa
1
Cymodocea serrulata
1
Enhalus acoroides
1
Halodule sp.
1
Halodule uninervis
1
Halodule wrightii
1
1
1
Halophila sp.
Halophila bailloniss
1
Halophila decipens
1
Halophila hawaiiana
1
Halophila ovalis
1
Halophila spinulosa
1
1
Luziola peruviana
1
Syringodium sp.
1
Syringodium filiforme
1
1
1
Thalassia sp. 1
Thalassia hemprichii
1
1
Thalassia testudinum
1
1
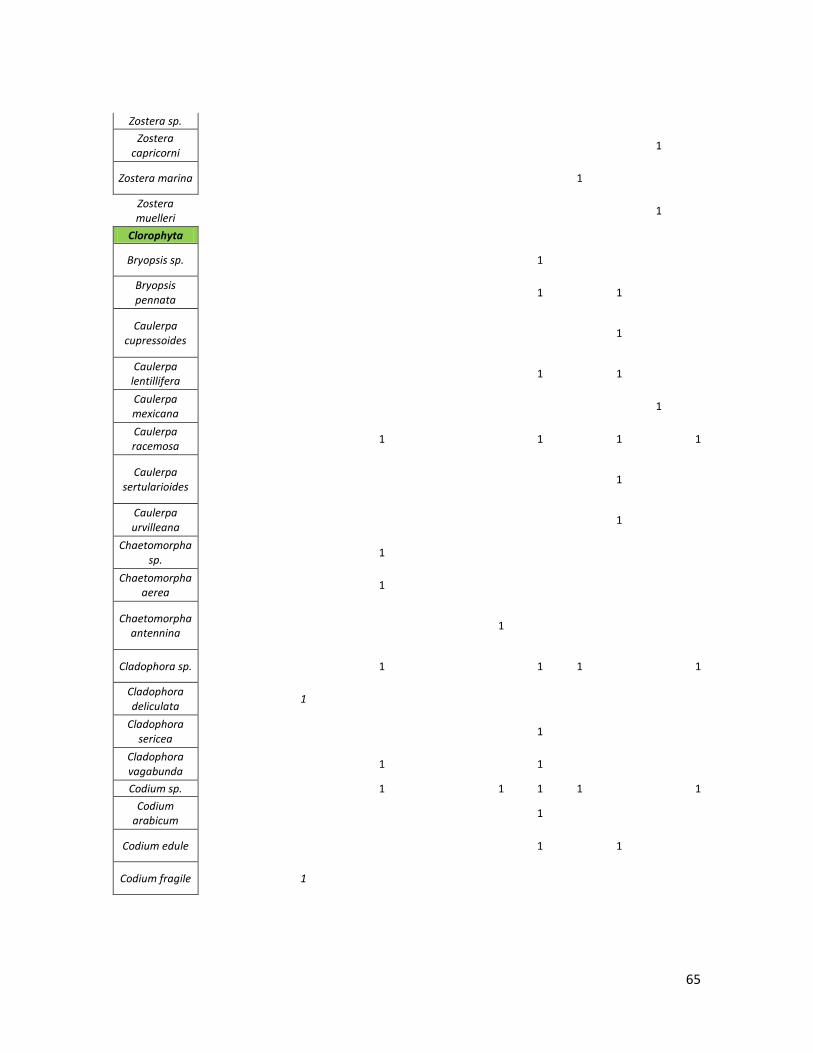
65
Zostera sp.
Zostera capricorni
1
Zostera marina
1
Zostera muelleri
1
Clorophyta
Bryopsis sp.
1
Bryopsis pennata
1
1
Caulerpa cupressoides
1
Caulerpa lentillifera
1
1
Caulerpa mexicana
1
Caulerpa racemosa
1
1
1
1
Caulerpa sertularioides
1
Caulerpa urvilleana
1
Chaetomorpha sp.
1
Chaetomorpha aerea
1
Chaetomorpha antennina
1
Cladophora sp.
1
1 1
1
Cladophora deliculata
1
Cladophora sericea
1
Cladophora vagabunda
1
1
Codium sp.
1
1 1 1
1
Codium arabicum
1
Codium edule
1
1
Codium fragile
1

66
Codium hawaiiense
1
Codium phasmaticum
1
Codium reediae
1
Codium spongiosum
1
Codium vermilara
1
Dictyosphaeria cavernosa
1
1
Dictyosphaeria versluysii
1
1
Ectocarpus siliculosus
1
Enteromorpha sp.
1
1
1
Enteromorpha acanthophora
1
Enteromorpha flexuosa
1
Halimeda gracilis
1
Halimeda opuntia
1
Halimeda tuna
1
Microdictyon setchellianum
1
Posidonia oceanica
1
Rhizoclonium grande
1
Rhizoclonium implexum
1
Ulothrix flacca
1
Ulva sp.
1
1 1
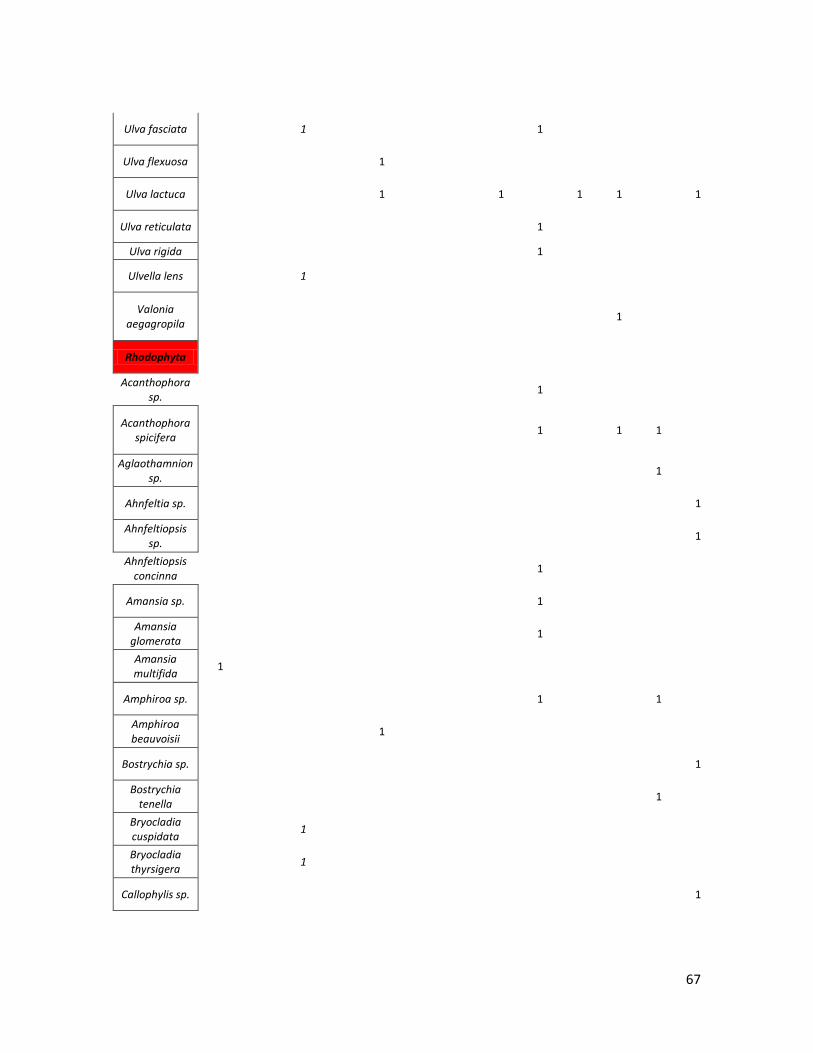
67
Ulva fasciata
1
1
Ulva flexuosa
1
Ulva lactuca
1
1
1 1
1
Ulva reticulata
1
Ulva rigida
1
Ulvella lens
1
Valonia aegagropila
1
Rhodophyta
Acanthophora sp.
1
Acanthophora spicifera
1
1 1
Aglaothamnion sp.
1
Ahnfeltia sp.
1
Ahnfeltiopsis sp.
1
Ahnfeltiopsis concinna
1
Amansia sp.
1
Amansia glomerata
1
Amansia multifida
1
Amphiroa sp.
1
1
Amphiroa beauvoisii
1
Bostrychia sp.
1
Bostrychia tenella
1
Bryocladia cuspidata
1
Bryocladia thyrsigera
1
Callophylis sp.
1

68
Caloglossa sp.
1
Catenella opuntia
1
Centroceros sp.
1 1
Centroceros clavulatum
1
1
1
Ceramium sp.
1
1
1
Ceramium byssoideum
1
Champia parvula
1
Chondracanthus sp.
1
Chondracanthus acicularis
1
Chondracanthus teedei
1
Chondria sp.
1
1
Chondria littoralis
1
Coelarthrum sp.
1
Corallina sp.
1
Corallina cubensis
1
Corallina officinalis
1
Corallina subulata
1
Criptopleura ramosa
1
Cryptomenia crenulata
1
Cryptomenia seminervis
1
Cryptopleura ramosa
1
Dasya sp.
1

69
Dictyosphaeria sp.
1
Dyctiota dichotoma
1
Euchema unicantum
1
Galaxura sp.
1
Gelidiella sp.
1
1 1
Gelidiella acerosa
1
1
Gelidium sp.
1 1
1
Gelidium crinale
1
Gelidium johnstonii
1
Gelidium pusillum
1
Gigartina sp.
1
1
Gracilaria sp. 1
1
1 1 1
1 1
Gracilaria debilis
1
Gracilaria edulis
1
Gracilaria foliifera
1
Gracilaria robusta
1
Gracilaria salicornia
1
Gracilaria spinigera
1
Gracilaria tikvahiae
1
Gracilaria verrocossa
1
Gracilariopsis laeminoformis
1
Grateloupia sp.
1
1
Grateloupia doryphora
1

70
Griffithsia sp.
1
Gymnogongrus sp.
1
Gymnogongrus griffithsiae
1
Halymenia sp.
1
Halymenia floresia
1
Halymenia refugiensis
1
Hypnea sp.
1
1 1
Hypnea cervicornis
1
Hypnea musciformis
1
1
1
1
Jania sp.
1
Jania rubens
1
Laurencia sp.
1
1
1
Laurencia johnstonii
1
Laurencia nidifica
1
Laurencia poitei
1
Leveillea sp.
1
Lithophyllum sp.
1
Lomentaria baileyana
1
Melanamansia glomerata
1
1
Plocamium brasilense
1
Polysiphonia sp.
1
1
1 1
Polysiphonia boldii
1
Polysiphonia echinata
1
Porphyra pujalsiae
1

71
Prionitis sp.
1
Pterocladia bartlettii
1
Pterocladia capillacea
1
Pterocladiella sp.
1
1 1
Pterocladiella acerosa
1
Pterocladiella capillacea
1
1
Rhodymenia sp.
Rhodymenia pseudopalmat
a
1
Rosenvingea intricata
1
Soliera tenera
1
Spyridia aculeata
1
Spyridia filamentosa
1
Tolypliocladia glomerulata
1
Vidalia obtusiloba
1
Ochrophyta
Chnoospora implexa
1
Colpomenia sp.
1
Colpomenia sinuosa
1
Cystoseira cf humilis
1
Dictyopteris delicatula
Dictyota sp.
1
1
1 1

72
Dictyota acutiloba
1
Dictyota ciliolata
1
Dictyota dichotoma
1
1 1
Dictyota flabellata
1
Hydroclathrus clathratus
1 1
Ishige sinicola
1
Levringia brasiliensis
1
Lobophora variegata
1
Myriotrichia subcorymbosa
1
Padina sp.
1 1
1
Padina australis
1
Padina durvillaei
1
Padina minor
1
Padina tenuis
1
Padina vickersiae
1
Sargassum sp.
1 1 1
1
Sargassum echinocarpum
1
Sargassum filipendula
1
Sargassum fluitans
1
Sargassum natans
1
Sargassum vulgare
1
Sporochnus bolleanus
1
Turbinaria sp.
1

73
Turbinaria ornata
1
1
Zonaria sp.
1
Cyanophyta
Hormothamnion sp.
1
1
Hormothamnion
enteromorphoides
1
Lyngbya majuscula
1
1
Lyngbya porphyrosipho
nis
1
Lyngbya semiplena
1
Oscillatoria sp.
1
Schizothrix sp.
1
Plantas Vasculares
Avicenna marina
1
Avicennia germinans
1
Ficus sp.
1
Hibiscus sp.
1
Laguncularia racemos
1
Ochroma sp.
1
Propagulos de Mangle
1
1 1
Rhizophora mangle
1
1
Scovea taccata
1
Triphora sp.
1
Vegetales de terrestres
1
Totales 12 5 33 5 35 2 3 18 66 20 26 51 28
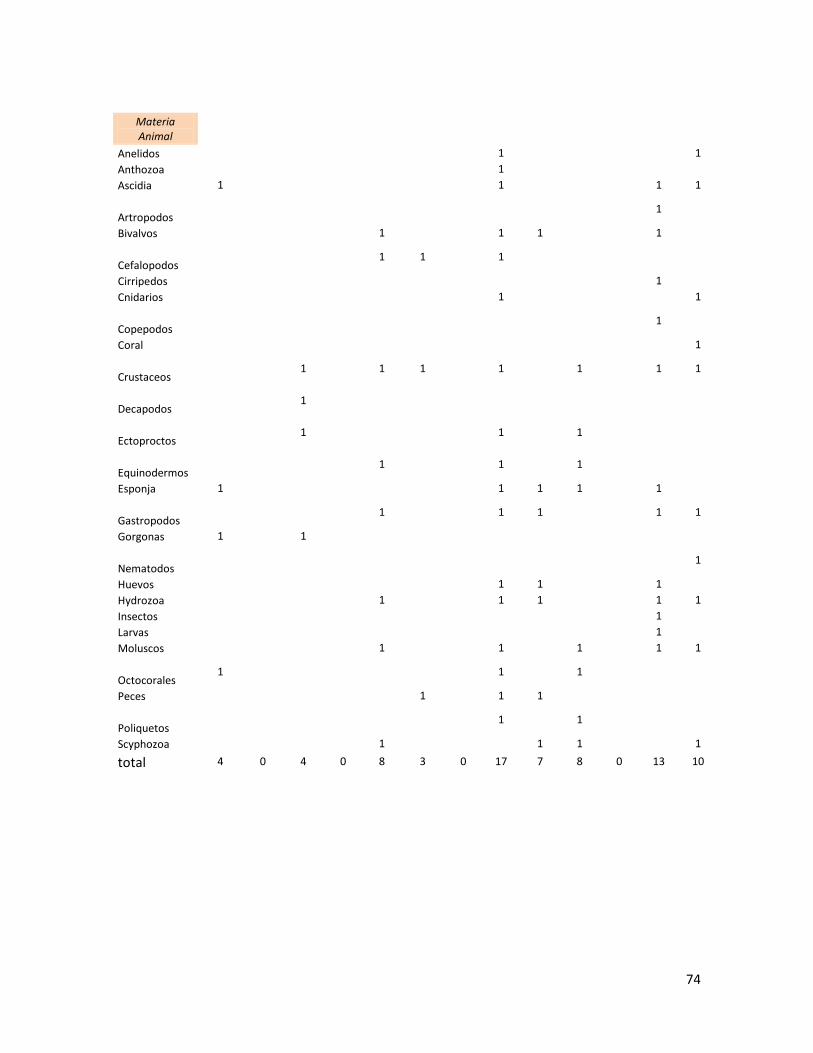
74
Materia Animal
Anelidos 1
1
Anthozoa 1
Ascidia 1
1
1 1
Artropodos 1
Bivalvos 1
1 1
1
Cefalopodos 1 1
1
Cirripedos 1
Cnidarios
1
1
Copepodos 1
Coral 1
Crustaceos 1
1 1
1
1
1 1
Decapodos 1
Ectoproctos 1
1
1
Equinodermos 1
1
1
Esponja 1
1 1 1
1
Gastropodos 1
1 1
1 1
Gorgonas 1
1
Nematodos 1
Huevos 1 1
1
Hydrozoa
1
1 1
1 1 Insectos
1
Larvas 1
Moluscos
1
1
1
1 1
Octocorales 1
1
1
Peces 1
1 1
Poliquetos 1
1
Scyphozoa 1
1 1
1
total 4 0 4 0 8 3 0 17 7 8 0 13 10

Especie Frec. Especie Frec. Especie Frec. Grupo Frec.
Pastos Marinos Rhodophyta Ochrophyta Materia Animal
Halodule wrightii
3Acanthophora
spicifera3 Dictyota sp. 4 Ascidia 4
Syringodium filiforme
3Centroceros clavulatum
3Dictyota
dichotoma3 Bivalvos 4
Clorophyta Ceramium sp. 3 Padina sp. 3 Cefalopodos 3Caulerpa racemosa
4 Gelidiella sp. 3 Sargassum sp. 4 Crustaceos 7
Cladophora sp. 4 Gelidium sp. 3Plantas
VascularesEsponja 5
Codium sp. 5 Gigartina sp. 2Propagulos de
Mangle3 Gastropodos 5
Enteromorpha sp.
3 Gracilaria sp. 7Rhizophora
mangle2 Huevos 3
Ulva sp. 3 Hypnea sp. 3 Hydrozoa 5
Ulva lactuca 5Hypnea
musciformis4 Moluscos 5
Cyanophyta Laurencia sp. 3 Octocorales 3Hormothamnio
n sp.2 Polysiphonia sp. 4 Peces 3
Lyngbya majuscula
2Pterocladiella
sp.3 Scyphozoa 4