Efecto de la lesión de nervio facial sobre la plasticidad ...
84
Efecto de la lesión de nervio facial sobre la plasticidad en la sinapsis comisural CA3-CA1 de hipocampo de rata. Rolando Andrés Torrado Arévalo Universidad Nacional de Colombia Facultad de Medicina, Maestría en Neurociencias Bogotá, Colombia 2019
Transcript of Efecto de la lesión de nervio facial sobre la plasticidad ...

Efecto de la lesión de nervio facial sobre
la plasticidad en la sinapsis comisural
CA3-CA1 de hipocampo de rata.
Rolando Andrés Torrado Arévalo
Universidad Nacional de Colombia
Facultad de Medicina, Maestría en Neurociencias
Bogotá, Colombia
2019

2
Efecto de la lesión de nervio facial sobre
la plasticidad en la sinapsis comisural
CA3-CA1 de hipocampo de rata.
Rolando Andrés Torrado Arévalo
Tesis presentada como requisito parcial para optar al título de:
Magister en Neurociencias
Directora:
Julieta Troncoso, PhD
Co-director:
Alejandro Múnera Galarza, MD, MSc, PhD
Línea de Investigación:
Fisiología del sistema motor facial
Grupo de Investigación:
Neurofisiología comportamental
Universidad Nacional de Colombia
Facultad de Medicina, Maestría en Neurociencias
Bogotá, Colombia
2019

3
A mi mamá, a mi papá y a mi hermana.

4
Agradecimientos
Primero que nada, quiero expresar mi más sincera gratitud a mi directora de
tesis, la Dra. Julieta Troncoso y a mi co-director, el Dr. Alejandro Múnera por
permitirme hacer parte de su grupo de investigación y por la paciencia e
invaluable apoyo que me brindaron durante toda mi maestría.
Agradezco también a la Dra. Marisol Lamprea por abrirme las puertas de su
laboratorio y permitirme hacer parte de su invaluable grupo de investigación,
conformado por magníficos estudiantes: Jeimmy, Ginna, Daniela, Luis,
Carlos, Moshe, Hans, Mónica, Jose y los que están por venir.
A mis compañeros de laboratorio Jonathan, Cécile y Manuel por su amistad,
por sus consejos y sus palabras de aliento en los momentos en los que los
experimentos no marchaban bien. Por las discusiones, las cervezas y los
almuerzos.
Finalmente quiero agradecer infinitamente a todos mis amigos y familiares
que estuvieron conmigo durante todo este tiempo dándome fuerzas para
continuar.

5
Resumen
Este proyecto de investigación se realizó en el laboratorio de Neurofisiología
Comportamental de la Facultad de Medicina de la Universidad Nacional, con el propósito de
observar la evolución en el tiempo de los cambios en la plasticidad sináptica del circuito
comisural CA3-CA1 del hipocampo tras una lesión del nervio facial en ratas.
Para tal fin se realizaron lesiones irreversibles de las ramas mandibular y bucal del nervio
facial (LNF) derecho, lo que provocó una parálisis del tercio inferior de la cara en los
animales experimentales. Esta parálisis imposibilitó el movimiento activo de las vibrisas del
lado lesionado.
Se registraron in vivo las amplitudes del potencial excitador post-sináptico poblacional
(fEPSP) de la región CA1 contralateral a la lesión tras una estimulación en la región CA3
ipsilateral, esto una vez hubieran transcurrido 1 (1D, n = 3), 3 (3D, n = 3), 7 (7D, n = 4) o 21
(21D, n = 2) días post-lesión y se compararon los valores obtenidos con respecto al grupo
control (Sham, n = 4).
Se observó que, aunque todos los animales presentaron una facilitación de la respuesta
mediante un protocolo de pulsos emparejados, la LNF disminuyó dicha facilitación en los
animales de los grupos 3D y 7D.
Se observó además que tras la LNF todos los grupos experimentales lograron potenciar la
respuesta post-sináptica tras un protocolo de estimulación por alta frecuencia (HFS), pero
evidenciando una disminución significativa de la expresión de la potenciación a largo plazo
(LTP) en los grupos 3D, 7D y 21D con respecto al grupo control Sham.
Con el protocolo de inducción empleado en esta investigación (6 trenes de 100 Hz) se logró
evidenciar un componente pre-sináptico en la LTP. Se observó que la LNF deterioró este

6
componente en los animales de los grupos 1D, 3D, 7D y 21D con respecto al grupo control.
Los animales del grupo control y en menor medida los animales del grupo 3D presentaron
una disminución en su facilitación de la respuesta tras la estimulación tetánica, lo cual no se
evidenció en los demás grupos experimentales.
Palabras clave: Lesión de nervio facial, hipocampo, plasticidad a corto plazo, plasticidad a
largo plazo, nervio periférico, registro in vivo.

7
Abstract
The experiments described here were conducted at the Behavioral Neurophysiology
Laboratory of Medicine School, Universidad Nacional de Colombia. The goal of this
research was to describe the evolution of changes in synaptic plasticity of hippocampal CA3-
CA1 commisural circuit, after facial nerve injury in rats.
For such end, irreversible injury on the mandibular and buccal branches of the right facial
nerve (FNI, facial nerve injury) were made. This injury provoked an irreversible paralysis of
the facial’s lower third of experimental animals. It means, the facial nerve injury produced
paralysis that lead to the impossibility of active movement of the vibrissae of the injured side.
The amplitudes of field excitatory potentials (fEPSP) were recorded in vivo in hippocampal
CA1 region (contralateral to injury) after electrical stimulation of crescent amplitude in the
CA3 region (ipsilateral to injury), at 1 (1D, n = 3), 3 (3D, n = 3), 7 (7D, n = 4) or 21 (21D, n
= 2) days post-lesion. The values obtained were compared with a sham-surgery group
(control, n = 4).
After the FNI we reported that all the experimental groups were able to potentiate the CA3-
CA1 synapsis, at least by one hour after high-frequency stimulation protocol (HFS);
however, we found a significant decrease in this potentiation in 3D, 7D and 21D groups,
compared with the control group.
Short-term plasticity was analyzed with a pair-pulse stimulation through the comparison of
the responses to two sequential identical stimuli, separated by an inter-stimuli interval of 100
ms. The ratio between the response of the second stimulus/response to the first stimulus
(R2/R1, registered in CA1 after stimulation in contralateral CA3) was evaluated in all
experimental groups. It was found that the control and all the experimental groups were able
to facilitate their response to a second stimulus in regard to the first one. Nonetheless, the
facilitation was significantly diminished in 3D and 7D groups when compared to the control

8
group. Furthermore, after a HFS, both control and 3D groups diminished significantly their
facilitation ratio.
Keywords: Facial nerve lesion, hippocampus, short term plasticity, long term plasticity, in
vivo recording.

9
Contenido
Resumen ............................................................................................................................ 5
Abstract ............................................................................................................................. 7
Lista de figuras ................................................................................................................. 11
Lista de símbolos y abreviaturas....................................................................................... 12
Lista de tablas .................................................................................................................. 14
Definición y planteamiento del problema ........................................................................ 15
1. OBJETIVOS ................................................................................................................ 17
1.1 General ................................................................................................................... 17
1.2 Específicos .............................................................................................................. 17
2. MARCO TEÓRICO ...................................................................................................... 18
2.1 Sistema sensoriomotor de las vibrisas mistaciales .................................................. 18
2.2 Sistema somatosensorial ........................................................................................ 18
2.3 Sistema motor ........................................................................................................ 22
2.4 Sistema hipocampal ................................................................................................ 23
2.5 Paradigma de lesión de nervio facial ....................................................................... 26
2.5.1 Efecto de la lesión de nervio facial sobre el sistema sensoriomotor de las vibrisas
..................................................................................................................................... 28
2.6 Efecto de la lesión del nervio facial sobre las células gliales. ................................... 29
2.7 Plasticidad sináptica................................................................................................ 30
3. MATERIALES Y MÉTODOS.......................................................................................... 34
3.1 Sujetos experimentales........................................................................................... 34
3.2 Consideraciones éticas............................................................................................ 34
3.3 Diseño experimental ............................................................................................... 34
3.4 Procedimiento quirúrgico ....................................................................................... 35
3.5 Peso de los animales antes y después de la lesión. ................................................. 36
3.6 Registros electrofisiológicos y procesamiento de datos. ......................................... 37
3.6.1 Curva estimulo respuesta o entrada por salida. ................................................ 37
3.6.2 Línea de base. .................................................................................................. 38

10
3.6.3 Plasticidad a largo plazo ................................................................................... 38
3.6.4 Plasticidad a corto plazo ................................................................................... 38
3.7 Electrodos............................................................................................................... 39
3.7.1 Electrodos de estimulación. ............................................................................. 39
3.7.2 Electrodo de registro. ....................................................................................... 40
3.8 Verificación histológica del emplazamiento de los electrodos y almacenamiento del
tejido. ........................................................................................................................... 41
3.9 Análisis estadístico .................................................................................................. 42
4. RESULTADOS ............................................................................................................. 43
4.1 Emplazamiento de los electrodos de registro y estimulación .................................. 43
4.2 Respuesta electrofisiológica de la región hipocampal CA3 tras la estimulación en la
región hipocampal CA1 contralateral. ........................................................................... 43
4.3 Lesión del nervio facial y parálisis facial. ................................................................. 45
4.4 Peso de los animales antes y después de la lesión de nervio facial. ......................... 46
4.5 Curvas entrada-por-salida pre-HFS ......................................................................... 48
4.6 Plasticidad a corto plazo. ........................................................................................ 52
4.5 Plasticidad a largo plazo. ......................................................................................... 53
4.6 Efecto de la HFS en la plasticidad a corto plazo ....................................................... 55
4.7 Curvas entrada-por-salida post-HFS ........................................................................ 57
5. DISCUSIÓN ................................................................................................................ 59
5.1 Plasticidad a corto plazo. ........................................................................................ 60
5.2 Plasticidad a largo plazo. ......................................................................................... 62
6. CONCLUSIONES ......................................................................................................... 67
PERSPECTIVAS .................................................................................................................. 68
7. BIBLIOGRAFÍA ........................................................................................................... 69

11
Lista de figuras
Figura 1. Rutas trigémino-tálamo-corticales de las vibrisas mistaciales en roedores.
Figura 2. Modelo gráfico del hipocampo en rata.
Figura 3. Diseño experimental.
Figura 4. Protocolo de estimulación.
Figura 5. Posición de los electrodos.
Figura 6. Corroboración del correcto posicionamiento de los electrodos y respuesta
electrofisiológica CA3-CA1.
Figura 7. Parálisis facial.
Figura 8. Peso de los animales.
Figura 9. Valores peso de los animales.
Figura 10. Estandarización curvas entrada-salida.
Figura 11. Curvas entrada-salida pre-HFS.
Figura 12. Plasticidad a corto plazo.
Figura 13. Plasticidad a largo plazo.
Figura 14. Plasticidad a corto plazo post-HFS.
Figura 15. Curvas entrada-salida post-HFS.

12
Lista de símbolos y abreviaturas
Abreviatura Término
LNF Lesión de las ramas bucal y mandibular del nervio periférico.
1D Grupo experimental con 1 día de recuperación post-LNF.
3D Grupo experimental con 3 días de recuperación post-LNF.
7D Grupo experimental con 7 días de recuperación post-LNF.
21D Grupo experimental con 21 días de recuperación post-LNF.
Sham Grupo control o falsa cirugía.
CA1 Cornus Ammonis 1.
CA3 Cornus Ammonis 3.
HFS Estimulación de alta frecuencia.
IEE Intervalo entre estímulos.
fEPSP Potencial excitador post-sináptico de campo.
mEPSC Corriente excitadora post-sináptica miniatura.
SNC Sistema nervioso central.
M Mediana.
X Media.
LTP Potenciación a largo plazo.
L-LTP Potenciación a largo plazo tardía.
E-LTP Potenciación a largo plazo temprana.
LTD Depresión a largo plazo.
AMPA α-ácido-3-hidroxi-5-metilo-4-isoxazolpropiónico.
AMPAr Receptor AMPA.
NMDA Ácido N-metil-D-aspártico.
NMDAr Receptor NMDA.
PrV Núcleo principal del trigémino.
SpV Núcleo espinal del trigémino.

13
VPM Núcleo ventral posterior medial del tálamo.
PoM Núcleo posterior medial del tálamo.
S1 Corteza somatosensorial primaria.
S2 Corteza somatosensorial secundaria.
M1 Corteza motora primaria.
M2 Corteza motora secundaria.
vS1 Área de las vibrisas en S1.
vM1 Área de las vibrisas en M1.
GCP Generador central de patrones.
MEA Área medial de la corteza entorrinal.
LEA Área lateral de la corteza entorrinal.
vIRt Área de las vibrisas de la formación reticular intermedia del trigémino.
GFAP Proteína acídica fibrilar glial.
Iba1 Molécula ionizada adaptadora de unión a calcio 1.
Gap-43 Proteína asociada al crecimiento 43.
ON Óxido nítrico.
FGF Factor de crecimiento fibroblástico.
IL Interleuquina.
TFG Factor de crecimiento transformante.
NFG Factor de crecimiento nervioso.
Cam-KII Calcio calmodulina quinasa II.
PSD Densidad post-sináptica.
TNF Factor de necrosis tumoral.
TARP Proteína transmembranal reguladora de AMPAr.
CA+2 Calcio.
VGCC Canales de calcio dependiente de voltaje.

14
Lista de tablas
Tabla 1: Emplazamiento de los electrodos…………………………………………………31
Tabla 2: Ecuaciones de las curvas de ajuste logarítmicas…………………………………..11
Tabla 3: Ecuaciones de las curvas de ajuste logarítmicas post-HFS………………………..11

15
Definición y planteamiento del problema
Una función primordial del sistema nervioso se centra en su capacidad de responder a
lesiones traumáticas, tanto de tipo sensorial como motor. Se ha reportado que, de 5.777
pacientes canadiense ingresados por lesiones traumáticas, 295 (5.1%) poseían lesión de
nervio periférico (Noble et al., 1998). Colombia es desafortunadamente el cuarto país del
mundo con más víctimas por minas antipersonas, muchas de ellas con amputaciones de una
o más extremidades, encontrado con 6.742 víctimas entre los años 1998 y 2008 (Ocampo et
al., 2010).
En estudios relativamente recientes, se ha evidenciado que tras una lesión de nervio ciático
hay un aumento en la expresión de citoquinas pro-inflamatorias en la región de la fascia
dentada del hipocampo de ratas y que esto está asociado con modificaciones en la plasticidad
de la sinapsis CA3-CA1 (Ren et al., 2011). Hasta la fecha no existen otros estudios que
evalúen las modificaciones en la plasticidad hipocámpica luego de lesiones en otros nervios
periféricos. En nuestro laboratorio utilizamos el modelo de lesión del nervio facial para
evaluar cambios estructurales y funcionales que ocurren en el sistema nervioso central (SNC)
luego de la lesión periférica. En este sentido, nos hemos enfocado en las modificaciones que
ocurren a nivel neuronal y glial en estructuras como la corteza sensorio-motora. En la región
de la corteza motora primaria (M1), hemos descrito una activación microglial que se hace
evidente desde los 7 y se presenta hasta los 21 días post-lesión, esto acompañado de
modificaciones morfológicas y fisiológicas en las dendritas de las neuronas piramidales
(Urrego et al., 2015). Para la región hipocampal hemos reportado un aumento bilateral en la
expresión Gap-43 (Troncoso et al., 2015), sin embargo, hasta el momento no hemos evaluado
si existen modificaciones a nivel funcional en el hipocampo tras la lesión de este nervio
periférico.
Los cambios electrofisiológicos que pueda llegar a presentar la sinapsis CA3-CA1 pueden
obedecer a varios factores, por ejemplo: la incapacidad del animal de realizar una exploración

16
activa del espacio circundante próximo y que repercuta en modificaciones en la actividad de
las redes neuronales a nivel de SNC (Grion et al., 2016); el estrés fisiológico que pueda
experimentar el animal dada la pérdida de un comportamiento vital como es el batido de las
vibrisas (Patarroyo et al., 2016) y las consecuencias que las hormonas del estrés pueden estar
teniendo sobre las redes neuronales del hipocampo (Kim & Diamond, 2002; Roozendaal,
2002); la activación microglial (Blinzinger & Kreutzberg, 1968; Urrego et al., 2015) o el
aumento de citoquinas pro-inflamatorias en SNC (Griffin et al., 2006; Kiefer et al., 1993),
con las consecuencias sobre los procesos de plasticidad.

17
1. OBJETIVOS
1.1 General
Caracterizar el efecto de la lesión unilateral irreversible de las ramas bucal y mandibular del
nervio facial sobre las propiedades electrofisiológicas basales y la potenciación a largo y
corto plazo de la sinapsis comisural CA3-CA1 del hipocampo en ratas.
1.2 Específicos
Caracterizar la respuesta post-sináptica poblacional en la región CA1 del hipocampo,
frente a estímulos de intensidad creciente en la región CA3, a distintos tiempos luego
de la lesión unilateral del nervio facial.
Caracterizar la evolución de los cambios en la plasticidad a corto plazo de la sinapsis
CA3-CA1 en el hipocampo, asociados a la lesión unilateral del nervio facial, a través
de un protocolo de estimulación con pulsos emparejados.
Caracterizar la evolución de los cambios en la plasticidad a largo plazo en la sinapsis
comisural CA3-CA1 del hipocampo, asociado a la lesión unilateral del nervio facial,
a través de un protocolo de estimulación tetánica (estimulación de alta frecuencia,
HFS).
Caracterizar las modificaciones en la probabilidad de liberación de la sinapsis CA3-
CA1 del hipocampo, asociadas a la lesión unilateral del nervio facial, antes y después
de la estimulación de alta frecuencia (HFS).

18
2. MARCO TEÓRICO
2.1 Sistema sensoriomotor de las vibrisas mistaciales
Las ratas son animales que desarrollan la mayor parte de sus actividades en ambientes
nocturnos (Norton et al., 1975). A corta distancia emplean el batido activo de las vibrisas y
el olfateo para el reconocimiento de objetos, el desplazamiento adecuado en su entorno, la
búsqueda de comida y refugio y la interacción social entre conespecíficos (Ahl, 1986)
En la rata, el parche de vibrisas mistaciales lo conforman aproximadamente 35 largas
vellosidades táctiles ubicadas a ambos lados del labio superior del animal y se encuentran
organizadas en varias filas. La fila superior (más dorsal) se denominó A, seguida de manera
descendente (más ventral) por las filas B, C, D y E. Cada fila posee entre 4 a 7 vibrisas
organizadas en arcos (Ahl, 1986; Brecht et al., 1997).
Las vibrisas se encuentran inervadas por proyecciones de las motoneuronas del nervio facial
y por aferencias del nervio trigémino, siendo estos la totalidad de su sistema sensoriomotor
(Hinrichsen & Watson, 1984). A continuación, una descripción de la conectividad cortical y
subcortical de estos nervios.
2.2 Sistema somatosensorial
El folículo piloso de cada vibrisa se encuentra inervado por neuronas del nervio trigémino
con diversos tipos de mecano-receptores, los cuales varían en su sensibilidad y en el tipo de
información sensorial que procesan. Se han reportado diferentes tipos de terminaciones
nerviosas, incluyendo: terminaciones nerviosas de Merkel, lanceoladas, reticuladas y de
Ruffini entre otras (Ebara et al., 2002; Takatoh et al., 2017).
El cuerpo celular de estas terminaciones sensoriales se encuentra principalmente en el
ganglio del trigémino y por norma cada neurona recibe información de solo una vibrisa,
organizándose de tal manera que se genera un mapa somatotópico (Kerr & Lysak, 1964;
Leiser & Moxon, 2006).

19
Las neuronas del ganglio del trigémino trasmiten la información táctil de las vibrisas a los
núcleos sensoriales del trigémino. Los núcleos del trigémino se conforman principalmente
en núcleo principal del trigémino (PrV) y el núcleo espinal del trigémino (SpV) (Friedberg
et al., 2004). La organización somatotópica presente en el ganglio se mantiene tanto en PrV
como en SpV, formando grupos discretos de neuronas que reciben el nombre de barriletes
(Belford & Killackey, 1979). Las neuronas de cada barrilete reciben información sensorial
de solo una vibrisa y mantienen confinado su árbol dendrítico dentro de los bordes del mismo.
Por otro lado, las neuronas que se encuentran entre los barriletes (tabiques o septa) presentan
arboles dendríticos mucho más extensos y reciben información sensorial de múltiples
vibrisas (Veinate & Dêschenes, 1999).
Los núcleos del trigémino poseen conexiones contralaterales con los núcleos pontinos, oliva
inferior, colículo superior y a la zona incerta. De manera ipsilateral se han descrito
conexiones con el núcleo olivar inferior, cerebelo, formación reticular pontomedular y núcleo
lateral facial (Bosman et al., 2011). Las proyecciones desde el PrV hacia las regiones lateral,
dorsolateral e intermedio del núcleo del nervio facial dan cuenta de la influencia
somatosensorial de las regiones mandibular y maxilar del rostro de la rata en el batido de las
vibrisas. Sin embargo, la mayor parte de las eferencias de los núcleos del trigémino se
proyectan hacia los núcleos ventral posterior medial (VPM) y posterior medial (PoM) del
tálamo (Pinganaud et al., 1999).
La organización somatotópica presente en los barriletes de los núcleos del trigémino se
mantiene en el núcleo VPM con el nombre de barriloides (Van der Loos, 1976), esta
estructuración anatómica no se presenta en el núcleo PoM. (Land et al., 1995, Sugitani et al.,
1990). El tálamo se considera la principal puerta de entrada de información a la región
cortical de varias modalidades sensoriales, entre ellas el tacto. La corteza somatosensorial
primaria y secundaria (S1 y S2, respectivamente) son las encargadas de procesar la
información somática proveniente de todo el cuerpo. En las ratas una gran parte de S1 se
encuentra dedicada exclusivamente a las vibrisas (vS1) y mantiene el arreglo topológico
presente en VPM, con el nombre en este caso de barriles (Woolsey & Van der Loos, 1970;
Welker, 1971). vS1 mantiene además el arreglo laminar presente en todas las cortezas, con 6
capas, siendo principalmente las capas 4 y 5 las encargadas de la entrada de información.

20
Desde las vibrisas hasta la corteza existen varias rutas eferentes, las más detalladas constan
de tres relevos neuronales: la neurona del ganglio del trigémino, la neurona de la ruta
trigémino-talámica y la neurona tálamo-cortical.
Hasta el momento se han descrito seis rutas trigémino-tálamo-corticales para la información
somatosensorial de las vibrisas, siendo las rutas lemniscal y paralemniscal de las más
detalladas (Figura 1). De estas dos, la ruta lemniscal es la principal encargada de conducir la
información sensorial individual de las vibrisas hacia la corteza, conectando los barriletes del
PrV con las capas 4, 5b y 6a de los barriles de vS1 a través de los barriloides del núcleo VPM
del tálamo (Veinante & Deschênes, 1999; Bureau et al., 2006). La ruta paralemniscal surge
desde el núcleo SpV del trigémino, proyecta en el núcleo PoM del tálamo y de ahí hacia vM1
y a las capas 5a y 1 de vS1 y vS2 (Bureau et al., 2006).
Los barriles de vS1 poseen aferencias hacia otras capas dentro de un mismo barril, a otras
regiones dentro de vS1, a otras regiones corticales fuera de vS1; como la vM1 (Izraeli &
Porter, 1995), insular, vS2 y perirrinal (Welker et al., 1988) y a los núcleos VPM y PoM en
el tálamo (Killackey & Sherman, 2003).
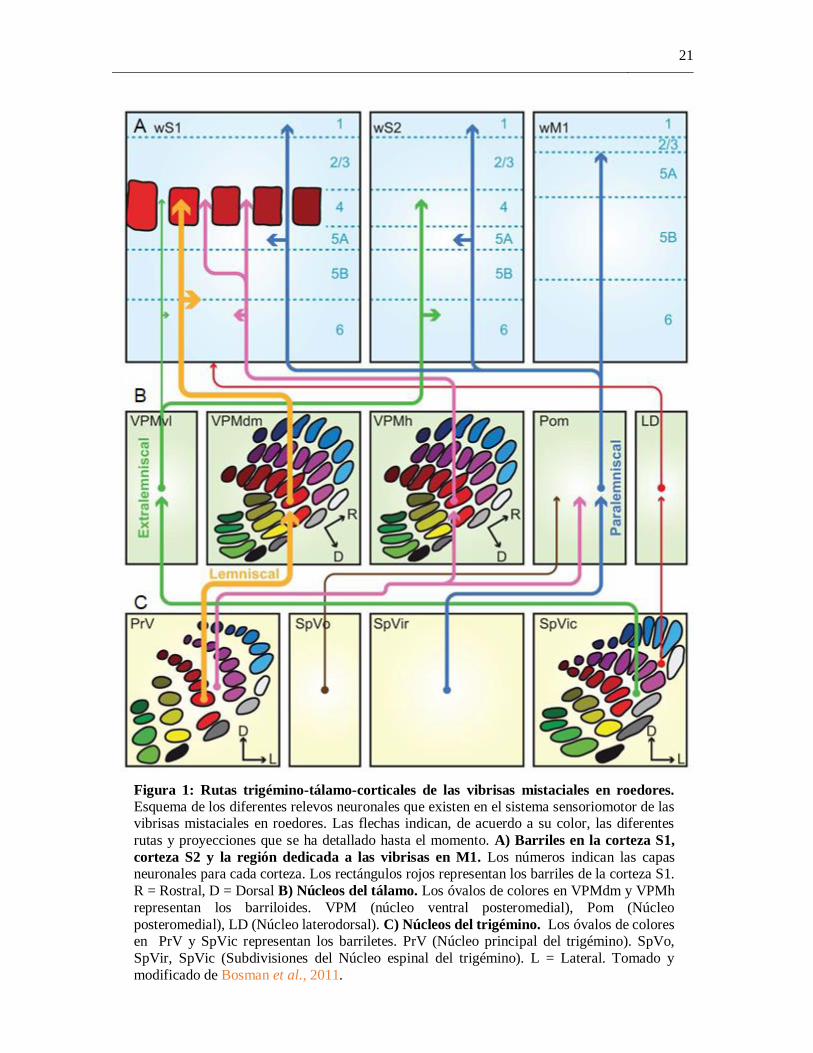
21
Figura 1: Rutas trigémino-tálamo-corticales de las vibrisas mistaciales en roedores.
Esquema de los diferentes relevos neuronales que existen en el sistema sensoriomotor de las vibrisas mistaciales en roedores. Las flechas indican, de acuerdo a su color, las diferentes
rutas y proyecciones que se ha detallado hasta el momento. A) Barriles en la corteza S1,
corteza S2 y la región dedicada a las vibrisas en M1. Los números indican las capas neuronales para cada corteza. Los rectángulos rojos representan los barriles de la corteza S1.
R = Rostral, D = Dorsal B) Núcleos del tálamo. Los óvalos de colores en VPMdm y VPMh
representan los barriloides. VPM (núcleo ventral posteromedial), Pom (Núcleo
posteromedial), LD (Núcleo laterodorsal). C) Núcleos del trigémino. Los óvalos de colores en PrV y SpVic representan los barriletes. PrV (Núcleo principal del trigémino). SpVo,
SpVir, SpVic (Subdivisiones del Núcleo espinal del trigémino). L = Lateral. Tomado y
modificado de Bosman et al., 2011.

22
2.3 Sistema motor
Las vibrisas mistaciales cuentan con dos tipos de músculos estriados que le permiten el
movimiento voluntario: los extrínsecos; aquellos que se originan fuera de la región del parche
de las vibrisas mistaciales, y los intrínsecos, asociados a los folículos de las vibrisas y a
diferencia de los extrínsecos no se encuentran anclados al cráneo. La protracción de las
vibrisas se da de manera activa y requiere exclusivamente de los músculos intrínsecos,
mientras que la retracción se da de manera pasiva (Dörfl, 1982; Haidarliu et al., 2010). Tanto
los músculos intrínsecos como extrínseco del parche de las vibrisas mistaciales son inervados
por motoneuronas colinérgicas del nervio facial, con su soma en el núcleo facial lateral
(Shohara & Sakai, 1983; Klein & Rhoades, 1985; Aldskogius & Thomander, 1986; Herfst &
Brecht, 2008; Watson et al., 1982).
El nervio facial hace parte junto con el nervio intermedio del séptimo par craneal. El nervio
intermedio es de carácter somatosensorial mientras que el nervio del facial es
predominantemente motor. A la salida del cráneo a través del foramen estilomastoideo, el
nervio facial se divide principalmente en cinco ramas, cuatro de las cuales son motoras: la
rama temporal, la cigomática, la bucal y las ramas mandibular marginal superior e inferior.
La quinta rama, la posterior auricular, es la primera en dividirse y es la única que posee fibras
eferentes. Las ramas bucal y mandibular marginal superior se encargan del movimiento de
las vibrisas, esto de acuerdo a estudios realizados con estimulación eléctrica y una
combinatoria de lesiones sobre las diferentes ramas (Semba & Egger, 1986; Henstrom et al.,
2011).
Las estructuras neuronales que originan y la regulan el batido de las vibrisas es aún tema de
investigación. El núcleo lateral del facial juega un importante rol debido a que es la única
estructura neuronal que posee una salida de información directa hacia ellas a través de sus
motoneuronas (Herfst & Brecht, 2008). Varias estructuras corticales y subcorticales
proyectan hacia el núcleo del nervio facial de manera monosináptica, entre ellas los núcleos
del trigémino, el colículo superior, la formación reticular pontomedular, el núcleo

23
mesencefálico profundo, el núcleo del rafé y el área de las vibrisas en la corteza motora
primaria (vM1) (Nguyen & Kleinfeld, 2005; Grinevich et al., 2005).
Se ha propuesto que un generador central de patrones (GCP) a nivel subcortical produce el
batido rítmico de las vibrisas. Esto con base en una serie de lesiones en vM1 que aunque
modifican de manera sutil el batido de las vibrisas no impide su generación (Welker, 1964;
Gao et al., 2003). Además, se ha visto tanto a nivel electrofisiológico como comportamental,
que la activación de la región vM1 produce un comportamiento inhibitorio y de regulación
fina del movimiento de las vibrisas de rata (Ebbesen et al., 2016; Stoltz et al., 1999).
Como GCPs se ha propuesto el núcleo del rafé debido a que sus proyecciones
serotoninergicas hacia el núcleo facial aumentan la probabilidad de disparo en las
motoneuronas durante estados de vigilia (Cramer et al., 2007). Recientemente y bajo la
evidencia de una cercanía anatómica del núcleo de las motoneuronas de las vibrisas y del
núcleo involucrado en el olfateo de la rata (Complejo pre-Bötzinger), así como la
coordinación comportamental entre el batido activo de las vibrisas y el olfateo, se ha
propuesto un oscilador neuronal para el movimiento rítmico de las vibrisas en la banda
intermedia de la formación reticular (vIRt), el cual no solo posee una actividad neuronal
coordinada con la periodicidad del batido de las vibrisas, también al verse lesionado o
inhibido farmacológicamente produce una ausencia de esta periodicidad y al activarlo de
manera artificial, la promueve (Moore et al., 2013, Deschênes et al., 2016).
Finalmente, se ha descrito que no solo vM1 posee la capacidad de modificar el movimiento
de las vibrisas. Los barriles de la corteza somatosensorial primaria ejercen también acción
directa sobre ellas y son en parte responsable de la retracción, pero no de la protracción de
las vibrisas (Izraeli & Porter, 1995; Matyas et al., 2010).
2.4 Sistema hipocampal
En el lóbulo medial temporal de los humanos se encuentran varias estructuras, entre ellas el
hipocampo, que cuentan entre sus funciones la formación de la memoria declarativa,
especialmente la memoria relacionada con eventos autobiográficos o memoria episódica y se

24
ha visto que esto se logra gracias a la capacidad que tiene el hipocampo para relacionar de
manera espacial y temporal estímulos sensoriales, cognitivos y emocionales del organismo
(Scoville & Milner, 1957; Eichenbaum et al., 1999). Para poder hacer esto el hipocampo
presenta una organización y conectividad particular.
En las ratas, la región hipocámpica se compone de dos estructuras corticales: la formación
hipocampal y la formación para-hipocampal. La formación hipocampal la componen el giro
dentado, el subículo y el hipocampo propiamente dicho, el cual a su vez se encuentra dividido
en tres regiones: CA3, CA2 y CA1 (Amaral & Witter, 1989). Todas las tres regiones de la
formación hipocampal comparten una tri-estratificación y poseen además una conectividad
unidireccional distintiva, diferente a las demás regiones corticales. El giro dentado a través
de sus fibras musgosas proyecta sobre la región CA3 del hipocampo, éste sobre el mismo a
través de sinapsis recurrentes, sobre CA1 a través de las colaterales de Schaffer y a CA2,
CA2 a su vez proyecta sobre CA1, el cual posee aferencias sobre el subículo (Ishisuka et al.,
1990). Este bucle de información en la red hipocampal concluye con las proyecciones que
emanan de CA1 y el subículo hacia la corteza entorrinal (Figura 2).
La corteza entorrinal provee al hipocampo de su mayor entrada de información cortical.
Clásicamente se ha dividido a la corteza entorrinal en área entorrinal medial (MEA) y área
entorrinal lateral (LEA). Existen dos vías de entrada hacia el hipocampo desde la corteza
entorrinal, la vía perforante que se proyecta hacia el giro dentado y CA3 y la vía temporo-
amónica que proyecta hacia la región CA1 y el subículo (Steward, 1976; Witter et al., 2000).
Debido a su rol en la formación de la memoria episódica, el hipocampo junto con la corteza
entorrinal fueron sujeto de estudios electrofisiológicos con animales no anestesiados en libre
movimiento, lo que permitió descubrir en un principio que en la región Cornus Ammonis
(CA) se encontraban grupos de neuronas con patrones de disparo que coincidían con la
posición en la que se encontraba el animal, por lo que se les denomino células de lugar
(O’Keefe & Dostrovsky, 1971; O’Keefe, 1976). Este descubrimiento modificó la concepción
del hipocampo como una estructura encargada exclusivamente de la memoria episódica y lo
posicionó como una red neuronal fundamental para la representación espacial y la memoria
espacial. Además de las células de lugar en la región CA del hipocampo, se encontró que en

25
la región dorsal de la corteza entorrinal existe otro grupo de células que también poseen un
patrón de disparo en sintonía con la posición del animal pero que, a diferencia de las células
de lugar, estas presentan varios lugares de disparo con regiones silenciadas entre ellas,
formando un patrón en forma de malla hexagonal, por lo que se les llamó células en red o
grid cells en inglés (Fyhn et al., 2004; Hafting et al., 2005). Junto con las células de lugar y
las células en red se han sumado las células de borde en el subículo (Barry et al., 2006; Lever
et al., 2009) y de posición de la cabeza en la corteza parahipocampal (Ranck, 1985), las cuales
han ampliado la diversidad de la red neuronal que permite la representación espacial en la
rata.
La región hipocampal cuenta con una simetría estructural en ambos hemisferios y posee
proyecciones recíprocas entre éstos. De manera septal, la región hipocampal, se agrupan
axones en una estructura llamada fimbria y que antes de entrar al fornix cruzan la línea media
generando fibras comisurales (Witter & Amaral, 2004).
El hipocampo no posee conexiones directas con las motoneuronas del núcleo facial; sin
embargo, existen varias rutas que el sistema somatosensorial usa para comunicarse con el
hipocampo. Se ha descubierto recientemente que la región CA1 recibe una entrada precoz de
información proveniente del núcleo reuniens del tálamo, el cual posee aferencias del núcleo
espinal (SpV) del trigémino (Krout et al., 2002, Varela et al., 2013). Contando un relevo
neuronal más, se ha descrito una entrada proveniente del sistema somatosensorial a la
formación parahipocampal a través de proyecciones del núcleo PoM del tálamo hacia la
corteza entorrinal y perirrinal (Groenewegen & Wittter, 2004; Deschênes et al., 1998).
De manera funcional se ha detallado que la frecuencia de disparo en las neuronas de la región
CA1 del hipocampo depende fuertemente de la información proveniente de las vibrisas y que
una inhibicion con muscimol de la corteza S1 y del nucleo VPM del talamo disminuye en
gran medida la actividad neuronal electrica evocada (Pereira et al., 2007), por lo que se ha
propuesto la ruta leminiscal como una entrada de informacion sensorial al hipocampo.

26
2.5 Paradigma de lesión de nervio facial
En caso de verse lesionado algún componente del sistema nervioso, los organismos cuentan
con mecanismos de contingencia que le permiten mantener la homeostasis y enfrentar la
perdida de funciones sensoriales y/o motoras que puedan llegar a experimentar (Waller,
1850). Para entender los mecanismos y limitaciones de dichas contingencias fisiológicas del
SNC en respuesta a una lesión periférica, ya sea motora o somatosensorial, ha sido necesaria
la implementación de procedimientos y técnicas que permitan una aproximación
experimental sin las restricciones de los modelos humanos. George Kreutzberg (Kreutzberg,
1986) formalizó un modelo in vivo en ratas en donde se genera una lesión del nervio facial a
la altura del foramen estilomastoideo, con lo que se ha logrado determinar mecanismo de
Figura 2: Modelo gráfico del hipocampo en rata. La línea negra representa la ruta canónica trisináptica del hipocampo. La línea roja muestra otras rutas importantes dentro del
hipocampo como las proyecciones recurrentes de CA3 o la entrada directa de la corteza
entorrinal hacia las áreas CA del hipocampo. (Tomado y modificado de Knierim, 2015). Abreviaturas: EC: Corteza entorrinal, DG: giro dentado (o fascia dentada), CA: cornus
ammonis.

27
regeneración y degeneración del nervio (Graeber et al., 1989; Hass et al., 1993; Hass et al.,
1994; McNamara et al., 2000), además de evaluar los efectos de diferentes factores
neurotróficos en la supervivencia de la motoneurona del nervio facial (Kloss et al., 1999;
Galiano et al., 2001; Raivich et al., 2004) y los cambios corticales y subcorticales
relacionados con el daño periférico (Franchi et al., 2006; Navarro et al., 2007).
La axotomía se realiza distal al foramen estilomastoideo, ya que una lesión a esta altura el
nervio facial es eminentemente motor, lo que produce una parálisis facial del lado lesionado
y que tiene como características la retracción de las vibrisas hacia la región preauricular de
la rata, la desviación de la nariz hacia el hemisferio lesionado (Urrego et al., 2011). La
axotomía del nervio facial posee varios beneficios en comparación con otro tipo de lesiones
periféricas más directas, entre ellas, la distancia que existe entre el lugar de la lesión y los
núcleos de la motoneuronas en el tronco cerebral garantiza que no se producirá una lesión
directa en el SNC y que no habrá una ruptura de la barrera hematoencefálica y por lo tanto
una invasión de células inmunes del torrente sanguíneo (Streit & Kreutzberg, 1988) o que
permite un rango de severidad en la lesión, lo cual ayuda al investigador a examinar
diferencias cualitativas y cuantitativas en los patrones de respuesta en diferentes niveles del
sistema sensorio-motor (Moran & Graeber, 2004).
Los tipos de lesión pueden clasificarse experimentalmente en quirúrgicos y farmacológicos,
y de manera funcional en reversibles e irreversibles. La lesión irreversible quirúrgica
involucra la remoción de una sección del nervio, impidiendo la regeneración axonal y, por lo
tanto, la recuperación de la función motora. De acuerdo a qué tan distal o proximal haya
ocurrido la lesión con respecto al núcleo del facial, la regeneración axonal y la muerte
neuronal serán menos o más severas, respectivamente (Mattson et al., 1998). Por otra parte,
una lesión reversible se logra, en la mayoría de los casos, realizando una compresión del
nervio, lo que conlleva un daño axonal no tan severo y con posibilidad de regeneración
estructural y funcional, la cual toma alrededor de dos a tres semana (Fawcett & Keynes,
1990). En una lesión reversible, la función motora comienza a recuperarse al décimo día,
evidenciando mejorías en el movimiento de las vibrisas tras dos semanas post-lesión y una
completa recuperación tras un mes (Torvik & Soreide, 1975).

28
Para el nervio facial se ha reportado que en ratas Wistar, aproximadamente el 25% de las
motoneuronas llevan a cabo muerte celular tras una axotomía. Este porcentaje se ve
incrementado si la axotomía se lleva a cabo de manera intracraneal y cerca al tronco del
encéfalo (Johnson & Duberley, 1998; Aldskogius & Thomander, 1986). El momento del
desarrollo en la rata tiene una gran importancia a la hora de evaluar la regeneración en las
motoneuronas. Los primeros indicios de esta reestructuración del plan fisiológico en la
motoneurona se lograron tras una compresión de nervio facial a diferentes tiempos
posnatales, en donde se reportó que la supervivencia de las motoneuronas se incrementa
conforme la rata envejece (Soreide, 1981). En estudios posteriores se cuantificó que hasta un
80% de las motoneuronas axotomizadas llevan a cabo muerte celular en neonatos (Kuno et
al., 1993).
2.5.1 Efecto de la lesión de nervio facial sobre el sistema sensoriomotor de las vibrisas
Mediante estimulación eléctrica intracortical se ha podido caracterizar una reestructuración
del mapa motor a nivel funcional en respuesta a lesión irreversible del nervio facial, en donde
la representación córtico-motora correspondiente al músculo denervado se torna silenciosa e
incapacitada de producir una activación del músculo en respuesta a una estimulación eléctrica
(Donoghue et al., 1990; Sanes et al., 1990). Pocas horas después de la lesión se puede
observar que una estimulación eléctrica en la misma región provoca la activación muscular
del párpado y antebrazo, evidenciando un proceso plástico en favor de la invasión del área
del músculo denervado por otras representaciones motoras intactas (Sanes et al., 1992). Se
ha reportado que esta rápida disminución del área de la corteza motora capaz de provocar
movimientos en las vibrisas tras un estímulo eléctrico continúa hasta por cuatro días (Toldi
et al., 1996). Se ha reportado, además, que la corteza M1 presenta una disminución de la
inhibición de manera bilateral tras una lesión del nervio facial (Farkas et al., 2000).
En nuestro laboratorio se ha observado que tras una lesión irreversible de las ramas bucal y
mandibular del nervio facial, el árbol dendrítico en neuronas piramidales de la capa 5 de M1
se retrae. Esta retracción difiere entre las dendritas apicales y basales de manera temporal y
en magnitud. Las ramificaciones apicales presentaron un pico de retracción a la semana de
haberse efectuado la lesión, con una regeneración que se prolongó por el resto del
experimento sin que estas llegasen a los niveles control. Para el árbol dendrítico apical se

29
observó una retracción permanente durante las cinco semanas que duró al menos por tres
semanas (Urrego et al., 2011). Mediante el uso de ratones transgénicos con marcadores
fluorescentes para proteínas de la microglía y de las neuronas piramidales de vM1, nuestro
laboratorio también ha podido caracterizar la proliferación de la microglía en vM1 tras una
lesión de nervio periférico (Urrego et al., 2015). Finalmente, se han reportado que este
paradigma de lesión produce cambios en las propiedades electrofisiológicas de las neuronas
piramidales de la capa 5 de vM1 (Múnera et al., 2012).
2.6 Efecto de la lesión del nervio facial sobre las células gliales.
La microglía es la célula inmune con capacidad macrófaga residente del SNC. Representa
aproximadamente un 20% del total de células gliales y se la clasifica morfológicamente entre
estado de reposo y estado activo. En su estado de reposo presenta ramificaciones que censan
de manera activa su entorno circundante. En su estado activo presenta una morfología
ameboidea, engrosando su soma celular y retrayendo sus ramificaciones, proliferando y
migrando hacia áreas del cerebro que presenten algún insulto fisiológico (Del Rio-Hortega,
1932; Kreutzberg, 1996), además de modificar su expresión proteica (Nakajima et al., 2007;
Kettenmann et al., 2012). En general la microglía propende por la supervivencia de la
neurona; sin embargo, es posible que, tras una activación muy prolongada ésta entre en un
círculo de citotoxicidad que puede llegar a ser perjudicial para las neuronas circundantes
(Aldskogius & Kozlova, 1997).
En el modelo de lesión de nervio facial se ha visto que la microglía prolifera y migra hacia
el sitio afectado, envolviendo a las motoneuronas y eliminando las terminaciones sinápticas
de aquellas que fueron axotomizadas, predominantemente en el soma celular (Blinzinger &
Kreutzberg, 1968). En ratones mutantes con expresión a la baja para la proteína MHC-I y
bajo un paradigma de lesión del nervio ciático, se encontró un aumento del desnudamiento
sináptico en las motoneuronas axotomizadas, además de inhibir la regeneración axonal hacia
el segmento distal del nervio escindido (Oliveira et al., 2004). La noción de que la microglía
actúa como agente causal del desnudamiento sináptico en las motoneuronas axotomizadas
parte de estas primeras observaciones llevadas a cabo por Kreutzberg (Blinzinger &

30
Kreutzberg, 1968), donde se puede observar una cercanía de los procesos microgliales
alrededor de la motoneurona previo a un desnudamiento sináptico. Sin embargo, se ha visto
que tras una lesión del nervio hipogloso e inhibiendo la proliferación microglial el
desnudamiento sináptico continúa presentándose, lo que pone en tela de juicio el rol de la
microglía como su principal agente causal (Svensson & Aldskogius, 1993; Perry &
O’Connor, 2010).
Los astrocitos al igual que la microglía, tras una lesión de nervio periférico, cambian su
fenotipo y se les considera activados, llevando a cabo una hipertrofia junto con una
sobreexpresión de la proteína acídica fibrilar glial (GFAP) y una reorganización de su
citoesqueleto. Los astrocitos sin embargo no proliferan y se ha visto que se encuentran
directamente relacionados con la generación de cicatriz astrocítica tras una lesión,
acompañado de una inhibición de la regeneración axonal. La causalidad de la cicatriz
astrocítica sobre la inhibición de la regeneración axonal aún es tema de estudio (Graeber &
Kreutzberg, 1988; Anderson et al., 2016).
Recientemente se ha visto que la activación de neuronas inhibitorias del estriado logra activar
receptores GABAb en astrocitos y que dicha activación astrocítica no solo produce cambios
comportamentales en el animal, también modifica los patrones de disparo y promueve la
formación de espinas dendríticas en las neuronas inhibitorias del estriado con las que tiene
contacto, con lo que se demuestra que las interacciones neurona-glía son complejas y parte
fundamental de la plasticidad sináptica neuronal (Nagai et al., 2019).
2.7 Plasticidad sináptica.
Tras una lesión de nervio periférico, ha sido de gran interés observar cambios en la
plasticidad en el cerebro. Esta puede definirse como los cambios en la fuerza sináptica de las
redes neuronales en respuesta a estímulos medioambientales entrantes, en función de
experiencias previas, lo que conlleva cambios funcionales entre las neuronas pre- y post-
sinápticas (Hebb, 1949, Wigström et al., 1986). Las sinapsis son particularmente susceptibles
a alteraciones dinámicas y cuentan con un papel fundamental en la plasticidad; éstas pueden
ver modificada su eficiencia en la transmisión sináptica en respuesta a múltiples fenómenos

31
patológicos o fisiológicos, como por ejemplo la recuperación tras una lesión de nervio
periférico (Doyére & Laroche, 1992). Así como para la axotomía de nervio periférico se
crearon modelos experimentales, la plasticidad cuenta con algunos paradigmas que tratan de
recrear procesos ocurridos de manera natural en la sinapsis, sea este el caso de la potenciación
a largo plazo LTP (Long Term Potentiation) (Bliss & Lomo, 1973). La LTP es un aumento
en la fuerza de la transmisión sináptica que puede ser inducida de manera artificial mediante
estimulación de alta frecuencia (tetánica o HFS por sus siglas en inglés High Frequency
Stimulation) o mediante ráfagas-θ (TBS por sus siglas en inglés Theta Burst Stimulation)
(Larson et al., 1986) y puede ser clasificada como de corta duración (E-LTP), la cual puede
durar hasta 1 hora, o de larga duración (L-LTP) y que puede durar desde unas pocas horas in
vitro, hasta algunos meses in vivo. Para que estos cambios se mantengan por más de una hora,
es necesario de procesos de transcripción y de síntesis de proteínas (Nguyen et al., 1994). Se
debe tener en cuenta que la expresión génica y síntesis de proteínas varía de acuerdo al patrón
de estimulación que recibe la neurona (Tyssowski et al., 2018).
Para la sinapsis CA3-CA1 se ha descrito que la transmisión sináptica depende de los
receptores post-sinápticos glutamatérgicos de tipo AMPA, mas no de los receptores
glutamatérgicos de NMDA. Sin embargo, estos últimos sí son necesarios para la inducción
de la LTP y por lo tanto la LTP en esta sinapsis es considerada dependiente de la activación
post-sináptica del receptor de NMDA (Collinridge et al., 1983; Harris et al., 1984).
Aunque la inducción de la LTP en la sinapsis CA3-CA1 dependa de receptores NMDA post-
sinápticos y de la entrada de Ca+2 a través de ellos (Müller & Connor, 1991; Malenka et al.,
1988; Nishiyama et al., 2000; Kauer et al., 1988) con la consecuente activación de la quinasa
dependiente de Ca+2/calmodulina CaMKII (Otmakhov et al., 2004; Fukunaga et al., 1993),
su expresión requiere de mecanismos moleculares que aún hoy son tema de estudio junto con
el lugar de la sinapsis en donde estos ocurren. Sin embargo hay un consenso acerca de ciertas
características: se sabe, por ejemplo, que la expresión de la LTP ocurre gracias a una
inserción de receptores AMPA (AMPAr), a través de exocitosis, en la membrana de las
dendritas de la región CA1 (Isaac et al., 1995; Jurado et al., 2013); que hay tráfico de AMPAr
desde regiones peri-sinápticas (Makino & Malinow, 2009); que existen modificaciones de
las subunidades que conforman los AMPAr, observándose por ejemplo, un aumento de la

32
conductancia en el canal mediada la fosforilación de GluA1 (Barria et al., 1997; Zamanillo
et al., 1999; Derkach et al., 1999; Derkach., 2003); que ocurre una fosforilación de proteínas
transmembranales reguladoras de AMPAr (TARPs) como la proteína stargazina (Nicoll et
al., 2006); y también que existen cambios estructurales en las espinas dendríticas a través de
la polimerización de filamentos de actina (Matsuzaki et al., 2004).
Además de la LTP, se sabe que una estimulación de baja frecuencia por periodos prolongados
resulta en una depresión a largo plazo en la fuerza de la transmisión sináptica (LTD) (Mulkey
& Malenka, 1992). La potenciación y la depresión son dos procesos que coexisten y que
permiten controlar la fuerza sináptica de las redes neuronales, generando un código
fisiológico y celular que, presumiblemente, permiten procesos complejos como el
aprendizaje y la memoria. Así como la LTP, la LTD depende del incremento de Ca+2
intracelular mediado por receptores NMDA, aunque se ha visto también formas de LTD que
dependen de la actividad de receptores metabotrópicos de glutamato (Citri & Malenka, 2008;
Kessels & Malinow, 2009; Fujii et al., 2003).
La plasticidad sináptica también puede ser de corta duración, evidenciando cambios en la
fuerza sináptica dentro de un dominio temporal del orden de las decenas de milisegundos
hasta algunos minutos. Un ejemplo de plasticidad a corto plazo, en el orden de los cientos de
milisegundos de duración, es la facilitación de la respuesta sináptica. Esta puede observarse
con un protocolo de estimulación de un par de estímulos de la misma intensidad separados
por un intervalo temporal corto. La facilitación consiste en un aumento en la amplitud de la
respuesta al segundo estimulo, en comparación con la respuesta al primer estimulo (Zucker
& Regehr, 2002). Un ejemplo de plasticidad a corto plazo en el orden de varios minutos de
duración es la potenciación post-tetánica (PPT). En la sinapsis CA3-CA1 se ha reportado una
PPT con una duración de 3 a 5 minutos independiente de receptores NMDA y dependiente
de canales de Ca+2 voltaje dependientes (Grover & Teyler, 1990).
El aumento de la fuerza sináptica en la plasticidad a corto plazo (tanto en la facilitación como
en la PPT) se le atribuye, principalmente, a cambios en la liberación de neurotransmisor por
parte de la neurona pre-sináptica. Este cambio necesita de un aumento acumulativo en la
concentración de Ca+2 intracelular (Katz & Miledi, 1968). El mecanismo de acción por el

33
cual el aumento de calcio intracelular regula la liberación de neurotransmisor ha sido
atribuido en parte a su interacción con varias proteínas con diferentes afinidades por el calcio
y que pueden tener su accionar en la movilización y anclaje vesicular; como por ejemplo
sinaptotagmina I, la PKA o la interacción Rab3A/RIM1α (Geppert et al., 1994; Trudeau et
al., 1996; Wang et al., 1997).

34
3. MATERIALES Y MÉTODOS
3.1 Sujetos experimentales
Se emplearon ratas macho (Ratus norvegicus) de la cepa Wistar, con pesos de 300 ± 10
gramos. Las ratas fueron adquiridas del Instituto Nacional de Salud (INS, Bogotá) y se
albergaron en el bioterio del Laboratorio de Aprendizaje y Comportamiento Animal (LACA)
de la Universidad Nacional de Colombia, sede Bogotá, con agua y comida ad libitum, ciclos
luz-oscuridad de 12-12 horas (luces encendidas a las 6 a.m) y temperatura controlada de 20
± 2˚C.
3.2 Consideraciones éticas
Los animales utilizados en este proyecto se manipularon de acuerdo a la constitución
colombiana en el título V de la resolución número 8430 de 1993 y a la ley 84 del 27 de
diciembre de 1989. De igual manera, se tuvieron en cuenta los procedimientos de manejo y
cuidado de animales de laboratorio recomendados por la normativa de la Unión Europea
(861609EU) y los institutos Nacionales de Salud de los Estados Unidos de América (National
Research Council, 1996).
Se utilizó la mínima cantidad de animales experimentales necesaria para obtener resultados
estadísticamente significativos. En todo momento se evitó el sufrimiento innecesario de los
animales, suministrando agentes anestésicos durante los procedimientos quirúrgicos y antes
del sacrificio.
3.3 Diseño experimental
Al llegar al LACA, todos los animales se acomodaron en grupos de a 3 animales por caja (38
cm de largo, 35 cm de ancho y 24 cm de alto, de policarbonato) y allí permanecieron por 7
días antes del comienzo de los experimentos (periodo de aclimatación). Durante este periodo,
los animales se sometieron a manipulación (handling) por parte del experimentador, dos

35
veces al día. Esto, con el fin de disminuir los niveles de estrés a un ambiente novedoso. Una
vez transcurrida la aclimatación, los animales se distribuyeron aleatoriamente en alguno de
los siguientes grupos experimentales (Figura 3):
Grupo control (Sham, n = 4), conformado por animales con falsa cirugía de lesión del
nervio facial.
Grupo lesión irreversible (n = 12), conformado por ratas con lesión irreversible del
nervio facial derecho. Este grupo de animales se dividió a su vez en 4 subgrupos: 1D
(n = 3), 3D (n = 3), 7D (n = 4) y 21D (n = 2), correspondientes a animales registrados
electrofisiológicamente a 1, 3, 7 o 21 días post-lesión, respectivamente.
3.4 Procedimiento quirúrgico
La lesión unilateral irreversible de las ramas mandibular y bucal se realizó bajo anestesia
general (ketamina 100 mg/Kg y xilazina 10 mg/Kg) administrada de manera intraperitoneal.
Posterior a la anestesia, se afeitó la región preauricular derecha de la rata, se realizó un corte
de 0,5 cm aproximadamente y se localizaron las ramas mandibular y bucal del nervio facial
derecho mediante estimulación eléctrica. Una vez localizadas las ramas se procedió a limpiar
el tejido conectivo alrededor del nervio y se escindió un segmento de aproximadamente 2
mm de cada una de las ramas, lo que imposibilitó la reconexión de los axones con su diana
sináptica muscular. La herida se suturó con dos puntos discontinuos de 4-0.
Figura 3: Diseño experimental. Esquema del diseño experimental mostrando la secuencia
temporal de los procedimientos e indicando cada uno de los grupos: Control (Sham) y lesión
del nervio con registro electrofisiológico a 1día (1D), 3 días (3D), 7 días (7D) o 21 días (21D)
post-lesión.

36
Después del periodo post-operatorio respectivo para cada subgrupo (de 1, 3, 7 0 21 días), los
animales se anestesiaron con uretano (1,5 g/Kg) intraperitoneal, para proceder al registro
electrofisiológico en el hipocampo. Una vez logrado el efecto de profundidad anestésica, se
posicionó al animal en un marco estereotáxico. Se expuso el cráneo a través de una incisión
fronto-occipital con bisturí. Se identificaron los puntos Bregma y Lambda y se corroboró que
el cráneo se encontrara correctamente nivelado en eje rostro-caudal. Se ubicaron los puntos
de inserción con respecto a Bregma para los electrodos de estimulación y de registro (Tabla
1). Después de perforar en los sitios indicados con un mototool Dremel modelo 225, se
introdujeron los electrodos mediante micromanipuladores. Las coordenadas utilizadas para
la inserción de los electrodos de registro (CA1) y estimulación (CA3) se indican en la Tabla
1, de acuerdo al Atlas estereotáxico de Paxinos y Watson (Paxinos & Watson, 2007)
3.5 Peso de los animales antes y después de la lesión.
Todos los animales fueron pesados durante los 7 días previos a la cirugía. Se calculó para
cada grupo experimental la tasa de aumento de peso mediante la pendiente máxima entre el
día de llegada al bioterio (-7) y el día de la cirugía (0). Tras la cirugía y a excepción del grupo
1D, se calculó la tasa de aumento de peso para el grupo Sham (día 1 a 7), 3D (día 1 a 3), 7D
Tabla 1: Emplazamiento de los electrodos. Coordenadas tomadas del atlas estereotáxico
Paxinos & Watson para la región dorsal del hipocampo de un mismo plano anteroposterior, -
3.8 mm con respecto al punto Bregma. El electrodo de estimulación se posicionó ipsilateral a la lesión, a la derecha de la línea media del cráneo, 3.5 mm. El electrodo de registro se
posicionó contralateral a la lesión, a la izquierda de la línea media del cráneo, - 2.5 mm. La
profundidad de ambos electrodos se tomó desde el contacto de los electrodos con las meninges
del animal, 3.0 y 2.5 mm para estimulación y registro respectivamente.

37
(día 1 a 7) y 21D (día 1 a 21). Se compararon las tasas de aumento de peso antes y después
de la cirugía.
3.6 Registros electrofisiológicos y procesamiento de datos.
Los registros electrofisiológicos se realizaron en el Laboratorio de Neurofisiología
Comportamental de la Facultad de Medicina, Universidad Nacional de Colombia, sede
Bogotá. Para el protocolo de estimulación se administraron pulsos cuadrados de corriente de
100 μs de duración a través de una unidad de aislamiento de estímulos modelo Isolator-11
(Axon instruments), contralada a su vez por un generador de pulsos modelo 9514 (Quantum
Composers). La señal fue digitalizada con una frecuencia de muestreo de 10 kHz, mediante
un convertidor análogo digital DigiData 1200 (Axon instruments). Los registros se guardaron
y se analizaron off-line mediante el software Clampfit (v.8.0).
3.6.1 Curva estimulo respuesta o entrada por salida.
Se realizó una curva estimulo respuesta o entrada por salida (I/O por sus siglas en ingles
Input/Output) con una frecuencia de estimulación de 0.33 Hz y de intensidad escalonada. Se
empleó la intensidad con un 50% de la respuesta máxima (I50) como valor de estimulación
estándar para los subsiguientes protocolos de estimulación para cada animal. Al final de los
demás protocolos de estimulación, se repitió la curva entrada por salida.
Debido a la variabilidad en la magnitud de la estimulación y la respuesta para cada animal,
los valores crudos fueron estandarizados a valores umbral en el caso de los valores de
estimulación eléctrica y en porcentaje para los valores de la respuesta electrofisiológica de la
siguiente manera: a) umbral = valor de estimulación en mA / valor de estimulación en mA
con el cual se obtuvo la menor respuesta b) porcentaje de respuesta = valor de la respuesta
en mV * 100 / valor de la respuesta máxima en mV antes de la inducción por alta frecuencia.

38
3.6.2 Línea de base.
La línea de base se realizó mediante pares de pulsos a intervalos de 100 ms y una frecuencia
de estimulación de 0.1 Hz durante media hora (180 estímulos, 6 cada minuto) o hasta que la
respuesta se hubiera estabilizado (Figura 4b). Se estandarizó cada valor de la línea de base
de la siguiente manera: Amplitud fEPSP = valor de la amplitud del fEPSP pre-HFS *100 /
promedio de los valores de la amplitud del fEPSP pre-HFS.
3.6.3 Plasticidad a largo plazo
Para la inducción de la LTP se utilizó un protocolo de estimulación de alta frecuencia (HFS
por sus siglas en ingles High Frequency Stimulation), el cual consta de 6 trenes de 100
estímulos a una frecuencia de 100 Hz, cada tren de estímulos separados por un intervalo de
60 segundos. Inmediatamente después de la HFS, se realizó nuevamente el protocolo para la
línea de base, con una frecuencia de estimulación de 0.1 Hz, durante 1 hora (Figura 4d).
Para el análisis de la LTP, los valores de la amplitud del fEPSP fueron estandarizados en
cada animal y expresados como porcentaje de cambio con respecto a la línea de base de la
siguiente manera: Amplitud fEPSP (% línea de base) = valor de la amplitud del fEPSP post-
HFS *100 / promedio de los valores de la amplitud del fEPSP pre-HFS.
3.6.4 Plasticidad a corto plazo
Para analizar la plasticidad a corto plazo en el circuito comisural CA3-CA1 realizamos un
protocolo de facilitación por pulsos emparejados. El análisis de los pulsos emparejados se
realizó a partir del cociente entre la amplitud del fEPSP de la segunda respuesta (R2) /
amplitud del fEPSP de la primera respuesta (R1) de dos estímulos separados entre sí por 100
ms. El protocolo de pulsos emparejados se realizó antes y después de la estimulación tetánica
(Figura 4b, 4d).
Tras una hora de registro de la LTP se realizó un protocolo de estimulación por pulsos
pareados con intensidad I50, una frecuencia de 0.33 Hz y una distancia interestímulo variable
de 50 ms, 100 ms, 150 ms, 200 ms y 500 ms.

39
3.7 Electrodos
3.7.1 Electrodos de estimulación.
La estimulación de la región CA3 del hipocampo derecho se realizó con electrodos bipolares
concéntricos, manufacturados en el laboratorio mediante agujas de odontología 30G y
alambre de cobre recubierto con laca.
Figura 4: Protocolo de estimulación. A) Curva entra por salida (I/O) pre-HFS Inmediatamente después de reconocer la respuesta electrofisiológica correspondiente a la
sinapsis comisural CA3-CA1 y tras haber establecido el valor de estimulación umbral
con el que se obtiene la mínima respuesta se comienza con el protocolo de curva I/O, el
cual consiste en pulsos cuadrados de 100 us de duración y valores de estimulación creciente. A partir de este protocolo se obtiene el valor de estimulación i50 B) Línea de
base. Previo a la estimulación por alta frecuencia se realizaron pulsos pareados con un
intervalo interestímulo de 100 ms y una frecuencia de 0.1 Hz durante media hora. C) Protocolo de estimulación por alta frecuencia (HFS). Para la inducción de la LTP se
aplicaron 6 trenes de 100 Hz a i50 D) Tras la HFS se repitió el protocolo de la línea de
base durante una hora E) Curva I/O post-HFS. Se repitió el protocolo de curva I/O
utilizado en la pre-HFS.

40
3.7.2 Electrodo de registro.
El registro de la región CA1 del hipocampo, contralateral a la lesión, se realizó con un
electrodo de platino/Iridio con 8 sitios de entrada, separados entre ellos cada 100 μm
(MicroProbes).
En este experimento se escogió realizar la estimulación (en CA3) y el registro (en CA1) en
hemisferios distintos del animal (derecho e izquierdo, respectivamente). Se realizó de este
modo dado que el posicionamiento de los electrodos de estimulación y registro en un mismo
hemisferio cerebral se dificulta cuando se realizan este tipo de experimentos en el animal in
vivo. La dificultad radica en los impedimentos estéricos del posicionamiento de estos
electrodos en sitios tan cercanos espacialmente. Por este motivo se escogió hacer la
estimulación en el hemisferio ipsilateral a la lesión (Figura 5a y 5b flecha roja) y el registro
en el hemisferio contralateral a la lesión (Figura 5a y 5b flecha azul), para sortear esta
dificultad.

41
3.8 Verificación histológica del emplazamiento de los electrodos y almacenamiento
del tejido.
Finalizado el registro electrofisiológico, se marcó el lugar de estimulación mediante el paso
de 25 µA por el electrodo de estimulación durante 2 minutos, utilizando un generador de
corriente (Stoelting Midgard No. 51595). Con el fin de fijar el cerebro, se realizó una
perfusión intracardiaca con la rata en estado de anestesia profunda. Se exanguinó la rata
A
Figura 5: Posición de los electrodos. Flecha roja: Electrodo de estimulación. Flecha
azul: Electrodo de registro. A) Se muestra el sitio de emplazamiento del electrodo de
registro (en la gráfica izquierda), en una diagrama de corte horizontal a una profundidad
dorsoventral de 3.0 mm y del electrodo de estimulación (en la gráfica derecha), a una profundidad dorsoventral de 2.5 mm. B) Se muestran los sitios de emplazamiento de los
electrodos de estimulación y registro en un diagrama de corte coronal a 3.8 mm de Bregma.
B

42
mediante un corte en el atrio derecho y se introdujo una cánula en el ventrículo izquierdo con
el fin de localizarla en la aorta. Después de canulada la aorta, se pasó solución salina hasta
que ya no se observó surgir sangre por el atrio derecho, una vez esto se hizo circular 200 ml
de paraformaldehído al 4%. Se decapitó la rata y se removió cuidadosamente el cerebro el
cual se depositó en un Falcon de 50 ml en post-fijación con paraformaldehído al 4%. Se
guardó el cerebro en un refrigerador hasta el momento de realizar los cortes. Los cortes
coronales en serie de la región de interés se realizaron de 50 μm de grosor en un vibrátomo
(Vibratome 1000 plus). Los cortes fueron almacenados en Eppendorfs de 1.5 mL en solución
anticongelante a una temperatura de 4˚C, con el objetivo de realizar futuras tinciones
inmunohistoquímicas.
El emplazamiento correcto de los electrodos se corroboró en un estereoscopio (Olympus
SZx16) y se tomaron fotografías con una cámara fotográfica (Sony cybershot DSCW7).
3.9 Análisis estadístico
Se empleó un análisis de varianza paramétrica (ANOVA una vía) y t-Student paramétricas
para la comparación entre los pesos de todos los grupos experimentales.
Debido a que los datos no presentan una distribución de probabilidad normal se empleó un
análisis no paramétrico de varianza (ANOVA) de medianas Kruskal-Wallis, con un análisis
post-hoc Dunn y t-Student no paramétricas.
Para el análisis de las curvas I/O pre y post-HFS, valores en porcentaje de cambio post-HFS
de la LTP, cociente R2/R1 de pulsos pareados pre-HFS y porcentaje de cambio post-HFS del
cociente R2/R1 se empleó el análisis de varianza no paramétrico.
Para el análisis entre los valores en porcentaje de cambio de la línea de base y post-HFS de
la potenciación a largo plazo y el cociente de los pulsos pareados se emplearon t-Student no
paramétricas para cada uno de los grupos experimentales.

43
4. RESULTADOS
4.1 Emplazamiento de los electrodos de registro y estimulación
Antes de analizar los datos de los registros electrofisiológicos de cada animal, se procedió a
la verificación del correcto emplazamiento de los electrodos de estimulación (en CA3) y de
registro (en CA1), dado que estos habían sido ubicados mediante coordenadas
tridimensionales, utilizando un atlas estereotáxico estándar del cerebro de rata. Para ello, una
vez acabados todos los protocolos de estimulación y registro en hipocampo, se tomaron los
cerebros perfundidos y fijados de cada animal experimental y se realizaron cortes coronales
de una zona que comprendiera +/- 0.5 mm respecto a la coordenada -3.8 mm anteroposterior
con relación al punto Bregma. En la figura 5a se observa una fotografía representativa de la
ubicación de los electrodos de estimulación (CA3, flecha roja) y registro (CA1, flecha azul).
En la figura 6b se observa la ubicación de los electrodos de estimulación (CA3, flecha roja)
y de registro (puntos azules) para todos los sujetos experimentales.
Del presente trabajo se retiraron de los análisis de resultados 8 animales experimentales
debido a que el electrodo de estimulación o el de registro se encontraron fuera de la región
CA3 o CA1 respectivamente, durante la verificación histológica del emplazamiento de los
electrodos.
4.2 Respuesta electrofisiológica de la región hipocampal CA3 tras la estimulación en
la región hipocampal CA1 contralateral.
La respuesta electrofisiológica obtenida en el stratum radiatum de CA3 tras una estimulación
en la región CA1 contralateral consistió en una deflexión positiva que comienza
aproximadamente a los 5 ms tras el estímulo, seguido de una deflexión negativa hacia los 10
ms en donde en algunos casos se puede llegar a presentar una espiga poblacional. Hacia los
20 ms la respuesta se torna nuevamente positiva (Figura 5c). Estudios previos de nuestro

44
laboratorio ya habían logrado caracterizar la respuesta electrofisiológica poblacional de los
diferentes estratos de la región CA3 del hipocampo tras una estimulación contralateral en la
región CA1 en ratón (Muñoz, J, 2015), arrojando resultados similares de respuesta trifásica
del presente trabajo.
A
Figura 6: Corroboración del correcto posicionamiento de los electrodos y respuesta electrofisiológica CA3-CA1. A) Fotografía representativa de la ubicación
de los electrodos de estimulación (CA3, flecha roja) y registro (CA1, flecha azul) de un
sujeto experimental. B) Representación de la ubicación final de los electrodos de
registro (puntos azules) y estimulación (puntos rojos) para todos los sujetos experimentales en un corte coronal -3.8 mm Bregma. C) Respuesta electrofisiológica
representativa obtenida en la región CA1 del hipocampo tras un estímulo en la región
CA3 contralateral.
B
C

45
4.3 Lesión del nervio facial y parálisis facial.
La lesión unilateral de las ramas bucal y mandibular del nervio facial produjo, de manera
inmediata una parálisis facial unilateral del tercio inferior de la cara de los animales
lesionados. Además, los animales mostraron una retracción total de las vibrisas del lado
lesionado y una desviación de la nariz hacia el lado contralateral a la lesión (Figura 7). La
parálisis facial se mantuvo hasta el final de los experimentos (es decir, al menor por 3
semanas). Por el contrario, los animales sometidos a la falsa cirugía de lesión (Sham) no
presentaron parálisis facial.
A
Figura 7: Parálisis facial. Representación gráfica del efecto de la lesión de las ramas bucal y mandibular del nervio facial en las vibrisas mistaciales. A) Animal control
con falsa cirugía de lesión (Sham): presenta batido rítmico de las vibrisas, de manera
bilateral. B) Animal experimental con lesión unilateral de las ramas bucal y mandibular del nervio facial: las vibrisas mistaciales del lado lesionado no presentan
batido activo y se encuentran retraídas. Imagen tomada y modificada de Moreno et
al., 2010.
B

46
Para mantener registro de la efectividad de la cirugía y de la presencia de la parálisis facial o
de su ausencia en el caso del grupo control, los animales fueron filmados justo antes de su
correspondiente sesión de registro electrofisiológico (video adjunto).
Tras la cirugía de lesión de nervio facial se prestó especial atención a la aparición de
inflamación de la glándula parótida, dentro de las 24 horas post-lesión, como consecuencia
de una posible complicación a consecuencia de la cirugía. En el presente estudio se presentó
un único caso de un animal con esta complicación post-quirúrgica, que se anestesió y se
sacrificó de inmediato para evitar su sufrimiento.
4.4 Peso de los animales antes y después de la lesión de nervio facial.
Todos los animales fueron pesados diariamente durante los 7 días de aclimatación y
manipulación previo a la cirugía y durante todo el periodo post-operatorio hasta el día del
registro electrofisiológico. Figura

47
Se calculó y comparó la tasa de aumento del peso (pendiente máxima gr/día) de todos los
animales durante el periodo de aclimatación al bioterio (Figura 8, días -7 a 0, siendo 0 el día
de la cirugía) y no se encontraron diferencias significativas entre ellas (F(4,11) = 1.445, p =
0.2836).
Tras la cirugía todos los animales continuaron aumentando de peso desde el segundo día. En
el primer día post cirugía todos los animales presentaron una pérdida transitoria de peso. Si
bien todos los animales continuaron aumento de peso, los animales del grupo 3D y 7D
presentaron una disminución significativa de dicha tasa tras la cirugía (Figura 9).
Figura 8: Peso de los animales. Cambios en el tiempo de los pesos antes (-7
a 0 días) y después (1 a 3, 7 o 21 días) de la cirugía (flecha negra en 0) para cada grupo experimental. Cada punto representa el promedio (±SEM) de ese
día para todos los animales de cada grupo experimental.

48
4.5 Curvas entrada-por-salida pre-HFS
Para comparar la fuerza sináptica basal en cada grupo experimental realizamos registros
extracelulares de campo en CA1 tras una serie de estímulos de corriente creciente en CA3
(Figura 10a y 10b). Se estandarizaron los valores de la respuesta post-sináptica para cada
animal (Figura 10c) y se agruparon de acuerdo a su respectivo grupo experimental, esto con
el fin de generar una curva de ajuste logarítmica representativa para cada grupo (Figura 11a
y Tabla 2).
Figura 9: Valores peso de los animales. A) Valores promedio de la pendiente
(gr/días ± SEM) antes (pre) y después (post) de la cirugía. B) Tabla con los valores de
la tasa de aumento de peso antes y después de la cirugía y resultado de la comparación estadística t-Student.
A
B

49
La comparación de la pendiente máxima para cada grupo experimental no arrojó diferencias
significativas entre los grupos experimentales (H = 4.063, p = 0.4377) (Figura 11b).
La comparación de la magnitud del porcentaje de la respuesta alcanzado a una intensidad de
estimulación estandarizada 10 umbral no arrojó diferencias significativas entre los grupos
experimentales (H = 6.04, p = 0.1942) (Figura 11c).
Tabla 2: Ecuaciones que describen las curvas de ajuste logarítmicas para cada grupo
experimental. R2 = Coeficiente de determinación.

50
A B
C
Figura 10: Estandarización de curvas entrada-salida. A) Respuesta electrofisiológica
escalonada. Trazos representativos de las respuestas electrofisiológicas de campo
obtenidas en la región CA1 del hipocampo tras estímulos escalonados en mA en la región CA3 contralateral. B) Representación de cada curva obtenida para cada animal con los
valores de la amplitud del fEPSP en mV y su respectiva estimulación en mA. C) Curva
estandarizada. Los valores de estimulación fueron convertidos a valores umbrales en
donde la estimulación con la que se obtuvo la mínima respuesta es 1. La respuesta fue convertida a porcentaje en base al mayor valor obtenido en la curva pre-HFS, la cual toma
un valor de 100%.
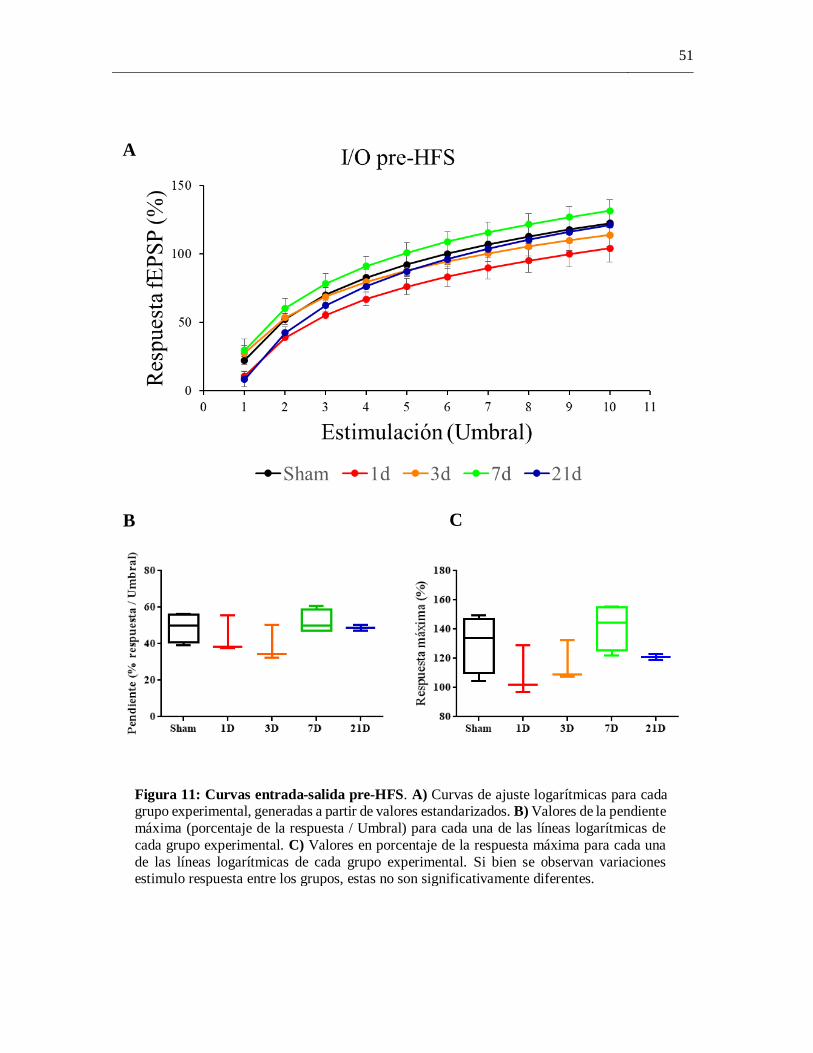
51
C B
Figura 11: Curvas entrada-salida pre-HFS. A) Curvas de ajuste logarítmicas para cada grupo experimental, generadas a partir de valores estandarizados. B) Valores de la pendiente
máxima (porcentaje de la respuesta / Umbral) para cada una de las líneas logarítmicas de
cada grupo experimental. C) Valores en porcentaje de la respuesta máxima para cada una
de las líneas logarítmicas de cada grupo experimental. Si bien se observan variaciones estimulo respuesta entre los grupos, estas no son significativamente diferentes.
A
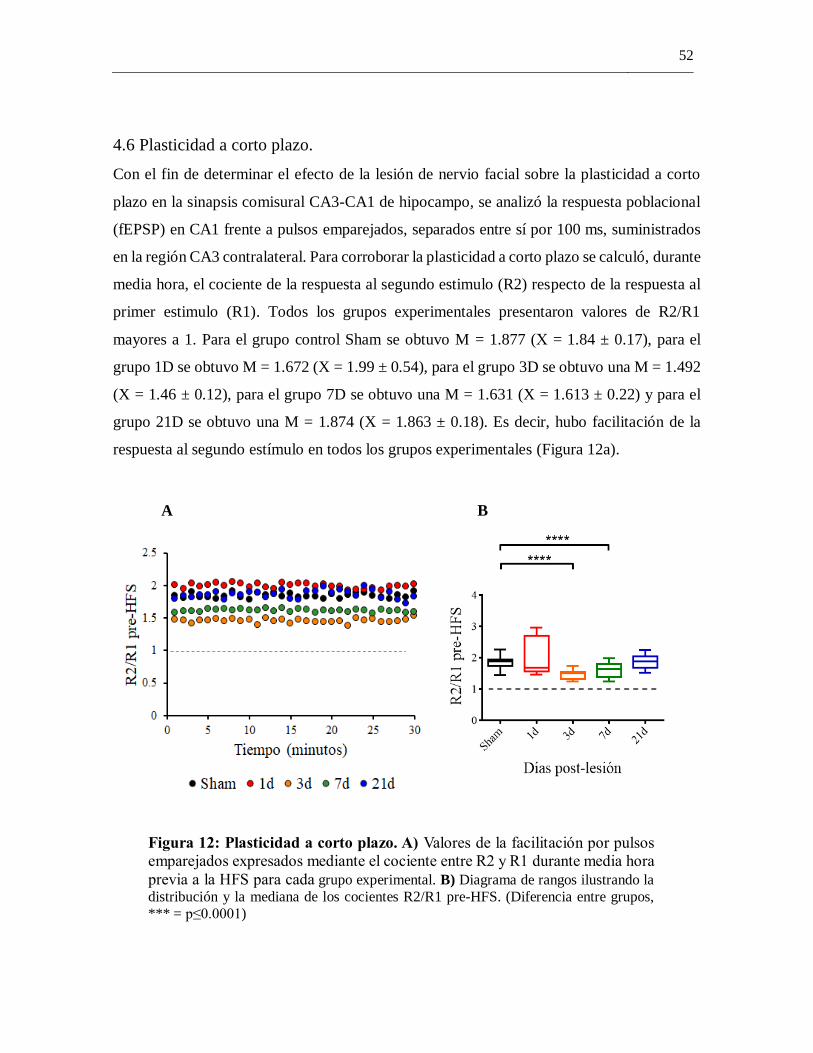
52
4.6 Plasticidad a corto plazo.
Con el fin de determinar el efecto de la lesión de nervio facial sobre la plasticidad a corto
plazo en la sinapsis comisural CA3-CA1 de hipocampo, se analizó la respuesta poblacional
(fEPSP) en CA1 frente a pulsos emparejados, separados entre sí por 100 ms, suministrados
en la región CA3 contralateral. Para corroborar la plasticidad a corto plazo se calculó, durante
media hora, el cociente de la respuesta al segundo estimulo (R2) respecto de la respuesta al
primer estimulo (R1). Todos los grupos experimentales presentaron valores de R2/R1
mayores a 1. Para el grupo control Sham se obtuvo M = 1.877 (Χ = 1.84 ± 0.17), para el
grupo 1D se obtuvo M = 1.672 (X = 1.99 ± 0.54), para el grupo 3D se obtuvo una M = 1.492
(X = 1.46 ± 0.12), para el grupo 7D se obtuvo una M = 1.631 (X = 1.613 ± 0.22) y para el
grupo 21D se obtuvo una M = 1.874 (X = 1.863 ± 0.18). Es decir, hubo facilitación de la
respuesta al segundo estímulo en todos los grupos experimentales (Figura 12a).
Figura 12: Plasticidad a corto plazo. A) Valores de la facilitación por pulsos
emparejados expresados mediante el cociente entre R2 y R1 durante media hora
previa a la HFS para cada grupo experimental. B) Diagrama de rangos ilustrando la
distribución y la mediana de los cocientes R2/R1 pre-HFS. (Diferencia entre grupos,
*** = p≤0.0001)
A B

53
El análisis estadístico arrojó diferencias significativas entre el cociente R2/R1 de los grupos
experimentales (H = 181.1, p < 0.0001). El análisis post-hoc mostró una disminución
estadísticamente significativa del cociente R2/R1 entre el grupo control Sham (M = 1.877) y
los grupos 3D (M = 1.492, p < 0.0001) y 7D (M = 1.631, p<0.0001), no así para los grupos
experimentales 1D (M = 1.672) y 21D (M = 1.874) (Figura 12b).
4.5 Plasticidad a largo plazo.
Se analizó el porcentaje de cambio de la amplitud del fEPSP tras un protocolo de
estimulación de alta frecuencia con respecto a la línea de base (Figura 13a). Todos los grupos
experimentales presentaron un aumento significativo del porcentaje de cambio de la amplitud
del fEPSP tras la HFS con respecto a la línea de base (100%). Sham (M = 203.1%, U = 0.0,
p < 0.0001), 1D (M = 199.5%, U = 0.0, p<0.0001), 3D (M = 135.1%, U = 4.0, p < 0.0001),
7D (M = 151.1%, U = 199.0, p < 0.0001) y 21D (M = 180%, U=6.0, p < 0.0001). Esta
potenciación de la respuesta en CA1, luego de la HFS en CA3 contralateral, se mantuvo
durante al menos una hora (Fig. 13b y 13c).
El análisis estadístico arrojo diferencias significativas entre los porcentajes de cambio de la
amplitud del fEPSP de los grupos experimentales (H = 371.7, p < 0.0001). Tras el análisis
post-hoc se reportaron diferencias significativas entre el grupo control Sham (M = 203.1%)
y los grupos experimentales 3D (M = 135.1%, p < 0.0001), 7D (M = 151.1%, p < 0.0001) y
21D (M = 180%, p < 0.0001). Esta diferencia no se encontró en los animales del grupo 1D
(M = 199.5%) (Figura 13b y 13c)

54
Figura 13: Plasticidad a largo plazo. A) Trazos representativos del fEPSP
antes (color claro) y después (color
oscuro) de la HFS, para cada uno de los
grupos experimentales. B) Porcentaje de cambio de la amplitud del fEPSP con
respecto a la línea de base. Se registró
una hora tras la HFS (Flecha negra) C)
Diagrama ilustrando rangos y mediana
del porcentaje de cambio con respecto a
la línea de base tras la HFS. (Diferencia
entre grupos *** = p≤0.0001. Diferencia con respecto a la línea de base δ =
p≤0.0001)
A
B
C

55
4.6 Efecto de la HFS en la plasticidad a corto plazo
Procedimos a evaluar el efecto de una estimulación tetánica en la liberación vesicular pre-
sináptica en la sinapsis CA3-CA1. Para este fin, evaluamos los cambios en la relación R2/R1
antes y después del protocolo de estimulación (Figura 14a).
Primero comparamos las relaciones R2/R1 antes y después de la HFS para cada grupo. Tras
la HFS solo los grupos experimentales 3D (M = 85.02%, U = 3869, p<0.0001) y el grupo
control Sham (M = 80.64%, U=53, p<0.0001) presentaron una disminución significativa
respecto de la facilitación antes de la HFS. Los grupos experimentales 1D (M = 98.79%, U
= 7195, p = 0.1349) y 7D (M = 100.6%, U=13386, p=0.2763) no presentaron diferencias
significativas con respecto a su línea de base. El grupo 21D (M = 102.4%, U=2909,
p=0.0358) presentó un aumento significativo respecto de la facilitación antes de la HFS
(Figura 14b).
Luego comparamos las relaciones R2/R1 entre grupos. Tras la HFS se encontraron
diferencias significativas entre los porcentajes de cambio del cociente R2/R1 de los grupos
experimentales (H=403.1, p<0.0001). El análisis post-hoc arrojó que los grupos 1D, 3D, 7D
y 21D presentaron diferencias significativas con respecto al grupo control Sham (p<0.0001)
(Figura 14b).

56
0
20
40
60
80
100
120
140
-30 -20 -10 0 10 20 30 40 50 60
R2
/R1
am
pli
tud
fE
PS
P (
%)
Tiempo (minutos)
Sham 1d 3d 7d 21d
A
Figura 14: Potenciación pre-sináptica.
A) Porcentaje de cambio del cociente R2/R1 con respecto a la línea de base.
Valores estandarizados con respecto al
promedio de los valores de la línea de
base. B) Diagrama ilustrando rango y mediana del porcentaje de cambio del
cociente R2/R1 tras la HFS. (Diferencias
entre grupos, *** = p≤0.0005. Diferencias con respecto a la línea de base, δδδ =
p≤0.0001, δ = p≤0.05).
B
HFS

57
4.7 Curvas entrada-por-salida post-HFS
Finalmente, tras una hora de registro de la inducción de la LTP por HFS, se repitió la curva
estímulo-respuesta y se caracterizó la excitabilidad de la sinapsis CA3-CA1 (Figura 15a y
Tabla 3)
La comparación de la pendiente máxima para cada grupo experimental no arrojo diferencias
significativas entre los grupos experimentales (H = 6.316, p = 0.1695) (Figura 15b).
La comparación de la magnitud del porcentaje de la respuesta alcanzado a una intensidad de
estimulación estandarizada 10 umbral no arrojó diferencias significativas entre los grupos
experimentales (H = 6.489, p = 0.1553) (Figura 15c).
Tabla 2: Ecuaciones que describen las curvas de ajuste logarítmicas para cada grupo
experimental. R2 = Coeficiente de determinación.
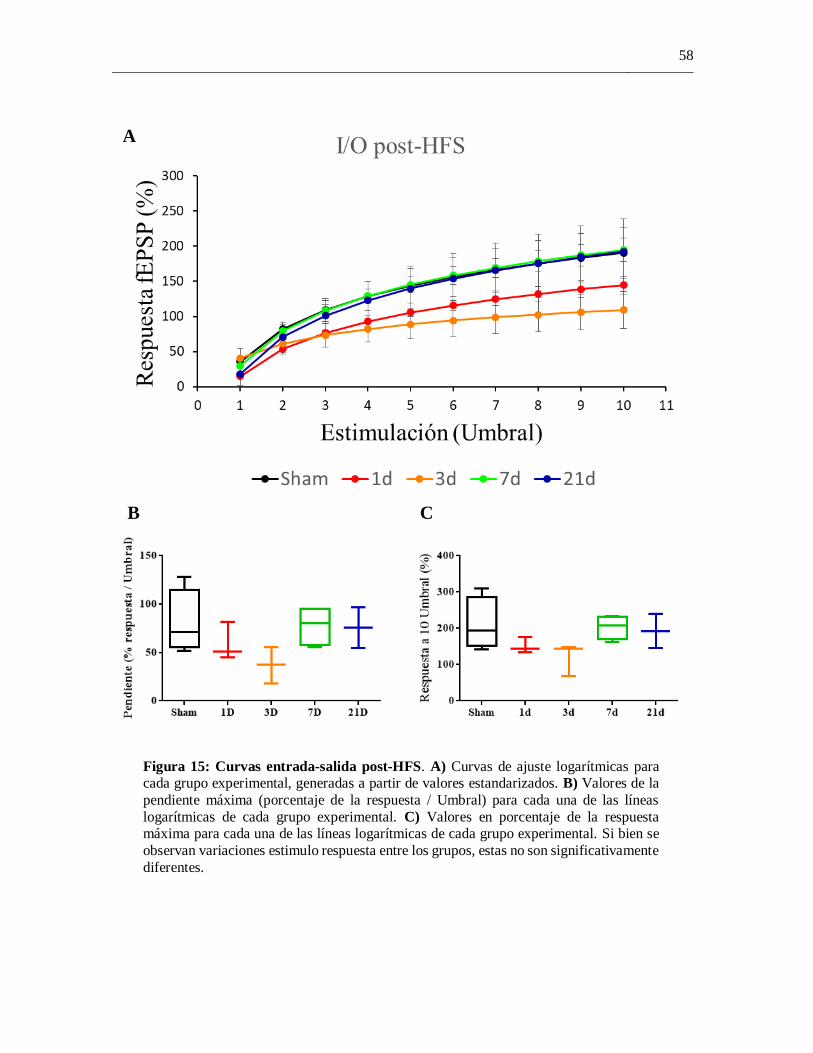
58
Figura 15: Curvas entrada-salida post-HFS. A) Curvas de ajuste logarítmicas para cada grupo experimental, generadas a partir de valores estandarizados. B) Valores de la
pendiente máxima (porcentaje de la respuesta / Umbral) para cada una de las líneas
logarítmicas de cada grupo experimental. C) Valores en porcentaje de la respuesta máxima para cada una de las líneas logarítmicas de cada grupo experimental. Si bien se
observan variaciones estimulo respuesta entre los grupos, estas no son significativamente
diferentes.
A
B C

59
5. DISCUSIÓN
En este estudio reportamos que una lesión unilateral e irreversible de las ramas bucal y
mandibular del nervio facial deteriora la potenciación a largo plazo de la sinapsis comisural
CA3-CA1. Este deterioro presenta componentes tanto pre-sinápticos como post-sinápticos.
A continuación, presentamos un resumen de los resultados obtenidos en esta investigación y
una discusión acerca de las posibles consecuencias fisiológicas y moleculares que surgen
como producto de la lesión del nervio facial y que podrían estar explicando los cambios
observados en la plasticidad a largo plazo de la sinapsis CA3-CA1 del hipocampo.
Nos interesamos en observar cambios funcionales en el hipocampo de la rata a través de
registros electrofisiológicos y bajo un paradigma de lesión irreversible del nervio facial dado
que en estudios previos realizados por nuestro grupo de investigación encontramos que las
ratas privadas del movimiento activo de las vibrisas presentaban una deficiencia en el recobro
de la memoria espacial en el laberinto de Barnes (Patarroyo et al., 2017), así como la
imposibilidad de reconocer un objeto novedoso en función de su textura en una prueba de
reconocimiento de objetos (Moreno et al., 2010). Tanto el reconocimiento de objetos
novedosos como la memoria espacial se han visto relacionados con la región hipocampal
(Morris et al., 1982; Moser et al., 1995; Tsao et al., 2013) la cual se ha considerado como la
región del SNC capaz de proveer un marco de referencia espacial para los eventos
experimentados por el individuo (Moser et al., 2015), integrando el estado emocional y
diferentes modalidades de información sensorial del animal, entre ellas información olfativa
(Eichenbaum et al., 1987), visual (O’keefe & Speakman, 1987) o táctil (Young et al., 1994;
Pereira et al., 2007).
En este estudio se encontró que una lesión unilateral e irreversible de las ramas bucal y
mandibular del nervio facial produjo una parálisis facial del tercio inferior de la cara en todos
los grupos de animales lesionados. Esta parálisis facial unilateral se presentó con retracción
e imposibilidad de movimiento de las vibrisas del lado lesionado y se mantuvo durante todo

60
el periodo post-operatorio para cada grupo experimental: 1D (1 día), 3D (3 días), 7D (7 días)
o 21D (21 días). El grupo control Sham, al cual se le realizó una falsa cirugía, no presentó
parálisis facial ni retracción de las vibrisas.
Durante el transcurso de todos los experimentos, se monitoreó el peso de los animales con el
fin de descartar la malnutrición como un factor que pudiera estar modificando la respuesta
electrofisiológica de la sinapsis comisural CA3-CA1 del hipocampo. No se reportaron
animales con un descenso constante de peso tras la cirugía; sin embargo, sí se observó una
disminución en la tasa a la cual los animales de todos los grupos experimentales aumentaban
de peso en el periodo post-operatorio. Este cambio fue significativo en los grupos 3D y 7D.
Se debe advertir que como era de esperarse todos los animales disminuyeron su peso el
primer día tras la cirugía y que los animales del grupo 1D fueron registrados justo en esta
ventana temporal de bajo peso (Figura 8).
A través del análisis de las curvas entrada-por-salida encontramos que la transmisión
sináptica no presentó diferencias entre los grupos experimentales producto de la lesión del
nervio facial (Figura 11).
5.1 Facilitación por pulsos emparejados en condiciones basales.
Se estudió la plasticidad a corto plazo mediante un protocolo de estimulación por pulsos
emparejados y se encontró que todos los animales presentaron un aumento de la amplitud del
fEPSP de la segunda respuesta con respecto a la primera. Normalmente, el cociente entre
pulsos emparejados es tomado como una relación inversa de la probabilidad de liberación
vesicular pre-sináptica, por lo que a un mayor valor R2/R1, una menor probabilidad de
liberación y viceversa (Debanne et al., 1996; Zucker & Regehr, 2002). Con base en esto,
todos los animales presentaron una facilitación por pulsos emparejados, sin embargo, los
animales que tuvieron un tiempo de recuperación post-lesión de 3 y 7 días presentaron un
deterioro significativo de dicha facilitación con respecto a los animales control (Figura 12a
y 12b). Se observó, además, que los animales con 21 días de reposo post-lesión recuperaron
los valores de la facilitación con relación a los animales del grupo control. Por estos motivos
consideramos que 1) tras una lesión del nervio facial se produce un aumento transitorio de la

61
probabilidad de liberación vesicular y 2) que este aumento de la liberación vesicular retorna
a valores control hacia la tercera semana post-lesión.
Para la sinapsis CA3-CA1 se ha reportado que la liberación vesicular pre-sináptica necesita
de canales de calcio dependientes de voltaje (VGCC) de tipo P/Q y N (Qian & Noebels, 2000;
Ahmed & Siegelbaum, 2009). Se sabe también que la facilitación en las neuronas piramidales
de la región CA3 del hipocampo disminuye bajo un bloqueo irreversible de los receptores de
rianodina, por lo que se cree que tras la entrada de calcio extracelular hay una liberación de
calcio por parte de reservas intracelulares lo que aumenta la probabilidad de liberación
vesicular (Emptage et al., 2001), sin embargo este fenómeno aún está siendo estudiado
(Carter et al., 2002).
En esta sinapsis se ha reportado además que la hormona corticosterona logra disminuir la
facilitación por pulsos emparejados y aumentar la frecuencia, pero no la amplitud de
mEPSCs, debido en parte a la activación no-genómica de receptores de mineralocorticoides
(MR) en la terminación pre-sináptica (Karst et al., 2005). Esto es para nosotros de especial
interés puesto que hemos reportado que la lesión del nervio facial aumenta los niveles
plasmáticos de corticosterona producto de la lesión (Patarroyo et al., 2017). Con respecto a
los efectos de la corticosterona sobre las corrientes de calcio en la región hipocampal, se ha
evidenciado que las corrientes de calcio mediadas por VGCC de tipo L en la región CA1
aumentan en presencia de dicha hormona (Chameau et al., 2007).
Por estas razones consideramos que la lesión del nervio facial podría estar modificando la
eficacia de la liberación vesicular a través de cambios en las dinámicas de calcio intracelular
pre-sináptico como producto de un cambio en la conductancia de los VGCCs y que la
activación de MR puede estar mediando este proceso.
Respecto de la facilitación o depresión por pulsos emparejados, se sabe que estas pueden
surgir debido a cambios post-sinápticos. Se ha reportado en cultivos celulares hipocampales
que el anclaje de receptores AMPA post-sinápticos mediado por la activación de CaMKII
promueve la depresión por pulsos emparejados (Opazo et al., 2010), por lo que los resultados
obtenidos mediante este protocolo de estimulación deben ser tomados con precaución ya que
pueden ser explicados por cambios tanto a nivel pre-sináptico como post-sináptico.

62
5.2 Plasticidad a largo plazo.
Procedimos luego a analizar el efecto de la lesión del nervio facial en la plasticidad a largo
plazo de la sinapsis CA3-CA1 mediante un protocolo de estimulación tetánica. Encontramos
que, aunque todos los animales potenciaron su respuesta con respecto a la línea de base, los
animales con 3, 7 y 21 días de recuperación post-lesión presentaron un deterioro significativo
con respecto a los animales control a los que se les realizó una falsa cirugía (Figura 13).
En los animales lesionados pudimos observar que la LTP se indujo y se mantuvo en el tiempo
puesto que todos los animales potenciaron su respuesta y mantuvieron esta potenciación hasta
por una hora tras la estimulación tetánica. Sin embargo, encontramos que la expresión de la
LTP se veía afectada por la lesión del nervio facial. En tal sentido, describimos que la
potenciación a largo plazo (porcentaje de cambio respecto de la línea de base) de los animales
lesionados es significativamente menor que la del grupo control. Este deterioro de la
expresión de la LTP se dio de manera gradual, manifestándose desde el primer día post-lesión
y haciéndose máxima el tercer día post-lesión, donde se observó el menor aumento en el
porcentaje de cambio. Se observó, además, una leve recuperación hacia la tercera semana
post-lesión pero sin alcanzar los valores del grupo control (Figura 13).
La sinapsis CA3-CA1 presenta varias formas de plasticidad a largo plazo y que varían de
acuerdo al tipo de estimulación empleada. De manera general se ha establecido que la
inducción y expresión de la LTP en esta sinapsis se lleva a cabo exclusivamente en la neurona
post-sináptica, siendo su expresión mediada por receptores AMPA (Citri & Malenka, 2008).
Sin embargo esto se ha descubierto a través de protocolos de estimulación que involucran
solo un tren de estimulación a 50 o 100 Hz (Nicoll & Malenka, 1999). Varios grupos de
investigación han encontrado que al inducir la LTP mediante varios trenes de estimulación
de 100 Hz, un solo tren de 200 Hz o un protocolo TBS, se hace evidente un componente pre-
sináptico adicional al post-sináptico (Yang & Calakos, 2013; Enoki et al., 2009; Zakharenko
et al., 2003).
Dado que en nuestra investigación empleamos varios trenes de estimulación de 100 Hz,
decidimos analizar el cambio en la relación entre pulsos pareados tras la estimulación de alta

63
frecuencia (HFS) y observar, de esta manera, si con nuestro protocolo de inducción la LTP
presentaba un componente pre-sináptico. Encontramos que la facilitación por pulsos
emparejados disminuye significativamente con respecto a los valores de la línea de base en
los animales del grupo control y en menor medida en aquellos que tuvieron 3 días de
recuperación post-lesión. Esta disminución de la facilitación no se observó en los animales
que contaron con 1 y 7 días de recuperación. Los animales con 21 días presentaron un
pequeño aumento en la relación de su facilitación (Figura 12). Con base en esto inferimos
que los animales control y en menor medida los animales del grupo 3D aumentan su
probabilidad de liberación vesicular tras un protocolo de estimulación tetánica, cosa que no
sucede con los animales de los grupos 1D, 7D y 21D. Dado que todos los grupos presentaron
una facilitación por pulsos emparejados significativamente menor al grupo SHAM creemos
que la lesión del nervio facial deteriora el componente pre-sináptico de la LTP, esto sin
descartar una explicación post-sináptica a través de cambios en las concentraciones de
receptores AMPA.
Debido a que la inducción de la LTP en la sinapsis CA3-CA1 requiere de la activación de
receptores de NMDA post-sinápticos, se ha especulado que una expresión de la LTP en la
neurona pre-sináptica debe de emplear una señal retrograda producto de la activación post-
sináptica de los receptores NDMA. Hasta el momento no se conoce con exactitud que
molécula que pueda estar mediando este proceso. Sin embargo, hay varias moléculas que se
han visto involucradas en la regulación de la sinapsis CA3-CA1, entre ellas la molécula
gaseosa óxido nítrico (Stanton et al., 2003) o la proteína Wnt 7 (Tabatadze et al., 2014).
Hay escasa información acerca de las consecuencias sobre la plasticidad en la sinapsis
comisural CA3-CA1 tras una lesión de nervio periférico. Ren et al., 2011 demostraron que,
bajo un paradigma de dolor neuropático, producto de una lesión del nervio ciático, la sinapsis
CA3-CA1 presenta un deterioro de la plasticidad a largo y a corto plazo, las cuales fueron
estudiadas mediante un protocolo de estimulación de alta frecuencia (100Hz, 50 pulsos, 4
trenes a intervalos de 15s) y de facilitación por frecuencia, respectivamente. En este estudio,
el deterioro de la LTP estaba mediado por la expresión de la proteína TNF-α.
Adicionalmente, se reportó una disminución de la expresión de la proteína sinaptofisina en
la región CA1 del hipocampo en los animales lesionados.

64
Si bien el modelo de lesión de nervio periférico de esta investigación y el usado por Ren y
colaboradores varía sustancialmente, en especial al tratarse de dos nervios con funciones
motoras y rutas sinápticas diferentes, consideramos que es importante prestar atención al
aumento de la expresión de TNF-α ya que podría ser un mecanismo de señalización
molecular compartido en el deterioro de la LTP producto de una lesión de nervio periférico.
¿Cómo afecta entonces la lesión del nervio facial la plasticidad a largo plazo la sinapsis CA3-
CA1 del hipocampo? En esta investigación hemos discutido que la lesión irreversible del
nervio facial afecta la plasticidad a largo plazo en el hipocampo probablemente a través de
dos eventos fisiológicos: 1) la modificación en el flujo de información en el circuito
sensoriomotor que se genera como consecuencia de la imposibilidad del batido activo de las
vibrisas; y 2) el estrés al que se ven sometidos los animales lesionados al no poder realizar
dicho comportamiento. Es probable, además, que células gliales como los astrocitos y la
microglía sirvan como intermediarios de los cambios funcionales y estructurales que se
puedan estar presentando en la sinapsis CA3-CA1 del hipocampo.
El flujo de información en el sistema somatomotor de las vibrisas en roedores ha sido
estudiado principalmente bajo los modelos de poda de vibrisas y de lesión del nervio facial.
Se ha reportado, por ejemplo, que tras la poda de una columna o de todo el parche de vibrisas,
los barriletes en el núcleo SpVi del nervio del trigémino y los barreloides en el núcleo VPM
del tálamo, los cuales se encuentran privados de entrada sensorial, presentan una alteración
en las propiedades de sus campos receptivos (Dolan & Cahusac, 2007; Jacquin et al., 1995).
En la corteza somatosensorial se ha evidenciado que la poda de las vibrisas produce una
reorganización de los patrones de disparo en varias capas de los barriles en S1 (Rema et al.,
2003; Jacob et al., 2016). Se ha visto, además, que una privación sensorial proveniente de las
vibrisas como producto de su poda bilateral, logra afectar la plasticidad y las propiedades de
la sinapsis CA3-CA1 en el hipocampo de ratones juveniles (Milshtein-Parush et al., 2017),
esto a través de una disminución de la excitabilidad en neuronas de la región CA3, una
disminución de la probabilidad de liberación pre-sináptica, cambios en las proporciones de
las subunidades de receptores NMDA y cambios en la proporción de receptores
NMDA/AMPA. En nuestro grupo de investigación hemos caracterizado cambios
morfológicos y funcionales en las neuronas piramidales de la capa V de la corteza motora

65
primaria (Munera et al., 2012; Urrego et al., 2015). Por último, se ha evidenciado que la
información táctil proveniente de las vibrisas de la rata logra modificar la actividad sináptica
de las neuronas de la región CA1 del hipocampo, esto a través del núcleo VPM del tálamo y
de la corteza S1 (Pereira et al., 2007).
Con respecto al estrés, nuestro grupo de investigación ha descrito que un evento
emocionalmente demandante, como lo es la lesión del nervio facial y su consecuente parálisis
facial, conlleva un aumento de los niveles de corticosterona en plasma de los animales
lesionados bilateralmente. (Patarroyo et al., 2017). La corticosterona y su interacción con los
receptores GR se ha visto involucrada en alteraciones neuronales morfológicas y funcionales.
En el hipocampos, morfológicamente se ha evidenciado que el estrés crónico producto de un
protocolo de restricción de movimiento (3 semanas) promueve la retracción dendrítica en la
región CA3 del hipocampo y cambios en la densidad y forma de las espinas dendríticas tanto
en CA3 como en CA1 (Magariños et al., 2011). De manera funcional se sabe que el estrés
comportamental y fisiológico afecta la LTP en la sinapsis CA3-CA1 (Foy et al., 1987; Artola
et al., 2006, Xu et al., 1997).
Finalmente, se ha reportado que las células gliales presentan cambios funcionales y
morfológicos tras una la lesión del nervio facial (Moran & Graeber, 2004), bajo un paradigma
de envejecimiento y deterioro de la percepción sensorial (Tremblay et al., 2012) o estrés
fisiológico y comportamental (Walker et al., 2013).
Para la microglía se ha reportado que en su estado de reposo expande y contrae sus procesos
celulares, vigilando activamente el estado de las sinapsis circundantes, haciendo breves
contactos de hasta por 5 minutos cada hora. La duración de dicho contacto puede aumentar
bajo un estado de insulto fisiológico, seguido de un desnudamiento sináptico en aquellas
neuronas con las que la microglía mantuvo contacto. Esto ha abierto la puerta a
especulaciones acerca de la participación de la microglía en procesos de plasticidad sináptica
puesto que se posiciona como una célula capaz de monitorear y responder a cambios en el
estado de la sinapsis (Wake et al., 2009; Eroglu & Barres, 2010).
En el hipocampo se ha observado que en presencia de la microglía, la frecuencia de las
mEPSCs disminuye en neuronas de la región CA1, así como la expresión de la sub-unidad

66
GluA1 del receptor AMPA (Ji et al., 2013). Se ha visto además que la microglía al ser
activada mediante lipopolisacárido, libera ATP y recluta astrocitos a través de sus receptores
purinérgicos P2Y1. Los astrocitos a su vez liberan glutamato, activando receptores
metabotrópicos de glutamato (mGluR5) lo que repercute en un aumento de la frecuencia de
las mEPSCs (Pascual et al., 2012). Esta liberación de glutamato por parte de los astrocitos y
la consecuente activación de mGluRs se ha visto relacionada con modificaciones en la
plasticidad sináptica a corto plazo de la sinapsis CA3-CA1 del hipocampo, a través de un
aumento de la probabilidad de liberación vesicular (Perea & Araque, 2007). En nuestro grupo
de investigación se encontró que tras una lesión del nervio facial, la microglía retrae sus
procesos celulares y adopta una morfología ameboidea, lo que se relaciona con un proceso
de activación microglial (Urrego et al., 2015), además, la proteína astrocítica GFAP aumenta
su expresión en el hipocampo (Troncoso et al., 2015).
Por todo lo anterior creemos que la glía tiene la capacidad de percibir cambios en el flujo de
la información táctil hacia el hipocampo o un evento fisiológicamente estresante como
producto de la lesión del nervio facial. Esta activación glial tiene la capacidad de afectar la
sinapsis hipocampal CA3-CA1 tanto en la neurona post-sináptica, como en la pre-sináptica,
a través de modificación en los receptores AMPA o mediante cambios en la probabilidad de
liberación vesicular.

67
6. CONCLUSIONES
1. La trasmisión basal de la sinapsis comisural CA3-CA1 del hipocampo no se vio
comprometida en animales con una lesión irreversible de las ramas bucal y
mandibular del nervio facial.
2. Aunque hubo facilitación por pulsos emparejados en los animales con lesión del
nervio facial, este fenómeno fue significativamente menor 3 y 7 días después de la
lesión en comparación con los animales control.
3. La estimulación de alta frecuencia en animales sin lesión del nervio facial provoca
potenciación a largo plazo de la sinapsis comisural CA3-CA1, con componentes pre
y post sinápticos. En todos los animales lesionados, se observó una disminución de
la potenciación a largo plazo a expensas del componente pre-sináptico,
principalmente.

68
PERSPECTIVAS
Los resultados de este trabajo de investigación permiten plantear varios experimentos a
futuro enfocados a esclarecer los mecanismos por los cuales se llevan a cabo modificaciones
a nivel del SNC tras lesión en nervios periféricos.
En primer lugar, podrían abordarse los experimentos con lesiones del nervio facial de tipo
reversible, para poder comparar la evolución de las modificaciones en la plasticidad a corto
y a largo plazo en el hipocampo, cuando hay la posibilidad de recuperación funcional tras la
lesión.
Además, sería de gran ayuda contar con los valores de los niveles de corticosteroides en la
sangre de los animales a diferente días post-operatorios, esto con el fin de corroborar el estado
del estrés fisiológicos tras la lesión del nervio facial a lo largo del tiempo post-operatorio. Si
queremos corroborar el efecto causal del estrés fisiológico en los cambios observados en este
estudio sobre la plasticidad sináptica, seria provechoso administrar antagonistas de los
receptores GR, como por ejemplo mifepristona, en paralelo con la lesión del nervio facial.
Resulta interesante también observar los efectos de la inhibición y activación de las células
gliales, esto con miras a establecer si la activación glial estaría jugando un papel protagónico
en los cambios en la plasticidad sináptica aquí reportados. Para la microglía existen varias
estrategias farmacológicas usadas para su activación, como por ejemplo lipopolisacárido
(LPS), o para su inhibición como por ejemplo la minociclina (Peña-Ortega, 2017). Es además
interesante detallar los patrones de expresión de ciertas citoquinas pro-inflamatorias liberadas
por la glía como Il-1 o TNF-α.
Finalmente, si se quiere ahondar en los mecanismos moleculares que podrían verse afectados
en la sinapsis CA3-CA1 del hipocampo tras la LNF, la marcación y cuantificación de los
receptores glutamatérgicos como AMPA o NMDA y de los canales VGCC es de vital
importancia.

69
7. BIBLIOGRAFÍA
1. Ahl, A. 1986. The role of vibrissae in behavior: A status review. Vet Res Commun
10 245.
2. Ahmed, M., Siegelbaum, S. 2009. Recruitment of n-type Ca+2 channels during LTP
enhances low release efficacy of hippocampal CA1 perforant path synapses. Neuron.
63, 372-385.
3. Aldskogius, H., Kozlova, N. 1997. Central neuron-glial and glial-glial interactions
following axon injury. Progress in Neurobiology. 55, 1-26.
4. Aldskogius, H., Thomander, L. 1986. Selective reinnervation of somatotopically
appropriate muscles after facial nerve transection and regeneration in the neonatal rat.
Brain. Res. 375, 126-134.
5. Amaral, D., Witter, M. 1989. The three-dimensional organization of the hippocampal
formation: A review of anatomical data. Neuroscience. 31(3), 571-591.
6. Anderson, M., Burda, J., Ren, Y., Ao, Y., O’shea, T., Kawaguchi, R., Copola, G.,
Khakh, B., Deming, T., Sofroniew, M. 2016. Astrocyte scar formation aids central
nervous system axon regeneration. Nature. 532, 195-200.
7. Artola, A., von Frijtag, J., Fermont, P., Gispen, W., Schrama, L., Kamal, A., Spruijt,
B. 2006. Long-lasting modulation of the induction of LTP and LTD in rat
hippocampal CA1 by behavioral stress and environmental enrichment. Eur J
Neurosci. 23, 261-272.
8. Barria, A., Muller, D., Derkach, V., Griffith, L., Soderling, T. 1997. Regulatory
phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term
potentiation. Science. 276, 2042-2045.
9. Belford, G., Killackey, H. 1979. Vibrissae representation in subcortical trigeminal
centers of the neonatal rat. J Comp Neurol. 183: 305-321.
10. Blinzinger, K., Kreutzberg, G. 1968. Displacement of synaptic terminals from
regenerating motoneurons by microglial cells. Z. Zellforsch. 85(2), 145-157.

70
11. Bliss, T., Lomo, T. 1973. Long-Lasting Potentiation of Synaptic Transmission in the
Dentate Area of the Anesthetized Rabbit following Stimulation of the Perforant Path.
J. Physiol. 232, 331-356.
12. Bosman, L., Houweling, A., Owens, C., Tanke, N., Shevchouk, O., Rahmati, N.,
Teunissen, W., Ju, C., Gong, W., Koekkoek, S., Zeeuw, C. 2011. Anatomical
pathways involved in generating and sensing rhythmic whisker movements. Frontiers
in Integrative Neuroscience. 5(53).
13. Brecht, M., Preilowski, B., Merzenich, M. 1997. Functional architecture of the
mystacial vibrissae. Behavioral Brain Research. 84: 81-97.
14. Bureau, I., Von Saint Paul, F., Svoboda, K. 2006. Interdigitated paralemniscal and
lemniscal pathways in the mouse barrel cortex. PLoS Biol. 4(12) e382.
15. Carter, A., Vogt, K., Foster, K., Regehr, W. 2002. Assesing the role of calcium-
induced calcium release in short-term presynaptic plasticity at excitatory central
synapses. J Neurosci. 22(1), 21-28.
16. Chameau, P., Spijker, S., Smit, A., Joëls, M. 2007. Glucocorticoids specifically
enhance L-type calcium current amplitude and effect calcium channel subunit
expression in the mouse hippocampus. 97, 5-14.
17. Citri, A., Malenka, R. 2008. Synaptic Plasticity: Multiple Forms, Functions and
Mechanisms. Neuropsycopharmacology. 33, 18-41.
18. Cramer, N., Li, Y., Keller, A. 2007. The whisking rhythm generator: a novel
mammalian network for the generation of movement. J Neurophysiol. 97, 2148-2158.
19. Debanne, D., Guérineau, N., Gähwiler, B., Thompson, S. 1996. Paired-pulse
facilitation and depression at unitary synapses in rat hippocampus: quantal
fluctuations affects subsequent release. Journal of Physiology. 491(1), 163-176.
20. Del Rio-Hortega, P. 1932. “Microglia” En: Cytology and cellular pathology of the
nervous system. (New York: Hoeber) 482-534.
21. Derkach, V., Barria A., Soderling T. 1999. Ca+2/calmodulin-kinase II enhances
channel conductance of α-amino-3-hydroxy-5-methyl-isoxazolepropionate type
glutamate receptors. Proc. Natl. Acad. Sci. 96, 3269-3274.
22. Derkach, V. 2003. Silence analysis of AMPA receptor mutated at the CaM-Kinase II
phosphorylation site. Biophysical Journal. 84, 1701-1708.

71
23. Deschênes, M., Veinante, P., Zhang, W. 1998. The organization of corticothalamic
projections: reciprocity versus parity. Brain res rev. 28, 286-308.
24. Deschênes, M., Takatoh, J., Kurnikova, A., Moore, J., Demers, M., Elbaz, M., Furuta,
T., Wang, F., Kleinfeld, D. 2016. Inhibition, not excitation, drives rhythmic whisking.
Neuron. 90, 374-387.
25. Dolan, S., Cahusac, P. 2007. Enhanced short-latency responses in the ventral
posterior medial (VPM) thalamic nucleus following whisker trimming in adult rat.
Physiology and Behavior. 92, 500-506.
26. Donoghue, J., Suner, S., Sanes, J. 1990. Dynamic organization of primary motor
cortex output to target muscles in adult rats II. Rapid reorganization following motor
nerve lesions. Exp. Brain. Res. 79, 492-503.
27. Doyére, V., Laroche, S. 1992. Linear relationship between the maintenance of
hippocampal long-term potentiation and retention of an associative memory.
Hippocampus. 2, 39-48.
28. Dörfl, J. 1982. The musculature of the mystacial vibrissae of the white mouse. J. Anat.
135: 147-154.
29. Ebbesen, C., Doron, G., Lenschow, C., Brecht, M. 2016. Vibrissa motor cortex
activity suppresses contralateral whisking behavior. Nature Neuroscience.
30. Ebara, S., Kumamoto, K., Matsuura, T., Mazurkiewicz, J., Rice, F. 2002. Similarities
and differences in the innervation of mystacial vibrissal follicle-sinus complexes in
the rat and cat: a confocal microscopy study. J. Comp. Neurol. 449, 103-119.
31. Eichenbaum, H., Kuperstein, M., Fagan, A., Nagode, J. 1987. Cue-sampling and
goal-approach correlates of hippocampal unit activity in rats performing and odor-
discrimination task. The Journal of Neuroscience. 7(3), 716-732.
32. Eichenbaum, H., Dudchencko, P., Wood, E., Shapiro, M., Tanila, H. 1999. The
hippocampus, memory and place cells: Is it spatial memory or a memory space?
Neuron. 23, 209-226.
33. Emptage, N., Reid, C., Fine, A. 2001. Calcium store in hippocampal synaptic boutons
mediate short-term plasticity, store-operated Ca+2 entry, and spontaneous transmitter
release. Neuron. 29(1), 197-208.

72
34. Enoki, R., Hu, Y., Hamilton, D., Fine, A. 2009. Expression of long-term plasticity at
individual synapses in hippocampus is graded, bidirectional, and mainly presynaptic:
Optical quantal analysis. Neuron. 62, 242-253.
35. Eroglu, C., Barres, B. 2010. Regulation of synaptic connectivity by glia. Nature. 468,
223-231.
36. Farkas, T., Perge, J., Kis, Z., Wolff, J., Toldi, J. 2000. Facial nerve injury-induced
disinhibition in the primary motor cortices of both hemispheres. Eur J Neurosci. 12,
2190-2194.
37. Fawcett, J., Keynes, R. 1990. Peripheral Nerve Regeneration. Annu Rev Neurosci.
13, 43-60.
38. Fioravante, D., Chu, Y., Myoga, M., Leitges, M., Regehr, W. 2011. Calcium-
dependent isoforms of protein kinase C mediate posttetanic potentiation at the calix
of held. Neuron. 70, 1005-1019.
39. Foy, M., Stanton, M., Levine, S., Thompson, R. 1987. Behavioral stress impairs long-
term potentiation in rodent hippocampus. Behavioral and Neural Biology. 48, 138-
149.
40. Franchi, G., Maggiolini, E., Muzzioli, V., Guandalini, P. 2006. The vibrissal motor
output following severing and repair of the facial nerve in the newborn rat reorganises
less than in the adult. Euro. J. Neurosci. 26, 1547-1558.
41. Friedberg, M., Lee, S., Ebner, F. 2004. The contribution of the principal and spinal
trigeminal nuclei in the receptive field properties of thalamic VPM neurons in the rat.
J. Neurocytol. 33: 75-85.
42. Fujii, S., Mikoshiba, K., Kuroda, Y., Ahmed, T., Kata, H. 2003. Cooperativity
between activation of metabotropic glutamate receptor and NMDA receptors in the
induction of LTP in hippocampal CA1 neurons. Neuroscience Research. 46, 509-521.
43. Fukunaga, K., Stoppini, L., Miyamoto, E., Muller, D. 1993. Long-term potentiation
is associated with an increased activity of Ca+2/Calmodulin-dependent protein kinase
II. The Journal of Biological Chemistry. 268(11), 7863-7867.
44. Galiano, M., Liu, Z., Kalla, R., Bohatschek, M., Koppius, A., Gschwendthner, A.,
Xu, S., Werner, A., Kloss, C., Jones, L., Bluethmann, H., Raivich, G. 2001.
Interleukin-6 (IL6) and cellular response to facial nerve injury: effects on lymphocyte

73
recruitment, early microglial activation and axonal outgrowth in IL6-deficient mice.
Eur. J. Neurosci. 14, 327-341.
45. Gao, P., Hattox, A., Jone, L., keller, A., Zeigler, H. 2003. Whisker motor cortex
ablation and whisker movement patterns. Somatosens. Mot. Res. 20: 191-198.
46. Geppert, M., Hammer, R., Li, C., Rosahl, T., Südhof, T. 1994. Synaptotagmin I: a
major Ca+2 sensor transmitter release at central synapse. Cell. 79(4), 717-727.
47. Graeber, M., Kreutzberg, G. 1988. Delayed astrocyte reaction following facial nerve
axotomy. Journal of Neurocytology. 17, 209-220.
48. Graeber, M., Raivich, G., Kreutzberg, G. 1989. Increase of transferrin receptors and
iron uptake in regenerating motor neurons. J. Neurosci. Res. 23, 342-345.
49. Grinevich, V., Brecht, M., Osten, P. 2005. Monosynaptic pathway from rat vibrissa
motor cortex to facial motor neurons revealed by lentivirus-based axonal tracing. The
Journal of neuroscience. 25(36), 8250-8258.
50. Griffin, R., Nally, R., Nolan, Y., McCartney, Y., Linden, J., Lynch, M. 2006. The
age-related attenuation in long-term potentiation is associated with microglial
activation. Journal of Neurochemestry. 99, 1263-1272.
51. Grion, N., Akrami, A., Zuo, Y., Stella, F., Diamond, M. 2016. Coherence between rat
sensorimotor system and hippocampus is enhanced during tactile discrimination.
PLoS Biol. 14(2).
52. Groenewegen, H., Witter, M. 2004 “Thalamus”, en Rat Nervous System, ed. G.
Paxinos, 3era Edn (Sydney: Academic Press), 408-441.
53. Haidarliu, S., Simony, E., Golomb, D., Ahissar, E. 2010. Muscle architecture in the
mystacial pad of the rat. Anat. Rec. 293: 1192-1206.
54. Hass, C., Donath, C., Kreutzberg, G. 1993. Differential expression of immediate early
genes after transection of the facial nerve. Neuroscience. 91-99
55. Hass, C., Dumoulin F., Lazar, P., Raivich, G., Reddington, M., Streit, W., Kreutzberg,
G. 1994. The role of calcitonin gene-related peptide in the regenerating facial nucleus.
Eur. Arch. Otorhinolaryngol. 71-72.
56. Hebb, D. 1949. The organization of behavior. Wiley.

74
57. Henstrom, D., Hadlock, T., Lindsay, R., Knox, C., Malo, J., Vakharia, K., Heaton, J.
2011. The convergence of facial nerve branches providing whisker pad motor supply
in rats: Implication for facil reanimation study. Muscle Nerve. 45, 692-697.
58. Hinrichsen, C., Watson, C. 1984. The facial nucleus of the rat: Representation of
facial muscles revealed by retrograde transport of horseradish peroxidase. 209(3),
407-415.
59. Herfst, L., Brecht, M. 2008. Whisker movements evoked by stimulation of single
motor neurons in the facial nucleus of the rat. J. Neurophysiol. 99: 2821-2832.
60. Isaac, J., Nicoll, R., Malenka, R. 1995. Evidence of silent synapses: Implications for
the expression of LTP. Nature. 15, 427-434.
61. Ishisuka, N., Weber, J., Amaral, D. 1990. Organization of intrahippocampal
projections originating from CA3 pyramidal cells in the rat. JCN. 295, 580-623.
62. Izraeli, R., Porter, LL. 1995. Vibrissal motor cortex in the rat: Connections with the
barrel field. Exp. Brain. Res. 104, 41-54.
63. Jacob, V., Mitani, A., Toyoizumi, T., Fox, K. 2016. Whisker row deprivation affects
the flow of sensory information through rat barrel cortex. J Neurophysiol. 117, 4-17.
64. Jacquin, M., Rhoades, R., Klein, B. 1995. Structure-function relationship in rat
brainstem subnucleus interpolaris. XI. Effects of chronic whisker trimming from
birth. J Comp Neurol. 356, 200-224.
65. Ji, K., Akgul, G., Wollmuth, L., Tsirka, S. 2013. Microglia actively regulate the
number of functional synapses. PLOS ONE. 8(2), 1-12.
66. Johnson, I., Duberley, R. 1998. Motoneuron survival and expression of neuropeptides
and neurotrophic factor receptors following axotomía in adult and ageing rats.
Neuroscience. 84, 141-150.
67. Jurado, S., Goswami, D., Zhang, Y., Molina, A., Südhof, T., Malenka, R. 2013. LTP
requires a unique postsynaptic SNARE function machinery. Neuron. 77, 542-
68. Karst, H., Berger, S., Turiault, M., Tronche, F., Schütz, G., Joëls, M. 2005.
Mineralocorticoid receptors are indispensable for nongenomic modulation of
hippocampal glutamate transmission by corticosterone. PNAS. 102(52), 19204-
19207.

75
69. Katz, B., Miledi, R. 1968. The role of calcium in neuromuscular facilitation. J.
Physiol. 195, 481-492.
70. Kauer, J., Malenka, R., Nicoll, R. 1988. A persistent postsynaptic modification
mediates long-term potentiation in the hippocampus. Neuron. 1, 911-917.
71. Kerr, F., Lysak, W. 1964. Somatotopic organization of trigeminal-ganglion neurons.
Arch. Neurol. 11: 593-602.
72. Kessels, H., Malinow, R. 2009. Synaptic AMPA Receptor Plasticity and Behavior.
Neuron. 61, 340-350.
73. Kettenmann, H., Kirchhoff, F., Verkhratsky, A. 2012. Microglia: New Roles for the
Synpatic Stripper. Neuron 77.
74. Kiefer, R., Lindholm, D., Kreutzberg, G. 1993. Interleukin-6 and transforming
growth factor-ß1 mRNAs are induced in rat facial nucleus following motoneuron
axotomy. European Journal of Neuroscience 5: 775-781.
75. Killackey, H., Sherman, S. 2003. Corticothalamic projections from the rat primary
somatosensory cortex. J Neurosci. 23: 7381-7384.
76. Kim, J., Diamond, D. 2002. The stressed hippocampus, synaptic plasticity and lost
memories. Neuroscience. 3, 453-462.
77. Klein, B., Rhoades, R. 1985. Representation of whisker follicle intrinsic musculature
in the facial motor nucleus of the rat. J Comp Neurol. 232, 55-69.
78. Knierim, J. 2015. The hippocampus. Current Biology. 25, 1116-1121.
79. Kreutzberg, G. 1986. Neurobiology of regeneration and degeneration. The Facial
Nerve. Thieme. 75-83.
80. Kreutzberg, G. 1996. Microglia: a sensor for pathological events in the CNS. Trends
Neurosci. 19, 312-318.
81. Krout, K., Belzer, R., Loewy, A. 2002. Brainstem projections to midline and
intralaminar thalamic nuclei of the rat. Journal of Comparative Neurology. 448(1),
53-101.
82. Land, P., Buffer, S., Yaskosky, J. 1995. Barreloids in adult rat thalamus: three-
dimensional architechture and relationship to somatosensory cortical barrels. The
Journal of Comparative Neurology. 355: 573-588.

76
83. Larson, J., Wong, D., Lynch, G. 1986. Patterned stimulation at the theta frequency is
optimal for the induction of hippocampal long-term potentiation. Brain Res. 368, 347-
350.
84. Leiser, S., Moxon, K. 2007. Responses of trigeminal ganglion neurons during natural
whisking behaviors in the awaken rat. Neuron. 53: 117-133.
85. Magariños, A., Li, C., Gal Toth, G., Bath, K., Jing, D., Lee, F., McEwen, B. 2011.
Effect of brain-derived neurotrophic factor haploinsufficiency on stress-induced
remodeling of hippocampal neurons. Hippocampus. 21(3), 253-264.
86. Makino, H., Malinow, R. 2009. AMPA receptor incorporation into synapses during
LTP: The role of lateral movement and exocytosis. Neuron. 64, 381-390.
87. Malinow, R., Tsien, R. 1990. Presynaptic enhancement shown by whole-cell
recordings of long-term potentiation in hippocampal slices. Nature. 346, 177-180.
88. Malenka, R., Kauer, J., Zucker, R., Nicoll, R. 1988. Postsynaptic calcium is sufficient
for potentiation of hippocampal synaptic transmission. Science. 242, 81-84.
89. Mattsson, P., Meijer, B., Svensson, M. 1998. Extensive neuronal cell death following
intracranial transection of the facial nerve in adult rat. Brain Res Bulletin. 49(5), 333-
341.
90. Matyas, F., Sreenivasan, V., Marbach, F., Wacongne, R., Barsy, B., Mateo, C.,
Aronoff, R., Petersen, C. 2010. Motor control by sensory cortex. Science. 330, 1240-
1243.
91. Matzusaki, M., Honkura, N., Ellis-Davies, G., Kasai, H. 2004. Structural basis of
long-term potentiation in single dendritic spines. Nature. 429, 761-766.
92. McNamara, R., Jiang, Y., Streit, W., Lenox, R. 2000. Facial motor neuron
regeneration induces a unique spatial and temporal pattern of myristoylated alanine-
rich C kinase susbtrate expression. Neuroscience. 97, 581-589.
93. Mulkey, R., Malenka, R. 1992. Mechanisms underlaying induction of homosynaptic
long-term depression in area CA1 of the hippocampus. Neuron. 9, 967-975.
94. Müller, W., Connor, J. 1991. Dendritic spines as individual neuronal compartment
for synaptic Ca2+ responses. Nature. 354, 73-76.

77
95. Múnera, A., Cuestas, D., Troncoso, J. 2012. Peripheral nerve lesions induce changes
in the firing properties of primary motor cortex layer 5 pyramidal cells. Neuroscience.
223, 140-151.
96. Muñoz, J., M. 2015. Efecto del bexaroteno sobre la plasticidad en la sinapsis
comisural CA3-CA1 en un modelo murino de enfermedad de Alzheimer (Tesis de
maestría). Universidad Nacional de Colombia, sede Bogotá.
97. Moran, L., Graeber, M. 2004. The facial nerve axotomy model. Brain Res Reviews.
44, 154-178.
98. Moore, J., Deschênes, M., Furuta, T., Huber, D., Smear, M., Demers, M., Kleinfeld,
D. 2013. Hierarchy of orofacial rhythms revealed through whisking and breathing.
Nature. 497, 205-210.
99. Moreno, C., vivas, O., Laprea, N., Lamprea, M., Múnera, A., Troncos, J. 2010.
Vibrissal paralysis unveils a preference for textural rather than positional novelty in
the oone-trial object recognition task in rats. Behav Brain Res. 211, 229-235.
100. Morris, R., Garrud, P., Rawlins, J., O’Keefe, J. 1982. Place navigation
impaired in rats with hippocampal lesions. Nature. 297, 681-683.
101. Moser, M., Moser, E., Forrest, E., Andersen, P., Morris, R. 1995. Spatial
learning with a minislab in the dorsal hippocampus. PNAS. 92, 9697-9701.
102. Moser, M., Rowland, D., Moser, E. 2015. Place cells, grid cells and memory.
Cold Spring Harb Perspect Biol. 7:a021808.
103. Nagai, J., Rajbhandari, A., Gangwani, M., Masmanidis, S., Fanselow, M.,
Khakh, B. 2019. Hyperactivity with disrupted attention by activation of an astrocyte
synaptogenic cue. Cell. 177(5), 1280-1292.
104. Nakajima, K., Tohyama, Y., Maeda, S., Kohsaka, S., Kurihara, T. 2007.
Neuronal regulation by which microglia enhance the production of neurotrophic
factors for GABAergic, catecholaminergic and cholinergic neurons. Neurochemistry
International. 50, 807-820.
105. Navarro, X., Vivó, M., Valero-Cabré, A. 2007. Neuronal plasticity after
peripheral nerve Injury and regeneration. Progress in Neurobiology. 82, 163-201.
106. Neniskyte, U., Vilalta, A., Brown, G. 2014. Tumor necrosis alpha-induced
neuronal loss is mediated by microglial phagocytosis. FEBS lett. 588(17), 2952-2956.

78
107. Nguyen, P., Abel, T., Kandel, E. 1994. Requirement of a critical period of
transcription for induction of a late phase of LTP. Science. 265,1104-1107.
108. Nguyen, Q., Kleinfeld, D. 2005. Positive feedback in a brainstem tactile
sensoriomotor loop. Neuron. 45, 447-457.
109. Nicoll, R., Malenka, R. 1999. Expression mechanisms underlying NMDA
receptor-dependent long-term potentiation. Ann. N. Y. Acad. Sci. 868, 515-525.
110. Nicoll, R., Tomita, S., Bredt, D. 2006. Auxiliary subunits assist AMPA-type
glutamate receptors. Science. 311, 1253-1256.
111. Nishiyama, M., Hong, K., Mikoshiba, K., Poo, M., Kato, K. 2000. Calcium
stores regulate the polarity and input specificity of synaptic modification. Nature.
408, 584-588.
112. Noble, J., Munro, C., Prasad, V., Midha, R. 1998. Analysis of the upper and
lower extremity peripheral nerve Injuries in a population of patients with multiple
injuries. J Trauma. 45, 116-122.
113. Norton, S., Culver, B., Mullenix, P. 1975. Development of nocturnal behavior
in albino rats. Behavioral Biology. 15, 317-331.
114. Ocampo, M., Henao, L., Vásquez, L. 2010. Amputación de miembro inferior:
Cambios funcionales, inmovilización y actividad física. Editorial Universidad del
Rosario. Documento de investigación 42.
115. O’Keefe, J. 1976. Place units in the hippocampus of the freely moving rat.
Experimental neurology. 51, 78-109.
116. O’Keefe, J., Speakman, A. 1987. Single unit activity in the rat hippocampus
during a spatial memory task. Exp Brain Res. 68(1), 1-27.
117. Oliveira, A., Thams, S., Lidman, O., Piehl, F., Hokfelt, T., Karre, K., Linda,
H., Cullheim, S. 2004. A role for MHC class I molecules in synaptic plasticity and
regeneration of neurons after axotomy. PNAS. 101(51), 17843-17848.
118. Otmakhov, N., Tao-Cheng, J., Carpenter, S., Asrican, B., Dosemeci, A.,
Reese, T., Lisman, J. 2004. Persistent accumulation of calcium/calmodulina-
dependent protein kinase II in dendritic spines after induction of NMDA receptor-
dependent chemical long-term potentiation. J Neurosci. 24(42), 9324-9331.

79
119. Olton, D., Walker, J., Gage, F. 1978. Hippocampal connections and spatial
discrimination. Brain Res. 139, 295-308.
120. Opazo, P., Labrecque, S., Tigaret, C., Frouin, A., Wiseman, P., De Koninck,
P., Choquet, D. 2010. CaMKII triggers the diffusional trapping of surface AMPARs
through phosphorylation of stargazing. Neuron. 67, 239-252.
121. Pascual, O., Achour, S., Rostaing, P., Triller, A., Bessis, A. 2012. Microglia
activation triggers astrocyte-mediated modulation of excitatory neurotransmission.
PNAS. 109(4), E197-E205.
122. Patarroyo, W., García-Perez, M., Lamprea, M., Múnera, A., Troncoso, J.
2017. Vibrissal paralysis produces increased corticosterona levels and impairment of
spatial memory retrieval. Behav Brain Res. 320, 58-66.
123. Paxinos, G., Watson, C. 2007. The rat brain in stereotaxic coordinates. 3rd
Edition.
124. Peña-Ortega, F. 2017. Pharmacological tools to activate microglia and their
possible use to study neuronal network patho-physiology. Current
Neuropharmacology. 15, 595-619.
125. Perea, G., Araque, A. 2007. Astrocytes potentiate transmitter release at single
hippocampal synapses. Science. 317, 1083-1086.
126. Pereira, A., Ribeiro, S., Wiest, M., Moore, L., Pantoja, J., Lin, S., Nicolelis,
M. 2007. Processing of tactile information by the hippocampus. PNAS. 104(46),
18286-18291.
127. Perry, H., O’Connor, V. 2010. The Role of Microglia in Synaptic Stripping
and Synaptic Degeneration: A revised perspective. ASN Neuro. 2(5), 281-291.
128. Pinganaud, G., Bernat, T., Buisseret, P., Buisseret-Delmas, C. 1999.
Trigeminal projections to hypoglosal and facial motor nuclei in the rat. J Comp
Neurol. 415: 91-104.
129. Qian, J., Noebels, J. 2000. Presynaptic Ca+2 influx at a mouse central synapse
with Ca+2 channel subunit mutations. J. Neurosci. 20(1), 163-170.
130. Raivich, G., Bohatschek, M., Da Costa, C., Iwata, O., Galiano, M., Hristova,
M., Nateri, A., Makwana, M., Riera-sans, L., Wolfer, D., Lipp, H., Aguzzi, A.,

80
Wagner, E., Behrens, A. 2004. The AP-1 transcription factor c-Jun is required for
efficient axonal regeneration. Neuron. 43, 57-67.
131. Rema, V., Armstrong-James, M., Ebner, F. 2003. Experience-dependent
plasticity is impaired in adult rat barrel cortex after whisker are unused in early
postnatal life. J Neurosci. 23, 358-366.
132. Ren, W., Liu, Y., Zhou, L., Li, W., Zhong, Y., Pang, R., Xin, W., Wei, X.,
Wang, J., Zhu, H., Wu, C., Qin, Z., Liu, G., Liu, X. 2011. Peripheral nerve injury
leads to working memory deficits and dysfunction of the hippocampus by up-
regulation of TNF-α in Rodents. Neuropsycopharmacology. 36, 979-992.
133. Roozendaal, B. 2002. Stress and memory: opposing effects of glucocorticoids
on memory consolidation and memory retrieval. Neurobiol Learn Mem. 78(3), 578-
595.
134. Soreide, A. 1981. Variations in the axon reaction in animals of different ages.
Acta anat. 110, 40-47.
135. Semba, K., Egger, M. 1986. The facial “motor” nerve of the rat: Control of
vibrissal movement and examination of motor and sensory components. The Journal
of Comparative Neurology. 247, 144-158.
136. Sanes, J., Suner, S., Donoghue, J. 1990. Dynamic organization of primary
motor cortex output to target muscles in adult rats I. Long-term patterns of
reorganization following motor or mixed peripheral nerve lesions. Exp Brain Res. 79,
479-471.
137. Sanes, J., Wang, J., Donoghue, J. 1992. Inmediate and delayed changes in
motor cortical output representation with new forelimb configurations. Cereb Cortex.
2, 141-152.
138. Scoville, W., Milner, B. 1957. Loss of recent memory after bilateral
hippocampal lesions. J Neurol Neurosurg Psychiatry. 20(1), 11-21.
139. Shohara, E., Sakai, A. 1983. Localization of motoneurons innervating deep
and superficial facial muscles in the rat: A Horseradish peroxidase and
electrophysiologic study. Experimental Neurology. 81, 14-33.

81
140. Svensson, M., Aldskogius, H. 1993. Synaptic density of axotomized
hypoglossal motoneurons following pharmacological blockade of the microglial cell
proliferation. Experimental Neurology. 120, 123-131.
141. Stanton, P., Winterer, J., Bailey, C., Kyrozis, A., Raginov, I., Laube, G., Veh,
R., Nguyen, C., Müller, W. 2003. Long-term depression of presynaptic release from
readily releasable vesicle pool induced by NMDA receptor-dependent retrograde
nitric oxide. J Neurosci. 23(13), 5936-5944.
142. Steward, O. 1976. Topographic organization of the projections from the
entorhinal area to the hippocampal formation of the rat. J Comp Neur. 167, 285-314.
143. Streit, W., Kreutzberg, G. 1988. Response of endogenous glial cells to motor
neuron degeneration induced by toxic ricin. 268, 248-263.
144. Stoltz, S., Humm, J., Schallert, T. 1999. Cortical injury impairs contralateral
forelimbs immobility during swimming: a simple test for loss of inhibitory motor
control. Behav. Brain. Res. 106: 127-132.
145. Sugitani, M., Yano, J., Sugai, T., Ooyama, H. 1990. Somatotopic organization
and columnar structure of vibrissae representation in the rat ventrobasal complex.
Exp Brain Res. 81:346-352.
146. Tabatadze, N., McGonigal, R., Neve, R., Routtenberg, A. 2013. Activity-
dependent Wnt 7 dendritic targeting in hippocampal neurons: Plasticity- and tagging-
related retrograde signaling mechanism? Hippocampus. 24(4), 455-465.
147. Takatoh, J., Prevosto, V., Wang, F. 2017. Vibrissa sensory neurons: linking
distinct morphology to specific physiology and function. Neuroscience. 368, 109-
114.
148. Toldi, J., Laskawi, R., Landgrebe, M., Wolff, J. 1996. Biphasic reorganization
of somatotopy in the primary cortex follows facial nerve lesions in adult rats.
Neurosci. Lett. 203, 179-182.
149. Toni, N., Buchs, P., Nikonenko, I., Bron, C., Muller, D. 1999. LTP promotes
formation of multiple spine synapses between a single axon terminal and dendrite.
Nature. 402, 421-425.
150. Torvik, A., Soreide, A. 1975. The perineuronal glial reaction after axotomy.
Brain Research. 95, 519-529.

82
151. Tremblay, M., Zettel, M., Ison, J., Allen, P., Majewska, A. 2012. Effects of
aging and sensory loss on glial cells in mouse visual and auditory cortices. Glia. 60,
541-558.
152. Troncoso, J., Buriticá, E., Múnera, A. 2015. Peripheral nerve injury induces
glial activation in primary motor cortex. Conference abstract: Latin-American school
on glial cells in the diseased brain (IBRO).
153. Trudeau, L., Emery, D., Haydon, P. 1996. Direct modulation of the secretory
machinery underlies PKA-dependent synaptic facilitation in hippocampal neurons.
Neuron. 17, 789-797.
154. Tsao, A., Moser, M., Moser, E. 2013. Traces of experience in the lateral
entorhinal cortex. Current Biology. 23, 399-405.
155. Tyssowski, K., DeStefino, N., Cho, J., Dunn, C., Poston, R., Carty, C., Jones,
R., Chang, S., Romeo, P., Wurzelmann, M., Ward, J., Andermann, M., Saha, R.,
Dudek, S., Gray, J. 2018. Different neuronal activity patterns induce different gene
expression programs. Neuron. 98, 530-546.
156. Urrego, D., Múnera, A., Troncoso, J. 2011. Retracción a largo plazo del árbol
dendrítico en neuronas piramidales cortico-faciales por lesiones periféricas del nervio
facial. Biomédica. 31, 560-569.
157. Urrego, D., Troncoso, J., Múnera, A. 2015. Layer 5 pyramidal neurons
dendritic remodeling and increased microglial density in primary motor cortex in a
murine model of facial paralysis. BioMed Research International.
158. Van der Loos, H. 1976. Barreloids in the somatosensory thalamus.
Neuroscience Letters. 2: 1-6.
159. Varela, C., Kumar, S., Yang, J., Wilson, M. 2013. Anatomical substrates for
direct interactions between hippocampus, medial prefrontal cortex, and the thalamic
nucleus reuniens. Brain Struct. Funct.
160. Veinante, P., Deschênes, M. 1999. Single- and multi-whisker channels in the
ascending projections from the principal trigeminal nucleus in the rat. J. Neurosci.
19: 5085-5095.

83
161. Wake, H., Moorhouse, A., Jinno, S., Kohsaka, S., Nabekura, J. 2009. Resting
microglia directly monitor the functional state of synapses in vivo and determine the
fate of ischemic terminals. The Journal of Neuroscience. 29(13), 3974-3980.
162. Waller, A. 1850. Experiments on the section of the glossopharyngeal and
hypoglossal nerves of the frog, and observations of the alterations produced thereby
in the structure of their primitive fibers. Philosophical Transactions of the Royal
Society of London. 140, 423-429.
163. Walker, F., Nilsson, M., Jones, K. 2013. Acute and chronic stress-induced
disturbances of microglial plasticity, phenotype and function. Current Drug Targets.
14, 1262-1276.
164. Wang, Y., Okamoto, M., Schmitz, F., Hofmann, K., Südhof, T. 1997. Rim is
a putative Rab3 effector in regulating synaptic-vesicle fusion. Nature. 388, 593-598.
165. Watson, C., Sakai, S., Armstrong, W. 1982. Organization of the facial nucleus
in the rat. Brain Behav. Evol. 20, 19-28.
166. Welker, C., Hoogland, P., Van der Loos, H. 1988. Organization of feedback
and feedforward projections of the barrel cortex: a PHA-L study in the mouse. Exp
Brain Res. 73, 411-435.
167. Welker, C. 1971. Microelectrode delineation of fine grain somatotopic
organization of SM1 cerebral cortex in albino rat. Brain. Res. 26: 259-275.
168. Welker, W. (1964). Analysis of sniffing of the albino rat. Behaviour. 22: 223-
244
169. Wigström, H., Gustafsson, B., Huang, Y. 1986. Hippocampal long-term
potentiation is induced by pairing single afferent volleys with intracellularly injected
depolarizing current pulses. Acta Physiol Scand. 126, 317-319.
170. Witter, M., Amaral, D. 2004 “Hippocampal Formation”, en The Rat Nervous
System, ed. G. Paxinos, 3era Edn (Sydney: Academic Press), 637-687.
171. Witter, M., Naber, P., Haeften, T., Machielsen, W., Rombouts, S., Barkhof,
F., Scheltens, P., da Silva, F. 2000. Cortico-Hippocampal communication by way of
parallel parahippocampal-subicuar pathways. Hipoocampus. 10, 398-410.

84
172. Woolsey, T., Van der Loos, H. 1970. The structural organization of layer IV
in the somatosensory region (S1) of mouse cerebral cortex: The description of a
cortical field composed of discrete cytoarchitectonic unit. Brain Ress. 17, 205-242.
173. Xu, L., Anwyl, R., Rowan, M. 1997. Behavioural stress facilitates the
induction of long-term depression in the hippocampus. Nature. 387, 497-500.
174. Yang, Y., Calakos, N. 2013. Presynaptic long-term plasticity. Front Synaptic
Neurosci. 5, 8.
175. Zakharenko, S., Patterson, S., Dragatsis, I., Zeitlin, S., Siegelbaum, S.,
Kandel, E., Morozov, A. 2003. Presynaptic BDNF required for a presynaptic but not
postsynaptic component of LTP at hippocampal CA1-CA3 synapses. Neuron. 39,
975-990.
176. Zamanillo, D., Sprengel, R., Hvalby, O., Jensen, V., Burnashev, N., Rozov,
A., Kaiser, K., Köster, H., Borchardt, T., Worley, P., Lübke, J., Frotscher, M., Kelly,
P., Sommer, B., Andersen, P., Seeburg, P., Sakman, B. 1999. Importance of AMPA
receptors for hippocampal synaptic plasticity but not for spatial learning. Science.
284, 1805-1811.
177. Zengel, J., Magleby, K. 1982. Augmentation and facilitation of transmitter
release. A quantitative description of the frog neuromuscular junction. J. Gen.
Physiol. 80, 583-611.
178. Zucker, R., Regehr, W. 2002. Short-Term Synaptic Plasticity. Annu. Rev.
Physiol. 64, 355-405.

![Tema 4 Tronco del Encéfalo: Visión anterior · Nervio óptico [Il] Nervio oculomotor [Ill] Protuberancia Nervio facial [Vll] Nervio vestibulococlear [VI' l] Nervio glosofaríngeo](https://static.fdocuments.ec/doc/165x107/5e17f5717125993d06498320/tema-4-tronco-del-encfalo-visin-anterior-nervio-ptico-il-nervio-oculomotor.jpg)