
EFECTO DE LA GLUCOSAMINA ORAL SOBRE LA …sappi.ipn.mx/cgpi/archivos_anexo/20050159_2336.pdf · a...
24
1 ADMINISTRACIÓN ORAL DE GLUCOSAMINA COMO UN MODELO PARA PRODUCIR RESISTENCIA A LA INSULINA EN LA RATA GESTANTE CLAVE CGPI 20050159 RESUMEN La glucosamina se ha utilizado como un fármaco que mejora los síntomas de la osteoartritis al disminuir la inflamación en las articulaciones, sin embargo, existen reportes de que produce resistencia a la insulina al disminuir la incorporación de la glucosa en los tejidos dependientes de insulina mediante la activación de la vía biosintética de las hexosaminas. Esta vía además tiene como función la síntesis de proteoglicanos. Estos estudios se han realizado en forma aguda, con una duración de hasta 6 horas de infusión de la glucosamina, por lo que si se administra a la rata la glucosamina de forma más parecida a la utilizada en la osteoartritis, esto es, por vía oral y durante varias semanas, entonces esperamos encontrar un estado de resistencia a la insulina con aumento en la glucosa en sangre, tanto en ratas nulíparas como en ratas gestantes. Resultados: La administración de 300 o 900 mg/kg peso de glucosamina disuelta en el agua de bebida durante once semanas o durante 15 semanas en la rata durante la gestación y la lactancia no provocó cambios en la ingesta de alimento, ni en el peso corporal de las ratas comparadas contra los grupos testigo, tampoco se encontraron cambios en los nivels de triglicéridos circulantes ni en la concentración de leptina, contrario a lo que se esperaba, ya que se ha demostrado que la glucosamina in vivo administrada durante 6 horas produce aumento en la síntesis de leptina. Las curvas de tolerancia a la glucosa y a la insulina muestran que la glucosamina produce resistencia a la insulina desde la quinta semana de administración y para la última semana de tratamiento (semana 11) se obtuvieron los efectos más pronunciados. Estos efectos aunque también se observaron en las retas durante la gestación y la lactancia, los cambios no fueron tan pronunciados, por lo que sugerimos que los cambios metabólicos propios del proceso reproductivo pueden estar aumentando el catabolismo de la glucosamina o disminuir su absorción. Por otro lado, la administración de 0.75 U/kg de insulina vía IP aunque no demuestra que los animales con glucosamina sean menos sensibles a esta dosis de insulina, las ratas tardan más tiempo en recuperarse de la hipoglucemia, indicando que se encuentra alterada la secreción de adrenalina o de glucagon, haciéndolos más propensos a periodos de hipoglucemia (Greenspan y Gardner, 2001). Se concluye que la glucosamina administrada por vía oral produce resistencia a la insulina en ratas nulíparas y en etapa reproductiva y que en las crías nacidas de las madres tratadas con glucosamina no se afecta su glucemia ni su incremento de peso corporal. INTRODUCCIÓN El sulfato de glucosamina es una molécula que se encuentra presente en el organismo humano en forma natural y es utilizada para la biosíntesis de los proteoglucanos de la sustancia fundamental del cartílago articular y del ácido hialurónico del líquido sinovial (Barclay et al; 1998). La glucosamina ha sido utilizada como droga antievolutiva de la osteoartritis (McAlindon et al, 2000). En la actualidad se utiliza ampliamente con estos fines, surgiendo la necesidad de revisar sus efectos adversos potenciales (Echard et al; 2001). La D-glucosamina de fuentes exógenas es incorporada a la vía metabólica de la síntesis de glucosaminoglucanos y la producción de proteoglucanos y aumenta la captación de sulfato por el
Transcript of EFECTO DE LA GLUCOSAMINA ORAL SOBRE LA …sappi.ipn.mx/cgpi/archivos_anexo/20050159_2336.pdf · a...

1
ADMINISTRACIÓN ORAL DE GLUCOSAMINA COMO UN MODELO
PARA PRODUCIR RESISTENCIA A LA INSULINA EN LA RATA GESTANTE
CLAVE CGPI 20050159 RESUMEN La glucosamina se ha utilizado como un fármaco que mejora los síntomas de la osteoartritis al disminuir la inflamación en las articulaciones, sin embargo, existen reportes de que produce resistencia a la insulina al disminuir la incorporación de la glucosa en los tejidos dependientes de insulina mediante la activación de la vía biosintética de las hexosaminas. Esta vía además tiene como función la síntesis de proteoglicanos. Estos estudios se han realizado en forma aguda, con una duración de hasta 6 horas de infusión de la glucosamina, por lo que si se administra a la rata la glucosamina de forma más parecida a la utilizada en la osteoartritis, esto es, por vía oral y durante varias semanas, entonces esperamos encontrar un estado de resistencia a la insulina con aumento en la glucosa en sangre, tanto en ratas nulíparas como en ratas gestantes. Resultados: La administración de 300 o 900 mg/kg peso de glucosamina disuelta en el agua de bebida durante once semanas o durante 15 semanas en la rata durante la gestación y la lactancia no provocó cambios en la ingesta de alimento, ni en el peso corporal de las ratas comparadas contra los grupos testigo, tampoco se encontraron cambios en los nivels de triglicéridos circulantes ni en la concentración de leptina, contrario a lo que se esperaba, ya que se ha demostrado que la glucosamina in vivo administrada durante 6 horas produce aumento en la síntesis de leptina. Las curvas de tolerancia a la glucosa y a la insulina muestran que la glucosamina produce resistencia a la insulina desde la quinta semana de administración y para la última semana de tratamiento (semana 11) se obtuvieron los efectos más pronunciados. Estos efectos aunque también se observaron en las retas durante la gestación y la lactancia, los cambios no fueron tan pronunciados, por lo que sugerimos que los cambios metabólicos propios del proceso reproductivo pueden estar aumentando el catabolismo de la glucosamina o disminuir su absorción. Por otro lado, la administración de 0.75 U/kg de insulina vía IP aunque no demuestra que los animales con glucosamina sean menos sensibles a esta dosis de insulina, las ratas tardan más tiempo en recuperarse de la hipoglucemia, indicando que se encuentra alterada la secreción de adrenalina o de glucagon, haciéndolos más propensos a periodos de hipoglucemia (Greenspan y Gardner, 2001). Se concluye que la glucosamina administrada por vía oral produce resistencia a la insulina en ratas nulíparas y en etapa reproductiva y que en las crías nacidas de las madres tratadas con glucosamina no se afecta su glucemia ni su incremento de peso corporal. INTRODUCCIÓN El sulfato de glucosamina es una molécula que se encuentra presente en el organismo humano en forma natural y es utilizada para la biosíntesis de los proteoglucanos de la sustancia fundamental del cartílago articular y del ácido hialurónico del líquido sinovial (Barclay et al; 1998). La glucosamina ha sido utilizada como droga antievolutiva de la osteoartritis (McAlindon et al, 2000). En la actualidad se utiliza ampliamente con estos fines, surgiendo la necesidad de revisar sus efectos adversos potenciales (Echard et al; 2001). La D-glucosamina de fuentes exógenas es incorporada a la vía metabólica de la síntesis de glucosaminoglucanos y la producción de proteoglucanos y aumenta la captación de sulfato por el

2
cartílago articular, e inhibe la respuesta inflamatoria contra agentes inespecíficos como el ácido acético, pero no tiene propiedades analgésicas. La administración de glucosamina exógena estimula la síntesis de matriz cartilaginosa y brinda protección inespecífica contra el daño químico, proporcionando al cartílago articular la hexosamina y el anión sulfato necesarios para la síntesis de glucosaminoglucanos. Estudios comparativos sugieren que la glucosamina tiene muchos menos efectos secundarios que los anti-inflamatorios no esteroidales, aunque tal vez interfiere en la tolerancia a la glucosa, por lo que la administración en pacientes diabéticos hace necesaria la medición de los niveles de glucosa en plasma (Barclay et al; 1998). ANTECEDENTES Como se mencionó anteriormente el consumo de la glucosamina para el tratamiento de la osteoartritis puede contribuir a la resistencia a la insulina, dado que la glucosamina tiene el potencial para interferir en el metabolismo de la glucosa (Echard et al; 2001). Existen tres eventos principales que regulan los niveles de glucosa plasmática, y que se presentan de manera coordinada después de la ingestión de carbohidratos: la estimulación de la secreción de insulina, la supresión de la producción endógena de glucosa por el hígado mediada por la insulina y la estimulación de la captura de glucosa dependiente de insulina por el músculo y por otros tejidos. La patogénesis de la diabetes mellitus tipo 2 (DM-2) es compleja y en la mayoría de los casos requiere la existencia tanto de defectos funcionales de la célula ß pancreática, como de alteraciones en la sensibilidad a la insulina. En conjunto, estas anormalidades producen un aumento de la glucosa liberada por el hígado y el riñón durante el ayuno, así como una disminución de la depuración de glucosa de la circulación, eventos que dependen de la sensibilidad de estos tejidos a la insulina. En la última década se han realizado numerosos estudios para discernir el papel de la resistencia a la insulina en el desarrollo y mantenimiento de un estado hiperglucémico. La hiperglucemia se desarrolla sólo cuando la cantidad de glucosa liberada a la circulación excede la velocidad de captación de la misma por los tejidos, independientemente de que la liberación se encuentre aumentada, de que la utilización se encuentre disminuida, o de que concurran ambos eventos (Racotta, 2002). La resistencia a la insulina se caracteriza por una secreción excesiva de insulina después de una carga de glucosa, para llevar a cabo la incorporación de la misma en los tejidos sensibles a ella, como son el muscular, adiposo y cardiaco. Por lo general se presenta en individuos obesos, que no son diabéticos o que sólo manifiestan ligeras anormalidades en la tolerancia a la glucosa. Se ha sugerido que esta hiperinsulinemia puede ser atribuible a una insensibilidad periférica a la insulina, y a una hipersensibilidad de los islotes pancreáticos a la glucosa (Harper, 1979). Recientemente, numerosos investigadores han propuesto que la resistencia a la insulina es un evento primario en la etiopatogénesis de la DM-2, que se ha tratado de explicar por los cambios en la regulación de la vía de las hexosaminas. Hipótesis de las Hexosaminas y la resistencia a la insulina. Uno de los tejidos más estudiados en cuanto al metabolismo de la glucosa es el muscular, en este se han descrito las vías metabólicas que puede tomar la glucosa una vez que ha sido transportada del plasma a las células (Hawkins et al, 1997). En células adiposas y de músculo esquelético (fig. 1) después del transporte y fosforilación de la glucosa a glucosa 6 fosfato (G-6-P), este producto es usado principalmente en dos vías: la vía de síntesis de glucógeno y la vía de la glicólisis y sólo del 1-3% de la glucosa que entra a la célula es convertida a fructosa-6 fosfato (F-6-P) la cual va a la vía biosintética de hexosaminas (VBH), cuyos productos finales son: el UDP-N-acetilglucosamina (UDP-Glc-N-Ac) y UDP-N-acetilgalactosamina (UDP-Gal-N-Ac), que sirven como sustratos en la síntesis de

3
glucoproteínas (Hawkins et al, 1997; Marshall et al, 1991; Ravussin, 2002; Traxinger et al, 1991; Buse, 2006 ). GLUCOSA GLUCOSAMINA G-6-P F-6-P GFAT Glc-N-6P G-1-P Glc-N-1P UDP-G Glc-N-Ac URIDINA GLICOLISIS UDP-Glc-N-Ac SINTESIS DE GLICOSILACION GLUCOGENO
Fig. 1.- Sitios de entrada de la glucosa, glucosamina y uridina en la vía biosintética de las hexosaminas (VBH) en músculo esquelético. Después de que es transportada la glucosa a las células es rápidamente fosforilada a Glucosa 6 fosfato (G-6-P), la glucosa es principalmente utilizada para la vía de síntesis de glucógeno y la glicólisis. La VBH recibe sólo del 1 al 3 % de la glucosa transportada vía la conversión de Fructosa 6 fosfato (F-6-P), a glucosamina 6 fosfato (Glc-N-6P) por la velocidad limitada de la enzima: fructosa 6 fosfato amindotransferasa (GFAT). Alternativamente la incorporación de la glucosamina exógena en las células es realizada por el sistema de transportación de la glucosa y es fosforilada a (Glc-N-6P). Posteriormente se forman metabolitos por subsecuentes acetilaciones y uridilaciones de la Glc-N-6P, después de que es infundida a la célula uridina se lleva acabo la formación de subproductos de la VBH. El principal subproducto es UDP-Glc-N-Ac.
En estudios realizados en preparaciones in vivo y en células aisladas se demostró que el incremento en el flujo en la VBH, antagoniza la habilidad de la insulina para la utilización de la glucosa (Marshall et al, 1991; Hawkins et al, 1997; Patti, 1999). Un aumento de las concentraciones de UDP-N-acetilhexosamina en músculo esquelético el cual es un índice del flujo hacia la vía de las hexosaminas, se relaciona positivamente con el grado de insulinoresistencia (Hawkins et al ,1996). Los últimos productos (UDP-N-acetilhexosamina) representan el mismo mecanismo por el cual la hiperglucemia crónica da como resultado una deteriorada estimulación del transporte de la glucosa mediado por la insulina (Rosseti, 2000). Los procesos bioquímicos que regulan la concentración de glucosa plasmática parecen depender de varios mecanismos. Por ejemplo, la respuesta inmediata se debe a la actividad de las células ß provocada por los cambios en la concentración de glucosa extracelular a través de la síntesis de hexosamina. La fructosa 6 fosfato (F-6-P) es convertida a glusosamina-6-fosfato por un paso limitante catalizado por la enzima glutamina:fructosa 6 fosfato amidotransferasa (GFAT), cuyo principal producto es la UDP-N-acetilglucosamina (UDP-Glc-N-Ac). La sobreexpresión de GFAT produce resistencia a la insulina en fibroblastos por estimulación de la glucógeno sintetasa, en tanto que la glucosamina causa resistencia a la insulina en el músculo (Marshall et al, 1991). La vía de las hexosaminas puede actuar como un sensor del estado nutricional de la célula. En otros estudios in vivo, se ha observado que el incremento de la glucosamina disponible induce resistencia periférica a la insulina en ratas normales, pero no tiene efectos adicionales inhibitorios en ratas diabéticas, implicando a la vía de la glucosamina como el mecanismo potencial por el cual la hiperglucemia pudiera inducir resistencia a la insulina. Por lo tanto, se ha planteado recientemente que

4
la sobreactividad de la VBH podría ser uno de los posibles mecanismos mediadores de la glucosa para inducir resistencia a la insulina (Patti, 1999; Virkamaki et al, 1997). Sin embargo, en ratas diabéticas la glucosamina no ha tenido efecto sobre la externalización de los transportadores de glucosa, sugiriendo que la resistencia a la insulina es secundaria a la activación de la VBH. Por lo cual, la hipótesis de que la glucosamina induce resistencia a la insulina en el metabolismo intracelular de la glucosa es controversial (Virkamaki et al, 1997). Otros estudios han demostrado que la sobreexpresión de la GFAT es responsable de varios trastornos metabólicos (Calzada, 2002) como son: el aumento de glucógeno hepático e incremento de los ácidos grasos exportados a los adipocitos, aún en presencia de concentraciones plasmáticas subnormales de glucosa. En el tejido adiposo produce aumento de la acreción grasa e incremento en los lípidos intracelulares. En el músculo produce disminución de la captura de glucosa estimulada por insulina. En las células ß produce aumento en la secreción de insulina. Ácidos grasos libres y resistencia a la insulina. Los niveles plasmáticos altos de ácidos grasos libres (AGL) se han relacionado con la resistencia a la insulina, lo cual se asocia con el decremento en el músculo esquelético de los niveles de G-6-P, indicando un defecto en el transporte y la fosforilación de la glucosa, y una marcada acumulación de los productos de la VBH, explicado de la siguiente manera: cuando existe un exceso de oferta de ácidos grasos libres en el músculo, se incrementa la concentración de acil-CoA derivada de su oxidación mitocondrial, el cual inhibe la deshidrogenasa pirúvica (DHP), disminuyendo la tasa de oxidación de piruvato, que junto a la inhibición de la fosfofructocinasa (PFC), que impide que la fructosa-6-fosfato ingrese a la vía glucolítica, determina una mayor concentración de ésta última para ingresar a la vía de las hexosaminas. La enzima GFAT cataliza la conversión de fructosa -6-fosfato a glucosamina-6-fosfato, siendo la enzima limitante de ésta vía, pudiendo por tanto existir un mayor flujo hacia la vía de las hexosaminas por la hiperglucemia crónica (aumento de la entrada de glucosa a la célula), por infusión de glucosamina (saltándose la enzima limitante GFAT) o bien por infusión de AGL (vía inhibición de la PFC). La activación de la vía de la hexosamina puede directamente modular eventos tempranos postreceptor de la vía de señalamiento de la insulina, observándose que el aumento de producción de glucosamina disminuye el transporte de glucosa y la translocación de los GLUT-4 (Marshall et al, 1991; Rossetti ,2000). Se ha descrito también que los mecanismos potenciales para ello, incluyen la activación de cinasas serina-treonina, las cuales fosforilan el sustrato para el receptor de insulina-1 (IRS-1), disminuyendo su actividad como sustrato para la actividad tirosina cinasa del receptor insulínico o bien mediante la activación de una proteína tirosina fosfatasa, pudiendo ser otro de los mecanismos por los cuales los ácidos grasos libres generan insulinorresistencia (Hawkins et al, 1997). Estos cambios son evidentes con aumentos dentro de límites fisiológicos. En el corto plazo, estas modificaciones parecen deberse a adaptaciones que permiten a la célula percibir que existe un exceso de calorías, que se acumulan como grasa para su uso futuro; pero cuando los cambios se hacen crónicos se producen resistencia a la insulina idéntica a la observada en DM-2, causada por disminución de la translocación de GluT-4, probablemente debido a una alteración en la actividad de la fosfatidilinositol-3 cinasa (Calzada et al, 2002). Vía de las hexosaminas y producción de leptina.
Varios estudios demuestran que la secreción de insulina y la leptina se autorregulan por mecanismos de retroalimentación negativa. La insulina estimula la secreción de leptina por los adipocitos y a su vez la leptina actúa en las células ß del páncreas inhibiendo la secreción de insulina (Kieffer et al, 1997). Por

5
otra parte, el mecanismo por el cual la insulina ejerce sus efectos sobre la síntesis y secreción de leptina no son bien conocidos. Se ha sugerido que el metabolismo de la glucosa en parte podría estar relacionado debido a que la secreción de leptina por los adipocitos de rata bajo estímulo de insulina no se observa con la presencia de 2–deoxy–D–glucosa (Mueller et al, 1998). La vía sintética de las hexosaminas también podría estar involucrada ya que se ha propuesto que esta vía sería un "sensor" intracelular de la energía disponible y que mediarían los efectos de la glucosa sobre la expresión de varios genes. Un aumento en las concentraciones tisulares de UDP–N–acetilglucosamina, el producto final de la vía de las hexosaminas, aumenta rápida y marcadamente la síntesis del RNA mensajero de leptina en células musculares y adipocitos de rata (Wang et al, 1998). La activación de la VBH en ratas por la infusión in vivo, por vía intravenosa de glucosa, uridina, ácidos grasos o glucosamina incrementa los niveles circulantes de leptina (Wang, et al 1998); además, se ha visto que la expresión de GFAT en el tejido adiposo y muscular tiende a incrementar los niveles de leptina en plasma (Obici et al, 2003). La acción de la insulina y el balance energético involucran múltiples y complejas cadenas en su regulación. Existe una asociación entre el peso y la homeostasis de la glucosa. Con la ingesta, la abundancia de nutrientes es percibida en sitios periféricos, en músculo esquelético y tejido adiposo y en sitios centrales, así como por el hipotálamo. En tejidos periféricos un incremento del flujo de lípidos y carbohidratos aumenta la actividad de la MalonilCoA y la Coenzima A de los ácidos grasos de cadena larga (LCFA-CoA) y la VBH. El aumento de cada uno de estos metabolitos tiende a participar en la resistencia a la insulina y a disminuir la oxidación de ácidos grasos. Así mismo, la activación de la VBH también disminuye el gasto de energía (muscular) e induce la expresión de leptina (en tejido adiposo y muscular) (Obici et al, 2003).
Efectos durante la gestación y lactancia Es sabido que, durante el embarazo se llevan acabo algunos cambios metabólicos en la madre desde el inicio de la gestación y continúan a lo largo del periodo, con el fin de optimizar la transferencia de nutrimentos al feto. Una alteración que se puede presentar es la secreción deficiente de insulina durante el embarazo, presentándose una clara intolerancia a la glucosa que se conoce como diabetes gestacional (DG). La DG es una intolerancia a los hidratos de carbono o glúcidos que se diagnostica en el curso del embarazo, sin haber pruebas de una diabetes previa y que habitualmente desaparece al cabo de un tiempo del postparto sin dejar secuelas. Es un trastorno metabólico pasajero ocasionado por las hormonas fetoplacentarias y la sobrecarga metabólica del embarazo. La DG constituye un problema de salud importante en las ciudades con mayor incidencia de factores de riesgo por tal alteración. Esta alteración cobra mayor importancia debido a las consecuencias a corto y largo plazo que se pueden presentar en el feto y en la madre. La repercusión de esta alteración en el ambiente hormonal materno puede afectar el funcionamiento del páncreas y el metabolismo de la glucosa en la cría, y pueden ocasionar el desarrollo de resistencia a la insulina. La DG se caracteriza por disminución de la sensibilidad a la insulina, aumento del 30 por ciento de la secreción de insulina y disminución de la sensibilidad a la glucosa por las células ß para secretar insulina (Gelardi et al, 1991, Butte et al, 2000). Los criterios definitorios de normalidad o anormalidad son arbitrarios y han cambiado a lo largo del tiempo. Para llegar al diagnóstico de diabetes gestacional se debe tener evidencia de que no existía la diabetes antes de la gestación, pero a veces una diabetes mellitus debuta en el embarazo y persiste para toda la vida. Por tanto, el segundo aspecto de la diabetes "gestacional" es su carácter transitorio durante el embarazo y su desaparición unas semanas después del parto. En cambio, la diabetes mellitus diagnosticada en el embarazo es un trastorno más grave del metabolismo y persiste con posterioridad al parto.

6
El riesgo de padecer diabetes gestacional no es igual para todas las embarazadas. Entre los factores de riesgo están la historia familiar de diabetes, la historia personal de glucosuria o intolerancia de los hidratos de carbono, obesidad, edad superior a 30 años, peso excesivo de la gestante cuando nació ella y antecedentes desfavorables en embarazos previos: aborto habitual, muerte fetal sin causa conocida, malformaciones fetales, peso fetal excesivo, hidramnios, preeclampsia o pielonefritis.
La diabetes gestacional no produce síntomas durante el embarazo, se puede sospechar cuando existen factores de riesgo y entonces se hacen pruebas a lo largo del embarazo para detectar las alteraciones metabólicas. Las embarazadas deben medir periódicamente su glucemia para detectar alteraciones que puedan ser motivo de estudio más profundo.
JUSTIFICACION La ingesta de glucosamina como un tratamiento suplementario para la osteoartritis puede ocasionar resistencia a la insulina en los pacientes que la consumen. Los efectos secundarios por su ingesta que han sido estudiados hasta ahora han demostrado que esta se encuentra relacionada con la resistencia a la insulina y posiblemente en la disminución de peso. Sin embargo, todos estos estudios se han realizado mediante su administración aguda, y no se ha establecido un estudio que pueda determinar estos efectos, al suministrarse en condiciones mas parecidas a las que utilizan los enfermos de osteoartritis. Es así que resulta importante el conocer si la glucosamina ocasiona cambios en la sensibilidad a la insulina.

7
OBJETIVO GENERAL Obtener un modelo de resistencia a la insulina mediante la administración crónica de glucosamina por vía oral en la rata hembra virgen y en la gestante.
Objetivos Particulares:
* Determinar la dosis de glucosamina oral que provoque la resistencia a la insulina en ratas vírgenes y en gestantes. * Determinar si la administración de la glucosamina durante la gestación tiene algún efecto sobre las crías (resistencia a la insulina). * Determinar si la activación de VBH aumenta la producción de leptina y su posible relación en la ingesta de alimento.
HIPÓTESIS Si el consumo de glucosamina por vía oral interviene en el metabolismo de la glucosa incrementando la vía biosintética de las hexosaminas, lo cual incrementaría la concentración intracelular de subproductos de esta misma, que ocasionarían la resistencia a la insulina así como la disminución de la ingesta de alimento y del peso corporal por producción de leptina. Entonces en la rata virgen como en la rata gestante, se obtendrán cambios en el metabolismo semejantes a los ocurridos durante un estado prediabético y diabetes gestacional respectivamente. MATERIAL Y METODOS Manteniendo una dieta estándar ad libitum se estudió el efecto de la administración de glucosamina oral en ratas hembras adultas, las cuales se mantuvieron en una cámara a 19 grados centígrados con ciclo de luz-oscuridad de 12 horas. Experimento 1. “Determinación de los efectos de la glucosamina”. Un lote de ratas hembras de la cepa Wistar de 180 g de peso promedio, se dividió en 2 grupos de 6 ratas cada uno y se les sometió a las siguientes condiciones: • Grupo uno, control siguió el proceso sin tratamiento. • Grupo dos, se administró una dosis de 300 mg/kg por día de glucosamina vía oral disuelta en el
agua de beber . Este tratamiento duró 11 semanas, durante las cuales se midieron los niveles de glucosa en sangre semanalmente. Se cuantificó el consumo de alimento y agua diarios y el peso de los animales dos veces por semana. Al inicio, a la quinta semana y al término del tratamiento se realizó su curva de tolerancia a la glucosa y concentración de insulina en plasma para medir la resistencia a la insulina; al final del experimento se realizó la curva de tolerancia a la insulina, por vía intraperitoneal, así como la medición de los niveles de leptina en plasma.

8
Experimento 2. “Determinación de los efectos de la glucosamina durante la gestación”. Se formaron dos grupos de ocho ratas hembra de la cepa Wistar de 180 g de peso en promedio y se siguió el siguiente protocolo. • Grupo 1, ratas gestantes sin tratamiento. • Grupo 2, ratas gestantes que recibieron una dosis de 900 mg/kg por día, de glucosamina por vía oral,
disuelta en el agua de beber. Las ratas se sometieron al tratamiento correspondiente y en la sexta semana se aparearon y continúo el tratamiento hasta la semana 15 después de iniciado el tratamiento. El día posterior al parto se ajustaron las camadas a un total de 8 crías, tratando de tener cuatro machos y cuatro hembras. Se realizaron las mismas mediciones que en el experimento 1, además, al inicio y al final de la gestación y lactancia se midieron las concentraciones de insulina en plasma y las curvas de tolerancia a la glucosa. En las crías se realizaron las mediciones de glucosa en plasma en los días 7, 14, 21 y 35 de edad. Se registró el peso corporal diariamente. Metodología. 1.- Medición de las concentraciones plasmáticas de Glucosa, Insulina y Leptina.
• Hembras adultas: • Se tomaron semanalmente muestras de sangre de la vena de la cola de las ratas. La concentración de glucosa se determinó empleando el método del glucosímetro. • Las muestras para las curvas de tolerancia a la glucosa y concentración de insulina en plasma se colectaron en tubos heparinizados y posteriormente fueron centrifugadas a 3000 rpm/10 min. a 2 °C, para la obtención del plasma. Las mediciones de las concentraciones plasmáticas de insulina y leptina en plasma se hicieron por métodos de inmunoensayo (Insulina y Leptina ELISA). • Crías: se tomó una muestra de sangre de la cola por punción para la medición de glucosa por el método del glucosímetro.
2.- Medición de la tolerancia a la glucosa.
Se tomaron las muestras de sangre de la cola de la rata en condiciones basales, posteriormente se administró por vía intraperitoneal 1 ml/kg de peso corporal de una solución al 35% de glucosa. Las muestras de sangre fueron colectadas en los tiempos 0, 15, 30, 90 y 120 minutos después de la administración. De estas muestras también se realizaron las mediciones de insulinemia. 3.- Medición de la glucosa en sangre después de la administración de insulina por vía intraperitoneal (curva de tolerancia a la insulina).
Para el experimento 1 se tomaron muestras de sangre de la cola de la rata, a los tiempos 0, 30, 60 y 120 minutos después de la administración de una solución de 3 unidades de insulina por kg de peso.

9
Mientras que en el experimento 2 para la curva de tolerancia a la insulina se tomaron muestras de sangre de la cola de la rata, la cual fue medida en los tiempos 0, 15, 30, 45, 60, 120 y 180 minutos después de la administración de una solución de 0.75 unidades de insulina por kg de peso.
RESULTADOS INGESTA DE ALIMENTO En las figuras 2 y 3 se muestran los cambios del aumento de peso de los dos grupos de ratas de cada experimento, durante el tiempo de tratamiento respectivo, en los cuales se observó un comportamiento semejante entre los dos grupos.
Fig. 2.- Aumento de peso en los dos grupos de ratas durante 11 semanas, el número de ratas para cada grupo fue de 6 (Experimento 1).
Fig. 3.- Aumento de peso en para los dos grupos en estudio durante 15 semanas, el número de ratas para cada grupo fue de 8 (Experimento 2). Semana 6 gestación, semana 9 lactancia, semana12 término de lactancia.
180
200
220
240
260
280
300
1 2 3 4 5 6 7 8 9 10 11Tiempo en semanas
Pes
o co
rpor
al (g
)
CONTROL GLUCOSAMINA
150
200
250
300
350
400
450
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Tiempo en semanas
Pes
o co
rpor
al (g
)
CONTROL GLUCOSAMINA

10
En las figura 4 y 5 se muestra la ingesta de alimento durante los periodos de tratamiento en las cuales no se observan diferencias importantes entre los grupos control y estudio en ninguno de los dos experimentos.
Fig. 4.- Ingesta de alimento durante el periodo de estudio de 40 días, el número de ratas para cada grupo fue de 6 (Experimento 1). Se muestra la media ± el error estándar. No hay diferencia entre grupos
Fig. 5.- Ingesta de alimento durante el periodo de estudio de 15 semanas, el número de ratas para cada grupo fue de 8 (Experimento 2). Semana 6 inicio de la gestación, semana 9 inicio de la lactancia, semana 12 término de lactancia. Se muestra la media ± el error estándar. No hay diferencia entre grupos.
10
12
14
16
18
20
22
24
2 3 4 5 6 7 8 9 10 11Tiempo en semanas
Inge
sta
de a
limen
to (g
r)
CONTROL GLUCOSAMINA
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Tiempo en semanas
Inge
sta
de a
limen
to (g
)
CONTROL GLUCOSAMINA

11
Los niveles de leptina plasmáticos en los dos grupos en estudio se muestran en la tabla 1, se observa que en cada experimento su concentración fue semejante entre grupos.
Concentración de leptina en plasma (ng/ml) Experimento 1 Experimento 2 CONTROL 0.73 + .111 4.94 +0.151 ESTUDIO 0.73 + .098 5.16 + .138
Tabla 1.- Niveles de leptina en plasma al finalizar el tratamiento de glucosamina (300 y 900 mg/kg de peso).Se muestra la media ± el error estándar. Grupo control (n=6) y el grupo estudio (n=7), p>0.05
La tabla 2 muestra los niveles de triglicéridos encontrados al final del experimento 2, los cuales disminuyeron significativamente a la administración de glucosamina.
Concentración de triglicéridos en plasma (mg/ml) C0NTROL 75.83 + 4.593 ESTUDIO 69.99 + 5.614
Tabla 2. Niveles de triglicéridos en plasma al finalizar el tratamiento (900 mg/kg de peso). Se muestra la media ±el error estándar. Grupo control (n=6) y el grupo estudio (n=7).
GLUCEMIAS La concentración de glucosa en sangre en ayuno de los grupos control y estudio se muestran en las figuras 6 (experimento 1) y 7 (experimento 2), la glucemia fue semejante entre los grupos en ambos experimentos.
Fig. 6.- Concentración de glucosa en sangre en ayuno, durante las once semanas de tratamiento, el número de ratas para cada grupo fue de 6 (Experimento 1). Se muestra la media + el error estándar.
50
55
60
65
70
75
80
85
90
95
1 2 3 4 5 6 7 8 9 10Tiempo en semanas
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA
12
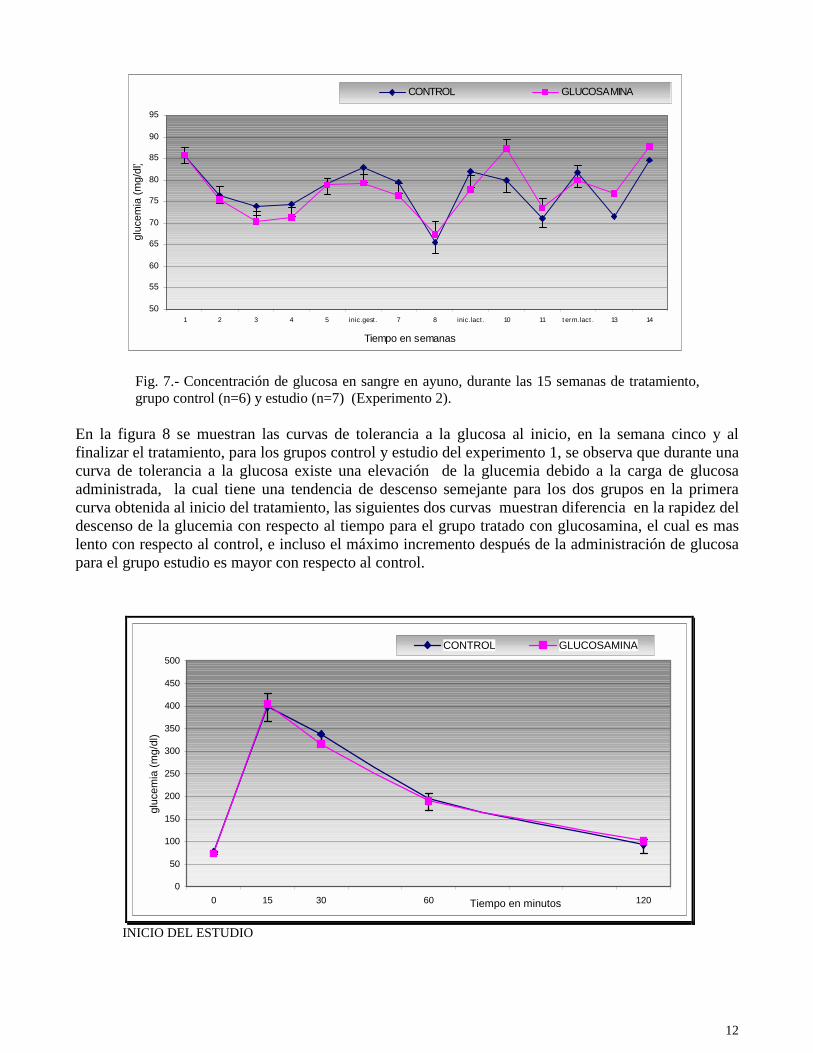
Fig. 7.- Concentración de glucosa en sangre en ayuno, durante las 15 semanas de tratamiento, grupo control (n=6) y estudio (n=7) (Experimento 2).
En la figura 8 se muestran las curvas de tolerancia a la glucosa al inicio, en la semana cinco y al finalizar el tratamiento, para los grupos control y estudio del experimento 1, se observa que durante una curva de tolerancia a la glucosa existe una elevación de la glucemia debido a la carga de glucosa administrada, la cual tiene una tendencia de descenso semejante para los dos grupos en la primera curva obtenida al inicio del tratamiento, las siguientes dos curvas muestran diferencia en la rapidez del descenso de la glucemia con respecto al tiempo para el grupo tratado con glucosamina, el cual es mas lento con respecto al control, e incluso el máximo incremento después de la administración de glucosa para el grupo estudio es mayor con respecto al control.
INICIO DEL ESTUDIO
50
55
60
65
70
75
80
85
90
95
1 2 3 4 5 inic.gest . 7 8 inic.lact . 10 11 t erm.lact . 13 14
Tiempo en semanas
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120Tiempo en minutos
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA

13
QUINTA SEMANA DE ESTUDIO
TERMINO DEL TRATAMIENTO * DS del área bajo la curva
Fig. 8.- Curvas de tolerancia a al glucosa en los dos grupos: estudio y control con tratamiento de 300 mg /kg de glucosamina administrado oralmente en el agua de beber durante 11 semanas de tratamiento. Se observa en todas las ratas una elevación de la glucemia a los quince minutos después de la administración de glucosa, así como una separación en las curvas al final del tratamiento entre el grupo control y el grupo de estudio. El área bajo la curva al inicio fue de 24436.5 para el grupo control (n=6) y 23512.5 para el estudio (n=6), para la quinta semana fue de 24003 para el grupo control (n=6) y 28548 para el estudio (n=6) y al finalizar el tratamiento fue de 30619.5 para el grupo control (n=6) y 39279 para el estudio (n=6). * p < 0.01
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120tiempo en minutos
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120Tiempo en minutos
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA

14
Curvas de tolerancia a la glucosa en los dos grupos: estudio y control con tratamiento de 300 mg /kg de glucosamina administrado oralmente en el agua de beber durante 11 semanas de tratamiento. El área (mg/dl*min) bajo la curva al inicio fue de 203.6 control y 195.9 estudio, para la quinta semana fue de 200 control y 237.9 estudio y al finalizar el tratamiento fue de 255.1 control (n=6) y 327.3 estudio (n=6). * p < 0.01. En el experimento 2 se observaron comportamientos similares en la curva de tolerancia a la glucosa administrada intraperitonealmente, solo que, ésta es después de la semana once y las áreas bajo la curva fueron menores (figura 14).
INICIO DEL TRATAMIENTO
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120Tiempo en minutos
glu
cem
ia (m
g/dl
)
CONTROL GLUCOSAMINA

15
SEXTA SEMANA DE ESTUDIO
ONCEAVA SEMANA DE ESTUDIO
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120Tiempo en minutos
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120Tiempo en minutos
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA

16
TERMINO DEL ESTUDIO * DS en el área bajo la curva
Fig. 9.- Curvas de tolerancia a al glucosa en los dos grupos: estudio y control con tratamiento de 900 mg /kg de glucosamina administrado oralmente en el agua de beber durante 15 semanas de tratamiento. Se observa en todas una elevación de la glucemia después de la administración de glucosa, así como una separación en las curvas al final del tratamiento entre el grupo control y el grupo de estudio. El área bajo la curva al inicio fue de: 25060 para el grupo control (n=6) y 23358.2 para el estudio (n=7), para la sexta semana fue de: 28899.6 para el grupo control (n=7) y 30025 para el estudio (n=7), para la onceava semana fue de: 25713.7 para el grupo control (n=6) y 26653.9 para el estudio (n=7) y al finalizar el tratamiento fue de: 27257.5 para el grupo control (n=6) y 34476.4 para el estudio (n=7). * p < 0.05
0
50
100
150
200
250
300
350
400
450
500
0 15 30 60 120Tiempo en minutos
glu
cem
ia (m
g/dl
)CONTROL GLUCOSAMINA
17
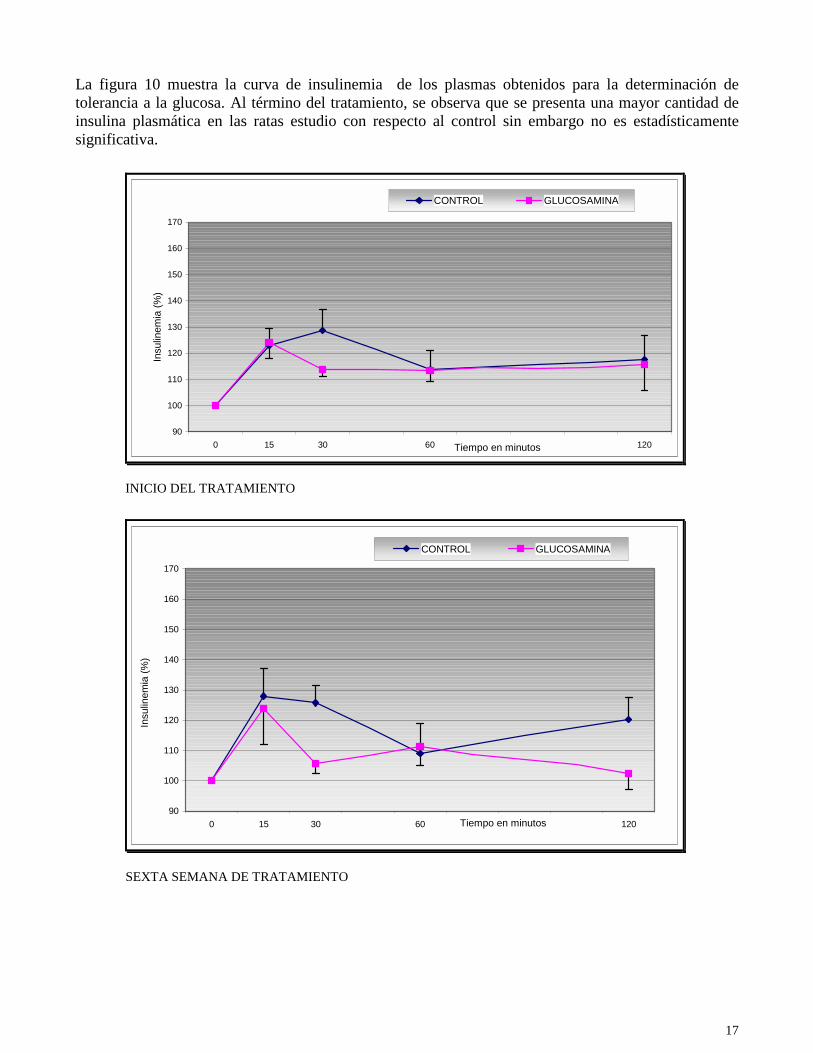
La figura 10 muestra la curva de insulinemia de los plasmas obtenidos para la determinación de tolerancia a la glucosa. Al término del tratamiento, se observa que se presenta una mayor cantidad de insulina plasmática en las ratas estudio con respecto al control sin embargo no es estadísticamente significativa.
INICIO DEL TRATAMIENTO
SEXTA SEMANA DE TRATAMIENTO
90
100
110
120
130
140
150
160
170
0 15 30 60 120Tiempo en minutos
Insu
linem
ia (%
)
CONTROL GLUCOSAMINA
90
100
110
120
130
140
150
160
170
0 15 30 60 120Tiempo en minutos
Insu
linem
ia (%
)
CONTROL GLUCOSAMINA

18
TERMINO DEL TRATAMIENTO
Fig. 10.- Curvas de insulinemia en los dos grupos: estudio y control con tratamiento de 900 mg /kg de glucosamina administrado oralmente en el agua de beber durante 15 semanas de tratamiento. Se observa en las curvas una elevación de los niveles de insulina plasmática después de la administración de glucosa, así como una separación en las curvas a la sexta semana y al final del tratamiento entre el grupo control y el grupo de estudio. El área bajo la curva al inicio fue de 14348.79 para el grupo control (n=6) y 13797.37 para el estudio (n=7), para la sexta semana fue de 14120.07 para el grupo control (n=6) y 12993.31 para el estudio (n=7) y al finalizar el tratamiento fue de 16701.92 para el grupo control (n=5) y 16914.43 para el estudio (n=7). P>0.05
La figura 11 muestra la curva de tolerancia a la insulina del experimento 1, al administrar 3 unidades por kilogramo de peso, para los dos grupos de ratas, en las cuales no se presentó ninguna diferencia entre ellos en la tendencia de descenso en la concentración de glucosa plasmática. En la figura 12 se observa que no existe una diferencia en el descenso de la glucemia sin embargo la recuperación es mas lenta para el grupo estudio mostrando sólo diferencia significativa en la última lectura (tiempo 180 minutos) .
90
100
110
120
130
140
150
160
170
0 15 30 60 120Tiempo en minutos
Insu
linem
ia (%
)
CONTROL GLUCOSAMINA

Curva de tolerancia a la insulina
0
20
40
60
80
100
120
0 30 60 120Tiempo en minutos
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA
Fig. 11.- Curva de tolerancia a la insulina (se administraron intraperitonealmente 3 U insulina/kg de peso al finalizar el tratamiento de 300 mg/kg de peso de glucosamina durante 11 semanas). El número de ratas para cada grupo fue de 5.
Figura 12. Curva de tolerancia a la insulina (se administraron iU insulina/ kg de peso al final del tratamiento de 900 mg/kg dedurante 15 semanas). Grupo control (n=6) y grupo estudio.(n=
0
20
40
60
80
100
120
0 15 30 45 60 90Tiempo en minutos
% d
e de
scen
so d
e gl
ucos
a (m
g/dl
)
CONTROL GLUCOSA MINA
*
19
ntraperitonealmente 0.75 peso de glucosamina 6). * p<0.01
150 180

20
CRIAS En las figuras 13 y 14 se muestran los cambios del aumento de peso y niveles de glucosa en sangre de las crías de los dos grupos de ratas del experimento 2 con ayuno de 8 horas, durante el tiempo de lactancia, en los cuales se observó aumento semejante entre las crías del grupos control y estudio.
Fig. 13. Peso corporal de las crías control y estudio durante el nacimiento y hasta los 35 días de edad, el número de ratas para cada grupo fue de 7 (Experimento 2).
Fig. 14.- Concentración de glucosa en plasma de las crías, en ayuno de 8 horas, del nacimiento y durante la lactancia, de las ratas que fueron tratadas con glucosamina 900 mg/kg el número de ratas para cada grupo fue de 7 (Experimento 2).
0
20
40
60
80
100
120
140
160
0 7 14 21 35
Tiempo en días
Pes
o co
rpor
al (g
).
CONTROL GLUCOSAMINA
50
60
70
80
90
100
110
120
0 7 14 21
Tiempo en días
gluc
emia
(mg/
dl)
CONTROL GLUCOSAMINA

21
DISCUSION Los resultados de los dos experimentos indican que no hubo diferencia en la ingesta de alimento entre ambos grupos y la oscilación en la cantidad de alimento ingerida no presentó diferencia significativa y sólo durante periodos cortos predominó una ingesta menor de alimento para las ratas tratadas con la glucosamina (figs 4y 5), estas observaciones concuerdan con los resultados obtenidos en los niveles de leptina en plasma al finalizar el tratamiento los cuales fueron semejantes en ambos grupos (fig 9), consecuentemente esto se reflejó en el aumento de peso corporal que para los dos grupos siguió el mismo comportamiento (figs 2 y 3), lo cual nos indica que la administración crónica de glucosamina a una dosis de 300 y 900 mg /kg de peso no afecta la ingesta de alimento ni el peso de los animales, siendo contrario a la hipótesis propuesta por Wang et al en 1998, y a los resultados encontrados en estudios con administración aguda de glucosamina en los que se encontró una disminución en la ingesta de alimento (Obici et al, 2003; Okabe et al 1988). Además, no se encontraron cambios en los niveles de triglicéridos de plasma en el grupo tratado con glucosamina (Tabla 2). La curva de glucemia para los dos grupos durante todo el tratamiento denota un aumento en la glucemia basal durante las últimas dos semanas en el grupo al cual se le administró la glucosamina vía oral (fig 6), este comportamiento muestra los efectos que se presentan en los inicios de la resistencia a la insulina, en el grupo tratado con glucosamina no sólo eran menores los niveles de glucosa basales al inicio del experimento, sino que estos muestran una tendencia a aumentar conforme transcurre el tiempo de la administración de la glucosamina; sin embargo, cabe mencionar que los valores no rebasan los rangos de glucemia normales, así mismo se observó esta tendencia en el experimento 2 donde en las primeras semanas se vió el mismo comportamiento que desaparece durante la gestación y lactancia (fig 12), estos resultados en un principio ayudan a sustentar que una dosis de 300 y 900 mg/kg de glucosamina por vía oral pueden ocasionar una resistencia a la insulina probablemente por el mecanismo planteado por muchos autores que proponen la activación de la vía de las hexosaminas (Kelley et al,1993; Patti, 1999; Sakai et al, 2003), con la acumulación de metabolitos de la biosíntesis de hexosaminas: Glucosamina 6 fosfato, UDP- GlcNAc, ya que el tratamiento con glucosamina mimetiza un estado de hiperglucemia debido a que aumenta el flujo a través de esta vía. La hiperglucemia y la glucosamina exógena conducen a un aumento marcado de los metabolitos de hexosaminas. El camino de las hexosaminas puede funcionar como un sensor de la glucosa y puede ser responsable de la resistencia a la insulina. Al comparar las curvas de tolerancia a la glucosa al inicio de los dos experimentos se observa que al administrarles la glucosa vía intraperitoneal los dos grupos presentaron niveles de glucosa en sangre con un comportamiento muy semejante durante los diferentes tiempos de medición. En la quinta y sexta semana respectivamente para el experimento uno y dos, aunque la elevación máxima no fue tan diferente en los dos grupos después de la administración de la glucosa, los niveles ya mostraron una tendencia mayor durante los últimos tiempos de las mediciones para el grupo que fue tratado con la glucosamina aunque no fue significativa. Es decir, la tendencia a la disminución de la glucemia hacia los niveles normales, a los 60 y 120 minutos fue más lenta para los grupos tratados con glucosamina con respecto a los control. En el humano durante una prueba de tolerancia a la glucosa, si los valores obtenidos de glucemia a los 120 minutos posteriores a la administración a la glucosa sobrepasa los 200 mg/ml es una prueba de diagnóstico positivo de diabetes tipo 2 (NDDI). Las curvas de tolerancia a la glucosa a la onceava semana de tratamiento con glucosamina muestran que los niveles de concentración de glucosa plasmática ya son mayores después de la administración de glucosa y a los 60 y 120 minutos posteriores, lo cual demuestra que se presenta una asimilación menos rápida de la glucosa por las células en comparación a los resultados del grupo control (experimento 1)

22
siendo esta una característica de la resistencia a la insulina, en la cual existe una alteración en la sensibilidad de los tejidos a esta hormona para el transporte de la glucosa. Este comportamiento se observó en el experimento dos hasta la semana quince, lo cual se piensa es atribuible al periodo de gestación y lactancia (figs. 8 y 9). Con respecto a los niveles de insulina que también fueron medidos en la muestra de sangre tomada para el inicio, sexta semana y término del tratamiento de la curva de tolerancia a la glucosa en el experimento dos se esperaba que el comportamiento fuera semejante a los de la glucemia, sin embargo, aunque si se muestra una tendencia en la ultima medición en el aumento de insulina para el grupo tratado con glucosamina la diferencia no es significativa (fig 10), sin embargo, los niveles de glucemia son altos, esto podría significar que las células ß del páncreas no responden adecuadamente a la señal de la glucosa ya que no produce exceso de insulina, o a que no es capaz de introducir la glucosa a las células dependientes de insulina (Hansen, 1999). Los niveles de glucemia, al administrar una carga de insulina para conocer el comportamiento del transporte de la glucosa a las células (curva de tolerancia a la insulina), no mostró diferencias entre los dos grupos de ratas al inicio de la curva (figs. 11 y 12). Sin embargo al aumentar los tiempos de medición en el experimento dos se observa una recuperación mas lenta hacia los niveles normales de glucemia para el grupo tratado con glucosamina lo cual sugiriere un efecto en las hormonas contra reguladoras, siendo este efecto una respuesta que no se tenía contemplado en este estudio, sin embargo, muestra que está afectado el sistema de regulación de la glucemia, probablemente afectando al páncreas lo cual apoya que durante la resistencia a la insulina observada en las curvas de tolerancia a la glucosa, la hiperglucemia es por falta de respuesta de las células hacia la insulina o en la secreción de la misma. Sin embargo, es probable que las dosis administradas de 3 y 0.75 Unidades de Insulina fueron muy grandes y no nos permitieron observar la diferencia entre grupos al inicio de su administración, o bien ya se esta presentando algún efecto a nivel pancreático. Con respecto a las crías no se observa ningún efecto en el peso y glucemia en plasma para ninguno de los dos grupos en estudio, solo se observó una tendencia en el aumento de peso para el grupo de crías de madres tratadas con glucosamina con respecto a los controles lo cual durante la lactancia no fue significativamente mayor. Es decir se sugiere que los efectos de la glucosamina son a largo plazo y no al inicio de su administración. Con estos resultados se puede establecer que la respuesta de los grupos tratados con glucosamina al final del periodo de tratamiento en la curva de tolerancia a la glucosa y en los niveles de insulina en plasma en comparación a los resultados de estos mismos grupos al inicio de la administración de la glucosamina demuestran dos importantes características de la resistencia a la insulina, una es la velocidad más lenta de regulación de los niveles de glucemia en los últimos tiempos en la curva de tolerancia a la glucosa y la segunda es el aumento de los niveles de la insulina para llevar acabo dicha acción.

23
CONCLUSIONES La glucosamina administrada vía oral diariamente durante once y quince semanas produce resistencia a la insulina en ratas. Las dosis usadas no afectan la ingesta de alimento ni el peso corporal de los organismos en estudio. No hubo cambio en los niveles de leptina basales. Durante la gestación no se presentan cambios en los niveles de glucosa en plasma en ratas ni en sus curvas de tolerancia a la glucosa con la administración de glucosamina vía oral Las crías de ratas que se les administra glucosamina vía oral no presentan ningún cambio en su glucemia y peso corporal. IMPACTO La capacidad de la glucosamina para producir resistencia a la insulina al ser administrada por vía oral, de forma y a una dosis parecidas a las que la toman los pacientes con osteoartritis con la finalidad de la disminución en la inflamación y el dolor articulares, la muestra como un fármaco peligroso, ya que si estos sujetos además tienen propensión genética a padecer diabetes, seguramente provocará un mayor riesgo a desarrollar la diabetes mellitus, por lo que se debería prohibir su utilización en estos sujetos así como en las mujeres en etapa reproductiva. BIBLIOGRAFIA
1. Barclay TS, Tsourounis C. Glucosamine.The Annals of Pharmacotherapy. 32:574-579.1998 2. Buse MG. Hexosamines, insulin resistance, and the complications of diabetes: current status.
Am J Physiol Endocrinol Metab. 290:E1-E8. 2006. 3. Butte NF. Carbohydrate and lipid metabolism in pregnancy: normal compared with gestattional
diabetes mellitus. Am J Clin Nutr. 71 (suppl):1256S-61S. 2000 4. Calzada LR, Altamirano BN, Ruiz R M: Etiología de la diabetes mellitus tipo 2 en pediatría:
evidencia en favor de resistencia primaria a la insulina. Acta Pediatr Méx; 23(4):232-237. 2002.
5. Echard BW, Talput NA, Funk KA, et al: Effects of oral glucosamine and chondroitin sulfate alone and in combination on the metabolism of SHR and SD rats. Mol Cell Biochem. 225(1):85-91.2001.
6. Gelardi NL, Chung-Ja MC, Oh William: Evaluation of insulin sensitivity in obese offspring of diabetic rats by hyperinsulinemic-euglycemic clamp technique. Pediatric Research. 30(1):40-44.1991.
7. Hansen, BC. The metabolic sindrome X. Ann NY Acad Sci.1999. ß 892:1-24. 8. Harper RM. Química fisiológica. 6a de. El Manual Moderno. 1979. México. 9. Hawkins M, Barzilai N, Lui R, et al: Role of the glucosamine pathway in fat-induced insulin
resistance. J Clin Invest 99(9):2173-2182. 1997. 10. Kann BB, Fler JS: Obesity and insulin resistence. J Clin. Invest. 106 (4):473-481.2000.

24
11. Kelley D, Mokan J, Simoneau A, et al: Interaction between glucose and free fatty acid metabolism in human skelet muscle. J Clin Invest 92:91-98. 1993.
12. Kieffer TJ, Séller RS, Habener JF, et al; Leptin supresion of insulin secretion by activation of ATP-sensitive K+channels in pancreatic beta cells. Diabetes. 46: 1067-1093. 1997.
13. Langer O, Levy J, Brustman L, et al. Glycemic control in gestational diabetes mellitus- How tight is tight enough: Small for gestational age versus large for gestational age?. Am J Obstet Gynecol. 161:643-53. 1989.
14. Marshall S, Bacote V, Traxinger RR: Discovery of the metabolic pathway mediating desensitization of the glucose transport system: role of hexosamine biosynthesis in the induction of the insulin resistance. J Biol Chem 266:4706-4712.1991.
15. McAlindon T E, La valley MP, Gulin JP, Felson DT: Glucosamine and chondroitin for treatment of osteoarthritis. A systematic quality assessment and meta-analysis. JAMA 283: 1469-1473. 2000.
16. Mueller WM, Gregoire FM, Stanhope KL, Mobbs CV, Mizuno TM, et al. Evidence that glucose metabolism regulates leptin secretion from cultured rat adipocytes. Endocrinology. 139: 551–558. 1998.
17. National Diabetes Data Group. Classification and diagnosis of diabetes mellitus and other categories of glucose intolerance. Diabetes. 1979. 28:1039-1057.
18. Obici S, Rossetti L: Minireview: Nutrient sensing and regulation of insulin action and energy balance. Endocrinology. 144(12): 5172-5178. 2003.
19. Okabe Y, Sakata T, Fujimoto K, et al: Differential mechanisms of feeding modulation induced by amino sugars in rats. Proc Soc Exp Biol Med. 188:23-29. 1988.
20. Patti ME: Nutrient modulation of cellular insulin action. Ann NY Acad. Sci. 892:187-203.1999. 21. Racotta P R. Metabolismo energético en el humano. 1a. Ed. Instituto Politécnico Nacional.
2002. México. 22. Ravussin E: Cellular sensors of feast end famine. J Clin Invest. 109(12):1537-1540.2002. 23. Rossetti L: Perspective: hexosamines and nutrient sensing. Endocrinology. 141(6):1922-
1925.2000. 24. Sakai K, Clemmons DR: Glucosamine induces resistance to insulin-like growth factor I (IGF-I)
and insulin in hep G2 cell cultures: biological significance of IGF-I/insulin hybrid receptors. Endocrinology. 144(6):2388-2395. 2003.
25. Traxinger RR, Marshall S: Coordinate regulation of the glutamine: fructose-6-P aminotransferase activity by insulin, glucose and glutamine. J Biol Chem. 266: 10148-10154.1991.
26. Virkamaki A, Daniels MC, Hamalainen S, et al: Activation of the hexosamine pathway by glucosamine in vivo induces insulin resistance in multiple insulin sensitive tissues. Endocrinology. 138(6):2501-2507.1997.
27. Wang J, Lui R, Hawkins M, et al: A nutrient-sensing pathway regulates leptin gene expression in muscle and fat. Nature. 393:684-688.1998.


















