análisis de los cambios epigenéticos en la metilación del dna ...
98
1 “ANÁLISIS DE LOS CAMBIOS EPIGENÉTICOS EN LA METILACIÓN DEL DNA INDUCIDOS POR LA DIABETES Y SUS POSIBLES MECANISMOS” ANGELINA PERNA CHAUX Microbióloga, MA. UNIVERSIDAD NUESTRA SEÑORA DEL ROSARIO FACULTAD DE MEDICINA MAESTRÍA EN CIENCIAS CON ÉNFASIS EN GENÉTICA HUMANA BOGOTÁ, 2016
Transcript of análisis de los cambios epigenéticos en la metilación del dna ...

1
“ANÁLISISDELOSCAMBIOSEPIGENÉTICOSENLAMETILACIÓNDELDNAINDUCIDOSPORLADIABETESYSUS
POSIBLESMECANISMOS”
ANGELINAPERNACHAUX
Microbióloga,MA.
UNIVERSIDADNUESTRASEÑORADELROSARIO
FACULTADDEMEDICINA
MAESTRÍAENCIENCIASCONÉNFASISENGENÉTICAHUMANA
BOGOTÁ,2016

2
Tesis presentada por ANGELINA PERNA CHAUX, para optar al grado de Maestría enCienciasconÉnfasisenGenéticaHumanadelaUniversidadNuestraSeñoradelRosario.
EltrabajoserealizóenelLaboratoriodeGenéticaHumanadelaUniversidaddelosAndes,bajoladireccióndelosDoctoresValerianoLópezSegurayHelenaGrootdeRestrepo.
DIRECTORDETESIS
ValerianoLópezSegura,M.Sc.,Ph.D.
CO-DIRECTORDETESIS
Dra.HelenaGrootdeRestrepo,M.Sc.
1.LaboratoriodeGenéticaHumana,UniversidaddelosAndes.
2.FacultaddeMedicina,UniversidadNuestraSeñoradelRosario.
Bogotá,2016

3
Dr.ValerianoLópezSegura,M.Sc.,Ph.D.
Biólogo
DIRECTORDETESIS
Dra.HelenaGrootRestrepo,M.Sc.
Bacterióloga
CO-DIRECTORADETESIS
Dra.ClaudiaTamarSilvaAldana,M.Sc.,Ph.D.
JURADO1
Dra.AlbaAliciaTrespalacios,M.Sc.,Ph.D.
JURADO2

4
Dedicatoria
ADiosporguiarycuidarcadaunodemispasos.Pordarmeunavidallenadeamorybendiciones.Amifamiliaporserelmotordemiexistirymiapoyoincondicional.AmihijaMaríaAntonellaporhacerdemis
díaslosmásalegres.Amipadrequeenpazdescanses,estoesparati.

5
AGRADECIMIENTOS
AmipapáelDr.AníbalPernaMazzeo+,porcultivarmeelamorporlanaturalezayenseñarmeelrespetoporlavida,losorganismosvivosylahumanidad.Ledoygraciasamipadreporinculcarmeestahermosapasión.
A la Universidad de los Andes por cobijarme de nuevo y permitirme realizar la investigación de mi tesis en susinstalaciones.
AlDr.ValerianoLópezporsuvaliosadirecciónydedicación.Lorecordarésiempreporenseñarmeepigenéticayampliarmivisióndelagenéticahumana.
A la Dra. Helena Groot por el apoyo incondicional de un gran ser humano que busca el bienestar de sus estudiantes.Graciasporlaoportunidadquemebrindóparahacerpartedeestegrupodeinvestigación.
AlprogramadeDiabetesdelaFundaciónSantaFeporestaoportunidad.Alospacientesporsuinterésenparticiparyserpartedeestainvestigación.
A la Universidad del Rosario por recibirme como su estudiante y enseñarme nuevos conocimientos, que no sólo mefomentaronmicrecimientoprofesionalsinotambiénmicrecimientopersonal.
AmisprofesoreselDr.CarlosRestrepo,laDra.NoraContreras,laDra.DoraFonseca,laDra.HeidiMateus,laDra.ClaudiaSilva,Dr.CarlosPrada,Dr.PaulL,DraLuisaMatheusylaDraSandra;acadaunodeustedesporsuvaliosadedicaciónyapoyocomodocentesquehicieronpartedemiformaciónacadémica.Siempreestarémuyagradecidaporsusenseñanzas.
AmiscompañerasdelaMaestríaenCienciasdelaUniversidaddelRosarioaLauraRico,MarcelaGálvez,KarinaMancera,yCatalinaHurtadoporeltiempocompartidoenlostresaños.
AmiscompañerosyamigosdelLaboratoriodeGenéticaHumanadelaUniversidaddelosAndesaDianaPolanía,NicolásRojas, Jairo Pinzón CarolinaMuñoz, Diana Narváez, Daniela Castiblanco, Sandra Camargo, Luz Adriana Pérez, MaríaFernandaJácome,KatherineDuque,AdrianaRobayo,AngélicaDíazyAndrésHernándezporelapoyoincondicionalyeltiempoquecompartimostrabajando.
AmigrupodetrabajoJairo,Nicolás,yDianaP,conustedesaprendínuevastécnicasylesagradezcoporsudedicaciónycompromiso,realizandoasíungrantrabajoenequipo.
A mi esposo Juan David Jaramillo por su incondicional apoyo durantemis estudios. Fuiste mi fuerza y empuje en losmomentosbuenosynotanbuenos,animándomesiempreencumplirmissueñosyalcanzarmismetas.AmihijaMaríaAntonella,iluminastemiexistirehicistedemisdíaslosmásfelices.
Amifamilia,mimamaLuzMaríaChaux,leagradezcoporenseñarmeperseverarylucharcadadía.MihermanaIsabellaPernaymisobrinaLucianaCárdenasporesperaryapoyarmeeneltiempodemisestudios.AYolandaMoncayo,EdgarJaramillo y Catalina Jaramillo por apoyarme en este proceso. A mi amiga y hermana del alma Julieta Villegas poracompañarme.

6

7
TABLADECONTENIDO
ÍNDICEDEFIGURAS10ÍNDICEDETABLAS12SÍMBOLOS,ACRÓNIMOSYABREVIATURAS13I. INTRODUCCIÓN16II. PLANTEAMIENTODELPROBLEMA18III. JUSTIFICACIÓN19IV. OBJETIVOS 204.1Objetivogeneral 204.2Objetivosespecíficos 20V. MARCOTEÓRICO 215.1EPIGENÉTICA 215.1.1Historiadelaepigenética 215.1.2Conceptodeepigenética 215.1.3Loscuatropilaresdelaepigenética 215.1.3.1Remodelacióndecromatina 225.1.3.2Modificacióndelascolasdehistonas 225.1.3.3RNAsnocodificantes 235.1.3.4ModificacióncovalentedelDNA 235.1.3.4.1ElpapeldelasCpGsylametilacióndelDNA 245.1.3.4.2EnzimasencargadasdelametilacióndelDNA 26A. DNAmetiltransferasas 26
a.DNMT1:DNA(citosina-5-)-metiltransferasa1 28B.UHRF1(Ubiquitin-like,containingPHDandRINGfingerdomains) 285.1.4Desmetilaciónactivaypasiva 295.1.4.1Maquinariadedesmetilaciónactiva 29
A.Glicosilasasten-eleventranslocasas(TETs) 30a. TET1 31
B.AID/APOBEC 31C.TDG 31D.MBD4 32
5.1.5Ciclodeuncarbono,claveenlametilación 325.2DIABETESMELLITUSCOMOMODELOBIOLÓGICO 335.2.1Conceptoactualdeladiabetesmellitus 335.2.2Causasdeladiabetes 335.2.3Mayorescomplicacionesenladiabetes 345.2.4Clasificacióndeladiabetes 35
A. DiabetesMellitustipo1 36B. DiabetesMellitustipo2 36
5.2.5CriteriosdeDiagnóstico 385.3OXIDACIÓNCELULARRELACIONADAALADIABETES 395.3.1Roldelestrésoxidativo 395.3.2Especiesreactivasdeoxígeno 395.3.3CadenatransportadoradeelectronesylaformacióndeROS 405.3.4Aumentodeoxidaciónencasodehiperglucemia 405.3.5ComplicacionesdelaDMporbajaregulaciónoxidoreducciónylasupervivenciacelular415.3.6VíasbioquímicasafectadasporROS 425.3.6.1PiezasdelrompecabezasdescritasporBrownlee 425.3.6.2Hiperglucemiaydañomicrovascular 43

8
A. Incrementodelavíadelospolioles 43B. PrecursoresdeAGEs 43C. IncrementodelaexpresióndereceptoresAGEsysusligando 44D. ActivacióndePKC 44E. Incrementodehexosaminas 45
5.3.6.3Lahiperglucemiainducelaactivacióndelascincovías,inhibiendoaGAPDHyactivandoaADP-ribosa. 45
5.3.6.4Resistenciaalainsulinaydañomacrovascular 465.4EPIGENÉTICAYDIABETES 485.4.1Laproblemáticadelaepigenéticaenladiabetes 485.5.2Memoriaepigenéticahiperglucémica 49VI. METODOLOGÍA 506.1POBLACIÓNDEESTUDIO 50A.Criteriosdeinclusión 50B.Criteriosdeexclusión 506.2RECLUTAMIENTOYRECOLECCIÓNDELAHISTORIACLÍNICADELOSPACIENTES 516.3MÉTODOSDELABORATORIO 516.3.1Obtencióndelamuestra 516.3.2Pruebasclínicasdelaboratorioyhemoglobinaglucosilada(HbA1c) 516.3.3ExtraccióndeDNA 516.3.4ExtraccióndeRNA 526.3.5CuantificaciónporNanodrop 526.3.6Microelisasdemetilaciónehidroximetilación 526.3.7RT-qPCR 546.3.7.1Diseñodecebadores 546.3.7.2EstandarizacióndecebadoresporPCRconvencional 556.3.7.3SíntesisdelaprimerahebradecDNA 556.3.7.4ReaccióndePCRconvencionalparaelcontrolpositivoGAPDH 566.3.7.5GeldeelectroforesisenagarosaparaloscontrolesdeGAPDH 576.3.7.6ReaccióndeqRT-PCRdelosgenesDNMT1yTET1 576.3.8Microarreglos 596.4DIAGRAMAMETODOLÓGICO 616.5ANÁLISISESTADÍSTICOSYBIOINFORMÁTICOS 626.5.1Análisisestadísticosdelasmicroelisas 626.5.2AnálisisestadísticosdeRT-qPCR 636.5.3Análisisestadísticosybioinformáticosdemicroarrgelos 63VII. ASPECTOSÉTICOS 65VIII. RESULTADOS 698.1EFECTODELCONTROLMETABÓLICOENLAMETILACIÓNEHIDROXIMETILACIÓNDELDNAEN
T2DM 698.2SEXOYFACTORESDEESTILODEVIDA 718.3EDADYDURACIÓNDELAENFERMEDAD 748.4qRT-PCRDELASENZIMASINVOLUCRADASENLADESMETILACIÓNDELDNA 758.5ANÁLISISINSILICODELOSNIVELESDEMETILACIÓNENTEJIDOSAFECTADOSPORLADM768.6HIDROXIMETILACIÓNYDIABETES:POSIBLESMECANISMOSDEDESMETILACIÓN 778.7GENESDIFERENCIALMENTEEXPRESADOS 81IX. DISCUSIÓN 829.1EFECTODELCONTROLMETABÓLICOENLAMETILACIÓNEHIDROXIMETILACIÓNDELDNAEN
T2DM 829.2EXPRESIÓNDEGENESINVOLUCRADOSENRUTASDEDESMETILACIÓN 83

9
9.3UNPOSIBLEMECANISMONUEVODEDESMETILACIÓNENESTADOSDEENFERMEDAD 849.4GENESDIFERENCIALMENTEEXPRESADOSYSUEFECTOSOBRELAPROGRESIÓNDELA
ENFERMEDAD 86X.CONCLUSIONES 88XI.RECOMENDACIONES 89XII.BIBLIOGRAFÍA 90XIII.ANEXOS 9413.1EPIDEMIOLOGIADELADIABETES 9413.1.1Datosmundiales 9413.1.2DatosenColombia 9413.1.3Posiblescausasepidemiológicas 9513.2GRANDESESTUDIOSDEDIABETESENLAPOBLACIÓNHUMANA 9513.2.1DiabetesControlandComplicationsTrial(DCCT) 9513.2.2EpidemiologyofDiabetesInterventionsandComplications(EDIC) 9613.2.3TheUnitedKingdomProspectiveDiabetesStudy(UKPDS) 97

10
ÍNDICEDEFIGURAS
Figura1.Modelogeneraldelaepigenética
Figura2a.EstructuramoleculardelasmodificacionesdebasesdelDNA.
Figura2b.Representaciónesquemáticadelatransferenciadelgrupometiloalcarbono5delacitosina
Figura3.Anatomíamoleculardelacromatina,lossitiosCpGsyelrolenlaexpresióngénica
Figura4.RepresentaciónesquemáticadelasestructurasdelasDNMT’s.
Figura5.Modeloclásicodelametilación
Figura6.DiagramadelasposiblesvíasdedesmetilacióndeDNA
Figura7.Reprogramaciónencélulasgerminalesypreimplantaciónembrionariaenratón
Figura8.Tresrutasmetabólicasquecomponenelciclodeuncarbono
Figura9.Modeloqueproponeelroldelaepigenéticaenlosmecanismospatogénicosdeladiabetes
Figura10.Mayorescomplicacionesdeladiabetes
Figura11.GenesasociadosavariantesdeT2DM
Figura12.VíasdelapatogeniadelaT2DMylascomplicacionesrelacionadas
Figura13.Cadenatransportadoradeelectrones
Figura14.Produccióndesuperóxidoporlacadenatransportadoradeelectronesmitocondrial
Figura15.Modelogeneralcomolahiperglucemiainducedañotisular
Figura16.Lahiperglucemiaincrementaelflujodelospolioles
Figura17.ConsecuenciaspatológicasporelincrementodeAGEs
Figura18.ConsecuenciadelahiperglucemiaporlaactivacióndePKC
Figura19.Incrementodelflujodelashexosaminasporlahiperglucemia
Figura20.Sobreproduccióndesuperóxidosmitocondrialesactivadosporlahiperglucemia
Figura21.Esquemadelosmecanismosafectadosporlahiperglucemia
Figura 22. Rol de la resistencia a la insulina y FFAs en la formación de ROS endotelial y aterogénesismacrovascular
Figura23.ResumenesquemáticodelasvíasquecontribuyenalascomplicacionesdelaT2DM
Figura24.Memoriametabólicaymecanismosepigenéticosenlacomplicacióndeladiabetes
Figura25.DiagramadeflujoparaelprocedimientosdelasELISAss
Figura26.AccióndelSYBRGreen
Figura27:DiagramadeflujometodológicodelestudiomoleculardepacientesdelHFSF
Figura28:Diagramadeflujometodológicodelestudioinsilico
Figura29.PorcentajedelametilaciónenhidroximetilaciónglobaldelDNA
Figura30.DiferenciasenlametilaciónehidroximetilaciónglobaldelDNA
Figura 31. Porcentaje de la metilación en hidroximetilación global del DNA, excluyendo datos deconsumidoresdealcoholytabaco

11
Figura32.Correlacióndelosporcentajesdemetilaciónehidroximetilaciónentrelaedad(AyB)ytiempodediagnóstico(CyD)
Figura33.ExpresióndegenesDNMT1yTET1entrepacientesycontroles.
Figura34.AnálisisdemicroarreglosdesangreperiféricaenpacientesT2DMyungrupocontrol
Figura35.AnálisisdelaexpresióndiferencialdegenesensangreperiféricadepacientesT2DMyelgrupocontrol
Figura36.AnálisisdeexpresiónentejidopancreáticoentrepacientesT2DMyelgrupocontrol
Figura37.AnálisisdeexpresiónentejidoadiposoentrepacientesT2DMyelgrupocontrol
Figura38.AnálisisdeexpresiónentejidomuscularentrepacientesT2DMyelgrupocontrol
Figura39.RepresentaciónesquemáticadelasposiblesvíasdedesmetilacióndelDNA.
Figura40.DCCTyEDIC

12
ÍNDICEDETABLAS
Tabla1:DescripcióngénicadelasDNMTs
Tabla2.DescripcióngénicadelasTETs
Tabla3:ClasificacióndelascategoríasprincipalesdelaDiabetesMellitus
Tabla4:CriteriosparaeldiagnósticodelaDiabetesMellitus
Tabla 5: Criterio para definir la población de alto riesgo de diabetes de la Asociación Americana deDiabetes
Tabla6:Especiesreactivas
Tabla7.Bufferdelavadoal1X
Tabla8.Preparacióndelacurvaestándarde5mCy5hmC
Tabla9.Dilucióndeanticuerpos,soluciónaumentadorayreveladoradefluorescencia
Tabla10.Cebadoresdecontrolyamplificación
Tabla11.VolumendePCRconvencional
Tabla12.PCRconvencionalGoTaqGreen
Tabla13.VolúmenesparalasíntesisdelaprimeracadenacDNA
Tabla14.SegundareacciónparalasíntesisdelcDNA
Tabla15.Condicionesdelasíntesisdeprimerahebraeneltermociclador
Tabla16.VolúmenesparalaPCRconvencional
Tabla17.CondicionesdelcontrolpositivoparaGAPDH
Tabla18.Estandarizacióndeconcentraciones
Tabla19.CondicionesdePCRcuantitativa
Tabla20.Concentracionesóptimasdecebadores
Tabla21.VolúmenesdelaqPCR
Tabla22.CondicionesdelaPCRcuantitativa
Tabla23:Tabladescriptivademicroarreglos
Tabla24.Tablacaracterísticadelosgruposdeanálisis
Tabla25.Rutasafectadasentejidopancreáticoysangreperiférica
Tabla26.Estimacióndeprevalenciadediabetesparael2030.
Tabla27.ResultadosdelaDCCT
Tabla28.ResultadosdeEDIC
Tabla29.Mayoresdescubrimientosporlosestudiosmundialesmásimportantes

13
SÍMBOLOS,ACRÓNIMOSYABREVIATURAS
• aa-aminoácido• ADA-AsociaciónAmericanadeDiabetes• ADCA-DN-Sordera-AtaxiaCerebelosaAutosómicaDominanteyNarcolepsia• ADP-ribosa-DinucleótidodeAdeninaCíclicaunidoaunaribosa• AID/APOBEC-DeaminacióndeCitosinaInducidaporActivacióndelaApolipoproteinaB• AGA-AlteracióndeGlucemiaenAyuno• AGEs-productodeGlicacionesAvanzadas• BER-SistemasdeReparaciónporEscisióndeBase• CA-Ciosina-Adenina• CAT-Catalasa• CD-DeaminasadeCitosina• cDNA-DNAcomplementario• CGI-IslasCpGs• CpGs-DinucleótidosdeGuaninasyCitosinas• CVD-EnfermedadCardiovascular• CTE-CadenaTransportadoradeElectrones• DANE-DepartamentoAdministrativoNacionaldeEstadísticas• DAG-Diacylglicerol• DCCT-DiabetesControlandComplicationTrial• DM-DiabetesMellitus• DNA-ÁcidoDesoxirribonucleico• DNMTs-DNAMetiltransferasas• DNMT1-DNA(citosina-5)-Metiltransferasa1• DNMT3A-DNA(citosina-5)-Metiltransferasa3A• DNMT3B-DNA(citosina-5)-Metiltransferasa3B• FSFB-HospitalUniversitarioFundaciónSantaFedeBogotá• EDIC-EpidemiologyofDiabetesInterventionandComplication• EDTA-ÁcidoEtildiaminotetraacético• eNOS-OxidoNítricoSintasaendotelial• FADH2-DinucleótidodeFlavinayAdenina• FFA-AcidosGrasosLibres• GAD65-AutoanticuerpodeAcidoGlutámico• GAPDH-Gliceraldehido-3-fosfatoDeshidrogenasa• GDM-DiabetesMellitusGestacional• GFAT-Fructose-6-fosfatoAminotransferasa• GLUT’s-TransportadoresdeGlucosa• GSH-PX-GlutatiónPeroxidasa• HAT-AcetilasasdeHistonas• Hb-Hemoglobina• HbA1c-HemoglobinaGlucosilada• HDAC-DeacetilasasdeHistonas• HLA-AntígenoLeucocitarioHumano• hLP-LactógenoPlacentarioHumano• HMG-1-High-mobilitygroupprotien1• HMT-MetiltransferasadeHistonas• HSN1E-NeuropatíaSensorialyAutosómicaHereditariaconDemenciayPérdidadeAudicióntipo1E• IAAs-AutoanticuerpodeInsulina

14
• IA-2yIA-2ª-AutoanticuerposdeFosfatasadeTirosina• ICAs-AutoanticuerpodeIslaLangerhans• IDF-FederaciónInternacionaldeDiabetes• IFG-AlteracióndeGlucosaenAyuno• IGT-ToleranciaAlteradadelaGlucosa• IHC-IntoleranciaalosHidratosdeCarbono• IMF-IntensidaddeFluorescenciaMedia• IR-ResistenciaalaInsulina• IR-S-ReceptoresdeSubstratodeInsulina• IRCT-InsuficienciaRenalCrónicaTerminal• LADA-DiabetesAutoinmuneLatentedelAdulto• LMRs-ZonasPobresenMetilación• MAT-MetioninaAdenosiltransferasa• MBDs-ProteínasdeUniónaGruposMetilos• meDNA-metilaciónenelDNA• MM-MemoriaMetabólica• MODY-DiabetestipoMody;MaturityOnsetDiabetesoftheYoung• miRNAs-MicroRNAs• MnSOD-ManganesoSuperóxidosDismutasa• MS-MetioninaSintasa• MTHRF-MetiltetrahidrofolatoReductasa• mTHF-metiltetrahidrofolato• NADPH-NicotinamidaAdeninaDinucleotidoFosfato• NDRs-SitiosLibresdeNucleosomas• NIDDK-InstitutodeDiabetesyDigestiónydeEnfermedadesdelRiñón• NIH-InstitutoNacionaldeSaluddelosEstadosUnidos• NFκB-FactorNucleardeTranscripciónκB(PotenciadordelasCadenasLigerasKappadelasCélulasB
Activadas)• nt-nucleótidos• qRT-PCR-ReacciónCuantitativaenTiempoRealdelaCadenadePolimerasa(PCR)• O2-Oxígenomolecular• OMS-OrganizaciónMundialdelaSalud• ORF-MarcodeAbiertodeLectura• PARP-PolyADP-ribosa• P.b-Paresdebases(p.b)• PCR-ReacciónenCadenadePolimerasa• PKC-ProteicinasaC• PTM-ModificacionesPost-traduccionalesdeHistonas• RAGE-ReceptoresdeAGEs• REDOX-Oxido-Reducción• ROS-EspeciesReactivasdeOxígeno• RNA-AcidoRibonucleótido• RNS-EspeciesReactivasdeNitrógeno• SAM-S-adenosilmetionina• SAT-TejidoAdiposoSubcutaneo• Set7-MetiltranferasadeHistonas• SAH-S-adenosilhomocisteina• SMRT-Singlemoleculereal-timesequencing

15
• SMUG1-GlicosilasasdeCadenaSencillaqueRemuevenUracilos• SOD-SuperóxidosDismutasa• SUMO-SUMOilacióndeProteínas;SmallUbiquitin-RelatedModifier• T1DM-DiabetesMellitus1• T2DM-DiabetesMellitus2• TET’s-TranslocaciónTen-Eleven• TET1-TranslocaciónTen-Eleven-1• TET2-TranslocaciónTen-Eleven-2• TET3-TranslocaciónTen-Eleven-3• TCA-AcidoTricarboxilico• TDG-DNAGlicosilasadeTimina• TF-FactoresdeTranscipción• TGF-FactordeCrecimientoTransformante• THF-Tetrahidrofolato• TSS-SitiodeInicioTranscripcional• TTS-SitioFinaldeTranscripción• UCP-ProteínasDesacoplante(Termogenina)• UCP-1-Termogenina-1• UDPGlcNAc-UDP-acetilglucosamina• UKPDS-TheUnitedKingdomProspectiveDiabetesStudy• VCAM-Moléculadeadhesiónencélulavascular• VEGF-FactordeCrecimientoVascularEndotelial• 5caC-5-carboxicitosina• 5hmC-5-hidroximetilcitosina• 5fC-5-formilcitosina• 5mC-5-metilcitosinas

16
I.INTRODUCCIÓN
Laepigenéticaeslacienciaqueestudialoscambiosheredablesenlafuncióndegenesquenoocurrenporcausadeuncambioenlasecuencianucleotídica(Ling&Groop,2009).Estasmodificacionesepigenéticaspuedenserheredadasdeunageneracióndecélulasaotracomoherenciamitóticayentregeneracionesdeuna especie como herencia meiótica. Sin embargo, a pesar de los numerosos estudios aún existe pocainformación sobre la herencia de rasgos epigenéticos entre generaciones. Hoy en día se sabe que laepigenéticapuedeserafectadaporelambiente,generandoalteraciónenlospatronescomosehavistoenlasenfermedadesmultifactorialescomplejascomoladiabetes(Ling&Groop,2009).
Losmecanismosepigenéticos incluyenmodificacionescovalentesdelDNA,modificacióncovalentede lascolasdehistonas,remodelaciónnocovalentedelacromatinayregulacióngénicaporRNAsnocodificantes(Gluckman et al., 2011). A través de todos estos mecanismos, la epigenética influye en procesosfundamentalesdelacélulacomoelsilenciamientotranscripcionaldegenes,lainactivacióndelcromosomaX, la impronta genómica, la regulación de elementos móviles transponibles, el establecimiento ymantenimientodelaidentidadcelularylaregulacióndelaestabilidadcromosómicaygenómica(Bhutani,Burns,&Blau,2011;Jaenisch&Bird,2003;Laurentetal.,2010)
La metilación del DNA es el mecanismo mejor estudiado y se ha considerado parte integral de lasdecisionesfundamentalesdelascélulas,jugandounpapelfundamentaleneldesarrollodelosorganismos(Bhutanietal.,2011).Enlosvertebrados,lametilacióndelDNAsedaporunprocesocatalíticoenzimáticoen la posición del carbono 5 de la citosina, principalmente en dinucleótidos CpGs. Las DNAmetiltransferasas(DNMTs)sonlasenzimasresponsablesderealizarlaaccióncombinadadeestablecerymantenerlospatronesdemetilaciónenelDNA(Pateletal.2013;Chenetal.2013).
DiversosestudioshandemostradoqueestasDNMTssonaltamenteexpresadaseneldesarrollocelularymenosexpresadasencélulasdiferenciadas(Jones&Liang2009).LosnuevospatronesdemetilaciónsonestablecidosporlasDNMT3ayDNMT3b,enelprocesoconocidocomometilacióndenovo.Porsuparte,lasDNMT1 son las enzimas responsables delmantenimiento de esos patrones tras las divisiones celulares,usandoparaellolashebrashemimetiladascomomolde.NuevosestudiossobrelasDNMT1handescritosuimportancia en el desarrollo embrionario y en cáncer, siendoprimordialespara elmantenimientode laidentidadcelular(Bhutanietal.,2011).
Estudiosrecienteshanevidenciadoque lascitosinasde losmamíferospueden tambiénserhidroxiladas.Estamarcasehavistoexpresadaabundantementeentejidocerebralycélulaspluripotentes.LafunciónyladistribucióndelDNAhidroximetiladoaúnnoestáclara.Sinembargo,hoyendía,lahidroxilacióndelas5mC, se ha postulado como el principal y único mecanismo de desmetilación del DNA basado en unprocesoenzimático,frentealmecanismomásclásicobasadoenlafaltademantenimientoporlasDNMT1.Elobjetivoprincipaldeestetrabajoesdeterminarsiexisteotraposiblevíaqueestéalterandolaoxidaciónde los grupos metilos y que permita entender los mecanismos de desmetilación en condicionespatológicas,comoenladiabetes.
La Diabetes Mellitus (DM) es una enfermedad metabólica crónica asociada a factores genéticos yambientales(Villeneuve,Reddy,&Natarajan,2011).Suorigenmultifactorialyheterogéneosecaracterizapor una desregulación homeostática de la glucosa y se correlacionada con el estilo de vida, unaalimentaciónhipercalórica,sedentarismoyobesidad(OnlineADA,2015).LaOrganizaciónMundialdelaSalud(OMS)estimaqueladiabetesafectaaproximadamentea346millonesdepersonasanivelmundialyqueestenúmeroaumentaráparael2030a400millones.Esconsideradaunaepidemiayunproblemadesaludpúblicaqueresultaen ladisminuciónde laexpectativadevidadadasuscomplicaciones (Intine&Sarras,2012;OnlineADA,2015).
Másalládelascausasdelaenfermedad,elestrésoxidativo,asociadoalaelevadaproducciónmitocondrialde especies reactivas de oxígeno, juega un papel fundamental en el desarrollo de las complicaciones

17
típicas tanto microvasculares y macrovasculares asociadas a la diabetes (Giacco & Brownlee, 2010). Sibien, hasta ahora nunca se ha asociado esta desregulación oxidativa con procesos epigenéticos, laevidencia ha demostrado la existencia de hipometilación del DNA en las células afectadas por lascomplicacionesdiabéticasyparalelamentesehaestablecidolaimportanciadeunbuencontrolglucémicoparaprevenirlaseveridaddelascomplicacionesdurantelavidadelpaciente(Villeneuveetal.,2011).
Hoyendíasehadescritoeldesarrollodenumerosasenfermedadesasociadasaalteracionesdepatronesdemetilación.Losestudiosdelepigenomaen trastornosmetabólicos,cáncer,Alzheimeryobesidadhansidolosmásestudiados,aunquenosetienemuchainformaciónalrespecto.Enesteproyectoutilizamosladiabetes como modelo biológico para relacionar el efecto de los estados hiperglucémicos y ladesmetilación del DNA. Es por ello nuestro interés en estudiar los mecanismos de desmetilación ehidroximetilacióndelDNAporprocesosoxidativosasociadosahiperglucemiamalcontroladaenpacientesdiabéticos.Esteprocesosupondríalaexistenciadeunavíaalternativadedesmetilaciónqueexplicaríalashipometilaciones encontradas enmuchas enfermedades complejas, no sólo la diabetes sino también lasenfermedades que presentan como elementos comunes oxidación celular, inflamación y muerte pordistintasvías.

18
II.PLANTEAMIENTODELPROBLEMA
Los patrones de metilación del DNA en células somáticas son generalmente estables y heredables. Encélulas germinales y durante el desarrollo embrionario temprano, el proceso de la metilación ydesmetilación del DNA ocurre principalmente demanera activa para establecer patrones demetilaciónespecíficos que luego le darán la característica de una célula somática diferenciada. Una de las grandespreguntasen laepigenéticaescómosegeneranestospatronesdemetilaciónydesmetilaciónenelDNAduranteeldesarrollodeunorganismoycómoéstossemantienenencélulasyadiferenciadas(Rodríguez-Dorantes,Téllez-Ascencio,Cerbón,Lez,&Cervantes,2004).
Estudios realizados en células humanas, ratas y peces cebra han documentado que los estados dehiperglucemia inducen la desmetilación de citosinas del genoma, generando cambios permanentes queafectan la memoria metabólica (Dhliwayo, Sarras, Luczkowski, Mason, & Intine, 2014), e inducen laaparicióndeenfermedadescomolaDiabetesMellitus.LaDMesuntrastornometabólicoquesecaracterizapor la deficiencia en la producción de insulina, la resistencia a su acción o ambas. Como consecuenciageneraunestadodehiperglucemiapersistenteyaltosnivelesdeoxidacióncelularqueconllevanaldañoen el tejido endotelial por la incapacidad de regular la entrada de glucosa. Esto a su vez causa lascomplicacionesmicrovascularesymacrovascularesqueafectandemaneracrónicayprogresivaaórganosdiana.
Sin embargo, aúnno es claro cómo la hiperglucemiapuede afectar varios tejidos y cómoésta genera elefectoacumulativoqueexplicalamemoriametabólica.Aesterespecto,laepigenéticapuedeserdeayudapara entender este tipo de cambios acumulativos. Por esta razón, este proyecto busca analizar losmecanismos moleculares que generan cambios en la metilación del DNA bajo condiciones dehiperglucemia.EsteproyectoproponecomohipótesisgeneraldetrabajounanuevavíadedesmetilacióndelDNAbasadaenlaoxidacióndirectadelasmetilcitosinasdesencadenadaporlasespeciesreactivasdeoxígeno.

19
III.JUSTIFICACIÓN
Desdequeiniciaronlosestudiosbioquímicos,molecularesygenéticosdeladiabetessefueronconociendolas múltiples dimensiones y la heterogeneidad de dicho desorden, del cual aún no se entiende porcompletosupatogenia.Apesardelconocimientoqueexistesobrelascomplicacionesdeladiabetes,desuefectoenelmetabolismodelaglucosayenlaproduccióndeespeciesreactivasdeoxígeno(ROS),noestáclarocómoestosmecanismospuedeninfluirenlamemoriametabólicayenlasalteracionesepigenéticasdelaenfermedadgenerandounamemoriametabólica.
LafuncióndelametilacióndelDNAendiversoscontextosseconocedesdelosañossetenta,sinembargopocosesabedelasdinámicasquesubyacenaloscambiosepigenéticos,comoenelcasodelospromotoresgénicos.Siporunladolametilacióntieneunafunciónclaraenelsilenciamientogénicoyenlaestabilidaddel genoma, está claro que la falta demetilos también tendría su propia importancia.Hoy en día se hacaracterizadomuybienlavíademetilaciónbasadoenDNMTs,peropocosesabesobrelosmecanismosdeladesmetilación.Recientementesehanpropuestodosmodelosdedesmetilaciónbasadosenunavíaactivay otra pasiva, aun así estos modelos no logran explicar las distintas situaciones fisiológicas de ladesmetilación celular y las situaciones en condiciones patológicas. En la actualidad se ha observadohipometilación global generalizada en patologías como diabetes, cáncer, enfermedades autoinmunes,inmunológicas,neurológicasentreotras.
EnesteproyectosehautilizadolaDiabetesMellituscomomodelobiológicoexperimentalparaestudiarycuantificar los mecanismos de metilación e hidroximetilación en un estudio de casos y controles depacientes con diabetes y sujetos sanos, con el objetivo de determinar la asociación de los estados dehiperglucemiaconloscambiosenlospatronesdemetilación.Secorrelacionaronlosresultadosclínicosymoleculares conel findegenerarunahipótesisalternativa referenteal estilodevidayasí identificar laposiblevíaafectada.

20
IV.OBJETIVOS
4.1OBJETIVOGENERAL
Evaluar los cambios en los niveles de metilación e hidroximetilación del DNA en sangre periférica,asociarlos con el control glucémico de pacientes diabéticos y comprobar in silico las alteracionesmolecularesqueexpliquenloscambiosepigenéticosenDiabetesMellitus.
4.2OBJETIVOSESPECÍFICOS
1. Determinar los niveles de metilación e hidroximetilación global del DNA en sangre periférica depacientesdiabéticosfrenteacontrolessanos.
2. Correlacionar las alteraciones en los niveles de metilación o hidroximetilación de los pacientesdiabéticosconsucontrolglucémico.
3. Demostrar la falta de desregulacióndeDNMT1yTET1 entre pacientes diabéticos y por lo tanto suindependenciadelprocesodedesregulaciónepigenética.
4. Analizar el efecto de la diabetes en diferentes tejidos afectados y los genes implicados a través delestudiodemicroarreglosdemetilaciónyexpresióninsilico.

21
V.MARCOTEÓRICO
5.1EPIGENÉTICA
5.1.1Historiadelaepigenética
Lahistoriadelaepigenéticaestáligadaalestudiodelaevoluciónyeldesarrollodelosorganismos.Hastaladécadadelos50lapalabraepigenéticaseutilizóparaclasificarloseventosdeldesarrolloembrionario.Seguidoesto,hubograndesdebatesentreembriólogossobrelanaturalezaylosmecanismosresponsablesdeldesarrollodelosorganismos.Dosescuelasconpensamientosdistintosseformaronintentandoexplicarlaepigenética.Unospensabanquecadacélulaconteníaelementospreformadosresponsablesen llevaracaboeldesarrollo,corrienteconocidacomoel“preformacionismo”ylosquepensabanqueeldesarrollosedaba por reacciones químicas de componentes solubles que ejecutaban el desarrollo, también conocidacomola“epigénesis”(Felsenfeld,2014).
ElembriólogoConradHalWaddington,establecióenladécadadelos40quelaepigenéticaeslaramadela biología que estudia las interacciones causales entre genes y su producto, generando un fenotipodeterminante (Goldberg, Allis, & Bernstein, 2007). Fue reconocido por establecer el paisaje epigenéticoparailustraryclasificartodosloseventosdeldesarrollodeunorganismo,desdelafertilizacióndelcigotoylosprocesosreguladoresquepartendelmaterialgenéticohastaeldesarrollodeunacélula(Felsenfeld,2014).
5.1.2Conceptodeepigenética
En1975sepublicarondosartículosdistintosconresultadossimilarespor losautoresArthurD.RiggsyRobinHolliday&FrankPugh.AmbosproponenquelametilacióndelDNApuedeactuarcomounamarcaepigenéticaquetieneunfuerteefectoenlaexpresióngénicayqueloscambiosdelametilacióndelDNAexplican la activación o inactivación de la expresión durante el desarrollo. A su vez establecieron quedistintasenzimaseranlasresponsablesdelametilación(Felsenfeld,2014).Estafuelaprimeravezqueselediounafunciónaciertasmarcascovalentesqueyaseconocían,comolametilación,peroquehastaesemomentonosesabíaparaqueseutilizaban.
Hoy en día la epigenética es descrita como la ciencia que estudia los patrones de expresión génicaheredables y cambios en el fenotipo que no se deben a la alteración en la secuencia del DNA. En otraspalabras,estudialoscambiosenelDNAquenoafectanlasecuencianucleotídicadeungenperoproducencambiosensuexpresión.Sesabequeloscambiosepigenéticossonheredablesysemantienendurantelameiosis.EstacienciasebasaenelestudiodelasunionesdegruposquímicosalDNAyalashistonasqueintervienen en la elección del destino celular, desarrollo embrionario y estabilidad genómica (Holliday,2006).
5.1.3Loscuatropilaresdelaepigenética
Diversas señalesepigenéticasparticipanen laexpresióndiferencial específicade cadacélula incluyendolasmodificaciones covalentes delDNA, lasmodificaciones post-traduccionales de colas de histonas (i.e.,acetilaciones,metilaciones,fosforilacionesyubiquitinaciones,entreotras.),laincorporacióndevariantesdehistonas, la regulaciónde la cromatinay la regulaciónporRNAsno codificantes (Maresca, 2015). Lafiguraacontinuaciónilustralosnivelesdelosmecanismosepigenéticos.

22
Figura1.Modelogeneraldelaregulaciónepigenética(Zaidietal,2010).
5.1.3.1Remodelacióndecromatina
La remodelación de la cromatina es unmecanismo importante ya que afecta a todos los estados de latranscripción. Se caracteriza por la remodelación transitoria y la apertura de la cadena de DNA de suoctámero de histonas para formar un loop de DNA y así mover los nucleosomas a una posicióntranslacional, todo para dar accesibilidad a los factores de transcripción (TFs). Los nucleosomas estáncompuestosde147pbdando1.65vueltasalrededordeunoctámerodehistonascon14puntosdeuniónalDNA.Estainteracciónmúltiplehacedelosnucleosomasunodeloscomplejosprotéicosmásestablesbajocondicionesfisiológicasnormales.Laformaenqueestácompactadalacromatina,leproveepropiedadesdinámicasquesonreguladasporvariosgruposprotéicos(Li,Carey,&Workman,2007).
Loscomplejosremodeladoresdelacromatinasoncomplejosprotéicoscapacesdemoverlosnucleosomasde su posición para dejar expuesta la región promotora y permitir la entrada de complejostranscripcionales para que se dé la expresión génica. Esta actividad está mediada por complejos conactividadATPasaenestecasoparticular,porelcomplejoSWI/SNFquefueelprimercomplejoidentificadoen levaduras. Su función es la de desestabilizar el nucleosoma al romper el contacto del DNA con eloctámero de histonas y así podermoverse libremente para asociarse a la región de interés. Todos loscomplejos remodeladores de nucleosomas pertenecen a la familia SNF2 de ATPasas y se dividen en 7subgruposdependiendodesusdominios
5.1.3.2Modificacióndelascolasdehistonas
Lamodificacióndelascolasdehistonasesunmecanismoimportanteenlamodificaciónpost-traduccionalde las histonas. Estas modificaciones incluyen metilación de los residuos arginina (R) y lisina (K),acetilaciónenlisinas,ubiquitinación,ADP-ribosilación,sumoilaciónde lisinasy fosforilacióndeserinasytreoninas. Muchas de estas marcas son señales que dirigen la remodelación hacia un estado concreto.Algunasmodificacionesestánasociadasaactividad transcripcional, como laacetilaciónde lahistona3ehistona4(H3yH4)o ladi-o tri-metilación(me)deH3K4.Estassonconocidascomomodificacionesdeeucromatina. Otras modificaciones están relacionadas a la inactivación de genes o regiones, como laH3K9meylaH3K27my,seconocencomomodificacionesheterocromáticas.Sinembargo,muchasdeellas

23
noinfluyenen la transcripción,perosien laestabilidaddelgenoma,oenreparación,comopuedeser lafosforilacióndelavarianteH2AXolaH3K79me(Lietal.,2007).
5.1.3.3RNAsnocodificantes
Los RNAs no codificantes conforman una extensa familia de moléculas que intervienen en distintosprocesos. Según su extensión se dividen en RNAs cortos no codificantes y RNAs largos no codificantes.Durantelaúltimadécadahansidomuyestudiados,enespeciallosmiRNAs,siendolosmásconocidoshoyen día y de los que se tiene mayor información. Los microRNAs son secuencias cortas de RNAs nocodificantes de aproximadamente 20-24 nucleótidos (nt) que están ampliamente involucrados en laregulación de la expresión de genes en organismos multicelulares, afectando así la estabilidad, latranscripción y la traducción de los mRNAs. Los miRNAs son transcritos por la RNA polimerasa IIgenerando un transcrito primario con su Cap y su cola poliadenilada (pri-miRNAs). Este transcritoprimarioescortadopor laribonucleasaDroshaIIIparageneraruntranscritode70nt(pre-miRNA),queluegoescortadoenelcitoplasmaporlaribonucleasaDicerparagenerarunmiRNAmaduroyunmiRNAantisentido(miRNA*).ElmiRNAmaduroseincorporaenelcomplejodesilenciamientoinducidoporRNA(RISC)yreconoceporemparejamientodebasesasusmensajerosdeRNAdiana,generandoinhibiciónenlatraducciónyladesestabilizacióndelmRNA(NBI,miRNA,2016).
Dentrodeltemadeestudiodeestatesis,cabedestacarunmiRNAenparticular,miRNA133a.Estudiosencélulas del miocardio (cardiomiocitos) han descrito la forma como dicho miRNA133a regulanegativamentelaactividaddeDNMT1yporlotanto,sudesregulaciónestaríaafectandoalametilacióndemantenimiento(NBI,miRNA,2016).
5.1.3.4ModificacióncovalentedelDNA
Gracias a los estudios SMRT (Singlemolecule real-time sequencing) en el cual se detectan las posiblesmodificaciones en las bases del DNA, se ha establecido que existen varias representaciones que sonilustradasenlasiguientefigura.
Figura2a.EstructuramoleculardelasmodificacionesdebasesdelDNA.
Entre las modificaciones del DNA, la metilación es la más conocida y estudiada. En los mamíferos, lametilación del DNA ocurre principalmente por lamodificación covalente en los dinucleótidos CpG y escatalizado por enzimas que transfieren el grupo metilo. Esta modificación no cambia la secuencia denucleótidosyactúacomounreguladorcríticodelgenoma(C.C.Chenetal.,2013;Maresca,2015;Zaidietal.,2010).
AunquelametilacióndelosresiduosdecitosinaenelDNAseconocedesdelosaños50,nofuehasta1975cuando se describió su función sobre el genoma, asociándola desde entonces de forma indisoluble alsilenciamiento génico. Se describió como una marca que podía ocurrir de novo y ser heredadamitóticamente en células somáticas a partir del reconocimiento enzimático de secuencias CpGpalindrómicas hemimetiladas, asímismopodía ser interpretada por proteínas de unión alDNApara su

24
silenciamiento.EvidenciasrecientessugierenqueestamarcadesempeñaotrasfuncionesenelgenomayaqueseencuentradistribuidaalolargodetodoelgenomaynoexclusivamenteenregionesCpGcercanasalsitiodeiniciodetranscripcióncomosecreía(Jones,2012).LasiguientefigurarepresentalatransferenciadelgrupometiloconS-adenosilmetionina(SAM)comodonadordelmismo.
Figura2b.Representaciónesquemáticadelatransferenciadelgrupometiloalcarbono5delacitosina(Maresca,2015)
Estamodificacióndelacitosinaesunreguladorepigenéticocríticoeneldesarrollodelosmamíferospuesjuega un papel importante en la diferenciación y desarrollo celular, así como en las enfermedades. Lacorrectametilacióndelgenomaesfundamentalenelmantenimientodelaidentidadcelular,lainactivacióndelcromosomaXylaimprontagenómica.Igualmenteprotegelaintegridadgenómicaporelsilenciamientodetransposonesyotroselementosmóviles(Denisetal.,2011;Laurentetal.,2010;Bhutani,Burns,&Blau,2011; Chen & Rigs, 2011). La metilación también juega un papel importante en el silenciamientotranscripcional de genes durante la formación de la heterocromatina, ya sea por la interferencia de launión de los factores de transcripción a su sitio diana o afectando las modificaciones en las colas dehistonas y la ocupación de nucleosomas dentro de las regiones promotores de genes, afectando así laestructuradelacromatina.AlgunasmetilacionessonreversiblesdebidoaquelascélulaspuedeneliminarfraccionesdemetilosdelasislasCpGsatravésdelareplicaciones(Zaidietal.,2010).
La metilación es importante para establecer las características fenotípicas de las células, tejidos yorganismos.Pocose sabedesuvariaciónnaturalen loshumanosy cómoesto influyeen lasdiferenciasfenotípicas entre individuos, así como en la susceptibilidad a ciertas enfermedades y sus patogenias,respuesta a medicamentos y agentes ambientales (Heyn et al., 2013; Horvath, 2013). En estudios decáncer, se ha demostrado que la generación de patrones aberrantes de hipermetilación conducen acambiosenlaestructuradelacromatinayalsilenciamientodegenessupresoresdetumores(Zaidietal.,2010;Laurentetal.,2010;Jones,2012).
Como modificación epigenética, la metilación puede ser heredada a través de múltiples divisionescelulares.Lospatronesdemetilacióngenómicossonestablecidosdurantedosperíodosdeldesarrollo,encélulas germinales y en la preimplantación embrionaria, generando células con un amplio potencial dedesarrollo (Branco, Ficz, & Reik, 2011; Denis et al., 2011; Volkmar, Dedeurwaerder, Cunha, Ndlovu,Defrance,Deplus,Calonne,Volkmar,Igoillo-Esteve,etal.,2012).
5.1.3.4.1ElpapeldelasCpGsylametilacióndelDNA
PormuchosañossehaestablecidoquelametilacióndelDNAenmamíferosocurresimétricamenteenlosresiduosde citosinaenambashebrasdedinucleótidosdeCpG,peroestudios recienteshandemostradoque esta marca no sólo se limita a las secuencias CpGs. En estudios in vitro, se ha observado que lametilacióntambiénocurrede formanosimétrica,comoporejemploennucleótidosCpXpG.(Denisetal.,2011).

25
La distribución de las CpGs en el genoma es heterogénea. Algunas áreas son ricas y otras pobres endinucleótidosCpG.Losnivelesypatronesdemetilaciónsontejidoespecíficoynotodaslascitosinasestánmetiladas.Estoúltimoesloquediferencia losestadosdeactividadgénicaentrecélulasdiferenciadas,yaque inducenprocesos fisiológicosclavesdeldesarrollo. (El-Ostaetal.,2008; Jones,2012).Enelgenomahumanohay56millonesdesitiosCpG,deloscualesel70-80%estánmetiladosycorrespondenal4-6%detodaslascitosinas.LosdinucleótidosdeCpGsestánconcentradosenlaspequeñasregionesllamadasislasCpGs(CGIs)queconformanel70%de lospromotoresdegeneshumanos(Laurentetal.,2010;Saxonov,Berg, & Brutlag, 2006). El 60% de los genes tienen CGIs en sus promotores y tienen sitios libres denucleosomas(NDRs)enelsitiodeiniciodelatranscripción(TSS).Enelrestodelgenomadelosmamíferoslas islasCpGsestánpobrementerepresentadas,este fenomenoestámuyposiblementeasociadoa laaltamutagénesisdela5-metilcitosinaencomparaciónconlacitosinanometilada(Jin,Kadam,&Pfeifer,2010).
EncélulassomáticaslagranmayoríadeCGIssemantienendesmetiladasensusTSS,sonmarcadasporlatrimetilaciónenlalisina4delahistonaH3(H3K4me3)ysecaracterizanporlaremodelaciónnucleosomalcon lahistonaH2A.Z, un antagónicode lasDNMTs, asociada a la transcripción.Generalmente, las zonascorriente abajo de los TSS son pobres en CGIs, aunque son ricas en dinucleótidos CpGs. Estos últimospuedenestaronometiladosysehavistosufunciónenelcontroldesplicing.Losnivelesdeexpresiónsoncontroladosporfactorestranscripcionalespertenecientesalasproteínaspolycomb,trayendonucleosomasal TSSmarcados con laH3K27me3. Sin embargo, se ha observado que lametilación de novo ocurre enausencia de H3K4me2 y H3K4me3 (Jones, 2012). En la siguente figura se puede observar la anatomíamoleculardelosCpGs.
Figura3.Anatomíamoleculardelacromatina,lossitiosCpGsyelrolenlaexpresióngénica(Jones,2012).

26
LosgenesconsitiosTSSpobresdeCGIsgeneralmenteestánnometiladosysonaltamenteexpresivos,talycomo ocurre en células primordiales. Hoy en día se sabe que la metilación no sólo se limita alsilenciamiento génico o a la expresión del gen. Diversos estudios han evidenciado que existe unacorrelación inversa entre la densidad de las islas CpGs y los estados demetilación en promotores. Losgenesactivosusualmenteestánhipometiladosensitiosde inicio transcripcional (TSS)yaltosnivelesdemetilaciónseencuentranenloscuerposgénicosparabloquearlainiciacióndetranscripcionesaberrantesdentrodegenes,previniendoasí laproduccióndemicroRNAs(mRNAs)yproteínastruncadas.LossitiosdecorteyempalmesonregionescaracterizadasporuncambioenlametilacióndelDNA,teniendomayormetilaciónlosexonesquelosintrones,sugiriendoasílaimplicaciónenlosprocesosdecorte,empalmeyprotección.(Chatterjee&Vinson,2013;Laurentetal.,2010;Heynetal.,2013).
La mayoría de los cuerpos génicos son pobres en CpGs aunque generalmente están extensamentemetilados.MetilacionesenCpGsdeexonessonlaprincipalcausademutacionestransicionalesC→T,queconllevan a enfermedades de las líneas celulares germinales y al cáncer. Sin embargo, lametilación encuerpos génicos no se asocia a represión ymás bien actúa como una protección de la transcripción deelementos móviles como LINEs, Alu, entre otros. Se ha visto que están más metilados los exones encomparaciónalosintrones,posiblementecomosemencionópreviamente,paralaregulacióncorrectaenelprocesodesplicing(Jones,2012).
Los promotores downstream son ricos en CpGs a diferencia de los promotores alternativos quegeneralmentesonpobreenCpGs.LametilaciónensitiosenhancerespobreenCpGsdadoquesonsitiosimportantes para la unión de complejos transcripcionales y es por esto que contienen zonas pobres enmetilación(LMRs).Enelcasodelos insulators,proteínasquesehandescritoporbloquearlainteracciónde los sitios enhacer y el promotor, se evita la transcripciónmediante la proteína CTCF, que se une enausencia demetilación y es por esto que estamarca es importante para la actividad de dicha proteína(Jones,2012).
5.1.3.4.2EnzimasencargadasdelametilacióndelDNA
A.DNAmetiltransferasas
En el genoma de los mamíferos, las DNA metiltransferasas (DNMTs) son pieza clave en la regulaciónepigenética. Las DNMTs catalizan la transferencia del grupo metilo de la S-adenosilmetionina (SAM)obtenidoenelciclodeuncarbonoalacitosinadelDNA.(Chen&Riggs,2011;Goll&Bestor,2005.Estasenzimas son las encargadas de establecer ymantener lametilación en la citosina de los CpGs del DNA.Existen tres enzimas funcionales como son las DNMT1, DNMT3a, y DNMT3b. Otra enzima como es laDNMT3Ltienefuncionesreguladoras,perocarecedesitiocatalítico.Porúltimo,existetambiénlaDNMT2,queenlaactualidadseconsideraqueejercesufuncióncasiexclusivamentesobreelRNA(Figura4).Todasson esenciales para establecer los patrones de metilación y para mantenerlos tras los procesos dereplicación(Denisetal.,2011).AcontinuaciónsemuestranlascaracterísticasgénicasdelasDNMTs(Tabla1).
DNMT1 DNMT3a DNMT3b
Posicióncromosómica 19p13.2 2p23.3 20q11.21
Transcritoprimario(pb) 5408 9501 4336
Transcritocodificante(pb) 4851 2739 2562
Proteína(aa) 1616 912 853
Fenotipo asociado a su Ataxiacereberal,sordera,narcolepsia
SíndromeTatton- Síndromedeinmunodeficiencia,

27
disfunción. autosómicadominante,
neuropatíatipoIEhereditaria
Brown inestabilidaddelaregióncentroméricayanomalíasfaciales
Tabla1.DescripcióngénicadelasDNMTs.(OMIM,CCDS,NCBI)
Pormuchos años se ha propuesto que elmodelo demetilación se basa en dos funciones,denovo y demantenimiento. Las enzimas DNMT3a y DNMT3b son las encargadas de establecer patrones nuevos demetilación. Están altamente expresadas en el desarrollo y la diferenciación celular y pobrementeexpresadasenlascélulasdiferenciadas.Apesardesubajaexpresiónencélulassomáticas,hoyendíasehadescritoquesonnecesariasparamantener lospatronesdemetilación juntoa laDNMT1. (Illingworth&Bird,2009;Rönnetal.,2013)
Figura4.RepresentaciónesquemáticadelasestructurasdelasDNMT’s(Denisetal.,2011).
El mantenimiento de los patrones de metilación se establece por el reconocimiento de la hebrahemimetilada durante la replicación del DNA, por medio de las DNMT1. Estudios genómicos en elmantenimiento de estos patrones han descrito la necesidad de más interacciones proteícas. Elmantenimientode lametilaciónnoes tansencillocomopareceynosóloestá involucrada laDNMT1,yaqueeldesarrolloyelcontroldelossistemasdereplicacióndebegarantizarquesemantengalafidelidaddedichospatrones.Estudiosdemuestranque lasDNMT3ayDNMT3bsonnecesariasparamantener lospatronesdemetilaciónjuntosalasDNMT1yqueelfielmantenimientodelospatronesnosólosemantieneporunafielcopiadurantelaembriogénesis,sinotambiénporlaayudaenlametilaciónporpartedeéstasDNMTs(El-Ostaetal.,2008;Rönnetal.,2013).Enlafigura5seobservaelmodeloclásicodelametilación.

28
Figura5.Modeloclásicodelametilación(Maresca,2015).
a DNMT1:DNA(citosina-5-)-metiltransferasa1
LaDNMT1es laprincipal enzimaparapreservarelmantenimientode lospatronesdemetilaciónde lascélulastraslareplicación.Existendoscaracterísticasimportantes,suexpresiónesubicuayaltadurantelaproliferaciónyvaríasegúnelestadodelciclocelulardemaneradependiente,siendomayorenlafaseSdesíntesis.Laotracaracterísticaessuescasezen lascélulasquenoestánendivisión.Encasospatológicoscomo el cáncer, se ha descrito actividad de metilación de novo por estas enzimas (Denis et al., 2011;Maresca, 2015) (Chen and Li 2006). Otros estudios han demostrado la capacidad de reparar lesionescausadasenelDNA(Mortusewicz,Schermelleh,Walter,Cardoso,&Leonhardt,2005).
En humanos, esta enzima se conforma de 1616 aminoácidos y es la más abundante en preservar ypropagarlaexistenciadeestospatrones.LaDNMT1muestrapreferenciaporregioneshemimetiladasyhamostradoserunaenzimaaltamenteprocesiva,capazdemetilarlargostramosdelacadenasindisociarsedeella(Hermanetal2004;(Denisetal.,2011;Reilingetal.,2010).AdemásinteractúaconelfactorPCNdelaDNApolimerasaqueasegurasulocalizaciónenlahorquilladereplicación(Jones,2012).
Los estudios knockdownde DNMT1muestran extensiva desmetilación del DNA y letalidad embrionariadurantelagastrulación.CélulasembrionariasconausenciadeDNMT1sonviablesdadolapocanecesidadde la enzima, pero mueren después de inducir la diferenciación. (Chen et al 2013; Maresca, 2015).Mutaciones que afectan el gen DNMT1, se han asociado a dos enfermedades neurodegenerativasautosómicas dominantes: neuropatía sensorial y autonómica hereditaria con demencia y pérdida deaudición tipo1E (HSN1E)y sordera-ataxiacerebelosaautosómicadominanteynarcolepsia (ADCA-DN).Unacaracterística importantede laDNMT1essudependenciaporeldominioSRAde laUHFR1parasureclutamientoenlossitiosCG(BashirAamiretal.,2012).
B.UHRF1(Ubiquitin-like,containingPHDandRINGfingerdomains)
ElgenUHRF1codificaparaunaproteínapertenecientealafamiliadeRING-fingertipoE3delaligasadeubiquitinaysecaracterizaporlaunióndeunasecuenciaespecíficaenelDNAparaelreclutamientodelaDNMT1.Dichauniónmedialaregulacióndelaestructuradelacromatinaylaexpresióndelgendiana.LaUHRF1ayudaalaDNMT1areconocerlahebrahemimetilada(Brancoetal.,2011;Z.Chen&Riggs,2011).

29
5.1.4Desmetilaciónactivaypasiva
Existendosmecanismosprincipalesdedesmetilación,lademetilaciónpasivayladesmetilaciónactiva.Ladesmetilaciónpasivaocurrepor lareducciónoausenciaen laactividadde lasDNMTs, enespecialde laDNMT1, en elmantenimiento de la 5mC tras la replicación celular, perdiéndose así lametilación en lahebrahemimetilada.Ladesmetilaciónactiva,porsuparte,ocurreenrespuestaacambiosdeseñalesenlacélula.Sebasaendosvíasprincipales,unavíaoxidativaquehastaelmomentosecreequeesgeneradaporlasglicosilasasten-eleventranslocasas(TET’s)queoxidanelgrupometilodela5-metilcitosina(5mC)a5-hidroximetilcitosina (5hmC) y otra vía de deaminación de citosina inducida por la activación de laapolipoproteínaB(AID/APOBEC)quepor losmecanismosdereparacióncelularsedesmetilande formaactivalas5-metilcitosinasgenerandoasíunacitosina(Bhutanietal.,2011).
Elpapeldeladesmetilaciónhasidoampliamentedocumentadoenpocesoscelularesimportantescomoeneldesarrollo temprano, lareprogramacióndurante lagametogénesisyen laclonación. Igualmentesehadescritosuimportanciaenlaformacióndelamemoriay laneurogénesis,enlasrespuestasinmunespormedio de las citoquinas y en la tumorogénesis. En diversas situaciones están involucradas estasmodificacionesqueafectanlaformaenlacualsevanaexpresaronolosgenes(Zhu,2009).
5.1.4.1Maquinariadedesmetilaciónactiva
Lamaquinaria de la desmetilación activa está representadapordioxigenasas y desaminasas que se venrepresentadas en la figura 6. La vía de la oxidación de la 5mC por las TETs, genera un compuestointermediario5hmCquepuedeseroxidadoporlamismaenzimayasíformarotrosintermediarioscomo5-formilcitosina (5fC) y 5-carboxicitosina (5caC). Aún no se ha demostrado si la 5caC puede serdescarboxiladaporlasDNAglicosilasasdetimina(TDG),perosehademostradoqueapartirdel5fC,lasTDGremuevenelgrupotiminaparaformarunacitosina.Lavíade ladeaminaciónesdescritacomootravíaactivaenlacualparticipancomplejosmoleculares,entreellas,laapolipoproteínaB(AID/APOBEC),laTDG, lasproteínasdeuniónagruposmetilos(MBD4)y lasglicosilasasdecadenasencillaqueremuevenuracilos (SMUG1), que pormecanismos de reparación por escisión de bases (BER) producen una baseapurínicaqueesreparadaporlabasecorrecta.Hoyendía,dichosmecanismossiguenenestudio(Brancoetal.,2011).
Diversosestudioshanpropuestoqueelprocesodedesmetilaciónesbidireccional.Estohasidoobservadoenestudiosdereprogramacióncelularyen laaccióndereguladoresasociadosalaplasticidadydestinocelular (Bhutani et al., 2011). Un estudio reciente, reveló una característica nueva de las DNMTs envertebrados, la capacidad de convertir el 5mC a 5hmC en presencia de iones de calcio y la ausencia deagentesreductores(Chenetal.,2013).

30
Figura6.DiagramadelasposiblesvíasdedesmetilacióndeDNA(Brancoetal.,2011).
A.Ten-eleventranslocasas(TETs)
Laexistenciadela5hmCfuereportadaenlos50porWyatt,aunquesufunciónesconocidahastahacemuypocotiempo.EldescubrimientodelasglicosilasasTET1,TET2yTET3fuedegraninterésporsucapacidadde catalizar la conversion del 5mC a 5hmC en diversos tipos de cáncer. Con el tiempo, se describió sufuncióndeactivarlarepresióndepromotoresengenesinactivos.Enestudiosenratonesseidentificósuactividadenelmantenimientodelapluripotenciayenladesmetilacióneneldesarrollotemprano(Figura7)yfueasícomoseconocióqueTET3estáinvolucradaenlapérdidarápidademetilaciónenelpronúcleomasculinodurantelafomacióndelcigotodespuésdeladivisióncelular(Bhutanietal,2011).
Figura7.Reprogramaciónencélulasgerminalesypreimplantaciónembrionariaenratón.Durantelosestadiosinicialesdeldesarrollo,lascélulasPGCsseencuentranaltamentemetiladasyentranen
desmetilacióndurantelapro-espermatogénesisylapro-ovogénesis,parafinalmenterestablecerse.Enelcasodelgenomapaterno(azul)despuésdelafertilizaciónocurreinmediatamenteunadesmetilación.Enelgenomamaterno(rojo),ladesmetilaciónsedaluegodepocasdivisionescelulares.Ambosgenomasson
remetiladosdurantelaimplantaciónembrionaria(Ding&Huang,2014).

31
Todas lasenzimasTETs tieneundominioCD(ricoencitosinasyregionesDSBH(Dominioconactividaddioxigenasa de metilcitosina de unión a metal)), que en presencia del ácido α-cetoglutárico y hierrodescarboxilanelO2paragenerar5hmCapartidel5mC.LosgenesTET1yTET3contienendominiosCXXCimportantesparalauniónalosdinucleótidosCpG,mientrasquelegenTET2carecedeestedominio(Heetal.,2011).AcontinuaciónsemuestranlascaracterísticasgénicasdelasTETs(Tabla2).
TET1 TET2 TET3
Posicióncromosómica 10q21 4q24 2p13.1
Transcritoprimario(pb) 9288 10166 11388
Transcritocodificante(pb) 6411 3498 5388
Proteína(aa) 2136 1165 1795
Fenotipo Leucemiaaguda Trombocitopeniaesencial,policitemiaverayleucemias.
Problemasendesarrollocelular
Tabla2.DescripcióngénicadelasTETs.(OMIM,CCDS,NCBI)
b. TET1
LaTET1estáaltamenteexpresadaencélulasembrionarias(ES)yestápobrementereguladaduranteladiferenciación.SudominioCXXCescaracterísticoyhacepartedelgrupodeproteínasqueloutilizanpara launiónalDNA,enestecasoseuneacitosinasmetiladasehidroximetiladas. (Xuetal.,2011).Estudios basados en inmunoprecipitación de anticuerpos 5hmC en DNA hidroximetilado (hMeDIP),revelaronunaltocontenidode5hmCencuerposgénicos,especialmenteenintronesyexones,ybajocontenidoenpromotores.Tambiénseconfirmólaaltadensidadde5hmCensitiosdeterminacióndelatranscripción(TTS)ypresenciadeTET1comoreguladordelatranscripción(Xuetal.,2011)
B. AID/APOBEC
Ladesmetilaciónmediadaporglicosilasashasidomuyestudiadaenplantas.Enhumanossecreequeporlomenosestáinvolucradaenunpasodeladesmetilación.Ladeaminasadecitidina(AID)juntoacomplejos protéicos contribuye a la remoción extensa demetilación del DNA en células germinalesprimordiales de humanos y en el núcleo paterno del cigoto (Sanz, Kota, & Feil, 2010). Aunque aúnfaltanestudiosparadeterminarsuverdaderafunciónenladesmetilación.
ElcomplejoAID/APOBEC(apoliproteínaB)comparte lamismacaracterísticadecontenerunmotivode zinc en su sitio catalítico. Existen diferentes variantes de esta enzima y se ha visto en diversostejidos que la presencia de ella conlleva a procesos de desmetilación por la vía de reparación porescisióndebasesBER(Ramiro&Barreto,2015).
C. TDG
LaTDGesunaglicosilasadetiminaquesecaracterizaporremoverlosmotivosdetiminademismatchde G/T, C/T y T/T por la hidrólisis de la unión del carbón-nitrógeno entre el azúcar-fosfato delesqueleto del DNA. También es capaz de remover uracilos y 5-bromouracilos de guaninas malpareadas.Estaenzimajuegaunpapelfundamentalenladefensacelularcontramutacionescausadasporladeaminaciónespontáneadela5-metilcitosinaylacitosina(NCBI,2016).

32
D. MBD4
Laproteínadeuniónagrupometilo(MBD)secaracterizaportenerundominiodeuniónametilosenelN-terminalyundominioconactividadglicosilasaelC-terminal,importanteenlareparaciónDNA(NCBI,2016).
5.1.5Ciclodeuncarbono,claveenlametilación
El metabolismo bioquímico de un carbono es clave para el proceso de metilación del DNA. La S-adenosilmetionina (SAM)y S-adenosilhomocisteína (SAH) son substratos yproductos esencialespara lametilación.SutasasehautilizadocomoindicadordelacapacidaddemetilacióncelularestableciendoqueunadisminucióndeSAMdisminuyeasuvezlametilación(Caudilletal.,2001).
El ciclo de un carbono recibe este nombre porque transporta un carbono durante su reacción y estáconformadoportresciclosbioquímicosqueparticipanconjuntamente.Latransmetilación,conocidocomoelciclodeSAM,es importanteporaportarelgrupometiloa laDNMT1.Elsegundoeselciclodel folato,fundamental para la síntesis de poliaminas donde el folato es el compuesto donador del grupometilo.Finalmente,elciclodelatransulfuración,queesesencialeimportanteparalageneracióndeprecursoresdeglutatiónytaurina.Evidenciasgenéticasrecientessugierenquelahiperactivacióndeestavíaconducealaoncogénesisyestableceestadosepigenéticosalterados(Locasale,2013).
Lametionina-sintasa(MS) juntoalcofactorvitamínicoB12,son importantesen lareacciónparagenerarmetionina.Apartirdeestecompuestojuntoeladenosiltrifosfato(ATP), laMATcatalizalareacciónparagenerar SAM, compuesto principal que dona su grupo metilo a las DNMT’s para producir S-adenosilhomocisteínayentraralciclodetransulfuraciónoremetilarse.Enlasiguienteimagenseobservanlostresciclosinvolucradoseneltransportedelgrupometilo.
Figura8.Tresrutasmetabólicasquecomponenelciclodeuncarbono(Locasale,2013).

33
5.2DIABETESMELLITUSCOMOMODELOBIOLÓGICO
5.2.1Conceptoactualdeladiabetesmellitus
Ladiabetessecaracterizaporunadeficienciadelpáncreasenproducirsuficienteinsulina,ladisminuciónde la respuestade los tejidosparautilizarla,oambas.La resistenciaa la insulinaesel resultadodeunapobrerespuestatisularalainsulinaaúnenpresenciadeconcentracionesnormalesdelahormona.Enladiabetessealteraelmetabolismodecarbohidratos,lípidosyproteínas(ADA&IDF,2016;Anexos1.)
La diabetes es una de las enfermedades crónicas más comunes en el mundo entero (Matough, Budin,Hamid, Alwahaibi, & Mohamed, 2012). La diabetes causa un desorden metabólico caracterizado porhiperglucemiacrónicagenerandoasídiversascomplicaciones.Suincidenciasehavistoenaumentoenlosúltimos 10 años y se cree que se duplicará para el 2030 (Forbes & Cooper, 2013). Se considera unproblema de salud pública grave y creciente a nivel mundial ya que provoca una reducción en laexpectativadevidayunaumentoenlamorbilidaddebidoalascomplicacionesespecíficasquegeneralaenfermedad(Ceriello,2008;Intineelal.,2012).
LaOrganizaciónMundialdelaSalud(OMS)estimaqueenelmundohaymásde347millonesdepersonascondiabetes.Másdel80%delasmuertesseregistranenpaísesdebajosymedianosrecursoseconómicos.Secalculaqueparael2030, ladiabetesserálaséptimacausademortalidadconmásde400millonesdeafectados(OnlineOMS,2015;Villeneuveetal.2011).SegúnlaFederaciónInternacionaldeDiabetes,másdelamitaddelosposiblescasosaúnnohansidoidentificados(OnlineIFD,2015).EnAméricaLatinalaDMTipo2eseltipomáscomúnylaincidenciavaenaumentocontasasdosvecesmásaltascomparadaconotrossitiosdistintos(OnlineIDF,2014).
EnColombia,suincidenciaanualvaenaumentoysuprevalenciaoscilaentreel4-8%enzonasurbanasyel2%enzonasrurales.Múltiplesfactoresgenéticosyambientalescomoelcambioenlosestilosdevida,elenvejecimiento, el mestizaje y la urbanización han impulsado el aumento de sobrepeso y el síndromemetabólico contribuyendo así a su patogenicidad y persistencia. En el 2007 el DepartamentoAdministrativoNacionaldeEstadísticas(DANE,2016),informóqueladiabetesseencuentradentrodelasprimerascincocausasdemuerteanualesyescuatrovecesmásfrecuenteenlapoblaciónurbanaqueenlarural.LaprevalenciadeDMTipo1esdel0.07%ysuincidenciaesde3a4por100,000niñosmenoresde15años(Aschner,2010).
5.2.2Causasdeladiabetes
La diabetes tiene un origen multifactorial en el que participan factores genéticos y factoresmedioambientales. El estilo de vida occidental, caracterizado por un desbalance de micronutrientes ydietasconaltovalorenergéticopromueveneldesarrollodeobesidadydiabetestipo2.Sesumaaestolainactividad física y el aumento considerable del peso. Todos estos factores son consecuencia deacontecimientos como la migración de poblaciones rurales a zonas urbanas, cambios de alimentosnaturales a alimentos procesados ricos en energía, exposición a agentes contaminantes orgánicos ysintéticos, el desequilibrio temprano en la microbiota, el uso indebido de antibióticos y el desarrollotecnológicoquereduceelgastoenergéticopararealizarunesfuerzo.Todosestosfactorescontribuyenalsedentarismo y al sobrepeso, promoviendo el desarrollo de diabetes en personas susceptibles (Nolan,Damm,&Prentki,2011),comoseresumeenlasiguienteimagen.

34
Figura9.Modeloqueproponeelroldelaepigenéticaenlosmecanismospatogénicosdeladiabetes(Ling&Groop,2009)
5.2.3Mayorescomplicacionesenladiabetes
Las mayores complicaciones de la diabetes se asocian a daños microvascular y macrovasculares,generados por una elevación crónica de los niveles de azúcar en sangre (Figura 10). Entre lascomplicacionesmicrovascularessedescribenlaretinopatíadiabéticaconpérdidapotencialdelavisión,lanefropatíadiabéticaqueproducedañorenaly lasneuropatíadiabéticaqueocasionadañodirectoen losnervios y alteración de los vasos que nutren a los nervios; vasa nervorum. Las complicacionesmacrovasculares por su parte, afectan principalmente la arteria coronaria e incluyen enfermedadesarterioescleróticas cardiovasculares prematuras, enfermedades cerebrovasculares y enfermedad arterialperiféricaconaltoriesgodeulceracionesyamputacionesdelasextremidades(Brownlee,2005;Forbes&Cooper, 2013; Giacco & Brownlee, 2010; Poretsky, 2010). En la siguiente figura se visualizan lascomplicacionesdeladiabetes.

35
Figura10.Mayorescomplicacionesdeladiabetes(OnlineIDF,2016).Complicacionesmicrovasculares:retinopatía diabética, enfermedad coronaria, nefropatía diabética y enfermedad periférica vascular.Complicacionesmacrovasculares:neuropatíadiabética,piediabéticoyenfermedadcerebrovascular.
Actualmente, la DM es la primera causa de insuficiencia renal crónica terminal (IRCT) en elmundo. Seestima que el 50% de los casos son causa de nefropatías diabéticas. Es la primera causa de cegueraadquirida enelmundoy laprimera causade amputacionesnodebidas a traumas. LaDMesunode loscuatro factores de riesgo clásico para aterosclerosis y enfermedades coronarias. Los otros trescorresponden al tabaquismo, la hipertensión y la dislipidemia. Es importante anotar que el 80%de losdiabéticos fallecen a causa de las complicaciones de la aterosclerosis por infartos almiocardio, eventoscerebrovasculares o enfermedad arterial periférica (Poretsky, 2010). Aunque la diabetes es unaenfermedad común en la mayoría de países, su patogenia aún no es clara debido a las múltiplesinteraccionesdefactoresgenéticosyambientales(OnlineIDF,2016).
5.2.4Clasificacióndeladiabetes
Esta enfermedad compleja y heterogénea contiene diversosmecanismos etiológicos. Su clasificación esarbitraria y está descrita en 4 categorías principalmente: DM tipo 1 (T1DM), DM tipo 2 (T2DM), DMgestacional (GDM) yDM causada por condiciones genéticas específicas (Tabla 3) (Poretsky, 2010;OMS,2016). Este último grupo es poco común y se caracteriza por desórdenes endocrinos, defectos en elpáncreas exocrino, endocrinopatías, defectos en la función de células β, defectos genéticos en elprocesamientodelainsulinaosuacción,porinfeccionesodrogas.Lasmásconocidasenestacategoríasonla DM tipoMODY, (MaturityOnsetDiabetesof theYoung) y la DMAutoinmune Latente del Adulto (tipoLADA)queapareceenlajuventudyenlaedadadulta(Poretsky2010).

36
CATEGORIASPRINCIPALES
I. DIABETESTIPO1 • Autoinmune• Idiopática
II. DIABETESTIPO2 • Insulinoresistenciaydefectoselasecrecióndeinsulina
• Defectosenlasecrecióndeinsulinaconosinresistenciaalainsulina
III. OTROSTIPOSESPECÍFICOS
• Defectosgenéticosdelafuncióndelascélulasbeta• Defectosgenéticosenlaaccióndelainsulina• Enfermedadesdelpáncreasexocrino• Endocrinopatías• Inducidasporfármacosoquímicos• Asociadaainfecciones• FormaspococomunesdeDiabetesmediadapor
inmunidad• Otrossíndromesgenéticosavecesasociadoscon
DiabetesIV. DIABETES
GESTACIONAL
Tabla3.ClasificacióndelascategoríasprincipalesdelaDiabetesMellitus(OMS,2016).
C. DiabetesMellitustipo1
Se conoce que la T1DM se genera por una respuesta directa o indirecta del sistema inmune en ladestruccióndelascélulasβdelpáncreas,productorasdeinsulina.Pormuchosañosseconsideróquelosafectados eranniños y adolescentes, pero esto ha cambiado en la últimadécada.Hoy endía la edaddeiniciodesíntomasyanoesunfactorrestrictivo.LaT1DMsecaracterizaydiagnosticaporuntríoclásicodepolidipsia, polifagia y poliuria junto a estados hiperglucémicos. Aunque aparece en edades tempranastambién puede darse en adultos. En estos casos, se debe hacer reemplazo de terapia hormonal deinmediato por la cual se requiere de mayor intervención en los tratamientos (Atkinson, Eisenbarth, &Michels,2014).
Lamayoríadeestudiosenladiabetestipo1hanreportadoquelaenfermedadsedebeaunadestrucciónautoinmunedelascélulasβproductorasdeinsulinaenelpáncreasoalaincapacidadderegeneracióndecélulasβ(Atkinsonetal.,2014).Porestarazón,lospacienteT1DMsoninsulinodependientes.
D. DiabetesMellitustipo2
LaT2DMesuntrastornometabólicode laenergíade lahomeostasis.Secaracterizaporhiperglucemiayalteraciónenelmetabolismodeloslípidoscausadosporlascélulasβdelosislotes,incapacesdesecretaradecuadainsulinaenrespuestaadiversosgradosdesobrealimentación,inactividadcomoconsecuenciadesobrepeso,obesidadyresistenciaalainsulina.Hoyendíaesunproblemagraveanivelmundialdebidoalrápidoaumentodesuprevalencia(Nolanetal.,2011).Eldevastadordañoalosórganosafectaelsistemadesaludgenerandocostosalarmantes.
SeconocencuatrocondicionesfisiopatológicasquedesencadenanlaT2DM;laobesidad,laresistenciaalainsulinaenmúsculoytejidoadiposo,ladisfunciónenlasecrecióndeinsulinaporcélulasbetayelaumentoen la producción de insulina hepática Su patofisiología se debe a un exceso crónico de la energía pornutrientes,quesedesarrollaenpersonasgenéticayepigenéticamentesusceptibles,afectandoasílasvíasneurohormonales. Individuos resistentes a la insulina son capacesde resistir a la sobrecarga crónicadenutrientesdebidoaunisloterobustodecélulasβ,manteniendolasecreciónadecuadadeinsulinaparala

37
compensaciónsegúnlorequeridoylaexpansiónsaludabledeltejidoadiposossubcutáneo(SAT).Enpocaspalabras el cuerpo es capaz de dispersar el exceso de calorías a la grasa subcutánea en lugar dealmacenarla.Deestamanerasemantienenlosnivelesdenutrientesenlasangreademásdeotrostejidoscomoelhígado,musculoesquelético,corazónyovarios(Nolanetal.,2011).
Encambio, los individuossusceptiblesdesarrollandiabetespor lafalladeestosmecanismosadaptativosen respuesta al exceso de energía (Figura 11). Las células β fracasan por el exceso de trabajo en laproduccióndeinsulinaysonincapacesdecompensardichoexcesoenergético.Segeneraunestréscelularcausado por un desbordamiento de nutrientes a tejido no adiposos, esto a su vez aumenta la secreciónendógenadeglucagónygeneraincapacidaddealmacenarenergíaeneltejidosubcutáneo.Resultandoasíen la inflamación de los tejidos, incremento de glucógeno hepático y resistencia a la insulina. Ladisminuciónenlafuncióndelascélulasβpuedeterminarensudestrucciónylanecesidaddeadministrarinsulina(Nolanetal.,2011).
Unestudiodedetoxificacióndenutrientes reveló elmecanismocomo las célulasβpancreáticaspuedendesregularse como medida preventiva de intoxicación. Describen como éstas censan los niveles denutrientes en sangre y modulado por señales neurohormonales, inducen la secreción de insulina deacuerdo a las necesidades del organismo. Esto genera una activación metabólica en los individuossusceptiblesadiabetesenquieneslascélulasβreducensupotencialcomomedidapreventivaparaevitarlaintoxicaciónpornutrientes,generandocomoresultadounafallaenlosislotes(Nolanetal.,2011).Enlafigura12seobserva ladiferenciade susceptibilidadesgenotípicasentre laspersonasquedesarrollan laenfermedadfrentealasqueno.
Figura11.VíasdelapatogeniadelaT2DMylascomplicacionesrelacionadas.(Nolanetal.,2011).

38
Figura12.GenesasociadosavariantesdeT2DM(Prasad&Groop,2015).
5.2.5CriteriosdeDiagnóstico
El diagnóstico de la diabetes se puede realizar con base en la medición de glucemia en ayunas comomuestra la tabla 4. La medición de la glucemia característica para diagnosticar la diabetes es que laglucemiaestéporencimade≥200mg/dlacualquierhoradeldíay≥126mg/dlenayunas.Existenotrasclasificacionessegúnlaintoleranciaalaglucosasinembargo,éstaeslamásutilizada.
Medicióndelaglucemiacausalacualquierhoradeldía ≥200mg/dl(11.1mmol/l)
Glucemiaenbasalenayunode8-14horas ≥126mg/dl(7.0mmol/l)
PruebadetoleranciaalaGlucosaenayunasya2horasdePoscarga
≥200mg/dl(11.1mmol/l)
75gdeglucosaoral
Tabla4.CriteriosparaeldiagnósticodelaDiabetesMellitus(Poretsky2010).
Tabla5.CriteriodediagnósticoparalosdiferentestiposdeDiabetesMellitus(Nolanetal.,2011).

39
5.3OXIDACIÓNCELULARRELACIONADAALADIABETES
5.3.1Roldelestrésoxidativo
Elroldelestrésoxidativoenlascomplicacionesdelasenfermedadesmetabólicashasidodegraninterésanivelcientíficoypúblico.Diversosestudiosclínicosyexperimentaleshanreportadoqueelestrésoxidativojuega un papel importante en la patogénesis y el desarrollo de las complicaciones de ambos tipos dediabetes. Sin embargo, el mecanismo por el cual el estrés oxidativo puede contribuir y acelerar eldesarrollo de las complicaciones de la diabetes aúnno es claro. Se sabe que la hiperglucemia induce laformación de radicales libres, afectando así el sistema de defensa antioxidante endógeno y las víasenzimáticasynoenzimáticasquecontrarrestanlasespeciesreactivasdeoxígeno(Matoughetal.,2013).
5.3.2Especiesreactivasdeoxígeno
Existen dos principales familias de especies reactivas; las especies reactivas de oxígeno (ROS) que seformanapartirdelareduccióndeloxígenomolecular(O2)ylasespeciesreactivasdenitrógeno(RNS)quesonproducidasporlaenzimaoxidonítricosintasa(eNOS)(Matoughetal.,2013).
LasROSsonespeciesaltamentereactivas(Tabla6),detiempodevidamediamuycortoyseproducenenelmetabolismonormaldeloxígeno,entodoslossistemasbiológicosaerobios.Estasreaccionancontodaslasmoléculasasualrededor,especialmenteconlasqueseencuentrancercadesusitiodeformación.Sureactividadesdadaalapareamientodesuúltimacapadevalencia,pasandodeunestadodeinestabilidadauno de equilibrio. Sabemos que el O2 que utilizamos para respirar proviene de manera exógena comoproducto de la oxidación del agua que se realiza en los fotosistemas II de las plantas. Pormedio de larespiracióny laalimentación, reducimoselO2enaguapara formarenergíaenATP(LunaLópez,2010).Contradictoriamentede la importanciafundamentaldeloxígenomolecularparasostener lavida,sesabeque el oxígeno es también tóxico a través de la formación de las especies reactivas de oxígeno (ROS)(Matoughetal.,2013).
Tabla6.Especiesreactivas(Matoughetal.,2013).
Como se ha visto en diversos estudios, la formación de ROS y RNS causa daño en macromoléculasbiológicas como las del plasma y células endoteliales. Igualemente son importantes en la señalización y

40
defensadelorganismo,enlafagocitosisyenlafuncióndelosneutrófilos,macrófagosycélulasdelsistemainmune. A su vez participan comomediadores de cascadas de señalización intracelular (Matough et al.,2013).
5.3.3CadenatransportadoradeelectronesylaformacióndeROS
La cadena de transporte de electrones es la principal generadora de ROS intracelular. Se compone decuatrocomplejosprotéicosI,II,III,yIV;NADHdeshidrogenasa,succinatodeshidrogenasa,citocromobc1,ycitocromocoxidasarespectivamente,comosepuedeapreciarenlafigura12.
Figura 13. Cadena transportadora de electrones. Se encuentra en la membrana interna de lamitocondria y su función es oxidar NADH y FADH2 y bombear H+ al compartimiento externo de lamitocondria.Lafosforilaciónoxidativaproduce32ATP’s(2013,García.Disponibleonline)
Esta cadena está organizada para regular la formación de energía en forma de trifosfato de adenosina(ATP)yparabombearprotonesatravésde loscomplejosI, IIIy IV,creandoasíunvoltajemitocondrial.Estevoltajepermite laentradadeproteínasdesacopladoras(UCP)a lamitocondriaparagenerarcalorymantenerconstantelageneracióndeATP(Brownlee,2005).
En el metabolismo normal, la glucosa entra al ciclo de Krebs para ser metabolizada y generar undinucleótidodenicotinamida (NADH), compuestoquedona loselectronesal complejo Ide la cadena.Elotrocompuestodonadorde la cadenaeseldinucleotidode flavinayadenina (FADH2), coenzimaque leaporta el electrón a la succianato deshidrogenasa, complejo II. Ambos electrones son transferidos a lacoenzimaQyesta los transfiereal complejo II, el citocromocycomplejo IVque finalmenteseuneaunoxígenomolecularyesreducidoaagua(Brownlee,2005).
5.3.4Aumentodeoxidaciónencasodehiperglucemia
Lageneracióndesuperóxidopormitocondriasdisfuncionalessehapostuladocomoelprocesoprincipaldel desarrollo de las complicaciones de la diabetes. Más del 90% del oxígeno en los humanos esmetabolizado durante la fosforilación oxidativa, en la cual la glucosa y otros metabolitos donan suselectrones para reducir el oxígeno molecular para la generación de ATP. A pesar de ser un procesoaltamenteregulado,entreel1al3%deesteoxígenoesparcialmentereducidoaO2-enlugardeserpartedegeneracióndeaguayútilparalasenzimasantioxidantes(Forbes&Cooper,2013;Browlee,2005).

41
Enladiabetes,lahiperglucemiainducelasuperproduccióndesuperóxidosmitocondrialescomoresultadodelexcesodeglucosaquedebesermetabolizada.Cuandolacadenatransportadoraalcanzasuumbraldetransportedeelectrones,elcomplejoIIIsebloquea,causandoqueloselectronesretrocedanalacoenzimaQ y esta termine donando el electrón a un oxígeno molecular, generando superóxidos. La isoformamitocondrial de la superóxidodismutasa convierte el superóxido a peróxidodehidrogenoque luego esconvertido a otras moléculas reactivas por diversas enzimas (Brownlee, 2005). Estudios in vitro hansugeridohipótesisdecómolasúperproduccióndeO2-yelcolapsodelpotencialdemembranageneranlafugadeelectronesen lugardegenerarATP(Copper&Forbes,2013).Lasiguiente imagenrepresenta lafosforilaciónoxidativa.
Figura14.Produccióndesuperóxidopor lacadenatransportadoradeelectronesmitocondrial.Elincrementodelosdonadoresdeelectrones(NADHyFADH2)porlosestadosdehiperglucemiaderivadosdelciclodeKrebsgeneranunpotencialdemembranamitocondrial(ΔμH+)mediantebombeodeprotonesporlamembranainternamitocondrial.InhibenasíeltransportedeelectronesalcomplejoIII,aumentadola vidamedia del producto intermediario coenzimaQ, reduciendoO2 a superóxido (Giacco&Brownlee,2013).
5.3.5Complicacionesdeladiabetesporbajaregulaciónoxidoreducciónylasupervivenciacelular
El balance entre la tasa de generación de radicales libres y su eliminación esmuy importante pues suacumulación es perjudicial. La etiología del estrés oxidativo en la diabetes surge de una variedad demecanismos tales como la excesiva producción de radicales libres por la auto-oxidación de la glucosa,glicacióndeproteínasylaglicacióndeenzimasantioxidantesquelimitansucapacidadparadesintoxicarlosradicaleslibres(Matoughetal.,2013).
El sistema oxido-reducción (REDOX) es muy importante para mantener la homeostasis celular. Bajocondiciones fisiológicas normales, las células mantienen la homeostasis generando un balance entre lageneración y eliminación de las especies reactivas de oxígeno y nitrógeno. Los niveles moderados deROS/RNS funcionancomoseñalesquepromueven laproliferacióny supervivenciacelular,mientrasqueun alto aumento puede inducir la muerte celular. En condiciones fisiológicas, el equilibrio entre lageneración y eliminación de ROS/RNSmantiene la adecuada función de la señalización de proteínas yasegura una respuesta correcta de los estímulos endógenos y exógenos. Cuando se perturba el sistemaREDOX, se genera estrés oxidativo conduciendo a aberraciones que generan la muerte celular ycontribuyenaenfermedadesdegenerativasycáncer(Trachootham,Lu,Ogasawara,Nilsa,&Huang,2008).

42
UnodelosfactoresclavesresponsablesenlacomplicacióncrónicadelaDiabetesMellituseslaproducciónexcesivaderadicaleslibresenlasmitocondriasdecélulasendoteliales,generandoasíunincrementoenelestrés oxidativo. Entre más azúcar hay en los espacios extracelulares hay más azúcar en las célulasendotelialesparametabolizaryportanto,másgeneraciónderadicales libres.Estacondicióngeneraunabajaactividaddelsistemaendógenoparaprotegerlasestructurascelularesdelosdañosocasionadosporlosperóxidosincrementadosdebidoalestrésoxidativo(Drzewoski,Kasznicki,&Trojanowski,2009).
Ya sea por un efecto dependiente o independiente de los estados de hiperglucemia, diversos estudiosapoyanque la superproduccióndeespecies reactivasdeoxígenoes laprimeraetapaenuna cascadadeeventosqueconducealascomplicacionescrónicas(Ceriello,2008a).
5.3.6VíasbioquímicasafectadasporROS
5.3.6.1PiezasdelrompecabezasdescritasporBrownlee
Los estudios de la Diabetes Control and Complications Trial (DCCT) y The United Kingdom ProspectiveDiabetes Study (UKPDS), establecieron la relación del daño tisular inducido por la hiperglucemia enpacientes diabéticos. En el siguiente diagrama se describe la relación entre los cambios agudos en elmetabolismo celular y la acumulación de cambios permanentes en lasmacromoléculas, causado por lahiperglucemiayeldañotisulargenerado.Esteprocesotambiénesafectadoporfactoresdesusceptibilidadgenética del individuo y factores independientes aceleradores como la hipertensión e hiperlipidemias(Brownlee,2005).
Figura15.Modelogeneralcomolahiperglucemiainducedañotisular(Giacco&Brownlee,2010).
Todaslasformasdeladiabetessecaracterizanporlahiperglucemia,lafaltarelativaoabsolutadelaacciónde la insulina, losmecanismosde resistenciaa la insulinayeldesarrollode las complicacionesmacroymicrovasculares. El estrés oxidativo es la causa esencial del desarrollo de las complicaciones diabéticascomo consecuencia de la sobreproducciónmitocondrial de superóxidos en las células endoteliales. Estegrupodecélulasafectadasincluyecélulasendotelialesdeloscapilares,célulasmesangialesdelglomérulorenal, las neuronas y las células de Shwann del nervio periférico (Brownlee, 2005; Giacco & Brownlee,2014).
ElcientíficoMichaelBrownleeunificócincoteoríasbioquímicasconocidasdesdelosaños60hastalosaños90sobreladiabetesylosprocesosdeoxidacióncelular.Distintosestudioscientíficosdemostraronlasvías

43
enquelahiperglucemiaintracelularaumentaexcesivamentelaproduccióndeROS,aumentadoasíelflujode lavíade lospolioles, la formacióndeproductosde laglicaciónavanzada(AGE’s),el incrementode laexpresión de receptores AGE y sus ligando, la activación de la proteinasa C (PKC) y el aumento de lashexosaminas.Comoconsecuenciadeesto,segeneranresiduosqueafectanpatológicamente laexpresióndegenesenladiabetes(Ceriello,2008a;Villeneuveetal.,2011).
5.3.6.2Hiperglucemiaydañomicrovascular
A.Incrementodelavíadelospolioles
En condiciones normales la enzima aldosa reductasa se encarga de la reducción de aldehídos tóxicos aalcoholes inactivos. En altos niveles de glucosa, esta enzima reduce la glucosa a sorbitol para luego seroxidada a fructosa (Figura 15). En este proceso también se consume el cofactor nicotinamida adeninadinucleótidofosfato(NADPH),importanteparalareduccióndelglutatiónporlaglutatiónreductasa.Estogenera un incremento en la susceptibilidad al estrés intracelular, lo que afecta células vasculares, denervios,retinayglomérulo(Giacco&Brownlee,2010;Brownlee,2005).
Figura16.Lahiperglucemiaincrementaelflujodelospolioles(Brownlee,2005).
B.PrecursoresdeAGE
Losprecursoresdelasglicacionesavanzadasseformanporlasreaccionesnoenzimáticasdelaglucosaconotros compuestos glicados derivados de la glucosa o de la oxidación de ácidos grasos en las célulasendotelialesarteriales.Entreellosestánlos3-deoxiglucosona,metilglioxalyelglioxal(Giacco&Brownlee,2010).
La formación de AGEs causa daño de las células por tres mecanismos principales (Figura 16). 1) Lamodificación intracelular de proteínas involucradas en la regulación de la transcripción, 2) la difusiónfueradelacélulaylainteracciónporproteínasreceptorasdelamatrizextracelular(integrinas)afectadolas vías de señalización celular y por último 3) la difusión al sistema circulatorio y modificación deproteínasen la sangree interacciónconmacrófagos, célulasendotelialesvascularesy célulasvascularesdelmúsculoliso(Giacco&Brownlee,2010;Brownlee,2005).

44
Figura17.ConsecuenciaspatológicasporelincrementodeAGEs(Brownlee,2005).
C.IncrementodelaexpresióndereceptoresAGEsysusligando
LosprecursoresdeAGEsfueradelacélulatambiénseunenalosreceptoresdeAGEs(RAGE),induciendolasuperproduccióndeROS,activandoel factorde transcripciónpleiotrópico (NF-κB)causandonumerososcambios patológicos en la expresión de genes asociados a la inflamación. Igualmente se incrementa laproducción de metilglioxal en células endoteliales interactuando así con otros factores epigenéticos(Giacco&Brownlee,2010).
D.ActivacióndePKC
Lahiperglucemiadentrodelacélulaincrementalasíntesismoleculardeldiacilglicerol(DAG).EsteesuncofactoresencialparaactivarlasdiferentesisoformasdeproteincinasaC(PKC).AlactivarseelPKCporlosaltosnivelesdeglucosa,éstealteramuchasvíasmetabólicas:disminuyelaexpresióndeprocesos“buenos”como del vasodilatador eNOS y aumenta lo “malo” como procesos inflamatorios, vasoconstrictores,oclusióncapilar,angiogénesisyanormalidadesenelflujosanguíneo(Figura17)(Brownlee,2005).
Figura18.ConsecuenciadelahiperglucemiaporlaactivacióndePKC(Brownlee,2005).

45
E.Incrementodehexosaminas
Elexcesodeglucosaparasermetabolizadaenlavíaglucolítica,incrementaelflujodelashexosaminas.Alformarse la fructosa-6-fosfato, ésta es convertida a glucosamine-6-fosfato por la enzima glucosamine--fructose-6-phosphate aminotransferasa (GFAT) y finalmente es transformada en UDP-acetilglucosamine(UDPGlcNAc).Éstaúltimaescapazdeentraralnúcleoeinteractuarconlosgruposdeserinaytreoninadelos factores transcripcionales, alterando la expresión génica. Lo anterior induce anormalidades en elglomérulo, disfunción de los cardiomiocitos y modificación endotelial (Brownlee, 2005). La figura 18representalavíaafectada.
Figura19.Incrementodelflujodelashexosaminasporlahiperglucemia(Brownlee,2005).
5.3.6.3Lahiperglucemiainducelaactivacióndelascincovías, inhibiendoaGAPDHyactivandoaADP-ribosa.
Durante la diabetes en humanos o animales, la hiperglucemia intracelular disminuye la actividad de laenzima glicolítica gliceraldehído-3-fostato deshidrogenasa (GAPDH). En presencia de las enzimastermogeninas UCP-1 óMnSOD, la inhibición de la actividad de GAPDH por hiperglucemia no genera lasobreproducción de superóxido mitocondrial. Como vimos en los mecanismos anteriores, todos losmetabolitos por encima de GAPDH están aumentados. El gliceraldehído-3-fosfato activa dos víasprincipales,lavíadelaPKCylavíadeformacióndeAGEs,generandoelprecursormetilglioxal,compuestoclaveenelincrementodelaexpresióndeRAGEqueactivaligandosdeS100calgranulinsyHMGB1(Giacco&Brownlee,2010;Brownlee,2005).
En estudios in vitro, se ha demostrado como el superóxido por si solo inactiva el GAPDH (Figura 19).Estudios en tejido endotelial demostraron que el superóxido inducido por hiperglucemia inhibe laactividad de GAPDH modificando la enzima ADP-ribosa. Esto se confirmó al inhibir la producción desuperóxidos con UCP-1 o MnSOD, previniendo así la modificación de GAPDH por la ADP-ribosa y ladisminuciónde laactividadpor lahiperglucemia. Estamodificaciónde laGAPDHpor laADP-ribosay lareducciónde suactividadpor lahiperglucemia también fueprevenidaporel inhibidorde lapolimerasapoli(ADP-ribosa)(PARP)(Brownlee,2005).

46
Figura 20. Sobreproducción de superóxidos mitocondriales activados por la hiperglucemia (Brownlee,2005).
En condiciones normales PARP permanece inactivo en el núcleo y se activa durante el daño del DNA.Cuandosegenera incrementodeROSmitocondrial, los radicales libres inducen rompimientodelDNAyposteriormente la activación de PARP (Figura 20). Éste a su vez crea polímeros de ADP-ribosa que seacumulanenGAPDHyotrasproteínasnucleares,modificandolaactividaddeGAPDHyalterandolasvíasyamencionadas(Brownlee,2005,Giacco&Brownlee,2010).
Figura21.Esquemadelosmecanismosafectadosporlahiperglucemia(Giacco&Brownlee,2010).
5.3.6.4Resistenciaalainsulinaydañomacrovascular
Adiferenciadelascomplicacionesmicrovascularesdondelahiperglucemiaeselprincipaldeterminantedeladiabetesyuncambiode laHbA1cde5.5%a9.5%aumenta10vecesel riesgo, en las complicacionesmacrovasculares,dichoriesgosóloaumentaeldoble.ElestudiodeSanAntonioHeartStudydemostróunincremento del 2.5 en las complicaciones cardiovasculares debido a resistencia a la insulina. Utilizandotejidocelularymodelosanimales,seencontróquelaresistenciaalainsulinageneraelaumentoenelflujodeácidosgrasoslibres(FFA)fueradelosadipocitosalascélulasendoteliales.ElincrementodeestosFFAen lascélulasendoteliales tambiénaumentasuoxidacióngenerandoaltonivelesdeROSyporelmismomecanismo que la hiperglucemia, se genera el desbalance mitocondrial y la activación de las 5 víasbioquímicasyamencionadas(Figura21)(Brownlee,2005;Giacco&Brownlee,2010).

47
Figura 22. Rol de la resistencia a la insulina y FFAs en la formación de ROS endotelial y aterogénesismacrovascular(Giacco&Brownlee,2010).

48
5.4EPIGENÉTICAYDIABETES
5.4.1Laproblemáticadelaepigenéticaenladiabetes
La “hipótesis unificadora” propuesta por Brownlee explica la fisiopatología de los efectos de lahiperglucemiaeneldañoprogresivoypermanentedelostejidosqueconllevanalareduccióndelacalidadde vida. La hiperglucemia intracelular persistente desregula procesos celulares vitales, generando lasobreproduccióndesuperóxidosanivelmitocondrial.Esteseríaelprimerpasocrucialenlaactivacióndelasvíasqueintervienenenlapatogeniadeladiabetes(Figura22)(Cooper&El-Osta,2010).
Figura23.ResumenesquemáticodelasvíasquecontribuyenalascomplicacionesdelaT2DM(Forbes&Cooper,2013).
Las modificaciones epigenéticas regulan funciones esenciales del metabolismo de los cromosomas.Conociendoloscambiosmolecularesdelaestructuradelacromatinaylarelacióndealteracionesdevíasmolecularessepuedeexplicarladiabetescomounfenómenodememoriametabólica.Aúnsedesconocenlosmecanismosmolecularesespecíficosqueconllevanaestamemoriahiperglucémicayaldesarrollodecomplicaciones vasculares que siguen siendo la principal causa demorbilidad ymortalidad de diabetes(Cooper&El-Osta,2010).
Eltérminodela“memoriametabólica”seutilizóparaexplicarunfenómenovascularalargoplazo,sobreelpobreefectoenelcontrolglucémico.PormuchotiempolosestudiosrealizadosporDiabetesControlandComplication Trial (DCCT) en 1983 y el seguimiento observacional de Epidemiology of DiabetesIntervention and Complication (EDIC) en 1994, demostraron como el control intensivo de la glucemiareduce la aparición y progresiónde complicacionesde la diabetes a largoplazo incluyendonefropatías,neuropatías y retinopatías. Los eventos cardiovasculares disminuyeron dando a conocer los efectossignificativosdelcontrolglucémico(Cooper&El-Osta,2010).Hoyendíaaúnnoseconoceelmecanismomolecularexactoquepuedeestarimplicadoendichamemoriametabólica.

49
5.4.2Memoriaepigenéticahiperglucémica
Estudios más recientes han planteado que no solo la hiperglucemia persistente causa cambios en lamemoriametabólica.Sehaplanteadolahipótesisdequela“memoriametabólica”eslacascadadelprocesodescritoinicialmenteporBrownlee,enelquelahiperglucemiaintracelularinducelasobreproduccióndesuperóxidos a nivelmitocondrial, proceso crucial en la activaciónde todas las vías que interviene en lapatogeniayenlascomplicaciones(Ceriello,2008a;Cooper&El-Osta,2010).
Hoyendíaseplanteaquelaproduccióndeespeciesreactivasdeoxígenonosóloestárelacionadaconlapresenciadehiperglucemiasinodependetambiéndelgradodeglicacióndeproteínasmitocondriales.Sehandeterminado intermediarios clavesquegenerandichamemoriaen laglicaciónde lasproteínas.Lasproteínasmitocondrialesquesufrenglicacionesavanzadasporefectode lahiperglucemia,pero tambiénsoncapacesdegenerarmásaniónsuperóxidosdentrodelasmitocondrias,apesardequelaglucemiasehayanormalizado,activandoasílasmimasvíasqueintervienenenlageneracióndecomplicacionesenladiabetes(Ceriello,2008a;Cooper&El-Osta,2010).
En la figura 23 se observa como la diabetes, la hiperglucemia y las glicación de proteínas avanzadas(AGE’s), activanmúltiples vías de transducción de señales que incluyen el estrés oxidativo que regulanfactores transcripcionales y factores epigeneticos. Estos factores son las metiltransferasas de histonas(HMT)ylasacetilasasdehistonas(HAT)yDNMT1quealteranmodificacionespost-traduccionales(PTM)de histonas y metilación en el DNA (meDNA) en promotores de genes inflamatorios y de la matrizextracelular, interleuquinas, factor transformante de crecimiento-B, inhibidor del activador deplasminógeno, cambiando la expresión.Lapersistencia en lasmarcasepigenéticas en regionesdegenestargetpuedenserunmecanismoimportanteenlamemoriametabólicayeldesarrollodecomplicacionesrenalesyvascularesapesardelcontrolglucémico(Villeneuveetal.,2011).
Figura 24.Memoriametabólicaymecanismosepigenéticos en la complicaciónde ladiabetes (Ceriello,2008a).

50
VI. METODOLOGÍA
6.1POBLACIÓNDEESTUDIO
Elpresenteestudiodecasosycontroles,seobtuvieroncuarentamuestrasdesujetoscondiagnósticodediabetesmellitustipo2yveintemuestrasdecontrolessanos,todoslosparticipantesasistenaconsultadeendocrinologíaenelHospitalUniversitarioFundaciónSantaFedeBogotá.LospacientesfueronreclutadoscondiagnósticopreviodediabetessegúnloscriteriosestablecidosporlaOMS(OrganizaciónMundialdelaSalud) y la ADA (Asociación Americana deDiabetes). Cada individuo aceptó participar en el estudio demanera voluntaria y firmó el correspondiente consentimiento informado (ver VII. Aspectos Éticos). Serealizó una historia clínica tipo encuesta para evaluar diferentes variables para el estudio comoinformacióndemográficarelevante,sexo,edad,tiempodediagnóstico,consumodemedicamentos,alcoholy tabaco. Los formularios y los consentimientos informados fueron archivados por el investigadorprincipaldelproyecto.
Loscriteriosdeinclusiónyexclusiónfueronlossiguientes:
A.Criteriosdeinclusión
PACIENTES
• Mayordeedad.
• DiagnósticopreviodeacuerdoalaAsociaciónColombianadeDiabetes.
• Noembarazo.
• Participaciónvoluntaria.
• Aceptacióndelascondicionesdelestudiopormediodelafirmadeunconsentimientoinformado.
CONTROLES
• Mayordeedad.
• Individuossanos,sinantecedentesfamiliaresopersonalesdediabetes.
• Sindiagnósticopreviodediabetesysinantecedentesdenivelesaltosdeglucosaensangre.
• Aceptacióndelascondicionesdelestudiopormediodelafirmadeunconsentimientoinformado.
B.Criteriosdeexclusión
• Menoresdeedad.
• Embarazadas.
• Diabetesgestacional.
• Pacienteseindividuoscontrolesquenodeseenparticiparenelestudio.

51
6.2RECLUTAMIENTOYRECOLECCIÓNDELAHISTORIACLÍNICADELOSPACIENTES
LospacientesfueronreclutadosyselesrealizóundiagnósticopreviodeDiabetesMellitusdeacuerdoalosestándaresinternacionales,AmericanDiabetesAssociationdelañocorrespondiente.Cadaindividuoaceptóparticiparenelestudiodemaneravoluntariaysefirmóelconsentimientoinformado.
La historia de los pacientes diabéticos fue realizada por el grupo de Endocrinología de la Clínica deDiabetesdelaFSFB.Serealizóunahistoriaclínicatipoencuestaparaevaluardiferentesvariablesparaelestudio.Los formatos fueronarchivadosenelLaboratoriodeGenéticaHumanade laUniversidadde losAndes.
Seobtuvo la informacióndemográficarelevante, incluyendosexo,edad, tiempodediagnóstico,consumodemedicamentos,alcoholytabaco.Sereclutaronlosindividuoscontrolesquecumplieranconloscriteriosdeinclusiónydesearanparticipardemaneravoluntariaenelestudio.
6.3MÉTODOSDELABORATORIO
6.3.1Obtencióndelamuestra
Seextrajeron5mldesangreperiféricaenuntuboconanticoagulanteEDTAacadaindividuo.Lasmuestrasse transportaron bajo los procedimientos adecuados en cadena de frío a 4°C hasta la realización de losprocedimientos.
Las cantidades necesarias para los diferentes ensayos fueron: 20μl para la medición de hemoglobinaglucosilada(HbA1c),250μlparalaextraccióndeRNAy3,5mldesangretotalparalaextraccióndelDNA.
6.3.2Pruebasclínicasdelaboratorioyhemoglobinaglucosilada(HbA1c)
Deacuerdoalasinstruccionesdelfabricanteserealizólapruebadelahemoglobinaglucósilada(HbA1c)conelkitcomercialNycoCardTMHbA1cenunlectorNycoCardTMREADERII.Estapruebasebasaenunatécnica de capilaridad que evalúa el control glucémico de los últimos 120 días de todos los individuossujetosalestudio.
De acuerdo a los resultados obtenidos se establecieron tres grupos de muestras: 1) grupo control, sinantecedentes de diabetes o factores influyentes, con niveles normales deHbA1, 2) pacientes diabéticoscontrolados(HbA1c≤7%)y3)pacientesnocontrolados(HbA1c>7%).Seestablecieronvaloresnormalesmenoresa7%deHbA1c.
6.3.3ExtraccióndeDNA
ParalaextraccióndelDNAseutilizóelkitcomercialdeextraccióndeDNACorpoGen2000®.Siguiendolasindicacionesdelfabricante,setrabajócon3,5mldesangretotal.
Serealizaroncuatrolavadosconlasolucióndelavadoconelfindeeliminarporlisiscelularlamayoríadeglóbulos rojos. Cada lavado se centrifugó (14,000 rpm por 1 minuto), se eliminó el sobrenadante y seresuspendieronlascélulasporvórtex.Setrabajóconunvolumenfinalde20μl.
Se adicionaron 500 μl de la solución de tapón de lisis y se mezcló por vórtex. Para desproteinizar lapreparación se prosiguió adicionando 500 μl de la solución salina, agitándolo vigorosamente hasta suhomogenización. Se incubó a -20°C por 30 minutos, se microcentrifugó (14,000 rpm) por 10 minutostomandocuidadosamente1mldelsobrenadantetransfiriéndoloauneppendorfde1.5ml.
ParalaprecipitacióndelDNAseadicionaron500μldeisopropanol,agitándolosuavementeporinversión.Secentrifugóyseeliminóelsobrenadantepor inversión.Se lavódosvecescon250μldeetanolal70%,centrifugandocadalavadoydesechandoelsobrenadanteporinversión.Lostubossesecaronenposicióninvertidasobreunpapelabsorbentepor5minutosyluegosedejaronalairepor15minutos.

52
Para lareconstitucióndelDNAseutilizaron100μlde lasolucióndereconstituciónysedejóa65°Cporunahora.Secuantificósuconcentraciónennanogramo(ng)pormediodelespectrofotómetroNanoDrop(ThermoSCIENTIFIC).TodaslasmuestrasdeDNAseguardarona-20°ChastalaspruebasdeELISA.
6.3.4ExtraccióndeRNA
LaextraccióndelRNAserealizóconelreactivocomercialTRIzol®LSReagentdeLifeTechnologies.Estemétodo se caracteriza por mantener la integridad del RNA por medio de la inhibición de RNasas. Porseguridadfrentealosreactivosqueconstituyenunriesgoparalasalud(trizol,cloroformoeisopropanol),enlafasedepreparacióndelamuestra,setrabajóencámaradeextracción.
Elprocedimientopartede laextracciónde250μldesangretotaladicionándole750μldeTRIzol.Estasehomogenizópor inversióny sedejóa temperaturaambientepor5minutosparapermitir ladisociacióncompletadeloscomplejosnucleoprotéicos.Seleadicionaron200μldecloroformo,seagitóvigorosamenteysedejóatemperaturaambientepor15minutos.Luegosecentrifugópor15minutosa4°Ca12,000gysetraspasólafaseacuosaauntubonuevo,evitandoarrastrarlainterfaseycompuestosorgánicos.
ParalaprecipitaciónyextraccióndelRNAseadicionaron500μldeisopropanolalafaseacuosa.Sedejóincubandoatemperaturaambientepor10minutosyluegosecentrifugópor10minutosa4°Ca12,000geliminandoelsobrenadanteydejandosóloelpellet.
Por último, éste se lavó con 1ml de etanol al 75%mezclándolo por vórtex. Luego se centrifugó por 5minutosa4°Ca7,500gysedescartóelsobrenadante.Parasecarelpelletsedejópor10minutosalaireyseresuspendióconagualibredeRNasasmezclándoloporinversión.Parafinalizarlaextracciónseincubóenbloquedecalorpor15minutosa56°C.
Inicialmente se guardó a -20°C hasta la realización del nanodrop para determinar su absorbancia yposteriormenteseguardóa-80°Cparasumejorconservación.
6.3.5CuantificaciónporNanodrop
Se realizaron las cuantificaciones de las concentraciones deDNA,RNA y cDNApara cadamuestra en elNanoDrop1000.Laslecturasserealizaronconlongitudesdeondade260/280nm.Seutilizarontasasdepurezasde1-8paraDNAyde ~2.0 paraRNA. Seguardaronen lasbasesdedatosdel laboratorioparaluegoserutilizadasenlasPCRs.
6.3.6Microelisasdemetilaciónehidroximetilación
Secuantificóelporcentajedemetilaciónehidroximetilacióndecadaindividuopormediodeunatécnicainmunológica de ELISA de fluorescencia procesada en microplacas. Para ello se utilizaron los kits deAbcam®: EpiSeeker Methylated DNA Quantification Kit, ab117129 y EpiSeeker Hidroxymethylated DNAQuantificationKit,ab117131.
EstosensayossebasanenadherirelDNAalfondodelpozodelamicroplaca.LafraccióndelDNAmetiladoo hidroximetilado es detectado utilizando un anticuerpo de captura y de uno de detección, cuyafluorescencia se cuantifica en un espectrofotómetro en unidades relativas de fluorescencia (RFU) conondas de excitación/emisión de 530/590nm. La cantidad de DNA metilado o hidroximetilado esproporcionalalaintensidaddefluorescenciamedida(Figura25)

53
Figura25.DiagramadeflujoparaelprocedimientosdelasELISA
En este experimento se partió con la extracción de DNA a partir de sangre periférica como se explicóanteriormente (Ver sección 6.3.3). El kit aportó los reactivos necesarios para la cuantificación de lametilaciónehidroximetilaciónglobaldecadaindividuo.SeajustaronlasconcentracionesdeDNAa100ngyseinicióconlasdilucionespreviasdelosbufferssegúnelprotocolo.
Preparacióndelbufferdelavadoal1X(BL1X):
Volumenbufferlavado(mL)
Aguadestilada(mL)
VolumenTotal(mL)
96Test 26 230 260
Tabla7.Bufferdelavadoal1X
Para cada ensayo se realizó una curva estándar de calibración con los controles positivos y negativosaportadosporelkit(Tabla8).
Tubos Control+(µl)
1XTE(µl)
Concentracionesdelcontrol+(ng/µl)
5mC(10ng/µl)
5hmC(5ng/µl)
5mC 5hmC 5mC 5hmC
1 1 0,5 19 12 0,5 0,22 1 1 9 9 1 0,53 1 1 4 4 2 14 2,5 2 2,5 3 5 25 4 5 0 0 10 5
Tabla8.Preparacióndelacurvaestándarde5mCy5hmC
Se preparó la dilución de los anticuerpos junto a la solución aumentadora. La solución reveladora defluorescenciasepreparóenuncuartooscuroalfinaldelprocedimientojustoantesdehacerlareacciónenlamicroplaca(Tabla9).
PreparacióndelADNgenómico
UnióndelADNalamicroplacasdeELISA
Adicióndelanticuerpodecapturayanticuerpodedetección.
Adicióndelasoluciónaumentadora
Agregarlasoluciónreveladorade�luorescenciaparalamedicióndeRFU.

54
REACTIVO DILUCIÓN VOLÚMENES(96POZOS)
Anticuerpodecaptura 1:1000 5µL 4,9995mLBL1X
Anticuerpodedetección 1:2000 2.5µL 4,997.5mLBL1X
Soluciónaumentadora 1:5000 1µL 4,999mLBL1X
Soluciónreveladorade
RFU*
10µLsoluciónreveladora*10µLslnaumentadoradefluorescencia*
5mLdiluyente*Tabla9.Dilucióndeanticuerpos,soluciónaumentadorayreveladoradefluorescencia(Estosreactivossepreparanalfinalantesdehacerlareacción)
La unión del DNA se inició con la adición de la solución de unión a cada pozo utilizando una pipetamulticanal.Seadicionaronlosbuffercontrolylos100ngdeDNAentre1-8μl.Setapólaplacaconparafilmyseincubóa37°Cpor90minutos.Alfinalizareltiemposeeliminólasolucióndeuniónyserealizarontreslavadosdelospozoscon150μldebufferdelavado.
Se adicionaron 50 μl del anticuerpo de captura de DNA a cada pozo y se tapó e incubó a temperaturaambiente por 60 minutos. Al finalizar el tiempo se eliminó la solución del anticuerpo de captura y serealizarontreslavadosdelospozoscon150μldebufferdelavado.
Seagregaron50μldelanticuerpodedetecciónacadapozo,setapóeincubóatemperaturaambientepor30minutos.Alfinalizareltiemposeeliminólasolucióndedetecciónyserealizaroncuatrolavadosdelospozoscon150μldebufferdelavado.
Seadicionaron50μldelasoluciónpotenciadoraacadapozo,setapóeincubóatemperaturaambientepor30minutos.Alfinalizareltiemposeeliminólasolucióndedetecciónyselavaronlospozoscon150μldebufferdelavado5veces.Finalmente,selavócadapozocon150μlconPBSal1X.
Sepreparóaoscuraslasoluciónreveladoraparadetectarlaseñaldefluorescencia.Sepreparóunamezcladevariosbuffersagregando50μldeestasoluciónacadapozo.Seincubópor4minutoshastasulectura.Lafluorescenciasemidióenunidaddefluorescenciarelativa(RFU)enunfluorómetroBioTekconondasdeexcitación/emisiónde530/590nm.
6.3.7RT-qPCR
6.3.7.1Diseñodecebadores
Se utilizó el programa bioinfomático Primer3 Plus para diseñar la secuencia de oligonucleótidos paraamplificar el fragmentode interés de la región codificante deGAPDH ,DNMT1 yTET1. Se utilizó el genGAPDHcomocontrolpositivodelareaccióndePCR.
Se consideraron parámetros importantes en el diseño como la longitud de nucleótidos entre 18-24,temperaturademelting,temperaturadeanillamiento,elcontenidodeGC’sentre40-60%,laestabilidaddelextremo 3’ con la formación de un GC clamp, así como la formación de estructuras secundarias o laformacióndedímerosdeprimers (cebadores) teniendoen cuenta losvaloresmenoresa4de losANYySELFANY.
LoscebadoresdelaqPCRsediseñaronapartirdelcDNAyseverificaronpormediodelprogramaBLASTNparaasegurarquenofuerancomplementariosconotrassecuenciasnucleotídicasenelgenoma(Tabla10).

55
Gen Secuenciadecebadores
GAPDH Forward 5’-CCACCCATGGCAAATTCCATGGCA-3’
Reverse 5’-TCTAGACGGCAGGTCAGGTCCACC-3’
DNMT1 Forward 5’-GCACGAATTTCTGCAAACAG-3'
Reverse 5’-CACTTCCCGGTTGTAAGCAT-3'
TET1 Forward 5’-TTCTGACAAGAGCTGGAGCA-3’
Reverse 5’-TGGAACCTTTTTGGATTTGG-3’
Tabla10.Cebadoresdecontrolyamplificación
6.3.7.2EstandarizacióndecebadoresporPCRconvencional
SeprepararonloscebadoresdelosgenesGAPDH,DNMT1yTET1aconcentracionesfinalesde500nm.Deesta dilución se utilizo 1 μl de cada cebador para un volumen final de 20 uL. Se utilizaron muestrasproblema para la estandarización. Se evaluó la actividad correcta de los cebadores en una PCRconvencionalutilizandoelkitdePromegaGoTaqGreen(Tabla11).
VolumenFinal20μl
Agua(ddH2O) 7μlMasterMix 10μlPrimerforward 1μlPrimerreverse 1μlcDNA 1μlTabla11.VolumendePCRconvencional
Lascondicioneseneltermocicladorseencuentranenlasiguientetabla.
Programadeltermociclador Temperatura TiempoDenaturacióninicial 95°C 2minutos30Ciclos
Denaturación 95°C 45segundosAnillamiento 55°C 1minutoExtensión 72°C 30segundos
Extensiónfinal 72°C 5minutosInfinito 4°C
Tabla12.PCRconvencionalGoTaqGreen
6.3.7.3SíntesisdelaprimerahebradecDNA
SeutilizóelkitdePromegaImProm-IITMReverseTranscriptionSystempara larealizaciónde laprimerahebradecDNAapartirdelRNAtotal.Estekit incluye la transcriptasareversay losreactivosnecesariosparaoptimizarlaeficienciaenlasíntesisdecopiasdelDNAcomplementario(cDNA).
En la síntesis de la primera hebra del cDNA se utilizaron los oligonucleótidos de timina (oligo dT),obtenidosenelkit.Serealizóunaprimerareaccióncon losvolúmenes indicadosen latabla13. Elaguautilizada era libre de RNasas. Esta mezcla se colocó en el bloque de calor a 70°C por 5 minutos yrápidamenteenhielopor5minutos.Seprosiguióconunspinparaprecipitartodalamezcla(Tabla13).

56
VolumenFinal5μlAgua 2μlOligodT*2 0.5μlARN 2μl
Tabla13.VolúmenesparalasíntesisdelaprimeracadenacDNA
Para la segunda reacción de la cadena semezcló la reacción principal con los reactivos en la tabla 14.Previamente se hizo la respectiva dilución del MgCl2 a 6mM. Se mezclaron los componentes para unvolumenfinalde20µlysecolocóeneltermocicladorparasuposterioramplificación(Tabla14).
VolumenFinal20μl
Agua 6.2μlBuffer de RXN Im-PromIITM 4μl
MgCl2(6mM) 2.8μldNTP’s 1μlRetrotranscriptasadeIm-PromIITM 1μl
Tabla14.SegundareacciónparalasíntesisdelcDNA
Lasiguientetablaresumelascondicionesestablecidaseneltermociclador.Estareacciónserealizóenuntermocicladorconvencional.
Programadetermociclador Temperatura Tiempo
Anillamiento 25°C 5minExtensión 42°C 60minInfinito Mantenidoen4°C
Tabla15.Condicionesdelasíntesisdeprimerahebraeneltermociclador
ElcDNAamplificadodeguardóa-80°Cparasuconservación.
6.3.7.4ReaccióndePCRconvencionalparaelcontrolpositivoGAPDH
SeutilizóelgenconstitutivoGAPDHcomocontrolpositivodelareaccióndelaPCR.Estapruebaserealizócon una PCR convencional con el kit de Promega Go Taq Green. Se realizó la desactivación de laretratransciptasaa70°Cpor15minutoseneltermociclador,antesdeiniciar.
Para la prueba de GAPDH se realizó la siguiente reacción presentada en la tabla 16. Los cebadores deGAPDHsereconstituyerona10mM.Paraunvolumenfinalde20µldelareacciónyunaconcentraciónfinalde500nMserealizaronalícuotasde1µly10µl.Setrabajóconunatemperaturademelting(Tm)de58°C.
VolumenFinal20μl
Agua 7μlPrimerF 1μlPrimerR 1μlMasterMixGoTaqGreen
10μl
cDNA 1μlTabla16.VolúmenesparalaPCRconvencional
SecolocólareaccióneneltermocicladorconlassiguientescondicionesdePCR.

57
Programadeltermociclador Temperatura TiempoDenaturacióninicial 95°C 2minutos30Ciclos
Denaturación 95°C 45segundosAnillamiento 58°C 1minutoExtensión 72°C 30segundos
Extensiónfinal 72°C 5minutosInfinito 4°C
Tabla17.CondicionesdelcontrolpositivoparaGAPDH
6.3.7.5GeldeelectroforesisenagarosaparaloscontrolesdeGAPDH
Se realizó un gel de agarosa al 1.5% para visualizar los resultados de la PCR realizada para el controlpositivo de la reacción de GAPDH. Para su gelificación, se utilizó el buffer TBE al 0.5X, los gramoscorrespondientedeagarosasegúneltamañodelacámaraquevaríanentre0.8-1.6gramosdeagarosayelbufferdecorridoTBEal0.5X.Lascondicionesdecorrido fueronde40-50minutoscon80voltiosy400MA.ParalavisualizaciónseutilizóelintercalateEzvision.
Se visualizaron las bandas del marcador de peso y las de GAPDH para cada individuo. Utilizando unfotodocumentadoryelprogramaGelDocseconfirmópormediodefluorescenciaeltamañodeinterésde496pb.
DelasmuestrasqueresultaronnegativasparaGAPDH,sevolvieronasintetizarsusrespectivoscDNA.
6.3.7.6ReaccióndeqRT-PCRdelosgenesDNMT1yTET1
PormediodetécnicascuantitativasdePCR,serealizólaamplificacióndelosfragmentosdeinterésconelkitPowerSYBR®GreenPCRMásterMixdeAppliedBiosystem®.EstemétodoutilizaunaAmpliTaqGoldDNApolimerasaquesecaracterizaporsualtasensibilidadyporsufuncionalidaddeHot-Start.
Estemétodo de amplificación se basa en la unión de la tinción SYBR Green a la doble cadena de DNA,proporcionandouna señal fluorescenteque refleja la cantidaddeproducto generadodurante laPCR.ElSYBRGreenesuncompuestoorgánicoqueinteractúaconlahendiduramenordelDNA, introduciéndoseenlaestructurasecundariadeladoblehéliceyacoplándoseenergéticamentealosácidosnucleicos.Estaunión emite fluorescencia por medio de un fenómeno llamado transferencia de energía medianteresonanciadefluorescencia(FRET).Lalecturaesrealizadaanalizandopicosdeabsorcióndelongitudesdeonda(λ),λ=498mndeabsorciónyλ=522nmdeemisión(Figura25).
Figura26.AccióndelSYBRGreen
Antesdeiniciarlascuantificacionesenelproyecto,serealizólaoptimizacióndelasconcentracionesdeloscebadoresparadeterminarlaconcentraciónmínimadeexpresiónquedécomoresultadoelmenorumbraldeciclo(CT)yelmáximo∆Rnparaasíreducirlasamplificacionesinespecíficas(Tabla18).
cDNA GAPDH,DNMT1&TET1(nM) GAPDH,DNMT1&TET1(nM)

58
(ng/μl) Forward Reverse25 50 100 250 500 50 100 250 50050 50 100 250 500 50 100 250 500200 50 100 250 500 50 100 250 500500 50 100 250 500 50 100 250 500
Tabla18.Estandarizacióndeconcentraciones
A partir de unamuestra problema ya conocida, se trabajó para buscar las concentraciones óptimas decDNA y cebadores para utilizar en el procedimiento. Se ajustó la concentración de cDNA a cuatroconcentraciones(25,50,200,500ng/μl)distintasyseevaluaroncadaunoconcuatroconcentraciones(50,100,250y500nM)distintasdecadaprimerparacadagen(GAPDH,DNMT1yTET1).
LosvolúmenesparalaPCRvariaronsegúnlasconcentracionesdeDNAycebadores.
Lascondicionesdeltermocicladorseencuentranenlasiguientetabla:
Programadeltermociclador Temperatura TiempoDenaturacióninicial 95°C 10min40Ciclos
Denaturación 95°C 30segAnillamiento 58°C 45segExtensión 72°C 45seg
Curvadedisociación Tabla19.CondicionesdePCRcuantitativa
Laspruebasconmejorperfildeexpresiónparacadaunadelasmuestrasygenesamplificadosfueron:
GEN Cebadores cDNAGAPDH 500nM 25ng/μlDNMT-1 500nM 25ng/μlTET-1 500nM 25ng/μl
Tabla20.Concentracionesóptimasdecebadores
Teniendo en cuenta las concentraciones de cDNA para cada individuo, se diluyeron a 100 ng/μl. Lasconcentraciones iniciales del cDNA eran de 2000ng/μl y se adicionaron 20 μl a cada tubo. Todos loscebadoressetrabaronaconcentracionesde500nM(Tabla20).
ParalareaccióndelaPCRcuantitativasetrabajóconunvolumenfinalde20µl.LosvolúmenessepuedenobservarenlaTabla21.EsteexperimentoserealizóenunaplacadePCRMicroAmp®optical96wellplatey se montó la curva de calibración por cada experimento. Cada gen de cada individuo se evaluó portriplicadoyelcontrolpositivoGAPDHseevaluóporduplicado.Seanalizaron16individuosporcadaplaca.
Volumen(µl)
Concentraciónfinal
PowerSYBR®GreenPCRMM 10 1X
PrimerForward 1 500ngPrimerReverse 1 500ngTemplado 1 100ngddH2O 7 -Total 20 -
Tabla21.VolúmenesdelaqPCR

59
LasamplificacionesentiemporealenelaparatodeAppliedBiosystem®7500FastReal-TimePCRde laUniversidaddelosAndes.Antesdeiniciarlaamplificaciónserequiere10minutosa95°CparalaactivacióndelaAmpliTaqGoldDNApolimerasa,UPparamejorsensibilidad.
Lascondicionesdelosciclosfueronson:
Programadeltermociclador Temperatura TiempoDenaturacióninicial 95°C 10min40Ciclos
Denaturación 95°C 30segAnillamiento 55°C 45segExtensión 72°C 45seg
Curvadedisociación Tabla22.CondicionesdelaPCRcuantitativa
Lamediciónsetratadelacuantificaciónrelativadelosamplicones.Sebasaenladetecciónsimultáneadedos segmentos de DNA en el cual uno representa el target, los genes de interés y el gen constitutivoGAPDH.Sehaceuna detecciónsimultáneadeambosgenesyal final la concentraciónsedeterminaconbasealaconcentracióndelamplificadointerno.
6.3.8Microarreglos
Se analizaron un total de 416 microarreglos extraídos de la base de datos de NCBI Gen ExpressionOmnibus (GEO) (Edgar et al., 2002). Se obtuvieron los resultados de distintos estudios con distintasplataformasparavariostejidosafectadosendiabetes.ParaanálisisdemetilacióndelDNAserecolectaron267 arreglos de la plataforma Illumina Human Methylation 27 BeadChip de los siguientes estudios:GSE21232,GSE34008,GSE38291,GSE20067; en losque seanalizaron casosy controles enmuestrasdesangre periférica, islotes pancreáticos, tejido adiposo y músculo. Por otra parte, para los análisis deexpresióngénica seobtuvieron125arreglos:45AffymetrixHumanGenomeU133Plus2.0Arrayde losestudios GSE27949, GSE19420, GSE55098; 63 Affymetrix Human Genome 1.0 ST Array del estudioGSE38642y17IlluminaHumanRef-8v3.0expressionbeadchipdelexperimentoGSE21321,enlosqueseanalizaron igualmente muestras de casos y controles de sangre periférica, islotes de páncreas, tejidoadiposoymúsculo(Tabla23).
TipodeEstudio Plataforma Tejidos #
Pacientes#
ControlesGEO
Accession#
Arreglodeexpresión
AffymetrixHumanGene1.0STArray
Islotespancreáticos 9 54 GSE38642
Arreglodeexpresión
AffymetrixHumanGene1.0STArray corazón 7 5 GSE26887
Arreglodemetilación
HumanMethylation27BeadChip
MúsculoesqueléticoyTejidoadipososubcutáneo
16 16 GSE38291
Arreglodemetilación
HumanMethylation27BeadChip
Islotespancreáticos 5 11 GSE21232
Arreglodemetilación
HumanMethylation27BeadChip Sangretotal 195 0
GSE20067,GSE34008,GSE57285,GSE61611

60
Arreglodeexpresión
ExiqonmiRCURYLNAmicroRNAArray
MúsculoesqueléticoyTejidoadipososubcutáneo
9 9 GSE26168
Tabla23.Tabladescriptivademicroarreglos
Paralosresultadosdelosmicroarreglosserealizóunanálisisdealgoritmosdeclusterizaciónquebuscalascaracterísticas similaresdentrodeunconjuntodedatos. Seanalizaron relaciones complejasde caráctermultivariadoeidentificaciónatravésdeespaciosdemúltipledimensiones,elementosanómalos(outliers).Losdatosmultivariadosobtenidosen lamatrizsemontaronalprogramaGeneCluster3.0.Latécnicadeanálisisdeagrupamientoutilizadafueladesimilaridadmétrica.Estatécnicaformagruposdeelementosdemaneraquelosdatosquepertenecenaungrupodeelementosseanparecidosentresi.EstosevisualizóenJavaView.
Para el análisis de expresión diferencial se realizó en el software Chipster. Este software contieneherramientasparaelanálisisdemicroarreglosdeúltimageneración.ElrestodelosanálisisserealizaronenelprogramadeR.
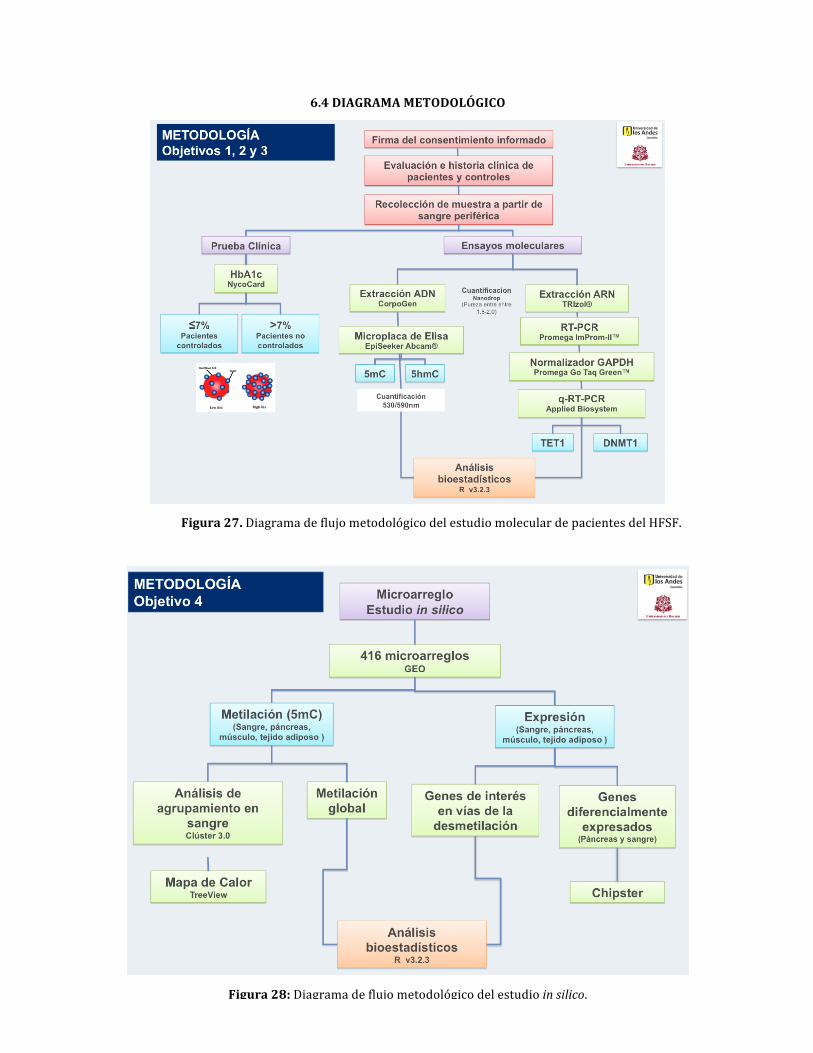
61
6.4DIAGRAMAMETODOLÓGICO
Figura27.DiagramadeflujometodológicodelestudiomoleculardepacientesdelHFSF.
Figura28:Diagramadeflujometodológicodelestudioinsilico.

62
6.5ANÁLISISESTADÍSTICOSYBIOINFORMÁTICOS
Elanálisisestadísticoinicióconlacaracterizacióndemográficadelossujetosyelcálculodelasfrecuenciasde lasvariablesestudiadasobteniendo lospromedios,desviacionesestándares,mediasymedianasparalosdatoscuantitativos.
6.5.1Análisisestadísticosdelasmicroelisas
Paraobtenerlosporcentajesdemetilaciónehidroximetilacion,seutilizoelcontrolnegativocomercialdelkitparaajustarlasunidadesdefluorescenciarelativa(RFU).Paracadaexperimentoserealizóunacurvade calibración con diluciones delDNA (0.5, 1.0, 2.0, 5.0 y 10.0 ng/µL para 5mc y 0.2, 0.5, 1.0, 2.0 y 5.0ng/µLpara5hmC).LacuantificaciónabsolutadelosresultadosserealizóapartirdelosdatosdeRFUdelacurvaestándarversuslacantidaddelamismadeloscontroles.Paradeterminarlapendientedelacurvaestándar se realizó una regresión lineal de la RFU/ng. Se prosiguió con el cálculo de la cantidad yporcentajedelDNAmetiladoportriplicadopormediodelassiguientesfórmulas.
Fórmula 1. Cálculos del porcentaje de 5mC S: cantidad de DNA en ng. *2: factor para normalizar elcontrol(+)de5mcal100%;elcontrolpositivotieneel50%de5mC.
Fórmula 2. Cálculos del porcentaje de 5hmC.S:cantidaddeDNAenng.*5: factorparanormalizarelcontrol(+)de5mhcal100%;elcontrolpositivotieneel20%de5mC.
Se crearon las tablas de los resultados de las ELISAS de metilación e hidroximetilación adicionandovariablespertenecientesa losdatosdel%de5mC,5hmCyHbA1c, controlglucémico,edady tiempodediagnóstico, consumo de alcohol y tabaco y frecuencias del consumo. Igualmente se formaron grupossegún el control glucémicode los individuos (1. controles, pacientes; 2. controles, pcientes controlados,pacientesnocontrolados;3.controlesypacientes,pacientesnocontrolados).ApartirdeestainformaciónserealizóunamatrizenformatoTXTyelanálisisestadísticoserealizóconelsoftwareRStatisticversión3.3.0.SecalculólanormalidaddetodaslasvariablesconeltestdeShapiro-Wilkysevisualizaronenformadehistograma.
Alevaluarlanormalidaddelosdatos,seobtuvieronvaloresp>0.05yseestablecióquelosporcentajesde5mCy5hmCnosiguenunadistribuciónnormalyporesarazónsetrabajóconpruebasno-paramétricas.Para el análisis de dos categorías (el efecto de 5mC y 5hmC según sexo, tiempo de diagnóstico, edad,consumodealcoholotabaco),serealizó lapruebadeWilcoxquecomparar lamedianadedosmuestras
5mC(ng)=RFUdecadamuestra–RFUcontrolnegativo Pendientex2
5mC%=Cantidadde5mCenng Sx100
5hmC(ng)=RFUdecadamuestra–RFUcontrolnegativo Pendientex5
5hmC%=Cantidadde5hmCenng Sx100

63
relacionadas y determinar si existen diferencias entre ellas. Para el análisis de dos o mas grupos seaplicaronlapruebadeKruskal-WallisyelensayodeDunnparacomprobarsiungrupodedatosprovienedelamismapoblación.Paraasociarla5mCy5hmCalasvariablesdelestudioseutilizaronlaspruebasdecorrelacióndeSpearmanyrangodeKendall.
6.5.2AnálisisestadísticosdeRT-qPCR
A partir de los resultados de las Ct obtenido de la RT-qPCR para cada gen, se realizó una matriz enMicrosoft Excel para sacar el promedio de cada muestra realizada por triplicado para DNMT1, TET1 yGAPDH. Se calculó la mediana de los sujetos controles y se utilizó ese valor como valor de referenciacontrol.ConlainformaciónobtenidadelaeficienciayelpromediodelosCtdecadagenserealizaronloscálculosparaelΔCt yΔΔCt,usandocomogennormalizadorelGAPDHparaasí calcular la cuantificaciónrelativa deDNMT1y TET1. Este último dato es adjuntado a la matriz general de los resultados de lasmicroelisas y todas las variables para llevar a cabo un análisis en conjunto. Se tomaron niveles deexpresión significativos según valores de p menor a 0.05. Los cálculos se realizaron con las siguientesfórmulas.
Fórmula3.FórmulasparalacuantificaciónrelativadelaRT-qPCR
Los análisis estadísticos se realizaron en R., para determinar la normalidad de los datos se realizó laprueba de Shapiro-Wilk y se visualizaron en histogramas. Los datos deDNMT1 se trabajaron como nonormalesmientrasquelosdatosdeTET1comonormales.
SerealizaronpruebasdecorrelacióndeSpearmanyKendallparalaasociacióndelaHbA1cylaexpresióndelosgenesdeinterésDNMT1yTET1.ParalacomparacióndelosgruposylasdiferenciasenlaexpresióndelosgenesdeinterésserealizólapruebadeWilcoxparacontrolesypacientesylapruebadeKruskal-Wallis para controles, pacientes controlados y pacientes no controlados. Se realizaron pruebas decorrelación de Spearman y Kendall para la cuantificación relativa y los porcentajes de metilación ehidroximetilación, edad y tiempo de diagnóstico. Por último, se analizó la cuantificación relativa segúnsexo,consumodetabacoyconsumodealcoholutilizandolapruebadeWilcoxparaDNMT1ylapruebaT-studentparaTET1.
6.5.3Análisisestadísticosybioinformáticosdemicroarreglos
El análisis de agrupamiento (clusterización) se realizó a partir de los datos de sangre periférica depacientesdiabéticosycontrolesconelsoftwareGeneCluster3.0.LasmatricesenformatoTXTsecargaronenelprogramayseestablecieronlascondicionesdelanálisisdejandolosfiltrosdelosdatospordefecto.

64
Estos remueven genes que han perdido valores y datos no informativos. Los datos se ajustaron contransformaciones Log2(X) y se realizó median-polish para centralizar los datos de genes y arreglosalrededorde lamediana.Estoúltimosehaceparaeliminar losposiblessesgosy losdatosextremosporfueradelasdistribuciones(outliers).Losdatosdegenesyarreglossenormalizaronparaajustarlasumadeloscuadradosa1.
Paraelanálisisdeclusterizaciónserealizóunagrupamientojerárquicoqueconsisteenobtenergruposdeelementosdelamatrizdeasociaciónenelcualseestructuranlosgruposdediferentesnivelesdesimilituden todas las posibles combinaciones. Este organiza los genes en un dendograma. El método deagrupamiento es de ligamiento centroide que se basa en agrupar elementos y variables utilizando ladistanciaeuclidianaentre loscentrosde losclusteracomparar.Estasecaracterizapor representarunasimplificacióndelpromedionoponderado.LosresultadosdeesteanálisissevisualizaronconelsoftwareTreeView1.1.6r4.paragenerarunmapadecalorqueesrepresentadoporcoloressegúnmetilación.
Paralacomparacióndelametilaciónglobaldelostejidosdesangreperiférica,músculoesquelético,tejidoadiposoytejidopancreáticoserealizóunasumadelosvaloresdemetilaciónparacadamicroarreglo.Conestos resultados se realizó el análisis diferencial de metilación que se visualiza en un boxplot en elsoftwareestadísticoRversión3.3.0(http://www.r-project.org).
ElanálisisdelosgenesasociadosalasrutasdeladesmetilaciónpasivayactivadelDNAserealizóapartirdelosmicroarreglosdemetilaciónyexpresión.SellevoacaboenelprogramaRenlacualsedeterminólanormalidaddelosdatospormediodeShapiro-WilkylahomogeneidaddevarianzasporlapruebaF.ParaelanálisisdelasvariablesserealizaronpruebasTdeStudent,TdeWelchopruebaderangosdeWilcoxonjuntoasusgráficascorrespondientes.
ElanálisisdemetilaciónyexpresióndiferencialdegenesserealizóenMicrosoftExcel2011v14.6.4pormedio de un análisis de varianzas ymodelamiento lineal a partir del paquete limma deBioconductor yEmpiricalBayes pormedio del software Chipster v.3.8.1 (http://chipster.csc.fi). Para la realización delanálisissecargarelarchivodeexpresióndiferencialdetejidopancreático.
Enlaherramientadenormalizacióndedatosseseleccionaelprocesamientodedatosprenormalizados,yaque los datos descargados de la base de datos de GEO ya se encuentran previamente procesados. Elresultado arroja un phenodata, el cual se edita para designar los pacientes y controles de cadamicroarreglo.Apartirdeestamodificaciónseescogeelmétodoestadísticodemodelo lineal, el cual fuerealizadopormediodelpaquetelimma,queescogelosgenesdiferencialmenteexpresados.Porúltimo,sevisualizanestosresultadosmedianteVolcanoPlots.Enestosgráficos,seescogierongenesconfoldchangessuperioresa1.00einferioresa-1.00.

65
VII. ASPECTOSÉTICOS
ElpresenteestudiofueaprobadoporelComitédeÉticaenInvestigacionesdelaUniversidaddelosAndesydelHospitalUniversitarioFundaciónSantaFedeBogotá.Losdatosobtenidosseorganizaronenunabasededatosqueesarchivadaycustodiadaporelinvestigadorprincipal.Losdatosgeneradoseneldesarrollodelproyectofueronregistradosyarchivadosporelresponsabledelprocedimiento,quienmensualmenteentregó una copia de los mismos al investigador principal. Cada participante recibió un código deidentificaciónyseprotegió laprivacidaddel individuo.EsteestudioesconsideradounainvestigaciónderiesgomínimosegúncriteriosestablecidosporlaResoluciónNo.008430de1993delMinisteriodeSalud“NormasCientíficas,TécnicasyAdministrativasparalaInvestigaciónenSalud”acercadelainvestigaciónensereshumanos.Enningúnmomentoseadvirtióningúnriesgoodañopara lasaluddeningunode lossujetosenquienesserealizólainvestigación.
Los estudios se realizaron con muestras de sangre periférica, lo cual no tiene ningún impacto en eltratamientode lospacientesquienes son seguidosenprotocolosyaestablecidos. La informaciónque sedesprendiódelestudiopuedegenerarinformaciónrelacionadaconeldiagnósticoyelpronósticotalcomolaliteraturalorefiereysepondráadisposicióndelosclínicos.
Paralarecoleccióndelmaterialbiológicodelosparticipantes,ellugardeejecucióndelproyectocontóconlas instalaciones y equipo de laboratorio de acuerdo con las normas técnicas del Ministerio, quegarantizaronelmanejosegurodelasmuestras.Ellaboratoriodisponíadeunmanualdeprocedimientosypersonaladiestradoparalamanipulacióndelasmuestras.
Todo procedimiento fue aprobado por el Comité de Ética el de la Universidad y así proceder con laautorizaciónparaseraprobadoporelComitédeÉticadelaFundaciónSantaFe.Setuvieronencuentalasnormas nacionales establecidas en la Resolución 8430 del Ministerio de Protección Social y normasinternacionalesbasadasenlaúltimarevisióndelaDeclaracióndeHelsinki2013.
Toda información se manejó de manera confidencial. Los investigadores no tuvieron acceso a laidentificacióndealgúnindividuo.Paraprotegerlaidentidaddelosparticipantes,secrearoncódigosylosresultadossepresentarondeformaanónima.Losparticipantesdeesteestudiofueroninformadossobreelmismoyselespidiósuconsentimientoinformadoatravésdelsiguienteformato:

66

67

68

69
VIII. RESULTADOS
8.1EFECTODELCONTROLMETABÓLICOENLAMETILACIÓNEHIDROXIMETILACIÓNDELDNAENT2DM
Paraanalizarloscambiosepigenéticoscausadosporladiabetes,secuantificaronlosnivelesdemetilaciónydehidroximetilaciónglobalenmuestrasdesangreperiféricadepacientescondiabetestipo2ycontrolessanos. Atodaslasmuestrasselesrealizóuncontroldelestadoglucémicoatravésdelamedicióndelosniveles de hemoglobina glucosilada (Hb1Ac), lo que permitió dividir a los pacientes en dos grupos: ungrupo de pacientes con niveles de glucemia controlados, que se caracterizan por presentar un valor deHb1Ac≤7 y un grupo de pacientes con un control metabólico deficiente, caracterizados por niveles deHb1Ac≥7(Vertabla24).
Tabla24.Tablacaracterísticadelosgruposdeanálisis
Al comparar los niveles de metilación global entre el grupo de pacientes y los controles sanos no seobtuvieron diferencias significativas (p=0.2204)(Figura 29A). Sin embargo, se observa una tendencia apresentar niveles superiores de metilación en el grupo de pacientes, resultado contrario a lo que seesperaríadeacuerdoconlosdatosconocidosenotrostejidos(Dhliwayoetal,2014;Ding&Huang,2014).
Teniendo en cuenta esta tendencia, se decidió separar y analizar a los pacientes según su estadometabólico, demostrando que el mal control metabólico está asociado con los cambios epigenéticos,provocandounaumentodemetilaciónestadísticamentesignificativofrenteacontrolessanosypacientesbiencontrolados(pacientesnocontroladosvs.controlessanos:p=0.007331;pacientesnocontroladosvs.controlados:p=0.002915)(Figura29C).Igualmenteocurriócuandoseagruparonloscontrolessanosylospacientesbiencontroladosenunsologrupoysecompararonconlospacientespobrementecontrolados.Estacomparacióndiocomoresultadounadiferenciaaltamentesignificativa(p<0.001)(Figura29E),loquedemuestra nuevamente que el control metabólico y los defectos oxidativos estarían en la base de losdefectosepigenéticosdetectadosenlaenfermedadyporlotanto,unpacientebiencontroladosecomportaaestenivelcomounapersonasana,contodoloqueestoconllevaeneldevenirdesuenfermedad.
Elestudiodelosnivelesglobalesdehidroximetilaciónfueaúnmásinteresante.Aunquenoseobservóunadiferencia estadísticamente significativa entre pacientes y controles al igual que con los niveles demetilación,latendenciaaserdiferentesesclaraconunvalorp=0.092(Figura29B).Alcompararlostresgruposporseparado,tambiénseobservóquelahidroximetilacióndelospacientescontroladosseasemejaal grupo control y ambos obtuvieron niveles de hidroximetilación estadísticamente menores que lospacientes no controlados (pacientes controlados vs. no controlados: p=0.01217, controles vs nocontrolados:p=0.006715)(Figura29D).Igualqueenlametilación,seanalizóelgrupodecontrolessanosjuntoalospacientescontroladosfrentealospacientesnocontrolados,comparaciónquediounadiferenciasignificativa(p=0.002326)(Figura29F),demostrandonuevamentequelafaltadecontrolglucémico,consusniveles elevadosdeoxidación celular, es labasede los cambioshalladosanivel epigenéticoenestaenfermedad.

70
A. Metilación B.Hidroximetilación
C. D.
E. F.
E. F.
Figura29.PorcentajedelametilaciónenhidroximetilaciónglobaldelDNA.A)Metilaciónglobalencontroles sanosvs.pacientes,B)Hidroximetilaciónglobalencontroles sanosvspacientes,C)Metilaciónglobal en controles sanos, pacientes controlados y pacientesno controlados,D)Hidroximetilación entrecontrolessanos,pacientescontroladosypacientesnocontrolados,E)Metilaciónglobalencontrolessanoypacientescontroladosvspacientesnocontrolados,F)Hidroximetilaciónentrecontrolessanosypacientescontroladosvspacientesnocontrolados.*=p<0.05,**=p<0.01,***=p<0.001.
**
***
**
*
**
**
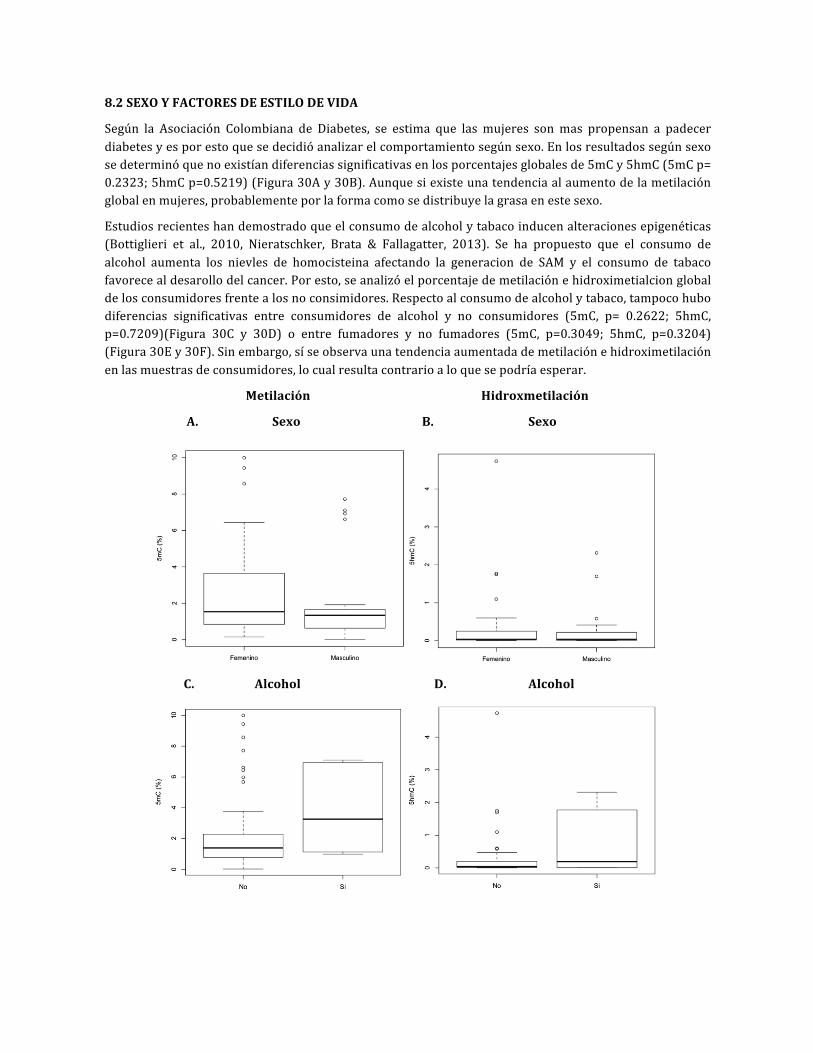
71
8.2SEXOYFACTORESDEESTILODEVIDA
Según la Asociación Colombiana de Diabetes, se estima que las mujeres sonmas propensan a padecerdiabetesyesporestoquesedecidióanalizarelcomportamientosegúnsexo.Enlosresultadossegúnsexosedeterminóquenoexistíandiferenciassignificativasenlosporcentajesglobalesde5mCy5hmC(5mCp=0.2323;5hmCp=0.5219)(Figura30Ay30B).Aunquesiexisteunatendenciaalaumentodelametilaciónglobalenmujeres,probablementeporlaformacomosedistribuyelagrasaenestesexo.
Estudiosrecienteshandemostradoqueelconsumodealcoholytabacoinducenalteracionesepigenéticas(Bottiglieri et al., 2010, Nieratschker, Brata & Fallagatter, 2013). Se ha propuesto que el consumo dealcohol aumenta los nievles de homocisteina afectando la generacion de SAM y el consumo de tabacofavorecealdesarollodelcancer.Poresto,seanalizóelporcentajedemetilaciónehidroximetialcionglobaldelosconsumidoresfrentealosnoconsimidores.Respectoalconsumodealcoholytabaco,tampocohubodiferencias significativas entre consumidores de alcohol y no consumidores (5mC, p= 0.2622; 5hmC,p=0.7209)(Figura 30C y 30D) o entre fumadores y no fumadores (5mC, p=0.3049; 5hmC, p=0.3204)(Figura30Ey30F).Sinembargo,síseobservaunatendenciaaumentadademetilaciónehidroximetilaciónenlasmuestrasdeconsumidores,locualresultacontrarioaloquesepodríaesperar.
Metilación Hidroxmetilación
A. Sexo B. Sexo
C. Alcohol D. Alcohol

72
E. Tabaco F. Tabaco
Figura30.DiferenciasenlametilaciónehidroximetilaciónglobaldelDNA.AyB)sexo,CyD)consumodealcoholyEyF)consumodetabaco.
Deacuerdoalosestudiosdealcoholytabaco,decidimoseliminarlosdatosdeindividuosqueconsumíanalcoholytabacoyserealizóunanálisisparaevaluarlosporcentajesdemetilaciónehidroximetilaciónqueposiblemente estuvieran afectando los resultados. No se observó una diferencia significativa en lametilación e hidroximetilación al comparar los pacientes y controles (p=0.6324 y p=0.09568). Sinembargo, seobsevóunadiferencia significativaen los tresgruposde tratamientospor separadopara lametilación(p=0.0048)yparalahidroximetilación(p=0.0056).Igualmentealagruparelgrupocontrolconlos pacientes controlados, se obtuvieron diferencias significativas para metilación (p=0.01137) ehidroximetilación(0.001939)(Figura31).
A. Metilación B. Hidroximetilación

73
C. D.
E. F.
Figura31.Porcentajede lametilaciónenhidroximetilaciónglobaldelDNA, excluyendodatosdeconsumidores de alcohol y tabaco. A) Metilación global en controles sanos vs. pacientes, B)Hidroximetilación global en controles sanos vs pacientes, C) Metilación global en controles sanos,pacientes controladosypacientesno controlados,D)Hidroximetilaciónentre controles sanos,pacientescontroladosypacientesnocontrolados,E)Metilaciónglobalencontrolessanoypacientescontroladosvspacientesnocontrolados,F)Hidroximetilaciónentrecontrolessanosypacientescontroladosvspacientesnocontrolados.*=p<0.05,**=p<0.01,***=p<0.001.
*** ***
***
***
*** ***

74
8.3EDADYDURACIÓNDELAENFERMEDAD
Debido a que la diabetes es una enfermedad crónica que empeora con el tiempo, es muy interesanteconocer como se correlacionan los cambios epigenéticos estudiados con el tiempo de duración de laenfermedadyconlaedaddelospacientes.Sesabequeelenvejecimientoseasociaacambiosdelmetilomaen los individuos (Heyn, 2013) y se espera que en pacientes diabéticos no sea la excepción. Para esteanálisis(Figura32)serealizóunacorrelacióndeSpearmanbuscandodeterminarsiexistíaunatendeciaenla metilación e hidroximetilación de DNA, según la edad y según el tiempo de diagnóstico de laenfermedad.Noobstante,enesteestudionoseencontróunacorrelaciónlinealentrelametilacióndelDNAy la edad (r= 0.07665, p=0.07592). En cuanto a la hidroximetilación, existe una tendecia, aunque no esestadísticamentesignificativa(r=0.1065,p=0.06228).Asuvez,noseobservóunacorrelaciónlinealentreladuraciónde ladiabetesy losnivelesdemetilaciónehidoximetilacióndelDNA(para5mCr=0.6131yp=0.5284;para5hmCr=0.3019y0.3827).
A. B.
C. D.
Figura32.Correlacióndelosporcentajesdemetilaciónehidroximetilaciónentrelaedad(AyB)ytiempodeldiagnóstico(CyD).

75
8.4qRT-PCRDELASENZIMASINVOLUCRADASENLADESMETILACIÓNDELDNA
ParaestudiarlaposiblevíaalteradasenlametilacióndelDNAensangreperiférica,serealizóunensayodeqRT-PCRparaasídeterminarlacuantificaciónrelativadelosfragmentosdeDNMT1yTET1.Elanálisisdeexpresióndeambasfragmentosserealizóporseparadoparacadagen.LaexpresióndeDNMT1(p=0.9506)yTET1(P=0.5758)seanalizóencontrolesypacientespormediodelapruebadeWilcoxysedeterminóquenoexistendiferenciasenlaexpresiónsegúnelcontrolglucémico.AlanalizarlaexpresióndeDNMT1delostresgruposporseparado,controles,pacientescontroladosypacientesnocontrolados,conlapruebadeWelch, tampoco se encontraron diferencias significativas en la expresión paraDNMT1 (p=0.9756) yTET1(p=0.8047).
A. B.
C. D.
Figura33.ExpresióndegenesDNMT1yTET1entrepacientesycontroles.A)ExpresióndeDNMT1enel grupo control sano ypacientes.B)ExpresióndeDNMT1 en controles sanos, pacientes controlados ypacientes no controlados. C) Expresión deTET1 en el grupo control sano y pacientes.D) Expresión deTET1encontrolessanos,pacientescontroladosypacientesnocontrolados.

76
8.5ANÁLISISINSILICODELOSNIVELESDEMETILACIÓNENTEJIDOSAFECTADOSPORLADIABETES
Teniendoencuentaresultadoslimitadosqueofrecetrabajarconsangreperiféricaenestaenfermedad,sedecidió llevar a cabo un estudio in silico con datos de metilación y expresión de microarreglos dediferentes tejidos afectados por la enfermedad. Para ello se analizaron un total de 416 microarreglosextraídosdelrepositorioGeneExpressionOmnibus(GEO)delNCBI(EdgarR,DomrachevM,LashAE.GeneExpression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res2002;30:207–10.).
Delosmicroarreglosutilizadosparaanalizarlosnivelesdemetilación,seestudiaronmuestrasdesangreperiféricadepacientes sin complicaciones,pacientes connefropatíasdiabéticasyungrupodecontrolessanos.TomandolosdatosdelamedidadeMFI(IntensidadMediadeFluorescencia)serealizóunamatrizde agrupamiento según individuos y genes. Para la clusterización de los datos se utilizó el programaCluster 3.0 que forma grupos según los niveles de similitud. En este caso, se realizó un análisis deagrupamientojerárquicoysevisualizaronlosresultadosenelprogramaTreeView.
Losresultadosobtenidosdelaclusterizaciónreflejanunagrupamientoclaroentrepacientesycontroles.(Figura35A).Enelanálisisestadísticodesangreperiféricaseobservógenesdiferencialmentemetiladosentre los pacientes y el grupo control. Al analizar la metilación global de estos grupos se obtiene unresultado idéntico al encontrado en los experimentosprevios, donde existe un aumento significativodemetilaciónenpacientesfrentealoscontroles(p=0.0000)(Figura34A).
Delmismomodo se analizaronmicroarreglos demetilación de otros tejidos de gran importancia en ladiabetes,concretamentetejidoadiposo,tejidomusculareislotesdelpáncreas,cadaunodeloscualesseveafectado de forma diferente. Mientras que en el caso del tejido adiposo y el tejido muscular no hubodiferencias significativas entre pacientes y controles, el páncreas mostró una hipometilación global enpacientes diabéticos frente al grupo control (p=0.0007), tal y como ha sido reportado en la literatura(Avrahami&Kaestner,2012)(Figura34C,DyE).
Figura34.AnálisisdemicroarreglosdesangreperiféricaenpacientesT2DMyungrupocontrol.A)Mapadecalordemuestrasdesangreperiféricadepacientessincomplicaciones,pacientesconnefropatíasy controles sanos procedentes de 317 microarreglos desmetilación (Illumina Human Methylation 27BeadChips). La escala semuestra en valores beta (relación de la intensidad de la sonda y la intensidad

77
global; la intensidadglobales lasumadeintensidaddelaondaynometilados;vadesde0(nometilado,verde)a1(completamentemetilado,rojo).B,C,DyE)ComparacióndelosnivelesdemetilaciónglobalenunidadesIMF(Intensidaddefluorescenciamedia)entrelospacientesconT2DMycontroles.Estoentejidodesangreperiférica,páncreas,tejidoadiposoymuscularrespectivamente.*=Valordep<0,05,**=valordep<0,01,***=valordep<0,001.
8.6HIDROXIMETILACIÓNYDIABETES:POSIBLESMECANISMOSDEDESMETILACIÓN
Considerandolosresultadosprevios,sequisoconocerquéenzimasovíaspodríanestarinterviniendoenlasalteracionesepigenéticasencontradas.ParaelloseanalizaronmicroarreglosdepacientesdeT2DMycontrolessanosensangreperiférica, islotespancreáticos, tejidoadiposoymúsculo.SeescogieronvariosgenesdeinterésinvolucradosenlasvíasdedesmetilacióndelDNAactualmentepropuestas,yseestudiósisus cambios de expresiónpodrían explicar los cambios en los niveles demetilación e hidroximetilaciónencontradospreviamente.Entre losgenesanalizadosestuvieron lasenzimasDNMT1,DNMT2,DNMT3A,DNMT3BYUHRF1porsupapelenlavíadedesmetilaciónpasiva,juntoalreguladornegativodelaDNMT1,miRNA-133a; las enzimas TET1, TET2 y TET3, por su papel en la hidroxilación de la 5mC, actualmenteconsiderada la vía de desmetilación activa primordial; y por último, las enzimas de reparación AID,APOBEC1, APOBEC2, APOBEC3A, APOBEC3B, APOBEC3D, APOBEC4, por su función final en ladesmetilaciónactivaaleliminarlas5hmCydarlugaraunDNAdesmetilado.
Elanálisisestadísticoparacompararlosnivelesdeexpresióndiferencialdelosgenescandidatosensangreperiféricadeterminóquenoexistendiferenciasquepuedanexplicarelaumentode lametilaciónyenelaumentodehidroximetilaciónentre lospacientesT2DMconglucemiano controlada, loque resulta aúnmásimportante,yaquenohaydiferenciasenlosnivelesdelaenzimaTET1,únicadelasTETsquepudoanalizarseenlosmicroarreglosdesangreperiférica(Figura35E).
Al analizar la expresión de las DNMTs, se encontraron diferencias en la expresión de DNMT2 entrepacientes T2DM y el grupo control sano (p = 0,031, Figura 35B). Sin embargo, teniendo en cuenta lafuncióndeestaenzima,queestámásasociadaalametilacióndelRNAynoaladelDNAyqueladiferenciaes por la disminución de la expresión, no parece ser que dicha enzima esté influenciando los cambiosencontradosen ladiabetes.El análisisdel reguladornegativode laDNMT1,miR-133, no reveló cambiossignificativosensuexpresión.
EncuantoalosgenesinvolucradosenlareparacióndelDNA,seencontrarondiferenciassignificativasenlosgenesAPOBEC3ByAPOBEC3D(p=0.017yp=0.042,respectivamente)(Figuras35Ky35L).Elaumentode APOBEC3B en el grupo control sano y de APOBEC3D en pacientes diabéticos tampoco explica loscambiosenlametilaciónyenunaposiblevíadedesmetilación.

78
Figura35.AnálisisdelaexpresióndiferencialdegenesensangreperiféricadepacientesT2DMyelgrupocontrol.A)DNMT1,B)DNMT2,C)DNMT3A,D)DNMT3B,E)TET1,F)UHRF1,G)AID,H)APOBEC1,I)APOBEC2, J) APOBEC3A, K) APOBEC3B, L) APOBEC3D, M) APOBEC4, N) miR-133a. *=p-value<0.05, **=p-value<0.01,***=p-value<0.001.
Posteriormente,seevaluólaexpresióndiferencialdegenesrelacionadosconladesmetilacióndelDNAentejido pancreático (Figura 36). Para la desmetilación pasiva se evaluaron DNMT1, DNMT2, DNMT3B yUHRF1;paraladesmetilaciónactivaseevaluaronlosgenesTET1,TET2,TET3yfinalmente,losgenesAID,APOBEC1, APOBEC2, APOBEC3A, APOBEC3C, APOBEC3D, APOBEC3F, APOBEC3G, APOBEC3H y CD,involucradosenlareparación.
AlrealizarlosanálisisestadísticosdelasenzimasinvolucradasenlametilacióndelDNA,noseobservarondiferencias significativas en los niveles de expresión de la DNMT1 que pudieran explicar un posiblemecanismopasivodedesmetilaciónen lospacientesdiabéticos.Enelcasode ladesmetilaciónactiva,seobservóunadisminuciónenlaexpresióndeTET2(p=0.031)enpacientesdiabéticos,locualseríacontrarioalprocesoobservadodedesmetilaciónysiendoinsuficienteparaexplicarlo.
Unode losdatosmás interesantesduranteel estudiodeeste tejido fue comprobar cómo la enzimaAIDaparece sobreexpresada en pacientes frente a controles (p=0.017), algo que no sucede en el caso de lasangre periférica y que podría explicar por qué este tejido, en concreto, puede llegar a desmetilar lasmetilcitosinaspreviamenteoxidadasporlosaltosnivelesdeROS.
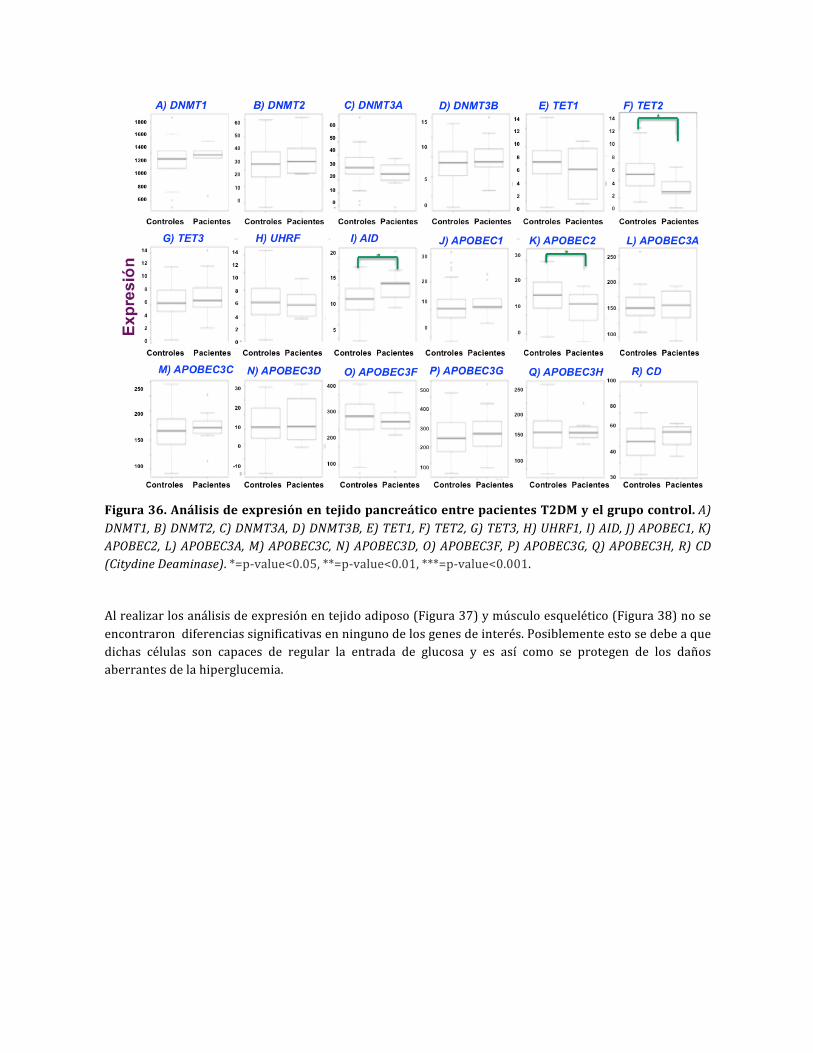
79
Figura36.AnálisisdeexpresiónentejidopancreáticoentrepacientesT2DMyelgrupocontrol.A)DNMT1,B)DNMT2,C)DNMT3A,D)DNMT3B,E)TET1,F)TET2,G)TET3,H)UHRF1,I)AID,J)APOBEC1,K)APOBEC2,L)APOBEC3A,M)APOBEC3C,N)APOBEC3D,O)APOBEC3F,P)APOBEC3G,Q)APOBEC3H,R)CD(CitydineDeaminase).*=p-value<0.05,**=p-value<0.01,***=p-value<0.001.
Alrealizarlosanálisisdeexpresiónentejidoadiposo(Figura37)ymúsculoesquelético(Figura38)noseencontrarondiferenciassignificativasenningunodelosgenesdeinterés.Posiblementeestosedebeaquedichas células son capaces de regular la entrada de glucosa y es así como se protegen de los dañosaberrantesdelahiperglucemia.

80
Figura37.AnálisisdeexpresiónentejidoadiposoentrepacientesT2DMyelgrupocontrol.A)AID,B)APOBEC1,C)APOBEC2,D)APOBEC3A,E)APOBEC3B,F)APOBEC33,G)APOBEC3D,H)APOBEC3F-3G, I)APOBEC3G, J) APOBEC4C, K) DNMT1, L) DNMT3A,M) DNMT3B, N)MBD4, O) TDG, P) TET1, Q) TET2, R)TET3,S)UHRF1.*=p-value<0.05,**=p-value<0.01,***=p-value<0.001.

81
Figura38.AnálisisdeexpresiónentejidomuscularentrepacientesT2DMyelgrupocontrol.A)AID,B)APOBEC1,C)APOBEC2,D)APOBEC3A,E)APOBEC3B,F)APOBEC33,G)APOBEC3D,H)APOBEC3F-3G, I)APOBEC3G, J) APOBEC4C, K) DNMT1, L) DNMT3A,M) DNMT3B, N)MBD4, O) TDG, P) TET1, Q) TET2, R)TET3,S)UHRF1.*=p-value<0.05,**=p-value<0.01,***=p-value<0.001.
8.7GENESDIFERENCIALMENTEEXPRESADOS
UtilizandoelprogramaChispterseanalizólaexpresióndiferencialdegenesenlosmicroarreglosdetejidopancreáticoysangreperiférica.Seescogieronaproximadamente50genesyutilizandolasbasesdedatosdeNCBI(NationalCenterforBiotechnologyInformation)yEnsembl,sebuscaronlasrutasmetabólicasenlaqueestáinvolucradocadagendiferencialmenteexpresado.Interesantemente,paraeltejidopancreáticoseencontraronalteradaslasvíasasociadasalmetabolismodelipidos.Porotraparte,ensangreperiféricalasvíasmásafectadassonlasdedivisióncelular,proteínasribosomalesyubiquitinligasas(Tabla25).
Tabla25.Rutasafectadasentejidopancreáticoysangreperiférica
Genesconexpresióndisminuida
Páncreas
-GTPasas-Reguladoresde
GTPasas-TirosinKinasa
-Proteínasdeuniónytransportede
metales
Sangre
-Divisióncelular-Reguladoresdetranscripción
-Transportadoresdesolutos
Genesconexpresiónaumentada
Páncreas
-Pepsinogenos3,4y5
-Moléculasdeadhesióncelularrelacionadasaantígeno
carcinoembriogénico6y7
-ApolipoproteínaD-Acil-coAsintasa
Sangre
-Transportadoresdesolutos
-Ubiquitinligasas-Divisióncelular
-Glutatiónperoxidasa-Proteínasribosomales

82
9.DISCUSIÓN
9.1EFECTODELCONTROLMETABÓLICOENLAMETILACIÓNEHIDROXIMETILACIÓNDELDNAENT2DM
Loscambiosepigenéticosasociadosaenfermedadescomplejasestánsiendoampliamenteestudiadostrasla confirmación de la importancia que su regulación tiene para la salud celular y fisiológica.Muchos deestas enfermedades, como la diabetes, el cáncer y enfermedades autoinmunes, inflamatorias yneurológicas,sehanrelacionadocondefectosdedesmetilaciónglobaldelDNA,locual,alalarga,conducea consecuenciasdesfavorablespara la célula (Cooper&El-Osta, 2010;Rodríguez-Dorantes et al., 2004).DurantemuchotiemponohuboclaridadsobrelosmecanismosdeladesmetilacióndelDNAyhastahacepoco solo se aceptaba la desmetilación pasiva relacionada con la pérdida del grupo metilo tras lareplicacióndelDNA.En2009sepropusounanuevavíaactiva lacual involucraenzimasquerealizanunprimerpasoenlaoxidacióndelmetilo(Brancoetal.,2011;Raiberetal.,2012).Graciasalaportedeestosdos estudios, en la actualidad se cuenta con ladescripcióndedosvías, activaypasiva. Sin embargo,noexiste explicación sobre cómo actúan estos mecanismos, debido a que las vías activas aún no se hancaracterizado bajo condiciones patológicas, por ejemplo, en la diabetes. En el presente estudio seanalizaron los cambios de los niveles demetilación del DNA en pacientes diabéticos frente a controlessanosconelfindeencontrarunaexplicaciónmecanísticaadichoproceso.
El estudio de los niveles globales de metilación e hidroximetilación asociados a los niveles de controlmetabólicomedidosatravésdelosporcentajesdeHb1Ac,hanpermitidodemostrarquenoesladiabetesper se, la causante de los cambios de metilación global. En el caso concreto de sangre periférica,encontramos una tendencia de aumento en la metilación e hidroximetilación global del DNA bajocondicionespatológicas.Unosde loshallazgosmássorprendentes fue laausenciadediferenciasentre lametilaciónylahidroximetilaciónentreelgrupocontrolsanoylospacientescontrolados.Estosugiereunarelación directa con la hiperglucemia crónica y persistente que generan cambios acumulativos en elepigenoma.Estohace referencia enparticular a resultadosde los estudiosde laDCCT (DiabetesControlandComplicationsTrial)yelestudiodelaEDIC(EpidemiologyofDiabetesInterventionsandComplications)(verANEXOS;13.2.1y13.2.3), loscualesdemuestranlaimportanciadelcontrolglucémicoparaprevenirlas complicaciones a largo plazo y paramejorar la calidad de vida de los pacientes. Específicamente, seencontró que los pacientes con un descontrol metabólico tendrán dramáticas repercusiones en elpronósticodelaenfermedad,provocandoladisminuciónenlacalidaddevidahastalamuerte.Elestudioprogresivo San Antonio Heart Study , aún en marcha, “San Antonio Heart Study”, demostró que unincremento de la HbA1c de 5,5% a 9,5% produce patologías microvasculares importantes (Brownlee,2005) y, por tanto, el buen control glucémico tendrá un efecto epigenético protector. Por otro lado, loscambios epigenéticos asociados al descontrolmetabólico explican su rol de lamemoriametabólica y suinfluenciaeneldesarrollodelascomplicacionesdiabéticas(Ceriello,2008;Intine&Sarras,2012).
Duranteelanálisisdenuestrosresultados,observamosunamayormetilaciónencondicionesdediabetes,locualresultacontradictorioparalamayoríadeestudiosepigenéticos,enloscualesexisteunprocesodedesmetilacióndelDNAdebidoa losaltosnivelesdeglucosaen la sangre (Giacco&Brownlee,2010;ASOlsen,SarrasJr.,&Intine,2010;AnsgarS.Olsen,Sarras,Leontovich,&Intine,2012).Enestecaso,podemosestablecerquelametilacióndelDNAesafectadaporlosestadosmetabólicosmásqueporladiabetescomotal.Sinembargo,diversosestudioshandemostradoquelospatronesdemetilaciónsontejido-específicosycada uno reacciona demanera distinta según diferentes estímulos;mientras que en tejido endotelial lahipometilaciónparecedominarenladiabetes,entejidosanguíneoresultadiferente(Heynetal.,2013;ASOlsenetal.,2010;AnsgarS.Olsenetal.,2012).
Debido asu efectosobre el estrés oxidativo, la disponibilidad de glucosa intracelular puede influir en lametilaciónporlavíadedesmetilación.Enesteestudioproponemosquealtosnivelesde5hmCensangreseestán dando por la oxidación celular generada por la hiperglucemia. Si bien, la falta de desmetilación

83
habríaqueexplicarlaporlafaltadeexpresiónenlasenzimasdereparaciónimplicadasenelprocesofinaldeeliminacióndemetilosdesdesusformasoxidadas.
Otraposiblerazónquepuedeexplicarlafaltadedesmetilaciónensangreesladinámicapropiadeltejido,quecuentaconunaltonivelderecambiodecélulasblancasencomparaciónconotrostejidosyquepuedenestar enmascarando los resultados, aunque para demostrar esta conjetura se requiere nuevos estudios.Adicionalmente, debemos tener en cuenta que el tejido sanguíneo está conformado por distintassubpoblacionesdecélulasquecontienendiferentesperfilesdemetilaciónydinámicasrespectoalusodelaglucosayportantoseríamuyinteresantepoderllevaracabounestudiopormenorizadodeloscambiosencada una de las estirpes celulares. La heterogeneidad del pool celular entre individuos puede tener unimpactoenlosestudiosepigenéticos.Elperiododevidadeestascélulasesdedías,mientrasqueenotrostejidoscomoelpáncreas,puedeserdeaños(Volkmar,Dedeurwaerder,Cunha,Ndlovu,Defrance,Deplus,Calonne, Volkmar, Igoillo-esteve, et al., 2012). Lametilación global también puede ser alterada por losmismosprocesosinmunológicoseinflamatoriospropiosdeladiabetes,afectandolosprocesoepigenéticosdel pool. A esto se le suman las condiciones medioambientales como el consumo de diferentesmedicamentosparaladiabetes,lainformaciónnoproporcionadaporlospacientesycontroles,losmaloshábitos de vida tales como el consumo de alcohol, tabaco y los excesos de la alimentación. Este últimohábito genera picos glucémicos transitorios, los cuales afectan los patrones de metilación de manerapermanenteygeneranlamemoriametabólica(El-Ostaetal.,2008;Ling&Groop,2009).
ElconsumodealcoholsehaasociadoaniveleselevadosdehomocisteinaquepuedencontribuirabajosnivelesdeSAMybajacapacidaddemetilación.Estosedebeaqueelalcoholafectaavariosintermediariosdel metabolismo del ciclo de un carbono, al consumir los cofactores B12 y B6 importantes en latransferenciadeesecarbonoaSAM.(Bottiglierietal.,2000;Iskandaretal.,2010;Nieratschker,Batra,&Fallgatter, 2013). Pacientes con alcoholismo crónico se han asociado con hipermetilaciones en sitios derestricciónespecíficosparalasenzimasHspIIyMspI.EstoconduceaunadisminucióndelaexpresióndelaDNMT3a y DNMT3b que finalmente conlleva a la hipometilación. A pesar de los diversos estudios delconsumodealcoholytabacoqueafectanlospatronesdemetilacióncelular,enesteestudionosepuedoestablecersuefectoenlospacientesdiabéticosensangreperiférica,sibien,laeliminacióndelasmuestrasconsumidoraspermitióobtenerdatosmuchomásrobustosenlascomparaciones,demostrandodeformaindirectaquesiestáninfluyendoenlasalteracionesepigenéticasencontradas.
9.2EXPRESIÓNDEGENESINVOLUCRADOSENRUTASDEDESMETILACIÓN
Al realizar el estudio de qRT-PCR para los genes de las enzimas DNMT1 y TET1 no encontramosdiferenciasenlaexpresióndeestosdosimportantesgenesimplicadosenladesmetilaciónpasivaoactiva.Elmecanismo clásico de desmetilación está asociado a cambios epigenéticos en la desmetilación pasivacaracterizada por la acción de la DNMT1. diferentes estudios han propuesto que existe otra víacaracterizada por la oxidación enzimática de los gruposmetilos por la familia TETs y la desaminaciónposterior de ellos (Branco et al., 2012). Como pudimos observar en nuestro estudio, las vías dedesmetilaciónactivaopasiva,aniveldeestesdosenzimas,noestánalteradasencasodeunpobrecontrolglucémico.Seobservóquelahidroximetilaciónestáaumentadaenpacientesconglucemianocontroladayque laTET1noestásobreexpresadabajo lamismacondición.Nohubodiferenciassignificativasentreelgrupocontrolylospacientes.
Añosatrás,unimportanteestudiorealizadoentejidomusculardescribióunmecanismodeprotecciónporpartedeltejidoylaentradaexcesivadeglucosa.Sehavistoqueeltejidonogeneraresistenciaalainsulinaaúnbajoaltosnivelesdeglucosaensangredebidoaunamenorexpresióndeltransportadordeglucosa4(GLUT4)encélulasnoafectadasporladiabetes(Dimitrakoudis,Ramlal,Rastogi,Vranic,&Klip,1992;Klipetal.,1992;Maretteetal.,1999).Estetransportadorescríticoparapermitirlaentradadeglucosa;células

84
comolasendoteliales,deretina,deShawn,neuronasycélulasdeglomérulo,quenosoncapacesderegularlaexpresióndeGLUT4,sonafectadasencasodeestaenfermedad.Permitirelexcesodeentradadeglucosaenlacélulageneraaltosnivelesdeoxidación,deestamaneraseafectanlasvíasbioquímicasqueconllevana las complicaciones (Brownlee, 2005). Por todo ello, postulamos la existencia de una nueva vía dedesmetilaciónalternaa lasyadescritas;unavíaqueoxidadirectamentea losgruposmetilosenelDNAdebidoaladesregulaciónenlosestadosdeoxidacióncelularporlaentradaexcesivadeglucosa.
9.3UNPOSIBLEMECANISMONUEVODEDESMETILACIÓNENESTADOSDEENFERMEDAD
Paracompararlosresultadosmolecularesobtenidospreviamente,serealizóunestudiodemicroarreglosinsilicoconelobjetodedemostrarquelametilaciónestejido-específicoyquelosresultadosobtenidosensangre concuerdan con los de losmicroarreglos. El análisis demicroarreglos demetilación para sangreperiféricadepacientes,pacientesconnefropatíasycontroles,confirmóquelasangreesuntejidoenelquehayunahipermetilacióndelDNA,frentealoqueseesperaba,apriori,endiabetes.Además,losresultadosde clusterizaciónmuestranque sin tener en cuenta el estadode controlmetabólico, dato con el quenocontamos para los microarreglos, no existen diferencias en la gravedad de la enfermedad a nivelepigenético.Portanto,lapresenciadecomplicacionesylosnivelesaltosdemetilaciónsonindependientes.Soloelbuencontrolglucémicopodríarevertirestecambioyposiblemente,detenereldeterioroprovocadoporlaenfermedad.
Adiferenciadelasangre,eltejidopancreáticosípresentahipometilaciónglobaldelDNA.Conelresultadodequeambostejidosrespondenaaltosnivelesdeglucosacirculante,elinterrogantequenosquedabaporresolver era conocer qué mecanismo subyace al proceso degenerativo que provoca el aumento dehidroximetilcitosinasensangre,ymuyposiblementeenelpáncreas,yporqué,apesardeeseaumento,lasangre no termina por desmetilarse, a diferencia de lo que ocurre en tejido pancreático. Como era deesperar, no encontramos diferencias en la metilación global de tejido adiposo y muscular, ya que sontejidosquecontrolanmucholaentredadeglucosayportantonoseencuentranbajoelefectooxidativodelahiperglucemia.
A travésdemicroarreglosdeexpresiónseanalizaronenzimas involucradasen lasvíasdedesmetilacióntantoactivacomopasivaensangreperiféricayentejidopancreático,enelcualsedeterminóquenohaydiferenciasenlaexpresióndeenzimasquepudieranexplicarloscambiosenlasmodificaciones.Enelcasodeltejidosanguíneo,noseencontróalteraciónenlaexpresióndeDNMT1yUHRF1,quepudieranexplicarel aumento en lametilación en el caso de pacientes.; tampoco se observó la presencia significativa delreguladornegativodelaDNMT1,elmiRNA133a.SeobservóunaumentosignificativodelaDNMT2enloscontroles, pero esto no explica la metilación en pacientes y tampoco es relevante ya que se relacionaprincipalmente con la metilación del RNA. Las enzimas TETs tampoco explican el aumento dehidroximetilosenlospacientes,motivoporelcualnuevamentenospreguntamoscómoestáocurriendolaoxidación. En el caso del tejido pancreático, no se encontraron diferencias en estas mismas enzimas,excepto en TET2 que está aumentada en los controles. Esto resulta contradictorio frente a losmismosresultadosdequeenelpáncreasexisteunahipometilaciónglobal,encasodediabetes.
La falta de cambios entre pacientes y controles en ambos tejidos parecen indicar que los procesos deoxidaciónahidroximetilcotosinasquesedanensangreyladesmetilacióncomprobadaenelpáncreas,nopuedenserdebidosalasvíasactualmenteestablecidas.Solamentepuedenexplicarelprocesolosnivelesaumentados de oxidación demostrados en diabetes, asociados a los altos niveles de hemoglobinaglicosiladademostradosenpacientesmalcontrolados.Deestemodo,proponemosunprimerprocesodeoxidación no enzimático de las 5-metilcitosinas que daría lugar a un aumento de 5hmC, tal como seencuentraensangreperiférica.

85
Adiferenciadeloqueocurreenelpáncreas,elhechodequelasangrenoterminepordemetilarsepuedeserdebido,comoseexplicóanteriormente,alapropiadinámicadeestetejido.Sinembargo,losdatosdelosmicroarreglosdeexpresiónpermitenintuirqueelmantenimientodelametilaciónensangrepuedeserayudadoporlafaltademaquinariaquereparelas5hmCacitosinasconvencionales.Entreestamaquinaria,en losúltimosaños,sehapropuestoun importantepapela lasenzimasAPOBECsyaAID.Laspequeñasdiferencias de APOBEC3B y APOBEC3D encontradas en sangre no pueden explicar lo resultados. LaAPOBEC3Bdisminuidaenpacientespodríaexplicarlafaltadedesmetilaciónencasodediabetes,quenotermina por desmetilarse, aunque se necesitan más estudios para corroborar la vía. Por otro lado, laAPOBEC3D no explica el aumento demetilación en pacientes y, por consecuencia, no es la causa de ladesregulaciónepigenética.
EldatoclavedenuestroestudiofueencontrarunaumentodeAIDeneltejidopancreáticodepacientes,yaqueestoindicaunaposiblevíadedesmetilaciónqueseasociaconlosaltosnivelesde5hmCqueterminaen la desmetilación global del genoma. La sobreexpresión de esta enzima en el páncreas podría ser larazón del aumento de 5hmC provocado por la oxidación celular generada por la glucosa, en la queterminanconvertidasencitosinasconvencionalesy,dandolugaraungenomadesmetilado.Porsuparte,lafalta omenor expresión deAID en sangre, restringida seguramente a su función sobre los genes de lasinmunoglobulinas y TCRs de los linfocitos, podría explicar por qué en sangre periférica semantiene lametilación,apesardelosaltosnivelesdehidroximeticitosinasexistentesenpacientesdiabéticos.Ademásdel papel principal de AID, se requiere un estudio de los mecanismos antioxidantes, como los de lasenzimasSODsyGSTsylosdelasenzimasencargadasenlareparaciónBER,yaquellasquedependendelaTDGyMBD4,cuyopapeltambiénhasidodescritocomocríticoparaelprocesodedesmetilación.
Enesteestudioproponemosunmecanismodedesmetilacióndistintoaloquesehadescritohastaahora.Un mecanismo que explica la hipometilación en la diabetes y en otras condiciones patológicas,independiente de enzimas, hoy en día caracterizadas en catalizar la primera reacción de oxidación del5mC, concretamente independiente de TETs. Este nuevo mecanismo de desmetilación se basa en ladesregulacióndelosestadosredoxcelulardelacélulaqueproponeunaoxidacióndirectaporpartedeROSalosgruposmetilosdelDNA(Figura40).EnlaactualidadsehadescritolamaneracómoafectalosestadosdeoxidaciónalDNA,yporquénoestamismavíapodríaserlacausantedelaoxidacióndelosmetilos.Deeste modo, el ambiente oxidante típico de la diabetes y de los procesos proinflamatorios, tambiénpresentesenenfermedadescomplejascomoel cánceryciertasenfermedadesautoinmunes,provocaríanesta primera oxidación de los 5-metilos a 5-hidroxilos. Posteriormente estas 5hmC se repararían pormediodeAIDoporTDGparaterminardandoungenomahipometilado.
Esevidentequeelmecanismooxidativopuedeserlacausademuchasdelasalteracionesepigenéticasenla diabetes. A la vez, es necesario tener en cuenta la predisposición genética de cada individuo, lostrastornos genéticos y bioquímicos de la maquinaria enzimática involucrada en este metabolismo, lascondicionesmedioambientalesyloshábitosdevida,comoelconsumodesustanciasalucinógenas,tabaco,medicamentos, alimentación y ejercicio. Todos estos factores afectan el epigenoma y la forma como seexpresanmuchosgenesinvolucradosenladiabetes.

86
Figura 39: Representación esquemática de las posibles vías de desmetilación del DNA.1) Vía dedesmetilación pasiva. Deficiencia de la función de DNMT1 causa falta en el mantenimiento de lasmetilacionesduranteladivisióncelular.2)Desmetilaciónenzimáticaactiva.LasTETsproducen5hmC,queluego es desaminada a 5hmC por el complejo AID o APOBEC. Esta base es retirada por el sistema dereparación por escisión de base deDNA (BER), por la glicosilasa de timina o porMDB4, la proteína deuniónamCpGs.3)Desmetilaciónactivanoenzimática.Víanuevapropuesta,unaumentoenROSyAGEsgeneraoxidacióndirectayespontáneadel5mC,generando5hmCysuposterioreliminación.
9.4 GENES DIFERENCIALMENTE EXPRESADOS Y SU EFECTO SOBRE LA PROGRESIÓN DE LAENFERMEDAD.
Por último, teniendo en cuenta los genes diferencialmente expresados bajo condiciones patológicas,pudimosencontrar lasposiblesvíasmetabólicasdesreguladasen los tejidospancreáticoy sanguíneo.Alanalizar los genes pobremente expresados del páncreas, pudimos determinar que se alteraron vías seseñalización como GTPasas y sus reguladores, tirosin-kinasas, proteínas de unión y de transporte demetalesysesobrerregularongenescomopepsinogeno3,4y5,moléculasdeadhesiónrelacionadaconelantígenocarcinoembriogénico,Acil-coAsintasayapolipoprotienaD.Estasdesregulación se correlacionacon vías que conllevan a la tumorogénesis y, en consecuencia, la alta susceptibilidad que tienen lospacientesapadecercáncer.
En el casode sangre, encontrados genespobremente regulados como genes involucrados en la divisióncelular, genes de reguladores de transcripción y genes de transportadores de soluto. Tambiénencontramosgenesaltamenteexpresados,asociadosaladegradacióndeproteínas,enzimasantioxidantesygenesdedetoxificación,como laglutatiónperoxidasa.Estohaceénfasisen lasobrerregulacióndeestaproteína en diabéticos, con el fin de neutralizar los radicales libres, indicándonos que, a pesar de noencontrar diferencias en patrones de metilación global en nuestro estudio, el tejido sanguíneo sí estásiendo afectado por los estados hiperglucémicos. También se encuentran desregulaciones en proteínasencargasdetrasporte,degradaciónydivisióncelularquepuedenllegararelacionarseconotraspatologíascomolasleucemias.

87
Porultimo, este estudio enpacientesdiabéticos esunprimerpasopara conocer y caracterizar las víasinvolucradas en la generación de sus complicaciones y la manera como es afectada por memoriametabólica.Estaultima,puedeserexplicadacomounefectodeladesregulaciónepigenéticayporlotanto,la caracterización epigenética de la patología puede tener beneficios sobre el tratamiento para lospacientescondiabetes.Todoslosresultadosobtenidossecorrespondenconlahipótesisdepartidayporelloapoyanlaideadelaexistenciadeunanuevavíadedesmetilacióniniciadaencondicionespatológicasymediada por los altos niveles de estrés oxidativo. Una nueva vía que prescinde de la familia TET,actualmente consideradas imprescindibles para el proceso de desmetilación activa, y que explicaría losprocesosdehipometilación encontrados enmuchas enfermedades complejas a través deunprocesodeoxidaciónespontaneadelas5mCa5hmCporefectodelosnivelesdeROS.

88
X.CONCLUSIONES
1. Losnivelesde5mCy5hmCensangresonafectadosporelcontrolglucémicoynoporlaDiabetesperse.
– Lametilaciónensangreesmayorenpacientesnocontroladosyestosecorroboróconelestudiodemicroarreglosinsilico.
2. La maquinaria descrita hoy en día no explica el proceso metilación del DNA bajo condicionespatológicas.
– Laexpresiónenlosnivelesdelasenzimasnoexplicanloscambiosobtenidosensangre.
3. Elanálisisensangredeexpresióndiferencial insilico tampocodemostrógenesdiferencialmenteexpresadosencasodediabetes,quepudieranexplicarladesregulacióndeestapatología.
4. Es por esto, se propone una vía alterna de desmetilación, basada un proceso de oxidación noenzimático.
5. Existen varias vías de regulación alteradas en diabetes, tanto en páncreas, como en sangre.Muchosdelosgenesalteradosestánrelacionadosconlascomplicacionesdeladiabetes.

89
XI.RECOMENDACIONES
ü Es importante ampliar el presente estudio con análisis in vitro de tejidos que se afectandiferencialmenteporlahiperglucemiaparacomprobarelefectodelasenzimassobrelosmecanismosdemetilaciónydesmetilación.
ü SeríainteresanterealizarunestudiodeMedChipporsualtaespecificidadysensibilidad,paraevaluarzonasespecíficasquesemetilanodesmetilandurante losestadoshiperglucémicos, loquepermitirácompararlosresultadosconlasvíasmetabólicasafectadas.
ü Para obtener resultados más predictivos, sería de gran utilidad ampliar el número de individuosevaluadosconelfindeeliminarposiblessesgosdeconfusiónenelestudio.

90
XII.BIBLIOGRAFÍA
Aschner,P.(2010).EpidemiologíadeladiabetesenColombia.Av.Diabetol.,26,95–100.http://doi.org/10.1016/S1134-3230(10)62005-4
Atkinson,M.a.,Eisenbarth,G.S.,&Michels,A.W.(2014).Type1diabetes.TheLancet,383(13),69–82.http://doi.org/10.1016/S0140-6736(13)60591-7
Avrahami,D.,&Kaestner,K.H.(2012).Epigeneticregulationofpancreasdevelopmentandfunction.SeminarsinCell&DevelopmentalBiology,23(6),693–700.http://doi.org/10.1016/j.semcdb.2012.06.002
BashirAamir,U.,Badar,N.,Mehmood,M.R.,Nisar,N.,Suleman,R.M.,Shaukat,S.,…Klimov,A.(2012).MolecularepidemiologyofinfluenzaA(H1N1)pdm09virusesfromPakistanin2009-2010.PLoSONE,7(8).http://doi.org/10.1371/journal.pone.0041866
Bhutani,N.,Burns,D.M.,&Blau,H.M.(2011).DNAdemethylationdynamics.Cell,146(6),866–872.http://doi.org/10.1016/j.cell.2011.08.042
Bottiglieri,T.,Laundy,M.,Crellin,R.,Toone,B.K.,Carney,M.W.P.,Reynolds,E.H.,&Hospital,P.(2000).monoaminemetabolismindepression,228–232.
Branco,M.R.,Ficz,G.,&Reik,W.(2011).Uncoveringtheroleof5-hydroxymethylcytosineintheepigenome.NatureReviewsGenetics,13(1),7–13.http://doi.org/10.1038/nrg3080
Brownlee,M.(2005a).Thepathobiologyofdiabeticcomplications.Diabetes,54(June2005),1615.http://doi.org/10.2337/diabetes.54.6.1615
Brownlee,M.(2005b).Thepathobiologyofdiabeticcomplications.Diabetes,54(June),1615.http://doi.org/10.2337/diabetes.54.6.1615
Caudill,M.A.,Wang,J.C.,Melnyk,S.,Pogribny,I.P.,Jernigan,S.,Collins,M.D.,…James,S.J.(2001).BiochemicalandMolecularActionofNutrientsIntracellularS-AdenosylhomocysteineConcentrationsPredictGlobalDNAHypomethylationinTissuesofMethyl-DeficientCystathionine�-SynthaseHeterozygousMice1,(May),2811–2818.
Ceriello,A.(2008a).La“memoriametabólica”inducidaporlahiperglucemia:elnuevoretoenlaprevencióndelaenfermedadcardiovascularenladiabetes.RevEspCardiol,8,12–8.Retrievedfromhttp://www.revespcardiol.org/es/linksolver/ft/id/13119904
Ceriello,A.(2008b).La“memoriametabólica”inducidaporlahiperglucemia:elnuevoretoenlaprevencióndelaenfermedadcardiovascularenladiabetes.RevistaEspañoladeCardiologíaSuplementos,8,12C–18C.http://doi.org/10.1016/S1131-3587(08)73550-7
Chatterjee,R.,&Vinson,C.(2013).NIHPublicAccess,1819(7),763–770.http://doi.org/10.1016/j.bbagrm.2012.02.014.CpG
Chen,C.C.,Wang,K.Y.,&Shen,C.K.J.(2013).DNA5-methylcytosinedemethylationactivitiesofthemammalianDNAmethyltransferases.JournalofBiologicalChemistry,288(13),9084–9091.http://doi.org/10.1074/jbc.M112.445585
Chen,Z.,&Riggs,A.D.(2011).DNAmethylationanddemethylationinmammals.TheJournalofBiologicalChemistry,286(21),18347–53.http://doi.org/10.1074/jbc.R110.205286
Cooper,M.E.,&El-Osta,A.(2010).Epigenetics:Mechanismsandimplicationsfordiabeticcomplications.CirculationResearch,107,1403–1413.http://doi.org/10.1161/CIRCRESAHA.110.223552
Denis,H.,Ndlovu,M.N.,&Fuks,F.(2011).RegulationofmammalianDNAmethyltransferases:aroutetonewmechanisms.EMBOReports,12(7),647–656.http://doi.org/10.1038/embor.2011.110
Dhliwayo,N.,Sarras,M.P.,Luczkowski,E.,Mason,S.M.,&Intine,R.V.(2014).Parpinhibitionpreventsten-eleventranslocaseenzymeactivationandhyperglycemia-inducedDNAdemethylation.Diabetes,63(9),3069–3076.http://doi.org/10.2337/db13-1916
Diabetes,N.,&Clearinghouse,I.(1993).DCCTandEDIC :

91
Dimitrakoudis,D.,Ramlal,T.,Rastogi,S.,Vranic,M.,&Klip,a.(1992).Glycaemiaregulatestheglucosetransporternumberintheplasmamembraneofratskeletalmuscle.TheBiochemicalJournal,284(Pt2,341–8.Retrievedfromhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1132643&tool=pmcentrez&rendertype=abstract
Ding,G.-L.,&Huang,H.-F.(2014).RoleforTetinHyperglycemia-InducedDemethylation:ANovelMechanismofDiabeticMetabolicMemory.Diabetes,63(9),2906–2908.http://doi.org/10.2337/db14-0675
Drzewoski,J.,Kasznicki,J.,&Trojanowski,Z.(2009).Theroleof“metabolicmemory”inthenaturalhistoryofdiabetesmellitus.PolskieArchiwumMedycynyWewnętrznej,119(7-8),493–500.Retrievedfromhttp://eutils.ncbi.nlm.nih.gov/entrez/eutils/elink.fcgi?dbfrom=pubmed&id=19776690&retmode=ref&cmd=prlinks\npapers3://publication/uuid/57906FCA-6EB0-4D6B-A6DC-3722E9385AE5
El-Osta,A.,Brasacchio,D.,Yao,D.,Pocai,A.,Jones,P.L.,Roeder,R.G.,…Brownlee,M.(2008).Transienthighglucosecausespersistentepigeneticchangesandalteredgeneexpressionduringsubsequentnormoglycemia.TheJournalofExperimentalMedicine,205(10),2409–2417.http://doi.org/10.1084/jem.20081188
Felsenfeld,G.(2014).Abriefhistoryofepigenetics.ColdSpringHarborPerspectivesinBiology,6(1).http://doi.org/10.1101/cshperspect.a018200
Forbes,J.M.,&Cooper,M.E.(2013).Mechanismsofdiabeticcomplications,137–188.http://doi.org/10.1152/physrev.00045.2011
Giacco,F.,&Brownlee,M.(2010a).Oxidativestressanddiabeticcomplications.CirculationResearch,107,1058–1070.http://doi.org/10.1161/CIRCRESAHA.110.223545
Giacco,F.,&Brownlee,M.(2010b).Oxidativestressanddiabeticcomplications.CirculationResearch,107(9),1058–1070.http://doi.org/10.1161/CIRCRESAHA.110.223545
Goldberg,A.D.,Allis,C.D.,&Bernstein,E.(2007).Epigenetics:ALandscapeTakesShape.Cell,128(4),635–638.http://doi.org/10.1016/j.cell.2007.02.006
He,Y.-F.,Li,B.-Z.,Li,Z.,Liu,P.,Wang,Y.,Tang,Q.,…Xu,G.-L.(2011).Tet-mediatedformationof5-carboxylcytosineanditsexcisionbyTDGinmammalianDNA.Science(NewYork,N.Y.),333(2011),1303–1307.http://doi.org/10.1126/science.1210944
Heyn,H.,Moran,S.,Hernando-herraez,I.,Sayols,S.,Gomez,A.,Sandoval,J.,…Esteller,M.(2013).DNAmethylationcontributestonaturalhumanvariation,1–11.http://doi.org/10.1101/gr.154187.112.Freely
Holliday,R.(2006).Epigenetics:Ahistoricaloverview.Epigenetics,1(2),76–80.http://doi.org/10.4161/epi.1.2.2762
Horvath,S.(2013).DNAmethylationageofhumantissuesandcelltypes.GenomeBiol,14,R115.http://doi.org/10.1186/gb-2013-14-10-r115
Illingworth,R.S.,&Bird,A.P.(2009).CpGislands-“Aroughguide.”FEBSLetters,583(11),1713–1720.http://doi.org/10.1016/j.febslet.2009.04.012
Intine,R.V.,&Sarras,M.P.(2012).Metabolicmemoryandchronicdiabetescomplications:Potentialroleforepigeneticmechanisms.CurrentDiabetesReports,12,551–559.http://doi.org/10.1007/s11892-012-0302-7
Iskandar,B.J.,Rizk,E.,Meier,B.,Hariharan,N.,Bottiglieri,T.,Finnell,R.H.,…Hogan,K.J.(2010).FolateregulationofaxonalregenerationintherodentcentralnervoussystemthroughDNAmethylation.JournalofClinicalInvestigation,120(5),1603–1616.http://doi.org/10.1172/JCI40000
Jaenisch,R.,&Bird,A.(2003).Epigeneticregulationofgeneexpression:howthegenomeintegratesintrinsicandenvironmentalsignals.NatureGenetics,33Suppl(march),245–254.http://doi.org/10.1038/ng1089
Jin,S.G.,Kadam,S.,&Pfeifer,G.P.(2010).ExaminationofthespecificityofDNAmethylationprofilingtechniquestowards5-methylcytosineand5-hydroxymethylcytosine.NucleicAcidsResearch,38,1–7.

92
http://doi.org/10.1093/nar/gkq223
Jones,P.a.(2012).FunctionsofDNAmethylation:islands,startsites,genebodiesandbeyond.NatureReviewsGenetics,13(7),484–492.http://doi.org/10.1038/nrg3230
Klip,A.,Marette,A.,Dimitrakoudis,D.,Ramlal,T.,Giacca,A.,Shi,Z.Q.,&Vranic,M.(1992).Effectofdiabetesonglucoregulation.Fromglucosetransporterstoglucosemetabolisminvivo.DiabetesCare,15(11),1747–1766.http://doi.org/10.2337/diacare.15.11.1747
Laurent,L.,Wong,E.,Li,G.,Hodges,E.,Smith,A.D.,Kendall,J.,…Sung,K.(2010).DynamicchangesinthehumanmethylomeduringdifferentiationDynamicchangesinthehumanmethylomeduringdifferentiation,320–331.http://doi.org/10.1101/gr.101907.109
Li,B.,Carey,M.,&Workman,J.L.(2007).TheRoleofChromatinduringTranscription.Cell.http://doi.org/10.1016/j.cell.2007.01.015
Ling,C.,&Groop,L.(2009).Epigenetics:Amolecularlinkbetweenenvironmentalfactorsandtype2diabetes.Diabetes,58,2718–2725.http://doi.org/10.2337/db09-1003
Locasale,J.W.(2013).NIHPublicAccess.NatureRevCancer,13(8),572–583.http://doi.org/10.1038/nrc3557.Serine
LunaLópez,A.(2010).MecanismosEndóGenosDeGeneracióNDeEspeciesReactivasDeOxíGenoYRespuestaCelularAntioxidanteAnteElEstréSOxidativo.EnvejecimientoHumano.UnaVisiónTransdisciplinaria,93–102.
Maresca,A.(2015).DnaMethyltransferase1MutationsandMitochondrialPathology:IsMtdnaMethylated?FrontiersinGenetics,6(March),1–15.http://doi.org/10.3389/fgene.2015.00090
Marette,a,Dimitrakoudis,D.,Shi,Q.,Rodgers,C.D.,Klip,a,&Vranic,M.(1999).GlucoserapidlydecreasesplasmamembraneGLUT4contentinratskeletalmuscle.Endocrine,10(1),13–8.http://doi.org/10.1385/ENDO:10:1:13
Matough,F.A.,Budin,S.B.,Hamid,Z.A.,Alwahaibi,N.,&Mohamed,J.(2012).فيةدسكلألةداضلماداولماو.5–18,(February)12,تافعاضمضرميركسلارودداهجلإايدسكأتلا
Mortusewicz,O.,Schermelleh,L.,Walter,J.,Cardoso,M.C.,&Leonhardt,H.(2005).RecruitmentofDNAmethyltransferaseItoDNArepairsites.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,102(25),8905–8909.http://doi.org/10.1073/pnas.0501034102
Nieratschker,V.,Batra,A.,&Fallgatter,A.J.(2013).Geneticsandepigeneticsofalcoholdependence.JournalofMolecularPsychiatry,1(1),11.http://doi.org/10.1186/2049-9256-1-11
Nolan,C.J.,Damm,P.,&Prentki,M.(2011).Type2diabetesacrossgenerations:Frompathophysiologytopreventionandmanagement.TheLancet,378(9786),169–181.http://doi.org/10.1016/S0140-6736(11)60614-4
Olsen,A.S.,SarrasJr.,M.P.,&Intine,R.V.(2010).Limbregenerationisimpairedinanadultzebrafishmodelofdiabetesmellitus.WoundRepairRegen,18(5),532–542.http://doi.org/10.1111/j.1524-475X.2010.00613.x
Olsen,A.S.,Sarras,M.P.,Leontovich,A.,&Intine,R.V.(2012).HeritabletransmissionofdiabeticmetabolicmemoryinzebrafishcorrelateswithDNAhypomethylationandaberrantgeneexpression.Diabetes,61,485–491.http://doi.org/10.2337/db11-0588
Patel,H.,Chen,J.,Das,K.C.,&Kavdia,M.(2013).HyperglycemiainducesdifferentialchangeinoxidativestressatgeneexpressionandfunctionallevelsinHUVECandHMVEC.CardiovascularDiabetology,12,142.http://doi.org/10.1186/1475-2840-12-142
Poretsky.(2010a).PrinciplesofDiabetesMellitus.
Poretsky,L.(2010b).Principlesofdiabetesmellitus.PrinciplesofDiabetesMellitus.http://doi.org/10.1007/978-0-387-09841-8
Prasad,R.B.,&Groop,L.(2015).Geneticsoftype2diabetes—pitfallsandpossibilities.Genes,6(1),87–123.http://doi.org/10.3390/genes6010087
Raiber,E.-A.,Beraldi,D.,Ficz,G.,Burgess,H.E.,Branco,M.R.,Murat,P.,…Balasubramanian,S.(2012).

93
Genome-widedistributionof5-formylcytosineinembryonicstemcellsisassociatedwithtranscriptionanddependsonthymineDNAglycosylase.GenomeBiology,13,R69.http://doi.org/10.1186/gb-2012-13-8-r69
Ramiro,A.R.,&Barreto,V.M.(2015).Activation-inducedcytidinedeaminaseandactivecytidinedemethylation,40(3).
Reiling,E.,Ling,C.,Uitterlinden,A.G.,Van’tRiet,E.,Welschen,L.M.C.,Ladenvall,C.,…’THart,L.M.(2010).Theassociationofmitochondrialcontentwithprevalentandincidenttype2diabetes.JournalofClinicalEndocrinologyandMetabolism,95(4),1909–1915.http://doi.org/10.1210/jc.2009-1775
Rodríguez-Dorantes,M.,Téllez-Ascencio,N.,Cerbón,M.A.,Lez,M.,&Cervantes,A.(2004).MetilacióndelADN:UnfenómenoepigenéticodeimportanciaMédica.RevistadeInvestigacionClinica.
Rönn,T.,Volkov,P.,Davegårdh,C.,Dayeh,T.,Hall,E.,Olsson,A.H.,…Ling,C.(2013).ASixMonthsExerciseInterventionInfluencestheGenome-wideDNAMethylationPatterninHumanAdiposeTissue.PLoSGenetics,9(6).http://doi.org/10.1371/journal.pgen.1003572
Sanz,L.a,Kota,S.K.,&Feil,R.(2010).Genome-wideDNAdemethylationinmammals.GenomeBiology,11(3),110.http://doi.org/10.1186/gb-2010-11-3-110
Saxonov,S.,Berg,P.,&Brutlag,D.L.(2006).Agenome-wideanalysisofCpGdinucleotidesinthehumangenomedistinguishestwodistinctclassesofpromoters.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,103(5),1412–1417.http://doi.org/10.1073/pnas.0510310103
Trachootham,D.,Lu,W.,Ogasawara,M.a,Nilsa,R.-D.V.,&Huang,P.(2008).Redoxregulationofcellsurvival.Antioxidants&RedoxSignaling,10(8),1343–1374.http://doi.org/10.1089/ars.2007.1957
Villeneuve,L.M.,Reddy,M.a.,&Natarajan,R.(2011).Epigenetics:Decipheringitsroleindiabetesanditschroniccomplications.ClinicalandExperimentalPharmacologyandPhysiology,38(January),401–409.http://doi.org/10.1111/j.1440-1681.2011.05497.x
Volkmar,M.,Dedeurwaerder,S.,Cunha,D.a,Ndlovu,M.N.,Defrance,M.,Deplus,R.,…Fuks,F.(2012).DNAmethylationprofilingidentifiesepigeneticdysregulationinpancreaticisletsfromtype2diabeticpatients.TheEMBOJournal,31(January),1405–1426.http://doi.org/10.1038/emboj.2011.503
Volkmar,M.,Dedeurwaerder,S.,Cunha,D.A.,Ndlovu,M.N.,Defrance,M.,Deplus,R.,…Eizirik,D.L.(2012).DNAmethylationprofilingidentifiesepigeneticdysregulationinpancreaticisletsfromtype2diabeticpatients,(January),1405–1426.http://doi.org/10.1038/emboj.2011.503
Zaidi,S.K.,Young,D.W.,Montecino,M.,Lian,J.B.,Stein,J.L.,vanWijnen,A.J.,&Stein,G.S.(2010).Architecturalepigenetics:mitoticretentionofmammaliantranscriptionalregulatoryinformation.MolecularandCellularBiology,30(20),4758–66.http://doi.org/10.1128/MCB.00646-10
Zhu,J.-K.(2009).ActiveDNAdemethylationmediatedbyDNAglycosylases.AnnualReviewofGenetics,43,143–166.http://doi.org/10.1146/annurev-genet-102108-134205
BibliografíaONLINE
ADA,2015.AsociaciónAmericanadeDiabetes.Disponibleonlineenhttp://www.diabetes.org/es/
CCDs,2016.ConsensusCDSproteindatabase.Disponibleonlineenhttps://www.ncbi.nlm.nih.gov/CCDS/CcdsBrowse.cgi
DANE,2016.DepartamentodeAdministrativosNacionaldeestadísticas.Disponibleonlinehttp://www.dane.gov.co
IDF,2016.FederaciónInternacionaldeDiabetes.Disponibleonlineenhttp://www.idf.org
NCBI,2016.NationalCenterforBiotechnologyInfomation.Disponibleonlineenhttp://www.ncbi.nlm.nih.gov/gene/406904
OMIM,2016.OnlineMendelianInhertanceinMan.Disponibleonlineenhttp://www.omim.org
OMS,2016.OrganizaciónMundialdelaSalud.Disponibleonlineenhttp://www.who.int/es/

94
XIII.ANEXOS
13.1EPIDEMIOLOGÍADELADIABETES
13.1.1Datosmundiales
Ladiabetesmellitusesunproblemadesaludpúblicagraveycrecienteanivelmundial,queprovocaunareducciónenlaexpectativadevidayaumentodelamorbilidaddebidoalascomplicacionesespecíficasquegenera la enfermedad (Ceriello, 2008). Es una de las enfermedades de mayor importancia en todo elmundo,porserunadelaspatologíasnotransmisiblesmásfrecuentesyporlaseveridadydiversidaddesus complicaciones crónicas. En los últimos 25 años se ha incrementado de manea dramática y suprevalenciavaríasegúnlapoblaciónyelnúmerodeestudiosquedescribenlasfrecuenciasenlosdistintosentornossocialesygeográficos(Sierraetal.2006).
El centrodeprensade laOrganizaciónMundialde laSalud(OMS),anunció losúltimosdatosycifrasdepersonas condiabetes en Septiembredel pasado año. LaOMSestimaque en elmundohaymásde347millonesdepersonascondiabetes.Másdel80%delasmuertesseregistranenpaísesdebajoydemediorecurso económico. Se calculó que en el 2004 fallecieron más de 3.4 millones de personas comoconsecuencia del exceso de azúcar en sangre y que para el 2030, la diabetes será la séptima causa demortalidadconmásde400millonesdeafectados(OnlineOMS,2015;Villeneuveetal.2011).
Según la Federación Internacional de Diabetes, más de lamitad de los posibles casos aún no han sidoidentificados,unaestimaciónde4.8millonesdepersonasmuerenalañoysunúmerodeafectadosvaenaumentoentodoslospaíses(OnlineIFD,2015).LaT2DM,eseltipomáscomúnentrelasdiferentesclasesde diabetes.Millones de americanos han sido diagnosticados con esta enfermedad ymuchos no se hanpercatadodelaltoriesgoqueseenfrentan.Existengruposconmayorriesgodepadecerdiabetestipodosqueotros,comoeselcasodeAméricaLatina.SegúnlaFederaciónInternacionaldediabetes,laincidenciade la comunidad latina va en aumento y tiene tasasdos vecesmás altas que los no latinos (Online IDF,2014).
Tabla26.Estimacióndeprevalenciadediabetesparael2030.
13.1.2DatosenColombia
EnColombia,suincidenciaanualvaenaumentoysuprevalenciaoscilaentreel4-8%enzonasurbanasyel2%enzonasrurales,enfuncióndelrangoyedaddelapoblaciónestudiada.Múltiplesfactoresgenéticos

95
yambientalescomoelcambioenlosestilosdevida,elenvejecimiento,elmestizajeylaurbanizaciónhanimpulsado el aumento de sobrepeso y el síndromemetabólico contribuyendo así a su patogenicidad ypersistencia.Enel2007elDepartamentoAdministrativoNacionaldeEstadísticas(DANE),informóqueladiabetes se encuentra dentro de la primeras cinco causas de muerte anuales y es cuatro veces másfrecuenteenlapoblaciónurbanaqueenlarural.LaprevalenciadeT1DMesdel0.07%ysuincidenciaesde3-4por100,000niñosmenoresa15años(Aschner,2010).
13.1.3PosiblesCausasepidemiológicas
Las posibles causas de la epidemia de la diabetes se deben a múltiples factores genéticos, sociales yambientales. Las modificaciones en los estilos de vida tras la industrialización y globalización de lassociedadeshanconllevadoaladecadenciaenlaactividadfísicanoprogramadayalfácilaccesoaalimentosricos en energía con un mínimo esfuerzo. Incrementando así la adiposidad corporal, desarrollandoresistenciaalainsulinaqueterminapordesbordarlacapacidadpancreáticaydesencadenarlaT2DMenindividuosgenéticamentesusceptibles.Elincrementodelalongevidadtraselcontroldelasenfermedadesinfecciosas entre otras, han permitido una mayor expectativa de vida y un envejecimiento relativo. Laprevalenciade laDMincrementacon laedadyamedidaquesomosmás longevossuprevalenciaglobalaumenta(Sierraetal.2006).
Las hipótesis del genotipo y fenotipo ahorrador, aumentan la prevalencia de la diabetes en personassusceptibles.Personasconungenotipoeconómicosehadescritoencomunidadesruralesquepasandeunestilo de vida marcado por la actividad física y la restricción calórica a otro caracterizado por elsedentarismoylaaltaingestacalóricaalmoversealaciudad.Lascomunidadeshumanashandepasarporépocas enfrentándose por medio hostil de escases de alimentos y necesidades de desplazarse a pie,cambiandolacapacidaddealmacenamientodelaenergíacaracterizándoseenindividuosquegastanpocoyalmacenaneficientementeentiemposdeescasez.Lahipótesisdelfenotipoahorradorsehaestudiadoenniños con bajo peso al nacer, que tienen mayor riesgo de desarrollar T2DM y obesidad dado a lascondicionesdeprivacióndenutrientesenelúteroquesobrevivenporgenerarunfenotipoahorrador.Alllegara laedadadultaposeeunaportecalóricosuperioralquesucuerporequiere,desarrollandoasí laobesidad,resistenciaalainsulinayT2DM(Sierraetal.,2006).
13.2GRANDESESTUDIOSDEDIABETESENLAPOBLACIONHUMANA
Una de las preguntas y problemas que han acompañado en el tratamiento de la diabetes se debe a larelaciónquehayentreelcontrolglucémicoyeldesarrollodelascomplicacionesdeladiabetes.Diversosestudioshanevidenciadoelpapeldeanomalíasmetabólicasylascomplicaciones,ysehaconcluidoqueuncontrol glucémico meticuloso podría prevenir el desarrollo de dichas complicaciones. Dos estudios decohortesmostraronelefectoquetieneestecontrolenlaprevencióny/oretrasodelascomplicacionesyenlamemoriametabólicadelindividuo(Poretsky,2010).
13.2.1 DiabetesControlandComplicationsTrial(DCCT)ElDiabetesControlandComplicationsTrial(DCCT)fueunestudiomulticéntricoimportantequeserealizóporelInstitutoNacionaldeSaluddelosEstadosUnidos(NIH)yelInstitutodeDiabetesyDigestiónydeEnfermedades del Riñón (NIDDK) entre 1983-1993. Se diseñó este estudio para evaluar si el controlestrictodelaglucosapuedepreveniroreducirlatasadeprogresióndelascomplicacionesdeladiabetesaunlargoplazo.ElDCCTinvolucroa1,441voluntariosentrelos13-39añosdeedadconT1DMy39centrosentre los Estados Unidos y Canadá. Los criterios de inclusión de los voluntarios es que debían tenerdiabetespor lomenos1añoynomásde15años,yquenotuvieranpresenciadepatologías..Elestudiocomparo los efectos estándares de los niveles de glucosa versus un control intensivo de glucosa con

96
valores de HbAc1 menores a 6.0 y mantener glucosa en sangre entre los 70-120mg/dL y un grupoconvencionalconcontrolnormalmanteniendolaglucosapordebajode180mg/dLpostprandial(Diabetes&Clearinghouse,1993;Poretsky,2010a).
Figura40.DCCTyEDIC(Pirolaetal.,2010).
En ambos tratamientos los valores medio de glucosa se mantuvieron en 155mg/dL en el tratamientointensivoy231mg/dLentratamientoconvencional.Losresultadossediscutenentablaacontinuación.
ResultadosdelaDCCT:
ControlIntensivodelaGlucosaReduce:
76%deriesgodeenfermedadesdelojo
50%deriesgodeenfermedadesenriñones
60%deriesgodeenfermedadesdelnervio
Tabla27.ResultadosdelaDCCT(Diabetes&Clearinghouse,1993).
13.2.2 EpidemiologyofDiabetesInterventionsandComplications(EDIC)Al finalizar elDCCT los investigadores continuaron evaluando las implicaciones a largoplazo y nació elestudio epidemiológico llamado Epidemiology of Diabetes Interventions and Complications (EDIC) quesiguióconmásdel90%delospacientes.Esteestudiodeseguimientoseenfocoenevaluarlaincidenciaypredecirloseventoscardiovascularescomoinfartocardiaco,accidentescerebrovascularesolanecesidadde cirugíade corazónasí como lasotras complicaciones relacionadas comodañoen la visión, riñonesynervios.Tambiénevalúael impactode lacalidaddevidayelcostoefectivoentre losdostratamiento.ElEDIC realizó el seguimiento cambiando a los del tratamiento convencional al tratamiento intensivo(Diabetes&Clearinghouse,1993;Poretsky,2010a).

97
ResultadosdelaEDIC
Controlglucémicointensivoreduce
42%decualquiereventocardiovascular
57% de ataque cardiaco, accidentescerebrovasculares
Tabla28.ResultadosdeEDIC(Diabetes&Clearinghouse,1993).
Tabla29.Mayoresdescubrimientosporlosestudiosmundialesmásimportantes(Nolanetal.,2011).
13.2.3 TheUnitedKingdomProspectiveDiabetesStudy(UKPDS)En1998serealizóelestudiomásgrande,TheUnitedKingdomProspectiveDiabetesStudy(UKPDS)quefuediseñadoparaobservarlosefectosdelcontroldelaglucemiaencomplicacionesalargoplazoenpacientesT2DM.Sereclutaron5,102pacientescondiagnósticorecienteyseleshizounseguimientode11años.Secomparó dos tratamientos, un tratamiento intensivo que consistía en aplicación de insulina oantidiabéticosoralesoambosmanteniendolosnivelesdeHbAc1pordebajode7.0%ysecomparóconlaterapiaconvencionalcondietaysieranecesario terapia farmacológicaynivelespromediodeHbAc1de7.9%.Losresultadoseneltratamientointensivomostraronunareducciónenelriesgodemuertesúbita,muerte por hiperglucemia, infartos miocardio, accidentes cerebrovasculares, insuficiencia renal,amputaciones, hemorragia vítrea, fotocoagulación de la retina, ceguera o cataratas en un 12 %. Lareducciónderiesgoderetinopatíasfuedel21%yelaspectodemicroalbuminuriaseredujoun30%. Lareducción en la presión arterial no disminuyó significativamente en comparación a las complicacionesmicrovascularesymacrovascularesqueseredujeronun37%y34%respectivamente(Poretsky,2010a).

98
Enresumentodoslosestudiosrealizadosendichascohortes,demostraronqueuncontrolglucémicoenlosestadios tempranode laenfermedad, juntoauncontrolen lapresiónsanguíneay losnivelesde lípidos,previene y reducen de manera significativa las complicaciones macro y microvasculares en la T1DM yT2DM.