Tema 6 cinética enzimática y regulación
28
Tema 6. Cinética enzimática. Ecuación de Michaelis Menten. Factores que afectan a la velocidad de reacción. Inhibición enzimática. Regulación de la actividad enzimática. Enzimas alostéricas. Modulación covalente DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR Prof. Juan Jiménez Carrasco
-
Upload
usfisioterapialuisfernandez -
Category
Education
-
view
1.855 -
download
3
Transcript of Tema 6 cinética enzimática y regulación

Tema 6. Cinética enzimática. Ecuación de Michaelis Menten. Factores que afectan a la velocidad de reacción. Inhibición enzimática. Regulación de la actividad enzimática. Enzimas alostéricas. Modulación covalente
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Catálisis enzimáticaPara que cualquier reacción química tenga lugar, los reactantes deben chocar con una energía y una orientación adecuadas. Las enzimas:
1. Permiten que los reactantes (sustratos) se unan a su centro activo con una orientación óptima para que la reacción tenga lugar.
2. Modifican las propiedades químicas del sustrato unido a su centro activo, debilitando los enlaces existentes y facilitando la formación de otros nuevos.
El producto tendrá menor energía interna que el sustrato. Esta diferencia de energía se denomina energía libre de Gibbs (∆Gº‘). Para que la transformación tenga lugar, el sustrato (S) debe activarse o tener suficiente energía interna para pasar a un estado de transición de alta energía (S*). La energía necesaria para que el sustrato se active se denomina energía de activación.Las enzimas aceleran la velocidad de la reacción disminuyendo la energía de activación, sin modificar las condiciones del equilibrio y, al final de ella, se recuperan sin modificarse.
El sustrato debe alcanzar un estado transitorio de alta energía que le permita transformarse en producto, sin pasar por este estado la reacción no tiene lugar.

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Velocidad de las reacciones enzimáticas
La velocidad de una reacción no catalizada es directamente proporcional a la concentración de sustrato activado (S*): v = k [S*]Cuanto mayor sea la energía de activación (EA) que necesite el sustrato (S) para alcanzar el estado transitorio (S*), más lenta será la reacción. Cuanto más bajo sea el salto energético, más rápida será la reacción.
En las reacciones catalizadas por enzimas, la formación del complejo enzima-sustrato (ES) tiene la ventaja de que estabiliza al sustrato en el estado transitorio (S*), con un nivel energético más bajo que si no estuviera catalizada. Es decir, necesitan menor energía de activación (EA), lo que le permite transformase en producto mucho antes: v = k [ES]La enzima crea un microentorno donde la configuración del sustrato activado (S*) es menos desfavorable energéticamente.
Con enzimas
Sin enzimas
EA
EA
Una enzima no tiene por qué actuar siempre a la misma velocidad. Su actividad puede estar modulada por factores como: el pH, la Tª o la presencia de cofactores o coenzimas.

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Cinética de las reacciones enzimáticaLa cinética enzimática estudia la velocidad de las reacciones catalizadas por los enzimas. Los estudios cinéticos proporcionan información directa acerca del mecanismo de la reacción catalizada y de la especifidad de la enzima. La velocidad de una reacción catalizada por un enzima puede medirse con relativa facilidad, ya que no suele ser necesario purificar o aislar la enzima. La medida debe realizarse utilizando siempre concentraciones de sustrato saturantes, una concentración de enzima constante y condiciones óptimas de pH y Tª. Bajo estas condiciones:
2.- La velocidad de la reacción enzimática puede determinarse midiendo tanto la aparición de la [P], como la desaparición de la [S].
Equilibrio
Tiempo
1.- La velocidad de una reacción enzimática (V), al igual que una reacción no enzimática, es proporcional a la [S], pero se diferencia en que es saturable y alcanza un máximo (Vmáx).
Reacción no catalizada
Reacción catalizada
Sustrato
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
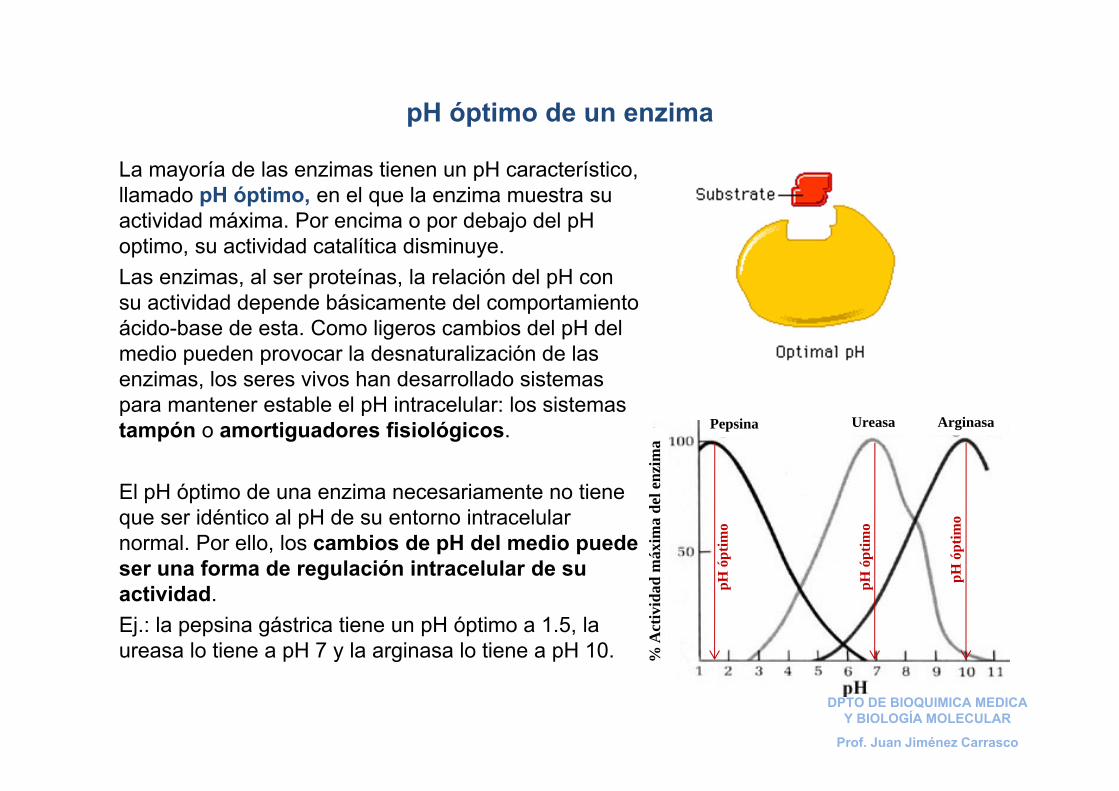
La mayoría de las enzimas tienen un pH característico, llamado pH óptimo, en el que la enzima muestra su actividad máxima. Por encima o por debajo del pH optimo, su actividad catalítica disminuye.Las enzimas, al ser proteínas, la relación del pH con su actividad depende básicamente del comportamiento ácido-base de esta. Como ligeros cambios del pH del medio pueden provocar la desnaturalización de las enzimas, los seres vivos han desarrollado sistemas para mantener estable el pH intracelular: los sistemas tampón o amortiguadores fisiológicos.
El pH óptimo de una enzima necesariamente no tiene que ser idéntico al pH de su entorno intracelular normal. Por ello, los cambios de pH del medio puede ser una forma de regulación intracelular de su actividad.Ej.: la pepsina gástrica tiene un pH óptimo a 1.5, la ureasa lo tiene a pH 7 y la arginasa lo tiene a pH 10.
pH óptimo de un enzima
pH ó
ptim
o
pH ó
ptim
o
pH ó
ptim
o
% A
ctiv
idad
máx
ima
del e
nzim
a
Pepsina Ureasa Arginasa

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
La Tª en la que la actividad catalítica de una enzima es máxima se denomina Tª óptima. En general, el aumento de Tª acelera las reacciones químicas. Las reacciones catalizadas por enzimas siguen la Ley general de Van´t Hoff: Por cada 10ºC de incremento de Tª, la velocidad de reacción se duplica.Pero las enzimas al ser proteínas, cuando superan ciertas Tª (50-60ºC), se desnaturalizan y dejan de funcionar. Existen algunas especies de bacterias y algas que habitan en fuentes de aguas termales muy cálidas y, en el otro extremo, ciertas bacterias árticas presentan Tª óptimas cercanas a 00 C.
Tª óptima de un enzima
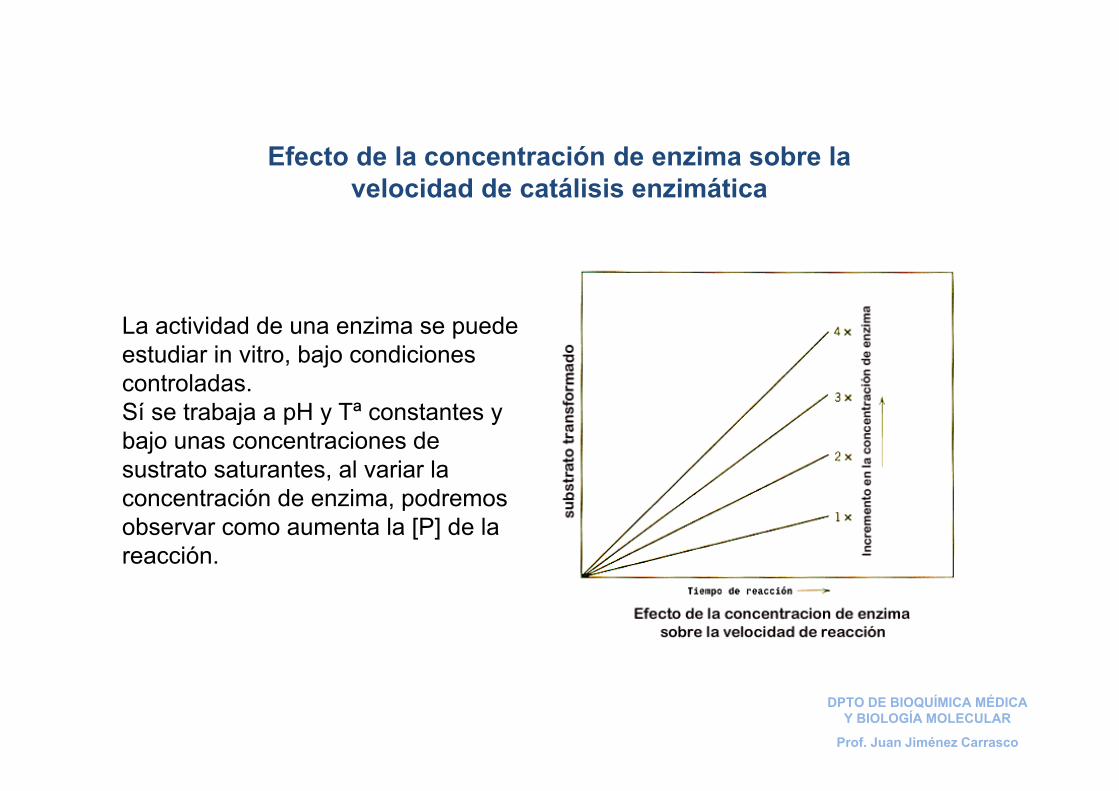
La actividad de una enzima se puede estudiar in vitro, bajo condiciones controladas. Sí se trabaja a pH y Tª constantes y bajo unas concentraciones de sustrato saturantes, al variar la concentración de enzima, podremos observar como aumenta la [P] de la reacción.
Efecto de la concentración de enzima sobre la velocidad de catálisis enzimática
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

Leonor Michaelis y Maud Menten (1913) propusieron unmodelo simple, que explica las características cinéticas deenzimas sencillas (no sujetas a regulación), considerando:
Cinética enzimática de Micaelis - Menten
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Leonor Michaelis y Maud Menten, en 1913 dedujeron la relación entre la velocidad máxima (Vmáx) de una reacción catalizada enzimáticamente, la formación del complejo enzima-sustrato (ES) y la formación de producto (P). En condiciones de alta [S], todos los sitios activos de la enzima se encontrarán saturados y, por tanto, la velocidad de reacción alcanza su Vmáx. En los enzimas que presentan este comportamiento se asume la existencia de un complejo ES intermediario. Así, al combinarse el enzima (E) con el sustrato (S) se forma rápidamente el complejo (ES), con una constante de velocidad k1. En ese momento, el complejo ES puede:1. Formar el producto (P) y regenerar el enzima (E), con
una constante de velocidad k3, en una reacción mucho más lenta que la anterior, por lo que es laetapa limitante de la reacción.
2. O puede disociarse de nuevo, generando nuevamente E y S, con una constante de velocidad k2.
Según este modelo, la velocidad (V0) de una reacción catalizada enzimáticamente será: V0 = k3·[ES]Como el valor [ES] no es fácil de obtener, será necesario una expresión equivalente, con valores conocidos, obtenidos por el desarrollo matemático o Ec. de Michaelis-Menten
Ecuación de Michaelis-Menten
Si hacemos que la V0 = ½ Vmáx
Km = [S]
½ Vmáx
Vmáx
Km
[Sustrato]
Ecuación de Michaelis- MentenV0 = Vmáx·[S]
Km+[S]

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Ecuación de Michaelis-Menten
Unión al sustrato Etapa catalítica• El modelo de Michaelis Menten supone que S se une a E, formando un intermediario ES, que
luego reacciona y se descompone en P + E.• También presupone que las [E], [S] y [ES] se encuentran todos en equilibrio, alcanzando una
concentración estacionaria de [ES] y que la descomposición del complejo [ES] en [E] + [P] es el paso limitante de la velocidad de catálisis, ya que la constante de disociación de este complejo [ES] k3 (o Kcat) <<< k1
• En esta reacción, la proporción de [ES], en relación con el nº total de moléculas de Enzima total [ET], es decir la relación [ES]/[ET], limita la velocidad inicial (V0) de la enzima, de manera que: V0 = Kcat ·[ES]
• Puesto que E, S, y ES se encuentran en equilibrio químico, la enzima alcanza una velocidad máxima (Vmáx) a concentraciones de sustrato [S] muy elevadas (saturantes), cuando [ES] [ET]. Por lo que: Vmax = Kcat ·[ET].
• Para la disociación del complejo [ES], la ley de acción de masas implica: Kd =• Teniendo en cuenta que: [ET] = [E] + [ES], • Puede demostrarse que: , por tanto: V0 =
• Puesto que Kcat corresponde a la Vmáx que se alcanza a altas [S], se obtiene la:Ec. de Michaelis-Menten: V0 =
[E]·[S][ES]
[ES][ET]
__[S]___, donde Km = Kd
Kd + [S])= Kcat [ET]·[S])Km+[S]
Vmáx·[S]Km+[S]
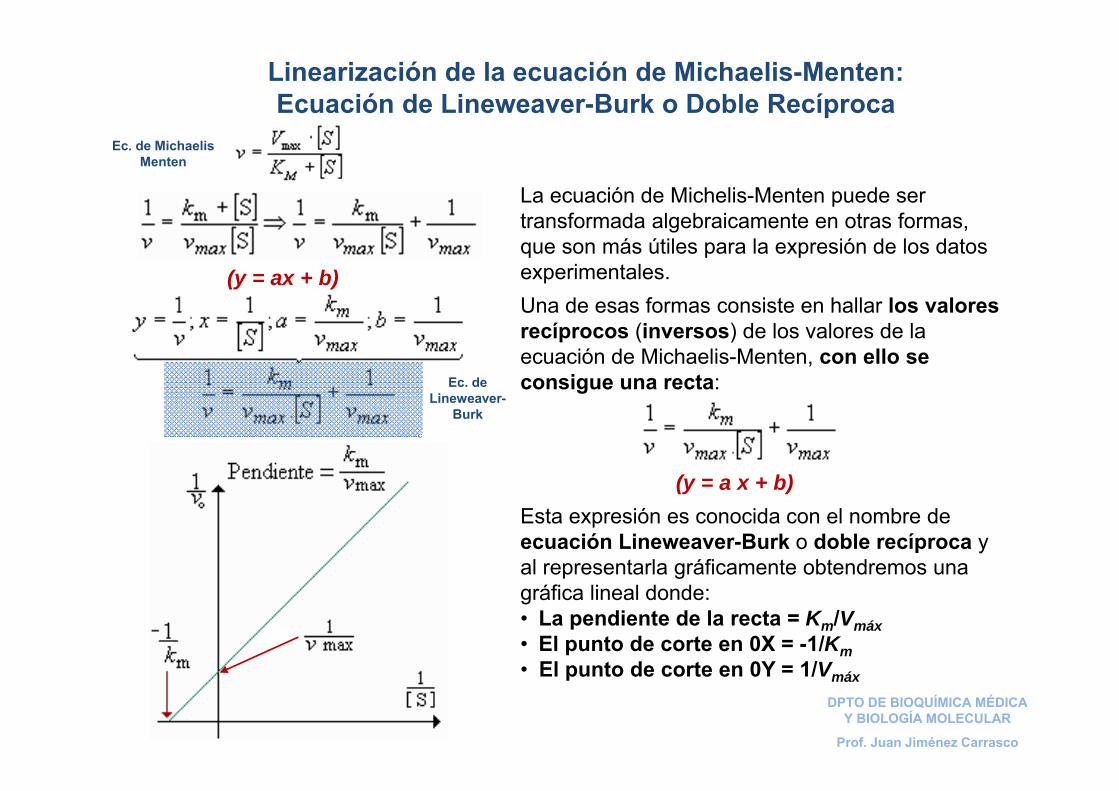
Linearización de la ecuación de Michaelis-Menten: Ecuación de Lineweaver-Burk o Doble Recíproca
(y = ax + b)
Ec. de Michaelis Menten
Ec. de Lineweaver-
Burk
La ecuación de Michelis-Menten puede ser transformada algebraicamente en otras formas, que son más útiles para la expresión de los datos experimentales. Una de esas formas consiste en hallar los valores recíprocos (inversos) de los valores de la ecuación de Michaelis-Menten, con ello se consigue una recta:
(y = a x + b)Esta expresión es conocida con el nombre de ecuación Lineweaver-Burk o doble recíproca y al representarla gráficamente obtendremos una gráfica lineal donde:• La pendiente de la recta = Km/Vmáx• El punto de corte en 0X = -1/Km• El punto de corte en 0Y = 1/Vmáx
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
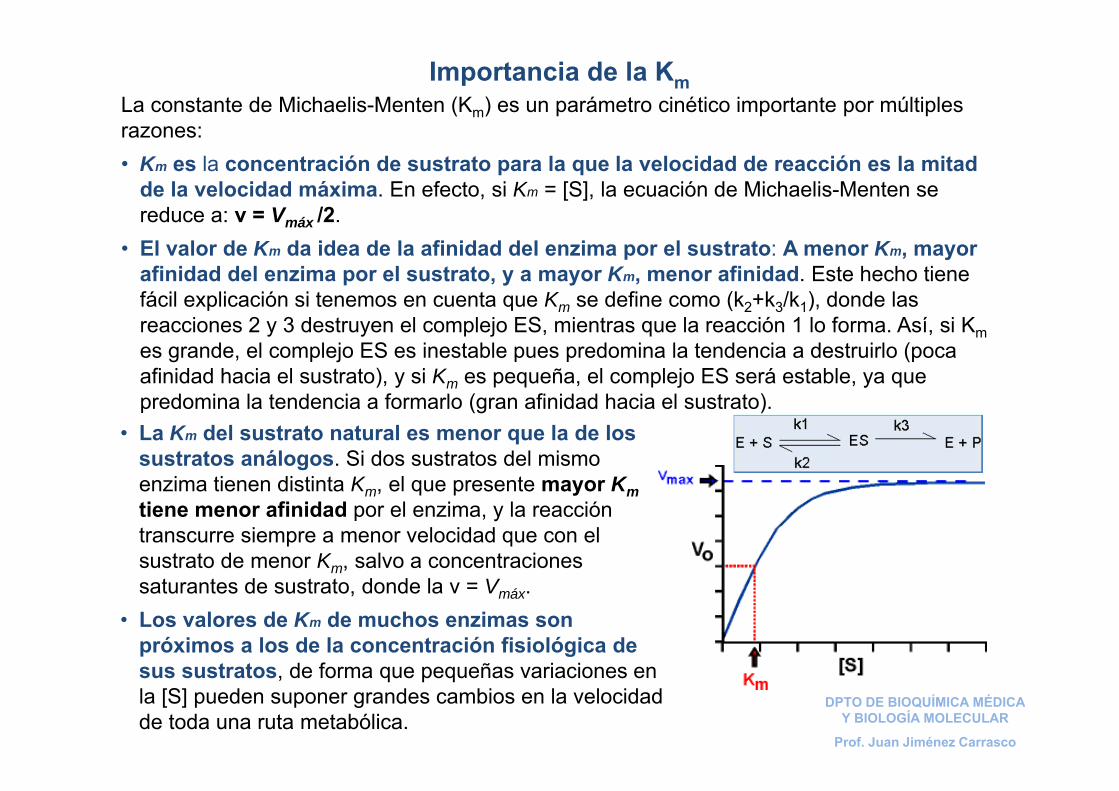
Importancia de la KmLa constante de Michaelis-Menten (Km) es un parámetro cinético importante por múltiples razones:• Km es la concentración de sustrato para la que la velocidad de reacción es la mitad
de la velocidad máxima. En efecto, si Km = [S], la ecuación de Michaelis-Menten se reduce a: v = Vmáx /2.
• El valor de Km da idea de la afinidad del enzima por el sustrato: A menor Km, mayor afinidad del enzima por el sustrato, y a mayor Km, menor afinidad. Este hecho tiene fácil explicación si tenemos en cuenta que Km se define como (k2+k3/k1), donde las reacciones 2 y 3 destruyen el complejo ES, mientras que la reacción 1 lo forma. Así, si Kmes grande, el complejo ES es inestable pues predomina la tendencia a destruirlo (poca afinidad hacia el sustrato), y si Km es pequeña, el complejo ES será estable, ya que predomina la tendencia a formarlo (gran afinidad hacia el sustrato).
• La Km del sustrato natural es menor que la de los sustratos análogos. Si dos sustratos del mismo enzima tienen distinta Km, el que presente mayor Kmtiene menor afinidad por el enzima, y la reacción transcurre siempre a menor velocidad que con el sustrato de menor Km, salvo a concentraciones saturantes de sustrato, donde la v = Vmáx.
• Los valores de Km de muchos enzimas son próximos a los de la concentración fisiológica de sus sustratos, de forma que pequeñas variaciones en la [S] pueden suponer grandes cambios en la velocidad de toda una ruta metabólica.

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
1. Velocidad inicial (v o V0): es el nº de moles de S transformados en P, por unidad de tiempo, medidos antes de que se haya formado suficiente P como para permitir que ocurra la reacción contraria (generalmente la [S] >>> [E]). La V0 es siempre proporcional a la [E], no es constante y depende del estado estacionario del complejo ES => V0 = k3 · [ES]
2. Nº de recambio o Turnover number (Kcat): es una constante que describe la rapidez con que un enzima cataliza una reacción. Se define como el nº de moléculas de S transformadas en P, por molécula de E y por unidad de tiempo, en condiciones de saturación de sustrato. Como k3 es la limitante de la reacción en las enzimas michaelianas, Kcat = k3. Es un valor constante para cada E y mide su eficiencia. Se expresa en μmoles/min. V0 = Kcat · [ES]
3. Velocidad máxima (Vmáx): es la velocidad en la que todos los centros activos están ocupados (saturados) con S. En la realidad no se alcanza al ser directamente proporcional a la concentración de enzima total [ET]. Así, [ET] = [ELibre] + [ES]. Al depender de la [ET], la Vmáx no es constante. Vmáx = Kcat · [ET]
4. Constante de Michaelis (Km): es la concentración de sustrato necesaria para alcanzar la mitad de la Vmáx. Mide la afinidad o tendencia de la enzima a reaccionar con un determinado sustrato. Puede variar mucho de un enzima a otro o en un mismo enzima, cuando actúa sobre distintos sustratos.
• Una Km alta: indica una baja afinidad de la enzima por su sustrato • Una Km baja: indica una alta afinidad de la enzima por su sustrato
¡Los valores de Kcat, Km y Vmáx son únicos para cada enzima!
Parámetros cinéticos característicos de una enzima

Unidades de actividad enzimática• Actividad de un enzima: son los moles de sustrato transformados por unidad de
tiempo. Se expresa en μmoles/min o unidades de actividad enzimática (U). Por lo tanto, la cantidad de una enzima se expresa usando unidades de Velocidad.
• Katal (kat): 1 katal = 60·106 U, es la unidad del Sistema Internacional de Unidades para actividad catalítica. Esta unidad es muy grande, de forma que suele utilizarse submúltiplos, como el µkat (10-6 kat) o el ηkatal (10-9 kat).
• Actividad específica de un enzima: es la actividad enzimática por mg de proteínas totales del tejido. Se expresa como μmoles/min/mg proteína o lo que es lo mismo U/mg prot.
• Concentración de un enzima: es la cantidad de enzima por unidad de volumen, o en su defecto la actividad enzimática por unidad de volumen U/ml. Muy útil la evaluación de enzimas séricas: U/ml de suero.
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

Hay enzimas cuya cinética no obedece a la ecuación de Michaelis-Menten. Se dice que su cinética es “no michaeliana”. Esto ocurre con los enzimas alostéricos, cuya gráfica de saturación no es hiperbólica, sino sigmoidal.La cinética sigmoidal de estas enzimas, se caracteriza por presentar una zona crítica (cercana a la Km), donde pequeñas variaciones en la [S] se traducen en grandes variaciones en la velocidad de reacción.
Cinética de los enzima no michaelianos
V
[S]Km
1/2Vmax
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
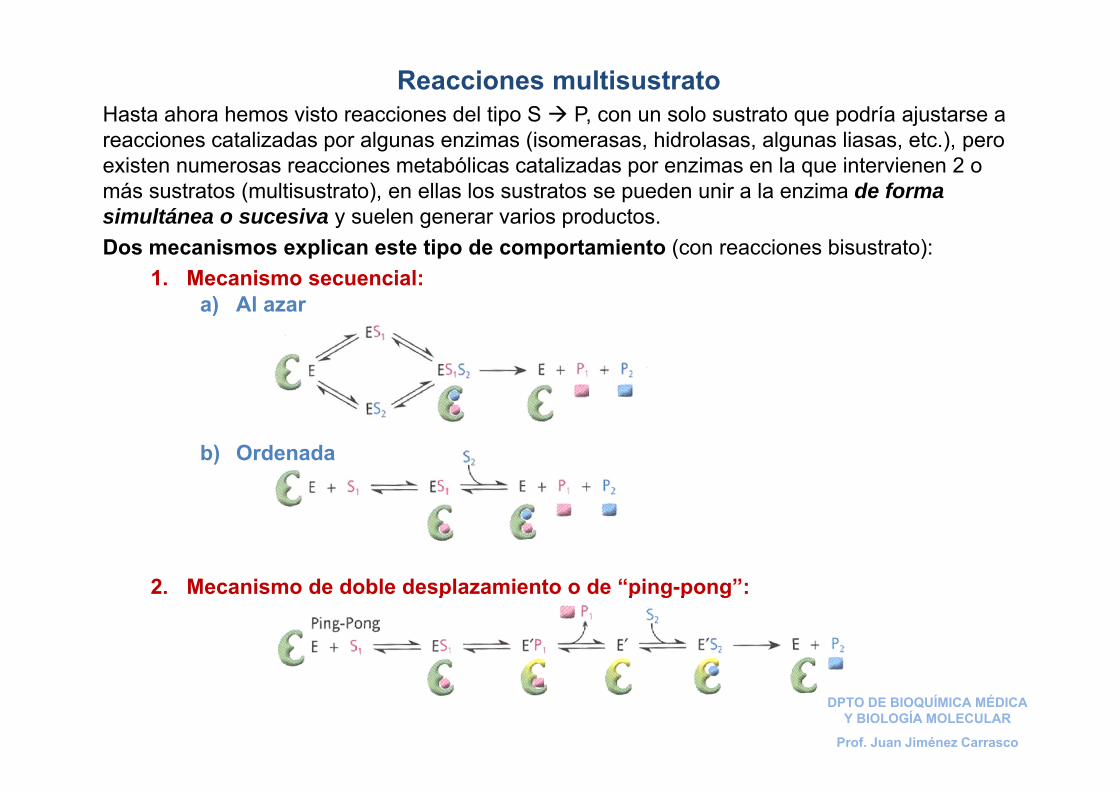
Reacciones multisustratoHasta ahora hemos visto reacciones del tipo S P, con un solo sustrato que podría ajustarse a reacciones catalizadas por algunas enzimas (isomerasas, hidrolasas, algunas liasas, etc.), pero existen numerosas reacciones metabólicas catalizadas por enzimas en la que intervienen 2 o más sustratos (multisustrato), en ellas los sustratos se pueden unir a la enzima de forma simultánea o sucesiva y suelen generar varios productos. Dos mecanismos explican este tipo de comportamiento (con reacciones bisustrato):
1. Mecanismo secuencial:
2. Mecanismo de doble desplazamiento o de “ping-pong”:
a) Al azar
b) Ordenada
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

Inhibición enzimáticaLos inhibidores son moléculas, naturales o sintéticas, que ralentizan o detienen las reacciones enzimáticas. Muchos fármacos actúan como inhibidores enzimáticos; por ejemplo: El sildenafilo (viagra), inhibidor la fosfodiesterasa 5, sirve para tratar la disfunción eréctil.Las formas de inhibición enzimática pueden ser:
1. Inhibición Reversible: El inhibidor se une a la enzima por enlaces débiles. Se caracterizan por una rápida disociación del complejo enzima-inhibidor, por lo que la inhibición es transitoria. Alterando los parámetros cinéticos (Km y/o Vmáx). Las principales formas de inhibición reversible son:
a) Competitiva b) Acompetitivac) No competitiva
2. Inhibición Irreversible: El inhibidor se une fuertemente al enzima de tal forma que el tiempo de disociación es muy largo, esto puede ocurrir tanto por uniones covalentes como no covalentes pero en cualquier caso, resultan muy difíciles de eliminar.
Cuando una enzima posee múltiples sustratos, sus inhibidores pueden mostrar distinto tipo de inhibición, dependiendo del sustrato que se considere, ya que el sitio activo posee diferentes lugares de unión, uno para cada sustrato y/o inhibidor.
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

Muchos fármacos son inhibidores enzimáticos
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Inhibición reversible competitiva
En la inhibición reversible, el inhibidor (I) se une de forma transitoria al enzima (E), mediante enlaces de tipo débil al enzima, por lo que puede disociarse rápida y fácilmente.En este tipo de inhibición, el lnhibidor (I) suele poseer una estructura similar al sustrato (S), compitiendo por acceder al mismo centro activo del enzima. Por ello solo puede unirse al enzima libre (nunca al complejo ES). Como el sustrato y el inhibidor no pueden unirse a la vez a la misma enzima:
• Si se eleva la [S], se puede superar la inhibición.
• No modifica la Vmáx (Vmáx(i) = Vmáx)• Aumenta la Km, (K’m(i) > Km)
Ej.: La intoxicación con metanol, se trata con etanol (mismo enzima: ADH)

En este tipo de inhibición, el I y el S no compiten por el centro activo del E (de ahí que se le llame inhibición acompetitiva). El inhibidor (I) solo se une al complejo ES ya formado, produciendo un complejo inactivo ESI estable.
• Como el complejo ESI no forma producto (P) disminuye la Vmáx
• El inhibidor y el aumento del complejo ESI, afectan a la disociación del sustrato disminuye la Km.
• En la representación de dobles inversos, como la Vmax y la Km se modifican en la misma proporción, se obtienen rectas paralelas y, por tanto, presentan la misma pendiente.
Inhibición reversible acompetitiva
complejo inactivo ESI
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
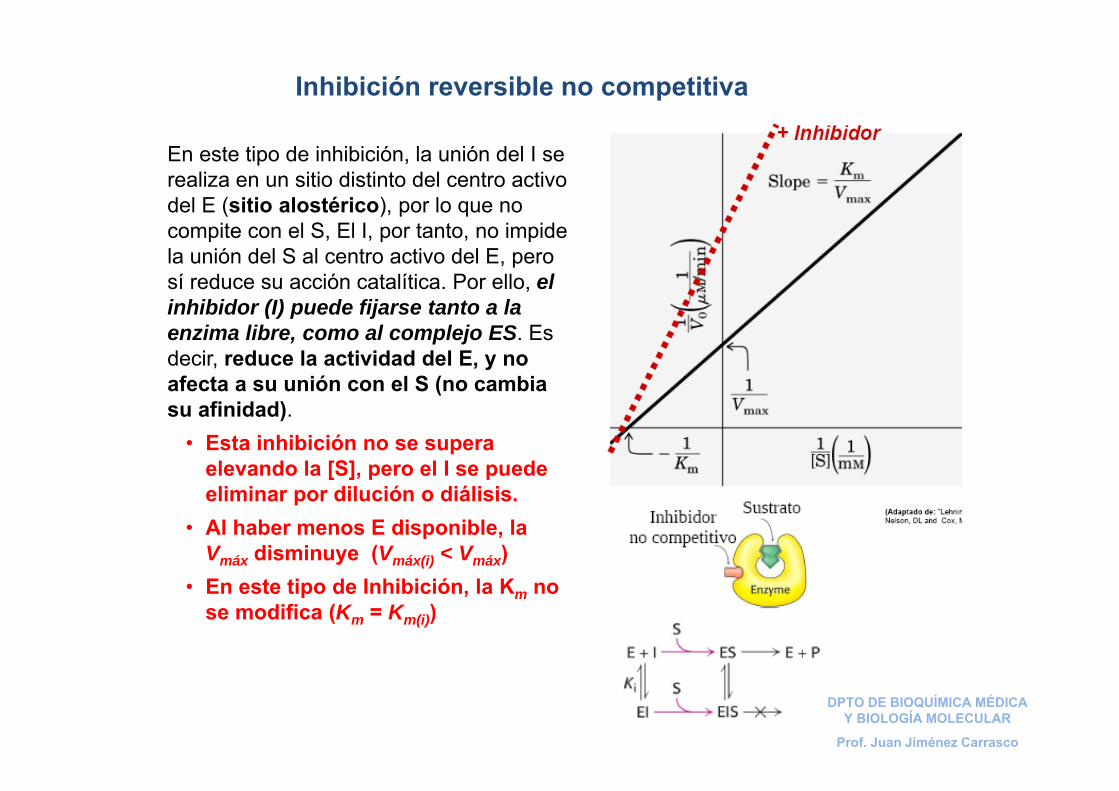
Inhibición reversible no competitiva
En este tipo de inhibición, la unión del I se realiza en un sitio distinto del centro activo del E (sitio alostérico), por lo que no compite con el S, El I, por tanto, no impide la unión del S al centro activo del E, pero sí reduce su acción catalítica. Por ello, el inhibidor (I) puede fijarse tanto a la enzima libre, como al complejo ES. Es decir, reduce la actividad del E, y no afecta a su unión con el S (no cambia su afinidad).
• Esta inhibición no se supera elevando la [S], pero el I se puede eliminar por dilución o diálisis.
• Al haber menos E disponible, la Vmáx disminuye (Vmáx(i) < Vmáx)
• En este tipo de Inhibición, la Km no se modifica (Km = Km(i))
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Inhibición irreversibleEn la inhibición irreversible, el inhibidor (I) se une tan fuertemente a la enzima que el tiempo de disociación del complejo (EI) es muy lento o no se produce. Generalmente ocurren por uniones covalentes, aunque también pueden ser no covalentes. Modifican la estructura química a nivel de los residuos aminoácidos, o los grupos funcionales, necesarios para la actividad catalítica. Suelen ser tóxicos naturales o sintéticos. En muchos casos la interacción del inhibidor se produce compitiendo con el sustrato, por el centro activo del enzima, lo que lleva a la disminución de la Vmáx y un aumento de la Km
aparente (del inhibidor). Este proceso es irreversible porque aunque se aumente la [S], este es incapaz de desplazar al inhibidor del centro activo.
Los inhibidores suicidas son un caso particular de inhibidores irreversibles, son sustratos modificados que unidos al centro activo del enzima lo transforma en una especie química muy reactiva que modifica covalentemente al enzima, inactivándola. Tienen, por tanto, la especificidad del inhibidor competitivo y la potencia de los inhibidores irreversibles. Actualmente se utilizan como fármacos dirigidos contra enzimas específicas. No presentan actividad hasta que se unen al centro activo de ellas, por lo que suelen ser muy efectivos.
• La penicilina modifica covalentemente una transpeptidasa responsable de la síntesis de la pared celular bacteriana.
• El alopurinol es un análogo estructural de la hipoxantina que inhibe la xantina oxidasa de forma irreversible, impidiendo que se forme ácido úrico, responsable de la gota
• La aspirina modifica covalentemente una ciclooxigenasa de la ruta de síntesis de las prostaglandinas involucradas en fenómenos de inflamación y dolor.

Enzimas reguladoresEn el metabolismo celular hay grupos de enzimas que funcionan conjuntamente, en rutas secuenciales, para realizar un proceso metabólico determinado (Por ej.: Conversión de glucosa en lactato en el músculo esquelético). En ellas, el producto de la reacción del primer enzima se convierte en el sustrato de la siguiente y así sucesivamente. La mayor parte de los enzimas de una ruta metabólica siguen las características hasta ahora descritas. Sin embargo, en cada ruta hay algún enzima que fija la velocidad de la secuencia global, ya que catalizan la reacción más lenta, constituyendo el paso limitante de la velocidad de la ruta. Estos son los enzimas reguladores, que además muestran una actividad catalítica mayor o menor, en función de ciertas señales o formas de síntesis.Enzimas reguladores:
• Enzimas alostéricos, por unión reversible, no covalente, de compuestos reguladores (moduladores o efectores alostéricos), metabolitos o cofactores.
• Enzimas regulados por modificaciones covalentes reversibles.• Enzimas regulados por su síntesis inactiva: Zimógenos
Los enzimas reguladores suelen estar formados por varias subunidades y en algunos casos el sitio o sitios reguladores y el sitio activo, se encuentran en subunidades diferentes.
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco

Las enzimas alostéricas son aquellos cuya actividad está regulada por la unión de distintos efectores a sitios diferentes (centros alostéricos) al que se une el sustrato (centro activo). Son muy importantes por estar situados en puntos clave (encrucijadas) de las rutas metabólicas.• Los efectores o moduladores los pueden activar
(positivos) o inhibir (negativos), induciendo un cambio conformacional del centro activo, que facilita o inhibe la actividad enzimática.
• La mayoría de los enzimas alostéricos son multímeros. Es decir, están compuestos por varias subunidades funcionales o protómeros..
• Presentan cooperatividad, el enzima puede estar en dos conformaciones distintas con diferente actividad enzimática (estados conformacionales R y T). La unión de un efector a una subunidad altera el equilibrio entre ambas conformaciones y el cambio conformacional se transmite a una o todas las subunidades vecinas. Como consecuencia de estos fenómenos los enzimas alostéricos presentan curvas sigmoidales, al representar V0 frente a [S], por lo que no se ajustan a la cinética de Michaelis-Menten(curvas hiperbólicas).
Enzimas alostéricas
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
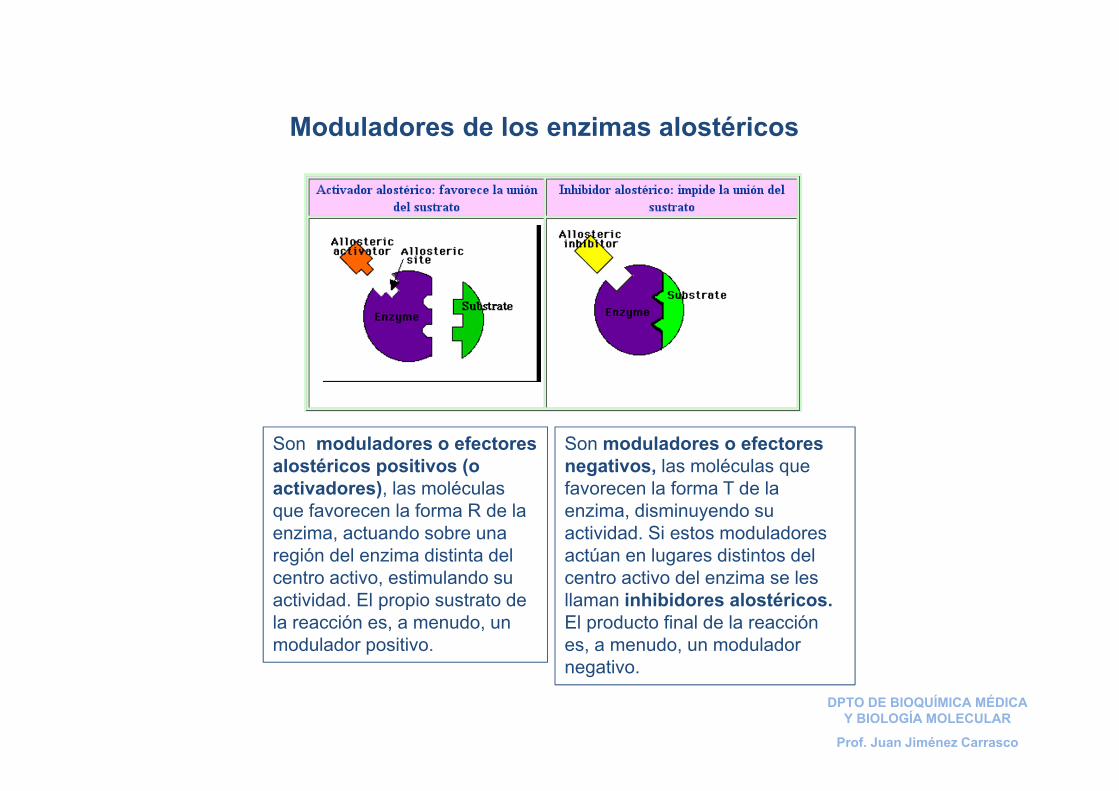
Son moduladores o efectores negativos, las moléculas que favorecen la forma T de la enzima, disminuyendo su actividad. Si estos moduladores actúan en lugares distintos del centro activo del enzima se les llaman inhibidores alostéricos. El producto final de la reacción es, a menudo, un modulador negativo.
Son moduladores o efectores alostéricos positivos (o activadores), las moléculas que favorecen la forma R de la enzima, actuando sobre una región del enzima distinta del centro activo, estimulando su actividad. El propio sustrato de la reacción es, a menudo, un modulador positivo.
Moduladores de los enzimas alostéricos
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
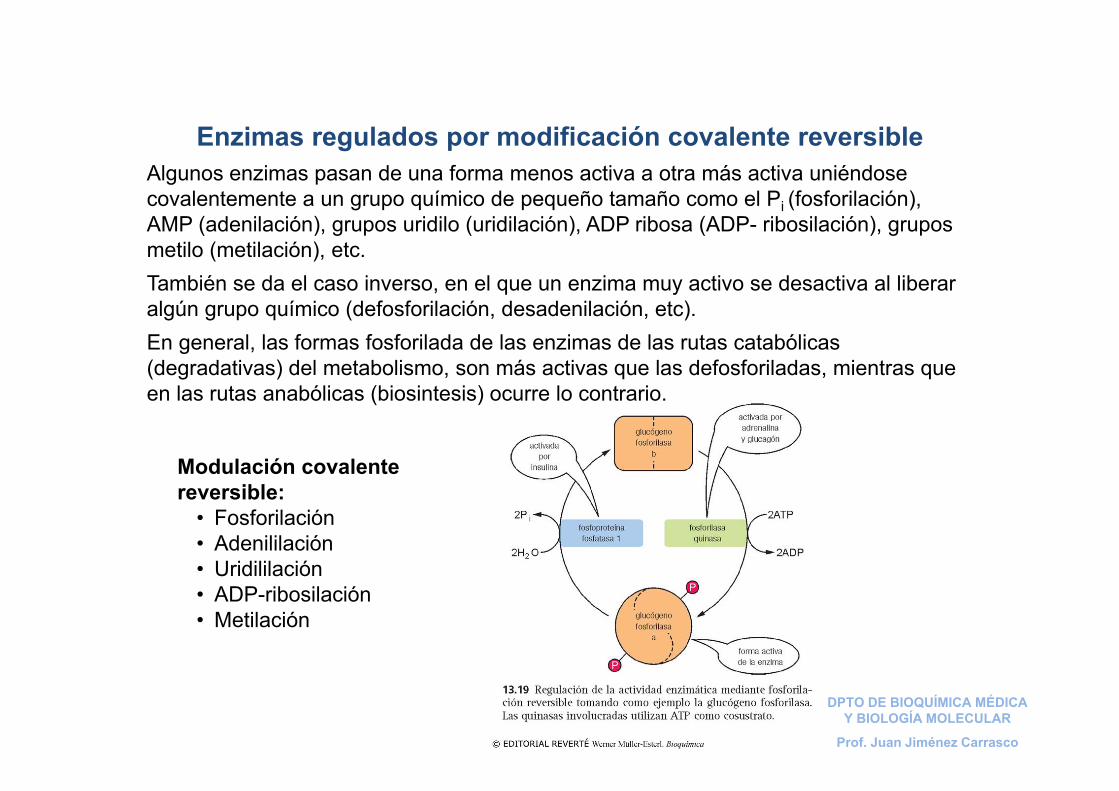
Enzimas regulados por modificación covalente reversibleAlgunos enzimas pasan de una forma menos activa a otra más activa uniéndose covalentemente a un grupo químico de pequeño tamaño como el Pi (fosforilación), AMP (adenilación), grupos uridilo (uridilación), ADP ribosa (ADP- ribosilación), grupos metilo (metilación), etc.También se da el caso inverso, en el que un enzima muy activo se desactiva al liberar algún grupo químico (defosforilación, desadenilación, etc). En general, las formas fosforilada de las enzimas de las rutas catabólicas (degradativas) del metabolismo, son más activas que las defosforiladas, mientras que en las rutas anabólicas (biosintesis) ocurre lo contrario.
Modulación covalente reversible:
• Fosforilación• Adenililación• Uridililación• ADP-ribosilación• Metilación

DPTO DE BIOQUÍMICA MÉDICA Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco
Activación proteolítica: ZimógenosAlgunos enzimas se sintetizan en forma inactiva, proenzimas o zimógenos, para evitar su acción en el propio tejido que lo sintestiza. Posteriormente, estos se activan en el lugar donde tienen que realizar su función catalítica. Es característico de los enzimas digestivos.
La enteropeptidasa inicia la cascada de activación de los zimógenos pancreáticos, en los primeros tramos del intestino delgado, activando al tripsinógeno en tripsina, que después será la encargada de activar al resto de zimógenos (proelastasa, quimotripsinógeno, procarboxipeptidasa, prolipasa, al propio tripsinógeno, etc), en sus correspondientes formas activas (elastasa, quimotripsina, carboxipeptidasa, lipasas, tripsina, etc).

Regulación de la actividad enzimática• Regulación general:
• Las propias concentraciones de S y P• El pH y la Tª• Los cofactores y coenzimas
• Regulación específica:1. Alosterismo y cooperatividad
• Moduladores positivos• Moduladores negativos
2. Modificación covalente:• Fosforilación• ADP-Ribosilación• Uridilación• Otros…
3. Síntesis de formas inactivas: Zimógenos4. Compartimentación celular: Isoenzimas5. Control de la expresión de Enzimas
• Enzimas constitutivos• Enzimas inducibles DPTO DE BIOQUÍMICA MÉDICA
Y BIOLOGÍA MOLECULAR
Prof. Juan Jiménez Carrasco