
Regulación de la densidad de tallos en el tiempo en...
55
Regulación de la densidad de tallos en el tiempo en cultivo de tomate bajo invernadero frío. Alumno: Ximena Alejandra Ibarra Leiva. Fecha: Mayo, 2003.
Transcript of Regulación de la densidad de tallos en el tiempo en...

Regulación de la densidad de tallos en el tiempo en cultivo de
tomate bajo invernadero frío.
Alumno: Ximena Alejandra Ibarra Leiva.
Fecha: Mayo, 2003.

1. INTRODUCCIÓN.
El tomate (Lycopersicon esculentum L.) es considerado, por superficie cultivada e
importancia económica, una de las principales hortalizas del país. Este cultivo es uno
de los que más tecnología utiliza en su proceso productivo, siendo también la
hortaliza más estudiada a nivel mundial, con un constante desarrollo de técnicas para
optimizar su producción (BRAVO y ALDUNATE, 1986).
Dentro de estas técnicas se encuentra la conducción de la planta, labor generalizada y
que se realiza de diversas formas según el criterio y experiencia del agricultor
(SALINAS, RAMÍREZ y OSPINA, 1994). La conducción de las plantas de tomate en
producciones bajo invernadero se realiza generalmente con el sistema de eje simple y
disposición alterna de las plantas (GONZÁLEZ, 1994). Otros sistemas de conducción
pueden implementarse aumentando el número de ejes en producción por planta. Un
ejemplo de esto es la conducción a dos o tres ejes, los cuales son originados a partir
de una yema axilar (DUIMOVIC, 1992).
Una opción para obtener mayores producciones o provocar una reducción en los
costos del cultivo es la implementación de un sistema de conducción que regule la
densidad de tallos por unidad de superficie en el tiempo de acuerdo a la intensidad
lumínica de la estación en que se desarrolla el cultivo. La implementación de este
sistema se traduce en numerosos beneficios como el aumento de la producción de
fruta comercializable, manteniendo los calibres preferidos por el mercado a lo largo
del cultivo, el ahorro de plantines por hectárea y la menor incidencia de desórdenes
de maduración provocados por un aumento en el área foliar de la planta
(COCKSHULL, HO y FENLON, 2000; COCKSHULL y HO, 1995).

En esta investigación se plantea un sistema de conducción en dos ejes, en el cual el
segundo eje se forma una vez concluida la fase de crecimiento vegetativo exclusivo.
Mediante este sistema de conducción se busca un aumento del rendimiento, sin la
disminución de la calidad del fruto.

2. REVISIÓN BIBLIOGRÁFICA.
2.1. Descripción de la especie:
El tomate pertenece a la familia Solanaceae y recibe el nombre científico de
Lycopersicon esculentum Mill. Es originario de la costa occidental de América del
Sur, extendiéndose desde Ecuador al norte de Chile (KINET y PEET, 1997).
Es una planta perenne que se cultiva casi universalmente como anual (KINET y
PEET, 1997). Posee un sistema radicular amplio, constituido por una raíz principal
que puede llegar hasta 1.50 m de profundidad y que está provista de un gran número
de raíces secundarias. Desde la base del tallo surge gran cantidad de raíces
adventicias (PICKEN, STEWART y KLAPWIJK, 1986).
El tallo es anguloso, alcanza 4 cm de diámetro en su base y está recubierto por
tricomas, en su mayoría de origen glandular, y que le otorgan el olor característico.
En las primeras etapas es erguido, luego debido al peso, toma un hábito rastrero
(MAROTO, 1994).
Las hojas están dispuestas alternadamente sobre el tallo, son compuestas e
imparipinnadas, generalmente tienen de siete a nueve foliolos lobulados o dentados y
también están cubiertas de tricomas (MAROTO, 1994).
La inflorescencia del tomate está formada en la parte terminal del brote, en forma de
racimos simples o ramificados, pudiendo tener entre tres a diez flores (MAROTO,
1994; ATHERTON y HARRIS, 1986).

2.2. Crecimiento del tallo:
La estructura de la planta corresponde a un conjunto de simpodios. El tallo principal
forma de seis a doce hojas que crecen lateralmente, antes que la yema terminal se
transforme en una inflorescencia. El crecimiento subsiguiente se produce de la yema
axilar de la última hoja, la cual desarrolla un tallo secundario que crece como una
prolongación del tallo primario y desplaza lateralmente a la inflorescencia. Los
sucesivos segmentos del tallo se desarrollan de forma similar, produciendo una
inflorescencia cada tres hojas. El aspecto es el de un tallo principal, que crece en
forma continua con inflorescencias internodales laterales cada tres hojas
(CHAMARRO, 1995).
El desarrollo del tallo es variable y está dado de acuerdo al cultivar, según esto
existen dos hábitos de crecimiento: i) de tipo determinado, en el cual el crecimiento
del eje principal está detenido por la formación de una inflorescencia terminal,
después de haber formado una cierta cantidad de inflorescencias (dos o tres), cada
una o dos hojas, y desarrolla fuertes brotes axilares desde la base del tallo y ii) de tipo
indeterminado, en el que alargamiento del eje principal es continuo y produce
inflorescencias laterales frecuentemente cada tres hojas, durante gran parte de la vida
de la planta (KINET y PEET, 1997).
En la punta del tallo principal se encuentra el meristema apical, región de división
celular activa, donde se inician nuevas hojas y estructuras florales. Tiene forma de
domo y es protegido por hojas recientemente formadas (PICKEN, STEWART y
KLAPWIJK, 1986).
Las hojas están ordenadas alternadamente con filotaxia de 2/5. Incluso en ambientes
fluctuantes ellas son producidas regularmente, por lo menos hasta florecer y crecen

con el tallo encorvado. Generalmente sólo seis a doce hojas son producidas debajo de
la primera inflorescencia (PICKEN, STEWART y KLAPWIJK, 1986).
Es común la aparición de hojas sobre la inflorescencia, a menudo adyacentes a la
primera flor. Además, brotes adventicios pueden potenciarse desde una variedad de
partes vegetativas, particularmente desde el raquis de la hoja madura (PICKEN,
STEWART y KLAPWIJK, 1986).
La fase vegetativa es usualmente corta en el tomate. La transición floral ocurre, en la
mayoría de los cultivares, cuando la tercera hoja está en expansión, lo que sucede
dentro de las tres semanas de la expansión del cotiledón (HURD y COOPER, 1970).
2.3. Particionamiento de fotoasimilados:
En la planta de tomate la distribución de los asimilados desde la hoja hacia órganos
en crecimiento se modifica a medida que la planta se desarrolla. La relación sink-
source de oferta y demanda de asimilados es compleja y dinámica (HO y HEWITT,
1986).
En una planta indeterminada de tomate, el 80 % de la ganancia de peso fresco de la
planta está dada por el crecimiento de los frutos. Como el fruto acumula más agua
que otros órganos, la diferencia en ganancia de peso seco entre órganos es la más
pequeña. Sin embargo, la acumulación diaria de materia seca del fruto es más alta que
en las hojas y tallo (COCKSHULL, GRAVES y CAVE, 1992).
Debido a que los órganos vegetativos son sinks mucho más débiles que los frutos, un
cambio en el número de frutos es principalmente compensado por un cambio inverso
en el tamaño del fruto, en vez de un cambio sustancial en la razón fruto/estructuras
vegetativas (COCKSHULL, GRAVES y CAVE, 1992).

2.3.1. Relación sink–source:
Se considera source a todo órgano capaz de exportar fotosintatos y sink a todo órgano
que los importe. Los principales sources son las hojas que alcanzan un cuarto a un
tercio de su tamaño final y los sinks son principalmente ápices de crecimiento, raíces,
tallos, hojas jóvenes y frutos en desarrollo (GIFFORD y EVANS, 1981). Cada sink
tiene una distinta capacidad para atraer asimilados (HO, 1988). La competencia entre
los diferentes sinks se ve facilitada por la existencia de vías comunes de transporte de
asimilados entre órganos sinks y sus hojas fuente compartidas (HO y HEWITT,
1986).
En general, la distribución de asimilados entre sinks es principalmente regulado por
los mismos sinks y el vigor de cada source es sólo de menor importancia (GIFFORD
y EVANS, 1981; HO, 1988). Sin embargo, el vigor del source puede influenciar
fuertemente la distribución de asimilados en tomate en el largo plazo, como el efecto
del número de frutos en la planta (PAPADOPOULOS y ORMROD, 1990). Por lo
tanto, un incremento en el vigor del source, resulta en un aumento del número de
frutos en simultáneo crecimiento e indirectamente influencia la asignación de
biomasa (HEUVELINK, 1995).
2.3.2. Cambios de prioridad durante el desarrollo de la planta:
Durante el desarrollo de la planta las prioridades de carbohidratos se modifican en
forma dinámica. Cuando la primera inflorescencia está en floración, el tallo y las
raíces son los principales sinks, en ese momento el tallo es alimentado por las hojas
superiores, el ápice por las hojas basales y las raíces por las hojas medias. La
inflorescencia atrae sólo una pequeña proporción de asimilados (HO, 1988). Una vez
que los tres primeros racimos ya han cuajado y están creciendo rápidamente son los
principales sinks y están siendo abastecidos por las hojas medias. Entonces el ápice es

alimentado por las hojas superiores y las raíces por las hojas basales (HO, 1996).
Posteriormente, los frutos se transforman en los principales sinks de asimilados, por
sobre las hojas jóvenes, flores y raíces (HO y HEWITT, 1986).
De acuerdo a HO (1996), al haber una disponibilidad limitada de fotoasimilados, los
frutos en crecimiento se convierten en el sink primario. Ello produce un retraso en el
desarrollo de las inflorescencias siguientes, una mayor demora en el crecimiento de
tallos y raíces, la senescencia temprana de hojas y por último la muerte de las raíces.
La capacidad sink de un fruto, la cual le permite importar asimilados desde las hojas,
se ve definida por: el control físico, el tamaño y número de células, y por el control
del metabolismo, almacenamiento y utilización de asimilados (HO, SJUT y HOAD,
1983; HELLMAN, BARKER y FROMEMER, 2000).
HEUVELINK (1997) demostró que la fuerza generativa del sink es proporcional para
el número de frutos en el rango de dos a siete frutos por racimo, y que la razón entre
la fuerza media del sink de una unidad vegetativa (tres hojas y los entrenudos entre
dos racimos), y la fuerza media del sink de un fruto tiene un valor constante de 2.96.
Los factores que determinan el particionamiento de fotoasimilados se pueden dividir
en factores endógenos y exógenos. Los factores endógenos se refieren principalmente
al sistema vascular y a la relación sink-source. Dentro de los factores exógenos se
encuentran la luz, la concentración de anhídrido carbónico, la temperatura, la
nutrición, las condiciones hídricas, la poda y las restricciones en las raíces (HO, 1988;
HO y HEWITT, 1986).

2.3.3. Efecto de la poda de tallos laterales sobre el particionamiento de asimilados:
SALINAS, RAMÍREZ y OSPINA (1994) afirman que la poda afecta el equilibrio
sink-source de la planta, puesto que, en una planta sin podar existe mayor
competencia interna por el llenado de los frutos, pues debe suplir a un mayor número
de éstos, lo que no sucede en una planta con uno o dos tallos. Un brote axilar joven,
como cualquier órgano joven, es un depósito y usa los asimilados producidos por las
hojas del tallo principal. Cuando se reduce la frecuencia de la poda de los brotes, cada
uno de estos crece más largo y se vuelve un competidor más fuerte por asimilados
para el tallo principal, las raíces y frutos. Finalmente, el brote axilar llega a ser una
fuente de carbono. Sin embargo, la relación entre el estado del carbono de los brotes
axilares y la frecuencia de desbrote no es conocida (NAVARRETE y JEANNEQUIN,
2000). En conclusión, en una planta a dos tallos, el equilibrio sink-source es más
estable y se suple mejor el llenado de frutos, obteniéndose una mejor calidad de éstos
(SALINAS, RAMÍREZ y OSPINA, 1994).
HARTMANN (1977) citado por NAVARRETE y JEANNEQUIN (2000), encontró
una correlación negativa entre el peso de los brotes axilares y la producción, lo que
confirma que el crecimiento de los brotes axilares implica una competencia con el
desarrollo de los frutos.
2.4. Regulación del crecimiento vegetativo:
La variación de almidón y sacarosa en condiciones normales son pocas en la hoja. El
transporte de sacarosa se realiza a través del floema y cada hoja suministra nutrientes
a determinados órganos de la planta (PICKEN, STEWART y KLAPWIJK, 1986).
El factor externo que más afecta al desarrollo vegetativo es la iluminación diaria total,
mientras que la calidad de la luz y el fotoperiodo tienen un rol secundario (HO y

HEWITT, 1986). El efecto de la radiación solar diaria en el alargamiento del tallo es
complejo. A valores cercanos a 2MJ m-2d-1 la altura de las plantas de un mismo peso
seco son constantes. Reduciendo la radiación solar diaria bajo estos niveles
generalmente incrementa la razón de elongación del tallo a expensas de otras partes
de la planta; el resultado es un tallo más frágil, delgado y alto (KINET, 1977). En
cambio, con reducciones mayores, la elongación del tallo es muy reducida debido a la
baja intensad de luz (PICKEN, STEWART y KLAPWIJK, 1986).
Hay efectos en la elongación del tallo debidos a la edad de la planta. El
sombreamiento antes de la fructificación reduce o no tiene efecto sobre la razón de
elongación del tallo, mientras que el sombreamiento después de la fructificación, lo
incrementa (COOPER, 1969). KINET (1977) comprobó que días largos aumentan el
alargamiento del tallo e incrementan el peso fresco de éste. Además, la proporción de
alargamiento del tallo generalmente aumenta con la temperatura, dando lugar a tallos
más delgados, con mayor proporción de tejido parenquimático y agua (CALVERT,
1964). Según COOPER (1969) el largo del día óptimo para la elongación del tallo
disminuye a medida que la temperatura aumenta o a medida que la planta crece.
La temperatura también puede ser importante para el desarrollo vegetativo del
tomate. Para determinadas condiciones de edad e iluminación, la temperatura diurna
óptima para el desarrollo vegetativo es de 23ºC y la nocturna de 17ºC (MAROTO,
1994). Cuando las temperaturas diurnas son elevadas, un descenso en las
temperaturas nocturnas puede traer beneficios, pero cuando las temperaturas diurnas
no son óptimas, la elevación de temperaturas nocturnas favorece el desarrollo
vegetativo (PICKEN, STEWART y KLAPWIJK, 1986).
Según HUSSEY (1963) citado por KINET y PEET (1997), existe una fuerte
competencia entre las hojas en desarrollo y el meristema apical. Una alta
disponibilidad de asimilados bajo condiciones de alta radiación estimula la actividad

del meristema y el crecimiento de la hoja, pero cuando las plantas están limitadas de
fuentes (altas temperaturas o baja luz), el crecimiento de hojas jóvenes es favorecido
a expensas del desarrollo apical. Los brotes axilares jóvenes compiten por asimilados
con el tallo principal, raíces y frutos. Así, una completa defoliación de una planta
joven favorece a los brotes laterales en detrimento del tallo principal. El desarrollo de
los brotes axilares es promovido por bajas temperaturas o días cortos, pero depende
de la respuesta del cultivar (AUNG, 1976).
De acuerdo a VERKERK (1955) citado por KINET y PEET (1997), el crecimiento
del sistema radicular es estimulado en relación con el crecimiento del brote en
condiciones de alta irradiancia. Como resultado, las condiciones que inducen una
proporción tallo/raíz elevada, son más detrimentales para el desarrollo reproductivo
de la planta (CALVERT, 1964).
2.5. Densidad poblacional:
Las densidades de tomate bajo invernadero en Chile varían, dependiendo del ciclo del
cultivo, en un rango que fluctúa comúnmente de 15.000 a 40.000 plantas·ha-1
(GIACONI y ESCAFF, 1994). De acuerdo a la encuesta realizada por GONZÁLEZ
(1995), en la provincia de Quillota se observa que en un invernadero tradicional con
pies centrales, se alcanzan densidades de 2.3 plantas·m-2 con tres mesas de plantación
y 18 cm entre plantas
En España la densidad de plantación más frecuente es de 2.5 plantas·m-2, oscilando de
2.0 a 4.0 plantas·m-2 de acuerdo al vigor varietal, fertilidad del sustrato, salinidad del
suelo y agua de riego. En invernaderos climatizados en Europa Occidental, las
densidades más frecuentes son de 2.5 plantas·m-2 en ciclo largo de transplante precoz
y de 3 a 3.25 plantas·m-2 en ciclo corto, aunque también se utilizan densidades
menores de 2.0 plantas·m-2, que luego se incrementan con transplantes

complementarios a mitad de ciclo (VAN DE VOOREN, WELLES y HAYMAN,
1986). En condiciones óptimas de fertilidad de suelo se recomiendan densidades de
1.0 a 1.3 plantas·m-2 en cultivares determinados, sin entutorar ni podar y de 1.8 a 2.1
plantas·m-2 en cultivos indeterminados, podados y entutorados (GEISEMBERG y
STEWART, 1986).
Con relación a las condiciones de precios del tomate en nuestro país, MENDOZA
(1992) afirma que, debido a la alta estacionalidad, se justifica plenamente el uso de
una alta densidad de plantas (35.000-40.000 plantas·ha-1) con una cosecha teórica de
245.000 a 280.000 racimos, en conducción a un eje y siete racimos por eje. La mayor
densidad permite concentrar la producción, de tal forma que el mayor número de
racimos cosechados se encuentre en los meses de mayor precio (agosto a noviembre)
(MENDOZA, 1992).
La densidad de plantas a utilizar en un invernadero depende del desarrollo vegetativo
de éstas, el cual es característico de cada cultivar. También se debe considerar el
hábito de crecimiento, época de cultivo, método de poda y entutorado, fertilización
del suelo, tipo de riego, exposición a la luz, ventilación, arquitectura del invernadero
y clima de la región (ESCAFF, 1993; CASTILLA, 1995).
De acuerdo a FERY y JANICK (1970), los incrementos de producción obtenidos con
altas densidades de población son principalmente una función del incremento del
número de frutos producidos por unidad de área. En consecuencia, el aumento de la
producción temprana es el resultado del incremento del número de primeras flores
cuajadas del racimo por unidad de área y no a un aceleramiento en la madurez. Por lo
tanto, una alta densidad provoca una disminución en el número de frutos por planta y
una disminución en el calibre. El menor número de frutos por planta se debe al menor
desarrollo de racimos, menor número de flores por racimo y por un bajo porcentaje de
frutos cuajados (GEORGE y PEIRCE, 1969). En general, a mayor densidad de

plantas de tomate por unidad de área el rendimiento aumenta en forma asintótica, es
decir, aumenta a una razón decreciente (FERY y JANICK, 1970).
Según CASTILLA (1995), los efectos destacables de una alta densidad de población
son el aumento de la producción temprana y el acortamiento del ciclo del cultivo, y
los efectos adversos serían una menor calidad del fruto, al haber mayor porcentaje de
calibres pequeños y el aumento del riesgo de enfermedades. Las condiciones
climáticas de nuestro país, especialmente en invierno, hacen que una alta densidad de
plantación provoque problemas como: falta de luz por un excesivo emboscamiento,
alta incidencia de enfermedades fungosas, debidas a la mala ventilación y a las
heridas provocadas por el deshoje y una mala cobertura de pesticidas por exceso de
canopia (MENDOZA, 1992). La escasez y elevados costos de la mano de obra
agrícola, son un importante factor al decidir la densidad poblacional, sobre todo al
tener en cuenta que la mano de obra aumenta linealmente con la densidad
(MAROTO, 1994).
Según FERY y JANICK (1970) el aumento de la densidad, en un rango de 3.000 a
41.000 plantas por unidad de superficie, no provoca una competencia por asimilados
que logre provocar un efecto de retraso sobre la inducción y desarrollo floral.
Además, al aumentar la población la producción del primer racimo es mayor.
HEUVELINK (1995) mostró que no hay ningún efecto directo en la asignación
proporcional de asimilados dentro del rango de 1.6 hasta 3.1 plantas·m-2.
De acuerdo a FAIGUENBAUM (1986), el rendimiento de una planta está
fuertemente correlacionado con su área foliar (AF). Altas densidades generan un alto
grado de competencia, provocando una disminución del AF por planta a través del
desarrollo del cultivo. Por el contrario, si las plantas se encuentran más espaciadas el
valor de su AF es más elevado, lo que se traduce en mayores rendimientos por planta.

Sin embargo, esta mayor producción individual no alcanza a compensar, en la
mayoría de los casos, la capacidad productiva de poblaciones mayores.
2.6. Distribución espacial de las plantas y ordenamiento de la canopia:
Existen dos sistemas de plantación: a hilera simple y doble (GONZÁLEZ, 1994), es
así como las plantas de tomate pueden estar distanciadas en camas a 1.7 m ó 1.9 m
entre sí y sobre ellas dispuestas a dos hileras a 0.6 m sobre ella. Las plantas se
localizan a 0.3-0.4 m en la hilera (ESCAFF, 1993). MENDOZA (1992) sugiere que
es más conveniente distribuir las plantas en hileras simples distanciadas a 1.1-1.2 m,
con distancias sobre la hilera de 20-25 cm.
Una forma de manejar la distancia sobre la hilera es modificarla de acuerdo al largo
del periodo de cosecha. En un periodo corto de cosecha y con cinco racimos, se usa
una distancia de 0.25 m y cuando se cosechan seis o más racimos se transplanta a 0.3
m (ROJAS y ALFARO, 1995).
El sistema convencional de disponer la canopia es colgar los tallos por medio de un
hilo al alambre en forma vertical. El hilo tutor es sujetado a la base de la planta o se
clava al suelo (ESCAFF, 1992). La disposición de los tallos puede hacerse no vertical
sino ligeramente inclinada, alternando las plantas adyacentes, formando una V, a fin
de conseguir la máxima intercepción de radiación solar por el cultivo (CASTILLA,
1995). Otra forma es permitir la mantención de la porción terminal del tallo junto al
alambre, desplazando la planta a lo largo de la línea de cultivo y dejar la parte baja
del tallo tumbado junto al suelo; esto se realiza preferentemente en cultivos de largo
aliento (GONZÁLEZ, 1994).

2.7. Efecto de la competencia entre tallos frente a un aumento en la densidad:
La luz, cuantificada como radiación fotosintéticamente activa (RFA), es el factor más
importante que afecta la productividad del cultivo de tomate bajo invernadero. Las
disminuciones en RFA y los cambios de duración del periodo de luz propia de cada
estación, reducen la producción de frutos de tomate (PAPADOPOULUS y
PARARAJASINGHAM, 1997).
De acuerdo a COCKSHULL y HO (1995), el rendimiento se determina por la
temperatura del aire del invernadero y tiene relación directa con la radiación solar a
una tasa cercana a dos kilos de peso fresco de fruta por 100 MJ de radiación solar
incidente sobre el cultivo. Por consiguiente, los cambios en el tamaño de la fruta son
causados probablemente por un desequilibrio entre la cantidad de asimilados
producidos en las diferentes estaciones del año y en el número de frutos disponibles
para usar los asimilados en su crecimiento.
RODRÍGUEZ y LAMBETH (1976) observaron que la intercepción de RFA incidente
en la parte superior de la canopia es igual a 76.5%, mientras que alrededor del 20% se
pierde a través de los pasillos e hileras. Además, estudios realizados por
PAPADOUPULUS y ORMOND (1988) indican que la proporción de RFA
disponible se incrementa con espaciamientos más cercanos, pero la penetración de
RFA aumenta y la distribución vertical mejora con el aumento del espaciamiento
entre plantas. Por ejemplo, en un cultivo de otoño plantado a una densidad de 2.5
plantas m-2, sólo el 30-40% del total de radiación solar fue interceptada por las
plantas. Sin embargo, cuando las densidades fueron aumentadas en cultivos de
primavera hasta 3.3 plantas·m-2, el 60-70% de la radiación solar disponible fue
interceptada por el cultivo (HARPER et al., 1979).

Los efectos del sombreamiento que afectan tanto a la producción fotosintética como a
la de auxinas, permitirían explicar por qué las densidades altas están asociadas a un
menor número de frutos por planta (FERY y JANICK, 1970).
2.8. Poda:
La poda es la remoción de estructuras de una planta, ya sean vegetativas o
reproductivas. Dentro de este concepto se incluyen el desbrote, deshoje, raleo de
flores y frutos y despunte o decapitación de la planta (DUIMOVIC, 1992).
El desbrote consiste en la eliminación de los brotes que nacen de las axilas de las
hojas en el tallo principal, de manera de ir formando la planta para conducirla en
altura con el sistema de entutorado seleccionado. Por lo tanto, la poda de brotes busca
un equilibrio entre el volumen de material vegetal y de aire disponible, aunque sea
con la disminución del rendimiento de cada planta, el cual se compensa porque se
puede establecer un mayor número de individuos (NAVARRETE y JEANNEQUIN,
2000).
El desbrote se debe realizar cuando éstos no han excedido los 5 cm, para así evitar
ocasionar una herida grande y no afectar el crecimiento del tallo principal
(DUIMOVIC, 1992). De acuerdo a NAVARRETE, JEANNEQUIN y SEBILLOTE
(1997), la frecuencia de desbrote óptima es entre siete y catorce días, dependiendo del
clima, estación y vigor del cultivar.
La poda de brotes laterales es un factor que afecta la calidad de los frutos. Cuando la
planta es podada hay un control en los calibres del fruto, mientras que una planta
puesta a crecer libremente esta variable no es manejable, afectándose las calidades de
mayor calibre, las cuales son de interés en el mercado especializado (SALINAS,
RAMÍREZ y OSPINA, 1994). NAVARRETE y JEANNEQUIN (2000) evaluaron el

desempeño de plantas de tomate con poda uno y dos ejes y observaron que las plantas
podadas inician su producción más temprano que las plantas sin podar. En las plantas
con poda se obtuvo el mayor porcentaje de frutos de primera calidad, pero con un
menor rendimiento total por unidad de área. En consecuencia, mayores rendimientos
se obtienen de las plantas no podadas, pero los frutos presentan un calibre menor.
2.9. Conducción:
El tomate es una especie decumbente. Es por eso que en el cultivo del tomate en
invernadero es necesario usar un sistema de conducción en altura de la planta, ya sea
con tutores verticales sujetos en el suelo o bien en las estructuras del invernadero, con
el objeto de guiar los tallos y brotes laterales de las plantas mediante amarras
(GONZÁLEZ, 1994; KRARUP, 1992)
ESCAFF (1993) indica que los objetivos de la conducción en altura son poder
desprender del suelo los frutos, reduciendo los riesgos de daños físicos por plagas y
enfermedades, mejorar la exposición a la luz, lo que permite una mejor coloración y
más rápida maduración. También se busca soportar una mayor densidad de
plantación, favoreciendo la aireación, minimizando los microambientes húmedos
generados por los emboscamientos y facilitar todas las labores que se realizan, desde
las aplicaciones de productos químicos a la cosecha.
2.9.1. Sistemas de conducción:
Entre los diferentes sistemas de conducción que existen destaca el “sistema holandés
o de hilo vertical”, que consiste en un alambre grueso que pasa paralelamente a la
línea de plantación a una altura de 2 m aproximadamente y del cual penden los hilos
de material plástico a los cuales van atadas las plantas. El alambre es soportado por
tutores colocados cada 5 m (GONZÁLEZ, 1994). Otro sistema es el inglés, que

consiste en un entramado de alambre colocado en V y sobre el cual van recostadas las
plantas. Es de un costo muy superior al anterior y no favorece mayormente el manejo
de las plantas y su aireación (GONZÁLEZ, 1995). La conducción danesa, consiste en
que, al llegar al cuarto racimo floral cosechado se deshoja la parte inferior de la
planta, la cual se baja dejando el quinto racimo a 0.5 m del suelo y amarrando
nuevamente la planta al alambre superior. Se utiliza para aumentar el lapso de
cosecha (10 racimos o más) (ESCAFF, 1993).
Los tipos más conocidos de conducción son dos: a un eje y a dos ejes. La conducción
a un eje consiste en eliminar todos los brotes axilares del tallo principal, logrando así
un crecimiento indefinido de éste, hasta su eventual despunte, el cual limita el ciclo y
contribuye a mejorar el tamaño del fruto (CASTILLA, 1995). Este tipo de
conducción es la más común de encontrar en la producción de tomates bajo
invernadero en nuestro país. Una modificación del eje simple, utilizada actualmente
en cultivos hidropónicos, es la llamada “cosecha del primer racimo”. Utiliza una
variedad precoz, de hojas pequeñas y una buena cuaja de frutos, con lo cual durante el
año se pueden llegar a realizar cuatro a cinco transplantes dejando un solo racimo
(WATANABLE et al., 1999). De acuerdo con estos autores las ventajas obtenidas
con este sistema serían la mayor posibilidad de ahorro en mano de obra, facilitar
labores de cultivo y obtener una mejor calidad del fruto.
El sistema que deja un brote lateral por planta, consiste en permitir el crecimiento de
uno de los brotes axilares a partir de la segunda y tercera hoja tras la primera
inflorescencia, pudiendo así disponer de dos tallos. Otra forma de realizarlo es a
través del método “Hardy”, en el cual se despunta el eje principal dos o tres hojas
arriba de la primera inflorescencia y se eligen los brotes laterales que salen de las
hojas opuestas (CASTILLA, 1995). Una variante de estos métodos es dejar el brote
sobre el primer racimo floral y cuando en este brote se ha formado la primera
inflorescencia se elimina la porción terminal del tallo sobre la segunda hoja del brote.

Esta modificación permite obtener una producción adicional entre el primer y
segundo racimo del eje principal (ESCAFF, 1992) (Figura 1). Otro sistema de
conducción a dos ejes es el utilizado en la región española de Murcia, en donde se
despunta el eje principal después de formadas dos hojas verdaderas y se dejan crecer
paralelamente los brotes axilares (ALLENDES, 2003)∗.
GONZÁLEZ (1994) señala que con el sistema de conducción a dos tallos se
disminuye el porcentaje de producción precoz, ya que lo habitual es que el primer
racimo del eje secundario recién se forma cuando el eje principal ya está formando el
tercer racimo. En cultivos hidropónicos conducidos a dos ejes se ha observado un
similar nivel de productividad de fruta en comparación a un cultivo guiado a un solo
eje, aunque el tiempo de cosecha fue más extenso y tardío (SAKAMOTO,
WATANABLE y OKANO, 2000).
SAKAMOTO, WATANABLE y OKANO (2000) coinciden en que la conducción a
dos brotes laterales es conveniente en reemplazo a la de un eje, lo que permite reducir
el número de plantines utilizados para establecer el cultivo. Esto se traduce en la
compra de menos plantas por hectárea (COCKSHULL, HO y FENLON, 2001).
WATANABLE et al. (1999) demuestran que los cultivares de verano-otoño son
ideales para un cultivo de verano conducido a múltiples ejes, ya que presentan un
mayor rendimiento de fruta que los cultivares de invierno-primavera. En un cultivo de
invierno la tendencia es opuesta.
Otra forma de conducir los brotes axilares es dejando tres ejes. HERNÁNDEZ
(2000), utilizando plantas injertadas sobre el portainjerto He-man, demostró que
plantas guiadas en tres ejes logran un 74 % de los frutos de calidad extra y primera,
en comparación a un cultivo guiado a un solo eje en el que se obtiene sólo un 67 %. ∗ ALLENDES, H. Ing. Agr. 2003. Universidad Católica de Valparaíso, Facultad de Agronomía. Comunicación personal.

FIGURA 1: Representación esquemática de diferentes sistemas de conducción a múltiples ejes: a) Conducción a eje simple. b) Conducción a doble eje tradicional. c) Conducción a doble eje modificado. d) Método de conducción Hardy. e) Conducción a tres ejes.
a b c d
e = Inflorescencia

Sin embargo, el inicio de cosecha de un cultivo guiado a tres ejes se retrasa una
semana y media con respecto al cultivo guiado a un eje.
Al dejar más de cuatro brotes laterales se afecta el crecimiento vegetativo y el
rendimiento del cultivo. También se disminuye el diámetro del tallo y su vigor, el
número de frutos por metro cuadrado decae, la cosecha comienza más tarde y lleva a
una significativa baja producción (NAVARRETE y JEANNEQUIN, 2000). Otros
tipos de problemas que se presentan al dejar más brazos por planta son: el aumento de
mano de obra para realizar las labores de deshoje y el aumento del volumen foliar y
por consiguiente, la mayor incidencia de enfermedades (ESCAFF, 1993).Según
GEINSENBERG y STEWART (1986) el aumento de ejes laterales reduce el tamaño
del fruto y se ve limitado por el vigor de la planta. Por esto, la densidad poblacional
debiera adecuarse al tipo de conducción previsto, aumentando el marco de plantación
si se conduce a más de dos ejes por planta.
2.9.2. Aumento del número de ejes durante el desarrollo del cultivo:
COCKSHULL y HO (1995) utilizaron densidades iniciales de 3.06 plantas·m-2 y a
través de la toma de un brote lateral en el tiempo llegaron a 4.07 tallos·m-2 en
sistemas hidropónicos. Dicho manejo indujo una disminución significativa del
promedio del peso del fruto y una reducción en los calibres de primera calidad,
mientras se incrementa la cantidad de frutos por unidad de superficie.
Una uniformidad del tamaño del fruto y un aumento del número de frutos cosechados
por unidad de superficie se logra plantando el cultivo a una baja densidad inicial (2.04
plantas·m-2) y aumentando la densidad de plantas (3.06 plantas·m-2) a través de la
toma de un brote lateral (COCKSHULL, HO y FENLON, 2001; COCKSHULL y
HO, 1995). Sin embargo, estos frutos adicionales no pueden ser cosechados hasta por
lo menos 80 días después que los brotes laterales han sido formados (COCKSHULL

y HO, 1995). Cuando los brotes laterales son formados tempranamente, alrededor de
la tercera semana del cultivo, una alta proporción de fruta comercializable fue de
primera calidad en todos los periodos de cosecha (COCKSHULL, HO y FENLON,
2001).
El número de frutos por unidad de superficie en un cultivo que se le toma un brote
lateral en el tiempo, aumenta en forma paralela con el incremento de radiación solar
de la estación. Además, en estos beneficios interviene el incremento del área foliar,
que contribuye a un aumento del índice de área foliar (IAF) produciendo más
asimilados por unidad de superficie. Esto último puede proteger a la fruta de la
radiación solar directa y por lo tanto, reducir desórdenes de madurez y permitir altas
producciones de fruta comercializable a lo largo del cultivo (COCKSHULL, HO y
FENLON, 2001).
2.10. Componentes de la calidad del fruto:
La calidad del fruto en función de la densidad de plantación y de tallos está
principalmente relacionada con su tamaño y peso. Otros factores que influyen la
calidad son: color, forma, ausencia de defectos, firmeza y sabor (CASTILLA, 1995).
En cultivos de tomate en alta densidad se ha observado que el diámetro y peso del
fruto son menores que en baja densidad (COCKSHULL, HO y FENOL, 2000) Un
método alternativo de cultivo es establecer la plantación en baja densidad y
posteriormente incrementar la densidad a través de la toma de un brote lateral en el
tiempo. Esto produce uniformidad en el peso y tamaño del fruto, disminuyendo
significativamente la cantidad de frutos de baja calidad (COCKSHULL y HO, 1995).
Es importante mencionar que la calidad organoléptica del fruto no se ha evaluado en
los ensayos llevados a múltiples ejes.

El color externo es el resultado de la pigmentación de la pulpa y la piel. El color
preferido por los consumidores es el rojo profundo y de tonalidad uniforme; a su vez,
la apreciación del estado de maduración se suele basar en el color (GRIERSON y
KADER, 1986). El deshojado en el periodo de maduración del fruto contribuye a
uniformar el color, induciendo precocidad en la recolección (CASTILLA, 1995).
Existen métodos subjetivos desarrollados para clasificar los estados de madurez del
tomate, tales como las escalas de clasificación y cartas de color, pero de muy difícil
aplicación (GRIERSON y KADER, 1986). KADER et al., (1978) señalan que los
métodos objetivos para evaluar el color de los frutos incluyen técnicas de medición de
tramitancia y reflexión de la luz.
La firmeza es muy variable entre cultivares, siendo más blandos los multiloculares
que los biloculares. Obviamente esta característica se ve influenciada por el estado de
madurez y por las condiciones del cultivo (CASTILLA, 1995).
El contenido de azúcares, ácidos y sus interacciones determinan el sabor del tomate.
Los principales azúcares son glucosa y fructosa que constituyen el 65 % de los
sólidos solubles (NUEZ, 1995). Dentro de los ácidos, los principales son el cítrico y
el málico. En conjunto representan entre un 80 a 90 % de la acidez total, ésta suele
oscilar entre 0.35 y 0.4 g/100 cc de jugo (DINAR y STEVENS, 1981). Altos niveles
de azúcares y relativamente altos niveles de ácidos se requieren para un buen sabor.
Altos contenido de ácidos y bajos de azúcares producen un tomate acre, mientras que
una situación inversa produce tomates de muy suave sabor. Cuando los azúcares y
ácidos son bajos, el resultado es un tomate insípido (GRIERSON y KADER, 1986).
Valores de pH inferiores a 4.4 y contenidos de azúcares superiores al 4-4.5 % son
necesarios para un buen sabor (DINAR y STEVENS, 1981).

3. MATERIALES Y MÉTODOS
3.1. Ubicación del ensayo:
El ensayo se realizó en la parcela 36 perteneciente a la Sociedad Agrícola Los
Rosales de Santa Julia, ubicada en la localidad de San Isidro 32º 50` latitud Sur, 71º
13` longitud Oeste, Provincia de Quillota, Quinta Región de Valparaíso, durante el
periodo comprendido entre el 3 de septiembre del 2002 y el 31 de enero del 2003.
3.2. Características de clima y suelo:
El clima de la zona de Quillota está clasificado como Mediterráneo Marino,
agroclima Quillota. La temperatura media anual es de 15.3 ºC, presentando una
máxima media del mes más cálido (enero) de 27°C y una mínima media del mes más
frío (julio) de 5.5 °C. El período libre de heladas aprovechable corresponde a nueve
meses, desde septiembre a mayo. La suma anual de temperaturas, base 5°C es de
3.700 grados días y 10°C, 1900 grados días. Las horas fríos de marzo a noviembre
llegan a 500. La temperatura media mensual se mantiene sobre los 10°C. El régimen
hídrico se caracteriza por una precipitación anual de 437 mm, siendo el mes de junio
el más lluvioso con 125 mm. La estación seca tiene un promedio de ocho meses
(NOVOA et al., 1989).
El suelo del lugar donde se desarrolló el ensayo pertenece a la serie San Isidro.
Corresponde a suelos de origen lacustrino, profundos; de textura franco arcillosa, de
color grisáceo muy oscuro en la superficie y textura arcillosa, de color pardo grisáceo
oscuro y negro en el tono 10 YR en profundidad. Suelo de topografía plana y
deprimida, de permeabilidad lenta a muy lenta y de drenaje imperfecto. Presenta
reacción violenta al HCl en todo el perfil (CIREN-CORFO, 1985).

3.3. Material vegetal:
Se utilizaron plantas de tomate de un cultivo comercial de la variedad ASTONA
perteneciente a la empresa Sunseeds Chile, híbrido de tipo indeterminado, larga vida,
vigoroso, de maduración media para invernadero o campo abierto. Sus frutos son
lisos, ligeramente achatados, con peso medio de 180-220 g y de un buen color rojo.
Es resistente a Verticillium 1, Fusarium 1 y 2, Virus del Mosaico del Tabaco (TMV)
y nemátodos (SUNSEEDS, 2003).
Las plantas se establecieron bajo las condiciones de un invernadero frío, a través del
sistema almácigo-transplante con un volumen de contenedor de 10 cm3. El
transplante se realizó el día 3 de septiembre del 2002. Las plantas se dispusieron en
V, la distancia sobre la hilera en la mesa de plantación fue de 0.23 m y la distancia
entre mesas de1.40 m.
Los invernaderos usados son de madera y están cubiertos con un polietileno de 200
micras, de dos temporadas. Corresponden al tipo dos aguas, de 4.6 m de alto, 30 m de
largo y 7.0 m de ancho.
Los manejos culturales se realizaron de acuerdo a las técnicas actuales que se utilizan
para la producción forzada de tomates bajo plástico en la región (MENDOZA, 1992).
La poda de brotes laterales se realizó de forma manual y cuando estos alcanzaban
entre los 3 y 6 cm aproximados de longitud. El despunte de las plantas fue de acuerdo
al tipo de tratamiento a que fueron sometidas. La fertirrigación, a través de dos cintas
de riego por mesa se realizó de acuerdo a la solución nutritiva modificada de Steiner.

Es importante señalar que el suelo de los invernaderos fue fumigado por última vez
con Bromuro de Metilo a inicios de la temporada 2001, lo que significa que para este
ensayo el suelo no ha sido fumigado.
3.4. Tratamientos:
El manejo de conducción consistió en dejar dos ejes por planta, a través de la toma de
un brote lateral desde la axila de la hoja dispuesta inmediatamente debajo de la
primera inflorescencia de la planta. Los brotes laterales no fueron podados hasta la
confirmación de la existencia de un brote lateral fuerte debajo de la primera
inflorescencia, para así asegurar su formación. El despunte del primer eje se realizó
sobre el quinto racimo floral y el despunte del segundo eje sobre el segundo racimo
floral. El primer eje se deshojó de acuerdo al sistema utilizado en el predio, el que
consistió en la eliminación de la segunda hoja después de cada inflorescencia. Al
segundo eje se le realizó una poda de hojas que consistió en arrancar todas aquéllas
que se encontraban debajo de la primera hoja dispuesta bajo el primer racimo floral.
Este manejo se realizó cuando los frutos del segundo racimo del segundo eje tenían
un diámetro ecuatorial promedio de 3.5 cm. Las plantas conducidas a un solo eje
fueron despuntadas sobre el séptimo racimo floral y los brotes laterales fueron
eliminados con los mismos criterios usados en las plantas conducidas a dos ejes.
Los dos tratamientos a que fueron sometidas las plantas, consistieron en conducir las
plantas a dos ejes para aumentar la densidad poblacional por unidad de superficie y
un tratamiento testigo conducido a un solo eje. Los tratamientos experimentados
fueron los siguientes:
T0: Testigo, plantas conducidas a un eje (2.5 ejes·m-2).
T1: Plantas conducidas a dos ejes intercaladas con plantas a un solo eje (3.8 ejes·m-2).
T2: Plantas conducidas a dos ejes (5.0 ejes·m-2).

3.5. Unidad experimental:
La unidad experimental fue representada por parcelas rectangulares de 29.9 m2 (4.6 m
de ancho y 6.5 m de largo), compuesta por dos mesas de plantación. En cada extremo
de las mesas se dejaron 2 m de borde. La unidad de muestreo consistió en una sección
comprendida por 14 plantas, distribuidas en la parte central de las dos mesas (siete
plantas en cada mesa) (Anexo 1).
3.6. Diseño experimental:
El diseño estadístico consistió en un Diseño Completamente al Azar, compuesto por
tres tratamientos y cuatro repeticiones.
3.7. Análisis de los resultados:
Se realizó un análisis de varianza, a través de una tabla ANDEVA. Cuando el valor P
asociado al estadístico de prueba fue ≤ 0.05 se utilizó el Test de Tukey, con un α =
0.05. Los datos se analizaron con el software Minitab (Minitab Inc., Pennsylvania,
EE.UU.).
3.8. Mediciones:
3.8.1. Rendimiento y calibre de los frutos:
La frecuencia de cosecha de los frutos de tomate fue de dos ó tres veces por semana,
desde los 92 días hasta los 144 días después del transplante. El índice de cosecha
correspondió a tomate rosado, lo que equivale a una presencia del 30 % a 60 % de la
superficie del fruto con una coloración anaranjada o roja. Internamente el fruto es
naranja o rojo y la matriz placental se hace acuosa (KRARUP, 2002). Los frutos

fueron pesados individualmente. La clasificación de calibres fue de acuerdo a los
siguientes criterios:
Extra: Fruto con un peso mayor a 250 g.
Primera: Fruto con un peso entre 150 y 250 g.
Segunda: Fruto con un peso entre 100 y 150 g.
Tercera: Fruto con un peso entre 80 y 100 g.
Rosca: Fruto no comercial con los lóculos marcados.
3.8.2. Parámetros vegetativos:
Altura de las plantas: Esta medición se realizó una vez por semana a cuatro plantas
por parcela, desde 19 días hasta 97 días después del transplante, a todos los
tratamientos. Se midió la altura de ambos ejes: desde el cuello del tallo principal hasta
su ápice y desde la bifurcación del eje secundario hasta el ápice. Las mediciones se
realizaron hasta los 20 días después de la fecha del despunte (24 de noviembre 2002).
Diámetro del tallo: La medición se realizó bajo la primera inflorescencia de cada
eje, semanalmente, a una muestra de cuatro plantas por parcela. La medición se hizo
desde 19 días hasta 125 días después del transplante.
Área foliar (A): Las mediciones se realizaron tres veces en la temporada, a un
número de cuatro plantas por parcela, basándose en el ancho de la hoja. Se midió la
expansión de la hoja del medio de cada simpodio, estimándose el área a través de la
siguiente fórmula, definida por SCHWARTZ y KLÄRING (2001):
Área foliar (A) = 0,203Lw1,674

Donde:
Lw: Ancho de la hoja.
3.8.3. Parámetros de calidad:
La medición de estos parámetros se obtuvo de una muestra promedio de siete frutos
por racimo, elegidos completamente al azar al momento de la cosecha y fueron
analizados en el Laboratorio de Post-Cosecha de la Escuela de Agronomía,
Universidad Católica de Valparaíso.
Diámetro de frutos: Se realizó una medición del diámetro polar y ecuatorial de los
frutos.
Color: La medición se realizó en el exterior del fruto y en su pulpa, la cual fue
obtenida procesando los frutos en una juguera. Los datos se obtuvieron de tres puntos
equidistantes y tres repeticiones en el ecuador del fruto y en la pulpa. El instrumento
utilizado fue un colorímetro por reflexión MINOLTA modelo CR-200, el que arroja
valores L*, a*, b*, los cuales están basados en la tabla de colores de Hunter. Estos
valores indican: rango de luminosidad (L) que va desde el negro al blanco, el rango
desde el color verde hasta el rojo (a) y el rango del azul hasta el amarillo (b)
(McGUIRE, 1992). Una vez obtenidos los valores promedio se calculó Croma que
equivale a la saturación y el Ángulo de Matiz al tono, a través de las siguientes
fórmulas (McGUIRE, 1992):
22)( baCCroma +=

Ángulo de matiz (hº) ( )abarctg /=
El ángulo de matiz tiene valores que van de 0º (rojo-púrpura), 90º (amarillo), 180º
(verde azuloso). Los sistemas computacionales arrojan los resultados en radianes, así
que para su correcta interpretación se debe ajustar la fórmula anterior a la siguiente
(McGUIRE, 1992):
Ángulo de matiz (hº) 360*2
⎟⎟⎟⎟
⎠
⎞
⎜⎜⎜⎜
⎝
⎛⎟⎠⎞
⎜⎝⎛
=π
abarctg
pH: Las mediciones se realizaron al jugo obtenido por decantación de la pulpa de los
frutos, por medio de un pH-metro SCHOTT modelo Handylab 1.
Sólidos solubles: La medición se le realizó al jugo decantado de los frutos, a través
de un refractómetro manual de 10° Brix, marca ATAGO modelo ATC-1.
Acidez titulable: Este parámetro fue obtenido midiendo el gasto de NaOH hasta
alcanzar un valor de pH 8,2 en una muestra de 20 ml de jugo decantado y 80ml de
agua destilada. Se utilizó la siguiente fórmula (ASSOCIATION OF OFFICIAL
ANALITYCAL CHEMISTS, 1990):
% Ac.Cítrico = gasto de NaOH * concentración NaOH (N) * 6.41
ml solución

4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Rendimiento:
El rendimiento de frutos es el factor clave que determina el éxito económico del
cultivo. El uso en cultivos de tomate bajo invernadero de tecnologías apropiadas ha
permitido incrementar los rendimientos en forma importante en nuestro país. Es así
que cultivos protegidos pueden alcanzar rendimientos que oscilan entre las 100 a 180
ton·ha-1 de acuerdo a su nivel de eficiencia, manejos culturales, condiciones
climáticas, sanitarias, entre muchas otras (URRUTIA, 2000). Por otro lado, el
rendimiento está directamente relacionado con el calibre y el número de frutos. A
medida que el calibre de los frutos es mayor el número de frutos va disminuyendo
(KRARUP, 1992).
En el presente ensayo al evaluar el rendimiento total de los diferentes tratamientos no
se encontró diferencias significativas entre ellos (Cuadro 1). Esta situación indica que
la producción de las plantas no varió al ser conducidas con uno o dos ejes. La
tendencia podría ser diferente si el número de racimos dejados en el primer o segundo
eje de los tratamientos hubiera sido mayor, es decir, aumentar el largo del cultivo.
En el ensayo realizado por MARCOTTI (1998) en la localidad de San Isidro, se
observa que el rendimiento de frutos, en plantas conducidas a eje simple y doble
manteniendo la densidad de plantas en 29.200 plantas·ha-1, se ve reducido en el
tratamiento conducido a eje doble. En cambio, a una densidad de plantas menor
(21.900 plantas·ha-1), la conducción a dos ejes presenta un mejor comportamiento en
el rendimiento total de frutos. Esto se explica por un aumento de la eficiencia
fotosintética de cada planta, el equilibrio sink-source de la planta es más estable que

en la conducción a un eje y de esta forma se suple mejor el llenado de los frutos
(SALINAS, RAMÍREZ y OSPINA, 1994).
De acuerdo a COCKSHULL y HO (1995), la toma de un brote lateral llega a
contribuir a una producción total adicional, pero el efecto no es significativo. Los
ensayos consistieron en la toma de un brote lateral en un cultivo de baja densidad
(2.04 plantas.m-2) hasta alcanzar una densidad de 3.06 ejes.m-2 y la toma de un brote
lateral a una alta densidad de 3.06 plantas.m-2 hasta alcanzar 4.07 ejes.m-2. La adición
de brotes laterales fue contrastada con densidades fijas de 2.04 plantas.m-2 y 3.06
plantas.m-2.
Similares resultados se obtuvieron en los ensayos realizados por COCKSHULL, HO
y FENLON (2000), en los cuales se evaluaron diferentes fechas para la toma de un
brote lateral en cultivos de largo aliento y fueron comparados con tratamientos
testigos de densidades bajas fijas (2.04 plantas.m-2) y densidades altas fijas (3.06
plantas.m-2) en el tiempo. Los resultados obtenidos mostraron un patrón de
producción consistente con los tratamientos testigos, pero la tendencia obtenida de
una mayor producción total de frutos comercializables no fue estadísticamente
significativa. Para los ensayos realizados por HERNÁNDEZ (2000), en plantas
injertadas y conducidas a dos y tres ejes, pero manteniendo las densidades de ejes por
unidad de área entre los tratamientos hasta el final del cultivo, los rendimientos
totales de frutos sí disminuyeron significativamente.
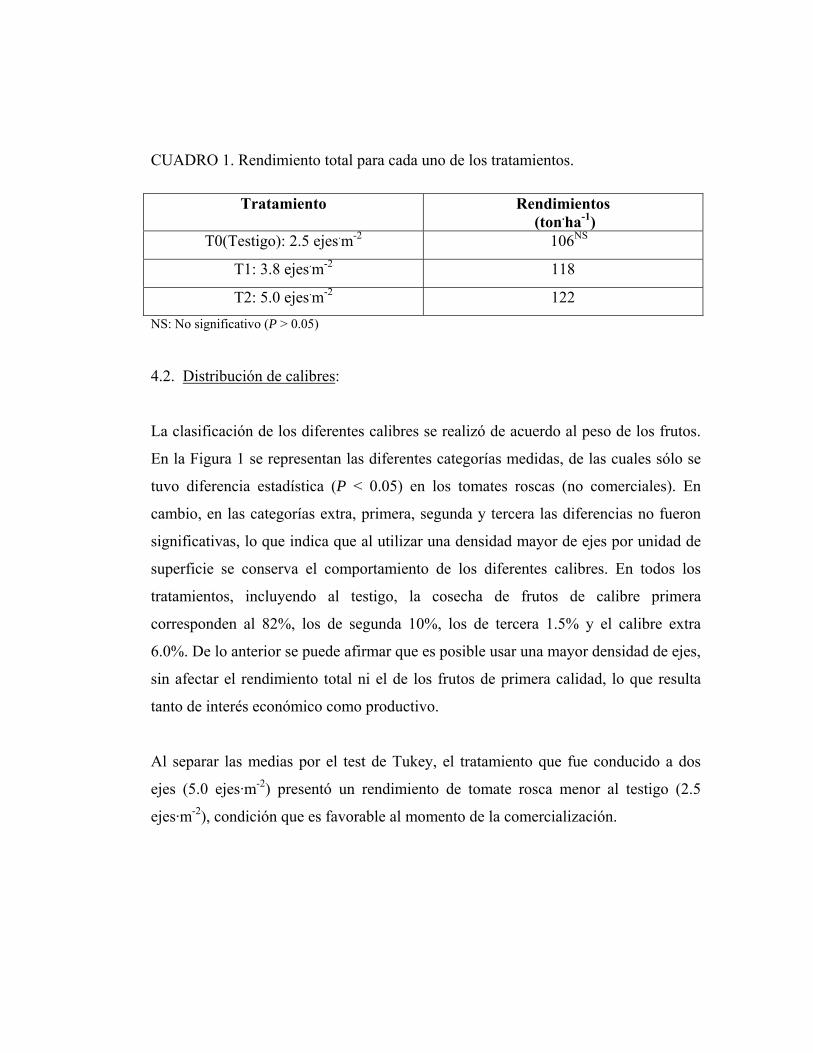
CUADRO 1. Rendimiento total para cada uno de los tratamientos.
Tratamiento Rendimientos (ton.ha-1)
T0(Testigo): 2.5 ejes.m-2 106NS
T1: 3.8 ejes.m-2 118
T2: 5.0 ejes.m-2 122 NS: No significativo (P > 0.05)
4.2. Distribución de calibres:
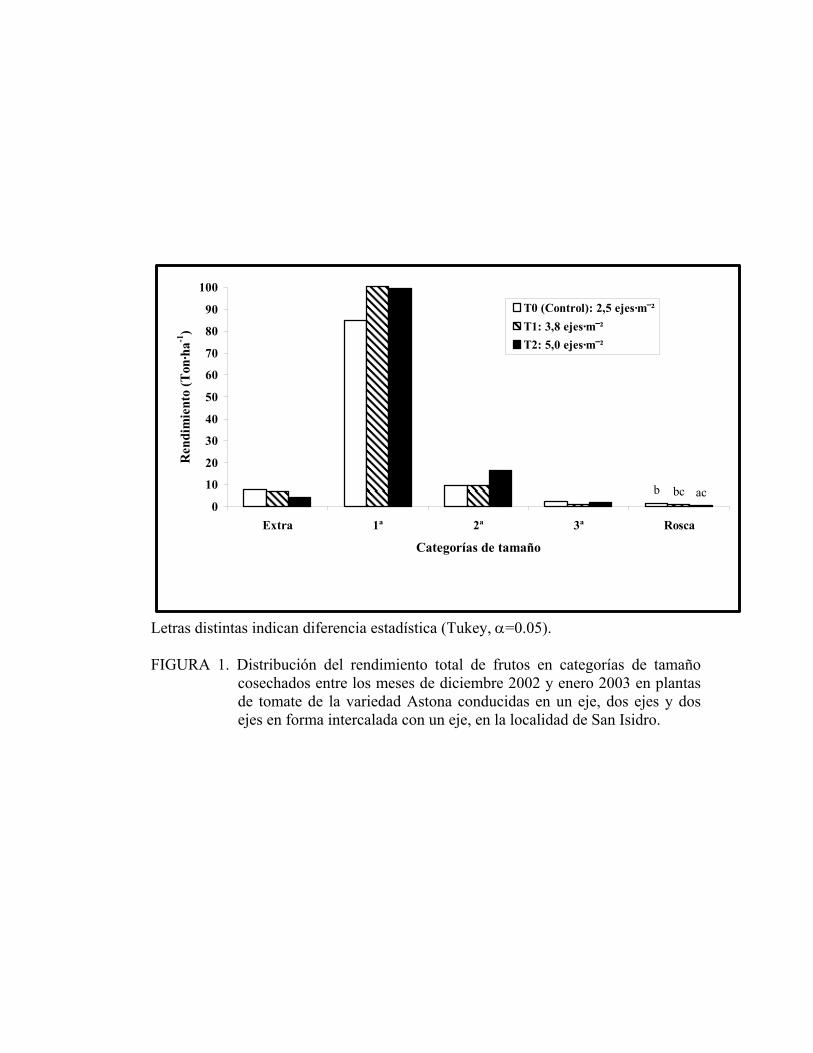
La clasificación de los diferentes calibres se realizó de acuerdo al peso de los frutos.
En la Figura 1 se representan las diferentes categorías medidas, de las cuales sólo se
tuvo diferencia estadística (P < 0.05) en los tomates roscas (no comerciales). En
cambio, en las categorías extra, primera, segunda y tercera las diferencias no fueron
significativas, lo que indica que al utilizar una densidad mayor de ejes por unidad de
superficie se conserva el comportamiento de los diferentes calibres. En todos los
tratamientos, incluyendo al testigo, la cosecha de frutos de calibre primera
corresponden al 82%, los de segunda 10%, los de tercera 1.5% y el calibre extra
6.0%. De lo anterior se puede afirmar que es posible usar una mayor densidad de ejes,
sin afectar el rendimiento total ni el de los frutos de primera calidad, lo que resulta
tanto de interés económico como productivo.
Al separar las medias por el test de Tukey, el tratamiento que fue conducido a dos
ejes (5.0 ejes·m-2) presentó un rendimiento de tomate rosca menor al testigo (2.5
ejes·m-2), condición que es favorable al momento de la comercialización.
b bc ac0
10
20
30
40
50
60
70
80
90
100
Extra 1ª 2ª 3ª Rosca
Categorías de tamaño
Ren
dim
ient
o (T
on·h
a-1)
T0 (Control): 2,5 ejes·m⎯²T1: 3,8 ejes·m⎯²T2: 5,0 ejes·m⎯²
Letras distintas indican diferencia estadística (Tukey, α=0.05). FIGURA 1. Distribución del rendimiento total de frutos en categorías de tamaño cosechados entre los meses de diciembre 2002 y enero 2003 en plantas de tomate de la variedad Astona conducidas en un eje, dos ejes y dos ejes en forma intercalada con un eje, en la localidad de San Isidro.

La toma de brotes laterales tuvo efectos beneficiosos sobre la reducción del desorden
fisiológico llamado rosca, que se caracteriza por frutos de un tamaño mayor al
normal, presentan hendiduras y protuberancias a lo largo del fruto. Es atribuido a una
baja temperatura (menor o igual a 10ºC) durante la formación de la flor, lo que induce
a una mala diferenciación de los carpelos, los que se unen en algunos sectores
(OYANEDEL, 2002). Por lo tanto, se sugiere que el aumento de la densidad de ejes
por metro cuadrado producida por el tratamiento 2, puede haber causado una mayor
estabilidad térmica dentro de la canopia y haber aumentado la temperatura mínima
media, produciendo así la disminución de este desorden en los periodos de mayor
susceptibilidad. Por otro lado, el mayor vigor expresado por el tratamiento de
conducción a un eje, bajo las mismas condiciones de baja temperatura que el resto de
los tratamientos, pudo haber influido en este resultado.
De acuerdo a los ensayos realizados por MARCOTTI (1998), no se detectaron
diferencias en el rendimiento de los calibres de primera (63%), segunda (35%) y
tercera (2%) categoría entre plantas conducidas a un eje y doble eje, en densidades de
plantación altas y bajas. Sin embargo, en ensayos realizados con plantas injertadas
conducidas a uno, dos y tres ejes producen un 67%, 71% y 74%, respectivamente, de
frutos de categoría extra y primera; a su vez las plantas sin injertar producen un 64%
de estas categorías (HERNÁNDEZ, 2000).
COCKSHULL y HO (1995) observaron que la producción de frutos de primera
disminuyó, principalmente porque la presencia de brotes laterales aumentó la
producción de frutos de segunda. En los estudios realizados por COCKSHULL, HO y
FENLON (2000), se encontró una disminución de los calibres de primera en los
tratamientos con toma de brotes laterales, pero se produjo una uniformidad del
tamaño de los frutos en todos los periodos de cosecha.

En estudios realizados por SALINAS, RAMÍREZ y OSPINA (1994) se observa una
respuesta favorable de las plantas conducidas a diferentes tratamientos de conducción
de ejes frente a la distribución de calibres. En plantas conducidas a uno y dos ejes se
aprecia el mayor porcentaje de calibres de primera calidad, en comparación a plantas
que no presentan ningún tipo de conducción al no haber sido podadas. La relación
entre el porcentaje de primera decrece a medida que el porcentaje de tercera aumenta,
para los tratamientos sin poda. En cambio, en plantas conducidas a uno y dos ejes el
porcentaje de primera se mantiene alto en relación con los porcentajes de segunda y
tercera.
4.3. Distribución de la producción en el tiempo:
El periodo de cosecha de frutos se extendió durante nueve semanas, comenzando los
primeros días de diciembre y culminado a fines del mes de enero. En las primeras
cuatro semanas de cosecha (diciembre) se cosechó el 40 % de los frutos, en las dos
semanas siguientes un 43 % y en las restantes el 17 %. El resultado del análisis
estadístico indica que en los primeros 15 días de cosecha no existen diferencias
significativas entre los tratamientos, en cambio en los tres periodos siguientes las
diferencias sí lo son (Figura 2).
En el periodo de 108 a 123 días después del transplante el tratamiento 2 es mayor que
el tratamiento 1 y el testigo. En los días 124 a 139 después del transplante se observa
que los dos tratamientos de conducción a múltiples ejes son mayores al testigo. Sin
embargo, en el periodo final de cosecha (140-155 días después del transplante), el
rendimiento del testigo fue mayor que los tratamientos de conducción. Por lo tanto, se
consiguió concentrar la cosecha en un periodo de tiempo menor.
En experimentos realizados en cultivos de largo aliento se ha observado que la toma
de brotes laterales baja la producción temprana de fruta comercializable, mientras que

incrementa el número de frutos producidos por unidad de superficie y aumenta la
producción final (COCKSHULL y HO, 1995). La pérdida de precocidad se ve
reflejada en la cosecha de frutos de los brotes laterales, la que se produce sólo
después de 80 días después de la formación del eje (COCKSHULL, HO y FENLON,
2001). De acuerdo a HERNÁNDEZ (2000) en plantas injertadas conducidas a uno y
dos ejes se aprecia una leve diferencia sólo en la tercera y cuarta semana de cosecha,
la que se debe al retraso de maduración de los frutos de los primeros racimos del
segundo eje.
4.4. Crecimiento vegetativo:
El crecimiento vegetativo es una interacción de las condiciones fisiológicas de la
planta y su respuesta a las condiciones medioambientales. El desarrollo vegetativo de
una planta depende de una serie de interacciones de factores, entre los que se pueden
mencionar la variedad, la radiación solar, la temperatura, la nutrición, el suministro
de agua, la concentración de CO2 y el estado sanitario de la planta (NAVARRETE y
JEANNEQUIN, 2000).
En este ensayo no se observaron diferencias estadísticas (P > 0.05) en los parámetros
de altura de la planta y diámetro del tallo, tanto para el primer eje como para el
segundo. En ambos caso se realizó un análisis de varianza para las medias de las tasas
de crecimiento relativo (TCR) en todas las fechas en que fueron medidas (Figuras 3 y
4). Esto apunta a que la alta densidad de ejes por metro cuadrado no afecta el
crecimiento vegetativo de las plantas de los tratamientos.
La tasa relativa de crecimiento de la altura y el diámetro del tallo disminuyen en el
tiempo. Esto se ve explicado porque a medida que los frutos cuajan, pasan a ser un
sink más fuerte de fotoasimilados que el tallo, el ápice o las raíces. Por lo tanto, una

a
b
bc c
a
b
c
a
a
0
10
20
30
40
50
60
92-107 108-123 124-139 140-155
Días después del transplante
% to
tal d
e co
sech
a
T0 (Control): 2,5 ejes·m⎯²T1: 3,8 ejes·m⎯²T2: 5,0 ejes·m⎯²
Letras distintas para cada rango de fechas indican diferencia estadística (Tukey, α=0.05) FIGURA 2. Porcentaje de frutos cosechados entre los meses de diciembre 2002 y enero 2003 en plantas de tomate de la variedad Astona conducidas en un eje, dos ejes y dos ejes en forma intercalada con un eje, en la localidad de San Isidro.

vez que los frutos comienzan a crecer la tasa de crecimiento vegetativo de la planta
llega a su valor mínimo (HO y HEWITT, 1986).
4.5. Área foliar:
Los valores obtenidos indican que el área foliar de los tratamientos en el día 50
después del transplante no presentan diferencias significativas con el testigo. En
cambio, en los días 104 y 135 después del transplante el tratamiento 1 presenta un
área foliar mayor al tratamiento 2 y al testigo (Figura 5).
La toma de brotes laterales en los tratamientos produjo un área foliar adicional en el
tratamiento 1, debido al tipo de conducción de la planta, la cual debe haber provocado
una competencia por radiación solar con las hojas del eje principal y además, en
estados tempranos de crecimiento de los brotes debe haber provocado una
competencia por asimilados entre el brote lateral y los frutos del tallo principal (HO,
1996). De la investigación realizada por COCKSHULL y HO (1995) se desprende
que el incremento del índice de área foliar, como resultado de la adhesión de brotes
laterales, fue probablemente la principal causa del incremento del número de frutos
producidos. De acuerdo a COCKSHULL, HO y FENLON (2000), uno de los
beneficios del incremento del área foliar es la mayor producción potencial de
asimilados por unidad de área y la protección de los frutos de la radiación solar
directa para poder reducir la incidencia de desórdenes de maduración.
4.6. Calidad del fruto:
La preferencia del consumidor por ciertos tipos de tomate se debe a percepciones de
apariencia, gusto, textura y valor nutricional, tanto como por el costo del producto
(HO, 2000).

0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
2,00
27 34 41 48 55 62 69 76 83*
Días después del transplante
Tas
a re
lativ
a de
cre
cim
ient
o de
altu
ra d
el
eje
prim
ario
(cm
·cm
-1·d
-1)
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
2,00
Tas
a rl
ativ
a de
cre
cim
ient
o de
altu
ra d
el e
je
secu
ndar
io (c
m·c
m-1
·d-1
)
T0 (Control): 2,5 ejes·m⎯²T1: 3,8 ejes·m⎯²T2: 5,0 ejes·m⎯²T1 (eje 2º): 3,8 ejes·m⎯²T2 (eje 2º): 5,0 ejes·m⎯²
FIGURA 3. Tasa relativa de crecimiento de la altura, medida entre los meses de octubre y noviembre 2002, en plantas de tomate de la variedad Astona conducidas en un eje, dos ejes y dos ejes en forma intercalada con un eje, en la localidad de San Isidro. * El despunte se realizó a los 83 días después del transplante.

0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
27 34 41 48 55 62 69 76 83 97 104 111 118
Días después del transplante
Tas
a re
lativ
a de
cre
cim
ient
o de
l eje
pr
imar
io (c
m·c
m-1
·d-1
)
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
Tas
a re
lativ
a de
cre
cim
ient
o de
l eje
se
cund
ario
(cm
·cm
-1·d
-1)
T0 (Control): 2,5 ejes·m⎯²T1: 3,8 ejes·m⎯²T2: 5,0 ejes·m⎯²T1 (eje 2º): 3,8 ejes·m⎯²T2 (eje 2º): 5,0 ejes·m⎯²
FIGURA 4. Tasa relativa de crecimiento de diámetro de tallo, medida entre los
meses de septiembre 2002 y enero 2003 en plantas de tomate de la variedad Astona conducidas en un eje, dos ejes y dos ejes en forma intercalada con un eje, en la localidad de San Isidro.

b
b
NS
a
a b
b
0
100
200
300
400
500
600
700
800
900
1000
1100
50 104 135
Días después del transplante
Áre
a fo
liar
eje
prim
ario
(cm
2 )
0
100
200
300
400
500
600
700
800
900
1000
1100
Áre
a fo
liar
eje
secu
ndar
io (c
m2 )
T0 (Control): 2,5 ejes·m⎯²T1: 3,8 ejes·m⎯²T2: 5,0 ejes·m⎯²
NS: No significativo (P > 0.05). Letras distintas indican diferencia estadística (Tukey, α=0.05). FIGURA 5. Área foliar promedio (cm2), medida entre los meses de octubre 2002 y enero 2003 en la localidad de San Isidro, en plantas de tomate de la variedad Astona conducidas en un eje, dos ejes y dos ejes en forma intercalada con un eje.

4.6.1 Peso promedio, diámetro polar y ecuatorial de los frutos:
En el parámetro de peso del fruto no se presentó diferencias estadísticas entre los
tratamientos (Cuadro 2), siendo el promedio de los frutos del ensayo de 179.76 g, el
cual se encuentra en el límite del rango esperado dado por la variedad utilizada,
ASTONA, la que fluctúa entre 180 a 220 g (SUNSEEDS, 2003). Debido a esto, no se
presentan signos de que el peso del fruto haya sido afectado por los tratamientos. Esto
no concuerda con lo expuesto por GEISEMBERG y STEWART (1986), los que
plantean que al aumentar el número de ejes por planta disminuye el peso del fruto.
CUADRO 2. Peso (gr) promedio del fruto para cada uno de los tratamientos.
Tratamiento Peso de frutos (g)
T0(Testigo): 2.5 ejes.m-2 182.38NS
T1: 3.8 ejes.m-2 181.42
T2: 5.0 ejes.m-2 175.48 NS: No significativo (P > 0.05).
Según COCKSHULL y HO (2000), la toma de brotes laterales disminuyó el
promedio del peso del fruto, redujo la variación mes a mes en el promedio del peso y
promovió un tamaño más uniforme durante el año. HERNÁNDEZ (2000) observó la
disminución del peso del fruto en plantas injertadas conducidas a dos y tres ejes con
respecto a plantas injertadas a un solo eje, pero el valor fue mayor que para plantas
sin injertar.
No se presentaron diferencias estadísticas entre los tratamientos con respecto a los
parámetros de diámetro polar y ecuatorial (Cuadro 3). El promedio del diámetro polar
de los frutos fue de 5.76 cm y el del diámetro ecuatorial de 7.5 cm.

CUADRO 3. Diámetro (cm) polar y ecuatorial promedio de los frutos para cada tratamiento.
Tratamiento Diámetro polar (cm) Diámetro ecuatorial (cm)
T0(Testigo): 2.5 ejes.m-2 5.8NS 7.6NS
T1: 3.8 ejes.m-2 5.8 7.5
T2: 5.0 ejes.m-2 5.7 7.4 NS: No significativo (P > 0.05).
4.6.2. pH y acidez titulable:
Los valores de pH no presentan diferencias significativas entre los dos tratamientos
de conducción y el testigo. Los valores obtenidos se encuentran dentro del rango
(menor a 4.4) necesario para un buen sabor (CASTILLA, 1995). En cuanto a la
acidez titulable, el tratamiento de 5.0 ejes.m-2 resulta ser significativamente superior
al tratamiento de 3.8 ejes.m-2 (Cuadro 4).
La acidez es un factor importante en el sabor del tomate. El tratamiento 2 presenta
una acidez titulable significativamente mayor al tratamiento 1, aunque las diferencias
entre estos valores se consideran insuficientes como para incidir en la calidad del
producto.
4.6.3. Sólidos solubles:
El típico sabor dulce está determinado principalmente por las concentraciones de
azúcares del fruto (sólidos solubles). La composición de azúcar del fruto depende del
cultivar y las concentraciones de luz y temperatura durante su crecimiento (HO,
2002). Los resultados promedios obtenidos de sólidos solubles presentan diferencias
estadísticas (CUADRO 5) entre el testigo y el tratamiento 1.
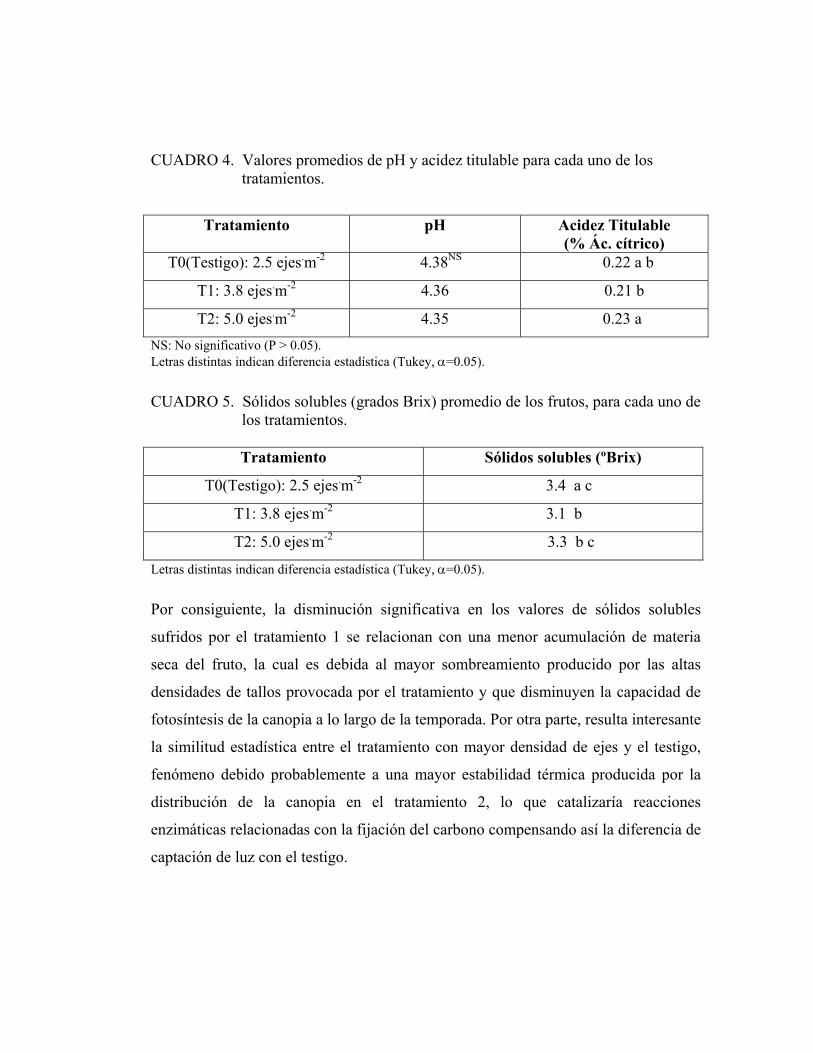
CUADRO 4. Valores promedios de pH y acidez titulable para cada uno de los tratamientos.
Tratamiento pH Acidez Titulable (% Ác. cítrico)
T0(Testigo): 2.5 ejes.m-2 4.38NS 0.22 a b
T1: 3.8 ejes.m-2 4.36 0.21 b
T2: 5.0 ejes.m-2 4.35 0.23 a NS: No significativo (P > 0.05). Letras distintas indican diferencia estadística (Tukey, α=0.05).
CUADRO 5. Sólidos solubles (grados Brix) promedio de los frutos, para cada uno de los tratamientos.
Tratamiento Sólidos solubles (ºBrix)
T0(Testigo): 2.5 ejes.m-2 3.4 a c
T1: 3.8 ejes.m-2 3.1 b
T2: 5.0 ejes.m-2 3.3 b c
Letras distintas indican diferencia estadística (Tukey, α=0.05).
Por consiguiente, la disminución significativa en los valores de sólidos solubles
sufridos por el tratamiento 1 se relacionan con una menor acumulación de materia
seca del fruto, la cual es debida al mayor sombreamiento producido por las altas
densidades de tallos provocada por el tratamiento y que disminuyen la capacidad de
fotosíntesis de la canopia a lo largo de la temporada. Por otra parte, resulta interesante
la similitud estadística entre el tratamiento con mayor densidad de ejes y el testigo,
fenómeno debido probablemente a una mayor estabilidad térmica producida por la
distribución de la canopia en el tratamiento 2, lo que catalizaría reacciones
enzimáticas relacionadas con la fijación del carbono compensando así la diferencia de
captación de luz con el testigo.

4.6.4. Color:
La apariencia del fruto, donde el color es uno de los principales parámetros, tiene el
efecto más inmediato y profundo sobre la elección del consumidor (HO, 2000). El
color del fruto del tomate está definido básicamente por dos pigmentos, ambos
carotenoides, el que representa mayor importancia es el licopeno y en menor medida
el β-caroteno, los cuales son producto de la degradación de la clorofila
(CHAMARRO, 1995).
El Cuadro 6 indica los resultados respecto a las mediciones del color externo e
interno efectuadas a cada tratamiento. El color externo no presentó diferencias
significativas entre los tratamientos. Con respecto al color interno, la luminosidad (L)
de los frutos del tratamiento 1 fue significativamente mayor al control (color más
claro). Esto representa una disminución de la calidad del fruto, lo que se traduce en
una pérdida de la preferencia por parte de los consumidores.
CUADRO 6. Análisis del color externo y color interno de los frutos de tomate en
cada tratamiento.
Color externo Color interno
Tratamiento L C hº L C hº T0: 2,5 ejes.m-2 208.7NS 49.5NS 30.5NS 59.4 a c 47.4NS 37.7NS
T1: 3,8 ejes.m-2 39.9 48.2 22.9 63.3 b 46.5 38.1 T2: 5,0 ejes.m-2 38.3 47.8 53.4 60.7 b c 47.5 36.9
NS: No significativo (P > 0.05). Letras distintas indican diferencia estadística (Tukey, α=0.05).

5. CONCLUSIONES
Al emplear dos sistemas de conducción a doble eje (3.8 y 5.0 ejes·m-2), se mantiene el
rendimiento total de los frutos con respecto a un cultivo de conducción simple (2.5
ejes·m-2) realizado bajo las condiciones edafo-climáticas de la localidad de San Isidro.
La distribución de las diferentes categorías de tamaño de los frutos no es influenciada
por el aumento de la densidad de ejes.
Los tratamientos con 3.8 y 5.0 ejes·m-2 produjeron una concentración de la cosecha,
con una mayor proporción de fruta cosechada en las semanas 5-6 que el testigo.
El crecimiento vegetativo, cuantificado como diámetro del tallo y altura de la planta,
no se ve afectado por el aumento de la densidad. Por el contrario, el área foliar es
mayor en plantas conducidas a doble eje, lo que sugiere una mayor producción de
asimilados por unidad superficie y la protección de los frutos de la radiación solar
directa.
La mayor parte de los parámetros de calidad del fruto (peso promedio, diámetro polar
y ecuatorial, pH y color externo) no son afectados por la toma de un brote lateral en el
tiempo. Por otro lado, la acidez titulable aumenta favorablemente al incrementar la
densidad a 5.0 ejes·m-2, mientras que los sólidos solubles y la luminosidad del color
interno del fruto disminuyen con densidades de 3.8 ejes·m-2, afectando
detrimentalmente la calidad del fruto.

6. RESUMEN
El ensayo se realizó en la localidad de San Isidro, provincia de Quillota, región de Valparaíso. El objetivo de este experimento fue evaluar el desempeño del cultivo de tomate de la variedad ASTONA frente a un sistema de conducción a dos ejes, en el cual el segundo eje es formado a partir de una yema axilar, una vez concluida la fase de crecimiento vegetativo exclusivo. Se evaluó el rendimiento, el crecimiento vegetativo de la planta (diámetro del tallo, altura de la planta y área foliar) y la calidad del fruto, que incluye factores como el pH, sólidos solubles, acidez titulable, color y calibre de los frutos. Los tratamientos a que fueron sometidas las plantas consistieron en dos tratamientos de conducción a doble eje. En el primero las plantas fueron conducidas a dos ejes intercaladas con plantas a un solo eje (3.8 ejes·m-2) y en el segundo todas las plantas fueron conducidas a dos ejes (5.0 ejes·m-2). Estos se contrastaron con un tratamiento control donde las plantas fueron conducidas a un solo eje (2.5 ejes·m-2). La toma del eje lateral, para aumentar la densidad poblacional por unidad de superficie, se realizó desde la axila de la hoja dispuesta inmediatamente debajo de la primera inflorescencia de la planta. El despunte del primer eje se realizó sobre el quinto racimo floral y el despunte del segundo eje sobre el segundo racimo floral. Las plantas conducidas a un solo eje fueron despuntadas sobre el séptimo racimo floral. Los resultados obtenidos por los tratamientos de conducción (3.8 y 5.0 ejes·m-2) no presentan un aumento del rendimiento total, pero sí producen una concentración de la cosecha en las dos primeras semanas de enero, lo que es económicamente favorable para el productor. La distribución de las diferentes categorías de tamaño de los frutos no fue influenciada por el aumento de la densidad de ejes. Con respecto al crecimiento vegetativo se observó que el área foliar final de las plantas de ambos tratamientos es mayor, lo que sugiere una mayor producción de asimilados por unidad superficie y la protección de los frutos de la radiación solar directa. El diámetro del tallo y la altura de la planta no se vieron afectados. Con respecto a la calidad del fruto, los factores peso promedio, diámetro polar y ecuatorial, pH y color externo no presentan diferencias. En cambio, la acidez titulable es mayor para el tratamiento de 5.0 ejes·m-2, mientras que los sólidos solubles y la luminosidad interna del fruto disminuyen en el tratamiento de 3.8 ejes·m-2. Por consiguiente, es factible el establecimiento de un cultivo de tomate conducido a doble eje para concentrar la cosecha, sin disminuir los rendimientos ni afectar la calidad organoléptica de los frutos.

7. LITERATURA CITADA. ATHERTON, J. and HARRIS, G. 1986. Flowering. In: Atherton, J and Rudich, J.
eds. The Tomato Crop. London, Chapman & Hall. pp. 167-200. ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS. 1990. Método
Nº942. 15 A. 15ª Edición. Virginia, Association of Official Analytical Chemists, Inc. 918 p.
AUNG, L. 1976. Effects of photoperiod and temperature on vegetative and
reproductive responses of Lycopersicon esculentum Mill. Journal of the American Society for Horticultural Science 101: 358-360
BRAVO, A. y ALDUNATE, P. 1986. El cultivo del tomate. El Campesino 117: 16-
47 CALVERT, A. 1964. The effects of air temperature on growth of young tomato
plants in natural light conditions. Journal of Horticultural Science 39: 194-211.
CASTILLA, N. 1995. Manejo del cultivo intensivo. In: Nuez, F. ed. El Cultivo del
Tomate. Madrid, Mundi-Prensa. pp.189-226. CENTRO DE INFORMACIÓN DE RECURSOS NATURALES. 1985.
Descripciones de suelos. Estudio agrológico de los valles de Aconcagua, Putaendo, Ligua y Petorca. Santiago, CIREN-CORFO. 317 p.
CHAMARRO, J. 1995. Anatomía y fisiología de la planta. In: Nuez, F. ed. El
Cultivo del Tomate. Madrid, Mundi-Prensa. pp. 43-92 COCKSHULL, K.; HO, L. and FENLON, J. 2001. The effect of the time of taking
side shoots on the regulation of fruit size in glasshouse- grown tomato crops. Journal of Horticultural Science & Biotechnology 76: 474-483
________. and HO, L. 1995. Regulation of tomato fruit size by plant density and
truss thinning. Journal of Horticultural Science 70: 395-407 ________.; GRAVES, C. and CAVE, C. 1992. The influence of shading on yield of
glasshouse tomatoes. Journal of Horticultural Science 67: 11-24 COOPER, A. 1969. Effects of shading on tomato stem extension. Journal of
Horticultural Science 44: 75-79

DINAR, M. and STEVENS, A. 1981. The relationship between starch accumulation
and soluble solids content of tomato fruits. Journal of the American Society for Horticultural Science 106: 415-418
DUIMOVIC, A. 1992. La poda, conducción y raleo en cultivos bajo plástico.
Fundamentos y usos. Universidad Católica de Valparaíso. Curso de producción de tomate bajo plástico, Ovalle 15 abril 1992. pp.G1- G4
ESCAFF, M. 1993. Tomate: variedades, almácigos y manejo de la planta. INIA La
Platina. Curso Internacional de Producción de Hortalizas Protegidas bajo Plástico. Santiago, 7, 8 y 9 noviembre 1993. pp. 4.21-4.29
________. 1992. El tomate para cultivo en invernadero. INIA Estación Intihuasi.
Seminario de Producción de Hortalizas bajo Plástico. La Serena, 10 y 11 julio, 1992. pp. 22-58
FAIGUENBAUM, H. 1986. Efecto de la densidad y espaciamiento sobre el área
foliar, duración del área foliar y producción de dos cultivares de frejol (Phaseolus vulgaris L.). Investigación Agraria 13: 91-101
FERY, R. and JANICK, J. 1970. Response of the tomato to population pressure.
Journal of the American Society for Horticultural Science 95: 614-624 GEISENBERG, C. and STEWART, K. 1986. Field crop management. In: Atherton,
J and Rudich, J. eds. The Tomato Crop. London, Chapman & Hall. pp. 511-557
GEORGE, B. and PEIRCE, L. 1969. The influence of population density and
competition on phenotypic stability of tomato plants. Journal of the American Society for Horticultural Science 94: 65-67
GIACONI, V. y ESCAFF, M. 1994. Cultivo de Hortalizas. Santiago, Editorial
Universitaria. 336 p. GIFFORD, M. and EVANS, L. 1981. Photosynthesis, carbon portioning and yield.
Annual Review of Plant Physiology 32: 485-509. GONZÁLEZ, L. 1995. Estudio de factibilidad técnica del cultivo del tomate de ciclo
largo en invernaderos de Quillota. Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 131 p.

GONZÁLEZ, M. 1994. Producción de tomates en invernadero. IPA Quilamapu 59: 21-26.
GRIERSON, D. and KADER, A. 1986. Fruit ripening and quality. In: Atherton, J
and Rudich, J. eds. The Tomato Crop. London, Chapman & Hall. pp. 240-279
HARPER, L.; PALLAS, J.; BRUCE, R. and JONES, J. 1979. Greenhouse
microclimate for tomatoes in the southeast. Journal of the American Society for Horticultural Science 104: 659-663
HELLMAN, H., BARKER, L., and FROMEMER, W. 2000. The regulation of
assimilate allocation and transport. Australian Journal of Plant Physiology 27: 583-594
HERNÁNDEZ, M. 2000. Evaluación de diferentes tipos de conducción, para tomate
injertado, cultivado en invernadero frío. Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 43 p.
HEUVELINK, E. 1997. Effect of fruit load on dry matter partitioning in tomato.
Scientia Horticulturae 69: 51-59 ________. 1995. Effect of plant density on biomass allocation to the fruits in tomato
(Lycopersicon esculentum Mill.). Scientia Horticulturae 64: 193-201 HO, L. 2000. Mejoramiento de la calidad del fruto de tomate a través del cultivo.
Universidad Católica de Valparaíso. Seminario Adaptaciones Tecnológicas para la Modernización de Cultivos Forzados. Quillota, 12-13 octubre 2000. s.p.
________. 1996. Tomato. In: Zamski, E and Schaffer, A. eds. Photoassimilate
distribution in plants and crops. Source-sink relationships. New York, Marcel Dekker, Inc. pp: 709-758
________. 1988. Metabolism and compartmentation of imported sugars in sinks
organs in relation to sink strength. Annual Review Plant Physiology 39: 355-378
________. and HEWITT, D. 1986. Fruit development. In: Atherton, J and Rudich, J.
eds. The Tomato Crop. London, Chapman & Hall. pp: 201-239

________.; SJUT, V and HOAD, V. 1983. The effect of assimilate supply on fruit growth and hormone levels in tomato plants. Plants Growth Regulation 1: 155-171
HURD, R. and COOPER, A. 1970. The effect of early low temperature treatment on
the yield of single-inflorescence tomatoes. Journal of Horticultural Science 45: 19-25
KADER, A.; MORRIS, L.; STEVENS, M. and ALBRIGHT-HOLTON, M. 1978.
Composition and flavor quality of fresh market tomatoes as influenced by fruit ripeness when harvested. Journal of the American Society for Horticultural Science 103: 6-13
KINET, J. and PEET, M. 1997. Tomato. In: Wien, H. ed. The Physiology of
Vegetable Crops. New York, CABI Publishing. pp 207-258 ________. 1977. Effect of light conditions on the development of the inflorescence
in tomato. Scientia Horticulturae 6: 15-26 KRARUP, C. 2002. Poscosecha de tomates. Universidad Católica de Valparaíso.
Curso producción de hortalizas: metodología de producción de calidad, post cosecha, valor agregado y comercialización. Quillota, 21 al 25 octubre,2002. pp. 1-24
________. 1992. Tomate: a la vanguardia tecnológica. Panorama Económico de la
Agricultura 84: 22-28 MARCOTTI, C. 1998. Evaluación económica de la disminución de densidad de
plantación para un cultivo de tomate cultivar FA-144 bajo invernadero frío durante el periodo de otoño. Taller de Licenciatura Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 57 p.
MAROTO, J. 1994. Horticultura Herbácea Especial. 4ª. ed. Madrid, Ediciones
Mundi-Prensa. 611 p. McGUIRE, R. 1992. Reporting of objective color measurements. HortScience 27:
1254-1255 MENDOZA, H. 1992. Densidades y marcos de plantación. Universidad Católica de
Valparaíso. Curso de Producción de Tomate Bajo Plástico. Ovalle, 15 abril 1992. pp. B1-B8

NAVARRETE, M. and JEANNEQUIN, B. 2000. Effect of frecuency of axillary bud pruning on vegetative growth and fruit yield in greenhouse tomato crops. Scientia Horticulturae 86: 197-210
________., ________. and SEBILLOTE, M.. 1997. Vigour of greenhouse tomato
plants (Lycopersicon esculentum Mill): analysis of the criteria used by growers and search for objective criteria. Journal of Horticultural Science 72: 821-829
NOVOA, R.; VILLASECA, S.; del CANTO, P.; ROUANET, M.; SIERRA, C.; del
POZO, A. 1989. Mapa agroclimático de Chile. Santiago, INIA. 221 p. OYANEDEL, E. 2002. Desórdenes del crecimiento vegetativo y reproductivo en
tomate. Avance Agrícola 107: 2-4 PAPADOPOULOS, A. and PARARAJASINGHAM, S. 1997. The influence of
plant spacing on light interception and use in greenhouse tomato (Lycopersicon esculentum Mill): a review. Scientia Horticulturae 69: 1-29
________. and ORMRON, D. 1990. Plant spacing effects on yield of the greenhouse
tomato. Canadian Journal of Plant Science 70: 565-573 ________. and ________. 1988. Plant spacing effects on photosynthesis and
transpiration of the greenhouse tomato. Canadian Journal of Plant Science 68: 1209-1218
PICKEN, A.; STEWART, K. and KLAPWIJK, D. 1986. Germination and
vegetative development. In: Atherton, J and Rudich, J. eds. The Tomato Crop. London, Chapman & Hall. pp 111-166
RODRÍGUEZ, B. and LAMBETH, V. 1976. Photosyntetic rate-yield component
relationship. Journal of the American Society for Horticultural Science 100: 694-697
ROJAS, L. y ALFARO, V. 1995. Densidad de plantación en tomate y pepino de
ensalada. Tierra Adentro 3: 47-49 SAKAMOTO, Y., WATANABLE, S. and OKANO, K. 2000. Multi-shoot training in
single-truss tomato cultivation. Bulletin of the National Research Institute of Vegetables, Ornamental Plants and Tea 15: 115-122.

SALINAS, O.; RAMÍREZ, O. y OSPINA, J. 1994. Efecto del sistema de tutorado, poda de tallos y poda de hojas sobre la calidad del fruto del tomate (Lycopersicon esculentum Mill.). Agronomía Colombiana 11: 184-189
SCHWARZ, D and KLÄRING, H. 2001. Allometry to estimate leaf area of tomato.
Journal of Plant Nutrition 24: 1291-1309 SUNSEEDS. 2003. Varieties, (on line). www.sunseeds.com
URRUTIA, G. 2000. Tomate para consumo fresco. Mercado y perspectivas.
Agroeconómico 54: 50-54 VAN DE VOOREN, J.; WELLES, W. and HAYMAN, W. 1986. Glasshouse
production. In: Atherton, J and Rudich, J. eds. The Tomato Crop. London, Chapman & Hall. pp. 580-623
WATANABLE, S.; SAKAMOTO, Y.; NAKASHIMA, T. and OKANO, K. 1999.
Varietal characteristics suitable for the cultivation of single- truss in hydroponics. Bulletin of the National Research Institute of Vegetables, Ornamental Plants and Tea 14: 165-175

ANEXO 1. Distribución espacial de las mesas de cultivo en invernadero frío con pies centrales y unidad experimental.
ME
SA
PASI
LL
O
7 m
1,4 m
30 m
2.3 m
1.4 m
Unidad de muestreo 6.5 m


















