Fisiología Resumen Medicina Segundo Año. Sistema Renal y Digestivo Universidad de Santiago
40
FUNCION RENAL El organismo tiende a mantener la constancia de su medio interno y la composición del líquido que rodea sus células. Para esto dispone de mecanismos que hacen posible el transporte y eliminación de los productos no utilizables del metabolismo celular. Los catabolitos inservibles no volátiles y las sustancias ingeridas en exceso (agua, sales, etc.) son transportadas por la circulación a, los órganos de su eliminación. Entre éstos, el de mayor importancia es el riñón. Los riñones elaboran la orina y depuran así la sangre de estas sustancias, regulando en esta forma la composición de la sangre y de los líquidos corporales en general. Los riñones cumplen con dos funciones principales: a) excretan productos finales del metabolismo celular; b) controlan a concentración en la mayoría de los componentes de los componentes de los líquidos del organismo. La cantidad y composición de la orina presenta amplias variaciones diarias. Se puede considerar, sin embargo, que el volumen de orina del adulto en 24 horas es alrededor de 1.5 litros. La orina es una solución que contiene sólo un 4% de sustancias sólida. Su peso especifico indica aproximadamente la concentración de los solutos y fluctúa entre 1.015 y 1.025. Tabla Nº3 PRINCIPALES COMPONENTES DE LA ORINA Componentes inorgánicos Componentes orgánicos Cloruro de sodio 10-15 g. Urea 20-30 g. Potasio 2-4 g. Creatinina 08-1.2 g: Acido sulfúrico 2-3 g. Acido úrico 0.6-0.8 g. Acido fosfórico 2-3 g. Indican 0.01-0.02 g. Amoniaco 0.5-1 g. Cuerpos cetónicos 0.04 g. Magnesio 0.1-0.2 g. . . Calcio 0.3-0.4 g. . . Otras sustancias 0.2-0.3 g. . . Fierro 0.005-0.010 g . . Su color amarillo se debe a la presencia de un Pigmento, el urocromo deriva de la hemoglobina. La Tabla 3 indica su composición. La reacción de la orina es normalmente ácida y su pH fluctúa entre 5 y 6, pero puede alcanzar va1ores tan bajos Como 4.7 o tornarse alcalina, pudiendo alcanzar un pH máximo de 8.2. Su pH depende de los alimentos ingeridos y de la Cantidad de ácidos o álcalis excretados. Los principales componentes urinarios, como puede apreciarse en la tabla 3, son el agua, la urea, los cloruros, sulfatos y fosfatos, el sodio y el potasio. Puede contener, además de la urea, cantidades relativamente pequeñas de otras sustancias nitrogenadas (sales amónicas, creatinina, ácido úrico y uratos).
-
Upload
claudio-alexis-cifuentes -
Category
Documents
-
view
42 -
download
2
Transcript of Fisiología Resumen Medicina Segundo Año. Sistema Renal y Digestivo Universidad de Santiago

FUNCION RENAL
El organismo tiende a mantener la constancia de su medio interno y la composición del líquido que rodea sus células. Para esto dispone de mecanismos que hacen posible el transporte y eliminación de los productos no utilizables del metabolismo celular. Los catabolitos inservibles no volátiles y las sustancias ingeridas en exceso (agua, sales, etc.) son transportadas por la circulación a, los órganos de su eliminación. Entre éstos, el de mayor importancia es el riñón.
Los riñones elaboran la orina y depuran así la sangre de estas sustancias, regulando en esta forma la composición de la sangre y de los líquidos corporales en general. Los riñones cumplen con dos funciones principales: a) excretan productos finales del metabolismo celular; b) controlan a concentración en la mayoría de los componentes de los componentes de los líquidos del organismo. La cantidad y composición de la orina presenta amplias variaciones diarias. Se puede considerar, sin embargo, que el volumen de orina del adulto en 24 horas es alrededor de 1.5 litros. La orina es una solución que contiene sólo un 4% de sustancias sólida. Su peso especifico indica aproximadamente la concentración de los solutos y fluctúa entre 1.015 y 1.025.
Tabla Nº3
PRINCIPALES COMPONENTES DE LA ORINA
Componentes inorgánicos Componentes orgánicos
Cloruro de sodio 10-15 g. Urea 20-30 g.
Potasio 2-4 g. Creatinina 08-1.2 g:
Acido sulfúrico 2-3 g. Acido úrico 0.6-0.8 g.
Acido fosfórico 2-3 g. Indican 0.01-0.02 g.
Amoniaco 0.5-1 g. Cuerpos cetónicos 0.04 g.
Magnesio 0.1-0.2 g. . .
Calcio 0.3-0.4 g. . .
Otras sustancias 0.2-0.3 g. . .
Fierro 0.005-0.010 g . .
Su color amarillo se debe a la presencia de un Pigmento, el urocromo deriva de la hemoglobina. La Tabla 3 indica su composición.
La reacción de la orina es normalmente ácida y su pH fluctúa entre 5 y 6, pero puede alcanzar va1ores tan bajos
Como 4.7 o tornarse alcalina, pudiendo alcanzar un pH máximo de 8.2. Su pH depende de los alimentos ingeridos y de la Cantidad de ácidos o álcalis excretados.
Los principales componentes urinarios, como puede apreciarse en la tabla 3, son el agua, la urea, los cloruros, sulfatos y fosfatos, el sodio y el potasio. Puede contener, además de la urea, cantidades relativamente pequeñas de otras sustancias nitrogenadas (sales amónicas, creatinina, ácido úrico y uratos).

ANATOMIA FUNCIONAL DEL RIÑON
La formación de la orina a partir del plasma está íntimamente relacionada con su estructura.
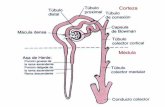
En un corte transversal del riñón se distinguen tres zonas: a) La zona cortical, que albergan los glomérulos: b) la Zona medular, integrada por las asas de Helen y por túbulos colectores que forman las pirámides de Malpighi ; c)La pelvis renal en que desembocan todo los túbulos colectores y que se continúa con los uréteres, que llevan la orina a la vejiga. Los riñones están arrigados por las arterias renales, ramas de la aorta abdominal . Son muy cortas y se dividen entre las pirámides de Malpighi, dando origen a las arterias interlaborales. De estas a su vez a nivel de la zona corticomedular, nacen las arterias arciformes, las cuales se continúan en las arterias interlobulillares que formarán final mente las arteriolas aferentes de los glomérulos. La figura 59 muestra un corte longitudinal del riñón (A) y el esquema de un glomérulo (B) con la cápsula de Bowman.
En el hombre, cada riñón está formado por 1.000.000 de nefrones, aproximadamente. Cada nefron es capaz, por si mismo, de formar orina.
Basta, por lo tanto, conocer el funcionamiento de un nefrón para comprender la función del riñón en su totalidad .
El nefrón está formado por dos partes funcionalmente diferentes: por el glomérulo que es el aparato de filtración y por el túbulo en el cual se absorbe parcialmente el filtrado y en que se realiza la secreción de ciertas sustancias.
Se puede resumir la anatomía funcional del riñón en la siguiente forma: sangre penetra al glomérulo por la. arteriola aferente para salir por .la arteria eferente .(Fig. 59B). El glomérulo, rodeado por la, cápsula de Bowman, es una red DE UNOS 50 capilares. La cápsula es un saco en que se colectan el liquido filtrado desde los capilares. La sangre sale del glomérulo por la arteriola eferente, la cual se capilariza prófusamente alrededo de los túbulos (red capilar peritubular).

Los capilares peritubulares con forman vénulas y venas de mayor calibre para desembocar finalmente en las venas renales . Una pequeña parte de la sangre fluye, sin embargo ,por, vasos que no pasan por los capilares que se dirigen; hacia la, parte distal del nefrón , para volver nuevamente hacia la parte proximal, reciben el .nombre de vasos rectos. La figura 60 presenta un diagrama simplificado de la estructura del nefrón y suirrigación.
El liquido que filtra a través de la pared capilar del glomérulo panetra la cápsula de Bowman y de aquí, a través del túbulo proximal al asa de Henle cuya parte inferior recibe el nombre dé segmento delgado. El filtrado pasa del túbuolo proxirnal al túbulo distal para llegar finalmente al túbulo colector, que reúne el filtrado. de varios nefrones: La mayor. parte del agua Filtrada, juntó con algunos solutos, son reabsorbidos a nivel tubular, de modo que la orina está constituida solo por la fracción no absorbida de agua y de solutos.
FUNCION DEL NEFRON
La función básica del nefrón consiste en depurar el plasma sanguíneo de catabolitos no utilizables. Las sustancias eliminadas son predominantemente productos finales del metabolismo, como urea, creatinina, ácido úrico, sulfatos. y fenoles. Además, materiales no metabolizables que se acumulan en cantidades excesivas en el organismo (iones de sodio, de potasio o de cloruro). El nefrón depura el plasma de estas sustancias por un mecanismo que puede esquematizarse en la forma siguiente: 1) aproximadamente la quinta parte del plasma que pasa por los glomérulos filtra por la membrana glomerular; 2) el líquido filtrado pasa a los túbulos en los que las sustancias que el organismo puede utilizar (glucosa, aminoácidos, agua y muchos electrólitos), retornan a los capilares peritubulares y las inservibles no se reabsorben. Hay pues una reabsorción tubular selectiva.
El estudio de la filtración glomerular es importante en la comprensión de la fisiología renal. Por este proceso el riñan elimina cierto volumen de líquido, una parte del cual se reabsorbe en los túbulos y el resto es eliminado por la orina.

La filtración qlomerular es totalmente pasiva, mientras que la reabsorción es en, su mayor parte un proceso activo. E1 mecanismo de filtración del liquido desde los capilares intraglomerulares hacia el espacio de la cápsula de Bowman, es similar al que rige el intercambio á través de la membrana capilar en el resto del organismo y está detalladamente descrito en un capituló precedente. Las presiones que tienden a expulsar líquido de los capilares son mayores que las que tienden á retenerlo, debió a lo cual filtra líquido a través de las páredes capilares glomerulares, hacia la cápsula de Bowman. La presión hidrostática en los capilares glomerulares es aproximadamente 60 mm de Hg (superior a la de los otros capilares del cuerpo), circunstancia que favorece la filtración. La presión hidrostática intracapilar, creada por la fuerza contráctil del corazón, es la que tiende a expulsar el líquido
hacia la cápsula de Bowman. A ella se Oponen dos presiones que tratan de retener el líquido en los capilares: una es la presión coloídoosmótica de las proteínas plasmáticas cuya magnitud, como se ha indicado anteriormente , (ver Líquidos corporales), es aproximadamente 32 mm de Hg; la otra es la presión hidrostática existente en el interior de la cápsula d Bowman, que es del orden de 14 mm de Hg. siendo la. presión hidrostática en los capilares') glomerulares alrededor de 70 mm de Hg y la presión coloidosmótica aproximadamente 24 mm de Hg, la presión de filtración es 28 mm de Hg. A la presión de filtración se opone la presión del líquido intracapsular (14 mm de Hg), de manera que la presión efectiva de filtración es de 14 mm de Hg (Fig. 6I ).
Los valores indicados son válidos siempre que la presión en la arteria renal sea de 100 mm de Hg..
Debido a que la presión efectiva de filtración en el riñón (24 mm de Hg) es sustancialmente superior a la presión de filtración capilar en otras partes del organismo, la velocidad de filtración renal es muy alta. La mayor presión hidrostática de los capilares glomerulares está condicionada por algunas particularidades de la circulación renal. Primero, a la reducida longitud y gran diámetro de las arterias renales. Estas nacen en ángulo recto, directamente de la aorta, lo que explica la pequeña diferencia de presión entre la aorta y tos capilares renales. Segundo, a la alta resistencia que encuentra el flujo de sangre en los capilares del glomérulo. Esta se debe, por una parte, a que el diámetro de la arteriola eferente es menor que el de la aferente; por otra parte a que la arteriola eferente se ramifica en una segunda red capilar que representa una resistencia adicional.
De esta manera, la sangre circula por los riñones a alta presión. Los riñones, en relación a su peso, son los órganos de más intensa irrigación del cuerpo humano, ya que pasa por ellos aproximadamente la cuarta parte del volumen-minuto, o sea, 1.2 litros/min. Esto significa que en 24 horas fluye por los riñones, cuyo peso conjunto no sobrepasa los 300 g, la extraordinaria cantidad de 1.728 litros de sangre. De los 1.200 ml de sangre que pasan en cada minuto por los riñones, alrededor de liso ml corresponden a plasma y 550 ml a los elementos figurados. Como sólo el plasma participa en la formación de la orina, para calcular el volumen total del líquido filtrado, la cantidad de los elementos figurados carece de interés. De los 1200 ml de plasma filtran hacia la cápsula de Bowman aproximadamente 125 ml de líquido por minuto, o sea, cerca de 180 litros en

24 horas. A primera vista aparece antieconómico que el riñón filtre un volumen de 180 litros, aproximadamente IS veces más que el volumen total del líquido extracelular, para eliminar en 24 horas unos 1.500 ml de orina. Pero esto les permite actuar rápida y eficazmente sobre la composición de este líquido, y regular así constantemente su volumen y composición.
Un factor de gran importancia en la filtración es la permeabilidad de la membrana filtrante. Esta membrana consta de varias capas cuya estructura histológica se ha llegado a conocer gracias a estudios realizados con el microscopio electrónico.
La membrana filtrante está formada por dos tipos de células: las células endoteliales de la pared capilar y las células epiteliales de la pared de la cápsula de Bowman, que se adosa a los capilares en toda su extensión. Entre ambas capas de células se encuentra la membrana basal o lámina densa, que representa en realidad el verdadero filtro. Esta lámina, que es la que realmente selecciona las sustancias que pasarán por el filtro, posee poros (orificios) funcionales de un diámetro de 80 Amstrongs (1 A = 0.1 milimicrón = diez millonésimas partes de un milímetro). Estos poros sólo permiten el paso de moléculas de agua y de aquellas substancias cuyo diámetro molecular no es mayor que 80 A (glucosa, urea, etc.) y retiene todas aquellas sustancias que tienen un diámetro molecular mayor. La existencia de estos poros no ha sido confirmada por el microscopio electrónico, pero las características funcionales de la membrana permiten afirmar su existencia y calcular su diámetro.
Tabla N ° 4
COMPOSICION RELATIVA EN EL PLASMA Y EN LA ORINA DE ALGUNAS SUSTANCIAS
Glucosa Plasma Orina Relación orina / plasma
mg/ 100 ml sodio
100 0 0
mEq/Litro Urea
140 20
140 800
1 40
Creatinina mg/ 100 ml
120 1 120
Proteínas g/ 100 ml
7 0 0
Magnesio mg %
2,5 6 2
Los elementos figurados de la sangre no pueden pasar por los poros de la lámina densa, como tampoco las proteínas, especialmente aquellas cuyo peso molecular es elevado (seroalbúminas). Por lo tanto, la composición del filtrado glomerular es idéntica a la del plasma, a excepción de los elementos figurados y proteínas. Cuando un líquido que contiene proteínas pasa por un filtro en tal forma que el filtrado resultante carece de proteínas, estamos frente a un proceso llamado ultrafiltración. Se puede, por consiguiente, definir el líquido en la cápsula de Bowman, como un ultrafiltrado del plasma.
La Tabla 4 compara la concentración de algunas sustancias en la sangre y en la orina. Hay que tener presente sin embargo, que la composición de la orina es muy distinta a la del ultrafiltrado glomerular, diferencia que se debe a los cambios que sufrí el ultrafiltrado durante su paso por los túbulos.

REGULACION DE LA FILTRACION
El volumen del filtrado depende del volumen y de la presión de la sangre que pasa por los riñones. El volumen del ultrafiltrado producido por los riñones en 24 horas y en condicionas fisiológicas, es constante. Esta constancia es el resultado de mecanismos de autorregulación, que hacen la irrigación de los riñones en cierto modo independiente de las variaciones circulatorias del resto del organismo.
La autonomía de la circulación glomerular resulta principalmente del ajuste de los diámetros de las arteriolas aferentes y eferentes, o sea, de la regulación del volumen de sangre que entra y salí de los glomérulos. Como ya se ha explicado, normalmente el diámetro de la arteriola aferente es superior al de la eferente, lo que contribuye a crear una alta presión en el interior del glomérulo. Si por alguna razón el flujo sanguíneo en el organismo disminuye o la presión arterial desciende, las arteriolas aumentan sus diámetros mediante los rnecanismos de autorregulación. Pero se mantiene la diferencia entre el diámetro de las arteriolas aferentes y eferentes, la que asegura en los capilares glomerulares la presión hidrostática necesaria y una filtración cuantitativamente adecuada.
Cuando la presión arterial o el volumen de sangre circulante es elevado, el ajuste se realiza en sentido opuesto. En condiciones fisiológicas los mecanismos de autorregulación actúan rápidamente, asegurando un volumen uniforme de filtrado, siempre que las variaciones de la irrigación y de la presión no sobrepasen ciertos límites. Cuando esta autorregulación deja de ser efectiva, disminuye el volumen de sangre que pasa por los glomérulos y consecutivamente la cantidad del filtrado. En estas condiciones la producción de orina decrece y en casos extremos puede hasta suspenderse por completo (anuria isquémica).
Se ha planteado que la actividad de las células epiteliales que forman la capa externa de la lámina densa o basal, podría jugar un papel en la regulación de la filtración glomerular a través del aumento o disminución del tamaño de sus poros. Sin embargo esta es sólo una hipótesis no confirmada.
METODOS DE ESTUDIO DE LA FUNCION GLOMERULAR
En general, se estudia la función renal por métodos indirectos, llamados clearance o depuración. Estos permiten calcular no sólo el volumen de líquido filtrado en los glomérulos, sino también la reabsorción y excreción tubular de sustancias. procesos que condicionan el volumen y composición final de la orina. Así por ejemplo, si 100 ml de plasma contienen 0.1 g de una sustancia que filtra libremente en los glomérulos y no se reabsorbe, ni se excreta en los túbulos renales, y en la orina formada en un minuto hay 0.1 g de la misma sustancia, habrá que admitir que en un minuto filtran 100 ml de plasma en los glomérulos. Se podría definir, por lo tanto, el concepto del índice de depuración o clearance plasmático, como el volumen de plasma depurado de sustancias por los riñones en la unidad de tiempo.
Ya hemos indicado que la sustancia utilizada para medir el volumen de filtrado debe reunir dos propiedades básicas: a) filtrar libremente en los glomérulos, para lo cual su tamaño molecular debe ser inferior a 80 A; b) que una vez filtrada no se reabsorba ni se excrete por los túbulos. Sólo en este caso estará su cantidad en la orina en relación directa con el volumen de filtrado glomerular. Una de las sustancias que posee estas calidades y cuya determinación cuantitativa en el plasma y en la orina no ofrece mayores dificultades, es la inulina. Esta sustancia, además de filtrar

libremente y no ser absorbida ni excretada por los túbulos, no se metaboliza en el organismo, no se une a las proteínas "plasmáticas, es fisiológicamente inerte y no es tóxica.
Se calcula el clearance de la inulina a base de que por cada miligramo de inulina que aparece en la orina, tiene que haber filtrado la cantidad de plasma que contenga en solución ese miligramo de inulina. Por lo tanto, si P representa la concentración de inulina en el plasma (expresada en mg/ml), U la concentración de inulina en la orinó (en mg/ml) y V el volumen de orina (en ml/minuto) la fórmula que permite calcular el clearance de inulina será la siguiente:
C = U x V ...........P
Repetidas mediciones han permitido establecer que el valor promedio del índice de depuración de la inulina en el hombre es de 127 ml/minuto para un adulto normal de 70 kg de peso y 1.73 m2 de superficie corporal. De esta cifra se deduce que diariamente filtran aproximadamente 183 litros de plasma. Esta cantidad equivaldría a 16 veces del volumen del líquido extracelular total y significa que el volumen total del plasma (que representa el 4.5% del peso corporal) filtra 58 veces al día y que cada 25 minutos se depura la totalidad del plasma circulante.
En resumen y para aclarar el concepto de clearance o índice de depuración de inulina, es conveniente reiterar que el valor de 127 ml/minuto significa que 127 ml de plasma deben ser filtrados en los glomérulos para proporcionar a la orina una determinada cantidad de inulina en un minuto.
FUNCION DE LOS TUBULOS RENALES
El túbulo renal consta de tres segmentos histológica y funcionalmente diferentes. En el primer segmento, denominado túbulo proximal, se efectúa la reabsorción activa de los solutos filtrados. El segundo segmento (asa de Henle) es largo y delgado y fue considerado antiguamente como funcionalmente inerte. Investigaciones recientes, que se explicarán más adelante, han revelado, sin embargo, que tiene un rol importante en el control del volumen urinario y del líquido corporal. Finalmente en el tercer segmento o túbulo distal, se realiza la reabsorción selectiva del sodio y la acidificación de la orina. Varios túbulos dístales desembocan en un túbulo colector y varios de estos túbulos colectores forman una papila renal. Las papilas renales confluyen para constituir la pelvis renal, que se continúa con el uréter, el cual conduce la orina a la vejiga. El túbulo colector llene además una participación activa en la reabsorción del agua y en el manejo de los electrólitos.
Una de las funciones importantes del riñón es la mantención de la constancia de la concentración de iones de H
+, función que será analizada al final de este capítulo. Sólo se mencionará en este
lugar que los túbulos proximales y dístales mediante la reabsorción de los bicarbonatos, formación de amoníaco y excreción de iones de H
+, contribuyen eficazmente a la conservación de la
constancia del medio interno.
El proceso principal que se realiza en los tres segmentos de los túbulos renales es la reabsorción, que afecta tanto al agua como los solutos. El mecanismo de absorción varía en cada segmento. En el túbulo proximal, por ejemplo, se reabsorben pasivamente agua y solutos, mientras en el asa de Henle y en el túbulo colector, por la acción de la hormona antidiurética, se reabsorbe activamente el agua.
MECANISMO DE SECRECION Y ABSORCION TUBULAR

En el túbulo proximal se reabsorben principalmente los solutos, junto con un volumen considerable de agua. La reabsorción del agua es totalmente pasiva, regida por las leyes de la osmosis y es sólo la consecuencia de la reabsorción de los solutos. Contribuye a la mantención de la osmolaridad del filtrado.
La capacidad del túbulo proximal para reabsorber líquido es casi ilimitada, si se considera que a su nivel, de los 185 litros filtrados en 24 horas, el 80-85% (alrededor de l50 litros) vuelve a la circulación. La reabsorción es un proceso complejo. El túbulo proximal, mediante procesos activos y muchas veces selectivos, cambia fundamentalmente la composición del ultrafiltrado. En consecuencia, el líquido que pasa del túbulo proximal a la asa de Henle es de composición diferente.
El análisis comparativo del líquido en el asa de Henle y del ultrafiltrado en la cápsula de Bowman, permite evaluar los cambios producidos en el túbulo proximal. Los 30 litros de líquido que fluyen en 24 horas por el asa de Henle (de los 180 litros filtrados 150 se reabsorben en los túbulos proximales), tienen la misma osmolaridad que el plasma sanguíneo, pero carecen de glucosa y contienen sólo cantidades insignificantes de bicarbonato, de fosfatos y de aminoácidos. Aproximadamente un 80-85% del Na
+ y Cl
- , la totalidad del K
+ , Ca
++ y Mg
++, el 50% de la urea y
del ácido úrico filtrado en el glomérulo, han sido reabsorbidos en el túbulo proximal.
La concentración de algunas sustancias (ácido hipúrico, urocromo, algunos fosfatos) es superior en el líquido del asa de Henle que el ultrafiltrado glomerular, a pesar de que la reabsorción en el túbulo proximal se efectúa en forma isoosmolar con el ultrafiltrado. Este hecho revela que el túbulo proximal es no sólo capaz de reabsorción, sino que tiene también capacidad excretora.
La reabsorción tubular se realiza mediante mecanismos similares a los que utilizan las membranas biológicas en general, o sea, mediante transporte activo o pasivo (ver Absorción Intestinal). Entre las sustancias reabsorbidas activamente son especialmente importantes la glucosa, fosfatos, ácido úrico, aminoácidos, vitaminas C y B12. La capacidad de transporte activo de los túbulos es limitada y cuando la cantidad de una sustancia por reabsorber sobrepasa esta capacidad, el exceso es eliminado por la orina. La cantidad máxima de una sustancia que las células tubulares son capaces de transportar del túbulo a la sangre, es designada como Tm. (transporte máximo) y es diferente para cada sustancia.
El transporte de glucosa es de especial interés clínico y merece una explicación detallada. La concentración de glucosa en la sangre, y por lo tanto en el filtrado glomerular, es normalmente alrededor de 1 g/litro. En condiciones fisiológicas, la glucosa es completamente reabsorbida en el túbulo proximal y, por lo tanto, la orina no contiene glucosa. En ciertas enfermedades (diabetes, por ejemplo) o debido a una ingestión excesiva de glucosa, su nivel en la sangre, y por lo tanto en el ultrafiltrado, puede elevarse en tal forma que sobrepasa el Tm. de las células tubulares. Una parte de la glucosa filtrada no será absorbida, sino eliminada por la orina. Se habla en este caso de glucosuria.
El Na+ , el Cl
- , el HC03
- y parcialmente el K
+, se reabsorben sin limitación alguna (hay que tener
presente que estas sustancias se reabsorben como iones y no como moléculas). Es importante señalar que, debido a la modalidad de la reabsorción y excreción renal, el líquido intratubular conserva su electroneutralidad. Así, por ejemplo, en el caso del NaCl, los iones Na
+ se transportan
en forma activa pero el Cl sigue pasivamente al sodio y se mantienen así constante la electroneutralidad del líquido reabsorbido y la del remanente en los túbulos. El 60-70% del Na
+
filtrado en los glomérulos, se reabsorbe obligatoriamente en el túbulo proximal, es decir, independientemente de las necesidades del organismo. La cantidad reabsorbida es, por consiguiente, siempre la misma. La causa de esta constancia no es conocida.
Existe una estrecha relación entre la reabsorción de Na+ y de agua. El agua sigue al NaCI de
acuerdo con la gradiente osmótica creada por la reabsorción de este último. Aproximadamente

siete octavas partes del agua filtrada se reabsorben en esta forma. La reabsorción es, por consiguiente, obligada, totalmente pasiva y se realiza por simple difusión osmótica. Como veremos más adelante, en los otros segmentos del nefrón la reabsorción acuosa es activa y controlada por la aldosterona y la hormona antidiurética.
De los bicarbonatos filtrados, aproximadamente un 99% se reabsorbe en forma activa. Esta reabsorción está regulada por el pH, es decir, por la concentración de iones hidrógeno en el interior de las células tubulares.
El proceso de reabsorción de los iones K+; es particularmente complejo. Normalmente casi la
totalidad del K+ filtrado se reabsorbe en el
túbulo proximal. Con la orina se elimina siempre, sin embargo, una cantidad mayor de K+ que la no
reabsorbida proximalmente. Hay que admitir, por lo tanto, que en algún otro segmento del nefrón se agrega K
+ al líquido tubular. Se acepta, actualmente, que las células del túbulo distal excretan
K+ en una cantidad que alcanza alrededor de 50 mEq/24 horas.
Entre las sustancias que se reabsorben pasivamente, la urea tiene particular importancia. Este catabolito difunde libremente a través de la membrana de todas las células del organismo y se encuentra, por lo tanto, distribuido uniformemente en todos los líquidos corporales. Como ya hemos explicado, la reabsorción activa de la glucosa y de algunos electrólitos arrastra agua, junto con la cual difunde la urea desde el lumen tubular hacia los capilares peritubulares. La difusión de la urea depende principalmente de la calidad y volumen del ultrafiltrado glomerular y de la orina excretada. Si el ultrafiltrado es escaso (caso en el cual el volumen de orina eliminada también lo es), puede reabsorberse hasta el 70% de la urea filtrada, en tanto que si la ultrafiltración es abundante, la reabsorción no sobrepasa el 40% (Fig. 62).
REABSORCION DE LAS PROTEINAS
Antiguamente se afirmó que el ultrafiltrado glomerular no contiene proteínas. Sin embargo, con el desarrollo de técnicas modernas, se ha demostrado su presencia en la orina. Se ha encontrado que pasan al filtrado sólo aquellas moléculas proteicas cuyo tamaño molecular es inferior a 80 A , o sea, menor que el probable diámetro de los poros de la membrana filtrante. De las proteínas plasmáticas filtran pequeñas cantidades de seroalbúmina (peso molecular 72.000), pero no lo hacen las seroglobulinas, cuyo peso molecular es alto. Algunas proteínas extrañas al organismo, como la albúmina de huevo y la gelatina filtran libremente, ya que sus pesos moleculares no son superiores a 35.000

Se ha determinado que en el filtrado glomerular hay alrededor de 10 mg% de proteínas. Por consiguiente, los 1800 litros filtrados en 24 horas, contendrían 18 g de proteínas. Sin embargo, no se excretan por la orina más de 0.07 g de proteínas por litro, o sea, aproximadamente, 0,11 g en 24 horas (diuresis diaria normal: 1.5 l), lo que indica que la mayor parte de las proteínas filtradas es reabsorbida en los túbulos. El mecanismo íntimo de esta absorción no se conoce, pero es, en todo caso, distinto de los procesos activos de transporte.
EXCRECIÓN TUBULAR
Hoy día este proceso, antiguamente negado, es aceptado por todos los investigadores. Se ha demostrado que normalmente se excretan diversos iones, especialmente por el túbulo distal. La excreción de hidrogeniones por el túbulo distal es de gran importancia, dado el papel que estos juegan en la regulación de la constancia del medio interno. Una cantidad elevada de iones H
+,
producidos en el metabolismo celular, es excretada por la orina. En este proceso hay competencia por los transportadores entre iones H
+, K
+ y Na
+ . El K
+ compite con el ion H
+ para ocupar los
mismos transportadores. Prima, sin embargo, el ion H+ debido a su mayor afinidad por los
transportadores. Las células tubulares dístales excretan también iones NH4+ , excreción que no es
activa como en el caso del K+, sino un simple proceso de difusión pasiva, cuya intensidad depende
de la gradiente de concentración osmótica entre líquido tubular y sangre. A través de la excreción de iones H
+ y NH4
+ , el túbulo distal cumple un papel importante en la regulación del pH de la orina
(ver Regulación del Equilibrio ácido-base).
Aparte de las sustancias normalmente excretadas por el riñón, hay una serie de otras, extrañas al organismo, que al ser suministradas son eliminadas a nivel tubular. Entre estas merecen mención algunas sustancias que circulan en la sangre unidas a proteínas y que por su gran tamaño molecular no filtran en el glomérulo (diuréticos mercuriales, rojo fenol, etc.). Como también, el ácido para-aminohipúrico (PAH), el ácido hipúrico, el diodrast (compuesto yodado), a penicilina. Estas y todas las sustancias extrañas al organismo son excretadas por el túbulo proximal y no por el túbulo dista¡, como es el caso del K
+, el H
+ y el NH4
+.
En resumen, en el túbulo proximal se reabsorbe la mayor parte del líquido ultrafiltrado en el glomérulo. Esta reabsorción es selectiva. El riñón elimina las sustancias inservibles para el organismo y se conservan (reabsorción total o parcial) las que el organismo puede utilizar. Son excretadas, además, las sustancias extrañas al organismo. Se reabsorbe en los túbulos proximales en total un 85% del ultrafiltrado, conservando el 15% restante la osmolaridad del ultrafiltrado (y del plasma). Su composición es, sin embargo, diferente. Se reabsorben en el túbulo distal Na
+ y H2O.
La reabsorción de ésta última es regulada por hormonas. Se excretan en el túbulo distal los iones K
+ + y NH4
+. Las dos funciones principales del túbulo distal son, por consiguiente, la regulación de
la reabsorción de Na+ por la aldosterona, hormona secretada por la corteza suprarrenal y la
regulación del pH de la orina.
El balance acuoso es mantenido por la excreción de una orina osmóticamente diluida (proceso de dilución o diuresis hídrica), o bien, por una orina osmóticamente concentrada (proceso de concentración o de antidiuresis). Normalmente prevalece el segundo proceso, pues la adaptación de la vida fuera del medio acuoso obliga al organismo a ahorrar constantemente agua. El mecanismo de dilución prevalece sólo cuando se ingiere líquido en exceso. Ambos procesos regulan también el balance osmolar, de acuerdo con la necesidad de eliminar o ahorrar osmoles (es decir, solutos). El principal soluto, para los efectos de la presión osmótica, es el Na
+: De
acuerdo con las necesidades del organismo, el riñón es capaz de eliminar orina cuya osmolaridad fluctúa entre 50 mmOsm/1 y 1.300 mOsm/l. La osmolaridad del plasma es de 300 mOsm/1.

En resumen, suponiendo que la ingestión de alimentos sea adecuada, tanto cualitativa como cuantitativamente, la menor ingestión de agua se compensa mediante la concentración de la orina y la sobrehidratación por eliminación de una orina diluida.
Nuestros conocimientos acerca de los mecanismos que regulan la concentración y dilución de la orina, han variado fundamentalmente con la introducción de la llamada teoría de contracorriente.
Esta teoría, comprobada experimentalmente casi en su totalidad, se basa en la peculiar arquitectura de la médula renal, en la que tanto el asa de Henle como los vasos, siguen recorridos en horquilla.
La figura 63 representa esquemáticamente un sistema de contracorriente. Como puede observarse, se trata de un sistema semejante al de un aparato destilador de agua de tubos adosados por uno de los cuales circula agua caliente y por el otro, en sentido contrario, agua fría. De acuerdo con las leyes de difusión del calor la temperatura de ambos juntos tiende a igualarse, y, a un nivel dado, el tubo de agua caliente entrega calor al tubo de agua fría. En consecuencia, la temperatura del agua caliente disminuye a medida que aleja de su punto de entrada al sistema. La del agua fría, que influye en sentido opuesto va aumentando a medida qué avanza. De este modo en el extremo del sistema de tubos, en el cual entra el agua caliente y sale la fría, el calor tiende a acumularse. En el otro extremo, donde sale el agua caliente y entra la fría, la cantidad de calor es mucho menor. Si se compara en un punto intermedio la temperatura de los dos sistemas, la diferencia es escasa y la cantidad de calor que pasa de un tubo a otro es pequeña. La gradiente de temperatura en los extremos es sin embargo, considerable. En líneas generales, los procesos de dilución y concentración de la orina se realizan de modo semejante. Las ramas ascendentes y descendentes de las horquillas túbulo-vasculares son comparables con los tubos de agua caliente y fría del sistema recién analizado.
Antes de entrar en la descripción detallada de los procesos de concentración y dilución de la orina, es necesario precisar algunos conocimientos básicos:
1. Tanto la rama ascendente, como descendente del asa de Henle son impermeables para el agua, en ausencia de la hormona antidiurética (ADH).
2. La acción de la ADH consiste en permeabilizar estas estructuras al paso del agua.
3. En el túbulo proximal, como hemos dicho anteriormente, se absorben en forma obligada las 7/8
partes del agua y del sodio filtrados en el glomérulo. O sea, de los 180 litros de agua filtrados se reabsorben 150 litros, de manera que sólo 30 litros llegan al asa de Henle. Como la reabsorción en el túbulo proximal se realiza en forma iso-osmótica, sólo estos 30 litros participan en los procesos de dilución y concentración.
MECANISMO DE CONCENTRACION DE LA ORINA

El Na+ es transportado hacia el espacio intersticial de la médula renal a través de la pared de la
rama ascendente del asa de Henle (que es imperable al agua) por un mecanismo activo desconocido. La electroneutralidad se mantiene porque el ion Cl
- sigue al Na
+ . El liquido Intersticial
se torna, así, hiperosmótico con respecto al líquido intratubular. Consecutivamente sale agua (a través de la pared) de la rama ascendente del Asa de Henle hacia le intersticio y aumenta la osmolaridad del filtrado a medida que avanza por Asa. Debido a esto se establece una gradiente osmótica en dirección de la papila renal y el líquido intratubular en la corteza esta más diluido que en las regiones más profundas, cercanas a la médula (fig.64).
La diferencia de osmolaridad entre el liquido intratubular y el del intersticio es insignificante cualquiera sea el nivel en que se analice. En otras palabras, la presión osmótica intersticial se modifica paralelamente con la intratubular. Por el contrario, la diferencia es considerable si se compara el líquido del túbulo proximal (isosmótico respecto al plasma) con el del asa de Henle (marcadamente hiperosmótico). La osmolaridad del líquido en los túbulos proximal y distal es muy semejante, pero en el túbulo distal es ligeramente hipoosmótico.
El intersticio en la médula (próxima a la papila) es altamente hiperosmótica ( osmolaridad 3-4 veces superior a la de la médula cercana a la corteza renal ) En la conservación de esta hiperosmolaridad participan los vasos rectos (vasa rec ta), que funcionan como otro de los elementos del sistema de contracorriente. En efecto, a medida que la sangre fluye hacia la
REGULACION RENAL DE LA COMPOSICIÓN Y VOLUMEN DEL Y DEL LIQUIDO EXTRACELULAR
Como ya hemos mencionado, a los riñones corresponda una participación relevante en la mantención de la constancia del medio interna a través de los siguientes mecanismos:

1. Regulación de la concentración de los electrólitos del líquido corporal. 2. Mantención de la osmolaridad del : medio interno. 3. Regulación del volumen del líquido extracelular. 4. Regulación del equilibrio ácido-base, o sea, la mantención de la concentración de iones H
+ r en
el líquido extracelular dentro de límites estrechos. Esta es una condición esencial para la conservación de la normalidad morfo-funcional de las células.
Como consecuencia del metabolismo celular, se produce en el organismo un exceso de iones H+,
que deben ser eliminados. Así, las proteínas, al ser metabolizadas, producen diariamente alrededor de 150 mEq de iones H
+. A su vez, el CO2 producido por las células se convierte en su
mayor parte en H2CO3, que se disocia y origina diariamente unos 12.500 mEq de H+, de los cuales,
sin embargo, sólo una pequeña cantidad es excretada por la orina (la mayor parte del CO2 se elimina por los pulmones).
Las células epiteliales de los túbulos dístales. proximales y colectores son capaces, si, las condiciones así lo requieren, de secretar iones H
+ hacia el líquido tubular. El mecanismo por el cual
se realiza este proceso, se ha esquematizado en la figura 66 de acuerdo con la cual: a) la excreción de hidrogeniones se inicia con la formación de H2CO3 en la célula tubular; b) en la formación de H2CO3 interviene una enzima, la anhidrasa carbónica, que cataliza la reacción C02 + H2O G H2CO3; el H2CO3 se desdóbla rápidamente en ion HCO3
- y ion H
+ y este último es
excretado a través de la membrana celular hacia el lumen tubular; c) la excreción de iones H+ está
estrechamente relacionada con la reabsorción del Na+; d) en forma análoga son secretados los
iones H+ en combinación con NH3
- para formar NH4
+, que se
intercambia con iones Na+ . Cabe señalar que el NH3
- se forma en la célula tubular por degradación
enzimática (glutaminasa y otros enzimos) del ácido glutámico y de otros aminoácidos.
Los iones H+ se combinan con los tampones que existen en la orina, de manera que el pH urinario
nunca se torna inferior a 4.0. Lo contrario ocurre cuando por algún motivo el pH de la sangre se eleva y se produce alcalosis. En este caso disminuye la excreción renal de hidrogeniones y se elimina una orina alcalina, compensándose así la tendencia a la acidosis.

FISIOLOGIA DE LA MICCION
La orina llega por los uréteres a la vejiga. Las contracciones rítmicas de la musculatura lisa de los cálices renales, expulsan la orina hacia los uréteres, cuyos movimientos peristálticos la impulsan rítmicamente hasta la vejiga.
La vejiga es un reservorio cuyas paredes tienen musculatura lisa dispuesta en 3 capas superpuestas, que forman alrededor del orificio uretral un anillo muscular que constituye el esfínter interno. En condiciones normales su capacidad es de 500-600 ml. Desde la vejiga la orina llega al exterior a través de la uretra. En el hombre existe además un esfínter externo, de musculatura estriada, ubicado alrededor de la uretra, inmediatamente por debajo de la vejiga.
Es importante mencionar que la vejiga no es un simple receptáculo, ya que es capaz de absorber tanto agua como ciertos solutos. Su musculatura conserva cierto grado de tono aun cuando esté vacía. Este tono depende de impulsos generados en el músculo liso mismo (tono miogeno), pero la micción está regulada por un centro medular. Especialmente importante es señalar que la vejiga se adapta continuamente, mediante cambios del tono de su musculatura, a los cambios de su contenido. Así, la presión intravesical sólo aumenta en grado insignificante al incrementarse su contenido. La presión se incrementa, sin embargo, durante la micción, lo que facilita el paso de la orina a la uretra y luego al exterior.
La contracción de la vejiga se produce por mecanismos nerviosos reflejos inducidos por la distensión de sus paredes. La contracción se acompaña de relajación refleja del esfínter interno. El esfínter externo, controlado por la voluntad, se relaja y la orina fluye por la uretra hacia el exterior. El esfínter externo, en condiciones normales, puede contraerse voluntaria y simultáneamente con la contracción de la musculatura vesical y con la relajación del esfínter interno, e impedir, dentro de ciertos límites, el vaciamiento. El vaciamiento de la vejiga es, por lo tanto, en último término, un fenómeno voluntario que se inicia con la contracción refleja de su musculatura y relajación del esfínter interno; luego aumenta la presión intra-abdominal, por contracción de los músculos del abdomen, y se relaja voluntariamente el esfínter externo.
La vejiga tiene doble inervación, simpática y parasimpática, cuyas fibras se distribuyen profusamente en la musculatura vesical. El sistema parasimpático, representado por los nervios pélvicos, formados por fibras parasimpáticas sacras, produce la contracción de la musculatura de la pared vesical y relajación del esfínter interno. El simpático, por su parte, disminuye el tono de la musculatura de la vejiga e incrementa el tono del esfínter interno. El esfínter externo está inervado por fibras nerviosas motoras somáticas, provenientes de la médula sacral.
Por lo tanto, el vago estimula y el simpático inhibe el vaciamiento de la vejiga. La influencia del simpático es de escasa importancia.
El centro nervioso que controla el vaciamiento vesical está ubicado en la médula sacra (Fig. 67). En condiciones fisiológicas el vaciamiento no se inicia por el aumento de la presión intravesical, sino por la distensión de sus paredes, la que estimula receptores allí ubicados. Estos receptores emiten impulsos hacia el centro medular, desde el cual se transmiten impulsos por las vías eferentes simpática y parasimpática, que van a producir el vaciamiento. A pesar de que al evacuarse la orina disminuye la distensión vesical, no cesa el vaciamiento, lo que se debe a la estimulación, por el flujo de orina, de receptores situados en la uretra. Estos receptores mantienen, por vía refleja, la contracción de la musculatura de la vejiga y la relajación de los esfínteres tanto interno como externo.

La destrucción del centro reflejo sacral produce retención de orina y aumento de la presión intravesical. En esta condición la vejiga no se distiende, por no producirse la disminución del tono que, como ya se ha señalado, ocurre normalmente al aumentar su contenido. En este caso el vaciamiento se hace por rebalse y en el momento en que la presión intravesical vence la resistencia ofrecida por el esfínter externo (incontinencia pasiva). Al prolongarse esta situación, la elevada presión intravesical repercute
sobre los uréteres, la pelvis renal y el riñón mismo, produciendo hidronefrosis que altera el funcionamiento renal.
Si se seccionan las raíces posteriores sacrales, interrumpiéndose así las fibras aferentes que transmiten los impulsos generados por la distensión de la vejiga y que van hacia el centro sacral, desaparece el vaciamiento reflejo, pero se conserva el voluntario. En este caso la micción puede realizarse, aunque con gran dificultad inicial, dificultad que disminuye progresiva y substancialmente con el tiempo.
Conjuntamente con el centro medular sacral, ciertos centros superiores, situados principalmente en la región bulbo-protuberancial, ejercen influencia sobre el vaciamiento vesical.

SECRECIONES EN EL TUBO DIGESTIVO
BUCAL
Como hemos expuesto, los alimentos provienen del medio externo y deben ser degradados antes de incorporarse al medio interno del organismo. El tubo digestivo es sólo una invaginación del medio externo en nuestro cuerpo, desde la cual los alimentos, previa preparación y degradación hidrolítica, llegan, atravesando la mucosa intestinal, a la sangre y a la linfa. La circulación lleva los materiales absorbidos hasta los capilares, desde los cuales pasan al espacio intersticial y de éste a las células.
La degradación de los alimentos, o sea, su digestión, se realiza bajo el efecto de los llamados jugos digestivos, secretados por diversas glándulas que constituyen un verdadero sistema glandular. La acción digestiva de estos jugos se debe esencialmente a los enzimos que contienen. La digestión se inicia como ya hemos dicho, en la boca, donde la saliva, producida por tres pares de glándulas (parótidas, sublinguales y submaxilares), ejerce su efecto. La saliva secretada por estas glándulas tiene diferente composición. La saliva de la glándula parótida es seroacuosa, mientras la de las otras dos glánulas, en que predominan las células mucígenas y serosas, es mucoserosa.
La saliva contiene 99.3% de agua y sólo 0.7% de residuo seco (amilana, maltasa, mucina, albúmina, globulina y una fracción ínfima de sales). La saliva humana contiene un enzimo, la amilana salival o ptialina, que es capaz de desdoblar el almidón. La acción de esta amilasa no es, sin embargo, indispensable. La ptialina es capaz de desdoblar el almidón hasta glucosa, pero requiere para este efecto un contacto prolongado con el subtrato. Como el bolo alimenticio pasa rápidamente de la boca al estómago, el tiempo que permanece en la cavidad bucal, faringe y esófago no es suficiente para que la ptialina ejerza su acción digestiva. En el hecho, la mayor parte de la digestión salival se realiza en el estómago, antes de que el bolo se haya acidificado, ya que la ptialina actúa sólo en un medio neutro o alcalino. La degradación del almidón por efecto de la amilasa salival es incompleta y su intervención no es indispensable. Puede ser reemplazada totalmente por la amilasa pancreática.
La acción amilolítica de la saliva es, por lo tanto, de importancia secundaria. La saliva cumple, sin embargo, otras funciones de gran importancia, entre las cuales caben señalar las siguientes:
a) desempeña un papel importante en la percepción del gusto, dado que las terminaciones de los nervios gustatorios en la mucosa bucal sólo se excitan si las sustancias están disueltas en la saliva;
b) humedece los labios y la cavidad bucal, facilitando en esta forma la articulación del
lenguaje y contribuyendo a mitigar la sensación de sed;
c) contribuye a la formación y lubricación del bolo alimenticio, embebiendo los materiales
triturados por la masticación, lo que facilita su aglutinación y su deglución.
Existe, durante la vigilia, una secreción básica y continua de saliva, cuya intensidad varía según las necesidades. La secreción aumenta con la ingestión de alimentos y puede alcanzar hasta 1.5 litros/24 horas. El 75% de la saliva es secretada por la glándula submaxilar. En general la saliva es

casi totalmente reabsorbida en el tubo digestivo, evitándose así la pérdida de una cantidad considerable de líquido.
El paso de los electrólitos del plasma a la saliva se realiza contra gradiente de concentración, lo que requiere energía.
Las glándulas salivales están inervadas tanto por el parasimpático como por el simpático. La estimulación de este último produce, en las glándulas submaxilares y sublinguales, una saliva escasa pero rica en sustancias orgánicas. El simpático carece de efecto sobre la glándula parótida.
El parasimpático induce la secreción de una saliva abundante pero pobre en sustancias orgánicas, tanto en las submaxilares y sublinguales, como también en las parótidas.
La ingestión de alimentos es el estímulo normal para la intensificación de la secreción salival. La secreción se produce por un mecanismo reflejo incondicionado que se inicia por la estimulación química y mecánica de receptores gustativos y de mecanorreceptores, ubicados en la mucosa de la cavidad bucal, especialmente en la lengua. Los impulsos generados en estos receptores son transmitidos a los centros de secreción salival, localizados en el bulbo raquídeo, por fibras aferentes de nervio lingual (rama del trigémino y gloso-faríngeo). Los centros bulbares envían a las glándulas impulsos secretorios por los nervios eferentes que ya han sido mencionados.
La secreción salival refleja puede iniciarse no sólo por reflejo incondicionado (ingestión de alimentos), sino también por estímulos visuales, acústicos, olfatorios, relacionados con la preparación de los alimentos o que se asocian con el acto de su ingestión (reflejos condicionados).
La intensidad de la secreción de saliva y su composición dependen de la calidad del alimento. Los alimentos secos, por ejemplo, producen mayor volumen de saliva que los líquidos. Los ácidos producen una salivación rica y abundante en prótidos, que actúan como tampones regulando el pH salival.
GÁSTRICA
El estómago desempeña en la digestión las siguientes funciones:
1. El fundus almacena los alimentos ingeridos. En esta parte continúa la acción de la
amilasa salival en el centro del bolo de alimentos (su superficie está en contacto con el HCl secretado por la mucosa, que inhibe a la amilasa).
2. El antro funciona como un esfínter que controla el vaciamiento gradual del estómago e impide su vaciamiento demasiado rápido. Facilita también su trituración y actúa al mismo tiempo como bomba (bomba antro-píloro-duodenal) que impulsa el contenido hacia el duodeno.
3. En el cuerpo del estómago se realiza la homogenización y cierto grado de isotonización de los alimentos. La mucosa del cuerpo secreta la pepsina, un enzimo que inicia la digestión de las proteínas. Las grasas son emulsionadas por la acción mecánica de las contracciones.
El estómago no es indispensable para el proceso digestivo, pero es, sin embargo, esencial para la neoformación de glóbulos rojos. En efecto, el factor intrínseco, producido por la mucosa gástrica es absolutamente necesario para la absorción del factor extrínseco (vitamina B12), elemento vital en

la hematopoyesis. Se puede considerar ésta como la única función importante, aunque no vital, del estómago. Aun esta función puede ser, como en las personas gastrectomizadas, por ejemplo, reemplazada por el suministro parenteral de la vitamina mencionada. E1 factor intrínseco es secretado por las células parietales.
La composición del jugo gástrico depende de la actividad de las células de su mucosa. Su pH es normalmente muy bajo (1.0 a 1.5), por el HCl secretado por la mucosa. La mucosa secreta también un enzima proteolítico llamado pepsina (ya mencionado) y dos diferentes tipos de mucus: uno soluble y otro insoluble en agua. El jugo gástrico contiene gran cantidad de agua, electrólitos (Na
+,
K+), el factor intrínseco, al cual nos hemos referido anteriormente, y dos enzimos de cierta
importancia, pero en modo alguno indispensables: la quimosina y la lipasa.
La pepsina, secretada por las células principales o zimógenas de la mucosa del fondo y cuerpo del estómago, es un enzimo proteolítico que rompe parcialmente las uniones peptídicas de las proteínas. La pepsina se encuentra en las células secretoras en su forma inactiva, denominada pepsinógeno, que se convierte en pepsina bajo el efecto del HCl. Es liberada por las células bajo la acción de estímulos vagales o del contenido gástrico mismo. El pH óptimo para su activación es de 2.00 menos. A un pH superior a 3.0 pierde su efecto proteolítico. Es absorbida en el intestino y circula con la sangre, siendo eliminada por los riñones como uropepsina. Su acción proteolítica consiste en la hidrólisis de las proteínas hasta la producción de polipéptidos, peptonas y proteosas y sólo en grado muy reducido, aminoácidos. E1 otro enzimo producido por la mucosa gástrica, la quimosina, se encuentra solamente en los lactantes y coagula (cuaja) la leche en presencia de Ca
++
La lipasa se encuentra en cantidad ínfima en el jugo gástrico. Es un enzimo lipolítico de importancia muy reducida.
Las células parietales u oxínticas del fundus y del cuerpo del estómago secretan HCl en cantidades tan elevadas que el órgano es una verdadera fábrica de este ácido. A excepción de su efecto activador del pepsinógeno, el HCl no tiene participación
en la digestión gástrica. Cabe mencionar que ejerce cierta acción antiséptica.

En cuanto al mecanismo mismo de la secreción clorhídrica, Bradford y Davies han demostrado que el líquido dentro de las células parietales es de reacción neutra, en tanto que el líquido en los canalículos intercelulares es fuertemente ácido. Los canalículos intercelulares representan la continuación de un sistema de conductos intracelulares (Fig. 69), cuyo contenido es también fuertemente ácido. Este hecho justifica la afirmación de los investigadores mencionados de que la secreción de HCl se realiza a través de la membrana que separa los canalículos intracelulares del citoplasma. La secreción misma es un proceso eminentemente activo, que involucra el paso de iones H
+ de la sangre a las células parietales y de aquí a los conductos intracelulares. La
concentración de iones H+ en estos conductos es de unos 4 millones de veces mayor (pH 2.0) que
en la sangre (pH 7.4). Este transporte se realiza en contra de una gradiente de concentración y de una gradiente eléctrica, y es evidentemente un proceso activo que consume una considerable cantidad de energía. Debido al paso de una gran cantidad de iones H
+ de la sangre a los
canalículos y al transporte simultáneo de iones HC03- en dirección apuesta, la sangre venosa que
sale del estómago, como asimismo la orina, se hacen alcalinas durante los períodos digestivos (marea alcalina post-prandial).
El mecanismo íntimo de la secreción no ha sido totalmente esclarecido, pero se conocen algunos de sus aspectos más importantes. Se ha demostrado que las células parietales contienen anhidrasa carbónica, enzimo que cataliza la reacción CO2 + H2O = H2CO3. Al estimular estas células con histamina, por ejemplo, se intensifica en el lado mucoso de la pared gástrica la secreción de HCl, y en el lado seroso la de cantidades equivalentes de iones HCO3
-. Sobre la base de estos resultados, se ha planteado la hipótesis de que en el
proceso de formación de HCl, los iones H+, producidos en ciertos procesos oxidativos de la célula
parietal, son excretados activamente hacia los canalículos intracelulares. El excedente de iones OH
- (alcalinos) son tamponados por el H2C03 intracelular formándose iones bicarbonato HCO3 (que
difunden al líquido extracelular que rodea la serosa) y H2O (HCO3 + OH- = HCO3
- + H2O). Esta
reacción es también catalizada por la anhidrasa carbónica.
La energía requerida para el transporte de los iones de H+ a los canalículos intracelulares proviene
de procesos oxidativos cuya supresión hace desaparecer la secreción del HCl.
El Cl- del HCl gástrico proviene del plasma sanguíneo, desde el cual las células parietales lo
transportan al lumen de los canalículos intracelulares. Este transporte se realiza en contra de la gradiente de concentración y del potencial negativo de la superficie de la mucosa. Las células parietales poseen una bomba de Na
+ que transporta este ion hacia la superficie serosa de la
mucosa. Resulta, en esta forma que, mientras en su polo seroso las células se liberan de Na+, o
sea de cargas positivas, en su polo mucoso expulsan Cl-, estableciéndose así una diferencia de
potencial entre la superficie de la mucosa (que es negativo) y serosa (que es positivo).
La mucosa gástrica y la del intestino secretan, bajo el efecto de glucosa suministrada por vía oral, una hormona hiperglucemiante. La glucosa así administrada estimula la mucosa gástrica e intestinal para secretar una hormona que intensifica la entrega de glucagón. Este, a su vez, eleva el nivel glucémico (ver Endocrinología). La hiperglucemia estimula las células beta del páncreas y aumenta consecutivamente la entrega de insulina a la circulación. Razón por la cual la glucosa administrada por vía oral tiene mayor efecto sobre la secreción de insulina que la inyectada por vía venosa.
La pancreozimina es otra hormona secretada por la mucosa duodenal y gástrica que regula, además de la secreción enzimática del páncreas, la de una hormona con propiedades similares al glucagon. Actúa probablemente también en forma directa sobre las células 13 del páncreas, aumentando la secreción y/o entrega de insulina.
REGULACION DE LA SECRECION GÁSTRICA

La secreción del jugo gástrico está controlada tanto por mecanismos nerviosos como humorales. Los experimentos del famoso fisiólogo ruso Iván Pavlov demostraron que se inicia ya antes que el bolo alimenticio llegue al estómago y es influida por numerosos factores. Este investigador observó que en un perro en el cual se realiza una alimentación falsa, o sea, se impide mediante una fístula esofágica que el alimento ingerido llegue al estómago (Fig. 70), se produce una secreción gástrica intensa al entrar el alimento a la boca. Comprobó, además, que después de repetir varias veces este experimento, no era necesario que el alimento penetrase a la cavidad bucal, bastaba la visión u olfación de la comida para provocarla. Estos resultados indican que la fase inicial de la secreción se produce por mecanismos incondicionados y condicionados. En el caso de los reflejos incondicionados (comida falsa), los impulsos que inducen la secreción gástrica parten desde receptores gustatorios de la cavidad bucal, los cuales son transmitidos por vías nerviosas aferentes hasta centros reflejos en la médula espinal. Desde estos centros, impulsos eferentes son transmitidos por fibras vagales al estómago. La secreción condicionada es inducida por impulsos generados en receptores situados fuera del sistema digestivo (visuales, acústicos, olfatorios, etc.). Las sensaciones desagradables, estados emocionales negativos que producen asco y repulsión, inhiben la secreción refleja condicionada del jugo gástrico. Las secreciones reflejas incondicionada y condicionada constituyen en conjunto la fase cefálica o vagal de la secreción gástrica.
Los alimentos, una vez en el estómago, intensifican la secreción, pero no por un efecto mecánico, sino por estimulación química de la mucosa, especialmente en la región pilórica. Son estímulos químicos especialmente efectivos el caldo de carne, el alcohol y ciertos condimentos. Tanto las proteínas como los derivados de su degradación (proteosas y otros) representan estímulos menos efectivos. Esta fase de la secreción, llamada fase gástrica o química, es la continuación de la fase vagal.
Se ha demostrado que la fase gástrica de la secreción se debe a una hormona producida por la mucosa pilórica. Esta hormona, llamada gastrina, llega por vía humoral a las células parietales y estimula intensamente su secreción. La secreción de gastrina, a su vez, es estimulada por las sustancias recién mencionadas (caldo de carne, condimentos, proteosas, etc.). Se pueden diferenciar en la secreción gástrica, tres fases: una primera inducida por la estimulación de receptores visuales, olfatorios, auditivos, y de los receptores que están en la cavidad bucal; una segunda fase inducida por la gastrina y finalmente una tercera fase llamada intestinal.
El alimento, parcialmente digerido en el estómago, penetra al duodeno donde estimula receptores, ubicados en la mucosa duodenal, e induce secreción de una hormona, probablemente idéntica con la gastrina y que por vía humoral estimula también la secreción de jugo gástrico. Esta secreción es especialmente intensa y rica en pepsina cuando el contenido duodenal es rico en proteínas y

compuestos resultantes de su degradación y menos intensa si es rico en grasas. Las grasas inducen en la mucosa del duodeno la secreción de otra hormona, la enterogastrona, que al llegar por la circulación a las células parietales del estómago, inhibe su secreción. La enterogastrona disminuye también la motilidad gástrica por un mecanismo reflejo viscero-visceral inducido por la distensión del duodeno. La distensión duodenal es, por consiguiente, un factor importante para la regulación del vaciamiento gradual del estómago. Una comida rica en grasas permanece, por lo tanto, un mayor tiempo en el estómago.
En resumen, se puede decir que la secreción del jugo gástrico se inicia antes que los alimentos sean ingeridos (estimulación visual, olfatoria, etc.), y se mantiene a un nivel adecuado por la presencia de los alimentos en la boca, en el estómago y en el duodeno, para volver a su nivel normal en los períodos interdigestivo.
En la segunda parte del duodeno, el contenido duodenal se torna alcalino, permitiendo así la acción del jugo pancreático y de otros jugos digestivos que requieren, para ser efectivos, un pH superior a 7.0.
Si se hace actuar proteínas, proteosas u otros secretagogos sobre la mucosa de un antro pilórico transplantado fuera de la cavidad abdominal y cuya inervación está eliminada, se produce secreción en el estómago. Este experimento demostraría que la mucosa antral ha secretado gastrina, que llega por vía humoral al estómago e induce la secreción.
Esta hipótesis se tornó, sin embargo, incompleta debido a diversos experimentos que han demostrado que la llegada de alimentos al estómago estimula terminaciones vagales en la mucosa gástrica y por vía refleja inducen al antro a secretar gastrina. Al mismo tiempo, el vago ejerce una acción permisiva sobre las células las parietales, capacitándolas para secretar, así que la separación entre una fase vagal (refleja) y otra antral (humoral) parece insostenible.
Ni la estimulación del vago, ni la distensión del antro, tienen separadamente influencia sobre la secreción gástrica, pero juntas potencializan mutuamente sus efectos e intensifican la secreción.
La secreción del HCl debe mantenerse dentro de ciertos límites ya que una acidez excesiva puede causar ulceraciones de la mucosa del estómago. Para este fin se dispone de varios mecanismos, entre los cuales la acidez misma es de gran importancia: una acidez excesiva inhibí la secreción.
La secreción del estómago está regulada, por lo tanto, por un conjunto de mecanismos como puede apreciarse en la figura 71.
En resumen, los alimentos, al distender el estómago, estimulan en la mucosa antral la secreción de gastrina, la cual al llegar por la circulación a las células parietales, incrementa la secreción del HCl. El contenido ácido del estómago y del antro, a su vez, inhibe la secreción de gastrina y, consecuentemente, la secreción de HCl. Como el contenido gástrico sigue distendiendo al estómago, en la mucosa antral se secreta nuevamente gastrina. Al iniciarse el vaciamiento del estómago, llega el contenido gástrico al duodeno y activa en éste otro mecanismo de regulación que consiste en la secreción de enterogastrona que actúa y participa en la modulación de la secreción y motilidad gástricas.
El contenido gástrico, una vez en el duodeno, estimula en la mucosa duodenal la secreción de varias otras hormonas: a) la secretina, que activa la secreción y entrega de jugo pancreático con un alto contenido de bicarbonato y agua; b) la pancreozimina, bajo cuya acción el páncreas secreta un jugo rico en enzimas digestivas.

La producción de pancreozimina es intensificada especialmente cuando el contenido duodenal es rico en proteínas. Esta hormona ejerce además los siguientes efectos: facilita el flujo de bilis, aumenta la secreción de mucus por las células de Brunner en el intestino e inhibe la secreción de las células parietales. La pancreozimina tiene efecto similar al glucagon, hormona hiperglucemiante, y al actuar además directamente sobre las células b del páncreas, aumenta la secreción y entrega de insulina.
La pancreozimina ejerce todavía otras acciones que anteriormente han sido atribuidas a otra hormona, a la colecistoquinina. Investigaciones posteriores revelaron, sin embargo, que esta última es idéntica con la pancreozimina. Por consiguiente, los efectos abajo enumerados y anteriormente atribuidos a la colecistoquinina, son causados por la pancreozimina. Entre estos efectos hay que destacar por su importancia, los siguientes:
1. Estimula la síntesis y entrega de los enzimos pancreáticos. 2. Al mismo tiempo relaja el esfínter de Oddi y produce la contracción de la vesícula y de los
conductos biliares. 3. Inhibe la secreción gástrica, cumpliendo así la función de una hormona reguladora de dicha
secreción. 4. Induce en las células a y b de los islotes de Langerhans pancreáticos la secreción de glucagón e
insulina (ver Páncreas Endocrino).
En resumen, la mucosa duodenal secreta 3 hormonas: la enterogastrona, poderosa inhibidora de la motilidad y secreción gástrica, cuya secreción se intensifica si el contenido gástrico es rico en grasas; la secretina, inhibidora de la secreción gástrica y activadora de la secreción de un jugo pancreático rico en bicarbonato y en agua; la pancreozimina cuyas acciones acabamos de enumerar.
Además de la acidificación de la mucosa antral, existen, por consiguiente, por lo menos tres mecanismos humorales de origen duodenal, que al mismo tiempo que modifican cuantitativa y cualitativamente la secreción de jugo pancreático, ejercen una acción inhibidora sobre la secreción de jugo gástrico y de gastrina: la enterogastrona, la secretina y la pancreozimina.
Se acepta que el término regulación enterogástrica comprende genéricamente el conjunto de los efectos de todas las hormonas duodenales tales como secretina, pancreozimina y otras.
INTESTINAL

Pancreas
El páncreas es de mucha importancia para la digestión de los alimentos, tanto por la acción de las enzimas de su secreción exocrina, como asimismo por su efecto neutralizante del contenido ácido del estómago que llega al duodeno. Su actividad secretora exocrina está controlada por mecanismos nerviosos y humorales. Existen en el páncreas del hombre y algunas especies animales, además de las células responsables de su secreción exocrina (digestiva), agrupaciones celulares que forman los islotes de Langerhans. La función de éstos no está relacionada con la digestión, sino que secretan ciertas hormonas (insulina y otras), que serán tratadas en el capítulo correspondiente.
Los jugos digestivos producidos por las glándulas del intestino mismo o anexas a él son en general ricos en bicarbonato que no sólo neutralizan el contenido duodenal, sino que lo tornan débilmente alcalino. Los jugos digestivos aumentan considerablemente el volumen del contenido intestinal (el volumen del jugo pancreático es alrededor de 1 l/24 hrs. y el de las otras glándulas puede alcanzar a 3 litros en el mismo tiempo).
El jugo pancreático contiene, además de agua, Na+ y K
+ en una concentración igual a la del
plasma. El ion de mayor importancia es el HCO3-, cuya concentración es aproximadamente 5 veces
mayor que en la sangre. Los componentes orgánicos del jugo pancreático son principalmente enzimos digestivos y/o sus precursores. Entre los enzimos son especialmente importantes la tripsina, la quimiotripsina y la carboxipeptidasa, La tripsina actúa lentamente sobre las proteínas (ovoalbúmina, colágeno, hemoglobina, etc.) y en forma más rápida sobre las proteínas desnaturalizadas por cocción. Los productos de degradación producidos por la tripsina, son digeridos por la quimiotripsina que los hidroliza gradualmente hasta peptonas, polipéptidos y finalmente aminoácidos. La carboxipeptidasa, rompe las uniones terminales peptídicas vecinas al grupo carboxílico.
El jugo pancreático contiene además otros dos enzimos relacionados con la digestión de las proteínas, la ribonucleasa y la desoxicarbonucleasa, cuya función es degradar a los polinucleótidos hasta mononucleótidos.
El páncreas interviene, mediante la lipasa pancreática, en la digestión de las grasas degradándolas hasta ácidos grasos y monoglicéridos. Los ácidos grasos se combinan con los álcalis intestinales para formar jabones, que son solubles en agua. Las sales biliares disminuyen la tensión superficial entre agua y grasas neutras, permitiendo una fina emulsión de estas últimas, lo que aumenta enormemente la superficie que presentan las grasas a la acción de la lipasa y facilita en esta forma su acción digestiva.
La amilasa, otro enzimo pancreático, interviene en la digestión de los hidratos de carbono, transformando el glucógeno y el almidón en maltosa. La maltosa, bajo el efecto de la maltasa pancreática, se convierte en glucosa y como tal pasará por la mucosa intestinal a la circulación. E1 organismo humano carece de b -amilasa, que en los herbívoros facilita la digestión de la celulosa. Por esta razón el ser humano es incapaz de aprovechar la celulosa.
Tres mecanismos regulan la secreción pancreática. Un mecanismo nervioso reflejo (incondicionado), por el cual el vago incrementa la secreción de un jugo pancreático rico en enzimos y de densidad alta. El simpático, no modifica el volumen de la secreción, pero disminuye su contenido en enzimos. Se ha demostrado que los receptores para este reflejo son quimioceptores (receptores gustatorios) localizados en la boca, su mecanismo de acción es igual al descrito para la secreción gástrica. E1 nervio aferente es el vago y los impulsos eferentes (secretores) llegan al páncreas también por vía vagal.

El segundo mecanismo nervioso es de carácter reflejo condicionado. Como la secreción del jugo pancreático se inicia casi simultáneamente con la del jugo gástrico, parece lógico suponer que es inducida en su etapa inicial por un reflejo condicionado, similar al que estimula la secreción gástrica.
Finalmente existe un mecanismo hormonal, demostrado por la observación de que el contenido gástrico, una vez en el duodeno, induce la secreción aun en un páncreas totalmente denervado. Pero la cantidad y composición del jugo pancreático así producido es diferente al que se obtiene por estimulación vagal. Su volumen es mayor pero su contenido de enzimos es menor. De acuerdo con los experimentos de Pavlov, el contenido ácido del duodeno induciría en éste la producción de una sustancia que llega por vía sanguínea al páncreas, estimulando su actividad secretora. Cualquier estímulo mecánico de esta mucosa puede causar el mismo efecto. La hipótesis de Pavlov fue comprobada a principios de este siglo por Bayliss y Starling, quienes observaron que la presencia de un ácido, o sea, la acidificación de la mucosa duodenal denervada, intensifica también la secreción en el páncreas de nervado. Este efecto no se obtiene por el suministro de ácido por vía endovenosa. La sustancia liberada en la mucosa duodenal bajo la acción de la acidez y que actúa sobre el páncreas por vía humoral, es la secretina. Como ya se ha mencionado, la secretina estimula la secreción de un jugo pancreático abundante, pobre en enzimos pero rico en álcalis. El jugo pancreático alcalino neutraliza la acidez del contenido duodenal, pero tiene escasa acción digestiva. La neutralización del contenido duodenal es de gran importancia, ya que los jugos digestivos sólo son efectivos en medio neutro 0 alcalino. Neutralizado el contenido duodenal, la secretina y con ella la secreción del páncreas se detiene por un mecanismo de retroalimentación negativa. Es decir, el contenido duodenal neutro inhibe la secreción del jugo pancreático. Los extractos de mucosa duodenal, suministrados por vía sanguínea, intensifican la secreción pancreática rica en enzimos. Esto demuestra que la mucosa del duodeno secreta, además de secretina, otra hormona, la pancreozimina, cuyo efecto sobre el páncreas es similar al obtenido por la estimulación vagal (jugo rico en enzimos). La secreción de la pancreozimina es intensificada por los productos de la digestión de las proteínas y grasas.
En resumen, se puede decir que la secreción pancreática es regulada por los siguientes factores:
1. Gastrina 2. Secretina 3. Pancreozimina 4. Actividad Vagal.
La importancia del factor vagal es muy discutida. Los experimentos de los últimos años parecen indicar que la acción del vago es sólo permisible, es decir, condiciona las células del páncreas para que, bajo el efecto de los factores humorales mencionados, inicie su secreción. El vago, por lo tanto, no es el nervio secretor del páncreas.
En resumen: la secreción pancreática, a semejanza de la del estómago, tiene una fase cefálica (refleja) y una fase intestinal (humoral). La secreción de la fase intestinal es cuantitativamente superior y de mayor duración que la inducida por los mecanismos reflejos. Sin embargo, el nervio vago parece tener una participación importante como factor permisible en la fase humoral ya que los enzimos y sus precursores no son excretados por la glándula privada de su inervación vagal. En la figura 72 se representan

en forma esquemática los diferentes factores que regulan la secreción del jugo pancreático.
La mucosa del intestino delgado tiene una doble función, en cuanto interviene no sólo en la digestión, sino también en la absorción de los alimentos. Estas funciones están reguladas y sincronizadas conforme a las necesidades del momento. La secreción del intestino delgado, a diferencia de las del estómago y duodeno, es continua, pero de intensidad variable. La distensión del intestino estimula la secreción (por un mecanismo nervioso reflejo cuya vía eferente es cl nervio vago) de un jugo digestivo rico en enzimos. El simpático tiene efecto inhibidor de la secreción. Hay que recalcar que este jugo digestivo permanece en el ribete estriado y pasa al lumen intestinal arrastrado por los alimentos. Se han obtenido evidencias experimentales importantes, aunque no irrefutables, de que existe también una regulación humoral de la secreción intestinal. Así, se ha logrado obtener de la mucosa intestinal un extracto cuyo suministro endovenoso aumenta no sólo el volumen del contenido intestinal, sino también sus enzimos. La mucosa elaboraría, por lo tanto, una hormona que se ha denominado enterocrinina, la cual induciría la secreción por vía humoral.
El intestino grueso tiene como función principal la formación de las heces. El contenido del intestino delgado pasa al ciego en forma periódica, debido, como ya hemos explicado, a las contracciones del intestino delgado y a la relajación periódica del esfínter íleo-cecal. El paso del alimento al intestino grueso se efectúa aproximadamente 4 a 6 horas después de su ingestión. El contenido del intestino grueso careo de sustancias aprovechables por el organismo. La función de este segmento del tubo digestivo es la formación de heces a partir de las sustancias no aprovechables y de los jugos digestivos no reabsorbidos. Además secreta mucina, secreción que es inducida por la distensión causada por la llegada de contenido desde cl íleon. A su nivel, se absorbe prácticamente sólo agua y sales. En el recto y en la última porción del sigmoídeo existe un plexo venoso submucoso, que permite la absorción de ciertos líquidos, sustancias alimenticias y fármacos suministrados por vía rectal. Estas sustancias pasan por la vena cava inferior directamente a la circulación general sin circular previamente por el hígado. Esto evita la acción inactivadora que el hígado ejerce sobre las sustancias absorbidas.
El intestino grueso tiene una rica flora bacteriana, especialmente coli y lactobacilos, que degradan la celulosa de los alimentos de origen vegetal. Estos pueden así ser digeridos. El papel de la flora

bacteriana es en el hombre de poca importancia, pero no así en los animales herbívoros, en los cuales cumplí una función indispensable para la digestión de los vegetales ingeridos. Los antibióticos de amplio espectro, pueden esterilizar el. intestino grueso y producir cuadros carenciales de cierta importancia.
ABSORCION INTESTINAL
Se entiende como tal el paso de las sustancias alimenticias desde el lumen del tubo digestivo (que es en realidad para del medio externo), a través de la mucosa a la sangre circulante, al espacio intersticial y a la célula, o sea, a nuestro medio interno. La velocidad de absorción depende del tamaño molecular, de la hidrosolubilidad y de la carga eléctrica de los alimentos absorbidos.
Salvo el caso de algunos fármacos (adrenalina,, nitroglicerina, etc.), no existe absorción a nivel bucal.
En el estómago la absorción es insignificante excepto al alcohol que, en presencia de CO2, se absorbe rápidamente y en volúmenes relativamente grandes.
La absorción se realiza casi en su totalidad en el intestino delgado. El gran número de vellosidades aumenta la superficie de la mucosa intestinal, lo que facilita la absorción. Por el centro de cada vellosidad corrí un vaso linfático rodeado por una arteria y una vena. La arteria da origen a una red capilar que desemboca en la vena central (Fig. 73). Las venas centrales de
las diferentes vellosidades confluyen para desembocar finalmente en la vena porta.
El vaso linfático central de la vellosidad lleva las sustancias absorbidas a través del conducto torácico, a la circulación general. Estas sustancias llegan a los tejidos, sin previo paso por el hígado.
Las vellosidades están, durante el paso de los alimentos, en movimiento constante. Sin embargo, ni estos movimientos, ni la gran superficie de la mucosa intestinal con la cual los alimentos están

en contacto, explican satisfactoriamente ni la gran velocidad, ni la selectividad con que se absorben los diversos materiales.
Las células epiteliales del intestino poseen, como todas las células del organismo, una membrana limitante formada por dos capas proteicas entre las cuales hay una sustancia fosfolipídica.
Las sustancias que pasan del lumen intestinal a la sangre y/o a la linfa, atraviesan por consiguiente las siguientes capas:
1. La capa proteica externa de la membrana celular. 2. La sustancia fosfolipídica que se encuentra entre 1 y 3. 3. La capa proteica interna de la membrana celular. 4. El protoplasma de la célula. 5. La membrana celular para salir de la célula. 6. Los espacios intercelulares. 7. La membrana basal. 8. El espacio intersticial. 9. La pared de los capilares sanguíneos y/o linfáticos.
Debido al carácter fosfolipídico de la membrana celular sólo las grasas y materiales liposolubles pueden atravesar la membrana libremente. Las sustancias hidrosolubles como los hidratos de carbono, por ejemplo, pasan libremente por los canalículos intercelulares, membrana banal, espacio intersticial y pared capilar, cuyos poros tienen un diámetro mayor que el tamaño molecular del agua y las sustancias hidrosolubles.
Las moléculas de agua y los iones inorgánicos son absorbidos de acuerdo con la gradiente de concentración, es decir, migran del lugar de mayor al de menor concentración. Por eso pasa, por ejemplo, el agua desde el lumen intestinal hacia el espacio intersticial, y luego a la sangre (el contenido intestinal es hipotónico, o sea, su concentración de agua es mayor en relación al líquido intersticial y al plasma). En el caso de que una solución en el lumen intestinal tenga mayor presión osmótica, es decir, que sea hipertónica, las sustancias disueltas migrarán, debido a la gradiente de concentraciones, desde el intestino hacia la sangre. Pero el agua debe migrar previamente de la sangre, (donde su concentración es mayor), al intestino, e igualar las concentraciones.
Dado que la diferencia de las presiones osmóticas es una de las fuerzas que regula la dinámica del intercambio de líquidos, para comprender el mecanismo de la absorción es indispensable conocer la magnitud de las presiones osmóticas que prevalecen en el lumen intestinal, en el espacio intersticial y en la sangre. Es importante recordar que la presión coloidosmótica del plasma sanguíneo es siempre superior a la del contenido intestinal. La presión sanguínea en los capilares de las vellosidades intestinales es relativamente alta, fluctuando alrededor de los 30 mm de Hg. La presión hidrostática intraluminal depende de la fuerza de las contracciones del intestino. En condiciones normales, mientras la intensidad de las contracciones no sobrepasa ciertos límites, la presión no es superior a los 10 mm de Hg.
En el intestino grueso, en el cual las contracciones son muy intensas, la presión hidrostática puede no sólo hacerse igual, sino superar a la presión capilar, produciendo así el paso de agua del lumen intestinal a la sangre. E1 volumen de agua reabsorbida es directamente proporcional a la diferencia de presiones entre el lumen intestinal y capilar arterial.
Se acepta actualmente que la absorción depende de la presión coloidoosmótica, siempre que la presión hidrostática del contenido intestinal es inferior a la presión coloidoosmótica de la sangre. En el paso de agua y el transporte de sustancias, influye también la gradiente de potencial electroquímico, dependiendo de la diferencia de concentraciones para las sustancias que carecen de carga eléctrica. Sin embargo, en algunos casos la absorción puede realizarse en contra de la gradiente de concentraciones. Además, se observa con cierta frecuencia que de dos sustancias

con estructura similar y a igual concentración, se absorbe más rápidamente aquella cuyo peso molecular es mayor. Estos hechos hacen necesario admitir que la absorción intestinal no se realiza siempre de acuerdo con las gradientes de concentración, sino que en algunos casos participan procesos de transporte activo, que requieren energía.
AGUA Y SALES
El agua migra, de acuerdo con lo dicho anteriormente, del lugar de su mayor al de su menor concentración. Por lo tanto, una solución hipertónica en el intestino se torna rápidamente isotónica por el paso de agua dude los capilares hacia el lumen intestinal. El agua puede, sin embargo, pasar del intestino a la sangre, siempre que su concentración intraluminal sea superior a la concentración que tiene en la sangre, o sea, que el contenido intestinal sea hipotónico respecto al plasma.
Las sales inorgánicas migran también de acuerdo con los principios arriba mencionados. La ingestión de agua puede tornar el contenido intestinal isotópico con el plasma. En el caso que el contenido intestinal es hipertónico, es posible que un transporte activo de sales hacia la sangre intervenga en su isotonización, así que no siempre es cl paso rápido de agua de la sangre al intestino causante de este proceso. Es posible que las sales sean transportadas activamente hacia la sangre y que el agua las siga pasivamente, de acuerdo con la diferencia de presiones osmóticas. Diversos experimentos han revelado que las sales son efectivamente transportadas en forma activa y selectiva. Así, se ha observado que si se introducen al intestino volúmenes iguales de soluciones isotópicas de NaCl y de Na2SO4, desaparece después de corto tiempo el NaCl y una cantidad considerable de Na
+ de la solución de Na2SO4, (al mismo tiempo aumenta, lógicamente, la
concentración de SO4-) lo que puede ser explicado por el transporta activo y selectivo del Na
+ y del
Cl-. El transporte de estos iones se efectúa en contra de las gradientes de concentración
respectivas. Es probable que, junto con estos iones, pase un volumen de agua suficiente para mantener la isoosmolaridad de las soluciones (respecto al plasma). El transporte del ion SO4
- debe
realizarse por un mecanismo diferente, ya que la intoxicación de la mucosa intestinal con inhibidores metabólicos hace que el transporte de Na
+ , C1
- y SO4
- se torne igual. Es importante
destacar que el agua y los iones no sólo son absorbidos desde el lumen intestinal hacia la sangre, sino que existe una migración simultánea en dirección opuesta. El resultado neto de este transporte, desde y hacia el lumen intestinal, depende de numerosos factores. En el intestino delgado los volúmenes intercambiados entre lumen y sangre son iguales, en tanto que en el colon el transporte es preferentemente hacia la sangre. En general la velocidad del transporte de agua y de iones del intestino hacia la sangre se incrementa en dirección caudal.
Como se ha dicho anteriormente, la velocidad de absorción en el intestino es selectiva y la comprensión de su mecanismo requiere el conocimiento de los productos que resultan de la degradación de los alimentos ingeridos y de la cinética de los procesos que facilitan su paso hacia la sangre.
En páginas anteriores hemos analizado el resultado de la acción de los enzimos digestivos sobre los alimentos, el que puede resumirse como sigue: a) los hidratos de carbono son ingeridos como polisacáridos (glucógeno y/o almidón). Las amilasas salival y pancreática los desdoblan en maltosa y en cantidades reducidas de isomaltosa y glucosa. La maltosa a su vez es desdoblada bajo la acción de maltasa a glucosa; b) La degradación de las proteínas se inicia en el estómago. Por acción de la pepsina son hidrolizadas hasta proteasas y peptonas de cadena larga. Estas, bajo el efecto de la tripsina y quimiotripsina, son degradadas hasta productos de cadena progresivamente más corta, convirtiéndose finalmente en tripéptidos y dipéptidos hidrosolubles y aminoácidos; c) La digestión de las grasas se realiza también en etapas hasta llegar progresivamente a compuestos que por ser liposolubles, pasan fácilmente por la membrana celular.

Los productos finales de la degradación intestinal son los monosacáridos, los di y tripéptidos, aminoácidos, ácidos grasos, glicéridos. Junto con éstos se encuentra en el contenido intestinal: agua, electrólitos (Na
+, CI
-, HCO3
-), vitaminas hidrosolubles (complejo B y C) y liposolubles (D, E, K
y A).
Las sustancias hidrosolubles, mientras su tamaño molecular no sea mayor que el diámetro de los poros, pasarán sin dificultades por los supuestos poros de la membrana basal y de la pared capilar. Aunque, como ya se ha explicado, la existencia de estos poros no ha sido comprobada por el microscopio electrónico, su existencia parece muy probable, dado que el paso de las sustancias hidrosolubles depende de su tamaño molecular y en consideración a que moléculas de cargas eléctricas iguales a la de los poros, no los pueden atravesar.
Se entiende por insorpción el paso de sustancias liposolubles del lumen intestinal a través de la célula al espacio intersticial. Por exsorpción el paso de sustancias desde el intersticio a través de la célula, al lumen intestinal. La absorción es la diferencia entre el volumen insorbido y exsorbido que, salvo en su parte inicial del intestino, es positiva. En la parte alta del intestino el volumen de agua que pasa del espacio intersticial a través de la célula hacia el intestino, es mayor que la cantidad insorbida.
La absorción en el tubo digestivo es el resultado de la acción de diversos mecanismos que facilitan el paso de sustancias lipo e hidrosolubles a través de las células de la mucosa gastrointestinal. Estos mecanismos son, en realidad, comunes a las células en general. Sin su intervención sólo las sustancias liposolubles podrían llegar al interior de las células. Debido a la estructura fosfolipídica de la membrana celular las sustancias insolubles en grasa no podrían atravesar la membrana celular.
Los mecanismos de transporte a través de la membrana celular pueden resumirse, en general, en la forma siguiente:
1. Arrastre por el solvente: los electrólitos y sustancias de bajo peso molecular son arrastrados por
el agua que pasa libremente por los poros.
2. Simple difusión que consiste en el paso de sustancias hidrosolubles (monosacáridos,
aminoácidos) por los poros, siempre que su concentración extracelular sea superior a la intracelular. La diferencia de concentraciones determinará la dirección y velocidad del flujo hacia y desde la célula. La velocidad de difusión dependerá además de las propiedades de la superficie y de la constante de difusión. Esta última depende del tamaño molecular de la sustancia y principalmente de la diferencia de concentraciones que es la fuerza motriz en este proceso. Interviene además la carga eléctrica de la sustancia por transportar. Se ha atribuido anteriormente a la simple difusión cierta importancia en la absorción intestinal, pero se acepta, hoy por hoy, que no interviene en este proceso.
3. El tercer mecanismo es la difusión facilitada, que juega un papel preponderante en la absorción intestinal. Consiste en la conversión transitoria de las sustancias hidrosolubles en liposolubles y hace uso de los llamados transportadores o carriers. Primeramente la sustancia hidrosoluble se une, en forma reversible, con su transportador, haciéndose liposoluble y atraviesa como tal la membrana celular. E1 transportador se libera luego de la sustancia transportada y vuelve a la superficie externa de la membrana donde queda disponible para nuevos transportes.
La difusión facilitada no consume energía ni requiere enzimos p la fuerza que determina la dirección del transporte es, como en las modalidades anteriores, la gradiente de concentraciones. Cuando la cantidad de sustancia a transportar es excesiva, los transportadores se saturan y el volumen transportado se mantiene constante a pesar de que la concentración de la sustancia

continúe aumentando. La velocidad de transporte es, por lo tanto, limitada, existiendo una velocidad máxima (V máx.).
La existencia de los transportadores ha sido postulada sobre la base de ciertas características de la transferencia de sustancias a través de las membranas biológicas. Es por lo tanto, hasta ahora, sólo una hipótesis de trabajo, pero ha sido de gran utilidad para el estudio de las funciones de la membrana celular.
Los transportadores son específicos, es decir, tiento preferencia por determinadas sustancias. Si esta preferencia es por dos o más sustratos, se produce competencia y el transportador se satura con la sustancia de mayor afinidad, lo que inhibe el transporte de las otras. Tal es el caso de la floridzina, que al ocupar los transportadores de la glucosa y galactosa, impide el paso de estas sustancias a la célula. La difusión facilitada requiere para su funcionamiento la presencia de ciertos iones, especialmente Na
+ y K
+. El transporte mediante este mecanismo es unidireccional, sólo
hacia el interior de la célula.
PROTEÍNAS
Los dipéptidos y aminoácidos resultantes de la digestión de las proteínas ingeridas, son absorbidos como tales. Después de la ingestión de proteínas, el nivel de los aminoácidos en la vena porta se eleva. Proteínas sólo parcialmente degradadas no pasan a la circulación. Su presencia en la sangre induciría la producción de anticuerpos y la aparición de fenómenos alérgicos que pueden producir trastornos graves. E1 mecanismo utilizado por los aminoácidos para atravesar la membrana celular no es bien conocido. En el feto y en el recién nacido la pinocitosis parece ser la modalidad más frecuente. En el adulto, sin embargo, este mecanismo es insuficiente para transportar la cantidad necesaria de aminoácidos y satisfacer los requerimientos.
Parece muy probable que la absorción de los aminoácidos se efectúe en el adulto por un mecanismo de transporta activo. A favor de esta hipótesis hablan los siguientes hechos:
1. Los aminoácidos requeridos en el metabolismo celular aparecen en la sangre antes que otros metabolitos del mismo tamaño molecular.
2. Su absorción intestinal depende sólo hasta cierto límite de la cantidad presente en el lumen intestinal.
3. El monoiodoacetato, que es un inhibidor metabólico, suprime el paso de los aminoácidos del lumen intestinal a la célula.
4. De una mezcla de aminoácidos levo y dextrógiros, los dextrógiros se absorben primero, lo que parece indicar la existencia de transportadores con capacidad selectiva.
GRASAS
Las grasas neutras ingeridas con los alimentos y absorbidas en el intestino representan un material indispensable para el funcionamiento normal del organismo. Además de formar parte de la

estructura de la membrana celular, de la envoltura de las fibras nerviosas y de diferentes estructuras intracelulares, aportan energía para el metabolismo celular y constituyen una importante reserva energética. Su absorción está dificultada por su insolubilidad en agua. En el tubo digestivo las grasas neutras se emulsionan, o sea, las gotas de grasa se fraccionan en gotas muy finas. Esta emulsificación se inicia, en pequeña escala, ya en el estómago y se completa en el duodeno bajo el efecto de los ácidos biliares. A nivel duodenal, las grasas neutras previamente emulsionadas son hidrolizadas por las lipasas pancreática e intestinal hasta di y monoglicéridos y ácidos grasos libres.
Se puede observar que cuando se ingieren cantidades elevadas de grasa, después de cierto tiempo la linfa del conducto torácico y el plasma sanguíneo se tornan opalescentes, debido a su contenido de finas gotitas de grasas neutras.
Estos hechos permiten afirmar que la mayor parte de las grasas, a diferencia de los hidratos de carbono y proteínas, no son absorbidas hacia los capilares sanguíneos, sino hacia los linfáticos. La presencia de grasas neutras en los vasos linfáticos, se debe a que las células del epitelio de las vellosidades intestinales resintetizan las grasas neutras a partir de monoglicéridos y ácidos grasos absorbidos desde el lumen intestinal. Una explicación alternativa sería que la resíntesis se produjera en el lumen intestinal mismo. Por otra parte, no se puede excluir la posibilidad de que una parte de las grasas neutras sea absorbida sin previo desdoblamiento. Parece probable que todos estos mecanismos intervengan, sin poder precisar la importancia relativa de cada uno.
Se atribuía antaño a las sales biliares y al bicarbonato del jugo pancreático un papel de gran significación en la absorción de las grasas. Se suponía que las sales biliares, provenientes del hígado, al llegar junto con la bilis al intestino delgado, formarían con las grasas una emulsión micelar, convirtiéndolas en hidrosolubles. Estas micelas ocultarían en su interior los mono y diglicéridos y las grasas emulsionadas y las harían pasar, junto con las vitaminas liposolubles, a través de la membrana celular.
Una parte de las grasas neutras, como asimismo los productos de su digestión (mono y diglicéridos y ácidos grasos libres), se encuentran en el intestino no sólo en forma de emulsión, sino también en estado de micelas. La relación cuantitativa entre ambas es siempre a favor de las micelas. Las sales biliares disminuyen la tensión superficial de las gotitas de grasa conjuntamente con la lipolisis efectuada por la lipasa pancreática, intensificando la emulsión. Las gotitas de la emulsión y las micelas se ponen en contacto con las vellosidades de la parte alta del intestino delgado y son absorbidas en presencia de sales biliares, de bicarbonato y de lipasa.
El esquema recién expuesto ha debido modificarse a la luz de evidencia experimental reciente. En efecto, se ha encontrado, por ejemplo, que cantidades elevadas de grasas pueden ser absorbidas en ausencia de lipasa y de sales biliares, lo que muestra la insuficiencia de nuestros conocimientos acerca de los mecanismos que estas sustancias utilizan para pasar del lumen intestinal a la circulación.
En cuanto a la pinocitosis, la enorme velocidad con que las grasas aparecen en la circulación parece indicar que tiene sólo una importancia secundaria.
Cabe señalar todavía que ciertos resultados experimentales parecen indicar la existencia de mecanismos de transporte activo de las grasas. El elevado número de hipótesis acerca del paso de las grasas del intestino a la circulación, indica claramente que nuevos experimentos son necesarios para resolver el problema.
HIGADO

El hígado es considerado con razón como el laboratorio de nuestro organismo y como tal el estudio de sus funciones pertenece más bien a la bioquímica que a la fisiología. Por consiguiente, expondremos sólo un breve resumen de sus funciones. Para mayores detalles, es recomendable recurrir a textos de bioquímica.
La irrigación del hígado, que es en realidad una glándula, está a cargo de dos sistemas íntimamente relacionados con las funciones del órgano. La arteria hepática representa uno de estos sistemas, mientras el otro está a cargo de la vena porta. Esta última colecta la sangre venosa del tubo digestivo y transporta al hígado las sustancias absorbidas en el intestino. Tanto la arteria hepática como la vena porta se ramifican dentro del hígado, formando las arterias y venas perilobulares respectivamente. Estas, a su vez, desembocan en vasos, llamados senos, entre las columnas de células hepáticas. Los senos contienen, por consiguiente, tanto sangre arterial como venosa. Entre las células hepáticas y los capilares hay un espacio, el espacio de Disse, que en condiciones fisiológicas es virtual. Sólo si cl volumen del plasma filtrado desde los capilares es abundante, llega a formar canalículos que conducen el filtrado a los vasos linfáticos. La sangre que circula por los senos pasa a las venas interlobulillares que desembocan en la vena central del lobulillo. Esta vacía su contenido en la vena sublobular. Las venas sublobulares desembocan en la vena cava inferior (Fig. 76).
En la parte central del lóbulo hepático se originan finos tubos capilares que forman el sistema conductor de la bilis. Estos tubos carecen de pared propia y su función consiste en llevar la bilis del centro hacia la periferia del lóbulo. Confluyen progresivamente para formar canalículos de diámetro cada vez mayor, que desembocan finalmente en el docto hepático. A través de este último la bilis pasa del hígado a la vesícula biliar y al duodeno.
VP = Vena Porta A = Arteria hepática VC = Vena Central Los vasos que se originan de VP y de A se unen en la periferia del lobulillo, formado entre las columnas celulares (doble línea) los senos, que se unen en la vena central (VC). Los capilares biliares (línea negra gruesa) se forman al otro extremo de la columna celular y conducen la bilis hacia la periferia del tobulillo.
La abundante irrigación del hígado está en relación con sus múltiples funciones. El 30% del volumen-minuto del corazón (aproximadamente 1.500 ml), pasa por el hígado. Del consumo de O2 total del organismo el 20% es utilizado por el hígado.
Las sustancias absorbidas en el intestino, a excepción de las absorbidas en el intestino grueso, pasan forzosamente por el hígado (vena porta), antes de pasar a la circulación general. El hígado, al retener algunas de estas sustancias y permitir el paso de otras, funciona como un filtro. Produce

además una serie de compuestos de vital importancia que son parcialmente entregados a la circulación o bien almacenados en el órgano mismo. El hígado es además el órgano de excreción de tos ácidos y sales biliares (cuya importancia para la absorción de las grasas .ya ha sido mencionada) y de otros componentes de la bilis que llegan con ésta al intestino. Es importante mencionar finalmente las células de Küpfer , ubicadas en los senos de los lobulillos, que forman parte del sistema retículoendotelial.
Las principales funciones hepáticas pueden describirse someramente como sigue:
1. Función secretora, que resulta en la producción de la bilis. Esta es primordialmente una función del sistema retículo-endotelial hepático (células de Küpfer). Consiste, por una parte, en la conjugación y excreción de la bilirrubina y de sus productos de degradación (ver Metabolismo de la Hemoglobina) que llegan al intestino con la bilis. Por otra parte, en la excreción de los componentes de degradación del colesterol y de algunas hormonas esferoidales. Finalmente, ciertas sustancias que circulan en la sangre unidas a proteínas, al llegar al hígado, son separadas del componente proteico.
Hay que mencionar todavía que algunas vitaminas son degradadas y excretadas por el hígado.
2. Función metabólica, tal vez la más importante, mediante la cual, el hígado participa en el metabolismo intermedio de casi todas las sustancias. Entre los compuestos que san absorbidos desde el intestino hacia la sangre, hay algunos todavía relativamente complejos (dipéptidos, disacáridos, monoglicéridos, grasas neutras). En el hígado se completa su degradación que termina con la formación de aminoácidos, monosacáridos, ácidos grasos y glicerina. Estos productos son finalmente utilizados como materiales energéticos en el metabolismo celular o empleados en la síntesis de diversas sustancias (hormonas, proteínas protoplasmáticas, glicógeno, grasas, y otros).
Es fácil comprender que a través de sus múltiples funciones el hígado participa en forma particularmente importante en la mantención de la constancia de nuestro medio interno. Es así como, por ejemplo, la participación hepática en el metabolismo de los hidratos de carbono condiciona en gran parte la constancia del nivel glucémico a pesar de la ingestión discontinua de cantidades variables de estas sustancias. Tanto la disminución (hipoglicemia) como la elevación (hiperglicemia) del nivel glucémico, ponen en marcha un conjunto de mecanismos que tratan de retornar la glicemia a su valor normal. La constancia del nivel glucémico es de vital importancia. Si bien su elevación transitoria no causa daño de consideración, su disminución, aún por corto tiempo, produce convulsiones intensas y, si es de larga duración, puede provocar la muerte (coma hipoglucémico).
El hígado participa en varias fases del metabolismo de los hidratos de carbono. Representa, desde luego, el lugar de su almacenamiento. La glucosa no utilizada en un momento dado, es polimerizada en el hígado hasta glucógeno, que es almacenado. A medida que el metabolismo celular lo requiere, el glucógeno hepático es despolimerizado hasta glucosa (proceso denominado glucogenolisis) y como tal pasa a la sangre. .
Todas las células contienen en realidad glucógeno, pero en concentración muy inferior a la de las células hepáticas. Así por ejemplo, las células musculares contienen, aunque en concentración menor que el hígado, una cantidad total mayor de glucógeno, debido a que la masa muscular es muy superior a la del hígado (100 a 120 g en el hígado, frente a 300 a 400 g en los músculos). El músculo es también capaz de desdoblar el glucógeno en glucosa, compuesto indispensable para su metabolismo energético. Pero existe una diferencia fundamental en este respecto entre hígado y músculo: el hígado posee la enzima glucosa-6-fosfatasa, capaz de convertir el primer producto de la glucogenolisis, la glucosa-6-fosfato, en glucosa que pasa fácilmente de la célula hepática a la sangre. El músculo carece de esta enzima y, por lo tanto, es incapaz de convertir la glucosa-6-fosfato en glucosa. La glucosa-6-fosfato en las células musculares es utilizada parcialmente en

forma anaeróbica como material energético, desdoblándose hasta ácido láctico. Este pasa libremente a la sangre y al llegar al hígado se convierte en glucosa.
El hígado es, por lo tanto, capaz tanto de sintetizar y almacenar glucógeno a partir de la glucosa absorbida en el intestino, como de
despolimerizar el glucógeno hasta glucosa. Esta capacidad homeostática es precisamente la que confiere al hígado importancia en la conservación de la constancia del nivel glucémico. En esta forma y, a pesar del consumo continuo y variable de glucosa, la glucemia se mantiene dentro de límites estrechos (80-120 Mg. % ).
La figura 77 muestra esquemáticamente la participación del hígado en la conservación de la constancia del nivel glucémico al suministras glucosa a un individuo por vía endovenosa.
El hígado juega también un papel importante en el metabolismo de las proteínas y de las grasas. Como hemos dicho, la mayor parte de las grasas ingeridas y absorbidas llega a la sangre por el conducto torácico, o sea, por vía linfática y sólo una pequeña parte pasa por la circulación portal al hígado. Pero las célula hepáticas son capaces de sintetizar grasas a partir tanto de proteínas como de hidratos de carbono y entregarlas a la sangre, en la cual circulan, unidas a proteínas, como lipoproteínas.
La sangre contiene grasas en las siguiente formas: a) como grasa neutra ( 400 a 800 Mg. %); b) como colesterol (180 a 240 Mg. %), que circula en la sangre en forma libre o formando un complejo con ácidos grasos; c) como fosfolípidos, predominantemente como lecitina (230 Mg. %). Los niveles de estos compuestos son constante debido a la capacidad de las células hepáticas de entregar y almacenar grasas. El hígado puedo transformar un ácido graso en otro, capacidad que en ciertas enfermedades, como la diabetes, adquiere gran importancia. Los productos intermedios de esta transformación, en condiciones fisiológicas no pasan a la sangre, pero en los diabéticos, circulan en la sangre y son utilizados como combustible en el metabolismo energético. En esta enfermedad, debido al déficit de insulina (ver Sistema Endocrino), la glucosa no pasa al interior de las células y disminuye, por lo tanto, su concentración intracelular. A causa de esto, incrementa la combustión de grasas. Los productos del metabolismo intermediario de las grasas, son ácidos que en cantidad elevada producen una disminución del pH sanguíneo, denominada acidosis metabólica.
Las proteínas ingeridas, degradadas y absorbidas en el intestino, llegan por la vena porta al hígado y pasan de aquí a la circulación general. Alrededor del 80% de los aminoácidos absorbidos son

retenidos en el hígado. El 2o% restante llega con la sangre de la vena cava a la circulación general, y son utilizados en la resíntesis de proteínas tisulares. Los aminoácidos retenidos son desaminados o transaminados. Los ácidos desaminados son utilizados directamente como combustible o bien convertidos en glucosa que es polimerizada luego hasta glucógeno 0 grasa, que son almacenados en las células hepáticas. La transaminación resulta en la síntesis de aminoácidos de diferente tipo.
Es importante mencionar la capacidad desintoxicante del hígado, mediante la cual ciertas sustancias tóxicas, absorbidas en el intestino junto con los alimentos, son conjugados con ácido gluconúrico y transformadas así en compuestos inocuos que son excretados con la bilis al intestino.
Una función importante del hígado es la inactivación de algunas hormonas, especialmente de aquellas de carácter esferoidal.
El hígado produce algunas sustancias indispensables para la coagulación de la sangre: fibrinógeno, protrombina, vitamina K.
Las células hepáticas participan también en el almacenamiento de la vitamina A. Su función de depósito no se limita a ésta y al glucógeno y otros compuestos mencionados anteriormente, sino que se extiende al almacenamiento de diversos metales (cobre, cobalto, manganeso, etc.). El papel del hígado en el metabolismo del fierro de la hemoglobina fue ya tratado.
VESICULA BILIAR
La secreción de la bilis por el hígado es continua, pero su intensidad varía de acuerdo con las diferentes fases de la digestión. La vesícula biliar es un receptáculo en que se almacena la bilis. Debido a la acción del esfínter, ubicado a nivel de la desembocadura del colédoco en el duodeno (esfínter de Oddi), la bilis no puede fluir al duodeno en forma continua. Al llegar el alimento al duodeno se relajan, por vía refleja, tanto el esfínter de Oddi como el del conducto cístico, y la bilis, proveniente tanto del hígado como de la vesícula, fluye hacia el duodeno.
La concentración de la bilis en la vesícula es 5 a 6 veces superior a la de la bilis recién secretada por el hígado. El aumento de la concentración se debe a la absorción de sales y agua por la mucosa vesicular que secreta además un líquido mucoso que altera la coloración de la bilis tornándola oscura y viscosa.
El alto contenido de Na+ y bicarbonato de la bilis contribuye eficazmente a la neutralización del
contenido gástrico ácido que llega al duodeno. La rapidez con que se realiza esta neutralización impide que la pepsina mediante su efecto digestivo destruya las enzimas pancreáticas de carácter proteico. La presencia de bilis en el intestino adquiere especial importancia cuando el alimento ingerido es rico en grasas, debido a la intervención de los ácidos y sales biliares en su digestión y absorción.
Fig. 78 Regulación del vaciamiento de la vesícula

Durante el ayuno la vesícula biliar está relajada y el esfínter de Oddi aumenta su contracción tónica. Como la secreción de la bilis es continua, durante el ayuno se almacena en la vesícula. La ingestión de alimentos es el estímulo que produce la contracción de la musculatura lisa vesicular, la relajación del esfínter de Oddi y el paso de la bilis al duodeno. Este ciclo de contracción vesicular y relajación esfinteriana es periódico, se produce cada 30 minutos aproximadamente y causa así un flujo intermitente de bilis al duodeno. Las contracciones vesiculares son de poca intensidad y tienen poca importancia. El vaciamiento intermitente está condicionado primordialmente por la abertura y cierre periódico del esfínter y por las contracciones rítmicas del duodeno que varían la presión intraduodenal. Si esta se hace superior a la presión intracoledociana, la bilis deja de fluir al duodeno aun cuando el esfínter de Oddi esté relajado.
Dos mecanismos regulan las contracciones de la vesícula, uno humoral y otro nervioso reflejo. Con la llegada de ciertos alimentos (grasas, huevos y otros) al duodeno, se inician las contracciones vesiculares.
A causa de que la presencia de alimentos en el duodeno estimula también la secreción de jugo pancreático, éste aparece en el duodeno simultáneamente con la bilis.
La musculatura lisa de la vesícula y la del esfínter de Oddi está inervada tanto por el vago como por el simpático. El vago contrae la vesícula y al mismo tiempo causa la relajación del esfínter, contribuyendo al vaciamiento vesicular Los efectos del simpático son opuestos y, por lo tanto, su estimulación impide, o por lo menos retarda, la llegada de bilis al intestino. La distensión del estómago y del duodeno activa un reflejo, cuya vía eferente está constituida por fibras vagales y que causa el vaciamiento vesicular. Sin embargo, la vagotomía bilateral no altera significativamente el vaciamiento. Se ha demostrado que éste es controlado por un mecanismo humoral además del mecanismo nervioso anteriormente descrito.
La regulación humoral se inicia en el duodeno, cuya mucosa, bajo la acción de los alimentos (especialmente grasas neutras y los productos de su digestión), secreta una hormona, pancreozimina (denominada antiguamente colecistoquinina), de efecto similar al obtenido por la estimulación del vago. La regulación humoral del vaciamiento supera en importancia a los mecanismos nerviosos. Se acepta en la actualidad que la contracción vesicular es inicialmente refleja y que sólo en una segunda fase interviene el mecanismo humoral.
El control del flujo de bilis y del vaciamiento de la vesícula está representado esquemáticamente en la figura 78.

El tono de la musculatura lisa de la vesícula depende predominantemente del vago y en menor grado del simpático. Las flechas del lado derecho indican las influencias que el estómago ejerce sobre la vesícula (reflejo gastrovesicular).
La figura 74, muestra esquemáticamente que en el caso de la glucosa, por ejemplo, sólo en presencia de Na
+ es el transportador capaz de hacerla pasar hacia el interior de la célula. Cuando
el lugar del Na+ está ocupado por el K
+, ion predominantemente intracelular, el transporte de
glucosa se detiene. Una vez dentro de la célula, la glucosa no puede salir por el mismo mecanismo.
4. Un cuarto mecanismo es el transporte activo, por el cual ciertas sustancias atraviesan la membrana celular en contra de la gradiente de concentración. Así por ejemplo, los monosacáridos y los aminoácidos atraviesan la membrana celular por difusión facilitada, sólo si su concentración extracelular es superior a la intracelular. Pero pueden pasar también al interior de la célula en contra de la gradiente de concentración, es decir, del lugar de su menor al de su mayor concentración. Se realiza este proceso mediana el transporte activo, que significa trabajo y, por lo tanto, consume energía. La energía es utilizada para la activación de una bomba biológica no específica, o sea, común para varias sustancias. El Na
+ se transporta también mediante un
mecanismo de este tipo (bomba de sodio).
Para la comprensión de este mecanismo es indispensable referirse a ciertas particularidades de la membrana celular, especialmente a las características de una membrana polar. Se entiende como membrana polar una capa delimitadora que posee bomba de Na
+, en general en cada punto de su
circunferencia, bomba que es utilizada para transportar este ion desde el interior de la célula hacia el espacio extracelular. La salida del Na
+ , de carga eléctrica positiva, produce un aumento de las
cargas eléctricas positivas extracelulares. Al mismo tiempo se incrementa la concentración relativa de las cargas negativas intracelulares. Se crea, en esta forma, una diferencia de potencial eléctrico

entre el interior y el exterior de la célula. La mantención de esta diferencia de potencial es de vital importancia.
Las células de la mucosa intestinal no están delimitadas por una membrana polar, es decir, no poseen »bomba de sodio« en toda su circunferencia. Son, por consiguiente, células asimétricas, en cuanto la bomba de sodio existe sólo en sus caras laterales y en la parte cercana a la serosa, careciendo de este mecanismo en la cara que mira hacia el lumen intestinal. Esto determina que el Na
+ puede salir de la célula sólo por las partes laterales y cercanas a la serosa. La distribución
intracelular de Na+ es en consecuencia desigual; su concentración es mayor en la parte mucosa
que en la parte serosa de la célula. De esta distribución resulta una diferencia de potencial entre ambos polos celulares, siendo la parte vecina a la mucosa positiva respecto a la serosa.
Como puede observarse en la Fig. 74 el Na+ pasa del lumen intestinal a las células que forman la
mucosa, por diferentes mecanismos: .difusión facilitada (acoplamiento a los transportadores de glucosa) y arrastre por el solvente. El Na
+ intracelular es eliminado hacia el espacio intersticial
mediante la bomba de sodio. Una vez entregada su carga (glucosa, aminoácidos, sales biliares, sodio), el transportador vuelve al lado mucoso de la membrana para iniciar un nuevo ciclo de transporte. El Na
+ no se acumula dentro de la célula, porque es eliminado hacia la serosa. No así la
glucosa y aminoácidos cuya concentración intracelular se eleva (atrayendo H2O y K+) y que pasan
luego por mecanismos de transporte activo y difusión facilitada (utilizando en este último caso los mismos transportadores que utiliza el Na+), de la célula de la mucosa al líquido intersticial y a la sangre capilar. En esta forma, la bomba de sodio interviene no sólo en el transporte de Na
+, sino
también en el de la glucosa, de los aminoácidos y otras sustancias.
Por las razones expuestas, en ausencia de Na en los espacios intercelulares, el transporte de glucosa es bloqueado. En consecuencia, aumenta la concentración de glucosa en estos espacios, lo que justifica la suposición que la bomba de sodio está ubicada en esta parte de la membrana celular. La concentración elevada de Na
+ en los espacios intercelulares atrae agua, lo que produce
su ensanchamiento durante la absorción, fenómeno que puede ser observado mediante el microscopio electrónico. El agua pasa del espacio intercelular a la célula, isotonizando el líquido intracelular. El contenido intestinal es hipertónico respecto al interior de las células de la mucosa. Por lo tanto, el agua, al ser absorbida, pasa del medio en que se encuentra en menor concentración (lumen intestinal) al medio intracelular, en el cual su concentración es mayor. Se trata, por consiguiente, de un transporte activo. Sin embargo, este transporte activo es más bien aparente, ya que en realidad el agua penetra a la célula acoplada al Na
+. En otras palabras, el Na
+
arrastra agua. El agua pasa por los poros de la membrana celular libremente, sin encontrar resistencia alguna. El Na
+, por su parte, no puede ser arrastrado por el agua debido a que su
tamaño molecular sobrepasa el diámetro de los poros.
En el caso de la absorción intestinal los mecanismos de transporte pueden ser resumidos en la forma siguiente:
1. Difusión facilitada que, como hemos visto, no consume energía, pero es Na+ dependiente y se efectúa mediante transportadores. Estos son saturables y pueden ser bloqueados por ciertas sustancias (floridzina). Son además específicos, es decir, cada sustancia utiliza un transportador específico. Esta modalidad de transporte es considerada como la de mayor importancia en el tubo digestivo.
2. Transporte activo, en el cual se utiliza la bomba biológica. Esta modalidad consume energía y no
es específica. Ciertos venenos que inhiben el metabolismo celular, como asimismo la falta de O2, bloquean este mecanismo de transporte.
3. Difusión simple, utilizada predominantemente por el agua y por algunos solutos. No interviene mayormente en la absorción intestinal.

4. Pinocitosis, en la cual la membrana celular introduce al interior de la célula ciertas sustancias, especialmente grasa, por un proceso de inclusión: la membrana celular forma una vesícula que rodea la gota de grasa , la vesícula se abre luego hacia el interior de la célula y la gota de grasa queda así incluída en el protoplasma celular (fig. 75)
HIDRATOS DE CARBONO
Los polisacáridos ingeridos deben ser primeramente degradados a monosacáridos ya que sólo así pueden ser absorbidos. La absorción de los monosacáridos en la unidad de tiempo es independiente de su peso molecular, pero es selectiva. Si se bloquea, mediante algún tóxico, la actividad de la mucosa intestinal, la velocidad de absorción de la glucosa y de la galactosa disminuye, pero no se modifica la de otros monosacáridos. Se ha podido demostrar que esto se debe a que la absorción de la glucosa y de la galactosa se efectúa por un transporte activo, en tanto que la de otros monosacáridos es pasiva. La glucosa es absorbida en contra de una gradiente de concentración, no así los, otros monosacáridos que pasan del lugar de mayor al de menor concentración. Ambas modalidades de transporte son saturables, es decir, la cantidad de monosacáridos absorbidos por cualquiera de los dos mecanismos depende sólo hasta cierto límite de su cantidad en el lumen intestinal. La absorción de glucosa en el intestino es limitada, limitación que aumenta por la competencia entre los diferentes monosacáridos por el transportador, lo que habla a favor del transporte activo. La glucosa, por ejemplo, al competir con la galactosa inhibe su absorción y viceversa.
La absorción se realiza en la parte apical de las vellosidades, en el ribete estriado y en menor grado en su tercio superior, disminuyendo gradualmente hacia su parte banal.
Los monosacáridos absorbidos pasan como glucosa o pentosa a la sangre y alcanzan, por la circulación portal, al hígado. La velocidad de absorción de los monosacáridos es variable, aumentando en el siguiente orden: arabinosa, xilosa, manosa, fructuosa, glucosa. La ingestión repetida de un monosacárido aumenta la velocidad de su absorción, probablemente por una activación progresiva de sus transportadores. La presencia de sodio durante el transporte de los monosacáridos es indispensable ya que su reemplazo por potasio o por otros iones, bloquea totalmente este proceso. Esta modalidad de transporte existe sólo en el intestino. Así por ejemplo, al suministrar una solución de monosacáridos por vía peritoneal, la absorción de los diferentes monosacáridos se efectúa con la misma velocidad, aun en ausencia de sodio, lo que indica que se efectúa por un mecanismo diferente al de la absorción intestinal.
En resumen, los hidratos de carbono ingeridos son hidrolizados en el intestino hasta monosacáridos y pasan como tales a través de la membrana al interior de las células de las vellosidades. El paso se efectúa por un mecanismo activo en que intervienen transportadores que requieren la presencia del ion Na
+. El monosacárido y el Na
+ transportados en forma activa,
atraviesan la membrana celular y penetran por el citoplasma hacia la cara serosa de la membrana, en la cual está ubicada la bomba de Na
+. Esta bomba, al eliminar Na
+, disminuye su concentración
intracelular. La concentración intracelular de K+ se mantiene elevada. Como el transportador, sin

Na+, es incapaz de retener el monosacárido, éste es retenido en la célula y aumenta su
concentración intracelular. El transportador se carga ahora con K+ y vuelve hacia la cara mucosa
de la membrana celular, o sea, hacia un medio pobre en K+ pero rico en Na
+. El K
+ se desprende
del transportador, pasa al espacio extracelular y es reemplazado por Na+, que lo capacita para
transportar nuevas cantidades de monosacáridos. En esta forma, los monosacáridos penetran a la célula en contra de la gradiente de concentración.
El paso de monsacáridos a la célula se realiza también por difusión facilitada. Ya hemos visto que este mecanismo si bien es Na
+ dependiente, no consume energía. Los monosacáridos utilizan
transportadores específicos, es decir, cada monosacárido tiene su transportador. Debido a la afinidad diferente de los monosacáridos hacia los transportadores, se establece cierta competencia.