DEGRABADA INMUNO 05
25
Academia Medics Science Página 1 FACULTAD DE MEDICINA HUMANA INMUNOLOGIA BASICA Clase:05 05/09/15 AUTOR: Medics Science COMPLEJO MAYOR DE HISTOCOMPATIBILIDAD (MHC) 1. GENERALIDADES Es complejo porque los receptores de linfocito T, son diseñados para reconocer antígenos que son lineales, presentados por moléculas de superficie de las células hospederas. Esas moléculas de superficie con el MHC tipo 1 o 2. La célula T no puede, o no está diseñada para nunca aceptar o unirse a antigenos tridimensionales, no se ligan directamente a antígenos que estén en estado libre, ellos se unen a antígenos que son procesados, para mantener una “linerealidad” de péptidos. No va a reconocer antigenos libres, deben ser péptidos antigénicos presentados por un MHC. • Los linfocitos T deben interactuar con otras células infectadas del hospedero y no directamente con microbios. • Los receptores de antígeno de los linfocitos T son diseñados para reconocer antígenos presentados por moléculas de superficie de en la célula hospedera y no directamente por antígenos en la superficie de los microbios o en estado libre fuera de la célula
-
Upload
enrique-moncada-mapelli -
Category
Documents
-
view
52 -
download
0
description
Inmunología
Transcript of DEGRABADA INMUNO 05

Academia Medics Science Página 1
FACULTAD DEMEDICINA HUMANA
INMUNOLOGIA BASICA
Clase:0505/09/15
AUTOR: Medics Science
COMPLEJO MAYOR DEHISTOCOMPATIBILIDAD (MHC)
1. GENERALIDADES
Es complejo porque los receptores de linfocito T, son diseñados para reconocer
antígenos que son lineales, presentados por moléculas de superficie de las
células hospederas. Esas moléculas de superficie con el MHC tipo 1 o 2. La
célula T no puede, o no está diseñada para nunca aceptar o unirse a antigenos
tridimensionales, no se ligan directamente a antígenos que estén en estado
libre, ellos se unen a antígenos que son procesados, para mantener una
“linerealidad” de péptidos. No va a reconocer antigenos libres, deben ser
péptidos antigénicos presentados por un MHC.
• Los linfocitos T deben interactuar con otras células infectadas del
hospedero y no directamente con microbios.
• Los receptores de antígeno de los linfocitos T son diseñados para
reconocer antígenos presentados por moléculas de superficie de en la
célula hospedera y no directamente por antígenos en la superficie de los
microbios o en estado libre fuera de la célula

Academia Medics Science Página 2
2. CARACTERISTICAS DE LOS ANTIGENOSRECONOCIDOS POR CELULAS T
Reconocer péptidos, ósea secuencias de aminoácidos que son
depositados en las hendiduras del MHC ya que están ligados a ellos.
No reconoce estructuras tridimensionales, péptidos lineales se unen en
la hendidura del MHC, perdiendo su conformación tridimensional. El
único receptor que reconoce estructuras tridimensionales es el BCR, que
es el receptor del linfocito B.
Linfocitos T reconocen antigenos asociados a células y no antigenos
solubles, los TCR solamente reconocen estructuras semejantes al MHC
e estas son exhibidas asociadas a péptidos en la membrana celular
Las células TCD4 y TCD8 reconocen al antígeno, que es presentado por
un MHC clase II y I, respectivamente. El MHC clase II, es presentado por
un APC, y las MHC clase I, que se expresa en todas las células
nucleadas.
3. MODELO DE RECONOCIMIENTO DEL COMPLEJO MHC-péptido POR EL TCR.
Tenemos una estructura para ver cómo se da esta unión, ustedes pueden
ver un TCR y un MHC, si ustedes se dan cuenta el espacio es muy
pequeño para que entre el péptido antigénico, es un espacio restricto
entonces 3 cosas:
Hay aminoácidos que son diferentes que son aa que estan en
contacto íntimo con el TCR.
Hay aminoácidos que forman una especia de ancla para que el
péptido se localice en el MHC.
Forman una bolsa o saco de MHC que son aa con carga
hidrofóbica, entonces buscan depositarse o que sea más estable en
la ligancion de péptido al MHC.

Academia Medics Science Página 3
4. FUNCIONES DE LAS DIFERENTES APC
Cuáles son las células que presentan estos péptidos lineales, son las
células presentadoras de antígeno (APC), y tenemos 3, célula dendrítica,
Macrófago y la célula o Linfocito B.
Cuando hablo de Celulas Dendriticas, es la mejor APC profesional
porque posee casi todos los toll like receptor (TLR), scavener, de
complemento de fracción constante, de Ig. es capaz de activar
linfocitos T vírgenes. Tiene moléculas estimulatorias. Como la B7 y
CD40, entonces lo que hace es activar linfocitos T que son vírgenes
pero maduros, y este linfocito T producirá citosinas realizando sus
funciones efectoras, como otro tipo células.
Macrofagos y Linfocitos B, estos activan células, solo que los Linf T
son previamente activados.
Tenemos al macrófago que fagocito su antígeno, vino una célula
efectora y se ligó, lo presento por MHC clase II. Las células efectoras
produce citosinas en este caso IFN, para inducir al macrófago que
realice la fagocitosis.
La células B reconoce péptidos, son tridimensionales, son captados y
procesados, y son presentados por MHC clase II, así activar al linfocito

Academia Medics Science Página 4
, este activara a esta célula B para que se convierta en plasmocito
(célula plasmática) y producirá anticuerpos que son importantes en la
inmunidad humoral, y en este caso para la células., en ambos casos
ayuda a captar los antígenos, por fagocitos o neutralización.
5. PROPIEDADES Y FUNCIONES DE LAS APC Celulas Dendriticas, Macrofagos, Células endoteliales con inducidas
por el INF-γ, esta citosina es la única que induce el MHC clase II.
En cambio el IFN alfa y beta, inducen el de MHC clase I, tiene la
propiedad de expresa moléculas coestimulatorias, son las moléculas
que van a inducir la mayor activación celular que tiene receptores
como CD40 con CD40L, también en ambos tiene la activación o
estimulación de proteínas coestimulatorias
En CD, iniciar la activación de los linfocitos T
En macrófagos, funciones efectoras como fagocitosis y
degracdacion de microoganismos
Los linfocitos B, se da una activación completa humoral
Academia Medics Science Página 5
6. VIAS DE ENTRADA DEL ANTIGENO
Donde se localizan las Células dendríticas, debes ver sus vías de entrada,
los antigenos entran por la piel, por los tejidos epiteliales, mucosa intestinal
respiratoria, genital o ganglios linfáticos y en el bazo, entonces todos estos
locales, son donde las dendríticas se localizan, entonces llegan al tejido
epitelial y lo capta una célula dendrítica inmadura, lo procesa, poco a poco
va madurando, sigue procesando el antígeno, y migrara a través de vasos
linfáticos yendo al bazo, donde también hay células dendríticas, pero
también habrá una presentación a los linfocitos T, generando una celula
dendrítica madura.
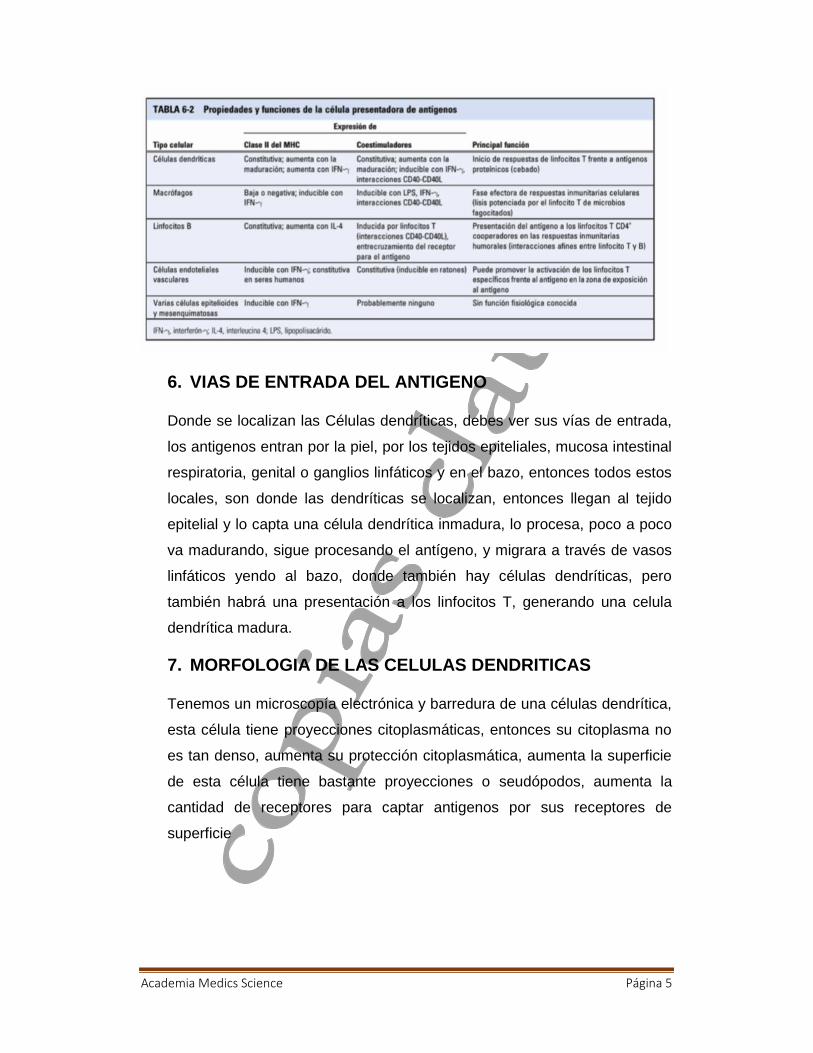
7. MORFOLOGIA DE LAS CELULAS DENDRITICAS
Tenemos un microscopía electrónica y barredura de una células dendrítica,
esta célula tiene proyecciones citoplasmáticas, entonces su citoplasma no
es tan denso, aumenta su protección citoplasmática, aumenta la superficie
de esta célula tiene bastante proyecciones o seudópodos, aumenta la
cantidad de receptores para captar antigenos por sus receptores de
superficie
Academia Medics Science Página 6
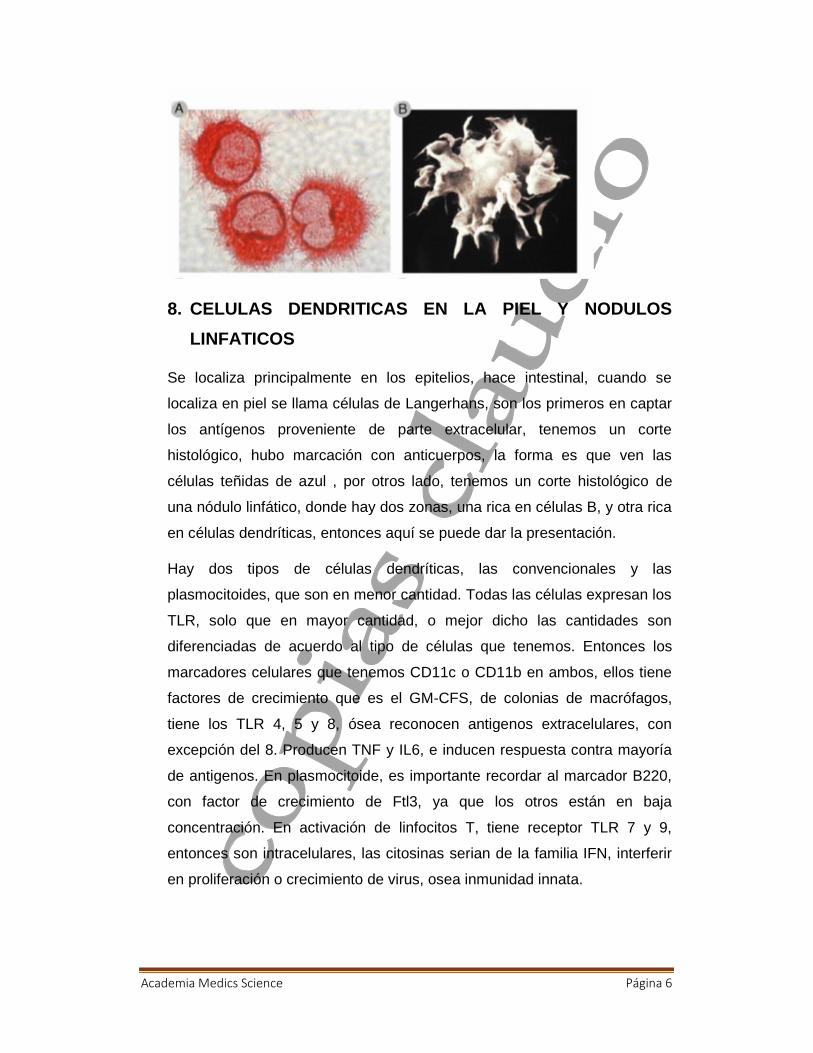
8. CELULAS DENDRITICAS EN LA PIEL Y NODULOSLINFATICOS
Se localiza principalmente en los epitelios, hace intestinal, cuando se
localiza en piel se llama células de Langerhans, son los primeros en captar
los antígenos proveniente de parte extracelular, tenemos un corte
histológico, hubo marcación con anticuerpos, la forma es que ven las
células teñidas de azul , por otros lado, tenemos un corte histológico de
una nódulo linfático, donde hay dos zonas, una rica en células B, y otra rica
en células dendríticas, entonces aquí se puede dar la presentación.
Hay dos tipos de células dendríticas, las convencionales y las
plasmocitoides, que son en menor cantidad. Todas las células expresan los
TLR, solo que en mayor cantidad, o mejor dicho las cantidades son
diferenciadas de acuerdo al tipo de células que tenemos. Entonces los
marcadores celulares que tenemos CD11c o CD11b en ambos, ellos tiene
factores de crecimiento que es el GM-CFS, de colonias de macrófagos,
tiene los TLR 4, 5 y 8, ósea reconocen antigenos extracelulares, con
excepción del 8. Producen TNF y IL6, e inducen respuesta contra mayoría
de antigenos. En plasmocitoide, es importante recordar al marcador B220,
con factor de crecimiento de Ftl3, ya que los otros están en baja
concentración. En activación de linfocitos T, tiene receptor TLR 7 y 9,
entonces son intracelulares, las citosinas serian de la familia IFN, interferir
en proliferación o crecimiento de virus, osea inmunidad innata.

Academia Medics Science Página 7
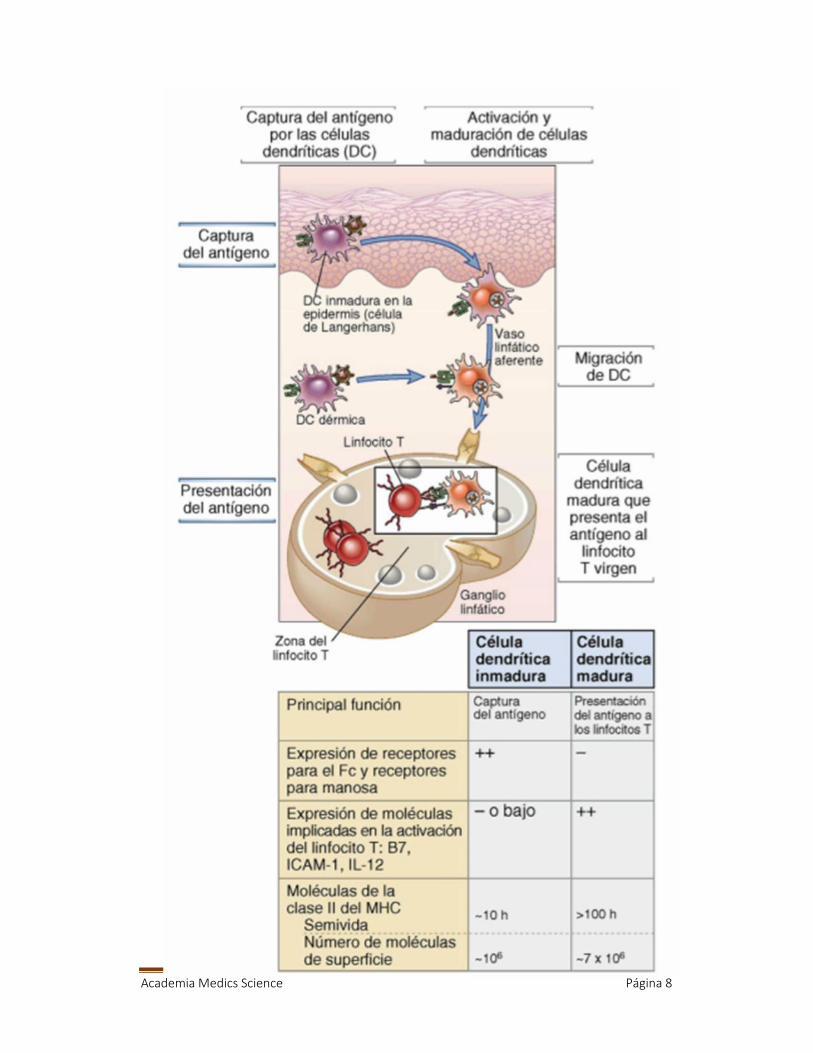
9. CELULAS DENDRITICAS Y LA CAPTURA YPRESENTACION DE ANTIGENOS
Entonces como se realiza esta captura de antígeno? Ustedes tienen un
células del Langerhans inmadura que tiene baja cantidad de MHC, a lo que
se le da un antígeno, viaja vía vaso linfático, el antígeno e procesado poco
a poco hasta llegar a un ganglio linfático, ya está madura, parece más
MHC, y moléculas coestimulatorias como B7 y CD40, y activa de forma
eficiente lo que es el linfocito T, sale a través de vías efectoras y se da la
activación celular
Entonces algunas diferencias entre células dendríticas maduras y no maduras
serian:
La inmadura capta antigenos, la madura los va a presentar a linf T
La inmadura debe tener receptores para capturar antigenos como TLR,
como fracción critalizable de Ig como la G o scavener o del
complemento o receptor de manosa. En maduras ya no es necesaria,
porque deben presentar entonces tienen MHC.
Una células inmadura tiene una vida media es menor encambio madura
no.
La expresión de moléculas como B7, ICAM, IL12 y asi la madura es la
que posee mas de esto. La madura debe tener un MHC II.
Academia Medics Science Página 8

Academia Medics Science Página 9
10. CD SON EFICIENTES INICIANDO LA ACTIVACIONDE LOS LINFOCITOS T, debido a:
• Las CD están localizadas en las regiones de entrada de los microbios, todos
los epitelios de piel intestinos, respiratorio, genital
• CD expresan receptores que los capacitan en la captura de Ag. Scavener
complemento CR3 TLR.
• CD pueden migrar de la zona donde están los Ag hasta la zona donde están
los linfocitos T, los antigenos estan en los ganglios linfaticos y bazo
• CD maduras expresan altas concentraciones de MHC, de proteínas co-
estimuladoras y citocinas importantes en la estimulación de los linfocitos T.
Como B7 y CD40.
11. COMPLEJO MAYOR DE HISTOCOMPATIBILIDAD(MHC)
Entonces que es un complejo mayor de histocompatibilidad? MHC,
complejo cuyo producto final es una proteína, osea una cadena, y tiene una
historia
• En los ratones el MHC esta denominado como complexo H-2
• La región genética responsable por el rechazo de enjertos de tejidos fue
denominada locus mayor de histocompatibilidad. Porque en la
donación había un rechazo codificado por este gen. Por compatibilidad
de tejidos. Los estudios siempre se realizan en ratones primero por ser
mamíferos eucariontes, mamíferos como nosotros.
• En los ratones esta localizado en el cromosoma 17 y codifica un grupo
sanguíneo denominado II, por eso su denominación histocompatibilidad
2 o H-2.
• Al analizar la región H-2 en ratones productos de muchos cruces se
observo que no es un gen, sino un conjunto de genes estrictamente
unidos envueltos en el rechazo de enjertos. Por eso se denomino:
COMPLEJO MAYOR DE HISTOCOMPATIBILIDAD.

Academia Medics Science Página 10
• Hay individuos que reciben un donación, y ellos antes esto producen
unos anticuerpos pero contra proteínas del donante, son las HLA, es la
cosa principal de rechazo de injerto, ya lo verán pronto.
• Se descubrieron al observar que moléculas de superficie de un individuo
son reconocidas como extrañas por otro individuo.
• Individuos que reciben transfusiones sanguíneas producen Anticuerpos
que reconocen las células y proteínas de los donantes, estas proteínas
se denominan Antígenos Leucocitarios Humanos (HLA), siendo estos la
causa principal del rechazo del enjerto.
• Estructural y Funcionalmente H-2 e HLA son idénticos ya que
determinan el futuro de los enjertos, denominándose de MHC.
12. FENOMENO DE RESTRICCION DEL MHC DE LOSLINFOCITOS T
Ustedes tienen un ratón, es de cepa A, hay diferentes linajes, ellos esta
infectada con el virus de la coriomeningitis linfocítica (LCMV), se espera
unos días, se mató al animal, y se sacó a los linf T citotóxicos específicos
para este virus. Por otro lado, yo tengo mi célula dendrítica de mi ratón de
cepa A, y presentara una proteína propia, y péptidos derivado del virus, y
también tengo mi cepa B, que también tiene sus antígenos propios y los
péptidos derivados del virus. Entonces cuando yo los coloco en un cultivo
habrá varias combinaciones, si pongo en un cultivo mi Linfocito citotoxico, y
este recibe o reconoce un péptido por un MHC clase II, e induce muerte de
células infectada.
Si mis células de las células A, no esta infectada no pasa nada porque no
hay reconocimiento, mi linfocito T citotóxico no va a reconocer péptidos
propios porque genéticamente tiene un TCR especifico para el virus
entonces no pasara nada, en otro caso tengo mi célula infectada por virus,
y no es reconocido por cepa A si no por cepa B igual no pasa nada porque
este linfocito T su TCR no esta capacitado para reconocer un antígeno
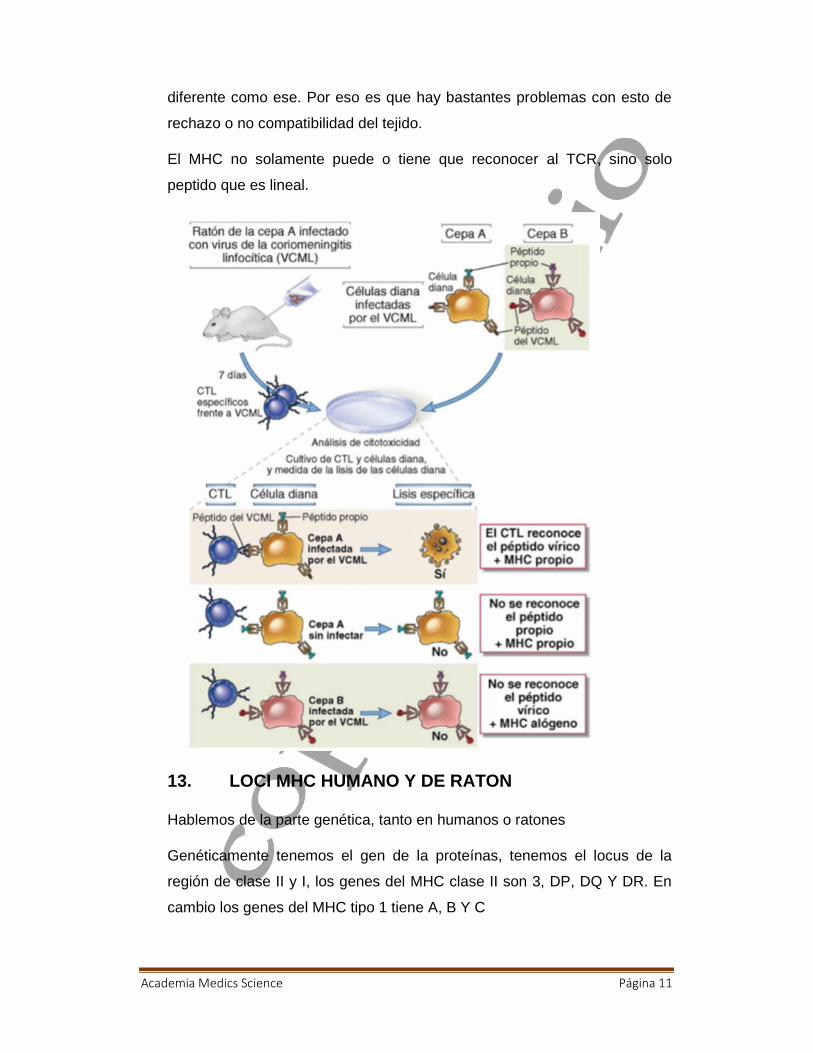
Academia Medics Science Página 11
diferente como ese. Por eso es que hay bastantes problemas con esto de
rechazo o no compatibilidad del tejido.
El MHC no solamente puede o tiene que reconocer al TCR, sino solo
peptido que es lineal.
13. LOCI MHC HUMANO Y DE RATON
Hablemos de la parte genética, tanto en humanos o ratones
Genéticamente tenemos el gen de la proteínas, tenemos el locus de la
región de clase II y I, los genes del MHC clase II son 3, DP, DQ Y DR. En
cambio los genes del MHC tipo 1 tiene A, B Y C
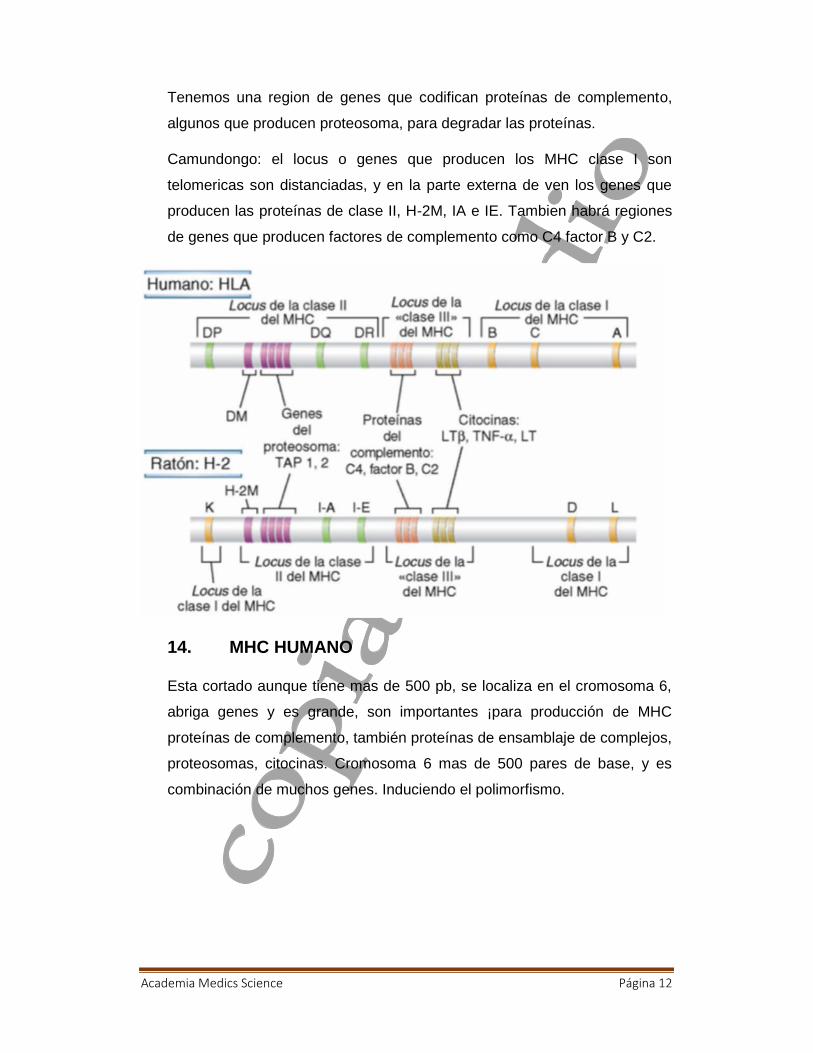
Academia Medics Science Página 12
Tenemos una region de genes que codifican proteínas de complemento,
algunos que producen proteosoma, para degradar las proteínas.
Camundongo: el locus o genes que producen los MHC clase I son
telomericas son distanciadas, y en la parte externa de ven los genes que
producen las proteínas de clase II, H-2M, IA e IE. Tambien habrá regiones
de genes que producen factores de complemento como C4 factor B y C2.
14. MHC HUMANO
Esta cortado aunque tiene mas de 500 pb, se localiza en el cromosoma 6,
abriga genes y es grande, son importantes ¡para producción de MHC
proteínas de complemento, también proteínas de ensamblaje de complejos,
proteosomas, citocinas. Cromosoma 6 mas de 500 pares de base, y es
combinación de muchos genes. Induciendo el polimorfismo.

Academia Medics Science Página 13
15. EXPRESION DE MHC e IFN- γ
Entonces la expresión de la proteína como es? Primero que esta es
ayudada por el IFN gamma, ayuda a producir proteínas MHC clase II, el de
clase I es con alfa y beta. Nuestra NK, va a reconocer y expresara IFN
gamma, que inducirá una alta expresión de MHC y moléculas
coestimulatorias para aumentar el chance de reconocimiento por la células
T, entonces esta células que comienza a ser madura comienza a expresar
grandes cantidades de MHC, presentado peptido antigenico que
presentara, la mayor APC o profesional o ligarse a varios linfocitos t??
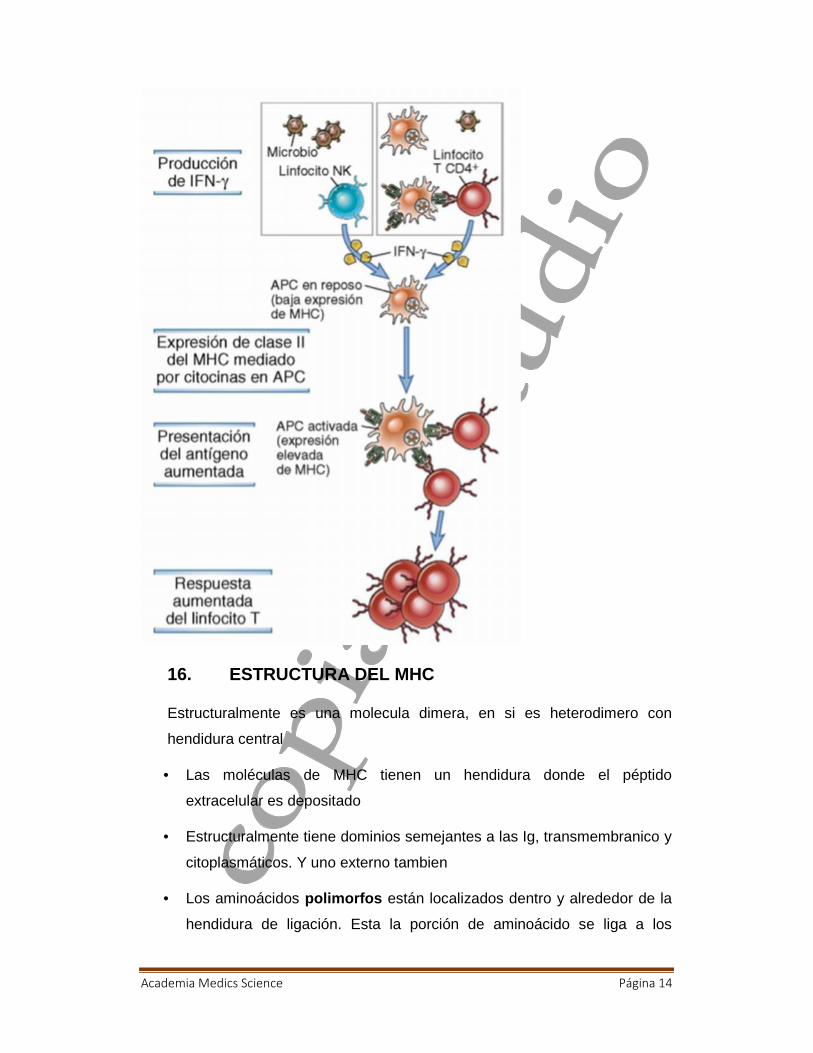
Academia Medics Science Página 14
16. ESTRUCTURA DEL MHC
Estructuralmente es una molecula dimera, en si es heterodimero con
hendidura central
• Las moléculas de MHC tienen un hendidura donde el péptido
extracelular es depositado
• Estructuralmente tiene dominios semejantes a las Ig, transmembranico y
citoplasmáticos. Y uno externo tambien
• Los aminoácidos polimorfos están localizados dentro y alrededor de la
hendidura de ligación. Esta la porción de aminoácido se liga a los

Academia Medics Science Página 15
péptidos que son presentados al TCR. Aminoácidos dentro de la
hendidura.
• Los dominios no polimorfos en los dominios de las Igs de las moléculas
MHC contienen sitios de ligación especificas para las moléculas CD4y
CD8 de las células T. son dominios constantes.
17. RESIDUOS POLIMORFICOS DE MHC
Entonces en los residuos polimórficos ustedes pueden ver la subunidad
alfa 1 y alfa 2, del MHC clase I, ven como un piso o una cama de estructura
beta plegada, y aca ven dos estructuras alfa hélice por alfa 1 y alfa 2, se
deposita el aminoácido o peptido antigenico, y ustedes se dan cuenta con
regiones de mucha variabilidad de rojo , o de poca variabilidad las azules
(pinten) cadena alfa dona un estructura alfa hélice, y dona 4 estructuras
beta plegadas, la estructura beta dona lo mismo. Esto será como una
camita donde de nuevo se deposita el péptido antigénico. Zonas de
aminoácido muy y poico variable. Aquí van a compartir la misma cantidad
en estructura en MHC clase II, recordar que si es en humanos es HLA
(Antigeno Leucocitico Humano).
18. MHC CLASE 1
La clase I es un heterodimero formado por una
cadena alfa y una cadena beta o mejor dicho
beta 2 microglobulina, y hay dos dominios. En la
beta 2 microglobulina es exclusivamente una
cadena con dominio de Ig, eran estructuras beta
antiparalelas por puentes disulfuro por ligación
con uniones convalente. Tiene un dominio
extracelular, transmembrana y citoplasmático.
Está formada por un domino de Ig osea un
domino alfa 3, y la hendidura de unión al
peptido antigenico es entre alfa 1 y alfa 2.

Academia Medics Science Página 16
19. MHC CLASE 2
Es diferente porque está formado por
dos cadenas alfa y beta, tiene
dominios de Ig unidas por puentes
disulfuro, tiene dominios de ligación
al antígeno, cada uno aporta una alfa
hélice y 4 secuencias beta plegada
antiparalelas. Estructuralmente son
diferentes, cadena beta tiene
variación hay beta 1 o 5, donde hay
regiones de variabilidad para que se
localice el antígeno. Donde ocupa
uno de 10 a 30 aminoácidos, en caso
de MHC I, será con antigenos cortos
que pesan de 8 a 11 aminoacidos.
Deben darse cuenta que tiene region
externa, transmembranica y citoplasmático.
20. UNION DE LOS PEPTIDOS A MHC
Como se unen molecularmente? Se ve un MHC clase I, que se deposita un
antígeno que es corto, no hay espacio para que entre un peptido tridimensional
o proteína grande por el espacio pequeño, se vene el grafico, en cambio en el
MHC clase II, no tanto porque el peptido es un poco mas grande 10 a 30 aa. El
mas eficiente seria de 16 aa dicen. El peptido en un corte ven como la
naturaleza del peptido que debe ser lineal, tiene aa de secuencia que forman
parte para depositarse en el MHC, y asi darse la ligación y asi formar buena
hendidura. Ayuda también a la unión con el TCR. Aunque no es lo mismo cada
uno que se una al MHC.
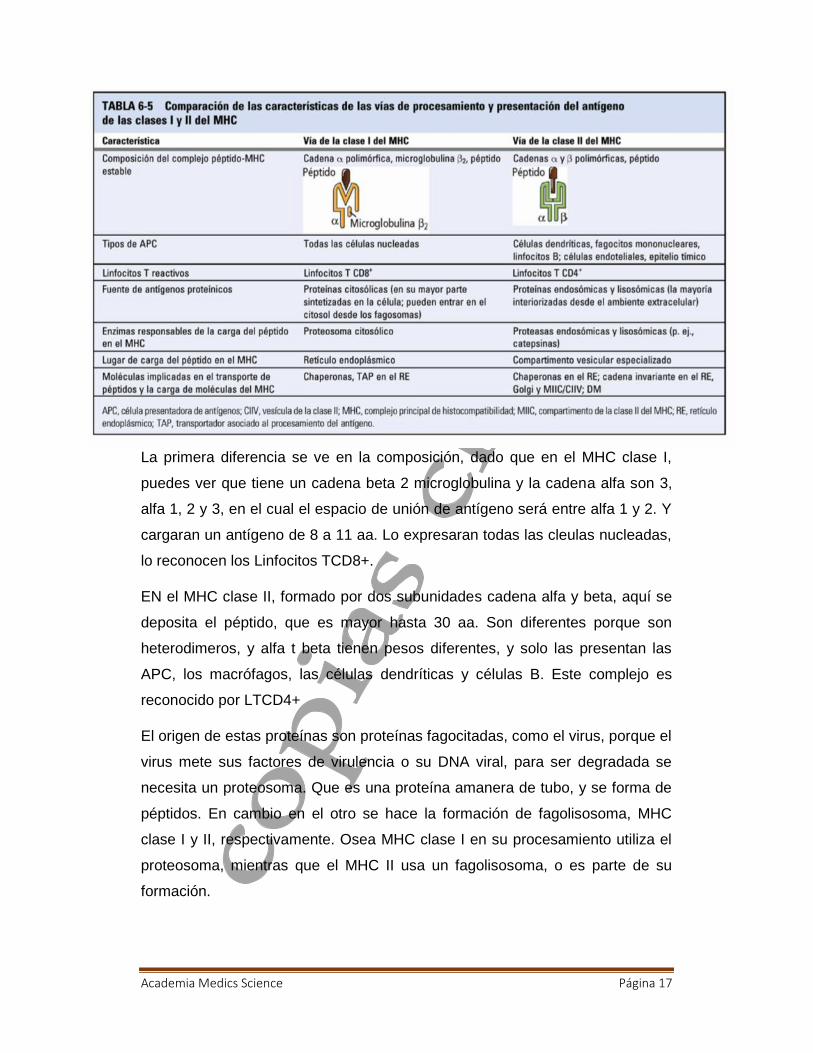
Academia Medics Science Página 17
La primera diferencia se ve en la composición, dado que en el MHC clase I,
puedes ver que tiene un cadena beta 2 microglobulina y la cadena alfa son 3,
alfa 1, 2 y 3, en el cual el espacio de unión de antígeno será entre alfa 1 y 2. Y
cargaran un antígeno de 8 a 11 aa. Lo expresaran todas las cleulas nucleadas,
lo reconocen los Linfocitos TCD8+.
EN el MHC clase II, formado por dos subunidades cadena alfa y beta, aquí se
deposita el péptido, que es mayor hasta 30 aa. Son diferentes porque son
heterodimeros, y alfa t beta tienen pesos diferentes, y solo las presentan las
APC, los macrófagos, las células dendríticas y células B. Este complejo es
reconocido por LTCD4+
El origen de estas proteínas son proteínas fagocitadas, como el virus, porque el
virus mete sus factores de virulencia o su DNA viral, para ser degradada se
necesita un proteosoma. Que es una proteína amanera de tubo, y se forma de
péptidos. En cambio en el otro se hace la formación de fagolisosoma, MHC
clase I y II, respectivamente. Osea MHC clase I en su procesamiento utiliza el
proteosoma, mientras que el MHC II usa un fagolisosoma, o es parte de su
formación.

Academia Medics Science Página 18
21. VIAS DEL PROCESO DE DEGRADACION YPRESENTACION DE ANTIGENO
Hay dos vías que vamos a estudiar:
MCH clase I, tenemos proteínas internas y necesitan ser linealizadas dentro del
proteosoma, y forman péptidos, y van a atravesar por las proteínas TAP, y se
va a anexar al MHC recién producido en retículo endoplasmatico y va a ser
exocitado, hacia la membrana y presentado a un LTCD8. Sucede todo lo
contrario con el MHC clase II, recordar que el MHC clase I, debio ser un
antígeno o péptido intracelular como un virus, en cambio MHC clase II de un
microrganismo extracelular, por eso será fagocitado.
En el MHC clase II, una endocitosis, adentro tiene un partícula y es degradado,
formación de fagolisosoma, y se una al RE, así se una al MHC clase II, tiene
una proteína o cadena invariante, para que le de estabilidad al MHC pero
luego cambiara de lugar con el antígeno y se va esta cadena, y asi presentar a
un LTCD4. (En el audio, enserio lo explico muy vagamente enserio no me ha
gustado nada :c lean el libro aunque les dejare una explicación de cada via de
procesamiento a mi estilo al final de la degra :3)
22. ANTIGENOS CITOSOLICOS Y EXTRACELULARES
Cuando hablamos de antígenos, podemos hablar de los citosolicos y los
extracelulares.
Si hubiera un antrigeno citosoilico puede ser antígenos virales, o producidos
externamente, por ejemplo si tengo un Gen de la albumina o lo coloca dentro
de una células eucariótica, y comenzara a producir ovoalbúmina, y esto va a
ser reconocida por el proteosoma, y será degradado y lo presentara un MHC
clase I.
Otro caso tengo de nuevo al gen de la ovoalbúmina, y nos damos cuenta que
entrara por cambios de temperatura o algo, y denuevo será reconocido
nuevamente sera degradado por el proteosoma y asi presentado este antígeno
mediante el MHC clase I.

Academia Medics Science Página 19
Ahora tenemos la ovoalbuimna y lo fagocita, y el APC fagocita esta albumina, y
será degradado por via lisosoma, y cuando ocurre esto, se presentan el péptido
proceso por via MHC clase II.
23. PRESENTACION DE ANTIGENOS INTRACELULARESVIA MHC DE CLASE I
Tienen un virus, el virua pego su ADN su material y lo coloca dentro de la
cleula hospedera, viene el ribosoma, y van a inducir la producción de proteínas,
en el ribosoma se encargara de producir la proteína y de ahí la formación
tridimensional, el punto es que forman una proteína, esta proteína DEBE SER
UBIQUITINIZADA. Son modificaciones postraduccionales. Y esta ubiquitinacion
es como una clave de reconocimiento, y asi en el Proteosoma, lo empieza a
lisar a la proteína formante, este proteosoma tiene partes beta y alfa, la beta se
encarga de dar los péptidos formados, y luego va a pasar que van a pasar a
través de una proteína de membrana que se llama TAP, cuando pasan vana
sufrir su ultimo corta, para estar en un tamaño ideal, por la enzima PARE,
entonces yo tengo aquí mi RE, y ahí tengo ribosomas que producirán mi
proteína que es un MHC clase I, y ahí tiene chaperonas, haciendo que mi MHC
no se desestructure, y tenga su forma tridimensional, luego se va

Academia Medics Science Página 20
transportando, y el MHC se une al TAP por medio de una enzima llamada
Tapasina. Entonces este complejo va a permitir que hay una mejor estabilidad
y no sea degradado, entonces cuando mi enzima pare, cliva mi péptido
antigénico y se depositara en el MHC, apenas se deposite, esta estructura de
libera. Y así ira a Golgi, y luego mediante vesicula exocitica. Asi será
presentado el MHC clase I al TCR pero del LTCD8.
24. PRESENTACION DE ANTIGENOSEXTRACELULARES VIA MHC DE CLASE II
En el caso de la presentación de MHC clase II, aquí tenemos la
partícula endocitada, aquí hay fusión con el lisosoma, porque el
lisosoma tiene enzimas líticas que harán que lo degrade a la
proteína, y asi tener antígenos, aquí pasan dos cosas, mis
antígenos son liberados, y que en el RE, tenemos un MHC clase II
por medio de ribosoma, tenemos una cadena alfa y beta, tenemos
proteína chaperona como calnexina que evitan el desemsamblaje
de la proteína, tenemos una cadena invariable, se depositara en la
hendidura de ligación con el antígeno, esto ayuda a la manutención
de la zona de unión del antígeno, luego se transporta al complejo
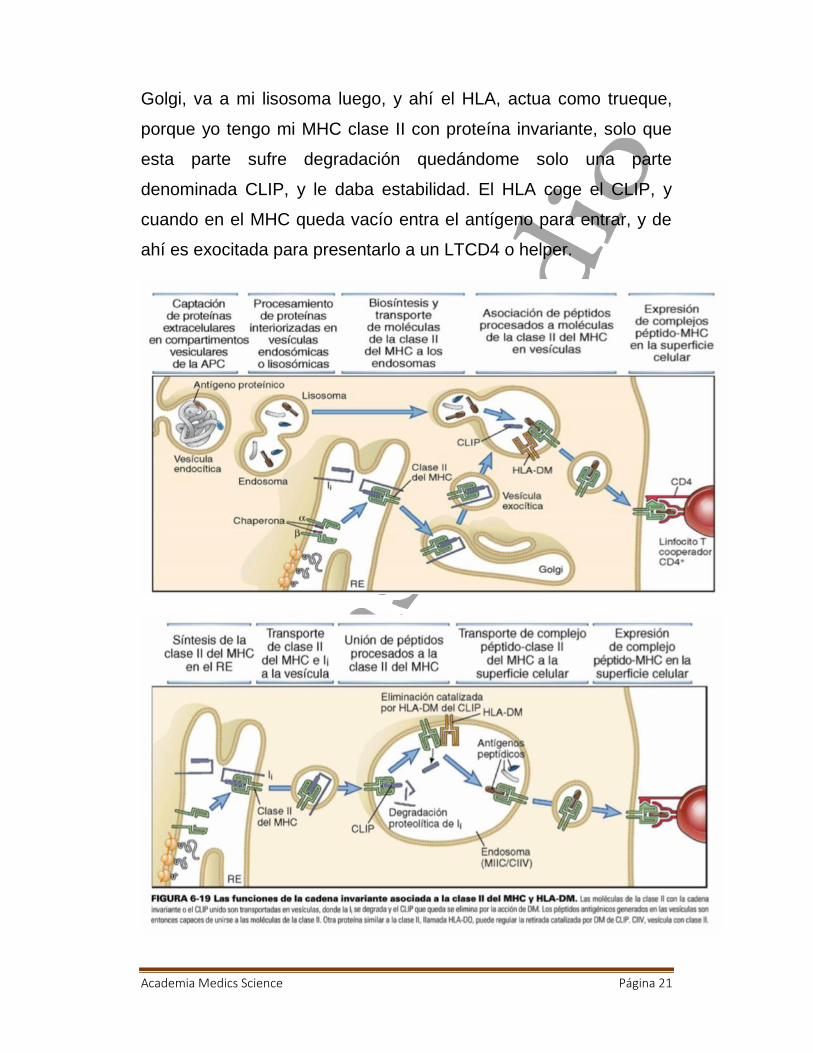
Academia Medics Science Página 21
Golgi, va a mi lisosoma luego, y ahí el HLA, actua como trueque,
porque yo tengo mi MHC clase II con proteína invariante, solo que
esta parte sufre degradación quedándome solo una parte
denominada CLIP, y le daba estabilidad. El HLA coge el CLIP, y
cuando en el MHC queda vacío entra el antígeno para entrar, y de
ahí es exocitada para presentarlo a un LTCD4 o helper.

Academia Medics Science Página 22
25. PRESENTACION CRUZADA DE ANTIGENOS A LASCELULAS TCD8
La presentación cruzada, cuando una células dendrítica que captura
antígenos externos, e imaginen el caso que fagocite una con una
bacteria intracelular, o que llega a fagocitar una celula infectada por
un virus o una tumoral. En eso caso se da un presentación cruzada.
( Lo que para es que si es intracelular actua MHC clase I, pero
como lo fagocito debería ser MHC clase I, entonces hay reacción
cruzada), aquí tu tienes tu APC, que es una célula dendrítica y
produce grandes cantidades de MHC, y no sabe que producir asi
que produce ambas, porque capta virus exocitadas, como proteínas
internas. Entonces son captados por unas células TCD8, que son
específicas para este virus, pero a su vez puede presentar via MHC
clase II, entonces se cruza. Osea tiene capacidad de producir los
dos MHC y activar dos tipos diferentes de células T.
26. PRESENTACION DE ANTIGENOS A CELULAS T
Cuando se da la presentación de antígeno, ustedes tiene la parte
que capta el antígeno puede ser propio citosolico, fagocitado o
receptor tipo TLR o de complemento o puede ser antígenos

Academia Medics Science Página 23
tridimensionales también captados pero por via BCR, osea tenemos
varios tipos de presentación de antígeno, y puede ser dado por
celula dendrítica, macrófago o células B.
Va a haber un reconocimiento e inducirá a la destrucción de linfocito
citotóxico, que botsa sus granulos
En macrófago habrá fagocitosis, y lo presenta al linfocito T, y asi
será activado para degradar al péptido.
Los lin B reconocen el antígeno, y lo presentan con MHC II, al
linfocito TCD4, y estas producen citosinas como IL2, Y ASI
ACTIVAR A LOS LINFOCITOS B PARA PRODUCIR
ANTICUERPOS ESPECIFICOS PARA ANTIGENO.
27. INMUNODOMINANCIA DE PEPTIDOS
Ustedes tiene un antígeno, es una estructura tridimensional con
diferentes seucencias de aa, todos pueden formar diferentes
epitopos, que serán reconocidas por el MHC, y viene la Células
dendrítica, y lo capta. En si TODO PEPTIDO ANTIGENICO TIENE
LA CAPACIDAD DE SER PRESENTADO A UNA CELULAS T, hay
una inmunológicamente dominanate y ese se presenta, y esto es LA
DOMINANCIA, ES EL PEPTIDO ANTIGENICO QUE DARA UN
RPTA INMUNA EFECTIVA, PARA DAR CLONACION DE
CELULAS T.
28. PRESENTACION DE ANTIGENOS NO PROTEICOS
• Estos Ag no proteico son reconocidos por algunos subtipos
de células B
• Las células NKT y la células Tγδ reconocen estos antígenos
no proteicos.

Academia Medics Science Página 24
• Estos lípidos e glucolípidos son presentados por MHC no
clásico parecido estructuralmente a la clase 1 denominado
de CD1.
• El complejo CD1-lipído en la membrana sufre endocitosis
por endosomos o lisosomas donde los lipídos ya habían
sido ingeridos son nuevamente endocitados para ser
reciclados.
Buenos chicos eso seria todo, disculpen las fallas ortográficas o
cualquier lapsus que haya tenido, es que la clase en si, a mi me
parecio un poco inconclusa, hasta casi confusa, no se dejo
entender tan bien. Bueno como se lo prometi, les dejare aquí,
una forma mas a mi estilo de explicar las vías de procesamiento.
PARA ESTE TEMA SE DEBE LEER EL CAPITULO 6 DEL
LIBRO ABBAS. LEANLO! ES ENSERIO! :3 un abrazo teletubie!
Como sabíamos existen patógenos intracelulares y extracelulares.
INTRACELULARES: En el citoplasma aparece un producto proteico delmicrooganismo, se procesa en el citosol y se asocia al REL, MHC I sesintetiza en el REL y Ag + MHC = reconocimiento del LTCD8+.
EXTRACELULAR: APC fagocita, degrada en pequeños fragmentos y almismo tiempo sintetiza MHC II. El MHC II se sintetiza en el REL y se une alendosoma. Antigeno + MHC = reconocimiento LTCD4+.
VIAS DE PROCESAMIENTO DE ANTIGENOS POR MHC TIPO 1
En primer lugar vamos a tener que el MHC tipo 1 va a ser reconocido porel LTCD8, osea reconoce microorganismo intracelulares, puedes ser queeste haya ingresado y haya sintetizado su proteínas, o que lo fagocite y lohaga entrar. El antigeno en forma de proteína, como tenemos dosproteínas, vamos a necesitar degradarlo en péptidos, pero toda aquella

Academia Medics Science Página 25
proteína extraña para nosotros o este mal plegada o parte demicroorganismo será UBIQUITINIADA, osea marcada con ubiquitina, va aservir de señal para que pase por una especia de licuadora denominadaProteosoma. Para hacer que esta proteína se vuelva péptidos pase através de una proteína TAP para que entre al REL, y asi entra estospéptidos al REL, por otro lado en el REL, se iba a dar el ensamblaje de lascadenas de MHC, alfa y beta o microglobulina, la cadena es inestable porlo tanto necesita una chaperona llamada calnixina, donde la va a llevar yobservamos que se pegue al lado de TAP en busca de un antigeno, estaforma de pegarse es gracias a la Tapasina. Una vez que este MHC se pegóal TAP, el antigeno va a pegarse por las chaperonas calreticulina y Erp57que van a hacer que ag se una al MHC una vez dado esto, este complejode unión va a agarrar y a través de una vesícula exocitica irse al Golgi yluego a la membrana celular para ser reconocida por el LTCD8.
VIAS DE PROCESAMIENTO DE ANTIGENOS POR MHC TIPO 2
El MHC tipo 2 va a estar reconocido por el LTCD4 y presentar los Agextracelulares. Vemos un ag extracelulares, se forma un fagolisosoma. Y sevuelve péptidos. Por otro lado en el REL, se va a sintetizar el MHC tipo 2, launión de las cadenas son inestables, necesitan chaperona llamada cadenainvariante o li, esta chaperona va a estabilizar complejo llevándolo alaparato de Golgi, y luego a través de vesícula exocitica, va a llegar alfagolisosoma, donde estaba el péptido, aquí ocurre un problema, lacadena invariante o li es grande, entonces lo que nosotros queremos esdeshacernos del li porque ahí está el ag, entonces reducimos al li, dondeserá reducida a CLIP, Esta molécula CLIP se va a ubicar en el lugar donde seubica el antigeno. Entonces como es que liberamos el MHC del CLIP, con elHLA-DM. Entonces el antígeno entra donde MHC, y se expulsa a lamembrana celular y así es presentado al LTCD4.