
Centro de Investigación en Alimentación y Desarrollo, … · realizada en la Facultad de Ciencias...
121
Centro de Investigación en Alimentación y Desarrollo, A. C. “RESPUESTAS BIOQUÍMICAS INDUCIDAS POR IRRADIACIÓN UV-C EN FRUTOS DE PAPAYA (Carica papaya L.) var. MARADOL ALMACENADOS A BAJAS TEMPERATURAS” POR M.C. DULCE MARÍA RIVERA PASTRANA TESIS APROBADA POR LA: COORDINACIÓN DE TECNOLOGÍA DE ALIMENTOS DE ORIGEN VEGETAL UNIDAD HERMOSILLO COMO REQUISITO PARCIAL PARA OBTENER EL GRADO DE: DOCTOR EN CIENCIAS HERMOSILLO, SONORA JUNIO DEL 2010
Transcript of Centro de Investigación en Alimentación y Desarrollo, … · realizada en la Facultad de Ciencias...

Centro de Investigación en Alimentación y Desarrollo, A. C.
“RESPUESTAS BIOQUÍMICAS INDUCIDAS POR IRRADIACIÓN
UV-C EN FRUTOS DE PAPAYA ( Carica papaya L.) var. MARADOL
ALMACENADOS A BAJAS TEMPERATURAS”
POR
M.C. DULCE MARÍA RIVERA PASTRANA
TESIS APROBADA POR LA:
COORDINACIÓN DE TECNOLOGÍA DE ALIMENTOS DE ORIGEN VEGETAL UNIDAD HERMOSILLO
COMO REQUISITO PARCIAL PARA OBTENER EL GRADO DE:
DOCTOR EN CIENCIAS
HERMOSILLO, SONORA JUNIO DEL 2010



iv
AGRADECIMIENTOS
Al CONACYT por el financiamiento otorgado para la realización de este trabajo.
Al CIAD A. C., unidad Hermosillo, por permitirme formar parte de su programa de
posgrado y de su comunidad.
A mi director de tesis, Dr. Gustavo A. González Aguilar, por recibirme en su equipo
de trabajo, por su contribución al proyecto, y por su apoyo y paciencia.
A los miembros del comité, los doctores: Alfonso Gardea Béjar, Elhadi Yahia Kazuz,
Miguel A. Martínez Téllez, Marisela Rivera Domínguez y Rogerio Sotelo Mundo,
gracias por su tiempo y disposición para enriquecer este trabajo.
Al Dr. Yahia y a la Universidad Autónoma de Querétaro por recibirme en la estancia
realizada en la Facultad de Ciencias Naturales y por las facilidades brindadas
durante ese tiempo.
A la Coordinación de Alimentos de Origen Vegetal por el respaldo académico y
administrativo.
A los académicos y técnicos de la CTAOV, en especial a: Dr. Fernando Ayala
Zavala, Q.B. Mónica A. Villegas Ochoa, M.C. Reynaldo Cruz, M.C. Chrystian M.
Rodríguez, gracias por el apoyo y consejos, pero sobretodo por su amistad dentro y
fuera del CIAD.
A mis compañeros de los laboratorios de Bioquímica y Fisiología de Frutas y
Hortalizas, y Frutos Frescos Cortados gracias por hacer tan agradable la
convivencia diaria.

v
Al personal administrativo y de biblioteca por facilitar la información necesaria para
este trabajo y en general por hacer posibles todos los trámites para la culminación
del grado.
A mi familia y amigos por estar ahí cuando más los necesito, incluso en el
laboratorio. Especialmente a mi esposo, Carlos M. Múzquiz, gracias por tu apoyo
incondicional y comprensión durante estos años, pero principalmente por estar
siempre conmigo amor.
…Muchas Gracias.

vi
CONTENIDO
Página
SINOPSIS . . . . . . . . . . . . . . . . . . . . . . . . vii
CONCLUSIONES GENERALES . . . . . . . . . . . . . . . xvii
CAPÍTULO I. Efectos Bioquímicos Postcosecha de la Ir radiación UV-C en Frutas y Hortalizas . . . . . . . . . . . . 1
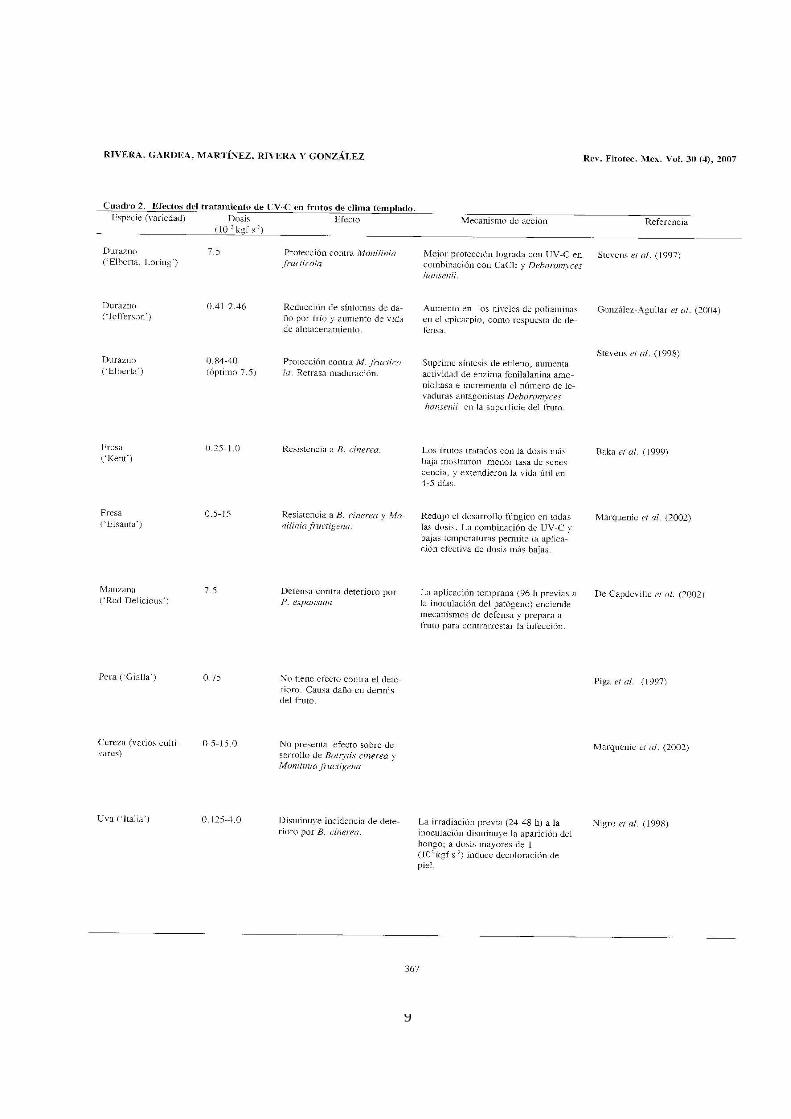
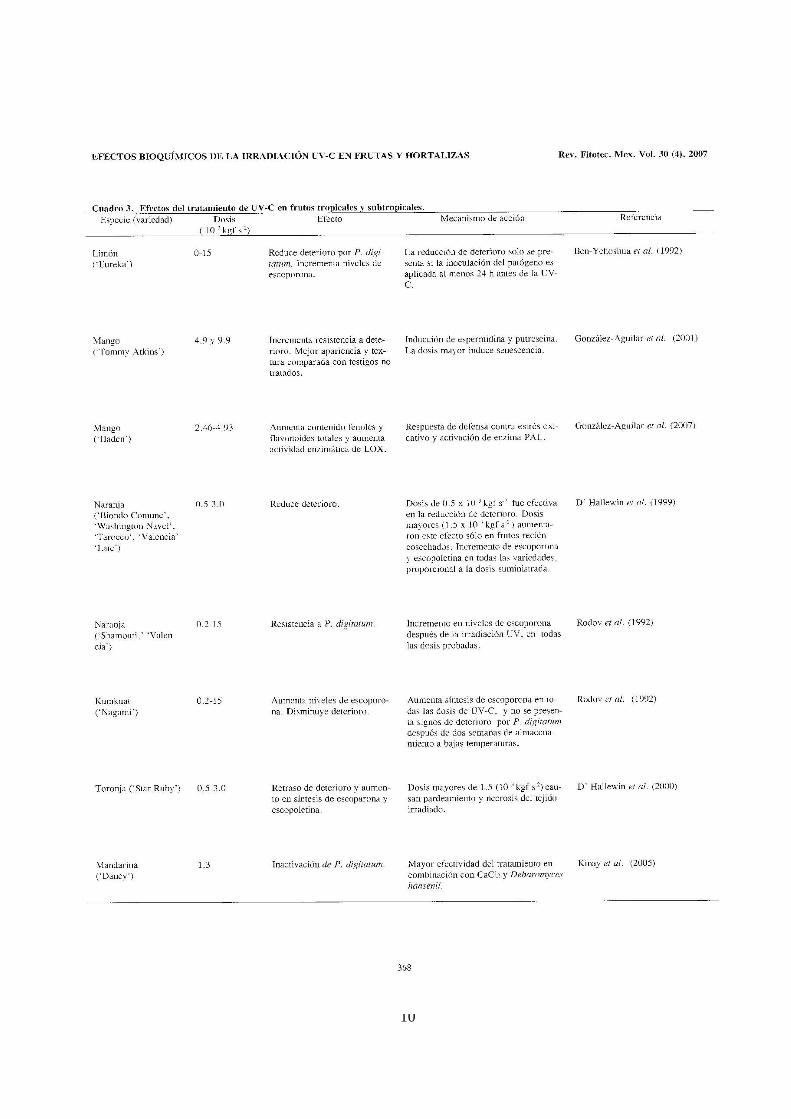
Rivera-Pastrana D M, Gardea Béjar A A, Martínez-Téllez M A, Rivera-Domínguez M, González-Aguilar G A. Efectos Bioquímicos Postcosecha de la Irradiación UV-C en Frutas y Hortalizas. Revista Fitotecnia Mexicana. 2007. 30 (4):361-371.
CAPÍTULO II. Perfiles de fenoles y carotenoides de papaya (Carica papaya L.) y su contenido a bajas temperaturas de almacenamiento . . . . . . . . . . . . . . . 15
Rivera-Pastrana, D.M., Yahia, E., González-Aguilar, G.A. Phenolic and carotenoid profiles of papaya (Carica papaya L.) and their contents under low temperature storage. Enviado a la revista Journal of the Science of Food and Agriculture, JSFA-10-0225. R1.
CAPÍTULO III. Efecto de la irradiación UV-C y temper atura de almacenamiento en la calidad postcosecha de papay a
‘Maradol’ . . . . . . . . . . . . . . . . . . . 49
Rivera-Pastrana, D.M., González-Aguilar, G.A., Rivera-Domínguez, M., Martínez-Téllez, M. A. Manuscrito para enviar a la revista Chapingo Serie Horticultura.
CAPÍTULO IV. Efecto de la irradiación UV-C y bajas temperaturas de almacenamiento sobre compuestos bioactivos, enzimas antioxidantes y actividad de inhibición de radicale s de frutos
de papaya . . . . . . . . . . . . . . . . . . . 74
Rivera-Pastrana, D.M., González-Aguilar, G.A., Ayala-Zavala, J.F. Gardea Béjar, A. A., Sotelo-Mundo, R. Effect of UV-C irradiation and low temperature storage on bioactive compounds, antioxidant enzymes and radical scavenging activity of papaya fruit. Manuscrito para enviar a la revista Postharvest Biology and Technology.
Producción Académica derivada de la tesis . . . . . . . . . . 102

vii
SINOPSIS
Los frutos tropicales a pesar de ser altamente perecederos, se han posicionado en
los mercados internacionales por su alta demanda debido a sus excelentes
características organolépticas y alto valor nutricional. Estudios epidemiológicos han
demostrado que su consumo previene o retrasa la aparición de enfermedades
crónico-degenerativas como el cáncer y la diabetes. La papaya es de los frutos
tropicales con mayor demanda en los mercados internacionales, ocupando el quinto
lugar en el mercado mundial. Sin embargo, su comercialización presenta tres
principales problemas: el desarrollo de hongos, su alta susceptibilidad a las bajas
temperaturas y la corta vida poscosecha. Considerando que México se ha
mantenido en la última década entre los principales productores y exportadores de
papaya a nivel mundial; la búsqueda de alternativas para el control de estos
problemas resulta no sólo pertinente, sino necesario.
Si se sobrepasa la temperatura óptima (15-16 °C) en el almacenamiento a bajas
temperaturas como herramienta para alargar la vida poscosecha, la papaya, puede
presentar diferentes fisiopatías como el desarrollo de síntomas de daño por frío, que
afectan la calidad y aceptabilidad del fruto. El daño por frío (DF) es un desorden
fisiológico que presentan los tejidos vegetales sensibles a las temperaturas bajas, y
los síntomas se hacen más evidentes al transferir el fruto a temperaturas
superiores. Se han propuesto diversos mecanismos para explicar el DF; la mayoría
de los autores coinciden en que los primeros cambios ocurren en la membrana
celular, afectación del metabolismo normal y un aumento en las especies reactivas

viii
de oxígeno, que pueden llegar a dañar los fosfolípidos de las membranas y otras
biomoléculas.
Para contrarrestar el estrés oxidativo, las plantas superiores poseen un sistema
antioxidante (enzimático y no enzimático), que involucra antioxidantes hidrofílicos
(compuestos fenólicos, ascorbato) y lipofílicos (carotenoides, vitaminas) así como
enzimas antioxidantes. Este sistema estabiliza los radicales superóxido (O2-),
hidroxilo (OH•) y peróxido de hidrógeno (H2O2), secuestrándolos o inactivándolos,
evitando el daño célular. Los aniones superóxido se eliminan mediante la acción de
la enzima superóxido dismutasa (SOD) que produce peróxido de hidrógeno. Las
enzimas catalasa (CAT) y peroxidasa (POD) descomponen el peróxido de
hidrógeno en agua y oxígeno, y pueden localizarse en los cloroplastos, mitocondria
y citoplasma de las células. Se ha observado en algunos frutos, que la reducción
de desórdenes fisiológicos ocasionados por estrés oxidativo como el DF, puede
estar relacionada con la activación del sistema antioxidante.
Se han utilizado distintas tecnologías postcosecha para el control del desarrollo de
los síntomas de daño por frío (DF) en papaya, que se basan en el control de la
maduración o en la inactivación de enzimas involucradas en la aparición de los
síntomas del DF. Entre las más empleadas se encuentran: los tratamientos
hidrotérmicos1, la aplicación de compuestos químicos como el 1MCP (metil-
ciclopropeno) empleado como retardador de maduración2; la combinación de
atmósferas modificadas con metil-jasmonato3 o fungicidas y altas/bajas
temperaturas4.
1 Huajaikaew, L., Uthairatankij, A., Kanlayanarat, S., and Gemma, H. 2005. Acta Hort.682:1063-1068. 2 Manenoi, A., Paull, R.E. 2007. Physiologia Plantarum. 131: 470-480. 3 González-Aguilar, G.A., Buta, J., Wang, C.Y. 2003. Postharvest Biol. Technol. 28: 361.370. 4 Pérez-Carrillo, E., Yahia, E.M. 2004. Food Qual. 27: 127-139.

ix
Una de las tecnologías emergentes, que podría ser utilizada en postcosecha para
reducir el daño por frío y el deterioro en frutos y hortalizas frescos es la irradiación
UV-C.
El CAPÍTULO I presenta una revisión de los últimos 20 años de los efectos del
tratamiento de UV-C en frutas y hortalizas en postcosecha. Aquí se describen los
fundamentos y principios de la irradiación UV-C, el concepto de hórmesis y el
mecanismo de acción biológico en microorganismos y en particular en plantas.
Enseguida se detallan los efectos del tratamiento en los cambios fisiológicos,
bioquímicos, nutricionales y los efectos antimicrobianos y adversos, en diferentes
productos hortofrutícolas. También se describen los mecanismos involucrados en la
reducción del deterioro y algunos síntomas de daño por frío en varios frutos en
postcosecha.
La revisión termina sugiriendo que el tratamiento de UV-C puede promover cambios
benéficos en el metabolismo de algunas frutas y hortalizas, los cuales están en
función del tipo de producto, dosis y factores ambientales. De acuerdo a los
resultados reportados concluye que el tratamiento de UV-C en postcosecha
aumenta en diferente medida los niveles de fitoalexinas, compuestos y enzimas
antioxidantes y vitaminas, los cuales se han relacionado con la reducción del
deterioro causado principalmente por microorganismos.
Por lo que consideramos que esta tecnología podría ser utilizada para reducir el
deterioro fúngico de frutos de alta importancia en México como la papaya Maradol,
así como profundizar en el conocimiento de algunos de los posibles mecanismos
por los cuales se induce estas respuestas en el fruto. Este cultivar es uno de los de

x
mayor producción, comercialización e importancia económica en México. Por lo que
el objetivo de este trabajo fue estudiar los posibles efectos fisiológicos y bioquímicos
de distintas dosis de UV-C en frutos de papaya almacenados a diferentes
temperaturas.
Con estos antecedentes, nos planteamos la hipótesis de que la irradiación UV-C
induce la síntesis de compuestos y/o enzimas antioxidantes en papaya Maradol,
que se relacionan con la reducción de fisiopatías desarrolladas en los frutos
almacenados a bajas temperaturas.
Para contrastar esta hipótesis, primeramente, determinamos el efecto del
almacenamiento en frío en el perfil y contenido de compuestos fenólicos y
carotenoides que constituyen los principales componentes del sistema antioxidante
no enzimático y verificar si su síntesis es alterada por el tratamiento. Después se
procedió a evaluar la efectividad del tratamiento de UV-C (1.76 kJ·m-2) para
mantener la calidad y reducir el daño por frío de los frutos de papaya almacenados
a 1, 5, 14 y 25 °C. En una tercera etapa, se invest igó si el tratamiento aplicado a
temperaturas bajas empleadas comercialmente (5 y 14 °C), inducía cambios en el
sistema antioxidante enzimático (medido como actividad de SOD, CAT y POD); y
no-enzimático (medido como contenido de fenoles y flavonoides totales). En este
último ensayo también se evaluó la capacidad antioxidante total (DPPH y TEAC),
para determinar si los cambios en algunos componentes del sistema antioxidante
por efecto del tratamiento, modificaban la calidad nutricional del fruto. Los
resultados de estas investigaciones son descritos en los Capítulos II al IV de esta
tesis.

xi
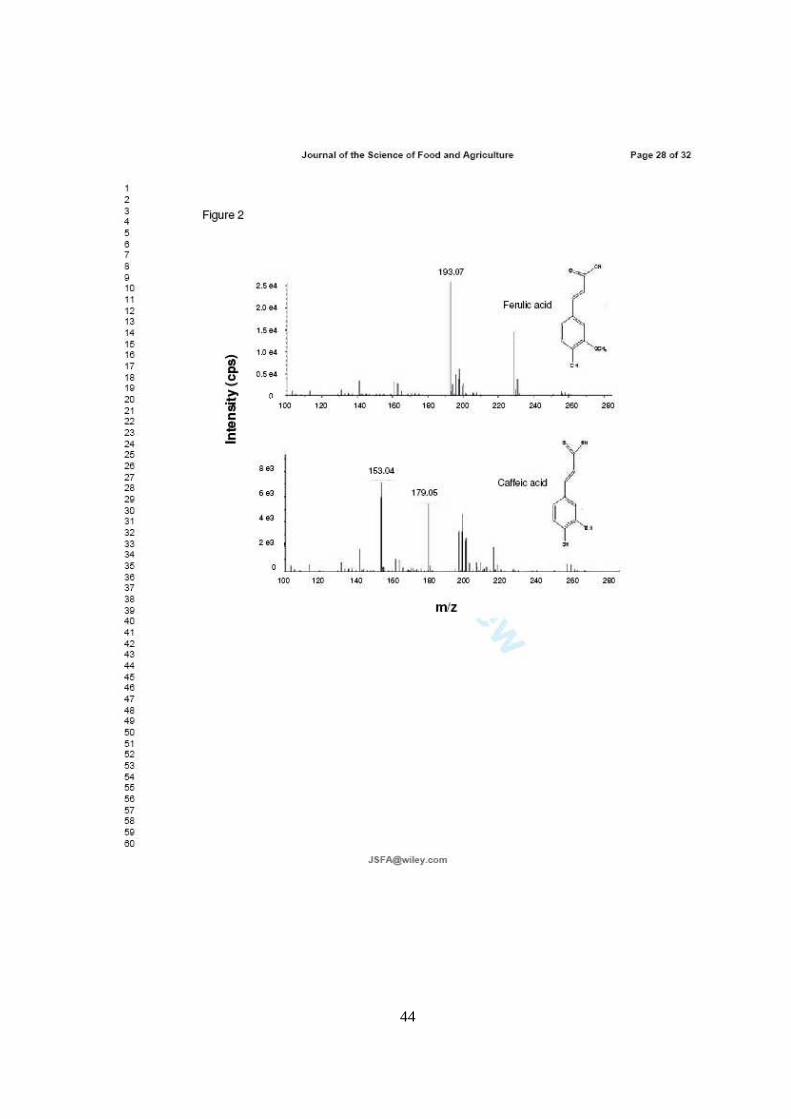
En el CAPITULO II se muestra la caracterización de los compuestos bioactivos
mayoritarios de los frutos de papaya Maradol, realizada por espectrometría de
masas, y el posible efecto del almacenamiento en frío. Las muestras en estado
verde-maduro fueron divididas en dos grupos al azar que fueron almacenaron a 1 ó
25 °C, y se evaluaron cambios en el perfil y conten ido de compuestos fenólicos y
carotenoides en tejido de cutícula y pulpa, cada 4 días durante 12 días. Para la
identificación de compuestos fenólicos, se analizaron extractos metanólicos crudos
e hidrolizados para identificar fenoles libres y conjugados, respectivamente. Se
logró identificar en la cutícula de los frutos 11 compuestos fenólicos, entre ácidos
fenólicos y flavonoides: derivados glucosídicos de los ácidos cafeico, ferúlico, gálico
y protocateico, así como los ácidos libres cafeico y ferúlico, y los flavonoides
quercetina, rutina, isoramnetina y kaempferol. Mientras que en la pulpa se
detectaron cuatro derivados glicosilados de ácido caféico y protocateico. Sin
embargo, las concentraciones en las que se encontraron estos compuestos fueron
muy bajas con excepción de los ácidos cafeico y ferúlico, y el flavonoide rutina en la
cutícula del fruto, que poseen un alto potencial antioxidante.
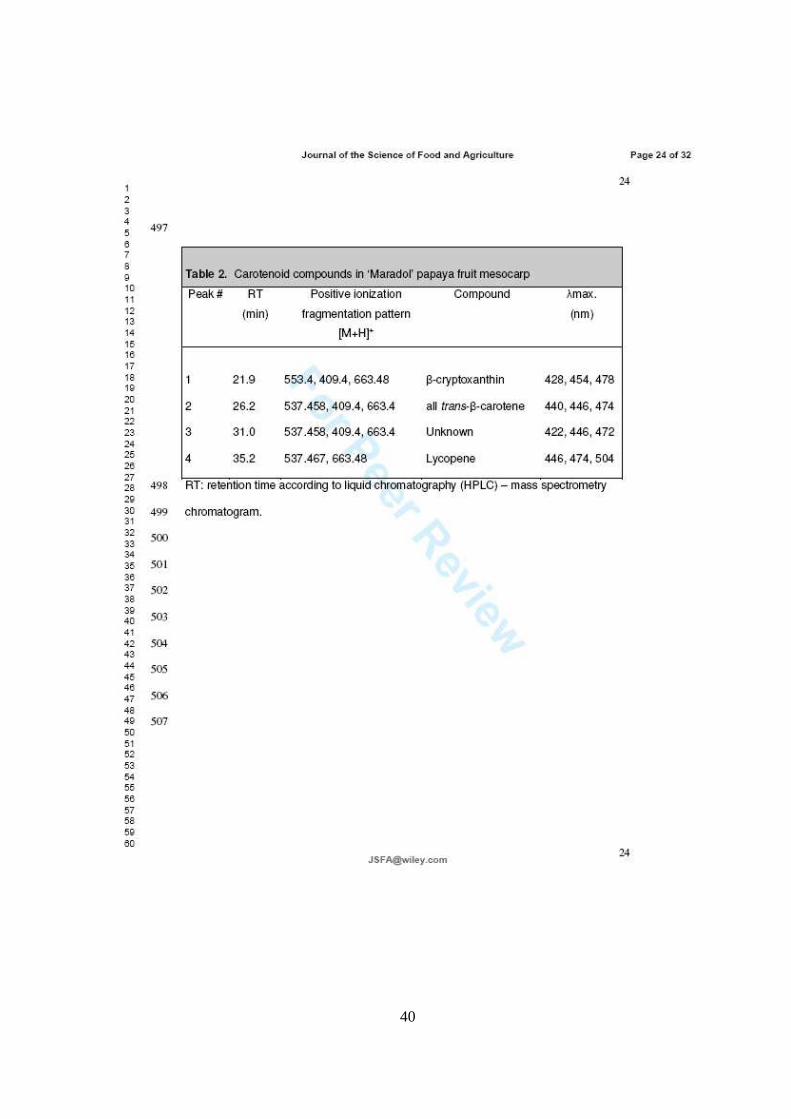
Por otra parte, el perfil de carotenoides se evaluó únicamente en la pulpa del fruto,
ya que es donde se encuentran en mayor concentración. La identificación y
cuantificación de carotenoides se realizó en extractos lipofílicos saponificados
identificándose como mayoritarios a licopeno, β-caroteno y β-criptoxantina.
Una vez identificados aquellos compuestos bioactivos que podían ser cuantificados
con buena reproducibilidad por cromatografía de líquidos, se determinó el efecto del

xii
almacenamiento en frío (1 °C) sobre su concentració n. No se encontraron
diferencias significativas (P> 0.05) en el contenido de los compuestos fenólicos
cafeico, ferúlico y rutina, ni de β-caroteno, por efecto de la temperatura de
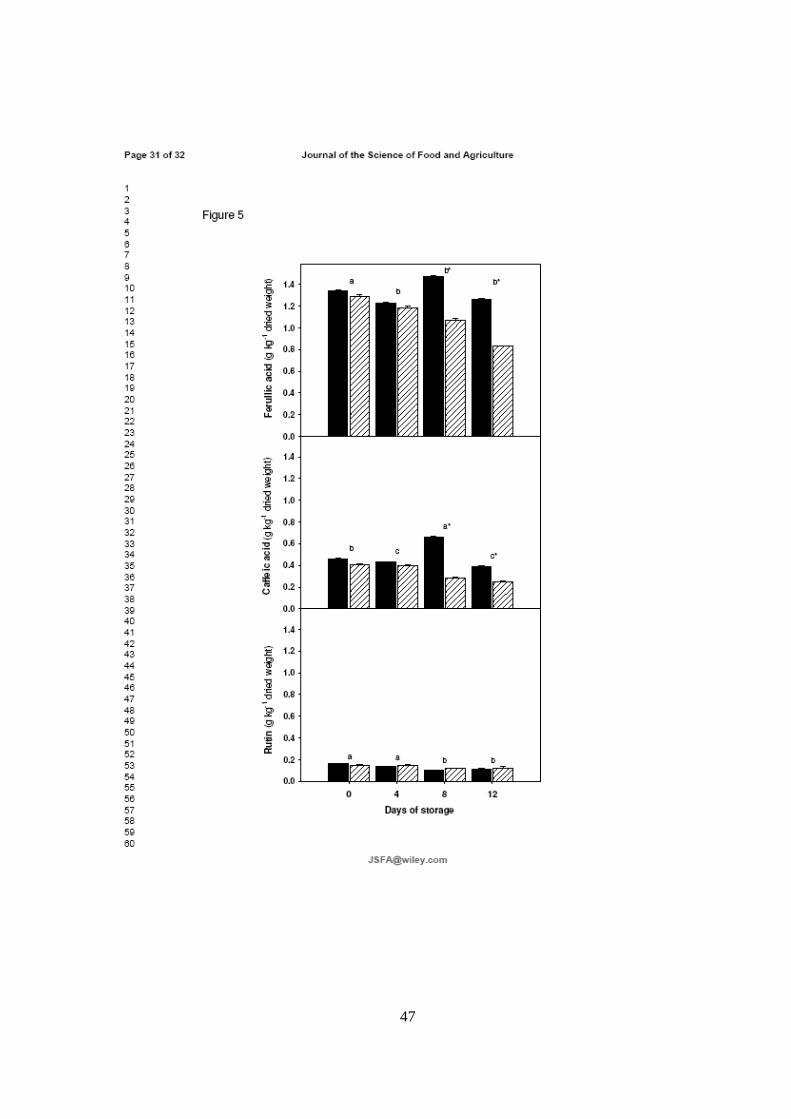
almacenamiento durante 12 días. Sin embargo, los niveles de ácidos ferúlico y
cafeico aumentaron significativamente entre los días 8 y 12 a 1 °C en comparación
con las muestras almacenadas a 25 °C. Por otra part e, se observó un claro efecto
inhibitorio de las bajas temperaturas en el contenido de licopeno y β-criptoxantina.
Estos resultados muestran que el estrés ocasionado por las bajas temperaturas
reduce la biosíntesis de carotenoides, afectando la calidad nutricional de la pulpa.
Mientras que la estabilidad en niveles de los compuestos fenólicos mayoritarios en
la cutícula del fruto, sugiere que este estrés no modifica su perfil a las temperaturas
evaluadas, e incluso incrementa intermitentemente el contenido de ácidos fenólicos
con alta capacidad antioxidante.
EL CAPITULO III presenta el efecto de la irradiación UV-C en la calidad de frutos de
papaya almacenados a bajas temperaturas. En este capítulo, se describen los
efectos del tratamiento de UV-C (1.76 kJ·m-2) en las variables de color (L, Chroma y
°Hue), pérdida de firmeza, pérdida de peso, sólido s solubles totales, deterioro de la
apariencia general y desarrollo de síntomas de daño por frío en los frutos. La dosis
de UV-C empleada en este ensayo fue determinada en experimentos preeliminares
que no son descritos en esta tesis, donde se seleccionó la dosis más alta dentro
del rango hórmico reportado en la literatura, que no provocó daño directo visible en
la cutícula de los frutos como quemaduras o abrasiones. Las muestras irradiadas

xiii
con UV-C y los testigos fueron divididos en grupos iguales, almacenados a cuatro
diferentes temperaturas 1, 5, 14 y 25 °C por 12 día s, + 2 días a 25 °C. Se
determinó el cambio en cada variable investigada como la diferencia entre los
valores iniciales y finales observados para un mismo grupo de muestras.
Al analizar los resultados, se encontró que el almacenamiento a bajas temperaturas
promueve un efecto negativo en los cambios de color y firmeza en los frutos testigo,
comúnmente asociados al desarrollo de síntomas de daño por frío. Durante el
almacenamiento a 1, 5 y 14 °C, los cambios en lumin osidad (L), en la intensidad de
color (Chroma) y en el ángulo de matiz (°Hue) dismi nuyeron significativamente y de
manera proporcional a la temperatura. Estos cambios en los parámetros de color
ocasionados por las bajas temperaturas concuerdan con los reportados en
estudios previos que argumentan un retraso de la maduración de los frutos. El
tratamiento de UV-C, retrasó el desarrollo de color del fruto maduro, y además,
aumentó el oscurecimiento en los frutos almacenados a 1 y 5 °C.
Por otra parte, se observó una disminución significativa en la pérdida de peso y el
deterioro en los frutos tratados con UV-C, respecto a los frutos testigo durante el
almacenamiento a todas las temperaturas evaluadas. El tratamiento en sí, no redujo
la pérdida de peso, pero los cambios ocurridos en este fruto podrían estar
relacionados con el retraso de la maduración y reducción de deterioro microbiano, lo
cual coincide con diferentes estudios reportados en la literatura y que se describen
ampliamente en el Capítulo I.
Los resultados obtenidos de esta parte del estudio nos indican que si bien la
irradiación UV-C induce un estrés oxidativo, la activación del sistema antioxidante

xiv
de papaya por el tratamiento de UV-C, no es suficiente para contrarrestarlo. Cada
especie y variedad de fruto responde de diferente forma a los distintos tipos de
estrés a los que son sometidos. Para el caso de la papaya Maradol, encontramos
que los efectos benéficos inducidos por la UV-C son inferiores a los observados en
otras especies.
Uno de los principales propósitos de esta investigación, consistió en probar la
efectividad de tratamiento de UV-C en la reducción del daño por frío en papaya, sin
embargo, se encontró que no reduce los síntomas de este desorden bajo las
condiciones de este ensayo, e incluso los incrementa en frutos almacenados a 5 °C.
Con el objetivo de conocer si la irradiación UV-C tiene un efecto significativo en el
sistema antioxidante en papaya, se evaluaron los cambios en el contenido de
compuestos antioxidantes medidos como fenoles y flavonoides totales, actividad de
las enzimas antioxidantes SOD, CAT y POD, y en la capacidad antioxidante medida
como inhibición de los radicales DPPH y ABTS (CAPITULO IV).
Se seleccionaron frutos en estado verde-maduro y se formaron dos grupos al azar.
El tratamiento con UV-C (1.76 kJ·m-2) fue aplicado a la mitad de los frutos de cada
grupo. Posteriormente, fueron almacenados a 5 y a 14 °C por 15 días junto con los
frutos testigo. Los frutos se muestrearon a intervalos de 5 días para evaluar el
sistema antioxidante.
En este ensayo se observó un aumento en el contenido de flavonoides en la
cutícula del fruto, como una respuesta de defensa contra el estrés oxidativo frente al
almacenamiento a bajas temperaturas (5 ° C).

xv
Por otra parte, el tratamiento de UV-C provocó un aumento significativo (P≤ 0.05)
aunque transitorio en el contenido de flavonoides totales en la cutícula de papaya
comparado con los frutos testigo; mientras que los niveles de fenoles totales no se
vieron afectados significativamente (P> 0.05).
En cuanto al sistema enzimático antioxidante, aunque se observó un aumento
significativo de la aplicación de UV-C en la actividad de SOD, éste fue dependiente
del tiempo y temperatura de almacenamiento. Mientras que el tratamiento de UV-C
solo o en combinación con las bajas temperaturas provocaron un incremento en la
actividad de la enzima CAT, lo cual nos sugiere que esta enzima podría estar
jugando un papel muy importante en el sistema de defensa de los frutos de papaya
contra diferentes tipos de estrés.
La actividad de POD fue inhibida por el almacenamiento a 5 °C y no se observaron
diferencias con respecto a las muestras tratadas con UV-C. En cambio, al aplicar el
tratamiento en frutos almacenados a 14 °C, la activ idad de POD se redujo
significativamente, lo cual coincide con un retraso en la maduración de los frutos
descrito en el Capítulo III.
En cuanto a la capacidad antioxidante, ésta fue mayor en la cutícula de frutos
almacenados a 5 °C, sin encontrarse diferencias sig nificativas entre los tratamientos
evaluados. Los frutos tratados con UV-C y almacenados a 14 °C presentaron un
aumento significativo en la capacidad antioxidante, pero sólo hacia el final del
almacenamiento.
Como pudimos corroborar, en este estudio, aunque la activación de la ruta de
síntesis de compuestos fenólicos por UV-C, ha sido ampliamente reportada en

xvi
tejidos vegetales, sus efectos varían en función del tipo de producto, de la
intensidad del tratamiento y condiciones de almacenamiento. Considerando que la
respuesta de defensa del sistema antioxidante de papaya es muy similar cuando el
fruto es almacenado en frío y cuando es irradiado con UV-C, se podría argumentar
que el estrés oxidativo inducido bajo estas condiciones y que activa el sistema
antioxidante, no es suficiente para reparar el daño generado, por lo que no se llega
a producir el efecto comúnmente conocido como hórmesis.

xvii
CONCLUSIONES GENERALES
La integración de los resultados de esta tesis permitió rechazar la hipótesis
propuesta. A pesar de que la mayoría de los compuestos fenólicos y carotenoides
identificados en el Capítulo II, poseen una alta actividad antioxidante. La
concentración más alta de fenoles fue encontrada en la cutícula del fruto, sin verse
afectada por el almacenamiento a baja temperatura, favoreciendo así la aplicación
del tratamiento de UV-C que es de baja penetración en el tejido. Sin embargo, al
evaluar el efecto de UV-C sobre los cambios en la calidad del fruto y su contribución
a la reducción del daño por frío (Capítulo III), se encontró que no fue efectivo en
este sentido, a pesar de reducir el deterioro y la pérdida de peso y retrasar la
maduración de los frutos. Finalmente, la respuesta del sistema antioxidante de
papaya ante UV-C, coincide con el efecto generado por las bajas temperaturas
(Capítulo IV). La respuesta de defensa más importante activada por UV-C es la
inducción de la síntesis de flavonoides. La aplicación de la irradiación UV-C como
tratamiento postcosecha en papaya podría ayudar a retrasar la maduración, reducir
el deterioro fúngico y aumentar el contenido de flavonoides y capacidad antioxidante
en la cutícula del fruto bajo ciertas condiciones de almacenamiento. Sin embargo, la
activación de estos mecanismos de respuesta al estrés, no es suficiente para lograr
mitigar el desarrollo de los síntomas de daño por frío, durante el almacenamiento a
bajas temperaturas.

CAPÍTULO I
Efectos Bioquímicos Postcosecha de la Irradiación UV-C en Frutos y Hortalizas

2

3

4

5

6

7

8
9
10

11

12

13

14

CAPÍTULO II Perfiles de fenoles y carotenoides de papaya (Carica papaya L.) y su contenido a bajas temperaturas de almacenamiento

16

17

18

19

20

21

22

23

24

25

26

27

28

29

30

31

32

33

34

35

36

37

38

39
40

41

42
43
44

45

46
47

48

CAPÍTULO III Efecto de la irradiación UV-C y temperatura de almacenamiento en la calidad postcosecha de papay a ‘Maradol’

50
EFECTO DE IRRADIACIÓN UV-C Y TEMPERATURA DE ALMACEN AMIENTO
EN LA CALIDAD POSTCOSECHA DE PAPAYA ‘MARADOL’
D. M. Rivera-Pastrana1; G. A. González-Aguilar1¶; M. Rivera-Domínguez; M. A.
Martínez-Téllez.
1Coordinación de Tecnología de Alimentos de Origen Vegetal, Centro de
Investigación en Alimentación y Desarrollo. Km. 0.6 Carretera a la Victoria.
Hermosillo, Sonora, C.P. 8300. México. Correo-e: [email protected]. (Autor
responsable) ¶
Autor responsable:
Gustavo A. González-Aguilar.
Teléfono: 00-52-6622-892400, Ext. 272.
Fax: 00-52-6622-80-0422.
Email: [email protected]

51
RESUMEN
Frutos de papaya var. ‘Maradol’ en estado verde-maduro, fueron expuestos a una
dosis de irradiación UV-C de 1.76 kJ·m-2, previamente al almacenamiento a 1, 5, 14
y 25 °C por 12 días. El efecto de los tratamientos fue evaluado después de transferir
los frutos a 25 °C por 2 días. El tratamiento de U V-C redujo el deterioro causado
por hongos sin afectar significativamente la firmeza de los frutos a las temperaturas
de almacenamiento analizadas. Sin embargo no fue efectivo para reducir los
síntomas de daño por frío de papaya, favoreciendo el oscurecimiento en los frutos
almacenados a temperaturas inferiores a 25 °C. Los parámetros L*, Hue°, Chroma*,
pérdida de peso (%) y deterioro general fueron disminuidos significativamente por
las bajas temperaturas de almacenamiento y el tratamiento de irradiación UV-C,
sugiriendo un retraso en la maduración o senescencia de los frutos. La irradiación
UV-C puede utilizarse para retrasar el deterioro fúngico y maduración de papaya
‘Maradol’ a temperaturas templadas (25 °C), aunque no disminuye el daño por frío
en este fruto.
PALABRAS CLAVE ADICIONALES : Carica papaya L., irradiación, deterioro, daño
por frío.
UV-C IRRADIATION EFFECT ON POSTHARVEST QUALITY OF ‘ MARADOL’
PAPAYA AT LOW STORAGE TEMPERATURES.
ABSTRACT
Green-mature ‘Maradol’ papayas were exposed to a UV-C irradiation dose of 1.76
kJ·m-2 before storage at 1, 5, 14 and 25 °C for 12 days . Treatment effects were
evaluated after transferring fruits at 25 °C for 2 d. UV-C treatment reduced general
decay related mainly to fungal attack, without significantly affecting fruit firmness at
analized storage temperatures. However, it was not effective to reduce chilling injury
symtoms in papaya fruit, promoting increased browning in fruits stored at

52
temperatures below 25 °C. L*, Hue°, Chroma*, weigh t loss (%) and general decay
parameters were significantly reduced by low storage temperatures and UV-C
treatment, suggesting a ripeness or senescence delay. UV-C irradiation could be
applied to delay fungal decay and ripening of ‘Maradol’ papaya at 25 °C, although it
doesn’t reduce chilling injury on this fruit.
ADDITIONAL KEY WORDS: Carica papaya L., Irradiation, decay, chilling injury.
INTRODUCCIÓN
En los últimos años, se ha incrementado la producción y exportación de papaya por
el creciente interés del consumidor a nivel mundial. México, es el quinto mayor
productor de papaya después de India, Brasil, Nigeria e Indonesia; y el mayor
exportador en el mundo (2009). Sin embargo, la calidad postcosecha de la papaya
es afectada por diversos factores como: condiciones ambientales, tratamientos
postcosecha, daños físicos, deterioro fúngico y daños por frío, (Proulx et al., 2005).
Los síntomas de daño por frío en papaya se presentan como picado y escaldado
de la cáscara, endurecimiento de la piel y maduración anormal con decoloración de
la pulpa, así como una mayor susceptibilidad al deterioro. Los frutos de papaya en
estado verde-maduro son más susceptibles al daño por frío que los frutos maduros
(Chen y Paull, 1986).
Para frutos cosechados en madurez fisiológica se ha reportado un amplio rango de
temperaturas de almacenamiento refrigerado (10-16 °C) en diferentes cultivares de
papaya. Entre las tecnologías postcosecha eficientes en la reducción del daño por
frío y preservación de la calidad de frutos de papaya se encuentran: los tratamientos
térmicos y aplicación de fungicidas (Perez-Carrillo y Yahia, 2004), tratamientos con
metil jasmonato y atmósferas modificadas (González-Aguilar et al., 2003), aplicación
de 1-metilciclopropeno (1-MCP) (Manenoi et al., 2007) e irradiación gamma
(Thomas y Moy, 1986).

53
La irradiación no ionizante UV-C se ha aplicado extensivamente para reducir el
deterioro microbiológico en frutas, ya que presenta un amplio efecto germicida
(Guerrero-Beltran y Barbosa-Cánovas, 2004). Otros efectos indirectos de su
aplicación en postcosecha son la activación de la ruta de síntesis de fitoalexinas,
aumento en la capacidad antioxidante y reducción de daño por frío en pimientos,
mangos y duraznos (Gonzalez-Aguilar et al., 2007, Gonzalez-Aguilar et al., 2004,
Gonzalez-Aguilar et al., 2001, Vicente et al., 2005a). Por lo que la aplicación de
irradiación UV-C podría contribuir a mejorar la calidad postcosecha de frutos de
papaya Maradol y a reducir el daño por frío ocasionado durante el almacenamiento
a bajas temperaturas. El objetivo de éste trabajo fue evaluar el efecto del
tratamiento de UV-C en la calidad postcosecha de papaya medida por cambios en
el color, pérdida de peso y firmeza, sólidos solubles totales, apariencia general y
daño por frío.
MATERIALES Y MÉTODOS
Frutos de papaya var. ‘Maradol’ fueron obtenidos de un campo comercial en
Tecomán, Colima, y transportados 48-72 horas después de cosecha a los
laboratorios del CIAD en en Hermosillo, Son., México. Los frutos se clasificaron y
seleccionaron en estado verde-maduro (<10 % de superficie amarilla), sin defectos
visibles y de tamaño uniforme. El experimental fue realizado en dos temporadas
(marzo-abril y octubre-noviembre 2009). Las muestras se separaron en 4 lotes de
24 frutos al azar, para cada temperatura de almacenamiento. La mitad de los frutos
de cada lote fueron tratados con una dosis de 1.76 kJ·m-2 de acuerdo a
preliminares, con lámparas GE de 15 W modelo G15T8 que emiten 95 % de
irradiación en el rango UV-C (100-280 NM), las cuales fueron situadas 15 cm arriba
y abajo de los frutos dentro de una campana de extracción. Las muestras fueron
rotadas 90 ° a la mitad del tratamiento para asegur ar una exposición homogénea de
todo el fruto. Como medida de seguridad se utilizaron durante la aplicación del
tratamiento, guantes, lentes y careta protectora. Después del tratamiento los

54
distintos grupos conteniendo frutos tratados y testigo se almacenaron por 12 días en
cajas de cartón corrugado a 1, 5, 14 y 25 ° ± 2 °C , y 80 ± 10 % HR. Posteriormente
se llevaron a cabo las mediciones de los parámetros de calidad en los frutos
almacenados a 1, 5 y 14 ° C y transferidos por 2 dí as a 25 °C. Se utilizaron seis
frutos por tratamiento para medir las variables de cambio en color, peso, firmeza,
sólidos solubles totales, deterioro de apariencia general y daño por frío para cada
temperatura.
Al inicio y al final del almacenamiento se midió el color de la cáscara de los frutos
en 3 puntos equidistantes de la superficie con un colorímetro Minolta CR-300
(Japón). Se obtuvieron parámetros de color L* (luminosidad), a*(color verde a rojo)
y b* (color azul a amarillo) de la escala CIELAB (Mcguire, 1992), y se calcularon el
ángulo de matiz (Hue°) y la saturación del color (Chroma) con las siguientes
fórmulas: Chroma = (a*2 + b*2)1/2, Hue° = arctang (b* / a*). Los datos se expresaro n
como el cambio de cada parámetro entre el valor inicial y final del almacenamiento:
∆L*, ∆Hue, ∆Chroma.
Para determinar la pérdida de peso se utilizó una balanza analítica digital modelo
PR 2003 Delta Range con capacidad de 2,100 g y aproximación de 0.01 g (Mettler-
Toledo, Greinfesee, Suiza); El porcentaje de pérdida de peso se obtuvo a partir de
la diferencia del peso inicial y final y su relación con el peso inicial.
La firmeza se determinó por el método de punción con un penetrómetro digital
Chatillón modelo DFM-50 con punzón cilíndrico de 0.8 cm de diámetro. En la parte
ecuatorial de cada fruto se retiraron 3 mm de la cáscara y se realizaron 3
mediciones equidistantes. Se registró la fuerza de oposición del tejido a la
penetración del punzón a 1 cm de profundidad expresada en Newtons (N) y se
calculó el porcentaje de pérdida de firmeza (% PF) hacia el final del
almacenamiento como: % PF= (firmeza inicial- firmeza final/firmeza inicial)*100.
Los sólidos solubles totales (SST) fueron analizados con un refractómetro digital
ATAGO modelo PR-101 (A.O.A.C., 1990) y los resultados en ° Brix fueron
expresados como cambio en el contenido de SST: ∆SST= SST final- SST inicial.
El grado de deterioro de la superficie de los frutos fue evaluado subjetivamente
usando una escala de 0 a 5 donde 0= 0 %, 1 = 20-30 %, 2 = 40-50 %, 3 =60-70 %,

55
4 = 80-90 % y 5 = 90-100 %. Dentro de la escala se consideraron signos de daño
físico, escaldado, ablandamiento excesivo, desarrollo de crecimiento fúngico y
oscurecimiento.
El grado de daño por frío se determinó de acuerdo a la escala subjetiva de 1-5
donde 1= no presenta, 2 = leve, 3 = moderado, 4 = severo y 5 = extremo (Perez-
Carrillo y Yahia, 2004); se calculó el índice de daño por frío (IDF) para cada
subgrupo de muestras por la sumatoria de los productos del número de frutos
dañados (n) y el grado de daño, entre el total de muestras (N) (Martínez-Téllez et
al., 2008) de acuerdo con la siguiente fórmula: IDF= (n(1) + n(2) + n(3) + n(4) +
n(5))/N.
El análisis de los datos se llevó a cabo bajo un diseño completamente al azar
mediante un análisis de varianza (ANOVA) y una prueba de Tukey-Kramer para la
comparación de medias (P ≤ 0.05) utilizando el paquete estadístico NCSS (2007).
RESULTADOS Y DISCUSIÓN
Cambios en el color
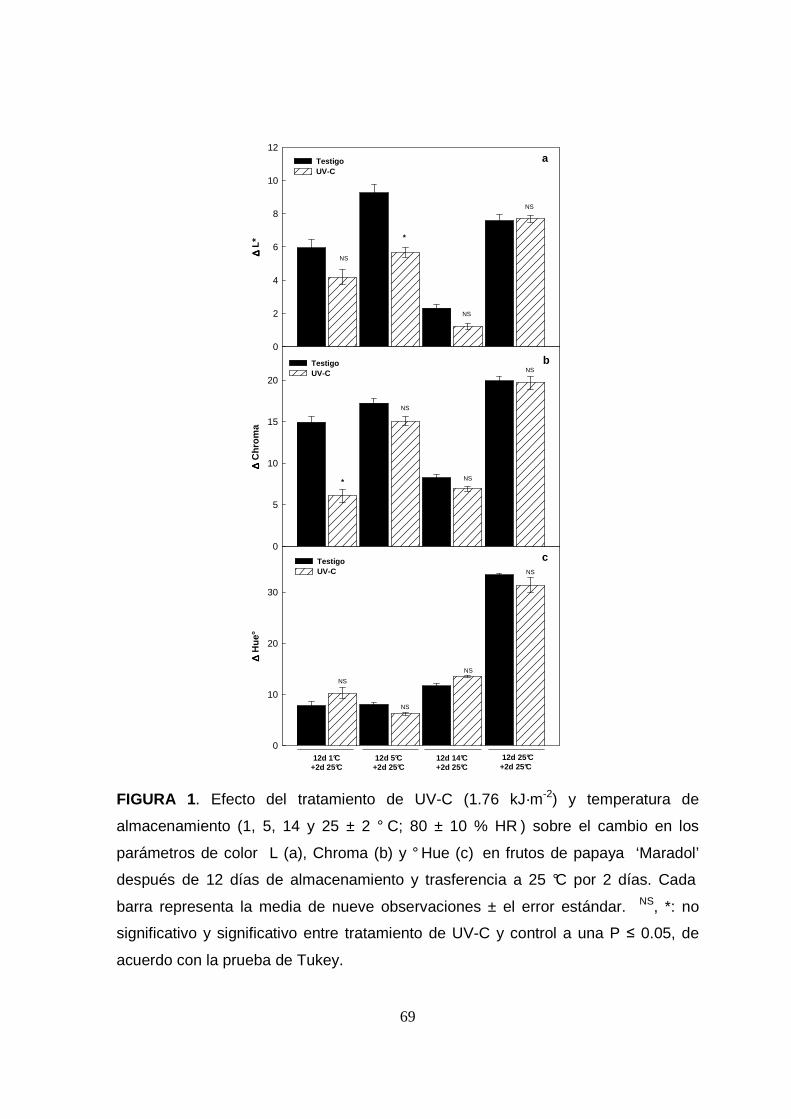
El tratamiento de UV-C redujo significativamente (P<0.05) la luminosidad en frutos
de papaya, resultando ineficiente en el control de daño por frío, observado como
obscurecimiento de la cáscara del fruto con un cambio negativo promedio en L*
(Sapers y Miller, 1995). La temperatura de almacenamiento también redujo
significativamente el cambio en luminosidad durante el almacenamiento (∆L*) a 1 y
14 °C (Figura 1a); lo cual podría relacionarse con una maduración retrasada en
respuesta al daño por frío, ya que el color de los frutos afectados no alcanza valores
de un fruto completamente maduro, aún al ser transferidos a una temperatura de 25
°C por 2 días, de acuerdo con lo reportado por (La m, 1990). Por otra parte, se ha
observado que a una dosis de 3.7 kJ·m-2 de UV-C, se presenta una menor

56
acumulación de cera epicuticular en frutos de tomate relacionada con daño a
células del epicarpio, lo que ocasiona una reducción del brillo así como cambios en
la reflectancia (Charles et al., 2008c). Estos cambios podrían contribuir a la
disminución de luminosidad en papayas tratadas con UV-C.
En la intensidad del color (Chroma*), se observó una reducción significativa
(P<0.05) por el tratamiento de UV-C y las temperaturas de almacenamiento 1 y 14
°C (Figura 1b). Mientras que en las muestras almac enadas a 5 y 25 °C no se
observaron diferencias en el cambio de intensidad del color, por lo que el efecto de
UV-C en papaya fue dependiente de la temperatura. Un menor cambio de Chroma*
durante el almacenamiento, concuerda con un retraso en la maduración por la
exposición a bajas temperaturas (Proulx et al., 2005).
La temperatura de almacenamiento también afectó significativamente (P<0.05)
ángulo de matiz (°Hue) en papaya, el cual disminuy e pasando de verde-amarillo a
naranja conforme avanza el proceso de maduración (Figura 1c). En este estudio, a
temperaturas inferiores a 25 °C, observamos un meno r cambio de °Hue ( ∆°Hue),
que también se atribuye a una maduración retrasada de los frutos.
A pesar de que los valores iniciales promedio de L*, a* y b*, de las muestras
analizadas en este trabajo corresponden con el estado de madurez fisiológica o
grado 1 de los índices de madurez propuestos en trabajos previos para papaya
‘Maradol’(Basulto et al., 2009), los frutos almacenados a 14, 5 y 1 °C no m aduraron
completamente una vez transferidos a la temperatura control (25 °C). Esta
observación hace evidente la dificultad de uniformizar la respuesta de este fruto
ante algunas condiciones de manejo postcosecha o tratamientos que basan su
efectividad en un estado de madurez particular como es el caso de la irradiación
UV-C.
Pérdida de peso y firmeza
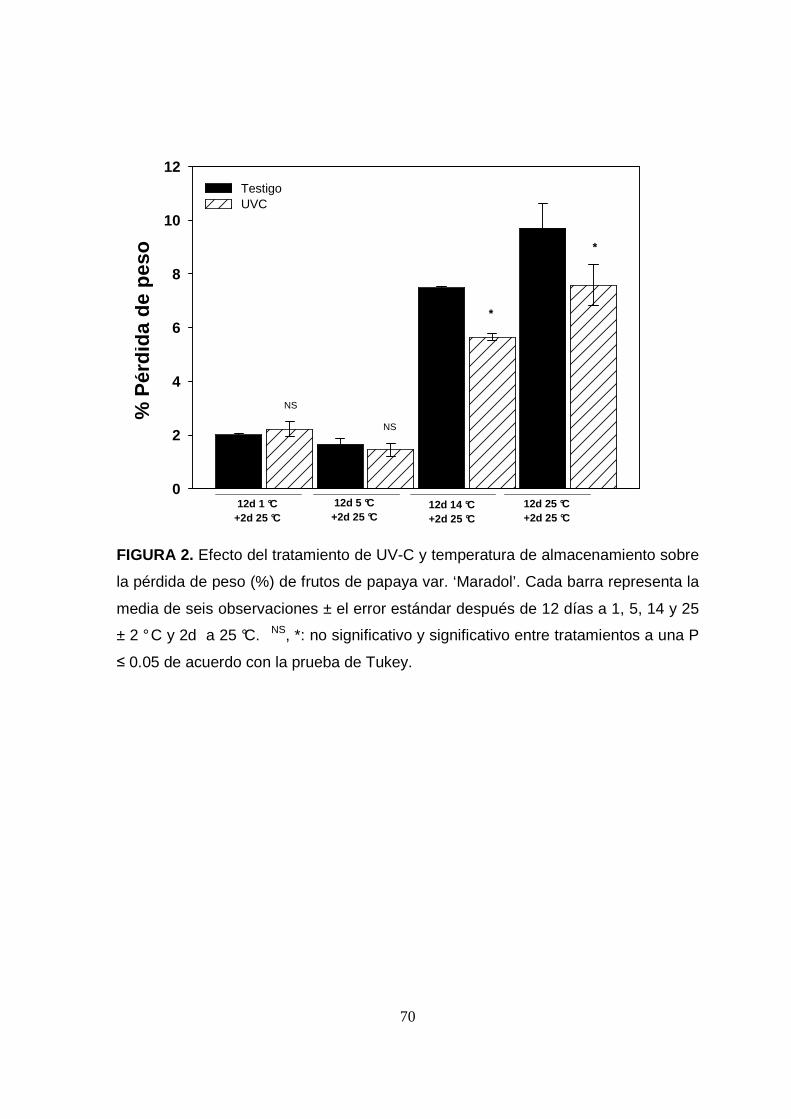
La pérdida de peso de los frutos testigo y tratados con UV-C fue afectada
significativamente por la temperatura de almacenamiento. El mayor porcentaje de

57
pérdida de peso (%PP) se observó en los frutos testigo almacenados a 25 °C por 12
días. Las papayas almacenadas a 14 °C perdieron ent re el 6 y el 7 %, mientras que
a 5 y 1 °C perdieron tan sólo entre el 1.5 y 2 % en peso, respectivamente (Figura 2).
El tratamiento de UV-C redujo significativamente (P≤0.05) la pérdida de peso en los
frutos almacenados a 14 y 25 °C, pero no así, en el resto de las temperaturas
evaluadas. (Paull y Chen, 1989) reportaron que una pérdida de peso del 8 % en
papayas en madurez fisiológica de las variedades ‘Sunset’ y ‘Sunrise’, ocasiona una
textura gomosa, pérdida de brillo y marchitamiento leve de la cáscara, los cuales
afectan negativamente la calidad del fruto. Por otra parte (Proulx et al., 2005)
observaron una pérdida de peso del 6 % en papayas de la variedad Exp.15,
almacenadas 6 días a 20 °C; y de 3 % después de 14 días a 0 °C (82-97 % HR).
Mientras que (Maharaj y Sankat, 1990) reportaron una pérdida de peso alrededor
del 7 % en papayas ‘Tainung' No.1 después de 14 días a 16 °C. La pérdida de
peso en los frutos de papaya es una de las principales causas de pérdidas
postcosecha; ya varía en función de la temperatura, la humedad relativa, la
madurez del fruto al momento de la cosecha, la variabilidad entre cultivares, y
diferencias entre temporadas de recolección (Nunes et al., 2006).
Se ha sugerido que la vía principal para la pérdida de peso en frutos de papaya es
la pérdida de agua a través de la cicatriz del pedúnculo, estomas y la cutícula (Paull
y Chen, 1989); y que el grosor de la cutícula disminuye después de alcanzar una
madurez de consumo (más del 50 % amarillo). Por lo que es posible que los frutos
testigo almacenados a 25 °C presentaron la mayor pé rdida de peso por encontrarse
en un estado más avanzado de madurez que el resto de los tratamientos.
La disminución de pérdida de peso por el tratamiento de UV-C observada a 14 °C
podría atribuirse al retraso de la maduración; efecto que se ha observado en otros
frutos y hortalizas como retrasos en la degradación de clorofila, en cambios en el
color, en ablandamiento del tejido y en el climaterio (Baka et al., 1999, Costa et al.,
2006, Maharaj et al., 1999, Stevens et al., 1998). Aunque algunos autores han
reportado un incremento en la pérdida de peso en tomate por la aplicación de UV-C,
también señalan que este tratamiento propicia cambios bioquímicos que llevan a un
incremento en la síntesis de compuestos fenólicos complejos como ligninas y

58
suberinas implicados en la formación de una barrera impermeable capaz de
prevenir la pérdida de agua y nutrientes (Charles et al., 2008b, Maharaj et al., 1999).
En este caso, aún cuando no existe un mecanismo propuesto por el cual el
tratamiento de UV-C reduce el %PP en papaya, éste efecto podría deberse a un
fortalecimiento de pared celular en las capas epicuticulares como resultado de una
lignificación, o por un retraso en la disminución del grosor del epicarpio relacionado
con un retraso en la senescencia de los frutos.
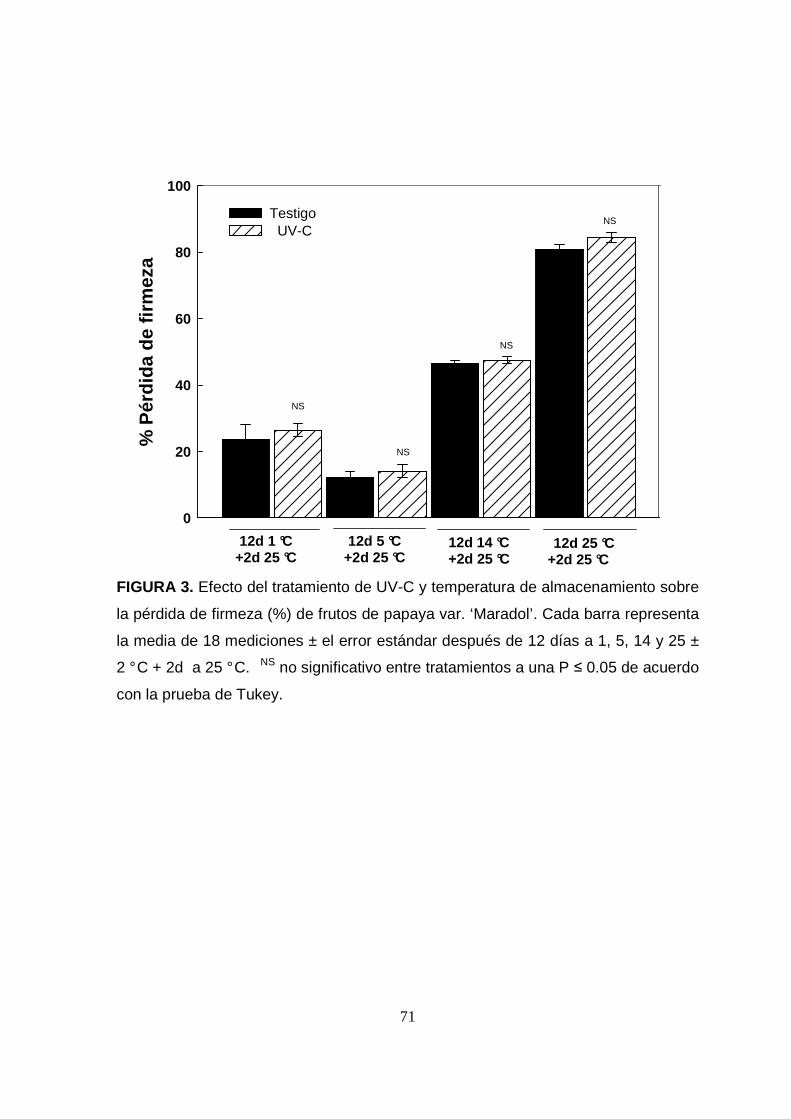
El porcentaje de pérdida de firmeza (%PF) fue afectado significativamente (P<0.05)
por la temperatura de almacenamiento. El %PF fue más alto en los frutos
almacenados a 14 y 25 °C (50 y 80 % respectivamente ) independientemente del
tratamiento (Figura 3). Mientras que en las papayas almacenadas a 1 y 5 °C por
12 días se observaron pérdidas de firmeza entre el 12 y 26 %, respectivamente.
Estos resultados coinciden con los reportados previamente para la misma variedad
de papaya, donde se observó una pérdida de firmeza del 90 % entre frutos en
estado verde-maduro y los estadíos 4-6 de madurez avanzada, almacenados a 23
°C y 70 % HR (Basulto et al., 2009).
En este trabajo no se encontró ningún efecto del tratamiento de UV-C en la pérdida
de firmeza. En un estudio con frutos de mango cv. ‘Tommy Atkins’ almacenados a
5 °C, se observó que el tratamiento de UV-C, dismin uyó significativamente el
ablandamiento aún después de transferir los frutos por 7 días a 20 °C (Gonzalez-
Aguilar et al., 2001). De la misma manera, dosis de 1.3-40 kJ m-2 y 0.8-40 x 10-3 kJ
m-2 de UV-C en tomate y duraznos respectivamente, resultaron en retraso de
maduración y disminución de ablandamiento de los frutos (Liu et al., 1993, Stevens
et al., 1996) En este sentido, (Barka et al., 2000) reportaron que la exposición a una
dosis hórmica de UV-C de frutos de tomate reduce la degradación enzimática de la
pared celular; por lo que las enzimas involucradas en este proceso:
poligalacturonasa, pectin metilesterasa, celulasa, xilanasa y β-galactosidasa,
pueden ser blanco de la irradiación UV-C induciendo proteólisis o inhibición de su
síntesis de novo. Recientemente, se ha asociado el retraso del ablandamiento en
fresas var. ‘Aroma’ inducido por el tratamiento de UV-C, con una disminución en la
transcripción de genes que codifican proteínas de pared celular y de enzimas

59
involucradas en la degradación de la misma (Pombo et al., 2009). En frutos de
papaya variedad ‘Maradol’, se ha sugerido que la actividad de las enzimas
poligalacturonasa y β-galactosidasa son las principales causantes del
ablandamiento del fruto durante la maduración (Barajas et al., 2008). La mayor
actividad de poligacturonasa (PG) en frutos de papaya se encuentra en el tejido
adyacente a la placenta seguido por el mesocarpio y exocarpio en ese orden en
frutos completamente maduros. El decremento gradual en la actividad de PG del
centro del fruto hacia la cáscara coincide con la dirección en que el fruto madura, es
decir de adentro hacia fuera (Chan et al., 1981). Por lo que la baja actividad de PG
en la parte más externa del fruto de papaya en un estado menos avanzado de
madurez, puede explicar que el tratamiento de UV-C con baja penetración en el
tejido no afectara significativamente la firmeza de fruto (Gardner y Shama, 2000).
Sólidos solubles totales
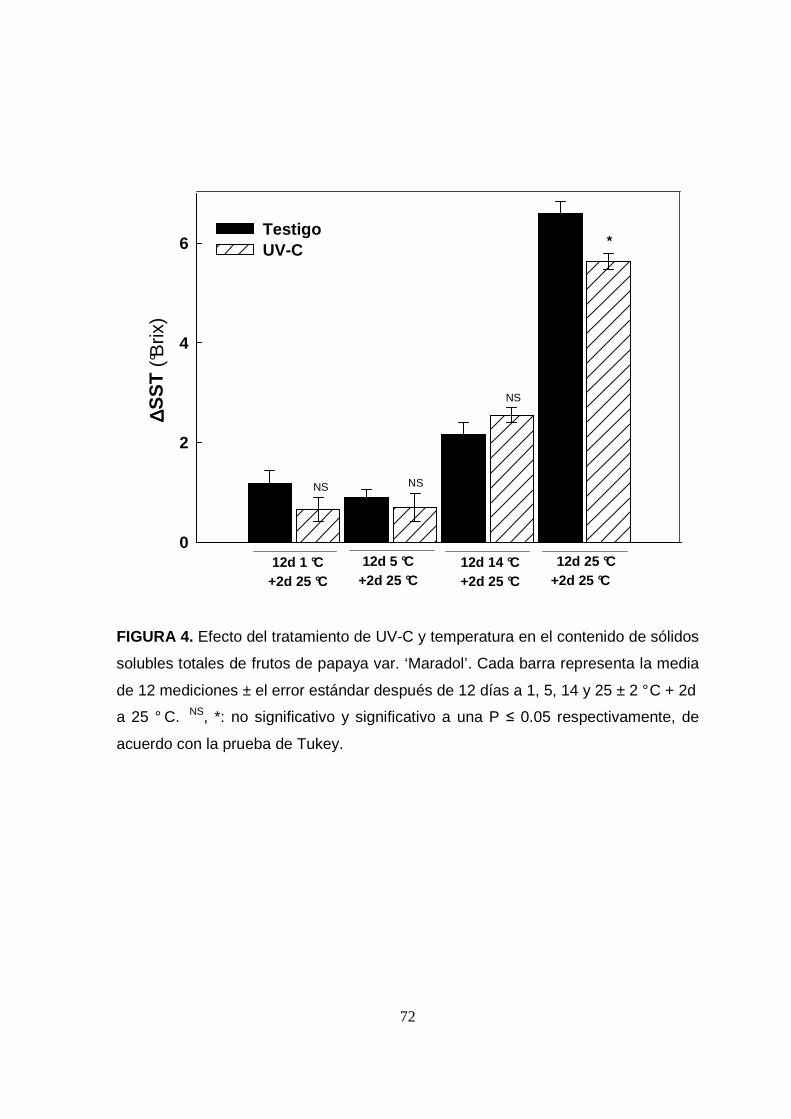
La figura 4 muestra el efecto del tratamiento de UV-C y temperatura en el contenido
de sólidos solubles totales (SST). La temperatura de almacenamiento afectó
significativamente (P<0.05) el contenido de SST a todos los niveles, mientras que el
tratamiento de UV-C fue significativo únicamente para la temperatura de 25 °C. El
contenido inicial de SST fue de 7.1 ± 0.5 ° Brix que coincide con el rango reportado
por (Basulto et al., 2009) para el estadío G (verde) y 1 (verde-maduro) en papaya
‘Maradol’. Sólo las muestras almacenadas a 25 °C al canzaron un contenido de SST
de un fruto de esta variedad completamente maduro (11-12.5 ° Brix). Mientras que a
14, 5 y 1 °C los frutos no maduraron por completo a ún después de transferirlos a 25
°C por 2 días; situándose entre los estados 2 y 4 s ugeridos para esta variedad
(Basulto et al., 2009). El bajo contenido de SST de las muestras al inicio del
experimento, sugiere que algunos de los frutos no se encontraban aún en la
madurez fisiológica, por lo que no alcanzaron la madurez comercial después de ser
expuestos a temperaturas de almacenamiento inferiores a 25 °C ± 2 °C. Los
resultados obtenidos en el presente estudio contrastan con trabajos previos, donde

60
se describe que frutos de papaya almacenados hasta por 14 días a temperaturas
inferiores a los 7 °C maduraron normalmente una vez transferidos a temperatura
ambiente (Chen y Paull, 1986), sin embargo se sugiere que las diferencias en
susceptibilidad al daño por frío están en función de la variedad.
En otros frutos el efecto de UV-C sobre el contenido de SST es contradictorio. Se
ha visto que el tratamiento de UV-C puede disminuir el contenido de SST como
resultado de un retraso en la maduración o senescencia en frutos de manzana var.
‘Red Delicious’, durazno var. ‘Loring’ y ‘Elberta’, en boysenberries y en frutos de ber
(Ziziphus Mauritania L.); o no afectarlo de manera significativa en frutos de tomate,
mango y manzana var. ‘Golden Delicious’ (Gonzalez-Aguilar et al., 2001, Hemmaty
et al., 2007, Lu, Jy et al., 2007a, Purohit et al., 2003, Vicente et al., 2004). La
temperatura de almacenamiento resultó ser una variable crítica en este estudio para
el efecto del tratamiento de UV-C, ya que se observó que los frutos tratados
tuvieron menores valores de SST que los frutos testigo, pero fueron
significativamente menores sólo a 25 °C. Nuestros r esultados coinciden con
reportes previos donde se describe que el efecto del tratamiento de UV-C sobre el
contenido de SST es inconsistente y dependiente del producto y condiciones de
almacenamiento, principalmente de la temperatura (Hemmaty et al., 2007, Lu et al.,
2007b).
Deterioro general y Daño por frío
El tratamiento de UV-C redujo significativamente (P< 0.05) el deterioro del fruto a
todos los niveles de temperatura evaluados (Figura 5). Los principales signos de
deterioro que presentó la papaya almacenada a 5 y 1 °C, fueron el escaldado y
oscurecimiento de la cáscara, síntomas que se pueden atribuir al daño por frío
(Chen y Paull, 1986). Por otra parte en frutos almacenados a 14 y 25 °C, las
principales causas de deterioro fueron el ablandamiento excesivo y crecimiento
fúngico respectivamente, que podrían estar relacionadas con una madurez
avanzada de los frutos y una mayor actividad de enzimas de degradación de pared

61
celular bajo estas condiciones de temperatura. La aplicación de UV-C retrasa la
senescencia y reduce el deterioro ocasionado por fitopatógenos y daño por frío en
otros frutos; al activar diferentes respuestas de defensa y sistema antioxidante que
involucran un aumento en la síntesis de compuestos antioxidantes, fitoalexinas,
poliaminas, inactivación de enzimas de degradación de pared celular, inducción de
una respuesta hipersensible y reforzamiento de la pared celular (Baka et al., 1999,
Barka et al., 2000, Charles et al., 2008c, Charles et al., 2008d, D'hallewin et al.,
1999, De Capdeville et al., 2002, Gonzalez-Aguilar et al., 2001, Maharaj et al., 1999)
Sin embargo, (Cia et al., 2007), reportaron que las dosis de 0.8 y 1.3 kJ·m-2 son
muy efectivas para inhibir la germinación de conidios y crecimiento del micelio de
Colletotrichum gloeosporiodes en ensayos in vitro; mientras que al aplicar el mismo
tratamiento en frutos de papaya ‘Golden’ previamente inoculados por inyección
subcuticular y almacenados a 25 °C, no se observó u n efecto inhibitorio del
desarrollo del hongo. En este contexto, en papaya Maradol, observamos que el
efecto de UV-C en el deterioro parece estar relacionado principalmente con la
reducción del desarrollo fúngico, el cual a su vez que puede estar influenciado por el
grado de madurez de los frutos tratados descrito en secciones anteriores.
El efecto del tratamiento de UV-C no fue significativo sobre el daño por frío (DF) en
frutos de papaya ‘Maradol’ (Figura 5). Sin embargo, en función de las temperaturas
de almacenamiento 1 y 14 °C, se observó una disminu ción por efecto del
tratamiento la cual fue significativa a 1 °C, mient ras que a 5 °C no se presentaron
diferencias entre frutos tratados y testigos. En este estudio, los frutos presentaron
síntomas de DF de leves a moderados de acuerdo con (Proulx et al., 2005) cuando
se almacenaron 12 días a 14, 5 y 1 °C. El grado de DF fue inversamente
proporcional a la temperatura de almacenamiento; acercándose al límite de calidad
comercializable al exponerlos a 1 °C, de acuerdo co n lo reportado para frutos de
papaya que presentan síntomas moderados de daño por frío (Nunes et al., 2006,
Proulx et al., 2005).
En estudios previos, la irradiación UV-C, se ha reportado eficiente en la reducción
de síntomas de DF en durazno, mango y pimiento rojo (Gonzalez-Aguilar et al.,
2004, Gonzalez-Aguilar et al., 2001, Vicente et al., 2005a, Vicente et al., 2005b);

62
correlacionando este efecto benéfico del tratamiento con la inducción del sistema
antioxidante del fruto, aumentando la actividad de enzimas involucradas en la
síntesis de compuestos fenólicos y el contenido de poliaminas. Por otra parte, las
bajas temperaturas por sí solas, también pueden inducir un aumento en la actividad
de la enzima fenilalanina amonioliasa (PAL), clave en la síntesis de compuestos
fenólicos y la acumulación de poliaminas (Gonzalez-Aguilar et al., 2001, Sanchez-
Ballesta et al., 2000).
La exposición de los tejidos vegetales tanto al tratamiento de UV-C como a bajas
temperaturas de almacenamiento puede aumentar el estrés oxidativo por
acumulación de especies reactivas de oxígeno (Frohnmeyer y Staiger, 2003)
(Karpinski et al., 2002). En este contexto, se ha descrito una mayor susceptibilidad
al ataque de B. cinerea en tomate inmediatamente después de aplicado el
tratamiento de UV-C que atribuyen a un superposición de estreses oxidativos
ocasionados por la dosis hórmica del tratamiento y la infección con el fitopatógeno
(Charles et al.,2008a). De la misma forma, ambos factores, UV-C y almacenamiento
a bajas temperaturas, pueden ocasionar un aumento combinado en el estrés
oxidativo del fruto de papaya, bajo el cual no se presenta un efecto benéfico del
tratamiento sobre el daño por frío.
CONCLUSIONES
El tratamiento de UV-C en frutos de papaya ‘Maradol’ redujo el deterioro general
relacionado principalmente con el ataque de hongos; sin afectar significativamente
la pérdida de firmeza de los frutos. Sin embargo, no se observó un efecto positivo
del tratamiento para contrarrestar el daño por frío ocasionado por el
almacenamiento a bajas temperaturas. Se concluye que el tratamiento de UV-C no
fue efectivo para mantener la calidad postcosecha de papaya ‘Maradol’
almacenada a bajas temperaturas posiblemente por una superposición de elicitores
del estrés oxidativo. Futuros estudios pueden enfocarse a evaluar el efecto del

63
tratamiento en combinación con otras tecnologías que reduzcan el daño por frío
para alargar la vida postcosecha de la papaya.
AGRADECIMIENTOS
Al financiamiento CONACYT, al apoyo técnico de Mónica A. Villegas Ochoa,
Chrystian M. Rodríguez Armenta y Manuel R. Cruz Valenzuela.
LITERATURA CITADA
A.O.A.C. 1990. Official Methods of Analysis. 15th. Association of Official Analytical
Chemists Inc. Arlington, VA., USA. 1006.
ANÓNIMO 2009. Production Yearbook. United Nations Food and Agriculture
Organization (FAO). Rome.
BAKA, M., MERCIER, J., CORCUFF, R., CASTAIGNE, F., ARUL, J. 1999.
Photochemical treatment to improve storability of fresh strawberries. Journal of Food
Science 64: 1068-1072.
BARAJAS, J., CEPEDA, J., ENCISO, T., RANGEL, D., ÁLVAREZ, G., LABAVITCH,
J. 2008. Control de la maduración en frutos de papaya (Carica papaya L.) con1-
metilciclopropeno y ácido 2-cloroetil sulfónico. Revista Fitotecnia Mexicana 31: 141-
147.
BARKA, E., KALANTARI, S., MAKHLOUF, J., ARUL, J. 2000. Impact of UV-C
irradiation on the cell wall-degrading enzymes during ripening of tomato
(Lycopersicon esculentum L.) fruit. J. Agric. Food Chem 48: 667-671.
BASULTO, F. S., SAURI, E. D., ESPADAS, F. G., DÍAZ, R. P., LARQUÉ, A. S.,
SANTAMARÍA, J. M. 2009. Postharvest ripening and maturity indices for Maradol
papaya. Interciencia 34: 583-588.

64
CIA, P., PASCHOLATI, S., BENATO, E., CAMILI, E., SANTOS, C. 2007. Effects of
gamma and UV-C irradiation on the postharvest control of papaya anthracnose.
Postharvest Biology and Technology 43: 366-373.
COSTA, L., VICENTE, A., CIVELLO, P., CHAVES, A., MARTÍNEZ, G. 2006. UV-C
treatment delays postharvest senescence in broccoli florets. Postharvest Biology
and Technology 39: 204-210.
CHAN, H. T., TAM, S. Y. T., SEO, S. T. 1981. Papaya polygalacturonase and its role
in thermally injury ripening fruit. Journal of Food Science 46: 190-191, 197.
CHARLES, M., BENHAMOU, N., ARUL, J. 2008a. Physiological basis of UV-C
induced resistance to Botrytis cinerea in tomato fruit:: III. Ultrastructural
modifications and their impact on fungal colonization. Postharvest Biology and
Technology 47: 27-40.
CHARLES, M., GOULET, A., ARUL, J. 2008b. Physiological basis of UV-C induced
resistance to Botrytis cinerea in tomato fruit - IV. Biochemical modification of
structural barriers. Postharvest Biology and Technology 47: 41-53.
CHARLES, M. T., MAKHLOUF, J., ARUL, J. 2008c. Physiological basis of UV-C
induced resistance to Botrytis cinerea in tomato fruit - II. Modification of fruit surface
and changes in fungal colonization. Postharvest Biology and Technology 47: 21-26.
CHARLES, M. T., MERCIER, J., MAKHLOUF, J., ARUL, J. 2008d. Physiological
basis of UV-C-induced resistance to Botrytis cinerea in tomato fruit - I. Role of pre-
and post-challenge accumulation of the phytoalexin-rishitin. Postharvest Biology and
Technology 47: 10-20.
CHEN, N., PAULL, R. 1986. Development and prevention of chilling injury in papaya
fruit. Journal of the American Society for Horticultural Science 111: 639-643.
D'HALLEWIN, G., SCHIRRA, M., MANUEDDU, E., PIGA, A., BEN-YEHOSHUA, S.
1999. Scoparone and scopoletin accumulation and ultraviolet-C induced resistance
to postharvest decay in oranges as influenced by harvest date. Journal of american
society for horticultural science 124: 702-707.
DE CAPDEVILLE, G., WILSON, C., BEER, S., AIST, J. 2002. Alternative disease
control agents induce resistance to blue mold in harvested'Red Delicious' apple fruit.
Phytopathology 92: 900-908.

65
FROHNMEYER, H., STAIGER, D. 2003. Ultraviolet-B radiation-mediated responses
in plants. Balancing damage and protection. Plant Physiology 133: 1420-1428.
GARDNER, D., SHAMA, G. 2000. Modeling UV-induced inactivation of
microorganisms on surfaces. Journal of Food Protection 63: 63-70.
GONZÁLEZ-AGUILAR, G. A., BUTA, J. G., WANG, C. Y. 2003. Methyl jasmonate
and modified atmosphere packaging (MAP) reduce decay and maintain postharvest
quality of papaya ‘Sunrise’. Postharvest Biology and Technology 28: 361-370.
GONZALEZ-AGUILAR, G. A., VILLEGAS-OCHOA, M. A., MARTINEZ-TELLEZ, M.
A., GARDEA, A. A., AYALA-ZAVALA, J. F. 2007. Improving antioxidant capacity of
fresh-cut mangoes treated with UV-C. Journal of Food Science 72: S197-S202.
GONZALEZ-AGUILAR, G. A., WANG, C. Y., BUTA, G. J. 2004. UV-C irradiation
reduces breakdown and chilling injury of peaches during cold storage. Journal of the
Science of Food and Agriculture 84: 415-422.
GONZALEZ-AGUILAR, G. A., WANG, C. Y., BUTA, J. G., KRIZEK, D. T. 2001. Use
of UV-C irradiation to prevent decay and maintain postharvest quality of ripe 'Tommy
Atkins' mangoes. International Journal of Food Science and Technology 36: 767-
773.
GUERRERO-BELTRAN, J. A., BARBOSA-CÁNOVAS, G. V. 2004. Advantages and
limitations on processing foods by UV light. Food Science and Technology
International 10: 137.
HEMMATY, S., MOALLEMI, N., NASERI, L. 2007. Effect of UV-C radiation and hot
water on the calcium content and postharvest quality of apples. Spanish journal of
agricultural research 5: 559-568.
KARPINSKI, S., WINGSLE, G., KARPINSKA, B., HÄLGREN, J.2002. Low
temperature stress and antioxidant defense mechanisms in higher plants pp. 69-
104. In:Oxidative stress in plants. Inzé, D. Van Montagu, M. E. (ed). Taylor &
Francis, London and New York.
LIU, J., STEVENS, C., KHAN, V. A., LU, J. Y., WILSON, C. L., ADEYEYE, O.,
KABWE, M. K., PUSEY, P. L., CHALUTZ, E., SULTANA, T. 1993. Application of
ultraviolet-C light on storage rots and ripening of tomatoes. Journal of Food
Protection 56: 868-872.

66
LU, J., STEVENS, C., KHAN, V., KABWE, M., WILSON, C. 2007a. The effect of
ultraviolet irradiation on shelf-life and ripening of peaches and apples. Journal of
Food Quality 14: 299-305.
LU, J. Y., STEVENS, C., KHAN, V. A., KABWE, M., WILSON, C. L. 2007b. The
effect of ultraviolet irradiation on shelf-life and ripening of peaches and apples.
Journal of Food Quality 14: 299-305.
MAHARAJ, R., ARUL, J., NADEAU, P. 1999. Effect of photochemical treatment in
the preservation of fresh tomato (Lycopersicon esculentum cv. Capello) by delaying
senescence. Postharvest Biology and Technology 15: 13-23.
MAHARAJ, R., SANKAT, C. K. 1990. Storability of papayas under refrigerated and
controlled atmosphere. 375-386.
MANENOI, A., BAYOGAN, E., THUMDEE, S., PAULL, R. 2007. Utility of 1-
methylcyclopropene as a papaya postharvest treatment. Postharvest Biology and
Technology 44: 55-62.
MARTÍNEZ-TÉLLEZ, M. A., BALANDRÁN-QUINTANA, R. R., SOTO-CERÓN, R.,
QUINTERO-RAMOS, A., MÁRQUEZ-MELÉNDEZ, R. 2008. Poliaminas y
acondicionamiento térmico reducen daño por frío y afectan actividad de β-
galactosidasa en calabaza zucchini Revista Chapingo Serie Horticultura 14: 243-
248.
MCGUIRE, R. G. 1992. Reporting of objective color measurements. HortScience 27:
1254-1254.
NUNES, M. C. N., EMOND, J. P., BRECHT, J. K. 2006. Brief deviations from set
point temperatures during normal airport handling operations negatively affect the
quality of papaya (Carica papaya) fruit. Postharvest Biology and Technology 41:
328-340.
PAULL, R. E., CHEN, N. J. 1989. Waxing and plastic wraps influence water loss
from papaya fruit during storage and ripening. J. Amer. Soc. Hort. Sci 114: 937-942.
PEREZ-CARRILLO, E., YAHIA, E. M. 2004. Effect of postharvest hot air and
fungicide treatments on the quality of 'maradol' papaya (Carica papaya L.). Journal
of Food Quality 27: 127-139.

67
POMBO, M. A., DOTTO, M. C., MARTÍNEZ, G. A., CIVELLO, P. M. 2009. UV-C
irradiation delays strawberry fruit softening and modifies the expression of genes
involved in cell wall degradation. Postharvest Biology and Technology 51: 141-148.
PUROHIT, A. K., RAWAT, T. S., KUMAR, A. 2003. Shelf life and quality of ber
[Zizyphus mauritiana Lamk.] fruit cv. Umran in response to post harvest application
of ultraviolet radiation and paclobutrazol. Plant Foods for Human Nutrition (Formerly
Qualitas Plantarum) 58: 1-7.
SANCHEZ-BALLESTA, M. T., ZACARIAS, L., GRANELL, A., LAFUENTE, M. T.
2000. Accumulation of PAL transcript and PAL activity as affected by heat-
conditioning and low-temperature storage and its relation to chilling sensitivity in
mandarin fruits. J. Agric. Food Chem 48: 2726-2731.
SAPERS, G. M., MILLER, R. 1995. Heated Ascorbic/Citric Acid Solution as
Browning Inhibitor for Pre-Peeled Potatoes. Journal of Food Science 60: 762-766.
STEVENS, C., LIU, J., KHAN, V. A., LU, J. Y., WILSON, C. L., IGWEGBE, E. C. K.,
KABWE, M. K., CHALUTZ, E., DROBY, S. 1998. Application of hormetic UV-C for
delayed ripening and reduction of Rhizopus soft rot in tomatoes: the effect of
tomatine on storage rot development. Journal of Phytopathology-Phytopathologische
Zeitschrift 146: 211-221.
STEVENS, C., WILSON, C. L., LU, J. Y., KHAN, V. A., CHALUTZ, E., DROBY, S.,
KABWE, M. K., HAUNG, Z., ADEYEYE, O., PUSEY, L. P., WISNIEWSKI, M. E.,
WEST, M. 1996. Plant hormesis induced by ultraviolet light-C for controlling
postharvest diseases of tree fruits. Crop Protection 15: 129-134.
THOMAS, P., MOY, J. H. 1986. Radiation preservation of foods of plant origin. III.
Tropical fruits: bananas, mangoes, and papayas. Critical Reviews in Food Science
and Nutrition 23: 147-205.
VICENTE, A. R., PINEDA, C., LEMOINE, L., CIVELLO, P. M., MARTINEZ, G. A.,
CHAVES, A. R. 2005a. UV-C treatments reduce decay, retain quality and alleviate
chilling injury in pepper. Postharvest Biology and Technology 35: 69-78.
VICENTE, A. R., PINEDA, C., LEMOINE, L., CIVELLO, P. M., MARTÍNEZ, G. A.,
CHAVES, A. R. 2005b. UV-C treatments reduce decay, retain quality and alleviate
chilling injury in pepper. Postharv. Biol. Technol 35: 69-78.

68
VICENTE, A. R., REPICE, B., MARTINEZ, G. A., CHAVES, A. R., CIVELLO, P. M.,
SOZZI, G. O. 2004. Maintenance of fresh boysenberry fruit quality with UV-C light
and heat treatment combined with low storage temperature. Journal of Horticultural
Science and Biotechnology 79: 246-251.
69
∆∆ ∆∆ L*
0
2
4
6
8
10
12TestigoUV-C
NS
*
NS
NS
*
NS
NS
NS
∆∆ ∆∆ H
ue°
0
10
20
30
TestigoUV-C
NS
NS
NS
NS
∆∆ ∆∆ C
hrom
a
0
5
10
15
20
TestigoUV-C
a
b
c
12d 5°C+2d 25°C
12d 14°C+2d 25°C
12d 25°C+2d 25°C
12d 1°C+2d 25°C
FIGURA 1. Efecto del tratamiento de UV-C (1.76 kJ·m-2) y temperatura de
almacenamiento (1, 5, 14 y 25 ± 2 ° C; 80 ± 10 % HR ) sobre el cambio en los
parámetros de color L (a), Chroma (b) y ° Hue (c) en frutos de papaya ‘Maradol’
después de 12 días de almacenamiento y trasferencia a 25 °C por 2 días. Cada
barra representa la media de nueve observaciones ± el error estándar. NS, *: no
significativo y significativo entre tratamiento de UV-C y control a una P ≤ 0.05, de
acuerdo con la prueba de Tukey.
70
% P
érdi
da d
e pe
so
0
2
4
6
8
10
12TestigoUVC
NS
NS
*
12d 1 °C+2d 25 °C
*
12d 5 °C+2d 25 °C
12d 14 °C+2d 25 °C
12d 25 °C+2d 25 °C
FIGURA 2. Efecto del tratamiento de UV-C y temperatura de almacenamiento sobre
la pérdida de peso (%) de frutos de papaya var. ‘Maradol’. Cada barra representa la
media de seis observaciones ± el error estándar después de 12 días a 1, 5, 14 y 25
± 2 ° C y 2d a 25 °C. NS, *: no significativo y significativo entre tratamientos a una P
≤ 0.05 de acuerdo con la prueba de Tukey.
71
% P
érdi
da d
e fir
mez
a
0
20
40
60
80
100
Testigo UV-C
NS
NS
NS
NS
12d 5 °C+2d 25 °C
12d 14 °C+2d 25 °C
12d 25 °C+2d 25 °C
12d 1 °C+2d 25 °C
FIGURA 3. Efecto del tratamiento de UV-C y temperatura de almacenamiento sobre
la pérdida de firmeza (%) de frutos de papaya var. ‘Maradol’. Cada barra representa
la media de 18 mediciones ± el error estándar después de 12 días a 1, 5, 14 y 25 ±
2 ° C + 2d a 25 ° C. NS no significativo entre tratamientos a una P ≤ 0.05 de acuerdo
con la prueba de Tukey.
72
∆∆ ∆∆SS
T (
°Brix
)
0
2
4
6TestigoUV-C
12d 5 °C+2d 25 °C
12d 14 °C+2d 25 °C
12d 25 °C+2d 25 °C
12d 1 °C+2d 25 °C
NS NS
NS
*
FIGURA 4. Efecto del tratamiento de UV-C y temperatura en el contenido de sólidos
solubles totales de frutos de papaya var. ‘Maradol’. Cada barra representa la media
de 12 mediciones ± el error estándar después de 12 días a 1, 5, 14 y 25 ± 2 ° C + 2d
a 25 ° C. NS, *: no significativo y significativo a una P ≤ 0.05 respectivamente, de
acuerdo con la prueba de Tukey.

73
∆∆ ∆∆ D
eter
ioro
(0-
5)
0
1
2
3
4
5
TestigoUV-C
* *
*
*
Índi
ce d
e da
ño p
or fr
ío (
1-5)
0
1
2
3
4
5
*
NS
NS
12d 5 °C+2d 25 °C
12d 14 °C+2d 25 °C
12d 25 °C +2d 25 °C
12d 1 °C+2d 25 °C
FIGURA 5. Efecto del tratamiento de UV-C y temperatura en el deterioro general y
daño por frío de frutos de papaya var. ‘Maradol’. Cada barra representa la media de
6 unidades experimentales ± la desviación estándar después de 12 días a 1, 5, 14 y
25 ± 2 ° C + 2d a 25 ° C. NS no significativo y * significativo a una P ≤ 0.05 * de
acuerdo con la comparación de medias por Tukey-Kramer.

CAPÍTULO IV Efecto de la irradiación UV-C y bajas temperaturas de almacenamiento sobre compuestos bioactivos, enzimas antioxidantes y actividad de inhibición de radicale s de frutos de papaya.

75
Effect of UV-C irradiation and low temperature stor age on bioactive
compounds, antioxidant enzymes and radical scavengi ng activity of
papaya fruit.
D. M. Rivera-Pastrana; G. A. González-Aguilar*; J. F. Ayala-Zavala; E. M. Yahia;
A. A. Gardea Béjar; R. Sotelo-Mundo.
Coordinación de Tecnología de Alimentos de Origen Vegetal, Centro de
Investigación en Alimentación y Desarrollo. Km. 0.6 Carretera a la Victoria.
Hermosillo, Sonora, C.P. 8300. México.
Corresponding Author:
Gustavo A. González-Aguilar.
Phone: 00-52-6622-892400, Ext. 272.
Fax: 00-52-6622-80-0422.
Email: [email protected]

76
Abstract
Mature green ‘Maradol’ papaya fruits were exposed to UV-C irradiation (1.76
kJ·m-2) and stored at 5 or 14 °C. Changes in total phenol s, total flavonoids,
enzymatic activities of superoxide dismutase (SOD), catalase (CAT) and
peroxidase (POD), as well as the scavenging activity against DPPH and ABTS
radicals were investigated in peel and flesh tissues during 15 days of storage.
UV-C irradiation increase flavonoid content and radical scavenging activity in
peel; and CAT activities in flesh. Flavonoid contents, CAT and SOD activities
were increased by low storage temperatures. UV-C irradiation effect on radical
scavenging of papaya peel could be attributed to increased flavonoid content.
Antioxidant system of papaya was activated by UV-C and cold storage by
increasing phenolic content and antioxidant enzymatic activities as a defense
response against oxidative-stress.
Keywords: Carica papaya L., antioxidant capacity, antioxidant enzyme, UV-C,
hormesis.

77
1. Introduction
The market for tropical fruits has increased in last years due to an increasing
consumer demand for exotic products, changes in diet habits, attractive sensorial
properties and because they supply an optimal mixture of antioxidants. Tropical
fruits contain polyphenols, carotenoids and vitamins C and E that could
contribute to improve health by reducing incidence of cronical degenerative
diseases (Yahia, 2010); and some of these phytochemicals are implicated in
reducing fruit postharvest decay (González-Aguilar et al., 2007b). Among tropical
fruits, papaya is one of the most popular because its characteristic taste and
nutraceutical value (González-Aguilar et al., 2008). However, papaya fruits are
very susceptible to deterioration and postharvest losses mainly by fungal decay,
physiological disorders such as chilling injury, pests, mechanical injury and over-
ripeness (da Silva et al., 2007; Perez-Carrillo and Yahia, 2004).
Most postharvest treatments involve the alterations of fruit natural
conditions of the fruit in order to induce an acclimation response that prolongs
postharvest life. Controlled postharvest abiotic stresses including temperature
and UV irradiation, among others, can be applied to fresh fruits in order to induce
the synthesis of specific secondary metabolites synthesis with antioxidant
properties (Cisneros-Zevallos, 2003).
Exposure to UV irradiation and low temperatures induce oxidative stress
in plant tissues by increasing reactive oxygen species (ROS). To cope with this
stress, the most effective protection mechanism stimulated by UV light is
flavonoids biosynthesis and other UV-absorbing phenolic compounds
(Frohnmeyer and Staiger, 2003). When exposed to low temperatures, the plant
antioxidant system is also activated at different levels, but the precise
mechanisms remain to be elucidated (Karpinski et al., 2002).
Therefore, some postharvest treatments could induce defense
mechanisms that affect produce metabolic activity, such as the triggering of fruit
antioxidant system by promoting an increase on healthy compounds, and
reducing decay susceptibility. UV-C irradiation and low temperatures abiotic
stresses could activate antioxidant defense system of papaya fruit. This study

78
evaluated the effects of UV-C treatment in conjunction with low storage
temperatures on bioactive compounds, antioxidant enzymes and radical
scavenging activity of papaya fruit.
2. Materials and Methods
2.1. Plant Material
Mature-green ‘Maradol’ papayas were obtained from a wholesale market in
Hermosillo, Sonora, Mexico. Fruits of uniform maturity stage (mature-green), size
and free from defects were selected and randomized in two groups of 48 fruits.
Half of samples on each group were treated with UV-C prior to storage for 15
days at 5 or 14 °C. Sampling was performed at five days intervals (0, 5, 10, 15).
Peel was separated manually with a sharp stainless steel manual peeler, and
immediately stored at -30 °C until use, to avoid sa mple degradation. Papaya
flesh was cut into cubes of 5 cm and also stored at -30 °C. Every measurement
was done by triplicate, where the experimental unit was the peel or flesh of three
fruits. The experiment was replicated twice.
2.2. Chemicals
Folin-ciocalteu reagent, aluminum chloride, sodium carbonate, sodium nitrate,
sodium hydroxide, potassium persulphate gallic acid, quercetin, β-
mercaptoethanol, tris-hidrochloride, sodium acetate, potassium phosphate, L-
metionine, EDTA, riboflavin, nitro blue tetrazolium (NBT), guaicol, bradford
reagent, bovine serum albumin, 1-diphenyl-2-picrylhydrazyl (DPPH), 2, 2’-
azinobis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS.-) were obtain from
Sigma-Aldrich (St. Louis, MO, USA). Methanol (HPLC grade) and ethanol
(reagent grade) were purchased from JT Baker (Xalostoc, Estado de Mexico,
Mexico).

79
2.3. UV-C treatment
Papaya fruits were washed and sanitized by immersion in hypochlorite solution
(200 ppm free chlorine) before exposure to a UV-C dose of 1.76 kJ·m-2, in order
to eliminate impurities and reduce fungal attack risk that could influence
treatment effect. This irradiation dose was selected since does not caused visible
scald according to preliminary experiments carried out in our laboratory (data not
shown). Irradiation was emitted by two unfiltered General Electric 15 Watts (G15
T8) germicidal lamps placed 15 cm above and below fruits. Irradiance emitted
(95%) by these lamps was in the UV-C (100-280 nm) region. After treatment
control and treated fruits were stored for 15 days at 5 or 14 °C in the absence of
light. A completely randomized experimental design was followed, with two
replicates per treatment and storage temperature.
2.4. Total phenols and flavonoids content
Phenolic compounds of papaya were extracted from 5 or 10 g of peel or flesh
tissue respectively. Those were homogenized in 20 mL of methanol solution
(80%). The homogenate was sonicated for 30 min at 40 °C, and then centrifuged
at 9400 xg for 15 min at 5 °C in an Allegra 64R Beckman Coult er centrifuge (Palo
Alto, Calif., USA). Supernatants were collected after filtered through Whatman
paper (grade1). Same procedure was repeated 3 times, these extracts were
used to measure total phenolics and flavonoid contents, as well as antioxidant
capacity.
Total phenols were measured as reported by (González-Aguilar et al.,
2007), where 50 µL of extract were mixed with 3 mL of H2O and 250 µL of Folin-
Ciocalteu phenol reagent (1 N). After equilibrating for 5 minutes, 750 µL of 20%
Na2CO3 and 950 µL of H2O were added to the reaction mixture. After vortexing,
the reaction was incubated for 30 minutes at room temperature protected from
light. Absorbance was then measured at 765 nm in a UV-Vis spectrophotometer
(Cary50 Bio model, Varian, Italy). Total phenol content was calculated based on

80
a gallic acid standard curve; results were expressed as mg of gallic acid per 100
g of fresh weight (mg GA /100gfw).
Flavonoid content was determined as described by (Zhishen et al., 1999)
with some modifications. One mL of methanolic extract was mixed with 4 mL of
H2O and 300 µL of 5% NaNO2. After agitation followed by a 5 min incubation,
300 µL of 10% AlCl3 were added and incubated for one more minute. Two mL of
NaOH 1 M and H2O were added to make up 10 mL of reaction volume. After
agitation, absorbance was measured at 415 nm, in a UV-Vis spectrophotometer
(Cary50 Bio model, Varian, Italy). Results were expressed as mg of quercetin
equivalents per 100 g of fresh weight (mg quercetin /100gfw).
2.5. Enzymatic activity assays
Catalase (EC. 1.11.1.6; CAT) activity was determined as described previously by
Blackwell et al. (1990), with the following modifications. The enzyme was
extracted from 0.2 or 0.5 g of acetone powder from peel or flesh tissue
respectively. It was homogenized in 10 mL of 0.1 M Tris-HCl, pH 8.5, containing
5 mM β-mercaptoethanol. The mixture was then agitated for 20 min at 4 °C and
centrifuged for 20 min at 12 000 xg and 4 °C. The reaction mixture contained 3
mL of 10 mM Tris-HCl, pH 8.5 and 0.1 mL of 0.88% H2O2 in 100 mM Tris-HCl; it
was started by adding 0.2 mL of enzyme extract. CAT activity was monitored at
240 nm for 5 min at room temperature (24-26 °C) wit h a UV-Vis
spectrophotometer. One unit of CAT specific activity was reported as the
decomposition of 1 µmol of H2O2/min·mg of protein.
Peroxidase (EC. 1.11.1.7; POD) activity was determined as described
Pérez-Tello et al. (2009) with some modifications. Enzyme was extracted from
0.1 or 0.2 g of acetone powder from peel or flesh tissue, respectively,
homogenized in 5 mL of 0.1 M Tris-HCl (pH 8.0), containing 5 mM β-
mercaptoethanol. The mixture was agitated for 20 minutes at 4 °C, centrifuged
for 30 min at 12 000 xg and 4 °C, and the supernatant was decanted. POD
activity was measured at 470 nm for 2 min at 30 °C in 2.15 mL reaction solution
with 10 mM sodium acetate (pH 5.3) containing 0.5% guaiacol, 0.25 mL of 0.1%

81
H2O2 and 0.1 mL of enzyme extract. POD specific activity was reported as the
decomposition of 1 mmol of guaicol/min·mg of protein.
Superoxide dismutase (1.15.1.1; SOD) activity was determined according
to Tejacal et al. (2005b) with modifications. The enzyme was extracted from 0.2 g
of acetone powder from peel or flesh tissue, homogenized in 10 mL of potassium
phosphate at pH 7.8. The mixture was agitated for 20 minutes at 4 °C,
centrifuged for 30 min at 12 000 xg and 4 °C and th e supernatant was decanted.
Reaction mixture consisted on 27 mL of 0.05 M phosphate buffer (pH 7.8)
containing 0.1 mM EDTA, 1.5 mL of L-methionine solution (30 mg mL-1), 1 mL of
nitroblue tetrazolium (1.41 mg mL-1) and 0.75 mL of X-100 triton solution (1%).
Following 0.03 mL of riboflavin solution (4.4 mg 100 mL-1) and 0.4 mL of enzyme
extract were added to 3 mL of reaction mixture and homogenized. Then the
reaction mixture was exposed to fluorescent light emitted by two lamps of 20 W
for 15 minutes, and absorbance was measured at 560 nm on a UV-Vis
spectrophotomer (Cary Bio50, Varian, Italy). Reaction velocity was determined
as absorbance increment due to nitroblue tetrazolium formazan formation per
unit of time. One unit of SOD was defined as enzyme extract concentration that
inhibits 50% of nitroblue tetrazolium formazan formation. Assays were performed
at room temperature (24-26 °C). SOD specific activi ty was expressed as units of
activity per gram of protein (U·min-1·gprot-1).
Total protein concentration was determined in all enzymatic extracts for
specific enzymatic activity calculation using bovine serum albumin as standard
(Bradford, 1976).
2.6. Radical scavenging activities, DPPH• and ABTS•+.
Radical scavenging activity of papaya was measured on methanolic extracts, by
the DPPH• (2,2-diphenyl-1picrylhydrazyl) and ABTS•+ methods, described by
Jimenez-Escrig et al. (2000) and Re et al. (1999) with some modifications.
For DPPH scavenging activity, 0.1 mL of methanolic extracts as used for
phenol and flavonoid analyses was added to 3.9 mL of DPPH• methanolic

82
solution (0.025 g/L). Fading of DPPH• color solution upon reduction was
measured in a UV-Vis spectrophotometer (Cary50 Bio model, Varian, Italy) at
515 nm after a 30 min room temperature dark incubation in darkness. Sample
was substituted with 80% methanol in the blank reaction, and methanol was used
for baseline correction. The radical scavenging activity was expressed as DPPH•
inhibition percentage = (blank OD – sample OD/ blank OD) x 100.
For the ABTS radical scavenging assay, 2.45 mmol of potassium
persulphate were added to 7 mM ABTS dissolved in water, stirred and incubated
for 12-16 h in darkness to give a dark blue solution. Solution was diluted with
ethanol until absorbance reached 0.7 at 734 nm. Extract (0.1 mL) was added to
3.9 mL of resulting radical solution in a quartz spectrophotometer cell. Reaction
was monitored for 5 min until end point, when absorbance became stable.
Sample was substituted with 80% methanol for blank reaction, and ethanol was
used for baseline correction. For comparison between methods, radical
scavenging activity was expressed as ABTS•+ inhibition percentage = (blank OD
– sample OD/ blank OD) x 100.
2.7. Statistical analysis
The effect of UV-C treatment, storage temperature, storage time and type
of tissue on total phenols, total flavonoids, CAT, POD and SOD enzymatic
activities, and DPPH and ABTS radical scavenging activities in papaya fruit were
evaluated by analysis of variance (ANOVA) of general linear models (GLM)
where differences were considered significant at a P< 0.05 based on a Duncan’s
multiple comparison test using the NCSS (2007) statistical software.
3. Results and Discussion
3.1. Phenolic and flavonoid contents
Total phenolic content (TPC) changes were evaluated in peel and flesh of
papaya fruit stored at 5 and 14 °C in response to U V-C treatment (Figure 1). The

83
TPC of fresh papaya was significantly affected (P<0.05) by UV-C irradiation
treatment, storage time and type of tissue analyzed. Highest phenolic contents
were found in control peel of papaya fruit. Although a decreasing tendency on
TPC was observed through storage time, no significant (P>0.05) effect of storage
temperature on TPC was found,
A similar tendency was found on TPC of peel and flesh tissues, at the
beginning of storage, controls at 5 °C had the high est phenolic content.
Increased TPC on UV-C treated papaya peel was observed only at 10 days of
storage at 14 °C, as compared with controls. In gen eral, the effect of UV-C
treatment on phenolic content of papaya fruit was tissue specific and dependent
upon storage temperature.
Accumulation of phenolic compounds varied strongly in relation to fruit
physiological state and results from the equilibrium between biosynthesis and
further metabolism including turnover and catabolism (Oufedjikh et al., 2000).
The effect of UV-C irradiation on phenylpropanoid metabolism has been
described previously in various fruit, where phenylalanine ammonia-lyase (PAL),
a key regulatory enzyme of phenylpropanoid biosynthetic route is activated by
UV-C light, leading to enhanced phenolic contents (Erkan et al., 2008; Gonzalez-
Aguilar et al., 2007a; González-Aguilar et al., 2007b; Stevens et al., 1999;
Stevens et al., 1998).
Total flavonoid content (TFC) of papaya fruit was significantly affected by
UV-C treatment, type of tissue analyzed, storage temperature and time (Figure
2). UV-C treated papaya, presented significantly (P<0.05) lower flavonoid
contents than controls. Cold storage (5 °C) had a p ositive effect on TFC despite
irradiation treatment or type of tissue, while a linear positive effect of storage time
was observed reaching highest mean values at day 15. An intermittent effect of
treatment was observed during storage at 14 °C in b oth tissues; were a positive
effect of UV-C treatment on TFC was found only at d 10. Thus, cold storage had
a bigger impact in TFC in both peel and flesh tissues of papaya than UV-C
treatment.

84
It had been demonstrated that flavonoids can act as UV protective agents,
participating in several plant resistance mechanisms by interfering with oxidative
processes both by chelating metal ions or by scavenging oxi-radicals, forming
less reactive “antioxidant radicals” that disappear by dismutation, recombination
or reduction (Bors et al., 1998; Bors et al., 1992). Our results are in accordance
to previous reports indicating that UV-C irradiation enhanced flavonoid content in
qumquats and oranges (D'hallewin et al., 1999; Rodov et al., 1992), grapes
(Cantos et al., 2003; Cantos et al., 2001), broccoli (Costa et al., 2006), and
mangoes (González-Aguilar et al., 2007b). However, treatment effect in papaya
was transitory. In contrast, no clear effect of treatment was found on total
phenolic contents, which is in agreement with previous studies findings in UV
irradiated shiitake mushrooms (Jiang et al., 2010).
In this study, low temperature storage affected positively some
components of papaya antioxidant system, particularly flavonoid compounds.
This is in accordance to previous findings describing a cultivar-dependent
increase in antioxidant capacity related to phenolic content of blueberry during
low-temperature storage (Connor et al., 2002). Similarly, cold storage retained or
increased phenolic contents and antioxidant capacity of strawberries and fresh-
cut mangoes (Ayala-Zavala et al., 2004; Robles-Sanchez et al., 2009). In this
context, in fruits were phenols and flavonoid compounds contribution to
antioxidant system is greater than that of ascorbic acid, low temperature storage
could increase fruit antioxidant capacity by increasing phenolic content
(Shivashankara et al., 2004).
3.2. Antioxidant enzymes (CAT, POD, SOD)
Catalase specific activities of papaya peel and flesh are shown in Figure 3. A
significant effect (P<0.05) of storage time and temperature on CAT activity was
observed in papaya. Fruit stored at 5 °C reached h ighest levels of CAT activity,
while an indirect linear effect of storage time was observed. UV-C irradiation
treatment and type of tissue did not affected CAT activity significantly (P>0.05);
however, UV-C treatment increased CAT activity in peel after five days at 5 °C

85
(688 µMH2O2/ min· mg protein). An inhibitory effect of 14 °C s torage temperature
on CAT activity was observed in control and treated samples. On the other hand,
UV-C treatment increased CAT activity (33%) in flesh of papaya during the first
ten days of storage at 5 °C compared to controls.
CAT protects cells against ROS because it catalyzes the decomposition of
hydrogen peroxide into oxygen and water, with high stability under cold storage
conditions (Sala and Lafuente, 1999). Our results are in agreement with those
observed in maize seedlings and mandarin fruits, were, increases in CAT activity
seems to respond to an induce-oxidative stress, promoted by low temperature
storage (Prasad, 1997; Sala and Lafuente, 1999).
POD activity of papaya peel was significantly (P<0.05) affected by UV-C
treatment, storage time and temperature (Figure 4). Decreased POD activity was
observed on papaya as a result of UV-C treatment and storage at 5 °C; also,
average activities diminished after storage. Contrary to what we found for the
other two enzymes, POD activity increased with storage temperature. At 14 °C,
POD levels remain constant or increased in UV-C treated and control samples.
Maximum activity was observed in control peel samples at day 10 (765 mM
guaicol/ min·mg protein). Activity of POD in papaya flesh was not detected.
Guaicol peroxidase and ascorbate peroxidase, are peroxidase enzymes
found in animal, plant and microorganism tissues, catalyzing oxidoreduction
between hydrogen peroxide (H2O2) and various reductants (Hiraga et al., 2001).
There is evidence suggesting that UV-C irradiation can induce a rapid
accumulation of photo-oxidation products. Plants respond by increasing POD
activity in grapes, fresh-cut melon and tomato, extending postharvest life of these
fruits (Barka et al., 2000; Lamikanra et al., 2005; Nigro et al., 1998).
Nevertheless, we found in papaya peel we found an inhibitory effect of cold
storage on POD activities and no significant effect of UV-C treatment despite
storage conditions that could be attributed to ripening or senescence delay, as it
has been described in sapote mamey fruits (Tejacal et al., 2005a).

86
SOD activity of fresh papaya was significantly affected by UV-C treatment,
temperature, time and type of tissue analyzed (Figure 5). UV-C irradiation
reduced SOD levels of papaya fruit at the storage conditions analyzed.
Diminished SOD levels were also accounted as effect of storage time with
respect to initial values in papaya peel; however, no mean differences were
found after day 5. Maximum SOD activity was found in control flesh samples
stored at 5 °C (16.8 Units· mg protein -1) and a linear increment was observed
with storage time. A positive effect of cold storage (5 °C) was observed in SOD
despite other factors.
Superoxide dismutases (SOD), a class of metal-containing proteins,
catalyze the dismutation reaction of superoxide radical anions into H2O2 and
molecular oxygen (Scandalios, 1993). SOD removes singlet oxygen, prevents
hydroxyl radicals formation and has been implicated as an essential defense
against oxygen toxicity (Fridovich, 1986; McCord, 1979). Since SOD is the first
line of cell defense, against free radicals, its high activity in fruits has been
related to a higher resistance to stress and a longer commercial life (Mondal et
al., 2004; Wang and Jiao, 2001). Other authors reported a decreased SOD
activity in strawberries fruits stored at 10 °C whe n compared to initial levels, but
after 15 d of storage, SOD activities of UV-C treated fruit were higher than
controls suggesting that treatment could activate fruit defensive responses
(Erkan et al., 2008). On the other hand, UV-C treatment reduced SOD activity
during ripening of tomato, but increased activity of other antioxidant enzymes as
a defense mechanism against oxidation (Barka, 2001). Therefore, increase in
SOD activities in papaya flesh could be a defense response against oxidative
stress caused by cold storage and in a minor degree to UV-C treatment.
3.3. DPPH· and ABTS radical scavenging activity
Figure 6 shows antioxidant capacity of peel and flesh tissues determined
as DPPH· and ABTS· radical inhibition percentage. A significant effect of tissue
type and storage time and temperature was observed in papaya fruit. Peel

87
radical scavenging capacity was twice as high as that in flesh (35 and 69% total
average respectively). In papaya peel, radical scavenging was increased in
samples stored at 5 °C, while at 14 °C, radical inh ibition was lower than initial
levels with no differences among treatments. Nevertheless, a positive effect of
UV-C treatment in peel antioxidant capacity was observed at 14 °C during the
storage period. In contrast, there was no effect of UV-C treatment in DPPH•
antioxidant activity of flesh samples at both storage temperatures.
A similar tendency was found for both antioxidant capacity methods,
however the values of antioxidant capacity determined with the ABTS method
were in average 20% higher and a significant (P<0.05) effect of UV-C treatment
was found. UV-C treatment, type of tissue, and storage time and temperature
had a significant effect on ABTS radical scavenging in papaya fruit. A reduction
on ABTS scavenging by effect of UV-C treatment and storage at 14 °C was
observed. Peel tissue average radical scavenging was 32 % higher than that of
flesh, however, irradiation treatment only increased radical inhibition significantly
in flesh tissue, in samples stored at 5 and 14 °C, radical scavenging activity
oscillated in a range of 45-57 and 51-84%, respectively.
It had been described that UV-C postharvest treatment in shiitake
mushrooms increased total flavonoids, ascorbic acid, antioxidant activities of
CAT, SOD, ascorbate-POD, and delayed the increase in singlet oxygen and
hydrogen peroxide production rate; indicating an improvement of antioxidant
capacity (Jiang et al., 2010). Also, an enhanced antioxidant capacity attributed to
total flavonoid content on UV-C treated fresh-cut mangoes and tomatoes was
reported earlier (Gonzalez-Aguilar et al., 2007; Vicente et al., 2005). However, in
this study, UV-C effect on papaya antioxidant capacity was intermittent during
storage, since, significant positive effect of UV-C on flavonoid and radical
scavenging activities was found at day 10, but not maintained until the end of
storage.
The delay in ripening and senescence of UV-C treated tomatos had been
attributed to significant increases in the levels of non-specific antioxidants such
as polyamines and phenols, suggesting that maybe the singlet oxygen

88
scavenging mechanism is not affected by UV treatment to the same extent as for
the superoxide scavenging mechanism (Maharaj, 1995; Maharaj et al., 1999). In
the same manner, in papaya, the antioxidant response against UV-C and cold
storage appear to be mediated mainly by flavonoids in peel and by CAT or other
antioxidants in flesh.
Since UV-C irradiation has a very low penetration energy, especially in
solids (< 5 µm), treatment effects are expected to appear mostly at surface level,
which was confirmed in this case for papaya peel. The more marked effect of
UV-C on papaya peel could also be explained by the fact that this particular
tissue contain higher levels of flavonoids and UV-C treatment, promoting the
activation of this particular biosynthetic route.
4. Conclusion
UV-C treatment and cold storage promoted changes in different components of
the antioxidant system of papaya fruit ‘Maradol’. Main effects of UV-C irradiation
were an increment on flavonoid contents in peel and higher CAT enzymatic
activity in flesh. While in response to cold storage, only SOD enzymatic activity
was also increased as a result of a defense response to oxidative stress. Our
study shows that UV-C irradiation affect antioxidant response of papaya,
however, this effect is dependent of storage conditions and type of tissue. Thus,
UV-C efficiency as a postharvest treatment to enhance antioxidant system of
papaya fruit is compromised by storage conditions.
Acknowledgements
The authors gratefully acknowledge financial support of CONACYT and technical
assistance of Mónica A. Villegas-Ochoa and Chrystian M. Rodríguez.

89
References
Ayala-Zavala, J.F., Wang, S.Y., Wang, C.Y., Gonzalez-Aguilar, G.A., 2004.
Effect of storage temperatures on antioxidant capacity and aroma compounds
in strawberry fruit. Lebensm-Wiss Technol 37, 687-695.
Barka, E.A., 2001. Protective enzymes against reactive oxygen species during
ripening of tomato (Lycopersicon esculentum) fruits in response to low
amounts of UV-C. Aust J Plant Physiol 28, 785-791.
Barka, E.A., Kalantari, S., Makhlouf, J., Arul, J., 2000. Impact of UV-C irradiation
on the cell wall-degrading enzymes during ripening of tomato (Lycopersicon
esculentum L.) fruit. J Agr Food Chem 48, 667-671.
Blackwell, R.D., Murray, A.J.S., Lea, P.J., 1990. Enzymes of photorespiratory
carbon pathway, In: Lea, P.J. (Ed.), Methods in plant biochemistry. Academic
Press, New York, New York, pp. 129-144.
Bors, W., Heller, W., Michael, C., 1998. The chemistry of flavonoids, In: Rice-
Evans, C.A., Packer, L. (Eds.), Flavonoids in health and disease. Marcel
Dekker New York.
Bors, W., Heller, W., Michel, C., Saran, M., 1992. Structural principles of
flavonoid antioxidants, In: Csomó, G., Fehér, J. (Eds.), Free Radicals and the
Liver. Springer, Berlin, Berlin, pp. 77–95.
Bradford, M., 1976. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding.
Anal Biochem 72, 248-254.
Cantos, E., Espin, J.C., Fernandez, M.J., Oliva, J., Tomas-Barberan, F.A., 2003.
Postharvest UV-C-irradiated grapes as a potential source for producing
stilbene-enriched red wines. J Agr Food Chem 51, 1208-1214.
Cantos, E., Espin, J.C., Tomas-Barberan, F.A., 2001. Postharvest induction
modeling method using UV irradiation pulses for obtaining resveratrol-
enriched table grapes: A new "functional" fruit? J Agr Food Chem 49, 5052-
5058.

90
Cisneros-Zevallos, L., 2003. The use of controlled postharvest abiotic stresses
as a tool for enhancing the nutraceutical content and adding-value of fresh
fruits and vegetables. J Food Sci 68, 1560-1565.
Connor, A.M., Luby, J.J., Hancock, J.F., Berkheimer, S., Hanson, E.J., 2002.
Changes in fruit antioxidant activity among blueberry cultivars during cold-
temperature storage. J Agr Food Chem 50, 893-898.
Costa, L., Vicente, A.R., Civello, P.M., Chaves, A.R., Martinez, G.A., 2006. UV-C
treatment delays postharvest senescence in broccoli florets. Postharvest Biol
Tec 39, 204-210.
D'hallewin, G., Schirra, M., Manueddu, E., Piga, A., Ben-Yehoshua, S., 1999.
Scoparone and scopoletin accumulation and ultraviolet-C induced resistance
to postharvest decay in oranges as influenced by harvest date. J Am Soc
Hortic Sci 124, 702-707.
da Silva, J., Rashid, Z., Nhut, D., Sivakumar, D., Gera, A., Souza Jr, M.,
Tennant, P., 2007. Papaya (Carica papaya L.) Biology and Biotechnology.
Tree and Forestry Science and Biotechnology 1, 47-73.
Erkan, M., Wang, S.Y., Wang, C.Y., 2008. Effect of UV treatment on antioxidant
capacity, antioxidant enzyme activity and decay in strawberry fruit.
Postharvest Biol Tec 48, 163-171.
Fridovich, I., 1986. Superoxide dismutase. Adv Enzymol 58, 61-64.
Frohnmeyer, H., Staiger, D., 2003. Ultraviolet-B radiation-mediated responses in
plants. Balancing damage and protection. Plant Physiol 133, 1420-1428.
González-Aguilar, G.A., Robles-Sánchez, R.M., Martínez-Téllez, M.A., Olivas,
G.I., Álvarez-Parrilla, E., de la Rosa, L., 2008. Bioactive compounds in fruits:
health benefits and effect of storage conditions. Stewart Postharvest Review
4, 1-10.
Gonzalez-Aguilar, G.A., Villegas-Ochoa, M.A., Martinez-Tellez, M.A., Gardea,
A.A., Ayala-Zavala, J.F., 2007a. Improving antioxidant capacity of fresh-cut
mangoes treated with UV-C. J Food Sci 72, S197-S202.

91
González-Aguilar, G.A., Zavaleta-Gatica, R., Tiznado-Hernández, M.E., 2007b.
Improving postharvest quality of mango 'Haden' by UV-C treatment.
Postharvest Biology and Technology 45, 108-116.
Hiraga, S., Sasaki, K., Ito, H., Ohashi, Y., Matsui, H., 2001. A large family of
class III plant peroxidases. Plant Cell Physiol 42, 462-468.
Jiang, T., Jahangir, M., Jiang, Z., Lu, X., Ying, T., 2010. Influence of UV-C
treatment on antioxidant capacity, antioxidant enzyme activity and texture of
postharvest shiitake (Lentinus edodes) mushrooms during storage.
Postharvest Biol Tec 56, 209-215.
Jimenez-Escrig, A., Jimenez-Jimenez, I., Sanchez-Moreno, C., Saura-Calixto, F.,
2000. Evaluation of free radical scavenging of dietary carotenoids by the
stable radical 2,2-diphenyl-1-picrylhydrazyl. J Sci Food Agr 80, 1686-1690.
Karpinski, S., Wingsle, G., Karpinska, B., Hällgren, J., 2002. Low-temperature
stress and antioxidant defense mechanisms in higher plants, In: Inzé, D., Van
Montagu, M. (Eds.), Oxidative Stress in Plants. Taylor & Francis, London and
New York, London and New York, pp. 69-103.
Lamikanra, O., Kueneman, D., Ukuku, D., Bett-Garber, K.L., 2005. Effect of
processing under ultraviolet light on the shelf life of fresh-cut cantaloupe
melon. J Food Sci 70, C534-C539.
Maharaj, R., 1995. The effect of ultraviolet radiation (UV-C) on the postharvest
storage behaviour of tomato (Lycopersicon esculentum Mill cv. Capello).
Université, Laval, Québec, Canada.
Maharaj, R., Arul, J., Nadeau, P., 1999. Effect of photochemical treatment in the
preservation of fresh tomato (Lycopersicon esculentum cv. Capello) by
delaying senescence. Postharvest Biol Tec 15, 13-23.
McCord, J., 1979. Superoxide dismutases: occurrence, structure, function and
evolution, In: Rattazi, M., Scandalios, J., Whitt, G.S. (Eds.), Isozyme: Current
Topics in Biological and Medical Research. Liss, New York, New York, pp. 1–
21.
Mondal, K., Sharma, N.S., Malhotra, S.P., Dhawan, K., Singh, R., 2004.
Antioxidant systems in ripening tomato fruits. Biol Plantarum 48, 49-53.

92
Nigro, F., Ippolito, A., Lima, G., 1998. Use of UV-C light to reduce Botrytis
storage rot of table grapes. Postharvest Biol Tec 13, 171-181.
Oufedjikh, H., Mahrouz, M., Amiot, M.J., Lacroix, M., 2000. Effect of gamma-
irradiation on phenolic compounds and phenylalanine ammonia-lyase activity
during storage in relation to peel injury from peel of Citrus clementina Hort.
ex. Tanaka. J Agr Food Chem 48, 559-565.
Perez-Carrillo, E., Yahia, E.M., 2004. Effect of postharvest hot air and fungicide
treatments on the quality of 'maradol' papaya (Carica papaya L.). J Food
Quality 27, 127-139.
Pérez-Tello, G., Martínez-Téllez, M., Vargas-Arispuro, I., González-Aguilar, G.,
2009. Chilling Injury in Mamey Sapote Fruit (Pouteria sapota): Biochemical
and Physiological Responses. American Journal of Agricultural and Biological
Science 4, 137-145.
Prasad, T., 1997. Role of catalase in inducing chilling tolerance in pre-emergent
maize seedlings. Plant Physiol 114, 1369-1376.
Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M., Rice-Evans, C.,
1999. Antioxidant activity applying an improved ABTS radical cation
decolorization assay. Free Radical Bio Med 26, 1231-1237.
Robles-Sanchez, R.M., Islas-Osuna, M.A., Astiazaran-Garcia, H., Vazquez-Ortiz,
F.A., Martin-Belloso, O., Gorinstein, S., Gonzalez-Aguilar, A., 2009. Quality
Index, Consumer Acceptability, Bioactive Compounds, and Antioxidant
Activity of Fresh-Cut "Ataulfo" Mangoes (Mangifera Indica L.) as Affected by
Low-Temperature Storage. J Food Sci 74, S126-S134.
Rodov, V., Ben-Yehoshua, S., Kim, J., Shapiro, B., Ittah, Y., 1992. Ultraviolet
illumination induces scoparone production in kumquat and orange fruit and
improves decay resistance. Journal of the American Society for Horticultural
Science (USA) 117, 788-792.
Sala, J.M., Lafuente, M.T., 1999. Catalase in the heat-induced chilling tolerance
of cold-stored Hybrid Fortune mandarin fruits. J Agr Food Chem 47, 2410-
2414.

93
Scandalios, J., 1993. Oxygen stress and superoxide dismutases. Plant Physiol
101, 7-12.
Shivashankara, K.S., Isobe, S., Al-Haq, M.M., Takenaka, M., Shiina, T., 2004.
Fruit antioxidant activity, ascorbic acid, total phenol, quercetin, and carotene
of Irwin mango fruits stored at low temperature after high electric field
pretreatment. J Agr Food Chem 52, 1281-1286.
Stevens, C., Khan, V.A., Lu, J.Y., Wilson, C.L., Chalutz, E., Droby, S., Kabwe,
M.K., Haung, Z., Adeyeye, O., Pusey, L.P., Tang, A.Y.A., 1999. Induced
resistance of sweetpotato to Fusarium root rot by UV-C hormesis. Crop Prot
18, 463-470.
Stevens, C., Khan, V.A., Lu, J.Y., Wilson, C.L., Pusey, P.L., Kabwe, M.K.,
Igwegbe, E.C.K., Chalutz, E., Droby, S., 1998. The germicidal and hermetic
effects of UV-C light on reducing brown rot disease and yeast microflora of
peaches. Crop Prot 17, 75-84.
Tejacal, I., León, M., Damián, M., Hernández, R., 2005a. Chilling in sapote
mamey (Pouteria sapota (Jacq.) H. E. Moore and Stearn). II. Changes in total
phenols and enzymatic activity. Rev Fitotec Mex 28, 25-32.
Tejacal, I.A., León, M.T.C., Damián, M.T.M., Hernández, R.M.S., 2005b. Chilling
injury in sapote mamey (Pouteria sapota (Jacq.) HE Moore and Stearn). I.
Changes in volatiles, firmness and total sugars. Rev Fitotec Mex 28, 17-24.
Vicente, A.R., Pineda, C., Lemoine, L., Civello, P.M., Martinez, G.A., Chaves,
A.R., 2005. UV-C treatments reduce decay, retain quality and alleviate chilling
injury in pepper. Postharvest Biol Tec 35, 69-78.
Wang, S.Y., Jiao, H.J., 2001. Changes in oxygen-scavenging systems and
membrane lipid peroxidation during maturation and ripening in blackberry. J
Agr Food Chem 49, 1612-1619.
Yahia, E., 2010. The Contribution of Fruit and Vegetable Consumption to Human
Health, In: de la Rosa, L.A., Alvarez-Parrilla, E., González-Aguilar, G.A.
(Eds.), Fruit and Vegetable Phytochemicals: Chemistry, Nutritional Value and
Stability. Wiley-Blackwell, Ames, Iowa p. 3.

94
Zhishen, J., Mengcheng, T., Jianming, W., 1999. The determination of flavonoid
contents in mulberry and their scavenging effects on superoxide radicals.
Food Chem 64, 555-559.

95
Figure legends
Fig. 1. Total phenolic content (mg gallic acid/ 100 gfw) of papaya peel and flesh
in control and UV-C treated samples during storage at 5 or 14 °C. Vertical
bars represent standard error.
Fig. 2. Total flavonoid contents (mg quercetin/ 100 gfw) of papaya peel and flesh
in control and UV-C treated samples during storage at 5 or 14 °C. Vertical
bars represent standard error.
Fig. 3. Specific CAT enzyme activities (µmol of decomposed H2O2/min·mg of
protein) of papaya peel and flesh in control and UV-C treated fruit during
storage at 5 or 14 °C. Vertical bars represent stan dard error.
Fig. 4. Specific POD enzyme activities (mmol of decomposed guaicol/min·mg of
protein) of papaya peel in control and UV-C treated fruit during storage at
5 or 14 °C. Vertical bars represent standard error.
Fig. 5. Specific SOD enzyme activities (Units/mg of protein) of papaya peel and
flesh in control and UV-C treated fruit during storage at 5 or 14 °C. Vertical
bars represent standard error.
Fig. 6. Changes on DPPH and ABTS scavenging activity of papaya peel and
flesh in response to UV-C irradiation treatment and storage temperature (5
or 14 °C). Vertical bars represent standard error.
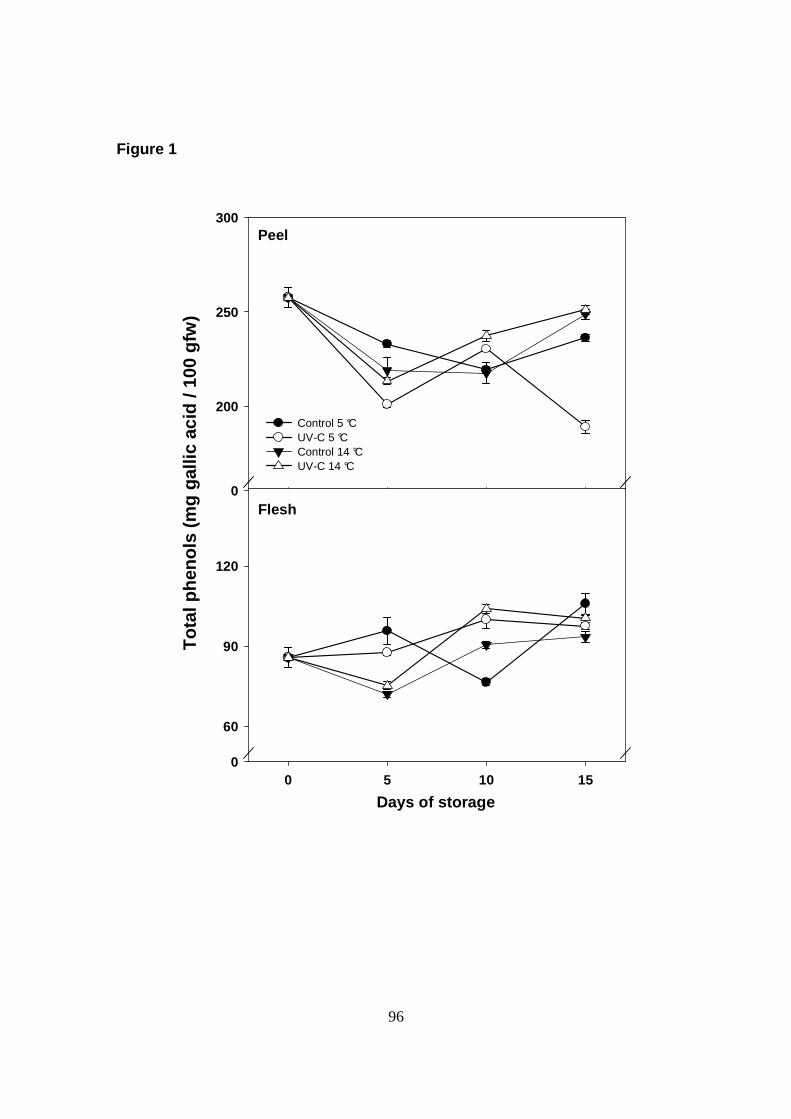
96
Figure 1
0 5 10 15
Tot
al p
heno
ls (
mg
galli
c ac
id /
100
gfw
)
0
200
250
300
Days of storage0 5 10 15
0
60
90
120
Control 5 °CUV-C 5 °CControl 14 °CUV-C 14 °C
Peel
Flesh
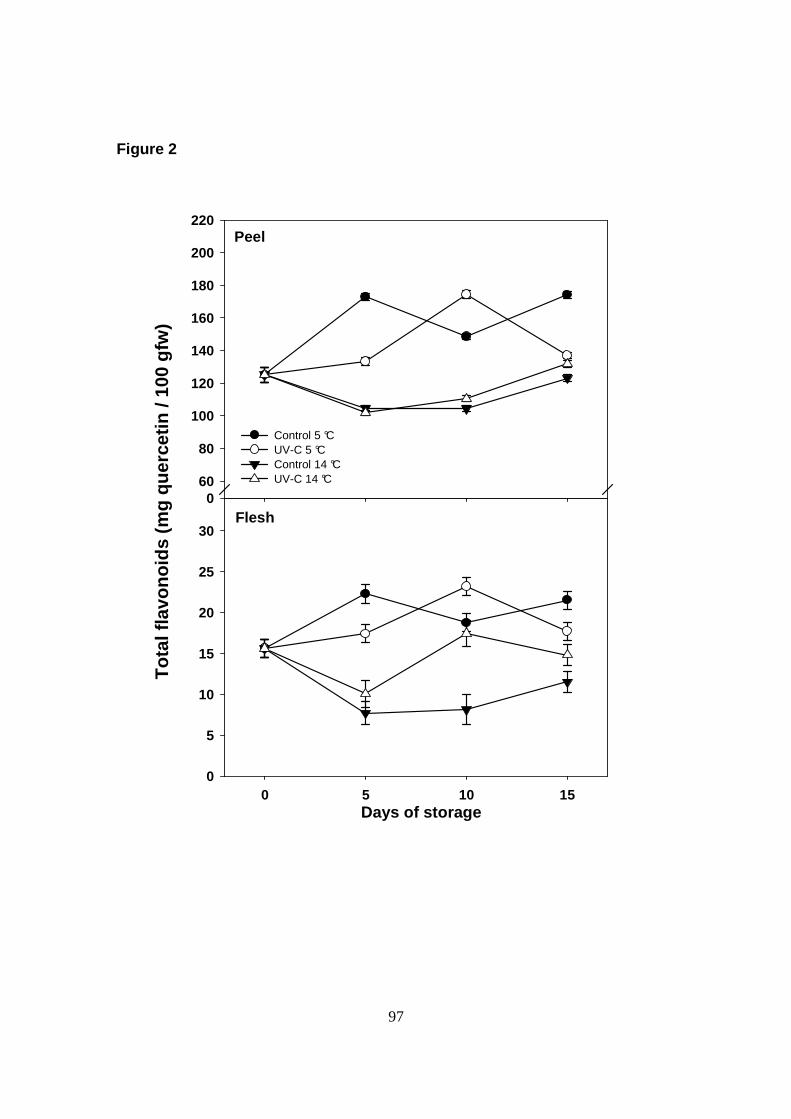
97
Figure 2
Days of storage
060
80
100
120
140
160
180
200
220
Control 5 °CUV-C 5 °CControl 14 °CUV-C 14 °C
Peel
0 5 10 150
5
10
15
20
25
30Flesh
Tot
al fl
avon
oids
(m
g qu
erce
tin /
100
gfw
)
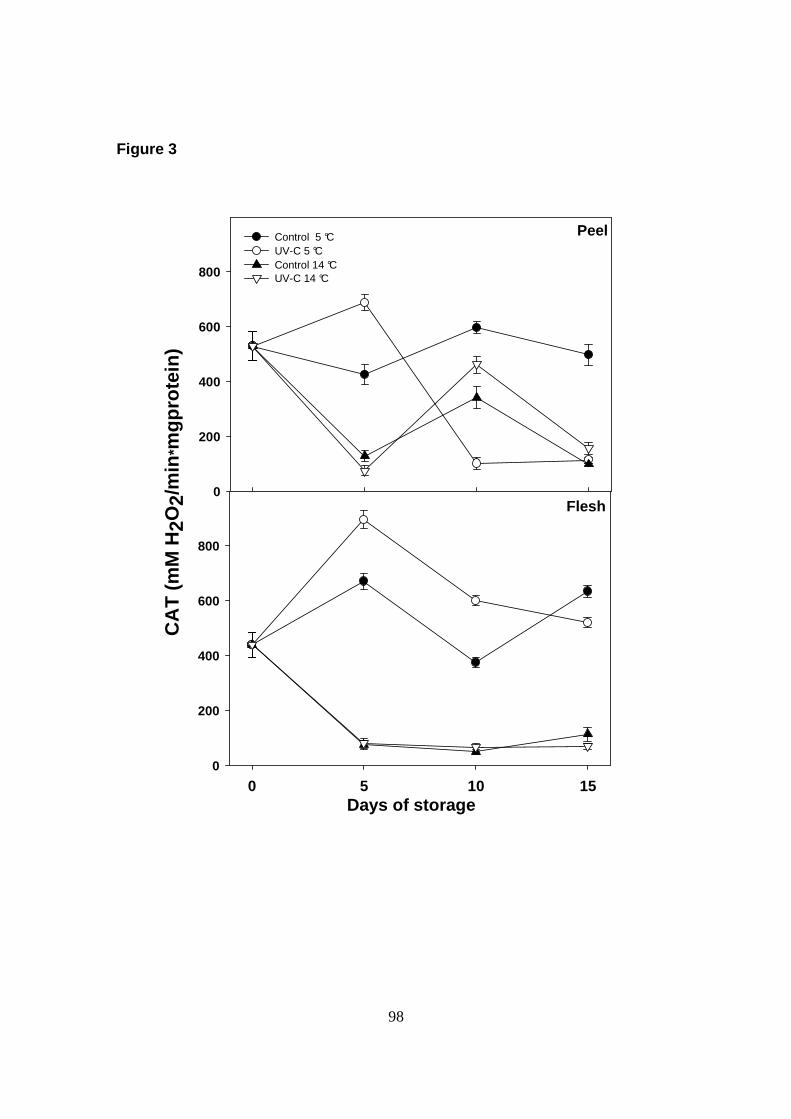
98
Figure 3
Days of storage
0 5 10 15
CA
T (
mM
H2O
2/m
in*m
gpro
tein
)
0
200
400
600
800
Control 5 °CUV-C 5 °CControl 14 °CUV-C 14 °C
Peel
0 5 10 150
200
400
600
800
Flesh
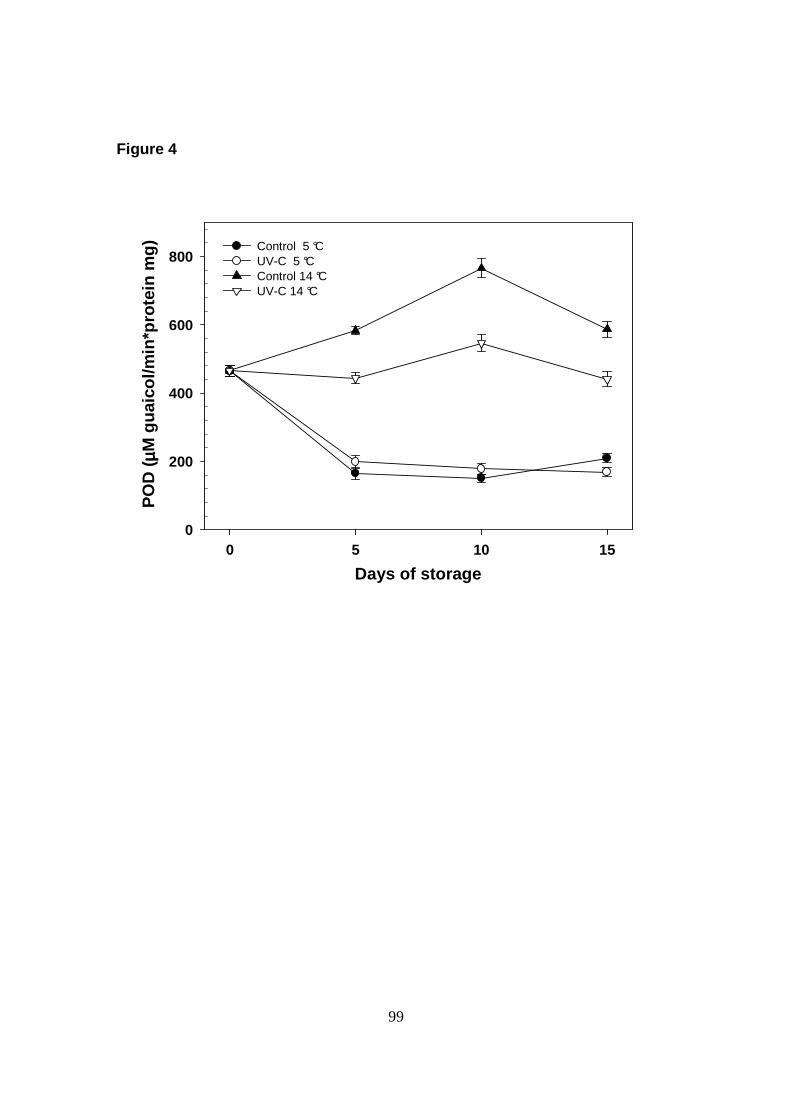
99
Figure 4
Days of storage0 5 10 15
PO
D (
µµ µµM g
uaic
ol/m
in*p
rote
in m
g)
0
200
400
600
800Control 5 °CUV-C 5 °CControl 14 °CUV-C 14 °C

100
Figure 5
Days of storage
0 5 10 15
SO
D (
Uni
ts/m
g pr
otei
n)
0
6
8
10
12
14
16Control 5 °CUV-C 5 °CControl 14 °CUV-C 14 °C
0 5 10 150
6
8
10
12
14
16
Peel
Flesh
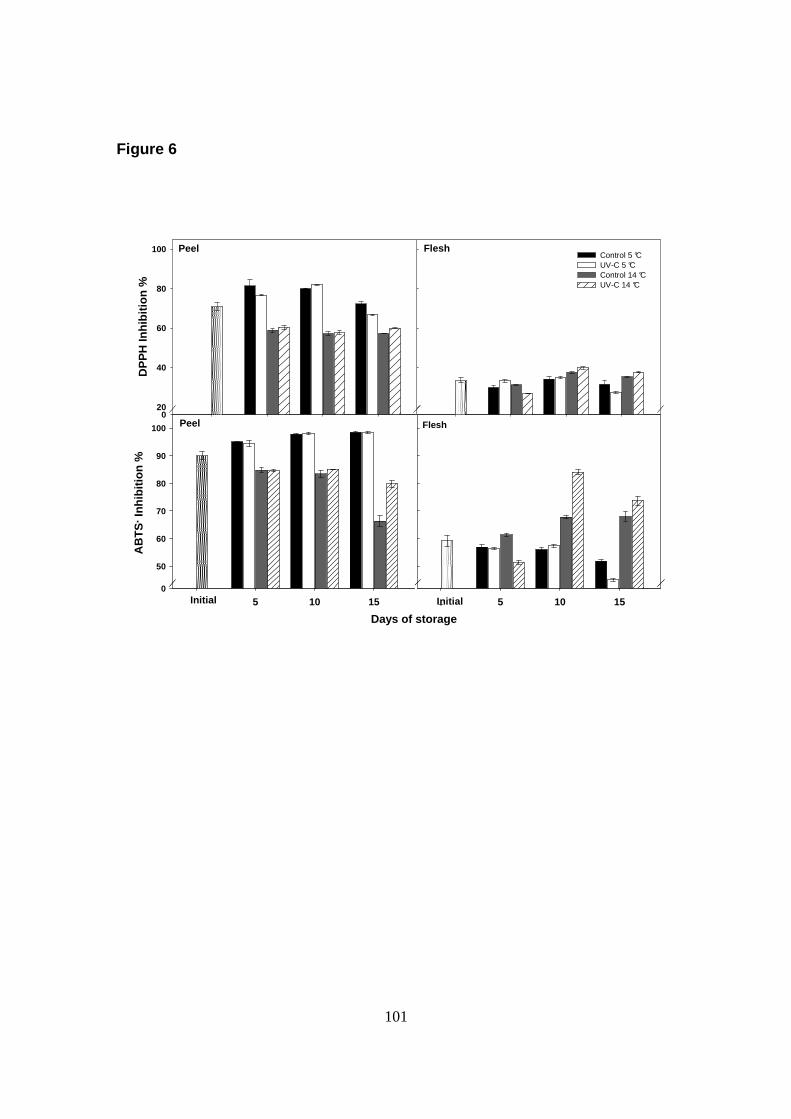
101
Figure 6
0 5 10 15
Days of storage
0 5 10 15
AB
TS
. Inh
ibiti
on %
0
50
60
70
80
90
100 Peel Flesh
DP
PH
Inhi
bitio
n %
020
40
60
80
100Control 5 °CUV-C 5 °CControl 14 °CUV-C 14 °C
Peel Flesh
Initial Initial

PRODUCCIÓN ACADÉMICA DERIVADA DEL TRABAJO DE TESIS

103
Artículos publicados
Rivera-Pastrana D M , Gardea Béjar A A, Martínez-Téllez M A, Rivera-
Domínguez M, González-Aguilar G A. 2007. Efectos Bioquímicos
Postcosecha de la Irradiación UV-C en Frutas y Hortalizas. Revista Fitotecnia
Mexicana. 30 (4):361-371.
Artículos aceptados y en revisión
Rivera-Pastrana D M , Yahia E M, González-Aguilar G A. 2010. Identification
of phenolic and carotenoid compounds in Carica papaya fruit by liquid
chromatography- mass spectrometry. Acta Horticulturae, Proceedings of VI
International postharvest symposium. In press.
Rivera-Pastrana, D.M. , Yahia, E., González-Aguilar, G.A. 2010. Phenolic
and carotenoid profiles of papaya (Carica papaya L.) and their contents under
low temperature storage. Journal of the Science of Food and Agriculture,
JSFA-10-0225. R1 (2nd round).
Artículos por enviar
Rivera-Pastrana, D.M. , González-Aguilar, G.A., Ayala-Zavala, J.F., Gardea-
Bejar, A and Sotelo-Mundo, R. 2010. Effect of UV-C irradiation and low
temperature storage on bioactive compounds, antioxidant enzymes and
radical scavenging activity of papaya fruit. Manuscrito para enviar al
Postharvest Biology and Technology.
Rivera-Pastrana, D.M. , González-Aguilar, Martínez-Téllez, M.A., Rivera-
Domínguez, M. 2010. Efecto de la irradiación UV-C y temperatura de
almacenamiento en la calidad postcosecha de papaya ‘Maradol’. Chapingo
Serie Horticultura.

104
Participación en congresos
Rivera-Pastrana D M , Gardea Béjar, A A, González-Aguilar, G A. “Efectos
postcosecha inducidos por irradiación UV-C en frutos de papaya (Carica
papaya) var. “Maradol” almacenados a bajas temperaturas”. IV Congreso del
Noroeste en Ciencias Alimentarias y Biotecnología (Noviembre, 2007).
Universidad de Sonora, Hermosillo, Sonora.
Rivera-Pastrana, D. M. , Yahia, E., González-Aguilar, G. A. “Identification of
phenolic and carotenoid compounds in Carica papaya fruit by liquid
chromatography- mass spectrometry”. 6th International Postharvest
Symposium. (Abril 8 - 12, 2009) Antalya, Turquía.
Rivera-Pastrana, D. M. , Yahia, E., González-Aguilar, G. A. “Effect of UV-C
irradiation treatment on antioxidant capacity of Carica papaya L. var. Maradol
fruit during cold storage”. Institute of Food Technologists 2009 annual
meeting (IFT09). Convention center, Anaheim, California (Junio 6-9, 2009).
Rivera-Pastrana, D. M. “Carotenoid and phenolic compounds of papaya“.
Symposium: The contribution of fruit and vegetable consumption in human
health. Facultad de Ciencias Naturales. Universidad Autónoma de Querétaro
(12 de Junio, 2009), Querétaro, Qro.


















