
CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C. · Objetivo general Objetivos específicos...
98
Transcript of CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C. · Objetivo general Objetivos específicos...



CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C.
UNIDAD DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR DE
PLANTAS
Regulación de la síntesis de alcaloides en
cumNos in vitro de Argemone mexicana
TESIS QUE PRESENTA
Q.B.B. MILDRED RUBÍ CARRILLO PECH
PARA OBTENER EL GRADO DE MAESTRÍA EN CIENCIASY BIOTECNOLOGÍA DE PLANTAS
MÉRIDA, yucATÁN, MÉXICO
2006•.: - .`.. .
a-\€.,_u
:: S::¡,^`
f-
(t=fflcy .¿,


DEDICATORIAS
A Dios, por todo lo que me ha permitido vivir.
A Miguel, por haberse cruzado en mi camino, por todo lo que representa en mi
vida y por todo el amor que me ha regalado.
A Alexa y Daniel, mis hijos, lo más valioso que Dios me ha dado.
A Lilí, mi madre, porque gracias a su ayuda y esfuerzo he llegado a este
momento de mi vida.
A Angela, Noemí y Eduardo, mis hermanos, por todo su cariño y comprensión.
A Manuela y Eduardo, mis abuelitos, por todos sus cuidados y sabios consejos.
A todos ustedes, gracias por creer en mi


RECONOCIMIENTOS
Este trabajo se realizó en la Unidad de Bioquímica y Biología Molecular de Plantas del
Centro de lnvestigación Científica de Yucatán, bajo la dirección del Dr Felipe Augusto
Vázquez Flota.
Parte de este trabajo fue fínancíado por el Consejo Nacional de Cíencia y Tecnología
(CONAcyT) a través del Programa lntegral para el Fortalecímiento del Posgrado
(P'FOP).
Se agradece la pamcipación del Dr. Peter Facchini del Departamento de Ciencias
Biológicas, Universidad de Calgary, Canadá, en la construcción de la genoteca de
ADNc.

Lista de abreviaturas
Cm
Alcaloides bencilisoquinolínicos
Ácido desoxirribonucleico complementario
Alcaloides mono-terpenoindólicos
Ácido naftalenoacético
Ácido ribonucleíco
Ácido ribonucleico mensajero
Alcaloides terpenoindólicos
Ríbonucleótido de trifosfato de adenosina
6-bencilaminopurina
Enzima del puente de berberina
Grado Celsius
Cromatografía de capa delgada
(S)-canadina oxidasa
Canadina sintasa
(S)-quelantifolina sintasa
Codeinona reductasa
Centímetro
CYP80BI W-metilcoclaurina 3-hidroxilasa
DEPC Dietil pirocarbonato
DMSO Dimetil sulfóxido
DBOX Dihidrobenzofenantridjna oxídasa
DRR 1,2-dehidrorretjculina reductasa
EDTA Ácido etilenodjamino tetraacético
9 Gramoh Hora
4-HFAA 4-hidroxifenilacetaldehído
ÍC Índice de crecimiento
L-DOPA 3,4-dihidroxifenilalanina
iii

11.3.1 Establecimiento y manejo de los cultivos celulares
11.3.2 Inducción de la síntesis de alcaloides en cultivos in vitro de A.mexicana
ll.3.2.A Efecto del régimen de iluminación sobre la síntesis de alcaloides encallos de A. mexicana
11.3.2.8 Efecto de la aplicación de MeJa sobre la acumulación de alcaloidesen cultivos en suspensión de A. mexicana
11. 4 Extracción, identificación y cuantificac'ión de alcaloides
11.5 Construcc.ión de una genoteca de ADNc a pahir de callos de A.
mexicana
Capítulo 111. Resultados y Discusión
111.1 Análisis del peml de acumulación de los alcaloides por CCD en
plantas de A. mex/.cana
111.2 Cuantificación de alcaloides en tejidos de A. mexí.cana
111.3 Efecto de la luz sobre el cultivo de callos
111.5 Acumulación de alcaloides
111.6 Establecimiento de los cultivos /.n v/.Íro
111.7 Efecto del MeJa en cultivos en suspensión de A. mex/.cana
111.7.1 Deteminación de la dosis de MeJa
111.7.3 Curso temporal de acumulación de alcaloides en cultivos inducidoscon MeJa
111,8 Construcción de una genoteca de ADNc de A. mex/.cana
Capítulo lv. Conclusiones y Perspectivas
Referencias
Apéndice A
il
27
28
29
32
35
35
39
42
45
49
51
52
55
58
61
65
77

TABLA DE CONTENIDO
Resumen
Abstract
lntroducción
Capítulo 1. Antecedentes
1.1 Descripción botánica de Argemor)e mex/.cana
1.2 Usos tradicionales de A. mex/.cana
1.3 Conocímiento fitoqui'mico e imponancia de A. mex/.cana
1.4 Ruta de biosíntesis de los ABl's
1.5 Enzi.mologja de la síntesis de los ABl's
1.5.1 Formación de norcoclaurina
1.5.2 Formación de reticulina
1.5.3 Formación de los alcaloides tipo benzofenantridina y protoberberina
1.5.4 Formación de los alcaloides tipo morfina
1.6 Regulación de la síntesis de ABl's
1.7 Producción de alcaloides en cultivo de tejidcts
1.7.1 Fuente de carbono
1.7.2 Fuente de fosfatos
1.7.3 Adición de precursores
1.7.4 Adición de reguladores del crecimiento vegetal
1.7.5 Aplicación de inductores
1.7.6 Efecto de la luz
Justificación
Objetivo general
Objetivos específicos
Capítulo 11. Materiales y Métodos
11.1 Diseño experimental
11.2 Colecta de plantas silvestres de A mex/.cana
11.3 Cultivos í.n v/'fro
1
3
3
3
4
4
5
5
7
8
10
12
16
17
18
18
18
20
22
23
24
24
25
25
25
26

STOX
STR
STS
TPNMT
TDC
TyDC
UV
VIH
(S)-tetrahidroprotoberberina oxidasa
Estrictosidina sintasa
Salutaridina sintasa
Tetrahidroprotoberberina ci.s-W-metiltransferasa
Triptofano descarboxilasa
Tirosina descarboxilasa
Ultravioleta
Virus de inmudeficiencia humana

Lista de figuras
Figura l.1 Ruta general de la biosíntesís de ABl's
Figura l.2 Etapas tempranas de biosíntesis de ABl's
Figura l.3 Ruta de biosíntesis de los alcaloides tipo benzofenantridina ytipo protoberberina
Figura l.4 Ruta de biosíntesis de los alcaloides tipo morfina
Figura ll.1 Diseño experimental del proyecto
Figura ll.2 Diseño experimental seguido para la inducción química de loscultivos de A. mex/.cana.
Figura ll.3 Curva de calibración promedio de berberina en el sistema dedisolventes J (n-butanol:ácido acético:agua 7:1 :2)
Figura ll.4 Curva de caljbración promedio de sanguinarina en el sistema dedisolventes J (n-butanol:ácido acético:agua 7: 1 :2)
Figura lll.1 Cromatogramas de extractos de diferentes tejidos de A.mexicana
Figura lll.2 Análisis de los alcaloides deA. mex/.cana por ccD de dosdimensíones
Figura lll.3 Cultivos de callos de A. mex/.cana en medjci pc
Figura lll.4 Crecimiento de callos deA. mex/.cana a lo largo de un ciclo decultivo de 50 días en presencia de luz evaluado como PF (.) yPS (D)
Figura lll.5 Crecimiento de callos deA. mexí.cana a lo largo de un ciclo decultivo de 50 días en oscuridad evaluado como PF (.) y PS (D)
Figura lll.6 Aspecto de las células en suspensión deA. mex/.canamantenidos en condiciones de luz continua a los 14 días decultivo en el medio 85 (A) y PC (8)
Figura lll.7 Aspecto de la suspensión celulardeA. mex/.cana (A, B, C y D) yde los medios libres de células (A', 8'. C' y D')
Figura lll.8 Células en suspensión de cinco días de cultivo expuestas aMeJa 100 HM durante 0 (A); 6 (8); 12 (C), y 24 (D) h.
Figura lll.9 Aspecto de una caja de petri usada para realizar la {itulación dela biblioteca de ADNc de A. mexí.cana
31
31
38
39
43
44
44
50
59

Cuadro 1.1
Cuadro 111.1
Cuadro 111.2
Cuadro 111.3
Cuadro 111.4
Cuadro 111.5
Cuadro 111.6
Cuadro 111.7
Cuadro 111. 8
Cuadro 111.9
Cuadro 111.10
Cuadro 111.11
Lista de cuadros
Enzimas de la ruta de bíosíntesis de ABl's cuyos genescorrespondientes han sído clonados en diferentes especies
Valores de Rf encontrados con las mezclas de disolventesempleados en este trabajo para la separación de losalcaloides en tejidos de A. mexi.cana
Contenido de berberina y sanguinarina en diferentes tejidosde A. mex/.cana repohados en Lig/g de PS
Parámetros de crecimiento de los callos de A. mex/.canacultivados bajo luz continua o en oscuridad
Contenido de sanguinarina en callos cultivados durante 50días en luz continua u oscuridad
Comparación del patrón de manchas fluorescentesobservado en cultivos de callos mantenidos bajo luzcontinua y en oscurídad total
Crecimiento evaluado como PF por matraz, de cultivos deA. mex/.cana mantenídos en medios líquidos
Efecto del MeJa sobre la acumulación de sanguinarina encultivos en suspensión de A. mex/.cana
Productividad de sanguinarina del cultivo de A. mex/.canaexpuesto a diferentes concentraciones de MeJa y DMSO
Curso temporal de la acumulación de sanguinarina encultivos en suspensión de A. mex/.cana tratadas con MeJa100 uM
Productividad de sanguinarina en cultivos de A. mex/.canaexpuestos a 100 uM de MeJa durante diferentes tiempos
Valores de RÍ del patrón de compuestos fluorescentes a365 nm, no identificados, observado en cultivos de célulasen suspensión en respuesta a diferentes concentracíonesde MeJa
vii
40
51
53
55
56
57
58

RESUMEN
Angemone mex/.cana es una planta Papaverácea que se ha utilizado en la medicina
tradicional desde tiempos prehispánicos por sus propiedades para curar diferentes
afecciones. Esta planta produce alcaloides del tipo bencilisoquinolínicos los cuales
tienen diferentes efectos fisiológicos y es probable que sus propiedades medicinales
se deban a estos compuestos. Sin embargo, los estudios que se han realizado en este
contexto son pocos, entre otros factores debido a que es una planta snvestre, lo cual
dificulta la dísponibilidad del material vegetal para la obtención de los productos
naturales de interés. Una alternativa para obtener estos compuestos es el cultivo de
tejidos vegetales bajo diferentes condiciones de induccíón. En este trabajo se
ensayaron diferentes condiciones de Íluminación y la exposición a jasmonato de
metilo. El régimen de iluminación tuvo un efecto sobre la capacidad de acumular
sanguinarina. Además, dependiendo de si los cultivos se mantuvieron en oscuridad o
luz continua se notaron algunos cambios en el patrón cualitativo de los alcaloides. El
tratamiento de los cultivos con 100 uM del inductor produjo un aumento de cerca del
200% de sanguinarina, respecto a los cultivos no expuestos después de 24 h. En
ningún caso se observó la presencia de berberina en los cultívos /.n v/tro, a pesar de
que este alcaloide se detectó en diferentes tejidos de plantas colectadas en campc). La
identificación de los alcaloides, así como su cuantíficación se llevaron a cabo mediante
criteríos de cromatografía de capa fina.
Por último, se construyó una genoteca de ácido desoxírribonucleico complementario
utilizando cultívos de callos de seis días. El título de la biblioteca fue del orden de 109
unidades formadoras de colonias/mL. Esta genoteca es útil debido a que contiene los
genes involucrados en las rutas de biosíntesis de los alcaloides bencílisoquinolínicos,
por lo que es una herramienta molecular lmportante para el estudio de las rutas de
biosíntesis de alcaloides en A. mexí.cana.
viii

ABSTRACT
Argemone mexí.cana (L) is Mexícan natíve plant belonging to the Papaveraceae family.
Because the medicínal properties attributed to it, this plant has been used to treat
different diseases since pre-Hispanic times. ln fact, tissues of this plant accumulate
diverse benzylísoquinoline alkaloids with physiological effects and which may explain
its medicinal properties. Nevertheless, there are only few studies on the biosynthesis
of such alkaloids in this plantt perhaps because it is an annual plant, growing in the
wild, which may limit the availability of fresh tissues to perform biochemical and
molecular analysis. /n v/.Íro cell cultures represent an alternative for the continuous
supply of tissues to perform such studies.
Different conditions were assayed to induce the synthesis of benzylisoquinoline
alkaloids in /.n v/.Íro cultures of Argemone mex/.car)a. Conditions included illumínation
regime and the exposure to methyl-jasmonate. Sanguinarine accumulation was
affected by the llluminatjon regime, and depending if cultures stayed in the dark or
under continuous light, some changes noticed Ín the qualitative pattern of the alkaloids.
Treatment of the cultures with methyl-jasmonate 100 tiM produced a 200% increase of
sanguinarine, with respect to non exposed cultures, after 24 h. Despite the culture
conditions, berberine, which was identified in different tissues of plants, was not
detected in /.n v/.Íro cultures. Alkaloids identification and quantification was carried out
based on criteria of thin layer chromatography.
A ljbrary of cDNA was constructed, using six-day old cultures. The title of the library
was in the order of 109 plate forming unit/mL This important molecular tool is useful for
containing the genes involved en biosynthesís pathway of benzylisoquinoline alkaloids.


lNTRODUCCIÓN
A lo largo de su ciclo de vida, las plantas están expuestas a diferentes condicionesambientales de estrés tales como la baja disponibilidad de agua y nutri.entes, la
temperatura y el ataque de patógenos, entre otras. Como resultado de su interaccióncon el medio ambiente, las plantas producen una serie de productos naturales que les
permíten adaptarse o defenderse; algunos de éstos pueden inducir respuestasfisiológicas en organi.smos diferentes de los que los producen, incluyendo el alivio de
algunos tipos de cáncer como la enfermedad de Hodkings. Es por esto que en losúltimos años, se han realizado estudios para tratar de conocer cuáles son los factores
que influyen en la biosíntesi.s de algunos de estos metabolítos en las plantas.
Las plantas penenecientes a la familia de las Papaveráceas tienen la capacidad desíntetizar una gran variedad de metabolitos secundarios, principalmente alcaloides, los
cuales tíenen actividades fisi.ológicas muy potentes como narcóticos, sedantes yantítusivos.
En la historia de la medicína y terapéutíca tradicional de muchas de las culturas
antiguas del mundo, existen evídencias del uso de plantas que pertenecen a la fami.li.a
de las Papaveráceas de la cual Argemone mexí.cana es una representante que se hautili.zado en la medicina tradicional mexicana para diferentes afecci.ones. Exjsten
algunas evidencias que sugieren que las propiedades medicinales que se le atribuyena esta planta se deben a la presencia de alcaloides bencilisoquinoli.nicos (ABl's) en
sus tejidos.
Debido a que Arigemone mex/.cana es una planta si.lvestre, la disponibilidad de materi.al
vegetal depende de las condiciones ambientales. Por otro lado, pocos han sido los
estudios que se han realizado en esta planta en el contexto de estudi.ar la regulaciónde la biosíntesis de los ABl's, así como de encontrar maneras alternativas deobtención de estos compuestos usando técnicas como el cultivo de tejidos vegetales,
por lo que en este trabajo se presenta un estudío sobre la síntesis de alcaloides enesta planta, incluyendo el establecimiento y caracterización de cultivos /.n v/.fn) de los
cuales se puedan obtener estos compuestos de i.nterés


CAPITUL0 I
ANTECEDENTES
1.1. Descripción botánica de Argemone mex/.cana
A. mexí.cana (Papaverácea) es una hierba anual que míde entre 30 y 90 cm de alto.Su tallo es solitario, glauco y ramoso en la base, con espinas escasas perpendiculareso ligeramente reflejas; sus hojas son alternas, sésiles y glaucas, lobuladas, deaproximadamente s cm de ancho y de 6 a 20 cm de largo. Las hojas basales sonovalolanceoladas y las superiores elípti.cas u ovobadas y espinosas. Sus flores
pueden ser amarillas o blancas de 3 a 7.5 cm de di.ámetro, las cápsulas tienen de 4 a6 carpelos y son dehiscentes por el vértice (Lozoya eí a/., 1982). Esta planta es
originaría de América y prefiere los cli.mas semicálidos, semisecos y templados desdeel nivel del mar hasta los 2,750 msnm. Crece en terrenos abandonados, en zonasagrícolas o a orillas de los caminos (Argueta eí a/.,1994). En Méxi.co, esta planta es
considerada como una maleza por los agricultores y está ampliamente distribuida en
los estados de Baja California, Michoacán, Morelos, Nuevo León, Oaxaca, Puebla,San Luis Potosí, Tlaxcala, Veracruz, Yucatán y Zacatecas (Lozoya eí a/.,1982)
1.2. Usos tradicionales de A. mex/.cana
En México A. mexí.cana fue utilizada por las culturas prehispánlcas con finesmedicinales, debido a sus propiedades sedantes, narcóticas y antitusivas. En las
antíguas culturas se usaban diferentes partes de la planta para curar afecci.ones talescomo las cataratas y la inflamación de los ojos, para lo cual se utilizaba el látex.
También era usado para alivjar la fiebre, para curar las Úlceras sexuales, la sarna,
como purgante, como diurético y en ciertas enfermedades cutáneas como lasverrugas. La presencia de alcaloides derivados de la tirosina en los tejídos de esta
planta podrían explicar las propíedades medicínales que se le atribuyen. Sin embargo,algunos de los compuestos identificados en esta planta incluyen alcaloides tóxi.cos,
como la sanguinarina, que se encuentra en las semillas y cuyos efectos secaracterizan por el dolor muscular, edema generalizado, aparición de sarcoides.
diarreas abundantes, y en casos extremos, aumento de la presi.Ón intraocular queconduce a la formación de glaucoma. Sin embargo, es importante mencíonar que
estos efectos solo se presentan cuando el alcaloide es consumi.do en dosjs altas, abajas concentraciones este compuesto es usado de manera comercial enformulaciones de dentríficos debido a sus propiedades como antibi.Ótico.

1.3. Conocimiento fitoquímico e importancia de A. mex/'cana
Las plantas del género Argemone sintetizan alcaloides del tipo benc.ilisoquinolínico
que incluyen entre los más abundantes a la berberina y la protopina. Otros alcaloides
presentes en menores cantidades .incluyen la coptisina, la sanguinarina, laalocriptopina, la queleritrina y la dih.idroqueleritrina (Lozoya eí a/.,1982).
Los alcaloides que producen estas plantas tienen diferentes efectos fisiológicos, según
los estudios realizados en famacología animal. La alocriptopina, protopina, berberina
y sanguinarina, entre otros, poseen la propiedad de modificar la función cardíaca,estimulando el tono y la fuerza de contracción lo que conduce a prolongar el período
refractario. Particularmente, la alocriptopina, posee propiedades antiarrítmicas que
han sido evaluadas como semejantes a las de ciertos compuestos usados en eltratamiento de la fibrilación auricular (Lozoya ef a/.,1982). Por otro lado, la protopina,
también actúa sobre la musculatura uterina con una potencia simnar a la de laoxitocina. La berberina también produce efectos simnares en animales, además de
que tiene propiedades antibióticas, por lo que se ha postulado su utilidad en eltratamiento de la leishmaniasis. Por último, también se ha descrito que la berberina
tiene propiedades coleréticas, por lo que se observa una disminución en los niveles de
bilirrubina y un aumento del volumen bil.iar a las pocas horas de haber suministrado el
producto por vía oral a pacientes con colecistitis crónica (Lozoya eí a/ ,1982).
Las semillas de Argemone contienen un 93% de ácidos grasos, de los cuales el 80%
corresponden a ácidos insaturados (linoléico, oléico, ricinoléico, palmitoléico y
linolénico), y el resto lo constituyen los ácidos saturados palmítico y esteárico (Lozoya
eí a/.,1982). Por sus altos contenidos de aceite en las semillas, la planta presenta un
buen potencial de explotación industrial con fines alimenticios y cosméticos; sin
embargo, el alto contenido de compuestos tóxicos ha impedido su utilización. Por otrolado, de las flores se han aislado tres derivados flavonoides: la isoharmnetina, el
glucósido de 3-isoharmnetina, y el glucósido de 7-isoharmnetina, aunque también sehan identificado otros flavonoides.
1. 4. Ruta de biosíntesis de los ABl's
Los ABl's son un grupo de metabolitos secundarios farmacológicamente activos quese encuentran predominantemente en las familias Berberidaceae, Fumariaceae,Papaveraceae, Menispermaceae y Ranunculaceae. Hasta ahora se han identificadoalrededor de 2,500 productos con este tipo de estructura, por lo que representan uno
de los grupos de alcaloides más numeroso (Facchini eí a/., 2000).
4

Algunos de estos compuestos presentan activídad biológica; por ejemplo, la morfina
es un potente analgésico, mientras que la tubocurarina es un bloqueadorneuromuscular (Facchini eí a/„ 2000).
Dada la imponancia comercial de algunos de los alcaloídes de este grupo, las rutasmetabólicas involucradas en su biosíntesis han sido estudiadas con detalle y en los
últimos años se han dilucidado algunas de éstas. Sin embargo, el conocimiento sobrelos mecanismos moleculares y los controles bioquímicos es limitado, debido a que
solo recientemente se han aplicado técnicas moleculares como herramientas para el
estudio de los mecanismos que las regulan. En este sentido, se han realizadoimponantes estudios en especies como la Amapola (Papaver somn/.Íewm) y laAmapola de California (Eschscho/zÍ.a ca/Í'fomí'ca) (Facchini ef a/. , 2000).
1.5. Enzimologia de la síntesis de los ABl's
De manera general, la ruta de biosíntesis de los ABl's se puede dividir en tres etapas.
En la etapa inicial, dos moléculas de tirosina, el aminoácido que da origen a todos los
alcaloides de este tipo, experimentan una serie de reacciones para producir dopamina
y 4-hidroxifenilacetaldehído (4-HFAA), dos intermediarios que se condensan paraformar norcoclaurina. Este compuesto es considerado como el intermediario central de
todos los alcaloides de este tipo (Fig. 1.1). La segunda etapa corresponde a la
formación de reticulina, el último intermediario común en la si'ntesis de todos los
alcaloides de esta ruta. En la etapa final, la ruta se ramifica para producir los tres
principales tipos de alcaloides de este grupo: los tipos benzofenantridina,
protoberberina y morfinano (Fig.1.1).
1.5.1. Formación de norcoclaurina
La bjosíntesís de los ABl's comienza con una seríe de descarboxilaciones, orto-
mdroxilaciones y desaminaciones que conviehen a la tirosina en dopamina y 4-HFAA.
La tírosina descarboxilasa (TyDC) es la enzima responsable de la formación de
tjramina y/o dopamina a partir de la L-tirosina y de su derivado dihidroxilado, la 3,4-
dihidroxifenilalanina (L-DOPA), respectivamente (Fig. 1.2). El siguiente paso es la
condensación de la dopamína con el 4-HFAA por la acción de la norcoclaurina sintasa
(NCS) para producír (S)-norcoclaurína, el cual es el precursor central de todos losABl's (Stadler eí a/„ 1989).

mfiff±wfin2."......""-wffL_",o8ina
'-cooii TyDC':',, -:
Sangu¡nar¡na
",amlna
'::mmz
Dopamina
:, (Oft::::
H,CO
BBE Ho
(Benzofenantridlna) (P[otoberberina) morfinano)
Figura 1.1. Ruta general de la biosíntesis de ABl's TyDC: tirosina descarboxilasa,
NCS: Norcoclaurina sintasa, BBE: Enzíma del puente de berberina.

HOJcrT:OOHL.Tlroslna
1
mzcooHLl)OPA
#
TyDC
TyDC
CH, 4'OMT=
Momm,"rsmlna
'::mH¡
DOpamina
";;g:
HOJH4_H FAA
NCS Ho
cHs CypSOB1= Ej
CH, NMT-";:#HWS.Reticulln a S-'H id roxlw-metll coclau.l na SW. Metl lcoclau.lna S-Cocl au rlna
Figura 1.2. Etapas tempranas de biosíntesis de ABl's. TyDC: Tirosina descarboxilasa.
NCS: norcoclaurina sintasa, 6'OMT: 6-O-metiltransferasa. NMT.. N-metiltransferasa,
CYP80B1 : N-metilcoclaurina 3-hidroxilasa y 4'OMT: 4-O-metiltransferasa.
1.5.2. Formación de reticulina
La (S)-norcoclaurina es convertida a (S)-retículina por la acción secuencial de una 6-
O-metiltransferasa (6'OMT) (Frick eí a/„ 1999), una W-metíltransferasa (NMT) (Frenzel
eí a/.,1990), una hidroxilasa dependiente del citocromo P450 (Pauli eí a/.,1998), y
una 4-O-metiltransferasa (4'OMT) (Frenzel ef a/., 1990). De estas enzimas, ya han
sido purificadas la 6'OMT y la 4'OMT en cultivos de CopÍÍ.s/.aponí.ca (Sato eí a/„ 1993),
y también ya han sido aislados y caracterizados los ADNc correspondientes(Morishige eí a/., 2000). Se sabe que estas enzimas tienen diferentes especificidadespor sus sustratos, entre los cuales se incluyen catecoles simples, fenilpropanoides yvarios ABl's. Lo anterior sugiere que algunas de las isoformas de estas enzimas esténinvolucradas en la biosíntesis de los fenilpropanoídes, indicando un posible origen
evolutivo común.
La (S)-reticulina es el punto intermedio en la ramificación de la ruta de biosíntesis para
muchos de los ABl's (Fig.1.3). Se han estudiado las rutas de ramificación que llevan a

la biosíntesis de alcaloides del tipo benzofenantridina, tales como la sanguinarina y lamacarpina, así como la que conduce a los alcaloides tipo protoberberina, como laberberina y la palmatina (Hashimoto eí a/., 1994). También se conoce de manera
detallada, la ruta de los alcaloides tipo morfina, de los cuales la moriina y la codeína
son los más conocidos (Facchini eí a/.,1998). Muchas de las enzimas involucradas en
estas rutas han sido purificadas, así como los ácidos desoxirr.ibonucleicos
complementarios (ADNc) correspondientes han sido clonados (Cuadro 11).
1.5.3. Formación de los alcalo.ides tipo benzofenantridina y protoberberina
El primer paso en la biosíntesis de los alcaloides de los tipos benzofenantridina,
protoberberina y protopina involucra la conversión del grupo W-metilo de la (S)-reticulina en un puente de metileno de (S)-esculerina por la enzima del puente de
berberina (BBE) (Fig. 1.3). Esta conversión es única en la naturaleza y no ha sido
posible llevarla a cabo por síntesis orgánica. Esta enzima se ha purificado de célulasen suspensión de Berberí.s beaní.ana (Steffens eí a/., 1985) y los ADNccorrespondientes han sido aislados de E. ca//.forníca (Dittrich eí a/.,1991) y Berben.s
sÍo/on/.fera (Chou eí a/., 1998). Los genes BBEl ya han sido aislados de P.
somn/.fewm (Facchini eí a/„ 1996) y E. ca//.Íorn/.ca (Hauschild eí a/„ 1998).
La (S)-esculerina, producto de la BBE, puede ser convehida a (S)-estilopina por dos
oxidasas dependientes del citocromo P450: la (S)-quelantifolina sintasa (CFS) y la
(S)-estilopina sintasa (SPS), lo cual da por resultado la formación de dos gruposmetilenodioxi. Ambas enzimas han sido caracterizadas de E. ca//.forr)/.ca (Bauer, eí a/„
1989 y 1991). La (S)-estilopina es N-metilada por una metiltransferasa que ha sido
purificada de células de Sanguí.nar/.a canadens/.s (O'Keefe eí a/.,1994), así como deCooÍc/a/i.s vag/.nans y de E. ca//.fom/ca (Rueffer ef a/.,1990). El paso de W-metilación es
seguido por una reacción catalizada por una tercera monooxigenasa dependiente delcitocromo P-450, la (S)-c/.s-W-metilestilopina-14-hidroxilasa (MSH), la cual cataliza la
formación de protopina. Esta enzima ha sido aislada de cultivos de C. vag/.nans
(Rueffer eí a/., 1987). La conversión de protopina a sanguinarina involucra unahidroxilación por parte de otra enzima dependiente de citocromo P450, la protopina 6-
hidroxHasa (PPH), seguida de un rearreglo intramolecular espontáneo para dar como
producto a la clihidrosanguinarina, el primer alcaloide con núcleo de benzofenantridina
(Tanahashi eí a/., 1990). La dihidrobenzofenantridina oxidasa (DBOX), una enzimacitosólica originalmente aislada de cultivos de E. ca/i.fom/.ca (Schumacher eí a/ ,1988),
cataliza la oxidación de dihidrosanguinarina a sanguinarina (Fig 1.3).

Por su parte, las enzimas que catalizan los dos pasos finales en la biosíntesis demacarpina, el alcaloide tipo benzofenantridina que posee el más alto estado de
oxidación en la naturaleza, son dihidroquelirrubina-12-hidroxilasa (una monooxigenasa
dependiente del citocromo P450) y la 12-hidroxidihidroquelirrubina-12-O-
metiltransferasa dependiente de la S-adenosil-metionina (SAM), las cuales fueron
descubiertas en cultivos de rha/Í.cíwm bu/garí.cum inducidos con levaduras (Kammerer
eí a/.,1994) (Fig.1.3).
En algunas familias de plantas como las Berberidáceas y Ranunculáceas, la
biosíntesis de berberina se inicia con la metilacíón de (S)-esculerina, para producir (S)-
tetrahidrocolumbamina. La reacción es catalizada por una enzima dependiente de
SAM, la esculerina-9-O-metiltransferasa (SOMT), la cual ha sido purificada de células
de C. /.apon/.ca (Sato eí a/., 1994). EI ADNc correspondiente también ha sido aislado
(Takeshita eí a/., 1995). El siguiente paso involucra la formación de un puentemetilenodioxi (Galneder eí a/., 1988; Hashimoto eí a/„ 1994) en una reacción de
desmetilación que originalmente se pensó que era catalizada por una peroxidasa noespecífica. Sin embargo, la enzima canadina sintasa (CDS) dependiente del citocromoP450, fue detectada en miembros del Tha//.cíwm y CopÍ/.s y se identificó como la
responsable de la formación del puente metilenodioxi de la (S)-tetramdrocolumbamina;
sin embargo, esta enzima no es la responsable de la misma reacción observada parala formación de columbamina, un alcaloide cuaternario (Fig.1.3), (Rueffer eí a/.,1994).
Por esta razón, se dedujo que la biosíntesis de berberina no procede por la vía de la
columbamina, como se había propuesto originalmente. La (S)-canadina, también
conocida como (S)-tetrahidroberberina, es oxidada a berberina ya sea por la (S)-canadina oxidasa (CDO) o por la (S)-tetrahidroprotoberberina oxidasa (STOX) (Fig.
1.3) (Amman ef a/., 1986). Aunque estas enzimas catalizan la misma reacción, sus
propiedades bioquímicas son bastante diferentes. La STOX de Benber/.s es unaproteína flavinilada con un amplio rango de preferencia por su sustrato, en tanto que laCDO de CopÍ/.s y rha//.círum que contiene hierro (no flavina), procede mediante unmecanismo diferente y acepta preferentemente a la (S)-canadina como sustrato,
(Hashimoto eí a/.,1994).

BemoG3nantr"Ina
Dlhldrosangulna.lna Sanguinarim
Figura 1.3. Ruta de biosíntesis de los alcaloi.des tipo benzofenantrídína y tipo
protoberberina. BBE: enzima del puente de berberina, CFS: quelantifolina síntasa,SPS: estilopina sintasa, TPNMT. tetrahidroprotoberberina c/.s-N-metiltransferasa,
MSH: W-metilestilopina-14-hídroxilasa, PPH: protopina 6-hidroxilasa, DBOX:
dihidrobenzofenantridina oxidasa, SOMT: esculerina-9-O-metiltransferasa, CDO.
canadina oxidasa y STOX: tetrahidroprotoberberina oxidasa.
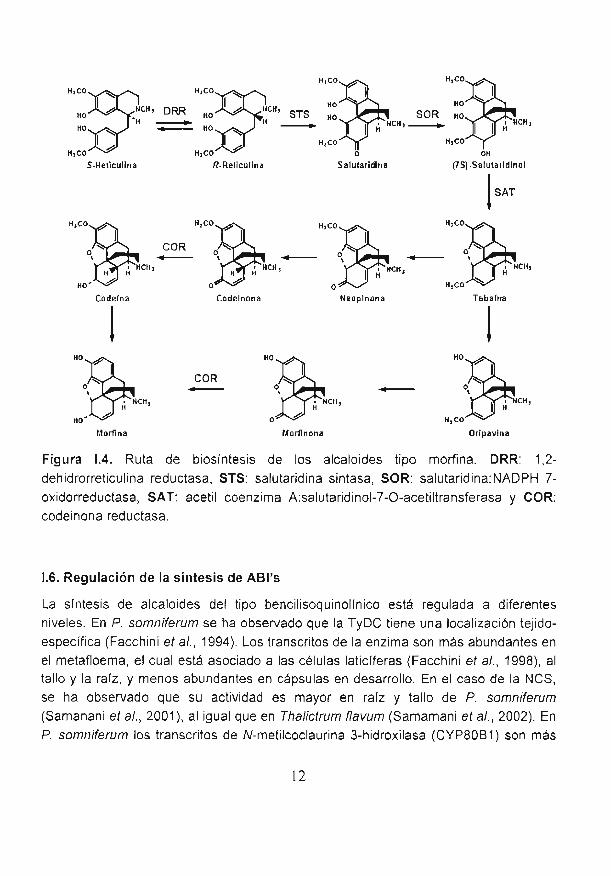
1.5.4. Formación de los alcaloides tipo morfina
En algunas especies del género Papaver se ha observado que la conversión de (S)-reticulina a su enantiómero (la forma R) representa el primer paso de biosíntesis de
los alcaloides de tipo momna (Fig. 1.4). La 1,2-dehidrorreticulina reductasa (DRR), una
enzima citosóli.ca dependiente de fosfato de dinucleótido de nicotinamida y adeninareducido (NADPH), es la que cataliza la i.educción estereoespecífica del 1,2-
dehidrorreticulina a (R)-reticulina; esta enzima que solamente ha sido encontrada en
plantas que sintetizan alcaloides del tipo morfinano, ha sido purificada de amapola
10

(De-Eknambul eí a/., 1992). El siguiente paso es un acoplamiento intramolecularcarbono-carbono de un fenol con la (R)-reticulina por la acción de la salutaridína
sintasa (STS), una enzima dependiente del citocromo P-450, para producirsalutaridina (Gerardy eí a/., 1993); la salutaridina se reduce a (S)-salutaridinol
mediante la salutaridina:NADPH 7-oxidorreductasa (SOR), una enzima citosólica
solamente encontrada hasta ahora en amapola y Papaver bracíeaíum (Gerardy eí a/.,1993). La transformación del (S)-salutaridinol en tebaína (un alcaloide del típo
morfinano) Ínvolucra el cierre de un puente de óxido entre el C-4 y el C-5 por la
enzima acetil coenzima A:salutaridinol-7-O-acetiltransferasa (SAT). Esta enzima ha
sido purificada a pahir de células de amapola (Lenz eí a/.,1995), y se ha detectado
también en P. bracíeaíum, pero no en aquellas especies que no producen alcaloidesdel tipo de la morfina (Fig. 1.4).
En los pasos restantes de la mayoría de las rutas que llevan a la producción demorfina, la tebaína es convertida a codeinona y ésta es reducida a codeína. Por últimola codeína es desmetilada para producir morfina. La codeinona reductasa (COR), que
cataliza la reducción dependiente de NADPH de (-)-codeinona a (-)-codeina (Fig.
1.4), ha sido purificada recientemente, y el ADNc correspondiente ha sido aislado de
amapola (Unterlinner eí a/., 1999). Cuatro ADNc que codifican para diferentes
isoformas de COR han sido clonados y expresados en E. co//.. Las cuatro isoformastienen propiedades catalíticas similares. La COR tiene un 53% de identidad de
aminoácidos con la 6'-desoxíchalcona sintasa de soya que partícipa en la ruta debiosíntesis de los fenílpropanoides, lo cual sugiere una conexión evolutiva entre las
enzimas de la ruta de biosíntesis de los fenilpropanoides y los alcaloides (Frick eí a/.,
1999; Unterlinner eí a/., 1999). Ambas enzimas son miembros de una familia de
aldo/ceto oxidorreductasas dependientes de NADPH encontradas en varias rutas del
metabolismo primario.
m
#HCtl¡ DFü
= - r± - :=- - - =Cod,eína
::,S"cH'Morfina
COR=
Neoplnona
H:offi-,
Morífnona
-\0'ipavína
Figura 1.4. Ruta de biosíntesis de los alcaloides tipo moriina. DRR: 1,2-
dehidrorreticulina reductasa, STS: salutaridína sintasa, SOR: salutaridina:NADPH 7-
oxídorreductasa, SAT: acetil coenzima A:salutaridinol-7-O-acetiltransferasa y COR:
codeinona reductasa.
1.6. Regulación de la síntesis de ABl's
La síntesis de alcaloides del tipo bencilisoquinolínico está regulada a diferentes
niveles. En P. somní.Íertjm se ha observado que la TyDC tiene una localización tejido-
específica (Facchini eí a/.,1994). Los transcritos de la enzima son más abundantes enel metafloema, el cual está asociado a las células laticíferas (Facchini eí a/„ 1998), altallo y la raíz, y menos abundantes en cápsulas en desarrollo. En el caso de la NCS,
se ha observado que su activídad es mayor en raíz y tallo de P. somn/ferum
(Samanani eí a/., 2001), al igual que en rha/Í.cfrum Wavum (Samamani eí a/., 2002). EnP. somní.Íerum los transcritos de W-metilcoclaurina 3-hídroxilasa (CYP80B1) son más
12

abundantes en tallo, raíz, hoja y tejido floral, respectivamente (Huang ef a/., 2000). Las
enzimas STS y SOR, las cuales convierten (R)-reticulina en salutaridinol, presentanmayor actividad en raíces y brotes (Gerardy eí a/.,1993a y b). En contraste, COR, lacual cataliza el penúltimo paso en la biosíntesis de morfina, está presente en toda la
planta, pero es más abundante en brotes (Unterlinner eí a/.,1999; Huang eí a/., 2000).Estos resultados sugieren que los intemediarios de la ruta son movilizados de un tejido
a otro. En un principio se pensó que la biosíntesis de morfina se llevaba a cabo en lascélulas laticíferas (Faccmni eí a/., 1998); sin embargo, ni STS ni SOR fueron
detectadas en una fracción de látex fresco proveniente de cápsulas de P. somn/.Íert/m
(Gerardy eí a/., 1993 a y b) sugiriendo que las células laticíferas son el sitio deacumulación, pero no de síntesis de este alcaloide. Por otro lado, algunas enzimascomo STS, SOR, SAT y COR han sido detectadas en cultivos de célulasdesdiferenciadas que carecen de células laticíferas (Gerardy eí a/.,1993 a y b; Lenz eía/.,1995 a y b). La acumulación de sanguinarina en raíz en P. somní.Íert/m (Faccmni eí
a/.,1995) sugiere que las enzimas involucradas en su biosíntesis solo están presentesen este tejido. Sin embargo, los niveles considerables del transcrito de BBE (Facchinieí a/„ 1996), así como su actividad enzimática en brotes (Steffens eí a/., 1985),
sugiere que los intermediarios de la ruta de biosíntesis de sanguinarina son
movilizados de brotes a raíces. Algo simHar ocurre en las raíces primarias de C.
/.apon/.ca, ya que los bajos niveles de actividad de SOMT sugieren que estos tej.idos noson el sitio principal de biosíntesis de berberina (Fujiwara eí a/.,1993). En este caso
tallos y raíces laterales presentan los más altos niveles de actividad de la SOMT.
En cultivos en suspensión de P. somní.fert/m algunos miembros de la familia de genes
de la TyDC, la enzima que canaliza la incorporación de la tirosina hacia la síntesis de
alcaloides (Fig. 1.2), se expresan en respuesta a tratamientos con homogenadosfúngicos (Facchini eí a/„ 1996 y 1998). Los niveles del transcrito de CYP80Bl
aumentan en respuesta a jasmonato de metilo (MeJa) en cultivos de E. ca//fomí.ca
(Pauli eí a/.,1998) y también en células de amapola tratadas con inductores fúngicos
(Huang eí a/., 2000). En células en suspensión de P. somn/.fert/m y de E. ca//.fomi.ca,la BBE, enzima clave en la formación de reticulina (Fig. 1.3), resulta activada
transcripcionalmente, tanto por la adición de homogenados fúngicos como de MeJa
(Dittrich eí a/„ 1991; Blechert eí a/.,1995, Facchini eí a/„ 1996). Más aún, en cultivosde E. ca/Í.fom/.ca, la actividad de las enzimas dependientes del citocromo P450 comoCFS, SPS, que están involucradas en la conversión de la esculerina en estilopina,
para la formación de los alcaloides tipo benzofenantridina (Fig. 1.3), se incrementandespués de la exposición a inductores fúngicos. Algo similar sucede con las
13

actividades de MSH y PPH, que partícipan en la conversión de S-c/.s-W-metilestilopinaa díhidrosanguinarina en esta misma ruta (Fig. 1.3), que también se activan en
respuesta a tratamientos de inducción (Tanahashi eí a/.,1990; Bauer eí a/.,1991;
Blecheh eí a/., 1995). Finalmente, la actívídad de DBOX, la enzima que cataliza la
conversión de dihídrosanguinarina en sanguinarina (Fig. 1.3), aumenta en cultivos de
S. canadens/.s tratados con MeJa o ácido salicílico (lgnatov ef a/.,1996).
La inducción de la biosíntesís de alcaloides tipo benzofenantridina en E. ca//.fom/.ca,
ocurre en concentraciones del inductor muy por debajo de los niveles requeridos paraactivar eventos asocíados con la respuesta hipersensible, tales como la producción decompuestos fenólicos (Roos eí a/„ 1998). Debido a que la producción de metabolitosfenólicos puede ser bloqueada selectivamente por una catalasa a altas
concentraciones del inductor, se sugiere que la biosíntesis de alcaloides no es
mediada por especies reactivas de oxígeno involucradas en la activación de larespuesta hipersensible (Facchini, 2001) Otros mecanismos de induccíón
relacionados con la fenilalanina amonio liasa (PAL), la TyDC y otras enzimas
involucradas en pasos posteriores de la biosíntesis de sanguinarina también han sido
demostrados en P. somn/.fert/m (Facchiní eí a/.,1996). Por otro lado, la induccíón de la
biosíntesis de los alcaloídes tipo benzofenantridína requiere una djsminución temporal
del pH citosólico causado por el fluio de protones de la vacuola (Roos eí a/.,1998), laacidificación artifícial del citosol produce la induccíón de la biosíntesis de alcaloides;
sin embargo, no produce la respuesta hipersensi.ble, que está relacionada con el
depósito de proteínas en las paredes celulares, la acumulación de compuestosfenólicos, la lignificación y la producción de exoenzimas antimicrobianas como las
quitinasas y de otras proteínas relacionadas con la patogénesis. La disminución de laconcentración de protones (H+) en la vacuola bloquea la acumulación de alcaloídes.La fosfolípasa A2, una protelna asociada a la membrana plasmáti.ca, podri'a activar la
transducción de señales que lleva al flujo de protones de la vacuola (Roos eí a/„1999). La función de las proteínas G en la inducción de las síntesis de alcaloides tipo
benzofenantridina fue estudjada en cultivos de S. canadensí.s, con moduladores de
proteínas que se unen a GTP y activadores de proteínas G (Mahady eí a/„ 1998). Porotro lado, la inducción de la biosi'ntesis de los alcaloides parece depender de fuentes
externas de Ca2+, sugiriendo que el calcio y posiblemente la calmodulina participan en
la ruta de transducci.Ón (Mahady eí a/.,1994).
14

Cuadro 1.1. Enzimas de la ruta de biosíntesis de ABl's cuyos genes correspondíenteshan sido clonados en diferentes especies.
| Enzíma Función Especie Tipo de enzima Rf
TyDC Tirosjna descarboxilasa P. somniferum Descarboxi.lasadependi.entede fosfatodepiridoxal
e1,2
6'OMT Norcoclaurina 6-0- C. japonica Metiltransferasa 3metiltransferasa dependiente de SAM
CYP80A1 Bermamuni.na sintasa 8. stolonifera Metiltransferasadependientede SAM 4
CYP80B1 W-metilcoclaurina 3'hidroxilasa P. somniferum Monooxigenasa 5E. califomica dependiente decitocromoP-450
CYP719 Enzima formadora del puente C. japonica Enzima dependiente 6metilenodioxi de citocromo P450
4'OMT 3'hidroxiw-metilcoclaurina C. japon`,ca MonooxigenasadependientedecitocromoP-450 3
BBE Enzima del puente de E. califomica Oxidorreductasa 7,8berberina P` somniferum8.stolonifera flavinilada
SOMT Esculerina 9-0- C. japonica Metiltransferasa 9metiltransferasa dependiente de SAM
COR Codeinona reductasa P. somniferum Aldo/ceto reductasa 10
Ref. (1) Facchini eí a/.,1994, (2) Maldonado-Mendoza eí a/„ 1996, (3) Morishige ef a/.,
2000, (4) Kraus ef a/.,1995, (5) Pauli eí a/„ 1998, (6) lkesawa ef a/,, 2003, (7) Dittrich
eí a/.,1991, (8) Hauschild e£ a/„ 1998, (9)Takeshita eí a/.,1995, (10) Unterli.nner ef a/„
1999.
15

Los ADNc correspondientes a los genes .involucrados en la síntesis de los ABl's hansido aislados en especies como P. somn/.ferL/m, C. /.apon/ca y E. ca//.fom/.ca, a partir de
genotecas preparadas con ácido ribonucleico (ARN) obtenido de tejidos qiie tienen lacapacidad de sintetizar los alcalo.ides en condiciones que promueven dicha síntesis,
como por ejemplo, cultivos /.n v/.Ím en condiciones de inducción.
En A. mex/.cana se han realizado pocos estudios dentro del contexto de la regulación
de las rutas de biosíntesis de los ABl's, por otro lado, tampoco existen muchosestudios con cultivos de tejidos vegetales /.n v/.Íro, es por esto, que el interés de estetrabajo está centrado en sentar algunas bases, así como desarrollar algunasherramientas moleculares, para empezar a realizar estudios a nivel bioquímico ymolecular en esta especie, relacionados con la regulación de la biosíntesis de los
ABl's.
Particularmente, las genotecas de ADNc son valiosas herramientas para elaislamiento e ident.ificación de genes de interés ya que, en principio, representa todos
los genes expresados en forma de transcritos al momento de realizar la extracción delARN, Contar con una genoteca de ADNc de A. mex/.cana, permitirá más adelante, Ia
búsqueda y el aislamiento de los genes involucrados en la biosíntesis de los ABl's, y
de esta manera, contribuir al conocimiento sobre los puntos de regulación de las rutasde síntesis. Por otro lado, el tener los genes de interés identificados, nos permitirá
realizar la comparación de éstos con los genes aislados en otras especies, así comorealizar estudios de expresión heteróloga en la continuidad de estos estudios.
1.7. Producción de alcaloides en cultivo de tejidos
Debido a que, en general, los metabolitos secundarios de origen vegetal se acumulan
en cantidades muy pequeñas, se han tratado de encontrar alternativas para laobtención de productos de importancia comercial. Una de estas alternativas es el
cultivo de tejidos vegetales. Una de las principales ventajas de este sistema es elsuministro continuo de materia prima, independientemente de las condiciones
climatológicas, ya que se realiza en condiciones controladas y reproducibles. Otra de
las ventaias de este sistema es que permite estudiar los mecanismos regulatorios debiosíntesis de estos metabolitos, lo cual es difícil de llevar a cabo a nivel de planta
(Chintalwar eí a/., 2003). El cultivo de tejidos vegetales se ha aplicado tanto a peciueñacomo a gran escala, con fines biotecnológicos. En algunos casos se ha logrado la
acumulación de los metabolitos de manera exitosa, por ejemplo, la producción de
shikonina a partir de cultivos en suspensión de LMospermum en4hrorh/.zon. Otras
16

especies que se han usado para la producci.Ón a gran escala de metabolitossecundarios a partir de cultivos vegetales son Mcoír.ana fabacum, CaíharaMusrioseüs y "a/í.ofrüm rugosüm, entre otras (Facchini eí a/.,1991 ).
Los estudjos que se han realizado para mejorar la acumulaci.Ón de los metabolitossecundarios de importancia comercial han demostrado que existen diversos factores
que afectan la producción de me{abolitos secundarios; éstos incluyen la fuente decarbono, Ia fuente de fosfatos, el suministro de precursores de la biosíntesís, losreguladores del crecimiento vegetal y el uso de inductores, además de las condiciones
de luz, ai.reacjón (Sato eía/.,1982), y agitación (Facchini eí a/.,1991). A contínuaci.ón
se presenta una breve revisi.ón del efecto de algunas condiciones de cultivo que
afectan la síntesis de los ABl's en cultivos í.n vÍ.Íno en algunas de las especies.
1.7.1. Fuente de carbono
El incremento en la producción de metabolitos secundarios, por aumento en laconcentraci.Ón de carbohidratos en el medio de cultivo, se le ha atribuído generalmente
a la mayor disponibilidad de precursores (Knobloch ef a/., 1983). Entre los
carbohídratos más empleados para aumentar la acumulación de ABI's, así como otrosmetaboli.tos se encuentran la sacarosa, la fructosa y la glucosa. En cultivos de C.
/.apon/.ca se observó que la sacarosa produjo el efecto más marcado en el crecimi.entocelular, y en la producción de berberina, en una concentración de 3%. En otrossistemas se ha usado tambíén la sacarosa al 2% (Byun ef a/„ 1990,. Jha ef a/.,1991;
Mahady eí a/.,1993); sin embargo, esta concentración no tuvo efectos significativos enla acumulación de alcaloides. El uso de concentraciones de sacarosa al 4% en
cultivos de S. canadens/.s, produjo una dismi.nución en el crecimi.ento celular y un
aumento en la biosi.ntesis de alcaloides del tipo benzofenantridina, posi.blemente
debi.do al estrés osmótico (Mahady eí a/., 1993). En otros sistemas de culti.vos de
tejidos, como en el caso de Cep^ae/í.s t.pecacuanha, se han probado concentracionesde hasta el 8% de sacarosa (Jha ef a/.,1991). Esta especie produce alcaloi.des comola cefaelina y la emeti.na, Ios cuales tienen propiedades eméti.cas y expectoran{es. EI
contenido de emetina aumentó de 0.22% en el medio de cultivo con 2% de sacarosa,a 0.32% con 6% de este azúcar, en tanto que la cefaelina aumentó de 0.1% a 1.68%.Esto demostró que la acumulación de estos dos alcaloides es dependiente de laconcentración de sacarosa.
17

1.7.2. Fuente de fosfaitos
Se ha reportado qiie la concentración de fosfatos en el medio de cultivo, (1 mMNaH2P04) favorece el incremento en biomasa celular de cultivos de S. canadens/.s,
pero .inh.ibe la biosíntesis de alcaloides del t.ipo benzonfenantridina al inhim a latetrahidroprotoberberina-N-metiltransferasa (Fig. 1.3), una enz.ima clave para su
síntes.is. Sin embargo, en células en suspensión mantenidas en medio de cultivo con
concentraciones bajas de fosfatos (0.5 mM NaH2P04), o en ausencia de éstos, se
pudo observar una mayor acumulación de sanguinarina y queleritrina, pero pocoaumento de biomasa (Mahady ef a/., 1991). Estos datos demuestran que lascantidades de fosfato en el medio de cultivo tienen una func.ión importante, tanto en la
proliferación celular, como en la biosíntesis de alcaloides. Se han observado efectossimilares respecto a los fosfatos, en cultivos de N. íabacum (Knobloch,1982) y C.roseus (Knobloch eí a/.,1983).
1.7.3. Adición de precursores
Entre las estrategias empleadas para obtener una mayor acumulación de metabolitossecundarios se encuentra la adición de precursores al medio de cultivo. En estud.ios
real.izados con cultivos en suspens.ión de A. mex/.cana para evaluar el efecto de laadición de diferentes concentraciones de fenilalanina y tirosina sobre la acumulación
de ABl's, se estableció que la t.iros.ina era el precursor específico en la biosíntesis de
berberina (Khanna eí a/ ,1982). Es importante mencionar que en este caso también
se pudo observar que la máxima acumulación de berberina no ocurre en cultivos conmayor proliferación celular, Io cual es comparable con lo observado al exponer los
cultivos a diferentes concentraciones de fuentes de carbono (Khanna eí a/.. 1982;Mahady ef a/., 1993). En cultivos de Covda/í.s yanhusuo, Ia adición de tirosina al
medio de cult.ivo resulta en un aumento de la acumulac.ión de alcaloides del tipo
protoberberina, tales como la tetrahidropalmatina y la coridalina (Staba ef a/.,1982;Lee eí a/., 2001).
1.7.4. Adición de reguladores del crecimiento vegelal
Entre los factores más analizados para aumentar la producción de metabolitossecundarios se encuentra la composic.ión de reguladores del crecimiento en el medio.En cultivos de C. /.apon/.ca se observó que una combinac.ión de ácido naftalenoacético
(ANA) (100 HM) y 6-bencilaminopurina (6-BAP) (1 LiM o 0.1 HM), se favorece tanto elcrecimiento celular como la producción de ABl's, a diferencia del uso de altas
18

cantidades de 6-BAP (10 HM), que resultaron en la disminución de la acumulación de
estos metabo" Un efecto similar se pudo observar al emplear una altaconcentración de cinetina (10 HM) (Fukui eí a/.,1982).
La biosíntesi.s y acumulación de alcaloides se ha relacionado con cierto grado dediferenciación celular. En cultivos de P. somn/.fert/m se han observado dos ni.veles deorganizaci.Ón celular; el pri.mero está relaci.onado con la formación de meristemos o
estructuras semejantes a órganos detenidas en un estadío temprano dereorganización, así como con la falta de células laticíferas. El segundo nivel se
reconoce por la presencia de tejidos que poseen brotes y raíces e incluyen la
presenci.a de cé" laticíferas típicas de una planta normal (Kamo eí a/.,1982). Elgrado de diferenciacjón de los culti.vos está, a su vez, relacionado con los reguladoresdel crecimiento vegetal.
Por otro lado, tambi.én se ha observado que la eli.mi.naci.Ón de los reguladores en
cultivos de P. somn/.fert/m induce la formación de estructuras organizadas o
diferenciadas, paralelamente con la i.nducción del metabolismo secundari.o (Sjah ef a/.,1991; Lee ef a/., 2001). Estudi.os realizados en P. óracfeaím demostraron quemientras los intermediarios tempranos de la ruta de biosíntesis, como la dopami.na, se
producen en cantidades sus{anciales en tejidos desdiferenciados, Ia acumulación dealcaloides de tipo morfinano está asociada con la citodiferenciación a células
laticíferas (Kutchan eí a/.,1983).
En culti.vos de rna/í.círt/m m/.nus se ha estudiado el efecto de la 6-BAP sobre elmetabolismo de la tirosina y la acumulación de berberina. Los cultivos de 7-. m/.nus
acumulan dopamina y ti.ramina, tanto en ausencía como en presencia de BAP; si.n
embargo, la conversión de estas aminas hasta berberi.na requiere la presencia de lacitocinina. Una vez que la biosíntesis de berberina se induce, las reacci.ones
enzimáticas intermedi.ari.as ocurren de una manera tan rápi.da que no permiten la
acumulación de los intermediarios (Hara ef a/.,1993). Los resultados del estudio del
efecto de BAP sobre la acumulación de intermediarios, sugieren que la ruta de
bi.osi'ntesis es ac{ivada a nivel de la formación de norcoclaurina o en algún paso
posterior a esta reacción. De este modo, la BAP parece activar las reaccionessubsiguientes a la descarboxilación de la tirosjna (Fig.1.2) (Hara ef a/.,1993).
19

1.7.5. Aplicación de inductores
El estudio de la biosíntesis de los alcaloides ha seguido la prem.isa de qiie estosmetaboritos son pahe del sistema de defensa química en respuesta al ataque de
patógenos y herbívoros. En algunos trabajos se ha logrado la acumulación de estosproductos mediante la manipulación de cienos factores como las concentraciones delos reguladores del crecimiento (Kutchan et a/.,1983, Cline ef a/.,1988), la adición de
inhibidores de la slntes.is de proteínas (Hsu, 1981; Mahady eí aL 1994), y la
exposición de los cultivos a temperaturas bajas (Cline ef aL 1988). Otra estrategiacomúnmente usada involucra el uso de inductores preparados a panir de hongosfitopatógenos (Heinste.in eí a/.,1985., Eilen ef a/.,1985 y 1986., Funk eí a/.,1987). Sin
embargo, en algunos casos se ha observado la acumulación de metabolitossecundarios, mientras que en otros no ha habido ningún tipo de respuesta altratam.iento. Esto sugiere que no todos los inductores preparados a paftir de cultivosfúngicos son capaces de iniciar los mecanismos de defensa. Por otro lado, tamb.ién seha observado que la inducción de la producción de metabolitos secundarios puedeaumentarse en los cultivos en fase de crecimiento tardla. Todas estas observacionesindican que no existe una regla general a seguir en la apl.icación de las técnicas de
inducc.ión para aumentar la acumulación de metabol.itos secundarios, ya que cadasistema de cult.ivo posee características diferentes (Byun eí a/.,1994).
Se han realizado diversos estudios en los cuales se han usado preparac'iones fúngicasderivadas de Oendn/ph/.on pen/.cwm un hongo patógeno específico de plantas del-*;.;¿_Ei:tiv-e-r.áti,'c-o'deveriici,,iumdah'.iae,o::.?_p?`_bg_e_r?_d_?_5_a_p_a_ve_r`_c.`I:^eí
a/., 1988). En el caso de E. ca//.Íom/.ca se han ut.ilizado preparaciones a partir de
Pen/cí'///.m (Schumacher eí a/.,1987) y Saccharomyces (Byun eí a/.,1994). En esteúlt.imo estudio, Ios cultivos se sometieron a diferentes concentraciones del inductor
(homogenado de levaduras) y se observó que la exposición a altas concentracionesde éste resultó en una menor acumulación de alcaloides del tipo benzofenantr.idina,
con respecto a los expuestos a menores concentraciones. La acumulación máxima dealcaloides totales (60 mg/L) se observó en cultivos expuestos a 60 Lig de la levadura
por gramo de peso fresco del cult.ivo celular (Lig levadura/g de PF). Por otra parte, enlos cult.ivos expuestos a 500 Ltg levadura/g de PF, Ia acumulación fue de alrededor de
40 mg/L (Byun e( a/., 1994). En cultivos de S. canadensi.s también ha sido usadoPen/.c/.///.um expansL/m como inductor (Mahady eí a/., 1994).
Otros productos utilizados como inductores de la biosíntesis de ABI's en cult.M de S.canadens/.s son los flavono.ides como la quercetina y la rutina En este caso, lainducción de los cultivos se realizó con Dend"ph/.on peni.c/.//i.um en la fase exponencial
20

del ciclo de cultivo (día 5). Las concentraciones que produjeron la mayor acumulación
de alcaloides fueron del orden micromolar. Altas concentraciones resultaron entoxicidad celular causando una djsmi.nución en el crecimiento celular, necrosis y una
menor acumulación de alcaloides (Mahady eí a/., 1994).
Las liasas de pectina son bien conocidas también por estimular la síntesis demetabolitos secundari'os (Nothnagel ef a/„ 1983; Davis eí a/„ 1984). Estas son liasas
de ácido a-1,4-endopoligalacturónico que en concentraciones nanomolares di.gieren
paredes celulares vegetales, y que son capaces de estimular la síntesis de berberinaen 7-. m/nus. En concentraciones menores también esti.mulan el crecimiento celular
(Smolko eí a/.,1994).
Los jasmonatos (ácido jasmónico y MeJa) son metabolitos que se han utilizado
regularmente para la inducci.Ón de metabolitos secundari.os en diversos sistemas de
culti.vo. Existen algunas evi.dencias acerca de la función de los jasmonatos enrespuestas de defensa por parte de las plan{as, particularmen{e, en las rutas debiosíntesis de metabolítos que participan en la defensa química, como las fitoalexinas,se ha demostrado que el ácido jasmónico activa la expresión de ciehos genes, comolos de la ruta de bi.osíntesis de alcaloides terpéni.cos (van der Fits eí a/„ 2000;
Memelink eí a/., 2001), así como la transcripción de los genes involucrados en la
biosíntesis de alcaloides del tipo benzofenantridina (Blechen eí a/.,1995; Pauw eí a/.,
1998,. Hajder eí a/., 2000).
En los últimos años se ha obteni.do evidencia de que el dimetilsulfóxido (DMSO), que
normalmente se utiliza como di.solvente y agente permeabilizante (Brodeljus eí a/.,
1983), puede tener propi'edades como inductor de la acumulaci.ón de metabolitos
secundari.os. En C. mseus se ha observado que al usar el DMSO como disolvente, enestudios con análogos de precursores de la ruta de bjosíntesi.s de alcalojdes
.monoterpeno-indólicos (AMl's), tales como la triptamina, el contenido de ajmali.cina seincrementa de manera significativa (Bonzom eí a/., 1997). Por otro lado, se ha
observado que al usar el DMSO al 1% (v/v) ocurre una disminución en la acumulación
de alcaloides en las rai'ces de plántulas de C. rosem las cuales acumulan ajmalicina,serpentina y yohimbina. Este efecto también se observó en las partes aéreas queacumulan vindoli.na y fue más notorio en plántulas de mayor edad. Por otro lado, la
acumulacjón de catarantina en los brotes no fue sensi.ble a este di.solvente. Dadas sus
características como agente permeabilizante, se sugiere que el DMSO podríafavorecer la excreción de alcaloides del tejjdo al medio de cultivo o bien, la entrada decomponentes del medi.o a las células (Bonzom eí a/„ 1998).
21

1.7.6. Efecto de la luz
Se han realizado algunos estudios para observar el efecto de la luz sobre laacumulación de diferentes tipos de alcaloides. Con respecto a los ABl's solamente seha estudiado en cultivos de callos de Che/i.don/.um ma/.us en los cuales se haobservado que cuando los callos son mantenidos en oscuridad acumularon una mayorvariedad de alcaloides. Al comparar el t.ipo de alcaloides presentes en estos cultivos
con los alcaloides acumulados en plantas diferenc.iadas de C. ma/.m se notó que loscultivos i.n vt.W no producen quelidonina, queleritrina ni berberina (Colombo ef a/.,
1991).
En cultivos de callos de M.coíi.ana g/a" la luz inhibió de manera significativa laacumulación de alcaloides como anabasina, nicotina y nornicotina, en particular, se
pudo observar que la cinética de acumulación de la anabasina, el cual fue el alcaloidemayoritar.io en los cultivos transferidos de la oscuridad (34 Lig/g de PF) a la luz ("
Hg/g de PF) o viceversa demostró que la inhibición de la acumulación de alcaloides
por efecto de la luz era reversible. En este mismo estudio, tamb.ién se usaron cultivosde raices de M.cofí.ana rtjsf/.ca, Ios cuales también acumularon un menor conten.ido dealcaloides cuando fueron mantenidos en luz. Sin embargo, en este caso
probablemente el efecto de la luz sobre la acumulación de alcaloides sea indirecto, al
parecer el contenido de alcaloides se redujo como resultado de un menor crecimientodel tejido ya que se inhibió 50%, debido a esto se redujo la cantidad de tejido capaz
de sintetizar alcaloides (Hobbs eí a/„ 1991 ).
En C. roseus se sabe que los alcaloides terpeno-indólicos (ATl's) son compuestos queabsorben la luz ultravioleta (UV), por lo cual se le ha conferido la función de
protectores de este tipo de radiación. Esto fue sugerido por la observación de que laluz UV .induce la formación de alcalo.ides d.iméricos en hojas de C. roseus (Hirata eí
a/., 1993). Además, se ha reportado también la regulación del gen de la triptofano
descarboxilasa (TDC) por luz UV, así como el aumento en la acumulación del ácido
ribonucleico mensajero (ARNm) de los genes Tdc y Sírque codifican para las enzimasTDC y estr.ictosidina sintasa (STR) involucradas en la ruta de biosíntesis de este tipo
de alcaloides. Por último, también se observó un aumento en la acumulación de
algunos ATl's en respuesta a luz UV, tales como estrictosidina, ajamlicina,catarantina, tabersonina, serpentina, vindolina y v.inblastina (Ouwerkerk eí a/.,1999).
En otro estudio realizado en C. roseus, cultivos de callos se crecieron en condicionesde luz continua y oscuridad total, y se observó que el contenido de v.indolina fue de 3 a
4 veces mayor en los callos mantenidos en luz respecto a los mantenidos enoscuridad. En el caso de serpentina, también se observó una mayor acumulación en
22

condiciones de luz. Por otro lado, también se detectó que los cultivos mantenidos enluz acumularon una mayor variedad de alcaloi.des que los culti.vos mantenidos en
oscuridad (Zhao eí a/., 2001).
JUSTIFICACIÓN
A. mexí.cana es una planta nativa de México que se ha utjlizado en la medicinatradicional. Esta planta produce alcalojdes simi.lares a los de otras Papaveráceas loscuales tienen importanci.a comercial debjdo a sus usos en tratamientos contra laleishmaniasis, así como algunas afecci.ones del corazón y el hígado. Más aun,
recientemente se ha postulado que algunos de estos alcaloides podri'an usarse entratamientos de fármaco-dependencia y contra el virus de i.nmunodeficiencia adquiri.da
(V'H).
Desafortunamente, en condjciones naturales estos metabolitos se producen encantidades muy pequeñas en la planta, aunado a esto A. mex/.cana es una planta quecrece de manera silvestre, Io cual dificulta la disponibilidad de material vegetal para la
obtención de los metabolitos de i.nterés ya que esto se encuentra limitado por lascondiciones ambientales. Todo esto hace necesario explorar otras alternativas deobtención de díchos metaboli.tos. Por lo tanto, generar un sistema de cultivos /.n vitro a
parti.r de esta planta que puedan además ser sujetos a tratamientos de inducción, nospermi.tiri'a disponer de un modelo con el que se puedan realízar estudios para elentendimiento de los posibles mecanismos de regulación de la síntesis y acumulaci.Ón
de estos compuestos. Esta información puede representar la base para, más adelante,tener la posibili.dad de mani.pular dlchos sistemas de cultivo con el fin de obtener los
metabolitos de jnterés farmacéutico.
23

OBJETIVOS
Objetivo general
Establecer un sistema de cultivos /.n v/.fro de A. mex/.cana y caracter.izarlo por sucapacidad de síntesis de alcalo.ides así como por su respuesta a la adición exógenade inductores del metabol.ismo secundar.io.
Objetivos específicos
• Analizar el patrón cualitativo de los alcaloides en los diferentes tejidos de
plantas maduras de A. mex/.cana.
Analizar el patrón cualitativo en cultivos /.n vÍ.Íro en diferentes condic.iones.
Desarrollar una metodología de inducción de la síntesis de alcaloides en
cultivos /.n v/.íro de A. mex/.cana.
Construh una genoteca de ADNc de tejidos con capacidad de síntesis dealcaloides.
24

CAPÍTULO 11
MATERIALES Y MÉTODOS11.1, Djseño experimental
Para cumplir con los objetivos propuestos, se planteó un diseño experimental dividido
en dos partes. En la primera parte se probaron diferentes condiciones de cuM queafectan la síntesi.s de alcaloides (Fig H.1). Estas condiciones incluyeron la iluminaci.Ón
y aplicaci.Ón de inductores químicos. Con base en estos resultados se seleccionó eltejido adecuado, por su respuesta de síntesis y acumulaci.Ón de alcaloi.des, para la
construcción de la genoteca de ADNc.
En la segunda parte, iina vez que se observó cuál de estas condici.ones produjo unarespuesta en la acumulacjón de alcaloides, se realizó un tratamiento a di.ferentestiempos de incubación bajo las condi.ciones de inducción selecci.onadas con base en la
respuesta.
11.2. Colecta de plantas silvestres de A. mex/.cana
Plantas maduras de A. mex/.cam con flores y cápsulas identjficables, fueroncolectadas completas (extraídas de raíz), en poblaciones silvestres localizadas en elestado de Yucatán. Las plantas fueron empacadas en bolsas de polietileno yconservadas en hielo para transportarlas al laboratorio en donde fueron lavadas conagua fría, secadas con toallas de papel absorbente y di.vjdjdas en hojas, tallos, raíces,flores y frutos (cápsulas). Porci.ones de cada tejido fueron pesadas, congeladas en
nitrógeno Iíquido y conservadas a -80 °C hasta su análisis.
La identi.fi.cación taxonómica de las plantas fue realizada por el Dr. Roger Orellana
Lanza, de la Uni.dad de Recursos Naturales de este Centro, y un ejernplar se depositó
en el herbario U NajH Tikín Xiw (Casa de la Hjerba Seca) de la misma Unidad bajo el
registro.. No de colecta 1, No. de catálogo 43761.
25

Figura m . Diseño experimental del proyecto. (Los detalles de cada componente sonespecificados en las secc.iones correspondientes).
11. 3. Cultivos i.n y/.fro
11.3.1. Establecimiento y manejo de los cultivos celulares
Para la generación de una línea de cultivos en suspensión se util.izó un cultivo de
callos inducidos en 1998, a partir de hojas jóvenes de A. mex/.cana Como medio de
mantenimiento basal se emplearon las sales del medio PC de Phillips y Collins (1979)
apH5.5quefueadicionadocon25g/Ldesacarosaycon0.5g/LdeANAy1.5g/Lde6-BAP (Campos-Tamayo, 2002). Para los medios semisólidos se agregaron 9 g/L deagar. Las líneas de callos y suspensiones celulares se han mantenido en lascondicionesdecuartodecultivo(25°C,baioluzcontinuadeentre40y50Hmolm-2s-'
prov.ista por lámparas fluorescentes de 39 W). Las suspensiones celulares se
26

mantuvieron además en agitación continua a 100 rpm. Los cultivos fueron transferidos
cada dos semanas.
11.3.2. Inducción de la síntesis de alcaloides en cultivos in vitro de A. mexicana
ll.3.2.A. Efecto del régimen de iluminación sobre la síntesis de alcaloides en callos de
A. mexicana
Para estudiar el efecto de la luz sobre los cultivos de callos, cultivos de 21 días
(Campos-Tamayo, 2002) se transfirieron al medio basal, y los frascos de cultivo fuerondepositados en el Ínterior de una caja que fue cerrada y cubi.erta con una manta negra.Para analizar el efecto del régimen de luz, se realizó un curso temporal manteniendo
un lote de callos en condiciones de iluminación continua, mientras que otro se sometió
a oscuridad completa. En ambos casos, el experimento se i.nició con 1 g de tejído decallos de 21 días, que se mantuvieron en las condiciones del cuarto de cultivo a lo
largo del experimento. De cada lote se tomaron muestras por triplicado cada 5 díashasta completar un ciclo de 50 días. Después de cosechar los callos, se les determinó
el PF, y fueron congelados inmediatamente en nitrógeno líquido y conservados a -80°C hasta ser procesados. Para cada cosecha, se colectaron al menos tres
repeticíones Índependientes de cada condición. El peso seco (PS) se determinó por
líofilización. A estas muestras se les determinaron los parámetros de Índice de
Crecimiento (ÍC) y % de PS al final del ciclo de cultivo.
El lc se determinó de la si.guiente manera:
Íc = pF' - PF'/PFI
En donde PFi y PFt representan los pesos frescos inicial y final
El porcentaje de PS final del ciclo de cultívo (% PS) se determinó en el día 50 del ciclo
de acuerdo a la sjguiente ecuacióm
% ps= (ps/pF) X 1oo
En donde PS representa el peso seco y PF el peso fresco
27

11.3.2.8. Efecto de la aplicación de MeJa sobre la acumulación de alcaloides encultivos en suspensión de A. mexicana
Estos experimentos se realizaron en dos etapas. La primera de ellas fue una dosis-respuesta evaluada después de un tiempo fijo de 24 h para una primera aproximaciónsobre una posible respuesta de los cultivos a la exposición con el inductor. Una vez
determinada la dosis a la que el inductor produjo la mejor respuesta en cuanto a la
acumulación de alcaloides, se realizó un curso temporal aplicando dicha
concentración (Fig.11.2).
Para el experimento de dosis-respuesta, cultivos de células en suspensión de 5 días
(5 g/35 mL de suspensión) fueron expuestos a tratamientos con 0 (testigo),10,100 y1000 LiM de MeJa disuelto en DMSO durante 24 h. Para la aplicación de los
tratamientos, se ajustó la concentración de una solución patrón de MeJa de tal manera
que cada matraz recibiera la dosis correspondiente en 1 mL del disolvente. Lostestigos recibieron el mismo volumen de DMSO Se preparó otra serie de testigos que
no recibieron ningún tratamiento. Se ajustó el pH de la solución de MeJa al del medio
de cultivo (5.5) y antes de su apl.icación la solución fue esterilizada por filtración a
través de una membrana Millipore (poro 0.22 L.m), resistente a disolventes orgánicos.
Cada tratamiento se aplicó por triplicado. Una vez transcurrido el tiempo del
tratamiento, el tejido fue cosechado por filtración al vacío, pesado, congelado ennitrógeno líquido y mantenido a -80 °C hasta su análisis. Para la extracción e
identificación de los alcaloides acumulados, el tejido fue liofilizado.
Una vez seleccionada la concentración que produjo tanto la mejor respuesta de
acumulación de alcaloides, como el menor daño al tejido tratado, ésta se empleó paraaplicarla a cultivos de la misma edad durante un periodo máximo de 24 h,
colectándose muestras por triplicado a las 0, 6, 12, y 24 h. El manejo de los cultivos
para la aplicación de los tratamientos y durante su cosecha fue como se describióanteriormente.
28

Figura 11.2. Diseño experimental seguido para la inducción química de los culti.vos de
A. mexicana.
11. 4. Extracción, identificación y cuantificación de alcaloides
Se obtuvo un extracto total de alcaloides mediante una extracción ácido-base
(Monforte-González eí a/„ 1992). Brevemente, 1 g de tejido liofilizado fuehomogeneizado en 50 mL de metanol e incubado a 55 °C con agitación durante 2 h.Después de este tiempo, la suspensión fue filtrada y el extracto metanólico se evaporóa sequedad a presión reducida. El resi.duo se resuspendió en 15 mL de áci.do sulfúrico
al 2.5% y esta suspensión fue extrai'da tres veces con volúmenes jguales de acetatode etilo, conservando la fase acuosa cada vez. Una vez recuperada la fase, se leajustó el pH a 9.5 con hi.dróxido de amonio concentrado; la fase acuosa basificada fueextraída tres veces con volúmenes iguales de acetato de etilo, conservando cada unade las fases orgánicas, que fueron mezcladas para evaporar a sequedad a presiónreducida. El residuo se disoMÓ en 1 mL de metanol. El contenjdo de alcaloídes totales
fue estimado por espectrofotometría. El contenido de alcaloides totales fue calculadode acuerdo a la siguiente fórmula:
tig/mL de alcaloides totales = (D028o) (100)/0.02289
Donde D028o es la densidad Óptíca a 280 nm; 100 es el factor de dilución y 0.02289 la
pendiente de una curva de calibración de una mezcla de alcalojdes.
29

La separación de los alcaloides se realizó mediante cromatografía de capa delgada
(CCD) utilizando placas de gel de silice (cromatofolios de gel de smce 60 F254 Merck)util.izando d.iferentes sistemas reportados en la literatura (Cuadro 111.1). Se probaron
diferentes mezclas de disolventes para obtener la mejor resolución en la separaciónde los alcaloides (ver resultados). Para la identificación de los alcaloides separados se
usaron estándares de berberina y sanguinarina, los cuales son representantes
principales de las rutas de síntesis de los alcaloides tipo protoberberina ybenzofenantridina, respectivamente. Los valores de Rf se calcularon de acuerdo a lasiguiente fórmula..
Ftf = DTIDF\
Donde Ríes el valor de migración de la mancha del estándar, DT es la distancia totalrecorr.ida por el sistema de disolventes y DR es la distancia recorrida por la mancha del
estándar.
La cuantificación de los alcaloides se realizó por densitometría /.n sÍ.Íu sobre las placas
cromatográficas, utilizando el método de estándar externo Wagner eí a/.,1984) y undens.itómetro Shimadzu CS-930 equipado con un graficador (DR-2). Las lecturas en el
densitómetro se hicieron por barrido lineal de cada carril y la concentración se
determinó comparando el área del pico de las muestras contra el área de los picosobtenidos aplicando concentrac.iones conocidas de los estándares de sulfato deberberina y cloruro de sanguinarina (Sigma-Aldrich Chemical Co.). Los alcaloides
fueron detectados por fluorescencia a longitud de onda (^) de excitación de 300 nm.
Esta ^ fue seleccionada experimentalmente con los estándares y en ella se logra el
pico de emisión máximo tanto para berberina como sanguinarina.
Se prepararon soluciones patrones de 1 mg/mL de ambos estándares que fuerondiluidas a una concentración de 10 ng/LtL (0.01Hg/tiL). Con estas soluciones se
realizaron las curvas de calibración usando un rango de 0 a 120 ng para berberina
(Fig 11.3) y de 0 a 160 ng para sanguinar.ma, con intervalos de 10 ng. (Fig.11.4).
30

20 40 60 80 100 120
Berberina (ng)
Figura 11.3. Curva de cali.bración promedio de berberina en el sistema de disolventes J
(n-butanol: ácido acético:agua 7.1:2). (Y = -3682.30 + 355.25X). Promedi.o de tresrepeticiones de cada punto.
20 40 60 80 100 120 140 160
Sanguinarina (ng)
Figura 11.4. Curva de cali.bración promedio de sanguinarina en el si.stema de
disolventes J (n-butanol:ácido acético..agua 7:1:2). (Y = -1485.303 + 245.518X).
Promedio de tres repeticiones de cada punto.
31

11.5. Construcción de una genoteca de ADNc a partir de callos de A. mex/.cana.
Para la construcc.ión de la genoteca de ADNc se seleccionaron callos de 5 díasmantenidos en luz continua.
La extracción del ARN total se realizó por el método de Logemann eí a/., (1987).
Brevemente, 1 g de tejido se congeló con nitrógeno Iíquido, se maceró hasta un polvofino y se homogeneizó con 2 mL del amortiguador de extracción (Guanidina-Hcl s M,Tris-HCI 50 mM, ácido etileno diamino tetraacético (EDTA) 20 mM, P-mercaptoetanol
50 mM, pH 7) y 1 mL de una mezcla de fenol.cloroformo:alcohol isoamílico 25:24:1.
Una vez separada la fase acuosa, los ácidos nucleicos contenidos en ésta se
precipitaron con 0.7 volúmenes de etanol absoluto frío y 0.2 volúmenes de ácidoacético 1 M. La pastjlla se lavó primero con 2 mL de acetato de sodio 3 M pH 5.2 y
después con 2 mL de etanol al 70%, y finalmente se resuspendió en 50 LtL de aguadestilada. La .integridad del ARN se verificó visualizándolo por fluorescencia en geles
de agarosa al 1% con formaldehído, teñidos con bromuro de etidio y se cuantificó porespectrofometría a 260 nm en un espectrofotómetro Smartspec 3000 de BioRad.
Como primer paso para la construcción de la genoteca, se purificó el ARN
poliadenilado poli(A+) a pafth del ARN total utilizando el paquete de Stratagene (SanDiego CA). Para ello, 500 Lig de ARN total se desnaturalizaron por calor a 65 °C
durante 5 min. Después de enfriar la muestra, se le adicionó el amortiguador decorrida (Tris-Hcl 10 mM pH 7.5, EDTA 1 mM y cloruro de sodio 5 M) y se cargó en
aproximadamente 700 uL en una columna de oligo(dT) celulosa, preequilibrada con
400 HL del amortiguador de corrida. EI ARN poli(A+) se eluyó con 800 LiL del
amohiguador de elución (Tris-Hcl pH 7.5 10 mM y EDTA 1 mM), precalentado a 65°C. El eluato que contenía el ARN pol.i(A+) fue precipitado con 2 volúmenes de etanol
absoluto frío toda la noche a -20 °C. El precipitado fue resuspendido en agua tratadacon dietil pirocarbonato (DEPC) y el rend.imiento de ARN se estimó por
espectrofometría a 260 nm; su calidad se verificó por visualización fluorescente en
geles de agarosa teñidos con bromuro de etidio.
Para la síntesis de la primera hebra de ADN se utilizó como cebador un
oligonucleótido d(T), que contiene un sitio de restricción para Xho 1. La mezcla de
reacción se preparó con 5 Lig de ARN poli (A+), 2.8 Hg del cebador, 0.5 HM de cada
uno de los desoxinucleótidos (dATP, dGTP, dlTP, dcTP) y 62.5 U de transcriptasareversa Stratascript en un amortiguador de reacción para un volumen total de 50 HL.
La reacción se incubó durante 1 hora a 42 °C y fue detenida en un baño de hielo. Parala síntesis de la segunda hebra, al ADN de cadena sencilla se le adicionaron 0.8 mM
32

de cada desoxinucleótido (dATP, dGTP y dTTP y dcTP mM), 0.9 U de ARNasa H y 90U del fragmento Klenow de la ADN polimerasa en un amortiguador de reacción para
un volumen final de 200 HL. La mezcla se incubó 2.5 horas a 16 °C y la reacción fuedetenida en un baño de hielo. Para finalizar el procesamiento del ADN, se adjcionaron
23 HL de una solución 2.5 mM de los cuatro desoxi.nucleótidos y 5 U de ADN
polimerasa PÍu. La mezcla se incubó por 30 min a 72 °C y la reacción fue detenida conun volumen de fenol:cloroformo 1 :1 v/v. La fase acuosa fue recuperada y después deuna extracción con un volumen de cloroformo, el ADNc fue preci.pitado con acetato desodio 3 M y etanol absoluto frío a -20 °C durante toda la noche. Se obtuvo el ADN
precipitado centrifugando durante 1 hora a 4 °C, la pastilla fue lavada con etanol al70% y secada a vacío.
Previ.o a la ligación del ADNc en el vector, se añadió un adaptador de EcoRl. Para
ello, el ADNc fue resuspendido en presencia de 3 6 Hg de los adaptadores e incubado
por 30 min a 4 °C. Después de esto, se adicionó 1. uL del amortiguador de li.gaci.ón, 1
HL de ATpr (10 mM) y 4 U de ADN ligasa T4. La reacción fue incubada por 2 días a 4°C h y se detuvo incubando a 70 °C por 30 min y después por 5 min a 4 °C.
Para insertar el ADNc de manera unidireccional, éste fue digerido con Xho 1 y despuésde ser precipitado con etanol, se mezcló con 5 LiL de amortiguador STE (cloruro desodio 100 mM, Trjs-Hcl 20 mM pH 7.5 y EDTA 10 mM) y se fraccionó en una columna
de sefarosa CL-2B de 30 cm de largo, pre-equilibrada con el amortiguador STE paraseparar por tamaño. EI ADNc fue eluído con 3 mL del mismo amortiguador, se
colectaron varias fracciones las cuales se extrajeron con fenol-cloroformo 1 :1 v/v y se
precipitaron con etanol a -20 °C toda la noche para recuperar el ADNc. El precipitadofue obtenido por centrifugaci.Ón a 12,000 X g por 60 min a 4 °C, y lavado con etanol al
8/NMovN.
Finalmente, la reacción de ligaclón del ADNc obtenido en el vector Uni-ZAP XR se
realizó Íncubando 1 uL de la muestra (~100 ng), 0.5 uL de un amortiguador de
reacción, 0.5 HL de ATpr 10 mM (pH 7.5),1 Ltg del vector de Uni-ZAP XR,1.5 HL de
agua y 2 U de ADN ligasa T4, para un volumen final de 5 LiL. La reacción se incubó
toda la noche a i2 °C.
La genoteca fue empacada en fagos ^ utilizando el sistema Gigapack 111 Gold
(Stratagene San Diego CA) y plaqueada en una línea celular de Escher/.ch/.a co//. XL1-Blue MRF'. El título de la genoteca fue determinado como recomienda el fabrjcante.
Brevemente, la cepa de E. co//. fue jncubada a 37 °C durante 15 mi.n con diferentesvolúmenes de la suspensión de fago para su infección y después plaquedas en cajas
33

de Petri con medio NZY (cloruro de sodio 85 mM; sulfato de magnesio s mM; 5 g/L
extracto de levadura; 10 g/L caseína hidrolizada, pH 7.5) después de 6 h de
incubación a 37 °C se contaron las placas de lisis, puesto que cada placa de lisiscorresponde a un evento de infección de un fago sobre una bacteria, el número totalde placas de lisis corresponde al número de fagos por unidad de volumen y fuecalculado de la siguiente manera:
TP= (PL) (10) (LLL producto empacado)/(Hg empacados) (HL plaqueados)
Donde TP es el título primario en Unidades Formadoras de Colonias por mL (UFC/ml).,
PL es el número de placas de lisis y 10 es el factor de dilución empleado.
Este parámetro es un indicio de la representación de todos los transcritos expresadosen las células en suspensión de A. mexí.cana, sistema que se utilizó para laconstrucción de la biblioteca. El valor del título primario de la genoteca fue de 848,000
un.idades formadoras de colonias (UFC). La genoteca fue amplificada para
incrementar dicho título y almacenada a -70 °C, en alícuotas en amortiguador SM
(cloruro de sodio 100 mM; sulfato de magnesio s mM; Tris-Hcl 50 mM pH 7.5., gelatina0.01 %) y cloroformo 20: 1.
34

CAPÍTULO 111
RESULTADOS Y DISCuslóN
Los resultados se han organizado para su presentacíón de la siguiente manera: en la
primera parte se describe el análi.sis del peml de alcaloides en los di.ferentes tejidos dela planta. Posteriormente se describe el proceso de obtención de los cultivos así comolas estrategias de inducción utili'zadas. Fjnalmente, en la tercera parte, se describe el
proceso de construcción de la genoteca de ADNc, a partir de los tejidos de callos.
111.1. Análisis del porfil de acumulación de los alcaloides por CCD en plantas de
A. mexicana
El trabajo se inició caracterizando el perfíl de alcaloides y su distribución en los
diferentes tejidos de la planta. Para ello, se analizaron extractos obtenidos a partir de
raíz, tallo, cápsula y hoi.as de plantas maduras, colectadas en campo. También secolectó el látex de los tallos. Este análisis se llevó a cabo por CCD, utilizando placas
de gel de sílice de 5 cm de largo, con soporte de aluminio e indícador de fluorescencia
(F254), (Merck). Como fase móvil se emplearon las mezclas de disolventes descri'tas enel Cuadro 111.1 de trabajos previos (referidos en el mismo cuadro). La identificación de
los alcaloides se Hevó a cabo por comparación d¡recta con muestra§ de estándares, y
en el caso de los componentes que por falta de estándares no pudieron seridentificados con certeza, se emplearon como cn.terios de identificación preliminar los
valores de Rf en diferentes disolventes y las propi.edades de fluorescencia. El trabajose concentró en dos de los alcaloides más abundantes en esta planta: la
sangui.narina, del tipo benzofenantridi.na, y la berberina, del tipo protoberberina. EI
primero de ellos produce una fluorescencía de color narani.a intenso; mientras que elsegundo genera una fluorescencia de color amarillo (Fig.111.1).
Para separar estos dos alcaloides, en un pri.ncipio se eli.gieron mezclas sencillas,
preparadas con dos di.solventes, por ejemplo las mezclas A, 8, C, D, E, F, G y H enCuadro lH.1. En todos los sistemas mencionados. la sanguinari.na se resolvió como
una mancha bien defini.da con los valores de Rf descritos en el Cuadro 111.1. Sin
embargo, la berberina siempre permaneció en el origen (Figs.111.1, paneles A, 8, C, D
E, F, G y H). Para lograr que la berberina migrara, se probaron mezclas máscomplejas, con al menos tres disolventes, como las mezclas 1 y L (Cuadro 111.1). No
obstante, con esta mezcla tampoco fue posible que la berberina migrara de su puntode origen, en tanto que la sanguinari.na se ubicó con un Rf de 0.91. Las mezclas J, K,
y M, (Cuadro 111.1) lograron que tanto la berberina como la sanguinarina migraran del
35

punto de origen. De éstas, en la mezcla J ambos alcaloides aparecieron como bandasadyacentes (datos no mostrados). Con el fin de lograr una mejor resolución, se realizóla elución 3 veces con la misma mezcla, Iográndose una separación y resolución
adecuada, con valores de RÍ de 0.75 para berberina y 0.52 para sanguinarina. (Fig.111.1 J). Con otras mezclas terciarias (K, M; Cuadro 111.1), también se separaron ambos
alcaloides; sin embargo, los valores de Rf para sanguinarina fueron muy altos en
ambos casos (0.95 y 0.96, respectivamente) y aunque los valores para berberinafueron de 0.29 y 0.75. respectivamente, ésta apareció como una mancha ligeramentebarr.ida. Se probaron algunos sistemas con cuatro disolventes, como las mezclas N y0 Cuadro 111.1). En ambos casos, la berberina no migró de su punto de origen en tanto
que la sanguinarina tuvo valores de Rf de 0.86 y 0.80 respectivamente.
La Figura 111.1 concentra las imágenes de los cromatogramas en las diferentes
mezclas de disolventes. De esta manera, y después de comparar la resolución logradaen cada uno de ellos, se seleccionó la mezcla J (n-butanol:AcOH:agua 7:1:2); Cuadro
111.1) haciendo la elución múltiple para analizar los extractos obtenidos a partir de los
diferentes tejidos. Para confirmar que las bandas identificadas correspondian a unsolo compuesto, éstos fueron extraidos del adsorbente y sometidos a unacromatografía bidimensional, utilizando la mezcla J para la primera separación y una
mezcla de cloroformo: acetona: metanol: ácido acético: agua 10:4:2:3:1; (Hara ef a/.,
1993) para la segunda. Estos análisis demostraron que la intensidad de la
fluorescencia de la manchas cromatográficas correspondientes a sanguinarina y
berberina se debia únicamente a estos compuestos (Fig. 111.2).
36

Cuadro 111.1. Valores de Rf encontrados con las mezclas de di.solventes empleados eneste trabajo para la separación de los alcaloi.des en tejidos de A. mex/.cana. En todoslos casos, las placas cromatográficas se cargaron con un volumen de extractoequivalente a 5 mg de PS. Las áreas sombreadas muestran las mezclasseleccionadas para los análisis de cuantificación de berberina y sanguinarina.
M
| Composición
Berborina 1 Sanguinarina lRof1
Rflo.oolo.oo'o.oo'o.oolo.oolo.oolo.oolo.oo'O.oo ' lRsl'+ 'Rflo86 RlA lcloroformo:metanol(9n
S1+
'
18 benceno:etanol(4: 1 ) - lo'81 +
1
lc benceno:metanol (6: 1 ) 081 +
1
D tolueno:metanol (9:1 ) - 056 +
1
E cloroformo..metanol (80:5) 082
2
F benceno:etanol (9:1) 053
++ 3
G cloroformo:etanol(9:1) - 081 +
4 1
H acetato de etílo:metanol (9: 1 ) . 088 +
4
1 cloroformo:acetato de eti.lo:metanol 0.91 +
11
(2:2: 1 )
J n-buta\nol:ácidoacético;aduat(7.1'2) o75 + o52 +`
K n-butanol:agua:hidróxido de amonio 0.29 + 0.95
51
8: 1 : 1
L benceno:acetona:metanol 7:2.1) o.oo l0.75 090 +MN cloroformo:methanol:ácido acético + 0.96
15
(25: 10: 1 )benceno:acetona:djetiléter:hi.dróxi.do de
'00.00 1
'
0.86 '
1
1amonío (4:6:1 :0.3) 1cloroformo:benceno:metanol:formamida
"011
0.80 1 '' 140:50:10:0.5)
M, mezcla: F\sl, resolución
(+) resolución adecuada o (-) inadecuada, definida como la migración de una manchacompacta y si.n di.fusión cuyo valor de Rf es fácilmente cuantjficable.
Reft (1) Baerheim ef a/.. (1983); (2) Wagner ef a/„ (1984); (3) Chen ef a/., (1996); (4)lsrailov eí a/.. (1986); (5) Sato eí a/., (1982).
37

E FL T H C 1
#!;3fto
ERTHCL
#i;3Íi
nE Ft T H C 1
#i:8Íi
E F` T H C L
%i:8±O
E Ft T H C L
#g:8£0
HE F` T H C L
#!:3É5
EE R T tl C L
%!:3fto
: ==-:1: -
E F` T H C L
#g::83:9
E FI T 11 C L
#::8Íi
ERTHCL
%!;3fio
E F{ T H C L
%i;3ko
HE F} T H C L
#!;#:5
|| B.rb.rlna|1 San.i.lmrlna
E F` T H C L
#!::£i
E R T 11 C L
%g:3ko
Figura lIH Cromatogramas de extractos de diferentes tej.idos de A. mexi.cana. A, 8,C, D, E, F, G, H, 1, J, K, L, M, N y 0 corresponden a las mezclas de disolventesdescritas en el Cuadro 111.1. Todas las mezclas fueron desarrolladas una sola vezexcepto J que fue desarrollada 3 veces. Cada carm fue cargado con un volu en deextracto equivalente a 5 mg de PS del tejido. E. Mezc`a de estándares de sanguinarina(fluorescencia naranja) y berberina (fluorescenc.ia amar.illa), R, T H, C y L, extractosde tallo, raíz, hoja, cápsula y látex. Los alcaloides fueron visualizados porautofluorescenc.ia bajo luz ultravioleta a 365 nm. Algunos de los colores pueden d.iferirde la apreciación que se describe en el texto debido a la fidelidad de la cámarafotográfica que se usÓ para tomar las imágenes.
38

| Berben.na|| SangLlinarina
Figura 111.2. Análisis de los alcaloides de A. mexí.cana por CCD de dos dimensiones.La primera separación de realizó con la mezcla J, mien{ras que la segunda con lamezcla cloroformo: acetona: metanol: ácido acético: agua 10..4:2:3..1. (A) Separaciónde una mezcla de estándares de sanguinaria (mancha naranja) y berberina (manchaverde) conteniendo 10 ng de cada alcaloide. (8) Separación de un extracto de raízcuyo volumen aplicado fue equivalen{e a 5 mg de PS. Los alcaloides fueronvisualizados por autofluorescencja bajo una lámpara de luz ultravioleta a 365 nm.
111.2. Cuantificación de alcaloides en tejidos de A. mex/.cana
Las canti.dades de sanguinarina y berberina en los djferentes tejidos se cuantificaron
por densitometría /.n s/.Íu. Para ello, los extractos fueron cromatografiados utilizando lamezcla J como fase móvil. Después de la separación, los alcaloides fueron analizados
por densitometría como se describió en la sección de materiales y métodos.
En los extractos de raíz, tallo, hoja y cápsula se detectó la presencia de sanguinarina
que fue identificada comparando las propiedades cromatográficas de la muestra conlas de un estándar comercial. Estas propiedades fueron los valores de Rf (0.52) y lafluorescencja naranja (Fig.111.1, panel J) La mayor cantidad de este alcaloide se
detectó en la raíz (50 Hg/g de PS). En los demás tejidos, este contenido fue unascinco veces menor (10 Hg/g de PS); Cuadro 111.2), mi.entras que no se detectó enextractos metanólicos del látex (Fig.111.1.J). La berberina, por su parte, se detectó en
todos los tejidos analizados, sjendo los extractos de tallo y látex los que mostraron elmayor contenido (125 ug y 6 mg/g de PS, respectivamente) (Cuadro 111.2).
39

Cuadro 111.2. Contenido de berberina y sanguinarina en dfferentes tejidos de A.
mex/cana reportados en Hg/g de PS. Promedio de tres repeticiones ± la desviaciónestándar.
Tejido Alcaloide (Hg/g PS)
Berberina Sanguinarina
RaízTalloHojaCápsulaLátex 48 ± 1 1 47 ± 3.6
125 + 4.8 11 ± 0.7
47 ± 1 .733±2 9 ± 0.3
11 ± 0.7
6000±620 ND
ND: No detectado
En otros trabajos, Ia acumulacm de berberina en raíces, tallos, hojas, frutos y flores
de A. mex/carffi fue del 0.10, 0.30, 0.50 y 1.30% de PS (Khana eí a/.,1982), estos
valores son superiores a los encontrados en este trabajo. Sin embargo, en el trabajo
cítado no reportan anális.S de látex, que fue donde se encontraron los mayoresnweles de berberina en nuestro trabajo.
Es interesante notar el elevado contenido de berberina presente en el látex. El látex es
una resina muy viscosa, que contk3ne azúcares, proteínas, lip.dos, terpenoides (Yoder
el aí.,1976) y, en el caso de las Papaveráceas, tambén cont.k3ne alcaloides (Facchini
ef a/,,1998). Se produce en células especializadas, llamadas laticíferas, que seencuentran asociadas con el tej.do vascular y ciue están d.Btribuk]os en dfferentes
órganos, como tallos y hojas (Yoder ef a/.,1976). De este modo, el alto contenido de
este alcaloide en tallos (Cuadro 111. 2) puede ser el reflejo de la incidencia de células
laticiferas en este tejido, como sucede en C. ma/.Lís (Tomé et a/.,1995). En contraste,
en r. Í/avum los mayores niveles de berberina (250 Lig/g PS) ocurren en raíces,
precisamente en la parte más madura, cerca de la base del tallo (Samanani ef a/.,2002), encontrándose menores niveles en otras regiones de la ra¡z, así como en elrizoma (entre 50-100 Lig/g de PS). En general, estos valores son comparables con los
encontrados en este trabajo para A mexí'cana (Cuadro 111.2).
En C./.apor¡/ca, Ios rizomas (tallos modfficados) representan el sitio de acumulación de
la berberina, aunque también se pueden encontrar en las raices principales y en
pequeñas cantidades en otras partes de la planta. Al parecer, las raices prmarias noson el principal sítio de sintesis de estos alcalok]es, puesto que algunas de las
40

enzi.mas i.nvolucradas, como la SMT, que cataliza la metilación de (S)-esculerina para
produci.r (S)-tetrahidrocolumbami.na (Fig. 1.3), ocurre principalmente en tallos y raíceslaterales (Fujiwara eí a/„ 1993) Estos resultados sugieren que en C. /.aponí.ca losalcaloides tipo protoberberjna son sintetizados en tallos o raíces laterales ytransportados para su acumulación a las raíces principales (Fujjwara eí a/.,1993). Losaltos contenidos de berberina en el látex, y en el tallo (en comparaci.ón con hojas y
raíces; Cuadro Hl.2), sugieren que algo simjlar podría ocurrir en A. mex/.cana.
A diferencia de la berberina, la sanguinarjna no se encontró en el látex, pero sÍ en
raíces, tallos, hojas y frutos (Cuadro 111.2). De este modo, se puede sugeri.r que estos
tejidos podrían ser sitios de síntesis y/o acumulación de este alcaloide, como ocurreen C. /.apon/.ca (Fujiwara ef a/.,1993) y P. somn/.rem (Facchini ef a/.,1995). En elcaso de P. somní.Íerum, exi.sten evidencias histoquímicas y enzimáticas de que el tallo
y la raíz son sitios de síntesis de alcaloides, pero no de acumulación (Facchini ef a/.,1995). En contraste, en S. canadensí.s se ha demostrado que el sitjo de síntesi.s y
acumulación de sanguinarina es la rai.z, pero el látex aislado de los tallos tiene un
color rojizo-naranja sugiriendo que puede ser transportado a las partes aéreas
(Mahady et a/.,1993; Schemeller ef a/.,1998). Combi.nando estos datos con nuestrosresultados, es posible sugeri.r que en A. mexí.cana la síntesis de sanguinarina ocurre
en la raíz, y que dado los bajos valores en tallo y su ausencia en el latex, no estransportado a los tejidos aéreos.
Por otro lado, además de la presencia de berberjna y sanguinari.na, los extractos de
los di.ferentes tejidos revelaron la presencia de una serie de componentes condiferentes propiedades fluorescentes. En este sentjdo, otros productos cuya presenciaha sido reportada en plantas del género Argemone como A. a/ba, A. p/aíyceras y A.h/brí.da incluyen alocriptopina, esculerina, quelantifolina, estilopina, queleritrina,
norqueleritri.na, re{iculina, coridina, isocoridina, magnoflorina, munitagina, platicerina,
metilplati.cerina y argemonina, así como algunos derivados de sanguinarina (por
ejemplo, norsanguinarina y dihjdrosanguinarina) (Krane eí a/„ 1984,. lsrailov eí a/.,
1986). Los patrones de los alcaloides presentes en los extractos de los diferentes
tejidos de A, mex/.cana revelaron la presencia de algunos componentes visibles condiferentes colores bajo luz UV a 365 nm. Los colores observados en el patrón debandas fueron amarillo, naranja, rojo, azul y morado. Desafortunadamente, no se
pudieron identificar con certeza muchos de los componentes que se observaron alseparar los extractos de los diferentes tejídos de A. mex/.cana, debido a que no sedisponían de estándares; sin embargo, se logró realizar una primera aproximaci.Ón enla identificación preliminar de algunos de estos metabolitos dado que existen datos en
41

la literatura para otras Papaveráceas, principalmente para P. somn/fertjm, en donde sehan identificado en análisis por CCD. Esta identificación preliminar se basó en las
características de migración de los compuestos en diferentes mezclas de disolventes
(valores de RO asi como por la fluorescencia que emiten cuando son observados bajoluz UV a 365 nm y por su reacción con algunas soluciones reveladoras desarrollandocolores característicos (Wagner ef a/„ 1984). De esta manera, los extractos de A.mexí.cana fueron separados en las diferentes mezclas de disolventes y tomando encuenta todos los parámetros mencionados arriba. Se observaron algunas manchascon fluorescencia en color amarillo que podrían corresponder a protopina, criptopina,
alocriptopina y argemonina, en tanto que algunas de las manchas en color azul que seobservaron podrian corresponder a quelidonina, homoquelidonina y norquelidonina.
Los valores encontrados en los extractos de los diferentes tejidos de A. mex/.cana eneste estudio es comparable a lo reportado para otras especies del género Argemone,como se mencionó anteriormente (Wagner eí a/.,1984; lsrailov ef a/.,1986).
111.3. Efecto de la luz sobre el cultivo de callos
Aún cuando se sabe que la luz altera el metabolismo de los alcaloides en diferentesespecies, como N. tabacum (Hobbs ef a/.,1991) y C. mseus (Vázquez-Flota ef a/.,1998), hasta ahora este efecto ha sido poco estudiado en la producción de alcaloidesderivados de tirosina.
En A. mex/.cana, previamente se había detectado una acumulación diferencial de
estos alcaloides en cultivos de callos tanto en respuesta a diferentes concentracionesde reguladores del crecimiento, como a diferentes condiciones de luz (Campos-Tamayo, 2002). Para profundizar en este aspecto, se d.iseñó un exper.imento concallos sometidos a condiciones de iluminación continua y de oscuridad a lo largo de un
ciclo de cultivo. Un lote de frascos con 1 g de callos de A. mex/.cana de 21 días de
cultivo fue mantenido bajo iluminación continua (50. umoles m-2 seg-'), mientras que
otro se mantuvo en oscuridad total. De cada lote se tomaron muestras cada 5 días yse les determinó el PF y PS, así como el contenido de alcaloides.
Los callos en luz mantuvieron su coloración verde, mientras que aquellos mantenidosen oscuridad perdieron este color, adquiriendo un tono ligeramente pardo (Fig. 111.3).
Los callos bajo iluminación continua presentaron iina etapa de retardamiento de 15
días, seguida de un rápido periodo de crecimiento que se extendió hasta 30 días deiniciado el experimento, no observándose mayor crecimiento durante el tiempo
restante (Fig.111.4). La acumulación de biomasa, medida como ganancia de PS, siguió
42

un compoftamjento similar (Fig. 111.4); sin embargo, el crecimien{o en PS fue
comparativamente menor que el PF, como lo demuestra la comparación de los ÍC'scalculados utjlizando ambos parámetros (Cuadro 111.3).
Por su parte, los callos mantenidos en oscuridad (Fig. 111.5) presentaron un menor
creci.miento durante el tiempo del experi.mento. Esto se refleja en los valores de ÍC y %de PS al final del ciclo (Cuadro 111.3).
Figura 111.3. Cultjvos de callos de A. mexi.oana en medio PC. (A) callos en luz
continua; (8) callos en oscuridad total. En ambos casos las fotografias se tomarondespués de 3 semanas de cultjvo.
43

10 20 30 40 50
DÍAS DE CULTIV0
Figura 111.4. Crecimiento de callos de A. mex/.cana a lo largo de un ciclo de cultivo de
50 días en presencia de luz evaluado como PF (.) y PS (E). Promedio de al menostres repeticiones .independientes, con la desviación estándar.
10 20 30 40
DÍAS DE CULTIVO
Figura 111.5. Crecimiento de callos de A. mex/.cana a lo largo de un ciclo de cultivo de
50 días en oscuridad evaluado como PF (.) y PS (D). Promedio de al menos tresrepeticiones independientes, con la desviación estándar.
44

Cuadro 111.3. Parámetros de crecimiento de los callos de A. mexi'cam cultivados bajoluz continua o en oscurídad.
Condición IC °/o PS al final del ciclo3.38LuzOscuridad 13.1 3
8.42 2.93
111.5. Acumulación de alcaloides
El análisís cromatográfico de los extractos de callos manteni.dos en ambascondiciones reveló la presencia de sanguinarina, pero no de berberi.na. La
acumulación de sanguinarina fue en promedi'o, mayor en los cultivos mantenidos bajoluz continua de acuerdo a los valores de Ltg/g de PS obtenidos (Cuadro 111.4). Más
aún, en estos cultivos mantenidos en luz se observó un ligero aumento en laacumulaci.Ón de este alcaloide a partir del día 35 y el máximo conteni.do se detectó en
el día 45 del ciclo de cultivo (929 Hg/g de PS) (Cuadro 111.4). Por su parte, en los
culti.vos mantenidos en oscuri.dad total la máxima acumulación de sanguinarina ocurrió
en el día 30 del ci.clo de cultivo (528 ug/g de PS). Estos contenidos se mantuvjeron
durante los si.guientes 10 di.as, disminuyendo ligeramente al final del ciclo (Cuadro
111.4). Estos valores son parecidos a los reportados en cultivos de P. somn/.fert/m (900
Hg/g de PS) mantenidos en condiciones normales. Sin embargo, en condiciones deinducción por BoÍo^/.s c/.nerea, los mi.smos culti.vos han alcanzado niveles de hasta 29
mg/g de PS (Eilert eí a/.,1984).
45

Cuadro 111.4. Conten.ido de sanguinarina en callos cult.ivados durante 50 dias en luz
cont.inua u oscuridad. Un volumen equivalente a 0.5 mg de PS se cargó en placas decromatografía y las muestras se separaron ut.ilizando la mezcla de disolventes J (vercuadro lH.1). Promedio de 3 repeticiones ± Ia desviación estándar.
Dias de cultivo Sanguinarina (Lig/g PS)
Luz Oscuridad
05 362 ± 9 278 ± 30
257 ± 5 224 ± 7
1015 262 ± 10 182 ± 9
357 ± 7 224 ± 40
20253035404550Promedio 345 ± 3 249 ± 30
252 ± 9 184 ± 10
180 ± 20 528 ± 70
430 ± 7 388 ± 80
394 ± 14 359 ± 20
929 ± 45 194 ± 40
221 ± 7 180 ± 20
362 271
Estos resultados sugieren que el efecto de la luz sobre la síntesis de sanguinarina
depende del crecimiento de los cultivos ya que la acumulación de este metabolito fuediferencial respecto al día de crecimiento de los callos (Cuadro 111.4); por otro lado, la
luz sÍ tiene un efecto sobre el patrón cualitativo de los ABl's, ya que puede definir la
presencia o ausencia de ciertos compiiestos.
Además de la presencia de sanguinarina, que es un producto final representativo de la
ruta de biosíntesis de alcaloides tipo benzonfentantridina, en los extractos de callosmantenidos en ambas condiciones de iluminación, Ios cromatogramas revelaron la
presencia de otras bandas con propiedades de autofluorescencia que no pudieron seridentificados debido a la carencia de estándares. No obstante, es interesante hacernotar que en los cultivos mantenidos en oscuridad, un total de tres manchas (connúmero asignado 2, 4 y 6) se encontraron ausentes en comparación con los cultivosbajo iluminación continua (Cuadro ln 5), sugiriendo que la iluminación tiene un papel
importante en la síntesis de estos compuestos. Más aún, en algunos casos la
46

intensidad de esas manchas mostró variaciones dependiendo del régi.men deiluminación bajo el que se mantuvieron los culti.vos (Cuadro 111.5 y ver apéndice A).
Se ha reportado que en cultivos de C. ma/.us manteni.dos en oscuridad, el peffn deABI's acumulados muestra la presenci.a de una mayor variedad de compuestosrespecto a cultivos de callos mantenidos en luz (Colombo eí a/., 1991); esto esopuesto a lo observado en los cultivos de A. mexí.cana. Esto sugi.ere que en cadaespecie, Ia luz tiene un papel diferencial en la regulación de la biosíntesis de los
alcaloides. Por otro lado, una comparación de los alcaloides presentes en plantasdiferenci.adas a partir de los cultjvos de callos de C. ma/.us mostró que en condi.ciones/.n v/.Íno no se producen quelidonina, queleri{rina ni berberina (Colombo eí a/., 1991);
esto concuerda con lo observado para A. mex/.cana ya que tanto en los cultivos decawos en luz como en oscuri.dad no fue posible detectar la presencia de berberina y
sugiere que, como en muchas otras especi.es, Ia organizacíón celular influye en el ti.po
de alcaloi.des que se producen (Facchi.ni, 2002). Cabe mencionar que se ha reportadola presencia de berberjna en células en suspensión de A. mex/.cana enconcentraciones de 0.030-0.051% de PS (Khanna eí a/.,1982).
Si bien es difícil establecer la identidad de las manchas observadas en loscromatogramas, debe comentarse que algunas características cromatográficas paraABI's han sido descri.tas en la li{eratura, incluyendo valores de Rf, propiedades de
autofluorescencia y reacción colorida a diversas soluciones reveladoras (Baerheim eía/., 1983). De este modo, es interesante notar la presencia de compuestos confluorescencia en azul (Cuadro 111.5), típica de los alcaloi.des del grupo de los
morfinanos (Wagner ef a/.,1984). Aquí debe mencionarse que, si bien existen reportesno confirmados de morfina en A. mexí.cana (Lozoya ef a/.. 1982), este alcaloide, al
igual que la codeína, solamente se acumula en P. somní.Íerüm, estando ausente aúnen otras especies del mismo género Papaver (Ei.len eí a/„ 1984). Es posible que la
presencia de alcaloídes relacionados expli.que esos reportes. Más aún, debido a losrequerjmientos de estructuras morfológicas especializadas, Ios cultivos /.n v/.ÍnJ de P.
somní.fewm no son capaces de produci.r morfina ni codeína (Facchini, 2001). Otrosalcaloides comunes en las Papaveráceas con fluorescencia azul incluyen a lanoscapina, la hidrastina y la papaverina (Wagner eí a/.,1984).
Como ya se comentó, en tejidos de díferentes géneros de Argemone, como A. a/ba, A.
p/aíyceras y A. h/br/da, se han encontrado alcaloides como protopína, alocrjptopina,esculerina, quelantifoli.na, estilopi.na, queleritrina, norqueleritri.na, reticulina, coridina,
isocoridina, magnoflorina, muni.tagina, platicerina, metilplaticerina y argemonina, así
como algunos derjvados de sanguinarina, incluyendo norsanguinarina y
47

dihidrosanguinarina (Krane ef a/., 1984; Israilov eí a/., 1986). No obstante, si estos
alcaloides también ocurren en cultivos /.n v/tro no ha sido ver.ificado. Sería interesante
determinar si alguna de las manchas cromatográficas presentes en los extractos delos callos, corresponde a alguno de ellos.
Cuadro m Comparación del patrón de manchas fluorescentes observado encultivos de callos mantenidos bajo luz continua y en oscuridad total. Un volumenequivalente a 0.5 mg de PS se cargó en placas de gel de sílice y las muestras sesepararon utilizando la mezcla de disolventes J (ver cuadro 111.1). Las placas fueron
visualizadas bajo luz UV a 365 nm.
Número Fluorescencia Rf Luz Oscuridad
asignado1 (365 nm)Naranja |Rojo1Azulclaro|Rojoladrillo|Azulvioleta|VerdeazullAzu,0.90 + +
0.78 + -2
0.66 + +34
0.62 + -
5 0.60 + +
678 0.49 + -
0.41 + +
NaranjaAmarilloAzulclaro 0,36 + +
0.21 - +910
0.19 + +
El efecto de la luz sobre la síntesis de alcaloides se ha documentado en diferentes-e=:¿:¿= -de Nicotiana (Hobbs et al., 1991)` =n Theob:?Ta Ca=a.=_!=U_r.n=! =t^na!.\.
1992), en C. ma/.us (Tomé eí a/.,1995), y en C. roseus (Vázquez-Flota eí a/.,1998).En cultivos de callos de W. g/auca, Ia luz disminuyó la acumulación de los alcaloides
mayoritarios anabasina, anabatina, nicotina y nornicotina, al igual que en cult.ivos de
raíces de N. rusí/.ca, sin embargo, el efecto de la luz pos.iblemente sea indirecto ya que
al parecer la acumulación de alcaloides se redujo como resultado del poco crecimientode las raíces que fue inhibido hasta en un 50% por efecto de la luz (Hobbs eí a/„1991). En cultivos de callos de 7-. cacao los alcaloides tipo purina, teobromina y
teofilina, se acumularon en mayor cantidad en condiciones de oscuridad que en
condiciones de fotoperíodo, en tanto que la acumulación de cafeína no tuvo
48

variacíones signjficativas en ambas condici.ones; sin embargo, los niveles de
acumulación observados fueron muy variables (Gurney ef a/., 1992). En C. ma/.us sedetectó la presencia de sanguinarina, quelidonina, queleri.tirina, berberina y coptisina
en extractos de planta asi como en el látex, y se observó que. excepto la berberinacuya acumulación no varió respecto a las condíciones de iluminación, los ni.veles de
acumulación de los otros alcaloides fueron mayores en los periodos de luz, en tanto
que en los períodos de oscuridad los contenidos de los mismos disminuyeron (Toméeí a/.,1995). En C. roseus la luz es indispensable para la síntesis de vindolína, una de
las subunidades de los alcaloides citotóxicos vinblastina y vincristina (Vázquez-Flota ef
a/.,1998a). En esta planta, este efecto se ejerce activando las tres últi.mas enzimas dela síntesis de este alcaloide (Vázquez-Flota eí a/.,1998b). Si bien este alcaloide está
ausente en los cultivos /.n vffno de esta especie, en callos la luz promueve latransformación de ajmalicina en serpentina (Monforte-González eí a/„ 1992). En
cuanto a los ABl's. no se tienen reportes del efecto de la luz sobre su síntesis.
111.6. Establecimlento de Jos cLiltivos /.A v/.Íro
El cultivo de células en suspensión se derivó de un cultivo de callos, previamente
obtenido de hojas (Campos-Tamayo, 2002). El cultivo de células en suspensión,
comparado con el de callos, presenta las ventajas de una mayor velocidad decrecimíento y la facilidad de manipulación. Con el objetivo de establecer una
suspensión a partir de los callos, matraces de 125 mL. conteniendo 25 mL de medio
PC, fueron inoculados con 1 g de callos de 21 días de cultívo. Al cabo de dos
semanas se pudo apreciar la disgregación del tejido, que seguía manteniendo lacoloración verde típica y un crecimiento notable (Cuadro 111.6; Fig.111.4). No obstante,
también se apreció una coloración grisácea en el medio residual, posiblemente poralgún tipo de estrés osmótico debido al alto contenido de sales en el medio de cultivo.
Por lo anterior, se decidjó resembrar parte de las células en un medío de menorcontenido se sales, como lo es el 85 de Gamborg (Gamborg e{ a/.,1968). Los cultivos
en el medio 85 mantuvíeron un color verde símilar al de los callos durante el primer
ciclo y también presentaron un marcado crecimiento (Cuadro Hl.6). Sin embargo, al
igual que en el medio PC, los cultivos adquirieron un color negro (datos no
mostrados), pero sólo después de períodos largos de subcultivo (entre 15 y 20resiembras quíncenales). En trabajos previos había sido detectada la rápida oxidaciónde los cultivos en suspensi.ón (Campos-Tamayo, 2002), aún utilizando aguadesionizada Tipo 1 (resistividad menor a 18 megaohm-cm), obtenida mediante la
resjnas EASYpure, Ultra Low Organic (Barnstead lnternational). Es posible que el
49

activo metabolismo de los derivados de tirosina de esta planta, combinado con lamayor disponibilidad de oxígeno ocasionada por la agitación, resulte en la oxidaciónde compuestos fenólicos, como se ha observado en callos de tabaco en los que estascondiciones promueven la actMdad de la polifenol oxidasa (PFO) y de las
peroxidasas, llevando a la acumulación de melaninas (Negrel eí a/.,1993). Más aun, latirosina puede dar lugar a la formación de productos, como los 2-(2-feniletil) cromonas
que, al oxidarse, adquieren iina coloración oscura (Qi ef a/., 2005). La aplicación deantioxidantes, como el ácido cítrico o el ácido ascórbico en diferentes dosis, solos o encombinaciones, así como de cisteína, no eliminó el oscurecimiento de los cultivos, porlo menos en las concentraciones utilizadas en este trabajo (datos no mostrados)
Figura 111.6. Aspecto de las células en suspensión de A. mex/.cana mantenidos en
condiciones de luz continua a los 14 días de cultivo en el medio 85 (A) y PC (8).
50

Cuadro 111.6. Crecimiento evaluado como PF por matraz, de cultivos de A. mex/.cana
manteni.dos en medios líquidos.
Resiembra lnóculo (g PF) Índjce de crecimiento
sÓ/„
Medio 85 1 Medjo PC3.71 *1.0 t23 1.5 7.0 4.2
2.6 4.4 NECallo de 21 días de edad, previamente mantenido en medio PC semi
s.up£mentado con 25 gft de sacarosa y 1.50 mgft de ANA y O.5ó mBAP
+ L_os cultivos fueron transferidos a medio 85 después de la primera resiembra
NE No evaluado
De este modo, con el fin de realizar los experi.mentos de inducción y dada la facilídad
con la que se disgregaban los callos en el medio líquido, se decidíó utilizar
suspensiones recién generadas, según se hiciera necesario.Es interesante notar que las células en sí no tienen este color negro, si bien nomantienen el verde típico de los callos. La tinción con colorantes vitales, como el azul
de tripano, ha demostrado que las células se mantienen viables, aunque no soncapaces de crecer (Campos-Tamayo, 2002). Se ha observado que en callos de tabacola tiramina puede dar lugar a compuestos tóxicos, como la indolquinonas, que se
forman a partir de la oxidacíón de ésta por acción de la PFO (Negrel eí a/„ 1993).
Puesto que los cultivos /.n vt.Íro de Argemone son capaces de sintetizar alcaloidesderivados de tirosi.na, también pueden acumular algunas cantidades de tiramina, que
es el primer paso en su síntesis (Figura 1.1). De este modo, las condiciones de excesode oxigenacjón, propi.o de la agitación de los cultivos en suspensión, puede promover
su oxidación en compuestos de este tipo, qLie son de color oscuro (Negrel ef a/.,1993). El cultivo de las células en suspensión en condiciones que inhiban la actividad
de la PFO podría reducir o evitar el oscurecimi.ento de los tejidos.
111.7. Efecto del MeJa en cultivos en suspensión de A. mex/-cana
Además de las condiciones de iluminacjón, se ensayó el efecto de inductores
químicos de la síntesjs de metabolitos secundarios. Se seleccionó el MeJa debido aque este inductor participa como medi.ador de la síntesis de alcaloides en respuesta adiferentes condiciones de estrés, como el daño mecánico y las infecciones fúngicas
51

(Weiler eí a/., 1993, lgnatov eí a/.,1996). Más aún, los efectos de este compuestosobre la producción de alcaloides isoquinolínicos está bien documentada (lgnatov eí
a/.,1996).
111.7.1. Deternriinación de la dosis de NleJa
Con el fin de encontrar la concentrac.ión de MeJa que permitiera la mayoracumulación de alcaloides, 5 g (± 0.75) de células de 5 días fueron expuestas duranteun periodo de 24 h. Para ello, se preparó una solución patrón 100 mM de MeJa
(Sigma-Aldrich) diluyéndolo en DMSO. El volumen de la solución patrón de MeJa seajustó de tal modo que los tratamientos, aplicados a 25 mL de suspensión celular,correspondieran a 10, 100 y 1000 L.M. Se realizaron dos tipos de testigos; en el
primero, una serie recibió los mismos volúmenes de disolvente, mientras que en elsegundo, los cult.ivos se mantuvieron en las condiciones habituales de cultivo. Los
diferentes tratamientos se aplicaron por triplicado y fueron cosechados después de 24h por filtración al vacío.
El tratamiento con MeJa, produjo un ligero oscurecimiento tanto en la células como en
el medio residual, que no se observó en los testigos (Fig. 111.7). La coloración que
adquirió el medio tuvo una tonalidad amarillo-naranja, por lo que se analizó el
contenido de sanguinarina en el medio (Cuadro 111.7), ya que este compuesto en
solución pudiera conferir esta coloración.
La mayor respuesta respecto al contenido de sanguinarina se obtuvo en los cultivosexpuestos a 100 y 1000 ttM, cuya acumulación aumentó aproximadamente 7 vecesen comparación con el testigo; no obstante, no se notaron diferencias entre estos dostratamientos (Cuadro 111.7). Por otra parte, la cantidad de sanguinarina liberada a los
medios de cultivo fue muy baja en todos los casos (del orden de 1 Hg/mL) por lo que lacoloración observada en el medio parece no deberse a la presencia de este alcaloide
(Cuadro 111.7).
52

`.,\,#
8,
Figura 111.7. Aspecto de la suspensi.Ón celular de A. mex/.cana (A, 8, C y D) y de los
medios libres de células (A', 8', C' y D'). Células de 5 días sometidas a 0 (testigo; (A),10 (8),100 (C) y 1000 (D) uM de MeJa durante 24 h.
Cuadro 111.7. Efecto del MeJa sobre la acumulaci.Ón de sanguinarina en cultivos en
suspensión de A. mex/.oana.
Tratamiento Contenido de sanguinarinaCélulas (LLg/g PS)Í42±3 1 Medio (Hg/mL)10.50±0.35
TestigoDMSOo.oio/oMeJa10LLM+ DMS0 0.01°/o|DMSO0.1%MeJalooLiM+DMSOO.1°/o |
33±1 0.50 ± 0.3636±7 1.21 ± 0.42
45±2 0.50 ± 0.10292±14 187±41 0.94 ± 0.45 10.32±0.011
DMS01°/o ÍMeJal000LLM+DMS01°/o|
295 ± 5 0.42 ± 0.01
53

Tanto en los cultivos expuestos a 1000 LiM de MeJa, como en los correspondientestestigos con el disolvente (1% DMSO v/v), se notó un rápido oscurecimiento, en las
primeras12h,yunadisminucióndelamasacelularinicialquenofuepercibidaenloscasos anteriores. Estos datos sugieren que el tratamiento estaba produciendo dañosfisiológicos, complicando las posibles interpretaciones de los aumentos observados
(Cuadro111.8).Másaún,aestasconcentracionesloscultivoscomenzaronamorirpocodespués de las 24 h, sugiriendo un efecto tóxico del inductor.
Es interesante observar que en altas concentraciones de DMSO (1 %), éste por sÍ solofue capaz de produch un aumento de casi 2 veces en el contenido de sanguinarina encomparación con el testigo, no expuesto a este disolvente (Cuadro 111.7). EI DMSO ha
sido utilizado como inductor en cultivos de otras especies como C. roseus (Bonzom e!a/., 1997 y 1998), en los que produce un aumento en la acumulación de ajmalicina,
cuando la dosis es de alrededor del 1% (v/v). En el caso de A. mex/.cana, se pudoobservar que concentraciones s.imilares a ésta producen un efecto en la acumulación
de sanguinarina. Sin embargo, estos efectos se pueden explicar debido a que elDMS0 funciona como un agente permeabilizante y solubilizante (Brodelius e! a/.,
1983), Io cual puede estar confiriendo algún tipo de estrés ine§pecifico a las células y
esto se refleja en el aumento de la acumulación de sanguinarina. Con el fin de
confirmar que los efectos observados eran debidos al inductor (MeJa) y no aldisolvente (DMSO), se realizaron experimentos diluyendo el inductor con etanol. Los
resultados obtenidos fueron similares a los observados con el DMSO, sugiriendo que,
en las concentraciones ensayadas, los efectos de deben al MeJa y no al disolvente.
Puesto que los tratamientos, aun en el corto periodo de exposición, afectan elcrecimiento, Ios resultados fueron recalculados como productividad de alcaloides por
matraz, al relacionar los valores en Lig/g de PS y Hg/mL de medio de cultivo (Cuadro
111.7)conelcontenidoenunmatraz.tantodePScomodevolumendemediodecultivo
(Cuadro 1" 8). Este parámetro considera no solo el efecto sobre la sinte§is dealcaloides, sino también sobre el crecimiento. Los datos asi obtenidos confirmaron quelos cultivos expuestos a 100 y 1000 HM de MeJa fueron los que indujeron la mayor
capacidad de sintesis de sanguinarina (Cuadro 111.8). Con base en estos resultados, y
debido a que con 100 LiM fue posible recuperar las células en buen estado y viables, adiferencia de las células tratadas con la mayor concentración, se seleccionó eltratamiento con 100 LiM para analizar con más detalle el efecto del inductor.
54

Cuadro 111.8. ProductMdad de sanguinarina del cultivo de A. mex/.cana expuesto a
diferentes concentraciones de MeJa y DMSO.
Tratamiento Contenido de sanguinarinaCélulas Medio* ProductividadLLg/matraz ltg/matraz Hg/matraz
Testigo 14.7 12.5 27.2DMSO 0.01% 10.6 12.7 23.3MeJa 10 üM + DMSO 0.01% 11.2 30.3 41.5DMS0 0.1% 19.4 12.5 31.9110. 942.8MeJa 100 HM + DMSO 0.1% 87.6 23.3DMSO 1% 34.8 8.0MeJa 1000 LLM + DMS01% 85.6 10.5 96.1*El volumen de la suspensión fue de 25 mL
111.7.3. Curso temporal de acumulación de alcaloides en cultivos inducidos conA,eJa
Se realizó un curso temporal exponiendo cultivos de 5 días a 100 uM de MeJa disueltoen DMSO durante 0, 6, 12 y 24 horas. Las condiciones fueron similares a lasanteriores y los tratamientos se aplicaron por triplicado. Después del perjodo de
exposición, los cultivos fueron colectados por filtración al vacío, conservando tanto eltejido como el medio residual. La Figura 111.8 muestra el aspecto de las células en
suspensjón después del tratamjento.
Figura 111.8. Células en suspensión de cinco días de cultivo expuestas a MeJa 100 uM
durante 0 (A); 6(8); 12, (C) y 24 (D) h.
55

Los efectos del inductor en cuanto al aumento de la acumulación de sanguinarina, se
presentaron a partir de las 12 h de exposición y llegaron al máximo después de 24 h
(Cuadro 111.9). Si bien en estas condiciones la inducción de la síntesis de sanguinarinaaumentó cerca de 10 veces respecto al testigo, estos valores fueron bajoscomparados con los observados en P. somni.fert/m, en donde el rango de acumulaciónde sanguinarina en condiciones de inducción llega al orden de mg (Eilert e{ a/.,1985;
Facchini ef a/.,1994).
Cuadro 111.9. Curso temporal de la acumulación de sanguinarina en cultivos en
suspensión de A. mex/.caria tratadas con MeJa 100 LiM.
Tratamiento Contenido d® sanguinarina
Cólulas (Hg/g PS) Medio (LLg/mL)
Oh 40 ± 0.6 0.33 ± 0.10
6h DMSO 0.1% 44 ± 1.6 0.48 ± 0.03
6 h MeJa + DMS0 0.1% 45 ± 2.2 0.29 ± 0.10
12 h DMSO 0.1% 33 ± 0.3 0.30 ± 0.05
12 h MeJa + DMSO 0.1% 227 ± 10 0.44 ± 0.11
24 h DMSO 0.1% 20 ± 0.5 0.24 ± 0.04
24 h MeJa + DMSO 0.1% 442 ± 13 0.35 ± 0.15
Es importante enfatizar que el experimento no pudo extenderse por mayores periodos
de tiempo debido a que los cultivos tomaban una coloración grisácea y finalmente se
oscurecían, lo cual era un indicio de necrosis debido al estrés al que estaban
sometidas las células, con la consecuente muerte de las mismas.
56

Cuadro 111.10. Productividad de sanguinarina en cultívos de A. mex/.cana expuestos a
100 LiM de MeJa durante diferentes tiempos.
Tratamiento Contenido de sanguinarinaCé'ulas Medio ProductividadtJg/matraz LLg/matraz ug/matraz
Oh 8.4 8 16.4
6h DMSO 0.1% 7.9 12 19.9
6 h MeJa + DMS0 0.1% 9.9 7.3 17.2
12 h DMS0 0.1% 7.6 7.5 15.1
12 h MeJa + DMSO 0.1% 45.4 11 56.424 h DMS0 0.1% 4.2 6 10.2
24 h MeJa + DMSO O.1°/o 97.2 8.8 106
*El volumen de la suspensión fue de 25 mL
La productividad del cultivo alcanzó valores similares a los del experimento de dosis
respuesta, y al igual que en éste, el mayor aporte de sanguinarina se encontró en lascélulas, siendo alrededor del 10% del total de alcaloides por matraz la cantidad
recuperada del medio (Cuadro 111.10).
Por otra pane, aún en las condiciones de inducción aplicadas, no se notó la presenciade berberina, si bien el patrón cualítativo de manchas cromatográficas sÍ mostró
algunas variaciones, en comparación con los cultivos no expuestos al inductor
(Cuadro lll.11 ).
No obstante, en algunos casos el patrón cualitativo mostró algunas variaciones tanto
en la presencia o ausencia de ciertas bandas, como en la intensidad de las mismas. Sibien estos compuestos no han podido ser identificados, mostraron tinción positiva al
reactivo de Draggendorff, sugiriendo que se trata de alcaloides. De este modo, Ias
manchas con número asignado 2, 6, 10, 13 y 14 desaparecieron el los cultivosexpuestos a MeJa (Cuadro HI.11). Por el contrario, la banda 5, que produjo una
fluorescencia típica naranja, sólo pudo observarse en los cultivos expuestos a 1 mM
de MeJa. Estos resultados podrían explicarse como variaciones en la cantidad deintermediarios biosintéticos, posiblemente por su conversión en los productos fínales.
Este típo de variaciones también fueron observadas en función del tiempo (datos nomostrados).
57

Cuadro 111.11. Valores de Rf del patrón de compuestos fluorescentes a 365 nm, noident.ificados, observado en cultivos de células en suspensión en respuesta a
diferentes concentraciones de MeJa.
Número Color R, T DMSO MeJa 10 MeJa MeJa
asignado (365 nm) 1% tJM 100 LJM 1000 HM
1 Naranja 0.76 + + + + +
2 Morado 0.69 + ++ + + -
3 Rojo naranja 0.69 ++ + ++ ++ +
4 Azul claro 0.65 + ++ + ++ ++
5 Naranja 0.64 - - - - +
6 Azul 0.64 + + ++ + -
7 Amarillo 0.62 + - - - -
8 Amarillo 0.53 + + + - -
9 Azul 0.52 ++ +++ ++ ++ +
10 Amarillo 0.50 + + + + -
11 Morado 0.45 + + + + -
12 Amarillo 0.35 + + + + +
13 Azul claro 0.29 + ++ + ++ -
14 Amarillo 0.22 + + + ++ -
T Testi9O
+ Presencia del compuesto- Ausencia del compuesto
111.8. Construccjón de una genoteca de ADNc de A. mex/.cana
Para la construcción de la genoteca de ADNc se seleccionaron cultivos de callos decinco días, mantenidos en condiciones de iluminación continua. Estos cultivos se
encontraban al inicio de la fase de crecimiento exponencial (Fig. 111.4) y fueron usados
con este fin debido a su capacidad de acumulación de alcaloides en tales condiciones
(Cuadro 111.4), lo que sugiere que las enzimas involucradas en la síntesis de losalcaloides se encuentran presentes, posiblemente al igual que los transcritoscorrespondientes. Si bien esta capacidad de síntesis puede ser incrementada cuandolos cultivos son sometidos a condiciones de inducción (Cuadro 111.7), la calidad del
ARN obtenido de los cultivos expuestos a MeJa no fue la adecuada para laconstrucción de la genoteca, debido al daño fisiológico que presentó el material
vegetal durante la exposición con el Ínductor.
58

Los detalles técnicos de la construcción de la genoteca están descritos en la seccióncorrespondiente a Materiales y Métodos. Siguiendo esos pasos, el título primario de la
genoteca fue de 848,000 UFC/mL, lo cual indica una alta probabilidad de que lostranscritos expresados en el ARN en el momento de la elaboración de la genoteca, seencuentren representados en ella. Con el fin de incrementar ese valor, la genoteca fueamplificada en fagos y después de un ciclo se logró elevar dicho valor al orden de 109UFC/mL (Fig.111.9) y se obtuvo un total de aproximadamente 150 mL del lisado en el
amortiguador SM que se mezclaron con un vigésimo de volumen de clorofomo y sealmacenaron en alícuotas a -70 °C. Considerando que A, mex/.cana es una especiediploide de 28 cromosomas (2n= 28); (Chaturverdi eí a/., 1999), en una genoteca coneste título existe una alta probabilidad de encontrar representados a la mayoría de lostranscritos presentes en el ARNm a partir del cual se construyó la genoteca, aún losde baja expresjón (Sibson eí a/.,1997).
El valor de esta genoteca de ADNc radica en que permite tener una colección de los
genes involucrados en la ruta de biosíntesis de ABl's en A. mexi.cana, a partir de lacual se pueden aislar con el fin de estudiar la expresión de genes en los estudios
posteriores sobre los mecanismos de la regulación del metabolismo de estosalcaloides.
Figura 111.9. Aspecto de una caja de Petri usada para realizar la titulación de la
genoteca de ADNc de A. mex/.cana.
59


CAPITUL0 lv
CONCLUSIONES Y PERSPECTIVAS
El método para la detección y cuantificación de los alcaloides se basó en unaseparación por cromatografía de capa fina, seguida por densitometría /-n s/.Íu. El
desarrollo de este método se basó en reportes previos sobre las propiedades deestos alcaloides, en diferentes fases móviles (Cuadro 111.1). El límite de detección
del método desarrollado fue del orden de nanogramos, en comparación con otrosbasados en cromatografía líquida donde es posible detectar niveles inferiores delorden de picogramos debido a la sensibilidad de los equipos utilizados para
desarrollar la metodología (Schieffer ef a/., 2002). Si bien el método resultó fácil de
implementar y adecuado para las necesidades del proyecto, tiene el inconveníente
de no ser posible de automatizar en las condiciones de análisis que se usaron
para este estudio (Cserhátí ef a/„ 1996).
El análisis de los alcaloides en los diferentes tejidos de la planta demostró la
presencia de sanguinarina y berberina, además de otros compuestos que nopudieron ser identificados, pero que fueron positivos a la tinción con el reactivo deDraggendorff, el cual es selectivo para alcaloides (datos no mostrados). No
obstante, es posible que sean del mismo grupo de las protopinas, ya que, a
diferencia de otras especies del género Angemone, la especíe A. mex/.canasolamente acumula alcaloides de este grupo (Schwarzbach ef a/.,1999). Si bien
exísten reportes sobre el perfil de alcaloides en esta planta (Lozoya,1982), se han
observado diferencias cualitativas que al parecer no son solo causadas por losfactores medioambientales, sino que también tienen un componente genéti.co, ya
que son específicos de ciertas poblaciones (Schwarzbach eí a/.,1999). Puestoque la mayoría de los estudios fitoquímicos en esta especie se han realizado conejemplares provenientes de las regiones del noroeste de México/suroeste deEstado Unidos, del Centro de México y de la lndia que difieren entre sÍ, debido a
díversos factores genéticos y a que están expuestas a diferentes condicic>nesambientales (Schwarzbach eí a/., 1999), es posible que pc>blaciones de la
Península de Yucatán presenten un patrón cualitativo distintivo. De este modo, la
identificación de los alcaloides en los díferentes tejídos de esta planta debe ser
consíderada en futuras investigaciones.
Por otra parte, se ha logrado desarrollar un sistema de cultivo /.n v/.Íro. No obstante,
ha sido complicado llevarlo al estado de células en suspensión, principalmente porel oscurecimiento de las suspensiones que logran establecerse (Figura 111.6). Si
bien los callos también muestran zonas de tejido oscuro, éste adquiere una
tonalidad rojiza, es necrótico y generalmente está en contacto directo con el medio
(Figura 111.3). A diferenci.a de esto, las suspensiones celulares adquieren un colornegro intenso y cesan su crecimiento (datos no mostrados)
61

En los cultivos /.n v/.fm se pudo identificar la presencia de sanguinarina, pero no de
berberina, que sí se encontró en los tejidos de la planta madura (Cuadros 111.2 y
111.4). Si bien para algunos alcaloides derivados de la tirosina, como la morfina y la
codeína, es necesario cierto nivel de organización tisular (Facchini ef a/.,1994),
éste no parece ser el caso para la síntesis de berberina, ya que su presencia hasido detectada en cultivos /.n vt.Írio de diferentes especies, incluyendo A. mex/.car)a
(Khanna eí a/., 1982; Amman eí a/„ 1986). De este modo, la pérdida de lacapacidad de síntesis de berberina parece ser una característica propia de la línea
celular generada en este trabajo.
En los cultivos mantenidos bajo diferentes condiciones, se lograron observaralgunas diferencias interesantes en los patrones de acumulación de alcaloides. De
este modo, en cultivos bajo diferentes condicíones de iluminación, aunque el
contenido de sanguinaria no varió de manera significativa en comparación con
aquéllos mantenidos en oscuridad continua (Cuadro 111.4), el periil cualitativo si
mostró algunas variaciones (Cuadro 111.5). La identidad de estos compuestos no
pudo ser analizada, pero debe ser considerada en futuros trabajos.
Además de los efectos de la luz, se pudo observar que los cumvos /.n vÍ.fro son
susceptibles de acumular alcaloides en respuesta a inductores químicos, como elMeJa (Cuadros 111.7 y 111.8). Cuando los cultivos de células en suspensión fueron
expuestos a este inductor, se observó una mayor acumulación de sanguinarina, elcual es un producto final representativo de la ruta de alcaloides tipo
benzofenantridina (Figura 1.1). Como ya se mencionó, aún en condiciones de
inducción no fue posible detectar la presencia de berberina, que es un alcaloide
del tipo protoberberina, ni tampoco alcaloides tipo morfinano (Cuadro 111.7). En
cultivos de otras especies, como S. canac/ensi`s (Mahady eí a/., 1993) y E.
ca//.forní.ca (Haider eí a/., 2000) la exposición de cultivos en suspensión a
jasmonato induce la acumulación de sanguinaria, lo que es precedido por laactivación transcripcional de algunas enzimas como la TyDC (Facchini eí a/.,
1995) y la BBE (Dittrich ef a/.,1991). El jasmonato es un medíador de la respuesta
a diferentes estímulos ambientales que incluye la síntesis de alcaloides y otros
productos naturales involucrados en la defensa química contra patógenos yherbívoros (Cheong eí a/., 2003). De hecho, en cultivos de C. mseus laacumulación de jasmonato ocurre en respuesta a la inducción con homogenadosfúngicos y ésta precede la activación de genes de la síntesis de alcaloides (Menke
eí a/.,1999). En A. mex/.cana, experimentos preliminares han demostrado que la
exposición de cultivos a homogenados fúngicos de BoÍvÍ/.s spp resultaron en el
aumento de la actividad de TyDC (datos no mostrados). Estas observacionessugieren que el aumento en la acumulación de sanguinarina en respuesta alinductor posiblemente fue consecuencia de un aumento de la actividad de
enzimas involucradas en su síntesis.
62

Finalmente, se logró la construccíón de una genoteca de ADNc de A. mex/.cana.
Después de amplificada, esta genoteca alcanzó un título en el orden de 109UFC/mL lo cual, junto con el hecho de que el tejido seleccionado tenía la
capacidad de acumular sanguínarina, permite que exista una alta probabilidad de
que se encuentran representados los transcrjtos de los genes involucrados en labíosíntesis de alcaloides, así como los que participan en su regulación. Si bien no
se realizó la búsqueda primaria de genes dentro de la genoteca, ésta representauna valiosa herramienta para futuros estudios, dadas sus características.
En resumen, en este trabajo se lograron establecer las bases para el estudio de A.mex/.cana, una planta de importancia en la medicína tradicional mexicana. Estas
bases incluyeron el desarrollo de valiosas herramientas, como un análisis
fitoquímico básico, un sistema de cultívo Í.r) vÍ.Íro y una genoteca de ADNc de la
cual se podrán aislar genes relevantes en la síntesis de alcaloides.
Este trabajo deja abiertas algunas interesantes posibílidades para realizar estudiosmás profundos, como la naturaleza de los alcaloides presentes en los diferentestejidos de la planta, o los que resultaron modificados en respuesta a las díferentes
condiciones de cultivos. Las bases fisiológicas del ennegrecimiento de los cultivos
también es un tema interesante para estudiar. Finalmente, si bien los alcaloi.des de
Aüemone tienen aplicaciones médicas, también son causa de seriasintoxicaciones debidas al consumo, voluntario o no, de sus semillas en algunos
países, como la lndia (Verma ef a/., 2001). Debe hacerse notar que las semillas deesta planta son una fuente de aceites de muy buena calidad que no puede serexplotada debido a la presencia de alcaloides que pueden ser tóxícos en altasconcentraciones, principalmente sanguinarina (Lozoya, 1982). De este modo, el
estudio del metabolismo de los alcaloides en esta planta podría llegar a desarrollar
estrategias para la reducción o eliminación de estos compuestos, y permitir su usocon fines comerciales.
63


REFERENCIAS
Amman M, Wagnner G, Zonk MH, (1986). lntracellular compartmentation of two
enzymes of berberine bíosynthesis in plant cell cultures. P/anfa.167: 310-320.
Argueta Villamar A, Cano, Asseleich LM, Rodarto ME, (1994). Af/as de /as p/aníasde /a med/'c/.na fradí.ci.ona/ mexí.cana /. lnstituto Nacional lndigenista. Primera edición,Méxíco D.F. Pp. 399-401.
Baerheim Svedensen A, Verpoorte R, (1983). Chromafography of a/ka/o/.ds. Pari A..77w.n /ayer chromafography. Department of Pharmacognosy, State University Leyden,Leyden. The Netherlands Elsevier Scientific Publishi.ng Company, Amsterdam-Oxford-
NY. Pp.171-220.
Bauer W, Zenk MH, (1989). Formation of both methylendíoxy groups in the alkaloid
(S)-stylopine is catalyzed by cytochrome P450 enzymes. refrahedron Le#. 30: 5257-5260.
Bauor W, Zenk MH, (1991). Two methylenedioxy bridge forming cytochrome P450
dependent enzymes are involved in (S)-stylopine biosynthesis. Phyfochem/.sÍv. 30:
2953-2961.
Blechert S, Brodschelm W, Hólder S, Kammorer L, Kutchan TM, Mueller MJ, XlaZQ, Zenk MH, (1995). The octadecanoíc pathway: Signal molecules for the regulatíon
of secondary pathways. Proc Naí/ Acad Sc/. USA. 92: 4099-4105.
Bonzom MP, Andary C, Guoiffior A, Roussel JL, Gargadennec A, (1997). Effect ofa tertiary amine on alkaloid accumulation in Cafharaníhus noseus. Phyfochem/.sfry. 46:235-239.
Bonzom MP, Gargadonnoc A, Andary C, Roussel JL, Gueiffier A, (1998). Alkaloidrepartition and DMSO effects on alkaloid accumulation in Cafharan£hus seedlings. J
Plant Physiol. 153.. 534-538.
Brodelius P, Nilsson K, (1983). Permeabjlization of immobílized plant cells, resulting
in release of intracellularly stored products with preserved cell viability. Eur J App/
Microbiol Biotechnol. 17.. 275-280.
Byun SY, Pedersen H, Chin CK, (1990). Two-phase culture for the enhanced
production of benzophenanthridine alkaloids in cell suspensions of Eschscho#z/.acalifornica. Phytochemistry. 29.. 3135-3139.
65

Byun SY, Pedersen H, (1994). Two-phase airlift fermentor operation with elicitation forthe enhanced production of benzophenanthridine alkaloids in cell suspensions of
Escherichia californica. Biotechol Bioeng. 44..14-20.
Campos-Tamayo FD, (2002). Efectos de los procesos de diferenciación celular sobrela síntesis de alcaloides en cultivos /.n v/.fro de Argemone mex/.cana y Caíharanfhusroseus. resí.s de //.cencí.afura. Químico Farmacéutico Biólogo. Facultad de Química.
UADY. Mérida, Yucatán, México. Pp.34-35.
Chaturvedi M, Datta K, Pal lvI, (1999). Pollen anomaly -a clue to natural hybridity in
Argemone (Papaveraceae). Grana. 38:339-342.
Chen YY, Chang FR, Wu YC, (1996). lsoquinoline alkaloids and lignans form Ro///.n/.a
mucosa. J Nat Prod. 59.. 904-906.
Cheong JJ, Choi YD, (2003). Methyl jasmonate as a vital substance in plants. TmndsGen.19: 409-413.
Chintalwar GJ, Gupta S, Roja G, Bapat VA, (2003). Protoberberine alkaloids fromcallus and cell suspension cultures of ri.nospora cordi.Ío/Í.a. Pharm Bi.o/. 41 : 81-86.
Chou WIvl, Kutchan TM, (1998). Enzymatic oxidations in the biosynthesis of complex
alkaloids. P/anf J. 15: 289-300.
Cline SD, Coscia CJ, (1988). Stimulation of sanguínarine production by combined
fungal elicitation and hormonal deprivation in cell suspension cultures of Papaver
bracteatum. Plant Physiol. 86..161-165.
Colombo ML, Tome F, (1991). Production of sanguinarine by Che/i.doní.um ma/.t/s
callus cultures. P/anía Med. 57:428429.
C§erháti T, Forgács E, (1996). lntroduction to techniques and instrumentation.
Practical Thin-Layer Chromatography. A Multidisciplínary Approach, (eds. Fr.ied 8,Sherma J). CRC Press lnc. Boca Raton, Florida. Pp: 1-18.
Davis KR, Lyon GD, Darvill AG, Albersheim P, (1984). Endopolygalacturonic acid
lyase from Erw/.n/'a caroíovora elicits phytoalexin accumulation by releasing plant cell
wall fragmen`s. Plant Physiol. 74.. 52-60.
De-Eknambul W, Z®nk MH, (1992). Purification and properties of 1,2-
dehydroreticuline reductase from Papaver somn/.fert/m seedlings. Phyfochem/.sfry. 31 :
81 3-821 .
66

Dittrich H, Kutchan TM, (1991). Molecular cloning, expression, and induction of
berberine bridge enzyme, and enzyme essential to the formation ofbenzophenanthridine alkaloids in the response of plants to pathogenic attack. ProcWaí/ Acad ScÍ.. USA. 88: 9969-9973.
Eilert U, Constabel F, Kuiz WGW, (1986). Elicitor-stimulation of monoterpene índole
alkaloid fomation in suspension cultures of Caíharaníht/s roseus. J P/aní Phys/.o/.126:11 -22.
Eilert U, Kurz WGW, Constabel F, (1985). Stimulation of sanguinarine accumulation
in Papaver somní.Íerum cell cultures by fungal elicitors. J P/aní Phys/.o/. 119: 65-76.
Facchini PJ, Bird DA, (1998). Developmental regulation of benzylisoquinoline alkaloid
biosynthesis in opium poppy plants and tissue cultures. /n VÍ.Íro Ce// Oev BÍ.o/.-P/aní.
34: 69-79.
Facchini PJ, De Luca V, (1994). Differential and tissue-specific expression of a gene
family for tyrosine/dopa decarboxylase in opium poppy. J 8/.o/ Chem. 269: 26684-
26690.
Facchini PJ, De Luca V, (1995). Phloem-specific expression of tyrosine/dopa
decarboxylase genes and the biosynthesis of isoquinoline alkaloids in opium poppy.
P/aní Ce//. 7: 1811-1821.
Facchini PJ, Dicosmo F, (1991). Plant cell bioreactor for the production of
protoberberine alkaloids from inmobilzed rha/Í.cfrum rt/gosum cultures. BÍ.ofechno/BÍ.oeng. 37: 397403.
Facchini PJ, Johnson AG, Poupart J, De Luca V, (1996). Uncoupled defense geneexpression and antimicrobíal alkaloid accumulation in elicited opium poppy cell
cultures. P/anf Phys/.o/. 111 : 687-697.
Facchini PJ, Penzes C, Johnson AG, Bull D, (1996). Molecular characterization of
berberine bridge enzyme genes from opium poppy. P/aní Phys/.o/.112: 1669-1677.
Facchini PJ, Bird DA, (1998). Developmental regulation of benzylisoquinoline alkaloid
biosynthesis in opium poppy plants and tissue cultures. /n Vítro Ce// Dev BÍ.o/.-P/anf.
34: 69-79.
Facchini PJ, Sang-Un P, Bird DA, Samanani N, (2000). Toward the metabolic
engineering of benzylisoquinoline alkaloid biosynthesis in opium poppy and related
spec.ies. Recent Res Dev Phytochem. 4.. 3147 .
67

Frenzel T, Zenk MH, (1990). Purification and characterízation of three isoforms of S-
adenosyl-L-methionine: (R, S)-tetrahydrobenzylísoquinoline-W-methyltransferase from
Berber/.s koeí/.neana cell cultures. Phyíochem/.sÍv. 29: 3491 -3497.
Frenzel T, Zenk MH, (1990). S-Adenosyl-L-methionine: 3'-hydroxy-W-methyl-(S)-
coclaurine-4'-O-methyltransferase, a region- and stereoselective enzyme of the (S)-reticuline pathway. Phyfochem/.sÍoÍ. 29: 3505-3511.
Frjck S, Kutchan TM, (1999). Molecular cloning and functional expression of 0-
methyltransferases common to isoquinoline alkaloid and phenylpropanoid
biosynthesis. P/aní J.17: 329-339.
Fujiwara H, Takeshita N, Terano Y, Fitchen JH, Tsujita T, Katagiri Y, Sato F,Yamada Y, (1993). Expression of (S)-scoulerine 9-O-methyltransferase in CopÍ/.s
/.apon/.ca plants. Phyfochem/.sÍv. 34: 949-954.
Fukui H, Nakagawa K, Tsuda T, Tabata M, (1982). Production of isoquinolinealkalo.ids by cell susper\s.ion cul\ures oi Coptis japonica. Proc 5th lntl Cong Plant TissCe// Cu/Í. En: P/aní Í/.ssue cu/Íure, (ed. Fujiwara A). Abe Photo Co. Ltd. Tokyo Japan.Pp: 313-314.
Funk C, Gügler K, Brodelius P, (1987). lncreased secondary product formation in
plant cell suspension cultures after treatment with a yeast carbohydrate preparation(elicitor). Phyfochemí.síry. 26: 401405.
Gamborg OL, Miller RA, Ojima K, (1968). Nutrient requirements of suspension
cultures of soybean root cells. Exp Ce// Res. 50..151-158.
Galdener E, Rueffer M, Wanner G, Tabata M, Zenk MH, (1988). Alternative finalsteps in berberine biosynthesis in Copí/.s/.apon/.ca cell cultures. P/aní Ce// Rep. 7: 14.
Gerardy R, Zenk MH, (1993a). Formation of salutaridine from (R)-reticuline by a
membrane-bound cytochrome P450 enzyme from Papaver somní.Íe/um.Phytochemistry. 32.. 79-86.
Gerardy R, Zenk MH, (1993b). Purification and characterization of
salutaridine: NADPH 7-oxídoreductase from Papaver somní.fe/um. Phytochemí.sÍoÍ. 34:125-132.
Gurney K, Evans LV, Robinson DS, (1992). Purine alkaloid production and
accumulation in cocoa callus and suspensíon cultures. J E*p Boíany. 43: 769-775
68

Haider G, von Schrader T, FüÍ)lein M, Blechert S, Kutchan TM, (2000). Structure-activity relationships of synthetic analogs of jasmonic acid and coronatine on induct.ion
of benzo[c]phenanthridine alkaloid accumulation in Eschscho/z/.a ca//.Íom/.ca cellcultures. B/.o/ Chem. 381 : 741-748.
Hara M, Kitamura T, Fukui H, Tabata M. (1993). lnduction of berberine biosynthesis
by cytokinins in Tha//.cfrum m/.nus cell suspension cultures. P/aní Ce// Rep.12: 70-73.
Hashimoto T, Yamada Y, (1994). Alkaloid biogenesis: Molecular aspects. Annt/ RevPlant Physiol Plant Mol Biol. 45.. 25]-285.
Hauschild K, Pauli HH, Kutchan TM. (1998). lsolation and analysis of a gene bbe7
encoding the berberine bridge enzyme from the California poppy Eschscho/zÍ.acalifornica. Plant Mol Biol. 36.. 473-4]8.
Heinstein PF, (1985). Future approaches to the formation of secondary natural
products in plant cell suspension cultures. J ~aí Proc/. 48: 1-9.
Hobbs MC, Yeoman MM, (1991). Effect of light on alkaloid accumulation in cell
cultures of WÍ.coÍ/.ana species. J Exp Boíany. 42: 1371-1378.
Huang FC, Kutchan TM, (2000). Distribution of morphinan and
benzo[c]phenanthridine alkaloid gene transcript accumulation in Papaver somn/.Íert/m.
Phytochemistry. 53.. 555-564.
Hsu AF, (1981). Effect of protein synthesis inhibitors on cell growth and alkaloid
production in cell cultures of Papaver somn/.fert/m. J Waí Prod. 44: 408414.
lgnatov A, Clark WG, Cline SD, Psenak M, Krueger RJ, Coscia CJ, (1996).Elicitation of dihydrobenzofenanthridine oxidase in Sangu/.nar/.a Canadens/.s cell
cultures. Phyfochemí.sfry. 43: 1141-1144.
lsrailov A, Yunusov MS, (1986). Alkaloids of four species of Argemone. Kh/.mí.yaPrirodnnykh Soedinenii. 2.. 204-206.
Jha S, Sahu NP, Sen J, Jha TB, Mahato SB, (1991). Production of emetine andcephaeline from cell suspension and excised root cultures of Cephae//.s /.pecacuanha.PAyíc>chemi.sfo/. 30: 3999-4003.
Khanna P, Prakash 0, Saluja M, (1982). Berberine from Argemone mex/.cana Linn.
tissue cultures. Proc 5th lntl Cong Plant Tiss Cell Cult. Er\.. Plant tissue culture, (ed.Fujiwara A). Abe Photo Co Ltd. Tokyo Japan.Pp: 311-312.
69

Kammerer L, De-Eknambul W, Zenk MH, (1994). Enzymic 12-hydroxylatjon and 12-
O-methylation of dyhydrochelirubine in dihydromacarpine formation by rna/Í.oírt/mbulgaricum. Phytochemistry. 36..1409-1416.
Kamo K, Kimoto W, Hsu AF, Mahlberg PG, Bills DD, (1982). Morphinane alkaloids
in cultured tissues and redífferentiated organs of Papaver somní.Íerum. Phyíochem/.sfv.21 : 219-222.
Knobloch KH, (1982). Uptake of phosphate and its effect on phenylalanine ammonia-lyase and cinnamoyl putrescine accumulation in cell suspension cultures of W/.coÍ/.ana
tabacum. Plant Cell Rep.1..128-130.
Knobloch KH, Berlin J, (1983). lnfluence of phosphate on the formation of indole
alkaloids and phenolic compounds in cell suspension cultures of Caíharaníhus roseus.Plant Cell Tiss Org Cult 2.. 333-340.
Krane BD, Fagbule MO, Shamma M, (1984). The benzophenathridine alkaloids. J WaíPnod. 47: 143.
Kraus PFX, Kutchan TIvl, (1995). Molecular cloning and heterologous expression of
cDNA encoding bermamuníne synthase, a C-O phenol-coupling cytochrome P150from the higher plant Berben.s sÍo/on/.Íera. Proc Waí/ Acad Sc/. USA. 92: 2071-2075.
Kutchan TM, Ayabe S, Krueger RJ, Coscia EM, Coscia CJ, (1983).Cytodifferentiation and alkaloid accumulation i.n cultured ceHs of Papaver bracfeaíum.
Plant Cell Rep. 2.. 281-284.
lkezawa N, Tanaka M, Nagayoshi M, Shinkyo R, Sakaki T, lnouye K, Sato F,
(2003). Molecular cloning and characterization of CYP719, a methylenedioxy bridge-forming enzyme that belongs to a novel P450 family, from cultured Copf/.s /.apon/.cacells. J BÍ.o/ Chem. 278.. 38557-38565.
Lee YL, Sagare AP, Lee CY, Feng HT, Ko YC, Shaw JF, Tsay HS, (2001). Formationof protoberberine-type alkaloids by the tubers of somatic embryo-derived plants of
Corydalis yanhusuo. Planta Med. 6].. 839-842,
Lenz R, Zenk MH, (1995a). Acetyl coenzyme A:salutaridinol-7-O-acetyltransferase
from Papaver somní.Íert/m plant cell cultures. J Bt.o/ Chem. 270: 31091-31096.
Lenz R, Zenk lvIH, (1995b). Purification and properties of codeinone reductase
(NADPH) from Papaver somn/.Íertjm cell cultures and differentiated plants. E.ur JBÍ.ochem. 233: 132-139.
70

Logemann J, Schell J, Willmitzer L, (1987). lmproved method for the isolation ofRNA from plant t.issues. Ana/ BÍ.ochem.163: 16-20.
Lozoya, X, Lozoya M, (1982). Flora medicinal de México. Plantas indígenas. Primera
pane. lnstituto Mexicano del Seguro Social. Pp.148-173.
Mahady GB, Schilling AB, Beecher CWW, (1993). Sangu/.nar/.a canac/ens/.s L.
(Sanguinarius): /n v/.íno culture at the production of benzophenanthridine alkaloids. En:Medicinal and Aromatic Plants. Biotechnology in Agriculture and Forestry, (ed. Bdia.iYPS). Springer-Verlag Berlin Heidelberg. 24: 313-328.
Mahady GB, Beecher CWW, (1991). The role of calcium in the induction of secondarymetabolism. En: /ní/ Res Cong Waf Prod. Chicago, lllinois. P. 20.
Mahady GB, Beecher CWW, (1994). Quercetin-induced benzophenanthridine alkaloid
production in suspension cell cultures of Sanguí.nar/.a canadens/.s. P/anía Mec/. 60: 553-557.
Mahady GB, Liu C, Beecher CWW, (1998). lnvolvement of protein kinase and G
proteins in the signal transduction of benzophenanthridine alkaloid biosynthesis.Phyíochemí.síry. 48: 93-102.
Maldonado-Mendoza lE, López-Meyer M, Galef JR, Burnett RJ, Nes§ler CL,
(1996). Molecular analysis of a new member of the opiumm poppy tyrosine/3,4-dihydroxyphenylalanine decarboxylase gene family. P/aní Physí.o/. 110: 4349.
Memelink J, Verpoorte R, Kijne JW, (2001). ORCAnizat.ion of jasmonate-responsive
gene expression in alkaloid metabolism. rrends P/aní ScÍ.. 6: 212-219.
Menke FLH, Parchmann S, Mueller MJ, Kijne JW, Memelink J, (1999). lnvolvementof the octadecanoid pathway and protein phosphorylation in fungal elicitor-induced
expression of terpenoid indole alkaloid biosynthetic genes in Caíharaníhus roseus.P/anf Physí.o/.119: 1289-1296.
Monforte-González M, Ayora-Talavera T, Maldonado-lvlendoza IE, Loyola-VargasVM, (1992). Quantitative analysis of serpentine and ajmalicine in plant tissues of
Cafharaníhus roseus and hyoscyamine and scopolamine in root tissues of Oaíurasframon/.um by thin layer chromatography-densitometry. Phyfochem Ana/ys/.s. 3: 117-121.
Morishige T, Tsujita T, Yamada Y, Sato F, (2000) Molecular characterization of theS-adenosyl-L-methionine: 3'-hydroxy-W-methylcoclaurine4'-O-methyltransferase
71

involved in isoquinoline alkaloid biosynthesis in Copí/.s /.apon/.ca. J BÍ.o/ Chem. 275:
23398-23405.
Negrel J, Javelle F, Paynot M, (1993). Biochemical basis of resistance of tobaccocallus tissue cultures to hydroxyphenylethylamines. P/aní Phys/.o/.103.. 329-334.
Nothnagel EA, MCNeil M, Albersheim P, Dell A, (1983). A galacturonic acid
oligosaccharide from plant cell walls elicits phytoalexins. P/aní Phys/.o/. 71 : 916-926.
Ouwerkerk PBF, Hallard D, Verpoorte R, Memelink J, (1999). Identification of UV-Blight-responsive regions in the promoter of the tryptophan decarboxylase gene from
Catharanthus roseus. Plant Mol Biol. 41.` 491-503.
0'Keefe BR, Beecher CWW, (1994). lsolation and characterization of S-adenosyl-L-methionine:tetrahydroberberine-cisw-methyltransferase from suspension cultures of
Sanguinaria canadensis L. Plant Physiol.105. 395-403.
Pauli HH, Kutchan TM, (1998). Molecular cloning and functional heterologous
expression of two alleles encoding (S)W-methylcoclaurine 3'-hydroxylase (CYP80B1),
a new methyl jasmonate-Índucible cytochrome P450-dependent mono-oxygenase ofbenzylisoqujnoline alkaloid biosynthesis. P/aní J. 13: 793-801.
Phillips GC, Collins GB. (1979). /n v/.íro tissue culture of selected legumes and plant
regeneration from callus cultures of red clover. Crop ScÍ..19:59-64.
Qi SY, He ML, Lin LD, Zhang CH, Hu LJ, Zhang HZ, (2005). Production of 2-(2-
phenylethyl) chromones i.n cell suspensi.on cultures of Aqu/./ar/.a s/.nens/.s. P/aní Ce//T/.ss Org Cu/í . 83: 217-221.
Roos W, Dordschbal 8, Steighardt J, Hieke M, Weiss D, Saalbach G, (1999). Aredox-dependent, G-protein-coupled phospholipase A of the plasma membrane js
involved in the elicitation of alkaloid biosynthesis in Eschscho/Íz/.a ca//.fom/.ca. B/.och/.m
B/.ophys Acía.1448: 390402.
Roos W, Evers S, Hieke M, Tschópe M, Schumann 8, (1998). Shifts of intracellular
pH distribution as a part of the signal mechanism leading to the elícitation ofbenzophenanthridine alkaloids. P/aní Phys/.o/. 118: 349-364.
Rueffer M, El-Shagi H, Nagakura N, Zenk MH, (1981). (S)-Norlaudanosolinesynthase: the first enzyme in the benzylisoquinoline biosynthetic pathaway. FEBS Le#.
129.' 5-9.
72

Rueffer M, Zenk MH, (1987). Enzymatic formation of protopines by a microsomal
cytochrome P450 system of Covda/Í.s vagí.nans. Teírahedron Leíí. 28: 5307-5310.
Rueffer M, Zenk MH, (1994). Canadine synthase from Tha//.círt/m ÍL/berosum cellcultures catalyses the formation of the methylenedioxy bridge in berberine synthesis.Phyfochemí.síry. 36: 1219-1223.
Rueffer M, Zumstein G, Zenk MH, (1990). Partial purification and properties of S-
ad en osy l-L-meth i o n i ne-( S) -tetra h yd ro proto be rberi n e-ci s-W-methyltra nsferase from
suspension-cultured cells of Eschscho/Íz/a and Coryda/i.s. Phyfochemí.sÍo/ 29: 3727-3733.
Samanani N, Facchini PJ, (2001). lsolation and partial characterization of
norcoclaurine synthase, the first commifted step in benzylisoquinoline alkaloid
bisynthesis, from opium poppy. P/anía. 213. 898-906.
Samanani N, Yeung EC, Facchini PJ, (2002). Cell type-specific protoberberinealkaloid accumulation in 77ia/Í.cfmm Í/avum. J P/aní Physí.o/. 159: 1189-1196.
Sato F, Endo T, Hashimoto T, Yamada Y, (1982). Production of berberine in cultured
Coptis japonica cells. Proc 5th lntl Cong Plant Tiss Cell Cult. Er\.. Plant tissue culture`
(ed. Fujiwara A). Abe Photo Co. Ltd. Tokyo Japan. 319-320.
Sato F, Takeshita N, Fitchen JH, Fujiwara H, Yamada Y, (1993). S-Adenosyl-L-methionine:scoulerine-9-O-methyltransferase from cultured Copf/.s /.apon/ca cells.Phytochemistry. 32.. 659-864.
Sato F, Tsujita T, Katagiri Y, Yo§hida S, Yamada Y, (1994). Purification andcharacterization of S-adenosyl-L-methionine:norcoclaurine 6-O-methyltransferase from
cultured Copí/.s/.apon/.ca cells. Eur L/. B/.ochem. 225: 125-131.
Schemeller T, Wink M, (1998) Utilization of alkaloids in modern medicine. En:
Alkaloids. Biochemistry, ecology and medicinal applications, (eds` F`oberis MF, W.inkM). Plenum Press. New York. P. 452.
Schumacher HM, Gundlach H, Fiedler F, Zenk MH, (1987). Elicitation of
benzophenanthridine alkaloid synthesis in Eschscho/Íz/.a cell cultures. P/aní Ce// Rep.
6: 410-413.
Schumacher HM, Zenk MH, (1988). Partial purification and characterization of
dihydrobenzophenanthridine oxidase from Eschscho/ízÍ.a ca//rorní.ca cell suspensioncultures. Plant Cell Rep. 7.` 4346.
73

Schwarzbach AE, Kadereit JW, (1999). Phylogeny of prickly poppies, Argemone
(Papaveraceae), and the evolution of morphological and alkaloid characters based onlTS nrDNA sequence variation. P/aní Sysí Evo/. 218: 257-279.
Siah CL, Doran PIvl, (1991). Enhanced codeine and morphine production in
suspended Papaver somní.rerum cultures after removal of exogenous hormones. P/aníCe// Rep.10: 349-353.
Smolko DD, Peretti SW, (1994). Stimulation of berberine secretion and growth in cell
cultures cif Thalictrum minus. Plant Cell Rep.14..131-136.
Staba EJ, Zito S, Amin M, (1982). Alkaloid production from Papaver tissue cultures. J
Nat Prod. 45.. 256-262.
Stadler R, Kutchan TM, Zenk MH, 1989). (S)-Norcoclaurine is the central
intermediate in benzylisoquinoline alkaloid biosynthesis. Phyfochem/.síry. 28: 1083-
1086.
Sibson DR, Starkey MP, (1997). lncreasing the average abundance of low-abundance cDNA's by ordered subdivision of cDNA population. En: cOWA //.bffiv
proíoco/s. Meíhods /.n mo/ecu/ar b/.o/ogy. Vol. 69, (eds. Cowell lG, Austin CA). HumanaPress lnc. Totowa New Jersey. Pp.13-32.
Steffens P, Nagakura N, Zenk MH, (1985). Purification and characterízation of theberberine bridge enzyme from Benben.s bean/.ana cell cultures. Phyíochem/.sÍv. 24:2577-2583.
Takeshita N, Fujiwara H, lvlimura H, Fitchen JH, Yamada Y, Sato F, (1995).Molecular cloning and characterization of S-adenosyl-L-methionine:scoulerine-9-O-
methyltransferase from cultured cells of CopÍÍ.s/.aponí.ca. P/aní Ce// Pnysí.o/. 36: 29-36.
Tanahashi T, Zenk MH, (1990). EIÍcitor induction and characterízation of microsomal
protopine-6-hydroxylase, the central enzyme in benzophenanthridine alkaloidbiosynthesis. Phyíochemí.síry. 29: 1113-1122.
Tomé F, Colombo M, (1995). Distribution of alkaloids in Che/Í.don/.um ma/.us and
factors affecting their accumulation. Phyfochem/.sÍo/. 40: 37-39.
Unterlinner 8, Lenz R, Kutchan TM, (1999). Molecular cloning and functionalexpressíon of codeinone reductase: the penultimate enzyme in morphíne biosynthesisin the opíum Papaver somní.Íert/m. P/aní J. 18: 465475.
74

van der Fits L, Memelink J, (2000). ORCA3, a jasmonate-responsive transcríptionalregulator of plant primary and secondary metabolism. Sc/.ence. 289: 295-297.
Vázquez-Flota F, De Luca V, (1998a). Jasmonate modulates development-and light-regulated alkaloid biosynthesis in Caíharaníhus roseus. Phyfochem/.síry. 49: 395-402.
Vázquez-Flota F, De Luca V, (1998b). Developmental and light regulation of
desacetoxyvindoline 4-hydroxylase in CaíharaMt/s noseus (L.) G. Don. P/aní Phys/.o/.117: 1351-1361.
Weiler EW, Albrecht T, Groth 8, Xia ZQ, Luxem M, Lip H, Andert L, Spengler P,
(1993). Evidence for the involvenment of jasmonates and their octadecanoid
precursors in the tendril coiling response of Bvon/.a d/.o/.ca. Phyfochem/.síoÍ. 32: 591-600.
Wagner H, Bladt S, Zgainski EM, (1984). P/aní Drug Ana/ys/.s. A Íh/.n /ayerchmmaíography aí/as. Springer-Verlag Berlin Heidelberg. Germany. Pp. 76-82.
Yoder LR, Mahlberg PG, (1976). Reaction of alkaloid and histochemical indicators in
laticifers and specialized parenchyma cells of Caíharaníhus roseus (Apocynaceae).Amer J Bof. 63: 1167-1173.
Zhao J, Zhu WH, Qiu H, (2001). Effects of light and plant growth regulators on the
biosynthesis of vindoline and other indole alkalo.ids in Cafharaní^us roseus callus
cultures. P/aní Growíh Regu/. 33.. 4349.
75


APENDICE A
No. Color Rf C Días de cu tivo0 5 10 15 20 25 30 35 40 45 50
1 Naranja 0.90 L + + + +++
0 + + + + + + + + +++
2 Rojo 0.78 L +
03 Azulclaro 0.66 L ++ + + + + + + + + + +
0 + + + +
4 Rojo'adri"O 0.62 L + + +
05 Azulvioleta 0.60 L + + + + + + ++
0 + + + +
6 Verdeazul 0.49 L + + + + + + + +
07 Azul 0.41 L + + ++ ++ +++ + ++ ++ +
0 +
8 Naranja 0.36 L +
0 + ++ + ++ + + ++ ++
9 Amarillo 0.21 L
0 + +
10 Azulclaro 0.19 L + + + + + ++ ++ ++ ++ + +
0 + + + ++ +
Comparación del patrón de manchas fluorescentes observado en cultivos de callosmantenidos bajo luz continua y en oscuridad total. Un volumen equivalente a 0.5 mg
de PS se cargó en placas de gel de sílice y las muestras se separaron utilizando lamezcla de disolventes J (ver cuadro 111.1). Las placas fueron visualizadas bajo luz
UV a 365 nm. No.: Número asignado, Color: Fluorescencia bajo luz UV a 365 nm, Rf
Valor de migración calculado, C: Condición, L: Luz continua, 0: Oscuridad total.
77

No. Color R, E OLLM 10 tlM 100 uM 1000 uMT T MeJa T lvleJa T MeJa
1 Naranja 0.76 C + + + + + + +
M + + + + + +
2 Morado 0.69 CM + + + ++ ++ ++
3 Rojonaranja 0.69 C + + + + +
M ++ ++ ++ + + +
4 Azulclaro 0.65 C + + + ++ ++ ++ ++
M5 Naranja 0.64 C
M +
6 Azul 0.64 C + + ++ + + +
M + + + + + +
7 Amarillo 0.62 CM +
8 Amarillo 0.53 C + + + +M
9 Azul 0.52 C ++ ++ + +++ ++ + +
M ++ ++ ++ ++ ++ ++
10 Amarillo 0.50 CM + + + + + +
11 Morado 0.45 CM + + + + + +
12 Amarillo 0.35 C + + + + + + +
M13 Azu'claro 0.29 C
M + + + ++ ++ +
14 Amarillo 0.22 C +M + + + + ++ +
Patrón de compuestos fluorescentes no Ídentificados, observado en cultivos de célulasen suspensión en respuesta a diferentes concentraciones de MeJa. Un volumenequivalente a 0.5 mg de PS se cargó en placas de gel de sílice y las muestras sesepararon utilizando la mezcla de disolventes J (ver cuadro 111.1). Las placas fueron
visualizadas bajo luz UV a 365 nm. No.: Número asignado, Color: Fluorescencia bajo
luz UV a 365 nm, Rf Valor de migración calculado, E: Extracto, T: Testigo, C: Células,
M: Medio de cultivo.
78

No. Color R, E Oh 6h 12h 24hT T MeJa T MeJa T MeJa
1 Naranja 0.76 CM + + + + + + +
2 Morado 0.69 CM + + + + + + +
3 Naranja 0.64 CM + + + ++ ++ + +
4 Azul 0.64 CM + + + + + + +
5 Azul 0.52 C + + + + + + +
M ++ ++ ++ ++ ++ +++
6 Amarillo 0.50 C + + + + + + +
M + + + + + + +
7 Moradoclaro 0.46 CM + + + + + + +
8 Moradoclaro 0.45 CM + + + + + + +
9 Azul 0.29 C + + + + + + +
M + + + + + + +
10 Amarillo 0.22 CM + + + + + + +
Patrón de compuestos fluorescentes no identificados, observado en cultivos de célulasen suspensión en respuesta a diferentes tíempos de incubación con 100 HM deMeJa. Un volumen equivalente a 0.5 mg de PS se cargó en placas de gel de sílice y
las muestras se separaron utilizando la mezcla de dísolventes J (ver cuadro 111.1).
Las placas fueron visualizadas bajo luz UV a 365 nm. No.: Número asignado, Color:
Fluorescencia bajo luz UV a 365 nm, Rf. Valor de migración calculado, E: Extracto, T:
Testigo, C: Células, M: Medio de cultivo.
79





















