
UNIVERSIDAD AUTONOMA METROPOLITANA …148.206.53.84/tesiuami/UAM7024.pdf · obtendrán las estirpes...
148
UNIVERSIDAD AUTONOMA METROPOLITANA UNIDAD IZTAPALAPA DIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD ,/' ,, / CARACTERIZACION DE FRACCIONES ENRIQUECIDAS EN CELULAS DE LEYDIG Y SERTOLI DE RATAS ADULTAS NORMALES TESIS QUE I?RESENTA EL BIOL. ENRIQUE M:ENDIETA MARQUEZ PARA OBTENER. EL GRADO DE 1 " " . MAESTRO EN BIOLClGIA EXPERIMENTAL DICIEMBRE 1992
Transcript of UNIVERSIDAD AUTONOMA METROPOLITANA …148.206.53.84/tesiuami/UAM7024.pdf · obtendrán las estirpes...

UNIVERSIDAD AUTONOMA METROPOLITANA
UNIDAD IZTAPALAPA
DIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD
,/' ,, /
CARACTERIZACION DE FRACCIONES ENRIQUECIDAS EN CELULAS
DE LEYDIG Y SERTOLI DE RATAS ADULTAS NORMALES
TESIS QUE I?RESENTA EL
BIOL. ENRIQUE M:ENDIETA MARQUEZ
PARA OBTENER. EL GRADO DE 1 "" .
MAESTRO EN BIOLClGIA EXPERIMENTAL
DICIEMBRE 1992

1 2 6 3 8 7 L ;-.Este trabajo experimental fue realizado en el Laboratorio de
L"iAndrología, Subjefatura de Investigación del Instituto Mexicano del
Seguro Social y en el Laboratorio de Mecanismos de Regulación i
Testicular, Departamento de Ciencias de la Salud, Universidad
Autónoma Metropolitana - Iztapalapa.
Este trabajo fue financiado parcialmente por PRONAES a través del
convenio C88-03-0049, y por DGICSA mediante el convenio C90-01-
0283.
La Maestría en Biología Experimental se encuentra registrada dentro
del Padrón de Excelencia de Programas de Posgrado del CONACYT, y es
financiada por éste a través del Convenio PFPN/66/91.

I
Dedico este trabajo a todos aquéllos que de una u otra manera me impulsaron a culminar la labor que se ve plasmada en esta tesis.
A mi familia, con todo el cariño que no es fácil expresar en palabras, pero que siempre está presente. Ellos han sitio la razón de mi esfuerzo y la fuente de mi inspiración.
A mis asesores, con gratitud, ya que sus valiosos comentarios y críticas me ayudaron a conjuntar mis ideas y a exponerlas de una forma más clara y precisa.
A mis compañeros, por su insistencia constante en lograr que terminara por fin con esta parte de mi formación.
y por último, aunque no al final, a todos mis amigos, quienes han demostrado que confían en mí y que me aceptan como soy.
GRACIAS.

INDICE
1. INTRODUCCION 1
1.1. ANATOMIA DEL APARATO REPRODUCTOR MASCULINO
1.2. ANATOMIA TESTICULAR
1.2.1. Túbulos Seminiferos
1.2.2. Espermatogknesis
1.2.3. Células de Sertoli
1.2.4. Compartimiento intersticial
1.3. FISIOLOGIA TESTICULAR
1.3.1. Eje hipotálamo-hipófisis- testículo
1.3.2. Regulación funcional de las células de Leydig
1.3.2.1. Vías esteroidogénicas
1.3.3. Regulación funcional de las células de Sertoli
1.3.4. Mecanismos intratesticulares de regulación
1.3.4.1. Interacciones células de Sertoli-células de la línea germinal
1.3.4.2. Interacciones células de Sertoli-células peritubulares
1.3.4.3. Interacciones células de Sertoli-células de Leydig
1.4. MODELOS DE ESTUDIO: OBTENCION DE FRACCIONES CELULARES ENRIQUECIDAS
2
4
5
7
9
13
15
15
17
19
23
25
25
28
29
31
". .

2. PLANTEAMIENTO DEL PROBLEMA 34
3. HIPOTESIS DE TRABAJO
4. OBJETIVOS
5. MATERIAL Y METODOS
5.1. OBTENCION DE FRACCIONES ENRIQUECIDAS
5.2. ENSAYO DE PROTEINAS UNIDORAS
5.3. EVALUACION DE LA :2STEROIDOGENESIS
5.4. SEPARACION Y CUANTIFICACION DE LOS ESTEROIDES
5.5. DETERMINACION CUANTITATIVA DE LOS ESTEROIDES
6. RESULTADOS
6.1. OBTENCION DE FRACCIONES ENRIQUECIDAS
6.2. CARACTERIZACION DE PROTEINAS UNIDORAS
6.3. ESTEROIDOGENESIS EN FRACCIONES ENRIQUECIDAS EN CIELULAS DE LEYDIG
6.4. ESTEROIDOGENESIS EN FRACCIONES ENRIQUECIDAS EN CELULAS DE SERTOLI
7. DISCUSION
7.1. ANALISIS DE LOS RESULTADOS DE LA OBTENCION DE FRACCIONES ENRIQUECIDAS EN COMPARACION CON DIVERSAS METODOLOGIAS PREVIAMENTE REPORTADAS
7.2. PRESENCIA DE PROTEINAS UNIDORAS DE ESTEROIDES EN LAS FRACCIONES ENRIQUECIDAS EN CELULAS DE LEYDIG Y CELULAS DE SERTOLI
7.2.1. Receptores de andrógenos
7.2.2. Receptores de estrógenos
36
36
37
37
39
41
43
45
48
48
49
51
53
55
55
61
61
64

7.2.3. Receptores de progestágenos
7.3. PATRONES ESTEROIDOGENICOS EN LAS FRACCIONES ENRIQUECIDAS EN CELULAS DE LEYDIG Y CELULAS DE SERTOLI
7.3.1. Fracción intersticial
7.3.2. Fracción tubular
8. CONCLUSIONES
9. BIBLIOGRAFIA
67
68
68
74
7 8
8 0

1
1. INTRODUCCION.
Durante mucho tiempo fue posible obtener información acerca del
funcionamiento del aparato reproductor masculino a través de la
utilización de metodologías que medían sus productos secretados
(espermatozoides y andrógenos) en medios externos a los órganos que
los componen. Conforme la naturaleza de los mecanismos
extragonadales de regulación fue mejor conocida, la utilización de
modelos diferentes de estudio se convirtió en una necesidad.
El desarrollo de técnicas de separación celular y de métodos más
sensibles de medición de la concentración de diferentes hormonas,
que permitieran la cuantificación in situ o en cantidades muy
pequeñas de tejido ha tenido un gran impacto en los niveles de
comprensión de la función testicular en los últimos años.
Para lograr conocer la respuesta específica de una estirpe celular
definida a la presencia de algún factor regulador, un modelo
experimental muy útil sería aquél que permitiera evaluar esta
respuesta en forma aislada a otros factores que pudieran estar
presentes en el tejido.
Para obtener a una estirpe celular en un grado confiable de pureza
y en un estado funcional adecuado, es necesario recurrir a
procedimientos mecánicos y/o enzimáticos, seguidos en ocasiones de
la utilización de diversas técnicas de centrifugación y de la

2
utilización de gradientes de densidad.
La edad y el estado funcional d.el animal a partir del cual se
obtendrán las estirpes celulares son factores que afectan
profundamente tanto el rendimiento y la pureza de las fracciones
separadas como el valor predictivo de los planteamientos
experimentales que las utilizan.
En vista de lo mencionado anteriormente, el interés de este trabajo
se centra en la validación de un esquema de obtención de fracciones
enriquecidas en células de Leydig y Sertoli de rata adulta normal,
que les permita mantener su integridad funcional durante
incubaciones a tiempos cortos así como servir de modelo para el
estudio in vitro de la fisiología testicular.
1.1. ANATOMIA DEL APARATO REPRODUCTOR MASCULINO.
El aparato reproductor masculino puede ser dividido en 4
principales componentes funcionales (1):
a. Los testículos o gónadas masc:ulinas, que son órganos pareados
ubicados en el interior del saco escrotal, y que son responsables
de la producción de los gametos masculinos (espermatozoides), así
como de la secreción de las hormonas sexuales,
.. .

3
b. Un sistema pareado de ductos, consistente en ductos eferentes,
epidídimos, ductos deferentes y ductos eyaculadores, l o s cuales
colectan, almacenan y conducen a los espermatozoides hacia el
exterior de cada testículo. Los ductos eyaculadores convergen en la
uretra, a partir de la cual los espermatozoides son expulsados
hacia el exterior del organismo,
c. Un par de glándulas exocri.nas (vesículas seminales) , una
glándula prostática y un par de glándulas bulbouretrales accesorias
(glándulas de Cowper), las cuales secretan un medio nutritivo y
lubricante llamado fluido seminal., que permite la supervivencia de
los espermatozoides durante su traslado al aparato reproductor
femenino, y
d. Un órgano de copulación (pene) .
Durante el desarrollo embrionario, cada testículo, junto con la
primera parte de su sistema de ductos, vasos sanguíneos, linfáticos
y nervios, desciende desde la pared posterior de la cavidad
peritoneal hacia el interior del escroto. Durante la migración, el
testículo acarrea con él una capa periférica de peritoneo, de forma
tal que en el interior del escroto, el testículo se encuentra casi
totalmente rodeado de una cavidad serosa que es una extensión de la
cavidad peritoneal y que se encuentra limitada por la llamada
túnica vaginalis.

4
1.2. ANATOMIA TESTICULAR.
El testículo es una glándula tubular compuesta, rodeada por ~. una
gruesa capa fibrosa llamada túnica albuginea. En la cara posterior
del órgano, un engrosamiento de tejido conjuntivo penetra en la
glándula y forma el mediastino. Unos delgados tabiques fibrosos
(septula testis) se extienden radialmente a partir del mediastino
hacia la túnica albuginea, y dividen al órgano en unos 200
compartimientos piramidales llamados lobulillos ( 2 ) . Cada lobulillo
está constituido por uno o más túbulos seminiferos, de una longitud
total de 30 a 60 cm. Estos túbu:tos contorneados convergen en el
mediastino, donde siguen un trayecto rectilíneo (túbulos rectos) y
se anastomosan para formar la rete testis, la cual a su vez
desemboca en los conductos eferentes, y eventualmente en la cabeza
del epidídimo.
Los túbulos seminiferos constituyen la porción exocrina del
testículo, que se ve ref lejada en su función gametogénica o de
formación de gametos maduros a partir de las células componentes de
su epitelio germinal, que es auxiliada por las células de sostén o
células de Sertoli (CS) que se encuentran formando la periferia de
los túbulos.
El tejido conectivo intertubular contiene grupos de células
poligonales de gran tamaño (célulaas de Leydig) (CL) que constituyen
el componente endocrino del testículo, y que son por lo tanto

5
responsables de la aparición y mantenimiento de las características
sexuales secundarias normales así como del comportamiento
típicamente masculino ( 3 ) .
Los testículos se encuentran irrigados por una arteria que penetra
por el hilio, continúa hacia el mediastino y se ramifica por la
cara interna de la túnica albuginea, mientras que otras ramas se
subdividen en arteriolas al interior de la glándula y se ramifican
en un capilar que rodea a l o s túbulos que irrigan; un sistema
colector venoso recoge la sangre y sus troncos son paralelos a las
arterias.
La innervación de las gónadas masculinas proviene de la cadena
simpática lumbar a través de fibras que innervan a los vasos
sanguíneos. Existen asimismo, terminaciones sensoriales por debajo
de la túnica albuginea y nervios vasomotores que penetran al
testículo (4).
1.2.1. Túbulos seminiferos.
Los túbulos seminíferos 'están constituidos por un complejo epitelio
estratificado constituido por dos; categorías celulares principales:
las CS y las células de la linea germinal. La apariencia que
presentan los túbulos en las di.ferentes etapas de la vida de un
animal depende fundamentalmente de los diversos grados de
diferenciación que presentan las células de la línea germinal, los
""

6
cuales se engloban dentro del proceso llamado espermatogénesis, el
cual es el responsable de la formación de los gametos maduros y
funcionales.
Cada túbulo se halla recubierb por una capa basal de tejido
conectivo laminar que contiene fibras elásticas y células
epitelioides aplanadas, cuya organización varía de acuerdo a la
especie de que se trate (2). En 1.0s roedores hay una capa única de
células aplanadas y poligonales que se unen borde con borde para
formar una lámina epitelioide continua que rodea al túbulo, y que
presentan una apariencia semejant.e a las de las células musculares,
por lo que se les ha denominado células mioides o peritubulares.
Se cree que las células peritubulares son las responsables de las
contracciones rítmicas y que, aunque poco fuertes, se observan en
los túbulos seminíferos de algunas especies de mamíferos (5).
En el caso del humano, el complejo peritubular se extiende desde la
membrana basal del túbulo hacia el tejido circundante, y consiste
de dos capas alternas de fibras de colágena y células mioides entre
las cuales pueden encontrarse capas delgadas de microfibrillas y
material semejante al de la membrana basal ( 6 ) . En casos de
infertilidad masculina se ha observado una conspicua
desorganización de la estructura. peritubular, detectable mediante
cortes histológicos seriados (5).

7
1.2.2. Espermatogénesis.
La espermatogénesis puede dividirse en 3 fases: a) una primera
durante la cual se producen espermatocitos como resultado de las
divisiones mitóticas de las espermatogonias; b) una segunda en la
cual la división meiótica de los espermatocitos genera cuatro
espermátides, y c) una última en la cual los espermatozoides se
forman como resultado de la metamorfosis de las espermátides a
través de un proceso denominado espermiogénesis ( 7 ) .
Existen diversas características del proceso espermatogénico que
son particulares de las diferentes especies, tales como las etapas
en que éste puede ser dividido y la duración de cada una de ellas.
Por otra parte, muchas Ivcélulas madrevv entran simultáneamente a una
línea de diferenciación sincrónica, por lo que múltiples
generaciones gaméticas evolucionan de forma paralela.
El epitelio seminífero se encuentra compuesto por 5-6 generaciones
de células germinales que no s e distribuyen al azar, sino que
forman asociaciones celulares de composición definida (8). Estos
agrupamientos representan etapas del ciclo del epitelio seminífero
definidas como series completas de asociaciones celulares sucesivas
que aparecen en un área cualquiera.
El número de etapas que comprende el ciclo varía, como ya se ha
mencionado, de acuerdo a la especie y a los criterios de

8
clasificación utilizados. La duración del ciclo ha sido determinada
en diversos mamíferos mediante la autorradiografía después de la
inyección de trazadores radiactivos (9) . Los valores obtenidos que oscilan entre 52-53 días (rata) y 64 días (hombre) son constantes
aún a nivel de la cepa particular en animales de laboratorio (7).
Las espermatogonias o células germinales se encuentran en la capa
basal del epitelio seminífero donde se dividen a través de mitosis
para dar lugar a otras espermatogonias (Tipo A) , y a
espermatogonias que proseguirán hacia meiosis (Tipo B). Las
espermatogonias tipo A se caracterizan por la presencia de un
núcleo grande, esférico o elíptico con una cromatina moderadamente
condensada, mientras que las de tipo B tienen un núcleo más pálido,
con nucleolos localizados en el (centro de la célula ( 3 ) .
Las espermatogonias tipo B al dividirse dan origen a los
espermatocitos primarios, fácilmente reconocibles por su gran
cantidad de citoplasma y núcleo conspicuo con cromatina condensada
en forma de hebras delgadas (10). La primera división meiótica se
lleva a cabo con relativa rapidez, generando a l o s espermatocitos
secundarios, y posteriormente a las espermátides, células que ya
poseen un complemento génico haploide.
En cada una de las espermátides ocurre un conjunto de
transformaciones que constituyen la espermiogénesis, consistentes
en la reducción y condensación de su núcleo, la pérdida de

9
prácticamente todo su citoplasma y la aparición del complejo
acrosomal y del flagelo, además de: una serie de cambios bioquimicos
que le permiten asumir la apariencia de un espermatozoide maduro.
Después de su diferenciación, los espermatozoides son liberados de
su contacto intimo con l o s elementos de sostén y se dirigen hacia
la luz de los túbulos seminíferos. De aquí en adelante adquieren
gradualmente su madurez funcional a través de una serie de procesos
de activación que preceden a la fertilización.
1.2.3. C é l u l a s de Sertoli.
Las CS se encuentran apoyadas en la membrana basal de l o s túbulos
seminiferos, y su citoplasma se extiende hacia la luz del túbulo,
ocupando todos los espacios libres entre las células de la línea
germinal (1). Debido a su intima asociación con estas células, se
ha postulado que funcionan como I1nodrizas1l, proporcionando soporte
estructural y metabólico para el desarrollo y diferenciación de las
células espermatogénicas (11).
El citoplasma de estas células contiene abundante retículo
endoplásmico tanto liso como rugoso; sus mitocondrias son largas y
delgadas, con crestas tubulares transversales. El aparato de Golgi
consiste en elementos múltiples dispersos, y sus estructuras
lisosomales parecen ayudar a la digestión de las células germinales
en degeneración y cuerpos residua.les liberados por las espermátides

10
durante la espermiación (12). Es posible encontrar también dentro
del citoplasma numerosas inclusiones, como p.ej., gotas lipídicas,
y en algunas especies como en el hombre, varios cristales
( c r i s t a l o i d e s de Charcot-Botcher) y diversos tipos de vesículas
(11) -
Las características nucleares incluyen una membrana nuclear
invaginada, un nucleoplasma relat.ivamente homogéneo, y un nucleolo
Único dividido en 3 partes, que aparece al microscopio electrónico
como una estructura reticular acompañada de 2 regiones
electrodensas pequeñas -compuestas de heterocromatina- llamadas
cuerpos heteropicnóticos, cuerpos juxtanucleares, esferas
perinucleares o cariosomas satélite (12).
Se ha reportado por diversos investigadores que la actividad
mitótica de las CS en roedores es muy pronunciada durante los
períodos fetal y postnatal temprano, pero cesa entre los 12 y 15
días después del nacimiento (13). No es claro si las células
maduras poseen la capacidad de reasumir su división bajo ciertas
condiciones; en algunos casos, no existen evidencias concluyentes
de síntesis continua de DNA o proliferación celular en cultivos de
células provenientes de ratas sexualmente maduras, aún en presencia
de mitógenos (14) .
La presencia de una barrera semipermeable entre la sangre
periférica y los testículos fue postulada con base en la

11
observación de que marcadores el.ectrodensos, colorantes y otras
moléculas pequeñas se veían excluí.das de los túbulos seminiferos en
los animales adultos (15).
En la mayoría de los vertebrados, esta barrera hematotesticular se
encuentra formada por los complejos de unión entre las membranas de
las CS (16). Sin embargo, en algun.os mamíferos, la barrera primaria
comprende una serie de uniones estrechas entre células mioides
adyacentes que rodean a los túbulos seminiferos, e impiden que
ciertas sustancias lleguen a l a s espermatogonias en el
compartimiento basal.
En algunas áreas de los túbulos seminíferos, las uniones entre las
CS se extienden hasta la lámina basal, con contactos más frecuentes
al nivel de espermatogonias. Las CS se separan de éstas por
espacios de 20 nm y entre sí por regiones de 9 nm de separación,
que pueden reducirse a 2 nm al nivel de las uniones estrechas, y en
otras uniones focales, que incluso permiten un fuerte acoplamiento
eléctrico por distancias variables a lo largo del epitelio (17).
Se considera en la actualidad, que la población de espermatogonias
se encuentra separada de las demás células germinales, formándose
un compartimiento basal, que contiene a las espermatogonias y
espermatocitos preleptoténicos, y un compartimiento adluminal, que
consiste de espermatocitos y espermátides ubicadas por arriba del
nivel de las uniones estrechas. Las sustancias que lograran
.". .

12
atravesar las células de la capa mioide tendrían acceso directo a
las células del compartimiento basal, pero para alcanzar la región
adluminal deberían a su vez atravesar las uniones entre CS. Como
resultado de esto, se producen ambientes diferentes en ambos
compartimientos de los túbulos seminíferos (12).
Como las células de la línea germinal deben realizar su proceso de
diferenciación en sentido transversal respecto de cada túbulo, y
éste las llevaría a pasar sucesivamente del compartimiento basal al
adluminal, es fácil suponer que este tránsito implicaría la
destrucción y reformación constante de las uniones estrechas entre
las CS. Observaciones en diferent.es modelos experimentales (18-20)
demuestran que las fases iniciales de la espermatogénesis preceden
siempre al establecimiento de la barrera hematotesticular.
El desarrollo de la barrera hematotesticular está muy
correlacionado con el desarrollo de la luz del túbulo, cuyo tamaño
aumenta rápidamente hasta los 2 5 días aproximadamente en el caso de
la rata (21) . La segunda fase de elevación del volumen tubular y por ende de la disminución de la permeabilidad de la barrera se
presenta hacia los 45 días.
Las CS son capaces de ingerir células germinales en degeneración,
cuerpos residuales o materia particulada inyectada. La actividad
fagocítica se hace más pronunciada después de dañado el epitelio
germinal y de una degeneración masiva de células germinales (12) .

13
El inicio y mantenimiento de la espermatogénesis en el interior de
l o s túbulos seminiferos es dependiente de la presencia de una serie
de hormonas, cuyos efectos directos se aprecian en las CS ( 2 2 ) , las
cuales modifican su metabolismo para producir l o s elementos que las
células de la línea germinal- requieren para completar su
diferenciación (23).
1.2.4. Compartimiento intersticial.
En muchas especies el tejido intersticial, localizado entre los
túbulos seminiferos, representa tan solo una pequeña porción del
volumen testicular, variando entre aproximadamente un 5-10% en la
rata hasta un 60% en el cerdo (24). De acuerdo a Fawcett y cols
(25), existen al menos 3 diferentes tipos de organización del
tejido intersticial en función d , e l volumen ocupado por las CL, la
masa de tejido conectivo intersticial y la localización y grado de
desarrollo de los vasos linfáticos intertubulares.
En el caso de la rata se encuentra un número reducido de CL (1-5%
del volumen total) , poco tejido lzonectivo y un endotelio linfático
continuo alrededor del tejido limitante y discontínuo u
ocasionalmente interrumpido alrededor de los grupos de CL, con
espacios llenos de fluido y sinusoides linfáticos. Las CL rodean a
l o s capilares y usualmente no están soportadas por mallas de fibras
de colágena ni por células del tejido conectivo, tales como
fibroblastos, macrófagos, células cebadas y células mesenquimatosas

14
indiferenciadas (4) .
En la mayoría de las especies, las C L se encuentran formando
racimos en el espacio intertubular triangular, y en forma de haces
entre los túbulos mu17 compactados. En el animal sexualnente maduro,
la membrana plasmática presenta muchas especializaciones,
incluyendo complejcs de unión,, proyecciones e indentaciones
superficiales lisas y recubiertas. Alrededor de las CL existe una
nembrana basal de grosor y disposición variables que parece
funcionar como soporte en combinación con fibras de colágena (24) .
Existen cuando mencs 3 tipos de uniones entre las C L : septadas de
20 nm, de 2 nm y hendidas , y desmosomas sedimentarios (26) . Las uniones hendidas son permeables, las septadas parcialmente
perneables y las estrechas impermeables a marcadores electrodensos.
Las proyecciones superficiales son microvellosas o bulbosas, y se
encuentran íntimamente relacionadas con las uniones.
La característica Iiltraestructur'ai más conspicua en estas células
es la presencia de cn retículo endoplásmico liso bien desarrollado,
típico de las células secretoras. Existe asimismo, una red de
filamentos abundantes que forinan haces , y que se encuentran
formados por diversos tipos de proteínas fibrilares tales como
actina, vimentina y desmina ( 2 7 ; 1 .
El citoplasma contiene numerosas inclusiones semejantes a gotitas

15
de lípidos, y en algunas ocasiones presenta cristales (cristales de
Reinke en el testículo humano) cuyas funciones son desconocidas
( 2 )
1.3. FISIOLOGIA TESTICULAR.
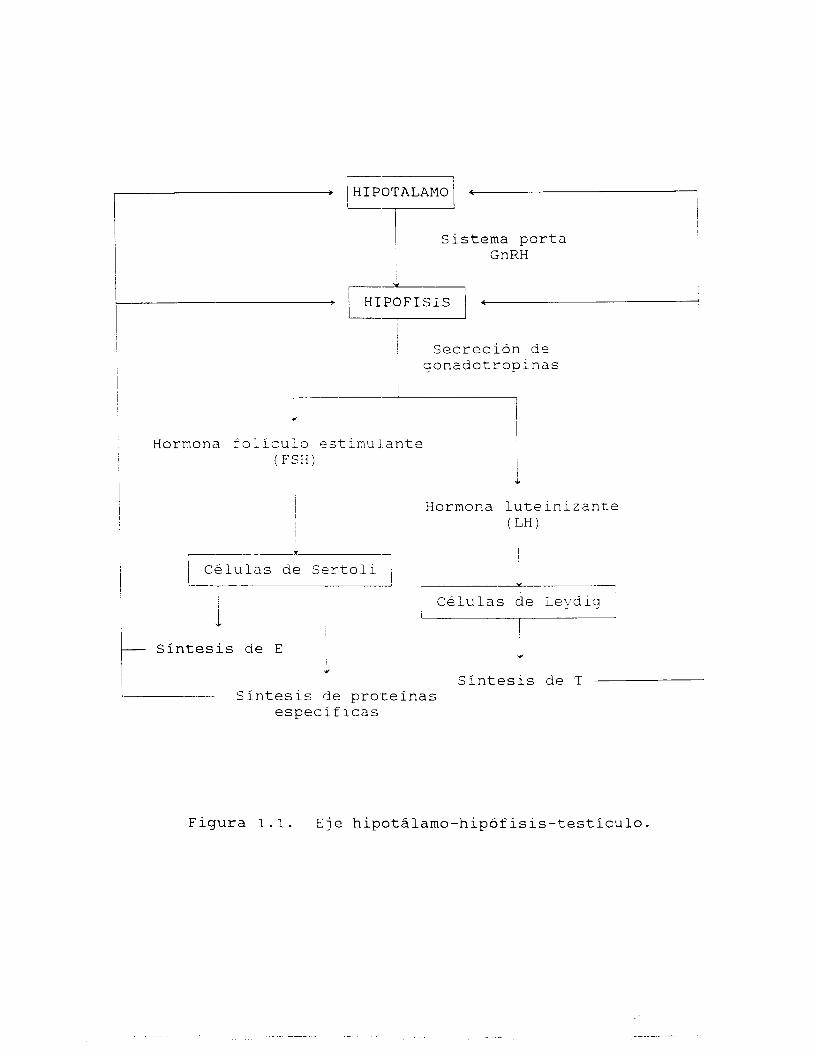
1.3.1. Eje hipotálamo-hipófisis-testículo (fig 1.1).
La actividad secretora de las CL se encuentra bajo el control de la
hormona luteinizante (LH), una d.e l o s gonadotropinas liberadas por
la adenohipófisis, mientras que la otra gonadotropina, la hormona
folículo estimulante (FSH), influye principalmente sobre la
actividad de las CS y por lo tanto, probablemente también sobre la
diferenciación de las células de la línea germinal ( 2 8 ) .

1 6
La liberación de las gonadotropinas por parte de l o s gonadotropos
hipofisiarios depende directamente de su estimulación por parte de
la hormona liberadora de gonadotropinas (GnRH), sintetizada en el
hipotálamo y enviada hacia la hipófisis a través de un sistema
porta especializado. Las cantidades de hormona liberadora presentes
en el sistema dependen de seAales reguladoras recibidas por
receptores específicos a nivel del. núcleo supraóptico hipotalámico,
a modo de un mecanismo de retroalimentación mediado parcialmente
por las hormonas esteroides.
Las gonadotropinas liberadas a la circulación periférica modulan la
actividad de sus células blanco a través de su interacción con
receptores membranales específicos, ligados al sistema de adenilato
ciclasa (29) . Así, las gonadotropinas unidas a sus receptores
incrementan la actividad enzimática, y por lo tanto, la
concentración intracelular del 3',5'-riboadenosilmonofosfato
cíclico (AMPc) , lo que a su vez desencadena cambios en patrones de
fosforilación proteica mediados a través de la acción de
proteincinasas de tipo A (30) , y modificaciones en l a s funciones
particulares de cada tipo celular.
Se ha demostrado que, a su vez, las hormonas esteroides tienen
diferentes papel en el control de la secreción de las
gonadotropinas (31). Así, la testosterona (T) suprime de igual
manera la liberación de LH y FSH, mientras que el estradiol (E)
preferentemente regula la secreción de FSH ( 3 2 ) .
Hormona
I HIPOYALAMO! 1
i !
Sistema porta GnRH
~ P ~ F I s I s 1 4
. I I i Secreción de
gonadotropinas
. folículo estimulante j
(FSH)
~ C G l u l a s de Sertoli i
i - Síntesis de E
*
i 1
Hormona luteinizante (LH)
1 w
I Células de Leydi.;
Síntesis de proteínas específicas
Síntesis de T
Figura 1.1. Eje hipotálamo-hipófisis-testículo.

17
Factores de naturaleza proteica secretados por las CS (inhibinas),
aparentemente sinergísticamente junto con el E se encargarían de
regular las concentraciones circulantes de FSH en animales adultos
sin afectar, más que en forma mínima, la concentración plasmática
de la LH (33).
La LH se une a l a s CL a través de receptores nembranales
específicos (34) que presentan elevadas constantes de afinidad y un
número limitado de sitios de unihn. Estas proteínas receptoras son
capaces de incrementar los niveles intracelulares de AMPc, activar
la función de la proteíncinasa A, e incrementar la tasa de
fosforilación de cuando menos 6 proteínas específicas ( 3 0 ) , que
presumiblemente tendrían un papel en la regulación de la
esteroidoqénesis, directamente como parte de las enzimas
biosintéticas de la vía de l o s andrógenos, o indirectamente a
través de diversos mecanismos de regulación autocrina o paracrina
( 3 5 ) *
Generalmente se acepta que el AMPc es el principal factor que media
la transducción de la señal evocada por LH, pero investigaciones
recientes han mostrado que otros tipos de segundos mensajeros
intervienen en la regulación de la función celular.
Es ampliamente aceptado que muchas hormonas de naturaleza proteica L

18
pueden ejercer sus efectos biolóqicos a través de la estimulación
de la hidrólisis del fosfatidilincsitol 4,5-bisfosfato (PIPz), que
produce inositol trifosfato (IP3) - encargado de la movilización de
Ca+2 a partir de sus fuentes intercelulares y de la modificación de
sus flujos transmembranales - y diacilqlicerol ( D A G ) que activa a
la proteíncinasa C (36).
Existen evidencias indirectas de que la fosfolipasa C que cataliza
la hidrólisis del "IP2está regulada por una proteína específica que
une nucleótidos de guanosina (Proteína G) , activada a su vez por
l o s estímulos provenientes de la estimulación a nivel membranal
(37) . De igual forna, la síntesis del AMPc se encuentra bajo el control dual de proteínas G estimuladoras ( G s ) e inhibidoras (Gi),
a su vez controladas por estímul.os extracelulares diversos (38) ,
que serían en Últina instancia conjuntamente responsables del nivel
global de estimulación de las fu:nciones celulares.
Otras variables que afectan la actividad de las vías
esteroidogénicas incluyen la formación de complejos de
Caf2/calmodulina (39) , y la unión de una variedad de factores de
naturaleza hormonal que cuentan con receptores específicos en las
membranas de las C L , tales como e l factor de crecimiento epidermal
(EGF) (40) , la hormona liberadora de LH (LH-RH) (41) , la insulina
y sus factores relacionados (IGFs) (42) , el factor de crecimiento
de fibroblastos (FGF) ( 4 3 ) , la vasopresina (44) y las catecolaminas
(45) -

19
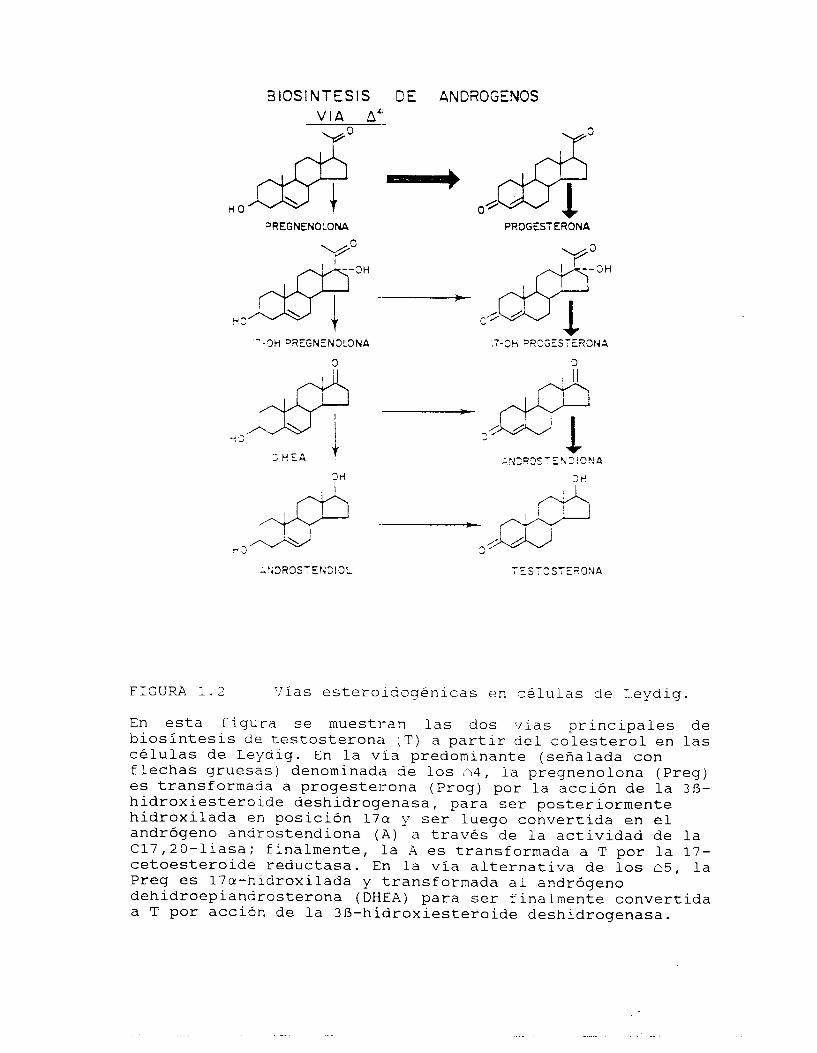
1.3.2.1. Vías esteroidoqénicas.
La esteroidogénesis en CL ha sido amplia y profundamente estudiada
para encontrar las rutas predominantes de síntesis de l o s
andróqenos, de forma tal que se conocen en la actualidad el
conjunto de intermediarios y l o s sistemas enzimáticos involucrados
en sus transformaciones, así como algunos de los factores
relacionados con su regulación (2!9) .
Recientemente se ha manifestad13 un punto de vista diferente
respecto a la evolución del proceso de la esteroiaoqénesis (46) , el
cual implica que las horr,onas es,teroides son biotransformadas en
llhormonadas'l o unidades ?e transformación en donde, de forma
organizada, se modifica un precursor hasta generar una hormona
final, sin que este compuesto o sus intermediarios puedan separarse
del agrupamiento.
Sin embargo, el esquema de producción de las hormonas esteroides
por lo general se considera agrupsdo en 2 vías diversas denominadas
de l o s c 4 y 2 5 en función de la posición del doble enlace en el
anillo A del ciclopentanoperhidrcfenantreno del mayor número de los
intermediarios que constituyen a cada una de las rutas metabólicas
(fig. 1.2).
La formación de la pregnenolona (Preg) a partir del colesterol
(Col) es el primer paso considerado como parte de las vías

2 0
esteroidogénicas. Esta reacción tiene lugar en las mitocondrias de
las CL, y requiere NADPH, oxígeno y una cadena de transporte
electrónico activa, donde es fundamental la presencia de un
citocromo P450 funcional ( 4 7 ) .
Se acepta por lo general, que el. 20a,22R-hidroxicolesterol es el
intermediario inicial del proceso de hidrólisis de la cadena
lateral del C o l mediado por la 20,22-desmolasa ( 4 8 ) , con la
consiguiente liberación de P r e q e isocaproaldehido hacia el
compartimiento citosólico (29).
Se ha propuesto que el paso limitante de la reacción Col a Preg es
la velocidad de asociación del primero con el sistema lítico
dependiente del citocromo P450, el cual está controlado a su vez
por el metabolismo del Ca+2 intra y extramitocondrial (49) . Por otra parte, la LH puede afectar también el proceso a cualquiera de l o s
siguientes niveles: disponibilidad del col y sus cofactores
reducidos, activación de la desmolasa propiamente dicha y/o
síntesis de novo de las enzimas del complejo ( 3 0 , 5 0 ) .
Existe un transportador inespecífico de lípidos llamado proteína
acarreadora de esteroles (SCP2: cuya localización y propiedades
estimulatorias parecen hacerh un candidato ideal para la
regulación de la actividad de la desmolasa de Col, ya que se
postula que podría participar en el aumento de su entrada a la
mitocondria, característico de la estimulación de las CL con LH, ya

21
que su síntesis se ve inducida por esta qonadotropina (51).
La producción de T a partir de Preg ha sido extensamente estudiada
desde mediados de la década de los 1950, cuando Slaunwhite y
Samuels (52) describieron la secuencia llamada de l o s 5 4 que
implica la serie de intermedizrios proqesterona (Proq) , 1 7 ~ -
hidroxiproqesterona (170HProg) y androstendiona (A) . Esta vía es predominante en un número importante de especies durante la etapa
adulta, incluyendo a la mayoría de l o s roedores, por lo que ha sido
utilizado como un indicador de la presencia de C L (29).
Certel & Eik-lIes (53) proporcionaron evidencias que apoyaban ia
existencia de una vía a1ter:nativa denominada de l o s 15,
caracterizada por la presencia de los intermediarios 17a-
kidroxipreqnenolona (170HPreq), dehidroepiandrosterona (DHEA) y
androstrendiol (Diol) , que tanbién puede operar en diversas
especies en mayor o menor proporción.
Se ha propuesto asimismo la existencia de algunas variantes a estas
rutas clásicas, tales como :La que involucra a esteroides
sulfoconjuqados en posición 30 (54) y algunos otros derivados
hidroxilados.
Las enzimas de la vía esteroidoqénica que catalizan la
transformación de Preg en T se encuentra localizadas en la fracción
citosólica y en el.. retículo end'oplásmico liso de las CL. Así, la

22
enzima 17a-hidroxilasa/C17,20 liasa requiere de la actividad del
citocromo P45017, asociada al retículo para generar a l o s
intermediarios A y DHEA ( 5 5 ) , la tranformación de A en T o de DHEA
en Diol es catalizada por la 17-cetoesteroide reductasa (no
dependiente de citocromo P450) ( 5 6 ) , y la transformación de Preg o
Diol en el correspondiente esterl3ide 2 4 es catalizada por la 3 0 -
hidroxiesteroide deshidroqenasa (29).
La expresión de varias enzinas de las vías esteroidoqénicas se ve
regulada por la presencia de una serie de factores de naturaleza
hornonal tales como LH, AMPc o E. Por ejemplo, la expresión de las
desmolasas dependientes de P4150 puede ser incrementada por
exposición crónica a LH o AMPc, y se ve inhibida por
concentraciones elevadas de T en CL normales o tumorales de ratón
(50) . Esta requlacidn varía de acuerdo a la edad del animal (57) ,
y se ha observado que las CL feta.les reaccionan en forma diferente
a la presencia de qonadotropina coriónica humana (hCG) ( 5 8 , 5 9 ) .
Finalmente, la prioridad que presente alguna determinada ruta
esteroidoqénica sobre l a s demás dependerá de la especie, la edad,
la estacionalidad reproductiva e incluso l o s ritmos circadianos en
los animales sensibles a estas variaciones, además de otra serie de
factores aún no claramente definidos.
SIOSINTESIS DE ANDiiOGENOS VIA A‘:
o
H O a!?$ 1REGNENOLON4 - o d $ PROGESTERONA
va
”3H PREGNENDLONA
3
FIGURA 1.2 ‘lías esteroidogénicas en c é l u l a s de Leydiq.
En esta figura se muestran las dos vías principales de biosíntesis de testosterona (T) a partir del colesterol en las células de Leydiq. En la vía predominante (señalada con flechas gruesas) denominada de los 34, la preqnenolona (Preg) es transformada a progesterona (Proq) por la acción de la 3R- hidroxiesteroide deshidrogenasa, para ser posteriormente hidroxilada en posición 17cr y ser luego convertida en el andrógeno androstendiona ( A ) a través de la actividad de la C17,20-liasa; finalmente, la A es transformada a T por la 17- cetoesteroide reductasa. En la vía alternativa de l o s n5, la Preq es 17a-hidroxilada y transformada al andrógeno dehidroepiandrosterona (DHEA) para ser finalmente convertida a T por acción de la 3f3-hidroxiesteroide deshidrogenasa.

2 3
1.3.3. Regulación funcional de las células de Sertoli.
La FSH juega un papel central en 1.a regulación de la diferenciación
de las CS y en el mantenimiento de su funcionamiento normal en las
células de animales adultos (60,61) . La unión de FSH a sus
receptores membranales específicos activa al sistema de adenilato
ciclasa, lo que incrementa la concentración intracelular de AMPc y
activa la proteíncinasa AMPc-dependiente, desencadenando una
cascada de eventos de fosforilación proteica (62).
Por otra parte, como resultado de la estimulación hormonal, la
actividad secretora de la CS se ve aumentada (63) . Dentro de las proteínas cuya secreción se VE? incrementada se encuentran la
proteína unidora de andróqenos (ABP) (ó4,65) , la transferrina
(66,67), el activador tisular de plasminóqeno (68,69), la inhibina
(70,71) , diversas qlucoproteínas ácidas (63,72) , proteínas que unen
a IGF (73), etc.
Diversos autores han demostrado que el incremento en la secreción
proteica de las CS está relacionado directamente con el aumento en
los niveles de RNAs mensajeros específicos de estas proteínas, al
menos en el caso de la ABP (74), transferrina ( 7 5 ) y activador de
plasminógeno (76) .
Al igual que en el caso de la regulación de las CL, la activación
de la adenilato ciclasa de las CS está mediada por algunas

2 4
proteínas Gs (77) , y quizá también con la participación de proteínas
del tipo Gi (78).
Por otra parte, la interacci6n de FSH con los receptores
membranales también determina un incremento en la producción in
vitro de E por parte de las CS provenientes de animales inmaduros
(79). Un incremento significativo en la producción de E se observa
desde el primer día de incubación en presencia de FSH (80), aunque
esta capacidad disminuye considerablemente hacia l o s 20 días de
edad del animal, al tiempo que la tasa mitótica de las CS hace lo
propio ( 8 1 , 8 2 ) .
Las CS in vitro poseen una alta tasa glucolítica (83) y producen
lactato y piruvato que puede ser detectado en los medios de
cultivo. Este proceso es estimulado por la presencia de FSH ( 2 3 , 8 4 )
y dibutiril AMPc (dbAMPc) (85). Este fenómeno se ha interpretado
como parte de la contribución que FSH realiza al proceso de la
espermatogénesis, ya que tanto el lactato como el piruvato
producidos permiten que l o s espermatocitos paquitenos y las
espermátides redondas, incapaces de realizar glucólisis por sí
mismas, completen su diferenciación (86).
Diversos factores hormonales solubles participan asimismo en la
regulación fUnCiOna1 de las CS, tales como EGF ( 8 7 ) , IGFs-I y I1
(88,89), andrógenos (go), FGF y somatomedina C (91) , los cuales
afectan tanto la actividad de aromatasas celulares, como la

25
síntesis de proteínas específic,ss, en cultivos primarios de CS
provenientes de animales inmaduros.
1.3.4. Mecanismos intratesticulares de regulación.
Existen evidencias cada vez más concluyentes de que l o s cambios
sutiles de la función testicu1a.r pueden ser regulados en forma
local, a través de interacciones de tipo paracrino y autocrino que
involucran contactos intercelulares, así como por interacciones
mediadas a través de factores sol.ubles de naturaleza diversa (92) .
Así, las hormonas hipofisiarias proporcionarían el estímulo basal
del cual depende en primera instancia el funcionamiento testicular,
mientras que la regulación fina del tiempo e intensidad de las
respuestas de estirpes celulares particulares, dependería de las
complejas interacciones inter e intracompartamentales.
Algunos de los mecanismos de regulación intratesticular para los
cuales diversos investigadores han obtenido pruebas experimentales
se muestran en la fig 1.3.
1.3.4.1. Interacciones células de Sertoli-células de la línea
germinal.
Debido a que la espermatogénesis se encuentra principalmente bajo
el control de T y FSH, y a que las células de la línea germinal

26
carecen de receptores específicos para estas hormonas, las
interacciones entre las CS y las células de la línea germinal
parecen jugar un papel esencial para la adecuada maduración de los
gametos masculinos.
La viabilidad de las células germinales disminuye rápidamente
cuando éstas son aisladas, mientras que en cultivos in vitro de
fracciones tubulares que incluyan CS, las células de la línea
germinal pueden sobrevivir estableciendo contactos con éstas ( 9 3 ) .
Estos cocultivos permiten incrementos en la síntesis del DNA y RNA
de las células germinales que no pueden ser mimetizados por
cocultivos en presencia de otras estirpes celulares de naturaleza
epitelial (94) .
El desarrollo de las células de la línea germinal está muy
precisamente determinado en términos cemporales y espaciales, y
esta organización parecería depender en primera instancia de la
serie de puentes citoplasmáticos que se establecen entre las CS y
ellas, si bien en número relativamente escaso (95) y de una forma
muy plástica ( 9 6 ) .
Es muy claro el papel de I1nodrizal1 que las CS cumplen con respecto
a las células de la línea germinal, ya que el establecimiento de la
barrera hematotesticular deposita en ellas la misión de canalizar
los diversos componentes metabólicos esenciales para la
supervivencia celular hacia los diferentes estados de

2 7
diferenciación con l o s cuales establecen contacto (97) . De igual manera, las grandes limitaciones metabólicas que presentan
especialmente los espermatocitos y l a s espermatides, determinan la
elevada dependencia de estas células de los productos que les
proporcionan las CS (98,99) .
Como ya se ha mencionado, las cs poseen una elevada tasa
glucolítica y producen lactato :y' piruvato que liberan hacia el
medio. Las células de la línea germinal, por su parte, son
incapaces de subsistir utilizando como sustrato de oxidación a la
glucosa a pesar de contar con todas las enzimas glucolíticas (99),
pero sí son capaces de utilizar al lactato y al piruvato para
mantener los niveles basales de ATP necesarios para sobrevivir
(100) .
Un posible mecanismo mediante el. cual las CS pudieran afectar la
diferenciación de las células germinales es la secreción de
factores paracrinos específicos,. Debido a que la organización
temporal y espacial del epitelio seminífero varía, se esperaría que
estos factores variaran asimismo en función de la etapa específica
del ciclo del epitelio seminífero en el cual se encontraran las
células germinales que rodean a las de Sertoli (101) , lo cual
determinaría que las células de la línea germinal tendrían también
la capacidad de influir sobre el. metabolismo de las CS.
LeMagueresse & Jégou (102) han mostrado que las células germinales

28
pueden afectar la producción de E y la síntesis de ABP de las CS de
ratas inmaduras i n v i t r o de u:na forma diferencial. De forma
equivalente, las células de la línea germinal son capaces de
estimular la actividad de la adenilatociclasa membranal de las CS
(103), de disminuir la secreción de transferrins (104) y de modular
la secreción de la proteína cíclica-2 (105) en cocultivos con
células de ratas inmaduras.
La presencia de células peritubulares en cultivos de fracciones
enriquecidas en CS incrementa la tasa de supervivencia de estas
últimas (106), y permite que las células se organicen en
estructuras tubulares, tal como se observa al cultivar las CS en
matrices extracelulares (107), y que incluso incrementen su
actividad secretora i n v i t r o (108).
Diversos estudios han demostrado que la membrana basal que limita
a los túbulos seminiferos está compuesta de moléculas tales como
fibronectina, coláqenas I y IV y proteoqlicanos ricos en condroitín
sulfato que son depositados por las células peritubulares, amén de
otros proteoglicanos ricos en heparán sulfato y laminina aportados
por las CS (109).
Por otra parte, las células peritubulares secretan una proteína
denominada P-Mod-S de un peso molecular de 70 KDa y cuya síntesis

2 9
es incrementada por la T (110), la cual estimula en forma máxima la
secreción de transferrina, ABP e inhibina, sin afectar los niveles
de activador de plasminógeno presentes en l o s medios de cultivo
(111,112) , a través del aumento de l o s niveles intracelulares de
GMPc (113).
Otras interacciones requlatorias entre células peritubulares y de
Sertoli parecen involucrar a los factores de crecimiento y
transformación Q y R (TGF-Q y TGF-6) y al EGF que son secretados
y/o presentan receptores específlcos en alguno o en ambas estirpes
celulares (114), y a la T que cuenta con receptores específicos en
ambos tipos celulares (115).
1.3.4.3. Interacciones células de Sertoli-células de Leydiq
(Interacciones intercompartamentales).
Como se ha señalado anteriormente, las CL son capaces de modular la
actividad funcional de las CS a través de su producción de T, la
cual es captada por éstas a través de receptores específicos, lo
que resulta en una estimulación de la actividad de RNA polimerasa
y la síntesis y secreción de AB€' y transferrina (116).
El concepto de que las CS pudieran a su vez influir sobre la
actividad de las CL ha venido afirmándose a partir de la
determinación de que daños estructurales a secciones particulares
de túbulos seminiferos pueden afectar secundariamente la morfología

Papadopoulos y cols (119), Verhoeven & Cailleau (120) y Yixun &
Dah1 (121) han descrito la activi'dad de un factor estimulador de CL
producido por túbulos seminiferos de testículos humanos o de rata
o por cultivos primarios de cs que parece incrementar la síntesis
de T en cultivos prirnarics de CL en períodos cortos de incubación
(4-5 horas) . Este factor parece estimular un paso temprano de la vía esteroidogénica, bloqueandc, a la vez en forma parcial la
conversión de los precursores C2len andrógenos ( 1 2 2 ) .
Además de estos efectos agudos sobre la esteroidoqénesis, las CS
parecen tener efectos tróficos a largo plazo sobre las CL. Así,
cuando se establecen cultivos primarios de CL de cerdo en medios
condicionados de CS (MCCS) cultivadas en presencia de FSH e
insulina, se observa en las primeras la disminución tanto en el
número de receptores para hCG como en la respuesta esteroidogénica
asociada a esta estimulación (125).

VASO SANGUINE0
TEJIDO INTERSTIC IAL TUBULO SEMlNlFERO
FIGURA 1-13 1,Tccanismos intratest iculares de regal.ación.
Diversos investigadores han postulado v a r i o s mecanismos de regulación paracrina de la función testicular que se ilustran en esta figura. De esta forma, se ha propuesto que la testosterona ( I r ) sintetizada por las células de Leydig puede ser tomada por las células de Sertoli en el túbulo seminífero a través de receptores específicos, modulando la síntesis de varias proteínas tales como la transferrina y la proteína unidora de andróqenos (ABP:, las cuales pueden influir sobre la diferenciación de l a s ciilulas de la línea germinal. Cabe mencionar que e s t a s últimar; podrían modificar a su vez la actividad funcional de las células de Sertoli, lo que no se muestra en la figura. Por otra parte, las cé1ul.a~ de Sertoli podrían modular la esteroidogénesis de las células de Leydig a través de su producción de 17B-estradiol (E) , a nivel de las enzimas de las vías biosintéticas de andrógenos y/o mediante su interacción con receptores específicos, o bien a través de la secreción de factores proteicos particulares. Las interrelaciones entre las células peritubulares y las células de Sertoli no se indican específicamente en esta figura.

126387 3 1
La diversidad de las respuestas evocadas por las CS sobre las CL
parece ser debida a la diversidad de condiciones experimentales
existentes entre los modelos utilizados por los diferentes grupos
de investigación, en términos d.e p.ej., edad de los animales,
composición de los medios de cultivo, etc (30) .
De igual manera, la naturaleza del o los factores involucrados en
estas interacciones queda por ser determinada. Algunos factores de
naturaleza inhibitoria ya descritos son presumiblemente de
naturaleza proteica y pesos moleczulares que oscilan entre 10 y 3 0
KDa (122,123,126,127), aunque diversos autores han intentado sin
éxito relacionar unívocamente alguna de las proteínas secretadas
por las CS con el factor moduladclr de la esteroidogénesis (72,128-
130).
1.4. MODELOS DE ESTUDIO: OBTENCION DE FRACCIONES CELULARES
ENRIQUECIDAS.
LOS sistemas originales de análisis concernientes a la función de
las CS comprendían el uso de túbulos seminíferos completos,
separados del compartimiento intersticialmediante disección húmeda
al microscopio estereoscópico (1:31). Estudios posteriores utilizan
una serie de tratamientos enzimáticos (DNAsa, colagenasa,
hialuronidasa, pancreatina y otras enzimas) para lograr la
disgregación de las asociaciones entre células germinales y CS, y

3 2
por lo tanto, la obtención de células individuales o de acúmulos de
un número celular limitado (132-1.37).
La mayoría de estas investigaciones han utilizado organismos
inmaduros o a lo sumo púberes para disminuir la contaminacijn
resultante de la presencia de una gama completa de estados de
diferenciación de las células de 1.a linea germinal. Algunos autores
han argumentado (138) que estas ciilulas se asemejan en cierto grado
a sus contrapartes adultas, una vez que la edad del animal hubiera
rebasado los 15-17 días, etapa en que se establecen las uniones
estrechas in vivo.
Por otra parte, la eliminación total del componente germinal para
crear un estado semejante al síndrome clínico de aplasia germinal
puede ser inducida por una variedad de métodos que incluyen la
administración de busulfán durante el desarrollo embrionario (139),
alimentación con dietas deficientes en vitamina A (140) ,
irradiación fetal (141), criptorquidia (142) o esterilización por
calor (143). Todos estos métodos dañan a las células germinales,
pero también afectan a las CS y al compartimiento intersticial en
grados variables (144), lo que pone en tela de juicio el valor de
estas aproximaciones para obtener fracciones celulares
enriquecidas.
En el caso de las células del compartimiento intersticial, se han
aplicado diversos tratamientos a las fracciones crudas obtenidas

33
por tratamientos iniciales con colagenasa para separar a los
túbulos seminiferos (145), que incluyen su paso por gradientes de
Ficoll (146), Percoll (147) o Metrizamida (148). Estos diversos
tratamientos pueden llegar a .modificar la viabilidad y las
propiedades funcionales de las cglulas intersticiales separadas,
particularmente para el caso de :-as CL (149).
Los cultivos primarios de l a s diferentes estirpes celulares del
testículo se han convertido en una metodología cada vez más
utilizada para el estudio de los diversos elementos de la
fisiología gonadal. Nediante el a.islamiento inicial de l a s células
y su mantenimiento durante períodos nás o menos prolongados en
condiciones i n v i t r o , los investigadores han podido analizar sus
respuestas directas a hormonas, nutrientes u otros factores.
. .

3 4
2. PLANTEAMIENTO DEL PROBLEMA.
Los trabajos que utilizan como modelo de estudio un órgano completo
i n vivo han sido en la mayoría de los casos la aproximación
experimental inicial a través de la cual se intenta dilucidar tanto
su fisiología como su participación en el funcionamiento global del
organismo.
Una preocupación presente en fcrma permanente en l o s diversos
investigadores es la validez que un modelo i n v i t r o tiene frente a
su referente original. Este cuestionamiento ha llevado a una
búsqueda continua de criterios de validación que permitan eliminar
todas las dudas planteadas a este respecto.
Para el caso del estudio de la fisiología testicular todas estas
inquietudes ya han sido consideradas, y los cuestionamientos han
girado principalmente respecto a si la edad de los animales o la
forma de obtención de las gónadas afectan o no el funcionamiento
normal de estos órganos, siendo por lo tanto necesario plantear

35
esquemas que permitan valorar dicho funcionamiento, teniendo
siempre en mente lo ya conocido para el animal completo.
En consecuencia, la utilizacitjn de metodologías simples y
fácilmente reproducibles para la obtención de fracciones
enriquecidas en determinadas est.irpes celulares del testículo en
animales adultos y normales, y' su correspondiente evaluación
funcional una vez aisladas, permitirá la validación de estos
modelos celulares i n v i t r o tanto para el estudio de l a s condiciones
normales de funcionamiento gonadal como también para la
determinación del efecto que diferentes factores endógenos y
exógenos presentan sobre éstas.
. -

3 6
3. HIPOTESIS DE TRABAJO.
Es posible obtener fracciones enriquecidas en células de Leydig y
Sertoli a partir de testículos de ratas adultas normales
preservando su integridad morfológica y funcional, de forma tal aue
puedan servir como un modelo de estudio in vitro de los mecanismos
de regulación paracrina y autocrina que participan en el control de
las funciones testiculares.
4. OBJETIVOS.
a. Diseñar una metodología que permita la obtención de fracciones
enriquecidas en C L y CS de ratas adultas normales a partir de un
mismo testículo.
b. Evaluar la pureza, viabilidad, y grado de contaminación cruzada
de las fracciones fracciones enriquecidas en C L y CS.
c. Determinar la presencia de componentes unidores de hormonas
esteroides en las fracciones enriquecidas en C L y CS.
d. Determinar l o s patrones esteroidogénicos presentes en las CL y
CS tanto en condiciones basales como de estimulación aguda.

3 7
5 . MATERIAL Y METODOS.
Se utilizaron ratas macho adultas normales de la cepa Wistar de
aproximamente 90 días de edad, las cuales fueron sacrificadas por
decapitación en grupos de 3 a 5 animales. Los testículos se
extirparon y la sangre secuestrada en el órgano se eliminó en la
mayor cantidad posible mediante varios lavados con una solución
amortiguadora Krebs-Ringer pH 7.4, adicionada con glucosa al 0.2%
(KRBG: 125 nM rJaC1, 5 mM KC1, 1.2 mM MgS04, 35 mM Tris, 1 m!'
NaH2P04).El peso promedio del pair de testículos fue de 3.5 g, con
un rango que oscil6 entre 2.8 a 4.1 g .
5.1. OBTENCION DE FRACCIONES ENRIQUECIDAS.
La metodología de separación est6 basada en la reportada por Welsh
& Wiebe (134) para las CS de animales inmaduros y por Janszen y
cols (146) para las CL, modiflcada de tal forma que pudieran
obtenerse ambos tipos de fracciones celulares a partir del mismo
par de testículos (fig 5.1).
Los testículos fueron procesados como se mencionó anteriormente
para eliminar la sangre retenida y fueron descapsulados, para ser
posteriormente tratados con una solución de colagenasa (Sigma, Tipo
Ia) a una concentración final de 1 mg/ml en un volumen total de 7
m1 por par de testículos, durante 18 minutos a 3 7 O C , con agitación
.

30
periódica.
La actividad de la colagenasa fue suspendida por la adición de 2
volúmenes de solución salina isotónica y dejando reposar la
suspensión durante 10 minutas a temperatura ambiente. A
continuación, dicha suspensión se filtró a través de una malla de
nylon con un poro de diámetro entre 50 y 100 pm, para retener a los
túbulos seminíferos, y separar las 2 fracciones crudas iniciales.
La fracción intersticial cruda SE! centrifugó a 1,500 x g durante 10
minutos a una temperatura de 1 O C , y la pastilla celular se
resuspendió en un volumen final de 1 m1 de KRBG, para ser
posteriormente colocada sobre una solución KRBG-Ficoll 400-Albúmina
sérica bovina fracción V (Sigma) , pH 6.5, a una concentración final
del 13% y 0.2% respectivamente, y se centrifugó a 1.000 x g durante
15 minutos a la temperatura anteriormente señalada, para recuperar
la fracción enriquecida en CL.
La fracción tubular cruda que había quedado retenida por la malla
fue recuperada y fragmentada con bisturí hasta quedar compuesta por
segmentos tubulares de 1-3 mm de longitud, para ser posteriormente
tratada con una solución de KRBG-pancreatina (Sigma) a una
concentración final de O. 2 mg/ml durante 20 minutos a 28OC, con
agitación periódica. La activida.d de la pancreatina fue suspendida
por enfriamiento a 4OC, y la suspensión fue nuevamente fragmentada
en forma mecánica para desprender los grupos de células germinales

Testículos descapsulados
KRBG-Colagenasa (1 mg/ml) 18', 37OC, c/aqitación
Adición de 2 vols. de NaCl 0.14M y dejar reposar a T ambiente durante 10'
Filtración a travc5s de una malla de diámetro de poro de 50-100 bm
Fracción tubular cruda Centrifuqación a 1500 x g durante 10' a 5OC
Resuspensióndelbotón celular en KRBG
Centrifugacióna 1000 x q durante 15' a 5OC a través de KRBG-Ficoll -BSA y recuperación del
botón celular
FIGURA 5.la Obtención de fracciones enriquecidas en células de Leydig a partir de testículos de ratas adultas normales.

Fracción tubular cruda
Fragmentación mecánica y con KRBG-pancreatina (0.2 mg/ml) durante 2 0 ' a 2 8 O C, c/aqitación
Suspensión de la actividad enzimática por enfriamiento a 5OC durante 10'
Fragmentación mecánica por agitación violenta y paso a través de una aguja de calibre 22 qa
Sedimentación a través de un gradiente discontinuo de sacarosa en KRBG 1 : 2 - 6 % ) durante 30' a 5OC
Recuperación de las fracciones correspondientes al 4 y 5% de sacarosa y filtración a través de una malla de diámetro de poro 50-100 pm
Centrifuqación a !500 x g 5' a 5OC y resuspensión del botón celular
FIG 5.lb Obtención de fracciones enriquecidas en células de Sertoli a partir de testículos de ratas adultas normales.

3 9
embebidos en l o s fragmentos tubulares.
La suspensión celular fue sedimentada a continuación a través de un
gradiente discontinuo de sacarosa en KRBG (2-6% en intervalos
simétricos, cada uno con un volumen de 5 ml) a 1 x q , durante 30
minutos a una temperatura de 4OC. Pasado este tiempo, se
recuperaron las fracciones correspondientes al 4 y 5% de sacarosa
y se centrifugaron a 500 x g durante 5 minutos a 4OC, para
recuperar la fracción enriquecida en CS.
La viabilidad celular resultante :fue evaluada utilizando la técnica
de exclusión del azul de tripano (151), con recuentos en al menos
3 campos diferentes con 100 células cada uno, por cada preparación
de un par de testículos. La pureza celular se evaluó con base en
criterios morfológicos, utilizando tinciones previas con fucsina
ácida, o bien microscopía de contraste de fases.
5.2. ENSAYO DE PROTEINAS UNIDORAS.
Las fracciones enriquecidas fueron centrifugadas a 1,500 x q
durante 5 minutos a 4 OC, y resu.spendidas mediante una agitación
vigorosa en 6 m1 de una solución amortiguadora Tris-EDTA (10 mM
Tris, 1 mM EDTA, pH 7.4). La suspensión se centrifugó a 105,000 x
g durante 60 minutos a 4OC en una ultracentrífuga Beckman L5-65 a
35,000 rpm en un rotor Tipo 65, p'sra obtener el citosol, el cual se

4 0
guardó en refrigeración hasta su posterior uso. Los citosoles
fueron ensayados a una dilución final de 1: 10, que estudios
preliminares mostraron ser la má.s adecuada.
Se afiadieron a las alícuotas de citosoles de ambos tipos de
fracciones enriquecidas (volumen final=500 pl) cantidades variables
de esteroides radiactivos ( 3H-l, 2,6,7-T 98.767 Ci/mmol;
3~-1,2,4,5,6,7-DHT123.10 Ci/mmo.L, 3H-6,7-E42.56 Ci/mmol o 3H-l,2-
Prog 56.08 Cilmmol) mezclados con cantidades variables del
correspondiente esteroide no radiactivo, con el objeto de variar la
actividad específica inicial de la hormona ensayada.
Los tubos se incubaron durante toda la noche (18-22 horas) a 4OC,
y l o s esteroides libres fueron separados por la adición de 200 p l
de una solucicn de carbón activado-dextrán T70 (Pharmacia 0 . 6 2 5 -
O. 0 6 2 5 % , seguida de agitación vigorosa y centrifugación a 3,000 rpm
durante 15 minutos a 4OC.
La especificidad de las proteínas unidoras se evaluó a través de un
análisis por saturación, mediante la incubación de los citosoles en
presencia o ausencia de concentraciones variables de diferentes
esteroides competidores (1 y 100 x con respecto a la concentración
utilizada para los experimentos iniciales de unión). Los esteroides
no radiactivos disueltos en etanol fueron añadidos previamente a
cada tubo, y el solvente fue (evaporado a sequedad para evitar
interferencias en el ensayo, antes de la adición de los citosoles. 5

126387 4 1
La incubación se realizó ba-jo las condiciones señaladas
anteriormente.
LOS sobrenadantes se decantaron a viales de vidrio y se les
adicionaron 5 m1 de Instagel (Packard) como líquido de centelleo.
La eficiencia de conteo fue del 51% para tritio en el rango de
desintegraciones por minuto detectadas.
La obtención de los parámetros de: unión de l a s proteínas unidoras
(Kd: Constante de disociación y Número de sitios de Unión) se
realizó mediante la expresión qráfica propuesta por Scatchard
(152).
5.3. EVALUACION DE LA ESTEROIDOGENESIS.
Se incubaron alícuotas con un volumen de 500 p 1 de las suspensiones
celulares correspondientes a las fracciones enriquecidas en CL y CS
durante períodos variables ( O a 3-80 minutos) a una temperatura de
37OC , con el objetivo de evaluar su producción esteroidogénica,
mediante la determinación de l as cantidades de precursores y
producto final de la vía de los n4 esteroides generados por sus
sistemas enzimáticos.
Para determinar la respuesta de las células a la estimulación
trópica, diversas dosis de hCG (0.0003 a 3 U/ml, Gonadotropyl-C,

42
i?.oussel, act. esp. aproximada de 70 U/mg) o de FSH (O. 1 a 100
nq/ml, Sigma) fueron añadidas respectivamente a las fracciones
enriquecidas en CL y CS inCubadams durante períodos de hasta 180
minutos y bajo las condiciones se:ñaladas en el párrafo anterior.
Adicionalmente, se determinó la capacidad de transformación de
diversos precursores de la vía me'diante la adición de 'H-Preg, 3H-
?rog, 3H-170HProg y 'H-A a las f:cacciones enriquecidas en C L en
condiciones basales, de l o s mismos esteroides además de 3H-C01 en
las fracciones estimuladas con hCG y de los mismos esteroides que
en las incubaciones basales además de 3Y-T, 3H-DHT y 3H-E para las
fracciones enriquecidas en CS.
Las mediciones de los esteroides; endógenos se llevaron al cabo
utilizando radioinmunoanálisis ( R I A s ) específicos j153-155), previa
purificación de los esteroides totales mediante sistemas de
cromatoqraf ía en capa fina, de acuerdo a lo señalado en el
siguiente inciso. En el caso de l o s esteroides generados a partir
de la transformación de los diversos precursores radiactivos, se
evaluó directamente la radiactividad presente en l a s zonas de las
cromatoplacas correspondientes a cada esteroide, y se determinó el
porcentaje de transformación con respecto a la cantidad inicial de
radiactividad añadida para cada precursor.
.. .

4 3
5.4. SEPARACION Y CUANTIFICACION DE LOS ESTEROIDES.
Las alícuotas de las fracciones enriquecidas incubadas de acuerdo
a lo señalado anteriormente fueron recuperadas en un tubo de
centrífuga de vidrio de 15 m 1 con tapón esmerilado, y se les
adicionaron 5 m1 de éter etílico, seguido de una agitación vigorosa
durante 1 minuto. Los tubos fueron posteriormente introducidos en
una mezcla hielo seco-acetona ccln el objeto de congelar la fase
acuosa y facilitar su separación de la fase orgánica, la cual se
recuperó en un tubo de centrífuga de igual volumen. Este
tratamiento se repitió una vez más para cada tubo procesado, previa
descongelación de la fase acuosa después de la primera extracción.
Para llevar un control de la pérdida de material durante los
procedimientos previos a la determinación cuantitativa de los
esteroides, se añadieron a tubos procesados de forma idéntica a la
mencionada anteriormente, aproximadamente 1,000 cpm de cada uno de
los esteroides; la misma alícuota se añadió a viales de conteo que
sirvieron como monitores de la radiactividad total inicialmente
adicionada.
Una vez reunidas las fases orgánicas de las 2 extracciones, éstas
fueron concentradas por evaporación a sequedad a un volumen final
de 0.75 ml, para ser posteriormente aplicadas a las cromatoplacas
con objeto de separar los esteroides.

4 4
Los extractos fueron resuspendidos con 10 gotas de una mezcla de
metano1 al 10% en éter etílico y fueron posteriormente transferidos
a una línea de origen en la cro~natoplaca (Silica Gel 6 0 , F-254,
O. 2 5 mm de espesor, 20 x 20 cm) , marcada a 2 cm de uno de los bordes, en fcrma tal que cada muestra r?o se extendiera más de 3 mm
con respecto al punto original de aplicación. Cada tubo fue lavado
repetidamente con la mezcla mencionada con el objeto de recuperar
la mayor parte posible de la muestra. Se colocaron adicionalmente
2 yg de estándares preestablecidos de los diferentes esteroides por
separar en carriles paralelos dentro de cada placa.
Las cromatoplacas se desarrollaron sucesivamente en 3 sistemas
diferentes: a) Benceno puro, b) Benceno-acetato de etilo ( 7 : 3 ) y c)
Benceno-Metano1 ( 9 5 : 5 ) . Una vez concluida la separación en el
tercer sistema, la cromatoplaca se secó y se reveló bajo luz
ultravioleta a una longitud de onda de 254 nm para determinar la
posición de l o s c4-3-ceto-esteroides; la Preq se reveló previa
adición del reactivo de Oertel (iicido sulfúrico-etanol 2:l).
Los esteroides fueron eluídos a tubos de ensaye de vidrio con 1 . 2
m1 de éter etílico-metano1 (9:l). Alícuotas de 500 1-11 cada una
fueron utilizadas para la determinación de los esteroides de la vía
mediante RIAs específicos, o bien fueron contadas directamente en
viales de vidrio adicionados con 5 m1 de Instagel (Packard) para
determinar los porcentajes de transformación de l o s precursores
añadidos.
. . . . . . . ""

4 5
5.5. DETERMINACION CUANTITATIVA DE LOS ESTEROIDES.
Las determinaciones de los esterlDides se realizaron por medio de
R I A s específicos, utilizando como fuente de los anticuerpos sueros
de conejos previamente inmunizados con esteroides unidos a través
de diferentes grupos funcionales (preferentemente en la posición
19-OH) , en forma de hapten0 a la albúmina sérica bovina en
diferentes proporciones (156) mediante residuos de ácido succínico.
Los marcadores radiactivos utili.zados fueron: ( 3H) -1,2-C01 (44.7
Ci/mmol) , ( 3H) -7-Preq (25 Ci/mmol) , ( 3H) -1,2,G17-Prog (100 Ci/mmol) ,
( 3H) -l12-170HProg ( 5 0 Ci/mmol) , ( 3H) -1,2,6,7-A (100 Ci/mmol) , ( 3H) -
1,2,G-T (100 Ci/mmol) y (3H) -6,7-E (45 Ci/mmol) , previamente
purificados por cronatografía en (capa fina de acuerdo a lo señalado
en el inciso anterior.
Tomando en consideración las cantidades de l o s diferentes
esteroides de la vía normalmente encontradas en homogenizados de
testículo completo de rata adulta. normal (157), se eligió un rango
de concentraciones de O a 1,000 pg para la curva patrón de cada
esteroide. Se partió de una soluc.ión madre de concentración 1 mg/ml
de cada esteroide puro disuelto en etanol absoluto; a partir de
ésta, se prepararon las siguientes diluciones: a) Solución N (1
ng/pl) , b) Solución E (20 pg/pl) , y c) Solución J (0.2 pg/pl).
La curva patrón para cada esteroide se preparó por quintuplicado,
de acuerdo a las proporciones sefialadas en la siguiente tabla:
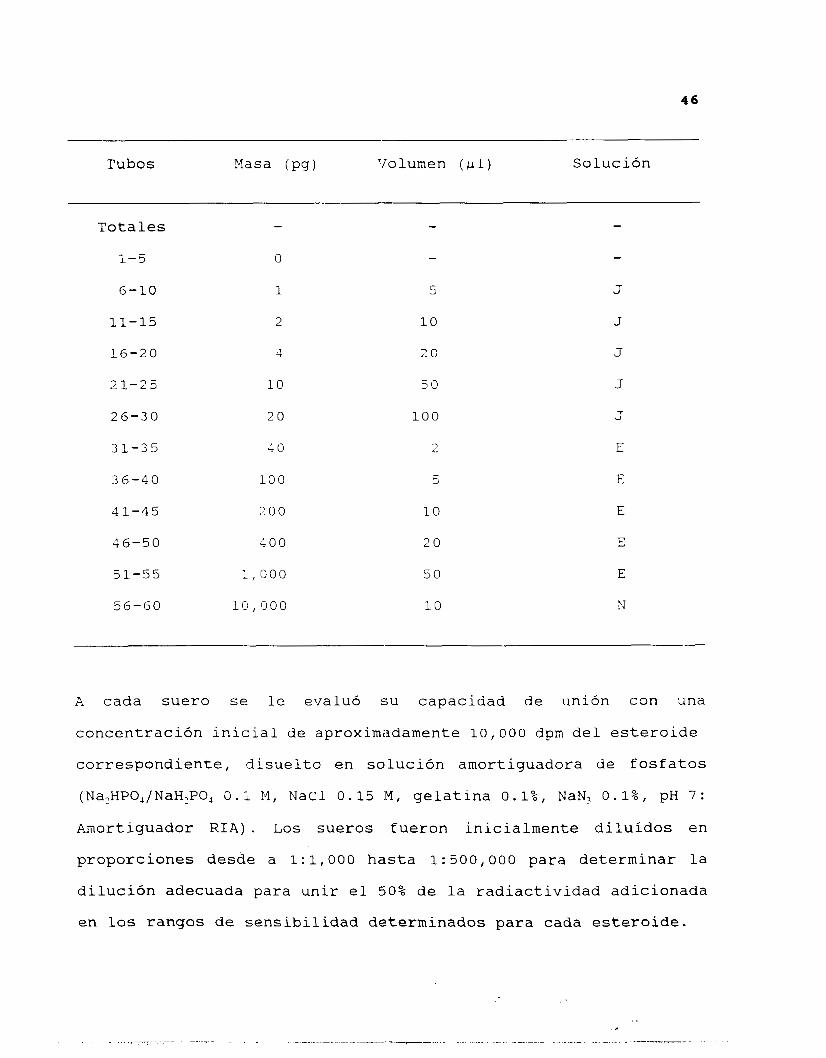
4 6
Tubos Masa (Pg) Volumen ( ,LL 1 ) Solución
Totales
1-5
6 - 1 0
11-15
16-20
2 1 - 2 5
2 6 - 3 0
31-35
36-40
41-45
46-50
51-55
56-GO
-
O
1
2
4
10
2 0
40
100
200
400
1,000
10, 000
5
10
2 0
50
100
2
5
10
20
50
10
J
J
J
J
J
E
E
E
E
E
N
A cada suero se le evaluó su capacidad de unión con una
concentración inicial de aproximadamente 10,000 dpm del esteroide
correspondiente, disuelto en solución amortiguadora de fosfatos
(Na,HPO,/NaH,PO, 0.1 M, NaCl 0.15 M , gelatina 0.1%, NaN, 0.1%, pH 7:
Amortiguador RIA). Los sueros fueron inicialmente diluidos en
proporciones desde a 1: 1,000 hasta 1: 500,000 para determinar la
dilución adecuada para unir el 50% de la radiactividad adicionada
en l o s rangos de sensibilidad determinados para cada esteroide.

4 7
Las diversas curvas patrón fueron incubadas por quintupiicado
durante 18-22 horas a 4OC para lograr el equilibrio de unión. Al
final de la incubación se agregaron 200 y1 de una suspensión de
carbón activado-Dextrán T70 (0.625-0.0625%) a cada tubo para
separar la fracción libre de la unida al anticuerpo, y se
centrifugó a 3,000 rpm durante 15 minutos a 4OC. El sobrenadante
que contenía la fracción unida se decantó a viales de conteo con 5
m1 de Instagel como líquido de centelleo y se contó en un
espectrofotómetro de centelleo ITquido Packard Tri-Carb 3390 con
una eficiencia en condiciones óptimas del 51% para 3H, como se ha
señalado anteriormente.
Las proporciones de sueros utilizadas fueron las siguientes: Preg,
Prog y 170HProg 1:10,000; A y T, 1:5,000, y E 1:3,000. Las
alícuotas obtenidas de las incubaciones de fracciones enriquecidas
en CL y CS mencionadas anteriormente, fueron procesadas de manera
semejante a lo señalado arriba, y las concentraciones de los
diversos esteroides presentes se determinaron por interpoiación en
las curvas patrón correspondientes.
Las concentraciones de los diferentes esteroides endógenos y
transformados, tanto en células estimuladas como en los controles
no estimulados, fueron comparadas mediante pruebas de T de Student,
y las diferencias entre dichas concentraciones fueron consideradas
significativas a un nivel de p menor de O. O 1 (p<O. 01) .

6 . RESULTADOS.
6.1. OBTENCION DE FRACCIONES ENRIQUECIDAS.
El procedimiento de obtención de fracciones enriquecidas permitió
separar adecuadamente los componentes de l o s compartimientos
tubular e intersticial. En el caso de la fracción intersticial fue
posible, partiendo de una composición inicial de aproximadamente el
6% de CL (fracción intersticial- cruda) obtener una preparación
final con el 50% de pureza y un 90% de viabilidad (tabla 6.1). La
principal fuente de contaminación celular en este caso correspondió
a espermatocitos primarios y espermatogonias en lo tocante a
células nucleadas, además de eritrocitos, señal de contaminación
por residuos de sangre presentes en la preparación. Debe
mencionarse que no fue apreciable la presencia de espermatozoides,
espermátides o CS, que mostrarían la presencia de componentes
tubulares en la fracción enriquecida en CL.
Con el objeto de intentar la disminución de la proporción de
eritrocitos que contaminaba la preparación final de CL, se analizó
el efecto de tratamientos hipotcjnicos sobre la viabilidad de las
células esteroidogénicas. A diferencia de lo que sucede cuando este
tipo de tratamientos se aplican al conjunto de las células
sanguíneas, l o s choques hipot6nicos afectan sensiblemente la
viabilidad de las CL, la cual disminuye de un valor inicial cercano

49
al 90% a uno equivalente al 60-70% de las células presentes.
En lo que corresponde a la fracción enriquecida en CS, el grado
máximo de pureza obtenido fue de aproximadamente un lo%, con las CS
formando acúmulos celulares, en asociación con células de la línea
germinal. La utilización de distintos procedimientos
suplementarios, tales como la centrifuqación a través de soluciones
de Ficoll de diferentes densidades no logró un enriquecimiento
sustancial en cuanto a pureza, pero sí una disminución notable de
la viabilidad celular (tabla 6.1) .
La principal fuenre de contaminación celular correspondió a las
espermátides, las cuales debido a la gran diversidad de formas que
adoptan en funcien de la etapa de diferenciación en que se
encuentran ubicadas, poseen densidades muy diversas que las colocan
a todos los niveles de los gradientes de sacarosa e impiden su
separación completa de otros tipos celulares.
6.2. CARACTERIZACION DE PROTEINAS UNIDORAS.
Se ensayaron por separado los citosoles tanto de hornogenizados
completos de testículo, como de fracciones enriquecidas en CL y CS
para buscar proteínas unidoras de T, DHT y Proq. Los estudios
iniciales se enfocaron hacia la determinación de las condiciones de
ensayo de las proteínas unidoras que permitieran su caracterización

5 0
en presencia o ausencia de las concentraciones endógenas de los
esteroides normalmente encontradas en los tejidos.
Citosoles obtenidos de hornogenizados de testículo completo fueron
incubados directamente en presencia de los ligandos radiactivos o
bien incubados previa adición de 500 p1 de una solución carbón
activado-Dextrán T70 y centrifugación a 3,000 rpm durante 15
minutos a 4 O C . Los resultados obtenidos mostraron que la diferencia
entre los parámetros de unión de ambos tipos de citosol era mínima,
por lo que se decidió trabajar en adelante con citosoles no
tratados en for7,a alguna, ya que no existia interferencia
apreciable en el ensayo por parte de los esteroides endógenos.
Los resultados obtenidos se muestran en la tabla 6.2. Es evidente
que l o s componentes unidores se encuentran localizados en la
fracción tubular; ningún compcnente unidor se encuentra en la
fracción enriquecida en CL. Todas las proteínas unidoras
presentaron constantes de disociación (K,,) en el rango nanomolar
(lO"'-lO"'h), y presentan un número limitado de sitios de unión (N,),
por lo que es muy poco probable que representen componentes
inespecíficos de unión. Es notoria la ausencia de componentes
unidores de E en ambas fracciones testiculares, si bien se encontró
capacidad unidora cie E en homagenizados de testículos completos
(datos no mostrados).
Tomando en consideración l a s características de unión de l o s

1 2 6 3 8 7 51
componentes localizados en las fracciones enriquecidas en CS se
procedió al análisis de su especificidad. Para este efecto, se
consideraron esteroides tanto del tipo 04-3-cetona (T, A y Prog),
comon5-3-hidroxi (Dehidroepiandrosterona:DHEA), Sa-reducidos (DHT)
o con aromatización en el anillo il (E).
En la tabla 6.3 se muestran los resultados obtenidos al poner en
competencia dichos esteroides a 2 concentraciones diferentes de
ligando frío (lx y 100 x con respecto a la cantidad inicial de
esteroide radiactivo añadido) con el ligando radiactivo. Debe
señalarse que el blanco del ensayo presenta una unión inespecífica
de aproximadamente el 2 0 % , porcentaje que se ha restado de los
resultados obtenidos.
6.3. ESTEROIDOGENESIS EN FRACCIONES ENRIQUECIDAS EN CELULAS DE
LEYDIG.
Las concentraciones de los diversos intermediarios y productos de
la vía esteroidogénica de los esteroides n4 se muestran en la tabla
6 . 4 y la fig 6.1. En todos l o s casos puede observarse que la
producción esteroidogénica se mantiene durante el período
analizado, lo que se refleja en e1 aumento en la concentración de
todos los intermediarios, desde el. primero (Preg) hasta el producto
final T.

52
Es asimismo evidente el cambio de la pendiente de la curva de
biosíntesis a partir de aproximadamente los 60 minutos de
incubación, lo que parecería reflejar cambios en las velocidades de
producción esteroidoqénica. Esta circunstancia se vuelve más clara
cuando se grafican las velocidades de producción de los diversos
esteroides ( p q / 1 0 6 céls/min) , como se aprecia en la fig 6.2. Cabe
mencionar que existe una correlación clara entre las
concentraciones de los esteroides y la velocidad de su síntesis.
Cuando l a s CL se ven sometidas a l a estimulación por diversas dosis
de hCG durante períodos de 180 minutos, se observa una clara
estimulación de la producción de T, la cual es dosis-dependiente en
el intervalo analizado (fiq 6.3) . La máxima estimulación (a una dosis de 3 Uinl de hCG) representa aproximadamente el 1,300 % de la
concentración control de T producida durante las 3 h de incubación.
El patrón de biosíntesis observado al medir las concentraciones de
esteroides correspondientes a sus pozas rnetabólicas se observa
también cuando se determina la transformación de 3H-Prega lo largo
de la vía (tabla 6.5 y fig 6 . 4 ) . De esta forma, hacia l o s 120
minutos de incubación, el 64.9% de la Preq añadida ha sido
transformada prácticamente en un 60% hacia ambos andróqenos ( A y
T) , y esta proporc.ión se incrementa conforme se considera a los
siguientes intermediarios.
El intermediario con'sumido en mayor proporción (170HProg) es

53
asimismo el que presenta una menor concentración dentro de las
pozas metabólicas celulares. Al final de la incubación puede
observarse que no menos del 45% de cualquiera de los esteroides
transformados fue convertido en T.
En presencia de una dosis máxima de hCG (3U/ml), puede observarse
que hacia los 120 minutos de incubación se consumió totalmente el
3 ~ - ~ 0 1 añadido, lo que no se logra en las CL no estimuladas (fig
G . 5) . Por otra parte, el Col consumido entra a la vía
esteroidoqénica y es secuencialmente convertido en los precursores
de manera semejante a lo que sucede en las células no estimuladas
(fiq 6.6).
6 . 4 . ESTEROIDOGENESIS EN FRACCIONES ENRIQUECIDAS EN CELULAS DE
SERTOLI.
La producción de E por las CS durante períodos de incubación cortos
(0-180 minutos) se muestra en la tabla 6.6. Si bien la cantidad de
E sintetizada es pequeña (un máximo de 135 pg/106CS) , la producción
de dicha hormona no tiende a disminuir, aún tratándose de su
síntesis basal.
Cuando las CS son estimuladas por diferentes concentraciones de FSH
puede observarse un incremento significativo para todas las dosis,
a excepción de la más baja (O. 1 ng/ml) , a partir de l o s 30 minutos

La evaluación de la transforma.ción de diferentes precursores
marcados con tritio en la fracción enriquecida en CS demostró la
utilización tan solo de l o s esteroides de la vía de l o s 9 4 en su
transformación hacia l o s productos finales T, DHT y E (tabla 6.7).
Si bien la 3H-Preg fue metaboliza.da en aproximadamente un 2 5 % , la
radiactividad que provenía de ella no pudo ser detectada en la zona
cromatográfica correspondiente a ninguno de los c4-intermediarios.
Por otra parte, la 'H-Tadicionada. a esta fracción fue transformada
hacia los productos finales E y DHT casi en su totalidad hacia l o s
120 minutos de incubación (datos no mostrados).
. ..

FIGURA 6.1 Contenido de los diversos esteroides de la vía de los intermediarios c4 en las fracciones enriquecidas en células de Leydig de ratas adultas normales, incubadas durante períodos de hasta 180 minutos a 37OC. Cada punto representa e.1 promedio de 3 experimentos diferentes, con triplicados para cada tiempo analizado ( n : = 9 ) .
pg:n>dlbn CL
2500
2000
1500
1000 I
ANDROSTENDIONA
,,/
/
/ i
I
O 50 1 O 0 150 200
MINUTOS
O 50 1 O 0 150 200
MINUTOS * S
O 50 1 O 0 150 200
MINUTOS

FIGURA 6.2 Velocidades de producción de los diversos esteroides de la vía de l o s intermediarios -3 en fracciones enriquecidas en células de Leydig de ratas adultas normales, incubadas durante períodos de hasta 180 minutos a 3 7 ° C .
Panel A. Esteroides con 21 átomos de carbono (pregnenolona, progesterona y 17 OH- proqesterona) .
Panel B. Est.eroides con 19 átomos de carbono (androstendiona y testosterona) .
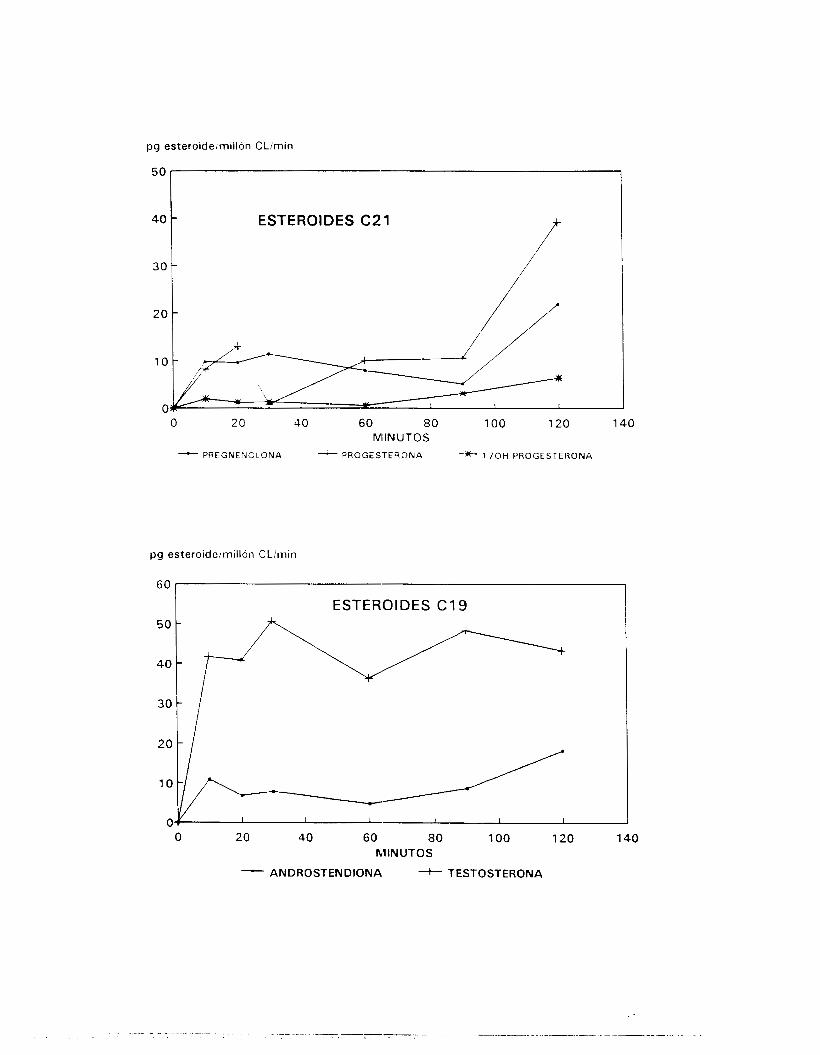
pg esteroide/rnillon CLimin
50 I 40 -
30 -
20 -
10 -
ESTEROIDES C21
/ .. , \
". - -4 I O I
O 20 40 60 a0 1 O0 120 140 MINUTOS - PREGNENCLONA PROGESTERONA 170H PROGESTERONA
pg esteroide:rnillón CLirnin
60
50
40
30
20
10
0.
ESTEROIDES C 1 9
I
O 20 40 60 80 1 O 0 120 140 MINUTOS
ANDROSTENDIONA TESTOSTERONA -

FIGURA 6.3 Síntesis de testosterona (T) por fracciones enriquecidas en células de Leydiq de ratas adultas normales, incubadas en presencia de diferentes dosis de qonadotropina coriónica humana (hCG) durante períodos de 130 minutos a 37OC. Cada punto representa el promedio de 3 experinentos diferentes, con tubos en triplicado para cada concentración de hCG utilizada (n:=9).
La producci'jn de T es siqnificativamente diferente al control no tratado con hCG (pCO.01) a pa.rtir de una concentración final de hCG de 0.03 U / m l .
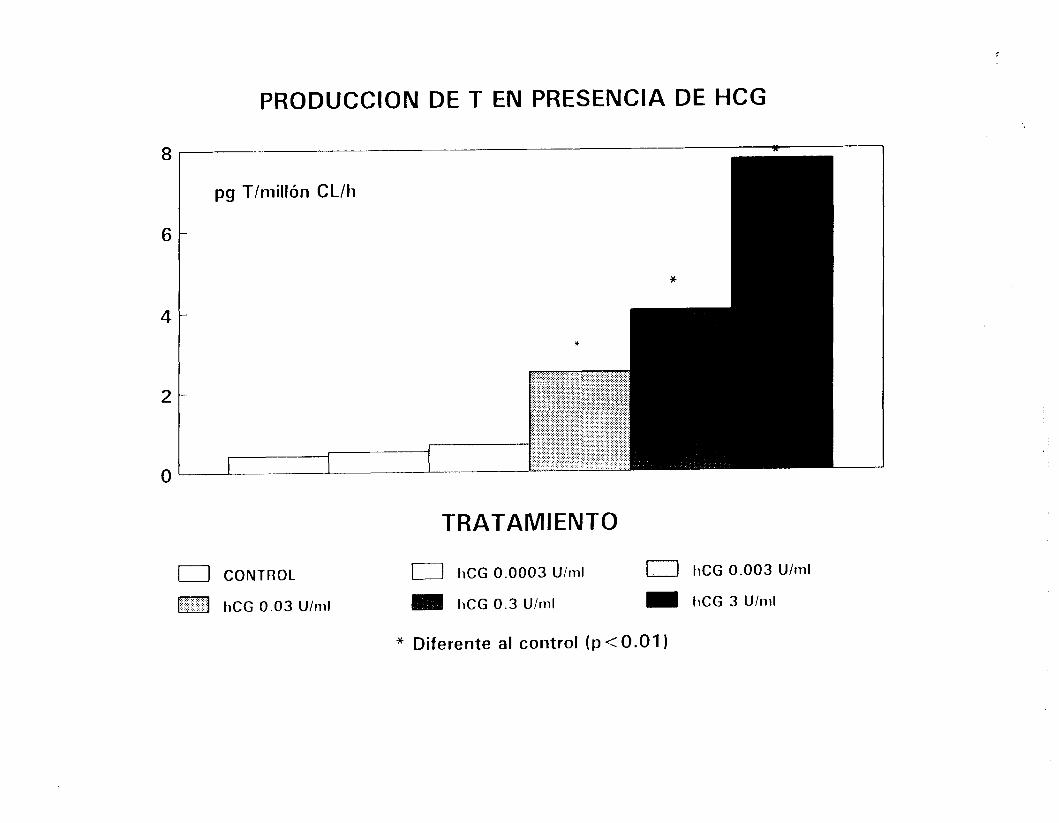
O L
PRODUCCION DE T
pg T/millón CL/h
.. .. . I
0 CONTROL n t ICG 0.03 Ulml
EN PRESENCIA DE HCG
7r
TRATAMIENTO
0 hCG 0.0003 UlmI 0 hCG 0.003 U/ml
hCG 0.3 UlrnI m hCG 3 Ulnd
* Diferente al control (p <0.01)

FIGURA 6.4 Transformacibn de l o s diversos 'H-esteroides adicionados a fracciones enriquecidas en células de Leydig de ratas adultas normales. En cada pane.1 se representa el porcentaje del esteroide correspondiente transformado a los diversos ~4-intermediarios de la vía.
Panel A. Transformación de la 'H-pregnenolona. Panel B. Transformación de la 'H-progesterona. Panel C. Transformación de la 'H-170H
Panel D. Transformaciónde la 'H-androstendiona. progesterona.
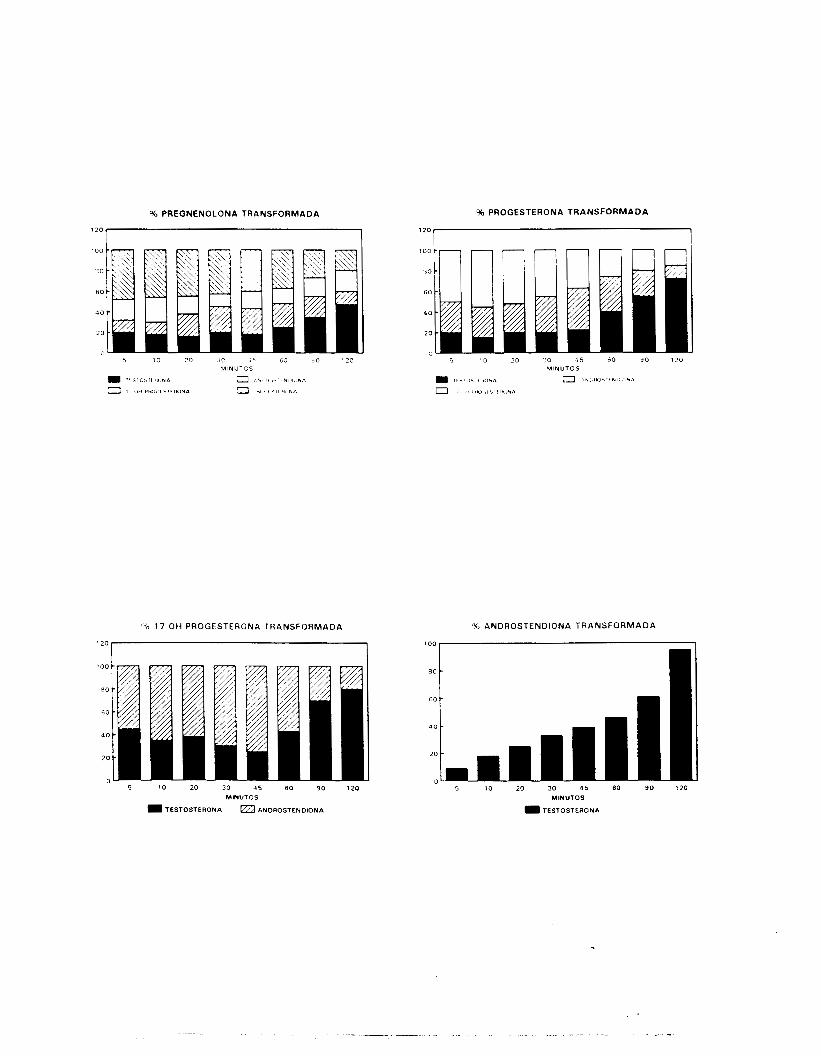
% PREGNENOLONA TRANSFORMADA
120!
% PROGESTERONA TRANSFORMADA
. O 0
S o YO
60 60
30 40
20 20
' 9 , 17 OH PROGESTERONA TRANSFORMADA
1 O 0
a o
60
40
20
0 5 10 20 30 1 5 60 90 120
MINUTOS
TESTOSTERONA ANOROSTENOIONA
1 O 0
80
60
40
20
o
'% ANDROSTENDIONA TRANSFORMADA
5 io 20 30 35 60 90 120 MINUTOS
TESTOSTERONA

FIGURA 6.5 Consumo de 'H--colesterol añadido a las fracciones enriquecidas en células de Leydig de ratas adultas normales, incubadas durante períodos de h.asta 180 minutos a 37°C. Cada punto representa el promedio de 3 experimentos diferentes, con tubos por triplicado para cada tiempo analiz.ado (n=9) .
Todos los porcentajes de transformación para las células estimuladas con gonadotropina coriónica humana (hCG) a una concentración final de 3 U / m l son significativamente diferentes con respecto a los controles no estimulados (p<O.Ol) a partir de l o s 10 minutos de incubación.
.
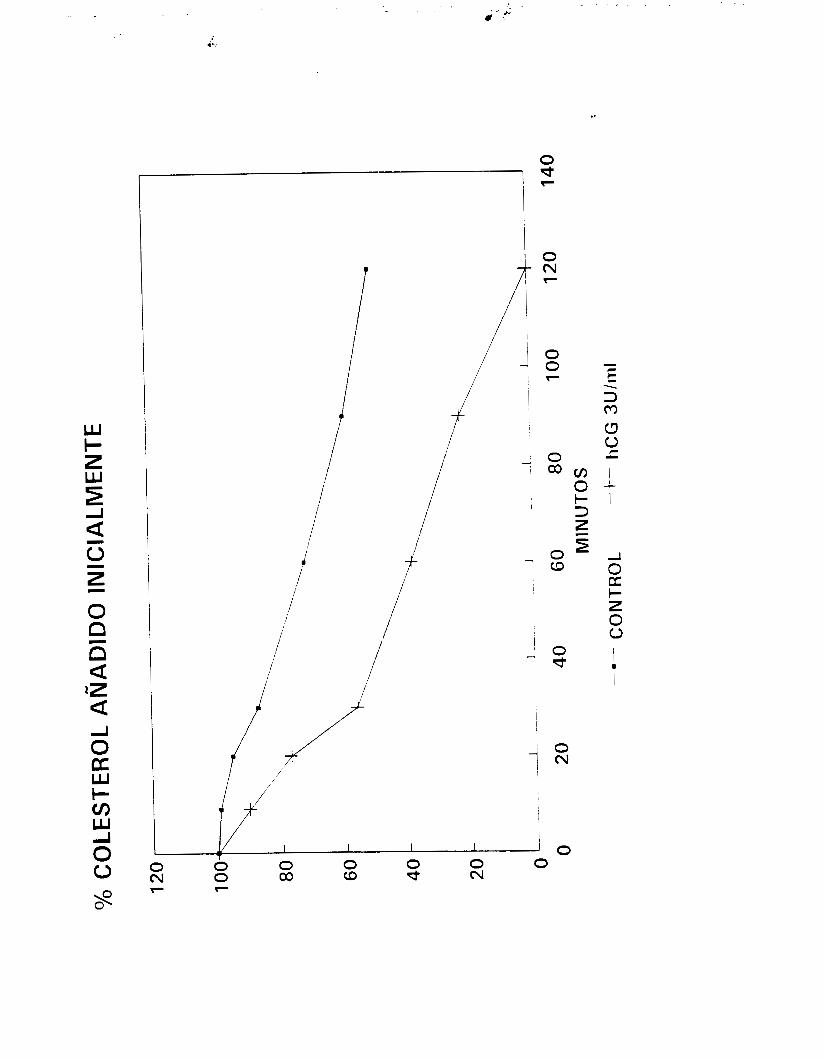
í 1
i
I
O
O O o O O O O O N O 03 CQ -3 cv

FIGURA 6.6 Porcentaje de radiactividad total añadida inicialmente en forma de 'H-colesterol a fracciones enriquecidas en células de Leydig de ratas adultas normales, que es transformada a los diverc,os c4-intermediarios de la vía esteroidoqénica, en células no estimuladas o estimuladas con una dosis de gonadotropina coriónica humana (hCG) de 3 U / m l , durante períodos de hasta 180 minutos a 37OC. Cada punto representa el promedio de 3 experimentos diferentes, con tubos por triplicado para cada tiempo analizado (n=9) .
Los porcentajes de 'H-colesteroi transformado a los diversos intermediarios es significativamente diferente en las células estimuladas con respecto a sus controles a partir de l o s 10 minutos de incubación para l o s intermediarios c21 (preqnenolona Y progesterona) , o a partir de l o s 3 0 minutos para l o s andrógenos (androstendiona y testosterona) (p<O. 01) .
Panel A. Trsnsformación a 'H-pregnenolona. Panel B. Tr3nsformación a 'H-proqesterona. Panel C. Transformación a 'H-androstendiona. Panel D. Transformación a 'H-testosterona.
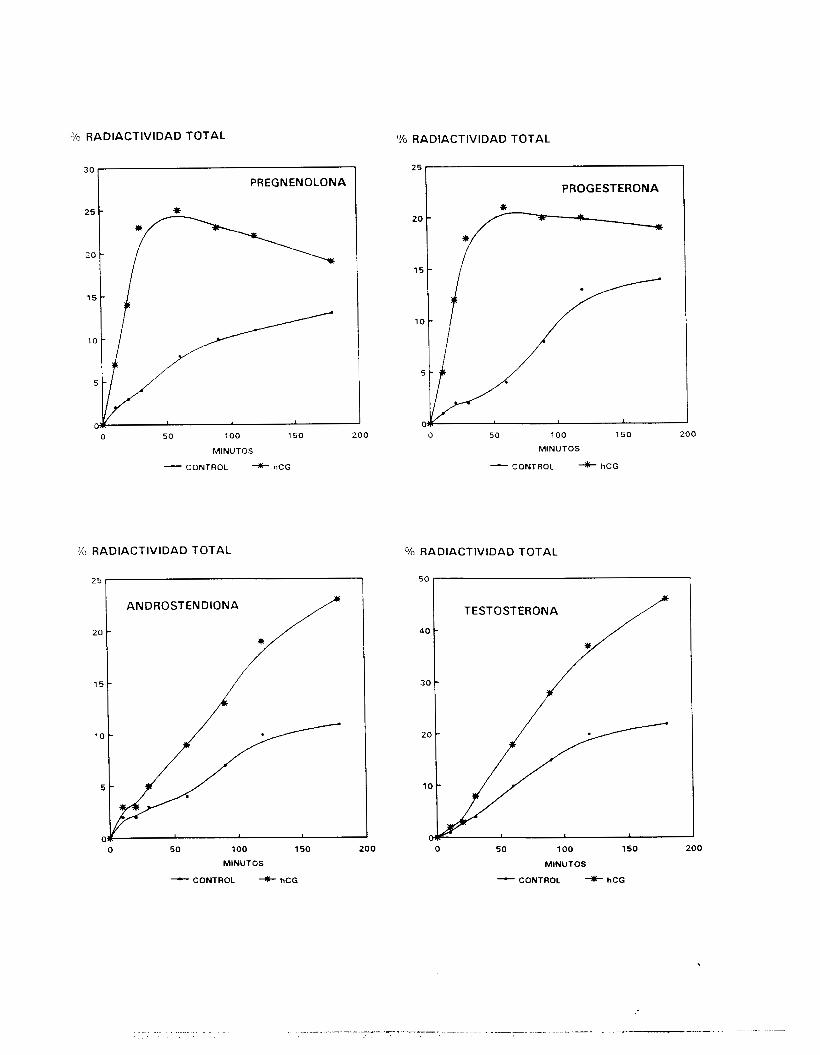
Y o RADIACTIVIDAD TOTAL
30
PREGNENOLONA
25 -
20 -
15 - f""--
O/o RADIACTIVIDAD TOTAL
- - I PROGESTERONA
O 5 0 1 O 0 150 200 O 50 100 150 200
MINUTOS MINUTOS - CONTROL -*- hCG - CONTROL --I(c hCG
'Yo RADIACTIVIDAD TOTAL
25
ANDROSTENDIONA
20
- / 15
-
'/o RADIACTIVIDAD TOTAL
TESTOSTERONA
40 -
30 -
O 50 1 O 0 150 200 O 50 100 150 200
MINUTOS MINUTOS - CONTROL hCG - CONTROL -*- hCG
. . .. . .

FIGURA 6.7 Síntesis de 1713-estradiol (E) por fracciones enriquecidas en células de Sertoli de ratas adultas normales, no estimuladas o tratadas con diferentes dosis de hormona folículo estimulante (FSH) durante períodos de hasta 180 minutos a 3 7 O C . Cada punto representa e.1 promedio de 3 experimentos diferentes, con tubos por triplicado para cada tiempo analizado (n=9).
Las cantidades producidas de E son significativamente diferentes para las células estimuladas 'con todas las concentraciones de FSH utilizadas, excepto ia más baja ( F S H 1 ) para todos l o s tiempos analizados (p<O.Ol).
FSH1: 0.1 ng/ml FSH2: 1 ng/ml
FSH3: 10 ng/ml FSH4: 100 ng/mi
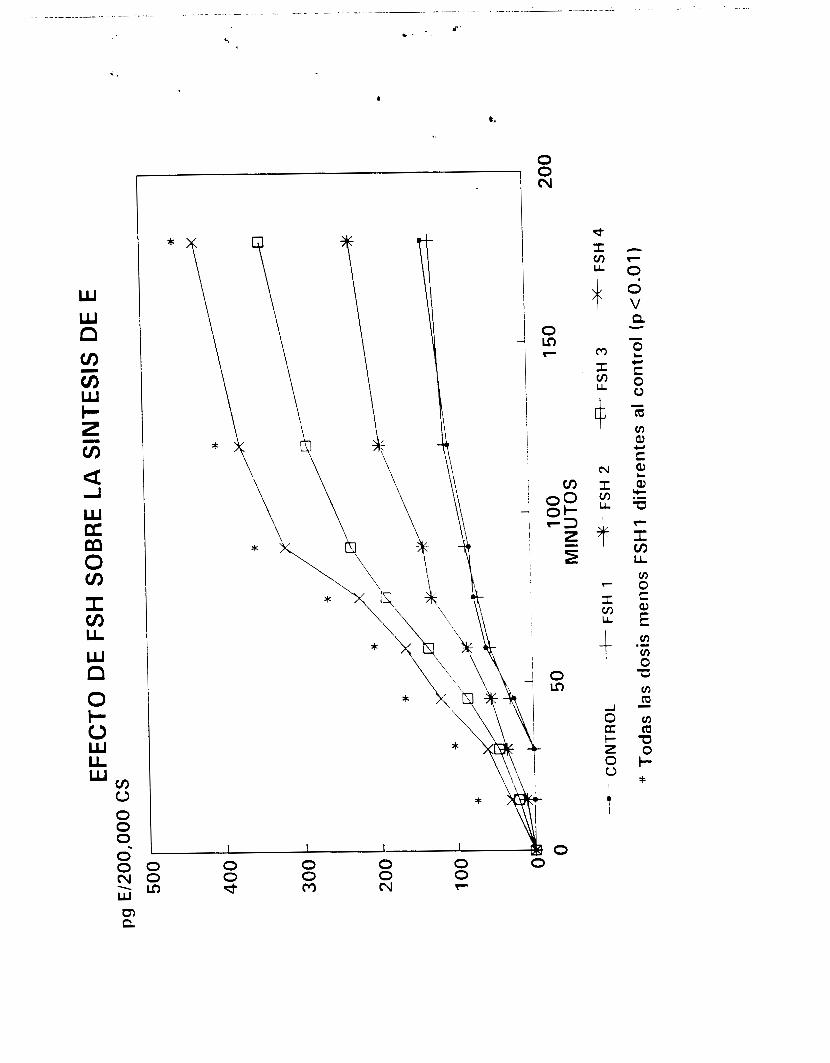
._
W W
v)
v) W I- z v)
I W ce m O v)
I v) LL W
n -
a
n O I-
LL
o O O O
O N W
o \
c.
O

FIGURA 6.8 Efecto de las diferentes dosis de hormona folículo estimulante (FSH) sobre la síntesis de 17B-estradiol en fracciones enriquecidas en células de Sertoli de ratas adultas normales, incubadas durante períodos de 180 minutos a 3 7 O C . Cada punto representa el promedio de 3 experimentos diferentes, con tubos por triplicado para cada dosis de FSH utilizada ( n = 9 ) .
La producción de E es significativamente diferente al control no estimulado ( ~ ~ 0 . 0 1 ) para todas l&s dosis de FSH excepto la más baja (FSH1) .
FSH1: O . 1 nq/ml FSH2: 1 ng/ml
FSH3: 10 nq/ml FSH4: 100 ng/ml
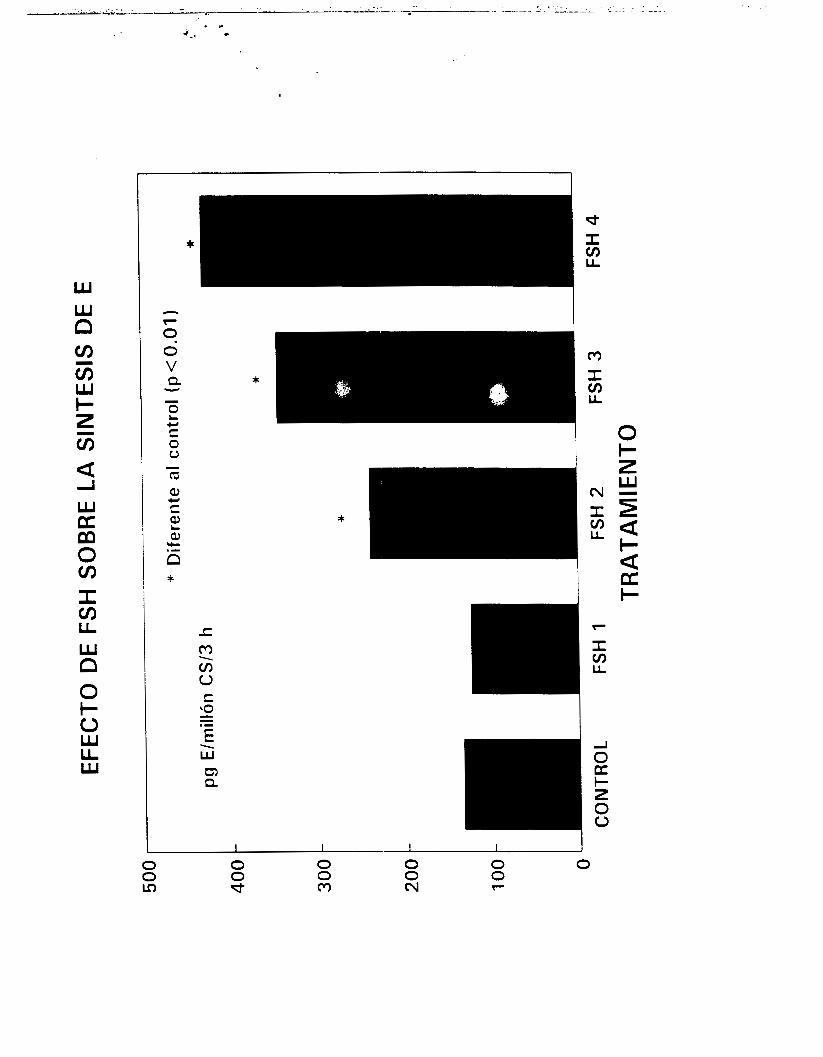
W W
cn cn W I- 7 cn
n
a A W U m O cn I v, LL W n O t- o W LL W
W m P
O O
O O
O O
O O
O O
O I- 7 W
2 a a t-
U I-
O

TABLA 6.1
CELULAS DE LEYDIG Y SERTOLI EN LAS FRACCIONES OBTENIDAS DEL TESTICULO DE RATA ADULTA
FRACCION CELULAR % PUREZA" % VIABILIDADb
Fracción de células de Leydig purificadas a través de Ficoll 5 0 f 7 ( 2 4 ) ' 90 5 3 ( 2 8 )
Fracción de células de Sertoli sedimentadas a través de un gradiente de sacarosa 7 2 3 ( 2 2 ) 92 i 2 ( 2 3 )
Fracción de células de Sertoli sedimentadas y posteriormente centrifugadas a través de Ficoll 11 2 2 ( 8 ) 80 k 12 ( 8 )
'Las células fueron teñidas con fucsina ácida e identificadas por su morfología. La contaminaci6n en la fracción intersticial corresponde a espermatocitos primarios y espermátides y en la fracción tubular las células diferentes a Sertoli son principalmente espermatocitos primarios.
bEstimada por exclusión de azul de tripán.
'Promedio 2 desviación estándar,; el número de observaciones se indica entre paréntesis.

TABLA 6 . 2
LOCALIZACION DE PROTEINA UNIDORAS DE ESTEROIDES EN FRACCIONES ENRIQUECIDAS EN CELULAS DE LEYDIG Y SERTOLI"
ESTEROIDE CELS. LEYDIGb CELS. SERTOLI'
TESTOSTERONA
Sa-DIHIDROTESTOSTERONA
PROGESTERONA
N D,
ND
ND
K,j 6.90 1 3 . 2 3 N, 1.20 i: 0 . 3 9
K, 1.00 2 0.11 N, 0.19 ? 0.06
KLi 5.50 ? O . 66 N, 0.15 k 0.01
"Los citosoles de las células aisladas fueron ensayados en cuanto a su capacidad de unión a los 3H-esteroides señalados mediante incubaciones a una temperatura de 4OC durante 18 horas y un tratamiento posterior con una solución de carbón-dextrán para separar l o s esteroides unidos tie los libres. Los datos fuercln procesados de acuerdo al análisis gráfico de Scatchard, y l o s valores de K, y N, representan los promedios k desviaciones estándar de los parámetros cinéticos de cada tipo de receptor, obtenidos a partir de un ajuste lineal de una curva de unión Construida con al menos 5 puntos diferentes, cada punto ensayado por triplicado.
bCélulas intersticiales (43-57% de células de Leydiq) . 'células tubulares (5-11% de células de Sertoli) . dND = No detectable.
'Promedio 2 desviación estándar expresado en M.
. .
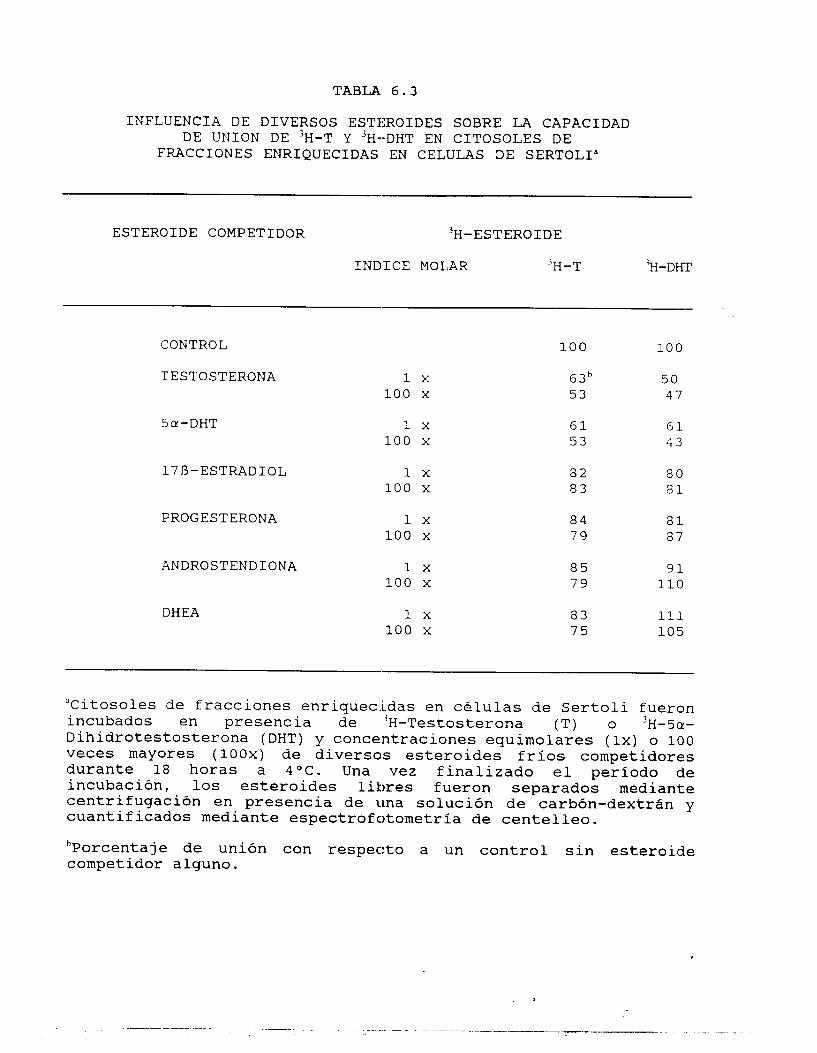
TABLA 6 . 3
INFLUENCIA DE DIVERSOS ESTEROIDES SOBRE LA CAPACIDAD DE UNION DE 3H-T Y 3H--DHT EN CITOSOLES DE
FRACCIONES ENRIQUECIDAS EN CELULAS DE SERTOLI'
ESTEROIDE COMPETIDOR 3H-ESTEROIDE
INDICE MOLAR 'H-T Ti-DHT
CONTROL
TESTOSTERONA
100 100
l x 100 x
6 3 b 5 3
5 0 47
5a-DHT l x 100 x
6 1 5 3
61 4 3
17R-ESTRADIOL l x 100 x
82 83
8 0 81
PROGESTERONA l x 100 x
84 79
ANDROSTENDIONA l x 100 x
85 79
91 110
111 105
DHEA l x 100 x
83 7 5
Titosoles de fracciones enriquecidas en células de Sertoli fueron incubados en presencia de 3H-Testosterona (T) o 3H-5cr- Dihidrotestosterona (DHT) y concentraciones equimolares (lx) o 100 veces mayores (1OOx) de diversos esteroides fríos competidores durante 18 horas a 4OC. Una vez finalizado el período de incubación, los esteroides libres fueron separados mediante centrifugación en presencia de una solución de carbón-dextrán y cuantificados mediante espectrofotometría de centelleo.
bPorcentaje de unión con respecto a un control sin esterolde competidor alguno.
+

TABLA!. 6 . 4
FRODL-CCI@?I ESTE401DOGENIC>A ZN FTWCCIOXES E:!RIQ!JECIDAS EN CELL;AS CE L Z i D I G DURANTE PERIODOS C3RTGS 3E INCUBACION"
-" LL EROIDE
TIEYPO DE IYCUB-&CION ( M I N )
3 0 G o 120 180
"2 x 10' zélulas de Le>-dig fueron incubadas por triplicado a una temperatcra de 3 7 ° C en un volumen total de 500 i ~ 1 durante períodos de 0-180 Tinucos . La actividad enzinática fue detenida por la adición ?e un volumen igual de N a O H 2N, l o s esteroides fueron extraídos con éter, separados psr cromatograf ía en capa fina y cuantificados a través de radioinmunoanálisis específicos.
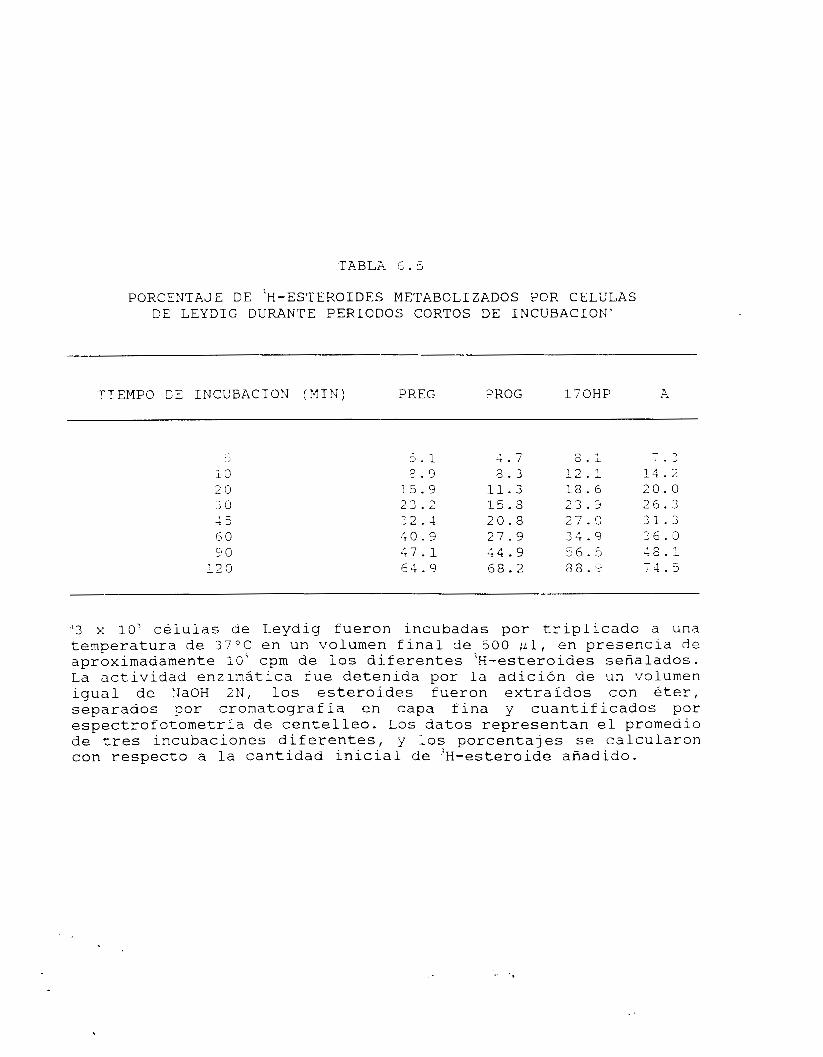
PORCENTAJE DE 'H-ESTEROIDES METABOLIZADOS POR CELULAS DE LEYDIG DURANTE PERICDOS CORTOS DE INCUBACION"
TIEMPO CE INCCBACION (YIN) ?REG PROG 170HP A
- 3
10 20 30 t 3 "
60 90
120
5.1 2.9 15.9 23.2 32.4 40.9 $7.1 54.9
4.7 8.3
11.3 15.2 20.8 27.9 44.9 68.2
- s . 1 I . " 12.1 14.2 18.6 20.0 23.3 26.3 2 7 . 0 31.3 34.9 36.0 56.5 -io. 1 8 2 . 9 14.5
" O
-
" 3 x 10' células de Leydiq fueron incubadas por triplicado a una temperatura de 3 7 O C en un volumen final de 500 p l , en presencia de aproximadamente 10' cpm de los diferentes 'H-esteroides señalados. La actividad enzimática fue detenida por la adición de un volumen igual de :JaOH 2N, los esteroides fueron extraídos con éter, separados por cromatoqrafía en capa fina y cuantificados por espectrofotometria de centelleo. Los datos representan el promedio de tres incubaciones diferentes, y los porcentajes se calcularon con respecto a la cantidad inicial de 'H-esteroide añadido.
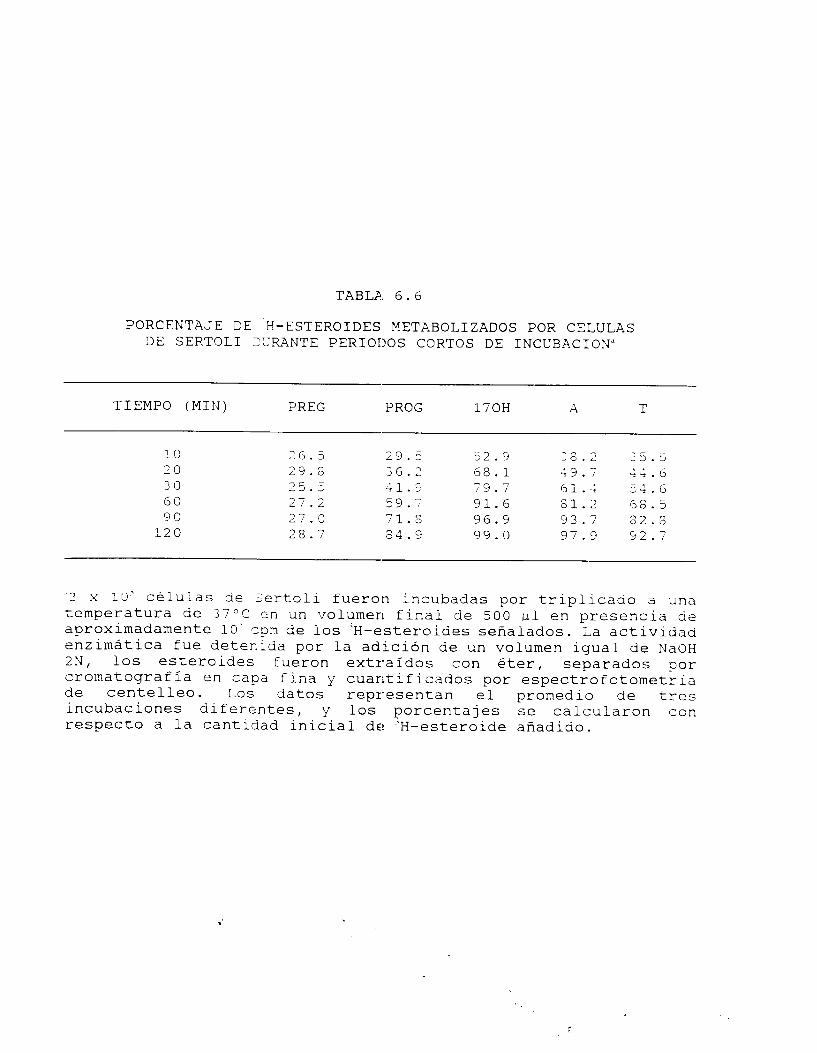
TABLA 6. G
PORCENTAJE DE 'H-ESTEROIDES METABOLIZADOS POR CELULAS DE SERTOLI SLRANTE PERIOIIOS CORTOS DE INCUBACION"
TIEMPO ( M I N ) ?REG PROG 170H A r
10 20 3 0 60 O 0 120
26.5 29.5 5 2 . 9 2 0 . 2 L 3 . 3
29.8 3 G . 2 68.1 49.7 44.6 25.5 l l . 5 79.7 61.4 54.6 27.2 59.7 91.6 31.2 6g.5 27.0 71.5 96.9 93.7 2 2 . 3 28.7 84.9 99.0 97.9 5 2 . 7
- m " -
'3 x 10' células de Sertoli fueron incubadas por triplicado a una temperatura de .37"C En un volumen final de 500 p l en presencia de aproximadamente 1O'sprn de los 'H-esteroides señalados. La actividad enzimática fue detenida por la adición de un volumen igual de NaOH 2N, los esteroides fueron extraídos con éter, separados por cromatoqrafía en capa fina y cuantificados por espectrofotometría de centelleo. Los datos representan el promedio de tres incubaciones diferentes, y l o s porcentajes se calcularon con respecto a la cantidad inicial de :H-esteroide añadido.

TABLA 6 . 7
EFECTO C E LA FSH SOBRE LA PROCLCCION DE ESTRADIOL POR FRACCIOPJES ENRIQUECII3AS EN CELULAS DE SERTOLId
XINUTOS TRAT.XMIENTOb
CONTROL FSHl FSH2 FSH3 FSH4
15
30
4 5
G O
7 - I 3
90
120
18 o
::D.
:iD
25113
G G r 2 0
7 5 ~ 1 9
50221
105127
1 3 5 ~ 3 7
: iD
r? D
30-9
55116
, Or15
85220
-
110?20
1 2 5 ~ 2 8
2 O T S
4 5 x 1 0
35x12
135123
1 9 0 ~ 2 5
235229
290t30
3i5r24
30?9
GOi10
120i23
165x23
225+34
320k22
3 7 5 2 3 2
330131
'Triplica volumen durante
.dos de tubos conteniendo 2 x 10' células de Sertoli en un final de 500 u1 fueron incubados a 3 7 O C con agitación l o s períodos señalados en la tabla. La incubación fue
detenida por adición de un volunen igual de una solución 2 N de NaOH, l o s esteroides fueron extraídos con éter y el estradiol cuantificado a través de un radioinmunoanálisis específico.
"Se añadieron las siguientes concentraciones finales de FSH: O . 1 ng/ml ( F S H l ) , 1 ng/ml ( F S H 2 ) , 10 ng/ml (FSH3) o 100 ng/ml (FSH4).
-ND = No detectable.

5 5
7 . DISCUSION.
7.1. ANALISIS DE LOS RESULTADOS DE LA OBTENCION DE FRACCIONES
ENRIQUECI2.4S EN CQMPARACION CON DIVERSAS METODOLOGIAS 'PREVIAMENTE
REPORTADAS.
Existen iiferentes netodologías [que han sido utilizadas para 1a
obtención je rracciones enriquecidas en diversas estirpes celulares
a partir 5e testículos de animales normales o alterados, adultos,
púberes c prepúberes, c u y o s rendinientos -.-arían ampliamente, al
iguai que io hacen las condiciones norfolóaicas y funcionales en
que las células se encuentran al L+lnal del procedimiento de
separacicn.
. .
LOS estudios iniciales desarrollados por Christensen y Yason (131)
permitieron la separación húmeda cruda de túbulos seminiferos y
compartimiento intersticial, no s610 en ratas adultas, sino también
en ratón, cuyo, hamster y otras Especies menores. Es, sin embargo
menester 7.encionar que las fracciones obtenidas de esta forma no
pueden ser distinguidas funcionalmente, ya que ambas son capaces de
metabolizar 4-''C-Prog con eficiencias semejantes, lo que revela una
contaminación de la fracción tubular por parte de C L .
\
Es posible obtener una mejor separación entre ambos compartimientos
cuando se utilizan enzimas proteolíticas que ayuden a la digestión

5 6
de l o s componentes que mantienen la integridad del ccnpartixiento
intersticial. Así, diversos investigadores utilizaron tratamientos
durante tiempos cortos con diferentes concentraciones de colagenasa
(134,145), DNAsa I (158) o tripsina-EDTA (137), para obtener
preferentemente a 'xna de las 2 fracciones, en szasiones en
detrimento de la integridad funcional de la otra.
Refinamientos técnicos posteriores incluyeron la ctilización de
tratamientos enzimáticos alternativos con ylicina/colagenasa (136) ,
pancreatina (134) o hialuronidasa (159) o tratamientos térmicos
post-siembra (160) para el caso de la separación para cultivo
primario de células del túbulo seminífero, así como la
centrifuqación a través de gradientes de densidad de Ficoll (146) ,
Percoll (147) o Metrizamida (148) en el caso de la obtención de
fracciones enriquecidas en CL.
La pureza de las fracciones obtenidas así como su viabilidad y
funcionalidad varían ampliamente de acuerdo a la técnica utilizada.
Así, las fracciones enriquecidas en CL pueden ilegar a tener una
pureza de cerca del 60% después de la centrifugación a través del
Ficoll (146), más del 90% cuando se utilizan gradientes de Percoll
(149), y un máximo del 94% en el caso de los gradientes de
Metrizamida (161), aún en el caso de que se manejen testículos de
animales adultos. Sin embargo, cabe mencionar que en algunos de
estos casos, particularmente cuando se utilizan gradientes de
Percoll, existen dudas fundamentadas acerca del efecto que esta
. . .

En el caso de las fracciones enriquecidas en CS, la situación varía
ampliamente dependiendo de si se utilizan animales inmaduros o
adultos para la separación de los compartimientos testiculares. De
esta forma, es posible obtener fracciones de CS prácticamente puras
cuando se trabaja con testículos de animales de 10-20 días de edad
(134), dada la ausencia de formas avanzadas de células de la línea
germinal y -3 la falta de una barrera hematotesticular
definitivamente constituida (164:. En cambio, la proporción de CS
en l o s testículos de animales adultos no llega al 1% del total
celular, y la presencia de uniones intercelulares bien
desarrolladas ccmplica los procesos de su separación y purificación
(150) .
En este trabajo se reportan Los resultados de combinar las
metodologías planteadas originalmente por Welsh & Wiebe (134) para
la separación de CS de ratas inmaduras, y por janszen y cols (146)
para la obtención de CL en ratas 'adultas, a ratas macho normales de
90 días de edad.
De acuerdo a lo señalado en la tabla 6.1, las fracciones
enriquecidas en CL tienen una pureza de 5 0 2 7 % con una viabilidad
final del 90+3%, a partir de una "pureza inicial" del 6% con

58
respecto al testículo completo. E s c o s resultados sGn comparables a
10s obtenidos por Janszen y cols (?.;6), los ccales reportan purezas
de 59?17% después de hacer pasar ].as células aisladas a través del
Ficoll, por una solución de dextr6n al 6%. Tratamientos semejantes
con soluciones de dextrán realizados a las CL purificadas por la
metodología planteada en este trabajo no incrementarcn
significativamente la pureza de la fracción, pero sí disminuyeron
la viabilidad aproxinadamente hasta un valor máximo del G O % .
En lo que toca al rendimiento cel.ular obtenido, Gale y cols (147)
utilizando centrifuqacien a travi!s de Percoll, reportan cifras de
1.5 x 10 células totales por testiculo, de l a s cuales el 74% serían
de Leydiq, mientras que Sharpe y Cooper (165) reportan un total 2e
27.824 .4 x lo6 CL de un total de 55 millones de células obtenidas
por testículo nediante tratamientcs únicamente con colagenasa.
G
El esquema de separación aquí reportado permite obtener
aproximadamente un total de 20 x 10Ucélulas intersticiales por par
de testículos, de las cuales aproximadamente la mitad serían CL
juzgadas por criterios morfológicos, previa tinción de
preparaciones fijas con fucsina ácida. Esta cifra representaría una
recuperación aproximada d e l 30% del número total de las CL, que
mediante criterios morfométricos ha sido estimada en 34 x 10'
células por testículo de rata adulta (166), y además serían viables
en una gran proporción, a diferencia de l o s casos anteriormente
mencionados, donde la viabilidad puede llegar a disminuir hasta un
r

59
70% en algunos casos.
Mendelson y cols (167) reportaron que las suspensiones crudas de
testículos de ratas adultas tratadas únicamente con colagenasa
contienen el 50% de CL libres de eritrccitos. Este reporte ha sido
cuestionado por diversos autores (p. ej. 147 j , y es muy claro que
las fracciones enriquecidas en C:L obtenidas como resultado del
desarrollo de esta tesis se ven fuertemente contaminadas con
eritrocitcs, a pesar de su centrifugación a cravés de la solución
de Ficoll-BSA 13%-0.2%. Janszen y cols (116) reportaron que la
densidad celular de los eritrocitos es aprDximadamente de 1.075
g/ml, por lo que sería imposible que éstos f,eran excluidos por la
solución de Ficoll, la cual presenta ana densidad aproximada de
1.053 g/ml.
Como resultado de la centrifugaci6n a través del Ficoll, la mayoría
de las células "ligeras1' (espermatozoides y espermátides) quedan
retenidas en la interfase, mientras que las células mayores o
"pesadas" sedimentan. Un análisis corfológico de la pastilla
celular (fracción enriquecida en CL) refleja que la principal
contaminación celular corresp0nd.e a espermatocitos primarios y
algunas espermátides, las cuales debido a sus formas muy variadas,
presentan un amplio espectro de ciensidades. Es prudente mencionar
que, al menos bajo un criterio puramente morfológico, la fracción
intersticial enriquecida parece estar libre de CS.

6 0
Cuando se comparan l o s resultados obtenidos con aquéllos reportados
por autores que utilizaron qradientes de densidad de Metrizamida o
Percoll para la separación de diferentes poblaciones de células de
Leydig, se puede observar que la densidad celular de la fracción
obtenida por centrifuqación a través de la solución de Ficoll-BSA
corresponde a las poblaciones con mayor capacidad de respuesta
esteroidogénica (ver más adelante).
En lo que toca a la fracción enriquecida en CS, iss datos
existentes en la literatura son escasos, y más bien se refieren a
sistemas de cuitivo prinario (150). Los datos presentados en la
tabla 6.1 reflejan una baja pureza (7i3%) con una elevada
viabilidad ( 5 2 t 2 % ) , una vez recuperadas l a s fracciones del
gradiente correspondientes originalmente al 4 y 5% de sacarosa.
Esta pureza representa un enriquecimiento de aproximadamente 5x con
respecto a la cantidad original de CS en un túbuio seminífero
adulto (98) , y no pudo ser incrementada sustancialmente por
centrifuqaciones posteriores a través de soluciones de Ficoll-BSA
que disminuyeron la viabilidad inicial al 80512%.

6 1
7.2. PRESENCIA DE PROTEINAS UNIDORAS DE ESTEROIDES EN LAS
FRACCIONES ENRIQUECIDAS EN CELULAS DE LEYDIG Y CELULAS DE SERTOLI.
La localización de componentes unidores de esteroides como un
parámetro para determinar el grac,o de contaminación por parte de
las fracciones enriquecidas de los diversos compartimientos
testiculares ha sido descrita anteriormente en la literatura. Es,
sin embarqo menester mencionar que no existe un total acuerdo con
respecto ni a la ubicación de 1.0s camponentes unidores en los
compartimientos testiculares, ni en lo que toca a la variación de
esta presencia en términos de la edad o condición de l o s animales
analizados.
7.2.1. Receptores de andrógenos.
La evidencia inicial de receptores de andróqenos en el testículo
provino de estudios in vivo realizados con marcadores radiactivos
en animales hipof isectomizados (168) , donde se ubicaron componentes
unidores tanto citosólicos como nucleares. Vernon y cols (169)
plantearon inicialmente la existencia de 2 diferentes componentes
unidores ubicados en l o s túbulos seminíferos de ratas adultas
hipofisectomizadas caracterizados por su alta afinidad a T y DHT,
uno de los cuales podía ser equivalente a la proteína unidora de
andróqenos (ABP) reportada originalmente por Ritzén y cols (170).
Wilson & French (171) describieron las parámetros cinéticos de

62
unión de los presuntos receptores testiculares de andrógenos, y los
compararon con aquéllos reportadcs para los receptores de
andrógenos descritos en epidídino y próstata, encontrando
constantes de equilibrio de disociación (Kd) para T y DHT de 2-5 x
IO-'%, y una mayor ?finidad por !:ET que por T. Otros receptores
citoplásmicos de andrógenos con características semejantes a las
descritas han sido caracterizados en testículos de otras especies
de mamíferos (172 , 173 j .
Estos receptores anarogénicos son claramente distinguibles de la
ABP tanto a través de sus propiedades fisicoquímicas tales como la
.Jida media de disociación del andrcjgeno unido (a T=O°C, t1/2=2 días
para el receptor y 6 xinutcs para . A B P ) , como por su comportamiento
en electroforesis en gel de poliacrilamida (Rf de 0.25 para el
receptor citoplasmático y de O. 5 para ABP en geles al 7%) , su
termosensibilidad y su patrón de especificidad (174,175).
Tindall y cols (176) describieron inicialmente la presencia del
receptor de andrógenos en fracciones enriquecidas en CS aisladas de
animales hipofisectomizados, y Sanborn y cols (177) localizaron
receptores citoplásmicos y nucleares en CS obtenidas de ratas
inmaduras y mantenidas en cultivo durante 2-3 días.
Aunque algunos autores reportan una unión muy débil al receptor de
andrógenos por parte de la cronatina de células de la línea
germinal (178), existe un acuerdo respecto a que las CS representan

6 3
el sitio principal de unión a andrógenos en el compartimiento
tubular, y de que no existe un significativo componente unidor en
la fracción intersticial del testículo, lo que convertiría a los
receptores de andrógenos en marcadores adecuados de la pureza de la
fracción tubular.
Los resultados obtenidos en este trabajo (tabla 6.2) rnuestran que
la fracción enriquecida en CS presenta componentes unidores de T y
DHT eon una Kdae 6.90t3.23 y 1.00:t0.11 nM respectivamente, dentro
del rango descrito por otros autores ya mencionados (171,176) para
receptores de andrógenos. Es claro, de acuerdo a lo señalado en
esta tabla, que no fue posible detectar componente unidor alguno en
la fracción intersticial, lo que podría representar una evidencia
de la no contaminación cruzada de l o s ccmpartimientos testiculares.
En io tocante a la especificidad de la unión, de acuerdo a lo
observado en la tabla 6.3, cuando 3H-T o 3 H - D H T funcionan como
ligandos, ambos andrógenos presentan una capacidad equivalente para
su desplazamiento de la unión, lo que se aprecia cuando actúan
Prog, A o DHEA como competidores. ;3s interesante observar que el E,
aunque en una proporción menor, logra desplazar también a la marca
radiactiva de los componences unidores, aunque su estructura
molecular no sugeriría la posibilidad de interaccionar con el
anillo A, el cual es la parte de la molécula receptora responsable
de la especificidad de unión (174).

6 4
Tomando en cuenta los resultados obtenidos, particularmente
aquéllos relacionados con la afinidad de l o s diferentes andrógenos
por los componentes unidores, y los relacionados con las constantes
cinéticas de unión, las evidencias parecerían apoyar que en este
nodelo de estudio, las especies nolecuiares que anen andrógenos
parecerían corresponder más bien a1 componente receptor que a la
proteína transportadora de andróge.nos ABP.
Sanborn 1; cois (177,179) han mostrado que ios andrógenos, en
términos generales compiten más fuertemente que los
qlucocorticoides o l o s estrógenos 'para desplazar l a narca unida en
pruebas de especificidad, resultad.os semejantes a los encontrados
por Schmidt y Danzo en cultivos de CS de animales inmaduros (180).
Si bien en este trabajo no se incluyeron a l o s qlucocorticoides
dentro de l o s esteroides competidores, datcs obtenidos por Ortlip
y cols (181) sugieren que estas 2 familias de esteroides no se
interfieren mutuamente en cuanto la unión a sus receptores
específicos.
7.2.2. Receptores de estrógenos.
Abney (182) describió un componente unidor de estrógenos en citosol
de homogenizado completo de testículo, con una Kdde 4.0721.05 x lolo
M-', cuyas características fisicoquímicas de unión difieren de
acuerdo a la edad del animal (1E83,184). Este componente unidor
parece ser muy lábil, ya que tanto la desensibilización por parte
, ..

6 5
de gonadotropinas (185), como pequeñas variaciones térmicas (186)
son capaces de inactivarlo.
Mulder y cols (187) reportaron inicialmente la presencia de un
componente unidor de E en l a s fracciones citoplasmática y nuclear
del tejido intersticial de testículo de rata aislado mediante la
técnica de disección húmeda, mientras que Van Beurden-Lamers y cols
(188) determinaron en estas mismas fracciones un intervalo de 100-
150 fmol E/mg proteína como la máxima capacidad de unión. A pesar
de esto, debe mencionarse que l o s receptores ubicados en la
fracción nuclear son muy difíciles de determinar, debido a la gran
cantidad de componentes no especificos de unión presentes en el
sistema (189).
Estudios realizados con fracciones intersticiales purificadas a
través de Metrizamida o Percoll (185,190), han permitido asignar a
las CL la ubicación de los receptores de estrógenos, l o s cuales
parecen ser muy específicos para E, ya que no permiten el
desplazamiento de l a unión de 3H-E por esteroides de ninguna otra
familia (182).
De acuerdo a l o s resultados obtenidos en este trabajo, no fue
posible detectar unión específica alguna a E en ninguna de las
fracciones testiculares aisladas. Aunque no se detalla dentro de
los resultados, l o s homogenizados de testículo completo presentan
capacidad de unión a 3H-E en el rango de Kd de lo%, capacidad que
_.

6 6
ya no puede ser detectada al ensayar las fracciones separadas.
Pueden existir varias razones que expliquen estos resultados. En
primer lugar, el tipo de 3H-E utilizado presenta una actividad
específica relativamente baja (42.56 Ci/mmol), lo cual permitiría
distinguir los componentes unidores tan solo en el caso de que se
presentaran en una concentración relativamente elevada, lo que
podría ser el caso en los homogenizados de testículo completo, pero
no necesariamente en los citosoles de las fracciones enriquecidas
en CL.
Por otra parte, la presencia de otros componentes unidores , o de
las mismas enzimas de l a vía esteroidogénica por l a s que el E
presenta afinidad ( 1 9 1 ) , podrían contribuir a "secuestrar la marcall
e impedir la adecuada visualización de un receptor específico.
Debe, sin embargo, mencionarse que la primera posibilidad señalada
es muy poco probable, ya que de acuerdo a los resultados obtenidos
en este trabajo, no parece haber componente unidor de esteroides
alguno en la fracción intersticial del testículo. En lo que toca a
la segunda posibilidad, no se tienen los datos suficientes como
para evaluar la importancia de su participación a este respecto.
Cabe aclarar por otra parte, que si bien diversos autores han
señalado la existencia de estos receptores en la fracción
intersticial del testículo, no ha podido establecerse hasta la
fecha con la suficiente claridad el papel de los estróqenos en la

7 . 2 . 3 . Receptores de proqestáqenos.
Son pocos los investigadores q u e han reportado la presencia de
eomponentes unidores de proqescáqenos en tescículos cie mamífero
(175,193). Schmidt & Danzo (180) fueron los prineros en reportar la
presencia de un componente uniaor de Proa en citosol de CS de ratas
inmaduras mantenidas en cultivo primario, el cual es relativamente
insensible a la competencia por parte del 25020, el zcetato de
nedroxiproqesterona y la T.
Estos datos sugieren que pueden existir 2 tipos diferentes de
moléculas unidoras en el citoso:L de las CS ya que, aunque las
proqestinas son capaces de interaccionar con l o s receptores de
andrógenos, no presentan una fuert.e capacidad de desplazar la unión
de 'H-T en ensayos de competencia, lo que podría representar la
existencia de un receptor específico para Proq presente en estas
células.
Los resultados obtenidos
existencia de componentes
nM ell la fracción tubular
en este trabajo (tabla 6.2) señalan la
unidores de Pro9 con una Kdde 5 . 5 0 2 0 . 6 6
enriquecida en CS, e indican que Prog no .

68
es capaz de desplazar a 3H-T o 3FI-DHT de su interacción con estos,
lo que parece estar de acuerdo con lo señalado por Schmidt & Danzo.
.l 1 . 3 . PATRONES ESTEROIDOGENICOS E:N LAS FRACCIONES ENRIQUECIDAS EN
CELULAS CE LEYDIG Y CELLLAS DE SERTOLI.
7.3.1. Fracción intersticial.
Diversos autores han reportado el estado funcional tanto basal como
posterior a estimulaciones con hClS de fracciones enriquecidas en CL
obtenidas por diferentes métodos con el objetivo de plantear cuál
de las metodologías empleadas permite obtener un mayor rendimiento
unido a un mejor mantenimiento del adecuado funcionamiento
metabólico celular.
Sharpe & Cooper (194) , al comparar los métodos de disección húmeda,
dispersión con colagenasa y una combinación de ambos, detectan una
producción basal de T de 2.920.2 :ng/106 células positivas a tinción

6 9
para 3D-hidroxiesteroide deshidrogenasa en un período de 4 horas,
la cual es Prácticamente igual a los 2 . 7 2 0 . 2 ng que se obtiene para
las fracciones resultantes de trat.amientos con colagenasa. Por otra
parte, Cooke y cols (149) determinaron que los tratamientos
mecánicos previos 3 la dispersión con colagenasa presentaban
efectos deletéreos sobre la capacidad esteroidoqénica celular.
Las células intersticiales dispersadas y posteriormente sometidas
a centrifugación a través de gradientes de densidad de Metrizamida
o Percoll se reparten en poblaciones celulares con diferencias de
densidad apreciables (195,136).
En el caso de la xtilización de gradientes de Metrizamida, la
población I (a=1.085-1.117 g/ml) presenta una producción de 22 nq
T , ! 1 0 4 células/3h, prácticamente la tercera parte de la
correspondiente a la población I1 (a=1.128-1.145 g / n l ) . Trabajos posteriores de este grupo de trabajo (197) señalan que las
diferencias en producción de T parecen reflejar al menos en parte
diferencias en la actividad de la C17,20-liasa.
Para el caso de las células separadas en gradientes de Percoll (15-
57% de pureza), Kerr y cols (196) han reportado la presencia de 3
fracciones diferentes de acu.erdo a su densidad, aunque
prácticamente no existen diferencias en cuanto a la producción
total de T, la cual es comparable a la correspondiente a células
disociadas por tratamientos con colagenasa. Cuando el gradiente
. - , .
. . . .

7 0
utilizado es más abierto (O-50%), la :.áxima producción de 7 de las
células purificadas es de aproximadamente 5 ng/106 células/2h
(198 , 199) .
Un parámetro inportante para eval,lar e1 estado funcional de las CT,
aisladas es determinar su capacidad de respuesta a la estimulacion
con diferentes dosis de LH o h C G , ya sea en términos de la
cuantificación de T y/o ANPc producidos como resultado de la
estimulación, o bien rnediante :la deterninación de I c s sitios
receptores y constantes de disociación de la unión del ligando con
sus receptores nembranales específicos.
Sharpe & Cooper (194) reportan una escimulación máxima de 10 x de
la producción de T por períodos de 4 horas en células aisladas
mediante tratamientos con colagenasa, eon respecto a la producción
basal del andrógeno, situación que también se presenta cuando la
estimulación proviene de dosis máximas de dbAMPc. Estos autores
reportan también la presencia de 3 nq de 125-I-hCGunida por 10'
células aisladas.
Las poblaciones celulares resultantes de la utilización de
gradientes de densidad para separar a las CL presentan variaciones
en términos tanto del número de receptores para hCG como de la
respuesta a la estimulación gonadotrópica que pueden generar
(195,196,200,201).

71
Las células purificadas a través de gradientes de Metrizamida
presentan un soio tipo de sitio unidor de h C G de acuerdo a sus
características fisicoquímicas (1'35), pero diferente capacidad de
respuesta en términos de producción de T y AMPc (200) . El número total de receptores para L H / h C G por célula también presenta
variaciones (197) .
las poblaciones celulares resultantes de la utilización de
qradientes de Percoll presentan elevada actividad unidora de 125"- -
hCG por célula (aproximadamente lo4 receptores/célula) , actividad
que no siempre se correlaciona c3n la capacidad de respuesta a una
estimulación qonadotrópica, medida ya sea en términos de la
producción de T o de AMPc (199 ,ZO:L) .
LOS resultados obtenidos en este trabajo señalan que la produccicn
kasal de T es de 6 . 9 7 k 0 . 4 6 ng/:L06 células/3h de acuerdo a lo
señalado en la tabla 6.4. Esta producción de T se encuentra en ei
yismo orden de rnaqnitud que reporta la mayoría de los autores
xencionados, inclusive para fracciones separadas a través de
gradientes de densidad, y es claramente semejante a l o s 5 . 4 2 2 . 0 n9
T / 1 0 6 células/3h reportado por Janszen y cols (146), cuya
netodologia fue la base para la utilizada durante este desarrollo
experimental.
LOS resultados que se mue'stran en la tabla 6.4 y la figura 6.1
indican que la vía preferencial de síntesis en las C L es la de los
" . i

72
llamados intermediarios 14 (Prog,, l7OHProg y A ) , como ha sido
reportado anteriormente (29) . Esta predominancia de la vía puede ser apreciada tanto en el caso de la determinación de las pozas
endógenas (tabla 6.4) como cuando se evalúa la transformación de
Frecursores radiactivos agregados excgenamente : T a b l a 6.5 y fig
6 . 4 ) .
Si bien existe una disminución aparente de la velocidad de
prOdUCCiÓn de los diversos interxediarios 5~ f ig 6.2) , las
concentraciones de todos los esteroides en las suspensiones
celulares continúan creciendo a lo larqo del 2eríodo de incubación,
lo que refiejaría una adecuada act:i.Jidad de las enzimas de la vía
involucradas, lo que permitiría a su vez suponer qae las células se
encuentran viables aún al final del período de incubación.
Resultados no incluidos anteriormente indican que la viabilidad
celular se Tantiene por encima del 8 0 % cuando se mantienen a las
células suspendidas en KRBG a 3 7 O C durante períodos hasta de 4
horas.
En términos de velocidades de transformación de los precursores
radiactivos, la vía biosintética parece encontrarse acelerada, al
menos en el segmento que abarca desde Preg hasta 170HProg; de ahí
en adelante la situación cambia, quizá debido a las características
de la 17a-hidroxilasa/C17,2O-liasa, que funciona como enzima
limitante de la vía (29) .

7 3
No se determinó como parte de este trabajo la presencia de
receptores a hCG en las fracciones enriquecidas en CL, por lo que
este parámetro no puede confrontarse con los resultados obtenidos
por otros grupos de trabajo. Sin embargo, una forma indirecta de
demostrar la existencia de un sistema funcional del receptor a LH
sería la capacidad de la hCG para estimular de forma dosis-
dependiente la producción de T en las fracciones celulares.
Como se obsertra en 13s figuras, dosis m6ximas de 3 U / n l de hCG son
capaces de estimular aproximadamente 15 veces la producción basal
de T medida c3mo r.7 T / 1 0 5 células/h (fig 6 . 3 ) , y de producir un
consumo total del '3-col añadido exógenamente en períodos máximos
de 90 minutos (fig G . 5 ) , el cual se reparte hacia la vía
esteroidogénica en forma equivalente a la que se presenta para las
pozas metabólicas de los intermediarios endógenos ( f i g 6.6). Esta
estimulación es dosis-dependiente en un intervalo de 3 x 3
U/ml (fiq 6 . 3 ) , y no parece aumentar a dosis mayores (resultados no
mostrados). Cabe mencionar que Janszen y cols (146) reportan que,
utilizando la netodología que sirvió de base para la manejada en
este trabajo, dosis de 1pg/ml de LH son capaces de estimular hasta
300% la producción de T / 1 0 6 células/2h.
-
Sharpe & Cooper (194) muestran una estimulación máxima de 900% en
el caso de CL purificadas tan solo mediante dispersión con
colagenasa, mientras que Cooke y cols (149) indican que la
población celular I11 de células de testículo de rata purificadas

1 2 6 3 8 7 7 4
a través de un qradiente de Percoil xkre incrementada 10 veces su
producción basal de T al ser tratadas las C L con una dosis máxima
de hCG. El factor de estimulación para las células purificadas a
través de gradientes de Metrizamida y mediante tratamientos
múltiples secuenciados se mantiene también dentro de este intervalo
(1,000-2,000%) (197,200-202).
7.3.2. Fracción tubular.
Estudios iniciales realizados por Christensen S( Mason (131)
utilizando túbulos seminiferos separados incompletamente del
compartimiento intersticial mediante técnicas de disección húmeda
demostraron que ambos compartimientos testiculares tienen la
capacidad de formar andróqenos i.n vitro a partir de la adición
exóqena de 14C-4-Proq,si bien las CL presentan una mayor capacidad
en este sentido.
Estudios posteriores (79,203) demostraron que la actividad
esteroidogénica de los tdbulos seminiferos aislados y en particular
de l a s CS se dirige fundamentalmente hacia la formación de
estrógenos y derivados Sa-reducidos tales como DHT a partir de 3H-T
adicionada exógenamente, aunque es también capaz de transformar
precursores 0 4 (Proq, l7OHProg y A ) más no 0 5 , debido a su carencia
de 3R-hidroxiesteroide deshidrogenasa (204).
La actividad de aromatasa es relativamente baja en CS aisladas de

7 5
ratas mayores de 20 días ce edad (205) , hasta llegar al punto en
que las C L concentran la mayor actividad de esta enzima en
testículos completos de animales adultos o en cultivos celulares
primarios (206) , lo que se ha su.gerido está relacionado con la
disminución de la capacidad mitótica <e las CS conforme el animal
va madurando (82) .
Dorrinqton y cols (205) reportan una producción máxima de E de
alrededor de 100 pg/mg de proteína/24h en cultivos no estimulados
de CS de animales de 20 días de edad, nientras que Rommerts y cols
(206) detectan una producción de 0.5C0.4 nu Preg/mg de proteína12h
y de 0.4110.21 nq E/mg de proteína/24h en fracciones enriquecidas
de animales de la misma edad, y Zna concentración de
aproximadamente 2 ng E/mg proteína/24h en fracciones enriquecidas
en CS de aninales adultos (70 días de edad) irradiados
prenatalmente para eliminar el componente germinal de sus túbulos
seminiferos.
Dorrinqton & Armstronq (79) demostraron originalmente la capacidad
de la FSH para estimular la producción de E a partir de T añadida
exógenamente en CS cultivadas in vitro, en concordancia con la
existencia de receptores específicos para esta hormona presentes en
las membranas celulares (207).
Rommerts y cols ( 8 0 ) , en cultivos de fracciones enriquecidas en CS
de ratas inmaduras, mostraron que tanto FSH como dbAMPc y T
c

7 6
añadidas a l o s medios de cultivo For 5-6 días, son capaces de
estimular la producción de E y de ABP hasta en 10 -leces con
respecto a los cultivos no tratados, siendo aparentemente no
sinérgico el efecto de FSH y T (206).
Los datos de producción de E por las fracciones enriquecidas en CS
reportados en esta tesis no pueden ser directamente comparados con
datos existentes en la literatura, ya que no se encuentran
reportadas las concentraciones producidas por células obtenidas de
ratas adultas normales durante períodos cortos de incubación, y la
única referencia para animales adultos sería el trabajo de Marcante
y cols ( 2 0 4 ) , el cual se enfoca má:s bien hacia la transformación de
los precursores adicionados exógenamente.
Los resultados obtenidos en cuanto a la capacidad de transformación
de l o s 3H-esteroides adicionados a fracciones enriquecidas en CS
(tabla 6.6) parecen avalar la no existencia de una actividad de 38-
hidroxiesteroide deshidrogenasa en estas células y por l o tanto la
ausencia de un contaminante apreciable de la fracción intersticial
del mismo testículo, ya que aproximadamente el 75% de la 3H-Preg
añadida no es transformada duranice períodos hasta de 60 minutos.
Debe señalarse que si bien el 25% de la 3H-Pregañadida inicialmente
sí es tranformada, no es posible detectar la radiactividad derivada
de ella en ninguna de las zonas cromatográficas correspondientes a
l o s c4-intermediariosr ni a los productos finales T, DHT o E, lo
que representaría su posible metabolización a través de vías

77
alternativas, tales c3mo la 20a-hidroxilación (63,208).
Cabe mencionar que los esteroides E y DHT no son metabolizados
durante la incubación, de acuerdo a su caracter de productos
sintéticos finales de la vía esteroidogénica en CS (datos no
mostrados).
En lo que toca a la estimulación de la producción de E de las CS
dependiente de FSH, los resulzados contradicen lo señalado
originalmente por Dorrinqton y cols (205), quienes demostraron que
CS de animales adultos eran incapaces de responder a la acción de
esta gonadotropina. Los resultados reportados (figs 6.7 y 6.8) en
este trabajo indican, sin embargo, que dicha estimulación existe al
menos para períodos cortos de incubación, si bien no al nivel
encontrado en testículos de animales inmaduros (206).
Aunque esta tesis no se avocó a la detección de la presencia de
receptores específicos de FSH en la fracción enriquecida de CS, los
resultados obtenidos permiten s'uponer de manera indirecta que
dichos receptores se encuentran presentes y funcionales en las CS
obtenidas bajo las condiciones reportadas en este trabajo
experimental.

8 . CONCLUSIONES.
2. Las fracciones celulares enriquecidas aparentemente no son
dañadas como resultado de su oktención, ya que presentan una
elevada viabilidad final, la cual se mantiene si se resuspenden en
amortiguadores adecuados durante períodos cortos.
3. Utilizando criterios morfológicos y fisio~ógicos, las fracciones
enriquecidas en CL y CS no se encuentran significativamente
contaminadas entre sí, aunque incluyen a otras estirpes celulares
propias del mismo compartimiento zesticular del cual se originan,
particularmente en el caso del compartimiento tubular.
4. La fracción enriquecida en CL presenta un funcionamiento normal,
tanto en condiciones basales como de estimulación con hCG, de las
vías esteroidoqénicas de síntesis de andróqenos, evaluadas tanto a
partir de las pozas metabólicas propias de las células, como
tomando en consideración la capacidad de transformación de
precursores e intermediarios añadiaos exóqenamente.

7 9
7. No es posible localizar componentes unidores de E en l a s
fracciones testiculares, aunque sí existan en citosoles de
hornogenizados de testículos completos.

80
9. BIBLIOGRAFIA.
2 . Fawcett DW. Tratado de Histología. , lla. Ed. , Ed.
Interamericana/McGraw-Hill, México, 1989, pp. 8 0 2 - 8 3 8 .
4. Grassé P-P (ed) . Traité de Zoologie. Anatomie, Systématique, Biologie. Tome XVI: Mammiferes. Mason & Cie, París, 1969, pp. 979-
1002.
6. Lesson ST & Lesson CR.
México, 1985, pp. 481-502 7
Histología, 4a.ed., Ed. Interamericana,
7 . Clermont Y . Kinetics of spermatogenesis in mammals: Seminiferous
epithelium Cycle and spermatogonial renewal. Physiol Rev 5 2 : 198-

81
236, 1972.
8. Leblond CP & Clermont Y. Definition of the stages of the cycle
of the seminiferous epithelium in the rat. Ann NY Acad Sci 5 5 : 5 4 8 -
5 7 3 , 1952.
9. Clermont Y. The cycle of the seminiferous epithelium in man.
Amer J Anat 1 1 2 : 35-51, 1963.
10. Huckins C . Cell cycle properties of differentiating
spermatogonia in adult Sprague-Dawley rats. Cell Tissue Kinet 3:
139-150, ;971.
11. Schulze C . On the morphology of the human Sertoli cell. Cell
Tissue Res 1 5 3 : 339-355, 1974.
12. Steinberger A & Steinberger E. The Sertoli cell. In: Johnson AD
& Gomes WR (ed) . The testis, vol IV, Academic Press, NY , 1977, pp.
371-399.
13. Fawcett DW. The ultrastructure and functions of the Sertoli
cell. In: Handbook of Physiology. Endocrinology, vol 5 , Sect 7:
Male Reproductive System, Washington, DC, American Physiological
Society, 1975, pp. 21-75.
14. Steinberger A & Steinberger E. Replication pattern of Sertoli
.." .

82
cells in maturing rat testis in vivo and in organ culture. Biol
Reprod 4: 84-97, i971.
15. Fawcett DW, Leak LV & Heidger PM. Electron microscope
observations on the structural components of the blood-testis
barrier. J Reprod Fertil Suppl 10: 105-122, 1970.
16. Iiussell LD & Peterson RN. Sertoli cell junctions: Morphological
and functional correlates. Int Rev Cytol 94: 177-211, 1985.
17. Dym M & Fawcett DW. The blood-t.estis barrier in the rat and the
physiological compartmentation of the seminiferous epithelium. Biol
Reprod 2: 3 0 8 - 3 2 6 , 1970.
18. Schulze W & Xehder U. Organization and morphogenesis of the
human seminiferous epithelium. Cel.1 Tissue Res 2 3 7 : 395-40’7, 1984.
19. Sun EL & Gondos B. Formation o:€ the blood-testis barrier in the
rabbit. Cell Tissue Res 2 4 3 : 575-578, 1986.
20. Buckland-Nicks S & Chia F-S. Fine structurz of Sertoli cells in
three marine snails with a discussion on the functional morphology
of Sertoli Cells in general. Cell. Tissue Res 245: 305-313, 1986.
21. Setchell BP, Pollanen P & Zupp JL. Development of the blood-
testis barrier and changes in vascular permeability at puberty in .

03
rats. Int J Androl 11: 225-233, 19138.
2 2 . Fritz IB. Sites of actions of androgens and follicle-
stimulating hormone on cells of the seminiferous tubule. In:
Litwack G (ed) . aiochemical actions cf hormones, vol 'i, Academic Press, NY, 1978, pp. 249-281.
23. Jutte NHPM, Jansen R, Grootegoed JA, Rommerts FFG & Van der
Molen HJ. FSH stimulation of the production of pyruvate and lactate
by rat Sertoli cells may be invo'lved in hormonal regulation of
spermatogenesis. J Reprod Fertil is: Z19-226, 1983.
2 4 . Connell CJ & Connell GM. The interstitial tissue of the testis.
In: Johnson AD & Gomes WR (eds) . The testis, vol IV, Academic Press, NY, 1977, pp.333-369.
25. Fawcett DW, Neaves W & Flores MN. Comparative observations on
intertubular lymphatics and the organization of the intesrstitial
tissue of the mammalian testis. Biol Xeprod 2: 500 -510 , 1973.
26. Nagano T & Suzuki F. Freeze-fracture observations on the
intercellulqr junctions of Sertoli cells and Leydig cells in the
human testis. Cell Tissue Res u!: 37-48, 1976. ' i .
27. Russell LD, Amlani SR, Voql AW & Weber JE. Characterization of
filaments within Leydig cells of the rat testis. Amer J Anat 178: .2

84
231-240, 1987.
28. Martin DW Jr, Mayes PA & Rodwell VW. Bioquímica de Harper, 9a. ed., Ed. El Manual Moderno, México, DF, 1984, pp.482-488.
29. Ewing LE & Brown BL. Testicular steroidogenesis. In: Johnson AD
& Gomes WR (eds) . The testis, vol I :V , Academic Press, N Y , 1977, pp.
239-287.
30. Rommerts FFG , Teerds K, Themnen APN & Van Noort M. Multiple
regulation of testicular steroidogenesis. J Steroid Eiochem 2 7 :
309-316, 1987.
31. Swerdloff RS & Walsh PC. Testosterone and estradiol suppression
of LH and FSH in adult male rats. Duration of castration, duration
of treatment and combined treatment. Acta Endocrino1 ,3: 11-18,
1973.
32. Chappel SC, Vlloa-Aguirre A 6: Ccucifaris C. Biosynthesis and
secretion of follicle-stimulating hormone. Endocrine Rev 4: 179-
211, 1983.
34. Catt KJ, Tsuruhara T, Mendelson C, Kietelslegers J-M & Dufau

85
ML. Gonadotropin binding and activation of the interstitial cells
of the testis. In: Dufau ML & Means AR (eds), Hormone binding and
target cell activation in the testis, Plenum Press, N Y , 1974, pp .
1-30.
35. Chaudhary LR & Stocco DM. Effect of different steroidogenic
stimuli on protein phosphorylati.on and steroidogenesis in MA-10
mouse Leydig tumor cells. Biochim Biophys Acta 1094: 175-184, 1991.
3 6 . Berridge MJ. Inositol triphosphate and diacylglycerol: Two
interacting second messengers. Annu Rev Biochem 5 6 : 159-193, 1987.
37. Exton JH, Taylor SJ, Auqert G & Socckino SB. Cell signaling
through phospholipid breakdown. Mol Cell Biochem m: 81-86, 1991.
38. Reithmann C , Gierschik P, Nerdan K & Jakobs KH. Role of
inhibitory G protein a-subunits in adenylyl cyclase
desensitization. Mol Cell Endocrinol 8 2 : C215-C221, 1991.
39. Hall PF, Osawa S & Mrotek J. The influence of calmodulin on
steroid synthesis in Leydig ce1:Ls from rat testis. Endocrinology
109: 1677-1682 , 1981.
40. Syed V, Khan SA & Nieschlag E. Epidermal growth factor
stimulates testosterone production of human Leydig cells in vitro. 1
J Endocrinol Invest 1 4 : 93-98, 1.991.

8 6
41. Sharpe RM & Fraser HM. Leydig cell receptors for LHRH(a) and
their modulation by administration or deprivation of the releasing
hormone. Biochem Biophys Res Commun 95: 256-262, 1980.
42. Bernier M, Chatelain P, Mather JP & Saez JM. Regulation of
gonadotropin receptors, gonadotropin responsiveness and cell
multiplication by somatomedin C an.d insulin in cultured pig Leydig
cells. J Cell Physiol m: 257-264, 1986.
43. Raeside JI, Berthelon M-C, Sanchez P & Saez JM. Stimulation of
aromatase activity in immature porcine Leydig cells by fibroblast
growth factor (FGF). Biochem Biophys Res Conmun m: 163-169, 1988.
44. Pointis G & Tahri-Joutei A. Controle intragonadique de la
fonction testiculaire par les peptides de type neurohypophysaire.
Ann Endocrino1 5 1 : 209-217, 1990.
45. Anakwe O 0 & Moger WH. Catecholamine stimulation of androgen
production by rat Leydig cells. Interactions with luteinizing
hormone and luteinizing hormone-releasing hormone. Biol Reprod 3 5 :
806-814, 1986.
46. Lieberman S, Greenfield NJ & Wolfson A . A heuristic proposal
for understanding steroidogenic processes. Endocrine Rev 5: 128-
1 4 6 , 1 9 8 4 .

48. Miller WL. Molecular biology of steroid hormone synthesis.
Endocrine Rev 9: 295-318, 1988.
49. Denton X4 Sr McCormack JC;. transport by mammalian
mitochondria and its roles in hormone action. Amer J Physiol m: E543-E554, 1985.
50. Payne AH. Hormonal regulation of cytochrome P450 enzymes,
cholesterol side-chain cleavage a:nd 17a-hydroxylase/C17-20 lyase in
Leydig cells. Biol Reprod u: 399-404, 1990.
52. Slaunwhite WR & Samuels LT. Progesterone as a precursor of
testicular androgens. J Biol Chem 2 2 0 : 341- 346, 1956.
53. Oertel GW & Eik-Nes KB. Toro pathways for the formation of
testosterone in vivo. In: Mowakowski H (ed) , Gewebs and
neurohormone, Springer-Verlag, E3erlin, 1962, p. 340.

88
of
dehydroepiandrosterone sulfate and androstenediol sulfate as
precursors of testicular androglens. Endocrinology 8 7 : 316-327,
1970.
54. Payne AH ;r: Jaffe RB . comparative roles
55. Fevold RH, Lorence MC, McCarthy JL, Trant JM, Kaqimoto M,
Waterman MR & Mason JI. Rat P450,,,1 from testis: Characterization of
a full-length cDNA encoding a unicpe steroid hydroxylase capable of
catalyzing both 6 4 - and 65-steroid-17,20-lyase reactions. :.lo1
Endocrinol 2: 958-975, 1989.
5G. Bogovich X & 'ayne AH. Purification of rat testicular
microsomal 17-ketosteroid reductase. Evidence that 17-ketosteroid
reductase and 17B-hydroxysteroid dehydrogenase are distinct
enzymes. J Biol Chen ,55: 5552-5559, 19so.
57. Lu Y, McDonough PA, Farley RA & Warren DW. Regulation of
testicular P-450 cholesterol sid.e-chain cleavage and P-450 C17-20
lyase/C17 hydroxylase enzymes in the neonatal and adult rat. Acta
Endocrinol 124: 449-454, 1991.
58. Warren DW, Huhtaniemi IT, Dufau ML & Catt KJ. Regulation of LH
receptors and steroidogenesis in the foetal rat testis in vivo. Acta Endocrinol m: 189-195, 1987.
59. Huhtaniemi I. Endocrine fun.ction and regulation of the fetal

89
and neonatal testis. Int J Dev Biol 3 3 : 117-123, 1989.
60. Means AR, Fakunding JL, Huckins C, Tindall DJ & Vitale R.
Follicle-stimulating hormone, the Sertoli cell and spermatogenesis.
Recent Prog Horn Res 3 2 : 477-527, 1976.
61. Orth JM. The role of follicle-stimulating hormone in
controlling Sertoli cell proliferation in testes of fetal rats.
Endocrinology 115: 1248-1255, 1984.
62. Fakundinq 3L & Means AR. Characterization and follicle
stimulating hormone activation of Sertoli cell cyclic AMP-dependent
protein kinases. Endocrinology 101: 1358-1368, 1977.
63. Griswold MD, Morales C & Sylvester SR. Molecular biology of the
Sertoli cell. Oxf Rev Reprod Biol. 10: 124-161, 1988.
64. Louis BG & Fritz IB. Follicle stimulating hormone and
testosterone independently increase ABP production by Sertoli cells
in culture. Endocrinology 104: 454-461, 1979.
65. Danzo BJ, pavlou SN & Anthony HL. Hormonal regulation of
androgen-binding protein in the rat. Endocrinology 127: 2829-2838,
1990.
66. Skinner MK & Griswold MD. Secretion of testicular transferrin

90
by cultured Sertoli cells is regulated by hormones and retinoids.
Biol Reprod 27: 211-221, 1982.
67. Pérez-Infante V, Bardin CW, Gunsalus GL, Musto NA, Rich KA &
Mather JP. Differential regulation of testicular transferrin and
androgen-binding protein secretion in primary cultures of rat
Sertoli cells. Endocrinology m: 383-391, 1986.
68. Lacroix M. Smith F & Fritz IB. The control of plasminogen
activator secretion by Sertoli cells in culture, and its possible
role in spermatogenesis. Horm Cell- Culture 6: 239-248, 1979.
69. Hettle JA, Walker EK & Fritz IB. Hormonal stimulation alters
the type of plasminogen activator produced by Sertoli cells. Biol
Reprod 3 4 : 895-904, 1986.
70. Morris PL, Vale WW, Cappel S & Bardin CW. Inhibin production by
primary Sertoli cell-enriched cultures: Regulation by follicle-
stimulating hormone, androgens and epidermal growth factor.
Endocrinology m: 717-725, 1988.
71. Grootenhuis AJ, Vanbeurden WMD, Timmerman MA & DeJong FH.
Follicle-stimulating hormone-stimulated secretion of an
immunoreactive 29-KDa inhibin-a-subunit complex, but not of 32-KDa
bioactive inhibin from cultured immature rat Sertoli cells. Mol
Cell Endocrino1 7 4 : 125-132, 1990.

9 1
7 2 . Cheng CY , Mather JP, Byer AL & Bardin CW. Identification Of
hormonally responsive proteins in primary Sertoli cell culture
medium by high performance liquid chromatography. Endocrinology
118: 480-488, 1986.
73. Smith EP, Dickson BA & Chernausek SD. Insulin-like growth
factor binding protein-3 secretion from cultured rat Sertoli cells.
Dual regulation by follicle stimulating hormone and insulin-like
growth factor-I. Endocrinology u[: 2744-2751, 1990.
74. Ya11 SH, Conti M, French FS & Joseph DR. Follicle-stimulating
hormone regulation of androgen-binding protein messenger RNA in
Sertoli Cell Cultures. No1 Endocrinol 2: 3:g-355, 1990.
75. Huqqenvik JI , Idzerda RL, Haywood L, Lee 3C, McKnight GS Sr
Griswold MD. Transferrin messenger ribonucleic acid: Molecular
cloning and hormonal regulation in rat Sertoli cells. Endocrinology
1 2 0 : 332-340, 1987.
76. Vihko KK, Penttila T-L, Parvinen M & Belin D. Regulation of
urokinase- and tissue-type plasmirlogen activator gene expression in
the rat seminiferous epithelium. Mol Endocrino1 3 : 52-59, 1989.
77. Dyrn M, Lamsam-Casalotti S, J i a M-C, Kleinman HK & Papadopoulos
V . Basement membrane increases G-protein levels and follicle-
stimulating hormone responsiveness of Sertoli cell adenylyl cyclase

92
activity. Endocrinology m: 1167--1176, 1991.
7 8 . Warren DW. Development of the! inhibitory guanine nucleotide-
binding regulatory protein in the :rat testis. Biol Reprod 40: 1208-
1214 , 1989.
79. Dorrinqton JH & Armstrong ET. Follicle-stimulating hormone
stimulates estradiol-17R synthesis in cultured Sertoli cells. Proc
Nat Acad S c i U S A 72: 6551-6555, 1975.
80. Rommerts FFG, Krüger-Sewnarain B Ch, Van Woerkom-Blik A,
Grootegoed JA & ['an der Molen HJ. Secretion of proteins by Sertoli
cell enriched cultures: Effects of follicle stimulating hormone,
dibutyryl CAMP and testosterone and correlation with secretion of
oestradiol and androgen binding protein. Mol Cell Endocrino1 10:
39-55 , 1978.
81. Pomerantz DK. Developmental changes in the ability of follicle
stimulating hormone to stimulate estrogen synthesis in vivo by the
testis of the rat. Biol Reprod 23: 948-954, 1 9 8 0 .
82. Payne AH, Perkins LM, Georgiou M & Quinn P G . Intratesticular
site of aromatase activity and possible function of testicular
estradiol. Steroids 5 0 : 4 3 5 - 4 4 8 , 1 9 8 7 .
8 3 . Robinson R & Fritz IB. Metabolism of glucose by Sertoli cells
. .

9 3
in culture. Biol Reprod 24: 1032-1-041, 1981.
85. Vivarelli E, Stefanini M Sr Siracusa G . Pyruvate and lactate
production by cultured Sertoli cells, fibroblasts and muscle
satellite cells, and the effect of hormonal and dcAMP stimulation.
Cell Biol Int Rep 8: 739-746, 1984.
87. Mallea LE, Machado AJ, Navarol F & Rommerts FFG. Epidermal
growth factor stimulates lactate production and inhibits
arOmatiZatiOn in cultured Sertoli cells from immature rats. Int J
Androl 9: 201-208, 1986.

91. Jaillard C , Chatelain PG & Sae'z JM. In vitro regulation of pig
Sertoli cell growth and function: Effects of fibroblast growth
factor and somatomedin-C. Bioi Reprod 3 7 : 665-674, 1987.
92. Saez JM, Perrard-Sapori MH, Chatelain PG, Tabone E & fiivarola
MA. Paracrine regulation of testicular function. J Steroid Biochem
27: 317-329, 1987.
93. Tres LL & Kierszenbaum AL. Viability of rat spermatogenic cells
- in vitro is facilitated by their coculture with Sertoli cells in
serum-free hormone-supplemented medium. Proc Nat Acad Sci USA 8 0 :
3377-3381, 1983.
94. Rivarola M A , Sanchez P & Saez JM. Stimulation of RNA and DNA
synthesis in spermatogenic cells by their coculture with Sertoli
cells. Endocrinology 117: 1796-1802, 1985.
95. McGinley DM, Posalaky z, Porvaznik M & Russell L. Gap junctions
between Sertoli and germ cells or rat seminiferous tubules. Tissue
Cell 11: 741-754, 1979.

95
cell communication. Ann NY Acad Sei 5 6 4 : 2 3 2 - 2 4 2 , 1989.
97. Skinner MK. Cell-cell interact,ions in the testis. Ann NY Acad
Sci 5 1 3 : 158-168, 1987.
98. Parvinen M. Regulation of the seminiferous epithelium. Endocr
Rev 3 : 404-417, 1982.
99. Grooteqoed JA, DenBoer PJ & Mac:kenbach P. Role of Sertoli cells
in spermatogenesis. In: Serio M (ed) . Ferspectives in Andrology.
Serono Symposia Publications frc,m Raven Press, vol. 53, Raven
Press, ??Y, 1989, pp. 167-173.
100. Grooteqoed JA, Oonk RB, [Tansen R & Van der Molen XJ.
Spermatogenic cells utilize metabolic intermediates from Sertoli
cells. Int Congr Ser Excerpta Med 6 5 8 : 225-237, 1985.
101. Wright W, Parvinen M, Nusto NA, Gunsalus GL, Phillips DM,
Mather JP & Bardin CW. Identif ic4ation of stage-specif ic proteins
synthesized by rat seminiferous tubules. Biol Reprod 2 3 : 257-270,
1983.
102. LeMaqueresse B & Jégou B. Possible involvement of germ cells
in the regulation of oestradiol-1~7B and ABP secretion by immature
rat Sertoli Cells ( I n vitro studies). Biochem Biophys Res Commun
m: 861-869, 1986.

9 6
103. Welsh MJ, Ireland ME & Treisman GJ. Stimulation of rat Sertoli
cell adenylate cyclase by germ cells in vitro. Biol Reprod 3 3 :
1050-1056, 1985.
104. LeMagueresse B, Pineau C, C-u.illou F & Jégou B. Influence of
germ cells upon transferrin.secretion by rat Sertoli cells in vitro. J Endocrino1 m: R13-Rl6, 1988.
105. Wright WW, Zabludoff SD, Erickson-Lawrence M & Karzai AW. Germ
cell-Sertoli cell interactions. An:? NY Acad Sci 5 6 4 : 173-185, 1989.
106. Tung PS & F r i t z IB. Interaction of Sertoli cells with myoid
cells ~.IJ vitro. Biol Reprod 2 3 : 207-217, 1980.
107. Tung PS & Frl’iz IB. Extracellular matrix promotes rat Sertoli
cell histotypic expression in vitro. Biol Reprod 3 0 : 2 1 3 - 2 2 9 , 1984.
109. Skinner MK, Tung PS & Fritz IB. Cooperativity between Sertoli
cells and testicular peritubular cells in the production and
desposition of extracellular matrix components. J Cell Biol m: 1941-1947, 1985.

97
110. Skinner MK &. Fritz IB. Testicular peritubular cells secrete a
protein under androgen control that modulates Sertoli cell
function. Proc Nat Acad Sci USA 82: 114-118, 1985.
111. Skinner MK & Fritz IB. Identification of a non-mitogenic
paracrine factor involved in i.nteractions between testicular
peritubular cells and Sertoli cells. Mol Cell Endocrinol 44: 85-97,
1986.
112. Skinner MK, McLachlan RI & Blremner WJ. Stimulation of Sertoli
cell inhibin secretion by the testicular paracrine factor PModS.
Mol Cell Endocrinol E: 239-249, 1989.
113. Norton J & Skinner MK. Regulaltion of Sertoli cell function and
differentiation through the actions of a testicular paracrine
factor, P-Mod-S. Endocrinology x%: 2711-2718, 1989.
114. Skinner MK. Mesenchymal (Stromal) -epithelial cell interactions
in the testis and ovary which regulate gonadal function. Reprod
Fertil Dev 2: 237-243, 1990.
115. Nakhla AM, Mather JP, Janne OA & Bardin CW. Estrogen and
androgen receptors in Sertoli, Leydig, myoid and epithelial cells:
Effects Of time in culture and cell density. Endocrinology 115:
121-128, 1984.
.- -

98
116. Sanborn BM, Lamb DJ, Tsai YH & Steinberger A. Androgen action
in the Sertoli cell. In: Mahesh S , Muldoon V I Saxena S & Sadler T
(eds) . Functional correlates of hormone receptors in reproduction, Elsevier North Holland, Amsterdam, 1980, pp. 205-220.
118. Syed V I Karpe P, Ploen I, & Ritzen EM. Regulation of
interstitial cell function by sem,iniferous tubules in intact and
cryptorchid rats. Int J Androl 9: 271-278, 1986.

9 9
122. Verhoeven G & Cailleau 2. Specificity and partial purification
of a factor in spent media from Sertoli cell-enriched cultures that
stimulate steroidogenesis in Leydig cells. J Steroid Biochem 2 5 :
393-402, 1986.
123. Syed :I , Khan SA, Lindh M & Ritzén EM. Mechanism of action of
the factor(s) secreted by rat seminiferous tubules and inhibiting
interstitial cell testosterone production in vitro. Acta Endocrinol 1 1 9 : 427-434, 1988.
124. Syed ‘ I , Khan SA & Ritzén E:M. Stage-specific inhibition of
interstitial cell testosterone secretion by rat seminiferous
tubules in vitro. Mol Cell Endocrinol S: 257-264, 1985.
125. Perrard-Sapori MH, Chatelain P & Saez JM. Modulation of Leydig
cell function coculture with pig Sertoli cell conditioned
medium:Effects of insulin, somatomedin-C and FSH. Mol Cell
Endocrinol 5 0 : 193-201, 1987.
126. Janecki A, Jakubowiak A & Lukaszyk A . Stimulatory effect of
Sertoli cell secretory products; on testosterone secretion by
purified Leydig cells in primary culture. Mol Cell Endocrinol 42:
235-243 , 1985.
127. Bartlett JMS & :Sharpe M. Effect of local heating of the rat
testis on the levels in interstitial fluid of a putative paracrine

1 0 0
regulator of the Leydig cells and its relationship to changes in
Sertoli cell secretory function. J Reprod Fertil 8 0 : 279-287, 1987.
128. Lin T, Harrington Calkins J, Morris PL, Vale W & Bardin CW.
Regulation of Leydig cell function in primary culture by inhibin
and activin. Endocrinology 125: 2134-2140, 1989.
129. Boujrad PJ, Fapadopoulos V , Drosdowsky MA 5. Carreau S . R61e
d’une protéine Sertolienne purifiile (CMB-21) dans la biosynthese de
testostérone Leydigienne chez le rat immature. Path Biol E: 819-
823 , 1989.
130. Grootenhuis AJ, Melsert R, Timmerman A, Hooqerbruqqe JW,
Rommerts FFG & DeJonq FH. Short-term stimulatory effect of Sertoli
cell conditioned medium on Leydig cell steroidogenesis is not
mediated by inhibin. J Steroid Biochem 3 6 : 445-449, 1990.
131. Christensen AK & Mason NR. Comparative ability of seminiferous
tubules and interstitial tissue of rat testes to synthesize
androgens from proge~terone-4-~~C!h vitro. Endocrinology 76: 646-
656, 1965.
132. Steinberger A, Heindel JJ, Lindsey JN, Elkington JH, Sanborn
BM & Steineberger E. Isolation and culture of FSH responsive
Sertoli cells. Endocr Res Commun 2: 261-272, 1975.

101
133. Dorrinqton JH, Roller NF & Fritz IB. Effects of follicle-
stimulating hormone on cultures o:€ Sertoli cell preparations. Mol
Cell Endocrinol 3 : 5 7 - 7 0 , 1975.
134. Welsh MJ & Wiebe JP. Rat Sertoli cells: A rapid method for
obtaining viable cells. Endocrinology x: 618-624, 1975.
135. Wilson RM & Griswold MD. Secreted proteins from rat Sertoli
cells. Exp Cell Res m: 127-135, 1979.
136. Mather JP & Phillips DM. Primary culture of testicular somatic
cells. In: Barnes DN, Sirbasku DB & Sato GH (eds) . Methods for serum-free culture of cells of the endocrine system. Cell culture
methods for molecular biology and cell biology, vol. 2, Alan R.
L i s s , NY, 1984, pp. 25-45.
137. Renier G, Gaulin J, Gibb W , Simard P, Leung TK, Collu R &
Ducharme JR. Isolation, purification and culture of Sertoli Cells
from immature piglet testes. Acta Endocrinol 111: 411-418, 1986.
138. Russell LD, Bartke A & Goh J. Postnatal development of the
Sertoli cell barrier, tubular lumen, and cytoskeleton of Sertoli
and myoid cells in the rat, and their relationship to tubular fluid
secretion and flow. Amer J Anat 2 8 4 : 179-189, 1989.
139. Heller RH & Jones HW. Production of ovarian dysgenesis in the

102
rat and human by busulphan. Amer J Obstet Gynec u: 414-420, 1964.
140. Rich KA & DeKretser DM. E:ffect of differing degrees of
destruction of the rat seminiferous epithelium on levels of serum
follicle stimulating hormone amd androgen binding protein.
Endocrinology 101: 959-968, 1977.
141. Fakundinq JL, Tindal DJ, Dedman JR, Mena DR & Means AR.
Biochemical actions of follicle-stimulating hormone in the Sertoli
cell of the rat testis. Endocrinology 98: 392-402, 1976.
142. Gupta D, Rager K, Zarzycki U & Eichner M. Levels of
luteinizing hormone, follicle-stimulating hormone, testosterone and
dihydrotestosterone in the circulation of sexually maturing intact
male rats and after orchidectomy and experimental bilateral
cryptorchidism. J Endocrino1 6 6 : 183-193, 1975.
143. Collins P & Lacy D. Studies on the structure and function of
the mammalian testis. 11. Cytological and histochemical
observations on the testis of the rat after a single exposure to
heat applied for different lengths of time. Proc Roy SOC Ser B 1 7 2 :
17-38 , 1969.
1 4 4 . Dierickx P & Verhoeven G . Effect of different methods of
germinal Cell destruction on rat testis. J Reprod Fertil 2: 5-9, 1980.

103
145. Moyle WR & Ramachandran J. Effect of LH on steroidogenesis and
cyclic AMP accumulation in rat Leydig cell preparations and mouse
tumor Leydig cells. Endocrinology 93: 127-134, 1973.
146. Janszen FHA, Cooke EA, Van Driel MZA & ':an der Xolen HZ.
Purification and characterization of Leydig cells from rat testis.
J Endocrinol 70: 345-359, 1976.
147. Gale JS, Wakefield JSJ & Ford HC. Isolation of rat Leydig
cells by density gradient centrifugation. J Endocrinol 9 2 : 293-302,
1982.
148. Conn M , Tsuruhara T, Dufau P.'[ & Catt KJ. Isolation of highly
purified Leydig cells by density gradient centrifugation.
Endocrinoloqy m: 639-642, 1977.
150. Russell LD & Steinberger A. Sertoli cells in culture: Views
from the perspectives of an in yivoist and an in vitroist. Biol Reprod 41: 571-577, 1989.
151. Kruse .PF & Patterson MK. Tissue culture: Methods and

1 0 4
applications. Academic Press, NY, 1973, pp. 268-273
152. Scatchard G . The attractions of proteins for small molecules
and ions. Ann N Y Acad Sci 5 1 : 660-672, 1949.
153. Bermúdez JA, León C & Herrera J. Fundamento y estudio
comparativo de dos métodos de análisis por saturación. Rev Méd IMSS
1 2 : 11-22, 1973.
154. Lee-Benítez D, León C, Herrera J, González-Bárcena D &
Bermúdez JA. Métodos de análisis por saturación para cuantificar
cortisol y testosterona. Arch Inv Méd 5 : 7-16, 1973.
155. Herrera 3 , Morán L, Rauda.les L, León C & Bermúdez JA.
Radioinmunoanálisis de pregnenolona, 17-hidroxipregnenolona y
dehidroepiandrosterona y sus formas sulfoconjugadas. Arch Invest
Méd 5 : 617-630, 1974.
156. Bermúdez JA, Coronado V, Mijares A, León C, Velázquez A, Noble
P b Mateos JL. Stereochemical approach to increase the specificity
of steroid antibodies. J Steroid Biochem 6: 283-290, 1975.
157. Podestá EJ & Rivarola MA. Concentration of androgens in whole
testis, seminiferous tubules and interstitial tissue of rats at
different stages of development. Endocrinology 95: 455-464, 1974.

105
158. Hadley M A , Byers SW, Suárez-Quián CA, Kleinman HK & Dym M.
Extracellular matrix regulates Sertoli cell differentiation,
testicular cord formation, and ge!rm cell development in vitro. J Cell Biol m: 1511-1522, 1985.
159. Caston LA & Sanborn BM. Regulation of testicular and Sertoli
cell gamma-glutamyltranspeptidase by follicle-stimulating hormone.
Biol Reprod 3 8 : 109-113, 1988.
160. Galdieri M, Zani B b Stefanini M. Effect of the association
with germ cells on the secretory activities of Sertoli cells in in
vitro cultures. In: Frajese G , Conti C, Hafez ESE & Fabbrini A
(eds). Oligozoospermia: Recent progress in andrology, Raven Press,
NY, 1981, PP. 95-102.
161. Aquilano DR & Dufau ML. Functional and morphological studies
on isolated Leydig cells: Purification by centrifugal elutriation
and Metrizamide fractionation. Endocrinology m: 499-510, 1984.
162. Wakefield JSJ, Gale JS, Berridge M V , Jordan TW & Ford HC. Is
Percoll innocuous to cells? Bioch.em J 2 6 2 : 795-797, 1982.
163. Laws AO, Wreford NGM & EleKretser DM. Morphological and
functional characteristics of rat Leydig cells isolated on Percoll
gradients: Is Leydig cell heteroqreneity in vitro an artifact? Mol Cell Endocrino1 42: 73-90, 1985.
a L

1 0 6
164. Hall SH, Conti M, French FS & Joseph DR. Follicle-stimulating
hormone regulation of androgen-binding protein messenger RNA in
Sertoli cell cultures. Mol Endocrinol 4: 349-355, 1990.
165. Sharpe FW & Cooper I. Variation in the steroidogenic
responsiveness of isolated rat Lebydig cells. J Reprod Fertil 6 5 :
475-481 , 1982.
166. Mori H & Christensen AK. Morphometric analysis of Leydig cells
in the normal rat testis. J Cell Biol 8 4 : 340-354, 1980.
167. Mendelson C, Dufau M & Catt I(. Gonadotropin binding and
stimulation of cyclic adenosine 3':5'-monophosphate and
testosterone production in isolated Leydig cells. J Biol Chem 2 5 0 :
8818-8823, 1975.
168. Hansson V, McLean SW, Smith A A , Tindall DJ, Weddington SC,
Nayfeh SN & French FA. Androgen receptors in rat testis. Steroids
- 23: 823-832, 1974.
169. Vernon RG, Kopec B & Fritz I E I . Observations on the binding of
androgens by rat testis seminiferous tubules and testis extracts.
Mol Cell Endocrinol 1: 167-187,1974.
170. Ritzén EM, Dobbins MC, Tinclall DJ, French FS & Nayfeh SN.
Characterization of an androgen binding protein (ABP) in rat testis

107
and epididymis. Steroids 2 1 : 593-607, 1973.
171. Wilson EM & French FS. I3inding properties of androgen
receptors. Evidence for identical receptors in rat testis,
epididymis and prostate. ; Biol Chem 2 5 1 : 5620-5629, 1376.
173. Monet-Kuntz C, Terqui M & Locatelli A. Characterization of a
cytoplasmic androgen receptor in the ram testis. Mol Cell
Endocrino1 16: 57-70, 1979.
174. Cunningham GR, Tindall DJ & Means AR. Differences in steroid
specificity for rat androgen binding protein and the cytoplasmic
receptor. Steroids 3 3 : 261-276, 1979.
175. Sanborn BM, Lamb DJ, Tsai YH & Steinberqer A . Androgen action
in the Sertoli cell. In: Manesh, Muldoon, Saxena & Sadler (eds) . Functional correlates of hormcne receptors in reproduction,
Elsevier North Holland, Amsterdam, 1980, pp. 2 0 5 - 2 2 0 . OJO CAMBIO
176. Tindall DJ, Miller DA & Means AR. Characterization of androgen
receptor in Sertoli cell-enriched testis. Endocrinology 101: 13-23,

108
1977.
177. Sanborn BM, Steinberger A, Tcholakian RK & Steinberqer E.
Direct measurement of androgen receptors in cultured Sertoli cells.
Steroids 2 9 : 493-502, 1977.
178. Tsai YH, Sanborn BM, Steinberger A & Steinberqer E. The
interaction Of testicular androgen-receptor complex with rat germ
cell and Sertoli cell chromatin. Biochem Biophys Res Commun 75:
3 6 6 - 3 7 2 , i977.
179. Sanborn BM, Tsai YH, Steinnerger A & Steinberger E.
Biochemical aspects of the interaction of androgens with Sertoli
cells. Ann Biol Anim Biochim Biophys 18: 615-622, 1978.
180. Schmidt WN & Danzo BJ. Androgen and progesterone binding
components in cytosol prepared from cultures enriched in Sertoli
cells from immature rat testes. Eiiol Reprod 2 3 : 495-506, 1980.
181. Ortlip SA, Li SA & Li JJ. Characterization of specific
glucocorticoid receptor in the Syrian hamster testis. Endocrinology
m: 1331-1338, 1981.
182. Abney TO. A cytoplasmic estradiol receptor in rat testicular
tissue. Endocrinology 9 9 : 5 5 5 - 5 6 6 , 1976.

1 0 9
183. Abney TO & Melner MH. Characterization of estrogen binding in
the developing rat testis ontogeny of the testicular cytoplasmic
estrogen receptor. Steroids 3 4 : 413-427, 1979.
184. Lin T, Chen CCC, Xurono EP, Osterman J & Nankin HR. Estradiol
receptors of two distinct populations of Leydig cells. Steroids 40:
53-63, 1982.
185. Nozu K, Dufau ML & Catt KJ. Estradiol receptor-mediated
regulation of steroidogenesis in gonadotropin-desensitized Leydig
cells. J Biol Chem 2 5 6 : 1915-1922, 1981.
186. Dickson RB & Clark CR. Estrogen receptors in the male. Arch
Androl z: 2 0 5 - 2 1 7 , 1981.
187. Mulder E, Brinkmann AO, Lamers-Stahlhofen GJM & Tian Der Molen
HJ. Binding of oestradiol by the nuclear fraction of rat testis
interstitial tissue. FEBS Lett 31: 131-136, 1973.
188. Van Beurden-Lamers WMO, Brinkmann AO, Mulder E & Van der Molen
HJ. High-affinity binding of oestradiol-17D by cytosols from testis
interstitial tissue, pituitary, adrenal, liver and accesory sex
glands of the male rat. Biochem J m: 495-502, 1974.
189. DeBoer W , DeVries J, Mulder E & Van der Molen HJ. Comparative ..
study of nuclear binding sites for estradiol in rat testicular and
. u

110
uterine tissue. Biochem J 1 6 2 : 331-339, 1977.
190. Lin T, Chen GCC, Murono EP, Osterman J & Nankin HR. The aging
Leydig cell: VII: Cytoplasmic estradiol receptors. Steroids 3 8 :
407-415, 1981.
191. Onoda M & Hall PF. Inhibition of testicular microsomal
cytochrome P-450 (17-hydroxylas~e/C17,20 lyase) by estrogens.
Endocrinology m: 763-770, 1981.
192. Damber J-E, 3ergh A, Daehlin L, Ekholrn C, Selstam G &
Soderqard R. The acute effect of oestrogens on testosterone
production appears not to be mediated by testicular oestrogen
receptors. Mol Cell Endocrino1 3 1 : 105-116, 1983.
193. Wilson EM, Lea OA & French FS. 9s binding protein for
androgens and progesterone. Proc Nat Acad Sci USA a: 1960-1964, 1977.
194. Sharpe RM & Cooper 1. Variation in the steroidogenic
responsiveness of isolated rat Leydig cells. J Reprod Fertil 6 5 :
475-481, 1982.
195. Payne AH, Downing JR & Wong Km-L. Luteinizing hormone receptors
and testosterone synthesis in two distinct populations of Leydig
cells. Endocrinology m: 1424-1429, 1980.

111
196. Kerr JB, Robertson DM & CeKretser DM. Morphological and
functional characterization of interstitial cells from mouse testes
fractionated on Percoll density gradients. Endocrinology 116: 1030-
1043, 1985.
197. O'Shaughnessy PJ, Wong K-L & Payne AH. Differential
steroidogenic enzyme activities in different populations of rat
Leydig cells. Endocrinology 1 0 9 : 1061-1066, 1981.
198. Bhalla VK, Rajan VP, Burgett AC & Sohal G S . Interstitial cell
heterogeneity in rat testes. I,, Purification of collagenase-
dispersed Leydig. cells by unit gravity sedimentatiion and
demonstration of binding sites for gonadotropin in light cells
versus enhanced steroidogenesis in heavier cells. J Biol Chem 2 6 2 :
5313-5321, 1987.
199. Browne ES, Flasch M V , SohaiL GS & Bhalla VK. Gonadotropin
receptor occupancy and stimulation of CAMP and testosterone
production by purified Leydig cells: Critical dependence on cell
concentration. Mol Cell Endocrinol 7 0 : 49-63, 1990.
200. Bhalla VK, Flasch M V , Browlne ES, Sohal GS & Sharawy MM. Interstitial cell heterogeneity in rat testes. 11: Purification of
cells by Percoll and Metr.izamide gradient centrifugation with
preferential localization of gonadotropin binding sites in light >
cell fraction and hormone-induced steroidogenesis in heavier cell

112
fraction. J Biol Chem 2 6 2 : 5322-5332, 1987.
201. Browne ES, Sohal GS & Bhalla VK. Characterization of
functional Leydig cells after purification on a continuous gradient
of Percoll. J A n u r o 1 11: 379-389, 1990.
202. Klinefelter GR, Hall PF & Elwing LL. Effect of luteinizing
hormone deprivation in situ on steroidogenesis of rat Leydig cells
purified by a multistep procedure, Biol Reprod 3 5 : 7 6 9 - 7 8 3 , 1987.
203. Payne AH, Kawano .A & Jaf fe RB. Formation of
dihydrotestosterone and other Sa-reduced metabolites by isolated
seminiferous tubules and suspension of interstitial cells in a
human testis. J Clin Endocrino1 Metab 3 7 : 448-454, 1973.
204. Marcante ML, Floridi A, Natali PG, Citro G , Cioli V & De
Martino C. Sertoli cells of adult rats in vitro. 11. Effect of
different steroid precursors on estradiol 17R-synthesis.
Experientia 3 5 : 563-565, 1979.
205. Dorrinqton JH, Fritz IB & Armstrong DT. Control of testicular
estrogen synthesis. Biol Reprod 1~: 55-64, 1978.
206. Rommerts FFG, De Jong FH, Brinkmann A 0 & Van der Molen HJ.
Development and cellular localization of rat testicular aromatase
activity. J Reprod Fertil 65: 281-288, 1982.

113
207. Ritzén EM, Hansson V & French FS. The Sertoli cell. In: Burger
H & De Kretser D (eds). The Testis, Raven Press, NY, 1981, pp. 171-
194.

EL JURADO DESIGNADO POR LA DIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD DE LA UNIVERSIDAD AUTONOMA METROPOLITANA, UNIDAD IZTAPALAPA, APROBO LA PRESENTE TIlSIS A LOS DIAS
DEL :.!ES DE DICIEMBRE DEL ARO DE MIL :;OVECIENTOS XOVENTA Y DOS.
-"-- DR. JOSE A URO BEW4UDEZ GOMEZ-LLANOS
AQUIN FERNANDO HERRERA MUÑOZ
FU Q. CANA ELENA LEMUS BRAVO


















