![I. REACCIONES SIGMATROPICAS - Yolanda Rios · Transposicion de Claisen Rearreglo de Cope Rearreglo Claisen-Cope sigmatropicas [3,3] con atomos de N o P en el anillo. Rearreglo de](https://static.fdocuments.ec/doc/165x107/5bb9c4e809d3f2fd488cc098/i-reacciones-sigmatropicas-yolanda-transposicion-de-claisen-rearreglo-de.jpg)
UNIVERSIDAD AUTONOMA METROPOLITAN4 UNIDAD …148.206.53.84/tesiuami/UAM3737.pdf · Constituida por...
31
UNIVERSIDAD AUTONOMA METROPOLITAN4 UNIDAD IZTAPALAPA CIENCIAS BIOLOGICAS Y DE LA SALUD DETEILWINACION DE HEMOGLOBINA GLICADA (HbAk ENRATAS CONDIABETES INDUCIDA ___~ CON ESTREPTOZOTOCINA TESINA PROFESIONAL para obtener el título de LICENCIATURA EN BIOLOGIA EXPEMMEAJTAL Realizada en la UNIDAD DE IN\?ESTIGACIOX ESPERlXlESTAL INSTITUTO NACIONAL DE LA NUTRICION SALVADOR ZUBIR4N Presenta &MA LILIA ROSALES HERNANDEZ Mkxico, D.F., 1994.
Transcript of UNIVERSIDAD AUTONOMA METROPOLITAN4 UNIDAD …148.206.53.84/tesiuami/UAM3737.pdf · Constituida por...

UNIVERSIDAD AUTONOMA METROPOLITAN4 UNIDAD IZTAPALAPA
CIENCIAS BIOLOGICAS Y DE LA SALUD
DETEILWINACION DE HEMOGLOBINA GLICADA (HbAk ENRATAS CONDIABETES INDUCIDA
_ _ _ ~ CON ESTREPTOZOTOCINA
TESINA PROFESIONAL para obtener el título de LICENCIATURA EN BIOLOGIA EXPEMMEAJTAL
Realizada en la
UNIDAD DE IN\?ESTIGACIOX ESPERlXlESTAL INSTITUTO NACIONAL DE LA NUTRICION
SALVADOR ZUBIR4N
P r e s e n t a
&MA LILIA ROSALES HERNANDEZ
Mkxico, D.F., 1994.

Con amor especial a mis padres por su contprensih y paciencia incondicional
La abundancia de salud, fe, amor y espercsnza sean devueltos a todas las persolras que vivieron y apoyaron mi reto
A mi amiga Claudia por compartir su linda y sincera fornta de ser

La determinación de hemoglobina glicada (HbA,) es de gran utilidad en el control metabdico a largo plazo del paciente diabético.
La hemoglobina es una proteína globular citoplasm5tica que constituye aproximadamente el 90% del peso seco de un eritrocito maduro, está Constituida por cuatro cadenas polipeptídkas, dos (a) y dos (p), unidas a un grupo heme.
En los humanos existen 3 tipos principales de hemoglobinas: la hemoglobina del adulto, HbA [2a,2p], que constituye aproximadamen- te el 97% de la hemoglobina total; la HbA, [26,2P], corresponde al 2.5% y la HbF [2a,2y] menos del 1 % (1).
La HbA est5 expuesta a los carbohidratos circulantes en la sangre, los cuales al unirse a ésta constituyen la hemoglobina glicada o HbA,, an- teriormente llamada glicosilada (2). En individuos no diabéticos se tie- ne 4 4 % HbA, (4). Los carbohidratos que se unen a fa tibA, son, entre otros, 1,6-difosfato de fructosa (HbAIal), C-fostato de glucosa (HbA,,,), glucosa (HbAJ (3,4); y lo hacen al amim terminal de la valina de la cadena beta de la hemoglobina, rnediant? una reacción no enzimática reversible que da lugar primeramente a ura Base de Schiff, que por un rearreglo de Amadori genera la estructlxa estable conocida como cetoamina (fig.1) (3,4,5).
O C "H H - C = N - P r 3teína C% - N - - P r o t e í n a I I ?
H-&"H H - k - 0 ~ L O Proteína-m, + OH-+-H 7 OH-A-H .- > O H - ~ "
H-C-OH H-C-OH €¡-¿!-OH de Amadori H-b-OH
I H-&"H Rrurreglo H-b-OH
h% " O H AH2 -GX 6% " O H
Figura 1. Representaci6n del aducto de glucosa al amino terminal de la cadena beta de la hemoglobina para formar WbA,,.
2

La hemoglobina glicada o HbA, está constituida por las hemoglobinas Alal, Ala2, A,, y A,c, las cuales son producto de una transformación no enzimática y postsintética de la HbA. Cuando se mide colectivamente como una sola fracción HbAlal+,a2+,b+,c o HbA, se denomina hemoglobina glicada y cuando se mide HbA,c es hemoglobina glucosilada (43).
El amino terminal de la cadena beta no es el Único sitio de glicación, sino también en el amino terminal de la cadena alfa, pero en menor proporción, tanto in vivo como in vitro. Varias lisinas con grupos amino en posición epsilon de las cadenas a y j3 de la hemoglobina, reaccio- nan con la glucosa.
El pK de cualquier grupo amino es el determinante de reactividad con la glucosa, pero sólo algunos grupos estarán accesibles a la glucosa libre, según la estructura conforrnacional de la hemoglobina. El orden t 5:
de reactividad de los grupos amino de la hemoglobina in vivo parece 5 ser p-Val-1, p-Lys-66, a-Lys-61, P-Lys-I 7, u-Val-l , a-Lys-40, p-Lys-8 y g; P-LYs- 144 (8). z
m El porcentaje de glicación de la hemoglobina está en función de las E v' concentraciones de carbohidratos circulantes en sangre y de la vida 9 2
!=E
media del eritrocito que es de 90-120 días en el humano (1,4,6) y de T"? . aproximadsrnente 45-50 dias en la rata (7).
En animales en los cuales la vida media del eritrocito sea menor com- parada con la del humano, el porcentaje de HbA, es menor. En babuinos no diabéticos se ha reportado que tienen la mitad del porcentaje de HbA,c que el humano, debido posiblemente a la vida media más corta del eritrocito, 30 a 60 dias (9,14).
En un estudio en ratones donde se sometieron eritrocitos a un gradien- te de densidad para separar los glóbulos jóvenes de los maduros, se observó que los más jóvenes tienen valores más bajos de HbAlc que las células rojas maduras. De esta forma, el tiempo de exposición a íos carbohidratos circulantes afecta los valores de HbA lo que debe ser' considerado en el caso de enfermedades hernatolÓ$kxs (1 O).
8 8
/
3

La Diabetes Melli.tus es una enfermedad caracterizada por una con- centración anormalmente elevada de glucosa en sangre, debido a la falta o resistencia a la insulina, la cual es secretada por las células beta de los islotes pancreáticos.
Los síntomas y signos clásicos de la diabetes insulino dependiente o no insulino dependiente, entre otros, son polifagia, poliuria y polidipsia. Un mecanismo para eliminar el exceso de glucosa en sangre es excretándola por la orina. Esta glucosuria se acompaña de diuresis osmótica, con pérdida excesiva de agua, lo que cierra el circulo del síndrome al causar poliuria y más tarde polidipsia en los diabéticos.
- 2 z z y j - j
Como la glucosa no puede ser utilizada por las células, sea por la falta o resistencia a la insulina, se produce un incremento en el catabolismo de proteínas y grasas. Los aminoácidos pasan de la sangre al hígado, sufren desaminación oxidativa y sus cadenas carbonadas contribuyen a la forrnación de nuwa gluccsa (gluconeogénesis).
Los cuerpos cetónicos, en ocasiones presentes en la orina de los dia- béticos, se originan de la degradación de los ácidos grasos libres del hígado para obtener fuentes hidrocarbonadas que produzcan energía. Hay pérdida global de nitrógeno, potasio y de otros iones intracelulares (11).
En pacientes diab6:icos con mal control, la glucosa sanguínea se en- cuentra elevada, favoreciendo su unión a las proteínas como es el caso de las hemoglobinas. El porcentaje de hemoglobina glicada (o hemoglobinas rápidas debido a sus movilidades electroforéticas), se incrementan con respecto a los individuos sanos (3,4,6). En ratas dia- béticas no tratadas se presenta este mismo proceso, los valores de HbA, están aumentados en relación a los controles (12,13,14).
Hay por lo rnenos 6 tipos diferentes de hemoglobinas en las ratas (1 5). De un cromatsgrama donde se observan seis picos de elución con un sistema de WPLC, denominados por las letras “a” a la I‘ f ‘ I , las fraccio-. nes a+b+c están incrementadas en las ratas diabéticas comparadas con las controles (E!), (fig. 2).
4

La Diabetes Mellitus es una enfermedad caracterizada por una con- centración anormalmente elevada de glucosa en sangre, debido a la falta o resistencia a la insulina, la cual es secretada por las células beta de los islotes pancreáticos.
Los síntomas y signos clásicos de la diabetes insulino dependiente o no insulino dependiente, entre otros, son polifagia, poliuria y polidipsia. UR mecanismo para eliminar el exceso de glucosa en sangre es excretándola por la orina. Esta glucosuria se acompaña de diuresis osmótica, con pérdida excesiva de agua, lo que cierra el círculo del síndrome al causar poliuria y m& tarde polidipsia en los diabéticos.
Como la glucosa no puede ser utilizada por las células, sea por la falta o resistencia a la insulina, se produce un incremento en el catabolismo de proteínas y grasas. Los aminoácidos pasan de la sangre al hígado, sufren desaminación oxidativa y sus cadenas carbonadas contribuyen a la formacibr: de nueva glucosa (gluconeogkcesis).
Los cuerpos cetónicos, en ocasiones presentes en la orina de los dia- béticos, se originan de la degradación de los &idos grasos libres del higado para obtener fuentes hidrocarbonadas que produzcan energía. Hay pérdida global de nitrógeno, potasio y de otros iones intracelulares (1 1).
En pacientes diabéticos con mal control, la glucosa sanguínea se en- cuentra elevada, favoreciendo su unión a las proteínas como es el caso de las hemoglobinas. El porcentaje de hemoglobina glicada (o hemoglobinas rápidas debido a sus movilidades electroforéticas), se incrementan con respecto a los individuos sanos (3,4,6). En ratas dia- béticas no tratadas se presenta este mismo proceso, los valores de HbA, están aumentados en relación a los controles (12,13,14).
Way por lo menos 6 tipos diferentes de hernoglobinas en las ratas (1 5). De un cromatograma donde se observan seis picos de elución con un sistema de HPLC, denominados por las letras "a" a la " f ' I , las fraccio-. nes a+b+c están incrementadas en las ratas diab6ticas comparadas con las controles ( A í ! ) , (fig. 2).
4

0.008 CI
0.007 - O
* m o
Y- - 0.006 - .- * OQ 0.005
- 8 0.003
- ,? 0.004
- U m m E
U Q
'1J m 0.002 -
E l = 0,000 r 3 I
I 0.000
R a t a N o r m a l
= inyección de la muestra
4 = cambio de amortiguador
O 20 40 60 80 I O0 120
m i n u t o s
Figura 2 Cromatograma de los componentes de la hemoglobina de rata, usando HPLC (Figura tornada de Blanc R1.H.I2)
La gficación de la hemoglobina se produce lentamente durante la vida media del eritrocito, de tal manera, que cambios repentinos en los nive- les de glucosa sanguínea, no afecta el porcentaje de HbA,, permitien- do que esta determinación sea de gran utilidad para el control glicémico a largo plazo del paciente diabético reflejando la concentración relativa de glucosa en sangre de 30-60 días previos a la determinación. (3,4,6,16). Otras proteinas piasrnáticas como la albúmina responderAn más rápidamente a estos cambios repentinos de glucosa en sangre. (17,18).
5

Estreptozotocina La estreptozotocina es un agente diabetogénico ampliamente usado para la producción de diabetes experimental (fig. 3). Es un antibiótico extraído de Streptomyces achromogenes que posee propiedades antitumorales y oncogénicas (19,20). Su acción citotóxica sobre las células beta pancreáticas es específica, rápida y~ aparente desde la primera hora de su administración, resultando en necrosis celular irre- versible (1 9,21). La estreptozotocina tiene actividad oncogénica sobre las células beta pancreáticas y en menor grado sobre el riñón (20).
I H k -H
¿!=O
cH3
I N-N=O
I
Figura 3. ilfolécula de estreptozotocina
Después de la adminstración de estreptozotocina, la glucosa en sangre presenta una respuesta trifásica: a las dos horas los niveles se elevan, posiblemente por un efecto glicogenolítico en el hígado, más tarde tie- ne un decaimiento muy marcado a las IO horas, para después incrementarse a las 24 horas originando hiperglicemia permanente (19,Zl).
La duraciún y severidad del estado diabético está relacionada con la dosis de estreptozotocina. Dosis elevadas (P.e. 100 mglkg) resultan' en una inmediata destrucción de las células beta, diabetes severa y más tarde la muerte por coma diabético (19,21,22).
6

La estreptozotocina puede producir una diabetes gradual ya que posee un amplio margen de diabetogenicidad (21,22). Otro factor a conside- rar es la sensibilidad a la estreptozotocina con la edad de los animales; a menor edad mayor la sensibilidad a la estreptozotocina (23,24).
El mecanismo de acción de la estreptozotocina aún no es claro, pero se han planteado dos probables procesos por los cuales ejerce sus efectos citotóxicos en la célula beta pancreática. 2 2 2 5 1 0 La estreptozotocina, una metilnitrosourea unida al carbono dos de la glucosa, se descompone sin metabolizarse en un compuesto isocianato y en un metildiazohidróxido. El compuesto isocianato sufre carbamilación intracelular o bien puede carbamilar otros componentes celulares. El metildiazohidróxido se descompone en ión carbonio que puede reaccionar con el agua para dar metano1 y participar en la forma- ción de una poza de carbonos, o bien puede alquilar proteínas u otros componentes celulares como el DNA (fig. 4).
El ion carbonio altamente reactivo, puede interactuar con centros nucleofilicos del DNA, como el nitrógeno de la posición 7 de la guanina. Estas alquilaciones son eliminadas por un proceso escisión-reparación. Parte de este proceso lo lleva a cabo la enzima poli-ADP-ribosa sintetasa que utiliza NAD como sustrato. Se cree que esta enzima se sobreactiva de tal forma que depleta críticamente el sustrato, provocando la cesa- ción de la función celular y más tarde la muerte.
Otros estudios sugieren otro proceso, en el cual la estreptozotocina además de alquilar el DNA, tambien alquila componentes celulares importantes necesarios para la producción de ATP, P.e. enzimas mitocondriales. También puede alquilar componentes relacionados con los mecanismos de secreción de insulina en la céiula beta. Por lo tanto, además de la sobreactivación de la enzima poli-ADP-ribosa sintetasa, que depleta críticamente el NAD, se presenta una baja en la formación de ATP, probablemente por la alquilación de enzimas clave para su producción (25).
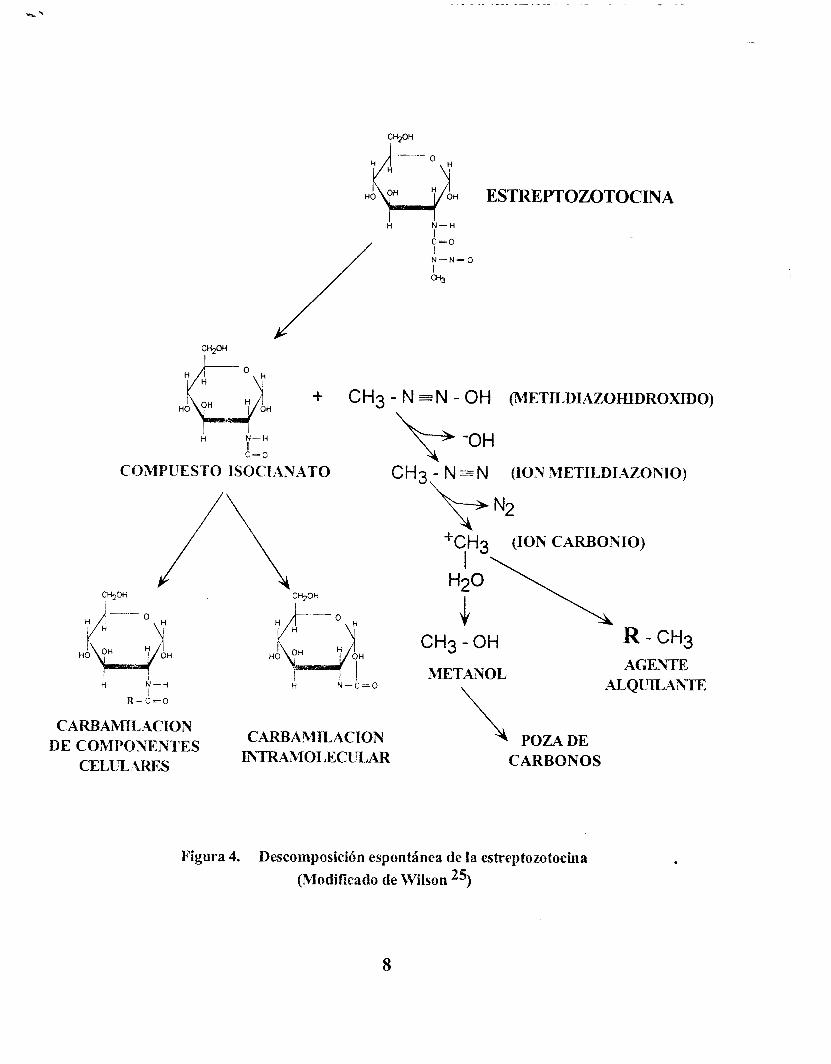
H H ESTREPTOZOTOCINA A N - H
I c=o /
C H l O H I
+ H H
CH3 - N =N - OH (METKDIAZOHIDROXIDO) c, -OH
'"I-. N2
CH3 - N E N (ION METII~DIAZONIO)
"CH3 (ION CARBONIO) I
- O
H CH3 - OH R - CH3 H H H
IVWTAVOL AGENIX H N-H H N-C=O
\ A L Q L K 4 N T E
R-L=O
CARBAML.4CION
CEL,LK-WS
CARBAMILACION POZADE DE C0,VPQNENTES ~ ~ ~ O L E C U L A R CARBONOS
Figura 4. Descomposición espontánea de l a estreptozotochla (Modificado de Wilson 25)
8

Metodos para determinar la HbA, Los métodos para cuantificar la hemoglobina glicada se pueden dividir en dos grandes grupos:
a) El primero depende del efecto que produce la glicación en la carga de la molécula de hemoglobina; como la cromatografía de intercam- bio iónico, HPLC, isoelectroenfoque y electroforésis en gel. La cromatografía de intercambio iónico sólo cuantifica la glicación en el amino terminal de la valina de la cadena beta de la hemoglobina, las glicaciones en otros sitios tienen un efecto insuficiente sobre la car- ga de la molécula de hemoglobina para permitir su separación por crornatografía (4).
b) El segundo grupo se basa en alguna propiedad específica del enla- ce cetoamino de la hemoglobina glicada y por lo tanto, mide la hemoglobina glicada total; ejemplos de estos métodos son hidrólisis en ácido débil y cromatografia de afinidad. El radioinmunoensayo también es utilizado p a a determinar la HbA,.
in ;.
Los factores principales que pueden afectar los resultados en la I
cromatografia de intercambio iónico son el pH del amortiguador y la y n ;:., I ,
temperatura, Por otro lado, enfermedades que producen uremia o icte- 9 ricia, concentraciones anormales de HbF, presencia de otras 9 m (.:. : i
hernoglobinas (HbS, HbC), enfermedades hematológicas pueden pro- ducir un aumento o disminución ficticio en el valor de HbA,; así como también el envejecimiento de las muestras, altas dosis de saliciiatos, transfusiones, el embarazo, etc. (3,4)
La determinación de HbA, ha probado ser útil en el control giicémico. Así, la elevación en los niveles de HbA,, podría estar relacionada con diabetes o intolerancia a la glucosa, pero valores normales de HbA, no excluyen la intolerancia a la glucosa (26). Asimismo, la HbA, incrementada no representa un marcador genético para la diabetes pero. sí refleja los niveles circulantes de glucosa en sangre en forma retros- pectiva por 1 o 2 meses (4).
U! r v
L.) ,
9

La Base de Schiff, conocida como la fracción iábil que se forma como un paso intermedio para dar la HbA,, puede incrementar el valor por- centual de la HbA,. En diabéticos con amplias fluctuaciones de gluco- sa sanguínea, es decir con mal control, podría ser relevante su partici- pación (3,4).
Algunos estuches de diagnóstico incluyen la eliminación de la fracción lábil. El valor %HbA, es afectado por la temperatura, pero al aplicar los factores de corrección de la temperatura (FCT) incluidos en el estuche, se obtiene el valor del %HbA, real.
10

. " . . ..
l . Determinar los valores de hemoglobina glicada (HbA,) en ratas con- trol (C) y experimental (DM).
2. Estudiar la correlación existente entre el porcentaje de hemoglobina glicada con los niveles de glucosa en sangre de las ratas DM.
i) LOS niveles elevados de glicemia en las ratas DM producen incre- mentos en el %HbA,.
.
11

Animales y Condiciones
14 ratas macho cepa Wistar entre 380-450 g de peso del Bioterio de la Unidad de Iwestigación Experimental, se dividieron en dos grupos:
I ) Experimer7tal (DM) con 5 ratas diabéticas, y
2) Control (C) con 9 ratas sanas.
bas ratas fueron alojadas individualmente en jaulas metabólicas 15 días antes de iniciar el estudio. Tuvieron acceso al agua y alimento (5001 de Bluebonnet) ad libidurn, bajo 12 hrs de luz y 12 hrs de obscuridad y a una temperatura de 20 a 22 O C .
Se transportaron en grupos de 4 ratas, al cuarto de procedimientos anexo, para iniciar el estudio y evitar el estrés.
Material para la inducción de la diabetes a) Estreptozotocha (Sigma, Catálogo no. S-0130), en dosis de 65 mg/
kg, colocada en tubos Eppendorf de acuerdo al peso corporal de la rata y cocservada a 4 O C por 18 hrs antes de la inyección.
b) Amortiguador de citratos 0.1 M y pH 4.5 (ajustado con NaOH 0.2M) recién preparado.
c) Balanza granataria para medición del peso corporal. d) Balanza analítica (Mettier H-20T) e) Cámara de anaestesia (Natge) con éter. f) Jeringas insuiínicas con capacidad de 1 mL. g) Tubos Eppandorf con EDTA al 6% como anticoagulante. h) Bísturi y cautín de punta fina.
La maniobra general en cada una de las ratas fue: i) Ayuno previo ii) Medición del peso corporal iii) Anestesia en cámara con éter iv) Toma de muestra sanguinea v) Inyección de estreptozotocina 6 amortiguador de citratos. vi) Recuperación y alojamiento.
12

Producción de Diabetes 1. Para producir la diabetes las ratas tuvieron ayuno de 15 a 17 hrs.
2. Se registró el peso antes de aplicar la anestesia en cámara con éter, una por una, iniciando por el grupo C y luego el grupo DM.
3. La muestra sanguínea de 250 pL se obtuvo cortando la punta de la cola y cauterizando con un cautín de punta fina; y recibida en tubos Eppendorf con anticoagulante.
4. Mediante inyección intravenosa, vena dorsal del pene, se administró al grupo C, 0.5 mL de amortiguador de citratos. En algunos casos fue necesario continuar la anestesia usando un cono con éter para realizar la inyección.
2 rJ 3 3 q t.J "8 * fJ
5. En la producción de la diabetes, el grupo DM recibió estreptozotocina mantenida en hielo y después disuelta con 0.5 mL de amortiguador de citratos, preparada inmediatamente antes de la inyección.
6. Ya recuperadas de la anestesia, fueron devueltas a sus jaulas para proporcionarles agua y alimento ad libidurn.
Determinación %HbA, a) Soportes y gradillas de acrilico con capacidad para 20 columnas.
b) Espectrofotómetro Beckman, Clinical System 700.
c) Micropipeta de 100 y 50 p t .
d) Pipetas de 10 mL (graduaciones de 1 y 0.5 mL).
e) Propipeta, vortex, vasos de precipitados (50 y 20 mL) y termómetro.
9 Estuche de diagnóstico para determinar hemoglobina glicada o %HbA,, (Sigma, Catálogo no. 440-8) que contiene:
i) Columnas de plástico con resina de intercambio catiónico y amor-+ tiguador pH 6.9 (Catálogo no. 440-4) ii) Amortiguador de fosfatos (solución eluyente) pH 6.9 y azida de sodio menos O. 1 % (Catálogo no. 440-3) iii) Solucibn hetnolizante (CatBlogo no. 440-2)
13

g) Preparaciones liofilizadas de sangre humana con concentraciones conocidas de HbA,, nivel normal (N), valor de referencia asignado 6.7 - 9.3% (Sigma, Catálogo no. G-2012); nivel alto (A), valor de referencia asignado 11 516.5% (Sigma, Catálogo no. G-I O1 2).
Procedimiento I. Las columnas insertadas en el soporte se drenan completamente en una charola de plástico extendida.
II. En tubos de ensaye con capacidad de 5 mL, se colocan IO0 pL de muestra sanguínea (sin duplicado). Las muestras fueron conservadas a 4 OC por 24 hrs para su procesamiento.
Ill. A cada uno de los tubos se adicionó 0.5 mL de solución hemolizante y posteriormente agitados en vortex.
IV. Las columnas ya drenadas y con tubos marcados con capacidad de I O ml, para recibir el eluído. se colocó suavemente 50 pL del hemolizado k I:
en la cama de resina, sin rebotarla. zc; V. La separación de HbA, se realizó con 5.0 mL dc solución de elución, que son deslizados lentamente por la pared de la columna. m y 5 2
VI. Registro de la temperatura durante la eluci6n. (Se recomienda que $ la temperatura permanezca constante hasta qcz la elución se haya completado). m!? 2 5
Vil. Una vez que drenó completamente la columna, el eluído, es decir la 92 HbA,, se cubrió con papel parafilm para homogenizarse.
Vlli. La hemoglobina adulta total (HbAJ fue preparada en otros tubos marcados con capacidad de 15 mL, con 20 pL del hemolizado y 10 mL de solución de elución, tapados y homogenizados.
IX. La lectura de la absorbancia de ambas fracciones fue leida en el espectrofotórnetro a 41 5 nm.
X. Elegicio el factor de corrección de la temperatura (FCT), se aplicó la fórmula para encontrar el %HbA,.
; S
m
Abs. HbA, x 100 x FCT
Abs. HbA, x 5 %HbA, =
14

La hemoglobina glicada o HbA, es eluída en una sola fracción y su porcentaje se calculó con respecto a la hemoglobina adulta total pre- sente. El diagrama de flujo de la figura 6, muestra la secuencia general.
El estuche de diagnóstico empleado no elimina la fracción lábil.
Se obtuvo el valor basal de HbA, y después a intervalos de 3 semanas. ~
Glicemia a) Autoanalizador Beckman, Glucose Analyser I I , que utiliza la tkcnica
de glucosa-oxidasa.
b) Centrífuga Beckman, Spinchron R Centrifuge
c) Pipetas Pasteur de punta larga y tubos Eppendorf
Procedimiento 1 O0 pL de sangre son centrifugados a 3000 rpm por 8 min para obtener el suero. Este último se trasladó con una pipeta Pasteur a otros tubos Eppendorf marcados. Se realizaron determinaciones de glicemia se- manalmente con 10 pL de suero (por duplicado), para documentar el estado diabético de las ratas.
Volumen de orha Se insertaron conos de metal en las jaulas metabólicas y frascos de pl6stico para colectar la orina de 24 hrs.
Antes de cuantificar el volumen de orina, los residuos de comida fueron. eliminados con una coladera fina.
La medición se hizo antes de la inyección de estreptozotocina y des- pués 2 veces a la semana.
15

Glucosa y cetonas en la orina Análisis semicuantitativo con tiras reactivas Keto-Diastix, (Ames Bayer, Catálogo no. 2883).
En el mismo frasco de plástico donde fue colectada la orina, (antes de cuantificar su volumen), se sumerje el área reactiva; y 1 minuto más tarde se compara con la &ta de colores y concentraciones (Tabla 1). Las cetonas son comparadas a los 15 seg y la glucosa a los otros 30 seg . Nuevamente, se obtuvo la medición basal y despues 2 veces a la se- mana.
" ""
GLUCOSA CETONAS
I
Negativo Negativo I00 mg/dL 5 mgldL
500 mgldL 40 mgldL 1 O00 mgldL 80 mgidL
2000 o más mgldL 160 mgldL
.* .. ~
r c .= , 250 mg/dL +I5 mgidL I
Tabla 1. Carta de colores y concentraciones, de glucosa y cetonas en la orina. (La alta concentración de cetona puede disminuir el color del área de glucosa).
Control de Calidad Se determinó el coeficiente de variación de muestras liofiliradas de concentracih conocida: intraensayo (una ocasibn), midiendo 1 O veces la misma muestra. El análisis interensayo se realizó construyendo grá- ficas de control, que demuestran los niveles promedio y rangos de acep-' tación de los niveles N y A. Ambos niveles fueron determinados por triplicado junto con las muestras de las semanas 0,3,6,9 y 12.
16

3'
Análisis Estadistica
Comparación de medias pareadas y no pareadas por la prueba de t (27), Coeficiente de Correlación de Rangos de Spearman (28), regre- sión lineal por Mínimos Cuadrados, Gráficas de Control (29) y el Coefi- ciente de Correlación de Pearson, utilizando el paquete estadístico Sigma Stat y graficación científica Sigma Plot (Jandel Scientific).
Figura 5. Minicotumna de cromatografia en soporte de acrílico.
17

,
Adicionar 0.5 rnL de hemolizante
100 pL de muestra. Agitar en Vortex
Insertar las columnas en el soporte, quitar los tapones superior
e inferior. Drenar totalmente la columna
1 Inmediatamente adi- cionar 50 pL del he-
molizado directo en la cama de resina, evitan. do "rebotaf la resina
~ ~~ ~ ~~~ ~~ ~
notar la temperatura Agregar lentamente
1
Figura 6. Determinación de Hemoglobina Glicada
Colocar gradilla y tubos marcados con capacidad de 10 mL para recibir el eluido
Marcar tubos de 15 m i
I solución de elución y
1
I Corregir el %HbA, con el Abs. HbA, x 100 x FCT Factor de Correccih de
Temperatura (FCT) Abs. HbA, x 5 I 18
- RESULTADOS
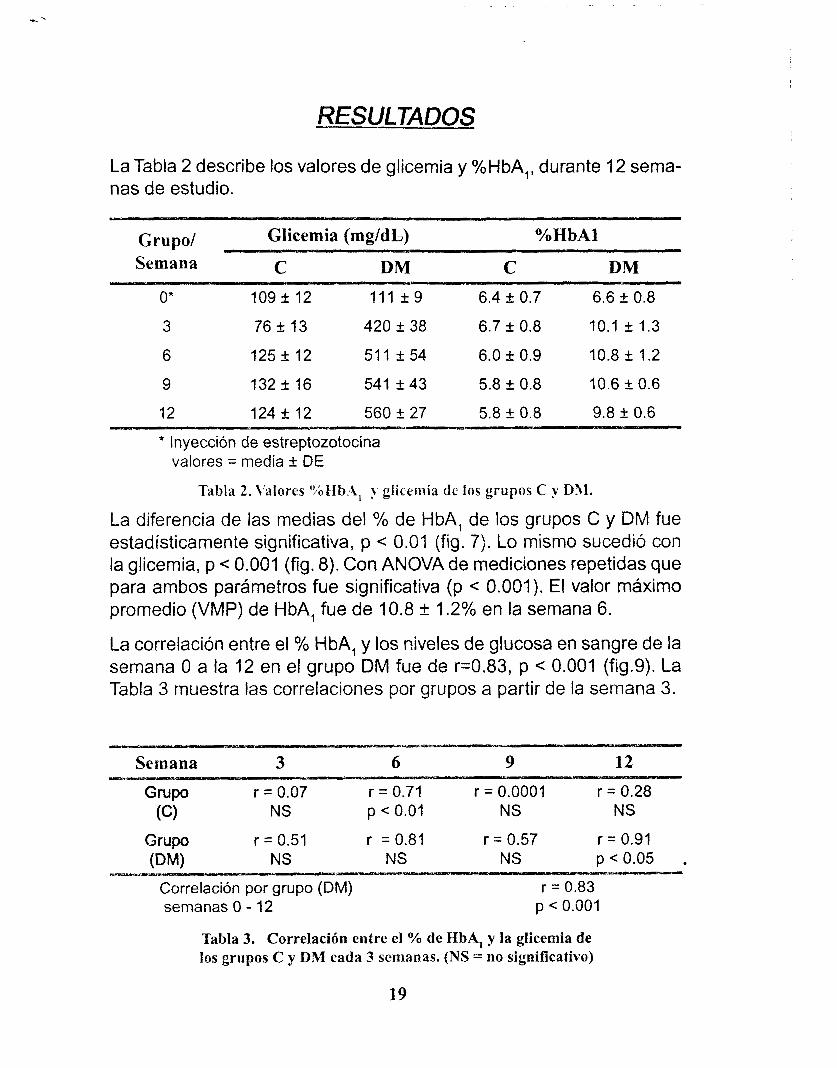
La Tabla 2 describe los valores de glicernia y %HbA,, durante 12 sema- nas de estudio.
Grupo/ Glicemia (mg/dL) YoHbAl Semana C DM C DM
O* 109 k 12 111 2 9 6.4 f 0.7 6.6 k 0.8
3 76 f 13 420 f 38 6.7 k 0.8 10.1 f 1.3
6 125 5 12 511 k54 6.0 2 0.9 10.8 2 1.2
9 132 k 16 541 2 43 5.8 k 0.8 10.6 +_ 0.6
12 124 k 12 560 k 27 5.8 k 0.8 9.8 & 0.6
* Inyección de estreptozotocina valores = media f DE
Tabla 2. \'alores O/;Hb.A, glicemia de los grupos C y D31.
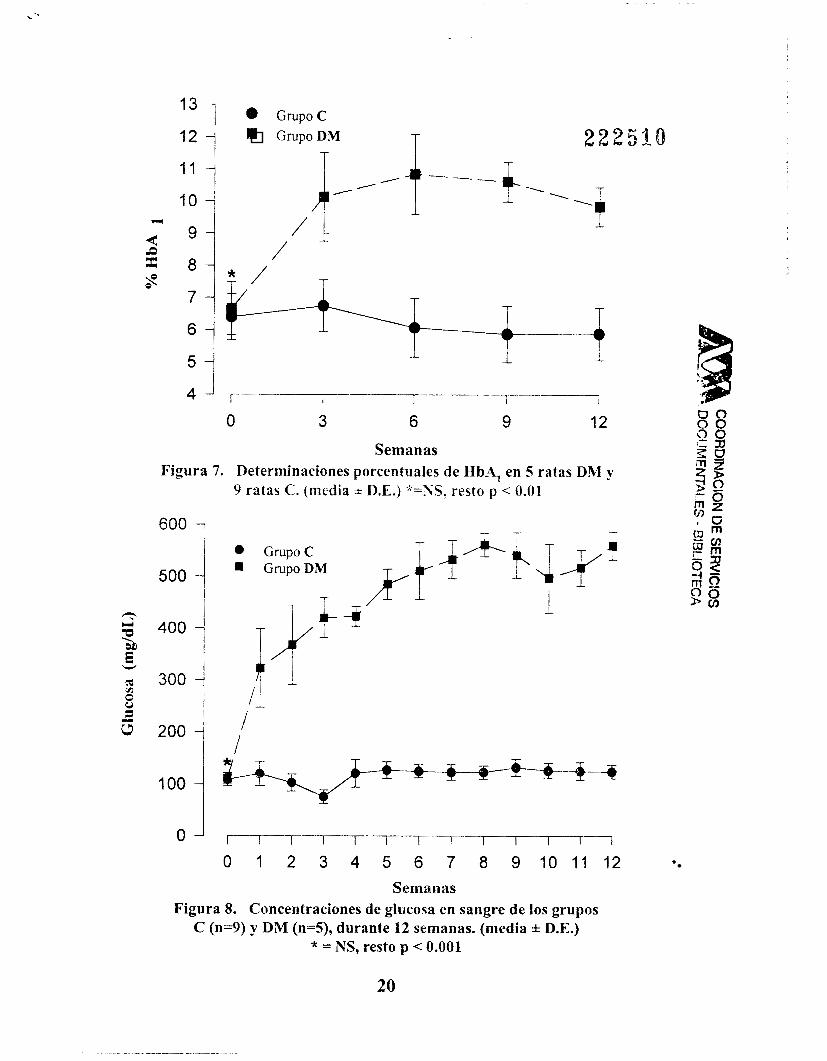
La diferencia de las medias del % de HbA, de los grupos C y DM fue estadísticamente significativa, p < 0.01 (fig. 7). Lo mismo sucedió con la glicernia, p < 0.001 (fig. 8). Con ANOVA de mediciones repetidas que para ambos parámetros fue significativa (p c 0.001). El valor máximo promedio (VMP) de HbA, fue de 10.8 k 1.2% en la semana 6.
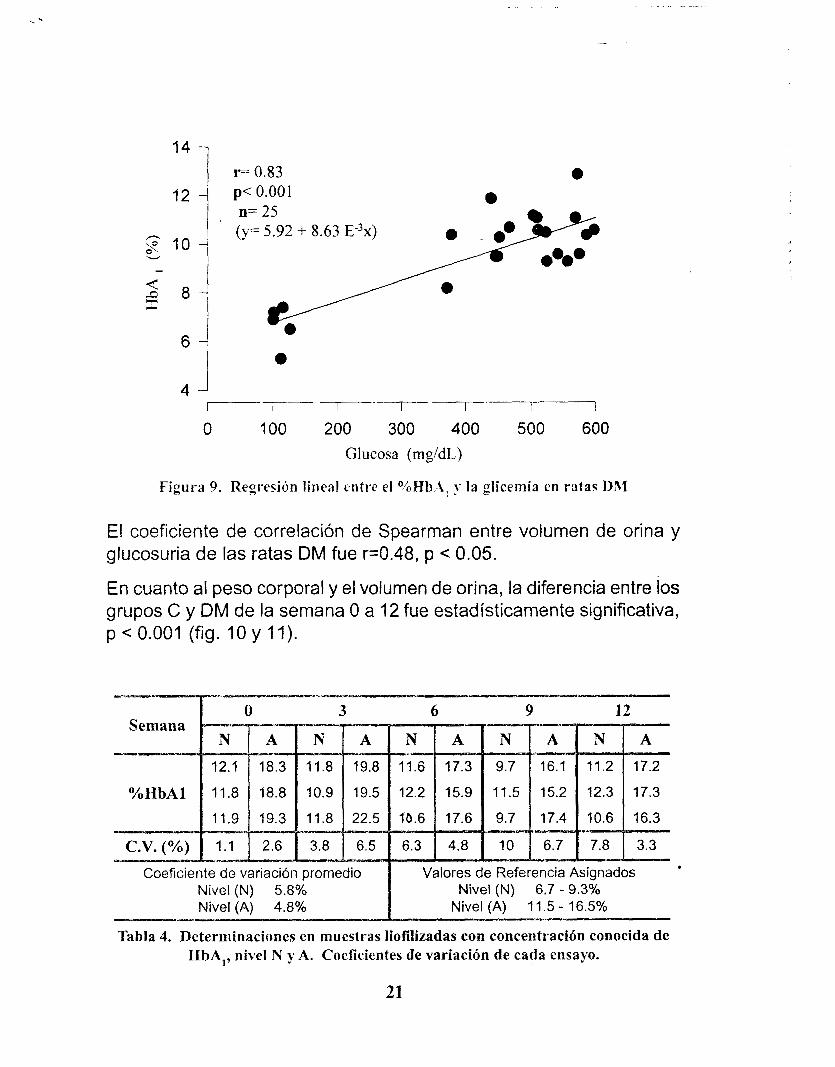
La correlación entre el % HbA, y los niveles de glucosa en sangre de la semana O a la 12 en el grupo DM fue de r=0.83, p < 0.001 (fig.9). La Tabla 3 muestra las correlaciones por grupos a partir de la semana 3.
Semana 3 6 9 12 ~ ~~~ _ _ ~ ~ -
Grupo r = 0.07 r = 0.71 r = 0.0001 r = 0.28 NS p c 0.01 NS NS ((3
(DM) NS NS NS P < 0.05 . Grupo r = 0.51 r = 0.81 r = 0.57 r = 0.91
Correlación por grupo (DM) semanas O - 12
r = 0.83 p c 0.001
Tabla 3. Correlación entre el YO de HbA, y la glicemia de los grupos C y DM cada 3 sernanas. (NS = no significativo)
19
l3 1 e Grupo c 12 "/ Grupo DM
I T T 10 l1 1
I
2 2 2
4 " r----r" -m O 3 6 9 12
Semanas Figura 7. Determinaciones porcentuales de HbA, en S ratas DM y
9 ratas C. (media X D L ) *=SS, resto p < 0.01
600
500
T F( 400 . W
1 O0
* - !
""
5 P o
O -I r-r-"--T-" I I I I I I I 1
O 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 Sf%I;anas
Figura 8. Concentraciones de glucosa en sangre de los grupos C (n=9) y DM (n=5), durante 12 semanas. (media f D.E.)
* = NS, resto p < 0.001
20
r= 0.83 e
n= 25 p< 0.001 e
h (y= 5.92 + 8.63 E - 3 ~ ) O ~
10
e 3
6 e
4
O 1 O0 200 300 400 500 600 Glucosa (mg/dL)
Figura ?. Regresihn lineal entre el OioHh,-i, y la glicemia en ratas DM
El coeficiente de correlación de Spearman entre volumen de orina y glucosuria de las ratas DM fue r=0.48, p c 0.05.
En cuanto al peso corporal y el volumen de orina, la diferencia entre los grupos C y DM de la semana O a 12 fue estadisticamente significativa, p c 0.001 (fig. 10 y 11).
~ ~~ ~~ ~
Tabla 4. Determinaciones en muestras liofilizadas con concentración conocida de HhA,, nivel N y A. Coeficientes de variación de cada ensayo.

550
250 "/ 200 '
r - " r - r Í 1 "-r-I-"
O 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
Semanas
Figura 10. Pesos promedio de ratas C (n=9) y D3f (n=5) (media f DE) * = M , ** p < 0.01, resto p < 0.001
I
240 4 ~
!
120 e
80
O 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
S e m a n a s Figura 11. Volumen de orina de 24 horas de ratas C (n=9) y DM (n=5)
(media f DE), * = M , resto p < 0.001
22

Control de Calidad: el coeficiente de variación intraensayo fue 5.5%. El coeficiente de variación interensayo, con muestras liofilizadas de con- centración conocida fue de 5.8% para el nivel N y 4.8% para el nivel A. La gráfica de control para las muestras liofilizadas N y A, muestra que todas las "corridas" están dentro de los limites de confianza (fig. 12-A y B). El análisis multivariado aplicado a los valores de ambos niveles en forma conjurrta estuvo dentro de los límites de control con T2 debajo de 5, siendo ei limite de rechazo T2 = 25.4 (fig. 12-C). La tabla 4 contiene los valores de referencia asignados para los niveles N y A, además de los datos obtenidos durante el estudio y sus correspondientes coefi- cientes de variación.
g t "-.---L""L--"-.- --"""" L
I I
I """"""""""" '
"""""
12 I I
2o t 5
O O 3 6 9 12
s e m a n a
.
Figura 12. Graficas de Control de Calidad. (A) Muestra liofilizada nivel N, (B) Muestra nivel A, y (C) análisis multivariado de los niveles N y A.
23

La giicación de la hemoglobina se lleva a cabo durante la vida media del eritrocito, aproximadamente 45-50 días en la rata (7), de esta forma fue posible aplicar las determinaciones %HbA, por intervalos de tiempo de 3 semanas, reflejando las concentraciones de glucosa en sangre a largo plazo, en el estado diabético de las ratas.
En el %HbA, y la glucosa en sangre de la semana O a la 12, la diferen- cia en las medias de los grupos C y DM, fue estadisticamente significa- tiva, p<O.OI y p<O.OOI, respectivamente. Los resultados son compara- bles a los de Blanc y cols. (1 2) y Vialettes y cols. (1 6).
AI estudiar la correlación existente entre el porcentaje de HbA, con los niveles de glucosa en sangre de las ratas DM, fue r=0.83, p<O.OOI. Gould y cols., en ratas de 15 semanas de edad, reportaron una correla- ci6n similar, r=0.90, p<O.OOI y en ratones de r=0.92, p<O.OOl.
El grupo de Vialettes, con ratas hembra, la correlación fue r=0.30, p4.05. A los 20 y 50 dias fue no significativo, pero a los 60 días fue r=0.58, ~ 4 . 0 5 .
De la regresión lineal aplicada al %HbA, contra la glucosa en sangre del grupo DM, demuestra que por cada incremento de 1 O0 mg/dL en la glucosa en sangre, la HbA, aumentó en 0.86 %.
Vialettes y cols., aunque aplica la correlación lineal, sugiere que la glicación de la hemoglobina es un proceso saturable con variaciones en el tiempo requerido, dependiendo del nivel de hiperglicemia.
El porciento de glicación de la hemoglobina está en función de las con- centraciones de carbohidratos circulantes en la sangre y de la vida media del eritrocito, que es de 45-50 días en la rata (7). En el grupo OM el %HbA, parece haber llegado a una saturación en la semana 6. En las ratas diabéticas del estudio de Blanc y cols., ocurrió en la semana 5, este intervalo de tiempo es similar al de la vida media del eritrocito de rata, que es aproximadamente de 6 semanas.
El valor máximo promedio (vrnp) de HbA, encontrado en la semana 6 fue de 10.8 k I .2%, mientras que con un sistema de HPLC (12), se obtuvo un vmp de 10.8 k 0.7% a la quinta semana.
24

Es importante señalar que la segunda parte del trabajo realizado por Vialettes et al, consistió en el transplante de islotes en ratas diabéticas, con el objeto de disminuir el %HbA, a valores comparables con los de las ratas no-diabéticas (de 17.6 2 I .4 a 9.4 2 0.9%); y encontró una velocidad de cambio de 5 semanas para normalizarla. Esto refuerza el hecho, de que, el tiempo aproximado de su formación es similar al de su normalización.
Las ratas DM, además de su marcada hiperglicemia, presentaron cetonuria aproximadamente de 60 mg/dL o más desde la primera se- mana de la inducción; proceso que era raro en el estudio realizado por Junod et al. El peso corporal de estas ratas durante el estado diabéti- co, permaneció similar o cerca del valor inicial en los 28 días de estudio (1 60-230 9).
En el grupo DM, las grasas fueron degradadas para obtener energía, produciendo la cetonuria. Su peso corporal disminuyó de 426 k 46 a 247 k 18 g, pc0.001, contra el grupo C.
La glucosuria apareció desde el primer día y se mantuvo hasta el final del estudio, entre 500-1 O00 mg/dL (0.5 - 1 .O%). En el trabajo de Junod y cols. se presenta este comportamiento pero en mayor cantidad (Z- I ) , de 2 a más del 10% de glucosuria desde el día 1 hasta el final (28 días).
Los coeficientes de variación obtenidos no son similares a los que usual- mente se manejan en la cromatografía de intercambio iónico: 2-3% en el intraensayo y de 4-5% en el interensayo (4), sin embargo, las gráfi- cas de control de calidad validan la aceptación de estos resultados.
25

CONCLUSlONES
Los valores de hemoglobina glicada o HbA, de las ratas DM, fueron más elevados que los de las ratas C, la diferencia en las medias de estos grupos a partir de la semana 3 fue estadísticamente significativa, pco.01.
Existió una buena correlación entre el %HbA, y la glicemia del grupo DM de la semana O a la 12, con r=0.83, pcO.001.
Cada incremento de 100 mg/dL de glucosa en sangre produjo un au- mento en 0.86% en el valor de la hemoglobina glicada.
26

BtBLlOGRAFtA
l . Lee GR, Bithell TC, Foerster J, Athens JW, Lukens JN. Wintrobe's Clinical Hematology. USA: Ninth Edition. Lea & Febiger. 1993:l-101-
2 . Roth M. cGlycated Hemoglobin)), not ((glicosylated)) or
128, 195-6.
((glucosylated)). Clin Chem 1983; 11: 1991.
3. Mayer TK, Freedman ZR. Protein glycosylation in diabetes mellitus: a review of laboratory measu- rements and of their clinical utility, Clin Chem Acta 1983; 127: 147-84.
4. Kennedy L. Glycation of hemoglobin and Serum proteins. In: Alberti KG", DeFronzo RA, Kenn H, Zimmet P, (eds). International Textbook of Diabetes Mellitus. England:John Wiley & Sons, 1992: 985-1 007. p _ _
5. Higgins PJ, Bunn HF. Kinetic Analysis of the nonenzymatic S
2 ; ) :. r ' .
glycosyiation of hemoglobin. J Biol Chem 1981 ; I O: 5204-8.
6. Gabbay KH, Hasty K, Breslow JL, Curtis Ellison R, Bunn HF, Gallop ? ' ~ PM. Glycosylated hemoglobins and Long-term blood glucose con- i trol in diabetes mellitus. J Clin Edocrinol Metab 1977; 44: 859-64. ! *
I CT 7
pT1' "
... r- . . ... - " . ._ . .. ", ,
7. Dukes y Swenson. Fisiología de los Animales Domésticos. España: Colecci6r: Ciencia y Técnica Aguilar, 1985: 1-45. 2- (ir;
8. Shapiro R, McManus MJ, Zalut C, Bunn HF. Sites of nonenzymatic glycosylation of human hemoglobin A. J Biol Chem 1980; 225: 31 20- 7.
9. Alperin JB, Dow PA, Sout LC. Comparison of hemoglobin A,c in human and baboon blood. Acta Haernatol 1979; 61: 334-8.
IO. Fitzgibbons JF, Koler RD, Jones RT. Red Cell Age-related changes of hemoglobins and A,c in normal and diabetic subjets. J Clin Invest 1976; 58: 820-4.
1 1. Tepperman J. Fisiología Metabblica y Endócrina. México. 1975: 172: 205.
27

12. Blanc MH, Rhie FH, Dunn PJ, Soeldner JS. The determination of glycosylated hemoglobins in rats, using high pressure liquid chromatography. Metabolism 1981 ; 4: 31 7-22.
13.Gould BJ, Flatt PR, Kotecha S, Collett S, Swanston-Flatt SK. Measurement of Glycosylated haemoglobins and Glycosylated plas- ma proteins in animal models with diabetes or inappropriate hypo- glicaemia. Horm Metabol Res 1986; 18: 795-99.
14. Higgins PJ, Garlick RL, Bunn HF. Glycosylated hemoglobin in Human and Animal Red Cells. Diabetes 1982; 31: 743-8.
15. Stein S, Cherian MG, Mazur A. Preparation and Properties of Six Rat hemoglobins. J Biol Chem 1971 ; 17: 5287-93.
16. Vialettes B, Vovan L, Simon MC, Lassmann V, Altomare E, Vague. Kinetics of fast haemoglobin in diabetic rats. Diabetologia 1982; 22: 264-8.
17. Sobe! DO, Abbassi Val. Use of fructosamine test in diabetic children. Diabetes Care 1991; 14: 578-83.
18. Day JF, lngebretsen CG, lngebretsen WR, Baynes JW, Thorpe RS. Nonenzymatic Glucosylation of serum proteins and hemoglobin: response to changes in blood glucose levels in diabetic rats. Diabe- tes 1980; 29: 524-7.
19. Rerup CC. Drugs producing diabetes through damage of the insulin secreting cells. Pharm Rev 1970;4: 485-51 8.
20. Kazumi T, Yoshino G, Fujii S, Baba S. Tumorigenic action of streptozotocin on the pancreas and kidney in male Wistar rats. Can Res 1978; 38: 2144-7.
21. Junod A, Lambert AE, Orci L, Pictet R, Gonet AE. Studies of the diabetogenic action of streptozotocin. Proc SOC Exp Biol Med 1967; 126: 201 -5.
22. Tancrede G, Rousseau-Migneron S, Nadeau A. Long-term change? in the diabetic state induced by different doses of streptozotocin in rats. Rr J Exp Path 1983; 64: 1 17-23.
28

23. Masiello P. Influence of age on the sensitivity of the rat to streptozotocin. Horm Res 1979; 5: 262-74.
24. Riley WJ, Mc Connell TJ, Maclaren NK, Mc Laughlin JV, Taylor G. The diabetogenic effects of streptozotocin in mice are prolonged and inversely related to age. Diabetes 1981; 30: 718-23.
25. Wilson GL, Letter EH. Streptozotocin Interactions with Pancreatic b cells and the Induction of Insulin-Dependent Diabetes. Curr Top in Microbio1 and lnmunol 1990; 156: 27-54.
26. Lev-Ran A. Glycohemoglobin: its use in the follow-up of diabetes and diagnosis of glucose intolerance. Arch Int Med 1981; 141: 747- 9.
27. Colton T. Statistics in Medicine. First Edition. USA: Little, Brown. 1974: 131 -45.
28. Siege1 S. Estadística no Paramétrica Aplicada a las Ciencias de la Conducta. h4éxico: Trillas, 1970: 233-45.
29. Dux PJ. Handbook of Quality Assurance for the Analytical Chemistry Laboratory. Second Edition. Van Nostrand Reinhold. USA. 1990: 31- 42.
29


















