
¿Pueden las poblaciones crecer infinitamente?Efecto beneficioso de las interacciones sociales...
50
Crecimiento poblacional ¿Pueden las poblaciones crecer infinitamente?
Transcript of ¿Pueden las poblaciones crecer infinitamente?Efecto beneficioso de las interacciones sociales...

Crecimiento poblacional
¿Pueden las poblaciones crecer infinitamente?
• Los individuos de una población no viven aislados y usan los mismos recursos limitados.
• Los recursos son finitos
COMPETENCIA INTRAESPECÍFICA
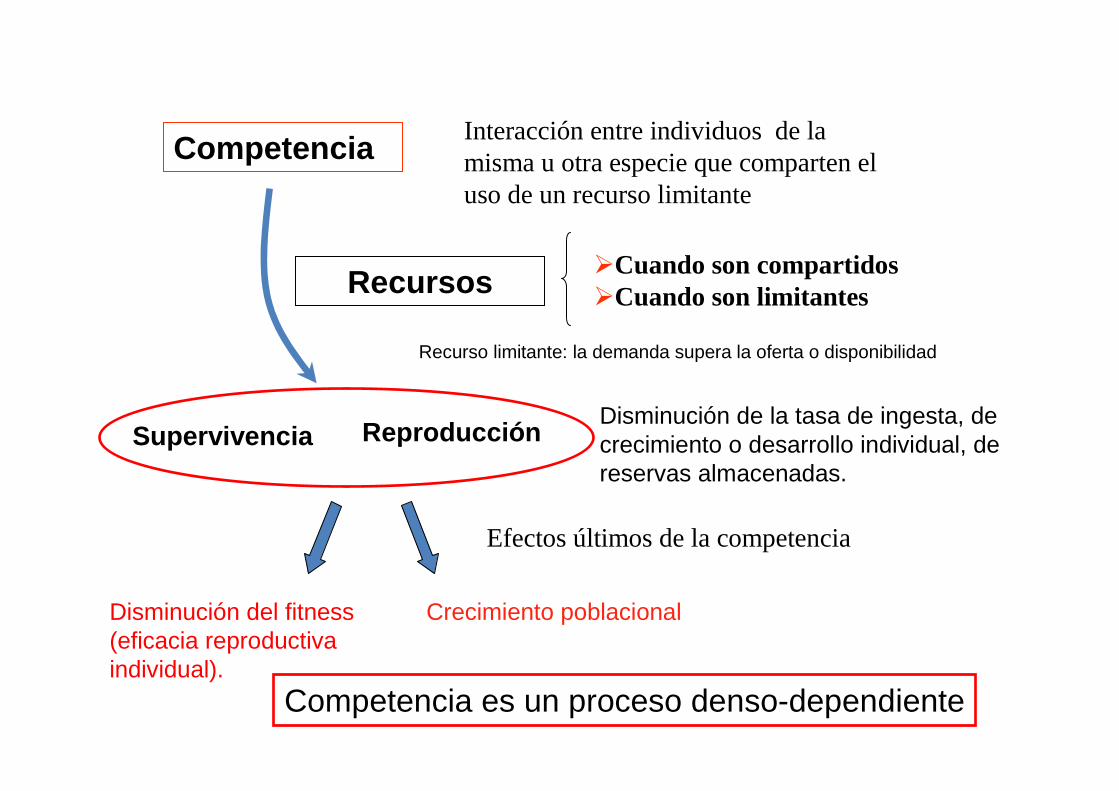
Efectos últimos de la competencia
Recursos
Competencia
�Cuando son compartidos�Cuando son limitantes
Interacción entre individuos de la misma u otra especie que comparten el uso de un recurso limitante
Recurso limitante: la demanda supera la oferta o disponibilidad
Disminución del fitness(eficacia reproductiva individual).
ReproducciónSupervivenciaDisminución de la tasa de ingesta, de crecimiento o desarrollo individual, de reservas almacenadas.
Competencia es un proceso denso-dependiente
Crecimiento poblacional
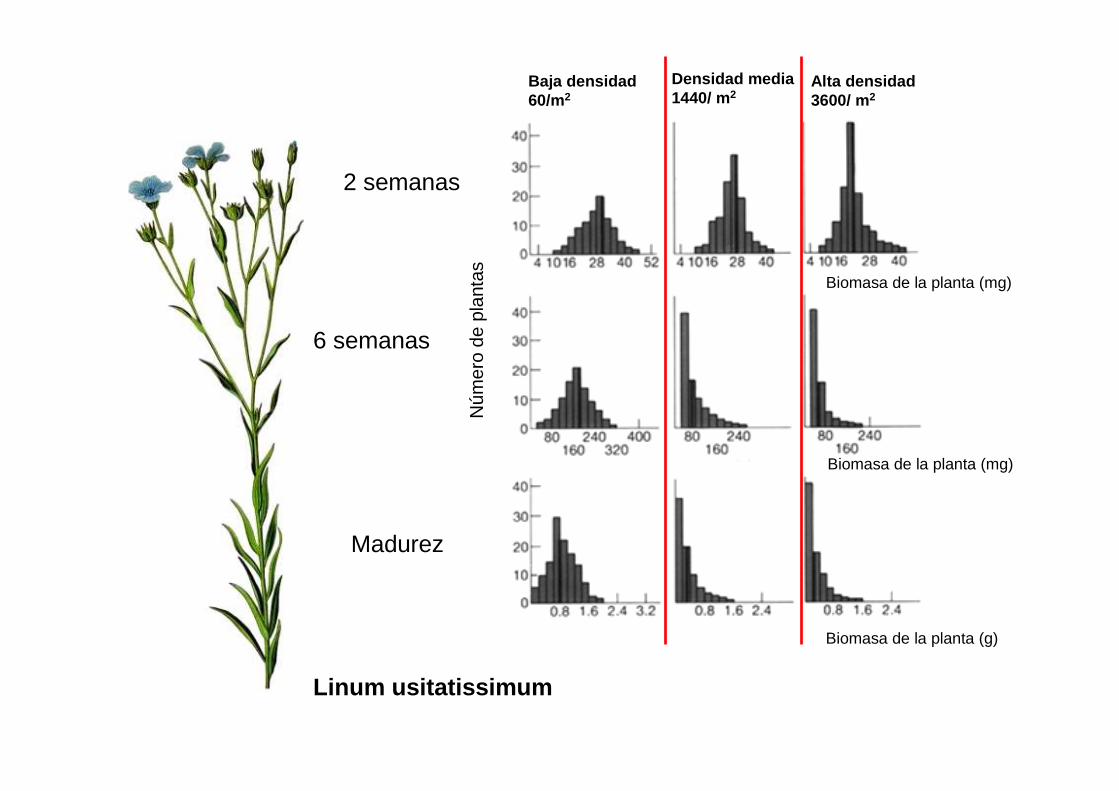
Linum usitatissimum
Baja densidad60/m2
Densidad media1440/ m2
Alta densidad3600/ m2
Biomasa de la planta (mg)
Biomasa de la planta (mg)
Biomasa de la planta (g)
2 semanas
6 semanas
Madurez
Núm
ero
de p
lant
as
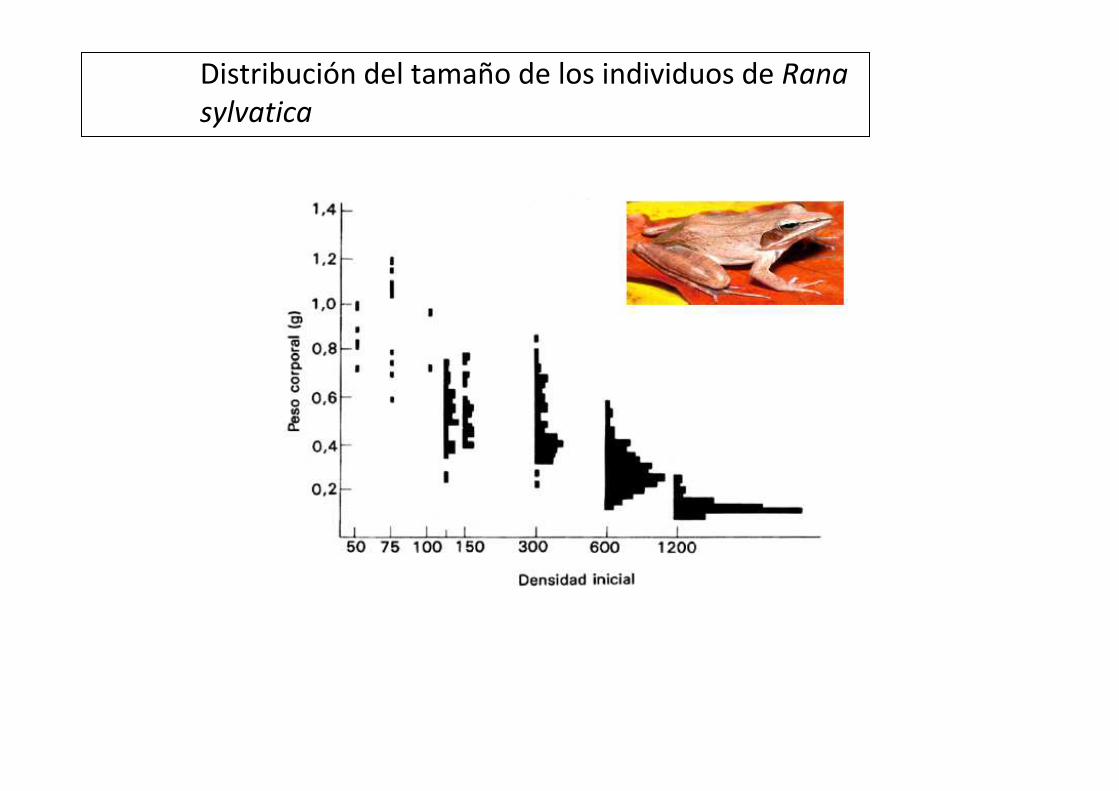
Distribución del tamaño de los individuos de Rana
sylvatica

Prokelisia marginata
(HomopteraDelphacidae) T
iem
po d
e de
sarr
ollo
(d
ías)
Larg
o de
l cue
rpo
(mm
)S
uper
vive
ncia
Densidad de chicharras

Efectos últimos
� Menor contribución a la próxima generación
� Disminución de supervivencia
� Disminución fecundidad
� Disminución de la eficacia biológica
Efectos próximos
� Cambios de tamaño (peso) individual y tiempo de desarrollo
� Disminución biomasa
� > Uso de hábitats pobres
� > Exposición a depredadores
� > Susceptibilidad enfermedades
Competencia intraespecífica
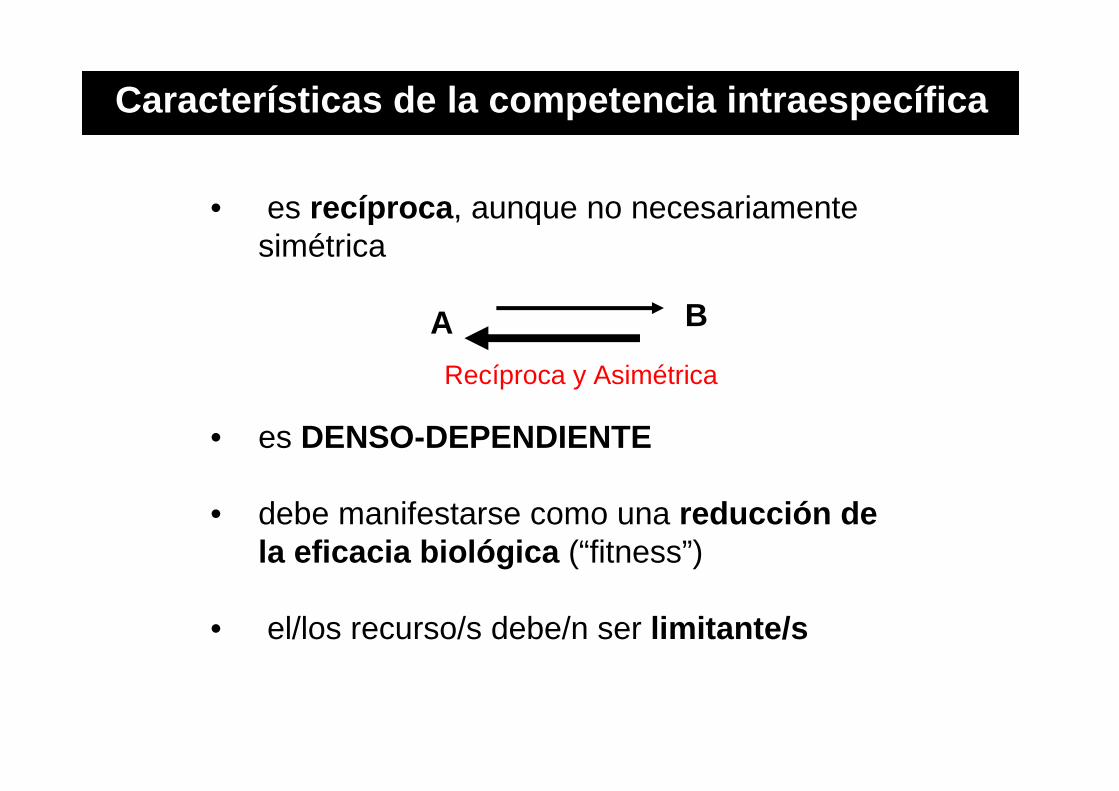
• es recíproca , aunque no necesariamente simétrica
• es DENSO-DEPENDIENTE
• debe manifestarse como una reducción de la eficacia biológica (“fitness”)
• el/los recurso/s debe/n ser limitante/s
Características de la competencia intraespecífica
Recíproca y Asimétrica
A B

Tipos de competencia intraespecífica
• de Interferencia (de torneo, jerarquizada, regulada, “contest”)
• por Explotación (anárquica, no jerarquizada, “scramble”)
Directa. Disminución de los recursos disponibles para un individuo debido a su interacción con otro individuo.
Indirecta. Mediada por el nivel del recurso limitante. Disminución de los recursos disponibles para un individuo debido a su consumo por parte de otro individuo.

Interferencia
Explotación
Efectos de la competencia por explotación y por interferencia
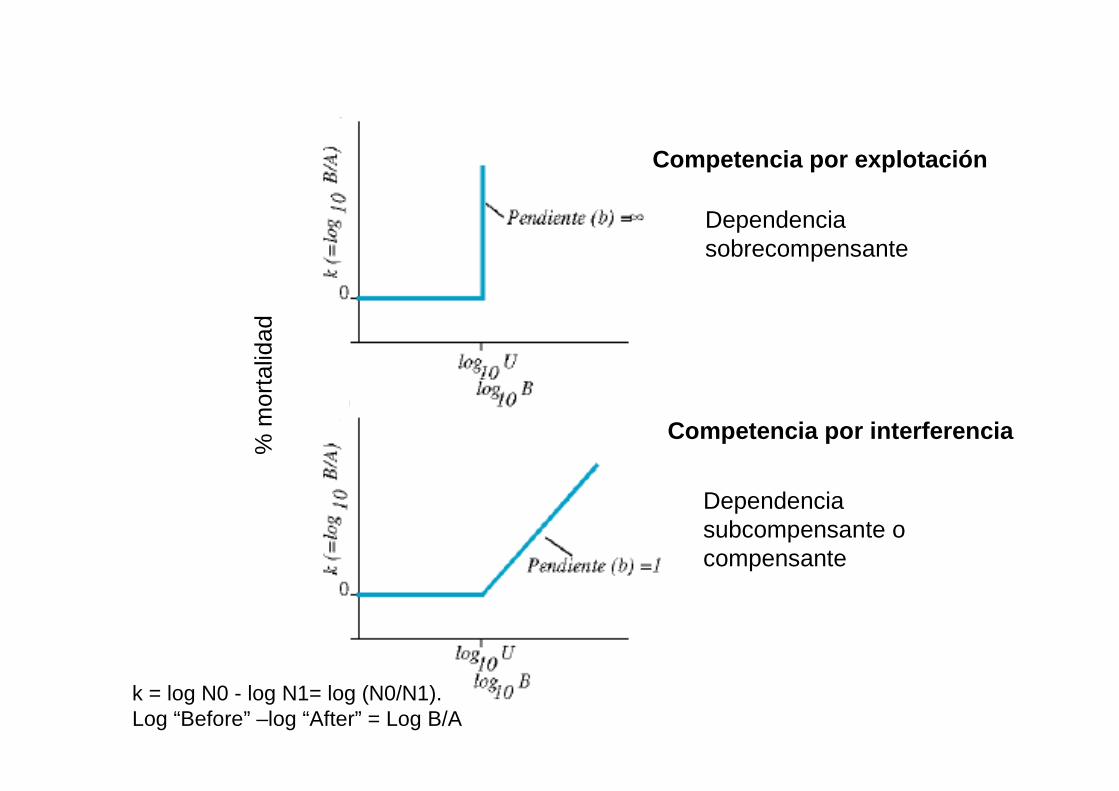
Efectos de la competencia por explotación y por interferencia
% m
orta
lidad
k = log N0 - log N1= log (N0/N1).Log “Before” –log “After” = Log B/A
Dependenciasobrecompensante
Competencia por explotación
Competencia por interferencia
Dependencia subcompensante o compensante
Número inicial de huevos
Tas
a de
mor
talid
ad
Núm
ero
de m
uert
esN
úmer
o de
sup
ervi
vien
tes
Número inicial de huevos
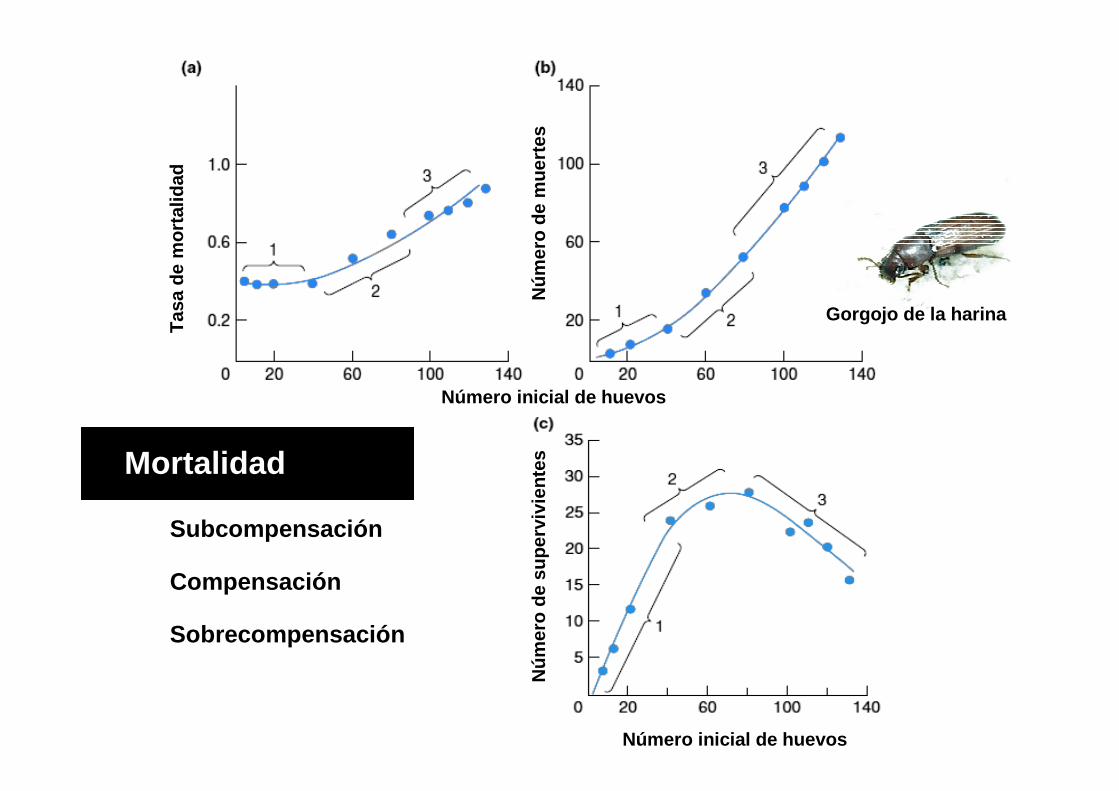
Gorgojo de la harina
Mortalidad
Subcompensación
Compensación
Sobrecompensación
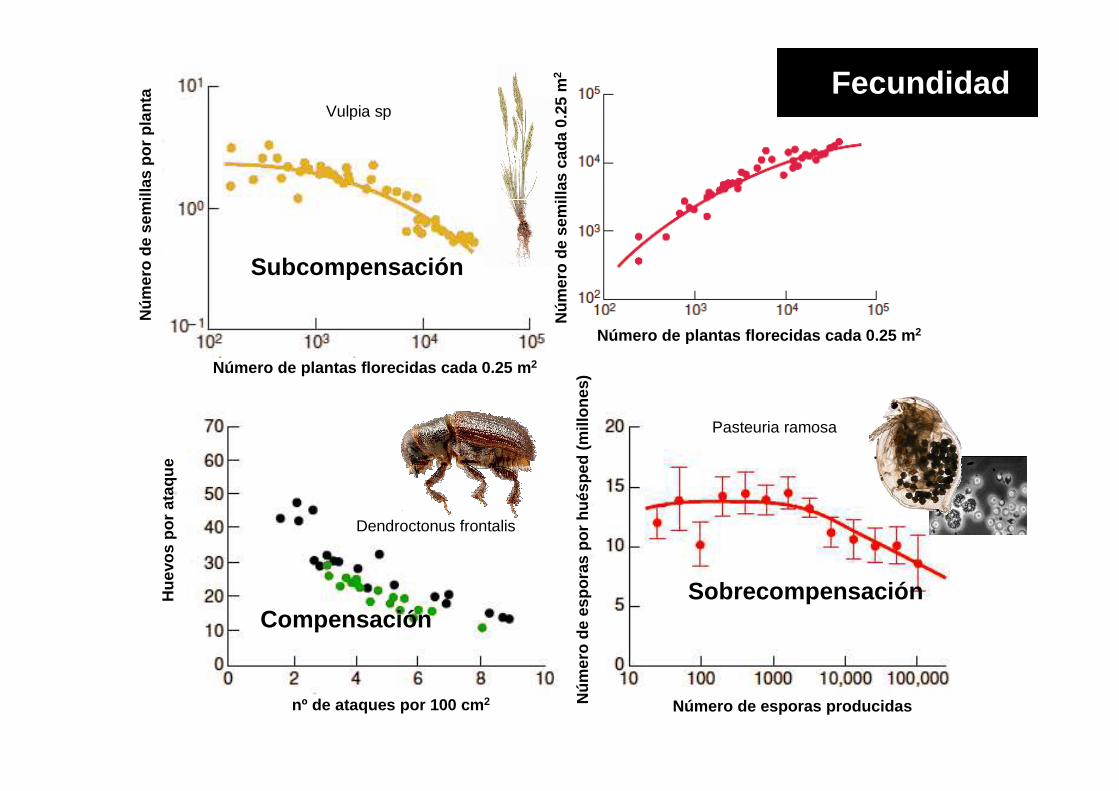
Vulpia sp
Dendroctonus frontalis
Pasteuria ramosa
Número de plantas florecidas cada 0.25 m 2
Núm
ero
de s
emill
as p
or p
lant
aH
uevo
s po
r at
aque
nº de ataques por 100 cm 2 Número de esporas producidasNúm
ero
de e
spor
as p
or h
uésp
ed (
mill
ones
)
Subcompensación
CompensaciónSobrecompensación
Núm
ero
de s
emill
as c
ada
0.25
m2
Número de plantas florecidas cada 0.25 m 2
Fecundidad
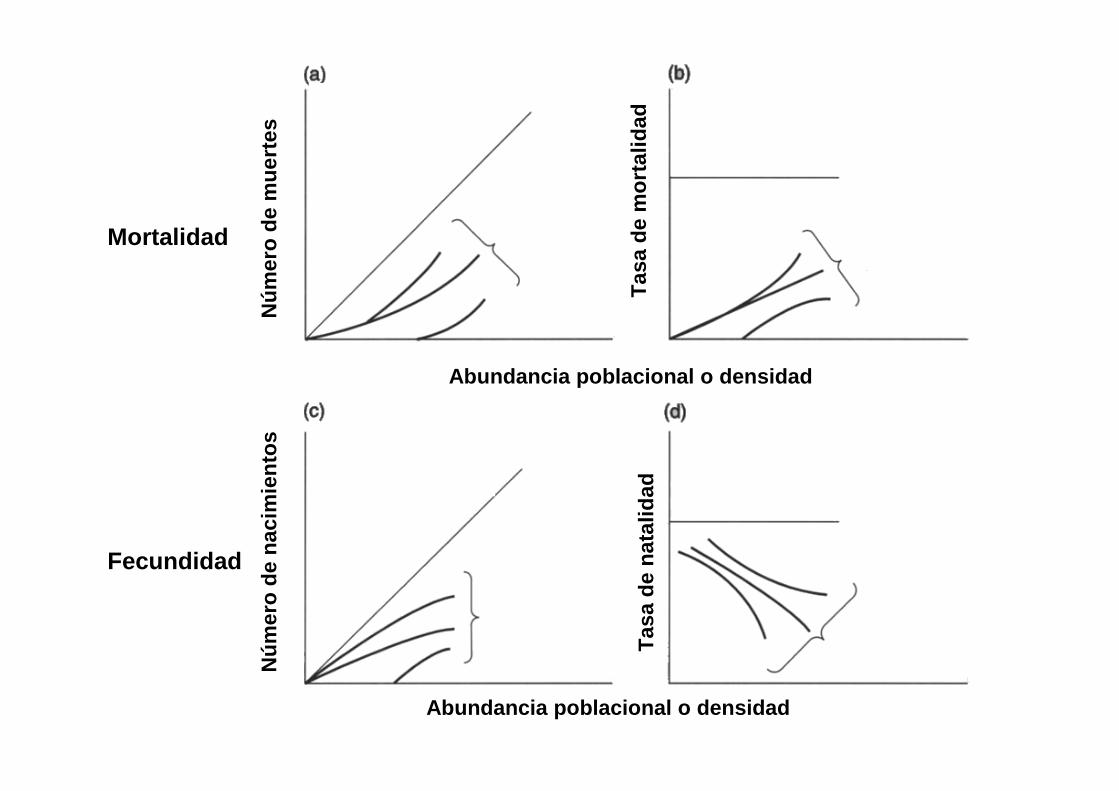
Mortalidad
Fecundidad
Núm
ero
de n
acim
ient
osN
úmer
o de
mue
rtes
Abundancia poblacional o densidad
Abundancia poblacional o densidad
Tas
a de
mor
talid
adT
asa
de n
atal
idad
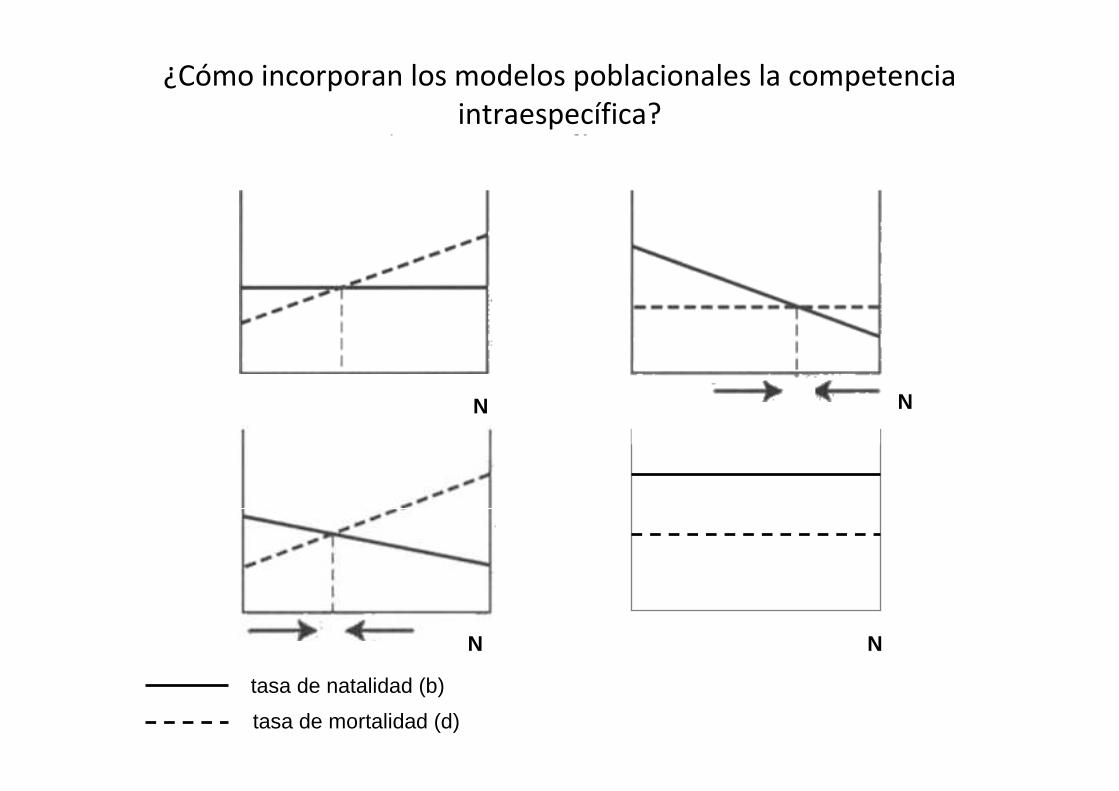
N
N
N
N
tasa de natalidad (b)
tasa de mortalidad (d)
¿Cómo incorporan los modelos poblacionales la competencia
intraespecífica?

MODELO DE CRECIMIENTO POBLACIONAL LIMITADO
�Cada individuo necesita una cantidad mínima l para reemplazarse a sí mismo.
�La cantidad limitada de individuos en el ambiente L/l = N* N de equilibrio (*) = Tamaño de la población en equilibrio (Crecimiento cero).
�El ambiente provee una cantidad limitada de recursos: L
�Si N = N* se dice que la población alcanzó su “K”
K= capacidad de carga o portadora de la población en un dado ambiente: Número de individuos de una población en equilibrio que puede soportar un ambiente.

�Si N x l < L, cada individuo puede producir más de un descendiente;
�Si N x l > L, cada individuo dejará menos de un descendiente.
�Si N x l = L, cada individuo dejará 1 descendiente.
La competencia intraespecífica tiende a
regular el tamaño poblacional.
Tiende a mantener el N dentro de ciertos límites.
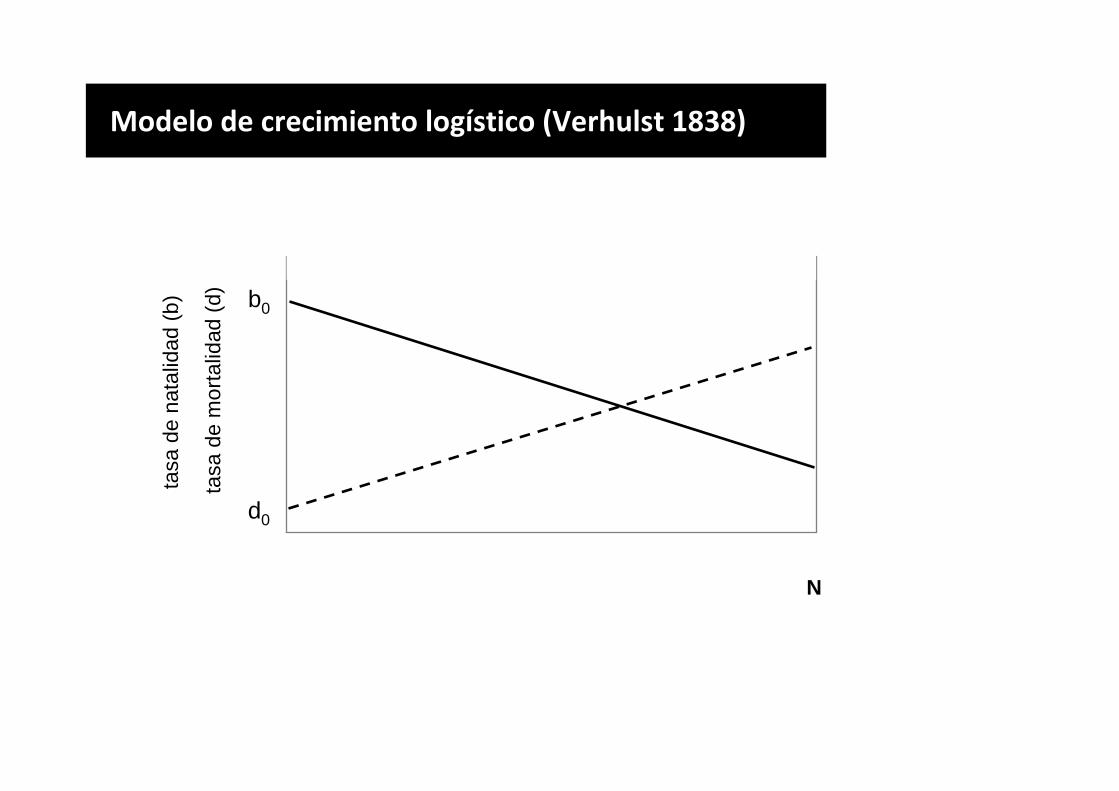
N
tasa
de
nata
lidad
(b)
tasa
de
mor
talid
ad (
d)
Modelo de crecimiento logístico (Verhulst 1838)
b0
d0
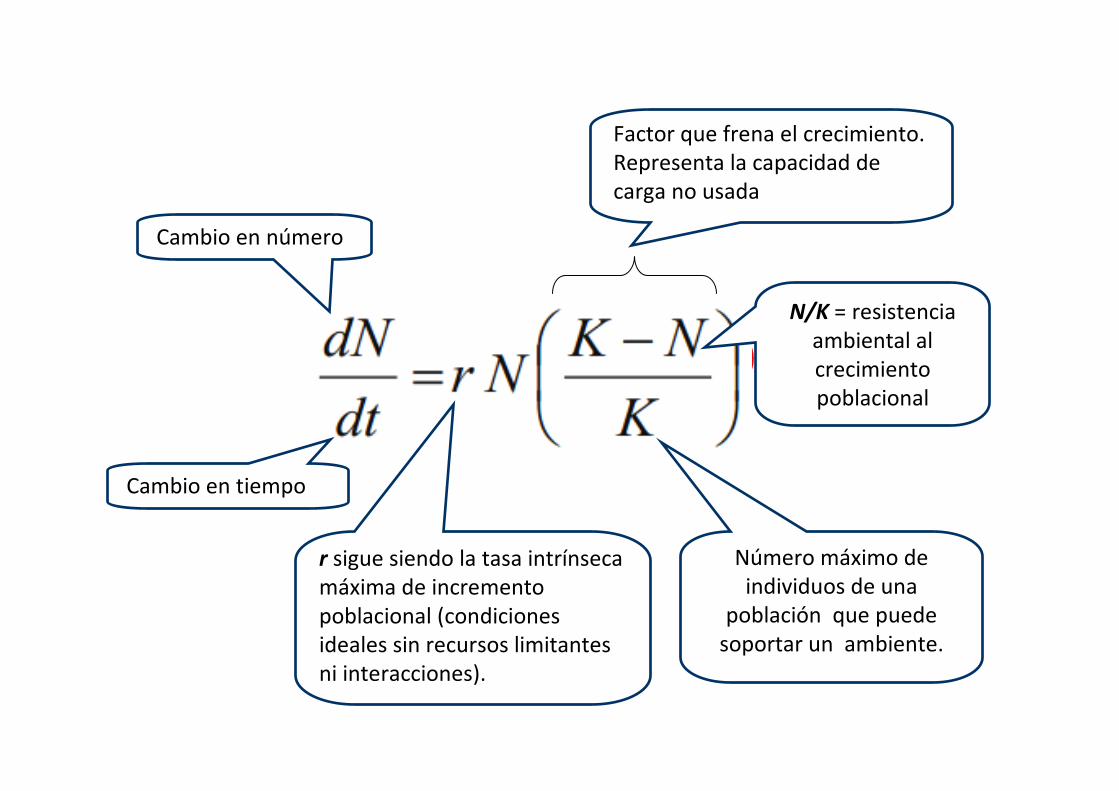
N/K = resistencia
ambiental al
crecimiento
poblacional
Factor que frena el crecimiento.
Representa la capacidad de
carga no usada
Cambio en tiempo
Cambio en número
r sigue siendo la tasa intrínseca
máxima de incremento
poblacional (condiciones
ideales sin recursos limitantes
ni interacciones).
Número máximo de
individuos de una
población que puede
soportar un ambiente.
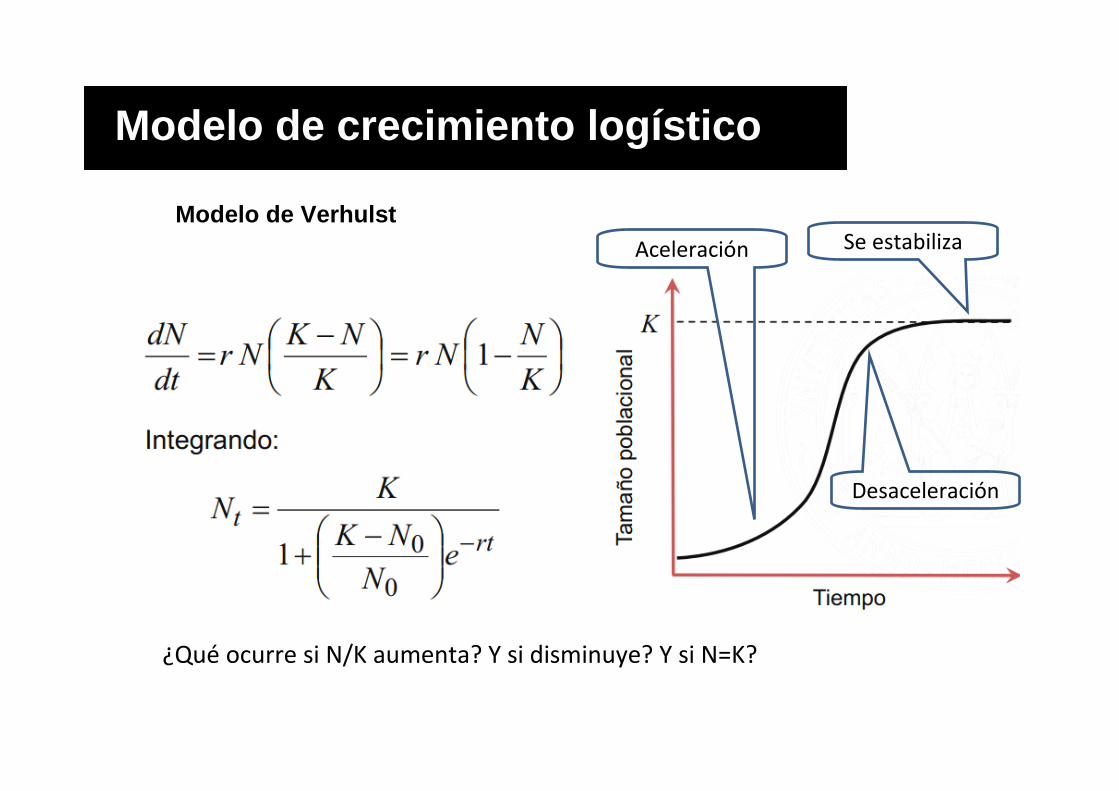
Modelo de crecimiento log ístico
¿Qué ocurre si N/K aumenta? Y si disminuye? Y si N=K?
Desaceleración
Aceleración Se estabiliza
Modelo de Verhulst
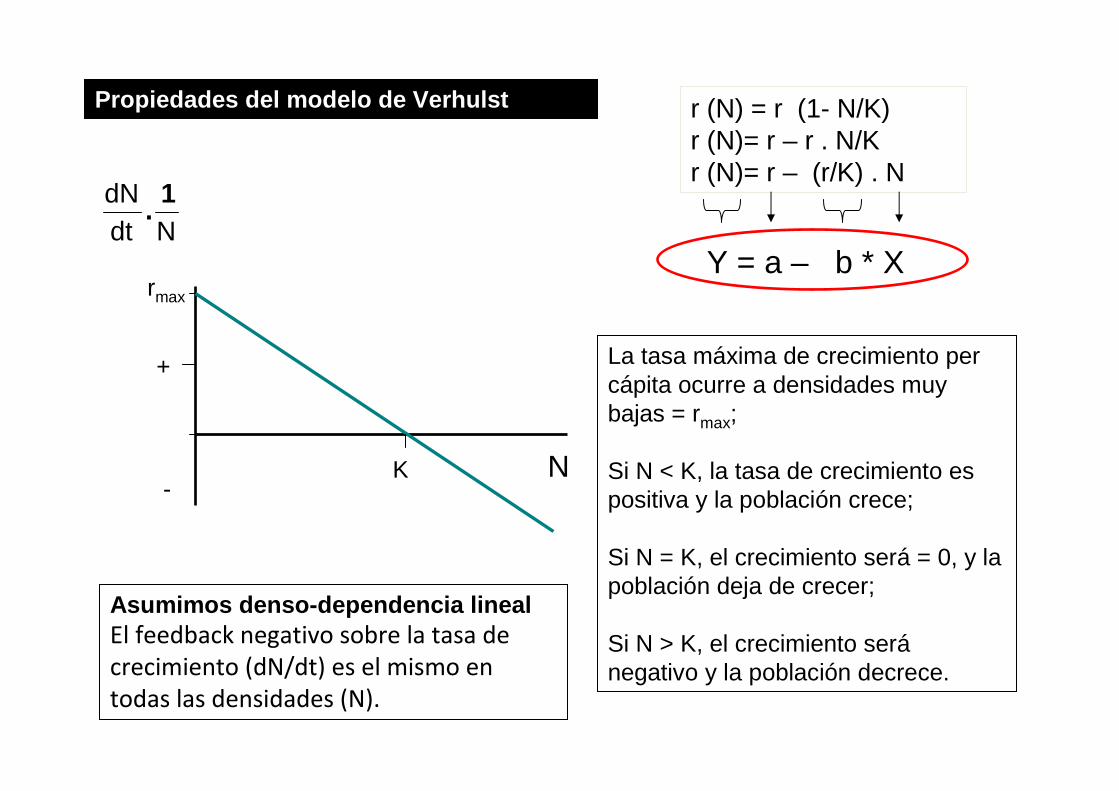
Propiedades del modelo de Verhulst
rmax
+
-K
NdtdN 1
.
N
r (N) = r (1- N/K)r (N)= r – r . N/Kr (N)= r – (r/K) . N
Y = a – b * X
La tasa máxima de crecimiento percápita ocurre a densidades muy bajas = rmax;
Si N < K, la tasa de crecimiento es positiva y la población crece;
Si N = K, el crecimiento será = 0, y la población deja de crecer;
Si N > K, el crecimiento seránegativo y la población decrece.
Asumimos denso-dependencia linealEl feedback negativo sobre la tasa de
crecimiento (dN/dt) es el mismo en
todas las densidades (N).
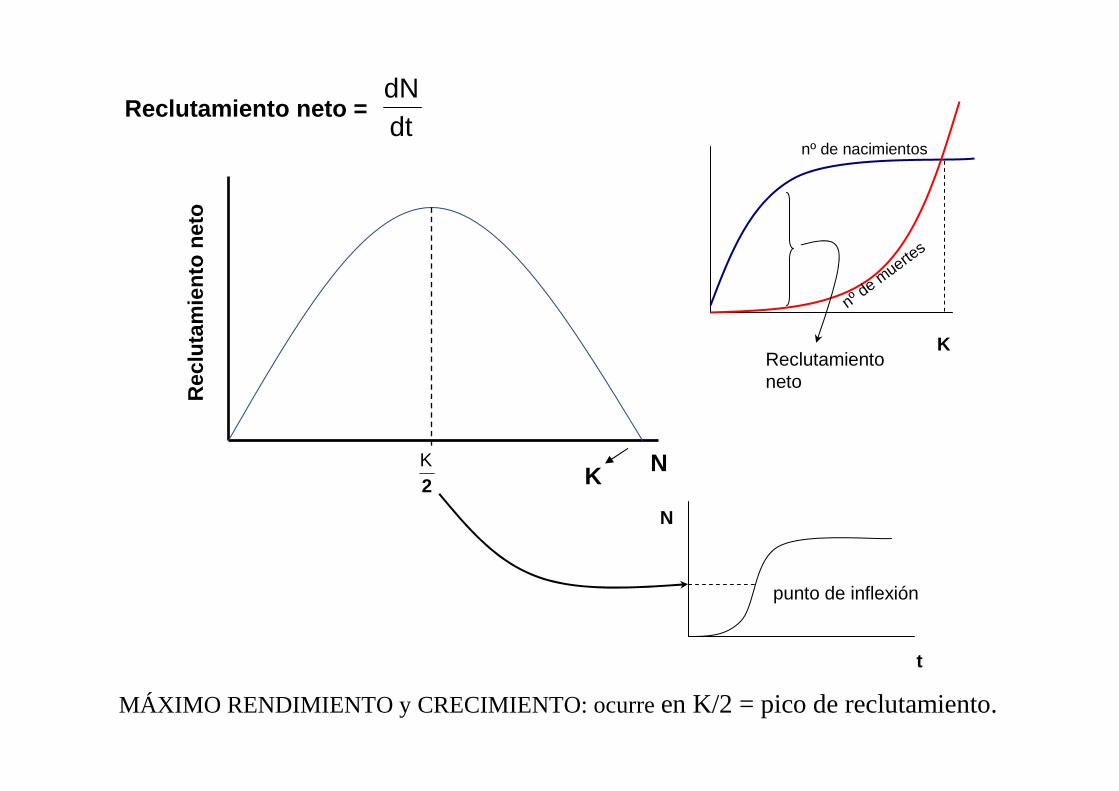
N
Rec
luta
mie
nto
neto
Reclutamiento neto = dtdN
K2K
KReclutamiento neto
nº de nacimientos
nº de muerte
s
MÁXIMO RENDIMIENTO y CRECIMIENTO: ocurre en K/2 = pico de reclutamiento.
N
t
punto de inflexión
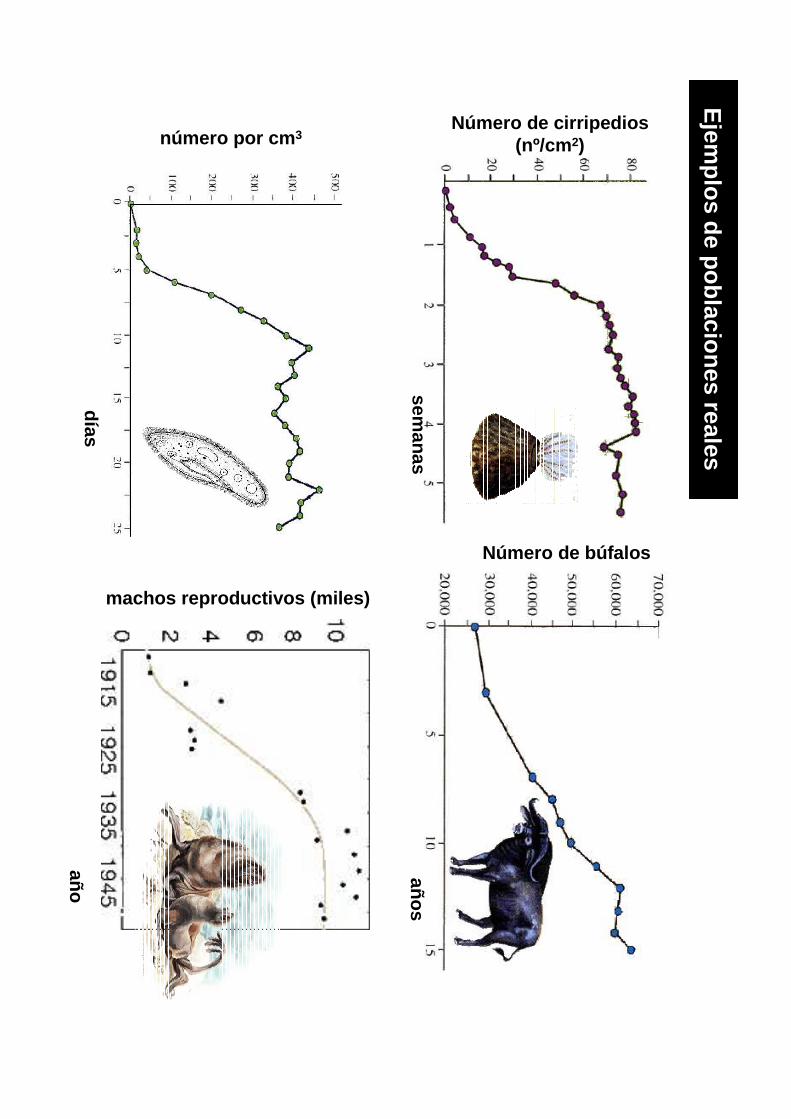
Número de cirripedios(nº/cm 2)
semanas
Número de búfalos
años
año
machos reproductivos (miles)
número por cm 3
días
Ejem
plos de poblaciones reales

�La disponibilidad de recursos impone un límite al crecimiento
�Las tasas de natalidad y/o mortalidad son dependientes de la densidad
�La tasa per capita de crecimiento poblacional disminuye con la densidad
�La tasa de reclutamiento neto muestra una curva parabólica (en u invertida)con la densidad
El crecimiento poblacional sigue un modelo logístico cuando:
Resumiendo

Supuestos del modelo crecimiento logístico
• No hay migración=población cerrada
• No hay variación genética
• No hay estructura de edades
• No hay tiempo de retardo
• Los recursos son limitados
• Capacidad de carga constante (modelo clásico de Verhulst)
• Denso-dependencia lineal de la tasa de crecimiento per capita con N.
Asume que cada individuo que se adiciona a la población produce un
decrecimiento en la tasa de crecimiento poblacional per capita.
• Las condiciones ambientales son constantes
• Hay varios supuestos que se pueden relajar para ganar más realismo.
Dado que deriva del modelo exponencial comparte algunos supuestos
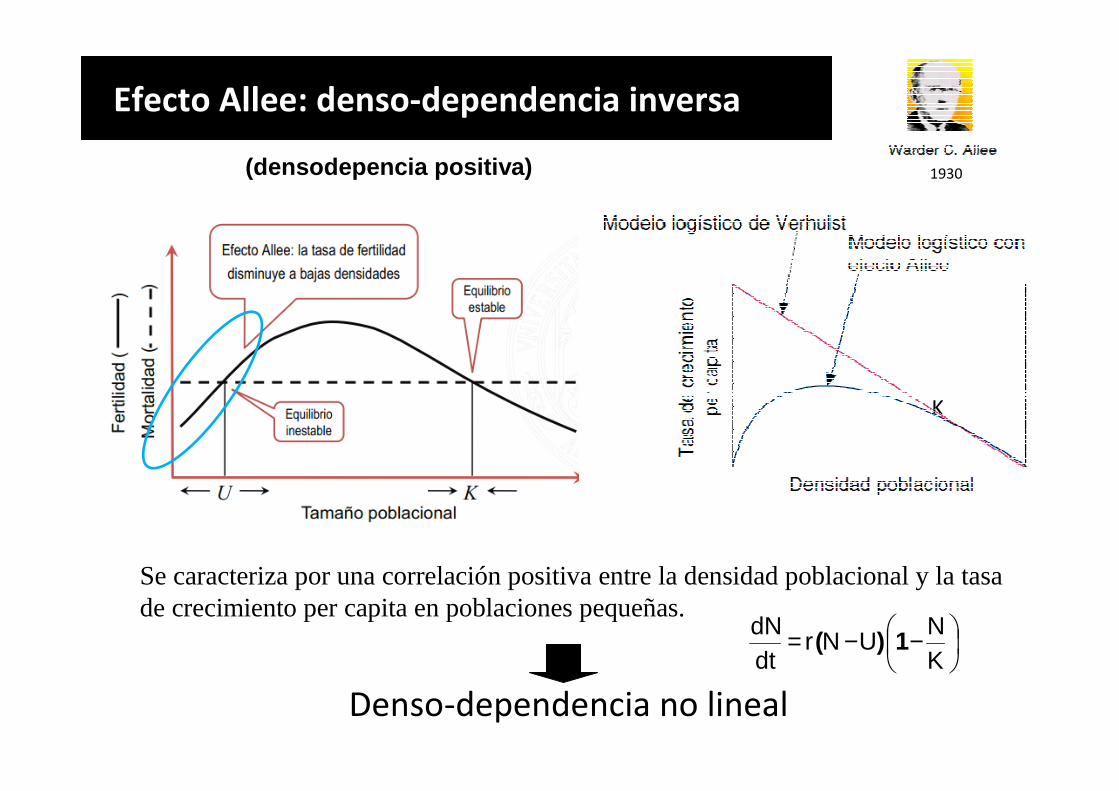
Denso-dependencia no lineal
1932
1930
Se caracteriza por una correlación positiva entre la densidad poblacional y la tasa de crecimiento per capita en poblaciones pequeñas.
K
Efecto Allee: denso-dependencia inversa
(densodepencia positiva)
−−=KN
UNrdtdN
1)(
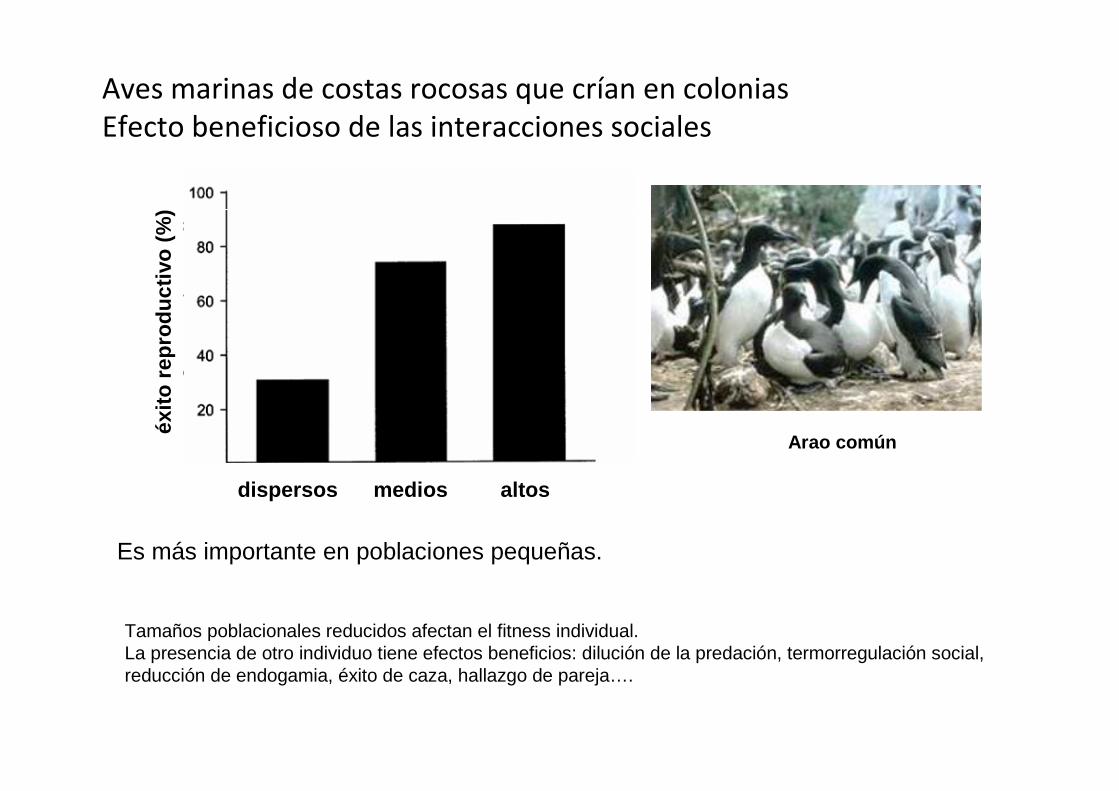
Es más importante en poblaciones pequeñas.
Aves marinas de costas rocosas que crían en colonias
Efecto beneficioso de las interacciones sociales
Tamaños poblacionales reducidos afectan el fitness individual.La presencia de otro individuo tiene efectos beneficios: dilución de la predación, termorregulación social, reducción de endogamia, éxito de caza, hallazgo de pareja….
Arao común
dispersos medios altos
éxito
rep
rodu
ctiv
o (%
)

Tamaño de la colonia (cm 2)
Mor
talid
ad a
nual
(%
)
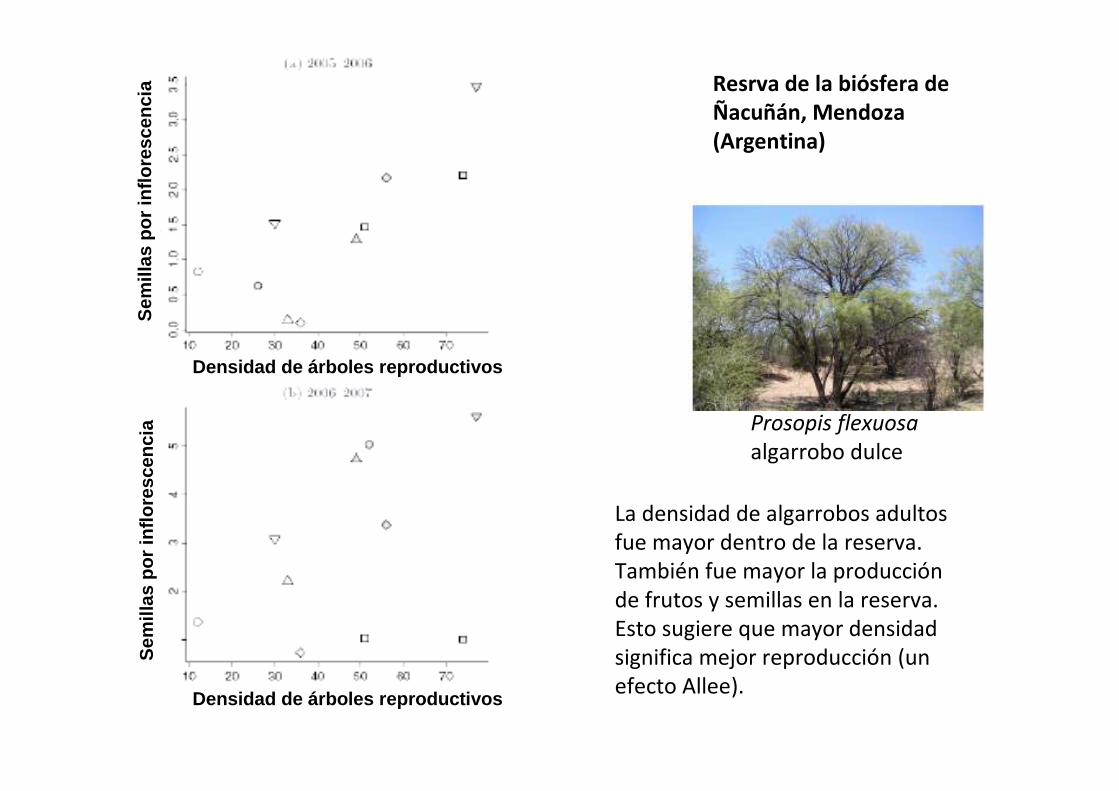
Prosopis flexuosa
algarrobo dulce
Resrva de la biósfera de Ñacuñán, Mendoza (Argentina)
La densidad de algarrobos adultos
fue mayor dentro de la reserva.
También fue mayor la producción
de frutos y semillas en la reserva.
Esto sugiere que mayor densidad
significa mejor reproducción (un
efecto Allee).
Sem
illas
por
inflo
resc
enci
a
Densidad de árboles reproductivos
Sem
illas
por
inflo
resc
enci
a
Densidad de árboles reproductivos

Población Mínima Viable (MVP)
Es el tamaño mínimo posible al cual una población natural puede existir sin extinguirse como consecuencia de desastres naturales, o estocasticidaddemográfica, ambiental o genética.
El tamaño de una población mínima viable es generalmente estimado (utilizando Análisis de Viabilidad Poblacional) como el tamaño necesario para obtener una probabilidad de supervivencia de la población del 90-95% en 100-1000 años.
Alternativamente puede inferirse a partir de datos de poblaciones de distinto tamaño que se hayan extinguido o que persistan.
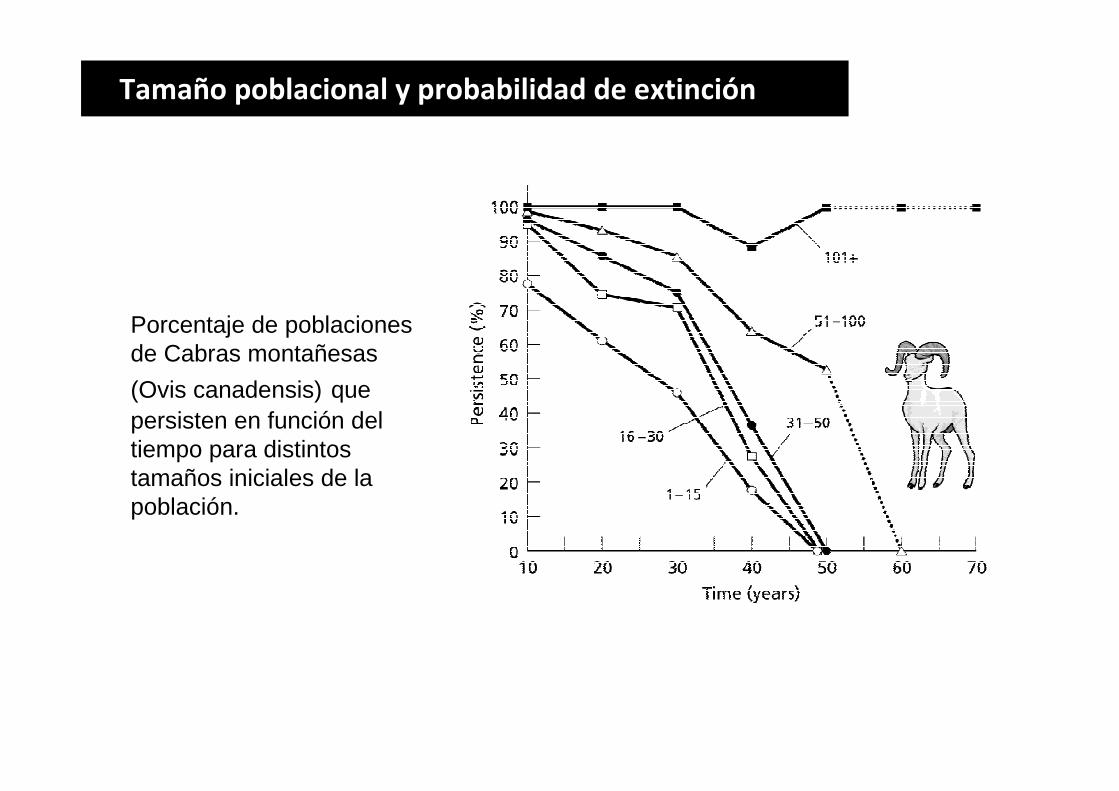
Tamaño poblacional y probabilidad de extinción
Porcentaje de poblaciones de Cabras montañesas
(Ovis canadensis) que persisten en función del tiempo para distintos tamaños iniciales de la población.
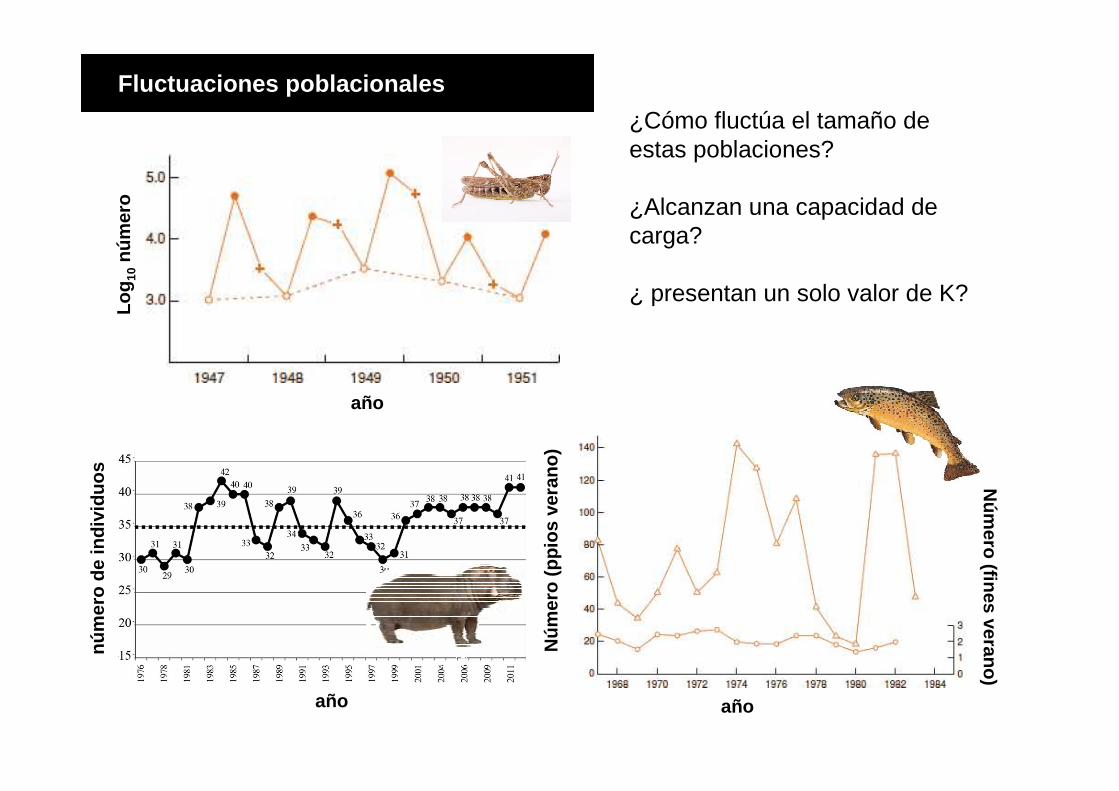
Log
10nú
mer
o
año
Núm
ero
(ppi
osve
rano
)
Núm
ero (fines verano)
año
¿Cómo fluctúa el tamaño de estas poblaciones?
¿Alcanzan una capacidad de carga?
¿ presentan un solo valor de K?
año
núm
ero
de in
divi
duos
Fluctuaciones poblacionales
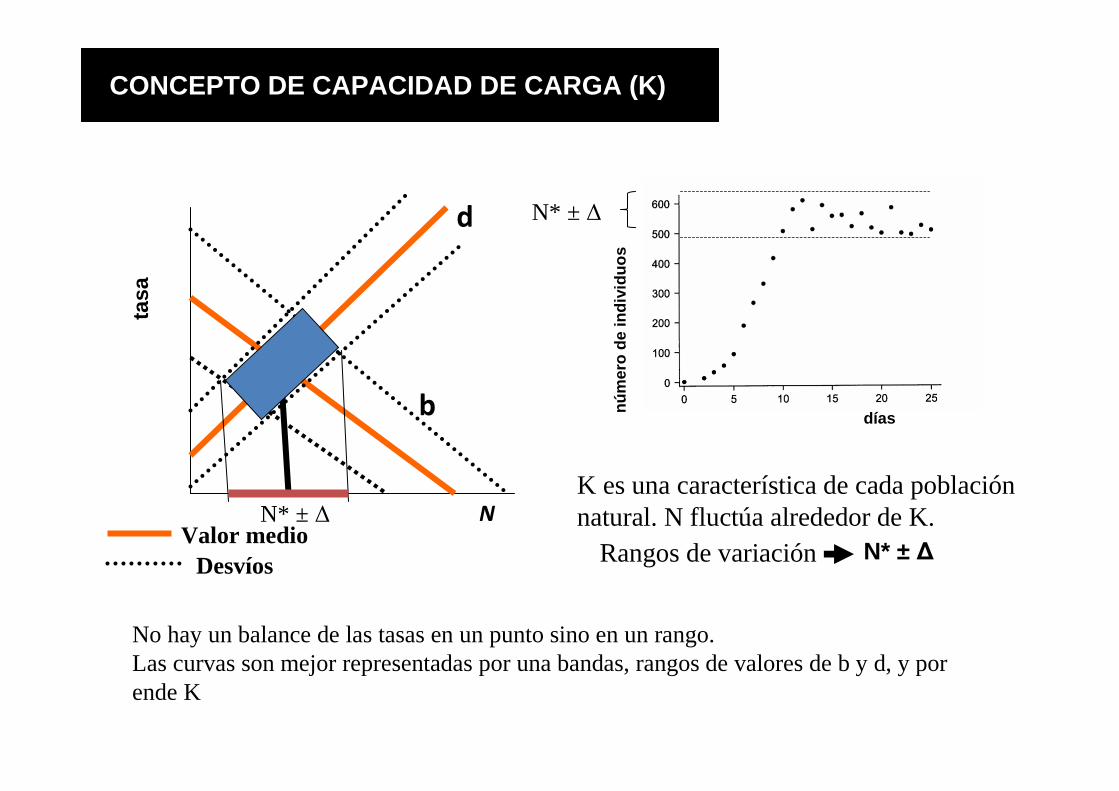
b
d
Valor medioDesvíos
N
N* ± ∆Rangos de variación
No hay un balance de las tasas en un punto sino en un rango. Las curvas son mejor representadas por una bandas, rangos de valores de b y d, y por ende K
K es una característica de cada población natural. N fluctúa alrededor de K.
N* ± ∆
CONCEPTO DE CAPACIDAD DE CARGA (K)
N* ± ∆
tasa
días
núm
ero
de in
divi
duos

Modelos estocásticos
año
año
Densidad relativa Densidad relativa
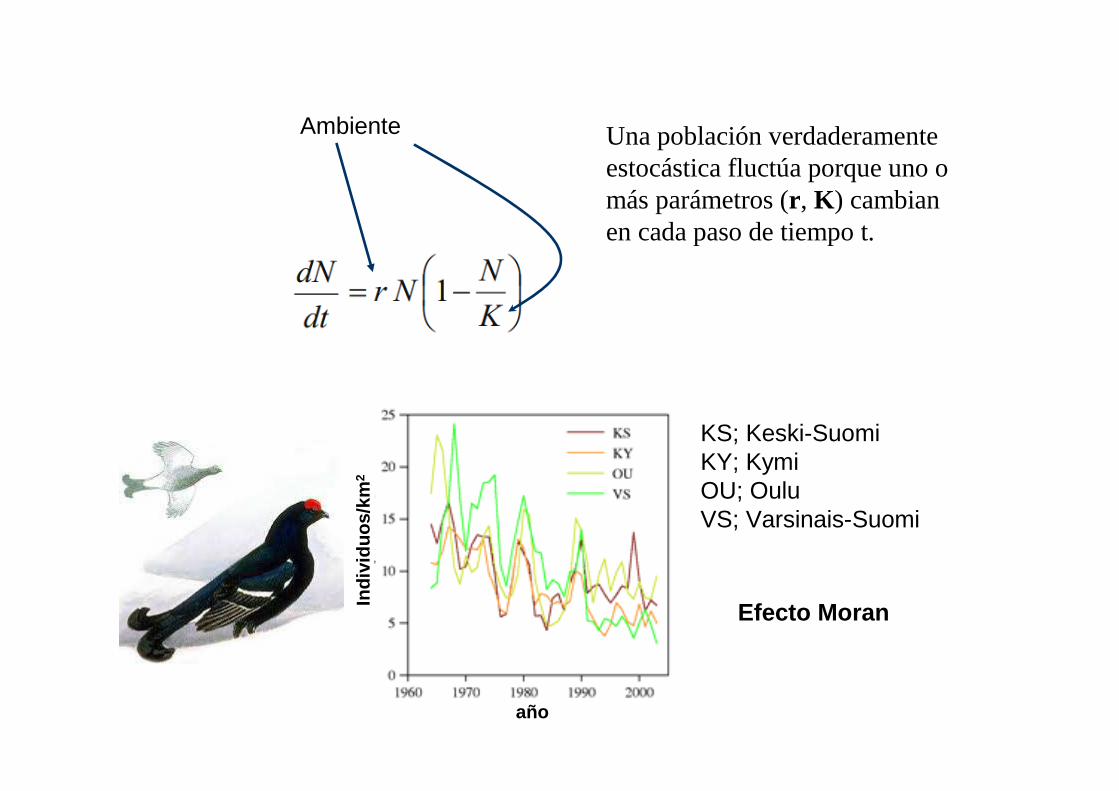
Ambiente Una población verdaderamente estocástica fluctúa porque uno o más parámetros (r , K ) cambian en cada paso de tiempo t.
KS; Keski-SuomiKY; KymiOU; OuluVS; Varsinais-Suomi
Efecto Moran
año
Indi
vidu
os/k
m2
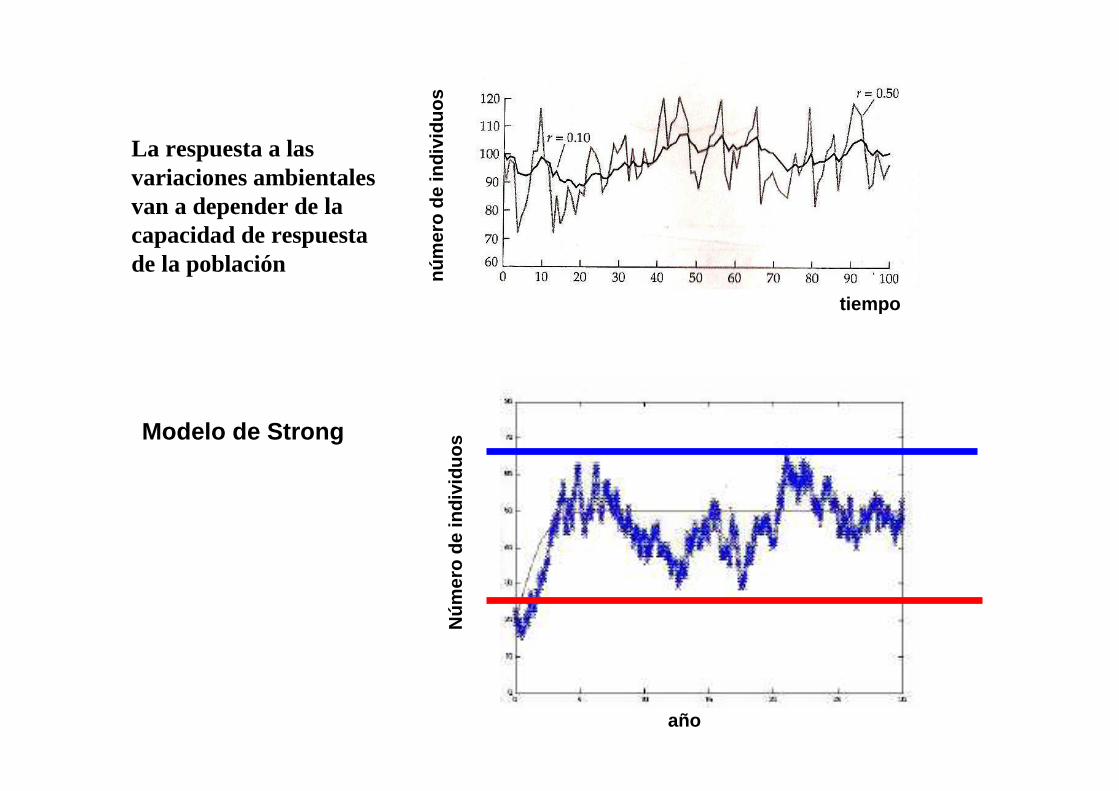
Modelo de Strong
año
núm
ero
de in
divi
duos
Núm
ero
de in
divi
duos
tiempo
La respuesta a las variaciones ambientales van a depender de la capacidad de respuesta de la población
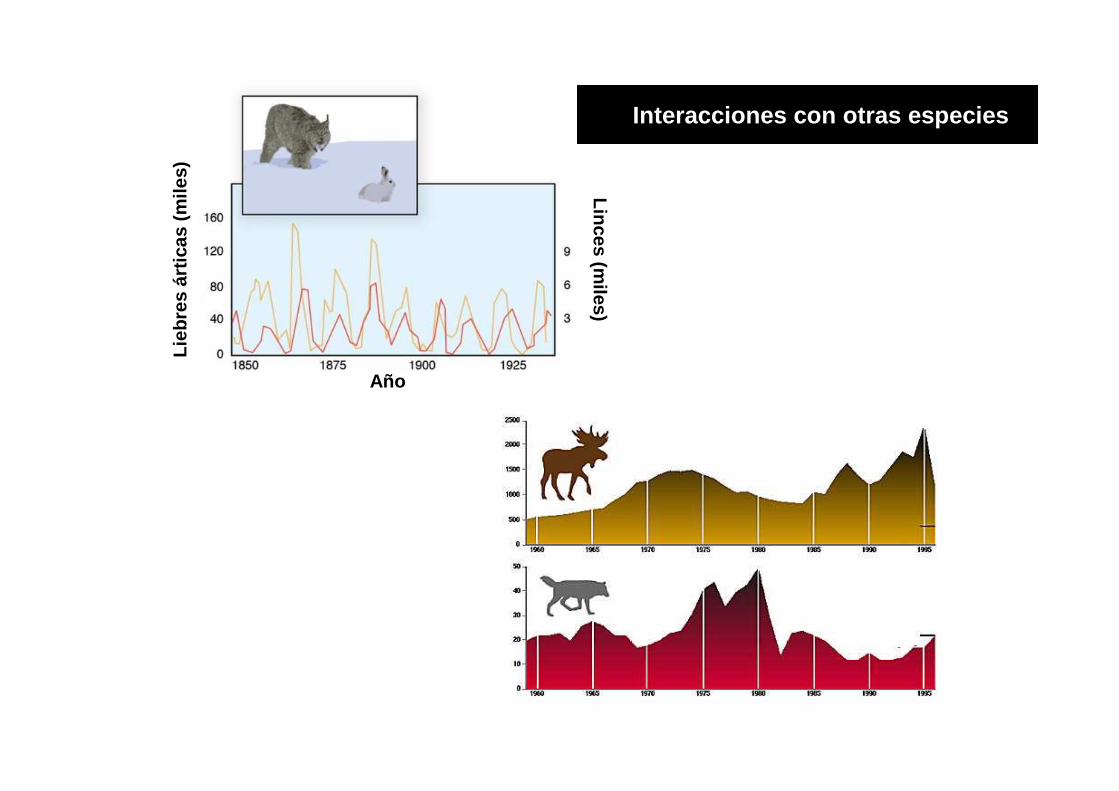
Linces (miles)
Lieb
res
ártic
as (
mile
s)
Año
Interacciones con otras especies

Los efectos de la densidad no son inmediatos:
MODELOS CON TIEMPO DE RETARDO
dNt /dt= r Nt (K-Nt)/K
No hay tiempo de retardo dNt/dt disminuye inmediatamente con N
dNt /dt= r Nt (K-Nt-τ)/K
Hay tiempo de retardo τ r (N) disminuye de acuerdo a N en t-τ
Los efectos de la densidad son inmediatos:
Cuando hay variabilidad estacional de recursos, respuesta predador presa….
Los individuos no ajustan en forma inmediata su crecimiento y reproducción cuando los niveles de recursos cambian y este retraso afecta la dinámica poblacional.
Hay un tiempo de retardo T entre el cambio en el tamaño de la población y sus efectos sobre la tasa de crecimiento.
Una población se incrementa en otoño y hasta la primavera no se expresaran los efectos denso-
dependiente, cuando las hembras dejen su descendencia.
Poco realista
(ejemplo: estacionalidad de los recursos, sistemas predador-presa)
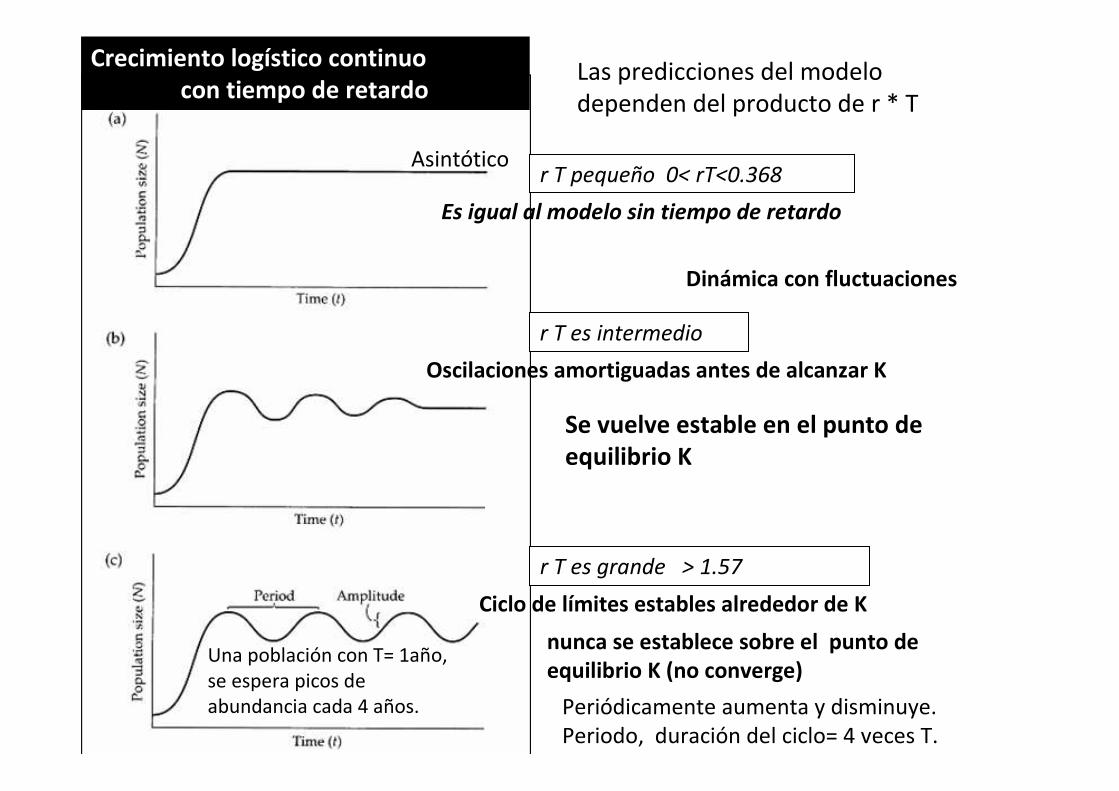
Crecimiento logístico continuocon tiempo de retardo
r T pequeño 0< rT<0.368
r T es intermedio
Oscilaciones amortiguadas antes de alcanzar K
Es igual al modelo sin tiempo de retardo
r T es grande > 1.57
Ciclo de límites estables alrededor de K
Se vuelve estable en el punto de equilibrio K
nunca se establece sobre el punto de equilibrio K (no converge)
Periódicamente aumenta y disminuye.
Periodo, duración del ciclo= 4 veces T.
Asintótico
Una población con T= 1año,
se espera picos de
abundancia cada 4 años.
Dinámica con fluctuaciones
Las predicciones del modelo
dependen del producto de r * T
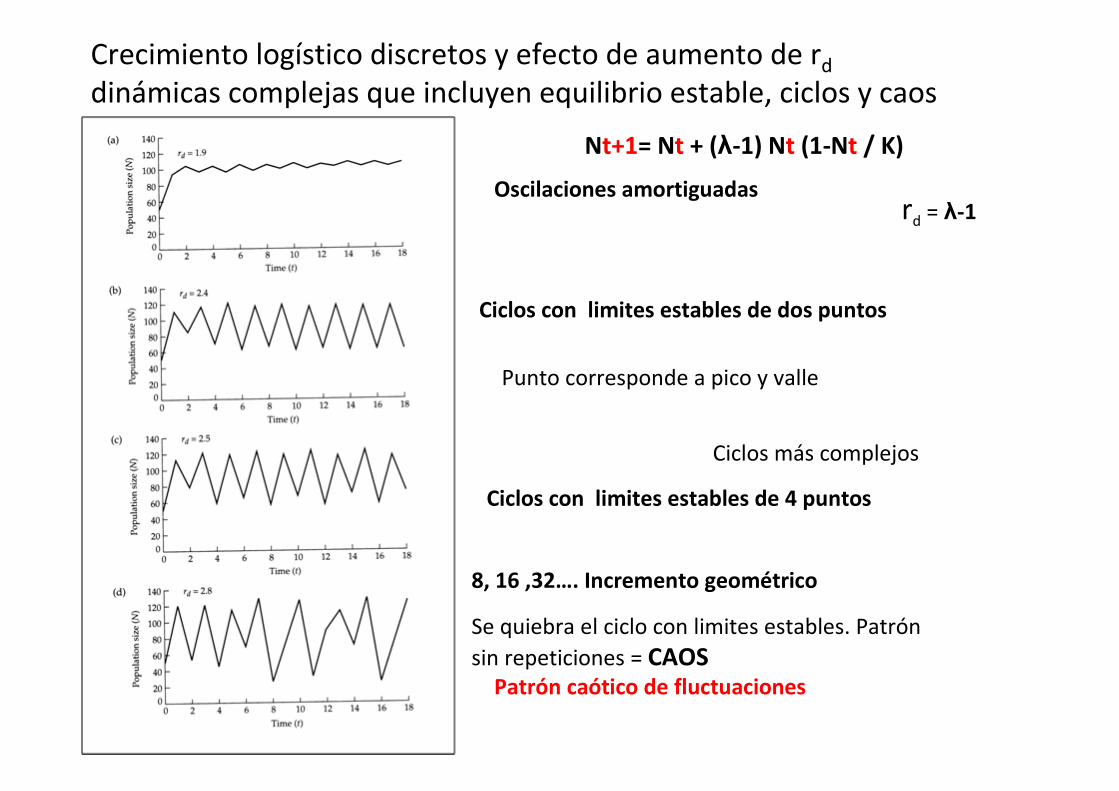
Nt+1= Nt + (λ-1) Nt (1-Nt / K)
Oscilaciones amortiguadas
Ciclos con limites estables de dos puntos
Ciclos con limites estables de 4 puntos
Patrón caótico de fluctuaciones
8, 16 ,32…. Incremento geométrico
Se quiebra el ciclo con limites estables. Patrón
sin repeticiones = CAOS
Crecimiento logístico discretos y efecto de aumento de rd
dinámicas complejas que incluyen equilibrio estable, ciclos y caos
rd = λ-1
Ciclos más complejos
Punto corresponde a pico y valle
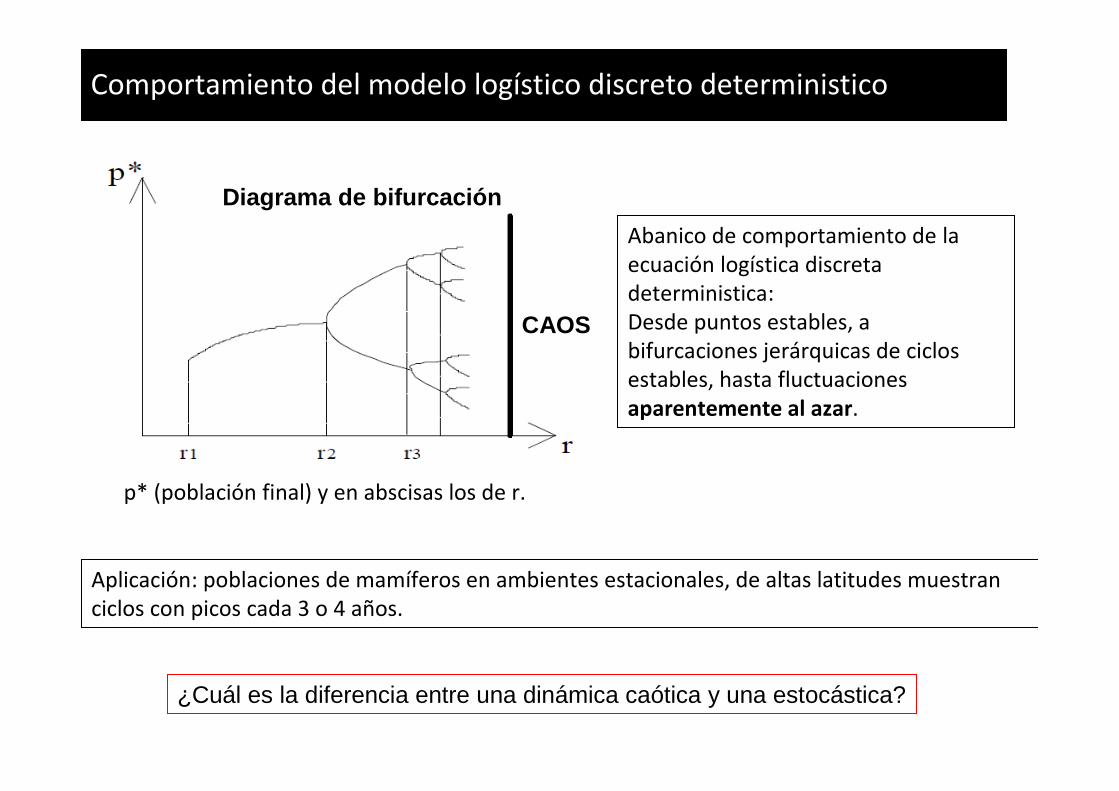
Comportamiento del modelo logístico discreto deterministico
p* (población final) y en abscisas los de r.
Abanico de comportamiento de la
ecuación logística discreta
deterministica:
Desde puntos estables, a
bifurcaciones jerárquicas de ciclos
estables, hasta fluctuaciones
aparentemente al azar.
Aplicación: poblaciones de mamíferos en ambientes estacionales, de altas latitudes muestran
ciclos con picos cada 3 o 4 años.
Diagrama de bifurcación
CAOS
¿Cuál es la diferencia entre una dinámica caótica y una estocástica?
Regulación y limitación poblacional
Factores limitantes:
• Factores bióticos : enfermedades, parasitosis, depredación, competencia• Factores abióticos : clima, inundación, temperatura extremas, heladas, salinidad
Factores denso-dependientesFactores denso-independientes
Ambos modifican las tasas de natalidad y mortalidad, pero ¿regulan?

Población humana

Densidad de población humana
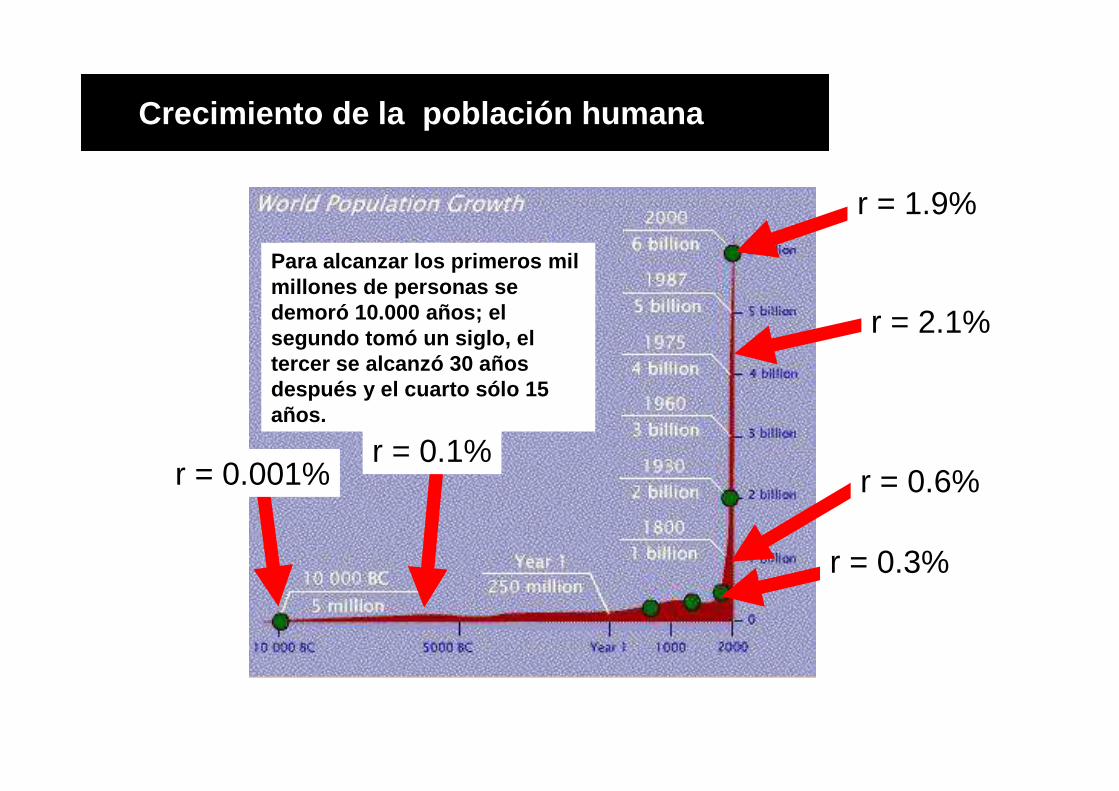
Crecimiento de la población humana
Para alcanzar los primeros mil millones de personas se demoró 10.000 años; el segundo tomó un siglo, el tercer se alcanzó 30 años después y el cuarto sólo 15 años.
r = 0.001%r = 0.1%
r = 0.3%
r = 0.6%
r = 2.1%
r = 1.9%

En base a datos directos e indirectos (arqueología, antropología) se estima que la
tasa de crecimiento (r) de la población humana aumentó progresivamente en
últimos 10.000 años.
Hasta hace 10.000 años (neolítico, pasaje de cazadores recolectores a agricultores):
N = 10 millones, r = 0.001% (r = tasa instantánea de crecimiento = natalidad-
mortalidad).
Con revolución agrícola se pasa rápidamente a N = 50 millones y r = 0.1% (aumenta
100 veces). Tiempo de duplicación poblacional (=70/r) = 700 años.
En siglo XVII: N = 500 millones, r = 0.3%
En siglo XIX: N = 1.600 millones y r = 0.6%.
Entre 1970 y 1990 aumenta de 3.600 a 5.400 millones (r = 2.1%, tiempo de
duplicación 33 años).
La población actual es aproximadamente 6.600 millones con un r = 1.9%
(declinando).
año
Tasa anual de crecim
iento
Pob
laci
ón g
loba
l (m
il m
illon
es)
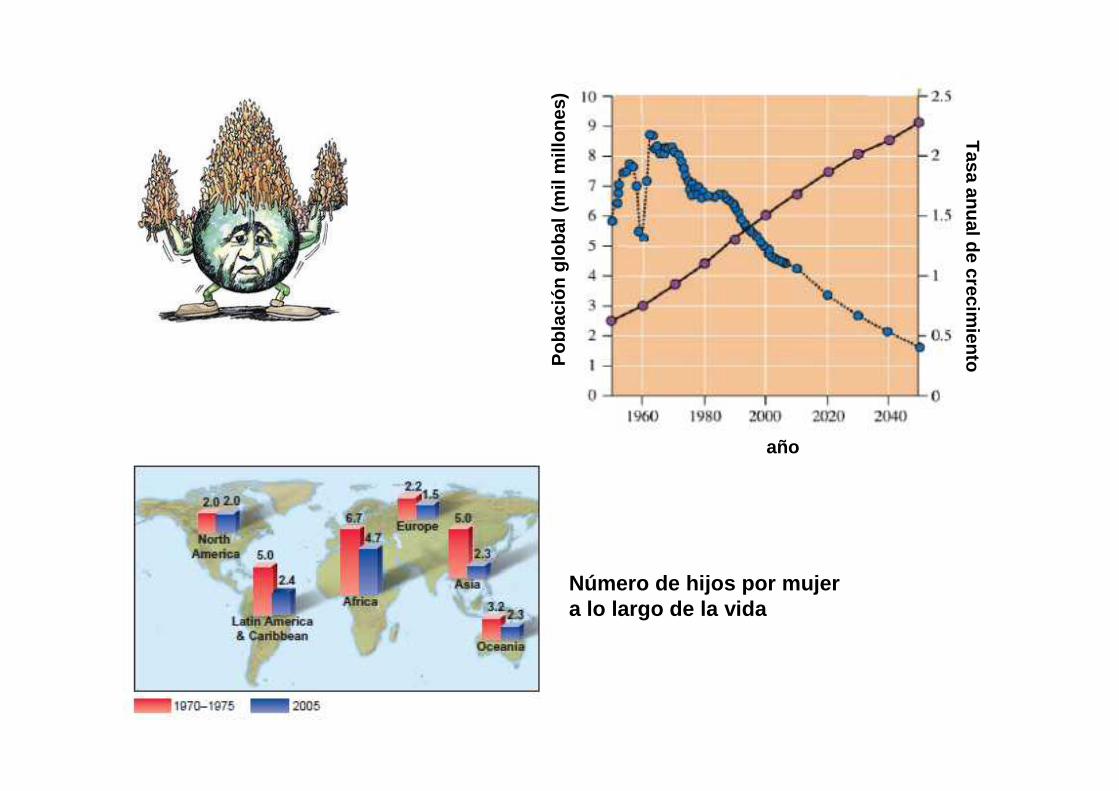
Número de hijos por mujer a lo largo de la vida

• Sociedades primitivas: tasas de natalidad y mortalidad muy altas, población estable.
• Sociedades modernas: tasas de natalidad y mortalidad muy bajas, población estable.
Al período de cambio de sociedad primitiva a moderna se llama “transición demográfica”.
MODELO TRANSICION DEMOGRAFICA
La transición demográfica es un cambio crítico en la tasa de crecimiento y en la estructura de edad.
Tasa bruta de
mortalidad
Tasa bruta de
natalidad
año
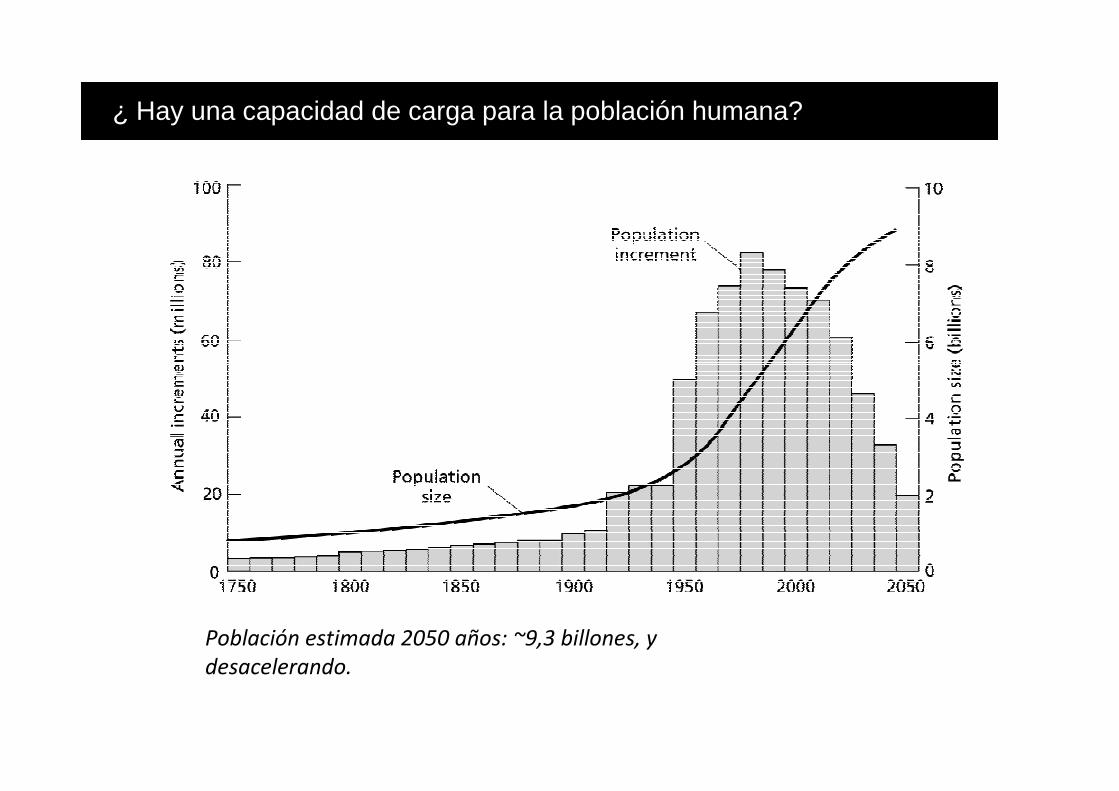
Población estimada 2050 años: ~9,3 billones, y
desacelerando.
¿ Hay una capacidad de carga para la población humana?
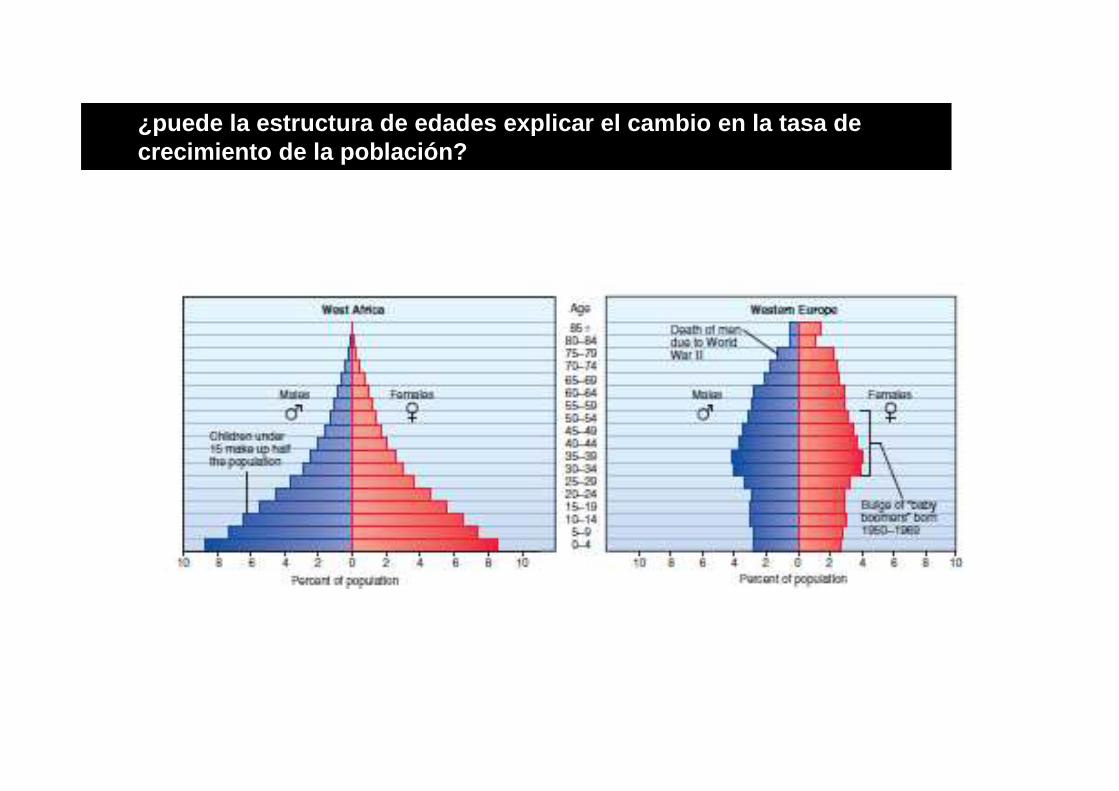
¿puede la estructura de edades explicar el cambio e n la tasa de crecimiento de la población?


















