
Page 1 Lípidos ULEAM 2010. Page 2 Generalidades Son un conjunto de moléculas orgánicas,...
122
Page 1 Lípidos ULEAM 2010
-
Upload
vasco-lovera -
Category
Documents
-
view
6 -
download
3
Transcript of Page 1 Lípidos ULEAM 2010. Page 2 Generalidades Son un conjunto de moléculas orgánicas,...

Page 1
Lípidos
ULEAM2010

Page 2
Generalidades
• Son un conjunto de moléculas orgánicas, biomoléculas, compuestas principalmente por C, H < O, aunque también pueden contener P, S y N.
• Característica principal el ser insolubles en agua y sí en solventes orgánicos (alcohol, benceno y cloroformo).
• A los lípidos se les llama incorrectamente grasas, ya que las grasas son sólo un tipo de lípidos procedentes de animales.

Page 3
Generalidades
• Los lípidos cumplen funciones diversas en los organismos vivientes, entre ellas la de reserva energética (triglicéridos), la estructural (fosfolípidos de las bicapas) y la reguladora (esteroides).
• Los Lípidos también funcionan para el desarrollo de la Materia gris, el metabolismo y el crecimiento
Page 4

Page 5
Características generales
• Los lípidos más abundantes son las grasas, que puede ser de origen animal o vegetal.
• Unos están formados por cadenas alifáticas saturadas o insaturadas, pero algunos tienen anillos (aromáticos).
• Algunos son flexibles, mientras que otros son rígidos o semiflexibles hasta alcanzar casi una total flexibilidad molecular; algunos comparten carbonos libres y otros forman puentes de hidrógeno.

Page 6
Características generales
• Tienen algún tipo de carácter polar, además de poseer una gran parte apolar o hidrofóbico.
• Otra parte de su estructura es polar o hidrofílica y tenderá a asociarse con solventes polares como el agua; cuando una molécula tiene una región hidrófoba y otra hidrófila se dice que tiene carácter anfipático.
• La región hidrófoba de los lípidos es la que presenta solo átomos de C unidos a átomos de H.

Page 7
Características generales
• La región hidrófila es la que posee grupos polares o con cargas eléctricas, como el hidroxilo (–OH) del colesterol, el carboxilo (–COO–) de los ácidos grasos, el fosfato (–PO4
–) de los fosfolípidos, etc.
• Como la larga "cola" alifática de los ácidos grasos o los anillos de esterano del colesterol.
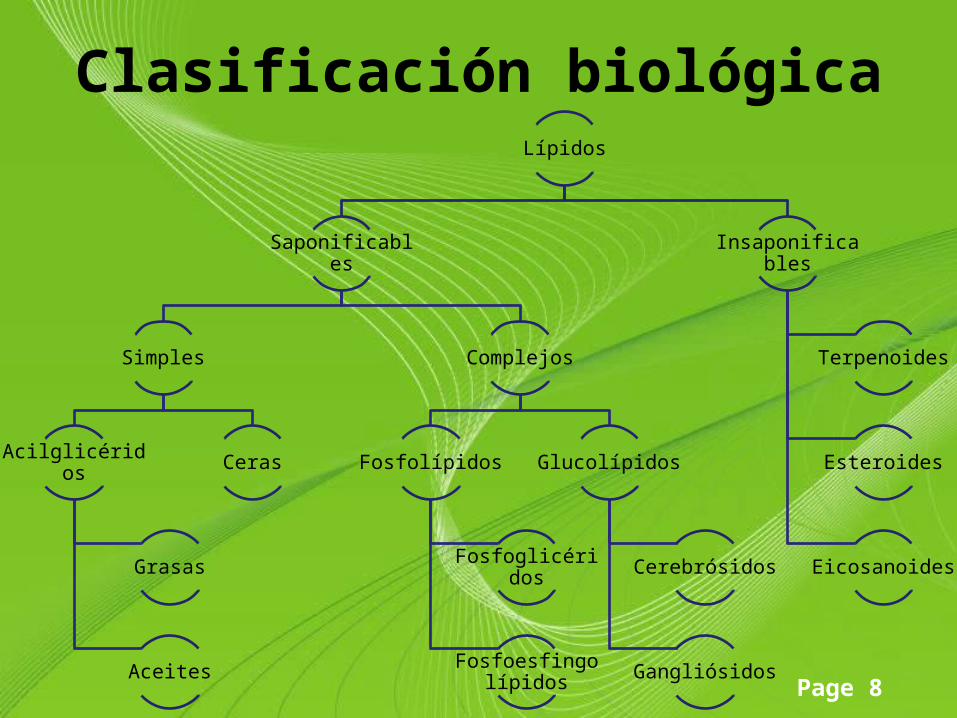
Page 8
Clasificación biológicaLípidos
Saponificables
Simples
Acilglicéridos
Grasas
Aceites
Ceras
Complejos
Fosfolípidos
Fosfoglicéridos
Fosfoesfingolípidos
Glucolípidos
Cerebrósidos
Gangliósidos
Insaponificables
Terpenoides
Esteroides
Eicosanoides

Page 9
Ácido graso• Formada por una larga cadena hidrocarbonada
lineal, de número par de átomos de carbono, en cuyo extremo hay un grupo carboxilo.
• Cada átomo de carbono se une al siguiente y al precedente por medio de un enlace covalente sencillo o doble.
• Al átomo de su extremo le quedan libres tres enlaces que son ocupados por átomos de hidrógeno (H3C-).
• Los demás átomos tienen libres dos enlaces, que son ocupados igualmente por átomos de hidrógeno ( ... -CH2-CH2-CH2- ...).

Page 10
Ácido graso• Se puede globalizar un ácido graso genérico como
R-COOH.• Los ácidos grasos forman parte de los fosfolípidos
y glucolípidos, moléculas que constituyen la bicapa lipídica de todas las membranas celulares.
• En los mamíferos la mayoría de los ácidos grasos se encuentran en forma de triglicéridos, moléculas donde los extremos carboxílico (-COOH) de tres ácidos grasos se esterifican con cada uno de los grupos hidroxilos (-OH) del glicerol (glicerina, propanotriol); los triglicéridos se almacenan en el tejido adiposo.

Page 11
Estructura química• Los ácidos grasos constan de una cadena
alquílica con un grupo carboxil (–COOH) terminal; la fórmula básica de una molécula completamente saturada es CH3–(CH2)n–COOH.
• Los ácidos grasos de los mamíferos tienen estructuras relativamente sencillas, pero los de otros organismos pueden ser muy complejos, con anillos ciclopropano o abundantes ramificaciones.

Page 12
Estructura química
Molécula de ácido esteárico, un ácido graso saturado, representada de manera esquemática.
La misma molécula con el grupo carboxilo disociado.

Page 13
Estructura química• Son frecuentes los ácidos grasos insaturados,
casi siempre de configuración cis; cuando hay más de un doble enlace por molécula, siempre están separados por un grupo metileno (–CH2–).
• Algunos animales, incluido el ser humano, también producen ácidos grasos ramificados, con uno o varios grupos metilo (–CH3) a lo largo de la cadena, como es el caso de las estructuras de ecolocalización de los cetáceos en que se hallan grandes cantidades de ácido isovalérico.

Page 14
Propiedades
• Son moléculas anfipáticas, de cadena corta, más solubles que los ácidos grasos de cadena larga porque la región hidrófoba es más corta.
• Si se colocan ácidos grasos en agua forman una capa superficial debido a su baja densidad; formarán una película con sus colas (la parte no polar) orientadas hacia arriba, fuera del agua, de manera que no quedan en contacto con la misma y la cabeza polar dentro del agua.

Page 15
Nomenclatura• Los átomos de carbono de los ácidos grasos se
numeran de dos maneras:– Números Árabigos: Empezando por el C carboxílico
(–COOH), que recibe el número 1; el C 2 es el que queda inmediatamente tras el 1 y así sucesivamente .
– Alfabeto Griego: El C carboxilico no recibe letra. Se empieza a nombrar desde el C 2, al cual se le asigna la letra α; al C 3 se le otorga la letra β (de donde proviene el término β-oxidación, ruta metabólica de degradación de los ácidos grasos en la matriz mitocondrial). el último C es el del extremo metilo (CH3–), al que se le asigna la letra ω (omega, la última letra del alfabeto griego).

Page 16
Nomenclatura• El modo oficial de denominar los ácidos grasos
consiste en el número de átomos de C seguido por dos puntos y el número de dobles enlaces; la localización de los mismos se designa por el número del átomo de C donde empieza, contando a partir del extremo carboxílico. – Así, el ácido oleico se designa 18:1(9); el número 18
nos indica el número de C, el 1 tras los dos puntos, el número de dobles enlaces y el 9 entre paréntesis que este doble enlace comienza en el 9º carbono (está entre el 9º y el 10º), contando desde el extremo –COOH.

Page 17
Nomenclatura• Otro modo de designación de los ácidos grasos
insaturados: – La posición que ocupan los dobles enlaces se indica
con respecto al último C de la cadena (el extremo CH3–), o sea, el C ω; de ahí derivan las denominaciones de ω-3, ω-6, etc.
– Un ácido graso ω-3 será el que tenga su primer doble enlace entre los carbonos 3 y 4, y un ácido graso ω-6 tendrá el primer doble enlace entre los carbonos 6 y 7, siempre a contar desde el extremo CH3–.
– Dado que el primer método empieza a contar desde el extremo –COOH y el segundo desde el extremo CH3–, puede producirse cierta confusión.

Page 18
Clasificación• Ácidos grasos saturados: Sin dobles enlaces
entre C; tienden a formar cadenas extendidas y a ser sólidos a temperatura ambiente, excepto los de cadena corta. – Cadena corta (volátiles)
• Ácido butírico (ácido butanoico)• Ácido isobutírico (ácido 2-metilpropionico)• Ácido valérico (ácido pentanoico)• Ácido isovalérico (ácido 3-metilbutanoico)
– Cadena larga: • Ácido mirístico, 14:0 (ácido tetradecanoico)• Ácido palmítico, 16:0 (ácido hexadecanoico)• Ácido esteárico, 18:0 (ácido octadecanoico)

Page 19
Clasificación• Ácidos grasos insaturados: Con dobles
enlaces entre C; suelen ser líquidos a temperatura ambiente.– Ácidos grasos monoinsaturados. Un solo doble
enlace. • Ácido oleico, 18:1(9) (ácido cis-9-octadecenoico)
– Ácidos grasos poliinsaturados. Con varios dobles enlaces. • Ácido linoleico, 18:2(9,12) (ácido cis, cis-9,12-
octadecadienoico) ( es un ácido graso esencial)• Ácido linolénico, 18:3(9,12,15) (ácido cis-9,12,15-
octadecatrienoico) (es un ácido graso esencial)• Ácido araquidónico, 20:4(5,8,11,14) (ácido cis-5,8,11,14-
eicosatetraenoico) (es un ácido graso esencial)

Page 20
Clasificación• Ácidos grasos insaturados.
– Ácidos grasos cis. Los dos átomos de hidrógeno del doble enlace están en el mismo lado de la molécula, lo que le confiere un "codo" en el punto donde está el doble enlace; la mayoría de los ácidos grasos naturales poseen configuración cis.
– Ácidos grasos trans. Los dos átomos de hidrógeno están uno a cada lado del doble enlace, lo que hace que la molécula sea rectilínea; se encuentra principalmente en alimentos industrializados que han sido sometidos a hidrogenación, con el fin de solidificarlos (como la margarina).

Page 22
Ácido graso esencial• Son aquellos que el organismo no puede sintetizar, por
lo que deben obtenerse por medio de la dieta. • Se trata de ácidos grasos poliinsaturados con todos los
dobles enlaces en posición cis.• Tanto la dieta como la biosíntesis suministran la mayoría
de los ácidos grasos requeridos por el organismo humano, y el exceso de proteínas y glúcidos ingeridos se convierten con facilidad en ácidos grasos que se almacenan en forma de triglicéridos.
• En el ser humano es esencial la ingestión un precursor en la dieta para dos series de ácidos grasos, la serie del ácido linoleico (serie ω-6) y la del ácido linolénico (serie ω-3).

Page 23
Ácido graso esencial• Los ácidos grasos esenciales se encuentran sobre todo
en el pescado azul, las semillas y frutos secos, como las de girasol o las nueces, en aceite de oliva o bacalao.
• La dieta de los animales para consumo también puede hacer que contengan gran cantidad de estos ácidos grasos.
• Por ejemplo la carne de los cerdos alimentados con bellota o las gallinas alimentadas con algas y harinas de pescado que ponen huevo con mayor cantidad estos ácidos grasos.

Page 24
Algunos ácidos grasos esenciales
• Ácidos grasos ω-6– Ácido linoleico, 18:2(9,12)– Ácido γ-linolénico, 18:3 (6,9,12)– Ácido eicosadienoico, 20:2 (11,14)– Ácido dihomo-gamma-linolénico, 20:3(8,11,14) – Ácido araquidónico, 20:4(5,8,11,14) – Ácido docosadienoico, 22:2(13,16) – Ácido adrénico, 22:4(7,10,13,16) – Ácido docosapentaenoico, 22:5(4,7,10,13,16) – Ácido caléndico, 18:3(8,10,12)

Page 25
Algunos ácidos grasos esenciales
• Ácidos grasos ω-3..– Ácido α-linolénico, 18:3(9,12,15) – Ácido estearidónico, 18:4(6,9,12,15) – Ácido eicosatrienoico, 20:3 (11,14,17) – Ácido eicosatetraenoico, 20:4(8,11,14,17) – Ácido eicosapentaenoico, 20:5(5,8,11,14,17) – Ácido docosapentaenoico (ácido clupanodónico),
22:5(7,10,13,16,19) – Ácido docosahexaenoico, 22:6(4,7,10,13,16,19) – Ácido tetracosapentaenoico, 24:5(9,12,15,18,21) – Ácido tetracosahexaenoico (ácido nisínico),
24:6(6,9,12,15,18,21)

Page 26
Biosíntesis de ácidos grasos• El primer paso en la biosíntesis de ácidos grasos es la
síntesis de ácido palmítico (ácido graso saturado de 16 C), los demás ácidos grasos se obtienen por modificaciones del ácido palmítico.
• El ácido palmítico se sintetiza secuencialmente en el citosol de la célula, gracias a la acción del polipéptido multienzimático ácido graso sintasa, por adición de unidades de dos carbonos aportadas por el acetil coenzima A; el proceso completo consume 7 ATP y 14 NADPH; la reacción global es la siguiente:
8 Acetil-CoA + 14 (NADPH + H+) + 7 ATP → Ácido palmítico (C16) + 8 CoA + 14 NADP+ + 7 (ADP + Pi) + 6
H2O

Page 27
Biosíntesis de ácidos grasos
• La fuente principal de acetil-CoA proviene del citrato que es transportado desde la matriz mitocondrial al citosol por un transportador específico de la membrana interna mitocondrial.
• Ya en el citosol, el citrato es escindido en oxalacetato y acetil-CoA, reacción que consume 1 ATP.
• El poder reductor, en forma de NADPH, lo suministra la ruta de la pentosa fosfato.
• En realidad, las unidades de dos carbonos que se añaden secuencialmente son aportadas por el malonil-CoA que, a su vez, es sintetizado por la enzima acetil-CoA carboxilasa, que adiciona un grupo carboxilo al acetil-CoA.

Page 28
Biosíntesis de ácidos grasos• El cuerpo humano puede sintetizar casi todos los ácidos
grasos que requiere a partir del ácido palmítico, mediante la combinación de estos mecanismos:– Alargamiento. Mediante este proceso, que tienen lugar en el
retículo endoplasmático y en la mitocondrias, se adicionan unidades de dos carbonos a la cadena de C16 del ácido palmítico, obteniéndose ácidos grasos de hasta C24.
– Desaturación. Mediante este proceso, que se produce en el retículo endoplasmático, se introducen dobles enlaces cis en la cadena hidrocarbonada de ácidos grasos suturados; el proceso es complejo e implica al NADPH, al citocromo b5 y diversos enzimas (como las desaturasas).

Page 29
Biosíntesis de ácidos grasos
• El principal precursor de los ácidos grasos es el malonil-CoA, una molécula que aporta dos de sus tres átomos de carbono al esqueleto carbonado del ácido graso en crecimiento.
• El malonil-CoA proviene, a su vez, del acetil-CoA.
• Todas las reacciones de síntesis de ácidos grasos tienen lugar en el citosol de las células.

Page 30
Síntesis de malonil-CoA
• En la síntesis de los ácidos grasos interviene un intermediario que no participa en la degradación (beta-oxidación), el malonil-CoA.
• El malonil-CoA se forma a partir de acetil-CoA y de bicarbonato, reacción que consume ATP y que está catalizada por la acetil-CoA carboxilasa, enzima que requiere biotina como cofactor.

Page 31
Síntesis de malonil-CoA; reacción de la acetil-CoA carboxilasa.Los colores corresponden a: enzima, coenzimas, substratos, iones metálicos, fosfato y bicarbonato

Page 32
Elongación
• Como en la β-oxidation, la elongación ocurre a través de cuatro reacciones recurrentes.
• Las unidades de acetil y malonil se muestran como sus tioésteres con su proteína transportadora de acilos (ACP); así es como los microorganismos y las plantas sintetizan sus ácidos grasos.
• En animales, esas mismas reacciones ocurren en una gran enzima dimérica, la ácido graso sintasa que tiene todas las actividades enzimáticas necesarias para la síntesis y libeación de ácidos grasos libres.

Page 33
Elongación
• El producto final del proceso es siempre ácido palmítico, un ácido graso saturado de 16 C, que es inmediatamente esterificado con el coenzima A, para formar palmitoil-CoA (lo mismo se hace con cualquier ácido graso proveniente de la dieta).
• A partir de él, una vez transportado al retículo endoplasmático, pueden sintetizarse otros ácidos grasos.

Page 34
Paso Descripción Reacción Enzima
Condensación
El primer paso es la condensación del acetil-ACP y el malonil-ACP, lo que conduce a la formación de acetoacetil-ACP con liberación de CO2.
β-Cetoacil-ACP sintase
Redución del acetoacetil-ACP
En este paso, el acetoacetil-ACP es reducido por el NADPH a D-3-hidroxibutiril-ACP. El doble enlace se reduce a un grupo hidroxilo. Solo se forma el isómero D.
β-Cetoacil-ACP reductasa
Deshidratación
En esta reacción, el D-3-hidroxibutiril-ACP es deshidratado a crotonil-ACP.
3-Hidroxiacil-ACP deshidratasa
Redución del crotonil-ACP
Durante es paso final, el crotonil-ACP es reducido por el NADPH a butiril-ACP.
Enoil-ACP reductasa

Page 35
Oxidación de ácidos grasos• Es un mecanismo clave para la obtención de energía
metabólica (ATP) por los organismos aeróbicos. • Los ácidos grasos son moléculas muy reducidas, su
oxidación libera mucha energía.• En animales su almacenamiento en forma de
triacilgliceroles es más eficiente y más importante que el almacenamiento de glúcidos en forma de glucógeno.
• La β-oxidación de los ácidos grasos lineales es el principal proceso productor de energía, pero no el único.
• Algunos ácidos grasos, como los de cadena impar o los insaturados requieren, para su oxidación, modificaciones de la β-oxidación o rutas metabólicas distintas.
• Tal es el caso de la α-oxidación, la ω-oxidación o la oxidación peroxisómica.

Page 36
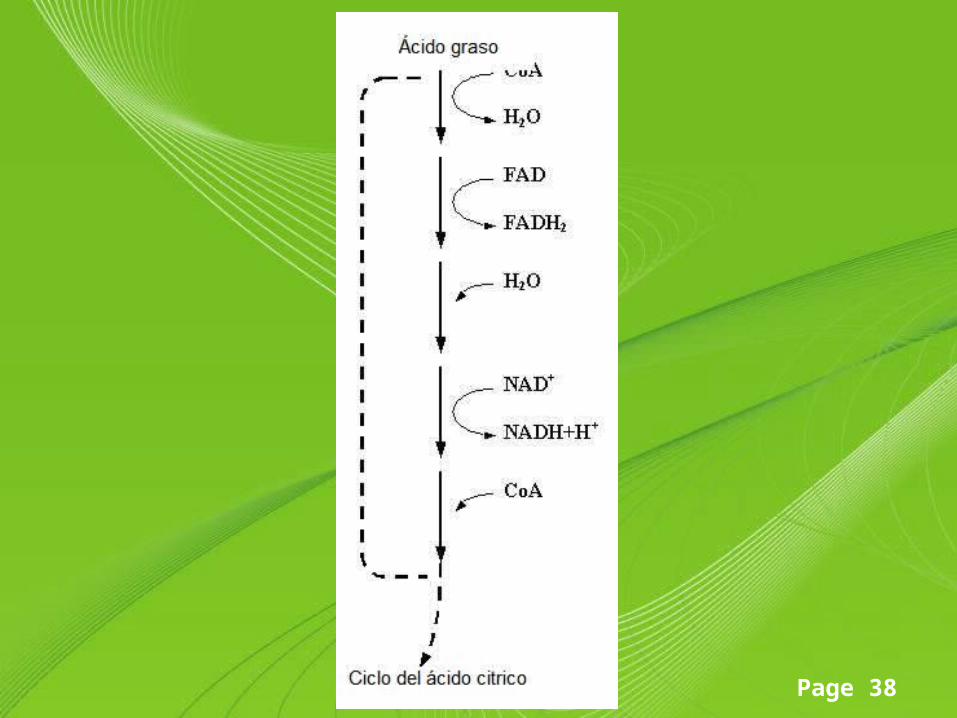
La β-oxidación de los ácidos grasos
• Sufren remoción, mediante la oxidación, de un par de átomos de carbono sucesivamente en cada ciclo del proceso, hasta que el ácido graso se descomponga por completo en forma de moléculas acetil-CoA, oxidados en la mitocondria para generar energía (ATP).
• La β-oxidación de ácidos grasos consta de cuatro reacciones recurrentes:– Oxidación por FAD– Hidratación– Oxidación por NAD+
– Tiólisis

Page 37
La β-oxidación de los ácidos grasos
• El resultado de dichas reacciones son unidades de 2 C en forma de acetil-CoA, molécula que pueden ingresar en el ciclo de Krebs, y coenzimas reducidos (NADH y FADH2) que pueden ingresar en la cadena respiratoria.
• No obstante, antes de que produzca la oxidación, los ácidos grasos deben activarse con coenzima A y atravesar la membrana mitocondrial interna, que es impermeable a ellos.

Page 39
Activación de los ácidos grasos• El paso previo a esas cuatro reacciones es la activación
de los ácidos grasos a acil coenzima A (acil CoA, R–CO–SCoA) grasos, que tiene lugar en el retículo endoplasmático (RE) o en la membrana mitocondrial externa, donde se halla la acil-CoA sintetasa (o ácido graso tioquinasa), la enzima que cataliza esta reacción:
R–COOH + ATP + CoASH →Acil-CoA sintetasa→ R–CO–SCoA + AMP + PPi + H2O
• El ácido graso se une al coenzima A (CoASH), reacción que consume dos enlaces de alta energía del ATP.

Page 40
Traslocación a la matriz mitocondrial
• Posteriormente debe usarse un transportador, la carnitina, para traslocar las moléculas de acil-CoA al interior de la matriz mitocondrial, ya que la membrana mitoncondrial interna es impermeable a los acil-CoA.
• La carnitina se encarga de llevar los grupos acilo al interior de la matriz mitoncondrial por medio del siguiente mecanismo.
• La carnitina es fuertemente inhibida por el malonil-CoA, uno de los pasos reguladores en el proceso de lipogénesis.
• La enzima carnitina palmitoiltransferasa I (CPTI) de la membrana mitocondrial externa elimina el coenzima A de la molécula de acil-CoA y, a la vez, la une a la carnitina situada en el espacio intermembrana, originado acilcarnitina; el CoA queda libre en el citosol para poder activar otro ácido graso.

Page 41
Traslocación a la matriz mitocondrial
• A continuación, una proteína transportadora, llamada translocasa, situada en la membrana mitocondrial interna, transfiere la acilcarnitina a la matriz mitoncondrial y, paralelamente, la carnitina palmitoiltrasnferasa II (CPTII) une una molécula de CoA de la matriz al ácido graso, regenerando así el acil-CoA .
• La carnitina se devuelve al espacio intermembrana por la proteína transportadora y reacciona con otro acil-CoA, repitiéndose el ciclo.
• La carnitina, también reconocida como vitamina B11, es un derivado aminoacídico que participa en el circuito vascular reduciendo niveles de triglicéridos y colesterol en sangre.
• Se produce naturalmente en el hígado a partir de los aminoácidos L-metionina y la L-lisina.
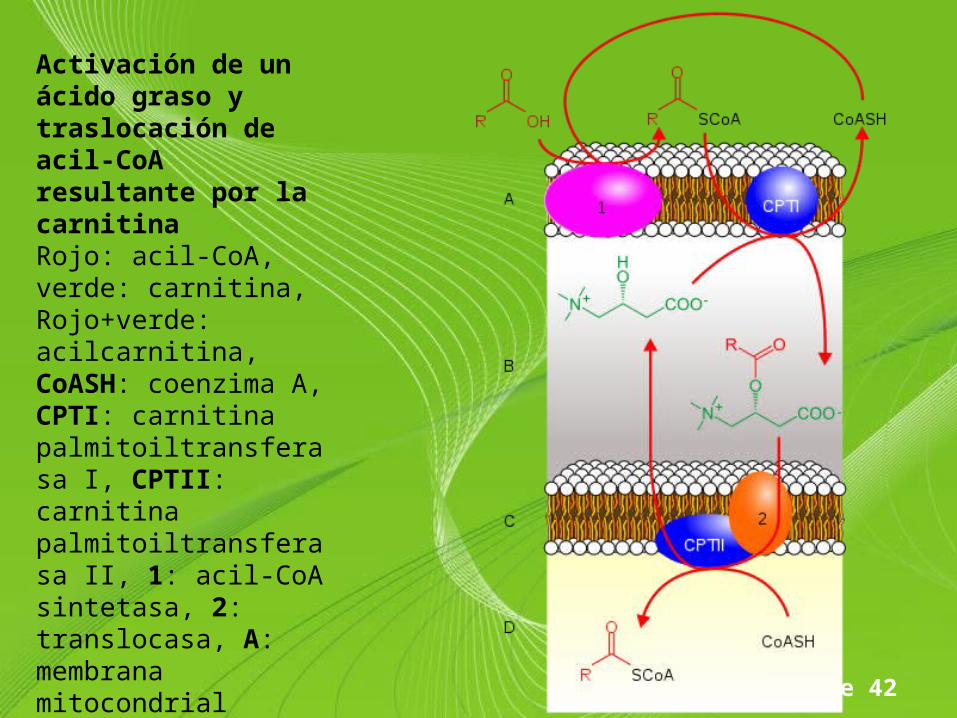
Page 42
Activación de un ácido graso y traslocación de acil-CoA resultante por la carnitinaRojo: acil-CoA, verde: carnitina, Rojo+verde: acilcarnitina, CoASH: coenzima A, CPTI: carnitina palmitoiltransferasa I, CPTII: carnitina palmitoiltransferasa II, 1: acil-CoA sintetasa, 2: translocasa, A: membrana mitocondrial extena, B: espacio intermembrana, C: membrana mitocondrial intena, D: matriz mitocondrial

Page 43
Descripcion Reacción Enzima Producto final
Oxidación por FAD: El primer paso es la oxidación del ácido graso por la acil-CoA deshidrogenasa. La enzima cataliza la formación de un doble enlace entre C-2 (carbono α) y C-3 (carbono β).
acil-CoA deshidrogenasa
trans-Δ2-enoil-CoA
Hidratación: El siguiente paso es la hidratación del enlace entre C-2 y C-3. Esta reacción es estereospecíca, formando solo el isómero L.
enoil CoA hidratasa
L-3-hidroxiacil CoA
Oxidación por NAD+: El tercer paso es la oxidación del L-3-hidroxiacil CoA por el NAD+, lo que convierte el grupo hidroxilo (–OH) en un grupo cetona (=O).
L-3-hidroxiacil CoA deshidrogenasa
3-cetoacil CoA
Tiólisis: El paso final es la separación del 3-cetoacil CoA por el grupo tiol de otra molécula de CoA. El tiol es insertado entre C-2 y C-3.
β-cetotiolasa
Una molécula de acetil CoA y una de acil CoA con dos carbonos menos

Page 44
Oxidación de los ácidos grasos insaturados
• La oxidación de los ácidos grasos insaturados requiere algunas variantes de la β-oxidación en la que participan algunos enzimas especiales, como la enoil-CoA isomerasa

Page 45
α-oxidación• Ruta catabólica de ciertos ácidos grasos ramificados
como el ácido fitálico procedente del fitol, cuyo grupo metilo localizado en el carbono 3 impide su catabolismo por beta oxidación.
• En esta vía catabólica la oxidación incial se produce en el carbono alfa.
• En la α-oxidación, que es especialmente importante para el metabolismo de ácidos grasos ramificados, se hidroxila el carbono α.
• Tiene lugar en el retículo endoplasmático y en la mitocondria, donde interviene la oxidasa de función mixta, y en el peroxisoma, donde interviene una hidroxilasa.

Page 46
Omega oxidación• Ruta metabólica del catabolismo de los ácidos grasos, donde se
oxida el carbono opuesto, el más alejado del grupo carboxilo (carbono ω).
• Este proceso es generalmente minoritario y afecta en especial a ácidos grasos de mediana longitud (10-12 átomos de carbono), pero puede ser importante cuando la β-oxidación está alterada.
• En vertebrados, los enzimas para la ω-oxidación se localizan en el retículo endoplásmico y en el citosol, especialmente del hígado y los riñones, en vez de en la mitocondria como en el caso de la β-oxidación.
• Se da en 3 pasos, cualquiera de los extremos del ácido graso puede unirse al coenzima A para formar un acil-CoA graso que puede sufrir la β-oxidación para producir ácidos de cadena más corta como el ácido succínico (C4), que puede ingresar en el ciclo de Krebs, y el ácido adípico (C6); este proceso tiene lugar principalmente en los peroxisomas.
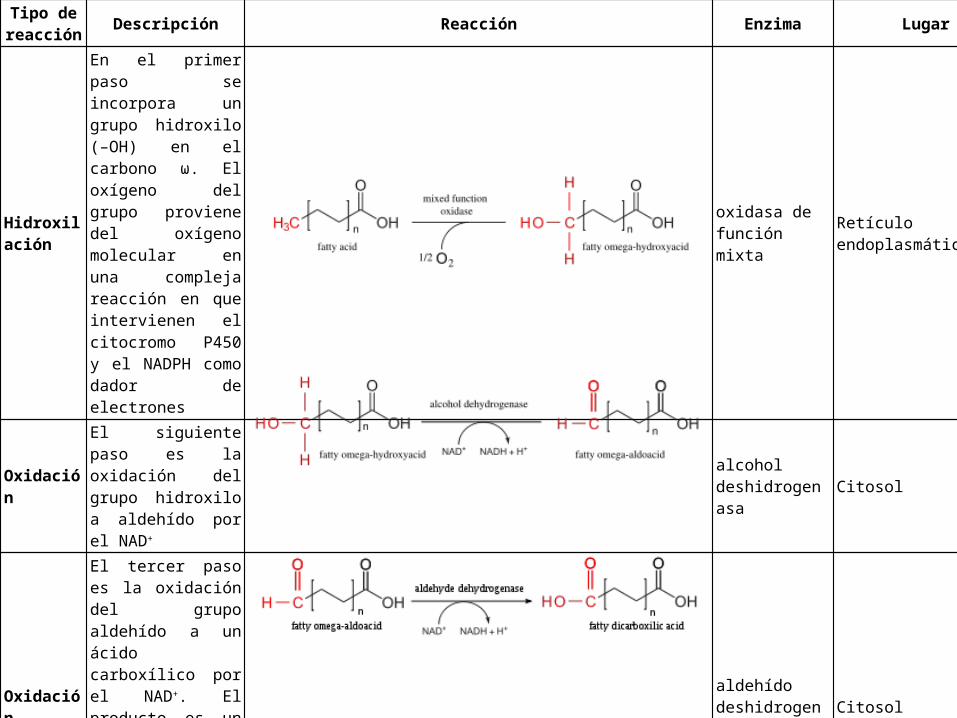
Page 47
Tipo de reacción
Descripción Reacción Enzima Lugar
Hidroxilación
En el primer paso se incorpora un grupo hidroxilo (–OH) en el carbono ω. El oxígeno del grupo proviene del oxígeno molecular en una compleja reacción en que intervienen el citocromo P450 y el NADPH como dador de electrones
oxidasa de función mixta
Retículo endoplasmático
Oxidación
El siguiente paso es la oxidación del grupo hidroxilo a aldehído por el NAD+
alcohol deshidrogenasa
Citosol
Oxidación
El tercer paso es la oxidación del grupo aldehído a un ácido carboxílico por el NAD+. El producto es un ácido graso con un grupo carboxilo en cada extremo (ácido dicarboxílico)
aldehído deshidrogenasa
Citosol

Page 48
Oxidación peroxisómicas de ácidos grasos
• Una fracción significativa de la oxidación de los ácidos grasos se produce en los peroxisomas, que contienen enzimas similares, aunque no idénticas, de los de la β-oxidación mitocondrial.
• Así, por ejemplo, en la deshidrogenación inicial, se forma H2O2 que es eliminado por la catalasa.
• Los peroxisomas tienen especificad para ácidos grasos de cadena más larga y a menudo no degradan totalmente la molécula, por lo que una posible función de este proceso sea el acortamiento de ácidos grasos de cadena larga hasta un punto en que la mitocondria pueda completar su β-oxidación.

Page 49
Papel biológico de los ácidos grasos• Función energética
– Los ácidos grasos son moléculas muy energéticas y necesarias en todos los procesos celulares en presencia de oxígeno, ya que por su contenido en hidrógenos pueden oxidarse en mayor medida que los glúcidos u otros compuestos orgánicos que no están reducidos.
– Cuando es demasiado bajo el nivel de insulina o no hay suficiente glucosa disponible para utilizar como energía en los procesos celulares, el organismo quema ácidos grasos para ese fin y origina entonces cuerpos cetónicos, productos de desecho que causan una elevación excesiva del nivel de ácido en la sangre, lo que podría conducir a la cetoacidosis, un problema importante y muchas veces ignorado o pospuesto hasta otra vez.
– Los síntomas de esta enfermedad van desde la presencia de un aroma a quitaesmalte en el aliento, hasta la aparición de pequeñas manchas de color amarillento (o verduzco) sobre la piel, y la ligera acidificación del semen, que conlleva un cierto dolor al eyacular.

Page 50
Acilglicéridos
• Los acilglicéridos o acilgliceroles son ésteres de ácidos grasos con glicerol, formados mediante una reacción de condensación llamada esterificación.
• Una molécula de glicerol (glicerina) puede reaccionar con hasta tres moléculas de ácidos grasos, puesto que tiene tres grupos hidroxilo.
• Las cadenas carbonadas de los ácidos que reaccionan con el glicerol, pueden ser saturadas o insaturadas.

Page 51
Tipos de acilglicéridos• Según el número de ácidos grasos que se unan
a la molécula de glicerina, existen tres tipos de acilgliceroles:– Monoacilglicéridos.
• Sólo existe un ácido graso unido a la molécula de glicerina. • Son los precursores de los siguientes.
– Diacilglicéridos. • La molécula de glicerina se une a dos ácidos grasos.• Son los precursores de los triglicéridos.
– Triacilglicéridos. • También se llaman triglicéridos, puesto que la glicerina está unida a
tres ácidos grasos.
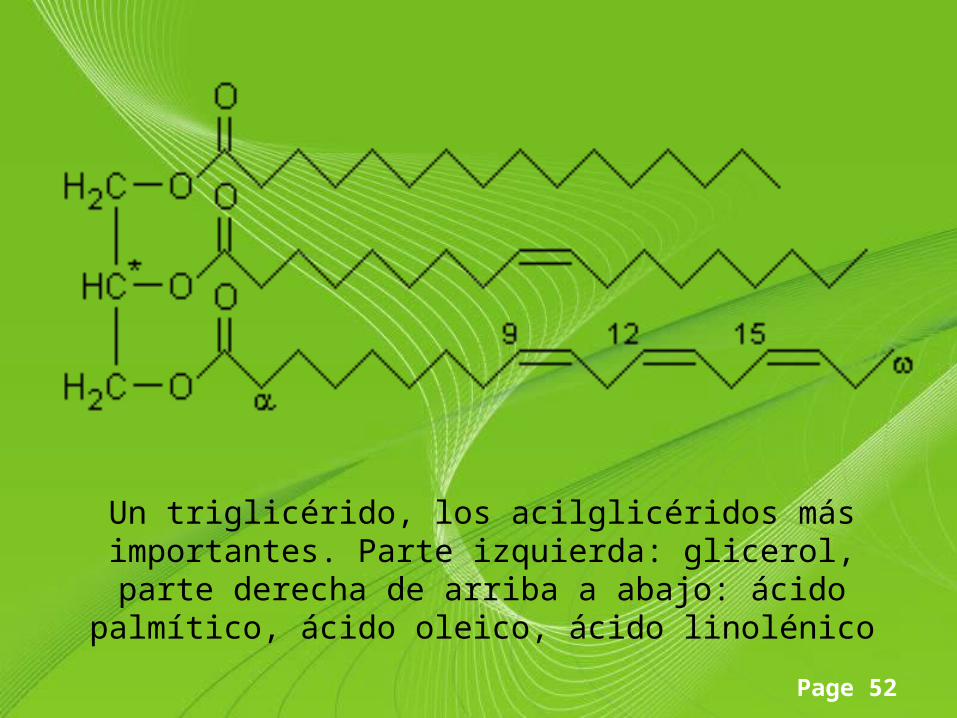
Page 52
Un triglicérido, los acilglicéridos más importantes. Parte izquierda: glicerol, parte derecha de arriba a abajo: ácido
palmítico, ácido oleico, ácido linolénico

Page 53Representación tridimensional de un triglicérido

Page 54
Céridos
• Son moléculas que se obtienen por esterificación de un ácido graso con un alcohol monovalente lineal de cadena larga (cera de abeja).
• Son sustancias altamente insolubles en medios acuosos y a temperatura ambiente se presentan sólidas y duras.
• En los animales las podemos encontrar en la superficie del cuerpo, piel, plumas, cutícula, etc.
• En los vegetales, las ceras recubren en la epidermis de frutos, tallos, junto con la cutícula o la suberina, que evitan la pérdida de agua por evaporación.

Page 55
Fosfolípidos• Los fosfoglicéridos o glicerofosfolípidos son
moléculas lipídica del grupo de los fosfolípidos. • Están compuestos por ácido fosfatídico, una molécula
compleja compuesta por glicerol, en el que se han esterificado dos ácidos grasos (uno saturado y otro insaturado) y un grupo fosfato.
• A su vez, al grupo fosfato se une un alcohol o un aminoalcohol.
• En los organismos vivos tiene función estructural puesto que son uno de los principales componentes de las bicapas de las membranas celulares y subcelulares.

Page 56
Fosfolípidos• Los fosfolípidos tienen un marcado carácter anfipático
consecuencia de la estructura de la molécula.• Las largas cadenas alifáticas de los ácidos grasos tienen
carácter hidrófobo (repelen el agua) y forman dos largas "colas" apolares, mientras que el grupo fosfato y el alcohol, cargados eléctricamente, son fuertemente hidrófilos (interaccionan con el agua) y constituyen la "cabeza" polar de la molécula; ello conduce a que, en un medio acuoso, se autoorganicen formando bicapas, con las cabezas polares en contacto con el agua y las colas hidrófobas "escondidas" y enfrentadas entre si.

Page 57
Ejemplos de fosfoglicéridos
• Según el alcohol que se una al ácido fosfatídico, se tienen diferentes fosfoglicéridos:– Fosfatidilcolina o lecitina. Posee el aminoalcohol
colina.– Fosfatidiletanolamina o cefalina. Posee el
aminoalcohol etanolamina.– Fosfatidilserina. Posee el aminoalcohol serina.– Fosfatidilinositol. Posee el alcohol inositol.– Fosfatidilglicerol. Posee el alcohol glicerol; si el grupo
OH esterifica otra molécula de ácido fosfatídico se origina el difosfatidilglicerol o cardiolipina (característico de las mitocondrias de las células musculares cardíacas).

Page 58
Fosfoglicéridos: 1: cabeza hidrófila; 2: colas hidrófobas. A: fosfatidilcolina; B: fosfatidiletanolamina; C: fosfatidilserina; D: representación esquemática de un fosfoglicérido con la cabeza hidrófila (1) y las colas hidrófobas (2).

Page 59
Estructura de un fosfoglicérido; X representa el alcohol o aminoalcohol que se esterifica con el grupo fosfato; el resto representa el ácido fosfatídico

Page 60
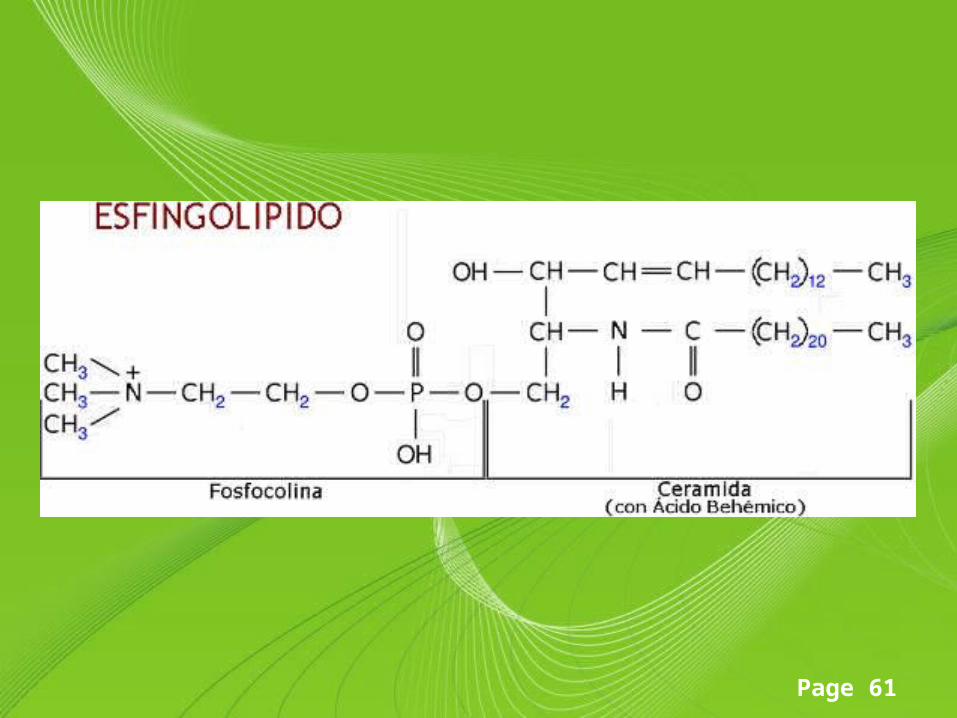
Esfingolípido
• Son lípidos complejos que derivan del alcohol insaturado de 18 C esfingosina; la esfingosina se halla unida a un ácido graso de cadena larga mediante un enlace amida formando la ceramida.
• Son una clase importante de lípidos de las membranas celulares de animales y vegetales y son los más abundantes en los tejidos de los organismos más complejos.

Page 62
Componentes estructurales• Todos los esfingolípidos poseen tres componentes básicos
característicos: un aminoalcohol de cadena larga llamado esfingosina (1,3-dihidroxi-2-amino-4-octadequeno).
• Los carbonos 1, 2 y 3 de esta molécula son portadores de grupos funcionales (-OH, NH2, -OH) y cuando un ácido graso saturado o insaturado de cadena larga se une al carbono 2 por medio de un enlace amida se obtiene una ceramida, la cual es la unidad estructural fundamental de todos los esfingolípidos.
• Este tipo de lípido se encuentra presente en cantidades especialmente grandes en el tejido nervioso y cerebral. La esfingosina, con su grupo amino libre no se encuentra en la naturaleza.
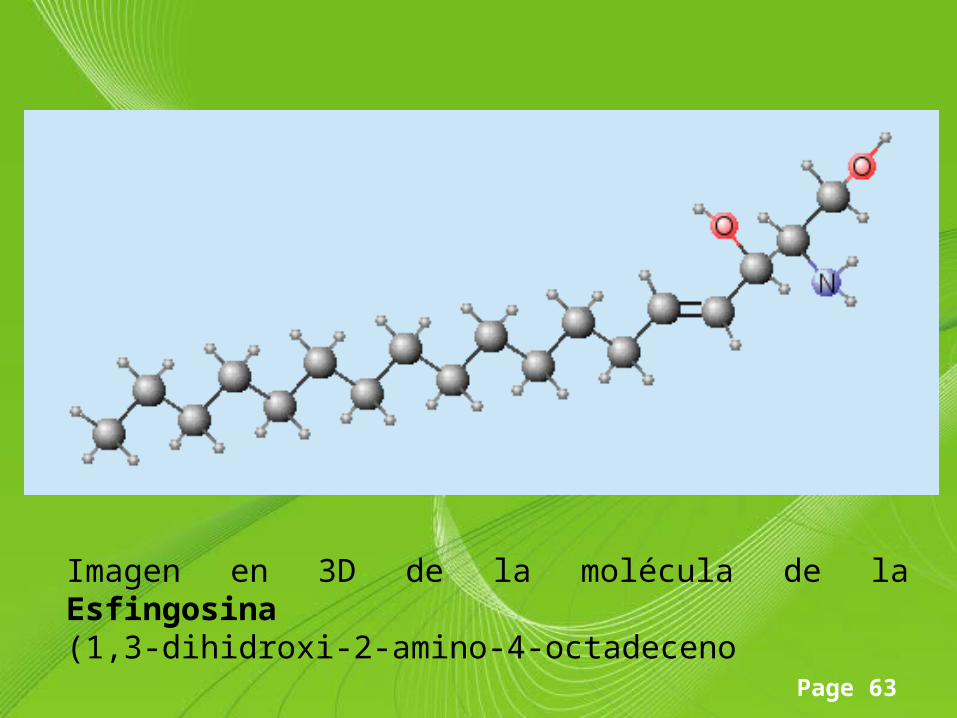
Page 63
Imagen en 3D de la molécula de la Esfingosina(1,3-dihidroxi-2-amino-4-octadeceno

Page 64
Grupos de esfingolípidos
• Los esfingolípidos se dividen en dos grupos clases, los fosfoesfingolípidos (esfingomielinas) y los glucoesfingolípidos (gangliósidos, cerebrósidos).– Esfingomielinas– Son fosfoesfingolípidos ya que contienen un grupo fosfato, unido
al grupo hidroxilo 1 de la ceramida, que se esterifica con la colina (fosfocolina) o etanolamina (fosfoetanolamina) para formar el grupo o "cabeza" polar de la molécula.
– Es el único esfingolípido que contiene fósforo y puede, por tanto, considerarse un fosfolípido, pero se acostumbra a clasificar como esfingolípido por sus componentes estructurales.
– Las esfingomielinas se hallan presentes en las membranas plasmáticas de las células animales, en la vaina de mielina que recubre y aísla los axones de las neuronas mielinizadas (de ahí procede su nombre), es uno de los principales lípidos estructurales de las membranas del tejido nervioso.

Page 65
Grupos de esfingolípidos
• Glucoesfingolípidos– O simplemente glucolípidos, carecen de fósforo; el
grupo hidroxilo 1 de la ceramida se une, mediante un enlace glucosídico a un monosacárido o a un oligosacárido, que normalmente se proyectan hacia afuera en la cara externa de la membrana plasmática.
– Los monosacáridos más frecuentes son la D-glucosa, D-galactosa y N-acetilgalactosamina. Se conocen diversos tipos:
• Gangliósidos. • Cerebrósidos .• Globósidos

Page 66
Grupos de esfingolípidos
• Gangliósidos. – Son los esfingolípidos más complejos en virtud de contener cabezas
polares muy grandes formadas por unidades de oligosacáridos cargadas negativamente ya que poseen una o más unidades de ácido N-acetilneuramínico o ácido siálico que tiene una carga negativa a pH 7. Los gangliósidos difieren de los anteriores por poseer este ácido.
– Están concentrados en gran cantidad en las células ganglionares del sistema nervioso central, especialmente en las terminaciones nerviosas.
– Los gangliósidos constituyen el 6% de los lípidos de membrana de la materia gris del cerebro humano y se hallan en menor cantidad en las membranas de la mayoría de los tejidos animales no nerviosos.
– Se presentan en la zona externa de la membrana y sirven para reconocer las células, por lo tanto se les considera receptores de membrana.

Page 67
Grupos de esfingolípidos
• Cerebrósidos. – Tienen un único azúcar unido mediante enlace β-glucosídico al grupo
hidroxilo de la ceramida; los que tienen galactosa (galactocerebrósidos), se encuentran de manera característica a las membranas plasmáticas de células del tejido nervioso, mientras que los que contienen glucosa (glucocerebrósidos) se hallan en las membranas plasmáticas de células de tejidos no nerviosos.
– Los sulfátidos poseen una galactosa esterificada con sulfato en el carbono 3.
• Globósidos. – Los globósidos son glucoesfingolípidos con oligosacáridos neutros
unidos a la ceramida.

Page 68
Lípidos insaponificables
• Terpeno – O isoprenoides son una vasta y diversa clase de
compuestos orgánicos derivados del isopreno (o 2-metil-1,3-butadieno), un hidrocarburo de cinco átomos de carbono.
– El nombre proviene de que los primeros miembros de esta clase fueron derivados del aguarrás ("turpentine" en inglés, "terpentin" en alemán).
– Cuando los terpenos son modificados químicamente, por ejemplo por oxidación o reorganización del esqueleto hidrocarbonado, suelen denominarse terpenoides (como la vitamina A o retinol que contiene un átomo de oxígeno).

Page 69
Modelo de bolas de la estructura molecular del isopreno, la unidad química de los terpenos

Page 70
Clasificación • Los terpenos son hidrocarburos que pueden verse como
una combinación de numerosas unidades isopreno, por lo general unidas de forma cabeza-cola, pero también pueden darse combinaciones cabeza-cabeza y algunos compuestos están formados por uniones cabeza-medio.
• Los terpenos pueden ser considerados como terpenos modificados donde grupos metilo han sido reacomodados o removidos, o a los que se les han añadido átomos de oxígeno.
• La clasificación de los terpenos según su estructura química, es similar a la de los terpenos, los cuales son clasificados en base al número de unidades isopreno presentes y en el caso de los triterpenos, si están ciclados. Se los clasifica en:

Page 71
Clasificación • Hemiterpenos.
– Los terpenos más pequeños, con una sola unidad de isopreno.– Poseen 5 carbonos. – El hemiterpeno más conocido es el isopreno mismo, un
producto volátil que se desprende de los tejidos fotosintéticamente activos.
• Monoterpenos. – Terpenos de 10 carbonos. – Llamados así porque los primeros terpenos aislados del
aguarrás en los 1850s, fueron considerados la unidad base, a partir de la cual se hizo el resto de la nomenclatura.
– Los monoterpenos son mejor conocidos como componentes de las esencias volátiles de las flores y como parte de los aceites esenciales de hierbas y especias, en los que ellos forman parte de hasta el 5 % en peso de la planta seca.

Page 72
Clasificación • Sesquiterpenos.
– Terpenos de 15 carbonos (es decir, terpenos de un monoterpeno y medio).
– Como los monoterpenos, muchos sesquiterpenos están presentes en los aceites esenciales.
– Además muchos sesquiterpenos actúan como fitoalexinas, compuestos antibióticos producidos por las plantas en respuesta a la aparición de microbios, y como inhibidores de la alimentación ("antifeedant") de los herbívoros oportunistas.
– La hormona de las plantas llamada ácido abscísico es estructuralmente un sesquiterpeno, su precursor de 15 carbonos, la xantosina, no es sintetizada directamente de 3 unidades isopreno sino producida por un "cleavage" asimétrico de un carotenoide de 40 unidades.

Page 73
Clasificación • Diterpenos.
– Terpenos de 20 carbonos. – Entre ellos se incluye el fitol, que es el lado hidrofóbico de la
clorofila, las hormonas giberelinas, los ácidos de las resinas de las coníferas y las especies de legumbres, las fitoalexinas, y una serie de metabolitos farmacológicamente importantes, incluyendo el taxol, un agente anticáncer encontrado en muy bajas concentraciones (0,01% de peso seco) en la madera del tejo ("yew"), y forskolina, un compuesto usado para tratar el glaucoma.
– Algunas giberelinas tienen 19 átomos de carbono por lo que no son consideradas diterpenos porque perdieron un átomo de carbono durante una reacción de "cleavage".

Page 74
Clasificación • Triterpenos.
– Terpenos de 30 carbonos. – Son por lo general generados por la unión cabeza-cabeza de
dos cadenas de 15 carbonos, cada una de ellas formada por unidades de isopreno unidas cabeza-cola.
– Esta gran clase de moléculas incluye a los brassinoesteroides, componentes de la membrana que son fitoesteroles, algunas fitoalexinas, varias toxinas y "feeding deterrents", y componentes de las ceras de la superficie de las plantas, como el ácido oleanólico de las uvas.
• Tetraterpenos. – Terpenos de 40 carbonos (8 unidades de isopreno). – Los tetraterpenos más prevalentes son los pigmentos
carotenoides accesorios que cumplen funciones esenciales en la fotosíntesis.

Page 75
Clasificación • Politerpenos.
– Los politerpenos, que contienen más de 8 unidades de isopreno, incluyen a los "prenylated quinone electron carriers" como la plastoquinona y la ubiquinona, también poliprenoles de cadena larga relacionados con las reacciones de transferencia de azúcares (por ejemplo el dolicol), y también a enormemente largos polímeros como el "rubber", usualmente encontrado en el látex.
• Meroterpenos. – Así se llama a los metabolitos secundarios de las plantas que tienen
orígenes sólo parcialmente derivados de terpenos. – Por ejemplo, tanto las citokininas como numerosos fenilpropanoides
contienen cadenas laterales de un isoprenoide de 5 C. – Algunos alcaloides, como las drogas anticáncer vincristina y
vinblastina, contienen fragmentos terpenos en sus estructuras. – Además algunas proteínas modificadas incluyen una cadena lateral
de 15 o 20 carbonos que es un terpeno, que es el que ancla la proteína a la membrana.

Page 76
Clasificación • Esteroides.
– Triterpenos basados en el sistema de anillos ciclopentanoperhidrofenantreno ("cyclopentane perhydro-phenanthrene ring system"). Buchanan et al. no los consideran terpenos.

Page 77
Biosíntesis • Si bien sus funciones son muy diversas, los terpenos
tienen en su origen biosintético común. • La biosíntesis de todos los terpenos a partir de
metabolitos primarios puede simplificarse a 4 pasos:– Síntesis del precursor fundamental, el isopentenil pirofosfato
(IPP).– Adiciones repetitivas del IPP hasta formar una serie de
homólogos del isoprenil difosfato, que sirve de precursor inmediato de muchas clases de terpenos.
– Elaboración de estos "allylic" prenil difosfatos por sintasa específicas de terpeno para dar con el esqueleto del terpeno, y
– modificaciones enzimáticas secundarias a los esqueletos (en general reacciones redox), para dar con las propiedades y la gran diversidad química de esta familia.

Page 78
Isopentenil pirofosfato, el precursor en la biosíntesis de los terpenos

Page 79
Esteroide
• Son derivados del núcleo del ciclopentanoperhidrofenantreno o esterano que se compone de carbono e hidrógeno formando cuatro anillos fusionados, tres hexagonales y uno pentagonal; posee 17 átomos de carbono.
• En los esteroides esta estructura básica se modifica por adición de diversos grupos funcionales, como carbonilos e hidroxilos (hidrófilos) o cadenas hidrocarbonadas (hidrófobas).

Page 80
Molécula de esterano o ciclopentano-perhidro-fenantreno.

Page 81
Estructura química
• El núcleo de esterano es bastante rígido con una estructura prácticamente plana.
• Las sustancias derivadas de este núcleo posee grupos metilo (-CH3) en las posiciones 10 y 13 que representan los carbonos 18 y 19, así como un carbonilo o un hidroxilo en el carbono 3; generalmente existe también una cadena hidrocarbonada lateral en el carbono 17; la longitud de dicha cadena y la presencia de metilos, hidroxilos o carbonilos determina las diferentes estructuras de estas sustancias
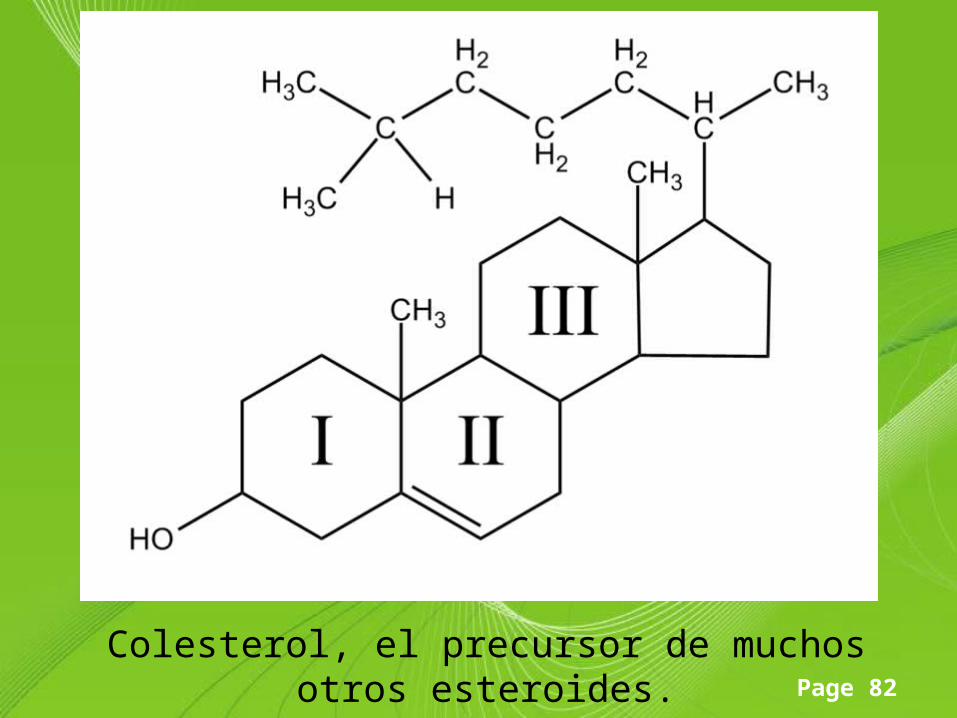
Page 82
Colesterol, el precursor de muchos otros esteroides.

Page 83
Funciones • En los mamíferos cumplen importantes funciones:
– Reguladora: Algunos regulan los niveles de sal y la secreción de bilis.
– Estructural: El colesterol es un esteroide que forma parte la estructura de las membranas de las células junto con los fosfolípidos. Además, a partir del colesterol se sintetizan los demás esteroides.
– Hormonal: Las hormonas esteroides son: • Corticoides: glucocorticoides y mineralocorticoides. Existen
múltiples fármacos con actividad corticoide, como la prednisona.• Hormonas sexuales masculinas: son los andrógenos, como la
testosterona y sus derivados, los anabolizantes androgénicos esteroides; estos últimos llamados simplemente esteroides.
• Hormonas sexuales femeninas.• Vitamina D y sus derivados.

Page 84
Funciones
• Las hormonas esteroides tienen en común que:– Se sintetizan a partir del colesterol.– Son hormonas lipófilas que atraviesan libremente la
membrana plasmática, se unen a un receptor citoplasmático, y este complejo receptor-hormona tiene su lugar de acción en el ADN del núcleo celular, activando genes o modulando la transcripción del ADN.
• Entre los esteroides se pueden destacar los esteroles. – Función hepática

Page 85
Eicosanoides • Son un grupo de moléculas de carácter lipídico originadas de la
oxigenación de los ácidos grasos esenciales de 20 C tipo omega-3 y omega-6.
• Cumplen amplias funciones como mediadores para el sistema nervioso central, los eventos de la inflamación y de la respuesta inmune tanto en vertebrados como en invertebrados.
• Todos los eicosanoides son moléculas de 20 átomos de C y están agrupados en prostaglandinas, tromboxanos, leucotrienos, y ciertos hidroxiácidos precursores de los leucotrienos.
• Constituyen las moléculas involucradas en las redes de comunicación celular más complejas del organismo animal, incluyendo el ser humano.

Page 86
Estructura química del ácido araquidónico, del cual derivan la gran mayoría de los eicosanoides

Page 87
Eicosanoides • De cada tipo de molécula agrupada dentro de los eicosanoides
derivan dos o tres moléculas producidas, bien sea a raíz de un ácido graso esencial tipo ω-3 o del tipo ω-6.
• Se notará que las actividades metabólicas de los derivados de eicosanoides dependen de su estructura bioquímica y, por lo general, su estructura individual explica los efectos orgánicos del omega-3 y del omega-6.
• Los eicosanoides son ácidos carboxílicos, virtud del cual obtienen sus propiedades metabólicas.
• Aquellos derivados del omega-3 (ω-3) por lo general tienen peculiaridades antiinflamatorias, no tanto así los derivados del omega-6 (ω-6).
• De los eicosanoides, las prostaglandinas y los tromboxanos incluyen anillos saturados y heterocíclicos, mientras que los leucotrienos y los hidroxiácidos son lineales (abiertas de principio a fin de su cadena).

Page 88
Eicosanoides • Todos ellos derivan del ácido araquidónico (AA), y son ácidos
grasos esenciales. • Además del ácido araquidónico, otros ácidos grasos esenciales
también pueden servir como precursores en la formación de estas moléculas, por ejemplo, el ácido linoleico y el ácido linolénico.– Prostaglandinas– Tienen 20 C, un grupo de ácido carboxílico y un anillo de 5 C como parte de su
estructura. – Todas las prostaglandindas tienen un ciclopentano, excepto la prostaglandina I2,
que tiene un anillo adicional.– Tromboxanos – Moléculas cíclicas (heterociclo) bien sea de 6 ó 5 C con 1 O, formando éste
último un pequeño anillo de oxano (anillo que contiene 5 C y un átomo de O. – Tienen estructuras parecidas a las prostaglandinas y siguen la misma
nomenclatura. – Constan de un anillo y dos colas. – Se encontraron primeramente en los trombocitos (plaquetas), de allí su nombre
tromboxano.

Page 89
Eicosanoides – Leucotrienos – Moléculas lineales. – Se identificaron en leucocitos y por ello se les conoce como leucotrieno. – Aunque tienen 4 enlaces dobles, inicialmente se pensaba que tenían 3 dobles
enlaces conjugados (de allí trieno). – Su producción en el cuerpo forma parte de una compleja secuencia metabólica
que incluye la producción de histamina.– Ácido linoleico – (un ω-6) tiene la última insaturación a seis posiciones del final y producen el
ácido araquidónico y pueden formar directamente la prostaglandina G2 (pro-inflamatorios).
– La palabra linoleico viene de la planta lino y -oleico relacionado con los aceites oleicos.

Page 90
Eicosanoides – Ácido linolénico – (un ω-3) tiene la última insaturación a 3 posiciones del final, impidiéndoles
producir ácido araquidónico por sí mismos. – Son capaces de producir la prostaglandina G3 (lo que les otorga propiedades
antiinflamatorias). – Debido a la carencia de las enzimas que sintetizan este ácido graso esencial, los
seres humanos no producen ácido linolénico y deben obtenerlo a través de la dieta.
– El peso de evidencias científicas sugiere que consumir una dieta rica en ácido linolénico (2-3 gramos cada día) es útil en la prevención de cardiopatías isquémicas.[

Page 91
Estructura de algunos eicosanoides
Prostaglandina E1: El anillo de 5 lados es característico de su
clase
Tromboxano A2: Los oxígenos se han adentrado en el anillo.
Leucotrieno B4: Note los tres doble enlaces conjugados.
Prostaciclina I2: El segundo anillo lo distingue de las prostaglandinas.
Leucotrieno E4: un ejemplo de un leucotrieno "cisteinilo“.

Page 92
Biosíntesis • La oxidación catalítica de ácidos grasos es un requisito
para la producción de eicosanoides, y tiene lugar gracias a dos familias de enzimas:– Ciclooxigenasa (COX) de tipos 1, 2 y 3; conducen a la formación
de prostanoides.– Lipooxigenasa, en sus variadas formas. La 5-lipoxigenasa (5-
LO) genera leucotrienos.

Page 93
Biosíntesis • El primer paso de la biosíntesis de eicosanoides ocurre
cuando la célula es activada por lesión celular, citocinas, factores de crecimiento u otros estímulos.
• La vía es tan compleja que el estímulo puede ser un eicosanoide mismo producido por una célula vecina.
• Esos estímulos provocan la liberación de fosfolipasa a nivel de la membrana celular el cual viaja a la membrana del núcleo celular.
• Una vez allí, este biocatalizador, dependiendo del isotipo, produce la hidrólisis (1) del enlace fosfodiéster de los fosfolípidos (por la enzima fosfolipasa A2) o (2) del diacilglicerol (por la fosfolipasa C).
• Esas interacciones libera de la membrana un ácido graso esencial de 20 C.

Page 94
Biosíntesis • La hidrólisis de los fosfolípidos parece ser la reacción
limitante, y por ende, la más regulada en la formación de eicosanoides.
• Aquellas células que no posean en su citoplasma la enzima fosfolipasa A2, por lo general son células deprovisatas de síntesis de eicosanoides.
• Es una fosfolipasa específica para ciertos fosfolípidos que contengan bien sea AA, EPA o DGLA en una posición determinada de la molécula.
• La fosfolipasa citoplasmática es la liberadora de precusores del factor activador de plaquetas (PAF).

Page 95
Vías de los eicosanoides Eicosanoides
'clásicos'Otras moléculas derivadas de ácidos
grasos esenciales de 20 carbonos•Los ácidos grasos tienendos destinos eicosanoides: •Vía de la 5-lipoxigenasa:
• Leucotrienos•Vía de la ciclooxigenasa("prostanoides"):
• Prostaglandinas• Prostaciclina• Tromboxanos
•Otras rutas oxidativas generan productos similares: •Vía de la 12-lipoxigenasa:
• Hepoxilinas• Lipoxinas
•Vía del citocromo P450: • Ácido
epoxieicosatrienoico
•Peroxidación por radicales libres:
• Isoprostanos• Isofuranos
•Está también la adición por etanolamina o glicerol: •Anandamida y otrosendocanabinoides

Page 96
Peroxidación lipídica y otras especies reactivas de oxígeno • En el paso siguiente, los ácidos grasos libres de la
membrana son oxigenados por cualquier vía reactiva del oxígeno.
• Las vías específicas de eicosanoides son las de la lipoxigenasa, de la ciclooxigenasa y del O2.
• A pesar de que el ácido graso es simétrico, los eicosanoides resultantes de él son compuestos quirales, pues el proceso de oxidación ocurre con alta estereoespecificidad.
• La oxidación lipídica es una reacción potencialmente peligrosa para la célula, en particular si ocurre cerca del núcleo.

Page 97
Peroxidación lipídica y otras especies reactivas de oxígeno • Sin embargo, la célula está provista de mecanismos que
evitan las oxidaciones innecesarias. • La ciclooxigenasa (COX), las lipoxigenasas y las
fosfolipasas son enzimas fuertemente reguladas; por ejemplo, existen al menos ocho proteínas activadas para coordinar la producción de leucotrienos, algunas en múltiples conformaciones.
• La oxidación tanto por COX como por lipoxigenasa libera especies reactivas de oxígeno (ROS), de hecho los productos iniciales de los eicosanoides son también altamente peroxidantes.
• El leucotrieno LTA2 puede formar aductos con tejido nuclear.

Page 98
Peroxidación lipídica y otras especies reactivas de oxígeno • Otras reacciones con lipoxigenasas generan lesión
celular: ciertos modelos con múridos implican a la 15-lipoxigenasa en la patogénesis de la aterosclerosis.
• La oxidación en la producción de eicosanoides está compartimentada, limitando aún más la posibilidad de lesión por peróxidos reactivos.
• Las enzimas biosintéticas de eicosanoides (como la glutatión S-transferasa, la epóxido hidrolasa y transportadores proteicos) pertenecen a familias cuyas funciones están involucradas en la desintoxicación celular.
• Esto sugiere que las señales eicosanoides pudieron haber evolucionado de la desintoxicación de los amenazantes ROS.

Page 99
Peroxidación lipídica y otras especies reactivas de oxígeno • De algún modo la célula encuentra ventajoso generar
hidroperóxidos lipídicos cerca de su núcleo. • En ese estado, las PG y los LT pueden enviar señales
intracelulares o bien regular la transcripción genética de ADN, mientras que LTB4 es el ligando del receptor nuclear llamado "receptores activados por la proliferación de los peroxisomas" (PPAR).
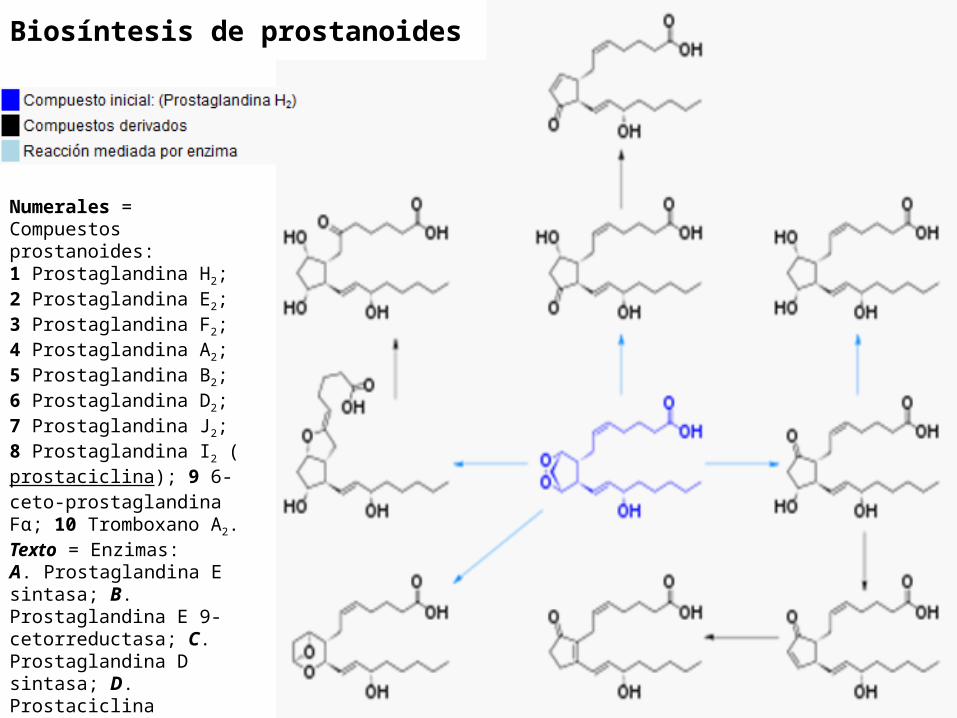
Biosíntesis de prostanoides
Numerales = Compuestos prostanoides:1 Prostaglandina H2; 2 Prostaglandina E2; 3 Prostaglandina F2; 4 Prostaglandina A2; 5 Prostaglandina B2; 6 Prostaglandina D2; 7 Prostaglandina J2; 8 Prostaglandina I2 (prostaciclina); 9 6-ceto-prostaglandina Fα; 10 Tromboxano A2.Texto = Enzimas:A. Prostaglandina E sintasa; B. Prostaglandina E 9-cetorreductasa; C. Prostaglandina D sintasa; D. Prostaciclina sintasa; E. Tromboxano-A sintasa

Page 101
Funciones de los lípidos
• Función de reserva energética– Los triglicéridos son la principal reserva de energía de los
animales ya que un gramo de grasa produce 9,4 kcal en las reacciones metabólicas de oxidación, mientras que las proteínas y los glúcidos sólo producen 4,1 kcal por gramo.
• Función estructural – Los fosfolípidos, los glucolípidos y el colesterol forman las
bicapas lipídicas de las membranas celulares. – Los triglicéridos del tejido adiposo recubren y proporcionan
consistencia a los órganos y protegen mecánicamente estructuras o son aislantes térmicos.
• Función transportadora – El transporte de lípidos desde el intestino hasta su lugar de
destino se realiza mediante su emulsión gracias a los ácidos biliares y a las lipoproteínas.

Page 102
Funciones de los lípidos
• Función reguladora, hormonal o de comunicación celular– Las vitaminas liposolubles son de naturaleza lipídica (terpenos,
esteroides); las hormonas esteroides regulan el metabolismo y las funciones de reproducción; los glucolípidos actúan como receptores de membrana; los eicosanoides poseen un papel destacado en la comunicación celular, inflamación, respuesta inmune, etc.
• Función Biocatalizadora– En este papel los lípidos favorecen o facilitan las reacciones
químicas que se producen en los seres vivos. – Cumplen esta función las vitaminas lipídicas, las hormonas
esteroideas y las prostaglandinas.

Page 103
Importancia para los organismos vivientes • Las vitaminas A, D, E y K son liposolubles, lo que
significa que estas solo pueden ser digeridas, absorbidas y transportadas en conjunto con las grasas.
• Las grasas son fuentes de ácidos grasos esenciales, un requerimiento dietario importante.
• Las grasas juegan un papel vital en el mantenimiento de una piel y cabellos saludables, en el aislamiento de los órganos corporales contra el shock, en el mantenimiento de la temperatura corporal y promoviendo la función celular saludable.
• Estos además sirven como reserva energética para el organismo.

Page 104
Importancia para los organismos vivientes • Las grasas son degradadas en el organismo para liberar
glicerol y ácidos grasos libres. • El glicerol puede ser convertido por el hígado y entonces
ser usado como fuente energética.• El contenido de grasas de los alimentos puede ser
analizado por extracción. • El método exacto varía según el tipo de grasa a ser
analizada, por ejemplo, las grasas poliinsaturadas y monoinsaturadas son analizadas de forma muy diferente.
• Las grasas también sirven como un buffer muy útil hacia una gran cantidad de enfermedades.

Page 105
Importancia para los organismos vivientes • Cuando una sustancia particular sea química o biotica,
alcanza niveles no seguros en el torrente sanguíneo, el organismo puede efectivamente diluir (o al menos mantener un equilibrio) las sustancias dañinas almacenándolas en nuevo tejido adiposo.
• Esto ayuda a proteger órganos vitales, hasta que la sustancia dañina pueda ser metabolizada y/o retirada de la sangre a través de la excreción, orina, sangramiento accidental o intencional, excreción de cebo y crecimiento del pelo.
• Aunque es prácticamente imposible remover las grasas completamente de la dieta, sería equivocado hacerlo.

Page 106
Importancia para los organismos vivientes • Algunos ácidos grasos son nutrientes esenciales,
significando esto que ellos no pueden ser producidos en el organismo a partir de otros componentes y por lo tanto necesitan ser consumidos en pequeñas cantidades.
• Todas las otras grasas requeridas por el organismo no son esenciales y pueden ser producidas en el organismo a partir de otros componentes.

Page 107
Tejido adiposo
• El tejido adiposo o graso es el medio utilizado por el organismo humano para almacenar energía a lo largo de extensos períodos de tiempo.
• Dependiendo de las condiciones fisiológicas actuales, los adipocitos almacenan triglicéridos derivadas de la dieta y el metabolismo hepático o degrada las grasas almacenadas para proveer ácidos grasos y glicerol a la circulación.
• Estas actividades metabólicas son reguladas por varias hormonas (insulina, glucagón y epinefrina).

Page 108
Tejido adiposo
• La localización del tejido determina su perfil metabólico.• La grasa visceral está localizada dentro de la pared
abdominal (debajo de los músculos de la pared abdominal) mientras que la grasa subcutánea está localizada debajo de la piel (incluye la grasa que está localizada en el área abdominal debajo de la piel pero por encima de los músculos de la pared abdominal).

Page 109
METABOLISMO DE LÍPIDOS
Ing. Leonardo Avellán VásquezULEAM
2010

Page 110
• Al igual que en el metabolismo de los carbohidratos, el metabolismo de lípidos consiste en:– Digestión– Transporte– Almacenamiento– Degradación– Biosíntesis
METABOLISMO DE LÍPIDOS

Page 111
• Los lípidos más abundantes en los alimentos son los aceites y las grasas.
• Ambos son triacilgliceroles (triglicéridos).• Los otros componentes más abundantes son
fosfolípidos.• Los lípidos de la dieta deben ser degradados en el
intestino a ácidos grasos para su absorción por el epitelio intestinal.
• La digestión de los lípidos ocurre en las interfases lípidoagua.
• En el lumen intestinal son incorporados en micelas formadas con la ayuda de las sales biliares.
• El enlace éster de los triacilgliceroles y fosfolípidos en las micelas está orientado hacia el exterior, permitiendo su hidrólisis por lipasas solubles secretadas por el páncreas (lipasa y fosfolipasa A2).
DIGESTIÓN DE LÍPIDOS

Page 112
• Son moléculas anfipáticas sintetizadas en el hígado a partir de colesterol y secretadas por la vesícula biliar.
• Facilitan la digestión de las grasas en el intestino.
SALES BILIARES

Page 113
• Las lipasa pancreática digiere los triacilgliceroles en ácidos grasos y monoacilglicerol, hidrolizando los enlaces ésteres.
• Su mecanismo catalítico es parecido al de las proteasas de serina.
DIGESTIÓN DE LÍPIDOS

Page 114
• Las micelas que contienen a los productos de la digestión son absorbidas por las células de la mucosa intestinal a través de la membrana plasmática.
• En estas células de la mucosa intestinal se resintetizan los triacilgliceroles.
• Para su transporte, los triacilgliceroles se empacan en partículas de lipoproteínas llamadas quilomicrones que se liberan al sistema linfático, desde donde pasan a la sangre.
• Los quilomicrones contienen fundamentalmente un tipo de proteína: la apolipoproteína B-48.
• Los quilomicrones también funcionan en el transporte del colesterol y de las vitaminas liposolubles (A, D, E y K) ingeridos en la dieta.
ABSORCIÓN Y TRANSPORTE DE LÍPIDOS

Page 115
DIGESTIÓN, ABSORCIÓN Y TRANSPORTE
DE LÍPIDOS

Page 116
• Los triacilgliceroles sintetizados en el hígado se transportan por la sangre en otro tipo de lipoproteínas llamadas lipoproteínas de muy baja densidad (VLDL)
• Los ácidos grasos se transportan en complejo con la albúmina sérica, que es la proteína má abundante del plasma sanguíneo.
• Las sales de Na+ o K+ de los ácidos grasos son jabones, por lo que si exceden una cierta concentración en forma libre son muy tóxicos.
• Los lípidos también se transportan a través de la sangre como cuerpos cetónicos, que son moléculas producto del catabolismo de los ácidos grasos y que se usan como fuente de energía en los tejidos periféricos bajo determinadas condiciones.
TRANSPORTE DE LÍPIDOS

Page 117
• Los quilomicrones y las VLDL se unen a las lipoproteín lipasas de la membrana plasmática de las células de músculo y tejido adiposo, fundamentalmente.
• De nuevo, los triacilgliceroles se degradan por una lipoproteínlipasa a ácidos grasos y monoacilglicerol para ser incorporados a las células.
• El glicerol se transporta al hígado o al riñón.• En las células de músculo y tejido adiposo se
resintetizan los triacilgliceroles y se almacenan.• La composición de la grasa almacenada, es decir su
proporción relativa en mono, di o triacilgliceroles y el tipo de ácido graso que contienen, depende del organismo.
• La transformación de las grasas de la dieta en las grasas características de cada organismo la realiza el hígado.
ALMACENAMIENTO DE LÍPIDOS

Page 118
• La mayor parte de la célula está ocupada por un gran depósito de triacilglicéridos.
ADIPOCITO

Page 119
• Se llama movilización al proceso de liberación de los ácidos grasos de la grasa almacenada cuando se necesita degradarla para producir ATP.
• La movilización de los ácidos grasos está regulada por una cascada controlada por hormonas (adrenalina y glucagón), semejante a la que regula el metabolismo de carbohidratos.
• Los triacilgliceroles se hidrolizan a glicerol y ácidos grasos por una triacilglicerol lipasa sensible a hormonas.
• Una vez que se liberan los ácidos grasos en el tejido adiposo, difunden a través de la membrana celular y se transportan al hígado unidos a la albúmina.
DEGRADACIÓN DE LÍPIDOS

Page 120
• Una excesiva movilización de los ácidos grasos puede llevar a la formación de un hígado graso, que contienen una gran proporción de tejido graso no funcional.
• El hígado graso también puede resultar de la exposición a químicos que destruyen las células hepáticas, las cuales son reemplazadas por tejido graso.
• Una deficiencia de colina y metionina puede también producir hígado graso, porque se produce una deficiencia en la síntesis de fosfolípidos y por tanto de lipoproteínas, afectándose así el transporte de los lípidos desde el hígado.
HÍGADO GRASO

Page 121
• La degradación y la síntesis de los ácidos grasos son
• procesos sencillos esencialmente uno el reverso del otro.
• Los cuatro pasos que son opuestos en la degradación y en la síntesis son:– Oxidación (reducción)– Hidratación (deshidratación)– Oxidación (reducción)– Ruptura de enlace (condensación)
• Se parte siempre de grupos acilo activados, por lo que ambos procesos comienzan con reacciones de activación.
DEGRADACIÓN Y SÍNTESIS DE ÁCIDOS GRASOS

Page 122
DEGRADACIÓN Y SÍNTESIS DE ÁCIDOS GRASOS