
NOMBRE: CASTILLO ROBLES FLOR PATRICIA EXPERIMENTAL DE Y148.206.53.84/tesiuami/UAM4166.pdf ·...
28
NOMBRE: CASTILLO ROBLES FLOR PATRICIA MATRICULA: 8525 1465 LICENCIATURA: BIOLOG~A EXPERIMENTAL DIVISI~N DE CIENCIAS BJOL~GICAS Y DE LA SALUD UNIDAD IZ’TAPALAPA TELÉFONO: 724-47-3 1 D~A/SEMANA 4/2 O TRIMESTRE: 97 - I TITULO: 6 6 ~ ~ Á ~ ~ ~ ~ ~ DE LOS COMPONENTES DE MEMBRANA DE ERITROCITOS DE NIÑOS QUE SUFREN DESNUTRICI~N CAL@~ICO PROTEICA SEVERA., .:,i,,b,’ ~, -.; I. t .- , . i“ *: . Nombre del asesor: CAROLINA CAMPOS MU PROFESOR TITULAR“B” DPTO. CIENCIAS DE LA S . ,. LUGAR DE I . REALIZACI~N: UNIVERSIDAD AUT~NOMA METROTTX,I’I’ANA UNIDAD IZTAPALAPA DIVISIÓN DE CBS DPTO. CIENCIAS DE LA SALUD LAB. DE NEUROFISIOLOGIAR- 1 18. FECHA DE INICIO: Septiembre 5 de 1996 FECHA DE TERMTNACIÓN: Marzo 5 de 1997 CLAVE: FIRMAS: ALUMNA: ASESOK!, /’ /
Transcript of NOMBRE: CASTILLO ROBLES FLOR PATRICIA EXPERIMENTAL DE Y148.206.53.84/tesiuami/UAM4166.pdf ·...

NOMBRE: CASTILLO ROBLES FLOR PATRICIA MATRICULA: 8525 1465 LICENCIATURA: BIOLOG~A EXPERIMENTAL
D I V I S I ~ N DE CIENCIAS B J O L ~ G I C A S Y DE LA SALUD UNIDAD IZ’TAPALAPA
TELÉFONO: 724-47-3 1
D~A/SEMANA 4/2 O TRIMESTRE: 97 - I
TITULO: 6 6 ~ ~ Á ~ ~ ~ ~ ~ DE LOS COMPONENTES DE MEMBRANA DE ERITROCITOS DE NIÑOS
QUE SUFREN DESNUTRICI~N CAL@~ICO PROTEICA SEVERA., . : , i , ,b , ’ ~, -.; I. t .-
, . i “ *: .
Nombre del asesor: CAROLINA CAMPOS MU PROFESOR TITULAR “B” DPTO. CIENCIAS DE LA S
. ,.
LUGAR DE I .
R E A L I Z A C I ~ N : UNIVERSIDAD A U T ~ N O M A METROTTX,I’I’ANA UNIDAD IZTAPALAPA DIVISIÓN DE CBS DPTO. CIENCIAS DE LA SALUD LAB. DE NEUROFISIOLOGIA R- 1 18.
FECHA DE INICIO: Septiembre 5 de 1996 FECHA DE TERMTNACIÓN: Marzo 5 de 1997
CLAVE:
FIRMAS:
ALUMNA: ASESOK!, /’ /

INDICE INTRODUCCI~N.
Tipos de desnutrición.
Efectos a nivel metabólico en la DCPS.
Composición de las membranas.
Fosfolípidos.
Colesterol.
Proteínas.
Carbohidratos.
Principales polipéptidos de la membrana del eritrocito. Espectrina y ancrina. Banda 3. Glucoforina. Distribución asimétrica de enzimas en la membrana.
ANTECEDENTES.
OBJETIVO GENERAL.
OBJETIVOS ESPECÍFICOS . M E T O D O L O C ~ .
Obtención de muestras biológicas.
Obtención de eritrocitos y membranas.
Cuantificación de proteínas.
Cuantificación de Colesterol en membranas plasmáticas.
Extracción de Fosfolípidos.
Separación de Fosfolípidos individuales.
Cuantificación de fosfolípidos por contenido de fósforo.
Medición de la fluidez membranal (anisotropía).
Cuantificación de la ATPasa de Na' / K'.
Cuantificación de la S'Nucleotidasa.
Estadística.
ACTIVIDADES RmLIZADAS . OBJETIVOS Y METAS ALCANZADAS.
RESULTADOS Y CONCLUSIONES.
RECOMENDACIONES.
BIBLIOCRA FIÁ.
TABLA I.
TABLA II.
TABLA III.
TABLA IV.
T4BL4 V.

A la Desnutrición Calórico Protéica Severa (DCPS) se le ha definido como el conjunto de síntomas, signos clínicos y alteraciones a nivel bioquímico que se observan a consecuencia de una ingestión deficiente y/o utilización de dietas en contenido variable calórico y bajo contenido protéico (Arrieta y Cravioto 1977).
Tipos de desnutrición:
En México se ha considerado de manera general la clasificación de la desnutrición sugerida por Gómez (1946) y Ramos-Galván (1 976), en la que se definen tres grados de desnutrición basados en la edad, peso y talla del niño: como primaria, secundaria, terciaria (o mixta).
En la desnutrición severa de tercer grado, se han observado dos formas identificadas por sus características anatomopatológicas las cuales son:
Marasmo: Esta se presenta en niños mayores de un año por la dieta baja en calorías y proteinas plasmáticas.
Kwashiorkor: Tiene un elevada incidencia en niños de 2 a 4 años de edad. Es un síndrome provocado por una deficiencia protéica severa y un aporte rico en carbohidratos.
Efectos u nivel metabólico en Iu DCPS.
Mediante el análisis fisiológico y bioquímico, se han demostrado cambios metabólicos que son típicos del Marasmo y Kwashiorkor.
La DCPS está caracterizada por la movilización de carbohidratos y triglicéridos almacenados. Una respuesta temprana a la privación de alimentos es la glucogenólisis a nivel hepático y la lipólisis en el tejido adiposos para generar glucosa y ácidos no esterificados, que ocurre como resultado de la disminución en la relación insulindglucagon que también influye en la lisis del tejido adiposos, los que se oxidan para obtener energía (Sugden, et al., 1989).
En la Rehabilitación Nutricional se lleva a cabo el restablecimiento de la tasa glucolítica, glucogenolítica y lipolítica normal y en el hígado en particular, es estimulada la c gluconeogénesis y glucogenogénesis (Holness y Sugden, 1987).

En niños con Kwashiorkor hay una disminución en el contenido de albúmina y globulinas séricas, en comparación con los niños que presentan Marasmo (Whithead, 1968). En niiios con Marasmo se ha observado baja la concentración de los aminoácidos esenciales, sin embargo en los no esenciales se encontró normal o elevada (Bjornesjo, et al., 1968).
Respecto a las concentraciones de aminoácidos libres en el plasma sanguíneo. los niños con Kwashiorkor muestran proporciones anormales en la relación de fenilalanindtirosina, este mismo efecto se observa en la orina (Cravioto, 1962).
En niños con DCPS se han reportado concentraciones de cortisol elevadas, indicando que este compuesto favorece la síntesis de proteínas y al mismo tiempo provoca un balance negativo de nitrógeno en todo el organismo. Los niveles de cortisol aumentan considerablemente con la DCPS, su tasa de degradación está reducida y existe una mayor exposición de los tejidos a este compuesto, lo cual permite asimilar mejor las proteínas de los tejidos (Waterlow, 1968).
Los niños con Kwashiorkor presentan infiltración de grasa en el hígado, esto altera el transporte de grasas del hígado a tejidos periféricos, lo cual probablemente tenga su origen en la reducida síntesis de proteínas o de sus precursores, las apoproteínas (Monckeberg, 1968).
Holman, et d . , (1981), cuantificaron el contenido de lípidos totales séricos en niños con Marasmo 1, Kwashiorkor, no encontrando diferencia entre ambos grupos. Sin embargo, detectan en los nifios con Kwashiorkor una mayor proporción de ácidos grasos saturados y sugieren una alteración en la actividad de las enzimas involucradas en la saturación.
Pineda (1968), observó en neutrófilos aislados de nifios con Kwashiorkor elevada la actividad de la hmarasa y un decremento en la actividad de la isocitrato deshidrogenasa y en la malato deshidrogenasa. Sin embargo, en niños con Marasmo disminuye la actividad de la fumarasa y la malato deshidrogenasa y hay un aumento en la isocitrato deshidrogenasa.
Hansen (1 968), reportó en niños con Kwashiorkor y con Marasmo una reducción en la secreción de quimiotripsina y tripsina y plantea que la DCPS puede producir daño pancreático irreversible.
A nivel celular se han reportado diversos daños en la DCPS, sin embargo, nuestro estudio se basa en las membranas de los eritrocitos, ya que es fundamental para las hnciones metabólicas que realizan, y es el organelo el que cumple las funciones de separar ambiente externo del interno y obtener información extracelular para inducir cambios.
Composicidn de Ius membranas.

Las membranas están constituidas por lípidos, proteínas y carbohidratos, en los modelos de organización de este organelo, a los lípidos (Spector y Yorek, 1985) se les ha asignado un papel estructural y de segundos mensajeros, sirviendo de barrera básica y matriz de anclaje para las proteínas, que cumplen diversas funciones y de carbohidratos cuyo papel es de receptores, ( Stubbs, 1983).
Fosfolbidos
Los fosfoacilgliceroles, son moléculas anfipáticas que contienen en su estructura, una región polar y otra no polar unidas por una región de polaridad intermedia. La mayoría de las membranas muestran heterogeneidad estructural, en las cabezas polares, longitud y grado de instauración de las cadenas de ácidos grasos (Yeagle, 1989).
Los fosfolípidos pueden establecer dominios hidrofilicos o hidrofóbicos que difieren en su composición dinámica. Estos dominios determinan agrupamientos conformando bicapas (Bennett, 1982).
Colesterol
La estructura del colesterol está formada por cuatro anillos fusionados, que producen una apariencia plana de la molécula. El sistema de cuatro anillos no está confinado a una conformación, ya que el primer anillo, puede adoptar una conformación alterna en cristal, además de haber una mayor flexibilidad en la cola no cíclica de la molécula (Voet y Voet, 1992).
Las propiedades de la membrana pueden ser alteradas por la disminución o aumento en la concentracih de colesterol, así como de ácidos grasos, ya que afecta la permeabilidad a la glucosa en sistemas naturales y artificiales (Yeagle, et al., 1977).
Así mismo, puede estar involucrado en la modulación de la actividad de proteínas integrales de la membrana. Por ejemplo, la ATPasa Na'K' y el intercambiador aniónico de membrana de los eritrocitos son inhibidos por el colesterol en sistemas reconstituidos. Mientras, el intercambiador ATP/ADP, el acarreador del lactato y la arabinosa son estimulados por el colesterol (Grunze, et al., 1980; Skou y Norby, 1979).
Proteirlas
Las Proteínas de las membranas pueden estar insertadas en la bicapa lipídica (integrales) o asociadas a ellas (periféricas). El interior de la bicapa es un ambiente energéticamente desfavorable para los enlaces peptídicos, por esta razón, las proteínas

insertadas en la bicapa presentan en las regiones transmembranales aminoácidos hidrofóbicos que adoptan preferencialnlente la confirmación de alfa-hélice (Spector, 1985), que es energiticamente favorable. Aunque hay casos excepcionales, como el de la porfirina, que presenta configuración de barril B (Jennings, 1989).
En los extremos transrnembranales, se presentan en su mayoría aminoácidos cargados, que mantienen a las proteínas en esta posición por el efecto hidrofóbico, ya que las regiones cargadas dificilmente pueden penetrar en el interior de la membrana (Singer and Nicolson, 1972).
Las proteínas integrales que realizan funciones de transporte transmembranal están colocadas asimétricamente en la membrana y su orientación es una dirección perpendicular al plano de la bicapa (Singer and Nicolson, 1979), y pueden interactuar con elementos del citoesqueleto durante la locomoción amiboidea, fagocitosis, pinocitosis y el movimiento de receptores durante la formación de los casquetes en linfocitos (Rothman y Jenard, 1977).
La mayoría de las membranas plasmáticas contienen carbohidratos unidos a proteínas y lípidos (glicoproteínas o glicolipidos) (Gitler, 1972), los cuales están orientados hacia el exterior de la célula. Los eucariontes presentan como componente principal ácido siálico que contribuye a la carga neta negativa típica en este tipo de célula (Yohe y Rosenberg 1977).
Las glicoproteínas membranales son capaces de reconocer carbohidratos extracelulares (Voet y Voet, 1992), por ejemplo, los hepatocitos se unen a geles con residuos de carbohidratos sobre superficies de plásticos cubiertas con análogos de glicolípidos (Blackbum y Schnaar, 1983).
Su localización y diversidad estructural hace que los carbohidratos actúen como antenas sensibles que le permite a la célula detectar señales extracelulares (Sharon, 1979). En general se considera que los carbohidratos participan en h c i o n e s celulares de polarización, comunicación, control de crecimiento celular, diferenciación y relación célula-célula (Spiegel, et al., 1985).
Principnles polipéptidos de la membrana del eritrocito.
Espectrina y ancrina.
La espectrina representa la masa más importante del citoesqueleto. Está compuesta por dos cadenas polipeptídicas no idénticas (a, de 240,000 dalton y p, de 220,000 dalton) y puede

encontrarse en forma de dímero (a, p) tanto como de tetrámero (a, p)2. Vista con el microscopio electrónico la molécula de espectrina aparece formada por dos cadenas entrelazadas en forma laxa y flexible de unos lOOnm (el tetrámero tiene 200nm de longitud). La espectrina se une a la ancrina (banda 2.1) que a su vez , está asociada con la banda 3 . De esta manera la espectrina del citoesqueleto queda anclada a la membrana. La espectrina tiene también sitios de fijación para la banda 4.1 y la actina (banda 5), que se encuentra en forma de oligómeros de aproximadamente 10 monómeros. De tal modo podemos decir que el citoesqueleto del eritrocito es una malla compuesta por tetrámeros de espectrina, junto con actina y la banda 4. l .
Es posible observar el citoesqueleto del eritrocito con el microscopio electrónico después de la fijación con el ácido tánico y glutaraldehído y también con el microscopio electrónico de barrido. Esta red filamentosa contiene estabilidad a la membrana plasmática y también puede controlar la forma característica del eritrocito.
Existen varias enfermedades hereditarias en las que hay una alteración de la forma bicóncava típica de los eritrocitos. Por ejemplo, los eritrocitos pueden ser esféricos (esferocitosis) o elipsoidales (elipsocitosis) (Cohen y Branton, 1981). Se ha descubierto que en dichas enfermedades existen alteraciones del citoesqueleto del eritrocito, por ejemplo, en la esferocitosis la espectrina tiene menor capacidad de experimentar la transformación de dímero a tetrámero. por otro lado, en algunos casos de elipsocitosis hay un defecto en la unión de la banda 4.1 a la espectrina, o aun la falta total de 4.1 (Fairbanks y Theodore 1971 ; Bennett, 1982).
Banda 3.
La proteína principal intrínseca (banda 3) tiene un peso molecular de 93,000 dalton, atraviesa el espesor de la membrana y tiene poca cantidad de hidratos de carbono en el polo de la superficie externa. Esta proteína forma dímeros unidos por uniones S-S. Hay 500,000 a 600,000 de tales dímeros por célula, número suficiente para explicar el de las partículas de 8nm que se observan por el método de congelación fractura. Esta proteína puede estar relacionada con el transporte de aniones (Fairbanks y Theodore, 197 1).
Glucoforina.
Esta proteína tiene un peso molecular de 55 Kd, el 60 % corresponde a los carbohidratos. Cerca del extremo CO-OH de la molécula existe una región muy hidrofóba que interactua con los lípidos de la membrana. En el extremo M;! es más hidrofilico, y está expuesto al medio externo. A éste se unen los oliogosacáridos que están en la superficie externa de la membrana.

Esta glucoproteína contiene determinantes antigénicos para los grupos sanguíneos A,B,O y otros, como los grupos M N que reaccionan con antisueros de conejo, los que reaccionan con el virus del germen de trigo. Se calcula que en cada eritrocito humano existen unas 500,000 copias de esta proteína. La glucoforina comprende el 80% de los carbohidratos y el 90?/0 del ácido slámico negativamente cargado presentes en la superficie celular.
Entre las proteínas más hidrofóbicas de la membrana se encuentran los proreolípidos, que se caracterizan por su fuerte asociación con los lípidos y por su solubilidad en los solventes orgánicos. Los proteolípidos fueron aislados al principio de la mielina por Folch y Lees. pero se los halla practicamente en todas las membranas celulares y en algunas representan proteínas receptoras para los transmisores sinápticos o .para establecer canales a través de las membranas (Fairbanks y Theodore, 197 1).
Distribución asimétrica de enzimas en la membrana.
En membranas plasmáticas aisladas se han detectado una amplia variedad de enzimas. Las que se encuentran con mayor constancia son la S'nucleotidasa, la Mg cc ATPasa, la Na'K' ATPasa, la fosfatasa alcalina, adenilciclasa, fosfomonoesterasa ácida y la RNAasa. Para algunas de estas enzimas se ha propuesto una localización específica, con una disposición en mosaico. La disacaridasa forma unidades globulares de 5 a 6 nm que cubren la membrana de las microvellosidades.
De todas las enzimas asociadas a la membrana, la ATPasa Na'K' es una de las más importantes por su papel en la transferencia de iones a través de la membrana plasmática.
La actividad de esta enzima depende de la presencia de lípidos y es inactivada cuando todos los lípidos son extraídos.
Las enzimas también muestran una distribución asimétrica. Por ejemplo, en la superficie externa se encuentran:aceticolinesterasa, nicotinamida, adeninadinucleotidasa y los sitios de unión de la ouabaína para la ATPasa Na'/K+ dependiente. En la superficie-intema se encuentran. NADH-diaforasa, adenilciclasa, proteinaquinasa, y ATPasa Mg'+ (J$rgensen, 1974; Walter, 1990).
ANTECEDENTES.
Las alteraciones clínico-bioquímicas observadas en niños, que han sido provocadas por ja falta de nutrientes esenciales para un desarrollo y crecimiento óptimo, han sido determinadas durante varias décadas (Krishnaswamy, 1989).

Sin embargo, las posibles modificaciones en membranas celulares de distintos tejidos, no ha recibido la misma atención. A pesar de la importancia que esta tiene en la funcionalidad de las membranas biológicas.
Las células que con mayor énfasis, se emplean en el estudio de las membranas biológicas, son de los eritrocitos de mamífero; estas catalizan diferentes tipos de reacciones enzimáticas y contiene un alto espectro de lípidos y proteínas, que pueden encontrarse en otros tipos celulares. Por su grado de especialización funcional, los eritrocitos no presentan organelos, por tanto sus membranas pueden ser aisladas con cierta facilidad. Balduini, et al., (1 976) sugieren que la composición y estructura de la membrana juegan un papel fundamental en la vida media del eritrocito y por tanto en su funcionalidad.
En algunos reportes sobre el efecto de la DCPS en eritrocitos de niños , se ha reportado un aumento en la resistencia a la lisis osmótica y aumento en la actividad de la ATPasa Na'/K' (Kaplay, 1978). En niños de tipo Marasmo y Kwashiorkor se ha observado una disminución en la actividad de la enzima glioxilasa, misma que está involucrada en la detoxificación metabólica, esta requiere de glutation como cofactor para que realice su función, sin embargo, esta no es afectada cuando existe el Síndrome Marasmo-Kwashiorkor (Ramdath y Golden, 1995).
Por otro lado, la glutation S-transferasa en eritrocito de niños con DCPS es activada, esta inducción puede ser el resultado de la exposición de los niños a un estres oxidante durante los meses en que se presenta la desnutrición (Ramdath y Golden, 1993).
La reducción del glutation juega un papel muy importante en el cuadro clínico .e historia natural de la desnutrición (Forrester-T., et al., 1990), esta disminución afecta también la actividad de la bomba de sodio.

OBJETIVO GENERAL.
Evaluar el estado nutricional de los niños que sufren Desnutrición Calórico Protéica Severa (DCPS), tomando como base la Membrana del Eritrocito.
OBJETIVOS ESPECÍFICOS .
Evaluar el estado nutricional de los niños desnutridos a nivel bioquímico en suero, tomando como parámetros: Proteínas totales, Albúmina, Urea, Lípidos y Colesterol.
Determinar el contenido de Fosfolípidos, Colesterol y fluidez en membranas de eritrocitos provenientes de niños que sufren desnutrición calórico protéica severa (DCPS).
METODOLOGÍA.
Obtención de muestrns biológicas.
La presente investigación fue realizada en 20 niños de entre seis meses y dos años de edad en la Ciudad de México en el Hospital de Recuperación Nutricional Cruz Blanca (Coyoacán) Los niños fueron agrupados en eutróficos (n = 10) y niños con DCPS (n = lo), de tipo Marasmo, según su estado nutricional, el cual fue establecido al comparar el peso y talla observado en ellos, con valores para sexo y edad cronológica de acuerdo con la tabla de medidas somatométricas propuesta por Ramos-Galván (1 976). Además, se realizó, un análisis químico en plasma, cuantificando proteínas totales, albúmina, lípidos totales, colesterol y urea. Todas las determinaciones heron realizadas usando equipos comerciales (Wiener Lab. Argentina).

Obtención de eritrocitos y membranas.
Por cada niño se colectaron 3 ml de sangre periférica con jeringa heparinizada, se sometió a centrifugación de un gradiente de densidad en Ficoll-Hypaque a 2000 rpm durante 15 min; se retiraron el plasma y los linfocitos. El botón con eritrocitos. se lavó dos veces con NaHP0124, (5mM) NaCl (150 IIM) y después, las membranas de los eritrocitos se centrifugando a 90,OOOXg durante 2.5 lus, (Dodge, et al., 1963).
Cuantificación de proteínas.
Una vez aisladas las membranas, se cuantificó el contenido protéico de acuerdo al método de Lowry modificado Markwell, et al., (1978) y el contenido de 1 mg de proteína sirvió como referencia para todas las mediciones, se realizó una curva de calibración de proteínas usando como proteína estándar, albúmina sérica bovina (BSA), y colocando en tubos de ensayo 25,50,75 y 100 pg/ml ajustando a un volumen final lml de agua destilada.
Se adicionó una mezcla de reacción conteniendo, carbonato de sodio (2%), hidróxido de sodio (0.4%), tartrato de sodio y potasio (0.16%), sdfato de cobre (0.4%) y dodecil sulfato de sodio (SDS) YO).
La disolución se agitó e incubó durante 10 minutos a temperatura ambiente. Después se adicionaron 0.3 ml. de reactivo de Folin-Cicalteau (2N) diluido 1 :1 con agua destilada; al final la solución se incubó a temperatura ambiente por 40 minutos. Se leyó la absorbancia a 660nm.
Para determinar el contenido en las muestras problema, se tomaron lop1 de la suspensión de membranas plasmáticas aisladas y se llevaron a 1 ml, siguiendo la adición de los reactivos antes descritos para la curva de albúmina.
CuantiJicación de Colesterol en membranas plasmáticas.
El contenido de colesterol en las membranas plasmáticas, se determinó según lo propuesto por Dittmer y Wells (1969). Se colocaron en tubos de ensayo alícuotas de las membranas plasmáticas a una concentración de lmg de proteína y como estándar una solución patrón de colesterol a concentración de 300 mddl.
A cada tubo se adicionó 1 m1 de mezcla de anhídrido acético (6.33 dl) en ácido acético (99%), se incubó a una temperatura ambiente durante 30 minutos y se añadieron 0.2 m1 de H2SO4 (97%), se colocó la solución en baño de agua a 25°C y se leyó la absorbancia de las muestras a 530 run.

Extracción de Fosfolbidos
Los fosfolípidos de membrana, se extrajeron por el método de Blingh y Dyer, (1959) de la siguiente manera: a lmg de suspensión membranal, se le adicionaron 5.0 ml. de la mezcla clorofomo: metano1 (2: 1 v/v), que se mantuvo durante 10 min. en agitación constante. Se centrifugó a 550 X g durante 10 min. a 4°C para eliminar las proteínas pecipitadas y otros residuos celulares. El sobrenadante se transfirió a otro tubo donde se adicionaron 1.25 ml de cloroforno y 0.5 m1 de agua destilada. La solución se mezcló y posteriormente se centrifugó a 550 X g durante 10 min. para separar las fases orgánica y acuosa (Marchellesi y Bazan, 1970).
La fase orgánica, se obtuvo con una pipeta Pasteur y se colocó en un tubo cónico graduado y se ajustó el volumen a 1 m1 con cloroformo. Los solventes se evaporaron bajo atmósfera de nitrógeno, para realizar la separación de los fosfolípidos.
Separación de FosfoI&idos individuales
Los fosfolípidos se separaron por cromatogafia en capa fina en dos dimensiones siguiendo el método de Rouser, et al., (1970). El procedimiento, se inició con la saturación de la cámara de cromatografía para la primera dimensión con c1oroformo:metanol:hidróxido de atnonio en proporción 65:25:5 . Se colocaron 50 m1 del extracto lipídico sobre las placas de vidrio de 5 x 5 cm y 0.25 mm de espesor de silica gel Sephadex G60, se corrieron en la cámara para desarrollar la primera dimension. Después la placa se puso en un desecador al vacío durante toda la noche y posteriormente se corrió en la segunda dimensión con c1oroformo:acetona:metanol: ácido acético:agua en proporción 30:40: 1 O: 10:5. Posteriormente la placa se colocó en un desecador durante 3 horas; los fosfolípidos, se revelaron con vapores de lodo (1% en metanol) y en un papel celofán, se marcó el patrón de separación de los fosfolípidos.
Cuantifcación de fosfol@idospor contenido de fósforo
Se cuantificaron los fosfolípidos de los eritrocitos según el método propuesto por Dittmer y Wells (1969). Los fosfolípidos individuales se obtuvieron raspando la placa y colocando el polvo de silica gel en un tubo de ensayo. Las muestras se hidrolizaron añadiendo 1 .O m1 de ácido perclórico concentrado e incubado a 80°C durante toda la noche. Se centrifugó la mezcla de silica gel a 3000 rpm durante 15 minutos, se obtuvo el hidrolizado, que fue ajustado a lml con agua desionizada y se añadieron 0.25 m1 de solución conteniendo molibdato de anlonio (2.7%) y de ácido sulfiírico (2.5N). Después de agitar, se agregaron 0.065 m1 de la solución reductora (ácido 1,2.4- arninaftol sulfónico al 8%). Paralelamente, se preparó una curva patrón de fosfato entre 0.005 a 0.6 pmol/ml bajo las mismas condiciones experimentales que las muestras problemas y se leyó la absorbancia a 660 nm.

Medicirin de la fluidez membranal (anisotropía).
La fluidez en las membranas de eritrocitos fue determinada por la polarización de fluorescencia del difenil-hexatrieno (DPH). Para esto, el volumen de 200 mg de membranas, se ajustó a 1 m1 con solución salina amortiguadora de fosfatos (PBS), entonces fue adicionado el volumen necesario de DPH disuelto en dimetilformamida para obtener la segunda concentración final de 2 mg. Se incubó esta mezcla a 25" C y fue leída la intensidad de fluorescencia en un espectrofluorometro SLMAMINCO equipado con polarizadores especiales tomando 365 nm de longitud de onda emisión y 430 nm de excitación. La polarización de fluorescencia para cada grupo de eritrocitos fue obtenido utilizando la fórmula propue,sta por Shinistzki e lnbar ( 1974).
Cuantificación de la A TPasa de Na+ / 6.
Se determinó en las membranas aisladas, la actividad de la ATPasa (Na'/K+) empleando el método propuesto por Swam y Albers (19Sl), la actividad es medida como la velocidad de la liberación de fosfato inorgánico. Las soluciones de reacción fueron 3n1M MgC12, 3 m M ATP, 13OmM NaC1, 20mM KC1 30 mM histidina a pH=7.5 (37' C). Se equilibró por 5 min a 37"C, se añadieron 2.5 m1 de la suspensión de enzimas. Medio de reacción ( 5 mg de proteína o suspensión celular), la reacción se lleva a cabo en 5 min después de haberse liberado 0.1 mM de fosfato inorgánico (que corresponde a menos del 10% del ATP total), la reacción se detuvo con O. lml de TCA al 50%, los tubos se transfirieron a baño de hielo y se anadió lml de HzO, se centrifugó a 3000 rpmll0 min., se les adicionó 1 m1 de molibdato en HzS04 y lml de reactivo reductor, las absorbancias se leyeron a 660 nm .
Clrnntifcación de la 5'Nncleotidnsa
L a 5' nucleotidasa fue medida en 200 mg de proteínas como liberación Pi, que se obtuvo utilizando como buffer Verona1 40 mmolíl a pH= 7.5, conteniendo lOmmol /I de Mg
y como sustrato lmmol / 1 de AMP, se incubaron 1 O min y se leyó a 660 nm (Bergmeyer, 1965)
++
Estadística
Se realizó con los valores obtenidos la prueba de U de Mann Whitney para establecer el grado de significancia estadística.

ACTIVIDADES REALIZADAS.
1.- Obtención de las muestras y conocimiento de los diferentes tipos de desnutrición en el Hospital de Recuperación Nutricional Cruz Blanca (Coyoacán).
2.- Aislamiento de las membranas de eritrocitos humanos, para confirmar que efectivamente eran las membranas en nuestro estudio, vía las enzimas marcadoras de membrana.(ATPasa de Na' / K' y 5 'Nucleotidasa).
3.- Ensayos de cuantificación de proteínas por diversos métodos (Biuret, Bradford) y el dominio del método de Lowry modificado
4.- Obtención, aislamiento y cuantificación de fosfolípidos.
5.- Determinación de la fluidez realizada en el laboratorio de Bioquímica del Lnstituto de Fisiología Celular.
6.- Revisión bibliográfica sobre el tema del proyecto realizado durante los meses del servicio social.
7.- En todos los puntos anteriores se realizó la interpretación de los datos.
8.- Introducción a la computación
a) Paquetes estadísticos : Se aprendió el uso de Software estadísticos (SAS y NCSS), para aplicarlos en la interpretación de los resultados. Utilizando los métodos paramétricos y no paramétricos.
b) Procesador de textos : Se aprendió el manejo de paquetes computacionales como : Word, Works, Word Perfect.
c) Paquetes graficadores y de hojas de cálculo: Aprendió a utilizar paquetes computacionales , tales como: Excell, Sigma Plo, Harvard Graphics.

OBJETIVOS Y METAS ALCANZADAS.
Las metas alcanzadas del proyecto fueron el conocer la dinámica membranal y la interrelación que existe entre lípidos y proteínas, observamos que cuando algun componente no se encuentra en concentraciones biológicamente adecuadas, la dinámica cambia y como consecuencia su función.
Cuando las proteínas insertadas en membrana (5'nucleotidasa y la ATPasa de Na'K') son alteradas, la dinámica entre sus componentes, afecta la actividad física y fisiológica de la membrana por la variación de sus elementos.
En cuanto a los objetivos del trabajo se estimuló al alumno a que hiciera sus propios proyectos y que analizara sus datos, ya que dentro del mismo trabajo se plantearon proyectos de investigación como: La separaciónde las proteínas del eritrocito y el análisis de la funcionalidad y concentración de las mismas, así como la interacción con el citoesqueleto.
Dentro del trabajo se propusieron dos objetivos, que se cumplieron teoricamente gracias a las bases de datos que se emplean actualmente (artículos de investigación en revistas especializadas), así como a las visitas a otros laboratorios para la implementación de técnicas y experimentalmente fué el dominio de las técnicas e interpretación de los resultados.
Por otro lado la actualización bibliográfica del tema, así como el contacto con investigadores del mismo, fue de suma importancia para su desarrollo académico, así como la integración a un grupo de trabajo (Instituto de Fisiología Celular, U.N.A.M. e Instituto Nacional de Diagnóstico y Referencias Epidemiológicos Dr. Manuel Martinez Baez S.S.A).

RESULTADOS Y CONCLUSIONES.
Las mediciones corporales permiten identificar a niños con riesgo de desnutrición y distinguir sus formas crónica o aguda . Sin embargo, no proporcionan indicio de una posible deficiencia nutricional en calidad (Polberger, et al., 1990). Para un mejor diagnóstico en una evaluación nutricional, Sharma y Mahajan (1987) proponen que los parámetros bioquímicos junto con las mediciones somatométricas, se pueden emplear para separar, y diagnosticar tempranamente la Desnutrición y sus grados.
Los resultados bioquímicos en este estudio muestran en los niños que sufren DCPS (Tabla I), una disminución en el contenido de proteínas totales í ü é 5.06 iO.10, en comparación a los niños control que resultó 6.75 It O. 10 y en albúmina fué 2.88 * 0.10 y en niños sanos 4.1 i O. 10, este comportamiento ha sido previamente observado por Whithead (1968) quien encontró que los niños con Kwashiorkor muestran valores más bajos en albúmina, que aquellos niños con Marasmo. La hipoalbulinemia sérica observada, puede deberse a varios factores como puede ser la disminución en la síntesis de proteínas hepáticas (Sharma y Mahajan, 1987) y en la vida media de la albúmina plasmática (Polberger, 19901.
La evaluación bioquímica del estado nutricional de los niños se muestra en la Tabla I, donde se observa que en los niños desnutridos, el contenido de proteínas totales mostró disminución en 25% y en albúmina en 30% en relación al valor promedio de 4.1 g/dl determinado en los niños eutróficos. En cuanto al peso corporal hubo, disminución mayor al 40%. Evidenciando con los parámetros bioquímicos y somatométricos la presencia de esta DCPS. En los niños con DCPS el contenido de urea fue 100% mayor al observado en los niños eutróficos. En relación al contenido de lípidos en suero fué de 476.48 f 181.72 y en los niños control 366.58 Zt128.68, el comportamiento es diferente, ya que fue mayor el contenido de suero en niños con DCPS, en tanto el contenido de colesterol no se modificó por la desnutrición .
En algunos de los niños estudiados en este trabajo, se observó ía presencia de edema, característica física típica de la DCPS, que concuerda con las observaciones de Edozien (1960), quien además mostró que la albúmina en niveles bajos está asociada con la generación de esta fisiopatología.
En cuanto al aumento de urea en el suero (Tabla I), se ha observado que los niños que padecen DCPS mostraron O. 139 f O. 10 y niños control 0.065 f O. 10, Waterlow y Alleige ( 1 97 1 ). Mostraron una mayor eficiencia en la utilización del nitrógeno para sintetizar proteínas. Los eritrocitos de niños con DCPS que además muestran alteraciones en la composición lipídica en sus membranas plasmáticas, que de alguna manera pueden influir con su capacidad de transporte de gases respiratorios o bien con su vida media, como ha sido

sugerido por Balduini, et al., (1976). Graham ( 1 982). considera que la actividad de enzimas membranales (ATPasa Na'/K' y S'nucleotidasa) proporcionan un modo de identificar diferentes tipos de membranas y además evaluar el estado metabólico en algún tiempo en particular.
La composición lipídica en las membranas de los eritrocitos fué modificada por la DCPS. En la Tabla II, se muestra que el contenido total de fosfolípidos de 5.2 -f 0.79 y en los niños control fué 4.7 * 0.6, en relación al colesterol la diferencia significativa fué de 0.71 * 0.65 y en las membranas de niños control fué de 1 .O1 * 0.19. Esta concentración mayor en desnutridos esta asociada a la expanción de la membrana, existiendo desorden.
En la Tabla III se muestra la actividad de la ATPasa Na' / K' en membranas de eritrocitos, también se vio modificada por la desnutrición severa, ya que esta aumentó 100% (6.72 * 1.3) en comparación a 3.30 * 0.6 U/mg proteína obtenida en las membranas aisladas de niños control. Se ha sugerido que la composición de ácidos grasos de la membrana del eritrocito, genera un cambio en el metabolismo lipídico y afecta las propiedades físicas y fisiológicas de la membrana, que contribuyen a cambios en la actividad de las enzimas que se unen a la membrana, este efecto se observó en nuestros resultados y se ha sido reportado en niños con DCP tipo Kwashiorkor en donde se ve incrementada la fluidez membranal (Vajreswari-A, et al., 1990).
Se ha observado, que la ATPasa Na'K' en eritrocitos humanos puede ser inactivada por tratamiento con fosfolipasa y reactivada por fosfatidilserina y ácido fosfatídico (Kaplay, 1978).
La actividad de las membranas de los niños con DCPS está relacionado con el hecho de que niños que presentan edema es más alta (Kaplay, 1978). Se han realizado estudios para determinar el flujo de sodio y/o potasio utilizando ouabaina, en niños con desnutrición, no existiendo diferencia (Harper-M. E, et al., 1990).
Sin embargo, se ha sugerido que en algunos transtornos metabólicos la actividad de la 5' nucleotidasa aumenta (Tabla III), en los niños en DCPS está elevada (1.7k0.54) y parece estar afectada por el aumento de fosfatasa alcalina, las proteínas tienen su actividad en un microambiente lipídico; que para esta enzima es esfingomielina. Este efecto parece deberse a la modificación, en la afinidad de la enzima por el sustrato. Sin embargo, este efecto parece reflejar cambios en el microambiente lipídico en la cercanía de la enzima, (Dipple, et al., 1982), el cambio observado en nuestro estudio hé fosfatidilserina.
Los resultados de la polarización de fluorescencia (Tabla N), representan una medida de la dinámica membranal, ya que muestran una mayor rigidez en las membranas eritrocitarias provenientes de niños que padecieron DCPS (O. 107 f 0.005) en comparación a los niños control (0.43 * 0.014) . A este cambio se le ha atribuido a la composición de la membrana, ya

que está alterada la fluidez de los eritrocitos sin embargo la DCPS presenta un efecto tolerante (asociada a deficiencias así como a infecciones).
Las membranas de eritrocito de mamífero no están en un estado completamente fluido. El contenido de colesterol en membranas de eritrocitos manifiesta una fluidez intermedia entre completamente fluida a un estado líquido-cristalino, que puede ser importante en la interacción lípido-proteína y en la función de enzimas membranales y sistemas de transporte (Marion, et d . , 1990), así como el tipo de fosfolípido que rodea a la proteína.
En nuestro estudio, las membranas aisladas por choque osmótico permitieron estimar el grado de desorden de los fosfolípidos (fluidez). Dickens, et al., (1988) sugirio que las membranas aisladas de eritrocitos por este procedimiento reflejan el estado in situ de la membrana. Ya que, la estimación de la polarización de fluoresencia en eritrocitos completos tienen la desventaja, que la dispersión de la l u z puede dar resultados falsos, dado que la hemoglobina no permite el paso de l u z impidiendo medir la fluorescencia del DPH.
En la Tabla V se muestra el contenido de fosfolípidos, Leichsenring, et al., (1995), observaron que en niños con DCPS disminuye la concentración de fosfaticolina en niños con Marasmo y Kwashiorkor en nuestro estudio encontramos un aumento en fosfatilserina en niños con DCPS de tipo Marasmo (12.15% en comparación con los niños control que fue de 9.50%). Se ha reportado que el efecto en la concentración de los lípidos puede ser debida al tipo de infección, tiempo de desnutrición, periodos largos con antibióticos y el tipo de la misma.
Se sugiere que los cambios en la dinámica de las membranas sean debidos al efecto de la DCPS, así como a la interacción de componentes exógenos (antibióticos) con la membrana. El estudio en la dinámica de las membranas es complejo, por su alta variabilidad de los efectos producidos en sus componentes, por otro lado un solo elemento puede ser el responsable para alterar la función de las membranas biológicas.

RECOMENDACIONES.
Dado que la fluidez membranal de los eritrocitos es un proceso biológico, donde interaccionan lípidos y proteínas, es posible que en este trabajo el cambio en la dinámica membranal, esté dado por la diferencia en la concentración de fosfolípidos, como primera posibilidad, sin embargo, los ácidos grasos juegan un papel muy importante en la dinámica.
Se han realizado estudios sobre ácidos grasos en eritrocitos de niños con DCPS en los cuales se ha observado una diferencia significativa en grasas saturadas e insaturadas, la recomendación a este trabajo es medir la fluidez membranal ya que los anteriores estan involucrados en la dinámica de la membrana (Marin-M.C., et al., 1991; Cuevas-Covarrubias, et C J / . , 1994).
Por otro lado, las proteínas insertadas en la membrana (5’nucleotidasa y ATPasa de Na-/K+) tienen su función, en relación a su anillo lipídico, además de que están intimamente relacionadas con el citoesqueleto del eritrocito (se cree que es la espectrina), otra recomendación sería el estudio de las proteínas en concentración, funcionalidad y su relación con las proteínas del citoesqueleto (Branton, et al., 1981)
Durante la elaboración del proyecto surgieron varias preguntas sobre el trabajo, mismo que es factible continuar, como en la actividad enzimática que no se conoce si hay disminución en el número de enzimas o la misma proteína esta afectada.
Como propuesta a los futuros trabajos de investigación, sugerimos la aplicación de técnicas de investigación para el estudio de nuevos proyectos, así como el uso y actualización de paquetes computacionales.

BIBLIOGRAF~A. Arrieta, R. M y Cravioto, J. (1977). Desnutrición calórico protéica en el niño. En: Nueva guía para el diagnóstico y tratamiento del paciente pediátrico., (Rodriguez, M. ed.) Hospital del niño DIF México, pp. 149-1 82.
Balduini, C.L., Tira, M.E., Ascari, E. and Balduini, C. (1976). Glycopeptides of erythrocyte membranes in some hematology disorders. Acta Haemat.. 55:282-288.
Bennett. V. (1982). The molecular basis for membrane-cytoskeleton association in human erythrocytes, J. Cell. Biochem., 18:49.
Bermeyer, H.U. ( 1965). Methods of enzymatic analysis. Ed. Academics Press.Vo1. 2 pag. 87 l .
Bjomesjo, K., Mellander, O. and Jageenburg.R. (1968). The distribution of aminoacids between plasma and red cells in protein malnutrition. I n : Calorie deficiences and protein deficiences. Ed. J & A Churchill Ltd. London. pp 135-144.
Blackbum, C.C. and Shnaar, R.L. (1983). Carbohydrates-specific cell adhesion is mediated by inmobilized glycolipid, J. Biol. Chem., 258:1180-1188.
Blingh. E. G. and Dyer, W. J. (1959). A rapid method of total lipid extraction and purification. J. Biochem. Phsyol., 37: 91 1-917.
Branton, D., Chun, C.M. and Tyler, J. (1981). Interaction cytoskeletal proteins on the human erithrocyte membrane. Cell., 24:24
Cohen, C.M. and Branton, D. (1981). The normal and abnormal red cell cytoskeleton TIBS. 6:266.
Cravioto, J. (1962). Apprasial of the effect of malnutrition on biochemical malnutrition. Am. J . C h . Nutr., 1 1 : 484-492.
Cuevas-Covarrubias, S.A., Rojo Callejas, F., Juarez Oropeza, M.A., Díaz Zagoya, J.C., (1994). Erythrocyte fatty acids in children with severe protein calorie malnutrition., Rev. Invest. Clin., 46: 197-20 l .
Dickens. D.F., Snow.T.R., Green, V. and Weglicki, W.B (1988). The effect of erythrocyte associates light scattering on membrane fluorescence polarization. Mol. Cell. Bioch., 79:91-94.
Dittmer, J. C. and Wells, M. A. (1969). Quantitative and qualitative analysis of lipids and lipid components. Meth. Enzymol., 16: 486-487.
Dipple, Y., Gordon, L. M. and Houslay, M. D. (1982). The activity of 5’ nucleoxidese in liver plasma membrane is affected by increasein bilayer fluidity a chievied by anionic drugs but not by cationic drugs. J. Biol. Chem., 257: 181 1-1815.
Dodge, J.T., Mitchell, C. and Honahon. D.J. (1963). The preparation and chemical characteristics of hemoglobin-free ghost of human erythrocyte. Arch. Biochem. Biophys., 100: 1 19-130
Edozien, J.C. ( 1 960). The serum proteins in Kwashiorkor. J. Pediat., 54:594.
Fairbanks, G. and Theodore, L. (1971). Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochem., 10: 13.
15

Forrester, T., Golde, M., Brand, S. and Swales, J. (1990). Reduction in vitro of red cell glutathione reproduces defects of cellular sodium transport seen in oedematous malnutrition. Eur. J. Clin. Nutr.. 44: 363-369.
Gitler, C. (1972). Plasticity of biological membrane. Ann. Rev. Biophys. Bioengin., 1:51-91 Gómez. F. ( 1 946). Desnutrición Bol. Med. Hosp. Inf. (Mex)., III, 543.
Graham, J. ( 1 982). Fractionation of mammalian cell membranes in iodinated density gradient media. J. Biol. Cell., 45: 476.
Grunze, M., Forst, B. and Deticke, B. ( 1 980). Dual effect of membrane cholesterol on simple and mediated transport process in human erythrocytes. Biochem. Biophys. Acta., 600:860-869.
Hansen, J. (1968). Features and tratament of kwashiorkor at the cape. Calorie deficiences and protein deficiences Ed. J&A Churchill Ltd. London. pp 34-48.
Harper, M. E., Patrick, J. and Willis, J.S. (1 990). The absence of adapted sodium and potassium . in erythrocytes of cerebral palsied children with secondary malnutrition. Eur. J. Clin. Nutr., 44: 549-55s.
Holman, R.T., Jonhson, S.B., Mercuri, O., Itarte H.J. Rodrigo, H.A. and De Thomas, M.E. ( 1 981). Essencial fatty acid deficiency in malnourished children. Am. J. Clin. Nutr., 34:1534- 1539.
Holness, M.J. and Sugden, M.C. (1987). Adaptation of plasma protein synthesis and breakdown to malnutrition. Biochem. J., 247: 627- 634.
Jennings, M. L. (1989). Topography of membrane proteins. Annu. Rev. Biochem., 58:999-1027.
JQrgensen, L.P. (1 974). Purification of ma' + K') ATPase active site determinations and criteria of purity. Ann. Proc. Nat. Acad. Sci., 242 :36-52
Kaplay, S.S. (1978). Erythrocyte membrane Na', K' activated adenosine triphosphatase in protein-calorie malnutrition., Am. J. Clin. Nutr., 3 1579-584.
Krishnaswamy, K. (1989). Drug metabolism and pharmacokinetics in mahourished children. Clinical Pharmacokinetics., 17: 68-88
Leichsenring, M., Sutterlin, N., Less, S., Baumann, K., Anninos, A. and Becker, K. (1995). Polyunsaturated fatty acids in erythrocyte and plasma lipids of children with severe protein energy malnutrition., Act. Pediatr., 84: 5 16-520.
Markwell, A.K., Jaos, S., Baiber, L.L. and Tolbert, N. E. (1978). Lipoprotein samples: modified Lowry method. Anal. Biochem., 72:296-299.
Marchellesi, V.L. and Bazan, N.G. (1 990). Quantitative analysis of fatty acids in phospholipids, diacylglicerol, free acids, and other lipids. Nutr. Biochem., 1 : 382-389.
Marin, M. C., De-Tomas, M. E. , Mercuri, O., Fernandez, A. (1991). Interrelationship between protein energy malnutrition and essential fatty acid deficiency in nursing infants., Am. J. Clin. Nutr., 53: 466-468.
Marion, A.H., Johan, W.M. and Horstra, G. (1990). Membrane fluidity of non-activated and activatet human blood platelets. Bioch. Biophys., 1025: 173-178.
2

Monckeberg, F. (1968). Adaptation to calories and protein restriction in infants.Calorie deficiences and protein deficiences Ed. J&A Churchill Ltd. London pp 91-108.
Pineda, O ( 1 968). Metabolic adaptation to malnutritional stress. Calorie deficiences and protein deficiences Ed. J&A Churchill Ltd. London pp 75-89.
Polberger, S.K.T., Fex, G.A., Axelsson, I.A. and Raiha, N.C.R. (1990). Eleven plasma protein as a indicators of protein nutritional status in very low birth weigth infants. Pediatrics., 86: 91 6.
Ramdath, D. D., Golden, M. H. (1995). Glyoxalase 1 activity in erythrocytes from severely malnourished children., Med. J., 44: 24-27.
Ramdath, D. D., Golden, M. H. (1993). Elevated glutathione S-transferase activity in erythrocytes from malnourished children. Eur. J. Clin. Nutr. 47558-665.
Ramos -Galván R. (1 976). Somatometría Pediátrica, Arch. Inves. Med., (México), 6: l .
Rothman, J; and Jenard, J. (1977). Membrane Asymmetry Sci., 195 : 743-753.
Rouser, G., Fleischer, S. and Yamamoto, A., (1970). Two dimensional thin-layer chromatographic separation of polar lipid and determination of phosphorus analysis of spot Lipids., 15: 494-496.
Sharma, G. and Mahajan, M. (1987). Biochemical asessement of protein calorie malnutrition., Ind. J. Nutr. Dietet., 24: 199.
Sharon., N. (1 979). Glicoproteins in membranes. In: membranes and intracellular comunication., (Balian, R., Chabre, M and Devaux, P.F. eds.). North Holland, Amsterdam. pp 177-182.
Singer, S. J. (1979). The membrane. In: Membranes and intercellular comunication (Balian, R., Chabre, M. y Devaux, P.F eds.) Nort-Holland Publishing Co. Amsterdam pp 5-14.
Singer, S. and Nicolson, G.L. (1 972). The fluid mosaic model of the structure of cell membranes., Sci. 175: 720-73 l .
Shinistzky, Inbar M. (1974). Difference in microviscosity induced by different cholesterol levels in the surface membrane lipid layer of normal lymphocytes and malignant lymphoma cell. J. Mol. Biol., 85: 603-5.
Skou, J.C., Norby, J.G. (1979)., Na' / K' ATPasa structure and kinetics. New York, USA Academic Press. pp 47-49
Spiegel, S., Yamada, K.M., Hom, B.E., Moss, J. and Fishman, P.H. (1 985). Fluorescent gangliosides as probe for the retention and organization of fibronectin by ganglioside deficient mouse cell., J. Cell Biol. 100:721-726.
Spector, A.A. and Yorek, M.A. (1985). Membrane Lipid Composition and Cellular Function., J. Lipid Res., 26: 1015.
Stubbs, C. D. (1983). Membrane Fluidity: Structure and dynamics of membrane lipids. Essays in Biochemistry., 19 : 1-39.
Sugden, M. C., Holness, M.J. and Palmer, T.N. (1989). Fuel selection and carbon fluxes during the starved - to - fed transition. Biochem. J., 263,313-323.
Swann, A.C., Alberts R.W. (1975). Sodium and potassium activated ATPase of mammalian brain. Regulation of phosphatase activity. Biochim Biophys. Acta., 382: 435-56.
3

Vajreswari, A., Narayanareddy, K., Roa, P. S. (1990). Fatty acid composition of erythrocyte membrane lipid obtained from children suffering from kwashiokor and marasmus., Hyderabad.: Metabolism., 39: 779-82.
Voet. D. y Voet, J. (1992) Bioquímica. Ed. Omega pp. 1315.
Walter A. (1990). Membrane solubilization with and reconstitution surfactant solutions. a comparison of phosphoatidylserina and phosphatidylcholine interactions with octyl glucoside. Mol. Cell. Biochem., 99: 11 7-123.
Waterlow, J.C. and Alleinge, G.A.O. (1971). Protein malnutrition in children advances in Knowledga in the last ten years. Adv. Prot. Chem., 25,177-241.
Waterlow, J.C. (1968). The adaptation of protein metabolism to low protein intakes. Calorie deficiences and protein deficiences. (McCance, R.A. and Widdoeson, E.M.eds). J&A Churchill Ltd. London pp. 61 -73.
Whitehead, R. ( 1968). Biochemical. Changes in Kwashiorkor and Marasmus. Calorie deficiences and protein deficiences (McCance,R.A. and Widdowson, E.M. eds). Ed. J&A Churchuill Ltd. Londo. pp 109- 1 18.
Yeagle, P.L., Martin, R.B., Lala, A. K., Lin, H. and Bloch, K. (1977). Differential effects of cholesterol and lanosterol on artificial membranes. Proc. Natl. Acad. Sci. USA., 74:4924-4926.
Yeagle, P.L. ( 1 989). Lipid regulation of' cell membrane structure and füction., FASEB. J., 3: 1833- 1842.
Yohe, C. and Rosenberg, A. (1977). Action of intrinsic of rat brain synaptic membranes on membrane sialolipid and component in situ. J. Bioi. Chem., 242:2412-2418.
4

TABLA 1.
Evaluación bioquímica del estado nutricional d e niños severamente desnutridos y niños control
Características analizadas
Niños Niños Desnutridos Control (Marasmo) (Eutróficos)
No. de sujetos Edad (meses)
Seso F:h4 Total de proteínas ( g / 1 O0 ml)
Albúmina ( d l 0 0 ml)
Total de lípidos (md100 ml) Colesterol (mg/100 ml)
Urea
10 6 - 24 2 : 8
5.059 * 0.10 2.879 * 0.10
0.1389 += 0.10 476.479 f 18 1.72 145.06 9 f 18.7
10 6 - 2 4 4 : 6
6.749 * 0.10 4.099 f 0.10
0.0649 f 0.10 366.579 f 128.68 192.909 f 20.0
5

TABLA 11. Contenido de lípidos en membranas de eritrocitos de niños desnutridos y control.
Total de fosfolípidos (nmol/mg proteína)
Colesterol (nrnoYmg proteína)
Desnutridos Control 3.80 4.00 4.50 4.2 1 4.73 4.28 4.82 4.3 1 5.10 4.37 5.40 4.55 5.45 4.65 5.67 5.40 6.20 5.50 6.40 5.63
Desnutridos 0.59 0.65 0.69 0.70 0.71 0.72 0.73 0.74 0.77 0.83
Control 0.72 0.78 0.92 0.93 1 .o0 1 .O8 1.10 1.10 1.1 1 1.36
Media * 5.2 4.70 * * 0.71 D.E. 0.79 0.60 0.65
1.01 O. 19
u = 10, p < 0.002 ** U = 10, p < 0.007
6
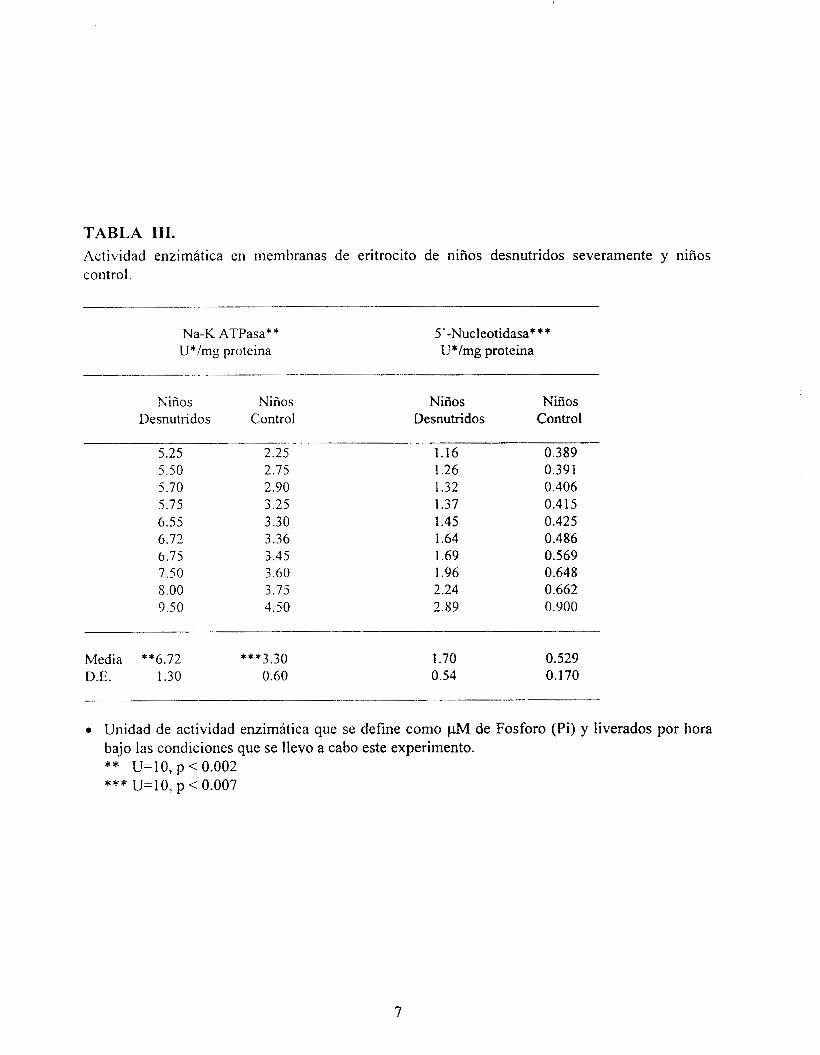
TABLA 111. Actividad enzimática en membranas de eritrocito de niños desnutridos severamente y niños control.
Na-K ATPasa** U*/mg proteina
5'-Nucleotidasa* * * U*/mg proteina
NiIios Niños Desnutridos Control
5.25 5.50 5.70 5.75 6.55 6.72 0.75 7.50 s.00 9.50
2.25 2.75 2.90 3.25 3.30 3.36 3.45 3.60 3.75 4.50
Niños Niños Desnutridos Control
1.16 0.389 1.26 0.39 1 1.32 0.406 1.37 0.4 15 1.45 0.425 1.64 0.486 1.69 0.569 1.96 0.648 2.24 0.662 2.89 0.900
Media **6.72 ***3.30 D.E. 1.30 0.60
1.70 0.529 0.54 O. 170
7
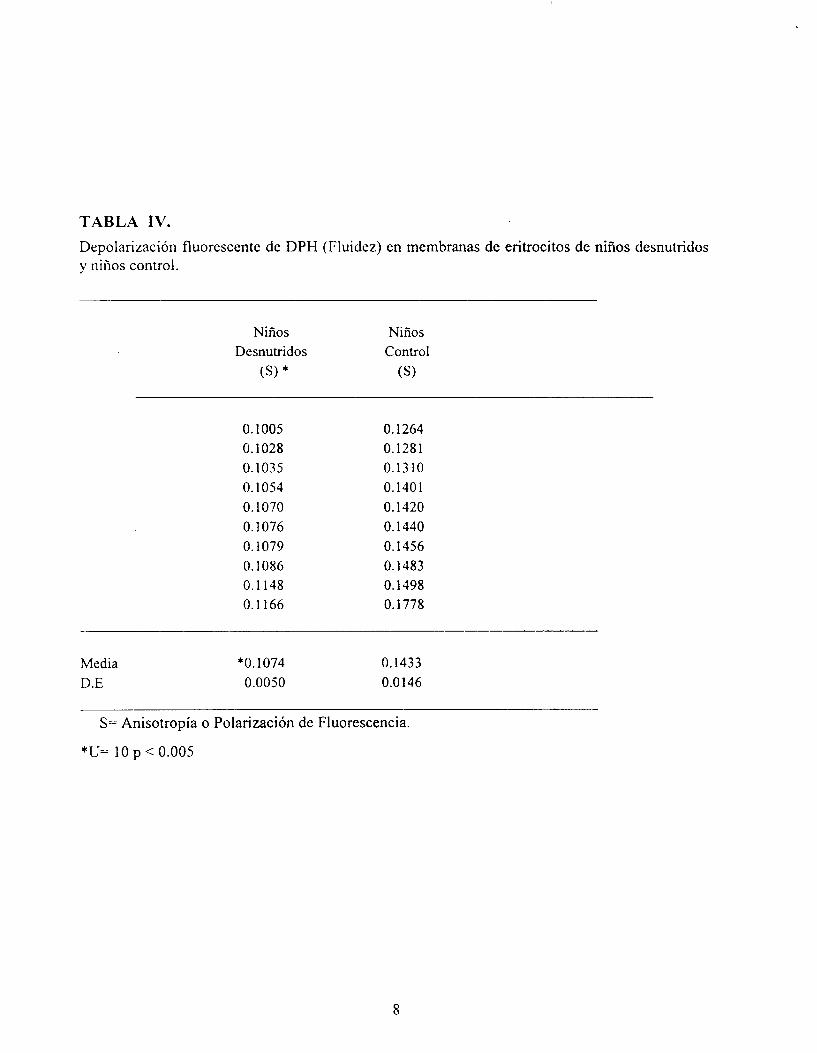
TABLA IV. Depolarización fluorescente de DPH (Fluidez) en membranas de eritrocitos de niños desnutridos y niños control.
Niños Niños Desnutridos Control
(S) * (S)
O. 1005 O. 1028 O. 1035 O. 1054 O. 1070 O. 1076 O. 1079 O. 1086 o. 1148 O. 1166
O. 1264 O. 128 1 0.1310 0.1401 O. 1420 O. 1440 O. 1456 O. 1483 O. 1498 O. 1778
Media D.E
*O. 1074 O. 1433 0.0050 0.0 146
S= Anisotropía o Polarización de Fluorescencia.
*u= 10 p < 0.005
8

TABLA V.
Composición individual de fosfolípidos en membranas de erítrocitos en niños desnutridos y niños control.
Desnutridos Control
(n = 5 ) (n = 5 )
% YO
Fosfatilserina *12.15 * * 9.50
Fosfatiletanolanlina 7.80 7.90
Fosfatidilcolina 9.00 7.60
Fosfatidilinositol 45.50 46.22
Otros 25.55 28.78
Total 105 .O5 98.02
u=5 p < 0.02
** U= 5 p < 0.05
9


















