Neurociencia y psiquiatría -...
34
Capítulo 1 Neurociencia y psiquiatría Steven E. Hyman, M.D. Josep T. Coyle, M.D. Los avances en las investigaciones del cerebro se han sucedido con un ritmo rápido y creciente du- rante los últimos veinte años y han alcanzado un punto en que la neurociencia puede considerarse justificadamente como el fundamento biomédico de la psiquiatría. El crecimiento logarítmico de nuestra comprensión de la organización y el fun- cionamiento del cerebro ha hecho posible empezar a analizar la conducta a nivel molecular. Estos avances han permitido la explotación fructífera de metodologías experimentales y diagnósticas cen- tradas en el cerebro como la resonancia nuclear magnética, la tomografía por emisión de positro- nes y el análisis topográfico de la actividad eléc- trica cerebral, para caracterizar irregularidades es- tructurales, metabólicas y electrofisiológicas en cerebros de pacientes psiquiátricos. Además, los avances paralelos en genética humana y en biolo- gía molecular nos permitirán definir las bases ge- néticas de las enfermedades hereditarias del com- portamiento, y fundamentalmente, determinar los mecanismos moleculares y celulares responsables de estos trastornos. Estos desarrollos están redu- ciendo progresivamente la separación cartesiana entre mente y cerebro, mejorando nuestra capaci- dad para correlacionar la experiencia mental con los procesos cerebrales. La investigación en el campo de la neurocien- cia ofrece importantes oportunidades a la psiquia- tría por su aplicación a la asistencia de los pacien- tes y, a largo plazo, porque aumenta la compren- sión de la experiencia y del comportamiento hu- manos. Por ello, es esencial para la psiquiatría aprovechar esta área de conocimiento de rápida evo- lución. Por consiguiente, la intención de este capí- tulo es revisar los aspectos moleculares y celulares de la investigación en neurociencia. No es posible, por limitación de espacio, tratar en profundidad la totalidad de los avances en la investigación del ce- rebro. Por tanto, hemos preferido abordar los temas principales y las estrategias más destacadas de la in- vestigación en neurociencia, que afectan directa- mente a la psiquiatría. Esperamos que este estudio servirá como fundamento para entender el cuerpo de conocimientos actuales y para evaluar crítica- mente los progresos futuros en esta rama de la cien- cia, así como su utilidad para los psiquiatras. @ ANATOMÍA FUNCIONAL DE LA NEURONA La neurona es una célula altamente especializada, tanto anatómica como bioquímicamente, para lle- var a cabo la función de procesamiento de la in- formación. En el sistema nervioso hay cientos de tipos de neuronas, cada una de las cuales se ocupa de funciones especializadas. A diferencia de mu- 3
-
Upload
trinhthien -
Category
Documents
-
view
218 -
download
0
Transcript of Neurociencia y psiquiatría -...

C a p í t u l o
1
Neurociencia y psiquiatríaSteven E. Hyman, M . D .
Josep T. Coyle, M . D .
Los avances en las investigaciones del cerebro sehan sucedido con un ritmo rápido y creciente du-rante los últimos veinte años y han alcanzado unpunto en que la neurociencia puede considerarsejustificadamente como el fundamento biomédicode la psiquiatría. El crecimiento logarítmico denuestra comprensión de la organización y el fun-cionamiento del cerebro ha hecho posible empezara analizar la conducta a nivel molecular. Estosavances han permitido la explotación fructífera demetodologías experimentales y diagnósticas cen-tradas en el cerebro como la resonancia nuclearmagnética, la tomografía por emisión de positro-nes y el análisis topográfico de la actividad eléc-trica cerebral, para caracterizar irregularidades es-tructurales, metabólicas y electrofisiológicas encerebros de pacientes psiquiátricos. Además, losavances paralelos en genética humana y en biolo-gía molecular nos permitirán definir las bases ge-néticas de las enfermedades hereditarias del com-portamiento, y fundamentalmente, determinar losmecanismos moleculares y celulares responsablesde estos trastornos. Estos desarrollos están redu-ciendo progresivamente la separación cartesianaentre mente y cerebro, mejorando nuestra capaci-dad para correlacionar la experiencia mental conlos procesos cerebrales.
La investigación en el campo de la neurocien-cia ofrece importantes oportunidades a la psiquia-
tría por su aplicación a la asistencia de los pacien-tes y, a largo plazo, porque aumenta la compren-sión de la experiencia y del comportamiento hu-manos. Por ello, es esencial para la psiquiatríaaprovechar esta área de conocimiento de rápida evo-lución. Por consiguiente, la intención de este capí-tulo es revisar los aspectos moleculares y celularesde la investigación en neurociencia. No es posible,por limitación de espacio, tratar en profundidad latotalidad de los avances en la investigación del ce-rebro. Por tanto, hemos preferido abordar los temasprincipales y las estrategias más destacadas de la in-vestigación en neurociencia, que afectan directa-mente a la psiquiatría. Esperamos que este estudioservirá como fundamento para entender el cuerpode conocimientos actuales y para evaluar crítica-mente los progresos futuros en esta rama de la cien-cia, así como su utilidad para los psiquiatras.
@ A NATOMÍA FUNC I O NAL DE
LA NEURO NA
La neurona es una célula altamente especializada,tanto anatómica como bioquímicamente, para lle-var a cabo la función de procesamiento de la in-formación. En el sistema nervioso hay cientos detipos de neuronas, cada una de las cuales se ocupade funciones especializadas. A diferencia de mu-
3

4 TRATADO DE PSIQUIATRÍA
Figura 1-1. Representación esquemática de una neurona. Un canal l i g a n d - g a t e d, posiblemente un receptor de gluta-mato, se muestra permitiendo la entrada de Na+ en el cuerpo de una neurona. Si el equilibrio de cargas positivas y ne-gativas es adecuado para despolarizar la neurona hasta llegar al umbral en la región del axón proximal o el engrosa-miento del axón, los canales de intercambio iónico de Na+ se abrirán, generando así un potencial de acción. El potencialde acción se propaga a lo largo del axón debido a la apertura secuencial de los canales Na+. Cuando el potencial de ac-ción invade la terminal presináptica, los canales de intercambio iónico de Ca+ + se abren, y la entrada de Ca+ + causa unaliberación de neurotransmisores (ver exposición en el texto). La repolarización de la neurona es el resultado de la aper-tura en rápida sucesión de los canales de intercambio iónico de K+ después de la entrada de Na+.
C a n a ll i g a n d - g a t e d
Na +
N ú c l e o
D e n d r i t a s
Axón hilloch
P e r i c a r i ó n
Potencial de acción
Na +
A x ó nVesículas de contenidon e u r o t r a n s m i s o r
Ca ++
Canales Na +de aperturasegún el voltage Na +
T e r m i n a lp r e s i n a p t i c o
chos tipos de células, como las que constituyen elhígado, la epidermis o el sistema hematopoyético,que son capaces de realizar la división celular du-rante toda la vida del individuo, el alto grado de es-pecialización de la neurona generalmente le impi-de llevar a cabo la división celular una vez estacélula es completamente madura. Esta incapacidadde la mayoría de las neuronas de experimentar unamitosis, influye de forma evidente en la irreversi-bilidad de las lesiones del sistema nervioso.
La neurona puede dividirse en cuatro compo-nentes distintos: el cuerpo celular o soma, las den-dritas, el axón y las sinapsis (ver Figura 1-1). En elsoma de la neurona tiene lugar la síntesis de prác-ticamente todas las proteínas y otros componentesestructurales. Situado en el soma se encuentra elnúcleo, que contiene el material genético en formade ácido desoxirribonucleico (ADN). La informa-ción para la síntesis de proteínas está codificadapor los genes que contiene el ADN: esta informa-ción genética se lee mediante un proceso llamadotranscripción en el cual el ADN sirve de plantillapara la síntesis del ácido ribonucleico (ARN). Lasprimeras transcripciones de ARN que resultan seprocesan y dan lugar al ARN mensajero (ARNm),que se exporta del núcleo al citoplasma del soma.Allí, el ARNm se traduce en proteínas gránulos lla-madas ribosomas. La rica concentración alrededordel núcleo de este sistema de síntesis de proteínaa través del ARNm explica la presencia de la
« s u stancia de Nissl», que se observa en el soma delas neuronas del tejido cerebral al emplear tincio-nes vitales. La síntesis de proteínas ocurre mayo-ritariamente en el soma, pero los ribosomas acti-vos se han detectado recientemente en dendritas,lo cual aumenta la posibilidad de que exista uncontrol local de síntesis de proteínas por estímuloneuronal.
El tamaño del soma neuronal es aproximada-mente proporcional a la extensión de las proyec-ciones de las dendritas y de los axones. Hay quedestacar que el soma tan sólo contiene un porcen-taje muy reducido de volumen neuronal; la parteprincipal del volumen celular se distribuye entreel axón y el árbol dendrítico. Por esta razón, los re-querimientos metabólicos y sintéticos del somaneuronal son inmensos, ya que el soma neuronalnutre al resto de la neurona. Los materiales sinte-tizados en el interior del soma son transportadosa lo largo de los axones y las dendritas por trans-porte axoplasmático para reemplazar los compo-nentes inactivados. Por el contrario, los productosde desecho de las proteínas metabólicas y estruc-turales de los axones y las dendritas, se transpor-tan en sentido inverso hacia el cuerpo celular paraser catabolizados.
El axón es una fina extensión tubular del cuer-po celular por el que circulan impulsos eléctricoshacia la terminación nerviosa. La neurona emitesolamente un axón, cuya longitud varía desde

NEUROCIENCIA Y PSIQUIATRÍA 5
menos de un milímetro en las interneuronas, hastamás de un metro en las neuronas motoras que iner-van las extremidades. El axón, cuando se acerca asu campo de inervación, puede ramificarse en di-versos grados, dependiendo del número de neuro-nas con las que establezca sinapsis. Algunas neu-ronas pueden tener uniones sinápticas muyespecíficas y restrictivas, mientras que los axonesde otras neuronas, como los de las neuronas dopa-minérgicas nigrostriadas, pueden ramificarse paraconectar con millones de neuronas en su zona ter-minal de inervación.
Las dendritas son extensiones tubulares múlti-ples del cuerpo celular neuronal que sirven de es-tructura primaria para la recepción de uniones si-nápticas procedentes de otras neuronas. (Algunasneuronas, como las células Purkinje en el cerebe-lo y algunos componentes del sistema reticular deltronco del cerebro, que poseen funciones integra-doras, poseen «á r b o l e s» dendríticos muy extensosque reciben input sináptico de miles de neuronas.La sinapsis es una estructura especializada en laque la terminación nerviosa de una neurona sepone en contacto con la parte receptora de la neu-rona adyacente; generalmente, la transmisión seconsigue por mensajeros químicos llamados neu-rotransmisores, pero en algunos casos puede tra-tarse de mensajeros eléctricos.
La sinapsis consiste en una evaginación de lasregiones terminales del axón denominada botónterminal, que está finamente sujeta a la membra-na dendrítica de la neurona adyacente por contac-tos especializados. La membrana dendrítica de lasinapsis es muy rica en receptores que respondenal neurotransmisor liberado por el botón terminal.El propio botón terminal contiene numerosas es-tructuras celulares que le permiten mantenerse re-lativamente independiente a nivel metabólico yfuncional del cuerpo celular neuronal. Así pues,contiene mitocondrias (unidades energéticas de lacelula que generan ATP (adenosín 5’ -trifosfato) apartir del metabolismo de la glucosa), enzimas im-plicados en la síntesis y en la degradación del neu-rotransmisor y vesículas de almacenamiento quemantienen importantes concentraciones de neu-rotransmisor en estado protegido. Cuando el teji-do cerebral se homogeniza cuidadosamente en unasolución isotónica de sacarosa, la sinapsis con elbotón terminal y la membrana postsináptica ad-yacente especializada se separan del resto para for-mar sinaptosomas. Estas estructuras se han utili-zado para estudiar aspectos bioquímicos de laneurotransmisión sináptica del cerebro.
La propiedad fundamental de todas las neuro-nas es la naturaleza excitable de su membrana, queles confiere la capacidad de generar y transmitiruna onda de despolarización electroquímica. Esta
propiedad deriva de la naturaleza especializada dela membrana neuronal, que mantiene un gradien-te de voltaje y posee canales de iones sensibles alvoltaje. Dos tipos de proteínas son mayoritaria-mente responsables de la regulación de iones y portanto del voltaje de la membrana neuronal. Estosson las bombas iónicas y los canales de sodio vol-taje. Basándose en la clonación molecular que tie-nen codificada algunas de estas bombas iónicas ycanales de sodio voltaje, parece ser que cada unorepresenta una familia de genes que deriva de sugen ancestral común. Aún más importante, la clo-nación molecular ha iniciado una era de estructu-ra y análisis detallados de la actividad que muyprobablemente conducirá a la creación de fárma-cos neuroactivos más eficaces.
Las bombas críticas para establecer el gradien-te fisiológico de iones que se encuentran a lo largode la membrana neuronal son una bomba de ATP-dependiente, la ATPasa que extrae dos iones deN a+ por cada ion de K+ que entra; y bombas que ex-traen Ca+ + de la neurona. Durante los descansos,hay altas concentraciones de Na+ y Cl– fuera de laneurona y una concentración relativamente alta deK+ dentro de la neurona. La fuente principal decarga negativa dentro de la neurona proviene deaminoácidos de carga negativa. La membrana sepolariza en su totalidad con una diferencia de vol-taje a lo largo de la membrana de aproximadamente–70mV, en comparación con el exterior de la neu-rona. Esto se llama el potencial de descanso de lam e m b r a n a.
Cuando la membrana neuronal se despolarizahasta llegar a aproximadamente –35mV, se produ-ce un «potencial de acción» que representa a la ac-tividad de «d i s p a r o» de la neurona, y es el meca-nismo fundamental de la estimulación neuronal.Concretamente, a medida que el interior de la neu-rona se vuelve positivo, los canales especializadosde sodio voltaje se abren, lo que permite que másiones positivos entren en la neurona (Figura 1-1).El potencial de acción representa la extensión dela despolarización por la apertura vectorial de loscanales de sodio. Debido a que cada canal de Na+
que se abre en sucesión proporciona la carga posi-tiva para que el siguiente segmento del axón al-cance el umbral para abrir sus canales de Na+, elpotencial de acción es autorregenerador y una vezha empezado, se propaga a lo largo del axón sin pro-blemas. Al llegar al terminal presináptico, el po-tencial de acción causa la apertura de los canalesde Ca+ + voltaje que se encuentran allí (denomina-dos canales de Ca+ + tipo N, en contraposición conlos canales Ca+ + tipo L, bloqueados por los blo-queantes de canales de Ca+ + tipo verapamino uti-lizados en la clínica). El Ca+ + entra e inicia unaserie de procesos bioquímicos complejos pero

6 TRATADO DE PSIQUIATRÍA
rápidos que causan una fusión de las vesículas con-tenedoras de neurotransmisores con la membranapresináptica y por tanto liberan su contenido en lasinapsis, permitiendo una transmisión sináptica.Debido a que la entrada de la carga positiva despo-lariza la membrana, acercando así la neurona al um-bral para «d i s p a r a r» un potencial de acción, los re-ceptores de neurotransmisores que permiten laentrada de cationes como Na+ o Ca+ + son excitado-res y los que causan la entrada de aniones como Cl–,o la salida de cationes, como K+, son inhibidores.
Las dendritas suman continuamente todos lospotenciales de las aferencias excitadoras e inhibi-doras, para determinar si la neurona generará o noun potencial de acción. La organización espacialde la inervación de la neurona no es casual sinoque está altamente organizada. Las aferencias ex-citadoras están generalmente concentradas en elextremo distal de las dendritas, mientras que lasaferencias inhibidoras están localizadas principal-mente en el extremo proximal de las dendritas yalrededor del soma. Esta distribución espacial sig-nifica que las aferencias inhibidoras tienen unpapel predominante al determinar si una neuronagenerará o no, un potencial de acción. Debido a queel potencial de acción es autorregenerador, la de-cisión de «d i s p a r a r» un potecial de acción es unproceso de todo o nada, y cuando el equilibrio sedecanta hacia la despolarización adecuada en la re-gión del axón proximal, donde la densidad de loscanales de sodio es alta, se genera un potencial deacción (Figura 1-1).
@ N E U ROT R A N S M I S O R E S
Aunque no suele apreciarse, cabe recordar que lapsiquiatría, especialidad médica que se ocupa demuchos aspectos de la comunicación, fomentó lamayor parte de las primeras investigaciones sobrelos mecanismos de la comunicación química entrelas neuronas. Debemos señalar que la existenciade neurotransmisores cerebrales fue propuesta porprimera vez con la hipótesis de que la esquizofre-nia sería el resultado de una anomalía del meta-bolismo del neurotransmisor adrenalina, por lo quese generaría un metabolito psicomimético, el adre-nocromo. Aunque esta hipótesis finalmente nopudo probarse, al principio de los años sesenta pro-movió numerosos estudios para caracterizar la dis-posición metabólica de las catecolaminas cerebra-les. Más recientemente, esta estrategia de estudiode la comunicación neuronal en el cerebro ha con-ducido a la identificación de un conjunto rápida-mente creciente de sustancias que actúan comoneurotransmisores. Así pues, a mediados de los
TABLA 1-1. CRITERIOS QUE DEBE CUMPLIR UN NEURO-T R A N S M I S O R
1 . La neurona contiene la sustancia.*2 . La neurona sintetiza la sustancia.*3 . La neurona libera la sustancia en la despolarización.*4 . La sustancia es fisiológicamente activa en las
n e u r o n a s .*5 . La respuesta fisiológica postsináptica a la sustancia es
idéntica a la del neurotransmisor liberado por la neu-rona.
* Todas las sustancias enumeradas en las tablas 1-2 y 1-3 cumplencon este criterio.
años sesenta se creía que sólo un reducido númerode sustancias satisfacían el criterio de neurotrans-misores cerebrales «p u t a t i v o s» . El término putati-vo se refiere al hecho de que es muy difícil satisfa-cer todos los criterios para que una sustancia seconsidere sin lugar a dudas un neurotransmisor ce-rebral (ver Tabla 1-1). Durante la última década, elnúmero ha crecido aproximadamente diez veces(ver una lista parcial en las Tablas 1-2 y 1-3). Ade-más, actualmente disponemos de un dato convin-cente y es que la mayoría de neuronas liberan másde un neurotransmisor, a menudo una pequeña mo-lécula neurotransmisora y uno o más péptidos.
La siguiente exposición se centra en ejemplosrepresentativos de los dos tipos principales de neu-rotransmisores en el cerebro: a) los neurotransmi-s o r e s «c l á s i c o s» como la adrenalina, que se sinte-tizan localmente en las terminaciones nerviosas,y b) los neurotransmisores neuropéptidos como labeta-endorfina, que se sintetizan en el soma neu-ronal. Para un estudio detallado, ver Nestler (1993)y Cooper et al. (1991).
C a tecolaminas
El sistema de neurotransmisores mejor estudiadodesde el punto de vista de la síntesis, el almacena-miento, la liberación y el metabolismo, es el sis-tema catecolaminérgico. Los principios estableci-dos para la neurotransmisión catecolaminérgicaperiférica y central tienen una aplicabilidad gene-ral al resto de los sistemas de neurotransmisoresclásicos. Los neurotransmisores catecolaminérgi-cos comprenden la dopamina, la noradrenalina y laadrenalina. Aunque cada uno de ellos actúa comoneurotransmisor diferenciado en los sistemas neu-ronales específicos del cerebro, todos ellos formanparte de una misma ruta biosintética (Figura 1-2).Las enzimas responsables de la síntesis de cateco-laminas se sintetizan en el soma de la neurona yse transportan a lo largo del axón hasta los termi-nales presinápticos. Las neuronas que usan dopa-mina como neurotransmisor poseen las primeras

NEUROCIENCIA Y PSIQUIATRÍA 7
TABLA 1-2. N E U R O T R A N S M I S O R E S «C L Á S I C O S »
A c e t i l c o l i n a Ácido aspárticoHistamina Ácido gamma-aminobutírico Serotonina Ácido glutámicoD o p a m i n a G l i c i n aN o r a d r e n a l i n a H o m o c i s t e í n aA d r e n a l i n a T a u r i n a
dos enzimas de la ruta, tirosina hidroxilasa y dopadecarboxilasa. Las neuronas que liberan noradrena-lina presentan una tercera enzima, dopamina-beta-hidroxilasa, y las neuronas que producen adrenali-na presentan una cuarta enzima, feniletanolaminaN-metil transferasa (PNMT). Debido a que la tiro-sina hidroxilasa es una enzima de flujo limitado yaltamente controlado, los mecanismos reguladoresgenerales que determinan la disponibilidad del neu-rotransmisor son comunes para todos los neuro-transmisores de este grupo.
Las rutas sintéticas de los neurotransmisoresclásicos, generalmente, aunque no de manera in-variable, implican la conversión de un precursorinactivo en cuanto a capacidad de transmisión deinformación en un neurotransmisor «c a r g a d o» d einformación. En el caso de las catecolaminas, actúacomo precursor el aminoácido no esencial L-tiro-sina. La tirosina hidroxilasa es el paso limitante dela tasa de síntesis y virtualmente se satura por losniveles cerebrales de este aminoácido. Por tanto,el incremento de los niveles de este aminoácido enel cerebro no afectará significativamente la bio-síntesis de las catecolaminas. Además, para pre-venir un círculo vicioso de síntesis y de degrada-ción de catecolaminas, la tirosina hidroxilasa estásujeta a un mecanismo de retroalimentación ne-gativo regido por el producto final. De este modo,cuando la concentración de catecolaminas en laterminación nerviosa sobrepasa su capacidad de al-macenamiento, el exceso de catecolaminas inhibela actividad de la tirosina hidroxilasa, evitando que
Figura 1-2. Vía de la biosíntesis de las catecolaminas. Ob-sérvese que la tirosina hidroxilasa se activa por la acciónde las proteín-quinasas y que la síntesis de feniletanola-mina-N-metil transferasa depende de los corticosteroides.
prosiga la síntesis de catecolaminas. Esta retroali-mentación asegura que siempre exista una concen-tración constante de catecolaminas frente a la de-manda de liberación de las mismas de la sinapsis.Por tanto, cuando las neuronas catecolaminérgicasno transmiten, la síntesis de catecolaminas se de-tiene. Por otra parte, cuando se liberan catecolami-nas y se agotan las reservas, esta retroalimentacióninhibidora desaparece y la síntesis se reanuda.
Se hace patente, sin embargo, que actúan tam-bién otros mecanismos durante los períodos demayor demanda en la liberación de catecolaminas.L o s «d i s p a r o s» repetidos de la neurona catecolami-nérgica producen la activación de sistemas de se-gundos mensajeros y por tanto de proteínas quina-sas (ver más adelante). La fosforilación de la enzimatirosina hidroxilasa por las proteínas quinasas re-duce su sensibilidad para la retroalimentación inhi-bidora e incrementa su afinidad por el cofactor crí-tico pterina (Nose et al., 1985). Los períodos deincremento prolongado de la actividad neuronal ca-tecolaminérgica provocan la puesta en marcha de unsegundo mecanismo, la síntesis de moléculas enzi-máticas adicionales en la vía de biosíntesis de las ca-tecolaminas. Este segundo proceso está regulado anivel del soma de las neuronas catecolaminérgicas,donde el ARNm adicional codificado por la tirosinahidroxilasa es copiado a partir del ADN del núcleo.Así, se hace patente que la síntesis de catecolami-nas está sometida a una regulación dinámica que, asu vez, está firmemente coordinada por la actividadeléctrica de la neurona catecolaminérgica. Despuésde que en el citosol del nervio terminal se haya pro-ducido la síntesis enzimática de las catecolaminas,estas sustancias se concentran en vesículas, peque-ños sacos membranosos localizados en la termina-ción nerviosa. El almacenamiento vesicular es unproceso activo que requiere energía y que resulta in-hibido irreversiblemente por el fármaco antihiper-tensivo reserpina. Las vesículas de almacenamien-to cumplen dos funciones. Primero, protegen a lascatecolaminas de ser degradadas por el enzima mo-noamino-oxidasa (MAO). Segundo, las vesículas in-fluyen en la descarga cuántica de catecolaminas porliberación mediante exocitosis, cuando un potencialde acción llega al nervio terminal.
Además de la enzima intraneuronal MAO, unasegunda enzima que inactiva catecolaminas se en-cuentra en la superficie externa de la membrananeuronal, así como en la superficie externa de mu-chos otros tipos de neuronas. Esta enzima, catecol-O-metiltransferasa (COMPT), cataliza la inactiva-ción de las catecolaminas por metilación de uno delos grupos co/anillo hidroxil.
Sin embargo, la degradación enzimática no esel mecanismo más significativo por el cual se ter-

8 TRATADO DE PSIQUIATRÍA
TABLA 1-3. NEUROPÉPTIDOS NEUROTRANSMISORES P U T A T I V O S
Hormona adrenocorticotropa (ACTH) Angiotensina II *Beta-endorfina Bombesina Bradiquinina Carnosina Colecistoquinina *Dinorfina Factor liberador de corticotropina Galinina Gastrina Glucagón Hormona liberadora de tirotropina (TRH) Insulina *Leu-encefalina *Met-encefalina Neuropéptido γNeurotensina Pancreostatina Péptido natriurético auricular Péptido relacionado con el gen de la calcitonina (CGRP) Péptido vasoactivo intestinal (VIP) Somatostatina Sustancia P Vasopresina
* Miembros de la familia de las endorfinas.
mina la acción de catecolaminas en la sinapsis. Elmecanismo más crítico es una recaptación activade las catecolaminas por la propia terminal ner-viosa que las ha liberado. La recaptación se lleva acabo gracias a una proteína transportadora especí-fica que cambia las catecolaminas con un procesodependiente de energía impulsado por el gradien-te de sodio a través de la membrana neuronal (Pa-cholczyk et al., 1991). Los transportadores dopa-minérgicos y la noradrenalina son miembros deuna misma familia genética de proteínas que tam-bién incluye los transportadores serotoninérgicosy de GABA (ácido gama-aminobutírico) (Revisadopor Giros y Caron, 1993).
Los procesos implicados en la síntesis, alma-cenamiento, liberación e inactivación de los neu-rotransmisores clásicos se resumen en la Figura 1-3. Estos procesos interrelacionados aseguran unadisponibilidad estable del neurotransmisor en laterminal nerviosa, sea cual sea la demanda y lainactivación del neurotransmisor liberado, a fin deque no pueda difundirse fuera de la hendidura si-náptica en cantidades significativas.
Ha sido demostrada la operatividad de los mis-mos principios en un grado importante, aunque va-riable, para los otros neurotransmisores clásicos,incluyendo la serotonina, la acetilcolina y la his-tamina. Los neurotransmisores de tipo aminoáci-do, sin embargo, representan una excepción im-portante del principio de acuerdo con el que los
neurotransmisores son sintetizados a partir de pre-cursores neurofisiológicamente inactivos. El ami-noácido glutamato parece ser el neurotransmisorexcitador predominante en el cerebro; y los ami-noácidos como el GABA (ácido gamma-aminobu-tírico), la glicina y posiblemente la taurina parecenser los neurotransmisores inhibidores más desta-cables. Estas moléculas están presentes en el plas-ma y son precursores importantes en la síntesis deproteínas. Estas características parecerían ser con-tradictorias con las de un neurotransmisor, quedebe poseer una acción altamente localizada sobrelos receptores apropiados. Sin embargo, se gasta unacantidad considerable de energía para mantener pro-cesos de transporte selectivo y la acción de enzimascatabólicos, con el objeto de mantener concentra-ciones de estos neurotransmisores aminoácidos ex-tremadamente bajas en el espacio extracelular delcerebro. La importancia de esta protección se ilus-tra por el hecho de que la concentración intracelu-lar de ácido glutamático en ciertas regiones del ce-rebro es mayor de 10 mM, mientras que laconcentración en el líquido cefalorraquídeo (CSF)es aproximadamente de 0,1 µM, lo que representaun gradiente 100.000 veces mayor.
Ne u r o p é ptidos
El hecho de que las proteínas pequeñas, como porejemplo los péptidos, sean utilizadas en el cuerpocomo señales, es bien conocido por su función comohormonas en la hipófisis y en varias glándulas en-docrinas. El posible papel de los péptidos comoneurotransmisores procede del descubrimiento deque los factores de liberación que controlan la se-creción de diversas hormonas hipofisiarias fueran,de hecho, péptidos sintetizados por neuronas enel núcleo arqueado del hipotálamo. Sin embargo,el descubrimiento decisivo que provocó un am-
Figura 1-3. Procesos que intervienen en la síntesis, la ac-ción sináptica y la inactivación de los neurotransmisoresc l á s i c o s .

NEUROCIENCIA Y PSIQUIATRÍA 9
plio interés en relación con los péptidos como po-sibles neurotransmisores fue el hallazgo de las en-dorfinas, péptidos opiáceos endógenos, que se en-cuentran ampliamente distribuidos en el sistemanervioso central (SNC) (Hughes et al., 1975). Desdeel descubrimiento de las endorfinas hace una dé-cada, el número de péptidos que se cree que actú-an como neurotransmisores en el cerebro se ha in-crementado considerablemente, y en la actualidades superior a cuarenta.
La investigación de los neuropéptidos (para unarevisión, ver Hokfelt, 1991) en la última década harevelado varios principios generales. A diferenciade los pequeños neurotransmisores molecularesque se sintetizan por procesos enzimáticos locali-zados dentro de la terminal nerviosa, los neuro-péptidos se sintetizan dentro del cuerpo neuronal(Figura 1-4). Esto refleja el hecho de que la síntesisde los neuropéptidos, que son pequeñas proteínas,se dirige por ARNm que ha sido transcrito delADN nuclear. Los niveles de neuropéptidos en laterminal nerviosa dependen completamente de lasíntesis, el procesamiento y el transporte de lospéptidos del soma neuronal. Asimismo, parece quelas neuronas peptidérgicas son menos propensas aresponder rápidamente a aumentos prolongados enla liberación a causa del retraso en la síntesis ytransporte de neuropéptidos suplementarios quedeben llegar desde el soma neuronal.
Figura 1-4. Secuencia de la síntesis de neuropéptidos.En el núcleo, el gen del neuropéptido precursor se trans-cribe en ARNm. EL ARNm se transporta desde el núcleoal citoplasma, donde se fija a los ribosomas. Posterior-mente, el ARN se transcribe merced a la síntesis protei-ca en los ribosomas del retículo endoplasmático rugoso.En el aparato de Golgi, el péptido precursor se modificaenzimáticamente para convertirse en neuropéptido, quese almacena en vesículas para su transporte axoplasmá-tico hasta las terminaciones nerviosas.
Uno de los hallazgos más sorprendentes en rela-ción a la síntesis de neuropéptidos es que un sologen a menudo da lugar a múltiples péptidos activosy, además, que la gama de péptidos producidos porun solo gen puede variar en cuanto al tipo de neu-ronas. La diversidad empieza después de la trans-cripción del gen que codifica el precursor del pépti-do. En neuronas eucariotas la mayoría de genestienen sus secuencias codificadas de proteínas (lla-mados *exones) interrumpidas por secuencias no-co-dificadas (llamadas *itrones). Cuando se transcribeun gen, la transcripción principal es co-lineal con elADN y por tanto contiene *exones e *itrones. Antesde que el ARN parta del núcleo para ser traducido,los *itrones se expulsan y los *exones se parten paraformar el ARNm. Se ha visto que la primera trans-cripción de ciertos genes neuropéptidos se parte demaneras distintas en diferentes tipos de neuronas.Al incluir o excluir exones concretos en el ARNmcitoplásmico, se pueden producir péptidos de ARNmcon diferentes codificaciones. Por ejemplo, los pép-tidos calcitronin y CGRP son productos del mismogen derivado de esta división alternativa.
La codificación de neuropéptidos maduros deARNm se traduce, en el retículo endoplasmáticorugoso para producir precursores grandes llamadosp o l i p r o t e í n a s. Estos precursores (es decir, pre-pro-opiomelanocortina o pre-proencefalina) contienenuna secuencia, casi siempre en el terminus deamino de la proteína, que dirige la proteína por lavía secretoria de la neurona. Esta secuencia líder opre-secuencia se parte rápidamente por una endo-peptidasa y deja el resto del precursor (pre-proopia-melano cortina o pre-proencefalina). Este precursorlleva a cabo más partición proteolítica y por lo tantoquímicamente modificada (es decir, glicosilación,amidación, acetilación o fosforilación) dentro delretículo endoplasmático rugoso y el aparato deGolgi para ceder numerosos péptidos biológicos ac-tivos para liberar.
Así como la división alternativa de ARNm per-mite la regeneración de múltiples moléculas de es-timulación a partir de un solo gen, el procesamien-to diferencial de los precursores de péptidos permitela regeneración de múltiples moléculas de estimu-lación de un mismo péptido precursor. Dentro de lospéptidos precursores, hay pares de residuos de ami-noácidos básicos (lisina o arginina) reconocidos porlos enzimas procesadores como zonas de división.La familia de enzimas responsable de procesar den-tro de las neuronas mamíferas ha sido descubiertarecientemente y por tanto no están completamentecaracterizadas. Sin embargo, parece ser que ciertasenzimas tienen mayor afinidad por algunos pares deresiduos dibásicos que por otros. Por tanto, depen-diendo de los enzimas que se expresan en un tipo

1 0 TRATADO DE PSIQUIATRÍA
neuronal particular o endocrino, los precursoresgrandes pueden ser divididos en diferentes pépti-dos activos con diferentes papeles fisiológicos.
Entre los neurotransmisores péptidos, los pép-tidos opiáceos endógenos son los más estudiadosy tienen una clara relevancia para la psiquiatría de-bido a su papel en la respuesta al estrés, en las con-ductas motivadas y en la analgesia. Hughes et al.(1975) identificaron los primeros opiáceos endóge-nos met y leu encefalina. Desde entonces, se hanaislado y caracterizado aproximadamente 20 pép-tidos opiáceos activos de cerebros de mamíferos yde glándulas pituitarias y adrenales. Todos estosopiáceos endógenos contienen los mismos cuatroaminoácidos cuyo grupo amino terminal es Tir-Gli-Gli-Fe, seguidos por Met o Leu (Met-encefali-na, Leu-encefalina). Todos los péptidos opiáceosderivan de uno de tres grandes precursores poli-proteínicos, cada uno codificado en un gen separa-do. Los precursores son la pro-encefalina (que tienecodificadas seis copias de la secuencia de Met-en-cefalina y una de las secuencias de Leu-encefali-na), la prodinorfina (que es el precursor de la *d i-norfina opiácea endógena y péptidos relacionados)y la pro-opiomelanocortina. La pro-opiomelano-cortina (POMC) es particularmente interesanteporque contiene las secuencias de péptidos activoscon funciones biológicas aparentemente diferen-tes, el péptido opiáceo beta-endorfina y el no-opiá-ceo, la hormona adrenocorticotropa (ACTH), y por-que se procesa para producir diferentes tejidos(Figura 1-5).
En el lóbulo anterior de la pituitaria, sólo unsubgrupo de las zonas de división dibásicas dentrode la pro-opiomelanocortina es reconocido por laenzima que está presente. En este tejido, el precur-sor produce un péptido terminal-N de función bio-
Figura 1-5. Procesamiento de la pro-opiomelanocorti-na (POMC). La proteína precursora POMC, que contiene165 aminoácidos, se divide enzimáticamente para con-vertirse en los péptidos fisiológicamente activos que seindican. En función de la localización (hipófisis anterior,hipotálamo, terminales nerviosas del mesencéfalo), al-gunos de estos neuropéptidos se expresan y otros no. (VerWatson et al., 1985).
lógica desconocida junto con ACTH y la hormonabeta-lipotropina. La beta-lipotropina se somete aotra división para procesar el péptido opiáceo beta-endorfina. Por tanto, cuando la ACTH es liberadapor la glándula pituitaria, las beta-endorfinas tam-bién son liberadas. Muchos mamíferos (pero no loshumanos) tienen un lóbulo pituitario intermedioque contiene una enzima procesadora adicional.En el lóbulo intermedio, la ACTH se somete a otradivisión para procesar un péptido llamado péptidodel lóbulo intermedio tipo corticotropina (CLIP) yla hormona melanocito estimulante (MSH). Esteproceso, sin embargo, no ocurre en los humanos.
Co-localización
Antes se creía que cada neurona usa un neuro-transmisor, y sólo uno. Sin embargo, en los últimosaños se ha asistido a un rápido crecimiento de lalista de excepciones a este principio, y se ha de-mostrado que las neuronas contienen y liberan másde un neurotransmisor. Generalmente, la demos-tración de la co-localización de dos o más neuro-transmisores en la neurona, comporta que uno deellos sea un neurotransmisor clásico y el otro unneuropéptido. Pero además en la actualidad existenejemplos en los que dos neurotransmisores clási-cos, como la serotonina y el GABA, coexisten en lamisma neurona. Es importante entender que losmismos neurotransmisores no están co-localizados.Por ejemplo, se ha demostrado que la encefalina,péptido opiáceo, está co-localizado en determina-das neuronas noradrenérgicas del sistema nerviososimpático y en algunas neuronas serotoninérgicasdel cerebro. Se ha demostrado también que en el ce-rebro la *colescistocinina está co-localizada en lasneuronas dopaminérgicas que inervan el sistemacorticolímbico, pero no en las neuronas dopami-nérgicas que inervan el estriado. Con el gran nú-mero de neurotransmisores putativos en el cerebro,el número de posibles combinaciones es inmensoy sugiere un grado mucho mayor de complejidad enla neurotransmisión sináptica del que había sidoconsiderado con anterioridad. Los mecanismos cla-ramente diferenciados implicados en la síntesis delos neuropéptidos comparados con los de los neu-rotransmisores clásicos sugieren que los neuro-transmisores co-localizados pueden desempeñarfunciones algo diferentes, pero complementarias.Algunos estudios sugieren que los neuropéptidos seliberan sólo durante los períodos de marcada acti-vidad neuronal, mientras que los neurotransmiso-res clásicos se liberan en proporción al flujo de losimpulsos. Sin embargo, actualmente la compren-sión de los mecanismos implicados en la comuni-cación compartida continúa siendo rudimentaria.

NEUROCIENCIA Y PSIQUIATRÍA 1 1
@ R E C E P TORES
La identificación, caracterización y, más reciente-mente, la cenificación molecular de los receptoresde los neurotransmisores, constituyen un granavance en la neurociencia, produciendo un impac-to considerable en la comprensión de la elabora-ción de información en el cerebro y los lugares deacción de las sustancias neuroactivas, incluyendolos fármacos psicotrópicos. Aunque habitualmen-te los neurotransmisores son descritos como exci-tadores o inhibidores, como si esta acción fuese in-herente a su estructura molecular, la naturaleza dela respuesta neuronal a un neurotransmisor de-pende en último término de la presencia de un re-ceptor unido a un transductor. Dependiendo deltipo de receptor-transductor que posea una neuro-na dada, un neurotransmisor puede ejercer efectosinhibidores, excitadores o moduladores.
Los receptores neuronales son proteínas situa-das en la superficie externa de la membrana neu-ronal. Estas proteínas tienen regiones de fijacióndel ligando accesibles a los mensajeros extra-celu-lares y a otras regiones involucradas en la trans-ducción de la interacción de unión a un efecto in-tercelular. La unión reversible del neurotransmisorcon la zona de reconocimiento del receptor causaun cambio de conformación que desencadena la po-sibilidad de que la membrana sea atravesada. Estehecho puede implicar la apertura de los canales ió-nicos que atraviesan la membrana neuronal o la ac-tivación de una proteína-G estimuladora de trans-ducción (ver más adelante) que se asocia a lasuperficie interna de la membrana. Los receptoresque actúan de vía de entrada en un canal iónico in-trínsico se llaman canales ligand-gated; los recep-tores que actúan vía las proteínas-G se llaman re-ceptores proteínas-G linked. Como en el caso deotros tipos importantes de moléculas mencionadasanteriormente en este capítulo, como los canalesiónicos sensibles al voltaje y los transportadores deneurotransmisores, los canales l i g a n d - g a t e d, los re-ceptores pro-lin-G y las proteínas-G forman gran-des familias independientes de moléculas con es-tructuras homólogas. Se cree que cada una de estasfamilias de genes empezó con un único predecesorprimitivo (es decir, un canal l i g a n d - g a t e d a n c e s t r a l )que, después de duplicaciones y mutaciones degenes dio lugar a un gran número de genes y portanto a proteínas con funciones específicas pero re-lacionadas, permitiendo una complejidad crecien-te de estimulación neuronal durante la evolución.
La fijación del ligando
Puesto que los receptores se unen con el neuro-transmisor con el que interaccionan, de forma
ávida, específica, reversible y saturable (por ejem-plo, el número de zonas receptoras está limitado),los neurocientíficos han aprovechado estas carac-terísticas al utilizar ligandos radioactivos para mar-car los receptores de una manera específica. Si laavidez de la interacción específica entre el ligandomarcado radioactivamente es suficientemente alta,dicho ligando puede quedar «a t r a p a d o» en el recep-tor de forma lo bastante intensa como para que elcomplejo radioactivo resultante pueda ser aislado.Esta estrategia ha facilitado enormemente los es-tudios orientados a examinar las características deestas interacciones entre receptor y neurotransmi-sor y su localización en el sistema nervioso. Porejemplo, la afinidad relativa entre los fármacos osustancias parecidas a los neurotransmisores y alreceptor, puede determinarse midiendo su poten-cia para la unión de un ligando radioactivo con sureceptor. Con este método, Snyder y sus colabora-dores demostraron una correlación precisa entre lasafinidades de los fármacos antipsicóticos con los re-ceptores dopamínicos tipo D2 y la potencia clínicade estos fármacos en el tratamiento de trastornospsicóticos (Figura 1-6). Los métodos de fijación delligando combinados con modelos tridimensionalesde receptores clonados deberían facilitar una com-prensión de la estructura precisa de las moléculasrequeridas para un reconocimiento óptimo en lazona del receptor y, por tanto, a la larga, un diseñofarmacológico racional.
Como ciertos radioligandos se unen estrecha-mente a sus receptores específicos, ha sido posiblevisualizar la distribución de los receptores en el ce-rebro mediante técnicas autorradiográficas. Coneste método, cortes finos de tejido cerebral son in-cubados con el ligando radioactivo en un tampónfisiológico. A continuación, el corte se lava con untampón libre de radioligando para eliminar la partede éste unida de forma no específica y laxa, que esfácil y rápidamente reversible. En el tejido quedael radioligando específicamente asociado con el re-ceptor. Esta asociación puede entonces ser revela-da por aposición del corte del tejido con una pelí-cula sensible a los rayos X, con lo que se producela precipitación de los gránulos de plata de la emul-sión de la fotografía en aquellas zonas donde estánsituados los ligandos radioactivos. La Figura 1-7presenta un autorradiografía de la distribución delos receptores muscarínicos evaluados por la fija-ción del antaponista muscarínico 3H específico ymuy potente denominado quinuclidinil benzilatoen una sección parasagital del cerebro de mono.
Con el desarrollo de los métodos de imagen asis-tida por ordenador capaces de localizar la fuente deemisiones radioactivas de positrones en un espaciotridimensional, ha sido posible emplear las técni-cas de fijación de ligando y receptor in vivo, con

1 2 TRATADO DE PSIQUIATRÍA
Figura 1-6. Correlación entre la potencia clínica de losneurolépticos y su afinidad por los receptores dopami-nérgicos D-2. Las ordenadas muestran la dosis media dia-ria de neurolépticos administrados para tratar la esquizo-frenia y las abcisas indican la afinidad de los neurolépticospor el receptor dopaminérgico D-2 (véase el texto). (Porcortesía de S. Snyder).
objeto de determinar la distribución de los recep-tores de los neurotransmisores en el cerebro delhombre. Por ejemplo, usando spiperona marcadacon carbono 11 y emisora de positrones, que es unneuroléptico con una alta afinidad por los recepto-res de dopamina-2, Wong y sus colaboradores (1986)han visualizado la distribución de estos receptorespara la dopamina en el cerebro humano (Figura 1-8). A medida que se vayan desarrollando ligandosemisores de positrones suficientemente ávidos paraotros receptores, podrán ser utilizados también parala visualización de las correspondientes zonas ce-rebrales con las técnicas tomográficas por emisiónde positrones. Estas tecnologías ofrecen la posibi-lidad de estudiar cambios en los receptores in vivoen estados patológicos y en respuesta a la ingestade fármacos.
Ne u r o fisiología
Los transductores, con los que interacciona el re-ceptor de un neurotransmisor, determinan en últi-mo término la respuesta fisiológica resultante de launión del neurotransmisor al receptor en una neu-rona dada. La relativa facilidad con que los estudiosde fijación de ligando son capaces de caracterizarlas interacciones entre los receptores y los neuro-transmisores, no elimina la tarea más ardua de de-finir los transductores acoplados con receptores es-
pecíficos. Esta dificultad deriva del hecho de quelas interacciones entre el receptor y el transductora menudo no pueden ser determinadas cuando seestudian al mismo tiempo amplias poblaciones dereceptores, como sucede en los estudios de fijaciónde ligandos, ya que pueden estar asociados con di-ferentes transductores en las distintas neuronas. Enconsecuencia, técnicamente es más complicadomedir la respuesta fisiológica provocada por la ac-tivación de receptores neuronales específicos.
Se han desarrollado varios métodos para medirlas consecuencias electrofisiológicas de la aplica-ción local de neurotransmisores en neuronas con-cretas. Con micropipetas múltiples formadas porun electrodo de registro y un conjunto de pipetasadosadas entre sí, colocadas en una neurona en elcerebro, el neurotransmisor o el fármaco es envia-
Figura 1-7. Corte coronal de un cerebro de mono quemuestra los receptores muscarínicos. El corte se incubócon (3N)-quinuclidinil benzilato, un potente antagonis-ta de los receptores muscarínicos. A continuación elcorte se colocó sobre una película de rayos X para reve-lar el autorradiograma. Esta imagen negativa pone de ma-nifiesto los receptores muscarínicos en forma de puntosde color blanco: las áreas de alta densidad de receptores,como el núcleo caudado (C), el putamen (P) y el córtex(Cx) aparecen de color blanco; las áreas de baja densidadde receptores como el cuerpo calloso (CC) aparecen decolor negro.

NEUROCIENCIA Y PSIQUIATRÍA 1 3
do desde las micropipetas hacia la neurona. Coneste procedimiento pueden ser determinados losefectos del neurotransmisor o del fármaco admi-nistrados tópicamente sobre la actividad espontá-nea de la neurona o la respuesta de ésta ante unaaferencia excitadora o inhibidora.
Para obtener una información más precisa acer-ca de los canales iónicos concretos implicados enlas alteraciones de la respuesta electrofisiológica,los investigadores han empleado cada vez más lastécnicas de registro intracelular, en las que una finamicropipeta es insertada en el cuerpo neuronal paramedir los cambios de voltaje a través de la mem-brana neuronal que se producen como respuesta ala aplicación del neurotransmisor sobre la superfi-cie de la neurona. Los estudios de registro intrace-lular en el animal intacto son sumamente difíci-les, es como pretender pinchar una uva colgada deun palo de 30 metros de altura. De acuerdo conesto, los investigadores están trabajando en la ac-tualidad con estudios de cortes finos del tejido ce-rebral que se conservan vivos durante varias horas,por perfusión mediante un medio fisiológico oxi-genado. En este tipo de preparaciones, las neuro-nas que nos interesan pueden observarse directa-mente con un microscopio de contraste de fases, yel electrodo de registro intracelular puede ser in-
Figura 1-8. Tomografía por emisión de positrones de losreceptores dopaminérgicos D-2 en un cerebro humano. Seha utilizado como marcador (11 C)-espiperona. Obsérvesela intensa fijación del caudado-putamen (C-P), una regiónnotablemente densa en receptores D-2. (Por cortesía de L.T u n e ) .
sertado en las neuronas previamente identificadas,bajo control visual directo.
Finalmente, un procedimiento nuevo para obte-ner información lo más precisa posible sobre el aco-plamiento entre receptores concretos con un canaliónico determinado, es el pinzamiento de canal enel que un fragmento microscópico de membrananeuronal es mantenido por medio de aspiración enla punta de una micropipeta. Con este método, elflujo de corriente a través de los canales iónicos in-dividuales puede ser controlado mediante la expo-sición del canal de la membrana a los receptores an-tagonistas. Modificando la composición iónica, esposible determinar los canales iónicos específicosque están afectados por la influencia del antagonis-ta (sodio, potasio, calcio) y la influencia cinética delantagonista sobre la dinámica del canal (tiempo deapertura, tiempo de circulación, diámetro del canal,etc.). Aunque las propiedades del canal iónico puedaparecer que están lejos de la psiquiatría clínica, pro-porcionan la base fisiológica para la estimulaciónneuronal. Por tanto, la función de los canales es unaconsideración importante cuando se habla de la fi-siopatología de los trastornos psiquiátricos y losefectos globales de fármacos psicotrópicos. Dehecho, el fármaco anticonvulsivo carbamacepina,utilizado en el tratamiento del trastorno bipolar,tiene como principal acción un efecto directo sobrela puerta de entrada de ciertos canales de Na+.
Canales Ligand-gated
Los canales l i g a n d - g a t e d son proteínas receptorasneurotransmisoras que contienen una zona de unióny un poro en el canal. Para formar el poro, cada sub-grupo de la proteína atraviesa la membrana cuatroveces. Los receptores parecen estar formados porcinco subgrupos diferentes ordenados en forma de ba-rril. La activación de este tipo de receptores, que con-tienen canales iónicos intrínsicos de rápida respues-ta, es responsable de la transmisión rápida deinformación de «extremo a extremo» del cerebro. Elneurotransmisor excitador más importante en el ce-rebro parece ser el glutamato. Un subgrupo de re-ceptores de membrana cierran directamente los ca-nales de Na+ de manera que cuando el glutamato seune al receptor, el canal trans-membrana de la mo-lecula receptora se abre para dejar entrar la afluenciade sodio, despolarizando así la neurona. Otros cana-les l i g a n d - g a t e d importantes en el sistema nerviosoincluyen los receptores de acetilcolina nicotínicos.El neurotransmisor inhibidor más importante en elcerebro es GABA, y en la médula espinal el aminoá-cido relacionado glicina. Los canales receptores deGABA y glicina admiten Cl–, lo que, en consecuen-cia, produce una hiperpolarización de la membrananeuronal.

1 4 TRATADO DE PSIQUIATRÍA
Receptores unidos a una proteína G
(G-protein linked)
La neurotransmisión excitadora rápida en el cere-bro parece ser llevada a cabo por una pequeña can-tidad de neurotransmisores, especialmente por glu-tamato. En comparación, con sólo una excepción,(el receptor serotoninérgico 5-HT3), los receptorespara todos los monoaminos y neuropéptidos no cie-rran directamente los canales iónicos, pero actúana través de proteínas estimuladoras de transduc-ción asociadas a la membrana llamadas proteínasG. Como se verá, los receptores unidos a las pro-teínas G están constantemente involucrados en unproceso de modulación de la respuesta de circuitosneurales. Esto añade una gran complejidad a la rá-pida transmisión de impulsos excitadores e inhi-bidores de glutamato, GABA y neurotransmisoresrelacionados por todo el sistema neural.
Los receptores unidos a las proteínas G que hansido analizados estructuralmente hasta ahora enestudios de clonación molecular tienen una es-tructura general común, que cruza la membrananeuronal siete veces (Kobilka, 1992). El dominiode fijación del ligando parece estar en un bolsilloproducido por estos dominios trans-membranadentro del plano de la membrana. El acoplamien-to a los mecanismos intracelulares de estimula-ción tiene lugar en el lado del citoplasma de lamembrana neuronal. Las proteínas G, así llamadasporque unen nucleótidos de guanina, se asociancon el interior de la membrana. La fijación del li-gando al receptor causa un cambio en la confor-mación del receptor que activa las proteínas G. Lasproteínas G, a cambio, transducen la estimulacióna efectos intracelulares.
Las proteínas G son heterotrímeras (es decir,proteínas hechas de tres subunidades diferentes),las subunidades de las cuales se denominan alfa,beta y gama. Con muy pocas excepciones, las su-bunidades alfa, que son muy diversas, son las quecausan específicamente la activación de proteínasG (Simon et al., 1991; Figura 1-9). En el estado inac-tivo, las unidades alfa, beta y gama están unidas yuna molécula de difosfato de guanosina (GDP) seune a la subunidad alfa. Cuando el GDP se activapor un receptor, es reemplazado por un trifosfatode guanosina (GTP) en la subunidad alfa, que luegose disocia del complejo con beta y gama. Esta su-bunidad activa permanece asociada con la mem-brana, donde puede causar la apertura o el cierrede canales de intercambio iónico específicos o laactivación o inhibición de enzimas que producensegundos mensajeros intracelulares.
La acción en particular depende del tipo de su-bunidad alfa que se activa por un receptor dado.
Por ejemplo, los receptores beta-adrenérgicos y do-pamínicos D1 activan una proteína G llamada Gs.La proteína G entera se nombra por su subunidadalfa. La subunidad alfa activa puede estimular cier-tos canales de intercambio iónico de calcio (los ca-nales tipo-L que son el tipo bloqueado por fármacostipo verapamino) y adenilciclasa, una enzima quecataliza la producción del segundo mensajero, AMPcíclico. La subunidad alfa activa tiene una activi-dad intrínseca GTPasa que conduce a hidrólisis deGTP a GDP. Cuando esto ocurre, la subunidad alfase reasocia con beta y gama y la acción finaliza.
Los efectos de las proteínas G en los canales ió-nicos alteran las respuestas de neuronas a la esti-mulación subsecuente a través de neurotransmiso-res excitadores o inhibidores, como el glutamato yel GABA. Por ejemplo, los péptidos opiáceos endó-genos pueden actuar a través de un tipo de receptor(designado mu) para activar un canal K+. Debido aque la fuerza de conducción electroquímica de K+
está fuera de las neuronas, estos opiáceos decrecenla carga positiva neta dentro de neuronas diana. Laneurona, por tanto, responde menos al glutamato (esdecir, se muestra menos inclinada a «disparar»). Estees un mecanismo por el que las proteínas G puedenalterar la respuesta de los circuitos neurales.
Además de sus efectos en los canales iónicos,las proteínas G regulan las enzimas que producenlos segundos mensajeros. Como ya se ha descrito,los receptores ligados a Gs inhiben la adenilcicla-sa para incrementar la producción de AMP cíclico.Los receptores ligados a Gi inhiben la adenilcicla-sa. Otra proteína G, llamada Gq, activa la enzimafosfolipasa C, que hidroliza ciertos fosfolípidos enla membrana para generar segundos mensajeros,diaciglicerol y trifosfato de inositol (IP3) (Figura 1-10). Otras vías importantes de segundos mensaje-ros parecen incluir metabolitos del ácido arachi-dónico y óxido nítrico.
Aunque la cantidad de segundos mensajeros quese encuentra en neuronas es grande, los mecanis-mos de acción pueden generalizarse conceptual-mente. Con pocas excepciones (por ejemplo, elAMP cíclico puede cerrar independientemente cier-tos canales iónicos del sistema olfativo), los se-gundos mensajeros ejercen sus efectos biológicosprincipales a través de protein-quinasas específicas.Las protein-quinasas son enzimas que mueven gru-pos de fosfato de ATP a substratos de proteína es-pecíficos. Basados en su carga y tamaño, los gruposde fosfatos alteran la conformación de proteínas ypor tanto su función. Debido a que la fosforilaciónes una modificación covalente, puede actuar du-rante una extensión de tiempo muy larga. Los subs-tratos para la fosforilización activada del segundomensajero incluyen canales iónicos, receptores, en-

NEUROCIENCIA Y PSIQUIATRÍA 1 5
Figura 1-9. Sistema de segundo mensajero de adenilato. En la parte superior se muestra un esquema de la membrananeuronal. Los receptores neurotransmisores (trama de puntos grises), los canales (se muestra un canal de Na+ en negro),y adenilciclasa (AC) son proteínas integrales de la membrana. Las subunidades de las proteínas G heterotriméricas (vertexto) α, β y γ están asociadas a la superficie interna de la membrana. Se muestra un receptor no ocupado a la izquier-da; en esta circunstancia, la subunidad alfa se une a GDP y las subunidades de proteína G se asocian totalmente. Conel ligando de un neurotransmisor (triángulo negro que se muestra a la derecha), el receptor puede activar la proteína G.La GDP se intercambia por GTP, y la subunidad alfa se disocia de beta y gama. Aquí, α-s se muestra en el centro acti-vando adenilciclasa que cataliza la síntesis del segundo mensajero intracelular c-AMP de la adenosina trifosfato (ATP).El c-AMP activa la proteín-quinesa A (que se muestra fosforilando el canal de calcio), la enzima sintetizadora de neu-rotransmisores tirosina hidroxilasa, y el factor de transcripción CREB en el núcleo de la neurona.
G D P
A T P c A M P
G T P
A R N
P O4
Ca ++
Proteinquiasa A P O4
T i r o s i n ah i d r o x i l a s a
N ú c l e oP O4
A D NC R E B
zimas sintetizadas por neurotransmisores, proteí-nas citosqueléticas y proteínas de control de latranscripción genética. A través de la activación dela fosforilización proteínica, los receptores ligadosa una proteína G regulan diversas funciones den-tro de la célula, y al regular la expresión genética,incluso regulan los constituyentes proteínicos dela célula. La fosforilización que puede, por ejem-plo, desactivar los receptores o aumentar o dismi-nuir la probabilidad de apertura de los canales ió-nicos de barrera de voltaje, alterarán el modo enque las neuronas procesan la información, por loque alterarán el comportamiento de los circuitoscerebrales de formas significativas. Claramente, enconsecuencia, el cerebro no es un simple sistemade alta tensión en donde la información se trans-mite a través de potenciales excitatorios o inhibi-torios. El cerebro está modificando constantemen-te la forma en que sus neuronas pueden procesar lainformación. Esta plasticidad del funcionamientoneuronal es fundamental en procesos como elaprendizaje y la memoria, aunque probablementetambién esté implicado en el origen de estados psi-copatológicos (por ejemplo, los cambios de estadoque aparecen al comienzo de la depresión) y, tal
c o m o se describirá más adelante, esta plasticidadpuede ser la base del mecanismo de acción de mu-chas drogas psicotrópicas.
@ N E U ROA NATO M Í A
M é to d o s
Si bien una descripción detallada de la neuroanato-mía del cerebro está fuera de la intención de estarevisión, merecen mencionarse algunos temas y es-trategias de investigación nuevos. Hasta hace quin-ce años, los procedimientos neuroanatómicos esta-ban restringidos a las técnicas de tinción que poníande manifiesto a las neuronas, en base a unas carac-terísticas químicas que no eran exclusivas de nin-guna clase particular de ellas. Las conexiones entreneuronas podían ser deducidas solamente a partirde métodos indirectos, por medio de estudios rea-lizados con microscopio electrónico, que resulta-ban muy laboriosos. Dos adelantos técnicos prin-cipales han facilitado un progreso enorme en elconocimiento de la organización funcional del ce-rebro. El primero emplea anticuerpos a fin de iden-

1 6 TRATADO DE PSIQUIATRÍA
Figura 1-10. Sistema de segundo mensajero fosfatidilinositol. Muchos receptores neurotransmisores están conectadosvía las proteínasG Gq y, en ocasiones Go a la enzima fosfolipasa C, que hidroliza fosfatidilinositol 4,5-bis-fosfato (PIP2)para generar segundos mensajeros, diaciglicerol e inositol 1,4,5-trifosfato (Ins 1,4,5 P3, más a menudo abreviado IP3). ElI P3 actúa en las neuronas liberando calcio de almacenamiento intracelular. Se metaboliza en formas que pueden serinactivas, incluyendo inositol 1,4,5-tetrafosfato (Ins 1,3,4,5 P4). Estas formas son metabolizadas eventualmente paraproducir tres monofosfatos de inositol diferentes que se distinguen sólo por el átomo de carbono al que el grupo de fos-fatos se une. La síntesis de inositol de glucosa-6-fosfato también debe pasar por un intermediario de monofosfato ino-sitol. Todos los monofosfatos de inositol se metabolizan por la enzima fosfatasa de monofosfato inositol. El litio enconcentraciones terapéuticas inhibe esta enzima. Como resultado, en presencia de litio, los monofosfatos de inositolno pueden desfoforilarse para producir el inositol libre que se necesita para regenerar fosfatidilinositol 4,5-bifosfato.También se muestra en la figura la habilidad del litio para inhibir una enzima adicional en este ciclo (inositol polifos-fato 1-fosfatasa), que se requiere para los dos pasos metabólicos anteriores de la vía de reciclaje. Adaptado de Hyman yNestler, 1993.

NEUROCIENCIA Y PSIQUIATRÍA 1 7
tificar las características químicas de las neuronas,y el segundo revela con facilidad las conexionesneuronales.
El alto grado de especificidad de las interaccio-nes entre antígenos e inmunoglobulinas, permitela tinción selectiva de partes concretas del tejido,como los neurotransmisores, las enzimas, o losmarcadores de superficie contra los que el antisueroha sido preparado. La eficacia y especificidad deestas técnicas han sido potenciadas por la intro-ducción de metodologías con capacidad de generaranticuerpos monoclonales, que reconocen una solaparte de un antígeno (es decir, un epitope).
Con los procedimientos inmunocitoquímicosestándar, se prepara el antisuero capaz de recono-cer un mayor número de epitopes. Aunque puedeperderse algo de especificidad, aumenta la posibi-lidad de detectar el antígeno de interés. Tanto si seutilizan anticuerpos monoclonales como policlo-nales, éstos son incubados con secciones de tejidoy unidos al antígeno que reconocen. Esta reacciónes después visualizada al incubar este corte con unsegundo anticuerpo, como la inmunoglobulinao-vina antirratón, a la que se le ha unido un marca-dor visualizable como la enzima peroxidasa. A con-tinuación, los cortes se revelan y puede serobservado el antígeno en neuronas concretas y ensus estructuras, ya sea con los procedimientos delmicroscopio óptico o electrónico.
La vía inmunocitoquímica ha sido explotada engrado considerable para visualizar la anatomía delas neuronas que utilizan neurotransmisores espe-cíficos, basándose en el desarrollo de anticuerposcontra el neurotransmisor, contra su enzima desíntesis específica, o contra un neuropéptido. A di-ferencia de las técnicas histológicas tradicionalesque revelan las neuronas partiendo de una base decaracterísticas comunes a todas ellas, como la pre-sencia de ácido nucléico cuando se emplean las tin-ciones de Nissl, o la capacidad de ser impregnadascon iones de plata en la tinción de Golgi, la tincióninmunoquímica para los antígenos relacionadoscon neurotransmisores permite la visualización deuna neurona y su axón basándose en el tipo de neu-rotransmisor utilizado (Figura 1-11).
El enfoque inmunocitoquímico está siendo ex-tendido para identificar otros componentes impor-tantes del sistema nervioso, con objeto de entendermejor su organización y función a nivel molecular.Un método adicional para identificar los compo-nentes específicos de las neuronas es la hibridaciónin situ. Esta técnica explota el principio del empa-rejamiento de base complementario que caracteri-za los ácidos nucléicos. Los mensajeros ARNm quecodifican proteínas celulares son de una hebra. UnARN de una sola hebra o sonda de ADN hi b r i d a r á
sólo con un ARNm complentario dentro de unasección de tejido bajo las condiciones experimen-tales correctas. De manera muy similar a lo quepasa en la autorradiografía del receptor, descritaanteriormente, la sección puede después yuxtapo-nerse sobre una película. Los granos plateados pue-den visualizarse describiendo el ARNm de interés.Combinada con la inmunohistoquímica, la hibri-dación in situ ha revelado muchas especies mole-culares que dan a neuronas determinadas sus iden-tidades específicas.
Figura 1-11. Tinción inmunocitoquímica de fibras se-rotoninérgicas en el córtex somatosensorial del mono. Seincubó un corte de 10 µm de córtex de mono previamentefijado, con un anticuerpo antiserotonina de cobaya. Loscomplejos anticuerpo-serotonina en los axones serotoni-nérgicos se visualizaron mediante un anticuerpo ovinoanti gamma-globulina de cobaya conjugado con fluores-ceína. Obsérvese la densa red de axones serotoninérgicosdispuestos aleatoriamente en el campo de observaciónentre las capas I y II.
P = superficie de la pía madre; V = vaso sanguíneo.(Por cortesía de M.A. Wilson y M.E. Molliver.)
1 8 TRATADO DE PSIQUIATRÍA
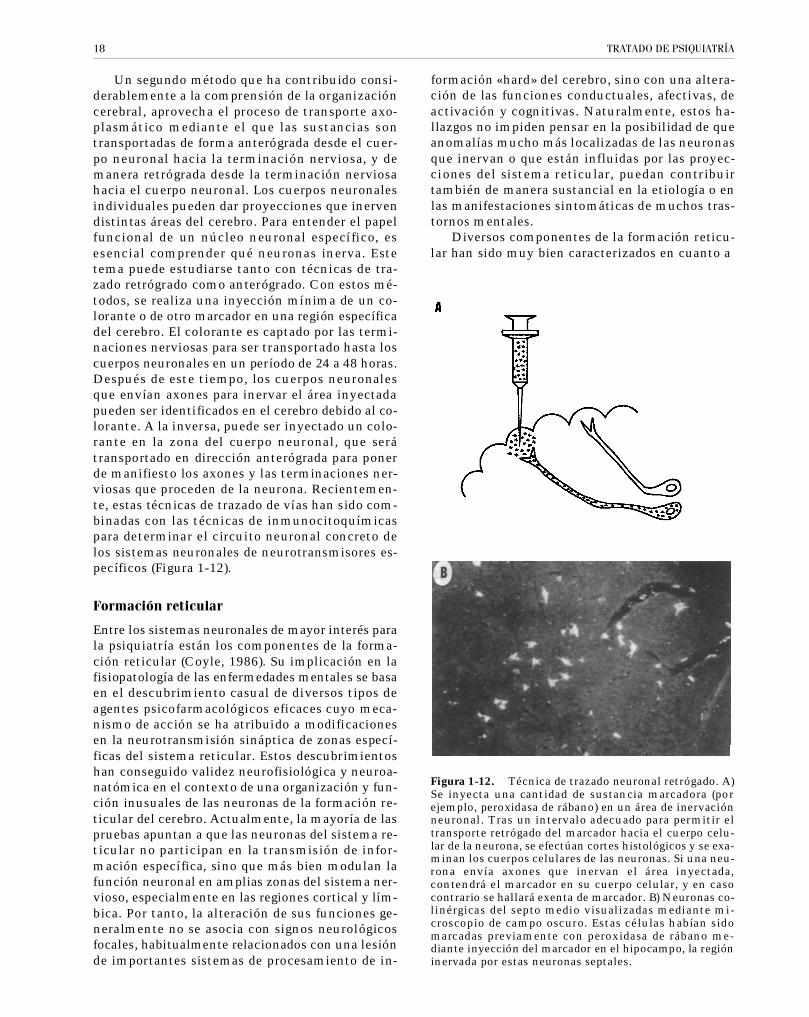
Un segundo método que ha contribuido consi-derablemente a la comprensión de la organizacióncerebral, aprovecha el proceso de transporte axo-plasmático mediante el que las sustancias sontransportadas de forma anterógrada desde el cuer-po neuronal hacia la terminación nerviosa, y demanera retrógrada desde la terminación nerviosahacia el cuerpo neuronal. Los cuerpos neuronalesindividuales pueden dar proyecciones que inervendistintas áreas del cerebro. Para entender el papelfuncional de un núcleo neuronal específico, esesencial comprender qué neuronas inerva. Estetema puede estudiarse tanto con técnicas de tra-zado retrógrado como anterógrado. Con estos mé-todos, se realiza una inyección mínima de un co-lorante o de otro marcador en una región específicadel cerebro. El colorante es captado por las termi-naciones nerviosas para ser transportado hasta loscuerpos neuronales en un período de 24 a 48 horas.Después de este tiempo, los cuerpos neuronalesque envían axones para inervar el área inyectadapueden ser identificados en el cerebro debido al co-lorante. A la inversa, puede ser inyectado un colo-rante en la zona del cuerpo neuronal, que serátransportado en dirección anterógrada para ponerde manifiesto los axones y las terminaciones ner-viosas que proceden de la neurona. Recientemen-te, estas técnicas de trazado de vías han sido com-binadas con las técnicas de inmunocitoquímicaspara determinar el circuito neuronal concreto delos sistemas neuronales de neurotransmisores es-pecíficos (Figura 1-12).
Fo rmación reticular
Entre los sistemas neuronales de mayor interés parala psiquiatría están los componentes de la forma-ción reticular (Coyle, 1986). Su implicación en lafisiopatología de las enfermedades mentales se basaen el descubrimiento casual de diversos tipos deagentes psicofarmacológicos eficaces cuyo meca-nismo de acción se ha atribuido a modificacionesen la neurotransmisión sináptica de zonas especí-ficas del sistema reticular. Estos descubrimientoshan conseguido validez neurofisiológica y neuroa-natómica en el contexto de una organización y fun-ción inusuales de las neuronas de la formación re-ticular del cerebro. Actualmente, la mayoría de laspruebas apuntan a que las neuronas del sistema re-ticular no participan en la transmisión de infor-mación específica, sino que más bien modulan lafunción neuronal en amplias zonas del sistema ner-vioso, especialmente en las regiones cortical y lím-bica. Por tanto, la alteración de sus funciones ge-neralmente no se asocia con signos neurológicosfocales, habitualmente relacionados con una lesiónde importantes sistemas de procesamiento de in-
f o r m a c i ó n «h a r d» del cerebro, sino con una altera-ción de las funciones conductuales, afectivas, deactivación y cognitivas. Naturalmente, estos ha-llazgos no impiden pensar en la posibilidad de queanomalías mucho más localizadas de las neuronasque inervan o que están influidas por las proyec-ciones del sistema reticular, puedan contribuirtambién de manera sustancial en la etiología o enlas manifestaciones sintomáticas de muchos tras-tornos mentales.
Diversos componentes de la formación reticu-lar han sido muy bien caracterizados en cuanto a
Figura 1-12. Técnica de trazado neuronal retrógado. A)Se inyecta una cantidad de sustancia marcadora (porejemplo, peroxidasa de rábano) en un área de inervaciónneuronal. Tras un intervalo adecuado para permitir eltransporte retrógado del marcador hacia el cuerpo celu-lar de la neurona, se efectúan cortes histológicos y se exa-minan los cuerpos celulares de las neuronas. Si una neu-rona envía axones que inervan el área inyectada,contendrá el marcador en su cuerpo celular, y en casocontrario se hallará exenta de marcador. B) Neuronas co-linérgicas del septo medio visualizadas mediante mi-croscopio de campo oscuro. Estas células habían sidomarcadas previamente con peroxidasa de rábano me-diante inyección del marcador en el hipocampo, la regióninervada por estas neuronas septales.

NEUROCIENCIA Y PSIQUIATRÍA 1 9
sus neurotransmisores. Entre ellos, son de parti-cular importancia para la psiquiatría las vías no-radrenérgicas, serotoninérgicas, dopaminérgicas yc o l i n é r g i c a s .
Ne u ronas nora d re n é rgicas
Las neuronas noradrenérgicas utilizan la noradre-nalina como neurotransmisor principal. En la Fi-gura 1-2 se muestra la vía de síntesis de la nora-drenalina. El núcleo noradrenérgico principal es ellocus ceruleus, llamado así por su color azul en laspreparaciones frescas de cerebro. El locus ceruleusse localiza bilateralmente en el suelo del cuartoventrículo (Figura 1-13). Existen otros núcleos no-radrenérgicos y adrenérgicos (neuronas liberadorasde adrenalina) esparcidos por la protuberancia, queinervan en primer lugar el tronco cerebral. Las40.000 neuronas que se estiman existen en el locusceruleus humano son la fuente principal de iner-vación noradrenérgica para la mayor parte del SNC,incluyendo la médula espinal, el cerebelo y las es-tructuras del prosencéfalo. Se calcula que una am-plia arborización axonal abarca más del 95 % delvolumen celular de cada neurona noradrenérgicadel locus ceruleus. Como los otros componentes delsistema reticular, los axones noradrenérgicos sonexpansiones finas y desmielinizadas, que contienenneurotransmisores en toda su longitud. Los abul-tamientos arrosariados distribuidos a lo largo de losaxones son zonas de contacto sináptico especiali-zado conocidas con el nombre de sinapsis de paso.Tal como queda especialmente bien representadoen el córtex cerebral, los axones noradrenérgicos in-dividuales establecen contactos sinápticos conmiles de neuronas, y el árbol axónico aparece comouna densa trama que se ramifica a través de todaslas capas del córtex cerebral (ver Figura 1-13). Ade-más, las neuronas noradrenérgicas individua-
Figura 1-13. Proyecciones primarias del locus ceruleusn o r a d r e n é r g i c o .
les envían axones que inervan regiones del cerebromuy diversas funcionalmente (por ejemplo, córtexcerebral y cerebelo).
En el cerebro, los efectos de la noradrenalinaestán mediados en gran parte por dos clases de re-ceptores: los receptores alfa y los receptores beta(Tabla 1-14). Estas categorías están subdivididas asu vez, en función de sus características farmaco-lógicas y sus efectos fisiológicos, en receptores alfa-1, alfa-2, beta-1 y beta-2. La estimulación de recep-tores alfa-1 se traduce en una excitación neuronalque puede ligarse parcialmente con el movimientode fosfoinosítidos. La clonidina y la alfa-metil-DOPA, son potentes agonistas de los receptoresalfa-2. La activación de los receptores alfa-2 se aso-cia habitualmente con una disminución en la acti-vidad noradrenérgica central y periférica. Los re-ceptores alfa-2 en el soma noradrenérgico, que sonestimulados normalmente a través de colateralesnoradrenérgicos recurrentes, inhiben la tasa de des-carga de las neuronas noradrenérgicas. Además, laactivación de los receptores alfa-2 en las termina-ciones noradrenérgicas reduce la cantidad de nora-drenalina liberada, seguramente por disminucióndel flujo de calcio durante la despolarización de laterminación nerviosa. Estos efectos inhibidores delos alfa-2 agonistas de la clonidina explican su ac-ción ansiolítica y su eficacia en la reducción de cier-tos síntomas en la abstinencia aguda de opiáceos.
Los receptores beta-1 y beta-2 se distinguen prin-cipalmente por una actividad noradrenérgica intrín-seca menor en estos últimos y por su diferente sen-sibilidad a los antagonistas. En el cerebro, losreceptores beta-1 parecen tener un alto grado de lo-calización en las neuronas, mientras que los recep-tores beta-2 se asocian predominantemente, aunqueno de manera exclusiva, con elementos no neurona-les tales como las células gliales. La activación de losreceptores beta se traduce en una estimulación de laadenil ciclasa a través de la proteína Gs, y en una ele-vación de los niveles intracelulares del AMP cíclico.Por tanto, las respuestas celulares de los agonistasbeta reflejan la activación de proteín-quinasas de-pendientes del AMP cíclico, que alteran transitoria-mente la actividad de enzimas específicas a través deuna fosforilación reversible. La desensibilización delos receptores beta corticolímbicos sa ha asociadocon el mecanismo de acción de los antidepresivos.
Neuronas serotoninérgicas
Las neuronas liberadoras de serotonina tienen sussomas localizados en un grupo de núcleos que ro-dean el acueducto en el cerebro medio, conocidoscomo Núcleos del rafe (Figura 1-14). Al igual quelas neuronas noradrenérgicas del locus ceruleus, las

2 0 TRATADO DE PSIQUIATRÍA
Figura 1-14. Vías de las neuronas serotoninérgicas del rafe.
neuronas serotoninérgicas proporcionan una iner-vación formada por numerosos colaterales que sedistribuyen hacia prácticamente todas las áreas delSNC. Sin embargo, los componentes de los núcle-os del rafe proporcionan una inervación más re-gionalizada.
Los efectos postsinápticos de la serotonina estánmediados por varios receptores pre y post-sinápticos(Tabla 1-4). Los estudios recientes farmacológicos yde clonación sugieren que existen al menos cuatrotipos de receptores serotoninérgicos (5-hidroxitrip-tamina (5-HT)) con múltiples subtipos. Los recepto-res 5-HT1 relevantes en la farmacología humana sonel receptor 5-HT1 a, un receptor mayoritariamentepresináptico, zona de acción del fármaco ansiolíticobuspirona; el receptor 5-HT1 c, que es tanto pre comopostsináptico; y el receptor 5-HT1 d, del cual es an-tagonista el nuevo fármaco sumatriptano. El recep-tor 5-HT2 es un receptor postsináptico que pareceser la zona de acción clave de la dietilamida del ácidolisérgico (LSD), la mescalina y otros alucinógenosrelacionados. Los receptores 5-HT1 a y 5-HT1 d i n h i-ben la adenilciclasa y activan el potencial iónico delos canales de K+ a través de Gi. Los receptores 5-H T1 c y 5-HT2 activan las vías del segundo mensaje-ro inositol trifosfato/diacilglicerol. El receptor 5-HT3
es el único receptor monoamino conocido que seaun canal l i g a n d - g a t e d. El fármaco reciente antie-mético odansetrón antagoniza los efectos excitado-res de la serotonina en la recepción 5-HT3 en la zonade la médula químicamente activada. (El otro tipode receptor significativo de estas neuronas es el re-ceptor dopamínico D2; por tanto, los antieméticosmás antiguos son antagonistas de receptores dopa-minérgicos.) No está clara la función de los recep-tores 5-HT4. Las neuronas serotoninérgicas, a travésde la inervación del córtex cerebral, han sido aso-ciadas con la regulación del estado de vigilia.
Los efectos serotoninérgicos en el sistema límbicopueden tener un papel en el control del estado deánimo, la ansiedad, la agresión, y en la modulacióndel dolor.
Neuronas dopaminérgicas
Para la investigación psiquiátrica, han sido de es-pecial interés tres grandes sistemas dopaminérgi-cos (ver Figura 1-15). Los cuerpos celulares dopa-minérgicos ampliamente pigmentados localizadosen la sustancia negra proporcionan una inervaciónconsiderablemente densa al caudado y al putamen,y constituyen aproximadamente un 15% de las si-napsis de dichas estructuras. Esta vía de axonesdesmielinizados, provista de un gran número decolaterales, se ramifica en una fina filigrana de axo-nes repletos de varicosidades, que proporcionanmillares de sinapsis de paso. Las proyecciones do-paminérgicas negrostriadas están íntimamente im-plicadas en la iniciación, mantenimiento y ejecu-ción ajustada de las actividades motoras, y puedeninfluir de forma similar en las funciones cogniti-vas, como reflejo de la gran conexión que va desdeel córtex frontal hasta el caudado. La degeneraciónde la vía dopaminérgica negrostriada provoca lossíntomas de la enfermedad de Parkinson, y el blo-queo de los receptores dopaminérgicos por los neu-rolépticos genera intensos efectos colaterales ex-trapiramidales, lo que indica una alteración en laneurotransmisión dopaminérgica del estriado.
Figura 1-15. Las tres principales vías dopaminérgicas.Comprenden la nigrostriada, la mesocorticolímbica (quetiene origen en «A-10») y la vía del núcleo arqueado ali n f u n d í b u l u m .

NEUROCIENCIA Y PSIQUIATRÍA 2 1
Los somas neuronales dopaminérgicos de la re-g i ó n «A - 1 0» , situados más centralmente, inervanel nucleus accumbens, la zona central del circuitolímbico, y también el córtex frontal, el córtex cin-gulado y el hipocampo. La inervación cortical estámucho más dispersa que en el estriado y en el ac-cumbens, y parece tener una distribución comple-mentaria a la de los aferentes noradrenérgicos (Le-vitt et al., 1984). La proyección dopaminérgica dela región A-10 al núcleo accumbens se ha impli-cado como un circuito cerebral de recompensa, quemediatiza los efectos positivos del abuso de dro-gas, incluyendo la cocaína, la anfetamina y proba-blemente los opiáceos. Las proyecciones dopami-nérgicas corticales pueden estar implicadas en laatención, la «memoria activa» y, por inferencia, laintegración cognitiva. Basándose en las propieda-des antagonistas dopamínicas de los fármacos an-tipsicóticos (véase más adelante), se ha supuestoque en la esquizofrenia y otros trastornos psicóti-cos existe una disfunción de los circuitos dopami-nérgicos mesocorticolímbicos.
Finalmente, un grupo de cuerpos neuronales do-paminérgicos, localizados en el núcleo arqueado delhipotálamo, emite axones que terminan en los senosvenosos de la hipófisis. Estas proyecciones dopami-nérgicas inhiben, en concreto, la liberación de la pro-lactina, una hormona de la hipófisis. Este papel fi-siológico ha sido ampliamente utilizado comomedida periférica de las alteraciones de la neuro-transmisión dopaminérgica central. Es el caso de losneurolépticos que, como potentes bloqueadores delos receptores D2 de dopamina, aumentan la libera-ción de prolactina. Los efectos sinápticos de la do-pamina parecen ser mediados por varios receptoresfarmacológica y fisiológicamente diferentes (Tabla1-4). Es probable que no se hayan descubierto todoslos tipos de receptores dopaminérgicos y, actual-mente, no existe una nomenclatura establecida. Enbase a su estructura (que se deduce de la clonaciónmolecular) y sus propiedades farmacológicas, los re-ceptores dopaminérgicos pueden agruparse en dos fa-milias, llamadas tipo D1 (los receptores D1 y D5, tam-bién llamados D1 a y D1 b por algunos) y las tipo D2 ( l o sreceptores D2, D3 y D4, también llamados D2 a, D2 b yD2 c por algunos). Además, se ha encontrado unaforma larga y una corta del receptor D2 a partir de ladivisión del receptor D2 ARNm, pero no se han acla-rado las diferencias funcionales de las dos formas.Los diferentes tipos de receptores tienen distribu-ciones que se solapan aunque no son idénticas en lasregiones del cerebro inervadas por las fibras dopami-nérgicas. Los receptores D1 y D5 activan la adenilci-clasa a través de Gs. El receptor D2 inhibe la adenil-ciclasa y activa el potencial iónico de un canal de K+
a través de Gi. Los efectos precisos de segundo
mensajero de los receptores D3 y D4 todavía no sonclaros. Los fármacos antipsicóticos son antagonis-tas de los receptores D2, D3 y D4. Una observaciónintrigante, que se comenta más abajo, es que la clo-zapina, un fármaco antipsicótico atípico, tiene unaafinidad particularmente alta para los receptoresD4 (Van Tol et al., 1991). Su baja afinidad al re-ceptor D2 probablemente explica la falta de efec-tos extrapiramidales.
Neuronas colinérgicas
La mayor fuente de inervación colinérgica que reci-be el córtex cerebral, el hipocampo y las estructuraslímbicas, es un complejo de grandes somas neuro-nales muy sensibles a la acetilcolinesterasa, locali-zados en la base del prosencéfalo (Figura 1-16). El nú-cleo basal de Meynert, un grupo algo disperso desomas colinérgicos localizado en la zona ventral ymedia del globus pallidus, envía axones que inervanel córtex cerebral. La banda diagonal de Broca, loca-lizada más hacia delante, y el núcleo septal medioinervan la formación hipocámpica y el córtex cin-gulado. Las ramificaciones terminales de los aferen-tes colinérgicos proporcionan una trama de fibrasorientadas al azar, distribuidas hacia todas las capasdel córtex cerebral. Por el contrario, en el hipocam-po, aparece una distribución específica mucho máslaminar, especialmente en la circunvolución denta-da. La inervación densa del caudado y del putamenno está proporcionada por estas proyecciones ascen-dentes, sino por neuronas de circuito local, y las ra-mificaciones de su axón se limitan al ganglio basal.
Los efectos postsinápticos de la acetilcolina enel prosencéfalo parecen estar mediados sobre todo
Figura 1-16. Neuronas colinérgicas prosencefálicas. Lasneuronas colinérgicas de la base del prosencéfalo (inclu-yendo el núcleo basal de Meynert, la banda diagonal deBroca y el núcleo septal medio) inervan estructuras del cór-tex cerebral, el hipocampo y el sistema límbico. El estria-do posee un circuito local de interneuronas colinérgicas.

2 2 TRATADO DE PSIQUIATRÍA
TABLA 1-4. FARMACOLOG´IA DE LOS RECEPTORES DE MONOAMINA Y ACETILOLINA
R e c e p t o r
A g o n i s t a sc l í n i c a m e n t er e l e v a n t e s
A n t a g o n i s t a sc í n i c a m e n t er e l e v a n t e s C o u p l i n g
Receptores adrenérgicos
α1-adrenérgicos (4 subtipos) F e n i l e f r i n a P r a z o s i n a I P3/ D Gα2-adrenérgicos (3 subtipos) C l o n i d i n a Y o h i m b i n a cAMP ⇓ ; K+ canal ↑ß-adrenérgicos (3 subtipos: ß1 en el I s o p r o t e r e n o l P r o p a n o l o l cAMP ↑
corazón; ß2 en los pulmones)
Receptores dopaminérgicos
D1 ( o D1 A) S F K 3 8 3 9 3 S C H 3 9 1 6 6 cAMP ↑D2 ( o D 2 A) B r o m o c r i p t i n a Antipsicóticos típicos cAMP ⇓ ; Κ+ canal ↑;
(selectivos raclopide) C a+ + canal ⇓D3 (o D2 B) K i n i p o r o l a Antipsicóticos típicos d e s c o n o c i d oD4 (o D2 c) K i n i p o r o l a C l o z a p i n a cAMP ⇓D5 (o D1 B) S F K 3 8 3 9 3 cAMP ↑
Receptores serotoninérgicos (5-HT)
5 - H T1 A Agonista parcial buspirona cAMP ⇓ ; K+ canal ↑(8-OH-DPAT selectivo)
5 - H T1 B cAMP ⇓5 - H T1 D S u m a t r i p t a n cAMP ⇓5 - H T1 E cAMP ⇓5 - H T1 F cAMP ⇓
5 - H T2 A α- M e t i l o - 5 - H T Ritaserin (selectivo) I P3/ D G5 - H T2 B α- M e t i l o - 5 - H T Clozapina (no selectivo) I P3/ D G5 - H T2c (antes 5-HT 1 C) α- M e t i l o - 5 - H T Clozapina (no selectivo) I P3/ D G
5 - H T3 2 - M e t i l o - 5 - H T O d a n s e t r o n Canal Catiónico intrínseco5 - H T4 5 - M e t o x i t r i p t a m i n a cAMP ↑
Receptores muscarínicos O x o t r e m o r i n a A t r o p i n a
M1 O x o t r e m o r i n a Pirenzepina selectivo I P3/ D GM2 O x o t r e m o r i n a Metroctamina selectivo cAMP ⇓ ; K+ canal ↑M3 O x o t r e m o r i n a H e x a h i d r o s i l a d i f e i d o l I P3/ D G
s e l e c t i v oM4 O x o t r e m o r i n a T r o p i c a m i d e cAMP ⇓
Nota: DG = Diacilglicerol; 8-OH-DPAT = hidroxi-2-(di-u-propilamino)tetralín.
por los receptores muscarínicos. Aunque en el ce-rebro puede ser demostrada la existencia de res-puestas nicotínicas, este tipo de receptores pare-cen ser distintos a los que median los efectos de laacetilcolina en la unión neuromuscular periférica.Aparte del propio efecto psicotrópico central de lanicotina, todavía se conoce poco la influencia delos receptores nicotínicos en el cerebro.
Por lo menos se han identificado cuatro tiposde receptores muscarínicos colinérgicos gracias alos estudios farmacológicos y de clonación (M1- M4)(Tabla 1-4). Un quinto receptor muscarínico iden-tificado por clonación no se ha establecido todavíacomo funcional. Los receptores muscarínicos quemedian los efectos de la acetilcolina en las pro-longaciones colinérgicas corticales y del hipocam-po tienen una función integradora en las funcionescognitivas más elevadas, especialmente en la me-
moria a corto plazo y su traducción en memoria alargo plazo. Así pues, los fármacos que bloqueanestos receptores, o la destrucción de las prolonga-ciones colinérgicas de la base del prosencéfalo enanimales de experimentación, provocan defectosselectivos en las funciones de la memoria recien-te. Hay que señalar que una pérdida importante deaxones colinérgicos del córtex y del hipocampo pa-rece ser una alteración habitual de la enfermedadde Alzheimer, y puede contribuir al deterioro cog-nitivo típico de esta enfermedad (Coyle et al.,1983). El sistema colinérgico se ha relacionadotambién con los estados de ánimo, ya que los an-tagonistas de los receptores muscarínicos mejoranel estado de ánimo en el hombre, mientras que lafisostigmina, un inhibidor de la acetilcolinestera-sa de acción central, se ha descrito que provoca unestado de ánimo depresivo. Finalmente, las pro-

NEUROCIENCIA Y PSIQUIATRÍA 2 3
yecciones colinérgicas tienen un papel en el sueño,especialmente en el sueño REM. Durante éste, lasneuronas noradrenérgicas del locus coeruleus estántónicamente inhibidas, mientras que las neuronascolinérgicas están activas. Los fármacos colinérgi-cos estimulan el REM, y los fármacos anticolinér-gicos lo antagonizan. El hecho de que la latenciadel REM disminuya durante una depresión mayor(consistente con una hiperactividad de las neuro-nas colinérgicas) hace más evidente que los siste-mas colinérgicos pueden tener un papel en la re-gulación y en los trastornos del estado de ánimo.
Aminoácidos
En el cerebro, los principales neurotransmisoresexcitadores e inhibidores son los aminoácidos L-glutámico y GABA. Su amplio papel en el proce-samiento de la información indica que están loca-lizados en un gran número de sistemas neuronalesdistintos en todo el cerebro, al revés que las neu-ronas de la formación reticular, cuyos somas estánconcentrados principalmente en núcleos concre-tos dentro del tronco cerebral.
G A B A
Las neuronas GABAérgicas son especialmente im-portantes para la psiquiatría porque numerosos se-dantes, ansiolíticos y anticonvulsivantes ejercen susefectos a través de la activación de los receptoresGABA (ver más adelante). En el córtex cerebral, elhipocampo y las estructuras límbicas, las neuronasGABAérgicas son predominantemente neuronas decircuito local que tienen tanto sus somas como susterminaciones axónicas dentro de dichas estructu-ras (ver Figura 1-17). En realidad, domina en ellas laneurotransmisión inhibidora GABAérgica, dado queel bloqueo farmacológico de los receptores para elGABA con el fármaco bicuculina provoca una de-sinhibición difusa y convulsiones. Las neuronasGABAérgicas pueden ser también neuronas con pro-yecciones de largo recorrido en otras áreas del ce-rebro. Por ejemplo, las eferencias principales quedesde el caudado-putamen se dirigen hacia el glo-bus pallidus y la sustancia negra, son neuronas GA-BAérgicas. Aparte de su vulnerabilidad en la enfer-medad de Huntington, lo que contribuye a lossíntomas discinéticos de esta enfermedad, descu-brimientos recientes indican que en la discinesiatardía se produce la pérdida de la eferencia GABA-érgica de largo recorrido. Los síntomas cerebelo-sos, tales como la ataxia que se manifiesta con unadosis excesiva de barbitúricos, reflejan segura-mente la potenciación de la neurotransmisión GA-BAérgica en este tipo de eferencias del cerebelo.
Glutamato
El glutamato es el neurotransmisor excitador másimportante del cerebro. Algunos ejemplos estudia-dos de neuronas que utilizan glutamato, incluyenlas células piramidales en el córtex cerebral, las cé-lulas hipocámpicas (Figura 1-18), y los aferentessensoriales primarios. La mayor parte de la neuro-transmisión excitadora en el cerebro utiliza recep-tores de glutamato que son canales l i g a n d - g a t e d.Estos receptores han sido llamados por sus antago-nistas farmacológicos, cainato, ácido-α- a m i n o - 3 - h i-droxil-5-metil-4-fosfonobutírico (AMPA), y N-metil-D-aspartato (NMDA). Los subtipos de estosreceptores son en la actualidad sujeto de intensa in-vestigación basada en la clonación molecular de lassubunidades receptoras nuevas (Nakanishi et al.,1990). También hay receptores de glutamato queactivan las proteínas G, actualmente conocidascomo receptores de glutamato «m e t a b o t r ó p i c o s» .Los estudios sobre clonación también han identifi-cado múltiples subtipos de estos receptores.
La unión del glutamato causa que los recepto-res de cainato y AMPA abran un canal intrínsicode Na+, aunque ciertos subtipos también puedenadmitir Ca+ +. Los receptores de NMDA son úni-cos porque su canal, que puede permitir la entra-da de Na+ y Ca+ +, es bloqueado por Mg+ en ciertascondiciones. La activación de los receptores deNMDA sólo puede ocurrir cuando dos hechos sedan simultáneamente: el glutamato debe adherir-se al receptor y la membrana debe despolarizarse(por activación de los receptores de glutamato no-NMDA que lo rodean), lo cual permite que el Mg+
salga por el canal. Debido a que hacen falta dos he-chos simultáneos para la activación del receptor
Figura 1-17. Principales vías GABAérgicas. El neuro-transmisor inhibidor GABA (ácido gamma-aminobutírico)se sintetiza en un circuito local de células estrelladas delcórtex cerebral, en las células cerebelosas de Purkinje y enlas neuronas nigrostriadas.

2 4 TRATADO DE PSIQUIATRÍA
NMDA (el receptor es un detector de coinciden-cias), los receptores NMDA se han consideradocomo un posible substrato para el aprendizaje aso-ciativo.
El glutamato ha sido implicado en un númerocreciente de trastornos neurológicos y psiquiátri-cos. Un hallazgo interesante para la psiquiatría esque los efectos psicotomiméticos de la droga PCPpenciclidina y los componentes relacionados sondebidos a la habilidad de estos para bloquear elcanal de receptores NMDA (Martin y Lodge, 1985).Debido a que el gutamato es el neurotransmisor delas neuronas piramidales corticales e hipocámpi-cas, se ha supuesto que los efectos disociativos ypsicotomiméticos del PCP pueden reflejar una in-terferencia con la neurotransmisión glutamatérgi-ca en esos sistemas.
Olney (1969) fue el primero en demostrar que lainyección periférica de glutamato en animales re-cién nacidos producía un modelo selectivo de de-generación neuronal que afectaba a las neuronas delnúcleo arqueado del hipotálamo, los órganos cir-cumventriculares del cerebro, y las capas más in-ternas de la retina. Propuso que la neurotoxicidaddel glutamato era el resultado de una excitación ex-cesiva de las neuronas, mediada por receptores deglutamato excitadores. Estudios posteriores pusie-ron de manifiesto que la inyección intracerebral deagonistas, de los tres subtipos principales de recep-tores glutámicos (ácido caínico, ácido quiscuálicoy receptores de NMDA) mataban las neuronas pró-ximas al lugar de la inyección pero no los axonesde las neuronas alejadas o elementos no neurona-
Figura 1-18. Vía glutamatérgica principal. El neurotrans-misor excitador ácido L-glutámico se libera en numerosasneuronas, como las células piramidales corticales e hipo-cámpicas, las células granulares cerebelosas, las fibras tre-padoras cerebelosas y las aferentes sensoriales primarias.
les tales como las células gliales. Según la localiza-ción de la inyección cerebral, estas «e x c i t o t o x i n a s»podían reducir la patología de diversas enfermeda-des neurodegenerativas, incluidas la enfermedad deHuntington, la epilepsia del lóbulo temporal y la de-generación espinocerebelosa (Schwarz y Meldrum,1985).
Estas observaciones llevaron a los científicos apreguntarse si el glutamato y otros neurotransmi-sores excitadores endógenos afines podían causaruna degeneración neuronal en el cerebro, en cier-tas circunstancias, como resultado de una libera-ción excesiva o de una inactivación insuficiente.Con el desarrollo de antagonistas potentes y espe-cíficos de los receptores de NMDA, ciertos hallaz-gos recientes muy esperanzadores han ratificadoesta hipótesis. El tratamiento con antagonistas dela NMDA previene la degeneración de las neuro-nas del sistema límbico causada por ataques per-sistentes en la degeneración a neuronas del estria-do como consecuencia de una hipoglucemia aguda,y del hipocampo como resultado de cinco minutosde isquemia completa. Estos resultados auguran eldesarrollo de nuevas clases de medicamentos neu-ropsicotrópicos que puedan prevenir o restringirsensiblemente el daño cerebral como consecuen-cia de la hipoxia y la isquemia, las causas más fre-cuentes de morbilidad aguda y muerte como con-secuencia de infartos de miocardio y enfermedadesvasculares (Robinson y Cole, 1988).
P u rinas
Así como ciertos aminoácidos que construyen gru-pos de proteínas (por ejemplo, el glutamato o la gli-cina) pueden actuar como moléculas estimulado-ras en el sistema nervioso, se ha visto que ciertosgrupos de purinas construidos a partir de ácidos nu-cléicos también pueden actuar como neurotrans-misores. La adenosin purina actúa a través de dostipos de receptores ligados a proteínas G. Se ha es-tablecido que los efectos estimulantes de la cafeí-na son el resultado de su acción como un antago-nista competitivo de los receptores de adenosina.Además, parece ser que el ATP, la principal fuen-te de energía de las neuronas, también puede ac-tuar como neurotransmisor. Un tipo de receptorATP ha mostrado ser un canal l i g a n d - g a t e d.
E n d o rfinas
Tal y como se describe en este capítulo, los neu-ropéptidos más importantes para la psiquiatría sonlas endorfinas. Esta familia de neuropéptidos fuedescubierta originalmente en estudios destinadosa determinar si existían sustancias cerebrales en-dógenas que sirvieran como agonistas de los re-

NEUROCIENCIA Y PSIQUIATRÍA 2 5
ceptores de narcóticos. Los pentapéptidos met— yleuencefalina fueron los primeros péptidos opiá-ceos endógenos descubiertos, aunque estudios pos-teriores han revelado la existencia de una familiacompleta de estos péptidos con distintos efectossobre las diversas subclases de receptores opiáce-os. En la Tabla 1-3 se indica una lista de los pépti-dos opiáceos identificados en el cerebro.
Las encefalinas se encuentran principalmenteen neuronas de circuito local en diversas áreas delSNC, pero también se encuentran en neuronas deproyección. Las encefalinas se encuentran en lasustancia gris situada alrededor del acueducto enel cerebro medio, que es una importante estaciónde relevo para los estímulos dolorosos. Las inter-neuronas que contienen encefalina que liberanneuronas dopaminérgicas VTA de la inhibición tó-nica a través de las neuronas GABAérgicas, pue-den presentar parte del sustrato de la recompensacerebral inducida por opio y, por tanto, de abuso yadicción opiácea. Las encefalinas se hallan tam-bién en altas concentraciones en el caudado, el pu-tamen y en el globus pallidus, áreas probablemen-te relacionadas con procesos motores.
El péptido opiáceo endógeno de mayor tamaño,la beta-endorfina, tiene una doble localización enel cerebro. La beta-endorfina se localiza en un grupode neuronas dentro del hipotálamo que emiten axo-nes que se proyectan hacia las áreas límbicas. Ade-más, la beta-endorfina es segregada por los cortico-tropos en la hipófisis anterior. La colocalización deACTH y beta-endorfina refleja el origen común deestos dos péptidos a partir del precursor POMC. Du-rante etapas de estrés, la liberación de ACTH y beta-endorfina por parte de la hipófisis se incrementa no-tablemente. Y los altos niveles de beta-endorfinacirculante pueden explicar la analgesia relacionadacon el estrés. Es más, las alteraciones hipotálamo-hipófisis-adrenales observables en las depresionesmayores, implican una secreción excesiva de beta-endorfina así como de ACTH y cortisol.
@ N E U RO B I O LOGíA MOLECULAR
Quizás los descubrimientos más interesantes de lainvestigación neurocientífica reciente procedan deluso creciente de las técnicas de la biología mole-cular (Hyman y Nestler, 1993). Estos métodos abrenconsiderables esperanzas para poder salvar el lapsoentre la genética clínica en psiquiatría, y los pro-cesos moleculares que regulan la estructura y lafunción cerebral. Estos métodos poderosos inclu-yen el análisis de acoplamiento con los polimor-fismos del DNA para localizar los genes que pue-den ser responsables de trastornos psiquiátricos,
métodos para identificar los genes que codificanlas proteínas implicadas en la estructura y funcióndel cerebro, y los métodos para estudiar la regula-ción de la expresión genética por estímulos am-bientales, incluyendo drogas. Con el conocimien-to, rápidamente creciente, acerca de la localizaciónde los genes importantes para el funcionamientocerebral en el genoma humano, habrá probable-mente en la próxima década una convergencia deinformación que aclarará la base molecular y ce-lular de muchas enfermedades psiquiátricas y deconductas no patológicas.
Clonación molecular
La aproximación más utilizada para identificar y lo-calizar los genes más importantes en relación con elcerebro requiere la purificación de la proteína que in-teresa. La proteína puede ser una enzima relaciona-da con la índole del neurotransmisor, un receptor, unneuropéptido o una proteína estructural. La proteí-na, o un fragmento importante de aproximadamen-te 20 aminoácidos de longitud, es secuenciada paradeterminar la serie de aminoácidos. Dado que se co-nocen los códigos del ADN para cada aminoácido enparticular, es posible sintetizar un filamento de ADNcapaz de codificar la secuencia entera de aminoáci-dos. Debido a que se conoce el código genético, la se-cuencia de proteína puede ser traducida a la inversaen una secuencia de ADN y ser utilizada para gene-rar sondas de ADN sintético marcadas radioactiva-mente (es decir, oligonucleótido) que pueden utili-zarse para identificar el gen que interese. Esto seconsigue utilizando la sonda para la hibridación deun fragmento de ADN complementario (ADNc) den-tro de lo que se llama la «biblioteca de ADNc» .
En resumen, el proceso de la clonación molecu-lar requiere enzimas llamadas endonucleasas de res-tricción que cortan el ADN en secuencias específi-cas, y enzimas llamadas ligasas que pueden unir losfragmentos de ADN. Una biblioteca de ADNc con-tiene una mezcla de fragmentos de ADN que repre-sentan todos los genes expresados en un determina-do tejido o tipo neuronal. Para desarrollar unabiblioteca de ADN, un tejido particular (por ejemplo,una región cerebral) se disecciona y homogeniza, ysu ARNm es extraído químicamente. Una muestrade tejido del ARNm da una representación de todoslos genes contenidos que han sido transcritos (o ex-presados). De manera que, por ejemplo, una muestrade estriatum contendría receptores dopaminérgicoscodificados, pero no así el ARNm que codifica la he-moglobina. Las copias de ADN complementarias decada ARN se producen luego en un proceso que in-cluye la enzima inversora transcriptasa (así llamadaporque transcribe a la inversa ARN a ADN).

2 6 TRATADO DE PSIQUIATRÍA
Cada uno de los ADNc que salen como resulta-do deben ser propagados (es decir, clonados) inser-tándolos en un plásmido bacterial o en virus bac-teriófago de ADN. Ambos pueden servir de vectorque puede replicarse de forma autónoma en bacte-rias como la Escherichia coli. Los plásmidos son pe-queños círculos de ADN que contienen solo unospocos genes; los plásmidos utilizados en la clona-ción se diseñaron a partir de plásmidos resistentesa los antibióticos que ya se encontraban en la bac-teria. La endonucleasa de restricción se utiliza paraabrir el vector de clonación. Cada vector se vuelvea cerrar con ligasas después de introducir un ADNc.La población de plásmidos o bacteriófaga, cada unocon un contenido de una copia de ADNc de un geneexpresado en el tejido de interés, se vuelve a intro-ducir en una Escherichia coli, donde se duplica.Cuando la bacteria se cultiva en un medio agar,cada una crecerá hasta formar una colonia que con-tiene múltiples copias de un solo ADNc, la llama-da biblioteca de ADNc. Se puede hacer un cribadoen la biblioteca para hallar genes concretos utili-zando sondas que detectarán secuencias particula-res por emparejamiento de base complementaria.
Un ejemplo del poder de este método era la clo-nación del gen que codifica la proteína amiloidea,el mayor componente de las placas seniles neurí-ticas en la enfermedad de Alzheimer. La proteínaamiloidea se purificó de los cerebros de pacientescon la enfermedad de Alzheimer determinándosela secuencia de aminoácidos. De esta secuencia, sesintetizó y utilizó una sonda para cribar una bi-blioteca de ADNc cerebral. Sin embargo, el poderde estos métodos va más allá. Pueden utilizarsepara la clonación de ADNc para proteínas quenunca han sido purificadas. Tal y como se ha des-crito anteriormente, muchas proteínas importan-tes en el sistema nervioso son miembros de fami-lias relacionadas evolutivamente. Éstas incluyenreceptores, transportadores y canales iónicos. Re-cientemente, los transportadores de GABA y la no-radrenalina se purificaron y clonaron (Pacholczyket al., 1991). Entonces fue posible comparar sus se-cuencias y determinar qué regiones se conservaronentre las dos proteínas transportadoras. Las regio-nes de conservación de secuencias compartidas porproteínas relacionadas generalmente representandominios funcionales críticos que han sido pre-servados de la tendencia mutacional por la selec-ción natural. Las sondas sintéticas de ADN que co-difican estas regiones conservadas se utilizan luegopara cribar bibliotecas bajo condiciones que tole-ran un pequeño grado de desparejamiento entre lasonda y su complemento; alternativamente, lasonda se puede utilizar en un proceso que amplifi-ca los ADNc a los que une una solución (una téc-
nica que se llama reacción en cadena polimerasa,o RCP). Explotadas las secuencias transportadorasconservadas derivadas de transportadores de GABAy noradrenalina, transportadores de serotonina ydopamina y otros transportadores desconocidos seclonaron. Se utilizaron técnicas similares para laclonación de la famila entera de receptores dopa-mínicos (Van Tol et al., 1991).
Po l i fo rmismos del ADN
Los avances recientes han proporcionado diferentesmétodos para localizar los genes responsables de en-fermedades hereditarias que no requieren ningúnconocimiento previo acerca de la expresión del pro-ducto genético defectuoso responsable de la enfer-medad (Wexler et al., 1991). Dado que en psiquia-tría habitualmente se conoce muy poco acerca delos productos genéticos alterados que pueden ser res-ponsables de un número creciente de enfermedadespsiquiátricas, tales como la enfermedad bipolar, laesquizofrenia o el síndrome de la Tourette, sobre lasque hay evidencia de una predisposición heredita-ria, este método representa una poderosa herra-mienta para aclarar los mecanismos involucradosen dichas enfermedades. La técnica permite la iden-tificación del «l o c u s» cromosómico relacionado es-trechamente con la manifestación fenotípica de undefecto genético, y por tanto resulta prometedorapara identificar de forma definitiva los genes alte-rados responsables de las enfermedades. Una vezidentificado el gen, puede ser caracterizada su fun-ción, lo que en muchos casos implicará un produc-to genético deficiente o alterado.
Estos métodos representan un tipo especial dea n álisis de la cadena genética, excepto que el vín-culo no es a un marcador fenotípico como el tipoHLA, sino a un marcador ADN polimórfico. Estosmarcadores son regiones detectables de cromoso-mas que contienen variaciones en la primera se-cuencia de ADN (es decir, polimorfismos) en la po-blación humana. Dos de los tipos más importantesde polimorfismos de ADN de uso común en estosanálisis son los polimorfismos de longitud de losfragmentos de restricción (RFLP) y los polimorfis-mos en números de secuencias repetidas, más a me-nudo repeticiones de dinucleótidos, pero tambiénsecuencias más largas llamadas números variablesde repeticiones en tándem (VNTR). Los marcadoresde ADN polimórficos pueden utilizarse en el análi-sis de cadenas, observando la co-segregación de unrasgo de interés (por ejemplo, el trastorno bipolar)dentro de un pedigrí con un marcador particular.
En el método RFLP, la demostración de la va-riación de secuencias entre individuos depende delas endonucleasas de restricción, las enzimas men-

NEUROCIENCIA Y PSIQUIATRÍA 2 7
cionadas anteriormente que reconocen secuenciasparticulares en el ADN y lo dividen dentro o cercade la secuencia (Botstein et al., 1980). Si ocurre unavariación dentro de la secuencia en el genoma hu-mano en la secuencia de reconocimiento de unaenzima de restricción, aunque altere tan sólo unasola base, la enzima ya no se dividirá en esa zona.Alternativamente, la variación puede crear una se-cuencia de reconocimiento donde no existía pre-viamente. Una pérdida o ganancia en la zona derestricción, producirá fragmentos de diferentes ta-maños cuando el ADN sea digerido por la enzima.Son estos fragmentos de diferentes tamaños los quese llaman RFLPs.
En la práctica, la enzima de restricción se utili-za para digerir ADN de los linfocitos provenientesde un sujeto concreto. Una enzima de restriccióndada puede dividir el ADN del genoma humano totalen más de un millón de veces y producir un núme-ro inicialmente no-interpretable de fragmentos deADN de muchos y variados tamaños. Sin embargo,los fragmentos pueden ordenarse por tamaños utili-zando electroforesis de gel. En la electroforesis, elADN, que tiene una carga negativa, migra hacia uncátodo en un campo eléctrico aplicado. Debido a queel ADN pasa por un gel que retrasa su movimiento,los fragmentos pequeños de ADN son los que mi-gran más rápido. Los fragmentos específicos de ADNpueden identificarse entonces por la técnica secan-te Southern. En este procedimiento, el ADN setransfiere por acción capilar, del gel al filtro de lamembrana al cual se adhiere irreversiblemente. Elfiltro pasa a incubarse con una sonda de ADN mar-cada radioactivamente desde una localización cro-mosómica conocida que hibridará sólo aquellas he-bras de ADN en el filtro que contengan secuenciascomplementarias a la sonda. El secante se exponeentonces a una película de rayos-X, que revela la lo-calización de los fragmentos de ADN marcados ra-dioactivamente. La ganancia o pérdida de la zona derestricción cambia los patrones migratorios porquecambia la longitud de los fragmentos de ADN (esdecir, los RFLP) detectados por la sonda radioactiva(Figura 1-19). Con la hibridación sistemática utili-zando un panel de sondas que cubre el genoma hu-mano, puede demostrarse que un patrón de RFLPen concreto (o, alternativamente, una repeticióndinucleótida) cosegrega con la enfermedad en unpedigrí dado. Si la cadena de unión es fuerte, sepuede obtener información de la localización ge-neral del gen vulnerable a la enfermedad dentrodel genoma. El siguiente paso es aislar el gen y de-terminar su función.
El interés de esta técnica radica en que no re-quiere una identificación previa de la patofisiolo-gía del trastorno, sino que depende más bien de la
herencia de secuencias de bases alteradas sufi-cientemente próximas al gen, de modo que puedeconsiderarse muy probable que sean heredadasjunto con el gen. La secuencia en cuestión ha sidoutilizada con éxito para encontrar los genes quecausan la enfermedad de Parkinson y la fibrosisquística. Este método, sin embargo, supone unagran cantidad de trabajo. Puede emplearse una es-trategia alternativa para identificar los genes en-fermos si se dispone de una buena informaciónpatofisiológica que proporcione pistas sobre lasproteínas específicas que pueden ser potencial-mente anormales en la enfermedad estudiada. Losgenes que codifican estas proteínas pueden serclonados y utilizados como «genes candidatos dela enfermedad» . Los poliformismos que se iden-tifican en o cerca de estos genes, se utilizan pos-teriormente en un análisis de cadena. Este análi-sis no sólo es más eficaz, sino que, si el gencandidato resulta ser el gen enfermo, no hace faltamás trabajo para pasar del marcador de uniónhasta el gen enfermo. En el caso de la enfermedadde Huntington, el intento continuado de ir desdeel marcador fuertemente ligado al gen enfermoduró una década. Una vez se ha identificado elgen enfermo, su acción en el cerebro debe ser in-vestigada. Existe una gran variedad de sistemas,incluyendo la posibilidad de expresar el gen hu-mano en otras especies, más comunmente en ratas,o inactivar el gen implicado en un modelo conratas con una técnica llamada recombinación ho-m ó l o g a. Estos métodos suelen permitir un est u d i o
Figura 1-19. Técnica de los polimorfismos de la longitudde los fragmentos de restricción. El ADN aislado de loslinfocitos se divide mediante incubación con una endo-nucleasa de restricción específica. Los fragmentos se se-paran según su tamaño (por ejemplo, la longitud) median-te electroforesis en gel de agarosa. A continuación, estosfragmentos de ADN se transfieren a nitrocelulosa, a la quese fijan firmemente, y se incuban con una sonda de cADNmarcada radioactivamente. La sonda de cADN se fijarásólo al fragmento(s) al que sea complementaria. La sondade cADN libre se eliminará por lavado y la(s) banda(s) a lasque se haya fijado el cADN radioactivo se identificará(n)mediante autorradiografía.

2 8 TRATADO DE PSIQUIATRÍA
Figura 1-20. Regulación de la expresión del gen. Se requieren ciertas regiones de ADN, los facilitadores y los promoto-res designados, para la transcripción precisa de los genes y para determinar la velocidad a la que se expresarán los genes.Una función específica del gen promoter es la de determinar la zona precisa en la que empezará la transcripción por laARN polimerasa. Los promoters y los enhancers son secuencias de ADN que funcionan sirviendo zonas específicas deligando para las proteínas involucradas en la regulación de la transcripción (es decir, los factores de transcripción).
FACTORES DE TRANSCIPCION
A D N P R O M O T O RF A C I L I T A D O R SECUENCIAS TRANSCRITAS
A R NARN POLIMERASA
detallado de la acción de la proteína codificada porel gen, lo que proporciona pistas sobre la patofi-siología del trastorno humano.
Regulación de la fo s fo rilación de proteínas y ex p resión neural del gen porn e u r ot ra n s m i s o res y fá rmacos
Una de las propiedades más importantes del siste-ma nervioso es su plasticidad: puede adaptarse alos cambios en el ambiente y puede formar re-cuerdos. Hay descrito un buen ejemplo de adapta-ción en la explicación de la hidroxilasa tirosina, laenzima que controla la velocidad en la biosíntesisde catecolaminas. En circunstancias en que lasneuronas de noradrenalina o dopamina deben dis-parar a velocidades rápidas, se adaptan incremen-tando la actividad de tirosina hidroxilasa y, ade-más, producen más moléculas de esta enzima. Laprimera adaptación se debe a la fosforilación deproteínas y la segunda a la regulación de la expre-sión genética. Juntas constituyen los mecanismosmás significativos que regulan la adaptación a largoplazo y, probablemente, toda forma de memoria enel sistema nervioso. La regulación de la expresiónneural genética por neurotransmisores, hormonasy fármacos puede producir potencialmente altera-ciones de larga duración en prácticamente todoslos aspectos del funcionamiento de las enzimassintetizadoras de neurotransmisores, neurotrans-misores péptidos, canales iónicos, proteínas esti-muladoras de transducción y otras proteínas neu-rales críticas.
La fosforilación de proteína y la regulación dela expresión genética a menudo están relacionadas.En la mayoría de los casos, es la fosforilación de-pendiente de proteín-quinasas la que acopla estí-mulos ambientales a cambios en la expresión ge-nética neural. (El otro mecanismo importante que
regula la expresión genética neural se produce através de hormonas esteroides, como se describemás adelante). Aunque la expresión genética se re-gula a muchos niveles, el control de la iniciaciónde la transcripción parece ser el mecanismo másimportante que regula la información desde el ge-noma a la producción de proteínas celulares. La re-gulación del inicio de la transcripción incluye dosprocesos críticos: 1) la colocación de la p o l i m e r a-s a, la enzima que transcribe el ADN en ARN, enla zona correcta de partida de los genes, y 2) el con-trol de la eficacia de las iniciaciones para producirla velocidad transcripcional apropiada. Estas fun-ciones de control son favorecidas por extensionescortas de ADN (elementos cis-regulatorios) dentrode los genes (Figura 1-20) que actúan como lugaresde fijación específicos para las proteínas que regu-lan la transcripción, generalmente llamados f a c-tores de transcripción. Los análisis de mutacioneshan demostrado que cada gen tiene una combina-ción particular de elementos cis-regulatorios. Lanaturaleza, el número y el arreglo espacial de loselementos cis-regulatorios determinan el patrón deexpresión único de cada gen, incluido los tipos deneuronas donde se expresarán, los niveles basalesen los que se expresarán y la respuesta a los estí-mulos del ambiente (Mitchell y Tijan, 1989).
Como se ha descrito en una sección anterior,la estimulación de los receptores ligados a pro-teínas G activa o inhibe los sistemas de segundomensajero y, en consecuencia, la proteín-quina-sa. Cuando son activadas, ciertas proteín-quina-sas no sólo actúan en el citoplasma neuronal sinoque se trasladan al núcleo de la neurona dondepueden fosforilar factores de transcripción. Estosfactores de transcripción que se activan (o de-sactivan) por la fosforilación pueden unir la esti-mulación de receptores neurotransmisores a cam-bios en la expresión genética.

NEUROCIENCIA Y PSIQUIATRÍA 2 9
Aquellos elementos cis-regulatorios que liganlos factores de transcripción que son regulados fi-siológicamente (es decir, por fosforilación) y que,por tanto, otorgan una capacidad de respuesta a unneurotransmisor, una hormona o un segundo men-sajero en los genes, a menudo se llaman «e l e m e n-tos de respuesta» . Quizás el ejemplo mejor caracte-rizado de la respuesta de un elemento de un segundomensajero es el que confiere la activación por AMPcíclico (y por tanto la proteín-quinasa dependientedel cíclico) en aquellos genes que la poseen (Combet al., 1987). El descubrimiento y análisis de los ele-mentos que responden al AMP cíclico (CRE) de mu-chos genes, depende de la habilidad de mutar las se-cuencias de ADN in vitro, y reintroducirlasposteriormente en neuronas en cultivo (un procesoque se llama corte transversal). Pueden observarseentonces los efectos de las mutaciones en las se-cuencias de ADN respecto a la habilidad del AMPcíclico para activar el gen. Utilizando tales méto-dos se descubrió que los CRE contienen la secuen-cia CGTCA de ADN o secuencias relacionadas, yque esta secuencia de nucleótidos puede ligar unaproteína llamada CREB (proteína ligadora CRE).Cuando se une a un CRE, la CREB activa la trans-cripción al ser fosforila da por una proteín-quinasadependiente del AMP cíclico. Muchos elementosde respuesta adicionales y factores de transcripciónhan sido caracterizados.
Re c e pto res de la hormona esteroide
Una familia importante de elementos de respues-ta del ADN que no incluyen necesariamente la fos-forilación son los elementos de respuesta de losgluocorticoides (GRE) y otros elementos de res-puesta de hormonas esteroideas. A diferencia deotros neurotransmisores u hormonas péptidas, quese unen a receptores en la superficie de las neuro-nas, los glucocorticoides y otras hormonas este-roides son hiposolubles y entran directamente enlas neuronas. Actúan uniéndose a receptores espe-cíficos dentro del citoplasma de la neurona. Los re-ceptores esteroideos citoplasmáticos incluyen re-ceptores glucocorticoides, receptores estrogénicos,receptores mineralocorticoideos y otros parecidos.Debido a que las hormonas esteroideas se expan-den ampliamente, la especificidad de la respuestadepende de la presencia o ausencia de receptoresespecíficos dentro de cada tipo de neurona. Cuan-do son activados por fijación hormonal, los recep-tores esteroideos pasan al núcleo, donde se unen aGRE (u otros elementos de respuesta a las hormo-nas esteroideas) contenidas en genes concretos. Launión del receptor al ADN aumenta o disminuyela velocidad con que estos genes diana se transcri-ben. Por tanto, los receptores de hormonas este-
roideas actúan como factores de transcripción sen-sibles a hormonas. La mayoría de los efectos co-nocidos de los glucocorticoides, los esteroides go-nadales, la hormona tiroidea y la vitamina D en lafunción neuronal, son mediados por sus accionesen la expresión genética.
@ P S I C O FA R M AC O LOGÍA MOLECULAR
El descubrimiento de medicamentos que reducende forma selectiva los síntomas de diversas enfer-medades psíquicas ha proporcionado una bateríaproductiva de pruebas farmacológicas para estudiarlas funciones potenciales de los sistemas neuralesespecíficos en relación con la patofisiología de lostrastornos psiquiátricos. Naturalmente, no puedeasumirse de forma directa que el lugar de acciónmolecular o celular de un medicamento psicotró-pico localice el defecto patofisiológico responsablede la enfermedad. Es posible que el sistema neu-ronal afectado por el fármaco esté implicado sólode forma secundaria, y que la alteración farmaco-lógicamente inducida de su función compense unaalteración básica en otros lugares del sistema ner-vioso. Sin embargo, nuestra capacidad para identi-ficar los puntos diana moleculares y los sistemasneurales en donde actúan los fármacos psicotrópi-cos, ha potenciado el desarrollo de teorías patofi-siológicas importantes y de descubrimientos fun-damentales en el campo de la neurociencia básica.En este apartado se revisan los conocimientos ac-tuales de los mecanismos de acción de tres gran-des grupos de fármacos psicotrópicos, haciendoparticular hincapié en los métodos de investiga-ción utilizados para aclarar sus funciones.
Ne u r o l é pticos
El descubrimiento de que la reserpina (un alcaloi-de de la rauwolfia) y la fenotiacina-clorpromazina,reducen drásticamente la agitación, las alucina-ciones y los delirios en las psicosis esquizofréni-cas agudas, abrió la moderna era de la psicofarma-cología hace 35 años. En la década siguiente, laindustria farmacéutica sintetizó un gran númerode fármacos con posibles efectos antipsicóticos.Aunque muchos de estos fármacos eran variacio-nes estructurales de la clorpromazina, se desarro-llaron también estructuras nuevas, tales como elgrupo de la butiroferona, con el haloperidol comoejemplo. Puesto que se desconocía la causa de laesquizofrenia, y que el mecanismo de acción deestos medicamentos era poco claro, el uso clínicose basaba en gran medida en la reducción empíri-ca de los síntomas evitando en lo posible los efec-tos secundarios.

3 0 TRATADO DE PSIQUIATRÍA
A principio de los años sesenta, la US VeteransA d m i n i s t r a t i o n y el National Institute of MentalH e a l t h emprendieron un ambicioso estudio parademostrar de manera inequívoca la eficacia clíni-ca de los medicamentos antipsicóticos. Para pre-venir las apreciaciones subjetivas, se utilizaron es-tudios con la técnica de doble ciego y el uso deplacebos, considerada hoy en día como la «regla deo r o» a la que se someten la mayoría de los fárma-cos nuevos (Kurland et al., 1961). Las drogas pre-suntamente activas se comparaban con el placebopara determinar si la droga era más efectiva queuna sustancia inerte, un objetivo de gran interéspara estudios relacionados con enfermedades cuyossíntomas aumentan y disminuyen a lo largo deltiempo. Para evitar que la propia situación de estaren tratamiento influyera positivamente sobre lossíntomas, no se informó a los pacientes acerca desi recibían una droga activa o un placebo. Para eli-minar factores conscientes o inconscientes que pu-dieran afectar la respuesta al tratamiento, tanto losmédicos como los evaluadores tampoco fueron in-formados acerca de si el paciente estaba siendo tra-tado con una sustancia activa o con un placebo.
Estos estudios generaron una gran cantidad deinformación que fue esencial para comprender elmecanismo de acción de los fármacos antipsicóti-cos. En primer lugar, establecieron que la sedaciónpor sí misma no explica la eficacia terapéutica, yaque el fenobarbital fue ineficaz. En segundo lugar,revelaron que la presencia de la estructura de la fe-notiazina no era suficiente ya que la prometazinaera relativamente ineficaz comparada con la clor-promazina. En tercer lugar, estos y otros estudiosposteriores que comparaban otros neurolépticoscon la clorpromazina, pusieron de manifiesto quetodos los antipsicóticos eran igualmente eficacesdesde el punto de vista cualitativo, aunque habíauna diferencia de casi 50 veces referida a su po-tencial clínico. Por tanto, dichos estudios genera-ron una herramienta decisiva, la relación entre es-tructura y actividad de los fármacos antipsicóticos,que podía ser utilizada para investigar los meca-nismos de acción del fármaco y diseñar nuevoscompuestos activos.
A principio de los años sesenta, varias obser-vaciones dispares implicaban a las neuronas dopa-minérgicas del cerebro anterior en el mecanismode acción de los neurolépticos. Hornyekiewicz(1966) acababa de demostrar que la enfemedad deParkinson se asociaba con una gran disminuciónde dopamina en la sustancia negra y el caudado-putamen. Se halló que la reserpina, un antipsicó-tico que no tenía relación estructural con las fe-notiazinas, causaba una marcada depleción deaminas biógenas, incluida la dopamina, en el ce-rebro de los animales de experimentación. Por úl-
timo, los efectos colaterales de tipo neurológicomás comúnmente observados en la administraciónde fármacos antipsicóticos eficaces, eran síntomasde tipo parkinsoniano.
En estudios realizados con clorpromazina y ha-loperidol, Carlsson observó que, mientras que estosantipsicóticos no mermaban la dopamina del ce-rebro, como hacía la reserpina, producían un in-cremento sustancial en el flujo de dopamina. En-lazando ambas evidencias, Carlsson fue el primeroen proponer que los fármacos antipsicóticos pue-den ejercer sus efectos terapéuticos bloqueando losreceptores cerebrales de la dopamina (Carlsson yLindqvist, 1963). Esta hipótesis recibió apoyo, aun-que algo indirecto, a través de los estudios psico-farmacológicos del comportamiento. De estemodo, fenómenos tales como la estereotipia y elvómito, que son inducidos en animales de experi-mentación por fármacos que aumentan directa oindirectamente la neurotransmisión dopaminérgi-ca central, pueden prevenirse con la administra-ción de neurolépticos.
Las especulaciones sobre las interacciones entrelos fármacos antipsicóticos y los receptores dopa-minérgicos quedaron a la espera de una validacióndirecta hasta que, en los años setenta, se desarro-llaron métodos para caracterizar bioquímicamen-te los receptores de los neurotransmisores. Con elprecedente de que los beta-receptores están ligadosa la adenil ciclasa, Kebabian et al. (1972) demos-traron que la dopamina estimulaba la formaciónde AMP cíclico en homogeneizados preparados apartir del caudado-putamen, un área cerebral ricaen inervación dopaminérgica, pero no noradrenér-gica. En estos homogeneizados, el propanolol, an-tagonista de los beta-receptores, mostraba unosefectos débiles bloqueando la formación de AMPcíclico, mientras que los neurolépticos del grupode las fenotiazinas eran potentes bloqueadores dela actividad de la adenilciclasa. Además, debido aque la potencia en este ensayo era proporcional ala eficacia clínica como antipsicóticos, se conclu-yó que las fenotiazinas actuaban sobre un receptordopaminérgico. Sin embargo, la conclusión de queeste receptor de la dopamina mediaba los efectosantipsicóticos de los neurolépticos, quedó socava-da por la observación de que potentes neurolépti-cos del grupo de las butiroferonas, tales como elhaloperidol, eran desproporcionadamente débilescomo antagonistas.
El uso de haloperidol radioactivo como ligandopara identificar los receptores de la dopamina portécnicas de fijación del ligando, reveló la existen-cia de un receptor claramente distinto. El 3H ha-loperidol se unía con una gran afinidad a una seriede zonas de reconocimiento que eran bastanteabundantes en regiones cerebrales que recibían

NEUROCIENCIA Y PSIQUIATRÍA 3 1
inervación dopaminérgica. A pesar de que la pro-pia dopamina tenía una afinidad 1.000 veces menorpor esa zona que el 3H haloperidol, la dopaminaera el neurotransmisor más activo en dicha zona.Es más, cuando se examinó el espectro total deneurolépticos clínicamente eficaces, se observóuna correlación más que notable entre su potenciaclínica como antipsicóticos y su afinidad por estazona de reconocimiento del receptor, indepen-dientemente de la clase de fármaco (Creese et al.,1976). Actualmente se sabe que estos dos tipos dereceptores son estructuralmente distintos: el pri-mero, ligado a la adenil ciclasa, se ha designadocomo receptor D1, y el segundo, que constituye ellugar de acción de los neurolépticos, se conocecomo receptor D2. La acción de las fenotiazinassobre los receptores D1 refleja la falta de especifi-cidad, más que una acción terapéutica crítica. Sub-secuentemente se demostró que el receptor D2 i n-hibe la adenilciclasa y activa un canal de K+ através de Gi. Como se ha descrito en una secciónanterior, sin embargo, estudios recientes de clo-nación molecular han identificado al menos tresreceptores dopaminérgicos adicionales. Estos es-tudios se plantean si el antagonismo D2 es nece-sario o tan siquiera suficiente para la acción delfármaco antipsicótico. Esto es debido a que el fár-maco antipsicótico atípico clozapina, que puedetener un efecto singular en la esquizofrenia peroestá relativamente libre de efectos extrapiramida-les, se une con baja afinidad a los receptores D2,pero con una alta afinidad a los recientemente des-cubiertos receptores D4. Resulta que casi todos losfármacos que interactúan con los receptores D2
también interactúan con los receptores D3 y D4. Laafinidad relativamente baja de la clozapina al re-ceptor D2 explica la falta de efectos secundarios ex-trapiramidales, y quizás la singular eficacia en eltratamiento de síntomas «n e g a t i v o s» de la esqui-zofrenia, tales como el repliegue emocional, quede hecho pueden estar causados por el antagonis-mo del receptor D2. La importancia relativa de blo-quear D2, D3, D4, o incluso los receptores dopami-nérgicos todavía desconocidos para la acciónantipsicótica es, en estos momentos, objeto de unainvestigación intensiva.
Existe una observación adicional sobre la ac-ción de fármacos antipsicóticos que es de impor-tancia crítica para entender el mecanismo. Todoslos fármacos antipsicóticos requieren unas sema-nas de administración antes de conseguir su má-ximo efecto terapéutico. Los pacientes que tomanclozapina pueden seguir mejorando incluso du-rante meses, lo que implica que el bloqueo de re-ceptores dopaminérgicos (el tipo resulta ser muyimportante) representa la interacción inicial de losfármacos antipsicóticos con el sistema nervioso.
Sin embargo, todo esto forma parte de la todavíadesconocida respuesta adaptativa de aparición lentadel sistema nervioso al bloqueo del receptor dopa-minérgico, lo que representa el actual mecanismopor el cual los síntomas psicóticos se alivian. Losmecanismos más probablemente involucrados secentran en la posibilidad de que el bloqueo de re-ceptores dopaminérgicos conduzca a cambios sig-nificativos en las proteínas (es decir, los canales ió-nicos, los receptores, las enzimas) contenidas porneuronas diana (que pueden ser la neurona dopa-minoceptiva misma u otras neuronas alejadas poruna o dos sinapsis). Se postula que tales cambiosconducen a funcionamientos alterados del circuitolímbico crítico. Esta aparición lenta y los cambiosde larga duración en el funcionamiento neuronalprobablemente incluyan cambios mediados por se-gundos mensajeros en la expresión del gen.
A n t i d e p re s i vos
Las primeras pistas sobre el mecanismo de acciónde los fármacos antidepresivos fueron resultado di-recto de los estudios fundamentales del Dr. JuliusAxelrod en el National Institute of Mental Health.En un experimento destinado a controlar el cata-bolismo de la noradrenalina radioactiva en vivo,Axelrod se dio cuenta de que una pequeña canti-dad de la noradrenalina administrada por vía sis-témica era retenida por los tejidos periféricos, sinser metabolizada (Axelrod et al., 1959). La canti-dad de noradrenalina radioactiva secuestrada porestos tejidos era proporcional al grado de inerva-ción simpática. En estudios posteriores se demos-tró que las neuronas noradrenérgicas tenían unp r oceso de transporte de alta afinidad para la no-radrenalina y que los antidepresivos tricíclicoseran potentes inhibidores de este proceso de trans-porte. Se observó que las neuronas centrales nora-drenérgicas, las dopaminérgicas y las serotoninér-gicas tenían proteínas transportadoras específicaspara sus neurotransmisores; estos transportadoresactualmente clonados, son el mecanismo princi-pal para que el neurotransmisor liberado en el es-pacio sináptico deje de ejercer su acción.
Estudios detallados desarrollados a lo largo delos años han puesto de manifiesto que los antide-presivos tricíclicos y los antidepresivos heterocí-clicos, introducidos más recientemente, son anta-gonistas poderosos de los transportadores de lanoradrenalina y/o la serotonina, y que de esta ma-nera potencian la acción de estos dos neurotrans-misores en el espacio sináptico. Más recientemen-te, se han introducido inhibidores selectivos de laserotonina como la fluoxetina y sertralina. Los es-tudios sobre los antidepresivos tricíclicos han con-firmado la importante distinción entre la i d e n t i f i-

3 2 TRATADO DE PSIQUIATRÍA
cación del lugar de acción de un fármaco y la com-prensión del mecanismo de su acción terapéutica.Mientras que la inhibición de la captación, y portanto, de la potenciación de la neurotransmisiónnoradrenérgica y serotoninérgica, debería ser unaconsecuencia bastante inmediata de la adminis-tración de antidepresivos, existe una considerableevidencia clínica de que se produce un retraso sus-tancial de dos o tres semanas antes de que aparez-can mejoras sintomáticas en una depresión mayor.La demora evoca la acción de los fármacos antip-sicóticos mencionada anteriormente.
La demora en la aparición de efectos clínicosllevó a los científicos a investigar sobre los efectosde los antidepresivos que sólo aparecían bajo unaadministración crónica de fármacos. El primer efec-to con inicio demorado que se observó fue una de-sensibilización de los receptores beta-adrenérgicosen el córtex cerebral en ratas. Es interesante elhecho que esta desensibilización acompaña prácti-camente todos los tratamientos eficaces de las de-presiones mayores, incluyendo los antidepresivostricíclicos, los inhibidores de la monoaminooxida-sa, diversos antidepresivos atípicos, y el tratamien-to electroconvulsivo. Subsecuentemente, se de-mostró que la destrucción selectiva de neuronasserotoninérgicas del cerebro causa una desensibili-zación de beta-receptores inducido por antidepresi-vos, lo que demuestra una unión funcional entre lossistemas serotoninérgicos y noradrenérgicos (Ja-nowsky et al., 1982). Subsecuentemente, se ha de-mostrado que algunos tratamientos antidepresivos,aunque no todos, desensibilizan los receptores adre-nérgicos alfa-2 y los receptores serotoninérgicos 5-H T2 (ver subsección anterior sobre neuronas sero-toninérgicas para un repaso de los receptoresserotoninérgicos).
A medida que se han acumulado pruebas, pare-ce posible que estas alteraciones en la sensibilidaddel receptor sean marcadores de los efectos antide-presivos crónicos, aunque probablemente no repre-senten el mecanismo de acción terapéutica. Los ha-llazgos sobre receptores, sin embargo, sugierenposibilidades para la investigación. Por ejemplo, sesabe ahora que la desensibilización del receptorbeta-adrenérgico se debe a los incrementos de acti-vidad del AMP cíclico dependiente de la proteín-quinasa y otras quinasas con alta especificidad parael beta-receptor mismo. La activación de estas qui-nasas es probablemente el resultado de una esti-mulación noradrenérgica aumentada de los beta-re-ceptores mismos, a consecuencia de la acción inicialde los antidepresivos (por ejemplo, el bloqueo de larecaptación o inhibición de la MAO). Por tanto, ladesensibilización de los beta-receptores es un mar-cador neuronal que muestra que la administracióncrónica de antidepresivos lleva a un aumento
de activación de la proteín-quinasa en las neuro-nas inervadas noradrenérgicamente. Ahora pode-mos investigar otras funciones de dichas quinasas,incluyendo factores de transcripción que puedenalterar la expresión genética neural. Tambiénhacen falta estudios complementarios a nivel delos sistemas, como por ejemplo análisis del estadode receptores de monoamina en pacientes depri-midos que pueden abordarse con técnicas de TEPo quizá con imágenes por resonancia magnética.Juntos, estos métodos prometen grandes posibili-dades para conocer aspectos de la acción de los an-tidepresivos en la próxima década.
Litio
El mecanismo preciso de la acción del litio en eltratamiento de la enfermedad maníaco-depresivaes desconocido, pero existen pistas científicas in-teresantes. En concreto, parece ser que el litio esel único psicofármaco que actúa directamentesobre las proteínas G y los sistemas de segundomensajero. Muchos receptores neurotransmisores,incluyendo los receptores adrenérgicos y los 5-HT2,están ligados a través de proteínas G (mayoritaria-mente Gq) a la activación de la fosfolipasa C, unaenzima que hidroliza un fosfolípido de la mem-brana, el fosfatidilinositol bisfosfato (PIP2) para pro-ducir dos segundos mensajeros, el diaglicerol y elinositol trifosfato (IP3). Como muestra la Figura 1-10, el litio inhibe ciertos pasos en el ciclo fosfati-dilinositol, y algunos autores han planteado la hi-pótesis de que estas acciones son responsables delos efectos antimaníacos y antidepresivos del litio.
El fosfatidilinositol bifosfato se sintetiza a par-tir de inositol libre y de grupos de lípidos para sersubsecuentemente fosforilatado. La mayoría de lascélulas obtienen inositol libre para la síntesis di-rectamente del plasma, pero las neuronas no pue-den porque el inositol no cruza la barrera de sangredel cerebro. Como resultado, las neuronas debenreciclar inositol o sintetizarlo de nuevo de la glu-cosa-6-fosfato, un producto de la glicolisis. Comomuestra la Figura 1-10, tanto el reciclaje como lasíntesis requieren desfosforilación de fosfatos deinositol. Sin embargo, utilizado en concentracio-nes terapéuticas, el litio inhibe ciertas fosfatasas.Por tanto, las neuronas expuestas al litio tienen unacapacidad disminuida de regenerar PIP2 después deque haya sido hidrolizado como respuesta a la ac-tivación del receptor neurotransmisor (Berriche etal., 1989). Se ha propuesto que cuando la velocidadde disparo de las neuronas es excesivamente alta,las neuronas tratadas con litio se vacían de PIP2
más rápidamente, y la neurotransmisión depen-diente de este sistema de segundo mensajero sefrena. Esta hipótesis de la reducción de inositol es

NEUROCIENCIA Y PSIQUIATRÍA 3 3
atractiva porque los efectos del litio pueden evi-denciarse sólo en las neuronas con velocidadesanormalmente altas, y porque el litio frena losefectos de múltiples sistemas de neurotransmiso-res, por lo que podría tratar tanto los estados ma-níacos como los depresivos. Sin embargo, aunqueestá hipótesis sea correcta permanece incompleta.Las neuronas críticas que son el blanco de la ac-ción terapéutica del litio siguen sin conocerse, yno está claro qué sistemas de neurotransmisoresdependientes de fosfatodilinositol deben ser fre-nados para que el litio tenga efectos terapéuticos.
Además de los efectos en el ciclo del fosfatidi-linositol, el litio altera el acoplamiento de algunosreceptores neurotransmisores a las proteínas G, al-terando así la función de múltiples vías de trans-ducción de neurotransmisores estimuladores en elcerebro (Avissar et al., 1988). A diferencia de la hi-pótesis de reducción de inositol, sin embargo, nohay actualmente una teoría que explique de quéforma los efectos del litio sobre las proteínas G pro-ducirá a efectos específicos en los estados manía-co y depresivo.
El litio también inhibe la adenilciclasa. Sin em-bargo, las concentraciones que se requieren paraejercer este efecto en el cerebro se encuentran a ni-veles más altos de los conseguidos clínicamente.
Ansiolíticos
A pesar de que la benzodiazepina prototípica, elclordiazepóxido, fue descubierta por azar, prontose observaron las notables propiedades de ésta yotras benzodiazepinas sintetizadas más tarde, y susuperioridad frente a los barbitúricos, utilizadoscomúnmente como sedantes y ansiolíticos. Lasbenzodiazepinas muestran una ratio mucho másfavorable, un mayor índice terapéutico y menosriesgo de dependencia y síntomas serios de absti-nencia comparadas con los barbitúricos.
La comprensión de las zonas moleculares de ac-ción de las benzodiazepinas, así como de los barbi-túricos, dependió de la elucidación de los meca-nismos fisiológicos y de los receptores quemediaban los efectos del GABA, el principal neu-rotransmisor inhibidor del cerebro. La aplicaciónlocal del GABA en neuronas concretas produce unainhibición de la descarga, causada por la hiperpo-larización que sigue a la apertura de los canales decloro en la membrana neuronal, hiperpolarizandoasí la neurona. Utilizando técnicas de fijación de li-gando con GABA radioactivo, se detectó un recep-tor de GABA entre las membranas cerebrales. Ade-más, utilizando diazepam radioactivo fue posiblemarcar directamente las zonas de reconocimientopara las benzodiazepinas. Después de inv e s t i g a c i o-nes adicionales, se estableció que el receptor de las
benzodiazepinas representaba una zona de uniónen el receptor GABAA, una gran proteína con múl-tiples subunidades que sirve como el principal re-ceptor del GABA en el sistema central nervioso(Levitan et al., 1988). El receptor GABAA está for-mado por múltiples subunidades, que han sido de-nominadas (basándose en estudios de clonación)alfa, beta, gama y delta. Los receptores GABAA s o nprobablemente pentámeros compuestos de dos uni-dades alfa, dos beta y una gama o delta. Las subu-nidades del receptor GABAA muestran un extraor-dinario grado de heterogeneidad; hasta la fecha, almenos siete subunidades alfa diferentes, cuatro su-bunidades beta diferentes y dos subunidades gamahan sido clonadas. Las diferentes subunidadesestán expresadas diferencialmente en varias regio-nes del sistema nervioso central y muestran dife-rentes propiedades funcionales.
El receptor GABAA contiene por lo menos treszonas de unión relevantes para la psicofarmacolo-gía. La zona de unión para el GABA está en la su-bunidad beta del receptor; las benzodiazepinas sejuntan con la subunidad alfa. Sin embargo, la subu-nidad alfa no es capaz de unirse a las benzodiazepi-nas a menos que una subunidad gama esté presen-te en el complejo, presuntamente, por la regulaciónalostérica de la subunidad alfa por la subunidadgama (Pritchett et al., 1989). Las benzodiazepinasno abren directamente el canal del receptor CL–.Más bien actúan aumentando la afinidad de la zonade unión de GABA en la subunidad beta de GABA,y por tanto realzan la acción sináptica de GABA.
Los barbitúricos también se unen al receptorG A B AA, pero en una zona que es físicamente dis-tinta de la de las benzodiazepinas, y por tantoambos fármacos pueden unirse al receptor a la vez.Los barbitúricos pueden ejercer una influencia enel receptor similar a la de las benzodiazepinas, au-mentando la afinidad del receptor para GABA y portanto aumentando la habilidad del GABA para ac-tivar el canal receptor de CL–. Sin embargo, a dife-rencia de las benzodiazepinas, altas dosis de bar-bitúricos pueden causar la apertura del canal CL–
directamente, en ausencia de GABA. Esto puedeexplicar por qué los barbitúricos pueden causar unadepresión más seria en el sistema nervioso central(con más probabilidad de ser letal) que una sobre-dosis de benzodiazepinas. Además de aumentar laafinidad del receptor GABAA para el GABA, lasbenzodiazepinas y los barbitúricos aumentan la afi-nidad del receptor mútuamente.
El alcochol etílico produce efectos en el siste-ma nervioso central a concentraciones de sangremucho más altas que cualquier otra sustancia psi-cotrópica ampliamente utilizada. En concentra-ciones de alcochol asociadas con intoxicación, lasalteraciones en las propiedades de la membrana

3 4 TRATADO DE PSIQUIATRÍA
neuronal probablemente afectarán el funciona-miento de una amplia gama de proteínas de lamembrana como receptores, canales y transporta-dores. Sin embargo, existen pruebas cada vez ma-yores de que un pequeño número de neurotrans-misores receptores y quizás de canales iónicos queson particularmente susceptibles a los cambios in-ducidos por el etanol en la membrana, son res-ponsables de los efectos conductuales del etanol.En concreto, en concentraciones farmacológicasrelevantes, el etanol inhibe la acción de un tipo dereceptor (es decir, el receptor NMDA) que hace demediador de los efectos importantes sobre el neu-rotransmisor excitador principal del cerebro, el glu-tamato. Todavía más pronunciados son los efectosfacilitadores del etanol sobre el receptor GABAA.En concentraciones bajas, el etanol altera el fun-cionamiento del receptor GABA tanto como lasbenzodiazepinas y los barbitúricos: el receptortiene una mayor afinidad para el GABA. Como losbarbitúricos, el etanol en grandes concentracionespuede causar la apertura del canal Cl– i n d e p e n-dientemente del GABA. Además, el etanol au-menta la aparente afinidad del receptor tanto paralas benzodiazepinas como para los barbitúricos.
Nuestra creciente comprensión de la farmaco-logía de este receptor racionaliza un número de ob-servaciones clínicas. La similitud de las accionesentre los fármacos con baja dosis de etanol, de losansiolíticos y los sedantes es consistente con el usode etanol que hacen muchos individuos para ali-viar la ansiedad, para sobreponerse a las inhibicio-ne sociales y para conseguir dormir. (Este uso esmal interpretado ya que, a medida que las concen-traciones de alcohol en la sangre caen, frecuente-mente tienen lugar síntomas de rebote reflectivosde tolerancia a la dosis. Por tanto, la ansiedad puedevolverse mucho peor que antes de haber tomado al-cohol y el sueño puede interrumpirse). La zona deacción común entre el etanol, las benzodiazepinasy los barbitúricos también explica por qué estos fár-macos producen tolerancia y dependencia cruza-das, características explotadas en el uso de benzo-diazepinas para la desintoxicación de etanol.También explica por qué estos fármacos potenciansus efectos mutuamente, haciendo que la sobredo-sis de una combinación de ellos sea tan arriesgada.
@ C O NC LUSIONES
La psiquiatría, como especialidad médica dedica-da principalmente al diagnóstico y el tratamientode los trastornos conductuales, debe incorporar ne-cesariamente la neurociencia en sus fundamentoscientíficos. Basándose en el creciemiento casi lo-garítmico de la investigación neurocientífica en la
última década, los progresos en el conocimiento dela estructura, de la organización y del funciona-miento del cerebro prometen ofrecer poderosos mé-todos nuevos para el diagnóstico de enfermedadespsiquiátricas, aclarando su fisiopatología y desa-rrollando terapéuticas más específicas y eficaces.
Parecen infundados los temores de que estosavances, que se basan en aproximaciones cada vezmás reduccionistas, pueden minar la tradición hu-manística de la psiquiatría y nieguen la importanciade la relación entre el paciente y el médico. En pri-mer lugar, incluso cuando sean factibles las técnicasdiagnósticas de base genética para diagnosticar al-gunas de las enfermedades mentales hereditarias másimportantes, el método clínico usado actualmentepara el desarrollo de diagnósticos provisionales to-davía será necesario para determinar qué personasdeben ser sometidas a pruebas. En segundo lugar, eldiagnóstico no puede hacerse ni aceptarse de mane-ra mecánica; elaborar un diagnóstico definitivo re-quiere un tratamiento psicológico humano y conti-nuado. En tercer lugar, la investigación clínica sobrelos tratamientos psicofarmacológicos de ciertas en-fermedades psiquiátricas está demostrando que lafarmacoterapia sola es insuficiente para el trata-miento completo y eficaz de los pacientes. Se va ha-ciendo cada vez más evidente que los métodos es-pecíficos conductuales, psicológicos y psicosocialesdeben asociarse a la farmacoterapia para conseguirresultados óptimos en el tratamiento de enfermeda-des psiquiátricas. En cuarto lugar, los métodos paraestablecer de forma rigurosa la eficacia de las tera-pias somáticas se aplicarán a las terapias conduc-tuales y psicológicas a fin de determinar las inter-venciones más eficaces en cada enfermedad enparticular. Así pues, los médicos podrán usar los tra-tamientos, sean somáticos o psicológicos, con unaconfianza creciente en su especificidad y eficacia.
Aunque los progresos en neurociencia y en bio-logía molecular pueden contemplarse acertada-mente como causantes de un cambio paradigmáti-co en la psiquiatría, hay que mirar más allá paraconsiderar la siguiente serie de cuestiones queestos conocimientos traerán consigo. Por lo que serefiere al cerebro, un órgano sensible de maneraúnica a la experiencia vital, es probable que se des-pierte de nuevo el intenso interés sobre los acon-tecimientos vitales durante el desarrollo, ya queéstos ejercen una influencia crítica en la expresiónfenotípica de los genes y en último término, afec-tan a los sistemas neuronales cerebrales relacio-nados con los impulsos, los afectos y las funcionescognitivas. Es más, será posible someter este tipode cuestiones a riguroso estudio. El desvelamien-to de los mecanismos moleculares implicados enla psicopatología originará una infinidad de cues-

NEUROCIENCIA Y PSIQUIATRÍA 3 5
tiones acerca de los mecanismos de protección quemodifican o previenen la expresión de enfermeda-des psiquiátricas en personas genéticamente vul-nerables. Por consiguiente, la profundización en lafisiopatología de la enfermedad mental y en la dis-paridad entre las características fenotípicas y ge-notípicas conducirá en última instancia a una mejorcomprensión de los factores que determinan tantoe l «bienestar mental» como la enfermedad mental.
@ B I B L I O G R A F Í A
Avissar S, Schreiber G, Danon A, et al: Lithium inhibitsadrenergic and cholinergic increases in GTP bindingin rat cortex. Nature 331:440–442,1988
Axelrod J, Weil-Malherbe H, Tomchick R: The physiolog-ical disposition of 3H-epinephrine and its metabolite,metanephrine. J Pharmacol Exp Ther 127:251–256, 1959
Berridge MJ, Downes CP, Hanley MR: Neural and devel-opmental actions of lithium: a unifying hypothesis.Cell 59:411–419, 1989
Botstein D, Whik R, Skolnick M, et al: Construction of agenetic linkage map in man using RFLPs. Am J HumGenet 32:314–331, 1980
Carlsson A, Lindqvist M: Effect of chlorpromazine andhaloperidol on formation of 3-methoxytyramine andnormetanephrine in mouse brain. Acta PharmacolToxicol 20:140–144, 1963
Choi DW, Rothman SM: The role of glutamate neurotox-icity in hypoxic-ischemic neuronal death. Annu RevNeurosci 13:171–182, 1990
Comb M, Hyman SE, Goodman HM: Mechanisms oftrans-synaptic regulation of gene expression. TrendsNeurosci 10:473–478, 1987
Cooper JR, Bloom FE, Roth RH: The Biochemical Basis ofNeuropharmacology, 6th Edition. New York, OxfordUniversity Press, 1991
Coyle JT: Aminergic projections from the reticular core,in Diseases of the Nervous System. Edited by AsburyA, McKhann G, McDonald W. Philadelphia, PA, WBSaunders, 1986, pp 880–889
Coyle JT, Price DL, DeLong MR: Alzheimer's disease: adisorder of cholinergic innervation of cortex. Science219:1184–1190, 1983
Creese I, Burt DR, Snyder SH: Dopamine receptor bind-ing predicts clinical and pharmacological potencies ofantischizophrenic drugs. Science 192:481– 483, 1976
Giros B, Caron MG: Molecular characterization of thedopamine transporter. Trends Pharmacol Sci 14:43–49, 1993
Hokfelt T: Neuropeptides in perspective: the last tenyears. Neuron 7:867–879, 1991
Hornykiewicz O: Dopamine and brain function. Phar-macol Rev 18:925–964, 1966
Hughes JU, Smith TW, Kosterlitz HW: Identification oftwo related penta peptides from the brain with potentopiate agonist activity. Nature 258:577–579, 1975
Hyman SE, Nestler EJ: The Molecular Foundations ofPsychiatry. Washington, DC, American PsychiatricPress, 1993
Janowsky AJ, Okada F, Applegate C, et al: Role ofserotonergic input in the regulation of the beta-adrenoreceptor coupled adenylate cyclase system inbrain. Science 218:900–901, 1982
Kebabian JW, Petzold GL, Greengard P: Dopamine-sensi-tive adenylate cyclase in the caudate nucleus of the ratbrain and its similarity to the “dopamine receptor.”Proc Natl Acad Sci U S A 79:2145–2149,1972
Kobilka B: Adrenergic receptors as models for G protein–coupled receptors. Annu Rev Neurosci 15:87–114, 1 9 9 2
Kurland AA, Hanlon TE, Tatom MH, et al: The compara-tive effectiveness of six phenothiazine compounds,phenobarbital and inert placebo in the treatment ofacutely ill patients: global measures of severity of ill-ness. J Nerv Ment Dis 133:1–18, 1961
Levitan ES, Schofield PR, Burt DR, et al: Structural andfunctional basis for GABAA receptor heterogeneity.Nature 335:76–79, 1988
Levitt P, Rakic P, Goldman-Rakic P: Region-specific dis-tribution of catecholamine afferents in primate cere-bral cortex: a fluorescence histochemical analysis. J Comp Neurol 227:23–36, 1984
Martin D, Lodge D: Ketamine acts as a non-competitiveN-methyl-D-aspartate antagonist on frog spinal cordin vitro. Neuropharmacology 24:999–1006, 1985
Mitchell PJ, Tjian R: Transcriptional regulation in mam-malian cells by sequence-specific DNA binding pro-teins. Science 245:371–378, 1989
Nakanishi N, Shneider NA, Axel R: A family of gluta-mate receptor genes: evidence for the formation ofheteromultimeric receptors with distinct channelproperties. Neuron 5:569–581, 1990
Nose PS, Griffith LC, Schulman H: Ca2 +-dependent phos-phorylation of tyrosine hydroxylase in PC 12 cells. J Cell Biol 101:1182–1190, 1985
Olney JW: Brain lesions, obesity and other disturbancesin mice treated with monosodium glutamate. Science164:719–721, 1969
Pacholczyk T, Blakely RD, Amara SG: Expression cloningof a cocaine- and antidepressant-sensitive human nor-adrenaline transporter. Nature 350:350–354, 1991
Pritchett DB, Sontheimer H, Shivers B, et al: Importanceof a novel GABAA receptor subunit for benzodiaze-pine pharmacology. Nature 13:338:582–585, 1989
Robinson, Coyle JT: Glutamate and related acidic neuro-transmitters: from basic science to clinical practice.FASEB J 1:446–455 ,1988
Schwarz R, Meldrum B: Excitatory amino acid antago-nists provide a therapeutic approach to neurologicdisorders. Lancet 2:140–143, 1985
Simon MI, Strathmann MP, Gautam N: Diversity of Gproteins in signal transduction. Science 252:802–808,1991
Van Tol HMV, Bunzow JR, Guan HC, et al: Cloning ofthe gene for a human dopamine D4 receptor withhigh affinity for the antipsychotic clozapine. Nature350:610–614, 1991
Watson SJ, Kelsey JE, Lopez JF, et al: Neuropeptide biol-ogy: basic and clinical lessons from the opioids, in Psychiatry Update: American Psychiatric AssociationAnnual Review, Vol 4. Edited by Hales RE, F r a n c e s

3 6 TRATADO DE PSIQUIATRÍA
AJ. Washington, DC, American Psychiatric Press,1985, pp 83–96
Wexler NS, Rose EA, Housman DE: Molecular ap-proaches to hereditary diseases of the nervous system:Huntington's disease as a paradigm. Annu Rev Neu-
rosci 14:503–530, 1991Wong DF, Wagner Jr HN, Tune LE, et al: Positron emis-
sion tomography reveals elevated D2 dopamine re-ceptors in drug-naive schizophrenics. Science234:1558–1563, 1986