H. A. Simon (1996)atta.labb.usb.ve/Klaus/art217.pdfPor ejemplo, se han encontrado patrones...
19
177 Modelos y simulaciones biológicas: ecología y evolución Harold P. de Vladar y Roberto Cipriani. (eds.) 2015 Impreso por Createspace. ISBN-13: 978-1516867561 / ISBN-10: 1516867564 https://goo.gl/kVfvnu Propiedades emergentes, eficiencia y redes de termiteros Diego Griffon Klaus Jaffe Carmen Andara La complejidad aparente de nuestro comportamiento en el tiempo es, en buena parte, un reflejo de la complejidad del ambiente en el cual nos encontramos. H. A. Simon (1996) Introducci ´ on Durante las ´ ultimas d´ ecadas del siglo veinte la investigaci´ on de los sistemas complejos se afianz´ o como un campo reconocido de investigaci´ on cient´ ıfica. Esta ´ area del conocimiento ha sido particularmente exitosa en proveer herramientas y aproximaciones epistemol´ ogicas tan novedosas (Capra, 1996) que algunas de ellas han dado paso a serios cuestionamientos y cambios de paradigmas (Gleick, 1997). Se considera particularmente importante dentro de este acercamiento, el abandono del reduccionismo, pr´ actica que ha permitido poner en relieve nuevamente la importancia que tienen las interacciones en la din´ amica a gran escala de los sistemas. Una importante consecuencia de estas interacciones es el surgimiento de propiedades emergentes (Goodwin, 1994), es decir aquellas que surgen a un nivel de organizaci´ on superior a partir de las interacciones que se dan entre los elementos de un nivel de organizaci´ on inferior (Sol´ e y Manrubia, 1996). Las propiedades emergentes explican una infinidad de patrones y procesos en las m´ as diversas ´ areas de la ciencia (Sol´ e y Goodwin, 2000). Copia personal del autor.
Transcript of H. A. Simon (1996)atta.labb.usb.ve/Klaus/art217.pdfPor ejemplo, se han encontrado patrones...

177
Modelos y simulaciones biológicas: ecología y evoluciónHarold P. de Vladar y Roberto Cipriani. (eds.) 2015Impreso por Createspace. ISBN-13: 978-1516867561 / ISBN-10: 1516867564
https://goo.gl/kVfvnu
Modelos y simulaciones biologicas: ecologıa y evolucion.Harold P. de Vladar y Roberto Cipriani (eds.) 2015.Impreso por CreateSpace. ISBN: 1516867564 978-1516867561.
Propiedades emergentes, eficiencia y redes determiteros
Diego Griffon Klaus Jaffe Carmen Andara
La complejidad aparente de nuestro comportamiento en eltiempo es, en buena parte, un reflejo de la complejidad delambiente en el cual nos encontramos.
H. A. Simon (1996)
Introduccion
Durante las ultimas decadas del siglo veinte la investigacion delos sistemas complejos se afianzo como un campo reconocido deinvestigacion cientıfica. Esta area del conocimiento ha sidoparticularmente exitosa en proveer herramientas y aproximacionesepistemologicas tan novedosas (Capra, 1996) que algunas de ellashan dado paso a serios cuestionamientos y cambios de paradigmas(Gleick, 1997).
Se considera particularmente importante dentro de esteacercamiento, el abandono del reduccionismo, practica que hapermitido poner en relieve nuevamente la importancia que tienen lasinteracciones en la dinamica a gran escala de los sistemas. Unaimportante consecuencia de estas interacciones es el surgimiento depropiedades emergentes (Goodwin, 1994), es decir aquellas quesurgen a un nivel de organizacion superior a partir de lasinteracciones que se dan entre los elementos de un nivel deorganizacion inferior (Sole y Manrubia, 1996). Las propiedadesemergentes explican una infinidad de patrones y procesos en las masdiversas areas de la ciencia (Sole y Goodwin, 2000).
Copia personal del autor.

178
Redes de termiteros
Una de las areas de investigacion activas en el estudio de lossistemas complejos, esta orientada a establecer hasta que punto unapropiedad emergente puede optimizar una funcion (Buhl et al., 2004;Perna et al., 2008). Es precisamente esta la lınea conceptual que sesigue en este capıtulo. Aquı analizaremos como una propiedademergente puede optimizar procesos y funciones especıficas.
Nos concentraremos en un caso particular de insectos eusociales.Los patrones conductuales de estos insectos son frecuentementeutilizados en la literatura como ejemplos de sistemas complejos(Sole y Manrubia, 1996) y han sido ampliamente estudiados(Theraulaz y Spitz, 1997).
Son particularmente interesantes los aportes realizados porBonabeau et al. (1999); Bonabeau y Theraulaz (1994); Theraulaz ySpitz (1997); Camazine et al. (2003) en los que se establecefirmemente el rol de las propiedades emergentes en loscomportamientos de enjambre de estos organismos (Theraulaz ySpitz, 1997; Bonabeau et al., 1999).
Investigadores como Buhl et al. (2004); Jackson et al. (2004);Goss et al. (1989); Perna et al. (2008) han encontrado evidencias quemuestran como del comportamiento de insectos eusociales surgenpropiedades emergentes que optimizan funciones importantes parasus colonias. En particular, utilizando un enfoque basado en la teorıade grafos Buhl et al. (2004); Perna et al. (2008) han mostrado comola estructura de estos nidos y sus galerıas optimizan elcomportamiento de forrajeo de los insectos.
La teorıa de grafos estudia, entre otras cosas, las propiedadesgeometricas de las redes, que son estructuras abstractas definidas porun conjunto de nodos conectados a traves de enlaces (Albert yBarabasi, 2002; Dorogovtsev y Mendes, 2002; Newman, 2003). Unaaproximacion interesante para estudiar la arquitectura de los nidos delos insectos eusociales y establecer cuan cercanas se encuentra estade optimos geometricos, utilizando esta teorıa, es considerar a losnidos como nodos y a las galerıas entre ellos como enlaces.
Copia personal del autor.

179
D. Griffon, K. Jaffe y C. Andara
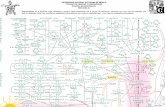
Figura 1: Red de termiteros y galerıas de N. ephratae luego que un incendio naturalhiciera su estructura visible. Derecha: fragmento de una red. Izquierda: detalle dela misma red en la cual se han resaltado sus elementos. Las redes estudiadas estancompuestas de 3 elementos basicos: nidos principales, nidos satelites y galerıas entrenidos. Lınea gris: nidos principales, Lınea blanca: nidos satelites, Lıneas Negras:galerıas.
En este capıtulo utilizamos esta aproximacion para estudiar laarquitectura de las redes de termiteros que Nasutitermes ephratae(Isoptera: Nasutitermitinae) construye sobre la superficie del suelo ensabanas herbaceas de la Gran Sabana (Venezuela) (Figura 1). Enparticular, este estudio se basa en informacion de campo obtenida de3 redes de termiteros: una gran red compuesta de 645 nidos y 707galerıas, a la que llamaremos Red Principal y dos redes maspequenas, una con 80 y otra con 70 nidos.
Redes complejas
Uno de los avances mas interesantes en el area de los sistemascomplejos, ha sido el descubrimiento de patrones de organizacionafines en redes de diferente naturaleza (Strogatz, 2001; Sole, 2009).Por ejemplo, se han encontrado patrones topologicos comunes enredes metabolicas, ecologicas, sociales e inclusive en la Internet(Sole, 2009). Hallazgos como estos han permitido pensar que es
Copia personal del autor.

180
Redes de termiteros
Figura 2: (a) Redes Poisson y (b) ley de potencia, con sus respectivas distribucionesde grados.
posible que existan patrones universales de organizacion en estasredes.
Distribucion de grados. Una de las caracterısticas que ha recibidomas atencion en el estudio de las redes complejas es la distribucionde grados (Sole, 2009). La distribucion de grados de una red es unhistograma que muestra la distribucion de frecuencias de nodos conun determinado numero de enlaces (i.e. grado del nodo). Dosdistribuciones de grado han recibido mucha atencion (Newman,2003): la distribucion tipo Poisson y la distribucion tipo ley depotencia (Figura 2). La distribucion Poisson caracteriza a redes cuyosenlaces han sido asignados al azar. En este tipo de redes el numero deenlaces por nodo es poco variable y puede ser caracterizado por lamedia. Por su parte, la distribucion tipo ley de potencia se encuentraen redes con un numero de enlaces por nodo muy variable y en lasque pocos nodos poseen multiples enlaces y mutiples nodos, pocosenlaces.
Una caracterıstica importante de estas distribuciones de grado esque son propias de redes cuyas topologıas optimizan procesosparticulares. En las redes Poisson, la estructura optimiza lavulnerabilidad de la red ante la perdida de nodos. Es decir, estasredes no pierden cohesion cuando se eliminan sus nodos. Por suparte, en las redes tipo ley de potencia, se optimiza la distancia entre
Copia personal del autor.

181
D. Griffon, K. Jaffe y C. Andara
Figura 3: Distribucion de grados de la Red Principal, ası como las distribucionespromedio resultado de la simulacion de redes Poisson y ley de potencia.
los nodos que no son vecinos (Albert et al., 2000). En este caso, seentiende por distancia la cantidad de enlaces que separan a dos nodosde la red.
La primera pregunta que abordamos en este estudio es establecersi las redes de termiteros de N. ephratae presentan distribuciones degrados similares a cualquiera de las antes descritas, Poisson o ley depotencia. Para ello se simularon 100 redes con topologıas acordes concada una de estas distribuciones. Las redes tipo Poisson se simularonmediante el mecanismo propuesto por Erdos y Renyi (1960) mientrasque las redes tipo ley de potencia mediante el algoritmo propuestopor Pennock et al. (2002). Todas fueron generadas con la restriccionde tener el mismo numero de nodos y promedio de grados, Kmed = 2.2,de la Red principal con la cual se las comparo.
En la Figura 3 se muestra la distribucion de grados real de la RedPrincipal, junto como las distribuciones promedio obtenidasmediante simulaciones. Si bien estos graficos tienen muy pocos
Copia personal del autor.

182
Redes de termiteros
Figura 4: Relaciones entre las redes regulares, de mundo pequeno y aleatorias.
puntos como para poder realizar un ajuste estadıstico riguroso, en lafigura se advierte que la distribucion de grados de la Red Principal nose asemeja a ninguna de las dos distribuciones teoricas.
Mundos pequenos. Otro patron arquitectonico comun a muchasredes complejas ha sido propuesto por Watts y Strogatz (1998). Estosautores han sugerido que la mayorıa de las redes observadas no lucencomo grafos con conexiones aleatorias, pero tampoco parecen mallasregulares (i.e., lattices o grillas) con igual numero de conexionesentre los nodos. Ellos proponen un modelo de formacion de redes enel cual a partir de una malla regular se reconectan los enlaces al azar,con una probabilidad P (Figura 4). Mediante este simple proceso seobtienen redes conocidas como de Mundo Pequeno. La caracterısticacentral de estas redes (y a la cual deben su nombre) es que paracualquier par de nodos escogidos al azar, se puede encontrar una rutacorta que los une, la cual involucra la intermediacion de un numeroreducido de nodos y enlaces.
Newman (2003) ha definido a una red Mundo Pequeno comoaquella en la que la distancia promedio es igual o menor al logaritmodel tamano del sistema (n). El tamano del sistema es igual al numerode nodos presente en la red. En nuestro caso, el tamano es 645 parala Red Principal. La distancia promedio de la red es el numero
Copia personal del autor.

183
D. Griffon, K. Jaffe y C. Andara
promedio de enlaces que separan a todas las parejas de nodos, 23 eneste estudio. Dado que el logaritmo de n es aproximadamente 3 en laRed Principal, difıcilmente esta puede ser considerada como una redde Mundo Pequeno.
Redes con restricciones dimensionales. Hemos visto que lasredes de termiteros no siguen patrones topologicos comunmentepresentes en otras redes complejas. Una posible explicacion de esteresultado es que las restricciones existentes en las redes de termiterosson diferentes a las de otras redes complejas. Veamos este argumentocon mas detalle. Una caracterıstica comun a todas las redes cuyasdistribuciones de grados se ajustan a distribuciones Poisson o ley depotencia, es que son redes en las cuales los enlaces no representandistancias reales. Por ejemplo, un enlace en una red trofica,metabolica o informatica no representa la distancia que la“informacion” debe recorrer en la red (i.e., los enlaces no sonestructuras espacialmente explıcitas). Sin embargo, los enlaces en lasredes de termiteros son estructuras reales, son galerıas cubiertas quese extienden por la superficie del suelo, en basicamente dosdimensiones. Las galerıas son utilizadas por las termitas con finesconcretos y su construccion involucra costos energeticos. Por lotanto, en las redes de termiteros operan restricciones espaciales yenergeticas evidentes que deben ser incluidas en su analisis.
Redes en dos dimensiones
Son muchas las redes reales construidas en dos dimensiones.Existen redes electricas, ferroviarias, de tuberıas y carreteras(Gorman y Kulkarni, 2004; Gastner y Newman, 2006a,b) que hansido bien estudiadas. En todos estos casos, el hecho de estarconstruidas sobre superficies ha mostrado tener importantes efectosen su topologıa, y todas poseen una estructura planar (Gastner yNewman, 2006a,b). Una red planar es aquella que puede ser
Copia personal del autor.

184
Redes de termiteros
Figura 5: Representacion esquematica de la Red Principal
representada en dos dimensiones sin que ninguno de sus enlaces sesuperponga (i.e., sus enlaces nunca se cruzan). La redes planarespueden ser entendidas como grafos que optimizan su estructura endos dimensiones, evitando que ocurran arreglos ineficientes como elpaso de dos enlaces por el mismo sitio (Gorman y Kulkarni, 2004).
Otra caracterıstica comun de estas redes es que los angulos entrelos enlaces optimizan alguna de sus funciones (Goss et al., 1989). Enel caso de las redes de termiteros, la funcion principal de las galerıases permitir la exploracion del area circundante a los nidos en buscade alimento, por lo tanto es posible que los angulos en estas redesoptimicen esta conducta. Es de esperar entonces que la estructura de lared permita explorar el area alrededor de los nidos de forma eficiente.
En las redes de termiteros, la construccion de galerıas involucracostos energeticos, por lo que una manera de optimizar suconstruccion es minimizar el numero de galerıas construidas. Ungrafo construido con el numero mınimo de enlaces es conocido comoun minimum spanning tree (MST) (Gastner y Newman, 2006a,b),Entonces, una manera de evaluar si las redes de termiteros optimizan
Copia personal del autor.

185
D. Griffon, K. Jaffe y C. Andara
Figura 6: Distribucion de grados de diferentes redes de termiteros. Notese el maximopresente en todas las redes en nidos con dos galerıas.
el numero de enlaces es comparando su estructura con la de MSTs.En los siguientes apartados de este capıtulo pondremos a prueba
si las redes de termiteros de N. ephratae son planares, optimizan losangulos entre los nidos, optimizan el area de forrajeo y, finalmente,son similares a MSTs particulares.
Redes de termiteros
Generalidades. Las redes de termiteros estudiadas pueden ocuparareas de hasta 2000 m2 y presentan un marcado sesgo hacia lapresencia de nidos con dos galerıas (i.e., grado dos). En efecto, elsesgo hacia nidos con dos galerıas es una caracterıstica comun atodas las redes de termiteros evaluadas. Esto se observa claramenteen la representacion de la Red Principal (Figura 5) y en sudistribucion de grados (Figura 6).
Copia personal del autor.

186
Redes de termiteros
1 2 5 10 20 50
25
1020
50100
200
500
Log Vecindario
Log
Rad
iopendiente= 1.35pendiente= 1.35pendiente= 1,35
Figura 7: Dimension de la Red Principal.
Redes de termiteros planares. Para establecer si una red es planarse debe calcular su dimension: redes con dimensiones menores a 2son planares. La dimension de una red puede ser calculada medianteel procedimiento propuesto por Gastner y Newman (2006a). Estametodologıa es conceptualmente similar al procedimiento de conteode cajas utilizado para calcular dimensiones fractales. El metodoconsiste en: (a) Utilizar cırculos de diferentes radios para establecerlos vecindarios de cada nodo. Un vecindario esta definido como elnumero de nodos con los cuales se conecta un nodo particular en unacircunferencia de radio dado; (b) Calcular la media de losvecindarios delimitados por circunferencias de diferentes radios; (c)Representar graficamente el logaritmo de la frecuencia de las mediasfrente al logaritmo de los radios de las circunferencias utilizadas paraevaluar la red (Figura 7); (d) Ajustar una recta de regresion demınimos cuadrados; (e) Obtener la pendiente de esta recta (Newmany Watts, 1999; Csanyi y Szendroi, 2004), que representa la
Copia personal del autor.

187
D. Griffon, K. Jaffe y C. Andara
Figura 8: Ejemplos de angulos entre galerıas (lıneas). Los cırculos grises representanterritorios de forrajeo de tamano constante. Angulos de 180o o de 90o (±45) impidenla sobreposicion de los territorios de forrajeo. Otros angulos producen sobreposicion,ver por ejemplo 30o.
dimension de la red. Para las redes de termiteros, esta pendiente es enpromedio 1.35 (± 0.03). Al ser este numero menor que 2, las redesde termiteros estudiadas pueden ser consideradas planares.
Angulos entre galerıas. En las redes de termiteros evaluadas, losangulos entre galerıas consecutivas no son aleatorios. En efecto,predominan ciertos intervalos de angulos. El 60% de los angulos seencuentran en el intervalo comprendido entre 60o y 120o, mientrasque el 25% de los angulos se encuentran entre 160o y 180o. Si sesupone que los territorios de forrajeo de esta especie tienen untamano constante, los angulos observados con mayor frecuencia enlas redes son precisamente aquellos que impiden la superposicion delos territorios de forrajeo (Figura 8), optimizando de esta manera laactividad de las termitas.
Copia personal del autor.

188
Redes de termiteros
Figura 9: Representacion a escala de un fragmento de la Red Principal, en gris serepresenta el area potencial de forrajeo. El cırculo es equivalente un area de 24 m2.
Area de forrajeo. Indudablemente, el fin ultimo de la conducta deforrajeo en las termitas es colectar la mayor cantidad posible dealimento. En el caso de N. ephratae, el alimento consiste en restosde hojas y tallos de gramıneas y ciperaceas que las termitas colectanen las areas circundantes a sus nidos. Por lo tanto, una estructura quepuede aumentar la eficiencia de esta actividad, es aquella que permitaa las termitas explorar y forrajear el maximo posible del areacircundante a la red. Ası, para establecer hasta que punto laestructura de la red ayuda al forrajeo, es necesario primero establecerque proporcion del area circundante a la red esta disponible paraforrajear.
Se conoce que N. ephratae puede forrajear a cielo descubierto auna distancia maxima de 0.5 m (Carmen Andara, datos nopublicados). Partiendo de esta informacion, se puede delinear sobredibujos a escala de las redes un perımetro maximo de 0.5 malrededor de la estructura mas proxima de los nidos y galerıas queforman la red, estableciendo ası el area de forrajeo (Figura 9).
En la Figura 9 se aprecia como la arquitectura de la red asegura unuso casi total del area alrededor de los nidos. En efecto, si dibujamos
Copia personal del autor.

189
D. Griffon, K. Jaffe y C. Andara
Figura 10: Eficiencia de la red. Izquierda: fragmento de la Red Principal. Derecha:MST obtenido a partir del fragmento de la Red Principal. Numero de galerıas enel fragmento de la Red Principal = 35, numero de galerıas en el MST = 31. Losvınculos representados con una lınea punteada en el fragmento de la Red Principal,son aquellos que deben ser eliminados para transformar esta red en un MST.
un cırculo de diametro de 5.5 m alrededor de los nidos principales(Figura 9) obtenemos en promedio un 77.8% de eficiencia en el usodel area.
Minimum spanning tree. Las redes de termiteros pueden sertransformadas en un MST (Buhl et al., 2004). El MST obtenidomediante esta transformacion representa la forma geometrica optimade conectar los nodos de la red original (i.e., utilizando el menornumero de enlaces posible). Para establecer si las redes de termiterostienen una estructura optima, se comparo el numero de enlacespresentes en ellas con el numero de enlaces presentes en MSTsobtenidos a partir de las redes originales. Se encontro que la relacion:numero de vınculos en la red de nidos / numero de vınculos del MST,es en promedio 0.85 (± 0.12). Esto demuestra que la estructura deesta red se encuentra muy cerca de su optimo. En la Figura 10 semuestra un ejemplo de las modificaciones necesarias para trasformarun fragmento de las redes de termiteros en un MST.
Copia personal del autor.

190
Redes de termiteros
Algoritmo 1 Red de Termiteros (Kmax,L,P, I )1: Kmax ← 8 (i.e. no mas galerıas/nido que el valor del grado maximo de las redes reales).
2: Se construye un nido en el centro de la grilla.
3: En el vecindario tipo Moore del nido se construye un nuevo nido con una probabilidad
homogenea para todas las 8 celdas. El nuevo nido se conecta con el anterior mediante una
galerıa. Si galerıas/nido > Kmax, se detiene el procedimiento.
4: Se repite (3) L−1 veces (i.e., caminata aleatoria bidimensional de L pasos).
5: Con probabilidad P, se repite (3) L veces partiendo del nodo inicial o final de la caminata
recien terminada (i.e., caminata aleatoria bidimensional de L pasos). Si galerıas/nido >
Kmax, se detiene el procedimiento.
6: Se repite (2)-(5) I veces.
¿Como construir una red de termiteros?
Hasta ahora se ha mostrado como ciertas caracterısticas de lasredes de termiteros de N. ephratae se encuentran muy cerca de susoptimos funcionales y geometricos. Esto nos lleva a preguntarnosacerca de los mecanismos que permiten que las termitas construyannidos con estas particularidades. Para contestar esta preguntaimplementamos un algoritmo simple, que supone que la longitud delos enlaces es uniforme y el ambiente homogeneo, restricciones queconcuerdan con la evidencia empırica (Algoritmo 1).
Como resultado de simulaciones efectuadas con el modelo, seencontro que bajo amplios intervalos de los valores de los parametrosL,P, I el modelo logra reproducir las caracterısticas basicas de lasredes de termiteros estudiadas (Figura 11).
Comentarios finales
En este capıtulo se ha mostrado que la especie de termitasNasutitermes ephratae construye redes de termiteros conarquitecturas sumamente eficientes en terminos del numero degalerıas utilizadas y del area de forrajeo generada por su estructura.
Copia personal del autor.

191
D. Griffon, K. Jaffe y C. Andara
Figura 11: Red simulada y su distribucion de grados.
Los resultados mostrados son particularmente interesantes cuando seconsidera que la construccion de estas redes es un ejemplo deautoorganizacion. Recordemos que los patrones arquitectonicosobservados en los nidos de los insectos eusociales emergen dedecisiones tomadas autonomamente por un grupo de agentes simples,sin que exista un plan general para su construccion (Buhl et al.,2004). Aun ası, muchas caracterısticas provistas por estas estructurasson consideradas optimas o cuasi-optimas, ejemplos los encontramosen su organizacion espacial, la ventilacion que proveen y latemperatura y humedad relativa que mantienen en su interior (Korb yLinsenmair, 1998; Cole, 1994; Farji-Brener, 2000; Nielsen et al.,2003).
Tambien se mostro que existen mecanismos de construccionsencillos que reproducen las caracterısticas generales de las redes determiteros de esta especie. Es interesante apreciar que en este casomediante la ejecucion de reglas simples, se pueden obtenerestructuras sumamente eficientes en terminos de la realizacion defunciones concretas, estructuras en efecto casi optimas. En estesentido Herbert Simon (1955) definio la palabra satisficing para
Copia personal del autor.

192
describir estrategias que alcanzan criterios de adecuacion, en vez deoptimos. Este tal vez sea un buen termino para definir la estructura delas redes de termiteros de N. ephratae, ya que su estructura esadecuada para llevar a cabo su funcion, aunque no es estrictamente laestructura optima.
Finalmente, se ha mostrado como una propiedad emergentepuede llevar a cabo funciones de una manera eficiente (satisficing).En este sentido, si consideramos que muchas propiedades emergentesconocidas surgen mediante mecanismos sencillos (no muy diferentesde los aquı descritos), no es descabellado especular que estastambien permitan realizar funciones de forma igualmente eficiente.
Agradecimientos Los autores deseamos agradecer a Glenda Briceno,
Dayaleth Alfonzo, Roberto Cipriani y a dos arbitros anonimos por sus sugerencias y
comentarios, los cuales ayudaron a mejorar significativamente la calidad de este
trabajo. De igual manera expresamos nuestro agradecimiento a Augusto Andara y al
personal de la Estacion Cientıfica Parupa (Gran Sabana-Venezuela), sin cuyo apoyo
no hubiese sido posible la realizacion del trabajo de campo.
Referencias
Albert, R. y Barabasi, A. L. (2002). Statistical mechanics of complex networks.Reviews of Modern Physics, 74(1):47–97.
Albert, R., Jeong, H., y Barabasi, A. L. (2000). Error and attack tolerance of complexnetworks. Nature, 406(6794):378–382.
Bonabeau, E., Dorigo, M., y Theraulaz, G. (1999). Swarm intelligence: from naturalto artificial systems. Oxford University Press, Oxford.
Bonabeau, E. y Theraulaz, G., editores (1994). Intelligence Collective. Hermes,Paris.
Buhl, J., Gautrais, J., Sole, R., Kuntz, P., Valverde, S., Deneubourg, J., y Theraulaz, G.(2004). Efficiency and robustness in ant networks of galleries. European PhysicalJournal B, 42(1):123–129.
Copia personal del autor.

193
Camazine, S., Deneubourg, J., Franks, N., Sneyd, J., Theraulaz, G., y Bonabeau,E. (2003). Self-organization in biological systems. Princeton University Press,Nueva Jersey.
Capra, F. (1996). The Web of Life. Anchor Books, Nueva York.
Cole, B. J. (1994). Nest architecture in the western harvester ant, Pogonomyrmexoccidentalis (cresson). Insectes Sociaux, 41(4):401–410.
Csanyi, G. y Szendroi, B. (2004). Fractal-small-world dichotomy in real-worldnetworks. Physical Review E, 70(1):016122.
Dorogovtsev, S. y Mendes, J. (2002). Evolution of networks. Advances in Physics,51(4):1079–1187.
Erdos, P. y Renyi, A. (1960). On the evolution of random graphs. Publications of theMathematical Institute of the Hungarian Academy of Sciences., 5:17–61.
Farji-Brener, A. (2000). Leaf-cutting ant nests in temperate environments: mounds,mound damages and nest mortality rate in Acromyrmex lobicornis. Studies onNeotropical Fauna and Environment, 35(2):131–138.
Gastner, M. y Newman, M. (2006a). The spatial structure of networks. EuropeanPhysical Journal B, 49(2):247–252.
Gastner, M. T. y Newman, M. E. J. (2006b). Shape and efficiency in spatialdistribution networks. Journal of Statistical Mechanics, p. P01015.
Gleick, J. (1997). Chaos: Making a new science. Penguin Books, Nueva York.
Goodwin, B. (1994). How the Leopard Changed Its Spots: The Evolution ofComplexity. Touchstone, Nueva York.
Gorman, S. y Kulkarni, R. (2004). Spatial small worlds: New geographic patternsfor an information economy. Environment & Planning B, 31:273–296.
Goss, S., Aron, S., Deneubourg, J. L., y Pasteels, J. M. (1989). Self-organizedshortcuts in the argentine ant. Naturwissenschaften, 76(12):579–581.
Jackson, D., Holcombe, M., y Ratnieks, F. (2004). Trail geometry gives polarity toant foraging networks. Nature, 432(7019):907–909.
Copia personal del autor.

194
Korb, J. y Linsenmair, K. (1998). The effects of temperature on the architectureand distribution of Macrotermes bellicosus (Isoptera, Macrotermitinae) mounds indifferent habitats of a West African Guinea savanna. Insectes Sociaux, 45(1):51–65.
Newman, M. E. J. (2003). The structure and function of complex networks. SIAMReview, 45(2):167–256.
Newman, M. E. J. y Watts, D. J. (1999). Scaling and percolation in the small-worldnetwork model. Physical Review E, 60(6):7332–7342.
Nielsen, M., Christian, K., y Birkmose, D. (2003). Carbon dioxide concentrationsin the nests of the mud-dwelling mangrove ant Polyrhachis sokolova Forel(Hymenoptera: Formicidae). Australian Journal of Entomology, 42(4):357–362.
Pennock, D., Flake, G., Lawrence, S., Glover, E., y Giles, C. (2002). Winners don’ttake all: Characterizing the competition for links on the web. Proceedings of theNational Academy of Sciences of the USA, 99(8):5207–5211.
Perna, A., Jost, C., Couturier, E., Valverde, S., Douady, S., y Theraulaz, G. (2008).The structure of gallery networks in the nests of termite Cubitermes spp. revealedby X-ray tomography. Naturwissenschaften, 95(9):877–884.
Simon, H. A. (1955). A behavioral model of rational choice. Quarterly Journal ofEconomics, 69:99–118.
Simon, H. A. (1996). The Sciences of the Artificial. MIT Press, Cambridge (MA).
Sole, R. V. (2009). Redes complejas. Del genoma a Internet. Tusquets, Barcelona.
Sole, R. V. y Goodwin, B. (2000). Signs of Life: How Complexity Pervades Biology.Basic Books, Nueva York.
Sole, R. V. y Manrubia, S. C. (1996). Orden y caos en sistemas complejos.Aplicaciones. Edicions UPC SL, Barcelona.
Strogatz, S. H. (2001). Exploring complex networks. Nature, 410(6825):268–276.
Theraulaz, G. y Spitz, F. (1997). Auto-organisation et comportement. Hermes, Parıs.
Watts, D. J. y Strogatz, S. H. (1998). Collective dynamics of ‘small-world’ networks.Nature, 393(6684):440–442.
Copia personal del autor.

195
ContactosDG: Laboratorio de Evolucion y Ecologıa Teorica. Instituto deZoologıa y Ecologıa Tropical, Universidad Central de Venezuela.Caracas, [email protected]
KJ: Laboratorio de Evolucion. Universidad Simon Bolıvar. Caracas,[email protected]
CA: Laboratorio de Neurociencias y Comportamiento. Departamentode Biologıa, Facultad Experimental de Ciencias y Tecnologıa,Universidad de Carabobo. Barbula, [email protected]
Copia personal del autor.