
Experimento de Anfinsen (1956-1957) - Apache2 Ubuntu...
27
Experimento de Anfinsen (1956-1957) Teoría “termodinámica” Christian B. Anfinsen (1916-1995)
Transcript of Experimento de Anfinsen (1956-1957) - Apache2 Ubuntu...
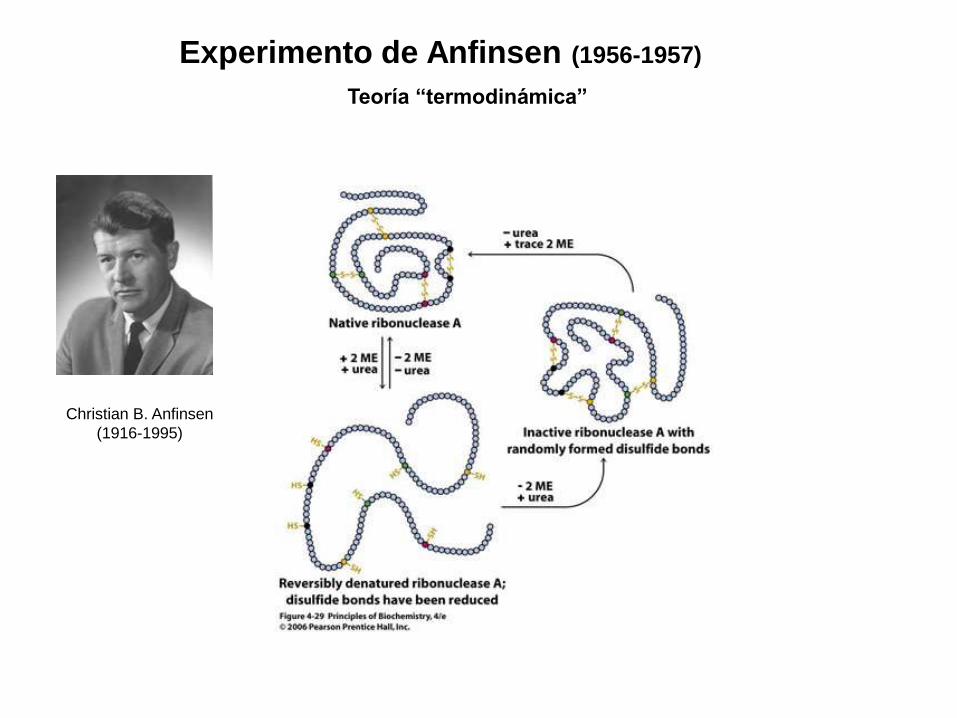
Experimento de Anfinsen (1956-1957)
Teoría “termodinámica”
Christian B. Anfinsen
(1916-1995)


Plegamiento de proteínas
Teoría cinética
Para describir la posición de un átomo en el espacio uno necesita
definir 3 coordenadas
Para una proteína de 100 aminoácidos necesitamos 6000
coordenadas (consideramos que un AA contiene en promedio 20
átomos)
Si cada una de esas coordendas puede tomar al menos 2 valores
distintos tendríamos que el espacio conformacional de la proteína
sería de 26000
Si cada transición entre cada una de éstas conformaciones distintas
requiere de un tiempo de por ejemplo 110-13 segundos….
…plegar una proteína requeriría
25987 segundos!!!
“Paradoja de Levintal”
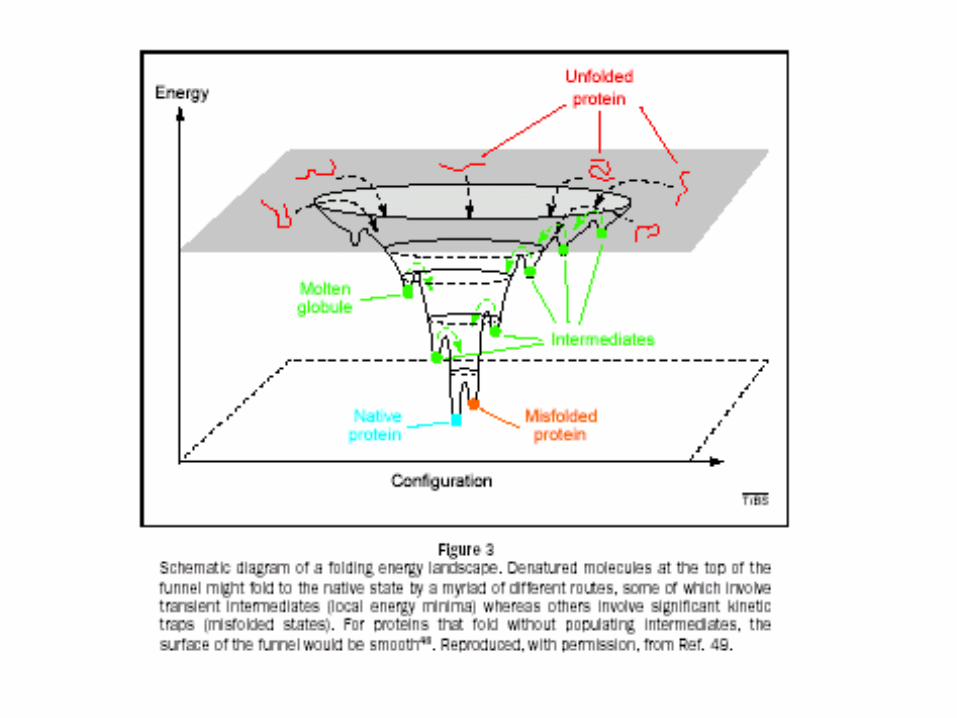

Una mirada a los factores contribuyentes.
Estructura nativa Conjunto desnaturalizado
Intra H-Bond (H) H-Bond to Water (H)
Hydrophobic Effect (S) Configurational Entropy (S)
Packing (H)
Energía libre del plegamiento = Gu – GN = H-TS ~ 10 kcal/mol
Número chico resultado de la diferencia de números MUY grandes.
Muy dificil para lograr un efecto particular!
Claramente hay una compensación entre entropía-energía.
A medida que T aumenta, G se vuelve cero y luego positiva
Protein Folding

Dos asunciones
• El plegamiento de una proteína encuentra el mínimo
global de energía libre en el paisaje de conformaciones
posibles.
• La estructura 3-D de una proteína es determinada
solamente por su estructura primaria (lista de
aminoácidos).
Anfinsen (Premio Nobel 1972)
Existe una única correlación entre estructura plegada y función
Protein Folding

Por qué es un problema
computacionalmente difícil?
Si “conocemos” los factores energéticos que controlan el sistema,
por qué no podemos encontrar el mínimo global de energía libre y
solucionar el problema?
Paradoja de Levinthal

El problema del golfista ciego
Intentar encontrar el agujero en un campo de golf plano.
Paradoja de Levinthal
El “Campo de golf” chato
Solución del camino de Levinthal
El “Campo de golf acanalado”
Energy D
Grooved golf
course
D
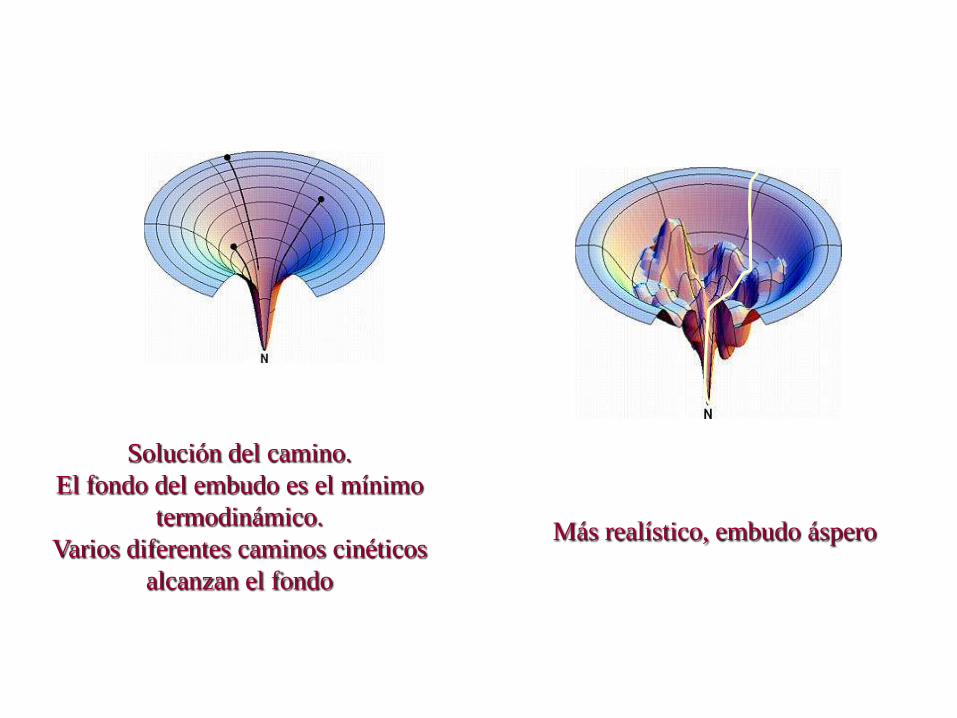
Solución del camino.
El fondo del embudo es el mínimo
termodinámico.
Varios diferentes caminos cinéticos
alcanzan el fondo
Más realístico, embudo áspero
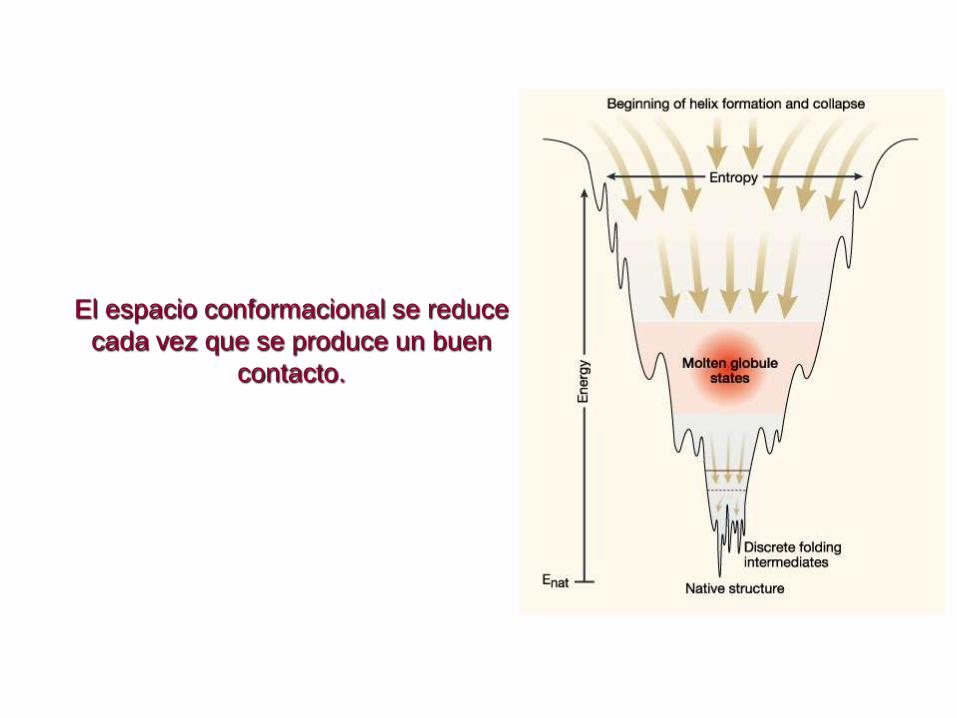
El espacio conformacional se reduce
cada vez que se produce un buen
contacto.

Protein Folding: Fast Folders
• Trp-cage, designed mini-protein (20 aa): 4μs
• b-hairpin of C-terminus of protein G (16 aa) : 6 μs
• Engrailed homeodomain (En-HD) (61 aa): ~27 μs
• WW domains (38-44 aa): >24 μs
• Fe(II) cytochrome b562 (106 aa): extrapolated ~5 μs
• B domain of protein A (58 aa): extrapolated ~8 μs
Folding MD Simulations Folding Experiments
ps ns ms ms ms sec
80’s 90’s 00’s 00’s 90’s 80’s
Time Scale:
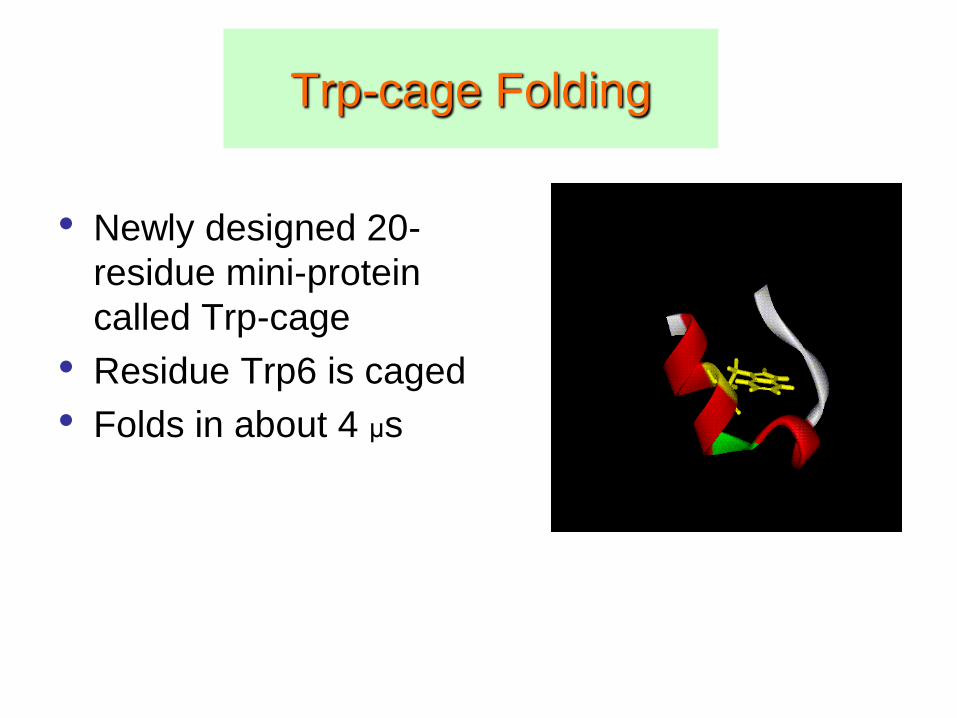
Trp-cage Folding
• Newly designed 20-
residue mini-protein
called Trp-cage
• Residue Trp6 is caged
• Folds in about 4 μs
Trp-cage

a helix
310 helix
polyproline II
i:i+10 H-bond
i:i-5 H-bond
(75%)
hydrophobic
core N
C
MD low energy
1L2Y model 1
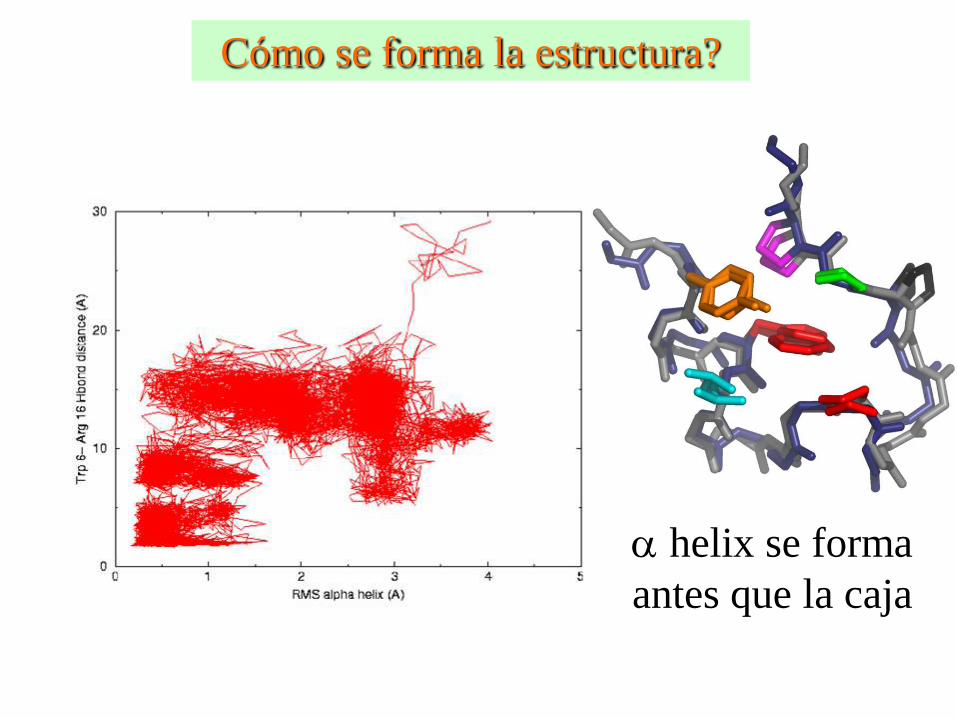
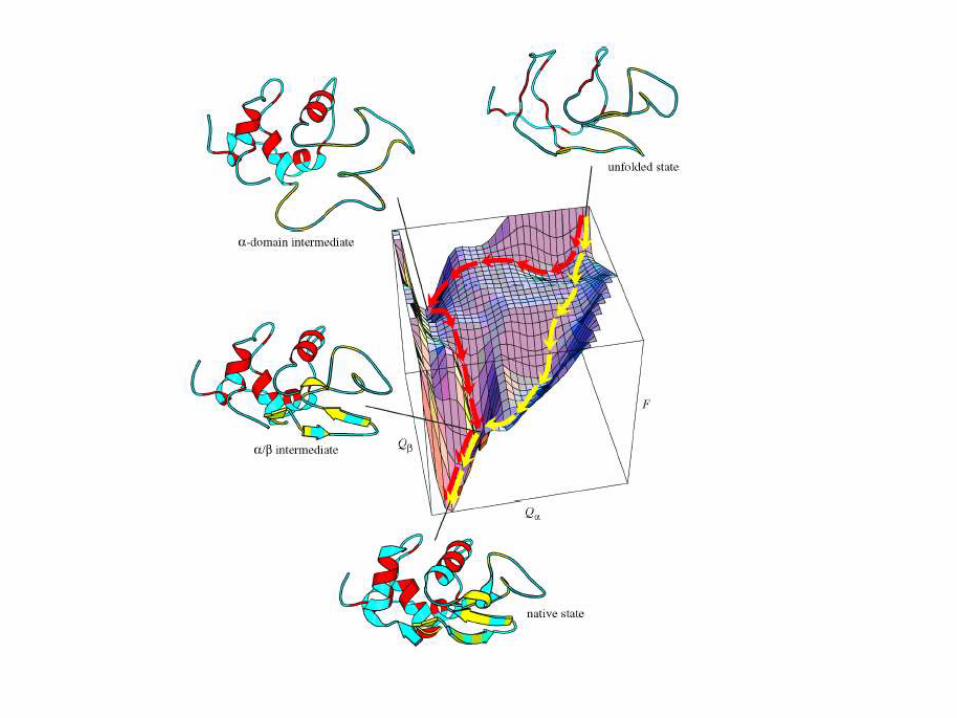
Cómo se forma la estructura?
a helix se forma
antes que la caja
Substituciones en la secuencia de aa
afectan a
función dinámica estructura
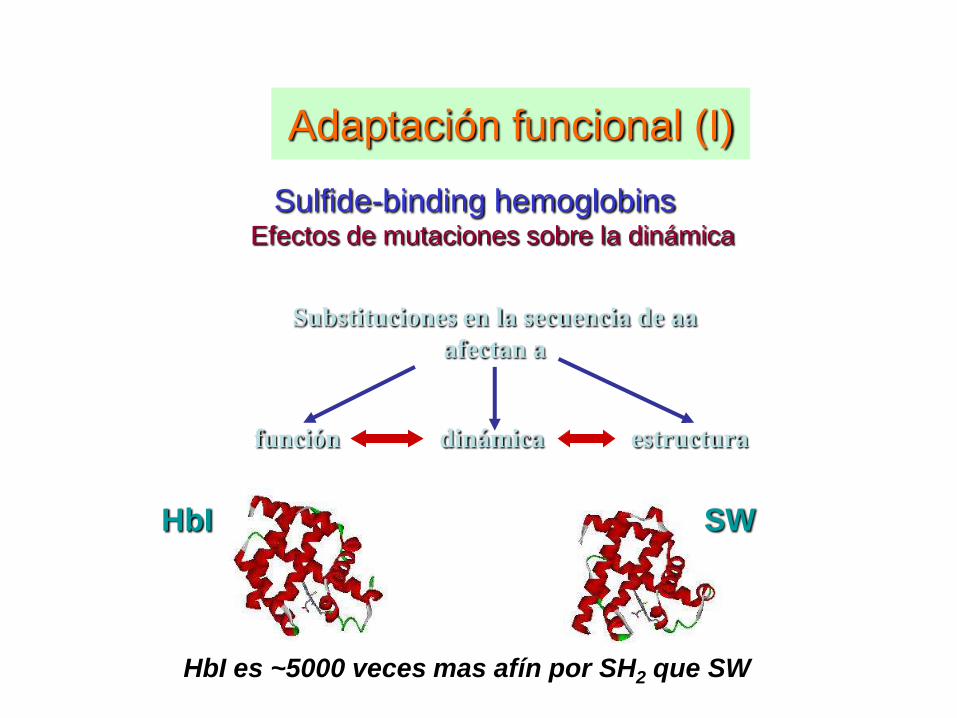
Adaptación funcional (I)
Sulfide-binding hemoglobins Efectos de mutaciones sobre la dinámica
HbI SW
HbI es ~5000 veces mas afín por SH2 que SW

Phe 43 Phe 43
Gln 64
His 64
Heme Heme
SH2 SH2
Leu 29 Phe 29
Phe 68 Val 68
HbI SW

His64(E7) Gln, Leu29(B10) Phe, Val68(E11) Phe
FQF es ~7 mas afín por SH2 que SW
¿Existe alguna propiedad dinámica que dá cuenta del resto?
HbI SW
FQF

HbI
His 97
Arg 45
Arg 99
SW
Ser 92
Rocking freedom inusual en el grupo heme de HbI que
facilitaria la entrada del ligando Puentes de H de heme-propionatos
Arg 99: lifetime = 1.7 ps
Ser92: lifetime = 3.4 ps
His97: lifetime = 2.2 ps
Arg45: lifetime = 4.1 ps

Enzimas psicrófilas evolucionaron para catalizar reacciones a bajas temperaturas
Compensación térmica (Ley de Arrhenius ( k=Ae-Ea/RT ))
para acomodar mejor a los sustratos y realizar los cambios conformacionales
Aumento de la flexibilidad estructural
Adaptación funcional
Adaptación al frío Efectos de mutaciones sobre la flexibilidad

Mesophile Psychrophile
SeqId %: 68.72
Bacillus Globisporus
(transferasa) Bacillus Subtilis
(transferasa)
Pig pancreatic (hydrolase) Alteromonas
haloplanctis
SeqId %: 46.53
Pseudoalteromonas
Haloplantis
(hydrolase)
Clostridium
Thermocellum
(hydrolase)
Adenilate Kinase
Amylase
Xylanase
SeqId %: 19.61

Factores estructurales en la adaptacion al frio
• Agregados de residuos de glicina (proveen mobilidad local)
• Disminucion el contenido de prolinas en loops y turns
(proveen flexibilidad entre estructuras secundarias)

• Disminución del número de argininas (disminucion Arg/(Arg+Lys))
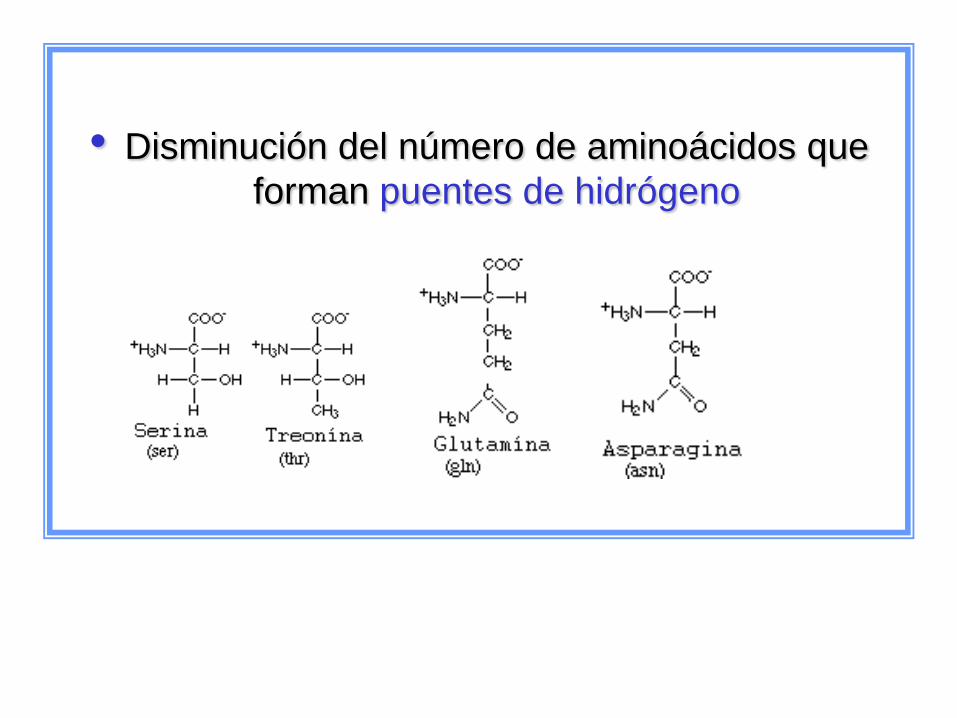
• Disminución del número de aminoácidos que
forman puentes de hidrógeno
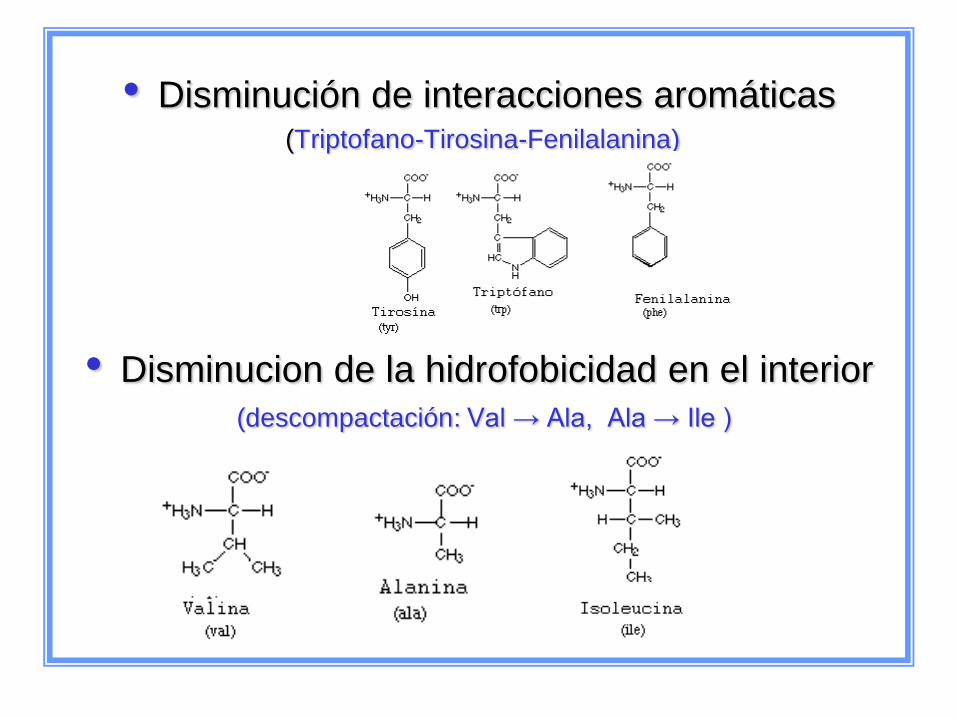
• Disminución de interacciones aromáticas (Triptofano-Tirosina-Fenilalanina)
• Disminucion de la hidrofobicidad en el interior
(descompactación: Val → Ala, Ala → Ile )

• Exceso de cargas negativas en la superficie
(cambio significativo del pI) (aumento de Aspartato)








![Cinética enzimática - Apache2 Ubuntu Default Page: It …ufq.unq.edu.ar/.../Teorias/T8-Cinetica-enzimatica.pdfActividad enzimatica •D istintas [E] •D istintas [S] •D istintos](https://static.fdocuments.ec/doc/165x107/5adda2867f8b9aeb668d2fab/cintica-enzimtica-apache2-ubuntu-default-page-it-ufqunqeduarteoriast8-cinetica-.jpg)
![Modulo2 1er2012.ppt [Modo de compatibilidad]ufq.unq.edu.ar/Docencia-Virtual/FQblog/Noelia/Modulo2_1... · 2015. 6. 29. · Más sobre variaciones de G dG SdT VdP Si no hay cambios](https://static.fdocuments.ec/doc/165x107/609e233236e5b474a2464cb9/modulo2-modo-de-compatibilidadufqunqeduardocencia-virtualfqblognoeliamodulo21.jpg)








