DE 1148.206.53.84/tesiuami/UAM7857.pdf · El descubrimiento de que las bases xanticas (corno la...
47
/NOMBRE : TELEFONO PARTICULAR: MATRICULA: CLAVE: /CARRERA : TRIMESTRE: LUGAR DE ACTIVIDADES: FEICHA DE INICIO: FECHA DE TERMINACION: NOMBRE DEL TUTOR: TITULO: LAURA FEIJOO SABORIT, 554-27-69 83237709 BIOLOGIA 8-9-1 (Crgditos Concluidos) Laboratorio de Genética Evolutiva. UAM - Iztapalapa 1 Diciembre 1987 15 Abril 1989 Biol. Alma G, Arellano M. Prof, Asistente "B",M.T. Depto. Ciencias de la Salud Aeterminación de la dura- .. . ción del ciclo celular en células de cebolla, Allium . cepa,- L. It \ LAURA FEIJOO ~ABORIT BIOL. ALMA G. ARELLANO MENESES
Transcript of DE 1148.206.53.84/tesiuami/UAM7857.pdf · El descubrimiento de que las bases xanticas (corno la...

/NOMBRE :
TELEFONO PARTICULAR:
MATRICULA:
CLAVE:
/CARRERA :
TRIMESTRE:
LUGAR DE ACTIVIDADES:
FEICHA DE I N I C I O :
FECHA DE TERMINACION:
NOMBRE DEL TUTOR:
TITULO:
LAURA FEIJOO SABORIT,
5 5 4 - 2 7 - 6 9
8 3 2 3 7 7 0 9
BIOLOGIA
8 - 9 - 1 ( C r g d i t o s C o n c l u i d o s )
L a b o r a t o r i o de G e n é t i c a E v o l u t i v a . UAM - I z t a p a l a p a
1 D i c i e m b r e 1 9 8 7
1 5 A b r i l 1 9 8 9
B i o l . A l m a G , A r e l l a n o M. P r o f , A s i s t e n t e " B " , M . T . D e p t o . C ienc ias d e l a S a l u d
A e t e r m i n a c i ó n d e l a d u r a - . . . c i ó n d e l c i c l o c e l u l a r e n
c é l u l a s d e c e b o l l a , A l l i u m . c e p a , - L. It
\
LAURA FEIJOO ~ A B O R I T
BIOL. ALMA G . ARELLANO MENESES

c, B
M é x i c o , D . F . 1 7 a b r i l 1989
M. EN C . BEATRIZ SILVA TORRES
D.C.B. S. P R E S E N T E
.. . S e s . r e t a z U h i c a
P o r m e d i o d e l a p r e s e n t e me p e r m i t o i n f o r m a r
a V d . q u e he. c o n c l u i d o m i S e r v i c i o S o c i a l , p o r l o q u e p r e -
s e n t o u n i n f o r m e s o b r e e l p r o y e c t o t i t u l a d o " D e t e r m i n a c i ó n
d e l a d u r a c i ó n d e l c i c i o c e l u l a r e n c é l u l a s d e c e b o l l a ,
A l l i u m c e p a , L.", q u e se r e a l i z ó e n e l L a b o r a t o r i o d e Ge-
n é t i c a E v o l u t i v a d e l a UAM I z t a p a l a p a .
M i a s e s o r a e n . d i c - h o p r o y e c t o f u e l a B i o l ,
A l m a G. A r e l l a n o M e n e s e s , P r o f . A s i s t e n t e "B", M e d i o
T i e m p o , d e l D e p t o . d e C i e n c i a s - d e l a S a l u d . '
A g r a d e c i e n d o d e a n t e m a n o l a a t e n c i ó n q u e
s e s i r v a p r e s t a r a l a p r e s e n t e , q u e d o d e Ud.
A t e n t a m e n t e
L a u r a F e i j ó o S a b o r i t Mat. : 8 3 2 3 7 7 0 9 Carrera : B i o l o g í a

Méxic0,D.F. 1 7 abril 1 9 8 9
M. EN C. BEATRIZ SILVA TORRES Secretaria Académica D.C. B. s. P R E S E N T E
Por este conducto me permito informarle que el proyecto de mi Servicio Social, titulado "Determinación de la duración del ciclo celular en células de cebolla, Allkum cepa, L.", se realizó en un tiempo mayor que el previsto de- bido a circunstancias fuera de mi control. La fecha de ini- cio fue el 1 de Diciembre de 1 9 8 7 , siendo la fecha prevista de terminación el 2 de Junio de 1 9 8 8 . Sin embargo, hubo una huelga y se remodeló el Laboratorio de Genética Evolutiva de esta Unidad, en el que se realizó el mencionado proyecbo. Por lo tanto, la fecha de terminación fue el 1 5 4e abril de 1989 .
b
. -
Agradeciendo de antemano la atención que se sirva prestar a la presente, quedo de Ud.
Atentamente
Laura Peijóo Saborit Mat.: 8 3 2 3 7 7 0 9 Carrera: Biología
. - "

INT-CION
- - -" _- *"""
La divisifin celular 5e considera -sensu strictom* como el
proceso mediante el cual una celula da lugar a otras dos. Este
proceso incluye tanto la divisibn nuclear (mitosis o
cariocinesis) como la d i v i s i h citoplasmatica. tcitocinesisl.
'Sensu lato', la divisibn celular se identifica con el proceso
completo de proliferacibn celular, incluyendo el crecimiento
durante la interfase y la divisibn en si. tGim&nez-Martin,ef al,
1977) . Para que una celula pueda dividirse, debe haber cumplido una
serie de requerimientos previos, los que estan asociados con la
duplicacibn exacta de la informacibn geneticia de la celula, asi
como una duplicacibn menos precisa de sus otros componentes.
(Gim&nez-M.,& aL,1977). Estos requerimientos premitbticos, junto
con la mitosis en si, conforman un proceso peri6dico que esta .
constituido por cuatru fases principales, denominadas de acuerdo
con una convencibn sugerida en 1953 por Howard y Pelc, de la
siguiente manera: e1 lapso entre el termino de la mitosis y el
comienzo de la replica del acid0 desoxirribonucleico (ADN) es la
fase G l i el periodo de replica del ADN se denomina periodo S; el
lapso que separa el fin de la rPplica del ADN del inicio de la
mitosis es la fase 62; y el tiempo dedicado a la dritosis es la
fase M. Se denomina entonces =inter+asem a 10 5 periodos Gl, S y
62 en conjunto, es decir, al periodo del ciclo en que no hay
mitosis. tAvers,1983).
1

2 Estas cuatro fases constituyen la porcibn nuclear del ciclo
celular. En la mayor parte de los sistemas eucaribticos existe
una divisibn concomitante de la celula, mediante la cual los
nuevos n&cleO5 son incluidos en sus propios limites celulares;
separados. Esta divisibn comienza normalmente durante la etapa de
telofase de la mitosis. Aunque no es una parte iptegral de la
mitosis misma, la citocinesis normalmente esta sincronizada con
los eventos mitbticos. (Gimenez-M.,Ek al,l977).
- .. - - - - - I_ . .
Se ha observado que los nkleos normalmente no entran a la
mitosis a menos que haya habido previamente replicacibn del ADN
cromosbmico, lo que ocurre durante el periodo S de la interfase.
La replicacibn del ADN produce el material genetic0 nuevo para la
siguiente qeneracih 'celular. La mitosis es el mecanismo para
distribuir este material a las cPlulas hijas.
Durante la mitosis se observan cuatro fases,
apreciarse distintivamente en observacibn directa al .-
de luz.
El primer signo evidente de mitosis que seirala el
que pueden
microscopio
comienzo de I
la profase es la condensacibn de los cromosomas (Foto 1) . A
medida que transcurre esta etapa los cromo~omas se hacen mas
cortos y gruesos; hacia el term'ino de la profase 105 cromosomas
contraidos se dirigen al plano ecuatorial del huso, dando
entonces lugar a la metafase (Foto 2) En esta etapa cada
cromosoma es una estructura replicada constituida por dos
cromhtidas cuyos centrtbmeros miran hacia polos opuestos de la
celula. La metafase es seguida por la anafase (Foto 3lS que
comienza cuando las cromatidas de cada cromosoma se separan. Las - .
, ..
cromatidas hermanas se mueven hacia polos opuestos de la celula,

3
Ya W e SUS centremeros se encontraban alineados de esta manera
durante la metafase. Cuando las cromatidas se separan, cada una
de ellas se transforma en un cromosoma integro e independiente.
Una vez que los cromosomas han llegado a sus respectivos polos
comienza la telofase (Foto 4 ) . Durante esta etapa .Cirial, los
cromosomas condensados comienzan a descondensarse y a adquirir
gradualmente la apariencia que. tenian durante la .-interfase. En
esta etapa ocurre la reorganizacibn nuclear, que incluye la
organizacibn de nuevos nucleolos y envoltura nuclear. (Avers,
1983).
" - " "i__ - . ""
..
Los inhibidores de los procesos que ocurren en la cClula
mientras transcurre el ciclo celular han permitido comprender
algunos de los rnecanismcrs de control que operan durante la
mitosis (especialmente en la profase), y el estado de
reorganizacibn nucleolar durante la telofase.
La aplicacibn de inhibidores en la solucibn de problemas
biolbgicos ha conducido a descubrimientos definitivos en cuanto a
la estructura y funcibn de los organismos vivos. El estado vivo
se caracteriza por una serie de reacciones enzimaticas que
ocurren a nivel celular, algunas simultaneamente y otras en
secuencia. Los inhibidores son muy &tiles cuando estas reacciones
resultan diflciles de separar unas de otras. Asl,. los inhibidores
especlficos de un proceso dado, especialmente aquellos cuyos
mecanismos de accibn a nivel molecular son conocidos, son muy
valiosos para establecer el papel que juega cualquier proceso en
particular en la fisiologla celular. ..
-.
"

4
El descubrimiento de que las bases xanticas (corno la cafeina,
por ejemplo) ejercen un efecto inhibitorio sobre la citocinesis,
ha permitido inducir la fornacilrrn de cPlulas binucleadas (Foto 5)
y polinucleadas en meristemos de raiz a voluntad. Este tipo de
c&lulas ofrece grandes posibilidades para el estudip de aquellos
mecanismos que gobiernan la regulacih genetica en cPlulas
."" . .
eucaribticas. En la5 plantas, el U50 de celulas polinucleadaa
parece ser una herramienta adecuada para la comprensibn de
mecanismos de proliferacibn, especialmente aquellos involucrados
en interacciones nacleo-n&cleo y n&cleo-citoplasma.
Bajo condiciones estables, la poblaci&n meristematica de una
ralz esta distribulda uniformemente a lo largo del ciclo celular.
La cafeina inhibe la citocinesis bloqueando 1a.fornacibn de la
pared celular en aquellas calulas espontaneamente sincrbnicas que
se encuentran en la telofase durante el tratamiento, por lo qua
se puede producir de esta manera una poblacibn de cPlulas
binucleadas. Esta constituye entonces una subpobracibn marcada y
puede distinguirse facilmente del resto de las cClulaa
meristematicas usando criterios morfolbgicos. Esta poblacibn
sincrbnica, por tanto, ha sido obtenida por sincronla
seleccionada y no inducida, y muestra un ciclo similar al de las
cClulas mononucleadas.
Para inducir la forrnaciBn de cClulas polinucleadas, la
poblacibn binucleada puede 5er tratada con cafeina una vez mas
durante su siguiente telofase (bi-telofase). Este segundo
tratamiento da lugar a celulas con un complemento cromosbmico 8n
que puede distribuirse en 2, 3 b 4- nbcleos dependiendo del nbmero . -
de fusiones nucleares. Ademas las celulas bi y polinucleadas
i

5
sirven como base para estudiar e1 desarrollo asincrbnico que
ocurre al inicio de la mitosis en diferentes nkleos presentes en
el mismo citoplasma de cglulas animales. En chlulas de cebolla se " - " - ".""
observ& que los n&cleos rodeados por un citoplasma mayor entran a
prof ase antes.
En cuanto a la divisihn celular de estas celulas bi y
polinucleadas, hay evidencia de la iniciacihn sincrhica de la
replicacih del ADN en los diferentes n&cleos que comparten el
mismo citoplasma, lo que apoya la hipbtesis de que existe un
factor citoplasmatico que controla el paso de G1 a S.'
Ya iniciada la replicacibn del ADN, 5e ha observado asinrronia
en la tasa de replicacibn del mismo en dos nacleos compartiendo
el mismo citoplasma en rPlulas binucleadas .de cebolla. Se
detectaron tambien diferencias en la tasa de replicacihn del ADN
durante el periodo S, con una disminucihn de la tasa
aproximadamente a la mitad del periodo.
En varias especies vegetales se ha observado un patrhn
especlfico para la replicacibn del ADN en cromosomas individuales
y en diferentes segmentos tromosbmicos, lo que indica, que el
tiempo ocupado por el periodo S no es otra cosa que la suma del
tiempo de replicacih de la5 diferentes porciones del genoma.
Evidencias experimentales muestran que la terminacibn del periodo
S en los diferentes n&cleos de una celula polinucleada no es
necesariamente un evento sincrbnico, y que los n&cleos no
interacttian de forma aparente entre ellos. (Girenez-tl.,eL a l r
1977) .

-
6
I Existen factores internos que modifican el tiempo necesario I
1
1 ! par a que se satisfagan los diferentes requerimientos
prenitbticos. Por ejemplo, 5e ha reportado una mutacibn que
elimina el requerimiento de replicacibn cromosbmica para que se
presente la mitosis. Esto confirma que la duplicacibn del genoma
y la mitosis pueden ser eventos independientes. Se ha mostrado
tambiPn que cuanto mayor sea el contenido de ADN, mayor sera la
duracian del ciclo celular. Estudios sobre el tiempo de interfase
en cPlulas polinucleadas confirman que el ciclo es mas largo
cuando la ploidia o el ndmero de ndcleos en la cPlula aumentan.
Ademas, el grado en el qua el ciclo mit&tico se alarga al
aumentar el contenido de ADN e5 menor en cPlulas poliploides que
.. - " - - __ "~ ",
.
. . en diploides..
Hay tambiPn
requerimientos
frecuentemente
pueden crecer
factores externos que modifican el tiempo para lor
premitaticos. Las ce!tulas vegetales se -ven
influenciadas por la temperatura, a pesar de que
a cualquier temperatura desde O hasta 3 5 O C. La i
progresien del ciclo completo es dependiente de este factor, lo
cual es de esperarse, ya que este tipo de .c&lulas - .. pueden
considerarse como 'poiquilotermas'. (L&per-Saer,& aL,1966).
I ! !
El estudio de la mitosis en celulas polinucleadas muestra que
al comienzo de la misma todas las profases son asincrbnicas,
mientras que la frecuencia de asincronla' disminuye
progresivamente en el tiempo. A partir de la retafase, todos IDS
nrlcleos son invariablemente sincrbnicos,
Se observa tambih un aumento en-el tiempo de sincronizacibn
al aumentar el ndmero de ndcleos por celula, lo que puede deberse

7
a que los nacleos =lentosm produzcan un inhibidor que evite que
los 'rapidos' entren en metafase. En .una calula nononucleada, la
importancia biolbgica de este mecanismo puede 5er que explica el
hecho de que todos los cromosomas terminen la profase
simultheamente, a pesar de que la duplicacibn firevia haya sido
asincrbnica, e incluso si se presenta una condensacibn
asincrbnica de cromo soma^ durante la profass.
- " I_ " ~
Durante una telofase normal, la reconstruccibn nuclear se
lleva a cabo gracias a un proceso que puede considerarse el
.reversoD de la profase. Se pueden observar dos eventos
morfolbgicos principales:
1 ) Fornacibn de membrana nuclear.
2) Descondensacibn cromosbmica.
Se piensa que son la base estructural para recomenzar los
patrones de transcripcih y replicacibn del ciclo, ya que en
Pstos se basa el que la celula recien formada entre de lleno en
la condicibn de interfase activa.
La descondensacit3n cromos&mica que ocurre durante la telofase
es el punto de partida para la iniciacibn de nueva actividad en
la nueva celula. En general, se piensa que la transcripcih de
genes no es compatible con la condensacih cromoshmica, aunque el
tPrmino del ciclo de descondensacibn cromosbmica no es en si
mismo el estlmulo inicial de la transcripcibn.
En una forma similar a la interfase, el tiempo mitbtico esta
claramente influenciado por factores externos e internos.
Estudios sobre la relacih entre la duracibn del ciclo y el I
contenido de.ADN sugiere que existe una correlacibn linear entre

8
ellos, siendo modificadas proporcionalmente todas las fases
Todo proceso biolbgico presenta una velocidad maxima de
desarrollo, asi como una 'temperatura bptina, , sobre la cual y
debajo de la cual el proceso es mas lento. El ciclo de divisien
celular es lauy complejo, conformado por toda una serie de
reacciones diferentes, y cada una de estas fases tiene su propia
temperatura eptima. (Lbpez-SAez,ek &,1966). El tiempo en que se
lleva a cabo el ciclo celular aumenta al disminuir la
temperatura, pero la duracibn relativa de las fases se mantiene a '
todas la5 temperaturas. (Gim&nez-tl.,rrf. &,19775 Lbpez-S&ez,rrf. al, 1966). Otro factor del que depende la duracibn del ciclo es la
tensi-bn de oxlgeno, aumentando la tasa del ciclo celular al r
aumentar la concentracibn de oxageno burbujeante hasta un 20%. .
Varios autores (Brown,l95l; Evans y Savage,l959; Van't Hoff y
Sparrow,l963; y Van't Hoff y Ying,lP64) Ran mostrado por
<'
diferentes nrPtodos que en vegetales la velocidad del ciclo de
divisian celular aumenta con ia temperatura hasta alcanzar su
maxim0 a los 25'C o 3OOC. Por otro lado, 5e ha demostrado tarnbien -. .-
la influencia de una temperatura especi+ica en ciertas etapas del
proceso, la cual modifica su velocidad relativa dentro de todo el
ciclo. (L&pez-SAez,rrf. al,1966). Evans y Savage (1959) mostraron
en el caso de Yicia que la metafase es la etapa de la
mitosis mas sensible a los cambios de temperatura- Sin embargo,
Lepez-Saez al mostraron en 1966 que los indices de fases
permanecen constantes a todas las.temperaturas, por lo que los
resultados obtenidos por Evans y Savage pueden deberse al mPtodo
. .

9
que utilizaron para determinar la frecuencia de fases, ya que la
modificacihn relativa de la metafase que ellos encontraron es muy
pequeña, y probablemente se deba a factores aleatorios. --. .. - “ _ _ ~ _L ””
Los diferentes metodos utilizados para medir la d9racih del
ciclo celular en cblulas meristematicas involucran trabajo I
experimental largo y laborioso que hace- dificil repetir el i
proceso cada vez para diferentes condiciones, por ejemplo,
diferentes temperaturas. Por tanto, es recomendable elegir una
fraccihn del ciclo de facil estudio pero que a la vez arroje 4
resultados que no varien considerablemente de un experimento a
otro en condiciones similares. Este es el caso de la B-interfase.
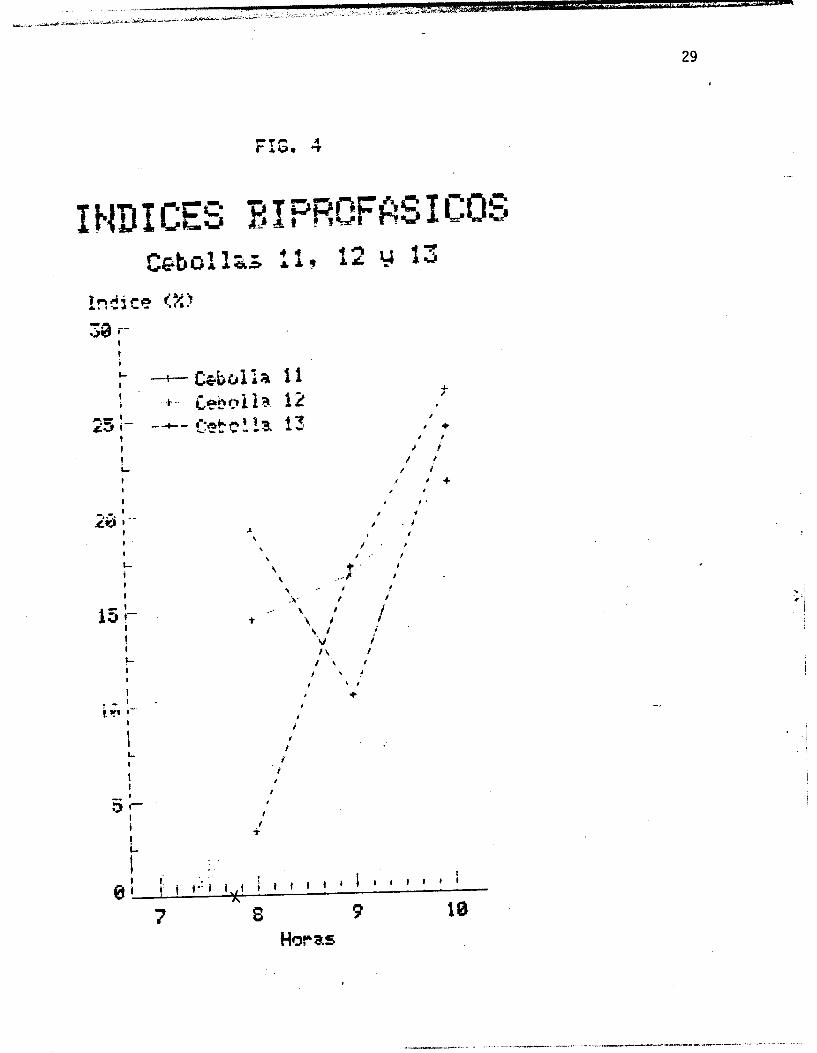
La B-interfase es el periodo que abarca desde la hora O (fin
dol tratamiento con cafejna) hasta ia aparicibn de las primeras
cPlulas binucleadas en biprafase (Foto 61. Su duracibn se calcula
tornando muestras de las raices cada hora y uniendo con una linea
recta las dos primeras lecturas positivas, de forma que corte el
eje de las abscisas. Este punto marca e1 periodo tebrico en que
todas las cblulas binucleadas estaban en interfase. El periodo
representa de hecho la interfase de aquellas cPlulas binucleadar -.
que recorrieron el ciclo mas rapidamente, y es una fraccibn mas
largo que la mitad del ciclo celular completo. Si su duracihn
relativa, como una parte proporcional del ciclo completo, es
constante a diferentes temperaturas, se tendrt un rnPtodo simple y
rapid0 para calcular la duracibn del ciclo bajo condiciones
dadas.
La duracien de la B-interfase est-& fuertemente influenciada
por las diferentes temperaturas de crecimiento. Dentro del amplio

10
rango de temperatura entre 0"C y 40°C, la temperatura bptima ! I
no muestran crecimiento significativo, aunque a ninguna de las
dos temperaturas es fatal el tratamiento.
La proliferacibn de celulas a las que da lugar el crecimiento
de las raices se lleva a cabo sin ningan cambio en el ndmero de
celulas neristematicas, que permanece practicamente constante por
varios dias despuk de la gerninaci6nl Mas adelante, cuando el
experimento se alarga de 4 a 6 dias a 25OC, el nbmero comienza a
decrecer poco a poco. Bajo estas condiciones, el niimero de
cPlulas meristematicas no aumenta .logarltmicamente como resultado
del ciclo de divisibri celular; ya que aunque' el nihuero de
celulas, maternaticamente hablando, sl aumenta a este ritmo, en
cada . ciclo de divisibn celular que da lugar .a' dos celulas hijas,
5blO una de ellas permanece como celula meristemktica, mientras
que la otra sufre diferenciacibn. Por lo tanto, el Indico
mitbtico parcial, definido 'como el porcentaje de cielulas
meristematicas en divisibn en un momento dado, e5 directamente
proporcional a la duracihn de la mitosis en relaci&n a la del
ciclo. Asi, el indice se mantiene constante para cada una de las
temperaturas a las que 5e realice el experimento. Lo mismo sucede
con los indices de fases, tomados como el porcentaje de C&lUlaS
en proceso de divisibn que se encuentran en las diferentes fases
de la mitosis. Como el indice mit&tico, el indice de fases
permanece constante a las diferentes temperaturas estudiadas.
1
-,
!

11
Se puede decir que el lndice sitbtico parcial expresa la
duracibn relativa de la mitosis en el ciclo. Por lo tanto, esa
duracih 5e puede expresar como un porcentaje, sin importar la. -:"
temperatura, siendo la intcrrfase el porcentaje restante para
completar el l O O X , es decir, el ciclo completo. Lo mismo se puede
aplicar para el indice de cada una de las fases, pudiendo
calcularse de esta manera la duracien de todo el ciclo, asi cono
la de cada fase. tL&pez-Saez,et u, 1966) . Existen otras tOcnicas para determinar la duracibn del ciclo.
Una de ellas es la cuantificacih del fujo celular, ya que Pste y
la duracibn del ciclo son inversamente proporcionales. (De la
Torre,l971). Sin embargo, este metodo involucra el conocimiento
de¡ porcentaje de cPlulas que pasa a traves de .cualquier fase del t
ciclo en un tiempo determinado, lo que implica el uso de tknicas
de autorradioqraf la, que son muy caras y complicadas.
Otro de estos metodos es el propuesto por Pincu ek al (1986) , que, trabajb con linfocitos humanos, pero puede ser efectivo
tambien en el caso de celulas vegetales. Esta tPcnica se basa en
la utilizacien de altas concentraciones de brornodeoxiuridina
(BrdU) seguida por la tecnica de tinciBn propuesta por Perry y
-,
. ,
Uolff (19741, que tiRe diferencialmente 10s ndcleos de las
celulas que se encuentran en diferentes estados de.proliferaci&n-
De esta forma es posible determinar las frecuencias relativas de
las celulas en mitosis. Como es evidente, este metodo involucra
trabajo laborioso, asi como la utilizacihn de sustancias caras y
de manejo delicado. "

12
Asi, el matodo de marcado con cafeina presenta muchas
ventajas, ya que es relativamente rapid0 y simple, proporciona u n ' .
alto grado de reproducibilidad, no es C O ~ ~ O I O y no involucra la .. .. - " ~ ".
utilizacian de sustancias para marcado radiactivo.

13
FOTOORAFIAS
.-
FOTO l. PROFASE
0
FOTO 2. METAFASE

14
. .
FOTO 3. A W A S E
FOTO 4. TELOFASE

15
FOTO J. CELULA BINUCLEADA
FOTO 6. CELULA EN BIRPOFASE

16
OBJETIVOS
- " - . _" . . . ."
Con base en los antecedentes e informaa'ih sobre el ciclo
celular, particularmente la mitosis en celulas vegetales, se
plantea el siguiente m -:
- Determinar la duracih del ciclo celular, y de cada una de
5us fases, en c+lulas neristematicas'de raiz de Eetbolla,
-, L., bajo las condiciones de trabajo en el Laboratorio de
Genatica Evolutiva de la UAM-Irtapalapa.
. . Para lograr el cumplimiento efectivo dei objetivo anterior, se
plantean los siguientes aid i v p q -ref:
1 ) Determinar la duraci&n de la B-interfase, segh la tecnica 7
propuesta P por Lbpez-Saez (19661, utilizando una temperatura de
crecimiento de 25OC. "
'e' ,
2) Determinar los indices mitbtiios y de fase &.meristemos de
cebo1 la (tratados con caf eina) . 3) Determinar la duracibn. relativa de cada fase (interfase y
mitosis), por medio de los lndices de fases.
4) Determinar la duracih absoluta de cada fase con base en la
duracih de la B-interfase y en las duraciones relativas de la5
fases. ' .
5 ) Determinar la duracihn total del ciclo a 25*CC$.:

MATERIAL Y METODO
17
"_.___
Se pusieron a germinar -¡¡ bulbos de c e ~ l ~ ~ c o m d n Allfun -, en dos lotes, y se controlaron los parhetros para su
crecimeinto. La temperatura 5e mantuvo a 25OC. Para esto, se
colocaron los bulbos del primer lote (cebollas NO. 1 1 , 12, 13,
14, 15 y 161, dentro de una estufa de germinacien, y esta a SU
vez dentro de un cuarto fr¶o. Esto se hizo con la finalidad de
que la temperatura del laboratorio, generalmente mayor de 2 5 O C,
no afectara la temperatura de crecirniento.de los bulbos. En el
caso del segundo lote (cebol.las No. 17, 18, 19, 24 y 211, no fue
necesario utilizar el cuarto fria, ya que la temperatura se
mantuvo constante dentro de la estufa' a 25% por haberse tratado
estos bulbos en invierno y ser la tewperatura dei laboratorio
menor de 25%. Las cebollas se compraron siempre en el mismo
?
puesto del mercado de Iztapalapa, esperando que de esta manera no
variaran significativamente sus condiciones de cultivo y
crecimiento antes del experimento. Se comenzb a numerar las
2' ;
cebollas =,partir del No. ' 11 y no del No. 1 debid0.a que ya se
habia iniciado este estudio, y se considera adecuado continuar la
rturneracibn ya existente.
La tensibn de oxigeno se mantuvo con un burbujeo constante
de aire de aproximadamente una burbuja por segundo. Ademas, 5e
cambib diariamente el agua de I 0 5 recipientes eh que germinaban
los bulbos. La germinacibn durb 5 dias en el caso del primer lote
y 3 dlas en el caso del segundo. Transcurrido este tiempo, las
raices ya tenlan un tamaño adecuado para su tratamiento (entre 15
y 20 mm. .de longitud).
- .

18
Las raices fueron entonces tratadas con cafeina al 0.1%
durante una hora, reincubandose despu&s en a w i e n t e con
aire burbujeante. Finalizado el tratamiento con cafeina se
tomaron muestras de las raices a intervalos de una hora; desde la
- ” I_ ”, ”
hora 7 hasta la hora 10 en el caso del primer lote, y desde la
hora 8 hasta la hora 10 en el caso del segundo, ya que los
resultados obtenidos con el primer lote, asi comd‘los arrojados
por la primera etapa del estudio (cebollas 1 a 101, mostraron que
en la hora 7 no habia todavia ninguna clSlula que se presentara en
biprofase. Se tomb como hora 0 el fin del tratamiento con la
cafeina. Se obtuvieron entre 3 y 5 raices por bulbo cada hora,
fijandolas en una solucibn de acido aceticrr’y- etanol absoluto
1:3. (Segdn Lbpez-SAez,& &,1966).
Se’ procedib entonces a elaborar preparaciones.de las raices
por el rnPtodo convencional de aplastado con orceina
acetoclorhidrica. Este rnetodo consiste en colocar las raices en
un vidrio de reloj, cubrihdolas con orceina acetoclorhidrica. Se
calientan despues sobre el mechero de alcohol, agitando levemente
el vidrio de reloj hasta observar vapor evitando la ebullician, y
dejando enfriar. Este procedimiento se repite tres veces, dejando
enfriar bien despuds del dltirno calentamiento. Enseguida se hacen
cortes de aproximadamente 1 mm. de largo de los meristemos de las
ralces, colocando el tejido sobre un portaobjetos limpio (una
raiz por cada portaobjetos), y cubriCndolo can una o dos gotas de .
orceina. DespuQs se coloca el . - cubreobjetos, realizando el I I
aplastado inicial con la punta de un lBpiz en movimientos
espirales de izquierda a derecha, y dejando girar libremente el
I

19
cubreobjetos sobre e1 tejido. El aplastado final se realiza con 1
I
el pulgar, ejerciendo presibn vertical de arriba .a. abajo. De esta
forma se'hicieron preparaciones de tres raices por bulbo por hora
i ~- . i - " __ ", - - 1
I Í
en el caso del primer lote, y de una r a i z por bulbo por hora en !
el caso del segundo. Las repeticiones en el primer caso se i
hicieron con el fin de afinar la tecnica, asi como obtener una !
mayor practica en la diferenciacibn morfolbgica de las diversas
fases de la mitosis. Asmimismo, los conteos se hicieron en ciego,
es decir, sin saber de antemano a que hora correspondia cada
preparacibn, de forma que los resultados obtenidos fueran
I
I
I
i ?
I
totalmente confiables y no se vieran influenciados por un
conocimiento previo de los valores esperados. i
. . j Una vez tratadas todas las raíces corno se indica arriba, se 4
procedi& a revisar las preparaciones en el microscapio de .luz, I registrando el nbmero de c&lulas que se encontraban en cada fase
del ciclo celular, asi como el de celulas binucleadas y en
biprofase. (Gim&ne~-Martin,l984).
*-
.

20
RESULTADOS
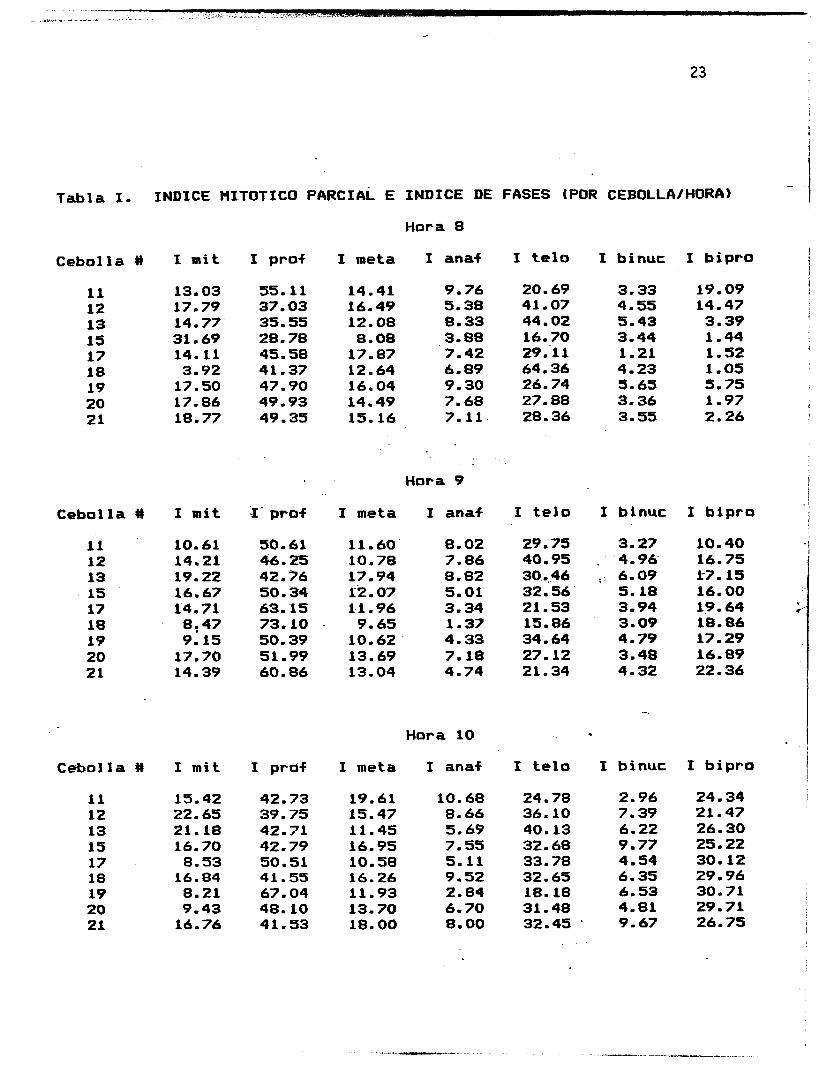
La Tabla I muestra los indices mitbticos parciales y los 1
Estos indices indican el porcentaje de celulas en.el meristemo que
se encontraban en cada una de las fases de la mitosis (o
simplemente en mitosis, en el caso del indice mit&tico parcial)
en el momento de la *f i.jaci&n de las ralces para la elaboracibn de
las subsecuentes preparaciones en las que 5ie llevb a cabo el
conteo celular. Estos indices fueron obtenidos. segbn lars
indices de fases obtenidos para cada uno de los bulbos estudiados. : !I
i/
:I
siguientes fbrmulas:
' No. de celulas en mitosis Indice mitCltico = x '100
Total de celulas .
. .
No. de celulas en cada fase
Total de celulas en mitosis Indice de fases = x 100
No. de celulas binucleadas
Total de cClulas Indice binucleadas = x 100
No. de celulas eib biprofase
Total de celulas binucleadas Indice biprofasico = x 100
I

21
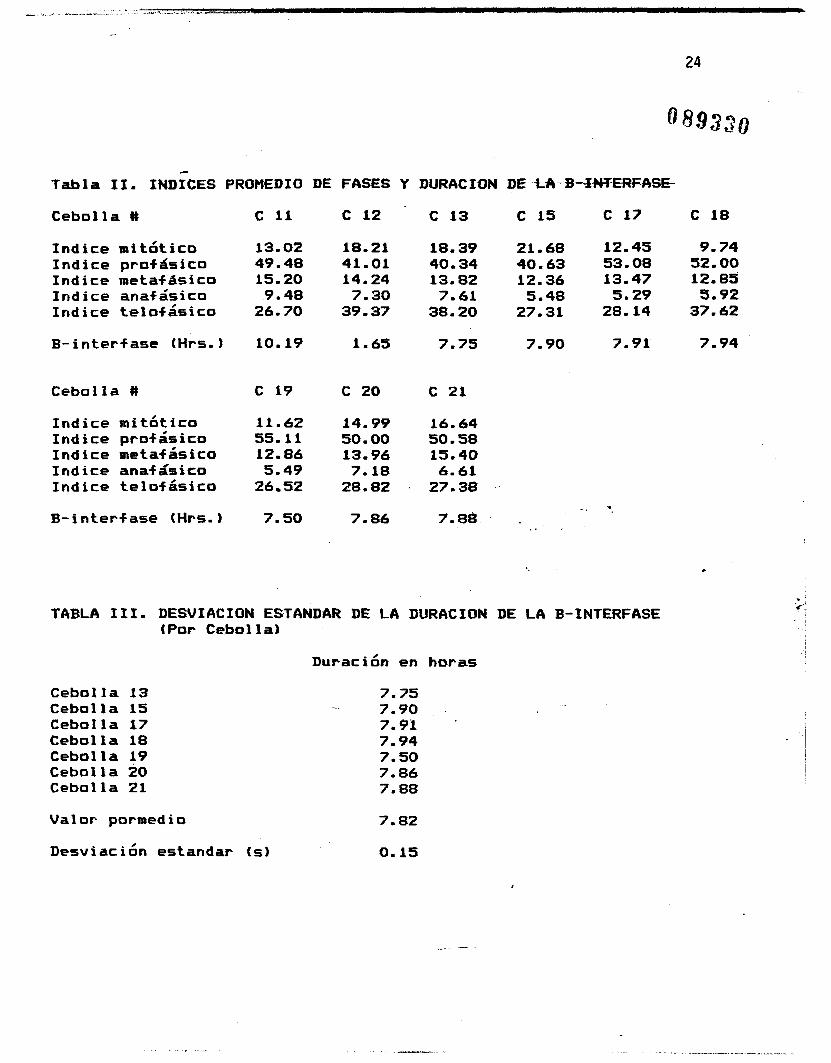
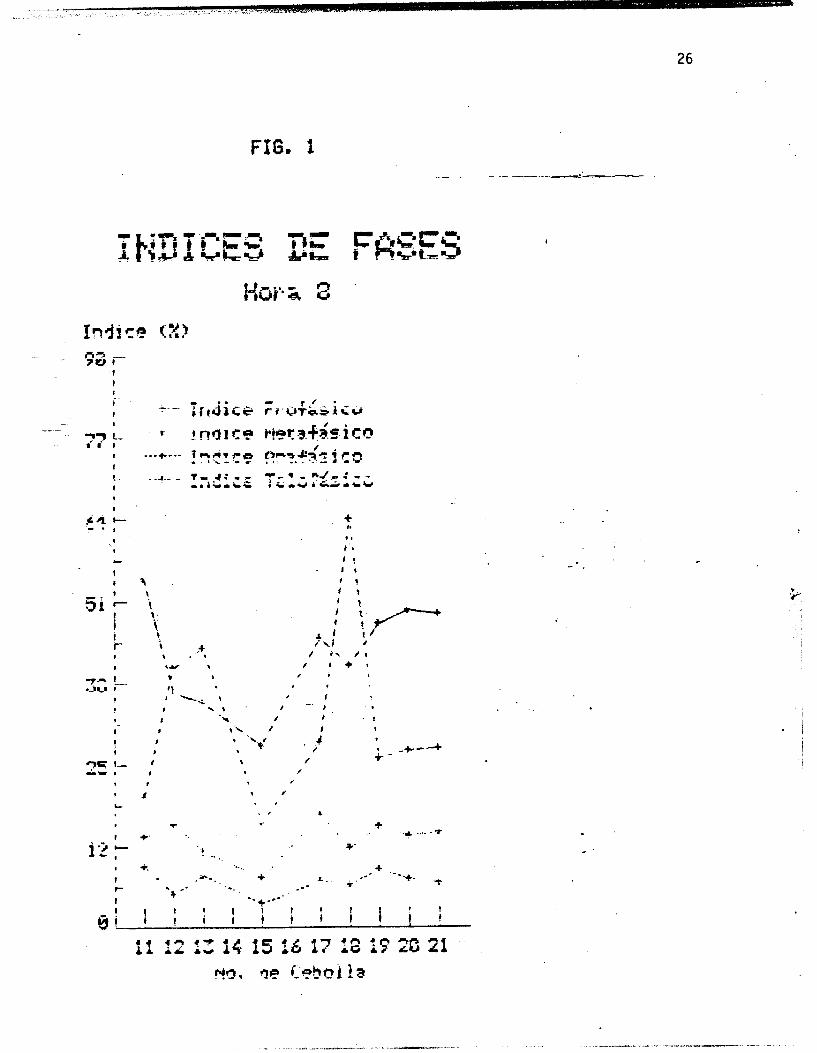
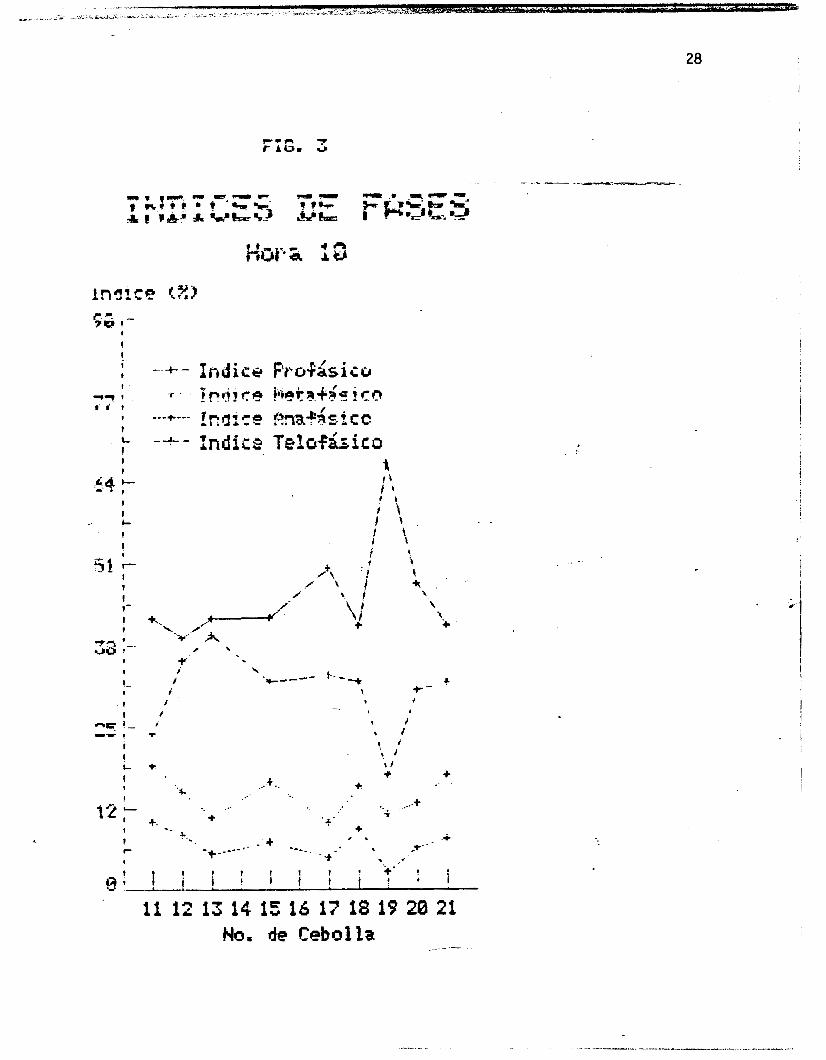
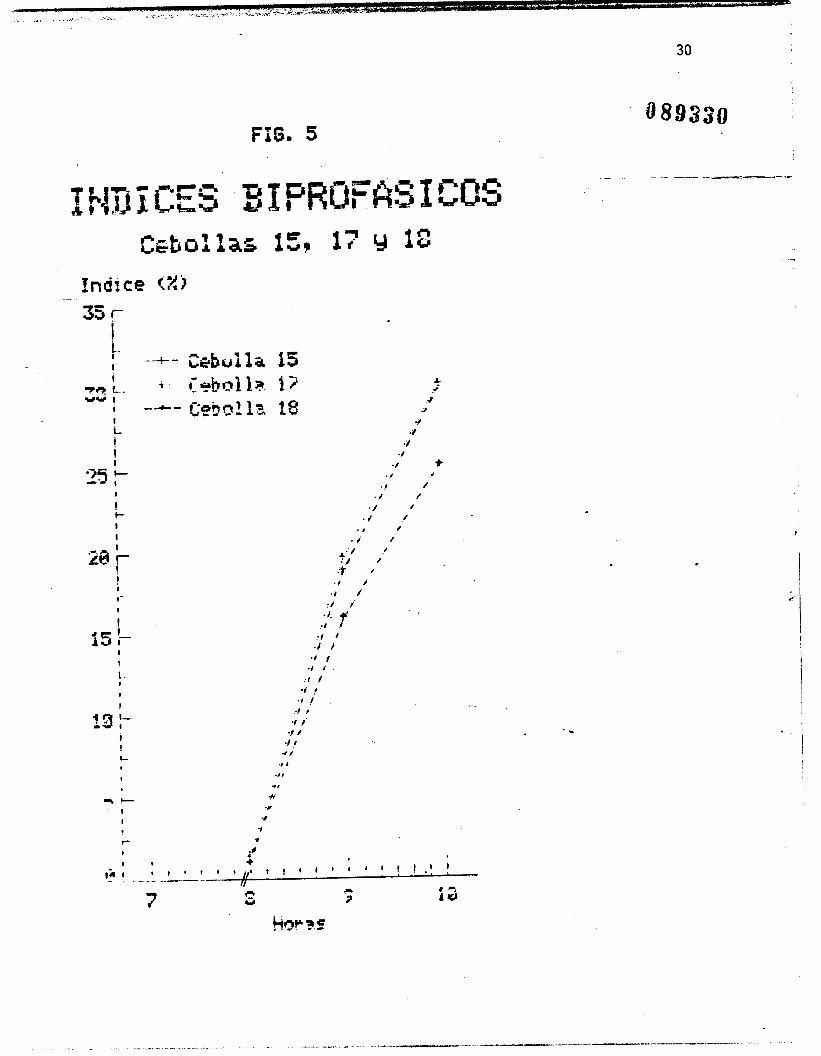
Las figuras I, 2 y 3 muestran los indices de fases por cebolla
esta linea hasta cortar el eje del abscisas, se obtiene un punto
que indica la duracien en horas de la B-interfase, Este calculo l
.~ se realiz& de forma gr8fica y utilizando las fhrmulas
correspondientes de la Geometria Analitica para hacer la
extrapolacibn y obtener asi resultados mas precisos. Los
resultados obtenidos se encuentran concentrados en la Tabla If.
Con el promedio de estos valores, 5e obtuvo un valor de 7.82
horas para la duracihn de la B-interfase (es decir, el periodo en
que todas las c+lulas binucleadas se encontraban en . . interfase).
La Tabla I1 muestra tambi.&n los valores promedio para el indice
mitbtico parcial y las indices de fases por cebolla, los que se
obtuvieron promediando los valores obtenidos para cada cebolla
cada hora. Estos valores promedio .,se encuentran tambien
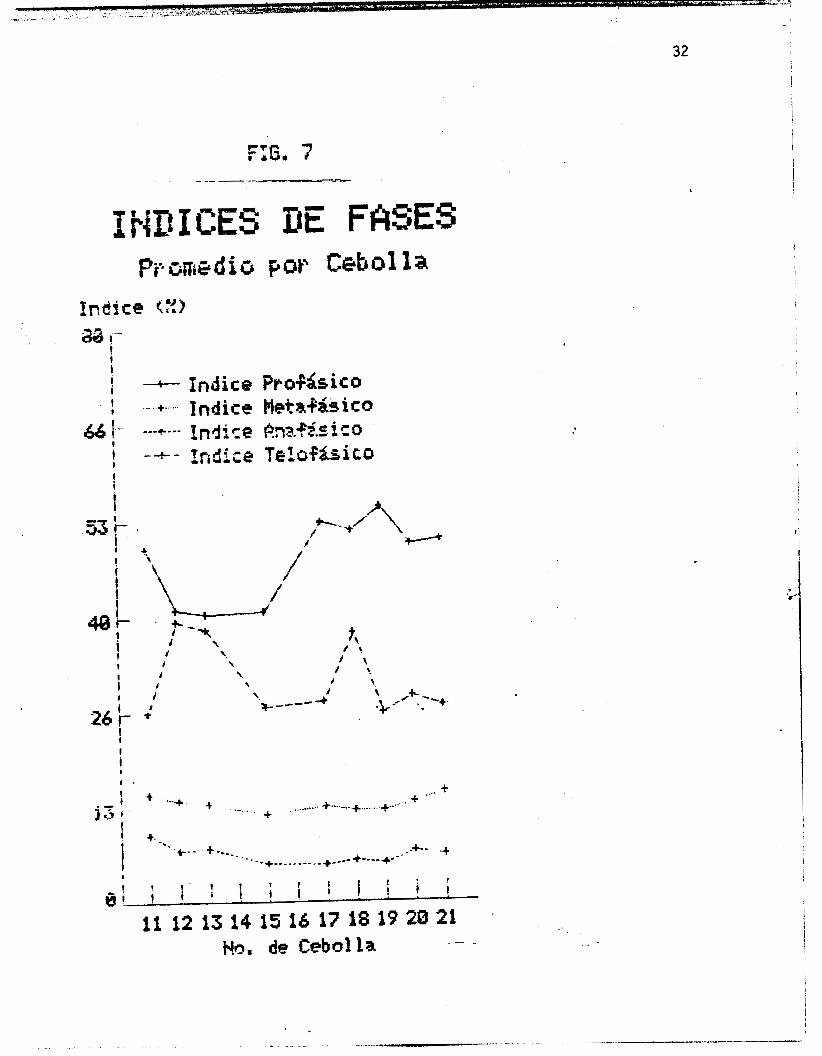
ilustrados en la figura 7, y representan la parte proporcional
de la mitosis ocupada por cada una de las fases. En la .Figura 8
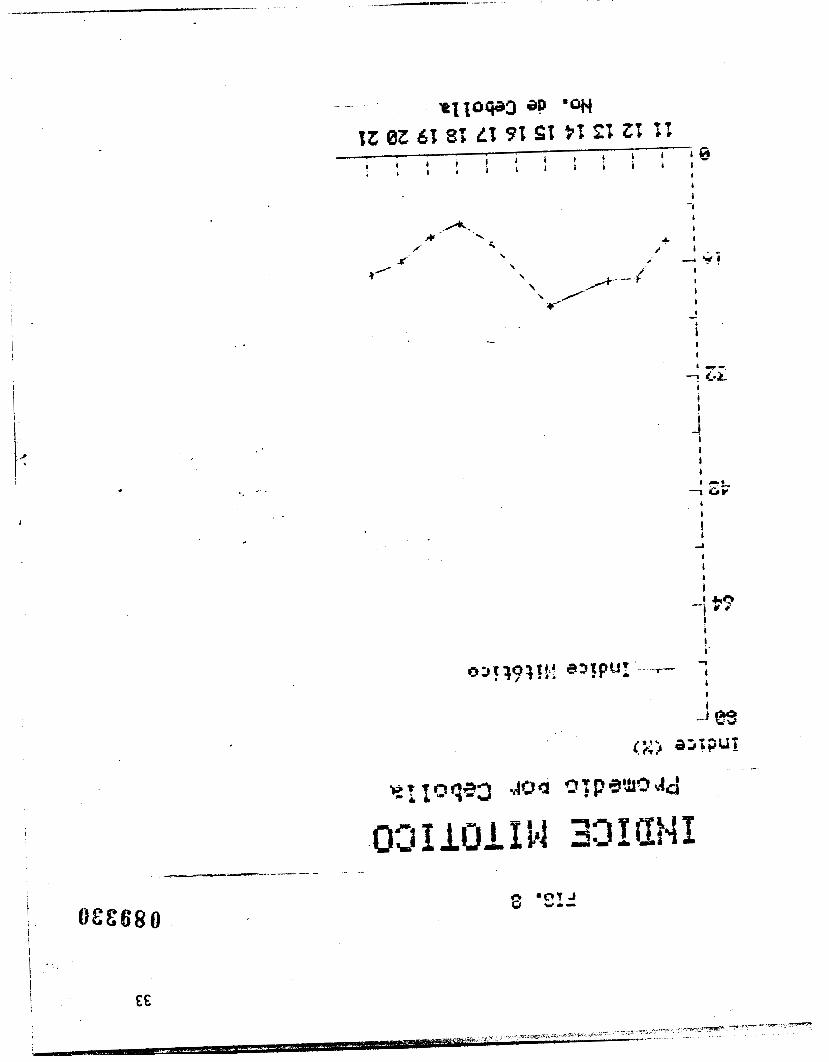
se ilustran los valores promedio para el lndice mitbtico, que
representan la parte proporcional del ciclo celular completo
ocupada por la mitosis.
El promedio global de los indices mitbticos representa la
parte proporcional del ciclo ocupada por la mitosis, es decir, el
19.19% del tiempo de duracibn total del ciclo celular. Por 10
tanto, las 7.82 horas que dura la interfase representan el

-.
22
porcentaje restante para completar el loo%, .es decir, el 84.81%
del ciclo completo, y el valor en horas representa el tiempo de .
duracibn del ciclo celular ocupado por Ea interfase. Asi,
" .. - " " ".
utilizando una simple regla de tres, se obtiene un valor de 1.4
horas para la duracibn de la mitosis. De la misma manera se
obtuvieron los valores en horas para la duracibn de cada fase de
la mitosis, tomando en este caso el valor. de 1.4 horas como el
100%, (es decir, la mitosis completa), y los promedios de los
lndices- de fases como l.as duraciones proporcionales de cada una
de ellas dentro de la mitosis. La Tabla I11 muestra la desviacian
estandar de la duracibn de la B-interfase (en horas), para la
que se obtuvo un valor de 0.15, es decir, la duracibn de la B-
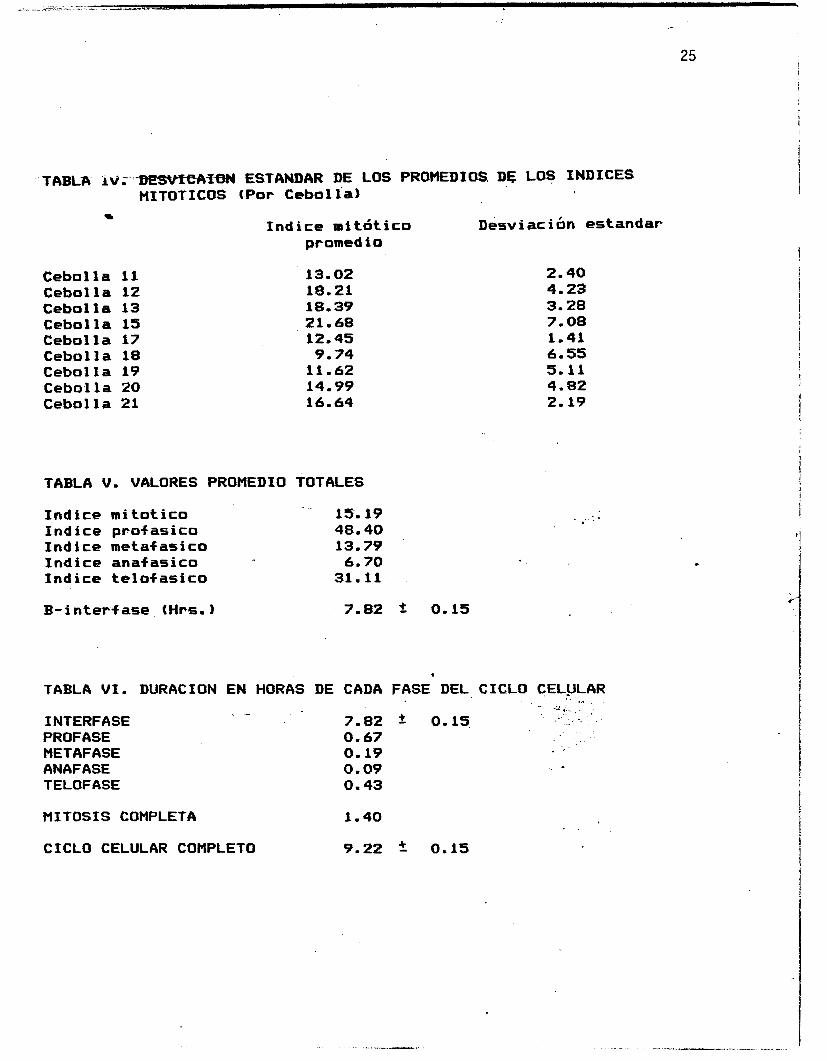
interfase es de 7.82k0.15 horae. En la Tabla I V se'observan las
desviaciones estandar de los promedios de los indices mitbticos
(por cebolla), las que se encuentran representadas graf icamente
en la figura 9.
Los valores promedio totales de los indices de fases y el del
indice mitbtico se encuentran concentrados en la Tabla V , donde
tambiPn se incluye el valor promedio 'de'la duracidn en horas de
la B-interfase, considerando su desviacibn estandar. Finalmente,
las valores en horas para la duracibn de cada fase de la mitosis,
a53 como de la interfase y de la mitosis completa, se muestran en
la Tabla.VI. En esta se incluye tambiPn el valor en horas del
ciclo celular completo, considerando la desviacien estandar
calculada para la interfase. Estos valores se ilustran en las
figuras 10, 11 y 12. En la figura 12 se muestran los valores
proporcionales de la interfase y de la5 fases de la mitosis "" -
referidos al ciclo celular completo.
23
Tabla I. INDICE IIITOTICO PARCIAL E INDICE DE FASES (POR CEBOLLA/HORA)
Hora 8
Cebolla
11 12 13 15 17 18 19 20 21
Cebolla #
11 12 13 15 17 18 19 20 21
Cebolla #
11 12 13 15 17 18 19 20 21
I m i t
13.03 17.79 14.77 31 . 69 14.11 3.92
17.50 17.86 18.77
I rnit
10.61 14.21 19.22 16.67 14.71 8,47 9.15
17.70 14.39
I m i t
15.42 22.65 21.18 36-70 8.53
16.84 8.21 9.43
16.76
I prof
95.11 37.03 35.55 28-78 45.58 41.37 47.90 49.93 49 . 3s
X- prof
50.61 46.25 42.76 50.34 63.15 73.10 50.39 52 . 99 60.86
I prof
42.73 39.75 42.71 42.79 50.51 41 . 55 67.04 48.10 41.53
I meta I anaf
14.41 16.49 12.08 8.08
17.87 12.64 16.04 14.49 15.16
9.76 5.38 8.33 3.88 7-42 4-89 9.30 7.68 7. 11
Hora 9
I meta I anaf
11.60 10.78 17.94 f2.M 11.96 9.69
10.62 13.69 13.04
8.02 7.86 8.82 5.01 3.34 1.37 4.33 7.18 4.74
Hora 10
I meta I ana+
19.61 15.47 11.45 16.95 10.58 16.26 11.93 13.70 18.00
10.6% 8.66 5.69 7.55 5-11 9.52 2.84 6.70 8.00
I te lo
20.69 41 . 07 44.02 16.70 29.11 64.36 26.74 27.88 28.36
I telo
29 . 7s 40 . 95 30 ...46 32. S6 21 . 53 15.86 34.64 27-12 21 . 34
I te lo
24 . 78 36.10 40.13 32.68 33.78 32.65 18.18 31 . 40 32.45
I binuc
3.33 4.5s 5.43 3.44 1.21 4.23 5.65 3.36 3.55
I binuc
3.27
.. .. 6.09 5.18 3.94 3.09 4.79 3.48 4.32
. 4.96-
I binuc
2.96 7.39 6.22 9.77 4.54 6.35 6.53 4.81 9.67
I bipro
19-09 14.47 3.39 1.44 1.52 1.05 5.75
2.26 1.97
I bipro
10.40 16.75 1-7.13 16.00 19.64 18.86 17.29 16.89 22.36
I bipro
24 I34 21 . 47 26.30 25.22 30.12 29.96 30.71 29.71 26.75
F
24
Cebolla #
Indice mitótico Indice profiisico Indice metafbsico Indice anafásico Indice telofásico
B-inter+ase (Hrs. 1
Cebolla #I
Indice mitótico Indice profásico Indice metafásico Indice anaf ásic0 Indice telofásico
B- i nterf ase (Hrs. 1
c 11
13.02 49.48 15.20 9.48
26.70
10. 19
C 19
11.62 55.11 12.86 5.49
26 52
7.50
c 12
18.21 41.01 14.24 7.30
39 . 37
1.65
c 20 14.99 50.00 13.96 7.18
28 . 82
7.86
TABLA 111. DESVIACION ESTANDAR DE LA (Por Cebol la1
Cebol la 13 Cebol la 1S Cebol la 17 Cebol la 18 Cebol la 19 Cebol la 20 Cebol la 21
Val or pormed i o
C 13 C 15
18.39 21 . 68 40.34 40.63 13.82 12.36 7-61 5.48
38.20 27.31
7.75 7.90
c 21
16.64 50 . 58 15.40 6.61
27.38 -
7.88 . . _ .
C 17
12.45 53.08 13.47 5.29
28.14
7.91
. .
DURACION DE LA B-INTERFASE
Duración en boras
7.75 7.90 .
7.91 7.94 7.50 7.86 7.88
-,
7.82
C 18
9.74 52-00 12.85 5.92
37.62
7.94
c
Y '
Desviación estandar ( S ) O. 15
"" -
25 I
I f i
TABLA iif;--%ESW€t%3~ ESTANDAR DE LOS PRO!IEDIOS.Dg LOS INDICES MITOTICOS (Por Cebol la)
Cebolla 11 Cebol la 12 Cebol la 13 Cebol la 15 Cebolla 17 Cebol la 18 Cebol la 19 Cebol la 20 Cebol la 2 1
Indice mitótico Desviación estandar promedio
13.02 18.21 18-39 21.68 12.45 9.74
11.62 14.99 16.64
TABLA V. VALORES PROMEDIO TOTALES
Indice mitotic0 Indice profasico Indice metafasico Indice anafasico Indice telofasico
2-40 4.23 3.28 7.08 1-41 6.55 5.11 4.82 2.19
.
INTERFASE PROFASE METAFASE ANAFASE
7.82 2 0.15, 0. 67 0.19 0.09
B-inter+ase tHr5.1 7.82 3 0.15;
TELOFASE 0. 43
MITOSIS COMPLETA 1-40
CICLO CELULAR COMPLETO 9.22 2 0.15
. . . . .
. .
26
l
T 1
8
8
I I .
x
, . 4 . / /
f
L
j_- -t"+
e .
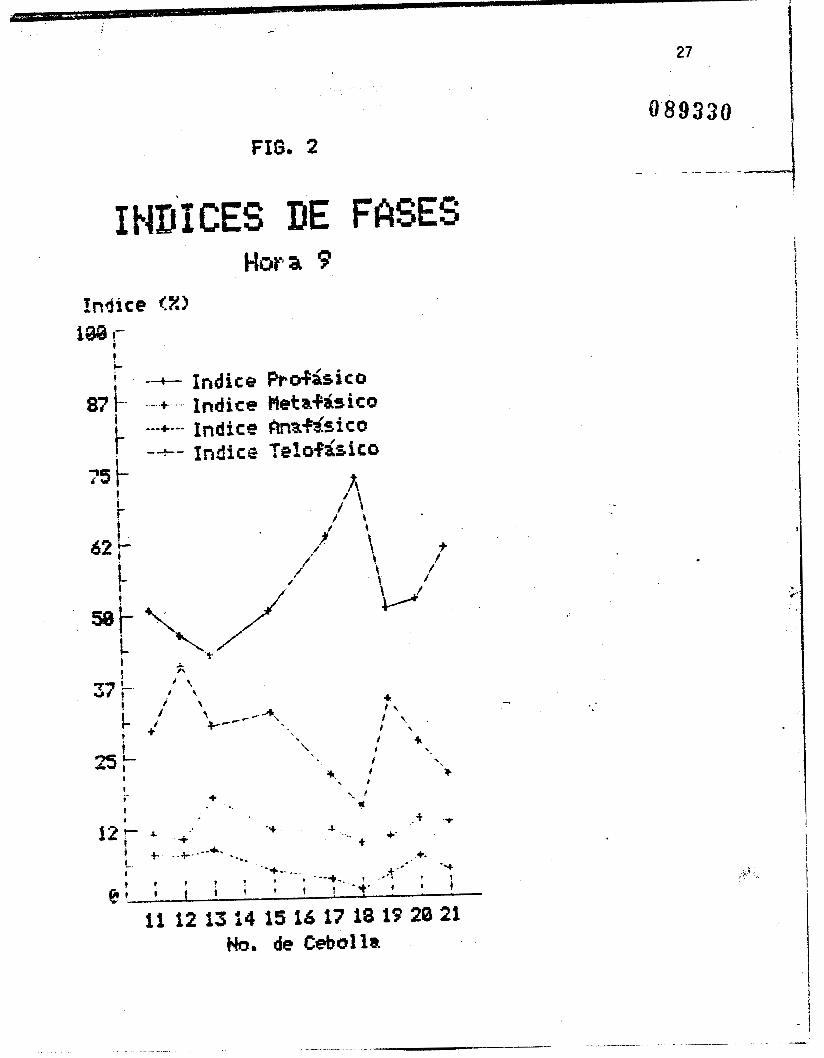
FIG. 2
11 12 13 14 15 16 17 18 19 20 21
-.
27
0.89330 1 i
28
. '. )-"" t"" A
- , c-
lf 12 13 14 15 16 I? 18 19 20 21 ).la. da Cebolla
!
" -
.. 7 3
29
30
FIG. 5
i 1
I> I
,

31 i I I I ! : I
i
I
f I+
t : I < i..
. .
32
t
53i- *
I
I i
1 I 1
26 t t
i 8 t
. . . . . . .
!
€ €
. , .
. .
. .. ".
I i
7
.- I
I
1 I i
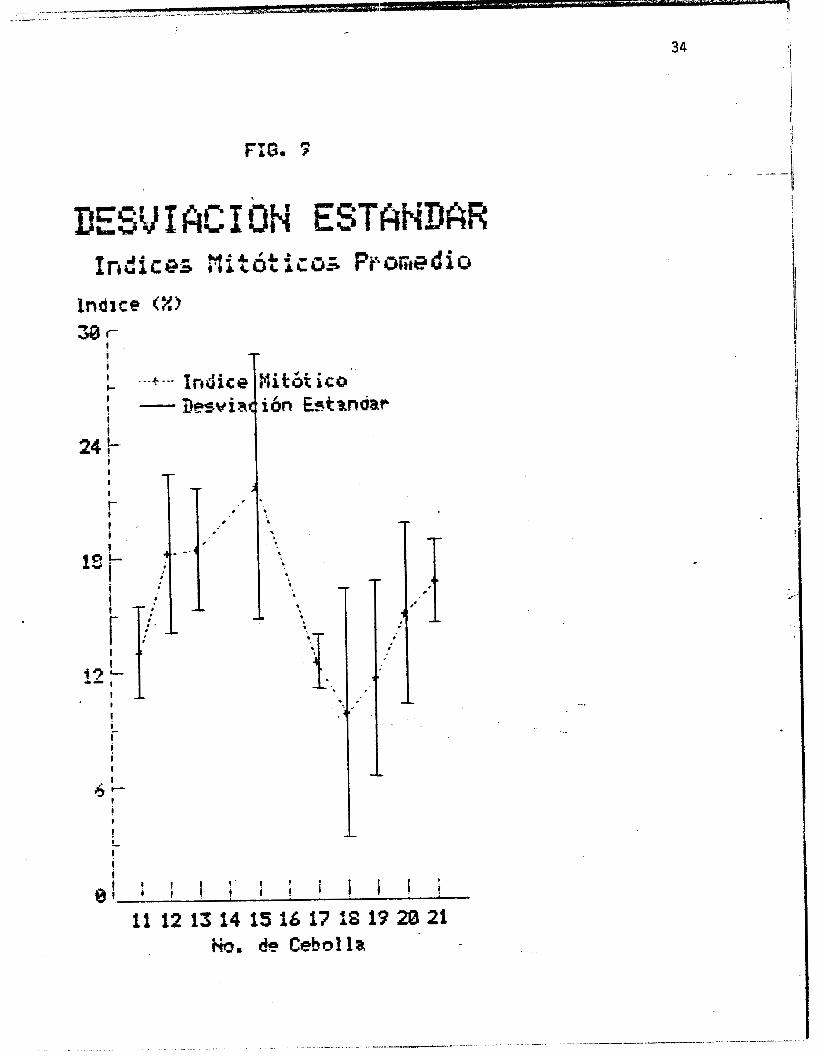
FIG. 7
34
:I; "

35
FIG. 10
INTERFASE
MITOSIS

36 ,:
TELOFASE
. . .
:1 'AFASE
ANAFASE
...

37 i/
. . ..
FIG. 12
INTERFASE
!
5% PROFASE
:f.@ METAFASE 8 ANAFASE
""

38
DISCUSION ""
Se esperaria que la represcntaci& grafica de 10s indices de
fases por cebolla y por hora (figuras 1, 2 y 3 ) fueran lineas
rectas paralelas al eje de las abscisar;, es decir, que todas las
cebollas presentaran indices iguales al encontrarse en igualdad
de condiciones. Sin embargo, pequeñas variaciones son de
esperarse entre una cebolla y otra, como siempre que se trabaja
'con organismos vivos. Como se observa en las figuras, la fase con t I
un 3ndice mayor +ue en todas los casos la profase, seguida de la
telofase, luego la metafase, y por dltimo la anafase. Se observa i
i tambiPn que la profase y la telofase presentan .variaciones
mayores, entre una y atra cebolla, que la metafase y la anafase.
La au~&ekia de' datos 'para las cdollas 14 y 16 se.;debe a que ,las . ..
* . ' rajces 'se perdieron accidentalmente.
En el caso de las cebollas 12, 13 y 18, .en la hora 8 , .en las
que los Indices telofasicos son mayores que los profksicos, puede
pensarse en un error de apreciacih al hacer el contea, aunque es
posible que' en estos casos realmente hubierada' telofases que
profases en la poblacih mononucleada total. A pesar de esto, se-
observa que en los promedios de los indices de fases por cebolla
(figura 71, las proporciones esperadas se mantienen, siendo los
Indices prof&sico5 s'iempre mayores que los telofasicos.
LOS indices bipro+&sicos presentan valores muy semejantes en
todas las cebollas, a axcepcian de los bulbos 11 y 12. En estos
dos casos, la gran diferencia entre los valores obtenidas y los
esperados se debe a un grave error de apreciacibn de las
I " . " .... ._ .. "_ " . -. ..~ ~.. " " . " .~ ". -
1
!
i

39
7.50 y las 8.00 horas. La desviacihn estandar encontrada de 0.15, -. .. :-cis decir '. 9 minutos, es realmente, pequeza s'i se considera el ., 1
tamaño de la muestra, ya que este tipo de estudios generalmente
se hace con tres bulbos (Lbpez-SAez,& aL,1966), y en este caso
se utilizaron 11, de 105 cuales 7 (cebollas 13, 15, 17, 18, 19,
20 y 21) arrojaron resultados muy parecidos a los esperados. La ?
I
muestra can la que se trabajtr representa por lo tanto un total
aproximado de 160,000 elulas. ,Considerando qua todo5 10s
calculos utilizados se hicieron con base en el ndmero de calulas
de cada preparacih, se puede considerar que las diferencias
encontr.adas entre los valores. son debidas al azar.
c
-.
. . . .
, El promedio de los lndices mitbticos por cebolla (figuras 8 y
9) muestra algunas varíaciones aparentemente significativas entre
uno y otro bulbo. Sin embargo, como se observa en la figura 9 ,
las desviaciones estandar para cada valor de la grafica (que a su
vez corresponde al promedio de los indices mitbticos de cada
hora) 5e traslapan, por lo que se puede concluir que las
diferencias son debidas al azar (Tabla I V ) , En esta figura se
utilizb una escala mayor en e1 eje de las ordenadas que en la

40 I
I I
+igura 8, para poder apreciar m i i s claramente los valores de las
desviaciones estandar. " ~ ?_.-
El promedio total de los indices mitbticos, de 15.19Xs es un
valor bastante cercano al reportado en la literatura, de 1 3 s
iL&pet-S&ez,& aL,19661. De la misma forma, los promedios totales
?Ga? 105 indices de fases son tambiPn normales segbn las reportados .
, El valor de la duraci&n. total del ciclos de 9.22kO.lJ horass
dif iere sigqif icativamente del reportado. por L&paz-Saez
(19661, de 13.5 horas, aunque el valor de la duracibn del periodo
mitbtico, . ,de 1.4 horass no difiere tanto d,el reportado por el
.-ni.smo -autor, de 1.75 haras. Por lo tantos . la diferencia r
I
-&gni$icatiua se: huentra en la duracih de la B-interfase, 'de
' . . 7."82,C0.1S horas en e,l . c a s o de este estudio, y 9.78)10.1 horas an
- . el caso del estudio rea.ii.r;ado par el .autor antes mencianado. Esta
a diferencia puedo deberse a que la variedad de las cebollas
utilizadas fuera 'distinta -.a pesar de tratarCe-.:*de .'la misma . .
especie, Allium -, L. Las condiciones ambientales generales
(altitud, latitud, humedad, etc.) pudieron tambiBn variar, a
pesar de que la temperatura se mantuvo constante en ambos
estudios, *e
L o s ' resultados obtenidos en el presente estudio son muy
similares a los que se reportaron en la primera fase del mismo
(cebollas 1 a 101, por lo que puede pensarse que las condiciones
se han mantenido estables, y que los resultados obtenidos hasta -- "
ahora son conf iablee.

41
La importancia del proceso nitthtico deriva del hecho de que de
61 depende el crecimiento y desarrollo de los organismos:
mientras que se lleve a cabo normalmente, el crecimiento y
desarrallo seran normales. Por tanto, debe 5er posible encontrar
claves sobre problemas de desarrollo y crecimiento a partir del
patrbn del ciclo mitbtico.
. - .. - - "
ASS, es importante establecer el patran del c i c l o mitetico
para cada especie de importancia econdmica. En 'este caso, el
. ciclo de.. . &l&hm cepa puede ser un htil punto .inicial que sirva
para hacer comparaciones o como alguna forma de control. En el
hombre puede incluso 5er &til el establecinieñto de patrones
mitbticos para los tejidos de los varios tirganos susceptibles de
,crecimiento canceroso o de alguna forma anormal, con. 1a.esperanza
de encontrar en el proceso alguna clave para su cura y control.
! sParece ser entonces *-e el estudio del ciclo; .celular de los
Y
C .
. 7rrrgan.Emncas presenta un futuro inpartanbe, 4
Por otra parte, el conocimiento de la duraci&n de las
diferentes fases que componen el ciclo celular puede qer de gran
Utilidad en estudios sobre la accidn de diferentes agentes
mutaghicos, que posiblemente afecten a la celula de manera
distinta dependiendo de la fase del ciclo en la que acttien.
.~
. * 4. * .
. .
. .
.

CONCLUSIONES
ciclo celular en cPlulas de meristeono de cebolla com?ln, Allium
-, L-# bajo las condiciones de trabajo en el Laboratorio de
GenCt.ica Evolutiva de la UAM Izapalapa.
De la misma manera, ~.~.obtuvo. un valor de 7.8220.15 horas para
1 . a duracim de la B-interfa-4 0467 .:haras para la profase; 0.19
horas para la metafase;. 0.09 horas para la anafasei y 0.43 horae
para la.te€ofase. La mitosis completa dura 1.4 horas, -,es decir,
ialrededor del 15% del tiempo total de duracih del ciclo celular.
2. El uso de la B-interfase es &til para establecer un metodo
rapid0 y confiable par.+ calcular la duracibn del ciclo ce.luIar en i . e .
I
l
una pohlacian l~?l_ulas meristembticas que se encuentren en un
estado de equilibrio dinbnica. . % I'
3. El conocimiento de 3a duracidn del ciclo celular y de cada .. I una de su5 fases en pob.lacianes de celulas meristematicas
.en, equilibrio din&mico-,: - ..permite elegir adecuadhente las %. condi.ci.an*s para cu,alquie,r experimento en particular-. '
. .. " . ". "-

43
i i
RESUtlEN
- - " " "- Se utiliza el marcado cpn cafeina para producir una poblacidn
de cClulas binucleadas altamente.sincronizadas en meristemos de
raiz de cebolla coman, -, y se realizaron preparaciones
utilizando el mbtodo convencional de tincibn con orceina
i
1 I
i acetoclorhldrica. De esta forma, fue posible hacer conteos al I
mkroscopio de:. Quz del ntlmere- de c&l,ulas que se encontraban en
cada fase. del .cicPa,- cefulsr,'-..tmto anmo como binucleadas,
calculando asl la duracien de 'la B-inter#ase, que es el periodo
[
i !
que abarca desde la hora O (fin del tratamiento con cafeina)
hasta la aparicibn de las primeras celulas binucleadas en . i
biprofase. La durac9-h d'e las fases mit&ticas se calculb a partir j
de sus frecuencias -:relativas en la poblacibn aeristematica.
I !
I
:I ? I
.. ).
1 .. 1 : ' ? . S - e ~ observb que el. c.iclo '&.lul?r completo en ALLUm tiene
una :. durtac.tbn de 9.22ifI.iS horas, ocupando la mi*tosis alrededor
. . del 15% 'da este tiempo, 'es. dec.ir, :1.4 horas. Lar 'fases mitbticas
muestran . siempre un patr&n .en a.1 que la profase es la fase que
ocupa un *apso mayor, ' seguida por la telofase, luego. la 'metafase,
y finalmente la anafase.
c
1 . . .
.
Se sugiere la utilizacibn de los valores obtenidos para el
estudio del efecto de agentes mutagPnicos en las diferentes fases
de la mito~is, as1 como estudios posteriores del ciclo nitbtico,
en la cebolla y otras especies de importancia comercial, ya que
estos pueden servir como base para comprender mejor los procesos
de crecimiento anormal en ibrganos de otros organismos. -

LITERAlURA CITADA
1. Avers,J. 1983. Bialaqia yolulPr.-Ln"m&-ditorial Iberoamerica, H&xicros pp. 426-441.
..aa.. De la Torre,C. 1971. 'Cell flow and its application to the e . ..
3.
, . - _ .
8.
9.
10.
11.
estimate of^ cell cycle parameters in,meristeas'. w. m. 90:707-710.
Evans,H.3. and 3.R.K. Savage. 19S9. 'The effect- Of temperature on mitosis and -on the action of colchicine in r&. incrister cells of l4,id.a m'. Ern, (Zall UL. '18: 51-61
Q i rrenez-Mar t-1-n de la Torrg,C,, L&pez-SBez,J. 1977. 'Cell division i'n higher plants. In: . in
nf diuisipn. Rost & Gilford Eda. .U.S.A.
4.
Lbpez-S2iez',3;F. ef. a. 1966. 'Duration of the cell division cycle arid its dependence on temperature'. fur 7", 75:591.
Perry,J.. and-*L.Wolff. 1.974. .. Citado en: Pincu,H. d. 1986 .
-.
Pi*ricu,H. filt al. 1986, "A modi+ied harlequin stain for cell .. kinetics?.. C u and t w . -tltr,s. Vol.19, No.3, 275-
287 . Vant*Hoff,J. and A.H.Sparron. 1963. 'The effect of mitotic
cycle duration on chromosome breakage in sqristematic cells of I ~ s L ~ m''. p s L nat. Acad, sci, 30:855- 860 .
Vant'Hoff,J. and U.K.Ying. 1964. 'Relationship between the duration of the mitotic cycles the rate of cell production and the rate of growth of roots at different temperatures..' -. Tokyo, 29:399-406=