Control de la actividad enzimática
43
CONTROL DE LA ACTIVIDAD ENZIMÁTICA
-
Upload
melissa-lds -
Category
Health & Medicine
-
view
208 -
download
3
Transcript of Control de la actividad enzimática

CONTROL DE LA ACTIVIDAD
ENZIMÁTICA

Control de la actividad enzimática
Enzimas reguladorasretroinhibición
Modificación covalente Enzimas alostéricas
Activacion por proteolisis


REGULACION ENZIMÁTICA
MECANISMO DE RETROINHIBICIÓN

ENZIMAS ALOSTERICAS
Existen sistemas enzimáticos en donde el producto de la reacción actúa como inhibidor especifico de una enzima situada al comienzo de la secuencia.Esta inhibición se asigna como:• Inhibicion por el producto• Inhibicion feed-back• Retroinhibición Estas enzimas reciben el nombre de enzimas alostericas propuestas por J. Monod,et.al y presentan un sitio diferente al centro activo al que se une reversiblemente


ENZIMAS REGULADORAS POR RETROINHIBICION
Retroinhibición de la conversión de L-Treonina en L-isoleucina, catalizada por una secuencia de 5 enzimas. La treonina deshidratasa (E1) es inhibida alostericamente por la L-Isoleucina, producto final de la secuencia.

Retroalimentación negativa en vías metabólicas
Asp + CP Carbamil aspartato CTP
Síntesis de pirimidinas
ATCasa
El producto final de la vía, CTP (citidin trifosfato) inhibe a laprimera enzima de la ruta metabólica, Aspartato transcarbamilasa
Al mismo tiempo, dicha enzima puede ser activada por ATP
ATP
+

Control por Retroalimentación Negativa
Síntesis de Iso a partir de Tre en bacterias
Las altas concentraciones de Iso inhiben a la Treonina
deshidratasa
Síntesis de AMP y GMPLas altas concentraciones de AMP
y/o GMP inhiben a la PRPP sintetasa

Modificación covalente

Enzimas reguladoras
2. Modulacion covalente reversibleFosforilacion AdenililacionUridililación ADP-ribosilaciónMetilación Carbamilación

CARACTERISTICAS • Esta enzima presenta 2 formas:
Fosforilasa a: forma activa
Fosforilasa b: forma menos activa
• La fosforilasa a es una proteína oligomérica con 4 sub-unidades, cada una con una Ser. Fosforilada en su OH.
• Estos grupos fosfato pueden separarse por una fosforilasa fosfatasa, para pasar a fosforilasa b
• La fosforilasa b puede activarse por acción de la fosforilasa quinasa.
• Por tanto, la actividad de la fosforilasa del glucógeno es regulada por enzimas que desplazan el equilibrio entre sus formas activas e inactivas,

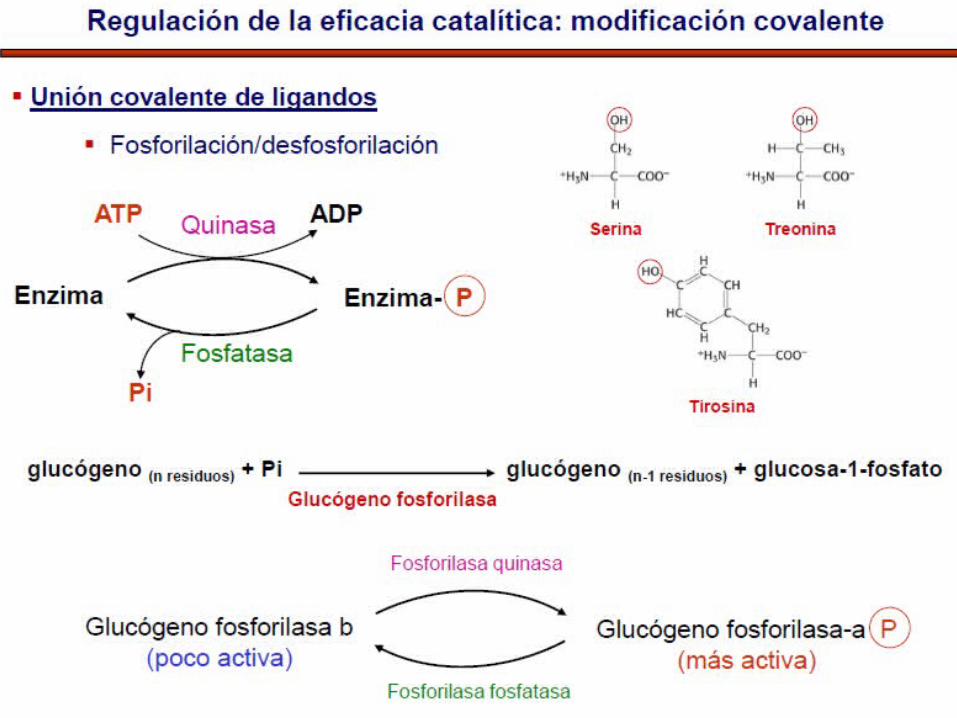
FOSFORILACIÓN - DEFOSFORILACIÓN
La forma Fosforilada y no Fosforilada son diferentes y dependen de fosforilasas y fosfatasas.
Estas enzimas son interconvertibles.
La actividad de la protein quinasa y protein fosfatasa es regulada por hormonas y neurotransmisores

Modificaciones covalentes de las enzimas
• Las formas activas e inactivas son interconvertibles por modificaciones covalentes de sus estructuras que son catalizadas por otras enzimas.
• Ej. Glucógeno fosforilasa de los tejidos animales que catalizan la degradación del polisacárido de reserva de glucógeno en glucosa mas fosfato.

Formas de modificación covalente, 1
Fosforilación: Protein kinasas
Sobre residuos de Ser, Thr y Tyr

La modificación estructural supone la esterificación de los términos hidroxilos de ciertos aa con el P del ATP

Regulación Enzimática
Modificación Covalente
2 ATP
2 ADP
Glucagon (H)Adrenalina (M)
↑[cAMP]
2 H2O
2 Pi
Insulina
Glucógeno Fosforilasa b
(menos activa)
- OH HO -
Glucógeno Fosforilasa a
(más activa)
- Pi Pi -
Fosfoprotein Fosfatasa 1
(PP1)
Fosforilasa b quinasa
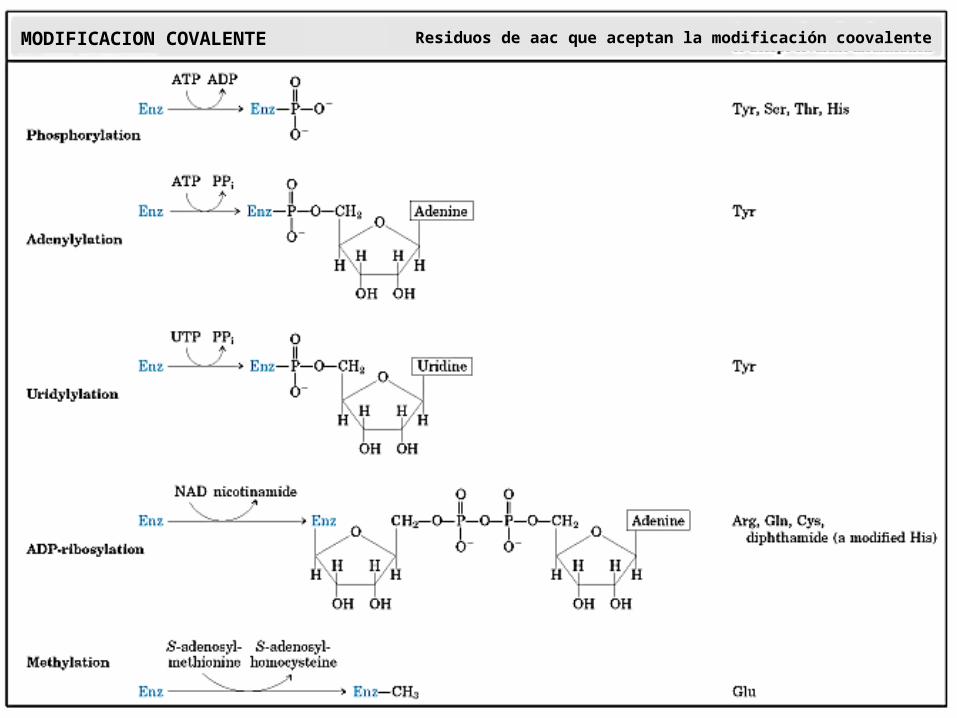
MODIFICACION COVALENTE Residuos de aac que aceptan la modificación coovalente

Formas de modificación covalente, 3
Adenilación: Sistema de la Glutamina sintetasa

Formas de modificación covalente, 4
ADP-ribosilación: actúa NAD+ como donador delgrupo ADP-ribosa


MODIFICACION ALOSTERICA

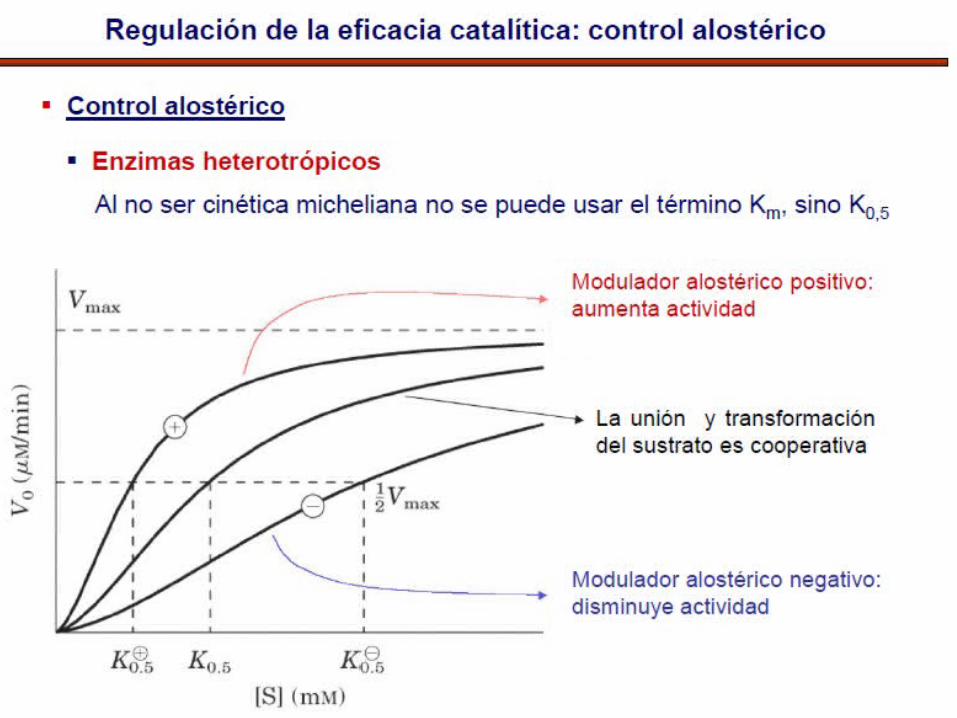
Características de las enzimas• Son oligoméricas• Estables a C°• Algunas enzimas son activadas o inhibidas por un efector
(modulador)• Cuando el efector es diferente al S: enz. Heterotrópicas• Cuando utilizan el S como modulador: enz. Homotrópicas.• No obedecen a la cinética de Michaelis-Menten• Frecuentemente muestran gráficas sigmoideas de velocidad• Porque la unión de la primera molécula se S incrementa la
velocidad de unión de las subsecuentes moléculas de S


• La cooperatividad positiva consiste en que la fijación de una molécula de sustrato favorece a la fijación del siguiente, y así, hasta ocuparse toda la molécula.
• Existe también cooperatividad negativa, cuando la fijación de una molécula de sustrato dificulta la fijación del siguiente
Modelo concertado
Modelo secuencial
La unión ligando a una sub-unidad induce la transición concertada de la otra (el efector cambia el equilibrio de T a R)
La unión ligando a cualquier sub-unidad induce un cambio conformacional a la sub-unidad siguiente y se transmite secuencialmente a las otras subunidades




Propiedad
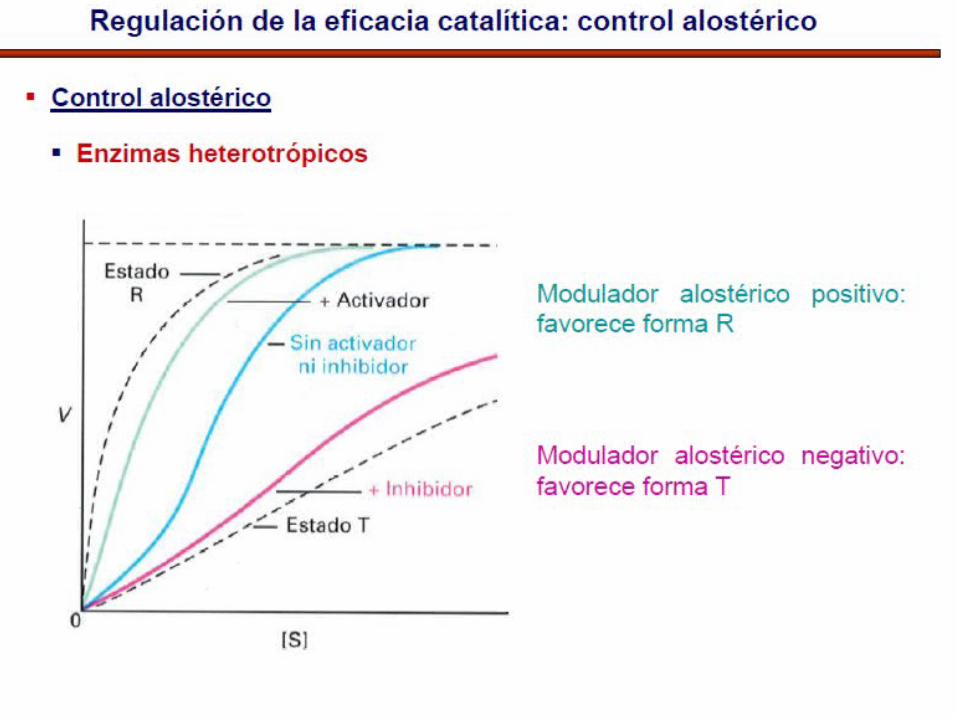
• La mayoría de enzimas alostéricas pueden ser activadas o inhibidas por efectores:
Los efectores (+) reducen el valor de S y la cooperatividadLos efectores (-) aumentan el carácter sigmoideo de la curva

Enzimas alostéricas
Sufren cambios conformacionales en respuesta a enlace de efectores o moduladoresEj • Inhibidores • Activadores


K enzimas- Efector modula km
V enzimas- Efector modula Vmax
Cinética sigmoidal: activación/
inactivación como interruptor (switch-like)

ACTIVACION POR PROTEOLISIS



ACTIVACIÓN DE ZIMÓGENOS
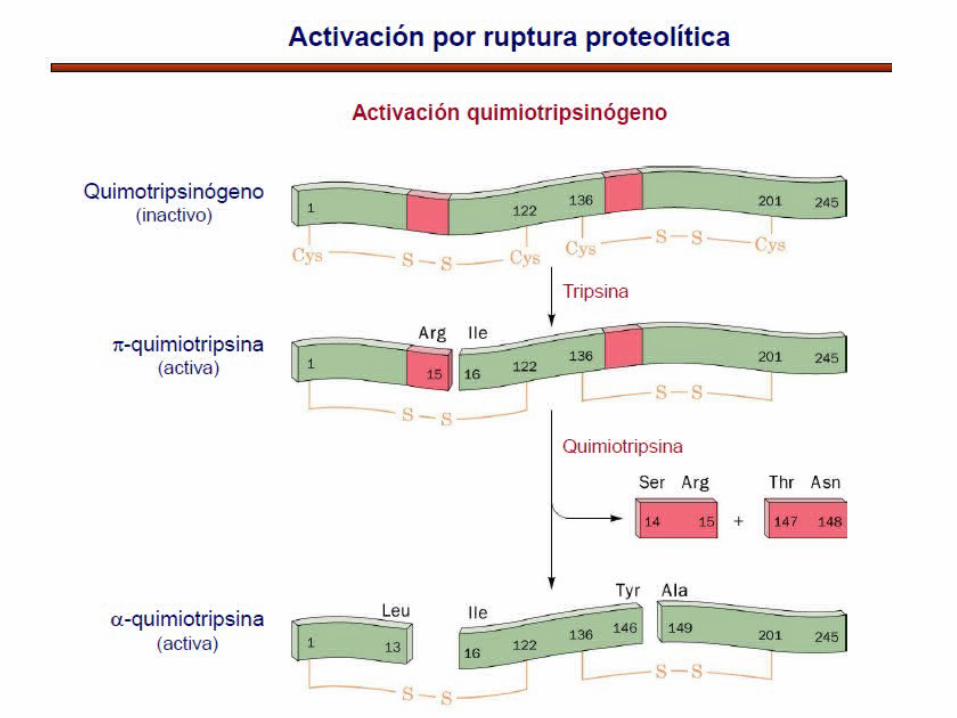

DEL QUIMITRIPSINÓGENO

Proteasa inactiva
Activación por proteólisis en
cascada

Proteolisis por trombina
A y B
Monómero de fibrina
Union β - βGHR Union γ - αGPR
βGHR
αGPR