'Consumo materno de alcohol: impacto en el desarrollo...
360
Dirección: Dirección: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293 Contacto: Contacto: [email protected] Tesis Doctoral Consumo materno de alcohol: Consumo materno de alcohol: impacto en el desarrollo embrio- impacto en el desarrollo embrio- trofoblástico temprano y en la trofoblástico temprano y en la gestación gestación a término a término Pérez Tito, Leticia Gabriela 2014-12-22 Este documento forma parte de la colección de tesis doctorales y de maestría de la Biblioteca Central Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe ser acompañada por la cita bibliográfica con reconocimiento de la fuente. This document is part of the doctoral theses collection of the Central Library Dr. Luis Federico Leloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the corresponding citation acknowledging the source. Cita tipo APA: Pérez Tito, Leticia Gabriela. (2014-12-22). Consumo materno de alcohol: impacto en el desarrollo embrio-trofoblástico temprano y en la gestación a término. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. Cita tipo Chicago: Pérez Tito, Leticia Gabriela. "Consumo materno de alcohol: impacto en el desarrollo embrio- trofoblástico temprano y en la gestación a término". Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 2014-12-22.
Transcript of 'Consumo materno de alcohol: impacto en el desarrollo...

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Consumo materno de alcohol:Consumo materno de alcohol:impacto en el desarrollo embrio-impacto en el desarrollo embrio-
trofoblástico temprano y en latrofoblástico temprano y en lagestación gestación a términoa término
Pérez Tito, Leticia Gabriela
2014-12-22
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Pérez Tito, Leticia Gabriela. (2014-12-22). Consumo materno de alcohol: impacto en el desarrolloembrio-trofoblástico temprano y en la gestación a término. Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires.
Cita tipo Chicago:
Pérez Tito, Leticia Gabriela. "Consumo materno de alcohol: impacto en el desarrollo embrio-trofoblástico temprano y en la gestación a término". Facultad de Ciencias Exactas y Naturales.Universidad de Buenos Aires. 2014-12-22.

UNIVERSIDAD DE BUENOS AIRES
Facultad de Ciencias Exactas y Naturales
Departamento de Biodiversidad y Biología Experimental
Consumo materno de alcohol:
impacto en el desarrollo embrio-trofoblástico temprano
y en la gestación a término
Tesis presentada para optar al título de Doctor de la Universidad de Buenos Aires
en el área Ciencias Biológicas
Lic. Leticia Gabriela Pérez Tito
Director de Tesis: Dra. Elisa Cebral
Consejero de Estudios: Dr. Dante Agustín Paz
Laboratorio de Reproducción y Fisiopatología Materno-Embrionaria
Departamento de Biodiversidad y Biología Experimental - FCEN - UBA
Instituto de Fisiología, Biología Molecular y Neurociencias - UBA - CONICET
Buenos Aires, 22 de diciembre de 2014

Resumen
Consumo materno de alcohol:
impacto en el desarrollo embrio-trofoblástico temprano y en la gestación a término.
El consumo de alcohol durante la gestación produce el Síndrome de Alcoholismo Fetal. Postulamos que
el consumo perigestacional de alcohol hasta períodos periimplantatorios, en el ratón, induce
alteraciones embrio-trofoblásticas que resultan en defectos feto-placentarios a largo plazo.
Utilizando modelos murinos (CF-1 y CD-1) y la administración oral de etanol (10%) previo a la
fecundación y hasta los días 4, 5 u 8 de gestación (dg), los objetivos fueron: 1) analizar parámetros
biométricos y el ciclo estral frente a la exposición pregestacional (17 días previos a la gestación); 2)
evaluar la diferenciación, crecimiento embrionario y capacidad invasiva trofoblástica luego del
consumo de alcohol hasta la etapa periimplantatoria (días 4 y 5 dg) y postimplantatoria temprana (día
8 dg); 3) analizar la incidencia de malformaciones fetales y anomalías placentarias en la gestación a
término (día 18 dg). La exposición pregestacional a alcohol en CF-1 indujo interrupción del ciclo estral,
resultados que determinaron la inducción de la ovulación. Al día 4 dg (preimplantación) en CF-1 y CD-1
se evidenció retraso en la diferenciación y crecimiento embrionario, alteración de la expresión y
localización de moléculas de adhesión (ZO-1, E-cadherina) y aumento de apoptosis. Al día 5 dg
(implantación), además de retraso de diferenciación, se halló desregulación en la dinámica de
crecimiento y expansión trofoblástica durante la implantación in vitro. Al día 8 dg, se evidenció menor
grado de diferenciación embrionaria, alteraciones histológicas junto a una alta tasa de apoptosis. La
reducida expresión de E-cadherina en ectodermo embrionario e incremento de metaloproteasas -2 y -9
posiblemente se relaciona con la expansión trofoblástica anómala observada en cultivos de conos
ectoplacentarios. Al día 18 dg, luego del consumo de alcohol hasta el día 4, 8 o 10 dg, se encontró
aumento de reabsorciones embrionarias, alta incidencia de malformaciones craneofaciales, osificación
fetal incompleta y alteraciones histológicas placentarias.
La ingesta perigestacional de alcohol hasta la preñez temprana altera la dinámica de diferenciación y
crecimiento del embrión periimplantatorio, las uniones celulares y la expresión de metaloproteasas en
los tejidos embrionarios conduciendo a malformaciones fetales y posible restricción del crecimiento
intrauterino al día 18 dg. La desregulación de la invasividad trofoblástica al día 5 y 8 dg podría
vincularse con pérdida temprana de la gestación y con alteraciones placentarias observadas en la
gestación a término.
Palabras claves: ingesta de alcohol, embriones de preimplantación, trofoblasto, gestación a término,
moléculas de adhesión, metaloproteasas, apoptosis, ratón.

Abstract
Maternal alcohol consumption:
impact on early embryo-trophoblast development and on gestation at term.
Alcohol consumption during pregnancy causes Fetal Alcohol Syndrome. We postulate that
perigestational consumption of alcohol to periimplantational periods in mouse, induces embryonic and
trophoblast changes that result in fetal-placental defects at long term.
Using murine models (CF-1 and CD-1) and oral administration of ethanol (10%) previously to
fertilization and up to days 4, 5 or 8 of gestation (g), the objectives were: 1) to analyze biometric
parameters and the estrous cycle upon pre-gestational exposure (17 days prior to gestation), 2) to
evaluate the differentiation, the embryo growth and trophoblast invasiveness after alcohol drinking up
to stages of peri-implantation (days 4 and 5 g) and early post-implantation (day 8 g), 3) to analyze the
incidence of fetal malformations and placental abnormalities in pregnancy to term (day 18 g). The pre-
gestational exposure to alcohol in CF-1 induced disruption of the estrous cycle, results that determined
the induction of ovulation. By day 4 g (preimplantation) in CF- 1 and CD-1 delayed embryo
differentiation and growth, disruption of expression and localization of adhesion molecules (ZO-1, E-
cadherin) and increased apoptosis were evidenced. By day 5 g (implantation), besides the delayed
differentiation, deregulation of trophoblast growth and expansion dynamics was found during in vitro
implantation. At 8th gd, it were evidenced lower degree of embryonic differentiation, and histological
anomalies with a high rate of apoptosis. The reduced E-cadherin expression in embryonic ectoderm
and metalloproteases -2 and -9 increase possibly is related to abnormal trophoblast expansion
observed in cultures of ectoplacental cones. At day 18 g, after alcohol consumption up to 4, 8 or 10 g,
increased embryo resorptions, high incidence of craniofacial malformations, incomplete fetal
ossification and placental histological abnormalities were found.
The perigestational alcohol intake up to early pregnancy alters the dynamics of differentiation and
growth of periimplantational embryo, the cell junctions and the expression of metalloproteinases in
embryonic tissues leading to fetal malformations and possible intrauterine growth restriction day 18 g.
Deregulation of trophoblast invasiveness on day 5 and 8 g could be linked to early pregnancy loss and
placental alterations observed in the pregnancy to term.
Keywords: alcohol intake, preimplantational embryos, trophoblast, term gestation, adhesion
molecules, metalloproteinases, apoptosis, mouse.

Agradecimientos
Agradezco profundamente a todos los que me acompañaron en este camino…
» Especialmente a la Dra. Elisa Cebral por su confianza, por su gran apoyo, sus consejos y cariño. Por
guiarme y acompañarme siempre.
» Especialmente a la Dra. Estela Bevilacqua por su inmensa generosidad, por su compromiso con
nuestros proyectos y su confianza. Por permitirme trabajar con absoluta libertad en su laboratorio,
por hacerme sentir “en casa”. Por su gran cariño y sabiduría.
» A Tamara Coll por compartir más que experimentos, haciendo lindo y entretenido el día a día. Al
Dr. Cristian Sobarzo por sus enseñanzas, su experiencia y por la ayuda técnica. A Martín Ventureira
por su buena predisposición y colaboración en este último tramo.
» A la Dra. Marta Mudry por su apoyo como Co Directora de Beca, por su predisposición y
enseñanzas. A la Dra. Nancy Andrioli por su entusiasmo y su colaboración en este proyecto.
» A la Dra. Telma Zorn, por su generosidad, por abrirme las puertas de su laboratorio, por su cariño y
grandes enseñanzas. A los Drs. Renato Mayrinck y Rodolfo Favaro por sus enseñanzas.
» A mis amigos del Trofo´s Lab: Alexandre U. Borbely, Aline Lorenzon, Camila F. Mendes, Carla
Bandeira, Fernanda Carini, Karen Prado, Karoll Nascimento, Mara Hoshida, Mariane Martucci,
Miriam R. Faria†, Rodrigo B. Weingrill, Sara Zago Gomes, Simone Correa, Sônia Giagioti. Gracias por
las horas de trabajo compartidas, por las enseñanzas, la compañía y el gran cariño que me brindan
siempre. Especialmente a Camila, Rodrigo y Sara gracias por su colaboración en este trabajo. Al Dr.
Sérgio de Oliveira, por permitirme el uso de su laboratorio, por su simpatía y cariño.
» A Rosângela Farias de Oliveira, no sólo por su valiosa ayuda técnica, sino por su contención y
cariño. A Fernanda Barrence por su ayuda y excelente predisposición. A Roberto Fernandez por su
ayuda en el confocal. A Flavio Mercau por su ayuda informática.
» A los vecinos y ex vecinos: Drs. Dante Paz, Lucas Jungblut, Fernando Meijide, Matías Pandolfi; Dras.
Andrea Pozzi, Evelin Elia, Carola Yovanovich, Carolina Pustovrh, Fabiana Lo Nostro, Paula Vissio; a
Jimena Frontera, Marilina Raices. Gracias! Por los préstamos, la ayuda, los consejos, enseñanzas y
los lindos momentos compartidos. Muy especialmente a la Dra. Nora Ceballos por su cariño y gran
apoyo, y al Dr. Rodrigo Da Cuña por su amistad.

Agradecimientos
» Al CONICET, UBA, Agencia Nacional de Promoción Científica y Técnica, y AUGM por el
financiamiento.
» Al IFIBYNE y DBBE por brindarme un espacio para formarme en la carrera científica.
» A la docencia en el DBBE… de la que tanto aprendí.
» A los secretarios Claudio Lutterbeck y Daniela De Luca, por su ayuda en lo administrativo y su
excelente predisposición siempre.
» A mis queridos amigos, más que amigos, hermanos: Dani Cecconi, Marian Desbats, Pau Sotelano
Sandra Bachmann, Renato Mayrinck y Simone Correa… infinitos gracias. Gracias por hacer más
llevaderas las angustias y más felices las alegrías. A Tatiani Donato, gracias por permitirme tantas
veces hacer de tu casa, mi casa.
» A mi mamá y a mi hermana, mis grandes compañeras. Un agradecimiento infinito por apoyarme
incondicionalmente. Por valorar mi trabajo y darme fuerzas. A mi cuñado Andrés por su apoyo,
contención y cariño.
» A Sávio por su inmenso amor.
» Y quien vive ahora en mi corazón… un infinito agradecimiento a mi papá.

Publicaciones
Parte de los resultados presentados en la presente tesis han sido publicados en revistas
científicas o están en proceso de publicación:
"PERI-IMPLANTATIONAL IN VIVO AND IN VITRO EMBRYO-TROPHOBLAST DEVELOPMENT AFTER
PERIGESTATIONAL ALCOHOL EXPOSURE IN CD-1 MOUSE". Pérez-Tito, L.G.; Bevilacqua, E; Cebral, E.
Drug and Chemical Toxicology (ISSN: 0148-0545) 37 (2): 184-197, 2014.
“LA INGESTA MODERADA PERICONCEPCIONAL DE ALCOHOL ALTERA LAS UNIONES
INTERCELULARES EN EMBRIONES PERI-IMPLANTATORIOS MURINOS”. Pérez-Tito, L.; Sobarzo,
C.M.A.; Coll, T.A.; Ventureira, M.; Cebral, E. Cs Morfol (ISSN 1851-7862) 15 (2): 1-16, 2013.
"FETO-PLACENTAL ABNORMALITIES INDUCED BY PERICONCEPTIONAL ALCOHOL INGESTION
DURING THE FIRST HALF OF PREGNANCY IN CD-1 MOUSE”. Pérez-Tito, L.G.; Mendes, C.F.; Moraes,
S.; Bevilacqua, E.; Cebral, E. -Enviado y en consideración- Cell Biology and Toxicology.CBTO-D-13-
00221

Abreviaturas
Lista de abreviaturas
ADH Alcohol deshidrogenasa
ALDH Aldehído deshidrogenasa
ATP Adenosín 5`-trifosfato
BAC Concentración de alcohol en sangre
BDO Baja densidad ósea
BrdU Bromodeoxiuridina
BSA seroalbumina bovina
CE Cono ectoplacentario
cf Capilar fetal
CG Célula glicogénica
CGT Célula gigante del trofoblasto
ct Citotrofoblasto o trofoblasto mononuclear
DAB 3-3´diaminobenzidina
DAM Decidua antimesometrial
De Decidua
DM Decidua mesometrial
ecf Endotelio de capilar fetal
eCG Gonadotrofina coriónica equina
EDC Calorías derivadas del etanol
EPG Célula espongiotrofoblástica
FAS Síndrome de alcoholismo fetal
FASD Fetal alcohol spectrum disorders
FITC Isotiosanato de fluoreceína
FSH Hormona folículo estimulante
GnRH Hormona liberadora de gonadotrofinas
HC Hembra control
hCG Gonadotrofina coriónica humana
HT Hembra tratada
IM Índice mitótico
InT-DA Interfase trofoblástico-decidual antimesometrial

Abreviaturas
InT-DL Interfase trofoblástico-decidual lateral
LH Hormona luteinizante
MCI Macizo celular interno
MEC Matriz extracelular
MMP Metaloproteasa
MN Micronúcleo
OH Alcohol
OI Osificación incompleta
PAS Ácido periódico de Schiff
PBS Buffer fosfato salino
PCE Eritrocito policromático
ROS Especies reactivas del oxígeno
Rta Reabsorción tardía
Rte Reabsorción temprana
SI Sitio de implantación
st Sinciciotrofoblasto
TA Temperatura ambiente
TE Trofoectodermo
TM Trofoectodermo mural
TP Trofoectodermo polar
TS Escala de Theiler
UA Unión adherente
UE Unión estrecha
UFP Unidad feto placentaria
ZE Zona de expansión del CE
ZL Zona laberíntica
ZO-1 Zonula occludens-1
ZP Zona proliferativa del CE
ZU Zona de unión
Otras abreviaturas se citan en el texto.


INTRODUCCION �
1. El alcohol y el alcoholismo materno 1
1.1. El alcohol como droga de abuso 1
1.2. Metabolización del etanol 2
1.3. Efectos metabólicos de la ingesta crónica de alcohol 4
1.4. Efectos de la ingesta crónica de alcohol en la función reproductiva femenina 6
1.5. Efectos teratogénicos de la ingesta materna de alcohol 7
1.5.1. Teratogénesis 7
1.5.2. El síndrome de alcoholismo fetal 9
1.6. Modelos experimentales para el estudio del los efectos de la ingesta materna de alcohol� 11
1.7. Efectos del consumo materno de alcohol en el desarrollo embrio-fetal: antecedentes 13
1.7.1. Efectos de la ingesta materna de alcohol en el sistema reproductivo femenino 13
1.7.2. Efectos de la ingesta materna de alcohol en el desarrollo embrio-fetal y placentario 14
2. Desarrollo embrionario, fetal y placentario murino 18
2.1. Desarrollo de preimplantación 18
2.2. Implantación embrionaria 24
2.3. Desarrollo de postimplantación temprano (TS6-TS11) 28
2.4. Desarrollo a término del feto y la placenta 34
2.4.1. Desarrollo a término de la placenta 34
2.4.2. Desarrollo a término del feto 40
3. Uniones intercelulares en el desarrollo embrio-trofoblástico: papel de las uniones estrechas y
adherentes
44
3.1. Uniones intercelulares 44
3.1.1. Uniones estrechas 46
3.1.2. Uniones adherentes 47
3.2. Uniones intercelulares en el desarrollo embrionario 49
3.2.1. Uniones estrechas y adherentes en el desarrollo de embriones de preimplantación 49
3.2.2. Uniones adherentes en el desarrollo embrionario de postimplantación temprano 52
3.3. Uniones adherentes en el embrión y consumo materno de alcohol: antecedentes 52
4. Matriz extracelular en el desarrollo embrio-trofoblástico: papel de las metaloproteasas 54
4.1. Matriz extracelular, su estructura y función 54
4.2. La matriz extracelular que recibe al embrión 54
4.3. Metaloproteasas en el desarrollo embrio-trofoblástico 56
4.4. Metaloproteasas y consumo de alcohol: antecedentes en otros modelos 60

5. Muerte celular y genotoxicidad embrionaria 62
5.1. Muerte celular: necrosis versus apoptosis 62
5.2. La apoptosis 64
5.3. La apoptosis durante el desarrollo embrionario 66
5.3.1. La apoptosis durante el desarrollo embrionario de preimplantación 66
5.3.2. La apoptosis durante el desarrollo embrionario de postimplantación 69
5.3.3. Factores que influyen en una apoptosis anormal: antecedentes 70
5.4. Genotoxicidad 72
5.4.1. Genotoxicidad del etanol: antecedentes 76
6. Modelos animales 79
HIPÓTESIS Y OBJETIVOS
Hipótesis de la investigación 84
Objetivos de la investigación 85
Objetivo general 85
Objetivos específicos 86
MATERIALES Y MÉTODOS
MATERIALES
1. Equipamientos principales 87
2. Insumos 87
3. Programas computacionales de análisis 89
MÉTODOS
1. Modelos animales 90
2. Diseños experimentales: tratamientos in-vivo 90
2.1. Tratamiento pregestacional con alcohol 90
2.2. Tratamiento perigestacional con alcohol: efectos gestacionales al día 4, 5 u 8 de preñez 91
2.2.1. Sin régimen de estimulación ovárica 91
2.2.2. Con régimen de estimulación ovárica 93
2.3. Tratamiento perigestacional con alcohol: efectos gestacionales al día 18 de preñez 94

3. Estudios realizados como controles del tratamiento con alcohol 96
3.1. Determinación de variaciones en parámetros biométricos 96
3.2. Determinación de los niveles de alcohol en sangre 96
CAPÍTULO I 97
Variaciones biométricas y del ciclo estral en dos modelos murinos exocriados frente al consumo
pregestacional de alcohol
I.1 MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo 97
2. Metodologías empleadas 98
2.1. Determinación de variaciones en parámetros biométricos y nivel de alcohol en sangre 98
2.2. Análisis del ciclo estral 98
3. Análisis estadístico 100
I.2 RESULTADOS
Efectos del consumo pregestacional de alcohol en las variaciones de parámetros biométricos y en el ciclo
estral en dos modelos murinos
101
1.1. Estudio de las variables biométricas en hembras murinas CF-1 y CD-1 durante el consumo
pregestacional de alcohol
101
1.2. Estudio del ciclo estral en hembras murinas CF-1 y CD-1 durante el consumo pregestacional de
alcohol
103
I.3. CONCLUSIONES PARCIALES 106
CAPÍTULO II 108
Consumo perigestacional de alcohol: efectos en la morfogénesis y en el crecimiento embrio-
trofoblástico periimplantatorio murino
II.1. MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo 108
2. Metodologías empleadas 111

2.1. Metodologías para el estudio del desarrollo embrionario de preimplantación: Día 4 de gestación 111
2.1.1. Recolección y procesamiento de embriones de preimplantación 111
2.1.2. Monitoreo de la preñez 111
2.1.3. Determinación de la diferenciación embrionaria in-vivo 111
2.1.4. Determinación de la viabilidad y evaluación de la morfología embrionaria 112
2.1.5. Determinación del grado de crecimiento embrionario e índice mitótico 113
2.2. Metodologías para el estudio del desarrollo embrio-trofoblástico durante la implantación:
Día 5 de gestación
115
2.2.1. Recolección y procesamiento de embriones implantativos 115
2.2.2. Monitoreo de la preñez 115
2.2.3. Determinación de la diferenciación embrionaria implantativa in-vivo 116
2.2.4. Cultivo de embriones implantativos 117
2.2.5. Determinación de la diferenciación embrionaria durante el cultivo 117
2.2.6. Evaluación de la morfología embrionaria implantativa 117
2.2.7. Determinación del grado de crecimiento de embriones implantativos 118
2.3. Metodologías para el estudio del desarrollo embrio-trofoblástico durante la fase de gastrulación:
Día 8 de gestación
120
2.3.1. Recolección y procesamiento de sitios de implantación 120
2.3.2. Monitoreo de la preñez 120
2.3.3. Procesamiento histológico de los sitios de implantación 120
2.3.4. Determinación del grado de diferenciación embrionaria en el sitio de implantación 122
2.3.5. Evaluación histológica embrio-trofoblástica al día 8 de gestación: estudio histopatológico 123
2.3.6. Cultivo de conos ectoplacentarios 125
2.3.7. Análisis de la viabilidad, desarrollo y capacidad invasiva de los conos ectoplacentarios en cultivo 126
2.3.8. Análisis de la morfología de los conos ectoplacentarios en cultivo 127
2.3.9. Análisis de la diferenciación de las células trofoblásticas del cono ectoplacentario en cultivo:
Inmunohistoquímica para BrdU
128
3. Análisis estadístico 128
II.2. RESULTADOS
1. Efectos del consumo perigestacional de alcohol en embriones de preimplantación: Día 4 de gestación 130
1.1. Efecto en el desempeño reproductivo 131

1.2. Efecto en la diferenciación embrionaria 131
1.3. Efecto en la morfología embrionaria 133
1.4. Efecto en la tasa de crecimiento embrionario 135
2. Efectos del consumo materno perigestacional de alcohol en el desarrollo embrio-trofoblástico durante
la implantación: Día 5 de gestación
137
2.1. Efecto en el desempeño reproductivo, en la diferenciación, morfología y crecimiento embrionario
implantativo (in-vivo)
137
2.2. Efecto sobre eventos de la implantación (estudios in-vitro) 140
3. Efectos del consumo perigestacional de alcohol en el desarrollo embrio-trofoblástico en la fase de
gastrulación: Día 8 de gestación
147
3.1. Efecto en el desempeño reproductivo 147
3.2. Efecto en la diferenciación embrionaria in-vivo 148
3.3. Efecto en la morfología embrio-trofoblástica: cambios histo-morfológicos 149
3.4. Efecto en la viabilidad y capacidad de desarrollo e invasión del cono ectoplacentario (estudios in-
vitro)
154
3.4.1. Viabilidad y capacidad de desarrollo del cono ectoplacentario 154
3.4.2. Crecimiento de los conos ectoplacentarios en cultivo 156
3.4.3. Morfología de los conos ectoplacentarios en cultivo 161
3.4.4. Diferenciación celular en los conos ectoplacentarios en cultivo 161
II.3. CONCLUSIONES PARCIALES 163
CAPÍTULO III 166
Consumo perigestacional de alcohol: efectos en las uniones intercelulares, metaloproteasas y
genotoxicidad en el tejido embrio-trofoblástico periimplantatorio murino
III.1. MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo 166
2. Recolección y procesamiento de embriones, sitios de implantación, conos ectoplacentarios y médula
ósea materna
169
2.1. Recolección y procesamiento de embriones de preimplantación (día 4 de gestación) 169
2.2. Recolección y procesamiento de sitios de implantación (día 8 de gestación) 169

2.3. Recolección y procesamiento de conos ectoplacentarios (día 8 de gestación) 170
2.4. Recolección y procesamiento de médula ósea 170
3. Metodologías empleadas 171
3.1. Metodologías para el estudio de moléculas de adhesión celular en embriones en el período
periimplantatorio: Día 4 y 8 de gestación
171
3.1.1. Inmunofluorescencia para ZO-1 en blastocistos en suspensión 171
3.1.2. Inmunofluorescencia para E-cadherina en blastocistos en suspensión 171
3.1.3. Doble inmunofluorescencia para ZO-1 y E-cadherina en blastocistos en suspensión 172
3.1.4a. Inmunofluorescencia para E-cadherina en sitios de implantación 172
3.1.4b. Análisis de imágenes y semicuantificación de la intensidad de inmunofluorescencia 173
3.2. Metodologías para el estudio de la expresión proteica y génica de MMP-2 y MMP-9 en embriones en
el período postimplantatorio: Día 8 de gestación
174
3.2.1a. Inmunohistoquímica para MMP-2 y MMP-9 en sitios de implantación 174
3.2.1b. Análisis de imágenes y semicuantificación de la inmunohistoquímica para MMPs 174
3.2.2. PCR en Tiempo Real para MMP-2 y MMP-9 de cono ectoplacentario 175
3.3. Metodologías para el estudio de la genotoxicidad materna y embrionaria, y de la muerte celular en
embriones en el período periimplantatorio: Días 4 y 8 de gestación
177
3.3.1. Genotoxicidad embrionaria preimplantatoria (día 4 de gestación) 177
3.3.2. Ensayo de TUNEL in-toto en embriones de preimplantación (día 4 de gestación) 177
3.3.3. Ensayo de TUNEL en sitios de implantación (día 8 de gestación) 180
3.3.4. Marcación nuclear con Hoechst 33342 en sitios de implantación (día 8 de gestación) 180
3.3.5. Test de micronúcleos para evaluación de genotoxicidad materna 181
4. Análisis estadístico 182
III.2. RESULTADOS
1. Efectos del consumo perigestacional de alcohol en moléculas de adhesión celular en embriones en el
período periimplantatorio: Día 4 y 8 de gestación
183
1.1. Estudio de las uniones intercelulares en embriones preimplantatorios: Día 4 de gestación 183
1.2. Estudio de las uniones adherentes en el ectodermo embrionario: Día 8 de gestación 189

2. Efectos del consumo perigestacional de alcohol en la expresión proteica y génica de MMP-2 y MMP-9
en embriones en el período postimplantatorio: Día 8 de gestación
190
2.1. Estudio del patrón de inmunomarcación de MMP-2 y MMP-9 en el tejido embrio-trofoblástico 190
2.2. Estudio de la expresión génica de MMP-2 y MMP-9 en el cono ectoplacentario 195
3. Efectos del consumo perigestacional de alcohol en la inducción de genotoxicidad materna y/o
embrionaria y en la muerte celular embrio-trofoblástica: Día 4 y 8 de gestación
196
3.1 Estudio genotóxico e incidencia de muerte celular programada en embriones de preimplantación: Día
4 de gestación
196
3.2. Estudio genotóxico e incidencia de muerte celular programada en embriones postimplantación
temprana: Día 8 de gestación
201
3.3. Estudio genotóxico materno luego del consumo de alcohol 205
III.3. CONCLUSIONES PARCIALES 207
CAPÍTULO IV 209
Consumo perigestacional de alcohol: efectos en el desarrollo feto-placentario en la gestación a término
IV.1. MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo 209
2. Recolección de fetos y placentas 210
3. Metodologías para el estudio del desarrollo feto-placentario a término: Día 18 de gestación 211
3.1. Monitoreo de la preñez 211
3.2. Análisis del crecimiento fetal y placentario 212
3.3. Análisis de la morfología fetal externa 212
3.4. Análisis de la morfología fetal interna: sistema esquelético 214
3.5a. Procesamiento histológico de las placentas 217
3.5b. Análisis morfométrico y cualitativo de las placentas 219
4. Análisis estadístico 219
IV.2. RESULTADOS

Efectos del consumo perigestacional de alcohol en el desarrollo feto-placentario a término:
Día 18 de gestación
220
1.1. Estudio de los efectos nutricionales 220
1.2. Efectos en el desempeño reproductivo al día 18 de gestación 221
1.3. Desarrollo a término del feto y la placenta 224
1.3.1. Estudio del crecimiento fetal y placentario 224
1.3.2. Estudio de las anomalías morfológicas fetales externas 225
1.3.3. Estudio de las anomalías morfológicas fetales internas: análisis del esqueleto 228
1.3.4. Estudio de la placenta a término 233
1.3.4a. Morfología de la cara fetal de la placenta: zona de unión y zona laberíntica 234
1.3.4b. Áreas relativas de la zona de unión y zona laberíntica: análisis morfométrico 234
1.3.4c. Análisis histológico de la zona de unión 237
1.3.4d. Análisis histológico de la zona laberíntica 240
IV. 3. CONCLUSIONES PARCIALES 243
DISCUSIÓN Y CONSIDERACIONES FINALES 244
BIBLIOGRAFÍA 312

��� ��� ���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���
���

Introducción: el alcohol y el alcoholismo materno
1
1. El alcohol y el alcoholismo materno
1.1. El alcohol como droga de abuso.
El alcohol es una de las drogas más consumidas en el mundo, cuyo abuso genera importantes
problemas médico-sociales (Ceccanti y col., 2007; Mancinelli y col., 2007; Aragón y col., 2008).
Como toxina, el alcohol puede producir daño en diversos órganos del cuerpo. Incrementa el riesgo
de aparición de desórdenes gastrointestinales, tales como, síndrome de mala absorción de
nutrientes, pancreatitis, erosión duodenal y gastritis, pudiendo inclusive producir cáncer en el tracto
gastrointestinal, hepatitis alcohólica y cirrosis; hipertensión arterial y alteraciones cardiovasculares;
alteraciones endócrinas; complicaciones neuropsiquiátricas, encefalopatías diversas (Schuckit, 2006).
Su consumo durante la gestación es capaz de generar anomalías congénitas estructurales, funcionales
y/o bioquímicas en el individuo en desarrollo (Abel, 1984; Sadler, 1995).
Luego de su ingesta, el etanol, molécula liposoluble y de bajo peso molecular, es absorbido
rápidamente en el tracto gastrointestinal. Cerca del 20% del etanol se absorbe a través del
revestimiento del estómago y aproximadamente un 80% es absorbido en el intestino delgado para
luego pasar a la circulación sanguínea, a través de la cual llega a los diversos órganos. Sólo un
pequeño porcentaje, entre el 2-10%, es eliminado directamente por orina, aire expirado y sudor.
Luego de beber una copa, la concentración de alcohol en la sangre alcanza su nivel máximo en 30-45
minutos y vuelve al nivel normal luego de una hora. Son diversos los factores que modifican la tasa de
absorción de alcohol, entre ellos la presencia de alimentos en el estomago, estando también
parcialmente determinada por factores genéticos (Crabb y col., 1987).
El aumento de la concentración de alcohol en la sangre (alcoholemia) depende de la dosis de
consumo. El nivel de alcoholemia en los humanos define las alteraciones funcionales y
comportamentales. En términos generales, cuando la concentración de etanol en la sangre alcanza
20-30 mg/dl se deteriora el tiempo de reacción, disminuye el control motor fino y las facultades
mentales, y se incrementa la euforia. Con 50-100 mg/dl se inicia el deterioro de la coordinación. Con
100-200 mg/dl se producen náuseas, vómitos, ataxia y deterioro mental (de memoria, de juicio y del
habla). Con 200-400 mg/dl, se produce una disminución en la respuesta a estímulos sensoriales,
pérdida de la capacidad de alerta, ataxia severa, relajación de esfínteres, alteraciones electrolíticas e

Introducción: el alcohol y el alcoholismo materno
2
hipotermia. Ya con valores superiores a 400 mg/dl, se produce falla respiratoria, coma, paro
respiratorio y muerte (Izquierdo, 2002).
1.2. Metabolización del etanol.
El etanol puede ser metabolizado en varios órganos incluyendo el estomago, el intestino delgado y el
cerebro, sin embargo, cuantitativamente, la mayor parte de este metabolismo se lleva a cabo en el
hígado, por medio de sistemas oxidativos presentes en las células (Figura 1.1). El producto de la
oxidación del etanol es el acetaldehído. La enzima principal que cataliza esta transformación es la
alcohol deshidrogenasa (ADH), enzima citosólica dependiente de la coenzima nicotinamida adenina
dinucleótido (NAD+), siendo sus productos el acetaldehído y el NADH, ambos responsables en gran
parte de los efectos tóxicos del alcohol. En simultáneo existen dos vías adicionales para la oxidación
del etanol, las cuales son minoritarias en relación a la de la ADH. Una de estas vías es la catalizada por
la enzima catalasa, que utiliza peróxido de hidrógeno como agente oxidante, y la otra es un sistema
enzimático microsomal hepático presente en el retículo endoplasmático liso, llamado sistema
microsomal de oxidación del etanol (MEOS, del inglés Microsomal Ethanol Oxidizing System), que
utiliza NADPH y oxígeno molecular. Su componente principal es CYP2E1, una citocromo P450, que es
directamente estimulada por etanol. La catalasa localizada en los peroxisomas hepáticos, pueden
interferir en el metabolismo del etanol, pero sólo en presencia de peróxido de hidrógeno (Handler
y Thurman, 1988). Por eso su participación como vía de metabolización, es minoritaria. El sistema
microsomal MEOS contribuye a la metabolización del etanol principalmente cuando la
concentración del mismo es elevada (Kato y col., 1988). En todas las vías, el producto inmediato de
la oxidación del etanol, es el acetaldehído, el cual es oxidado a acetato por acción de la aldehído
deshidrogenasa (ALDH), enzima mitocondrial y dependiente de NAD+. De esta forma, el
acetaldehído que es formado por la oxidación del etanol en el citosol de las células, difunde al
interior de las mitocondrias donde por acción de la ALDH es oxidado a acetato (Crabb y col., 1987).
Como consecuencia de la metabolización del etanol, a su vez, se originan moléculas oxidantes
altamente reactivas, las especies reactivas del oxígeno (ROS, del inglés Reactive Oxygen Species),
compuestos sumamente agresivo en su forma de reaccionar (Wu y Cederbaum, 2003). Las ROS,
como el anión superóxido y el radical hidroxilo, se generan de forma natural a partir del
metabolismo basal, la respiración celular, los ciclos de oxidación microsomales y durante los

Introducción: el alcohol y el alcoholismo materno
3
procesos inflamatorios. Sin embargo, al igual que lo que ocurre con otros tóxicos, el etanol puede
sufrir ciclos de oxido-reducción que incrementan esta generación natural produciendo un
desequilibrio, que si se sostiene en el tiempo y/o los mecanismos naturales de defensa
antioxidantes no pueden contrarrestar el efecto, adquiere protagonismo el estrés oxidativo (Wu
y Cederbaum, 2003).
Figura 1.1. Representación esquemática de las vías de metabolización celular del etanol. El etanol es
metabolizado principalmente por la enzima citosólica alcohol deshidrogenasa (ADH) en acetaldehído el
cual es oxidado a acetato por la aldehído deshidrogenasa mitocondrial (ALDH). Dos vías alternativas para la
metabolización del etanol son las mediadas por el sistema microsomal MEOS y por la catalasa presente en
peroxisomas. Como resultado de la metabolización del alcohol se generan compuestos nocivos para la
célula: el acetaldehído, las especies reactivas del oxígeno (ROS) y una modificación en el potencial redox.
Modificado de www.javeriana.edu.co/Facultades/Ciencias/neurobioquimica/libros/perinatal/alcoholed.
La velocidad de absorción del alcohol, la distribución y eliminación difieren según varios factores,
entre ellos, el sexo. En tal sentido, la cantidad de agua en el organismo influye en la velocidad de
absorción del alcohol. En general, las mujeres tienen mayor proporción de grasas en el cuerpo y
por lo tanto menor contenido de agua; mientras que los varones tienen mayor masa muscular y,
por lo tanto, mayor proporción de agua, motivo por el cual en ellos el alcohol se metaboliza de
forma más rápida que en las mujeres. Sin embargo, el principal motivo por el que las mujeres
metabolizan el alcohol de forma diferente se debe a una menor expresión de enzima ADH.

Introducción: el alcohol y el alcoholismo materno
4
Demostrando este hecho que el factor genético influye también. Así, los patrones genéticos están
relacionados con la expresión de las enzimas que metabolizan el alcohol. En humanos la ADH tiene
8 isoformas cada una con diferentes propiedades metabólicas y diferente patrón de expresión
según la etnia. La expresión está genéticamente regulada. Si bien, esta enzima se encuentra
principalmente distribuida en el hígado, existen pequeñas cantidades en el tracto gastrointestinal
y en el riñón. Se ha localizado también ADH en páncreas, corteza cerebral, cerebelo, hipotálamo,
testículos, ovario y útero en menor grado (Crabb y col., 1987). La ALDH tiene al menos 4 isoformas.
Existen polimorfismos genéticos en las deshidrogenasas de alcohol y aldehído, con propiedades
catalíticas diferentes y con frecuencias distintas en las diversas poblaciones raciales (Bosron y Li,
1986). Las variaciones en la tasa de metabolización del etanol entre los individuos también están
dadas por el patrón de consumo y el grado de ayuno. Los alimentos ingeridos influyen en la
cantidad y velocidad de alcohol que puede absorber el organismo. Si el estómago está lleno, se
enlentece la absorción de alcohol (Crabb y col. 1997).
El consumo crónico de alcohol puede generar mecanismos de tolerancia metabólica producida en
parte por un incremento en la actividad de la ADH y del sistema MEOS, dado que se incrementa la
habilidad para metabolizar el etanol consumido en forma prolongada (Jaffe, 1985).
1.3. Efectos metabólicos de la ingesta crónica de alcohol.
Los efectos metabólicos del alcohol, además de la dosis, dependen del tipo de consumo: agudo,
semicrónico (o subcrónico) o crónico. Se considera intoxicación aguda cuando los síntomas clínicos
aparecen tras la exposición a alcohol corta y a una dosis potencialmente tóxica y capaz de inducir
un daño. Obedecen, generalmente, a dosis únicas o a dosis múltiples absorbidas en un período de
tiempo breve que normalmente se fija en un máximo de 24 horas. La intoxicación crónica se
considera cuando la ingesta de la sustancia se mantiene a lo largo del tiempo resultando, en
general, en una acumulación de la sustancia tóxica en el organismo.
En particular, en el presente trabajo, nos interesó abordar el estudio de los efectos de la ingesta
prolongada de alcohol en el modelo experimental animal utilizado entendiendo que esta
exposición podría contribuir a efectos metabólicos nocivos.
En el caso del etanol, el responsable de la toxicidad no es en sí el producto original. Especialmente
en el alcoholismo crónico la toxicidad es debida al acetaldehído, verdadero metabolito reactivo

Introducción: el alcohol y el alcoholismo materno
5
producto de la oxidación hepática del etanol, y a la formación excesiva de NADH y de ROS. Tanto
el acetaldehído como las ROS pueden interactuar con macromoléculas como proteínas, ácidos
nucleicos y lípidos, generando grandes desequilibrios bioquímicos (Wu y Cederbaum, 2003). A
nivel celular, el alcohol altera la composición lipídica y proteica de las membranas celulares,
disrumpe las interacciones entre lípidos y proteínas, genera alteraciones permanentes en la
conformación de ciertas proteínas (Waring y col., 1982). Así, está bien establecido que el consumo
de etanol daña las mitocondrias de las células hepáticas de tal manera que la síntesis de ATP se ve
comprometida (Cunningham y col., 1990). Más recientemente, Young y colaboradores (2006)
demostraron que el consumo crónico de etanol provoca una disminución de la producción de ATP
tanto mitocondrial como glicolítica en hepatocitos.
Como se mencionó, como consecuencia del consumo crónico, se altera significativamente la
relación NADH/NAD+, generando esto importantes efectos negativos. El contenido de glucógeno
en el hígado disminuye intensamente, lo que limita la producción de energía. El aumento de
NADH, junto con la actividad de la lactato deshidrogenasa, aumentan la tasa de producción
hepática de lactato, contribuyendo a la acidosis, reduciendo en consecuencia la capacidad renal de
excreción de ácido úrico (hiperuricemia). Esta reacción a su vez desvía el piruvato de la
gluconeogénesis, reduciendo la capacidad del hígado de entregar la glucosa a la sangre,
generando un estado de hipoglucemia (Grimberg y col., 1998). El NADH derivado de la
metabolización del etanol en el hígado conlleva a la desinhibición de la vía de lipogénesis, la cual
se activa, generando una notable alteración en el metabolismo de los lípidos, con desarrollo de
hipertrigliceridemia, la cual se manifiesta a nivel hepático por la aparición de pequeñas
acumulaciones de grasa en el interior de los hepatocitos (Carmona, 2008), que en condiciones
severas pueden originar la esteatosis hepática (hígado graso) y un aumento de los niveles de
lípidos en sangre (hiperlipidemia). Esto está a su vez reforzado por la producción de acetato a
partir del acetaldehído en la mitocondria, conduciendo a mayores niveles de acetil-CoA. Dado que
la mayor generación de NADH también reduce la actividad del ciclo de Krebs, el acetil-CoA se
desvía a la síntesis de ácidos grasos. La reducción citosólica de NAD+ conduce a la reducción de la
actividad de la glicerol-3-fosfato deshidrogenasa que resulta en un aumento de los niveles de
glicerol-3-fosfato, columna vertebral de la síntesis de la triacilglicéridos. Este estado
hipermetabólico del hígado generado por el consumo de alcohol, aumenta notablemente el
consumo de oxigeno, resultando en una hipoxia localizada. La primera etapa del daño hepático

Introducción: el alcohol y el alcoholismo materno
6
tras el consumo crónico y excesivo de alcohol es la aparición de hígado graso, seguido por la
inflamación, la apoptosis, la fibrosis y la cirrosis por último. La Figura 1.2, muestra de manera
resumida las alteraciones recién descriptas en una célula hepática.
Figura 1.2. Representación esquemática de los trastornos metabólicos hepáticos asociados a la
metabolización del alcohol. El etanol genera un gran desequilibrio óxido-reducción. La metabolización
hepática del alcohol resulta en la generación de grandes cantidades de NADH citosólica y mitocondrial,
conduciendo a alteraciones en la gluconeogénesis llevando a una hipoglucemia potencialmente grave.
La desviación del piruvato para la síntesis de lactato resulta en excesiva liberación de este compuesto a
la sangre, generando acidosis láctica. La incapacidad para convertir el lactato a piruvato representa la
interrupción en la gluconeogénesis hepática normal. Aumentan los triglicéridos causando incremento
en las grasas. Las ROS causan estrés oxidativo activando la vía apoptótica mitocondrial en hepatocitos.
Modificado de http://themedicalbiochemistrypage.org/es/ethanol-metabolism-sp.php
1.4. Efectos de la ingesta crónica de alcohol en la función reproductiva femenina.
Los efectos del alcohol a nivel reproductivo femenino, además del efecto directo sobre el ovario,
pueden deberse a acciones sobre el sistema hipotálamo-hipofisario. Así, alteraciones en la secreción
de gonadotrofinas podrían generar indirectamente alteraciones fisiológicas en la gónada (Yung y
Russfield, 1972). Se sabe que la secreción pulsátil de hormona liberadora de gonadotrofinas (GnRH,

Introducción: el alcohol y el alcoholismo materno
7
del inglés Gonadotropin-Releasing Hormone) se encuentra alterada en mujeres alcohólicas que
presentan enfermedad hepática, alterándose e incluso suprimiéndose la liberación de la hormona
luteinizante (LH, del inglés Luteinizing Hormone) y de la hormona folículo estimulante (FSH, del inglés
Follicle-Stimulating Hormone) (Mendelson y Mello, 1988). En algunos casos aparecen severas
anomalías en el ciclo menstrual, incluyendo amenorrea, anovulación y disfunción de la fase lútea
(Valimaki y col., 1984). Esto, en su conjunto, deteriora notablemente la fertilidad y, si ocurriera
embarazo, aumentaría las posibilidades de abortos espontáneos (Hugues y col., 1980; Valimaki y col.,
1984; Mello, 1988).
1.5. Efectos teratogénicos de la ingesta materna de alcohol.
1.5.1. Teratogénesis.
La teratología (del griego tératos, monstruo) es la ciencia que estudia las desviaciones del
desarrollo normal que se encuentran en un individuo al momento del nacimiento y que se
desarrollaron o produjeron durante la gestación. La causa de las alteraciones puede ser ambiental
y/o genética, pudiendo estos factores actuar independientemente o en forma simultánea.
Existen diversos tipos de alteraciones estructurales o anomalías congénitas: (1) las malformaciones
son alteraciones que se producen durante la formación de las estructuras, es decir, durante la
embriogénesis, pudiendo ocurrir la falta completa o parcial de una estructura o la alteración de su
morfología normal; (2) las desorganizaciones (disrupciones) provocan alteraciones morfológicas de
las estructuras ya formadas, y en general, se asocian a procesos degenerativos; (3) las
deformaciones obedecen a fuerzas mecánicas que moldean una parte del feto durante un período
de tiempo prolongado. Con frecuencia las deformaciones afectan al sistema musculoesquelético y
pueden, generalmente, ser reversibles en el período postnatal (Sadler, 1995)
En relación a los factores ambientales o externos que inducen anomalías congénitas, ya las
observaciones de Lenz en 1961, que vinculaban las anomalías de los miembros con el consumo
materno de talidomida, permitieron aclarar que las drogas podían atravesar la placenta y provocar
defectos congénitos. A partir de entonces, se han identificado muchos agentes de diversos
orígenes, que son teratogénicos, entre ellos agentes infecciosos, hormonas, alteraciones

Introducción: el alcohol y el alcoholismo materno
8
metabólicas maternas (como la diabetes y las carencias nutricionales), la hipoxia, sustancias
químicas ambientales (como los plaguicidas) y las drogas de abuso como el alcohol, entre otros.
Como consecuencia de numerosos estudios de laboratorio y observaciones clínicas, se han
definido los factores que determinan la capacidad de un agente de provocar trastornos
congénitos. Estos principios de teratología fueron formulados por primera vez por Wilson en 1959
y resistieron el paso de tiempo. Los principios comprenden los siguientes criterios: (1) la
susceptibilidad al teratógeno depende del genotipo del individuo expuesto, de las propiedades del
producto y de su concentración. También es importante el papel del genoma materno con
respecto a la metabolización del agente, su resistencia al mismo y en la inducción de procesos
bioquímicos y moleculares que podrían generarse en el embrión. (2) la susceptibilidad al
teratógeno varía según la etapa de desarrollo del individuo al momento de la exposición y el
tiempo de exposición al teratógeno. De forma general, el período más sensible en la inducción de
defectos de nacimiento es el correspondiente al período de embriogénesis. Sin embargo, cada
sistema orgánico puede tener una etapa de mayor susceptibilidad. Por lo tanto, si bien, la mayoría
de las anomalías se originan durante la embriogénesis, antes o después de esta etapa pueden
producirse defectos, de manera que ningún período del desarrollo está completamente seguro
ante los efectos adversos de la exposición a un tóxico. (3) los teratógenos actúan a través de
mecanismos específicos sobre las células y tejidos en desarrollo para dar lugar a una
embriogénesis anormal. Ellos ejercen sus efectos interfiriendo con los mecanismos normales de
desarrollo, produciendo en las células del organismo expuesto, entre otras consecuencias,
genotoxicidad con función alterada de ácidos nucleicos e interferencia mitótica (pudiendo generar
alteraciones en el número cromosómico), inhibición enzimática, deficiencia de sustancias
esenciales, suministro energético alterado, daño en las membranas y muerte celular. Lo referente
a la muerte celular y genotoxicidad se desarrollará en la sección 5. (4) las manifestaciones del
desarrollo anormal incluyen: malformaciones, retraso del crecimiento, trastornos funcionales e
inclusive la muerte. En tal sentido, muchos factores pueden interactuar con el embrión durante los
procesos de diferenciación y de crecimiento. Sin embargo, el resultado no siempre es una
malformación macroscópica. En algunos casos, el agente teratógeno es tan agresivo, o puede
afectar de manera tan significativa un órgano vital del embrión o del feto, que provoca su muerte.
En otros casos, la influencia del agente es más leve y el embrión o el feto logra sobrevivir, aunque
resulten afectados algunos de sus sistemas orgánicos (Sadler, 1995). Para entender esto, debemos

Introducción: el alcohol y el alcoholismo materno
9
considerar que el crecimiento embrionario/fetal normal depende de un suministro adecuado de
nutrientes y de una relación temporal estricta entre la aparición de un tejido y su diferenciación.
Cuando estos procesos resultan alterados por algún factor, las consecuencias negativas podrán
afectar o no la supervivencia embrionaria, fetal o posnatal. Así, muchas de las anomalías del
desarrollo que se originan durante el período embrionario se manifiestan tardíamente incluso
durante la vida postnatal, particularmente, si los defectos son relativamente leves. Defectos en
cerebro, pulmones, intestino y riñones durante su desarrollo, no comprometen la supervivencia
fetal dado que ellos no son esenciales para la vida intrauterina. Un ejemplo de ello son las
alteraciones del tubo neural originadas en fetos expuestos a ciertos teratógenos. Por el contrario,
dado que la supervivencia intrauterina depende de la adecuada función del saco vitelino, de la
placenta, del hígado fetal (sitio de hematopoyesis) y del sistema cardiovascular, la gran mayoría de
los casos de muerte embrionaria/fetal puede explicarse por defectos en uno de estos sistemas. La
formación de estos órganos y estructuras, por lo tanto, representan puntos de control para
desarrollo embrionario y fetal temprano (Fowden, 2001).
1.5.2. El síndrome de alcoholismo fetal.
Está bien establecido que el consumo materno de alcohol altera el desarrollo embrionario/ fetal
constituyendo, aún hoy, una de las complicaciones médicas y sociales de mayor relevancia
(Ceccanti y col., 2007; Mancinelli y col., 2007; Aragón y col., 2008). El alcohol, consumido durante
la gestación, es capaz de interferir en la morfogénesis de diversos órganos y generar, en
consecuencia, alteraciones físicas, fisiológicas, cognitivas y comportamentales permanentes e
irreversibles (Jones y Smith, 1973; Jones y col., 1973; Clarren y Smith, 1978; Streissguth y col., 1980;
Warren y Bast, 1988; Cook, 2003). La más severa de las manifestaciones teratogénicas del alcohol es
el denominado síndrome de alcoholismo fetal (FAS, del inglés Fetal Alcohol Syndrome), cuadro
patológico descripto en 1973 por Jones y Smith (1973) y observado por primera vez por Lemoine y
colaboradores en 1968, a partir un patrón común de defectos al nacimiento hallados en 127 niños
nacidos de madres alcohólicas en Francia (Lemoine y col., 1968). El FAS se puede producir en el
humano cuando la madre fue consumidora de alcohol antes de la gestación (Lee y Wakabayashi,
1985), durante la misma (Samson, 1981) o en días específicos de la preñez (Brannigan y Burke, 1982;
Brannigan y Cottel, 1984), por ejemplo durante la gastrulación y organogénesis embrionaria (Randall,

Introducción: el alcohol y el alcoholismo materno
10
1987; Ledig y col., 1991; Webster y Ritchie, 1991). El diagnóstico del FAS es realizado a través del
reconocimiento de ciertas características morfológicas y conductuales distintivas en el nuevo
individuo, entre ellas, (1) la deficiencia de crecimiento prenatal (restricción del crecimiento
intrauterino) y postnatal, con evidente reducción del peso y tamaño corporal (Abel, 1982); (2) la
presencia de anomalías craneofaciales características, tales como, microcefalia, microoftalmia, fisuras
palpebrales cortas, surco nasolabial poco desarrollado, labio superior delgado, micrognatia
(mandíbula poco desarrollada) (Rosett, 1980) (Figura 1.3); (3) la presencia de anomalías neurológicas,
tales como, deficiencia mental, déficit de atención, hiperactividad y comportamiento anormal (Becker
y col., 1996; Sampson y col., 2000; Hoyme y col., 2005). También están asociadas disfunciones
hormonales y metabólicas (inhibición de la síntesis proteica, desbalances hormonales, hipoglucemia,
disturbios vitamínicos, cambios en el balance ácido base) y orgánicas (defectos cardíacos, genitales
externos anormales y anomalías del oído medio, alteración del sistema muscular y esquelético) (Day y
Richardson, 1991; Froster y Baird, 1992). Actualmente el término FASD (del inglés, Fetal Alcohol
Spectrum Disorders) incluye y describe el amplio espectro de efectos que pueden aparecer en un
individuo luego de la exposición prenatal a alcohol (Riley y McGee, 2005).
Diferentes mecanismos se proponen para explicar los efectos teratogénicos del alcohol en el embrión
en desarrollo. Estos incluyen el aumento del estrés oxidativo; metabolismo alterado de glucosa,
proteínas, lípidos y ADN; deterioro de la neurogénesis y aumento de la apoptosis celular, en especial
de las células de la cresta neural; efectos endocrinos; efectos en la expresión génica (Ornoy y Ergaz,
2010); y alteraciones epigenéticas (Haycock, 2009; Liu y col., 2009), entre otros.
Figura 1.3. Representación esquemática de los rasgos craneofaciales característicos de un niño con FAS.
Modificado de Streissguth, 1994.

Introducción: el alcohol y el alcoholismo materno
11
Como con cualquier otro teratógeno, la severidad del FAS está dada por la combinación de la ingesta
materna de alcohol y factores de riesgo, tales como la cantidad y frecuencia de consumo, el período
de gestación en que se ingiere la droga, la edad materna, aspectos genéticos, nutricionales y físicos
(May y col., 2006). Varios estudios demuestran que la exposición prenatal a bajos niveles de alcohol
puede afectar negativamente el desarrollo fetal (Charness y col., 1994). Sin embargo, aún no se ha
podido establecer cuál es el nivel “seguro” de ingesta de alcohol durante la gestación que impida
modificaciones significativas en desarrollo embrio-fetal y placentario. Por lo tanto, se ha
recomendado que tanto mujeres gestantes como aquellas que deseen un embarazo eviten todo
consumo de bebidas alcohólicas, más aún considerando que los defectos pueden ser mayores si el
consumo ocurre en las fases iniciales del desarrollo (Cook, 2003).
1.6. Modelos experimentales para el estudio del los efectos de la ingesta materna de alcohol.
Debido a la gran importancia de los efectos adversos del consumo de alcohol a nivel médico-social, los
investigadores han concentrado esfuerzos en dilucidar los efectos y mecanismos de acción del alcohol
en el embrión/feto en desarrollo. Mucho de lo que se conoce sobre el FAS y sobre las consecuencias
de la exposición prenatal al alcohol deriva de estudios en animales de experimentación. Estos
modelos in-vivo han contribuido y aún contribuyen al entendimiento de los mecanismos que
conducen a los cambios morfológicos y de comportamiento postnatales que se originan en los
embriones/fetos expuestos al alcohol (Randall y col., 1987; Becker y col., 1996). En tal sentido, la
mayoría de las anomalías clínicas observadas en asociación al consumo materno de etanol, pudieron
ser replicadas en animales de experimentación de varias especies (ratón, rata, pollo, cerdo, perro,
mono). En particular, el ratón resulta apropiado para el estudio de los efectos teratogénicos del
alcohol donde tanto los rasgos faciales característicos del FAS (Figura 1.4), así como, otras
malformaciones estructurales, el retraso en el crecimiento pre y postnatal, la disfunción del sistema
nervioso central y las alteraciones conductuales han podido ser recreadas (Randall y col., 1987; Sulik y
col., 1988; Driscoll y col., 1990; Becker y col., 1996).
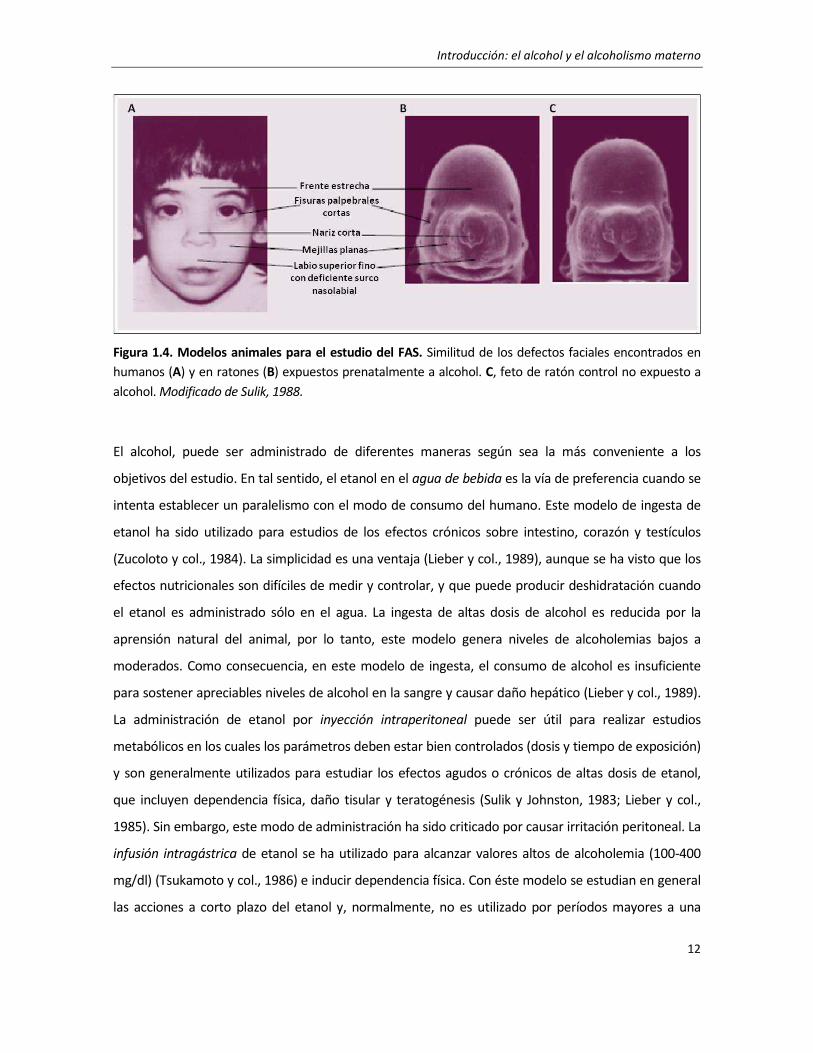
Introducción: el alcohol y el alcoholismo materno
12
Figura 1.4. Modelos animales para el estudio del FAS. Similitud de los defectos faciales encontrados en
humanos (A) y en ratones (B) expuestos prenatalmente a alcohol. C, feto de ratón control no expuesto a
alcohol. Modificado de Sulik, 1988.
El alcohol, puede ser administrado de diferentes maneras según sea la más conveniente a los
objetivos del estudio. En tal sentido, el etanol en el agua de bebida es la vía de preferencia cuando se
intenta establecer un paralelismo con el modo de consumo del humano. Este modelo de ingesta de
etanol ha sido utilizado para estudios de los efectos crónicos sobre intestino, corazón y testículos
(Zucoloto y col., 1984). La simplicidad es una ventaja (Lieber y col., 1989), aunque se ha visto que los
efectos nutricionales son difíciles de medir y controlar, y que puede producir deshidratación cuando
el etanol es administrado sólo en el agua. La ingesta de altas dosis de alcohol es reducida por la
aprensión natural del animal, por lo tanto, este modelo genera niveles de alcoholemias bajos a
moderados. Como consecuencia, en este modelo de ingesta, el consumo de alcohol es insuficiente
para sostener apreciables niveles de alcohol en la sangre y causar daño hepático (Lieber y col., 1989).
La administración de etanol por inyección intraperitoneal puede ser útil para realizar estudios
metabólicos en los cuales los parámetros deben estar bien controlados (dosis y tiempo de exposición)
y son generalmente utilizados para estudiar los efectos agudos o crónicos de altas dosis de etanol,
que incluyen dependencia física, daño tisular y teratogénesis (Sulik y Johnston, 1983; Lieber y col.,
1985). Sin embargo, este modo de administración ha sido criticado por causar irritación peritoneal. La
infusión intragástrica de etanol se ha utilizado para alcanzar valores altos de alcoholemia (100-400
mg/dl) (Tsukamoto y col., 1986) e inducir dependencia física. Con éste modelo se estudian en general
las acciones a corto plazo del etanol y, normalmente, no es utilizado por períodos mayores a una

Introducción: el alcohol y el alcoholismo materno
13
semana por la irritación asociada a la vía de aplicación (Lieber y col., 1989). Con las dietas líquidas es
posible estudiar los efectos crónicos del etanol cuando son necesarios elevados niveles de
alcoholemia, ya que con alrededor de un 35% de calorías derivadas del etanol se alcanzan 100-150
mg/dl de alcoholemia en los roedores (Lieber y col., 1989; Lieber y DeCarli, 1991). Sin embargo, se ha
criticado este régimen líquido ya que forzaría al animal a consumir cantidades preestablecidas de
ingredientes/suplementos alimentarios (Zucoloto y col., 1984).
1.7. Efectos del consumo materno de alcohol en el desarrollo embrio-fetal: antecedentes.
El consumo materno de alcohol es referido a la ingesta de alcohol por parte de la mujer durante la
gestación. Sin embargo si la ingesta ocurre en la etapa previa al embarazo, esta situación podrá
generar efectos deletéreos tanto a nivel reproductivo como a nivel gestacional. Por tal motivo se
hará mención en primer lugar de los efectos que produce el consumo de alcohol durante la fase
reproductiva (antes de la gestación) en el ciclo estral, la ovulación y sobre la gameta femenina; y
luego se abordan los antecedentes relativos a los efectos específicos del consumo materno de
alcohol en la gestación.
1.7.1. Efectos de la ingesta materna de alcohol en el sistema reproductivo femenino.
Considerando la importancia del normal desarrollo de la gameta femenina en la determinación del
desarrollo embrionario y fetal a término, se mencionan brevemente los efectos en el sistema
reproductivo femenino como consecuencia de la exposición materna a alcohol
Las acciones directas del alcohol sobre la gameta femenina se han estudiado exponiendo los
oocitos a distintas concentraciones de etanol. Así, los oocitos de los mamíferos, arrestados en la
segunda metafase meiótica, pudieron ser activados por exposición directa a etanol, conduciendo
al desarrollo de un embrión partenogenético. La activación de los oocitos por el etanol causa un
aumento transiente de calcio intracelular, que dura mientras permanece la exposición a etanol
(Cuthbertson, 1983; Kaufman, 1983; Shiina y col., 1993). El tipo de tratamiento o la exposición a
etanol podrá determinar la aparición de aberraciones cromosómicas. Así, se demostró que la
exposición a alcohol en oocitos de ratón no fecundados afecta directamente al huso durante la

Introducción: el alcohol y el alcoholismo materno
14
primera división meiótica causando no-disyunción cromosómica (Kaufman y Bain, 1984; Kaufman,
1985, 1997).
Por otro lado, se ha visto que la administración crónica de alcohol in-vivo, altera la ovulación y el
ciclo estral en animales de experimentación (Krueger y col., 1982; Mello y col., 1983). También, se
ha observado que ratas tratadas con etanol presentan marcada disrupción de la estructura ovárica
y de su función (Valimaki y col., 1984; Mello, 1988). Cebral y colaboradores, han demostrado que
la exposición de ratones hembras híbridos (C57/BL x CBA) a dosis bajas y moderadas de etanol en
el agua de bebida en forma prolongada (30 días) durante la etapa pregestacional resulta en una
marcada alteración de la ovulación, del ciclo estral (Cebral y col., 1998a) y un deterioro de la
calidad oocitaria (Cebral y col., 1998b).
1.7.2. Efectos de la ingesta materna de alcohol en el desarrollo embrio-fetal y placentario.
En la población humana, el consumo de alcohol pregestacional (antes de la gestación) en
condición de adicción crónica y comúnmente en forma severa, está asociado con la continuidad de
la ingesta de alcohol durante el embarazo, caso que determina el llamado consumo
perigestacional de alcohol. En esta problemática, es probable que, a partir del reconocimiento del
embarazo, la mujer alcohólica reduzca la ingesta de alcohol, aunque no lo evite completamente y
por lo tanto, la ingesta durante la gestación continuaría aunque de modo moderado. Dado que el
reconocimiento del embarazo, en la mayoría de los casos, no se produce hasta la 4ta-5ta semana
de gestación, la exposición a la droga se producirá así durante las etapas críticas del desarrollo
temprano del embrión. Por la importancia que tiene la ingesta de alcohol desde antes de la
gestación hasta los primeros meses de embarazo, en el desarrollo fetal, nuestro interés se centró
en abordar el estudio de las consecuencias en la formación del embrión y en el inicio de la
formación de la placenta luego de consumo materno de alcohol.
Es bien conocido que el etanol es tóxico y teratogénico para el desarrollo embrio-fetal, siendo sus
efectos dependientes de la dosis, la vía de administración y el período gestacional de exposición
(Chernoff, 1980; Randall, 1987; Blakley, 1988). En relación con lo último, no son pocos los estudios
realizados en cuanto a los efectos de la ingesta de alcohol en diferentes y específicos períodos de
la gestación. Considerando las fases de preimplantación, postimplantación temprana
(embrionaria) y fetal, los estudios fueron enfocados a dilucidar los efectos, en el desarrollo

Introducción: el alcohol y el alcoholismo materno
15
embrionario, de la exposición durante estos períodos de la gestación, tal como se describe a
continuación.
Las consecuencias teratogénicas de la exposición a etanol durante el período de preimplantación
han recibido una atención llamativamente escasa. Sin embargo, hoy en día, las investigaciones en
ratones, demuestran que la exposición a etanol in útero durante el período de preimplantación sí
se manifiesta posteriormente en resultados adversos. Ejemplo de ello es el estudio llevado a cabo
en ratones MF1, donde se demostró que la administración intraperitoneal de 5,8 g/kg de etanol en
cualquier día del período de preimplantación (días 1-4) da lugar a malformaciones graves similares
a las halladas en el FAS así como retraso en el crecimiento embrionario en el 80% de los embriones
viables, según la evaluación al día 15 de la gestación; efectos que se manifestaron también a nivel
del peso placentario (Padmanabhan y Hameed, 1988). Checiu y Sandor (1986) también
describieron la reducción en la tasa de implantación y características morfológicas anormales en el
embrión y feto del ratón expuesto al alcohol durante el período preimplantativo. Por otro lado,
numerosos trabajos basados en estudios in-vitro, también proveen evidencias sobre las
alteraciones en el crecimiento embrionario cuando los embriones de preimplantación son
expuestos directamente al etanol (Winston y col., 1991; Leach y col., 1993; Stachecki y col., 1994a,
1994b; Kowalczyk y col., 1996). Más aún, Cebral y colaboradores en diversos trabajos demostraron
que la ingesta crónica de dosis bajas a moderadas de etanol durante el período pregestacional
(solamente antes de la gestación) induce deterioro de la tasa de fecundación (Cebral y col., 1997) y
del desarrollo del embrión de preimplantación (Cebral y col., 1999; Cebral y col., 2000; Cebral y
col., 2001), en ratones híbridos (C57/BL x CBA).
Son numerosos los trabajos realizados en cuanto a los efectos de la exposición en el período de
postimplantación, viéndose las consecuencias de la exposición aguda o crónica. La administración
de dosis agudas (2,9 hasta 5,8 g/kg de etanol) en distintos momentos del desarrollo resultó en una
amplia gama de anomalías morfológicas, incluyendo malformaciones esqueléticas y de órganos
junto con aumento de las tasas de reabsorción embrionaria (Becker y col., 1996). En este sentido,
las anomalías craneofaciales, muchas de las cuales son sorprendentemente similares a los rasgos
faciales del FAS, resultan principalmente de dosis agudas aplicadas en los días 7, 8 y 9 de
gestación. Las anomalías en el cerebro parecen surgir después de tratamientos agudos en los días
7 y 8 (Becker y col., 1996). Sulik y colaboradores (1981), también demostraron que la

Introducción: el alcohol y el alcoholismo materno
16
administración aguda en un solo día de preñez en el ratón (día 7) fue suficiente para inducir en las
crías defectos faciales análogos a los observados en el FAS.
Los estudios que emplean regímenes de dosificación crónicos, con exposición a lo largo de toda la
gestación o durante varios días de ella, han informado de fenotipos característicos de todo el
espectro de FAS en el ratón. Estos estudios se han basado generalmente en dietas líquidas en las
que el alcohol proporciona el 15%-30% de las calorías de la dieta, aunque algunos estudios han
empleado también vía de administración intragástrica e intraperitoneal (Becker y col., 1996). Al
igual que en los estudios que emplean regímenes de dosis agudas, fueron observadas asociaciones
entre momento del desarrollo en que se administra el alcohol y las malformaciones halladas. El
sistema ocular, cardiovascular y esquelético parecen particularmente vulnerables a la exposición a
alcohol entre los días 4-12 de gestación, mientras que el sistema urogenital es particularmente
vulnerable entre los días 4-10; y el retraso del crecimiento parece fuertemente asociado a la
exposición crónica a alcohol durante la gestación tardía (12-17 días de gestación en ratón). El
sistema nervioso central, es sensible a la exposición crónica a alcohol a lo largo de todo el período
de gastrulación y organogénesis, fetal e inclusive posnatal (Becker y col., 1996). En modelos
animales, las anomalías derivadas de la exposición crónica al alcohol han incluido exencefalia,
hidrocefalia, microcefalia y varios defectos estructurales (Becker y col., 1996). Inclusive la
existencia de anomalías estructurales menos severas, como la reducción del volumen cerebral en
los casos de FAS, pueden delinear las bases de las distintas características cognitivas y
comportamentales de los individuos expuestos prenatalmente al alcohol (Spadoni y col., 2007).
Según lo expuesto antes y en relación con la ingesta materna de alcohol en un período
perigestacional, tal como puede ocurrir en el humano, la exposición desde antes de la gestación y
hasta diferentes períodos gestacionales no fue abordada completamente. Pocos autores, como
Soltes y colaboradores (1996), demostraron que la exposición crónica a etanol en ratones desde
antes y durante la gestación, produce alteraciones anatómicas fetales. Al presente, poco se sabe
acerca de la susceptibilidad del modelos murinos exocriados (CF-1, CD-1, entre otros) en relación a
la exposición a alcohol en la gestación. Recientemente, en nuestro grupo de trabajo hemos
demostrado que la ingesta oral considerada baja a moderada de etanol (10%) administrada
perigestacionalmente a ratones hembras CF-1 induce un significativo retraso de la diferenciación,
disminución de la viabilidad y el crecimiento, y defectos en la morfogénesis de los embriones
organogénicos al día 10 de gestación, particularmente a nivel del cierre del tubo neural (Coll y col.,

Introducción: el alcohol y el alcoholismo materno
17
2011). A pesar de estos antecedentes, tanto los efectos como los mecanismos, en el desarrollo
embrionario temprano inducidos por la ingesta oral periconcepcional, desde antes de la gestación
y hasta los primeros estadios de la preñez, incluyendo la organogénesis, en modelos murinos
exocriados, aún no han sido completamente clarificados.
Por otro lado, la exposición a alcohol tiene múltiples y complejos efectos adversos sobre el
crecimiento y función de la placenta, que en consecuencia pueden actuar indirectamente sobre el
desarrollo embrio-fetal. Por un lado, esto es debido a que el acetaldehído es capaz de atravesar la
placenta, la que posee la capacidad mínima de metabolización del alcohol, sumándose a que el
hígado fetal tampoco es un sistema eficaz para el clearance de alcohol (Burd y col., 2007). Por otro
lado, la exposición a alcohol afecta directamente a la estructura y función de la placenta.
Gundogan y colaboradores (2008, 2010) han demostrado, en ratas, que la exposición gestacional
crónica a etanol deteriora ampliamente el desarrollo de la placenta, produciendo reducción en el
espesor del órgano por un aumento en la necrosis celular. Dado que el intercambio de nutrientes
entre la madre y el feto se produce dentro de la barrera placentaria, las alteraciones inducidas por
el alcohol podrían poner en riesgo el suministro de nutrientes al feto contribuyendo así a la
restricción del crecimiento intrauterino luego de la ingesta de alcohol.
Por lo expuesto, se consideró la necesidad de investigar los efectos y algunos mecanismos de
inducción de potenciales anomalías embrionarias de preimplantación, del embrión en gastrulación
y evaluar una posible generación de anomalías feto-placentarias a término, luego de la ingesta
perigestacional de alcohol en el modelo murino exocriado.

Introducción: desarrollo embrionario, fetal y placentario murino
18
�
2. Desarrollo embrionario, fetal y placentario murino.
2.1. Desarrollo de preimplantación.
Durante la mayor parte del desarrollo embrionario en los mamíferos, el embrión se desarrolla
unido al útero materno. Sin embargo, el desarrollo de preimplantación abarca un período de vida
libre durante la embriogénesis, etapa que culmina con la formación del blastocisto (Watson,
1992), cuya extrusión de la zona pelúcida (hatching) es el prerrequisito para que ocurra
implantación (Brison y Leese, 1994).
Una de las características del desarrollo de preimplantación es la generación de dos linajes
celulares con distinto destino, el linaje embrionario y el linaje extraembrionario (trofoectodermo),
este último requerido para la implantación del embrión en el útero (Sasaki, 2010).
En el ratón, luego de la cópula, los oocitos son fecundados cerca de la medianoche del día de la
ovulación. Durante la interacción gamética, el espermatozoide descondensa su cromatina para dar
lugar al pronúcleo (PN) masculino. El oocito detenido en metafase II libera su bloqueo meiótico, el
segundo cuerpo polar es extruido y se forma el PN femenino (Payne y col., 1997). Tras la aposición
de los dos PNs, se organiza la primera división mitótica del embrión. Para ello, se desorganiza la
envoltura nuclear de cada PN y el material genético se condensa formando los cromosomas, los
cuales se orientan en un huso mitótico común (singamia). Los centríolos arrastran las cromátides
hacia los polos opuestos y luego se produce la división del citoplasma, con la formación de dos
células hijas diploides que contienen aproximadamente la mitad del contenido citoplasmático
oocitario. Las divisiones mitóticas sucesivas en el embrión, resultan en un rápido aumento en el
número de células, pero sin un incremento del tamaño embrionario, hasta la formación de la
cavidad blastocélica (Johnson y McConnell, 2004). La duración total del primer ciclo de división
celular, desde la fecundación hasta el primer clivaje, es de aproximadamente 18 hs. Después de
otras 36 hs y dos divisiones sincrónicas más, se alcanza al día 3 de gestación, el estadio de 8
células. A partir de este momento, las divisiones sucesivas comienzan a ser asincrónicas (Kaufman
y Bard, 1999). El segundo ciclo de división, como el primero, es relativamente largo (~18 hs),
mientras que el tercero y los subsiguientes adoptan la duración más convencional observada en
las células somáticas (~12 hs) (Pratt, 1987). Hasta el estadio de 8 células, los límites de las
blastómeras son evidentes, siendo cada célula claramente individualizada. Hasta este estadio es

Introducción: desarrollo embrionario, fetal y placentario murino
19
�
posible demostrar que cada blastómera es totipotente siendo capaz, bajo condiciones
experimentales adecuadas, de formar un feto a término normal y viable (Kelly, 1975, 1979).
Dos eventos morfogenéticos particularmente importantes ocurren durante el desarrollo de
preimplantación, estos son: la compactación y la cavitación, eventos que llevarán a la formación
del blastocisto, embrión preparado para iniciar la implantación (Watson, 1992).
El primer evento, la compactación, surge en el ratón luego del comienzo del estadio de 8 células
(4to ciclo de clivaje), e involucra un período de aproximadamente 4 a 5 hs desde el inicio. La
compactación se manifiesta por un incremento en los contactos intercelulares que llevan a la
pérdida de definición de los límites blastoméricos, dando lugar a la formación de la mórula
(Watson, 1992). El término mórula se refiere a aquellos embriones cuyas blastómeras se
encuentran con cierto grado de agregación/compactación. Esto abarca desde embriones de 8
células compactados (mórula temprana) hasta 16 y 32 células (mórula tardía). La compactación
representa el inicio de la diferenciación celular durante el desarrollo embrionario de los mamíferos
(Pratt y col., 1982), y está asociada al establecimiento de la polaridad celular en las blastómeras
más externas (Schultz, 1986). El embrión continúa dividiéndose hasta alcanzar las 16 células.
Durante dicha mitosis, la mórula compactada sufre una descompactación transitoria, para luego
compactarse nuevamente. En el estadio de mórula compactada, el embrión presenta dos capas,
una capa de células polarizadas externas que encierra un pequeño grupo de células apolares
internas (Hogan y col., 1986; Johnson y McConnell, 2004). Esta posición relativa diferencial se
refuerza por propiedades adhesivas distintivas entre las dos poblaciones celulares. De tal manera
que las células externas tienden a rodear y encerrar, mediante uniones estrechas (principalmente
moléculas zonula occludens, ZO-1) (Fleming y col., 1989) y uniones adherentes (E-cadherinas)
(Hyafil y col., 1980; Watson y col., 1990b), a las blastómeras apolares internas, entre las cuales
predominan la uniones de tipo comunicante (gap). Aquellas células polarizadas externas
contribuirán a la progenie del linaje trofoblástico, mientras que las apolares internas, con gran
plasticidad, contribuirán a la progenie del linaje pluripotente embrionario (Johnson y McConnell,
2004).
La compactación es crucial para el desarrollo, dado que ella contribuye al inicio y progresión del
segundo gran evento del desarrollo preimplantacional: la cavitación. La cavitación ocurre entre el
5to y 6to ciclo de división celular, y se manifiesta tempranamente por la acumulación de fluido
entre las blastómeras internas cuando el embrión tiene aproximadamente 32 células y se
encuentra en tránsito hacia el útero (Pratt, 1987; Motosugi y col., 2005). Para que la cavitación

Introducción: desarrollo embrionario, fetal y placentario murino
20
�
ocurra, es requisito indispensable que el embrión haya desarrollado un epitelio polarizado,
constituido por las blastómeras externas, así como la presencia de mecanismos para el transporte
de fluido para formar la cavidad. En tal sentido, la distribución polarizada de bombas Na+/K
+-
ATPasas desempeña un papel fundamental en el establecimiento de un gradiente de iones Na+,
que resulta finalmente en la movilización osmótica de agua a través del epitelio y su acumulación
en la cavidad blastocélica naciente (Borland, 1977; Wiley, 1988; Biggers y col., 1988, 1991;
Gardiner y col., 1990a). El mantenimiento de dichas bombas Na+/K
+-ATPasas en el dominio
basolateral de las blastómeras externas está influenciado por la estabilización de las uniones
estrechas y adherentes (Fleming y col., 1989; Watson y col., 1990b). A su vez, factores de
crecimiento, tales como, TGF-α (del inglés, Transforming Growth Factor-alpha) y EGF (del inglés,
Epidermal Growth factor) estimulan el desarrollo de dicha cavidad mediando, en parte, la
expresión de segundos mensajeros que resultan en un aumento de la actividad de la Na+/K
+-
ATPasa. Todo esto conduce a la formación de una cavidad central llamada cavidad blastocélica o
blastocele y a la diferenciación de dos linajes: el trofoectodermo (TE) y el macizo celular interno
(MCI), momento en el que el embrión pasa a ser denominado blastocisto. En el estadio de 32
células, ocurre por lo tanto, la formación del blastocisto inicial o temprano (día 3,5-4 de gestación
murina). En el estadio de 64 células, el blastocisto inicial se expande por acumulación continua de
líquido en la cavidad, la cual se presenta bien definida, con un MCI protruyendo hacia la cavidad y
un anillo de células TE alrededor, las cuales son designadas como células trofoectodérmicas
polares (TP) si se encuentran cubriendo al MCI o como trofoectodérmicas murales (TM) si se
encuentran recubriendo al blastocele. El embrión en este estadio adquiere el nombre de
blastocisto expandido (Johnson y McConnell, 2004).
Con aproximadamente 128 células, además de mostrar signos de adelgazamiento de la zona
pelúcida, una capa de células derivadas del MCI, forman el hipoblasto (o endodermo primitivo),
mientras que las células remanentes del MCI, mantienen su pluripotencialidad y son ahora
llamadas epiblasto (o ectodermo primitivo). El blastocisto, en ese momento (día 5 de gestación),
como consecuencia de la acumulación de fluido y por la acción de enzimas proteolíticas, se libera
de la zona pelúcida (hatching) (Johnson y McConnell, 2004), estando ya preparado para iniciar la
adhesión en el epitelio uterino y comenzar así el proceso de implantación.
En la Figura 2.1 y en la Tabla 2.1, se resumen los eventos principales del desarrollo de
preimplantación, indicando el timing de cada uno de ellos. La Figura 2.2, muestra en forma
esquemática, los linajes celulares derivados del embrión temprano.

Introducción: desarrollo embrionario, fetal y placentario murino
21
�
Figura 2.1. Representación esquemática de los principales eventos del desarrollo de preimplantación
murino. Se indica el número de ciclos celulares desde el primer clivaje hasta la formación del
blastocisto; los días de gestación (D), considerando D1 al día de la observación del tapón vaginal; los
estadios embrionarios (E), considerando 0,5 el día de observación del tapón (según Kaufman y Bard,
1999); y la clasificación según la escala de Theiler (TS; Theiler, 1989) basada en la morfología de los
embriones. Bc: blastocele, Embr: embrión, FEC: fecundación, IMPL: implantación, MCI: macizo celular
interno, TE: trofoectodermo, TM: trofoectodermo mural, TP: trofoectodermo polar. Modificado de Saiz
y Plusa, 2013; Czechanski y col., 2014.

Introducción: desarrollo embrionario, fetal y placentario murino
22
�
Figura 2.2. Diagrama que ilustra los linajes celulares derivados del embrión de preimplantación. TE:
capa externa de tipo epitelial. TM: progenitor de las CGTs1rias
involucradas en la implantación en la
zona antimesometrial del útero. TP: progenitor del ectodermo extraembrionario y del cono
ectoplacentario involucrado en la implantación en la zona mesometrial y en el desarrollo de la
placenta. MCI: se diferencia en el epiblasto, grupo de células pluripotentes, progenitoras de las tres
capas germinales del feto y de las membranas extraembrionarias; y en el hipoblasto, quien formará
parte de los tejidos extraembrionarios. (*): será reemplazado por endodermo definitivo. CGTs: células
gigantes del trofoblasto. Modificado de Kaufman y Bard, 1999.
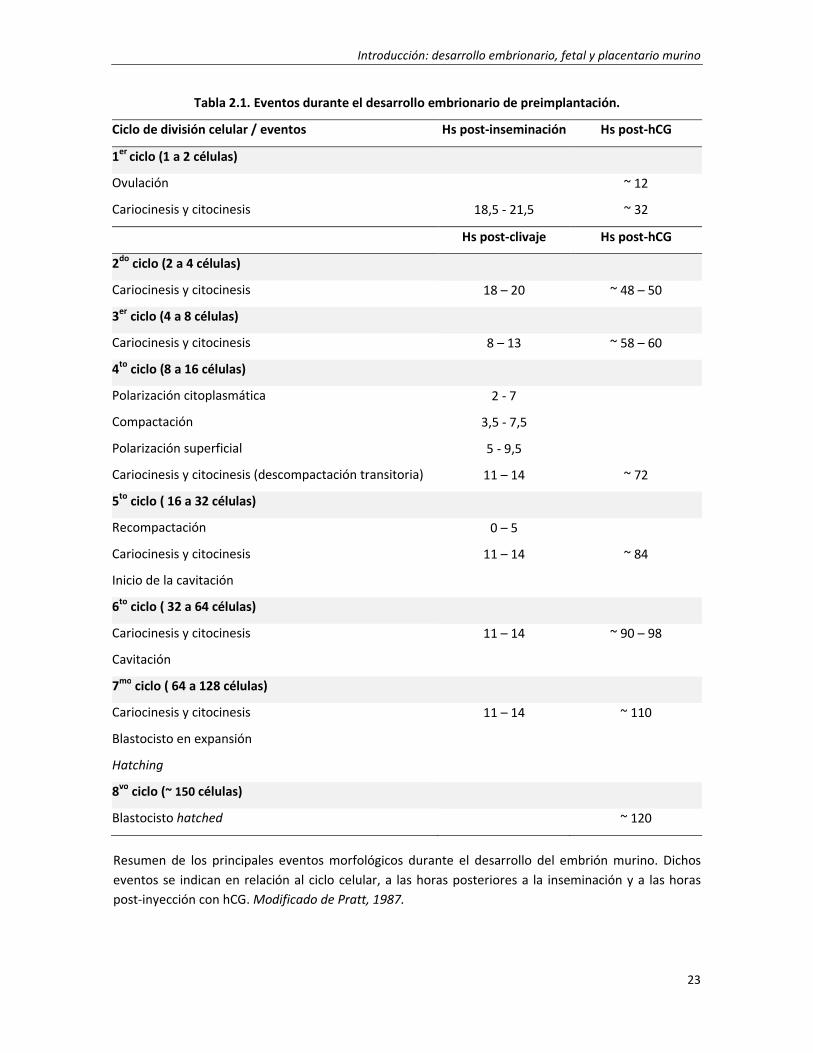
Introducción: desarrollo embrionario, fetal y placentario murino
23
�
Tabla 2.1. Eventos durante el desarrollo embrionario de preimplantación.
Ciclo de división celular / eventos Hs post-inseminación Hs post-hCG
1er
ciclo (1 a 2 células)
Ovulación ~ 12
Cariocinesis y citocinesis 18,5 - 21,5 ~ 32
Hs post-clivaje Hs post-hCG
2do
ciclo (2 a 4 células)
Cariocinesis y citocinesis 18 – 20 ~ 48 – 50
3er
ciclo (4 a 8 células)
Cariocinesis y citocinesis 8 – 13 ~ 58 – 60
4to
ciclo (8 a 16 células)
Polarización citoplasmática 2 - 7
Compactación 3,5 - 7,5
Polarización superficial 5 - 9,5
Cariocinesis y citocinesis (descompactación transitoria) 11 – 14 ~ 72
5to
ciclo ( 16 a 32 células)
Recompactación 0 – 5
Cariocinesis y citocinesis 11 – 14 ~ 84
Inicio de la cavitación
6to
ciclo ( 32 a 64 células)
Cariocinesis y citocinesis 11 – 14 ~ 90 – 98
Cavitación
7mo
ciclo ( 64 a 128 células)
Cariocinesis y citocinesis 11 – 14 ~ 110
Blastocisto en expansión
Hatching
8vo
ciclo (~ 150 células)
Blastocisto hatched ~ 120
Resumen de los principales eventos morfológicos durante el desarrollo del embrión murino. Dichos
eventos se indican en relación al ciclo celular, a las horas posteriores a la inseminación y a las horas
post-inyección con hCG. Modificado de Pratt, 1987.

Introducción: desarrollo embrionario, fetal y placentario murino
24
�
2.2. Implantación embrionaria.
En mamíferos, el sistema reproductor femenino está anatómicamente adaptado para que durante
la gestación el embrión sea recibido en un medio interno anatómica y funcionalmente permisivo, y
adecuado para su desarrollo. Esto ocurre gracias a un proceso conocido como implantación
embrionaria. Durante esta fase, el embrión establece un posicionamiento fijo en el útero, que
favorece la formación de una superficie de intercambio metabólico para la adquisición de
nutrientes del ambiente materno, independientemente del medio externo.
En el ratón, el proceso de implantación embrionaria es precoz en la gestación y se caracteriza por
la formación de una íntima relación entre los tejidos maternos y embrionarios. Estas interacciones
evolucionan a la largo de la gestación, se tornan cada vez más complejas y culminan con la
formación de la placenta.
A diferencia de las especies cuyo desarrollo embrionario se da en el lumen uterino, el desarrollo
de los embriones de primates y roedores ocurre totalmente en el intersticio uterino. La eficiencia
de esto depende en gran medida de la actividad altamente invasiva exhibida por las células
gigantes del trofoblasto (Bevilacqua y Abrahamsohn, 1989), así como también del desarrollo de
mecanismos facilitadores o permisivos por parte del organismo materno. Aunque las células
trofoblásticas comparten grandes similitudes con las células malignas en la forma en la que
invaden los tejidos y rompen los vasos sanguíneos, en condiciones normales, no comprometen la
fisiología uterina.
Si bien el detalle del proceso de implantación es variable en las diferentes especies, de modo
general, este proceso puede ser subdividido en las siguientes fases: aposición y adhesión, para
embriones que se desarrollan en el lumen uterino (placentación epiteliocorial), e invasión, para
aquellos que presentan alguna interacción con estructuras del estroma endometrial (placentas
sindesmocoriales, endoteliocoriales y hemocoriales) (Pijnenborg y col., 1985; Pratt, 1987) (Figura
2.3). En primates y roedores, durante el desarrollo de la placenta hemocorial, con el avance de la
gestación, el embrión penetra el estroma endometrial, rompe vasos sanguíneos endometriales e
interactúa directamente con la sangre materna.
Para que el proceso de implantación se inicie es necesario que el embrión y el organismo materno
estén perfectamente sincronizados. El endometrio debe haber sido sensibilizado por una
secuencia hormonal específica que le permita tornarse receptivo al embrión y éste, a su vez, debe

Introducción: desarrollo embrionario, fetal y placentario murino
25
�
haber alcanzado la etapa de blastocisto expandido, mostrándose morfológica y funcionalmente
preparado para la interacción con el organismo materno (Weitlauf, 1994).
A partir del día 5 de gestación, los blastocistos ya sin zona pelúcida se encuentran distribuidos a lo
largo de los cuernos uterinos, alojados en la cripta uterina antimesometrial, preparados para
iniciar el proceso de implantación embrionaria. Es a través de las células trofoblásticas que hacen
contacto e interactuarán con el organismo materno durante toda la gestación (Billington, 1971).
Figura 2.3. Representación esquemática de las fases de implantación del blastocisto murino. Luego
del hatching, mediado por señales químicas el TE del blastocisto inicia la orientación y contacto con el
epitelio uterino. Le sigue la aposición, en la que el TM se asocia de manera inestable al útero. Prosigue
la fase de adhesión, donde la unión entre el TM y el epitelio uterino es firme, surgiendo
interdigitaciones entre ambos. Las células del TE han iniciado su diferenciación a células trofoblásticas
invasivas, conduciendo a la siguiente fase, la invasión, donde el blastocisto penetra el epitelio luminal y
el estroma uterino. LB: lámina basal, MCI: macizo celular interno, TE: trofoectodermo, TM:
trofoectodermo mural, TP: trofoectodermo polar, ZP: zona pelúcida. Modificado de Bazer y col., 2009.
En la fase de aposición, el blastocisto sin zona pelúcida se apoya sobre el epitelio uterino, donde
las superficies de las células trofoblásticas murales y del epitelio uterino se aproximan (Potts,
1969). Un gradual cierre de la luz uterina alrededor del embrión lleva a un contacto más íntimo
entre las superficies trofoblásticas y epiteliales uterinas (Enders y Schlafke, 1967). Luego la
siguiente fase es la de adhesión, que se caracteriza por una notable disminución del espacio entre
los organismos, materno y embrionario (Potts, 1966; Reinius, 1967). A medida que esta fase

Introducción: desarrollo embrionario, fetal y placentario murino
26
�
progresa, las proyecciones de las membranas del trofoblasto y del epitelio uterino se interdigitan,
estableciéndose una relación cada vez más íntima y compleja (Reinius, 1967; Potts, 1968). Dada la
importancia de la adhesión para el éxito de la implantación, un amplio espectro de moléculas
participan de este evento. A su vez, en el organismo materno el desarrollo de la decidua es parte
fundamental de este proceso de implantación e involucra numerosas transformaciones en el
estroma uterino que llevan a la formación de un tejido morfológica y funcionalmente diferenciado
del tejido conjuntivo que le dio origen. Dichas modificaciones que ocurren en el endometrio,
preparándolo para recibir al embrión se denominan reacción decidual, donde los fibroblastos del
estroma uterino adquieren una forma poliédrica y disposición típicamente epiteloide,
transformándose en células deciduales (Abrahamsohn, 1983). La gran proliferación de estas
células deciduales ocurre hasta el noveno día de gestación (Finn y Martin, 1967), luego del cual,
entran en un masivo proceso degenerativo (Katz y Abrahamshon, 1987). El conjunto de
modificaciones morfológicas y bioquímicas observadas en el endometrio, contribuyen a la
instalación de un período transitorio de receptividad uterina, donde el embrión y los tejidos
maternos, fisiológicamente sincronizados, participan del proceso de recepción del embrión,
período denominado ventana de implantación (Rogers y Murphy, 1989).
Como se mencionó anteriormente, en especies que desarrollan una placenta hemocorial, el
proceso de implantación prosigue a una fase de invasión, que lleva al embrión a alojarse en el
estroma endometrial, perdiendo su contacto con el lumen uterino (Mossman, 1987). En roedores,
esta fase se caracteriza por el desplazamiento del epitelio uterino de su membrana basal y la
gradual fagocitosis por parte de las células trofoblásticas murales (presentes en la región
antimesometrial), hasta que esa capa no se interponga más entre el embrión y el estroma
endometrial (Bevilacqua y Abrahamsohn, 1989). Evidencias morfológicas muestran que dichas
células trofoblásticas murales sufren un proceso de diferenciación pasando a presentar enormes
dimensiones, amplios núcleos poliploides (Barlow y col., 1972) y características estructurales de
células fagocíticas y migratorias, adquiriendo el nombre de células gigantes del trofoblasto
primarias (CGTs 1rias
) (Dickson, 1963; Bevilacqua y Abrahamsohn, 1988, 1989). El desplazamiento
del epitelio uterino está a su vez favorecido por un proceso pre-programado de muerte celular por
apoptosis (Parr y col., 1987; Parr y Parr, 1989a, 1989b). Al atravesar el epitelio uterino, las células
trofoblásticas que envuelven al embrión, inician el acceso al estroma uterino donde se
establecerán definitivamente y mantendrán una interacción con la sangre materna hasta el final
de la gestación (Bevilacqua y Abrahamsohn, 1989). Las células trofoblásticas que permanecen en

Introducción: desarrollo embrionario, fetal y placentario murino
27
�
el polo mesometrial, al contrario de las demás, mantiene su capacidad proliferativa y
gradualmente forman una estructura saliente denominada cono ectoplacentario, que dará origen
a la cara fetal de la placenta. Las células más periféricas del cono ectoplacentario también se
diferencian, aunque más tardíamente, en células gigantes del trofoblasto secundarias (CGTs 2rias
)
(Gardner y col., 1973; Copp, 1979), se disponen periféricamente y participan de la implantación
del embrión en la región uterina mesometrial (Kaufman, 1983).
A medida que avanza la invasión, y una vez alcanzados los vasos endometriales, las células
trofoblásticas emiten prolongaciones entre las células endoteliales, las que al ser desplazadas,
permiten el acceso a la sangre materna (Bevilacqua, 1984; Welsh y Enders, 1987). La superficie de
contacto entre la sangre materna y el trofoblasto aumenta a medida en que ésta pasa a circular
entre las células trofoblásticas, células que se disponen en forma de red alrededor del embrión
(Welsh y Enders, 1987). Esta interfase trofoblástico-decidual, gradualmente, se especializa en la
función de intercambio metabólico entre los dos organismos, materno y fetal. En las fases iniciales
de esta interacción, una intensa actividad fagocítica por parte del trofoblasto es observada en
relación a las células sanguíneas (Bevilacqua y Abrahamsohn, 1988). Se cree que esta actividad
contribuye al suministro de los requerimientos básicos para el desarrollo embrionario hasta que la
placenta este completamente madura (Billington, 1971; Bevilacqua y Abrahamsohn, 1988, 1989).
En las fases más avanzadas, los vasos sanguíneos uterinos son parcialmente revestidos por las
células trofoblásticas (Bevilacqua, 1984; Welsh y Enders, 1987), y como consecuencia, la sangre
materna extravasada pasa a circular entre los espacios de la capa de CGTs que recubre al embrión.
El proceso de diferenciación de las CGTs 1rias
y 2rias
no involucra sólo un evento morfológico, sino
que también está asociado con la adquisición de innumerables funciones por parte de estas
células, tales como, las mencionadas capacidades adhesivas, invasivas, fagocíticas y endócrinas
(Soares y col., 1991, 1993). Diversas evidencias indican que el aumento de las dimensiones
celulares y nucleares, ocurre como consecuencia del proceso de poliploidización, el cual permitiría
la maximización y concomitancia de las actividades de estas células (Copp, 1979; Zybina y Zybina,
1996). En el caso de la endorreduplicación, el aumento en la cantidad de ADN en la célula no lleva
a cambios estructurales en los cromosomas. Las cromátides formadas en cada cromosoma por la
sucesión de endociclos (replicación periódica del ADN) no se separan, permaneciendo el número
de cromosomas inalterado. El ciclo celular corto e independiente de mitosis aumenta los niveles
de ploidía (D`Amato, 1989) otorgando a estas células trofoblásticas una ventaja en relación a otras
células que necesitan concluir ciclos de división celular y nuclear para lograr producir el mismo

Introducción: desarrollo embrionario, fetal y placentario murino
28
�
número de productos. Por lo tanto, la endorreduplicación en este tipo de células, ha sido referida
como un mecanismo adaptativo para adecuar las necesidades de una rápida diferenciación y
favorecer la invasión, produciendo hormonas polipeptídicas y otras proteínas regulatorias,
esteroides, diversos receptores, mediadores de intercambios metabólicos relacionados con la
nutrición, protección e inmunomodulación (Sherman, 1983; Bevilacqua y Abrahamsohn, 1988; Lin
y col., 1993), durante la gestación.
2.3. Desarrollo de postimplantación temprano (TS6-TS11).
Alrededor del estadio E3,5/TS4, las células de la superficie inferior del MCI se delaminan para
formar el hipoblasto o endodermo primitivo (Figura 2.1). El resto de las células del MCI forman el
epiblasto o ectodermo primitivo, las cuales retienen su pluripotencialidad. Luego en la transición
del blastocisto (E3,5/TS4) al embrión en estadio de huevo cilíndrico temprano-TS7, ya implantado,
dos cambios importantes son observados en la configuración del embrión. En primer lugar,
durante el E4/TS5, existe una migración periférica de células del endodermo primitivo desde la
zona de delaminación, en la región subyacente a la superficie del MCI con la cavidad blastocélica
(Figura 2.1; Figura 2.4). Así, el endodermo primitivo delimitará o cubrirá la cavidad blastocélica en
toda su extensión, incluida la superficie lindante con el TE mural, donde formara el endodermo
extraembrionario parietal, con un aspecto aplanado característico. Luego de completar esta
migración, la cavidad blastocélica cambia su nombre por el de cavidad del saco vitelino (Snell y
Stevens, 1966). En segundo lugar y en simultáneo, las células del epiblasto cubiertas por una capa
de endodermo visceral, proliferan notablemente, debido a una intensa actividad mitótica, con el
concomitante aumento de volumen. Este crecimiento, provoca el desplazamiento o expansión de
la masa de células internas del epiblasto hacia la cavidad de saco vitelino. Este evento determina
un cambio en la forma del embrión, asemejándose a un cilindro, y adquiriendo entonces el
nombre de embrión en estadio de huevo cilíndrico.
Durante el estadio de huevo cilíndrico (E6/TS8) (Figura 2.4), algunos cambios ocurren en la
configuración del concepto, siendo el más obvio, la aparición de la cavidad proamniótica dentro
del epiblasto. El epiblasto, ubicado en la región antimesometrial del concepto, en relación a sus
características histológicas, presenta células de tipo cuboidal/columnar. La superficie externa del
epiblasto está recubierta por una capa de endodermo visceral, mientras que las células que
delinean el trofoblasto mural constituyen el endodermo extraembrionario parietal, anteriormente

Introducción: desarrollo embrionario, fetal y placentario murino
29
�
mencionado. En este estadio se evidencia por primera vez la membrana de Reichert. Esta consiste
en un material acelular secretado por el endodermo parietal, el cual separa al endodermo parietal
de la subyacente capa de células trofoblásticas murales, muchas de las cuales, en este estadio, ya
se han diferenciado en CGTs 1rias
. Simultáneamente, la parte central del trofoectodermo polar
comienza a proliferar, formando el cono ectoplacentario (CE), el cual delimita la superficie
mesometrial de huevo cilíndrico y actúa como una punta de lanza avanzando en el tejido materno.
Esta estructura será parte, posteriormente, de la placenta corioalantoidea. Las células más
periféricas del CE, se diferencian, como se mencionó anteriormente, en CGTs 2rias
, y en este
estadio comienzan a migrar dentro del tejido decidual en dirección mesometrial. En el margen
periférico del trofoblasto polar, cercano al CE, la membrana de Reichert se une con el endodermo
visceral. En esta región, el endodermo presenta una morfología columnar y una alta actividad
metabólica, comparada con el endodermo visceral de tipo escamoso del embrión (Gupta y col.,
1982).
Durante el estadio E6,5-7/TS9, la cavidad proamniótica ya se ha extendido en dirección al CE,
formando el canal proamniótico, estando limitado en su región distal, por ectodermo embrionario
y en su región proximal por ectodermo extraembrionario derivado del trofoblasto polar (Copp,
1979), cuyas células presentan una morfología cuboidal. Por lo tanto, en este estadio se
evidencian claramente las dos regiones diferenciadas una región distal embrionaria y una región
proximal extraembrionaria. Este estadio es denominado como estadio de huevo cilíndrico tardío.
Las dos partes del concepto (la parte embrionaria y la extraembrionaria) continúan cubiertas por
una capa simple de endodermo visceral, que se formó previamente (TS5). Las células que
delimitan el polo embrionario tienden a mostrar una morfología principalmente escamosa
mientras que aquellas que delimitan el ectodermo extraembrionario del concepto, presentan una
apariencia cuboidal o, inclusive, columnar. Estas células endodérmicas extraembrionarias, como se
describió previamente, poseen vacuolas apicales y presentan microvellosidades en el borde apical.
La zona de transición entre la región cubierta por células de morfología escamosa y aquellas de
morfología cuboidal/columnar, no siempre coincide exactamente con el límite entre los
componentes embrionarios y extraembrionarios del concepto.
El embrión, localizado en el polo embrionario del huevo cilíndrico, está enteramente formado por
el epiblasto o ectodermo primitivo, siendo esta capa la progenitora del ectodermo neural así como
del ectodermo superficial del embrión (Lawson y col., 1991).

Introducción: desarrollo embrionario, fetal y placentario murino
30
�
El CE derivado del trofoblasto polar, en expansión, invade los vasos sanguíneos maternos, y en
consecuencia las células trofoblásticas se bañan de sangre materna. Este evento representa, por lo
tanto, el primer paso en la formación de un sitio especializado de intercambio metabólico entre la
madre y el embrión, y posteriormente el feto, cuando la placenta definitiva este establecida.
En los estadios TS8-9, por lo tanto, el componente embrionario así como el extraembrionario son
bilaminados, constando de una capa interna de ectodermo y una capa externa de endodermo.
Particularmente, los roedores, son la excepción comparado con otros mamíferos en cuanto a la
orientación de las capas germinales, dado que en esta etapa, las capas están invertidas en relación
a lo normalmente observado en el resto de los mamíferos (por ejemplo en el embrión humano).
Así, las capas germinales de los embriones de roedores logran invertirse y alcanzar su disposición
definitiva (con el ectodermo orientado hacia el exterior) durante el proceso de rotación, evento
posterior a la gastrulación.
Hacia el final del estadio TS9, y asociado con un incremento en el volumen del canal proamniótico
que se formó en el TS8, ambas, la región embrionaria y extraembrionaria adquieren una definición
más precisa. Mientras el componente ectodérmico de ambas regiones tiende a ser de morfología
columnar, aquellas pertenecientes al ectodermo embrionario tienden a ser más elongadas aún.
En etapas más avanzadas de este mismo estadio, es posible reconocer evidencias del inicio de la
gastrulación, con la delaminación intra-embrionaria del mesodermo (Figura 2.4, E6,5-7/ST9a)
(Reinius, 1965). Durante el proceso de gastrulación, estas células migran desde el ectodermo
embrionario hacia y a través de la línea primitiva, y a medida que se separan de la línea media
embrionaria, van separando el ectodermo del subyacente endodermo. Una vez que se evidencia el
movimiento del mesodermo intra-embrionario, se considera que el embrión ha alcanzado el
estadio de estadio de línea primitiva temprana (Downs y Davies, 1993), estadio denominado
TS10a (Figura 2.4).
Durante el estadio de línea primitiva (E7-7-5/TS10), es el momento en que transcurre la mayor
parte del proceso de gastrulación, siendo muy difícil de seguir en detalle en el ratón. El surco
primitivo, el cual aparece por primera vez en el TS7 en la región posterior/caudal del epiblasto, se
extiende anterior/rostralmente hacia la región cefálica presuntiva proveyendo la primera
evidencia morfológica de un eje embrionario. Allí, en el extremo rostral de la línea primitiva, se
forma el nodo que si bien en el ratón no está tan claramente diferenciado y evidente como el
nodo de Hensen en el pollo, éste juega un papel similar controlando la subsecuente morfogénesis
del embrión murino.

Introducción: desarrollo embrionario, fetal y placentario murino
31
�
Alrededor del TS10a, los movimientos celulares que caracterizan la gastrulación comienzan e
involucran tres actividades coordinadas. Una proporción de células dentro del epiblasto se
mueven hacia el surco primitivo y a través del nodo, el que migra caudalmente. A través del nodo,
estas células se desprenden del ectodermo y se convierten en células endodérmicas definitivas
que desplazan a las primitivas, mientras que las células que migran más tardíamente, forman el
mesodermo embrionario. Algunas de estas células del nodo migran rostralmente para formar el
proceso notocordal, que más tarde será reemplazado por la notocorda definitiva.
El proceso de gastrulación continúa por algunas horas hasta que el nodo se mueve caudalmente.
Otro evento importante durante el inicio del estadio TS10ab, es la aparición del pliegue amniótico
posterior, el cual se forma y crece a partir de una masa de células ectodérmicas extraembrionarias
ubicadas en la línea media dorsal, justo en el límite entre el ectodermo embrionario y el
extraembrionario, y en posición caudal a la región de la línea primitiva. Posteriormente, una
cavidad se forma dentro del pliegue, la que finalmente constituirá la cavidad exocelómica, al
fusionarse el pliegue amniótico posterior con un segundo pliegue llamado pliegue amniótico
anterior. Este se desarrolla en la línea media del lado opuesto del concepto, en el ectodermo
extraembrionario proximal a la región donde presuntivamente se formará el pliegue cefálico de
ectodermo neural. Esta cavidad quedará delimitada por una capa de mesodermo
extraembrionario delaminado del ectodermo.
En esta etapa, el canal proamniótico presenta una región central estrecha, que se cerrará una vez
que los pliegues amnióticos se encuentren y se fusionen formando la cavidad exocelómica. Este
evento ocurre hacia el final del TS10 y tales embriones se designan en estadio de línea primitiva
tardía. En el estadio TS10b, en el concepto quedarán definidas, entonces, tres cavidades siendo de
proximal a distal, la cavidad ectoplacentaria, la exocelómica y la amniótica, separadas
respectivamente por el corion y el amnios. Como se mencionó anteriormente, existe un cuarto
compartimento, la cavidad del saco vitelino, delimitada por endodermo visceral y endodermo
parietal, que rodea a todo el huevo cilíndrico.
El saco vitelino es una importante membrana extraembrionaria que se forma en TS10. Está
localizada en el límite lateral de la cavidad exocelómica (pero no en el límite proximal ni en el
distal). Es exclusivamente de origen extraembrionario y bilaminada, formada por endodermo
extraembrionario visceral que forma el límite externo de la parte media del concepto y por
mesodermo extraembrionario que delinea el margen lateral de la cavidad exocelómica.

Introducción: desarrollo embrionario, fetal y placentario murino
32
�
El corion es una estructura bilaminar, formada completamente por la aposición y fusión de
componentes extraembrionarios, mesodermo y ectodermo extraembrionario del pliegue
amniótico posterior y anterior. Una vez que se fusionan estas láminas, quedan delimitadas la
cavidad ectoplacentaria y la exocelómica.
El amnios, por su parte, si bien también es una estructura bilaminar, está constituida por un
componente mesodérmico extraembrionario y un componente ectodérmico que se continúa con
el ectodermo embrionario.
A medida que avanza el desarrollo las cavidades amniótica y exocelómica se expanden
comprimiendo a la cavidad ectoplacentaria, la cual disminuirá de tal manera que las paredes de la
misma se contactaran y se fusionaran formando la placa coriónica, desapareciendo totalmente el
lumen de la cavidad ectoplacentaria (TS12-13/E8-8,5).
Durante el estadio TS10b-11 (E7,5) (Figura 2.4), el alantoides, enteramente formado por
mesodermo extraembrionario, crece dentro de la cavidad exocelómica y más adelante en el
desarrollo, se fusionará con la placa coriónica (TS12-13/E8-8,5) permitiendo que su vasculatura se
amalgame para dar lugar a la formación de la placenta corioalantoidea.
En cuanto al origen del amnios, Pereira y colaboradores (2011), plantean un origen distinto al
descripto por Kaufman y Bard (1999). Dichos autores descartan la existencia de un pliegue
amniótico anterior y atribuyen la formación de las tres cavidades a la aparición de un pliegue
amniocoriónico, llamado así porque daría lugar tanto al corion como al amnios (que coincidiría
histológicamente con el pliegue posterior descripto por Kaufman). El ectodermo de este único
pliegue amniocoriónico tendría un componente derivado del epiblasto que formará parte de
ectodermo del amnios, y un componente derivado del ectodermo extraembrioanrio que formará
el ectodermo del corion. Siendo en ambos casos, tanto para el corion como para el amnios, el
mesodermo que los constituye de origen extraembrionario (Pereira y col., 2011).
Al final del estadio E7,5 (TS11), el embrión que antes de la gastrulación era bilaminar, ya ha
completado la diferenciación de las tres capas germinales con lo cual se ha convertido en
trilaminar. Esto marca el fin de la etapa de gastrulación, final que se superpone con el inicio de la
organogénesis, la cual se manifiesta por la aparición de los pliegues neurales en la zona cefálica
(Kaufman y Bard, 1999).

Introducción: desarrollo embrionario, fetal y placentario murino
33
�

Introducción: desarrollo embrionario, fetal y placentario murino
34
�
Figura 2.4. Representación esquemática de los principales eventos del desarrollo de
postimplantación temprana. Luego de la implantación del blastocisto ocurren cambios en el embrión,
que por su aspecto, éste se denomina huevo cilíndrico. En este estadio el embrión es bilaminado, con
un core ectodérmico rodeado en todas las regiones, exceptuando el cono ectoplacentario (CE), por
endodermo (TS8). El hipoblasto o endodermo primitivo del blastocisto implantativo da lugar a un tejido
extraembrionario que no debe confundirse con el endodermo definitivo. En el huevo cilíndrico, el
hipoblasto da origen al endodermo extraembrionario parietal (EndoEx parietal), al endodermo visceral
extraembrionario (EndoEx visceral) y al endodermo visceral embrionario (EndoEmb visceral), que si
bien es extraembrionario se lo llama embrionario porque que recubre el futuro embrión (el epiblasto).
El trofoblasto polar (TbP) da lugar al ectodermo extraembrionario (EctoEx). En el estadio TS9a, puede
distinguirse mesodermo, indicando el inicio de la gastrulación. El embrión pasará secuencialmente del
estadio de huevo cilíndrico al de línea primitiva. La línea primitiva (LP) en la unión embrionaria-
extraembrionaria define el lado posterior del futuro feto y se alarga anteroposteriomente. La cavidad
del saco vitelino (SV) es la primera en surgir, entre el EndoEx parietal que tapiza la ex cavidad
blastocélica (Bc) y el endodermo visceral que rodea al concepto. Luego surge el canal proamniótico
(Proam), que formará la cavidad amniótica (CA). Del surgimiento y fusión de los pliegues amnióticos
posterior (PAP) y anterior, o de un único pliegue amniocoriónico (PAC), quedan delimitadas las tres
cavidades, de proximal a distal: cavidad ectoplacentaria (CEP), exocelómica (CEx) y CA, y se conforman
las membranas coriónica, amniótica y del saco vitelino (TS10b). Modificado de Downs, 2011.
2.4. Desarrollo a término del feto y la placenta.
2.4.1. Desarrollo a término de la placenta.
Con el avance de la gestación, la decidua y trofoblasto van sufriendo modificaciones para
finalmente constituir la placenta, un órgano vital e híbrido, con una porción materna y otra fetal.
La placenta, así constituida proporcionará funciones altamente especializadas durante la
gestación. Dentro de estas funciones placentarias críticas para el desarrollo normal del
embrión/feto se destacan el anclaje del concepto, la prevención del rechazo del feto por sistema
inmunológico materno, la protección, la producción de hormonas y de moléculas señalizadoras
entre la madre y el feto, así como el transporte de nutrientes, desechos y gases entre ambos
organismos. La placenta murina, como la humana, es de tipo hemocorial, por lo tanto, la sangre
materna está en contacto directo con las células trofoblásticas. Estructuralmente, la placenta
madura está constituida por la cara fetal, formada por dos compartimentos, el laberinto y la zona
de unión, y por la cara materna, distal al feto, formada por la decidua (Coan y col., 2006).
Precozmente en el desarrollo, desde la región trofoectodérmica polar adyacente al MCI, emerge
un grupo de células que mantiene su capacidad proliferativa, que originará, por un lado el

Introducción: desarrollo embrionario, fetal y placentario murino
35
�
ectodermo extraembrionario, y por otro, formará un aglomerado celular que se proyecta hacia la
región mesometrial del sitio de implantación, estableciendo el cono ectoplacentario (CE). Esta
estructura se mantiene completamente revestida por las células CGTs 2rias
(Copp, 1979), que
aunque se diferencien más tardíamente en relación a las CGTs 1rias
, presentan las mismas
características morfológicas y propiedades funcionales en lo que respecta a la invasión y
fagocitosis. Por lo tanto, como se describió anteriormente, las CGTs 2rias
también están
involucradas en la implantación del embrión, pero en la región mesometrial.
A medida que avanza la gestación, aproximadamente, luego del noveno día de gestación, el
estroma endometrial mesometrial ya ha sufrido profundas modificaciones, mostrándose
completamente decidualizado y exhibiendo una rica y peculiar disposición vascular. Los vasos
mesometriales se disponen de manera radial en relación al embrión. Esta disposición,
característica de la placenta coriovitelina, permite que la sangre extravasada, así como ocurre en
la región antimesometrial, se filtre entre los espacios celulares, a medida que la diferenciación
placentaria avanza. Este tipo de placenta es fundamental entre la implantación y la mitad de la
gestación, y está compuesta por un solo tipo de células del trofoblasto diferenciadas, las CGTs
(Soares y col., 1996).
Desde mediados de la gestación en adelante, la placenta corioalantoidea asume importantes
responsabilidades en el desarrollo del feto (Soares y col., 1996). En tal sentido, la cara fetal de la
placenta inicia su desarrollo y se torna gradualmente más compleja entre los días 8,5 y 14,5 de
gestación. Comienza su formación cuando el alantoides, porción vascular de la placenta derivado
del mesodermo extraembrionario, que se extiende desde el extremo posterior del embrión (E7,5),
alcanza la zona más basal del CE, la placa coriónica (formada por ectodermo extraembrionario),
uniéndose en un proceso llamado fusión corioalantoidea (~E8) (Müntener y Hsu, 1977). Poco
después de la fusión, el corion comienza a plegarse para formar ramificaciones, uniformemente
espaciadas a través de la superficie coriónica, creándose así un espacio dentro de ellas para la
formación de vasos sanguíneos fetales que crecen a partir del mesénquima alantoideo e invaden
profundamente el interior del CE entre las células trofoblásticas preexistentes, proceso llamado
invasión vascular del corion (~E8,5-10,5). En ese momento, el trofoblasto comienza a organizarse
en capas alrededor del mesénquima (Cross, 2000). Entre las células trofoblásticas circula sangre
materna mientras que en el interior del mesénquima del alantoides se forman los vasos fetales.
Durante la morfogénesis de esta región de la placenta, las ramas de mesénquima se alargan y
ramifican progresivamente, y distintos tipos celulares comienzan a diferenciarse. La continua

Introducción: desarrollo embrionario, fetal y placentario murino
36
�
ramificación, en última instancia, produce una compleja estructura vellosa que proporciona la gran
superficie que se requiere para el intercambio de nutrientes que sustenta el crecimiento
embrionario y fetal pasado el E10,5 en el ratón (Simmons y col., 2008). En esa instancia, esta
región pasa a denominarse zona laberíntica, llamada así por la intrincada red de espacios de
sangre materna y vasos fetales, que posibilitan los intercambios metabólicos entre ambos
organismos (Müntener y Hsu, 1977; Bevilacqua y Abrahamsohn, 1988; Bevilacqua, Faria e
Abrahamsohn, 1991). Las células trofoblásticas coriónicas que forman parte del laberinto se
diferencian en dos tipos: las células sinciciotrofoblásticas multinucleadas, formadas por la fusión
de células trofoblásticas, y las células trofoblásticas de tipo mononuclear o citotrofoblásticas, un
subtipo de células gigantes del trofoblasto, llamadas también células trofoblásticas sinusoidales. A
su vez, las células trofoblásticas diferenciadas, en los roedores, no se disponen al azar en el
laberinto, sino que se organizan de una manera muy particular. Tres láminas o capas de células
trofoblásticas separan los espacios de sangre materna y fetal, por eso este tipo de placenta recibe
el nombre de hemotricorial. El trofoblasto trilaminar incluye una sola capa de células sinusoidales,
que mantienen contacto directo con la sangre materna extravasada en los espacios o senos
maternos (Coan y col., 2005; Simmons y Cross, 2005; Simmons y col., 2007) y cuya función es
principalmente endocrina; y dos capas de células sinciciotrofoblásticas, fuertemente adheridas
una a la otra, estando la última en contacto directo con la membrana basal que la separa del
endotelio de los capilares fetales (Davies y Glasser, 1968; Peel y Bulmer, 1977; Simmons y Cross,
2005; Watson y Cross, 2005), formando una estructura muy delgada involucradas en transporte
bidireccional materno-fetal de gases, nutrientes y desechos (Soares y col., 1996). Así, el laberinto,
constituye la mayor parte de la placenta madura, donde la sangre materna (único elemento
materno presente en el laberinto) y fetal fluyen en dirección contracorriente, para así maximizar
los intercambios entre ambos organismos (Adamson y col., 2002). Si el laberinto no está
apropiadamente vascularizado, con una adecuada ramificación, la perfusión placentaria se
deteriora, resultando esta situación en una mala oxigenación y difusión de nutrientes (Pardi y col.,
2002).
La región de la placenta fetal situada entre el laberinto y la decidua, recibe el nombre de zona de
unión. Esta región, normalmente, presenta un aspecto reticulado, con cordones espesos de células
epiteloides que delimitan espacios vasculares amplios e irregulares. Estos espacios contienen
sangre materna extravasada. Allí es posible distinguir, principalmente, tres poblaciones celulares,
las CGTs, las células espongiotrofoblástica y las células glicogénicas.

Introducción: desarrollo embrionario, fetal y placentario murino
37
�
Las CGTs, de grandes dimensiones (poliploides) y con varios nucléolos (Ilgren, 1983), en la
gestación a término se mantienen en contacto directo con la decidua. También, se las encuentra
revistiendo, de modo endoteliforme, la luz de los espacios sanguíneos maternos. En tal sentido, en
ocasiones, es posible percibir en las áreas más próximas al endometrio, que los espacios
sanguíneos están parcialmente revestidos por células endoteliales típicas (en la cara limitante con
la decidua) y parte revestidos por trofoblasto (en la cara hacia la parte fetal). Estas células pueden
presentar características invasivas, pero a su vez, poseen importantes actividades endocrinas
(Soares y col., 1996). También se les atribuye una posible función de defensa actuando como
barrera placentaria. Si bien, luego de la formación de la placenta, estas células gradualmente
pierden las características de células fagocíticas, se cree que esta actividad puede ser reactivada
en situaciones de desafío, por ejemplo, si el organismo materno fuera invadido por patógenos
diversos.
Las células espongiotrofoblásticas (EPG), con características de células indiferenciadas,
principalmente basófilas y con grandes núcleos, se agrupan en la zona de unión formando grupos
o clusters de células, así como, cordones epiteloides. Estás células poseen principalmente
funciones endócrinas relacionadas con la síntesis y secreción de hormonas, como la hormona
lactógeno placentaria, prolactina, hormona de crecimiento, siendo también productoras de
citocinas (Ain y col., 2003; Malassiné y col., 2003; Soares, 2004). Ellas ya se encuentran presentes
en la placenta murina en el E12,5, las cuales proliferan notablemente en el E16,5, siendo
responsables en parte del aumento de tamaño de la zona de unión, que alcanza su máximo en
este estadio, lo cual tiene relación directa con el requerimiento hormonal y de otros factores para
el éxito y continuidad del desarrollo (Coan y col., 2006).
En la placenta madura se encuentra un tipo de células que se caracteriza por presentar citoplasma
con un alto número de vesículas, que aparecen ópticamente vacías luego de las técnicas de
coloración convencional, pero que coloreadas por la técnica de PAS (ácido peryódico de Schiff),
resultan fuertemente PAS-positivas, con gránulos color fucsia, indicando que sus vesículas
corresponden a depósitos de glicógeno. Estas células reciben entonces el nombre de células
glicogénicas (CG), por su capacidad de acumular glicógeno, otorgando a la zona de unión la
característica distintiva de ser rica en material PAS-positivo. La función principal de estas células
placentarias es la de ser un reservorio potencial de energía (Davies y Glasser, 1968). Las CGs
surgirían en la zona de unión, posteriormente, a las células EPG (Redline y col., 1993; Adamson y
col., 2002), y si bien se pensó que derivaban del linaje de células EPG, por expresar genes

Introducción: desarrollo embrionario, fetal y placentario murino
38
�
específico expresados en dichas células, actualmente se cree que derivan de un linaje diferente
(Coan y col., 2006). En el estadio E12,5 pocos y pequeños clusters de células PAS-positivas son
observados en la zona de unión, siendo en esta etapa morfológicamente similares a las células EPG
PAS-negativas, excepto por presentar un tamaño reducido respecto de las EPG. Posteriormente,
entre E13 y E14,5, tiene lugar un significativo evento proliferativo de CGs. Ya en el E13,5, las CGs
son claramente identificadas, con su citoplasma vacuolado y su inconfundible marca PAS-positiva.
En E16,5 gran parte de las CGs invaden la decidua, y una vez que lo hacen, dejan de verse en
cluster, siendo frecuentemente observadas de manera aislada, mientras que ya en el E18,5, se ve
una gran reducción en el número de CGs tanto en la zona de unión como en la decidua.�Siendo la
glucosa la fuente principal de energía para la placenta, el glucógeno almacenado en dichas células
podría ser utilizado en la etapa final de la gestación (E18,5) (Barash y Shafrir, 1990). Las reservas
de glucógeno estarían destinadas al crecimiento del feto al final de la gestación, debido a la
notable demanda de éste de los recursos maternos para su desarrollo final. El glucógeno de
reserva, en este momento, puede ser convertido en glucosa, ingresar en la circulación materna y
eventualmente pasar al feto a través de su circulación en el laberinto (Barash y Shafrir, 1990).
La porción materna de la interface materno-fetal también sufre importantes modificaciones a lo
largo de gestación. A medida que la porción fetal de la placenta se diferencia, la decidua
mesometrial inicia un proceso de involución. Alrededor del día 13 de gestación esta estructura ya
se encuentra reducida y es casi vestigial a los 17 días de gestación (Katz y Abrahamsohn, 1987). El
tejido endometrial más distal en relación al embrión abriga células inmunológicamente
competentes entre las cuales se destacan las células natural killer uterinas. Estas células tienen
una función primordial en la gestación, dado que limitan la invasión de las células trofoblásticas
(Kiso y col., 1992) por medio de su actividad citotóxica (Manaseki y Searle, 1989) y producción de
factores tales como el IFN-γ (del inglés, Interferon-gamma) (Marzusch y col., 1997) y a su vez
contribuyen al normal desarrollo vascular de la decidua (Croy y col., 1997a).
En la Figura 2.5, se describen los eventos que llevan a la formación de la placenta madura, y en la
Tabla 2.2 se presenta un resumen de los tipos trofoblásticos presentes en la placenta definitiva,
con su localización y función.

Introducción: desarrollo embrionario, fetal y placentario murino
39
�
Figura 2.5. Representación esquemática de los eventos principales involucrados en el desarrollo de la
placenta hemotricorial murina. A, el linaje celular que será parte de la placenta surge en el blastocisto.
En E7,5, surge el brote alantoideo, que aportará el mesodermo extraembrionario. B, detalle del avance
del desarrollo placentario de la zona recuadrada en A. En E8, el alantoides contacta con la placa
coriónica. En E8,5, el corion comienza a plegarse como consecuencia de la invasión vascular. En E10,5,
la ramificación del mesénquima alantoideo contribuye a la formación de vellosidades uniformemente
distribuidas dentro de la placa coriónica. Los vasos fetales se diferencian en el interior del mesénquima.
La morfogénesis de las vellosidades lleva a la formación del laberinto; C, en E14,5 se distinguen los tres
compartimentos diferenciados de la placenta madura: la decidua (De), distal al feto; el compartimento
intermedio, la zona de unión con células gigantes trofoblásticas (CGTs), espongiotrofoblásticas (EPG) y
glicogénicas (CGs); y la zona más proximal al feto, el laberinto, red de espacios de sangre materna y
vasos fetales. En detalle se muestra la disposición de las células: sinusoides maternos (m, *: glóbulo
rojo materno) limitados por trofoblasto mononuclear (ct, capa I), los cuales contactan con el
sinciciotrofoblasto (st, capa II y III) quien rodea a la vasculatura fetal (cf: capilar fetal, ecf: endotelio de
capilar, **: glóbulo rojo fetal). D, se muestran las proporciones de los tres compartimentos en E18,5 y
los tipos trofoblásticos presentes. CU: cordón umbilical. Modificado de Watson y Cross, 2005; Soares
Lab Home KU Medical Center-The University of Kansas.

Introducción: desarrollo embrionario, fetal y placentario murino
40
�
Tabla 2.2. Tipos de células trofoblásticas en la cara fetal de la placenta corioalantoidea murina.
Tipo celular Localización Función
Células gigantes trofoblásticas
Zona de unión Endócrina, invasiva
Zona laberíntica Endócrina, invasiva
Células espongiotrofoblásticas Zona de unión Endócrina
Células glicogénicas Zona de unión Reservorio de energía
Células sinciciotrofoblásticas Zona laberíntica Barrera de transporte
Resumen de los tipos de células trofoblásticas presentes en la cara fetal de la placenta madura, se
indica su localización en la placenta y su función.
2.4.2. Desarrollo a término del feto.
La gastrulación es la etapa del desarrollo embrionario en que se forma el esquema corporal. Las
células del blastocisto se reorganizan espacialmente para formar tres capas de células,
endodermo, ectodermo y mesodermo embrionario. Cada capa germinal formada en este estadio,
se diferencia y crece para formar finalmente los diferentes órganos y sistemas del feto (Figura 2.6).
Este proceso de desarrollo y formación de órganos ocurre durante la etapa llamada
organogénesis, la cual en el ratón, se extiende desde el final de estadio E7,5 (TS11) hasta el E14
(TS22). El período que le sigue y que transcurre entre el E14 hasta el final de la vida intrauterina
E18,5-19 (TS26), es el período fetal, fase en la que los tejidos y órganos, formados en la etapa
anterior, maduran y tienen un rápido y significativo crecimiento. El incremento en el tamaño
corporal también ocurre principalmente en esta etapa. Sin duda, este crecimiento fetal
intrauterino implica un coordinado aumento en la masa de una amplia variedad de diferentes
tejidos y órganos, por lo que está cuidadosamente regulado, es decir, debe existir una relación
temporal muy estricta entre la aparición de un tejido y su diferenciación.
La etapa fetal es también una fase crítica del desarrollo, dado que los sistemas del cuerpo que
sustentarán la vida del nuevo individuo, comienzan a prepararse para el desafío de la vida fuera
del útero. En este sentido, la evolución del desarrollo fetal a término puede ser monitoreado por
análisis de su crecimiento. Generalmente, el crecimiento se mide en términos de cambios en el

Introducción: desarrollo embrionario, fetal y placentario murino
41
�
peso o tamaño corporal, siempre en relación a la edad gestacional, y si bien, depende de muchos
factores, como por ejemplo, factores genéticos y/o ambientales, el estado nutricional materno es
clave para el crecimiento normal del feto. En modelos animales, la evaluación del crecimiento fetal
básicamente se realiza a través de la medición del peso y largo corporal total, aunque otros
estudios como la evaluación del desarrollo de ciertos sistemas y órganos internos, completa el
análisis del crecimiento fetal (Fowden, 2001).
En la Figura 2.7, se muestra un resumen del desarrollo embrionario / fetal murino con sus etapas
morfogenéticas más importantes.
�
Figura 2.6. Diagrama que ilustra los tejidos derivados de las tres capas germinales. Se muestran los
principales tejidos y órganos derivados del ectodermo, mesodermo y endodermo embrionario
formados durante la gastrulación. El mapa de diferenciación de tejidos y órganos muestra, en parte, la
gran complejidad que existe en el desarrollo de los mamíferos. Modificado de Life Map Science, 2012.
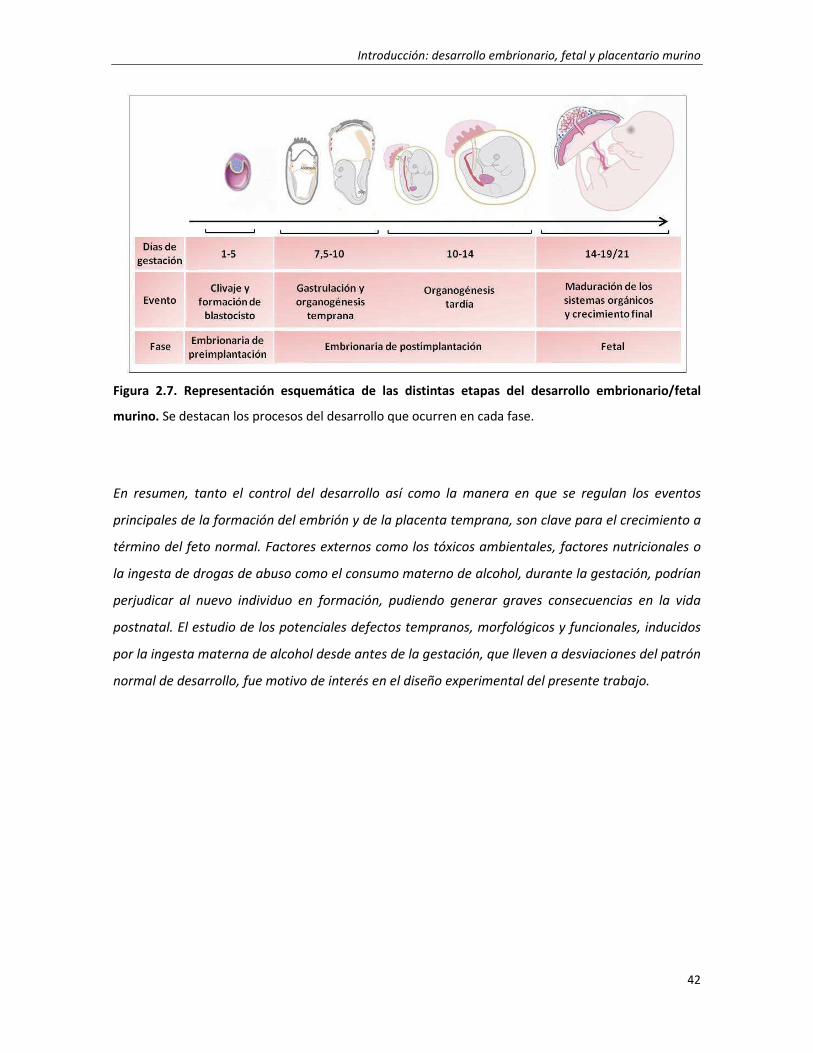
Introducción: desarrollo embrionario, fetal y placentario murino
42
�
Figura 2.7. Representación esquemática de las distintas etapas del desarrollo embrionario/fetal
murino. Se destacan los procesos del desarrollo que ocurren en cada fase.
En resumen, tanto el control del desarrollo así como la manera en que se regulan los eventos
principales de la formación del embrión y de la placenta temprana, son clave para el crecimiento a
término del feto normal. Factores externos como los tóxicos ambientales, factores nutricionales o
la ingesta de drogas de abuso como el consumo materno de alcohol, durante la gestación, podrían
perjudicar al nuevo individuo en formación, pudiendo generar graves consecuencias en la vida
postnatal. El estudio de los potenciales defectos tempranos, morfológicos y funcionales, inducidos
por la ingesta materna de alcohol desde antes de la gestación, que lleven a desviaciones del patrón
normal de desarrollo, fue motivo de interés en el diseño experimental del presente trabajo.

Introducción: uniones intercelulares
44
�
3. Uniones intercelulares en el desarrollo embrio-trofoblástico: papel de las uniones
estrechas y adherentes.
3.1. Uniones intercelulares.
La uniones celulares son regiones especializadas de la membrana plasmática en las que se
concentran proteínas transmembrana especiales, mediante las cuales se establecen conexiones
entre dos células contiguas o entre una célula y la matriz extracelular adyacente. En la Tabla 3.1,
se resumen los tipos de uniones celulares según tipo de unión, proteínas transmembrana que
participan, vínculo con el citoesqueleto y su función. En la Figura 3.1 se muestra un esquema de
las uniones, así como su localización celular.
Tabla 3.1. Clasificación de las uniones celulares.
Unión Tipo de unión Proteínas de
membrana
Vínculo con el
citoesqueleto Función
De oclusión Estrecha
Célula-célula
Ocludina
Claudina
JAM
Microfilamentos
de actina
Sellan el espacio intercelular
hacia la luz (superficie luminal
de células epiteliales vecinas),
evitando el pasaje de
sustancias por ese espacio
De anclaje
Adherente
Célula-célula
Complejo
Cadherina/
Catenina
Microfilamentos
de actina Si bien la función varía de
acuerdo al tipo de unión,
estas uniones mantienen la
ubicación entre células
vecinas, y entre células con la
MEC, a través de uniones al
citoesqueleto
Focal
Célula-MEC Integrina
Microfilamentos
de actina
Desmosomas
Célula-célula Cadherina
Filamentos
intermedios
Hemidesmosomas
Célula-MEC Integrina
Filamentos
intermedios
Comunicantes Nexo
Célula-célula Conexinas ---
Permiten el pasaje de
pequeñas sustancias entre
células contiguas
Moléculas de adhesión intercelulares, sus principales componentes y funciones. JAM: molécula
adhesiva de la unión (del inglés, Junctional Adhesion Molecule), MEC: matriz extracelular.

Introducción: uniones intercelulares
45
�
Figura 3.1. Representación esquemática de la distribución
de las uniones celulares. En la zona más apical de la
membrana apicolateral de las células, se encuentran las
uniones estrechas formando una barrera impermeable que
impide la difusión de proteínas y lípidos de membrana. En
ubicación más basal, se encuentran las uniones adherentes,
compuestas por proteínas transmembrana en relación con el
citoesqueleto, manteniendo a las células estrechamente
unidas. Los desmosomas, con la placa proteica interactuando
con filamentos intermedios, dan gran resistencia mecánica a
los tejidos. Las uniones nexo compuestas por conexinas,
proteínas integrales de membrana dispuestas circularmente,
forman poros transmembranosos entre las células. En el
dominio basal y en contacto con la matriz extracelular se
encuentran los hemidesmosomas y los contactos focales, los
que anclan filamentos intermedios y microfilamentos de
actina, respectivamente, a la membrana basal. SA: superficie
apical, SB: superficie basal, SL: superficie lateral.
En el proceso de diferenciación epitelial, las células adquieren contactos celulares polarizados,
apicolaterales y basolaterales con la consecuente formación de complejos de uniones
apicolaterales (CUA), que involucran uniones estrechas y, más basalmente, uniones adherentes.
Estas uniones intercelulares contribuyen a la adhesión e integridad epitelial necesaria para el
transporte transepitelial. En particular las uniones estrechas contribuyen al mantenimiento de la
polaridad de la membrana celular y regulan la permeabilidad paracelular. Ambos tipos de uniones,
a su vez, funcionan como mediadores en la señalización intracelular, controlando aspectos como
la proliferación celular y la expresión génica (Alexander y col., 1996). Por el importante papel en la
diferenciación epitelial durante el desarrollo temprano del embrión, particularmente en el
presente trabajo estudiamos las uniones oclusivas o estrechas (UE) y las uniones adherentes (UA)
en diferentes etapas de la diferenciación embrionaria murina, con el fin de evaluar el impacto de
la ingesta materna de alcohol en dichas uniones y su consecuencia en la formación de estructuras
embrionarias.

Introducción: uniones intercelulares
46
�
3.1.1. Uniones estrechas.
Como se mencionó, las uniones oclusivas (zonula occludens) o UE están compuestas por complejos
multiproteicos que contribuyen a la adhesión celular. Proteínas transmembrana, tales como
ocludinas, proteínas JAM, y claudinas, interactúan directamente con un grupo de proteínas
andamio que pertenecen a la familia MAGUK (del inglés, Membrane-Associated Guanylate Kinase
homologs), colectivamente conocidas con el nombre de proteínas zonula occludens o ZO
(proteínas ZO-1, ZO-2, y ZO-3; donde el número revela el orden en que fueron descubiertas)
(Stevenson y col., 1986; Gumbiner y col., 1991). Las proteínas ZO se caracterizan por presentar
dominios de unión a otras proteínas de las uniones estrechas. En tal sentido, se observó que ZO-1
o ZO-2 son requeridas para el agrupamiento de claudinas y para que las UE funcionen
eficientemente como barrera. También presentan dominios de unión a proteínas de señalización
intracelular y a actina, por lo que están anatómica y funcionalmente asociadas al citoesqueleto
(González-Mariscal y col., 2008; Paris y col., 2008). Las proteínas que forman el complejo de UE se
disponen en cadena a lo largo de la línea de unión para formar una barrera en la zona más apical
de las células. De esta manera, este tipo de unión impide la libre difusión de moléculas
hidrosolubles, regulando la barrera de permeabilidad paracelular para iones y moléculas de
acuerdo a su tamaño y carga. A su vez, mantiene la polaridad de la membrana celular de los
epitelios impidiendo que los lípidos y proteínas de membrana difundan libremente de un dominio
celular a otro. De esta manera, establecen regiones funcionales especializadas en la membrana
plasmática. Es decir, contribuyen a la división del plasmalema en un dominio apical y otro
basolateral con distinta composición de proteínas de membrana (las cuales son incapaces de
atravesar la unión oclusiva por difusión lateral). Las UE participan también en la estabilización de
los epitelios, dado que en dichos puntos de unión, las células se encuentran fuertemente unidas
(Figura 3.2. A y B).
Proteínas ZO: para todas las proteínas ZO se ha postulado una función reguladora durante la
formación de la unión zonula occludens. Diversos agentes patógenos y químicos actúan sobre
estas proteínas permeabilizando, en consecuencia, la unión. ZO-1 es una proteína de 220 kDa y
forma parte de las UE de las células epiteliales y endoteliales. En las células epiteliales, ZO-1 se
encuentra asociada a las UA cuando la UE aún no se ha establecido, como ocurre en las primeras
etapas del desarrollo embrionario (antes de la compactación de la mórula de 8 células) (Fleming y

Introducción: uniones intercelulares
47
�
col., 1989). Se han detectado tres dominios de procesamiento alternativo en ZO-1 (α, β y γ), donde
el dominio α es una región de 80 kDa y está presente sólo en las UE y no en las adherentes. La ZO-
1 desempeña un papel fundamental en la fisiología de las UE. Así, se ha visto que agentes que
modulan la expresión de ZO-1 (por ejemplo, los glucocorticoides) alteran, en consecuencia, la
funcionalidad de las UE (Underwood y col., 1999). La baja expresión de ZO-1 en los epitelios se
relaciona con la progresión tumoral (Hoover y col., 1998), sugiriendo que las uniones estrechas y/o
sus componentes desempeñarían un papel en la regulación del crecimiento, proliferación y/o
migración celular. ZO-1 contiene en su secuencia dos señales de direccionamiento nuclear. En
relación a esto, se la ha visto en el núcleo de monocapas epiteliales subconfluentes o en células
con contactos célula-célula inmaduros. Esta localización nuclear sugiere nuevos papeles para esta
proteína, y la observación de su unión a un factor de transcripción que regula la progresión de
ciclo celular, refuerza esta idea (Balda y Matter, 2000). ZO-2 es una proteína de 160 kDa que está
presente también en las UE de células epiteliales y endoteliales. Contiene en su secuencia, señales
de importación y exportación nuclear, y es capaz de moverse entre el núcleo y las UE de acuerdo
al estado de confluencia del epitelio (Islas y col., 2002). De la misma manera, ZO-3 (de 130 KDa) en
su secuencia posee dos señales de localización nuclear (González-Mariscal y col., 2008).
Utilizando diversas técnicas se logró determinar que el ensamble de las UE es precedido por la
formación de UA (Gumbiner, 1987; Gumbier y col., 1988).
3.1.2. Uniones adherentes.
Las uniones de anclaje, en particular las UA (zonula adherens), son uniones célula-célula que se
extienden a lo largo de todo el perímetro celular. Se encuentran en posición basal respecto de las
UE y al igual que ellas, están formadas por complejos multiproteicos, en los que participa una
proteína transmembrana, la cadherina. Las cadherinas de células vecinas se unen entre sí por su
dominio extracelular (unión homofílica y dependiente de iones calcio), y por su extremo citosólico,
se unen a los microfilamentos de actina a través de proteínas fijadoras que están asociadas
formando placas de anclaje (cateninas, vinculinas, α-actininas). Las cadherinas median así la
verdadera unión entre células contiguas, y a través de la placa de anclaje, median la fijación al
citoesqueleto. Por lo tanto, las uniones de anclaje, proveen estabilidad mecánica a las células
epiteliales mediante la vinculación del citoesqueleto de una célula al citoesqueleto de la célula
vecina, creando la unidad estructural del epitelio manteniendo a las células estrechamente unidas

Introducción: uniones intercelulares
48
�
(Figura 3.2. C y D). Dado que si bien, en la zonula occludens hay una unión de membranas celulares
contiguas, su resistencia ante un estrés mecánico es limitada. El refuerzo de esta región depende
de un sitio de unión fuerte (UA) en posición basal a la UE. A su vez, mediante la capacidad en la
transducción de señales mediada por el complejo cadherina/catenina, las UA también cumplen
funciones importantes en el reconocimiento célula-célula, en la morfogénesis, diferenciación,
migración, proliferación, y supervivencia de las células epiteliales (Ross, 2007).
Cadherinas: Las cadherinas constituyen una superfamilia de glicoproteínas, donde la E-, N- y P-
cadherina son las que, habitualmente, se encuentran en las UA y regulan la adquisición del
fenotipo epitelial. Derivan el nombre de la inicial del tejido en que fueron descriptas (epitelial,
neural y placentario, respectivamente) (Ross, 2007). Las cadherinas, para funcionar
adecuadamente, necesitan estar completas, ya que al expresar experimentalmente cadherinas
que sólo codifican los dominios extracelular y transmembrana, o sólo el citosólico, las células no se
adhieren y en consecuencia se generan serias anomalías en el desarrollo (Levine y col., 1994). La
adhesión intercelular, por lo tanto, es un fenómeno complejo en el que además del contacto
homotípico entre las cadherinas de dos células vecinas se necesita el anclaje de estas moléculas al
citoesqueleto, requiriéndose también de la participación de las cateninas.

Introducción: uniones intercelulares
49
�
Figura 3.2. Representación esquemática de la organización molecular de los complejos de unión
intercelulares. A y B, complejo de unión estrecha, con sus principales componentes, donde las
proteínas transmembrana (ocludina, Claudina y JAM) interactúan con el citoesqueleto de actina a
través ZO-1 y ZO-3; C y D, complejo de unión adherente con sus principales componentes, donde los
filamentos de actina de células contiguas están vinculados con el complejo E-cadherina/catenina a
través de α-actinina y vinculina. El complejo E-cadherina/catenina interactúa con moléculas idénticas
incluidas en la membrana plasmática de la célula vecina, mediante una unión dependiente de calcio
(Ca2+
). JAM: molécula adhesiva de la unión. Modificados de Ross, 2007.
3.2. Uniones intercelulares en el desarrollo embrionario.
3.2.1. Uniones estrechas y adherentes en el desarrollo de embriones de preimplantación.
En mamíferos, el embrión inicialmente es una estructura indiferenciada. El primer tipo celular
diferenciado que surge durante la embriogénesis es un epitelio, el trofoectodermo (TE), el cual se
forma en la parte más externa del embrión. Este epitelio está involucrado en el transporte
polarizado para generar la cavidad blastocélica contribuyendo así a la formación del blastocisto y
proporcionando de esta manera un espacio que permitirá reordenamientos celulares y
morfogénesis, que darán posteriormente forma al embrión en desarrollo (Eckert y Fleming, 2008).

Introducción: uniones intercelulares
50
�
Este epitelio trofoectodérmico comienza a formarse en el estadio de 8 células y completa el
proceso en la fase de 32 células, abarcando un período de aproximadamente 24 hs. Durante ese
período, ocurren divisiones celulares asimétricas, donde las células que quedan en el interior no
presentan características epiteliales y forman el macizo celular interno (MCI). El proceso de
diferenciación del TE se caracteriza por el ensamblaje secuencial de proteínas en los complejos de
uniones apicolaterales durante los estadios de 8, 16 y 32 células. Este proceso de ensamblaje
culmina con la segregación de los CUA en dos dominios diferenciados: las UE y las UA (Sheth y col.,
2008). La E-cadherina es una de las moléculas clave en todo este proceso, y ya se encuentra
presente en el oocito fecundado como ARNm y proteína (Kemler y col., 1977; Ohsugi y col., 1997).
Esta molécula de adhesión es vital para el proceso de compactación en la fase de mórula y la
subsiguiente formación del TE, por lo tanto, esta molécula de adhesión juega un papel crucial en la
diferenciación temprana trofoectodérmica. En tal sentido, en la etapa de 8 células se encuentra
uniformemente distribuida en las regiones de contacto célula-célula, lo que ocasiona que las
blastómeras se aplanen y adhieran fuertemente entre sí y formen una mórula compacta (Kan y
col., 2007).
Una vez que el embrión de 8 células sufrió el proceso de compactación, las proteínas
constituyentes de las UE se reúnen en la zona de contacto apicolateral de las blastómeras. Este
proceso de montaje de las UE se produce secuencialmente, durante el período de 24 hs, entre la
compactación y la formación de la cavidad blastocélica (Eckert y Fleming, 2008). Inmediatamente
después de la compactación, la isoforma α- de la ZO-1 se ensambla en la membrana periférica en
los contactos intercelulares de las blastómeras ricas en cadherinas (Fleming y col., 1989; Sheth y
col., 1997; Sheth y col., 2000). La isoforma α+ a diferencia de la α−
, aparece más tarde, cuando se
forma el blastocele y se establecen las UE (Sheth y col., 1997). Durante la etapa de 16 células, las
proteínas de membrana cingulina y ZO-2, se reúnen en el sitio de contacto apicolateral por
primera vez, co-localizando con E-cadherina en un complejo transitorio que comprende UA y UE
(Javed y col., 1993; Sheth y col., 2008). Por lo tanto, estas primeras etapas en la biogénesis de las
UE dependen de la adhesión de E-cadherina. Se ha visto, que embriones que no poseen E-
cadherina logran la compactación gracias a la presencia E-cadherina de origen materno, sin
embargo, son incapaces de formar un TE funcional (Larue y col., 1994; Ohsugi y col., 1997). A su
vez, la inhibición de la E-cadherina provoca el montaje al azar de las UE en la membrana,
resultando en una pérdida completa de la adhesión celular (De Vries et al., 2004), de modo que la

Introducción: uniones intercelulares
51
�
E-cadherina es quien asegura el correcto establecimiento de la UA y en consecuencia de las UE en
el TE en desarrollo.
Por último, durante la etapa de 32 células, ZO-1α+ y las proteínas transmembrana ocludina y
claudina se ensamblan (Fleming y col., 2001). Es sólo después de la fase de montaje final, que el
embrión genera un sello de permeabilidad entre las células del TE, para formar finalmente la
cavidad blastocélica (Sheth y col., 2000).
La formación gradual primero de las UA y luego de las UE en las blastómeras externas de la mórula
determina el establecimiento de la capa de TE y contribuye a la retención de fluidos en el interior
del embrión. Por lo tanto, la alteración o inhibición de alguno de estos componentes, afectará al
desarrollo del blastocisto (Wang y col., 2008). En relación a esto, distintos estudios demostraron
que embriones que carecen de la proteína ZO-2 en las UE del TE, la formación de la cavidad
blastocélica resulta retrasada. Los embriones deficientes en ZO-2, mostraron a su vez, un mayor
ensamblaje de ZO-1 pero no de ocludina, lo que indicaría una redundancia de la proteína ZO-1
como un mecanismo de compensación que contribuye al fenotipo leve observado por ausencia de
ZO-2. En contraste, la deficiencia de ZO-1, conduce a fenotipo más severo que la ausencia de ZO-2,
generando una significativa inhibición en la formación de blastocele. A partir de esos resultados,
se le ha asignado a ZO-1 un papel crítico en la morfogénesis del blastocisto (Sheth y col., 2008). La
expresión de ZO-1α+ en la fase de mórula tardía indica que, probablemente, esté involucrada en
los eventos finales de la transición de mórula a blastocisto (Sheth y col., 1997, 2000). El konckdown
del gen de ZO-1 en embriones de ratón utilizando ARN de interferencia, resultó en el desarrollo de
los mismos sólo hasta la fase de mórula, no prosiguiendo hasta la de blastocisto. Estos resultados
sugieren que ZO-1 sería necesaria para mantener la integridad de la capa de TE, que es esencial
para la formación del compartimento celular y blastocélico que define a los blastocistos (Wang y
col., 2008).
Por lo tanto, la construcción de una capa epitelial requiere en primer lugar de la formación de UA
(basadas en E-cadherina) las cuales ayudan a ensamblar las UE (Gumbiner y col., 1988). Estas dos
redes de unión regulan conjuntamente la fuerza de adhesión entre las células que permite la
formación de epitelios. En ausencia de ZO-1, las uniones basadas en E-cadherina no proporcionan
suficiente sostén a las células externas de la mórula para iniciar la acumulación de líquido en el
interior del embrión (Wang y col., 2008). De la mima manera, en ausencia de E-cadherina, como se
mencionó, las UE no se establecen adecuadamente. Por lo tanto, ambas moléculas son
fundamentales durante la diferenciación epitelial del embrión.

Introducción: uniones intercelulares
52
�
3.2.2. Uniones adherentes en el desarrollo embrionario de postimplantación temprano.
Así como las moléculas de adhesión son fundamentales en los estadios iniciales de la
embriogénesis, también lo son en estadios más avanzados, donde predominan eventos de
diferenciación celular, crecimiento y proliferación. En estas fases, las cadherinas son críticas para
la determinación, el mantenimiento de la diferenciación y la arquitectura de tejidos epiteliales
(Takeichi, 1991). La clásica subfamilia de las UA que actúa en estadios tempranos de la
embriogénesis, comprende las E-, N- y P-cadherinas que interactúan intracelularmente con �-
catenina y con catenina p120 (Gumbiner, 2000). Los cambios en el patrón de expresión espacio-
temporal de E-cadherina durante la embriogénesis tienen un marcado impacto en la mecánica de
adhesión de las células y por lo tanto, en su destino (Wheelock y col., 2008), dado que la expresión
de la E-cadherina está involucrada en la transición epitelio-mesenquimal (TEM) y la adquisición de
un fenotipo apto para la migración (Savagner, 2001). En tal sentido, la pérdida de E-cadherina es
una fuerza motriz para la transformación de las células epiteliales en mesenquimáticas durante la
formación de mesodermo en la gastrulación. El epitelio epiblástico, antes de la formación del
mesodermo, varía en las distintas especies de mamíferos, siendo cilíndrico pseudoestratificado, en
ratón. Estas células epiblásticas están apoyadas en una lámina basal y presentan uniones
intercelulares del tipo comunicantes, desmosomas, UE y UA (Burdsal y col., 1993). En la TEM que
ocurre durante la gastrulación, las células del epiblasto que ingresan por la línea primitiva, pierden
la E-cadherina, y experimentan cambios fundamentales en la forma y organización celular,
pasando de una célula epitelial polarizada a una mesenquimática no polarizada. Por lo tanto, a
medida que las células embrionarias se van diferenciando en varios tipos, la E-cadherina
desaparece de algunas capas de células y permanece en otras. En embriones que se encuentran en
etapas tardías, esencialmente todas las células epiteliales que proliferan derivan del ectodermo y
el endodermo, expresan E-cadherina, de alguna forma quedando restringida únicamente a tejidos
epiteliales en adultos. El tejido neural y mesodérmico no presentan E-cadherina (Viebahn, 1995)
3.3. Uniones adherentes en el embrión y consumo materno de alcohol: antecedentes.
Dada la importancia de las cadherinas en la determinación de la morfogénesis y arquitectura de
los tejidos (Tunggal y col., 2005), recientemente estudiamos la expresión de E- y N- cadherina en
embriones al día 10 de gestación (organogénesis), luego de la ingesta materna de alcohol al 10%

Introducción: uniones intercelulares
53
�
desde antes de la gestación y hasta el día 10 de preñez (Coll y col., 2011). En nuestro modelo CF-1,
la exposición a etanol no afectó la localización o distribución de E- y N-cadherina en los embriones.
A pesar de la similar inmunorreactividad tanto de E- como N-cadherina entre el grupo control y el
tratado, se detectó por immunoblotting un aumento de la expresión de la E- y N-cadherina en
embriones expuestos al etanol. Este incremento podría deberse a un efecto negativo del etanol en
la adhesión celular del embrión expuesto. El potencial daño en las uniones adherentes como
consecuencia de la exposición a etanol podría inducir mecanismos compensatorios de regulación
positiva de la expresión de las cadherinas en los tejidos embrionarios. Dado que la N- y E-
cadherina juegan un papel fundamental en el desarrollo normal de los órganos, participando en
procesos de diferenciación, segregación y migración de las células embrionarias, la regulación
espacio-temporal anormal de la expresión de cadherina puede ser causa de las anomalías de los
embriones organogénicos expuestos a alcohol. Es posible que la desregulación o la sobreexpresión
de E- y N-cadherina jueguen un papel importante en el retraso de crecimiento, la reducción de la
viabilidad y los defectos en el desarrollo del tubo neural después de la exposición a etanol (Coll y
col., 2011).
Por lo expuesto, y dada la importancia de la expresión y ensamblaje de las cadherinas y proteínas
ZO en las UA y UE en el desarrollo de embriones tempranos, abordamos el estudio particular de la
E-cadherina y ZO-1 en blastocistos y embriones en gastrulación, para evaluar los efectos de la
exposición materna a alcohol en dichos períodos.

Introducción: metaloproteasas
54
�
2. Matriz extracelular en el desarrollo embrio-trofoblástico: papel de las
metaloproteasas.
4.1. Matriz extracelular, su estructura y función.
La matriz extracelular (MEC) está compuesta por una gran variedad de macromoléculas, cuya
distribución y organización varía con el tipo de tejido en que están presentes. En el tejido
conectivo, estas moléculas están organizadas en forma de red tridimensional rellenando los
espacios intercelulares, confiriendo cohesión, rigidez y estructura al tejido.
En relación a la composición química, las moléculas de la MEC pueden ser agrupadas en 4 clases:
proteínas colágenas, proteínas del sistema elástico, glicosaminoglicanos (GAGs) y proteoglicanos
(PGs), y otras glicoproteínas. Por presentar propiedades químicas y estructurales diferentes, el
predominio de un componente sobre otro determina muchas de las características de los tejidos
donde están presentes. Brevemente, proteínas colágenas y elásticas determinan rigidez, forma y
flexibilidad en un tejido; GAGs y PGs, por ser polianiones altamente hidratados, confieren al tejido
una consistencia gelatinosa; las glicoproteínas, ligan células y otros componentes de la MEC
contribuyendo a la cohesión del tejido como un todo (Hay, 1991).
Si bien durante muchas décadas a la MEC se le atribuyó un papel fundamentalmente de soporte y
relleno, actualmente, son ampliamente reconocidas las funciones ejercidas por moléculas de la
matriz en relación al comportamiento celular. La interacción MEC-célula, mediada por receptores
específicos, puede inducir no sólo modificaciones en las propiedades adhesivas de las células sino
también intervenir en su migración, proliferación y diferenciación.
4.2. La matriz extracelular que recibe al embrión.
En las primeras etapas del proceso de implantación embrionaria en roedores, el tejido conjuntivo
endometrial se transforma en un tejido totalmente modificado. Esos cambios morfológicos y
funcionales que ocurren en el endometrio llevan en su conjunto el nombre de reacción decidual, y
preparan al endometrio para recibir al embrión. Durante dicha reacción, los fibroblastos aumentan
intensamente en número (Martin y Finn, 1968), se transforman en células poliédricas y establecen
contactos intercelulares de tipo nexo y adherentes (Abrahamsohn, 1983) confiriendo a la decidua
un aspecto epiteloide. La decidualización se origina en la región antimesometrial del sitio de

Introducción: metaloproteasas
55
�
implantación y progresa en dirección mesometrial. Debido a su intensa actividad proliferativa, la
decidua adquiere un aspecto celular denso y compacto. La proximidad entre las células deciduales
resulta en una drástica reducción del espacio extracelular, donde a su vez, grandes alteraciones
cuantitativas y cualitativas de los componentes de la MEC son observadas (Zorn y col., 1986).
Particularmente en roedores, el estroma uterino que recibe al embrión, como se mencionó, es un
tejido morfológica y funcionalmente modificado que se caracteriza por la presencia de células
deciduales y de una MEC de composición peculiar (Abrahamsohn y Zorn, 1993) que sufre
significativas modificaciones en cuanto a sus componentes: laminina, fibronectina, colágenos,
glicosaminoglicanos, proteoglicanos (Wewer y col., 1986; Abrahamsohn y Zorn, 1993) y ácido
hialurónico (Zorn y col., 1995). Estos cambios contribuyen a la formación de un medio interno más
adecuado para la penetración y sobrevida del embrión en la pared uterina. Es decir, la
remodelación de la MEC, tiene un carácter estructural permitiendo que el útero adquiera las
características necesarias para soportar el crecimiento del embrión. La remodelación de los
componentes de la MEC se inicia en la fase de preimplantación. Un ejemplo interesante de las
modificaciones que ocurren durante la decidualización es el cambio en relación a la cantidad y
distribución de fibras colágenas en el endometrio (Zorn y col., 1986). Alrededor del día 4 de
gestación en el ratón, hay una drástica reducción en la cantidad de fibras colágenas, que son
gradualmente reemplazadas por fibras reticulares. Sin embargo, ya durante la fase de
implantación, día 5 de gestación, se ha descripto la aparición de fibras colágenas muy espesas
entre las células deciduales, donde el diámetro fibrilar se incrementa significativamente hasta el
día 7 de gestación (Zorn y col., 1986). Estos cambios están en parte relacionados con la capacidad
atribuida a la decidua de “contener” la invasión trofoblástica (Amoroso, 1952). Así, ha sido
comprobado que las células trofoblásticas cuando son implantadas en lugares extrauterinos como
testículos, riñones y bazo (tejidos donde no ocurre decidualización) poseen una intensa actividad
fagocitaria, invasiva y destructiva (Amoroso, 1952). La decidua, entonces, regularía los
movimientos del trofoblasto formando una barrera física durante su penetración y generando un
medio enriquecido en citocinas que promueven la adhesión del trofoblasto más que su migración
(Kirby, 1960), así como la liberación de inhibidores de proteasas que controlan la invasividad del
trofoblasto (Alexander y col., 1996).
Como contrapartida, durante la invasión trofoblástica, sea para alojar al embrión en el
endometrio, para alcanzar una fuente nutricional para el desarrollo embrionario (la sangre
materna) o para aumentar el área funcional de la placenta, el trofoblasto debe ser capaz de

Introducción: metaloproteasas
56
�
adherirse e invadir a través de diferentes componentes de la MEC del estroma decidualizado,
atravesando las láminas basales vasculares ricas en laminina, colágeno de tipo IV, entactina, y
fibronectina (Damsky y col., 1993). Así, la invasión endometrial no es tanto una consecuencia de la
expansión provocada por el crecimiento embrionario, sino por el contrario, es un proceso
bioquímicamente activo de las células trofoblásticas, regulado en parte por la decidua. Este
proceso invasivo trofoblástico está asociado tanto con la expresión de moléculas y receptores
específicos para componentes de la MEC como con la secreción de proteínas capaces de degradar
estos componentes (Bischof y Martelli, 1992). Una serie de enzimas proteolíticas de acción
específica sobre los componentes de la MEC, como colagenasas y gelatinasas, han sido asociadas
al trofoblasto con fenotipo invasivo (Bischof y col., 1994).
Asimismo, las interacciones adhesivas entre las células y la MEC son de vital importancia para el
desencadenamiento de otros eventos, como la diferenciación y proliferación celular durante el
desarrollo del embrión (Hay, 1990). Por lo tanto, el papel de los componentes de la MEC, y en
particular la expresión y actividad de las metaloproteasas, son de gran relevancia y por ello dichas
proteasas son frecuentemente estudiadas.
2.3. Metaloproteasas en el desarrollo embrio-trofoblástico.
El éxito de la implantación y de la penetración del embrión en el tejido materno depende del
desarrollo sincronizado tanto de un fenotipo invasivo por parte del embrión como de la
receptividad del endometrio. Estos procesos son acompañados por una extensa remodelación de
los componentes de la MEC, involucrando un delicado equilibrio entre la síntesis y degradación de
diversas moléculas (Zhang y col., 2004). Adquieren gran importancia entonces las metaloproteasas
(MMPs, del inglés Matrix Metalloproteinases), una familia de enzimas proteolíticas zinc-
dependientes, capaces de degradar componentes de la MEC. La mayoría de las MMPs son
secretadas como proenzimas inactivas (zimógenos), las cuales son procesadas a la forma activa
por otras proteasas en el compartimento extracelular. Las MMPs participan de la remodelación de
la MEC degradando sus componentes proteicos (Bischof y col., 2000). La expresión de MMPs
normalmente es baja en los tejidos y es inducida cuando la remodelación de la MEC es requerida.
Bajo condiciones fisiológicas, las MMPs se encuentran finamente reguladas, a diferentes niveles: a
nivel transcripcional, a nivel de secreción y a nivel proteico por la activación de la proforma
inactiva o la inhibición de la forma activa (dado que su actividad es específicamente bloqueada por

Introducción: metaloproteasas
57
�
los inhibidores TIMPs, del inglés Tissue Inhibitors of Matrix metalloproteinases, o por estabilización
de MMPs por glicosilación, por ejemplo) (Cohen y col., 2006).
Miembros de la familia de las MMPs han sido implicados tanto en proceso fisiológicos como
patológicos (como la progresión tumoral) (Hahn-Dantona y col., 1999). Las MMPs juegan un papel
fundamental en procesos de invasión, migración y proliferación celular, no sólo por el clivaje
directo de componentes “barrera” de la matriz, sino por liberar factores de crecimiento presentes
en la MEC aumentando así su biodisponibilidad. De esta manera, regulan el comportamiento
celular por diferentes vías, incluyendo interacciones MEC-célula y célula-célula, así como a través
de la liberación, activación o inactivación de moléculas de señalización (autócrinas y parácrinas) y
de receptores celulares de membrana (Staun-Ram y Shalev, 2005).
Diversos estudios en roedores, han implicado a la MMP-2 y MMP-9 como las principales enzimas
de las células trofoblásticas encargadas de la remodelación de la MEC durante la implantación e
invasión trofoblástica a través de la decidua y de la vasculatura materna (Zhang y col., 2004), de
modo que su expresión, espacial y temporal, está finamente regulada a lo largo de la gestación
(Bischof y col, 2000; Cohen y col., 2006).
Las MMP-2 y MMP-9 son proteasas que degradan fundamentalmente colágeno de tipo IV, el
principal componente de las membranas basales, así como colágeno desnaturalizado (por lo que
son consideradas gelatinasas). Sin embargo, la MMP-2 también es capaz de degradar colágeno de
tipo I, V, VII, X y XI, fibronectina, laminina, agrecan, elastina y vitronectina, mientras que la MMP-9
degrada colágeno V y XIV, agrecan, elastina y vitronectina (Cohen y col., 2006).
La estructura básica de las MMPs consiste en una serie de dominios característicos: un péptido
señal (amino-terminal) que dirige la secreción al exterior de la célula, un propéptido que mantiene
a la enzima inactiva hasta que sufre un corte proteolítico, y un dominio catalítico carboxilo-
terminal que une zinc (Figura 4.1.A). Sobre esta estructura básica aparecen diversas variantes,
como es el caso de la MMP-2 y MMP-9 (también llamadas gelatinasas A y B, respectivamente)
donde a la estructura básica se suma un dominio de tipo hemopexina (en el extremo carboxilo-
terminal), mediador de la especificidad del sustrato y las interacciones con inhibidores endógenos,
unido por una bisagra al dominio catalítico, el cual a su vez presenta un serie de repeticiones tipo
fibronectina (Egeblad y Werb, 2002; Folgueras y col., 2004) (Figura 4.1.B).

Introducción: metaloproteasas
58
�
Figura 4.1. Representación esquemática de la estructura de las metaloproteasas de la matriz
extracelular. A, representación de los miembro más simple de la familia de MMPs (matrilisinas: MMP-7
y MMP-26) conteniendo como unidad básica un péptido señal, un propétido (de alrededor de 80
aminoácidos) en la región amino-terminal (N-T) y un dominio catalítico en el extremo carboxilo-
terminal (C-T) de alrededor de 170 aminoácidos, con un sitio fijador de zinc (Zn2+
); B, representación de
la estructura de MMP-2 y MMP-9, donde además de la estructura básica, presentan un dominio
formado por cuatro repeticiones de hemopexina en el extremo C-T unido al dominio catalítico por una
pequeña bisagra (B), y un dominio tipo fibronectina (Fi) sólo presente en estás MMPs. Modificado de
Folgueras y col., 2004.
Diversos estudios han demostrado que la MMP-2 y la MMP-9 se expresan en blastocistos de ratón,
en el momento en que las células trofoblásticas penetran la membrana basal de las células
epiteliales de la luz uterina e invaden el estroma decidual subyacente. A través de estudios in-
vitro, fue demostrado que en blastocistos en expansión durante el cultivo existe expresión de
ARNm de MMP-2 luego de 72 hs de cultivo, si bien, la misma mostró ser débil comparado con el
nivel de expresión de MMP-9, la cual ya se detectó a partir de las 48 hs de cultivo (Sharkey y col.,
1996). A su vez, asociado con el alto nivel de expresión, se detectó una fuerte actividad MMP-9 en
blastocistos en expansión in-vitro, medido por zimografía, mientras que no fue detectada actividad
MMP-2 (Behrendtsen y col., 1992). Llamativamente, la depleción génica de MMP-9 no condujo a
fallas en la implantación (Vu y col., 1998), sugiriendo que si bien MMP-9 juega un papel crucial en
la degradación del estroma decidual, no es un requisito indispensable para la implantación,
probablemente, debido a que el embrión murino expresa otras proteasas que pueden compensar
la ausencia de MMP-9.
Como contrapartida, el estroma endometrial, al día 4 de gestación, antes del comienzo de la
implantación ya presenta inmunorreactividad positiva para MMP-2 y MMP-9. Al día 6 de
gestación, luego de la implantación, la inmunomarcación para MMP-2 y MM-9 se describe
principalmente fuera de la zona decidualizada en el estroma endometrial indiferenciado (exterior

Introducción: metaloproteasas
59
�
a la zona primaria de decidualizacion), con fuerte marcación en el estroma adyacente al
miometrio. Al día 8 de gestación, la inmunorreactividad para MMP-2 se detectó principalmente en
el área de las células gigantes trofoblásticas primarias (CGT 1rias
), en la capa delgada de células no
diferenciadas del estroma adyacente al miometrio, y en las células del estroma del polo
mesometrial, donde posteriormente se formará la placenta. Una fuerte marcación para MMP-9
permaneció en la capa delgada de células del estroma endometrial no diferenciadas adyacente al
miometrio y en el área de las células gigantes trofoblásticas (Alexander y col., 1996; Bany y col.
2000) (Figura 4.2). Estos estudios de inmunolocalización proteica, coincidieron con los de
hibridación in-situ en cuanto a la distribución de ARNm que codifica para MMP-2 y MMP-9 en
sitios de implantación (Harvey y col., 1995; Alexander y col., 1996; Das y col., 1997). Asimismo,
transcriptos de ambas MMPs fueron detectados por RT-PCR, tanto en embrión como en cono
ectoplacentario aislados (Bany y col., 2000), siendo también demostrada la expresión génica de
MMP-9 en cultivos de células de cono ectoplacentario de rata y su actividad proteolítica verificada
por zimografía (Peters y col., 1999). El fenotipo invasivo de las células trofoblásticas está asociado
con la activación transcripcional de MMP-2 y principalmente de MMP-9, la cual tiene su
contrapartida a nivel uterino, y en estrecha asociación con la expresión de los TIMPs, quienes
regulan el avance de las células trofoblásticas invasoras. En tal sentido, la primera ola de células
trofoblásticas gigantes emerge de la zona mural del blastocisto e invade la zona antimesometrial
de la decidua en formación para formar una red de células interconectadas de células
embrionarias en el endometrio. La zona antimesometrial es donde se inicia la reacción decidual.
Tanto ARNm como proteína de MMP-9 se expresan en dichas células CGT 1rias
. La diferenciación
del estroma materno a células deciduales va acompañada de una llamativa inducción de la síntesis
de TIMP-3. Así, la expresión de MMP-9 por parte de las CGT 1rias
se encuentra con una síntesis
elevada de TIMP-3 en las células deciduales adyacentes, quien en parte limita la expansión del
trofoblasto. La segunda ola de crecimiento trofoblástico, la expansión y avance del cono
ectoplacentario en la decidua mesometrial, al día 7,5-8 de gestación, ocurre justamente cuando la
expresión de TIMP-3 decae a niveles indetectables (Alexander y col., 1996).
Todas estas evidencias, en conjunto, sugieren un papel crítico de las MMPs y de sus inhibidores en
el proceso de invasión trofoblástica, el cual en parte se asemeja al proceso de invasión tumoral
(Cross y col., 1994). Sin embargo, bajo condiciones fisiológicas adecuadas (no patológicas), la
compleja y sincronizada serie de eventos moleculares y celulares entre el útero receptivo y el

Introducción: metaloproteasas
60
�
trofoblasto invasivo, permite la penetración “controlada” del embrión en la pared uterina (Staun-
Ram y Shalev, 2005).
Figura 4.2. Sitio de implantación entre los días 7,5 y 8 de gestación. Se muestran las distintas zonas
del sitio de implantación en base a las cuales se describió el patrón de distribución de MMP-2 y MMP-
9. Modificado de Alexander y col., 1996.
2.2. Metaloproteasas y consumo de alcohol: antecedentes en otros modelos.
Existen diversos estudios que demuestran el efecto del consumo de alcohol en las MMPs, y en
consecuencia en el fenotipo de las células que las expresan, en relación a la migración e invasión
celular. Muchos de estos estudios, conducidos en pacientes con patologías tumorales o en
modelos que las reproducen, sientan las bases para el entendimiento del comportamiento de
células con características invasivas que, por analogía, pueden en parte extrapolarse a las células
embrionarias en desarrollo. En tal sentido, existen evidencia que el consumo de alcohol
incrementa la metástasis en pacientes con carcinoma hepatocelular (CHC) (Hsiang y col., 2007). Si
bien el mecanismo aún no es del todo claro, se ha demostrado que el acetaldehído, principal
metabolito de la metabolización del alcohol, podría ser el factor involucrado en la invasividad del
CHC en pacientes alcohólicos, dado que dicho compuesto incrementa tanto la actividad gelatinasa
de la MMP-9 como la expresión de su transcrito en células HepG2 promoviendo así la invasión de
dichas células (Hsiang y col., 2007). A su vez, estudios realizados en modelos in-vitro demostraron
que el etanol puede activar a la MMP-2 en fibroblastos de manera dosis dependiente, y que el
medio condicionado proveniente de fibroblastos expuestos a etanol altera significativamente el

Introducción: metaloproteasas
61
�
comportamiento invasivo de células de cáncer de mama. A una concentración fisiológica (50–200
mg/dl), el etanol aumentó notablemente la invasividad de las células, sin embargo, a
concentraciones mayores (400 mg/dl), la inhibió; indicando un efecto bifásico del etanol en
relación a la activación de dicha MMP dependiendo de la concentración de exposición (Aye y col.,
2004). De la misma manera, estudios in-vivo en ratas (Lois y col., 1999; Partridge y col., 1999)
demostraron que el consumo de alcohol altera la expresión y/o actividad tanto de MMP-2 como
de MMP-9, indicando que dichas MMPs son target del etanol. Así, las MMPs, ya sea
incrementando la degradación de la MEC o interactuando con integrinas de la superficie celular,
pueden incrementar la movilidad celular y por lo tanto favorecer la invasión. En otros estudios del
desarrollo del sistema nervioso fetal, se sugiere que uno de los procesos que estaría alterado
como consecuencia de la exposición a etanol es la migración neuronal, donde la proteólisis de la
MEC regulada por MMPs, juega un papel fundamental en dicha migración y en el crecimiento de
neuritas del cerebro en desarrollo (Ismail y col., 2010). En el mismo contexto, se han encontrado
TIMPs down-regulados como resultado de la exposición murina in-vivo a etanol durante los días 7
y 9 de gestación (Hard y col., 2005), y en consecuencia desregulación de la migración de las células
neuronales asociada a mayor remodelación de la MEC. Por otro lado, estudios conducidos en
modelos de fibrosis hepática como resultado del consumo de alcohol, muestran que un
incremento en la actividad de determinados TIMPs, y en consecuencia una reducción del pool
activo de MMPs, son en parte responsables del desbalance entre la fibrinogénesis y fibrinólisis en
la MEC (Thompson y col., 2011).
Dados estos variados antecedentes que vinculan la desregulación en la actividad y/o expresión de
MMPs con el consumo de alcohol, el estudio de los potenciales defectos en la expresión de estas
proteasas de la matriz en el tejido embrio-trofoblástico temprano, como mecanismo asociado a las
anomalías del desarrollo luego de la exposición materna a alcohol fue de primordial interés para
entendimiento de la patogenia embrionaria temprana inducida por el alcohol.
�

Introducción: muerte celular y genotoxicidad
62
5. Muerte celular y genotoxicidad embrionaria.
5.1. Muerte celular: necrosis versus apoptosis.
La muerte celular puede producirse mediante dos mecanismos distintos, la necrosis y la apoptosis,
cuyas características morfológicas y bioquímicas son distintivas en cada uno de los procesos
(Figura 5.1).
La necrosis es la muerte de un conjunto de células o de un tejido provocada por un agente nocivo
que causa una lesión de tal gravedad que no puede ser reparada. Este proceso se produce
generalmente cuando las células se exponen a agresiones físicas o químicas graves, a variaciones
extremas de las condiciones fisiológicas (por ejemplo, hipotermia, hipoxia) o a agentes tales como
virus líticos, que generan un daño directo en la membrana plasmática y/o producen un cese del
metabolismo celular. La necrosis comienza con un deterioro de la capacidad de la célula para
mantener la homeostasis, que conduce a un ingreso de agua y de iones extracelulares por
aumento de la permeabilidad de la membrana dañada, y en consecuencia la célula se hincha. Las
organelas intracelulares y la célula en su conjunto se lisan. Durante este proceso, el ADN de las
células se escinde al azar. Debido a la descomposición final de la membrana plasmática, el
contenido citoplasmático se libera al fluido extracelular. Por lo tanto, in-vivo, la muerte celular
necrótica se asocia a menudo con daño tisular extenso, dando lugar a una importante respuesta
inflamatoria en el tejido sano adyacente (Wyllie y col., 1980; Majno y Joris, 1995; Chi y col., 2011).
Por el contrario, la apoptosis, constituye un tipo de muerte celular fisiológica, regulada y
autodirigida, por lo que es llamada muerte celular programada. Durante este proceso fisiológico
una célula específica se elimina en un momento preciso y de manera controlada, siendo la propia
célula un participante activo de su muerte, ya que depende de la activación de la propia
maquinaria interna de muerte celular (White, 1996). Es decir, la apoptosis tiene su base en un
mecanismo genético y está caracterizada por una compleja cascada de eventos bioquímicos que
darán como resultado los signos típicos de la muerte celular programada.
La apoptosis está regulada a tres niveles. A nivel de membrana, existen receptores específicos
mediando señales de muerte. A nivel nuclear, el genoma contiene genes, relacionados con la
misma, que se transcriben en respuesta a la iniciación del proceso apoptótico. A nivel
citoplasmático, existen vías de señalización que median la transducción de señales, las cuales

Introducción: muerte celular y genotoxicidad
63
pueden variar según el receptor involucrado. Sin embargo, las etapas finales son similares en la
mayoría de los casos e involucra a un grupo de proteasas, las caspasas (Enari y col., 1998).
La apoptosis es frecuente en las células de los individuos adultos. A menudo es un proceso
deseable para eliminar células dañadas o mutadas (células pretumorales) y generar el recambio
celular normal para mantener la homeostasis de los tejidos. Durante el desarrollo, la apoptosis es
un proceso natural necesario para la formación de estructuras y órganos en embriones y fetos, y al
igual que en el adulto, en necesaria para la remodelación de tejidos eliminando poblaciones de
células no deseadas o dañadas (Smith, 1997).
Figura 5.1. Representación esquemática que ilustra las principales diferencias entre la muerte celular
por necrosis y por apoptosis. Como resultado de una injuria o lesión grave que no puede ser reparada
ocurre muerte celular por necrosis causando el hinchamiento de la célula y la rotura de membranas,
con consecuente respuesta inflamatoria en el tejido sano adyacente. En contraste, durante la apoptosis
no hay inflamación asociada. En la apoptosis, inicialmente la cromatina se condensa alrededor de la
membrana nuclear interna, el citoplasma se contrae, el núcleo se fragmenta, y luego lo hace la célula
formando cuerpos apoptóticos rodeados de membrana, los cuales son fagocitados por células vecinas.

Introducción: muerte celular y genotoxicidad
64
5.2. La apoptosis.
El término apoptosis apareció por primera vez en la literatura biomédica en 1972 para delinear un
modo estructuralmente distintivo de muerte celular, responsable por la eliminación de células
aisladas dentro de un tejido vivo (Kerr y col., 1972).
Los signos típicos de la apoptosis son el resultado de una compleja cascada de eventos. A nivel
bioquímico, ciertas señales conducen a la activación de una familia proteasas intracelulares, la
caspasas, las cuales desempeñan un papel fundamental en la iniciación y ejecución de la apoptosis
inducida por diversos estímulos (Wyllie, 1997). A nivel morfológico, las características distintivas
de este tipo de muerte celular incluyen un cambio en la morfología celular. La célula se redondea
indicando alteraciones en la adhesión celular (Wyllie y col., 1980; Collins y col., 1997) y aunque la
membrana plasmática mantiene su integridad, ya en las primeras etapas de la apoptosis se
producen cambios en la superficie celular. Entre ellos, ocurre una redistribución de fosfolípidos de
la membrana plasmática (Hardy, 1999), caracterizada principalmente por una translocación de la
fosfatidilserina desde el lado interno al externo, quedando expuesta en la superficie celular
(Martin y col., 1995). A nivel nuclear ocurre la marginación, condensación, y finalmente
fragmentación de la cromatina, siendo este último, un acontecimiento irreversible que
compromete a la célula a morir. Se ha demostrado que esta fragmentación del ADN es el resultado
de la activación de endonucleasas dependientes de Ca2+
y Mg2+
, enzimas que escinden
selectivamente el ADN en sitios ubicados entre unidades nucleosomales generando así fragmentos
de ADN oligonucleosómicos de tamaño bastante específico (Wyllie y col., 1980; Arends y Wyllie,
1991; Hardy, 1999). A nivel citoplasmático ocurre una evidente contracción celular acompañada
de una transitoria pero significativa formación de “ampollas” en la superficie. La fisiología
mitocondrial se altera en las células apoptóticas, evidenciada por una alteración en la
permeabilidad de la membrana mitocondrial y consecuente liberación de proteasas activadoras
específicas de la apoptosis. El proceso apoptótico culmina con la fragmentación del citoplasma y
del núcleo formando pequeños cuerpos subcelulares delimitados por membrana, los cuerpos
apoptóticos. In-vivo, las modificaciones en diversas moléculas de superficie, incluyendo a la
fosfatidilserina de la membrana plasmática, actúan como señal en los cuerpos apoptóticos,
promoviendo su fagocitosis por células vecinas sanas y/o macrófagos. Debido a este mecanismo
eficiente para la eliminación de células apoptóticas, este tipo de muerte celular no conlleva una

Introducción: muerte celular y genotoxicidad
65
respuesta inflamatoria asociada. Si, eventualmente, los cuerpos apoptóticos no son fagocitados
sufren un aumento de la permeabilidad de la membrana, una distensión citosólica, se hinchan y,
finalmente, se lisan. Esta fase terminal de la muerte celular se denomina necrosis secundaria
(Hardy, 1999; Chi y col., 2011).
La apoptosis es una característica esencial de muchos procesos normales, sirviendo a una variedad
de propósitos como ya fue mencionado (Wyllie y col., 1980). Puede ser el resultado de la
activación de un programa endógeno y/o puede ser inducida por estímulos externos. Si bien, este
proceso está finamente regulado, en ciertos casos, cuando esta regulación falla la apoptosis puede
desencadenar procesos patológicos.
Diversos son los métodos utilizados para el análisis de la muerte celular por apoptosis. Las
primeras etapas de la apoptosis pueden ser detectadas por marcación con anexina V, que pone en
evidencia los cambios ocurridos en la membrana plasmática, por poseer alta afinidad por la
fosfatidilserina de las membranas celulares (Eijnde y col., 1997). También son utilizados los
sistemas de marcación de caspasas u otros componentes de la cascada de apoptosis (Spanos y
col., 2002). Las etapas posteriores de la apoptosis pueden identificarse por marcación directa del
ADN. Los compuestos fluorescentes con afinidad por el ADN pueden poner en evidencia núcleos
con morfología condensada o fragmentada. Por lo tanto, mediante la marcación del ADN con
fluorocromos específicos tales como el Hoechst o el 4`, 6-diamino-2-fenilindol (DAPI), la
morfología nuclear típica de la apoptosis puede ser identificada. Compuestos como el Hoechst
33342 penetran la membrana plasmática y marcan el ADN, tanto de las células vivas como de las
apoptóticas, cuyas membranas plasmáticas se encuentran intactas. En contraste con las células
normales, los núcleos de las células apoptóticas presentan su cromatina altamente condensada y
coloreada de manera uniforme y azul intenso luego de una corta incubación con Hoechst 33342
(Dive y col., 1992). Sin embargo, para la identificación específica de células apoptóticas es
ampliamente utilizada la técnica de TUNEL (del inglés, terminal deoxynucleotidyl transferase dUTP
nick end labeling), la cual permite poner en evidencia la fragmentación del ADN nuclear, revelando
una coloración focal en el interior del núcleo apoptótico y/o de los cuerpos apoptóticos (Gavrieli y
col., 1992). Esta técnica se basa en la marcación enzimática, con nucleótidos modificados, de los
extremos 3`OH libres producto de la fragmentación del ADN debido a la acción de endonucleasas
durante el proceso de apoptosis. En contraste con las células apoptóticas, los núcleos de células
normales o en proliferación, poseen un número insignificante de extremos 3`OH.

Introducción: muerte celular y genotoxicidad
66
5.3. La apoptosis durante el desarrollo embrionario.
Los mecanismos fisiológicos controlados para la eliminación de células que se encuentran en
exceso, que son anormales o potencialmente peligrosas, son fundamentales para el normal
desarrollo embrionario. En tal sentido, el fenómeno de la muerte celular está establecido
firmemente en el desarrollo del embrión de mamíferos ya desde etapas tempranas (Hardy y col.,
1997). Es por ello, que la apoptosis ha recibido y aún recibe gran atención por su destacado papel
en la embriogénesis, como partícipe necesaria de eventos que requieren una eliminación selectiva
de células, por ejemplo antes de la implantación, tan pronto como las células totipotentes se
empiezan a diferenciar en el blastocisto (Hardy, 1997); durante ingreso a través del epitelio
uterino e implantación en el estroma endometrial (Straszewski-Chavez y col., 2005); durante la
formación de la cavidad amniótica poco después de ocurrida la implantación (Coucouvanis y
Martin, 1995); durante la formación de los dígitos en los esbozos de las extremidades, entre otros.
De la misma manera, es llamativa su participación en condiciones patológicas donde según su
severidad, la apoptosis, puede estar involucrada en la pérdida embrionaria (Gjørret y col., 2003).
5.3.1. La apoptosis durante el desarrollo embrionario de preimplantación.
Desde mediados de la década de 1970, han habido descripciones ultraestructurales y de
microscopía convencional de células muertas y en proceso de muerte en embriones de
preimplantación en varias especies de mamíferos incluyendo ratón (El-Shershaby y Hinchliffe,
1974; Copp, 1978), vaca (Mohr y Trounson, 1982), mono (Enders y Schlafke, 1981) y humano
(Lopata y col., 1982; Mohr y Trounson, 1982; Sathananthan y col., 1982). Los rasgos principales
fueron la formación de ampollas en la membrana, la cromatina condensada, la fragmentación
nuclear y la fragmentación citoplasmática (Handyside y Hunter, 1986; Hardy y col., 1989; Brison y
Schultz, 1997; Hardy, 1997). Estas observaciones y el creciente interés en la importancia de la
apoptosis en el desarrollo temprano, guiaron numerosas investigaciones en el área. Así, la
presencia de diversos componentes moleculares de la cascada de apoptosis fue demostrada en
embriones de preimplantación de ratón, humano y bovinos (Warner y col., 1998; Exley y col.,
1999; Gutierrez-Adan y col., 2004; Jurisicova y Acton, 2004). Las células embrionarias poseen

Introducción: muerte celular y genotoxicidad
67
entonces, constitutivamente, la maquinaria para ejecutar la muerte celular programada (Fabian y
col., 2007).
Las primeras evidencias de apoptosis espontánea fueron observadas a partir de la etapa de 2
células (fin del primer ciclo celular) y 4 células (fin del segundo ciclo celular) en embriones de ratón
y humanos, respectivamente, probándose con estos resultados que las primeras evidencias de
apoptosis coinciden con las etapas siguientes a la activación del genoma embrionario en cada
especie (Schultz, 1993; Jurisicova y col., 1996; Warner y col., 1998; Byrne y col., 1999). La función
de esta primera onda de apoptosis estaría relacionada con la eliminación de embriones anormales
o defectuosos, con anomalías cromosómicas, mosaicismo o que han sufrido una activación
inapropiada del genoma embrionario (Byrne y col., 1999).
Las características morfológicas distintivas de la muerte celular apoptótica han sido observadas en
mórulas y blastocistos (Handyside y Hunter 1986; Hardy y col., 1989). Llamativamente, las
evidencias muestran en una variedad de especies incluyendo ratón, rata y humanos, que un
marcado aumento en la incidencia de la apoptosis surge durante la etapa de blastocisto con
respecto a las etapas iniciales del desarrollo (El-Shershaby y Hinchliffe, 1974; Copp, 1978; Hardy y
col., 1989; Pampfer y col., 1990; Jurisicova y col., 1995; Byrne y col., 1999). La formación del
blastocisto involucra, entonces, procesos de proliferación, diferenciación y también de muerte
celular, como parte del programa normal del desarrollo que dará lugar al establecimiento de los
linajes celulares, el MCI y el TE. En la mayoría de las especies mencionadas, la apoptosis
predomina en el MCI respecto a lo observado en el TE (Hardy, 1997; Byrne y col., 1999). Ha sido
reportado que el porcentaje de células muertas en el MCI de embriones de ratón entre los días 4 y
5 de gestación es del orden del 4% (Jurisicova y col., 1998; Liu y col., 1999), o inclusive mayor,
entre el 10 y 20% (Hardy, 1997). Por el contrario, en el TE se reportaron valores menores, que
variaron entre el 1 y 3% (Hardy y Handyside, 1996; Byrne y col., 1999). La mayor incidencia de
apoptosis observada en el MCI estaría relacionada con la diferenciación celular que ocurre durante
el estadio de blastocisto. Se postula que la muerte celular programada, en este caso, eliminaría
células del MCI que aún mantienen potencialidad para formar TE después de la expansión del
blastocisto (Handyside y Hunter, 1986; Pierce y col., 1989).
Este control de calidad celular para eliminar células que están dañadas, con anomalías
cromosómicas o en exceso dentro del MCI, es crítico, ya que a partir de este linaje se formará el
feto (Hardy y col., 1993; Jamieson y col., 1994). Sin embargo, una muerte celular desregulada y

Introducción: muerte celular y genotoxicidad
68
exacerbada puede comprometer el desarrollo tardío, dado que un número mínimo de células en el
MCI es requerido para el normal desarrollo de postimplantación (Tam, 1988). Una reducción
significativa en este linaje celular puede causar una disminución en la viabilidad embrionaria (Tam,
1988; Pampfer, 1990; Kelly y col., 1992). Es decir, los procesos apoptóticos que se produzcan
durante la fase de blastocisto podrán tener un efecto beneficioso como perjudicial, dependiendo
del grado de incidencia (Fabian y col., 2005).
Desde el aspecto morfológico, la fragmentación en el desarrollo de embriones de ratón se asoció
con la apoptosis (Jurisicova y col., 1998a). También en humanos, donde un alto porcentaje de
embriones de 2 a 8 células fragmentados resultaron TUNEL-positivos, mientras que embriones no
fragmentados resultaron negativos para la misma reacción (Yang y col., 1998). A nivel
ultraestructural, blastómeras apoptóticas por lo general muestran contracción y mayor densidad
citoplasmática, hacinamiento de orgánulos, mayor cantidad de vacuolas y de lisosomas. Cuando se
realiza marcación del ADN, los núcleos de blastómeras apoptóticas muestran condensación o
fragmentación nuclear y positividad para TUNEL (Fabian y col., 2005). Las etapas terminales del
proceso se caracterizan por el deterioro celular en cuerpos apoptóticos, los cuales pueden ser
eliminados por fagocitosis de blastómeras vecinas o por extrusión al espacio perivitelino o al
blastocele, seguida de necrosis secundaria. Este último caso es más frecuente en las células de la
capa TE. Por el contrario, las células apoptóticas del MCI son fagocitadas por células vecinas
(Fabian y col., 2005). El escaso potencial de embriones con amplia fragmentación para formar
blastocistos de buena calidad morfológica e implantar (Giorgetti y col., 1995) indica que la mayoría
de los embriones con blastómeras fragmentadas tienen un defecto intrínseco. Ciertamente, un
alto grado de fragmentación se ha asociado también con una mayor incidencia de anomalías
cromosómicas (Pellestor y col., 1994), tema que se detallará más adelante.
5.3.2. La apoptosis durante el desarrollo embrionario de postimplantación.
La apoptosis desempeña un papel clave en la formación de las estructuras extraembrionarias y del
propio embrión/feto. Se ha demostrado apoptosis en las membranas fetales (Runic y col., 1998).
Estudios de microscopía electrónica revelaron cambios ultraestructurales en el amnios y en las
células trofoblásticas coriónicas consistentes con apoptosis.

Introducción: muerte celular y genotoxicidad
69
La apoptosis en embriones de postimplantación, así como durante la etapa de preimplantación,
está involucrada en procesos tales como el control del número de células, la eliminación de células
anormales, células deslocalizadas, no funcionales, o perjudiciales, pero también está involucrada
en la formación de diversas estructuras. Se ha demostrado que la formación del canal
proamniótico ocurre debido a la muerte por apoptosis de las células ectodérmicas en el core del
embrión en desarrollo. La apoptosis también contribuye a la formación de vesículas y tubos, por
ejemplo, el tubo neural. Se sabe que ocurre apoptosis durante y después de la neurulación en el
neuroepitelio. En tal sentido, se ha postulado que se requiere apoptosis para la formación de los
pliegues neurales y para la remodelación epitelial una vez que los pliegues neurales entraron en
contacto y se fusionaron (Jacobson y col., 1997; Copp y col., 2003). Las neuronas y
oligodendrocitos, que son producidos en exceso durante el desarrollo del sistema nervioso,
también son eliminados por apoptosis (Narayanan, 1997). Avanzado en el desarrollo, otro ejemplo
de la función de la apoptosis, es el observado en la placa de la mano durante la formación de los
dígitos. Inicialmente, los dedos comienzan a formarse como condensaciones de tejido
mesenquimático. Estas condensaciones son los signos primarios de la ubicación del dígito futuro
(Zakeri y Ahuja, 1997). Allí, posteriormente, las células mesenquimáticas interdigitales se eliminan
por apoptosis (Mori y col., 1995), la cual normalmente procede y se limita a determinadas áreas
del mesénquima (Hurle y col., 1996). La importancia de una fina regulación en la muerte celular
programada para el desarrollo normal se pone de manifiesto en mutantes de ratón en que se
produce “dedo en martillo”, en los que una insuficiente apoptosis subyace en el desarrollo
anormal de miembros (Zakeri y col., 1994). Otras estructuras embrionarias que se eliminan por
apoptosis son los conductos de Müller y de Wolff, según el sexo (Lee y col., 1988; Larsen, 1993).
5.3.3. Factores que influyen en una apoptosis anormal: antecedentes.
Lo expuesto anteriormente demuestra el papel importante de la apoptosis en el desarrollo
normal. A diferencia de la necrosis, la apoptosis juega un papel tanto protector en el desarrollo del
embrión (mediante la eliminación de células defectuosas) como estructural (participando en el
desarrollo de estructuras) (Fabian y col., 2005). Sin embargo, bajo ciertas condiciones, el curso
normal de la apoptosis durante la etapa embrionaria o fetal puede ser perturbado. En tal sentido,
evidencia acumulada demuestra que el arresto embrionario, la pérdida de la gestación temprana,

Introducción: muerte celular y genotoxicidad
70
así como el nacimiento de progenie con defectos congénitos, puede estar asociado a distorsiones
del adecuado patrón espacio-temporal de apoptosis (Brill y col., 1999).
Cada vez es más reconocido que la cascada de eventos intracelulares que conducen a la
eliminación de células raramente se produce en un contexto estrictamente endógeno estando el
entorno o medio ambiente embrionario/fetal involucrado como disparador de la muerte celular.
Por lo tanto, la regulación de la apoptosis implica un delicado equilibrio de las señales externas y
expresión de genes endógenos (Hardy, 1999), de tal manera que un aumento de la incidencia de la
muerte celular puede ser un indicador del inadecuado entorno in-vivo o in-vitro al que están
sometidos los embriones. Desequilibrios en los niveles de factores de crecimiento, de
supervivencia y/o de hormonas, pueden incidir en la tasa de apoptosis embrionaria (Byrne y col.,
2002; Hardy y Spanos, 2002; Fabian y col., 2004).
El aumento de la muerte celular también ocurre en respuesta a la exposición a agentes
teratogénicos. Llamativamente, los teratógenos son capaces de inducir la muerte celular en zonas
donde, como parte del programa normal de desarrollo, ya ocurre apoptosis, sugiriendo que
existiría un vínculo entre la muerte celular programada y la “inducida” por un teratógeno (Sulik y
col., 1988).
Numerosos estudios han mostrado que diversos teratógenos inducen una excesiva apoptosis en
embriones tempranos produciendo una deficiencia en el desarrollo (Hsuuw y col., 2005; Huang y
col, 2003) y conduciendo, muchas veces, a la aparición de anomalías estructurales en el feto e
inclusive a la muerte embrionaria. Es decir, el desarrollo de postimplantación es potencialmente
propenso a ser afectado por una alta apoptosis en la fase de preimplantación (Gjørret y col.,
2003). A su vez, varios trabajos han demostrado también que los diferentes tejidos después de la
implantación del embrión de los mamíferos son diferencialmente sensibles a la inducción de la
muerte celular por teratógenos, con una llamativa sensibilidad de las células del sistema nervioso
central en desarrollo. Una muerte celular excesiva en el neuroepitelio craneal puede alterar el
cierre del tubo neural anterior dejando al embrión con un número insuficiente de células para que
ocurra un cierre adecuado, lo que en muchos casos resulta en exencefalia fetal (Copp y col., 2003).
Diversos estudios han puesto de manifiesto que el efecto de la mayor parte de los teratógenos
químicos y físicos se asocia con la inducción de la apoptosis en órganos blanco.
Varias publicaciones concuerdan en que la apoptosis es el tipo de muerte celular que ocurre en
embriones de ratón como consecuencia de la exposición al alcohol. Los fundamentos se basan en

Introducción: muerte celular y genotoxicidad
71
que las células embrionarias expuestas a alcohol presentan varias características estructurales y
bioquímicas que son exclusivas de la apoptosis (Sulik y col., 1988). Otras pruebas que apoyan esta
idea surgen a partir de estudios con inhibidores específicos de caspasas (Cartwright y col., 1998).
En conjunto, estos hallazgos sugieren que la exposición prenatal al alcohol puede dar lugar a la
eliminación apoptótica de células embrionarias; sin embargo, está comprobado que no todas las
células se ven afectadas por igual. Muchas sustancias tóxicas, incluyendo al alcohol, son
específicas en sus acciones y afectan sólo a un subconjunto de células en el embrión. El momento
de la exposición a alcohol juega un papel importante en la inducción del efecto negativo sobre el
organismo en desarrollo, dado que los tejidos poseen picos de mayor susceptibilidad a una posible
disrupción en determinadas etapas del desarrollo. En relación a este punto, es ampliamente
reconocida la vulnerabilidad del embrión a sufrir malformaciones faciales como consecuencia de la
exposición a etanol. El análisis de estos defectos faciales inducidos por el alcohol reveló ciertos
puntos comunes entre las estructuras afectadas. En particular, muchas de estas estructuras
derivan del mismo linaje celular: las células de la cresta neural craneana. Este linaje se origina en
el neuroectodermo del tubo neural primitivo y migra del mismo para luego diferenciarse en una
amplia variedad de estructuras craneofaciales (Noden, 1991). Por lo tanto, estas células fueron y
son actualmente el foco de investigaciones que muestran lo particularmente vulnerables que son
a la muerte celular inducida por el alcohol. Numerosos estudios tanto en roedores como en pollos
son muestra de ello, donde la exposición a alcohol en etapas específicas del desarrollo
embrionario temprano resultó en la muerte significativa de células de la cresta neural craneana, a
través de la activación de la propia maquinaria de apoptosis presente en ellas (Smith, 1997).
A partir de estos estudios se postuló a la apoptosis inducida por alcohol como uno de los posibles
responsable de la producción de las anomalías características del FAS (Smith, 1997), y se señaló
como ventanas de sensibilidad o períodos críticos para la inducción de defectos faciales, al corto
momento en el cual se diferencian las tres capas germinales básicas del embrión (la gastrulación) y
durante la formación de las primeras estructuras embrionarias (organogénesis), en roedores
(Webster y col., 1980; Dunty y col., 2006) y en pollos (Cartwright y Smith 1995).

Introducción: muerte celular y genotoxicidad
72
5.4. Genotoxicidad.
La genotoxicidad es la capacidad de ciertos agentes físicos, químicos o biológicos, considerados
genotóxicos, de causar daño al material genético al interactuar química o físicamente con la
secuencia de bases del ADN, alterar su estructura física y/o su funcionalidad. Por lo tanto, dicho
daño no incluye sólo al ADN, sino también a todos aquellos componentes celulares que se
encuentran relacionados con la funcionalidad de los cromosomas, entre ellos, proteínas que
participan de la reparación del material genético dañado, proteínas que intervienen en los puntos
de control del ciclo celular, en la condensación y descondensación del ADN, u otras estructuras
como el huso mitótico. Dichas alteraciones pueden resultar en mutaciones, desembocando las
mismas en anomalías genéticas, procesos tumorales o muerte celular (Mudry y Greizerstein,
2006).
La probabilidad que un agente particular cause daño genético está dada por su concentración, por
la vía de entrada y la distribución en el organismo, por los sistemas metabólicos de los tejidos en
los cuales se distribuye, por la reactividad del compuesto original y/o de sus metabolitos con
moléculas de las células blanco (especialmente el ADN) y por la capacidad de la célula para
rectificar o amplificar el daño (Casarett y Doull`s, 2001). Las alteraciones genéticas van desde
cambios en uno o pocos pares de bases (mutaciones genéticas) hasta cambios importantes en la
estructura de los cromosomas (aberraciones cromosómicas) o en el número (aneuploidías).
Para evaluar la potencial mutagenicidad de un agente son utilizados diferentes ensayos. Así, la
evaluación de la actividad genotóxica de una sustancia in-vivo puede realizarse a través del test de
micronúcleos (MN). Un MN es literalmente un núcleo pequeño y claramente reconocible al
microscopio óptico por su coloración, tamaño, forma y por estar cercano al núcleo principal. Su
formación puede ser inducida por ciertos agentes. Este ensayo permite censar daños producidos
por agentes aneunógenos (agentes capaces de inducir aneuploidías, es decir, variaciones en el
número cromosómico por afectar al huso mitótico/meiótico, por ejemplo) y clastógenos (agentes
capaces de inducir rupturas en los cromosomas e incrementar la tasa normal de aberraciones
cromosómicas). Por lo tanto, el MN es un indicador indirecto de daño en el material genético
(pérdidas y roturas cromosómicas). Esta estructura sólo se pone de manifiesto luego de la división
celular, donde tanto fragmentos acéntricos, cromátides enteras e inclusive cromosomas que no
han podido ser incorporados en los núcleos hijos al momento de la división se muestran formando

Introducción: muerte celular y genotoxicidad
73
parte del MN. La formación de MN ocurre, particularmente, en la transición de la metafase a la
anafase, momento de la migración cromosómica hacia los polos, donde por efecto de la
exposición a un genotóxico se pueden producir cromosomas o cromátides rezagadas (evento
aneugénico) o fragmentos acéntricos que se alejan del cromosoma de origen después de la rotura
del mismo (evento clastogénico), quedando excluidos del núcleo luego de la división. Sin embargo,
la agresión genotóxica puede ocurrir en momentos previos a la fase S (Figura 5.2).
Figura 5.2. Representación esquemática de la formación de un micronúcleo. A, los micronúcleos (MN)
se forman como consecuencia de la exposición de la célula a un agente genotóxico; B,
estructuralmente, son pequeños cuerpos extra-nucleares originados a partir de fragmentos de
cromátides, cromátides o cromosomas enteros que debido el daño originado por el agente no son
incluidos en el núcleo principal en la telofase. Están envueltos por membrana nuclear y se asemejan a
la estructura del núcleo hijo, a pesar de ser más pequeños. Modificado de Luzhna y col., 2013.
Los genotóxicos también pueden afectar la meiosis generando, en consecuencia, aneuploidías en
las gametas. Cuanto mayor sea la diferencia en el número cromosómico en las células
aneuploides, ya sea por demás o de menos, mayor será el desequilibrio producido y, por tanto,
menor su viabilidad.
La mitosis, con sus estrictos puntos de control, garantiza en cierta forma que luego de cada ciclo
de división cada célula contenga el mismo número de cromosomas que la célula original. Los
puntos de control del ciclo celular inhiben el comienzo de una etapa hasta que la etapa anterior no
haya sido completada con éxito. Si el proceso se desarrolla de forma errónea, las consecuencias
para las células pueden ser negativas: muerte o el origen de anomalías severas.
Si al ciclo celular se le adjudicara un comienzo, este se ubicaría al final de la fase G1. El estadio G1
ocupa la mayor parte del ciclo celular. Este es el estado en el cual la célula produce las sustancias

Introducción: muerte celular y genotoxicidad
74
necesarias para crecer. Las diferencia en la tasa de proliferación celular y duración del ciclo están
correlacionadas con la duración de la fase G1. El pasaje por un punto de restricción indica que la
célula está preparada para dividirse y, por lo tanto, comienza la replicación de su ADN.
Exceptuando a aquellas células en que ocurre endorreduplicación del ADN, por varios ciclos de
replicación del ADN sin la subsiguiente división celular, las células prosiguen la división celular. La
salida del estadio S y la entrada al estadio G2 está regulada por un punto de control de la etapa S.
Una rotura de simple cadena sin reparar es suficiente para detener el ciclo en este estadio. Tan
pronto como la replicación del ADN se ha completado exitosamente, la célula ingresa en G2. Para
el pasaje de G2 a M se debe activar el factor promotor de la mitosis. Una vez en la fase M, ocurrirá
la desorganización de la membrana nuclear, la condensación cromosómica y la formación del huso
mitótico, para luego dividir el material genético apropiadamente, de manera equitativa y
equilibrada en los núcleos de cada célula hija. Si este proceso es alterado como consecuencia de la
exposición a un agente genotóxico, siendo el huso mitótico o los cromosomas blanco del ataque,
los cromosomas podrán verse afectados en parte o en su totalidad y será dificultosa su inclusión
en el núcleo de cualquiera de las células hijas, originando este material resultante no incorporado
al nuevo núcleo, un MN (Mudry y Abrevaya, 2006).
El ensayo de MN en eritrocitos policromáticos de médula ósea de ratón es frecuentemente usado
para monitorear la potencialidad genotóxica de la exposición a alcohol, como un método directo
(in-vivo) rápido y sencillo de detectar daño genético que no implica cultivo celular ni tratamientos
in-vitro. Nuestro laboratorio, recientemente ha demostrado por medio de este test, que la ingesta
materna de alcohol induce incremento de MN en hembras murinas expuestas desde antes de la
gestación y hasta el período de organogénesis (día 10 de preñez) (Cebral y col., 2011).�
Otro indicador de daño nuclear es el grado de fragmentación en embriones de preimplantación,
siendo dicha fragmentación distinta de la fragmentación del ADN asociada principalmente con la
apoptosis a finales del desarrollo de preimplantación (Hardy y col., 2001). A su vez, la
fragmentación celular embrionaria ha sido correlacionada negativamente con el potencial de
implantación (Alikani y col., 1999), de modo que diversos trabajos estudiaron la ocurrencia de
fragmentación celular y aneuploidía, e intentaron establecer una correlación. Mediante marcación
con DAPI o con sondas específicas para determinados cromosomas que normalmente son
afectados durante el desarrollo embrionario en el humano, se demostró que efectivamente
algunos fragmentos presentaban ADN del núcleo principal (Grifo y col., 1996; Chavez y col., 2012).

Introducción: muerte celular y genotoxicidad
75
Adicionalmente, en estos casos, se encontró en forma asociada con los fragmentos
inmunomarcación positiva para LAMIN-B1, marcador de la envoltura nuclear. Por lo tanto, fue
sugerido que estas estructuras correspondían a MN, dónde por alguna razón, los cromosomas
faltantes del núcleo principal se encapsulan en micronúcleos antes de ser secuestrados en
fragmentos citoplasmáticos, estableciéndose entonces una correlación positiva entre embriones
aneuploides y grado de fragmentación (Chavez y col., 2012).
Otro parámetro utilizado como indicador de la actividad proliferativa celular y cuya modificación
respecto del patrón esperado sugiere alteraciones en el ciclo celular, es el índice mitótico. Los
agentes genotóxicos pueden modificar este índice llegando incluso hasta su máxima disminución
en el caso de la citotoxicidad en tanto que la disminución del índice mitótico en el tiempo respecto
de un valor considerado control, es indicativo de citoestaticidad.
5.4.1. Genotoxicidad del etanol: antecedentes.
Diversos estudios señalan el potencial genotóxico del etanol mostrando efectos cromosómicos en
linfocitos de individuos alcohólicos: como los intercambios de cromátides hermanas (Obe y
Anderson, 1987), la inducción de aberraciones cromosómicas (López y col., 2001) y aumento de la
incidencia de aneuploidía (Kucheira y col., 1986).
Las pruebas de genotoxicidad del etanol fueron revisadas por primera vez por la Comisión
Internacional para la Protección contra Mutágenos Ambientales y Carcinogénesis (Obe y
Anderson, 1987), sin llegar a un acuerdo sobre los efectos del etanol en relación al daño genético.
En 1995, el Departamento de la Comisión de Salud del Reino Unido en Mutagénesis de Productos
Químicos en Alimentos, Productos de Consumo y Medio Ambiente examinó la mutagenicidad del
etanol, del acetaldehído y bebidas alcohólicas. El Comité sugirió que el consumo de bebidas
alcohólicas no presentaba un importante potencial mutagénico. Sin embargo, en 2001, Phillips y
Jenkinson (Phillips y Jenkinson, 2001) hicieron una nueva revisión de la información disponible
sobre la genotoxicidad del etanol llegando a la conclusión de que los datos derivados de los
estudios, que utilizaron métodos estándar de genotoxicidad, eran incompletos. Más aún, los
resultados de algunos ensayos in-vivo en roedores fueron contradictorios. Las pruebas reportadas
para aberraciones cromosómicas in-vivo fueron todas negativas y una minoría de los test de MN
habían dado resultados positivos. Si bien, la capacidad de etanol para producir cambios

Introducción: muerte celular y genotoxicidad
76
cromosómicos ha sido motivo de controversia, en los últimos años, estudios más recientes han
demostrado que tanto el etanol por sí mismo como el producto de su oxidación, el acetaldehído,
producen efectos genotóxicos (Kayani y Parry, 2010; Kotova y col., 2013). Así, Kayani y Parry
(2010), mediante estudios in-vitro, han demostrado que tanto el etanol como el acetaldehído
generan aumentos significativos dosis-dependientes de la frecuencia de MN, sugiriendo además,
que la inducción mediada por el etanol sería principalmente a través de un mecanismo
aneugénico, mientras que la del acetaldehído lo haría a través de uno clastogénico, demostrando
el comportamiento genotóxico de ambos compuestos. La posibilidad que tiene el acetaldehído de
formar lesiones del ADN complejas, tales como entrecruzamientos ADN-ADN y aductos de ADN-
proteína, representa un aspecto importante de la genotoxicidad inducida por la exposición a
alcohol (Lorenti Garcia y col., 2009; Kotova y col., 2013).
Las alteraciones cromosómicas en células somáticas y germinales, generadas como consecuencia
de la exposición a genotóxicos en modelos experimentales y en humanos, tienen consecuencias
negativas importantes que van desde una posible teratogénesis (al generar defectos evidentes al
momento nacimiento), hasta neoplasias o incluso procesos que pueden terminar en muerte
perinatal e infantil. Diversos estudios han demostrado anomalías morfológicas, aberraciones
cromosómicas y formación de MN en embriones de preimplantación luego de la exposición de
ratones a agentes genotóxicos (Pacchierotti y col., 1994), lo cual puso en evidencia la alta
susceptibilidad que tienen estos embriones a dichos agentes. Sin embargo, poco se conoce acerca
de los efectos genotóxicos de la exposición a alcohol en los embriones de preimplantación. Al
respecto, Cebral y colaboradores (2000, 2001), analizaron los efectos genotóxicos de la
administración oral de etanol al 10% a hembras de ratón sólo durante 30 días previos a la
gestación. Al día 4 de gestación encontraron retraso en la diferenciación embrionaria, embriones
con un menor número de células respecto del control, y un elevado índice mitótico. De modo que
si bien el alcohol está asociado a arrestos embrionarios, estos resultados indicarían que dicho
efecto no sería a través de una acción citoestática del alcohol, dado que en blastocistos
provenientes de hembras durante el período pregestacional se registraron células mitóticas. Por
otro lado, el hecho que las mórulas y un elevado porcentaje de blastocistos pertenecientes a las
hembras tratadas presentaron un alto grado de fragmentación junto con anomalías morfológicas
(Cebral y col., 2000, 2001) sugieren que la exposición materna a alcohol tiene una acción
genotóxica.

Introducción: muerte celular y genotoxicidad
77
Apoyados en las alteraciones nucleares observadas en las cigotas luego de la fecundación in-vivo e
in-vitro, de animales tratados en forma prolongada con alcohol en concentraciones bajas y
moderadas, Cebral y colaboradores (2000, 2001), postularon que el retraso en el desarrollo podría
deberse a alteraciones nucleares gaméticas/embrionarias inducidas por la exposición a etanol. En
este sentido, reportes previos también habían demostrado que la exposición periovulatoria a
alcohol producía tanto embriones monosómicos como trisómicos capaces de alcanzar, aunque
más lentamente, el estadio de mórula (Kaufman y Bain, 1984b). Estos antecedentes sugieren que
el consumo materno de manera crónica de cantidades moderadas de alcohol hasta la etapa previa
a la gestación puede comprometer la competencia de los oocitos para el desarrollo del blastocisto
a través de la inducción de daño cromosómico durante la maduración oocitaria.
Por lo expuesto y según los antecedentes encontrados en la literatura de la temática de
investigación, aún hoy es escaso el conocimiento sobre los efectos genotóxicos de la ingesta
materna perigestacional de alcohol en embriones tempranos en cuanto a la incidencia de
apoptosis, fragmentación celular, frecuencia de MN, y velocidad de división celular embrionaria,
por lo que fue de interés analizar los efectos de la ingesta perigestacional de alcohol sobre los
parámetros mencionados en embriones de preimplantación como en gastrulación.

Introducción: modelos animales
79
6. Modelos animales.
Debido a múltiples consideraciones éticas, entre otras, que restringen o prohíben la investigación
en el humano, sin duda, los modelos animales son piezas cruciales para llevar a cabo objetivos
experimentales.
El uso de animales de experimentación tiene como ventaja la posibilidad de controlar las
condiciones experimentales, y esto posibilita el testeo de diversas variables. Sin embargo, deben
reconocerse sus limitaciones como representantes de condiciones similares o parciales a los casos
estudiados en seres humanos. Por eso, la elección del modelo más apropiado dependerá, en
parte, de los interrogantes planteados. La interpretación, la relevancia o validez de los resultados
obtenidos con un determinado modelo, que reproduzca una situación particular de la condición
humana, puede depender de la especie animal elegida (Tabakoff y Hoffman, 2000; Cudd, 2005).
El estudio de los efectos del consumo materno de alcohol es en sí mismo complejo, dada la
multiplicidad de mecanismos a través de los cuales el alcohol media sus efectos, ya que estos son
dependientes de la dosis, del patrón de consumo, del momento y período de exposición, entre
otros. Ningún modelo animal recrea exactamente la condición del ser humano. Por lo tanto, es
esencial ser cuidadosos al momento de la elección del modelo dependiendo del tipo de estudio e
interrogante (Cudd, 2005). En general, las especies estrechamente relacionadas con los seres
humanos, genética o evolutivamente (por ejemplo, primates no humanos), proporcionan una
mayor validez a los resultados. Sin embargo, los estudios de los efectos del consumo de alcohol en
primates no humanos son costosos y técnicamente dificultosos. Es por ello que se han utilizado
ampliamente a los roedores para el estudio del alcoholismo humano (Tabakoff y Hoffman, 2000).
Así, ratas y ratones son especies de elección para el estudio del FAS, no sólo porque recrean gran
parte de las alteraciones inducidas por el consumo de alcohol en humanos, sino porque además
presentan un período de gestación corto (19-21 días), son de fácil adquisición, manteniendo y
manipulación (Cudd, 2005). Sin embargo, debido a las diferencias intrínsecas entre los seres
humanos y los roedores, no existe un único modelo de rata/ratón que represente en su totalidad
las características de anormalidad que se producen en el síndrome de alcoholismo humano. Por
eso, se han desarrollado modelos parciales y según el foco de los interrogantes se elegirá uno u
otro (Rhodes y col., 2005). Por ejemplo, los ratones son particularmente útiles para el estudio de
los efectos físicos craneofaciales, mientras que las ratas para estudios neuroconductuales y

Introducción: modelos animales
80
neurofisiológicos, asociados al FAS. Aunque actualmente se ha conseguido recrear también las
alteraciones conductuales en ratones (Becker y col., 1996).
Los modelos in-vitro, como ser la utilización de células y tejidos ya sean de origen humano o no,
tienen como desventaja la pérdida del contexto fisiológico general, de modo que, sin duda la
utilización de los modelos animales in-vivo constituye un procedimiento más adecuado para el
estudio de las alteraciones fisiológicas, aproximándose a una situación o contexto fisiológico más
real (Tabakoff y Hoffman, 2000).
En este contexto, se debe considerar entonces la importancia de la elección del modelo animal, en
cuanto a su origen y background genético, características que influirán en el resultado o efecto
esperado.
La posibilidad de contar con líneas de roedores de laboratorio que están genéticamente definidas
ha sido uno de los principales avances de la ciencia de los animales de laboratorio. Las líneas
consanguíneas (inbred strains) de ratones, ratas y cobayos, ya hace más de 80 años, son el
prototipo de las líneas genéticamente estandarizadas. Su constitución genética fijada en forma
casi definitiva brinda gran valor experimental porque permiten realizar experimentos eliminando
la variabilidad de origen genético. La consanguinidad es el apareamiento entre individuos
emparentados. Por lo tanto, una línea consanguínea, también llamada endocriada, endogámica o
cepa resulta entonces del apareamiento sistemático e ininterrumpido entre hermanos y
hermanas, por más de 20 generaciones. Para asegurar la consanguinidad de la línea, es muy
importante que todos los animales desciendan de un único par de progenitores. La práctica
sistemática de la consanguinidad disminuye la frecuencia de los genotipos heterocigotas y genera
una progresión hacia el estado homoalélico (una sola variante alélica presente), donde algunos
alelos (en principio al azar) son fijados en ese grupo de animales. De esta forma, cada línea
consanguínea representa una colección única de genes (alelos) imposible de repetir. Brevemente,
las principales características de las líneas consanguíneas son: (i) la isogenicidad, (ii) la
homocigosis, (iii) la individualidad, (iv) la uniformidad fenotípica y (v) la estabilidad genética a largo
plazo. La isogenicidad, o igualdad genética, es sin dudas la característica más importante de estas
líneas. El alto porcentaje de homocigosis (mayor al 98%) es otro rasgo particular de estos
animales. La asociación de los caracteres fijados en cada línea particular genera una individualidad
con respecto a sus cualidades, rasgo que debe ser tenido en cuenta a la hora de la elección de una
línea para investigación. La sensibilidad a ciertas enfermedades infecciosas, la tendencia a

Introducción: modelos animales
81
desarrollar enfermedades autoinmunes (como lupus y diabetes) y la susceptibilidad a los
carcinógenos son sólo algunos ejemplos de la gran utilidad de estas líneas como animales de
laboratorio de uso particular. Cada línea consanguínea tiene entonces sus características
particulares, desde la predisposición al desarrollo espontáneo de ciertas patologías, la habilidad
para aprender, la vida media, la respuesta reproductiva, el comportamiento y la susceptibilidad a
los agentes infecciosos. Por esto mismo, la elección de la línea más apropiada es una parte crítica
del diseño y la planificación de un experimento. Dado que se trata de animales genéticamente
idénticos, la uniformidad fenotípica de las líneas consanguíneas es otro rasgo importante que nos
permite afirmar que la variabilidad en los parámetros experimentales se debe exclusivamente a
factores no genéticos (ambientales o metodológicos). Esta uniformidad permite lograr precisión
estadística con un menor número de animales que los necesarios al trabajar con grupos no
consanguíneos. A su vez, la uniformidad hace posible la comparación de resultados
experimentales entre animales de diferentes laboratorios y además, debido a su relativa
estabilidad genética, a lo largo del tiempo (Tabakoff y Hoffman, 2000; Benavides y Guénet, 2003).
Sin embargo, las cepas (líneas consanguíneas) poseen una desventaja. Resulta difícil generalizar
los resultados obtenidos a partir de una cepa endogámica a la condición de la población humana
(Tabakoff y Hoffman, 2000).
Las primeras cruzas consanguíneas de ratones fueron comenzadas por Clarence C. Little en 1909 y
tuvieron como consecuencia la creación de la línea DBA. En ese mismo año, Helen D. King inició las
primeras líneas consanguíneas de rata, las cepas albinas WKA y PA, en el Wistar Institute. En el año
1918, en el Cold Spring Harbor Laboratory, New York, Little comenzó los cruzamientos que darían
origen a las líneas de ratones más famosas, incluyendo C57BL/6, C57BL/10, C3H, CBA y BALB/c,
muchas de las cuales tienen en la actualidad más de 150 generaciones de endocría. A pesar de la
existencia de cientos de líneas consanguíneas, en la actualidad, el 70-80% de los trabajos de
experimentación se basan principalmente en el uso de diez líneas consanguíneas. Entre ellas, la
línea de ratón más común es BALB/c, debido a que es una excelente línea de uso general. Las
líneas C3H, C57BL/6, CBA y DBA/2 también son muy aceptadas (Benavides y Guénet, 2003).
Por otro lado, también se han empleado líneas murinas híbridas. Los ratones Híbridos F1,
corresponden a la primera generación obtenida de la cruza de dos líneas consanguíneas. El ratón
híbrido F1 hereda un juego de cromosomas completo de cada progenitor, por lo tanto, es

Introducción: modelos animales
82
heterocigota para todos aquellos loci en los cuales difieran las líneas parentales. Además, los
ratones F1 generados a partir de las mismas líneas serán genéticamente idénticos (isogénicos).
El vigor híbrido es un rasgo sobresaliente de estos animales, respecto al de las líneas parentales,
junto con una mejor adaptación al medio (Benavides y Guénet, 2003).
A pesar del empleo generalizado de las cepas consanguíneas o híbridos como modelos animales
corrientes, ratas y ratones no consanguíneos exocriados o exogámicos (random bred o outbred),
son ampliamente usados en investigación en todo el mundo. Su gran popularidad se debe
especialmente a su menor costo respecto al de las líneas consanguíneas y a su vigor reproductivo.
Debido a la falta de homogeneidad genética y a que no están definidos genéticamente, estos
animales son nombrados como grupos o colonias, y no como líneas o cepas. Para mantener
correctamente estos grupos de roedores, no debe superarse el 1% de endocría por generación.
Esto se logra cruzando animales no emparentados. Estos grupos de roedores son los que mejor
representan la variabilidad genética de una población humana típica, característica que puede ser
confirmada por estudios de frecuencia génica de, por ejemplo, genes de isoenzimas, o por el
porcentaje de heterocigosis de marcadores moleculares (microsatélites). El polimorfismo
generado en estos grupos es alto y se acerca al de las poblaciones humanas. Por esta
característica, los grupos de ratas y ratones exocriados son muy usados en toxicología y
farmacología, siendo también valiosos modelos para el estudio de enfermedades humanas
(Benavides y Guénet, 2003).
La mayoría de los ratones outbreds tienen un origen común. Descienden de una sola colonia Swiss
de la cual, un grupo de 9 ratones fueron llevados, por Clara Lynch en 1926, desde Suiza hacia
Estados Unidos, donde posteriormente se originaron las colonias de ratones hoy conocidas, entre
ellas: MF1 (Mike Flack 1) y CD-1 (Caesarian Derived 1). Sin embargo, no todos los animales
outbreds descienden de ratones Swiss, entre ellos, la colonia CF-1 (Carworth Farms 1) que
probablemente es de origen albino salvaje (Yalcin y col., 2010). También las ratas Wistar
Hannover, Wistar Kyoto, Sprague-Dawley, Long Evans y Zucker son animales no consanguíneos.
Las primeras colonias de ratas albinas fueron comenzadas en 1906 en el Wistar Institute, donde se
originaron la mayoría de estos grupos exocriados.
A continuación, en la Tabla 6.1, se aprecia un resumen de las características distintivas de cada
modelo.
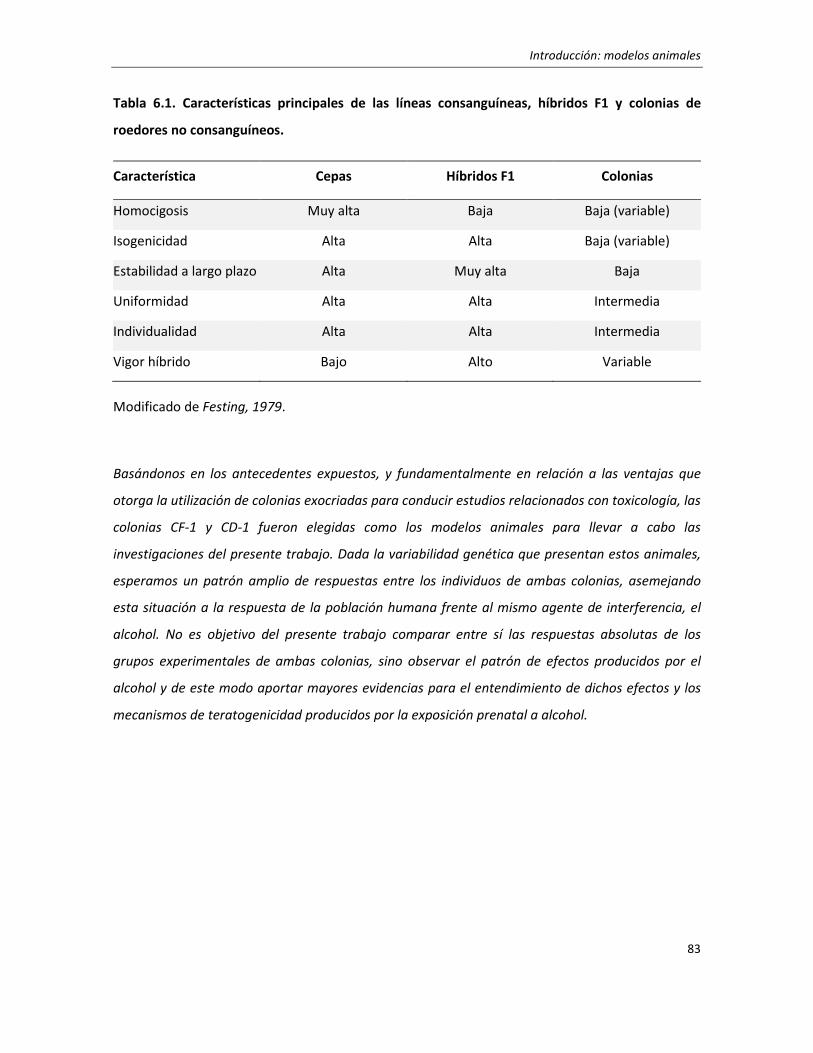
Introducción: modelos animales
83
Tabla 6.1. Características principales de las líneas consanguíneas, híbridos F1 y colonias de
roedores no consanguíneos.
Característica Cepas Híbridos F1 Colonias
Homocigosis Muy alta Baja Baja (variable)
Isogenicidad Alta Alta Baja (variable)
Estabilidad a largo plazo Alta Muy alta Baja
Uniformidad Alta Alta Intermedia
Individualidad Alta Alta Intermedia
Vigor híbrido Bajo Alto Variable
Modificado de Festing, 1979.
Basándonos en los antecedentes expuestos, y fundamentalmente en relación a las ventajas que
otorga la utilización de colonias exocriadas para conducir estudios relacionados con toxicología, las
colonias CF-1 y CD-1 fueron elegidas como los modelos animales para llevar a cabo las
investigaciones del presente trabajo. Dada la variabilidad genética que presentan estos animales,
esperamos un patrón amplio de respuestas entre los individuos de ambas colonias, asemejando
esta situación a la respuesta de la población humana frente al mismo agente de interferencia, el
alcohol. No es objetivo del presente trabajo comparar entre sí las respuestas absolutas de los
grupos experimentales de ambas colonias, sino observar el patrón de efectos producidos por el
alcohol y de este modo aportar mayores evidencias para el entendimiento de dichos efectos y los
mecanismos de teratogenicidad producidos por la exposición prenatal a alcohol.

Hipótesis y objetivos
84
Hipótesis de la investigación
Estudios previos de nuestro laboratorio han evidenciado que la ingesta de alcohol desde un
período corto previo a la gestación (17 días) y hasta la etapa de organogénesis temprana (día 10
de gestación) induce, en hembras sexualmente maduras de la colonia murina CF-1, aumento de
reabsorciones embrionarias, retrasa la diferenciación de los embriones, aumenta la frecuencia de
defectos de cierre del tubo neural y reduce la viabilidad embrionaria (Coll y col., 2011). En virtud
de esos hallazgos, postulamos que estas alteraciones encontradas en la formación embrio-
placentaria luego del consumo perigestacional de alcohol hasta el día 10 de gestación en ratones
CF-1, tienen origen en días previos, durante la ingesta de alcohol en los días 7-8 de gestación
(postimplantación temprana) o aún antes, durante los días 4 (preimplantación) y/o día 5 (inicio de
la implantación). Así, planteamos como hipótesis que la inducción de anomalías histomorfológicas
y/o funcionales en el linaje celular del propio embrión y trofoblástico lleva al arresto del desarrollo
embrionario en períodos periimplantatorios y, en consecuencia, a la reabsorción al día 10 de
gestación. Para probar estas hipótesis, se administró alcohol desde un período previo a la
gestación y hasta los días 4, 5 u 8 de preñez, momento en los cuales se analizaron los efectos en el
desarrollo y la morfogénesis embrio-trofoblástica, esperando demostrar que la ingesta
perigestacional de alcohol hasta dichas fases de la preñez afecta negativamente la diferenciación,
el crecimiento, la morfología embrionaria y los procesos morfogenéticos de los tejidos
embrionarios y trofoblásticos. Postulamos que estas anomalías son causadas, al menos en parte,
por acciones adversas del alcohol sobre moléculas claves que participan del desarrollo embrio-
trofoblástico durante la etapa de preimplantación y gastrulación. Así, alteraciones en ciertas
moléculas de las uniones intercelulares (adherentes y estrechas) del blastocisto y de la gástrula
temprana; además de la disrupción de la expresión, tanto proteica como génica, de
metaloproteasas (MMP-2 y MMP-9) relevantes para el proceso de implantación e invasión
trofoblástica temprana en el tejido uterino (fase postimplantatoria temprana), podrían
representar parte de los mecanismos moleculares afectados en el embrión y en el tejido
trofoblástico. Dado que la ingesta de alcohol puede inducir eventos genotóxicos en el organismo
expuesto, se postuló que, complementariamente a la generación de genotoxicidad materna,
aumentaría el riesgo genotóxico embrionario, así como la muerte celular por apoptosis en los
tejidos embrio-trofoblásticos de preimplantación y postimplantación temprana.

Hipótesis y objetivos
85
Por otra parte, las publicaciones sobre el tema demuestran que el consumo materno de altas dosis
de alcohol durante la gestación puede producir un espectro de alteraciones: malformaciones
fetales, restricción del crecimiento, muerte intrauterina y alteraciones postnatales a corto y largo
plazo. En nuestro modelo murino, postulamos que el consumo de una dosis moderada de alcohol
desde antes de la preñez y hasta fases tempranas de la gestación (preimplantación, gastrulación u
organogénesis), y luego de suprimir la administración de alcohol desde dichos períodos hasta la
fase fetal a término, produciría malformaciones fetales y anomalías placentarias al día 18 de
gestación, las que serían similares a las encontradas en el síndrome de alcoholismo fetal.
Objetivos de la investigación
Objetivo general
Fue de interés analizar las alteraciones en la diferenciación y morfogénesis del embrión y la
placenta en estadios tempranos, así como estudiar algunos de los mecanismos involucrados en la
inducción de anomalías del linaje embrionario y trofoblástico, luego de la ingesta perigestacional
de alcohol. Para ello, utilizando como modelo hembras murinas adultas de dos colonias
exocriadas, CF-1 y CD-1, nos planteamos estudiar los efectos de la ingesta de alcohol en el
desarrollo embrio-trofoblástico durante la fase de preimplantación y postimplantación temprana,
evaluando aspectos morfológicos de los embriones/tejidos embrio-trofoblásticos, la expresión de
moléculas de adhesión intercelular y de metaloproteasas, la apoptosis, y genotoxicidad en dichas
estructuras. También fue de interés investigar la existencia de posibles consecuencias adversas a
largo plazo luego del consumo de alcohol. Para lo cual se planteó el estudio de los efectos fetales y
placentarios en la gestación a término producidos por la ingesta perigestacional de alcohol hasta la
primera mitad de la preñez.

Hipótesis y objetivos
86
Objetivos específicos
I. Estudiar los efectos del consumo pregestacional de alcohol en cuanto a las variaciones en
parámetros biométricos (grado de ingestas sólidas y líquidas, y niveles de alcohol en sangre) y del
ciclo estral. -Capítulo I-
II. Estudiar los efectos del consumo perigestacional de alcohol -previo a la gestación y hasta el día
4, 5 u 8 de preñez- en la tasa reproductiva, diferenciación, crecimiento y morfología embrio-
trofoblástica in-vivo; y evaluar los efectos en la capacidad implantativa e invasiva de blastocistos
(día 5), e invasiva del trofoblasto de conos ectoplacentarios (día 8) en estudios in-vitro. -Capítulo II-
III. Estudiar los efectos del consumo perigestacional de alcohol -previo a la gestación y hasta el día
4 u 8 de preñez- en la expresión de moléculas de las uniones adherentes y estrechas de embriones
y tejido trofoblástico en el período periimplantatorio (días 4 y 8); en la expresión proteica y génica
de MMP-2 y MMP-9 en el tejido embrio-trofoblástico en el período postimplantatorio (día 8); en la
inducción de genotoxicidad y apoptosis embrio-trofoblástica en el período periimplantatorio (días
4 y 8) y en la inducción de genotoxicidad materna. -Capítulo III-
IV. Estudiar los efectos del consumo perigestacional de alcohol -previo a la gestación y hasta el día
4 (fase preimplantatoria), 8 (fase postimplantatoria temprana, gastrulación) o 10 de preñez (fase
postimplatatoria media, organogénesis temprana)- en el desempeño reproductivo y en el
desarrollo fetal y placentario en la gestación a término (día 18 de preñez). -Capítulo IV-


Materiales y métodos
87
�
MATERIALES
1. Equipamientos principales
Lupa binocular estereoscópica (Arcano), microscopio confocal (Olympus), microscopio de
fluorescencia (Axiophot Zeiss, Carl Zeiss, Inc. Oberkochen, Alemania), microscopio invertido
(Labomed), microscopio óptico de campo claro (Leica Microsystems, Wetzlar, Alemania);
microscopio óptico equipado con cámara de video (Axiophot Zeiss, Carl Zeiss, Inc. Oberkochen,
Alemania), termociclador Mastercycler ep (Eppendorf AG, Hamburgo, Alemania),
termociclador StepOnePlus™ Real-Time PCR Systems (Applied Biosystems, Foster City, CA,
EEUU).
2. Insumos
2.1. Reactivos varios.
Colorantes:
Alizarina red-S (MERCK, SP, Brasil), azul de toluidina (BIOPACK, Bs As, Argentina), Giemsa
(MERCK, Bs As, Argentina), hematoxilina de Harris (BIOPACK), hematoxilina de Mayer (MERCK),
eosina (BIOPACK), reactivo de Schiff (BIOPACK����
Sales:
Bicarbonato de sodio (NaHCO3), citrato de sodio dihidratado (NaH2(C3H5O(COO)3), cloruro de
calcio dihidratado (CaCl22H2O), cloruro de potasio (KCl), cloruro de sódio (NaCl), dextrosa
(C6H12O6), fosfato de potasio monobásico (KH2PO4 anhidro), fosfato de sodio dibásico
(PO4HNa2), fosfato de sodio monobásico (PO4HNa), sulfato de magnesio heptahidratado
(MgSO4 7H2O); (ANEDRA, Bs As, Argentina).
Solventes:
Acetona (ANEDRA), ácido acético glacial (SINTORGAN S.A., Bs As, Argentina), alcohol bencílico
(MERCK), alcohol etílico absoluto (BIOPACK), alcohol metílico (BIOPACK), cloroformo
(BIOPACK), éter etílico (MERCK), xilol (BIOPACK).

Materiales y métodos
88
�
Drogas, hormonas y otros reactivos:
Antibióticos (GIBCO-BRL, EEUU), Glicerol (SIGMA, St. Louis, EEUU), gonadotrofina coriónica
equina (eCG, Novormon, Bs As, Argentina), gonadotrofina coriónica humana (hCG, SIGMA),
líquido de montaje DPX (SIGMA), DNAsa (SIGMA), insulina (SIGMA), lactato (SIGMA), mitos (BD
Biosciences, San Diego, CA, EEUU), papaverina (Lab Monserrat y Eclair S.A., Bs As, Argentina),
parafina (Paraplast, Leica Microsystems, Bs As, Argentina), Paraformaldehído (ANEDRA),
peróxido de hidrogeno (SIGMA), piruvato (SIGMA), poli-L-lisina (SIGMA), proteinasa K (SIGMA),
seroalbumina bovina (BSA, SIGMA), solución de Formalina 37% (J.T. BAKER, México), suero de
cabra (Vector Labs, Burlingame, CA, EEUU), suero fetal bobino (SFB, GIBCO), sulfato de amonio
(MERCK), tritón X-100 (BIOPACK), vaselina líquida (SuFarma, Bs As, Argentina).
2.2. Anticuerpos, fluorocromos, kits comerciales y primers.
Anticuerpos y complejos:
Anticuerpo primario anti bromodeoxiuridina: mouse anti BrdU biotinilado (BD Biosciences),
anticuerpo primario anti E-cadherina: mouse anti E-Caderina (BD Biosciences), anticuerpo
primario anti metaloproteasa-2: goat anti MMP-2 (Santa Cruz Biotechnology, Inc.), anticuerpo
primario anti metaloproteasa-9: rabbit anti MMP-9 (Calbiochem, Darmstadt, Alemania),
anticuerpo primario anti zonula occudens-1: rabbit anti ZO-1-FITC (Zymed Labs, Inc. San
Francisco, CA, EEUU), anticuerpo secundario anti-IgG de cabra: rabbit anti goat biotinilado
(Vector Labs), anticuerpo secundario anti-IgG de conejo: goat anti rabbit biotinilado (Vector
Labs), anticuerpo secundario anti-IgG de ratón: donkey anti mouse-Rodamina (Millipore),
anticuerpo secundario anti-IgG de ratón: goat anti mouse-Alexa 488 (Vector Labs), anticuerpo
secundario: anti FITC biotinilado (Vector Labs), neutral avidine-FITC (Souther Labs),
streptoavidin horseradish peroxidase conjugate (HRP, Molecular Probe, Inc., Eugene, OR,
EEUU).
Fluorocromo:
Hoechst 33342 (SIGMA)
Kits comerciales:
Kit para determinación de alcoholemia (Roche), Kit de revelado 3,3'-diaminobencidina (DAB,
Cell Marque), ApopTag® Plus Peroxidase In Situ Apoptosis Detection Kit (Millipore), Power
SYBR® Green Cells-to-CtTM (Ambion® RNA by life technologies, Carlsbad, CA, EEUU).

Materiales y métodos
89
�
Primers:
Mus musculus MMP-2 (NCBI Ref. Seq. mRNA: NM_008610.2); Mus musculus MMP-9 (NCBI
Ref. Seq. mRNA: NM_013599.3); Mus musculus Ywhaz (NCBI Ref. Seq. mRNA:
NM_001253805.1) (Exxtend Biotecnologia Ltda., SP, Brasil).
2.3. Medios de incubación/cultivo.
Medio Dulbecco´s Modified Eagle´s Medium (D-MEM, SIGMA), medio Human Tubal Fluid (HTF)
con HEPES (GMHH-100, Life Global), medio HTF con gentamicina (Irvine Scientific, Santa Ana,
CA, EEUU), medio Krebs-Ringer bicarbonato salino con capacidad buffer: 11 mM C6H12O6, 145
mM Na+, 2,2 mM Ca2+, 1,2 mM Mg2+, 127 mM Cl-, 25 mM HCO3-, 1,2 mM SO4
2- y 1,2 mM PO43-.
3. Programas computacionales de análisis
3.1. Análisis de imágenes.
Adobe Photoshop CS2 (Adobe Systems Inc., San Jose, CA, EEUU), Image ProPlus (Media
Cybernetics, Inc., Rockville, MD, EEUU), Scion Image, versión 4.0.3.2 (Scion Corporation,
Frederick, MD, EEUU).
3.2. Análisis estadístico.
GraphPad InStat v2.05a (GraphPAD Software Inc., San Diego, CA, EEUU).

Materiales y métodos
90
�
MÉTODOS
1. Modelos animales
Se utilizaron ratones de la especie Mus musculus, sexualmente maduros (60 días de edad) con
un peso corporal promedio de 30 g (hembras) y 33 g (machos). Los animales de la colonia CF-1
se obtuvieron del Bioterio Central de la Facultad de Ciencias Exactas y Naturales (FCEN) de la
Universidad de Buenos Aires (UBA - Buenos Aires, Argentina); y los animales de la colonia CD-1
se obtuvieron del Bioterio del Instituto de Ciencias Biomédicas (ICB) de la Universidad de São
Paulo (USP - São Paulo, Brasil). Las hembras fueron alojadas en grupos de 5 animales por jaula
y los machos individualmente. Todos los animales fueron mantenidos bajo un régimen
constante de 12 hs de luz y 12 hs de oscuridad, y temperatura controlada de 22°C. Durante los
experimentos fueron alimentados ad libitum con alimento balanceado de origen comercial.
Los ratones de la colonia CF-1 recibieron alimento con un aporte calórico de entre 2.900 y
3.100 Kcal/kg de alimento (Balanceado Cooperación Rata-Ratón, Asociación Cooperativa de
Alimentos, S.A. Buenos Aires, Argentina). Los animales de la colonia CD-1 recibieron alimento
Nuvilab CR-1 con un aporte de 3.976 Kcal/kg de alimento (Nutrivital, Curitiba, Brasil).
2. Diseños experimentales: tratamientos in-vivo
Los experimentos fueron realizados según lo estipulado por la Guía para el Cuidado y Uso de
Animales de Laboratorio (8th Ed. 2010, NRC, EEUU), según Res CD 140/00 de la CICUAL-FCEN,
UBA para el Cuidado y Uso de animales de laboratorio, y según protocolos del Comité de Ética
de la Sociedad Brasilera de Ciencias de Animales de Laboratorio (SBCAL) aprobados por el
Comité de Ética para Animales de Experimentación (EAEC), USP, Brasil.
Se realizaron tres diseños experimentales diferentes, los cuales se detallan a continuación. En
cada grupo experimental y por tipo de estudio se utilizó un mínimo de 5 animales.
2.1. Tratamiento pregestacional con alcohol.
Hembras de la colonia murina CD-1 y CF-1 recibieron etanol 10% (m/v) en el agua de bebida ad
libitum durante un total de 17 días (hembras tratadas, HT). La solución de alcohol fue
renovada cada dos días a lo largo del período exposición. En paralelo, el grupo control
(hembras controles, HC) recibió sólo agua como bebida. La Figura 1, muestra de manera

Materiales y métodos
91
�
esquemática, el diseño experimental. Diariamente se determinó el grado de ingesta sólida
(gramos y calorías) y líquida (ml y calorías de la bebida con etanol), y el estadio del ciclo estral.
A finalizar el tratamiento, se determinó la variación del peso corporal, el nivel de alcohol en
sangre y la variación del ciclo estral, como se detalla en el Capítulo I.
Figura 1. Representación esquemática del diseño experimental. Tratamiento pregestacional con
alcohol. Hembras murinas CD-1 y CF-1 adultas recibieron etanol 10% en el agua de bebida durante
17 días (hembras tratadas, HT). Las hembras controles (HC) recibieron agua durante el mismo
período de tiempo. Al día 17 de tratamiento fueron sacrificadas por dislocación cervical para
continuar con los estudios detallados en el Capítulo I.
2.2. Tratamiento perigestacional con alcohol: efectos gestacionales al día 4, 5 u 8 de preñez.
2.2.1. Sin régimen de estimulación ovárica.
Un grupo de hembras CD-1 recibió etanol 10% (m/v) en el agua de bebida (hembras tratadas,
HT), siendo la solución de alcohol renovada cada dos días a lo largo de todo el tratamiento. El
grupo control (hembras controles, HC) recibió sólo agua como bebida. El tratamiento con
alcohol comenzó 15 días previos al apareamiento. Al día 15 de tratamiento, las HC y las HT
fueron alojadas con machos de la misma colonia aislados previamente, en relación 1:1, por
tres días consecutivos hasta lograr cópula positiva. Cada mañana posterior al apareamiento, la
cópula fue confirmada mediante la observación del tapón vaginal. Cuando el tapón vaginal no
fue detectado, tanto las HC como las HT fueron retiradas de las jaulas de apareamiento y

Materiales y métodos
92
�
colocadas en sus propias jaulas, hasta el fin de la tarde de ese mismo día en que fueron
alojadas con los machos nuevamente durante toda la noche. Así, las HT continuaron
recibiendo alcohol hasta el momento en que fueron colocadas nuevamente en las jaulas de
apareamiento. Aquellas hembras que no presentaron tapón vaginal luego de tres días
consecutivos de contacto con el macho fueron excluidas del experimento. Aquellas hembras
que sí presentaron tapón vaginal (día 1 de gestación) fueron separadas y continuaron
recibiendo alcohol (HT) o agua (HC) hasta el día 4 (D4), 5 (D5) u 8 (D8) de gestación. A la tarde
del día 4 de gestación (entre las 13:00 y 16:00 hs), a la mañana del día 5 o del día 8 de
gestación (entre las 9:00 y 12:00 hs), las HC y HT fueron sacrificadas por dislocación cervical
para extraer los embriones o los sitios de implantación y realizar los estudios correspondientes
al Capítulo II y III. La Figura 2, muestra de manera esquemática el diseño experimental
descripto.
Figura 2. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, sin régimen de estimulación ovárica. Hembras murinas CD-1 adultas recibieron etanol
10% en el agua de bebida durante 15-17 días previos al apareamiento (hembras tratadas, HT). Las
hembras controles (HC) recibieron agua durante todo el experimento. A partir del día 15 de
tratamiento y durante tres días consecutivos hasta lograr cópula positiva, las hembras fueron
alojadas con machos previamente aislados. Cada mañana fue observada la presencia de tapón
vaginal. Las hembras con tapón (día 1 de gestación) continuaron recibiendo alcohol (HT) o agua
(HC) hasta el día 4 (D4), 5 (D5) u 8 (D8) de gestación, momentos en que fueron sacrificadas por
dislocación cervical para realizar los estudios detallados en los Capítulos II y III.

Materiales y métodos
93
�
2.2.2. Con régimen de estimulación ovárica.
Un grupo de hembras CF-1 recibió etanol 10% (m/v) en el agua de bebida durante un total de
21 o 25 días (hembras tratadas, HT), siendo la solución de alcohol renovada cada dos días a lo
largo de todo el experimento. El grupo control (hembras controles, HC) recibió sólo agua como
bebida. A las 13:00 hs del día 15 de tratamiento cada hembra recibió por vía intraperitoneal 5
UI de gonadotrofina coriónica equina (eCG, del inglés, equine Chorionic Gonadotropin). A las
13:00 hs del día 17 de tratamiento (transcurridas 48 hs de la primera inyección), cada hembra
recibió por la misma vía, 5 UI de gonadotrofina coriónica humana (hCG, del inglés, human
Chorionic Gonadotropin). Inmediatamente después de la inyección de hCG, las hembras fueron
alojadas con machos de la misma colonia aislados previamente, en relación 1:1. En la mañana
posterior, la cópula fue confirmada mediante la observación del tapón vaginal. Aquellas
hembras que no presentaron tapón vaginal fueron excluidas del experimento. Aquellas
hembras que sí presentaron tapón vaginal (día 1 de gestación) fueron separadas y continuaron
recibiendo alcohol (HT) o agua (HC) hasta el día 4 (D4) u 8 (D8) de gestación. A la tarde del día
4 de gestación (entre las 13:00 y 16:00 hs, 96-99 hs post-hCG) o a la mañana del día 8 de
gestación (entre las 9:00 y 12:00 hs), tanto las HC como las HT fueron sacrificadas por
dislocación cervical para extraer los embriones o los sitios de implantación y realizar los
estudios que se describen en los Capítulos II y III. La Figura 3, muestra de manera esquemática
el diseño experimental descripto.

Materiales y métodos
94
�
Figura 3. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, con régimen de estimulación ovárica. Hembras murinas CF-1 adultas recibieron etanol
10% en el agua de bebida durante 17 días previos al apareamiento (hembras tratadas, HT). Las
hembras controles (HC) recibieron agua durante todo el experimento. Al día 15 y 17 de tratamiento
recibieron estimulación ovárica mediante inyección de eCG y hCG, y luego fueron alojadas con
machos previamente aislados. Al día siguiente fue confirmada la cópula observando la presencia de
tapón vaginal (día 1 de gestación, D1). Las hembras con tapón continuaron recibiendo alcohol (HT)
o agua (HC) hasta el día 4 (D4) u 8 (D8) de gestación, momentos en que fueron sacrificadas por
dislocación cervical para realizar los estudios detallados en los Capítulos II y III.
2.3. Tratamiento perigestacional con alcohol: efectos gestacionales al día 18 de preñez.
Un grupo de hembras CD-1 recibió etanol 10% (m/v) en el agua de bebida (hembras tratadas,
HT), siendo la solución de alcohol renovada cada dos días a lo largo de todo el tratamiento. El
grupo control (hembras controles, HC) recibió sólo agua como bebida. El tratamiento con
alcohol comenzó 15 días previos al apareamiento. Al día 15 de tratamiento, las HC y las HT
fueron alojadas con machos de la misma colonia aislados previamente, en relación 1:1, por
tres días consecutivos hasta lograr cópula positiva. Cada mañana posterior al apareamiento, la
cópula fue confirmada mediante la observación del tapón vaginal. Cuando el tapón vaginal no
fue detectado, tanto las HC como las HT fueron retiradas de las jaulas de apareamientos y
colocadas en sus propias jaulas, hasta el fin de la tarde de ese mismo día en que fueron
alojadas con el macho nuevamente durante toda la noche. Así, las HT continuaron recibiendo
alcohol hasta el momento en que fueron colocadas nuevamente en la jaula de apareamiento.
Aquellas hembras que no presentaron tapón vaginal luego de tres días consecutivos de
contacto con el macho fueron excluidas del experimento. Aquellas que sí presentaron tapón

Materiales y métodos
95
�
vaginal (día 1 de gestación) fueron separadas y continuaron recibiendo alcohol (HT) o agua
(HC) hasta el día 4 (D4), 8 (D8) o 10 (D10) de gestación. Transcurridos dichos períodos de
exposición, en los 3 grupos tratados, la bebida alcohólica fue discontinuada y reemplazada por
agua hasta el final del experimento al día 18 de preñez (gestación a término). Los grupos
controles recibieron agua durante los mismos períodos y hasta el día 18 de gestación. A la
mañana del día 18 (entre las 9:00 y 12:00 hs), tanto las HC como las HT fueron sacrificadas por
dislocación cervical para extraer los fetos y placentas para realizar los estudios
correspondientes al Capítulo IV. La Figura 4, muestra de manera esquemática el diseño
experimental descripto.
Figura 3. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol: efectos en la gestación a término. Hembras murinas CD-1 adultas recibieron etanol 10%
en el agua de bebida durante 15-17 días previos al apareamiento (hembras tratadas, HT). Las
hembras controles (HC) recibieron agua durante todo el experimento. A partir del día 15 y durante
tres días consecutivos hasta lograr cópula positiva, las hembras fueron alojadas con machos
previamente aislados. Cada mañana fue observada la presencia de tapón vaginal. Las hembras con
tapón (día 1 de gestación) continuaron recibiendo alcohol (HT) o agua (HC) hasta el día 4 (D4), 8
(D8) o 10 (D10) de gestación, momento en que la bebida alcohólica fue reemplazado por agua
hasta alcanzar el día 18 de preñez (D18). Al D18, las HC y HT fueron sacrificadas por dislocación
cervical para realizar los estudios detallados en el Capítulo IV.

Materiales y métodos
96
�
3. Estudios realizados como controles del tratamiento con alcohol.
3.1. Determinación de variaciones en parámetros biométricos.
En los grupos experimentales de ambas colonias, CF-1 y CD-1, se registró diariamente la
cantidad de alimento y bebida ingeridos por hembra. Para ello, se calculó la diferencia entre el
peso de alimento (g) ofrecido y el remanente no consumido, y la diferencia entre el volumen
(ml) de bebida ofrecida y el volumen final no consumido. Estos datos, junto con los valores de
aporte calórico de cada tipo de alimento y las calorías consumidas con la bebida alcohólica
(etanol 10% m/v), se utilizaron para calcular la ingesta calórica (Kcal) diaria y el porcentaje de
calorías derivadas del etanol (%EDC, del inglés, Ethanol Derived Calories). Se registró el peso de
los individuos, al inicio, durante y al término del tratamiento con alcohol.
3.2. Determinación de los niveles de alcohol en sangre.
Para determinar la concentración de alcohol en sangre (BAC, del inglés, Blood Alcohol
Concentration) o alcoholemia, un grupo de animales de cada protocolo experimental, fue
decapitado a las 6:00 hs am (horario en que finaliza el período de oscuridad) del último día de
tratamiento con alcohol. La sangre troncal fue recolectada en tubos Eppendorf heparinizados y
mantenida a 4oC hasta la realización del ensayo (máximo una semana). Las determinaciones
fueron realizadas utilizando un kit comercial, con el que se procedió según las indicaciones del
fabricante. Los resultados fueron expresados como mg etanol/dl sangre.


Capítulo I
97
�
Variaciones biométricas y del ciclo estral en dos modelos murinos
exocriados frente al consumo pregestacional de alcohol.
I.1 MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo.
1.1. Tratamiento pregestacional con alcohol.
Hembras de 60 días de edad de las colonias murinas CD-1 y CF-1, recibieron etanol 10% en el agua
de bebida por 17 días (Figura I.1). Durante el tratamiento se realizaron los estudios que se detallan
a continuación. Al día 17 de tratamiento, las hembras fueron sacrificadas y se determinó la
concentración de alcohol en sangre (BAC).
Figura I.1. Representación esquemática del diseño experimental. Tratamiento pregestacional con
alcohol. Hembras CD-1 y CF-1 recibieron etanol 10% durante 17 días (hembras tratadas, HT). Las
hembras controles (HC) recibieron agua durante el mismo período. Diariamente se analizaron
parámetros biométricos y el ciclo estral, y al día 17 se determinó el BAC.

Capítulo I
98
�
2. Metodologías empleadas.
2.1. Determinación de variaciones en parámetros biométricos y nivel de alcohol en sangre.
En las HT y HC de ambas colonias, CD-1 y CF-1, se registró diariamente la cantidad de alimento (en
gramos, g) y de bebida (en mililitros, ml) ingeridos por hembra, se calcularon las calorías de
alimento y líquido ingeridas (Kcal diarias) y el porcentaje de calorías derivadas del etanol (EDC) y
se registró el peso corporal de los individuos al inicio y al término del tratamiento. Luego de
sacrificar las hembras por dislocación cervical, los valores de alcoholemia (BAC) alcanzados el
último día de tratamiento fueron determinados (kit comercial) y los resultados se expresaron
como mg etanol/dl sangre.
2.2. Análisis del ciclo estral.
Con el fin de estudiar posibles alteraciones en el ciclo estral durante el período de alcoholización,
se procedió a analizar la citología vaginal mediante la coloración de Shorr. Para ello, desde el inicio
del tratamiento, diariamente y en el mismo horario de la mañana, se tomaron muestras de la
mucosa vaginal tanto de las HC como de las HT mediante un hisopado vaginal. Los extendidos
celulares obtenidos fueron fijados durante 10 minutos en alcohol 70% - éter etílico (1:1), y
posteriormente coloreados con hematoxilina de Harris seguida por el colorante de Shorr. Luego,
los extendidos fueron deshidratados en alcoholes de gradación creciente, diafanizados en xilol y
montados en un medio anhidro. Bajo microscopio óptico, se observaron los tipos celulares del
epitelio vaginal: células parabasales, intermedias, superficiales y queratinizadas (Figura I.2). Como
se muestra en la Tabla I.1, cada fase del ciclo se caracteriza por presentar una determinada
proporción de las células mencionadas. Por lo tanto, luego de registrar diariamente las
proporciones de los tipos celulares encontrados en el frotis vaginal, se estableció la fase o estado
del ciclo estral para cada hembra. Con estos datos, se calculó el número promedio de estados del
ciclo durante los 17 días de tratamiento, la duración promedio de cada estado (en días), el
porcentaje de hembras con alteraciones de la ciclicidad al término del tratamiento y el momento
(día de tratamiento) en el que se iniciaron alteraciones en la ciclicidad, vistas por la repetición del
mismo estado del ciclo. Para esto último se consideró ciclo estral alterado o irregular siempre que
un mismo estado se presentara más de tres días consecutivos.

Capítulo I
99
�
Figura I.2. Citología vaginal. Foto representativa de un frotis vaginal de ratón, coloreado según el
método de de Shorr, mostrando los distintos tipos celulares del epitelio vaginal. 1: célula parabasal, de
tamaño uniforme, forma redonda u oval, citoplasma pequeño de aspecto homogéneo y con bordes
lisos, núcleo redondo, central y relativamente grande ocupando casi todo el citoplasma; 2: célula
intermedia, representa la transición entre las parabasales y las superficiales, presentan un citoplasma
con bordes lisos y, tamaño y forma variable según el grado de maduración, usualmente son grandes y
poligonales, con núcleo redondo u oval, central y grande, aunque no ocupa gran parte del citoplasma;
3: células superficiales, representan la transición entre las intermedias y las queratinizadas, con forma
más o menos poligonal (según el grado de maduración) y núcleo redondo y pequeño; 4: células
queratinizadas, células poligonales con bordes angulosos y frecuentemente doblados, con citoplasma
altamente queratinizado, y núcleo ausente. Aumento 10x.
Tabla I.1. Proporción de tipos celulares según la fase del ciclo estral en el ratón.
Fase del Ciclo Estral
Tipos Celulares Proestro Estro Metaestro Diestro
Parabasales 30% 0% 10% 70%
Intermedias 60% 0% 30% 30%
Superficiales 5% 20% 30% 0%
Queratinizadas 5% 80% 30% 0%
Leucocitos 0% 0% Escasos Abundantes
Proporciones (expresadas en porcentaje, %) de los tipos celulares observados en un frotis vaginal de
ratón, en base a lo cual, se estableció diariamente la fase del ciclo estral para cada animal. En negrita
se destacan el/los tipos celulares predominantes en cada fase.

Capítulo I
100
�
3. Análisis estadístico.
Los valores expresados como promedios + SEM (del inglés, Standard Error of the Mean) o + SD (del
inglés, Standard Deviation), fueron comparados mediante la prueba T de Student. Los valores de
frecuencias, expresados como porcentajes, fueron comparados mediante la prueba exacta de
Fisher. Para el análisis estadístico se utilizó el programa estadístico GraphPad Instat software Inc.
v2.05a. Un valor de p<0,05 fue considerado estadísticamente significativo.

Capítulo I
101
�
I.2 RESULTADOS
En primera instancia nos propusimos evaluar si la ingesta de alcohol semicrónica, durante 17 días y
previo a la gestación, inducía alteraciones en algunos parámetros biométricos, así como en el ciclo
estral, con la finalidad de establecer las estrategias reproductivas más adecuadas para la
obtención de preñez en los dos modelos murinos de consumo pregestacional de alcohol utilizados,
CF-1 y CD-1.
Efectos del consumo pregestacional de alcohol en las variaciones de parámetros biométricos
y en el ciclo estral en dos modelos murinos.
1.1 Estudio de las variables biométricas en hembras murinas CF-1 y CD-1 durante el consumo
pregestacional de alcohol.
El objetivo planteado en primer lugar fue conocer el efecto del consumo de alcohol en los dos
colonias murinas CF-1 y CD-1 en cuanto al grado de ingesta líquida y sólida, calorías ingeridas y
cantidad de calorías derivadas de la bebida alcohólica y su posible influencia en el peso corporal,
parámetro considerado indicador del estado nutricional del animal.
Durante los 17 días de consumo de alcohol, las HT-CF-1 tuvieron una ingesta líquida
significativamente menor que el consumo registrado para las HC-CF-1 (p<0,001; Tabla I.2). En las
HT-CF-1, este consumo líquido representó la ingesta de 130,0 + 7,5 Kcal/kg/día. Cuando se analizó
la ingesta sólida de CF-1, no se encontraron diferencias significativas en el consumo diario de
alimento en las HT versus las HC y la ingesta de Kcal totales entre ambos grupos fue similar. En
este modelo, las HT consumieron diariamente 17,6% de EDC (Tabla I.2).
Las HT-CD-1 también mostraron una ingesta líquida significativamente menor respecto del
consumo registrado para las hembras controles, HC-CD-1 (p<0,01; Tabla I.2), representando una
ingesta de 125,9 + 21,7 Kcal liquidas/kg/día y un 13,6% de EDC (Tabla I.2). En esta colonia tampoco
se encontró una diferencia significativa en la ingesta sólida y la cantidad de calorías totales entre
las HT y las HC (Tabla I.2).

Capítulo I
102
�
La concentración de alcohol en sangre fue determinada tanto en las HT como en las HC. El valor
promedio de BAC luego de 17 días de consumo de alcohol fue de 24,5 + 5,0 mg/dl para las HT-CF-1
y 27,8 ± 4,1 mg/dl para las HT-CD-1. Como era esperado, en las HC dicho parámetro resultó nulo
(Tabla I.2). Para determinar si el consumo de alcohol afecta o modifica el peso corporal de las HT,
el mismo fue registrado al término del período de alcoholización, partiéndose de un peso corporal
inicial homogéneo entre todos los animales utilizados. No se encontraron diferencias significativas
entre los pesos de las HC y las HT después del consumo de alcohol (peso final), en los grupos de
hembras CF-1 y CD-1. A su vez, en ninguna de las colonias se encontraron diferencias significativas
entre el peso inicial y el final de las hembras controles y tratadas. Es decir, luego de 17 días de
tratamiento, no se registraron ganancias ni pérdidas de peso en las hembras tratadas (Tabla I.2).
Tabla I.2.�Variables biométricas durante 17 días de tratamiento con alcohol.
CF-1 CD-1
HC (13) HT (16) HC (5) HT (6)
Ingesta líquida
ml alcohol 10%/Kg/día 264,1 + 17,6 183,1 + 10,6 *** 244,3 + 45,6 177,4 + 30,7 **
ml alcohol 10%/ratón/día 7,5 + 0,8 5,1 + 0,3 7,5 + 1,4 5,2 + 0,9
ml alcohol/kg/día 0 23,3 0 22,4
ml alcohol/ratón/día 0 0,64 0 0,65
g alcohol/Kg/día 0 18,3 0 17,7
Kcal líquidas/kg/día 0 130,0 + 7,5 0 125,9 + 21,7
Ingesta sólida
g/Kg/día 264,1 + 28,1 212,7 + 21,3 241,0 + 55,3 201,3 + 61,4
g/ratón/día 7,5 + 0,5 5,7 + 0,6 7,4 + 1,7 5,9 + 1,8
Kcal sólidas/kg/día 792,2 + 84,3 638,1 + 63,9 958,2 + 219,8 800,4 + 244,1
Kcal totales
Kcal totales/Kg/día 792,2 + 84,3 768,1 + 71,4 958,2 + 219,8 926,3 + 265,7
% EDC 0 17,6 0 13,6
BAC (mg/dl) 0 24,5 + 5,0 0 27,8 ± 4,1
Peso corporal (g)
Inicial (D1) 28,5 + 1,2 27,1 + 0,8 30,4 + 2,5 29,3 + 2,8
Final (D17) 28,4 + 1,0 28,1 + 0,9 31,1 + 2,2 29,6 + 1,8

Capítulo I
103
�
Parámetros biométricos calculados a lo largo de 17 días de tratamiento en hembras controles (HC) y
tratadas con alcohol (HT), CF-1 y CD-1. Entre paréntesis se indica el número de hembras de cada
grupo. BAC: concentración de alcohol en sangre, D1: día 1 de tratamiento, D17: día 17 de tratamiento,
EDC: calorías derivadas del etanol. Los valores se expresan como promedios + SEM o + SD (pesos
corporales). **p<0,01 y ***p<0,001 versus su control (Prueba T de Student).
1.2. Estudio del ciclo estral en hembras murinas CF-1 y CD-1 durante el consumo pregestacional
de alcohol.
El estudio del ciclo estral durante el período de alcoholización fue fundamental para determinar el
estado reproductivo de las hembras, que en el caso del tratamiento pregestacional con alcohol
pudo resultar alterado, afectándose en consecuencia, el rendimiento reproductivo en cuanto a la
ocurrencia de apareamientos y preñez. Por lo tanto, surgió la necesidad de evaluar la ciclicidad de
las hembras tratadas en comparación con las hembras controles, para posteriormente decidir y
adoptar las estrategias reproductivas adecuadas para el logro de preñez.
Diariamente se realizó el registro de los estados del ciclo estral en las HC y HT de ambas colonias a
lo largo de 17 días de tratamiento. Se cuantificó el número de hembras que presentaron alguna
irregularidad o alteración en la ciclicidad cuando se presentaron estados persistentes o repetidos
en tres o más días consecutivos. Mientras que ambas colonias CF-1 y CD-1 tuvieron alrededor de
un 40% de hembras controles con ciclo irregular, la cantidad de hembras tratadas con ciclo
alterado, desde la primera semana y hasta la finalización del tratamiento, se incrementó a 80% en
CF-1 (p<0,05) y a 65% en CD-1 (Figura I.3).
Figura I.3. Porcentaje de hembras controles (HC) y tratadas con alcohol (HT) con alteraciones en el ciclo
estral a lo largo de 17 días de tratamiento para las colonias CF-1 y CD-1. *p<0,05 versus HC-CF-1 (Prueba
exacta de Fisher).
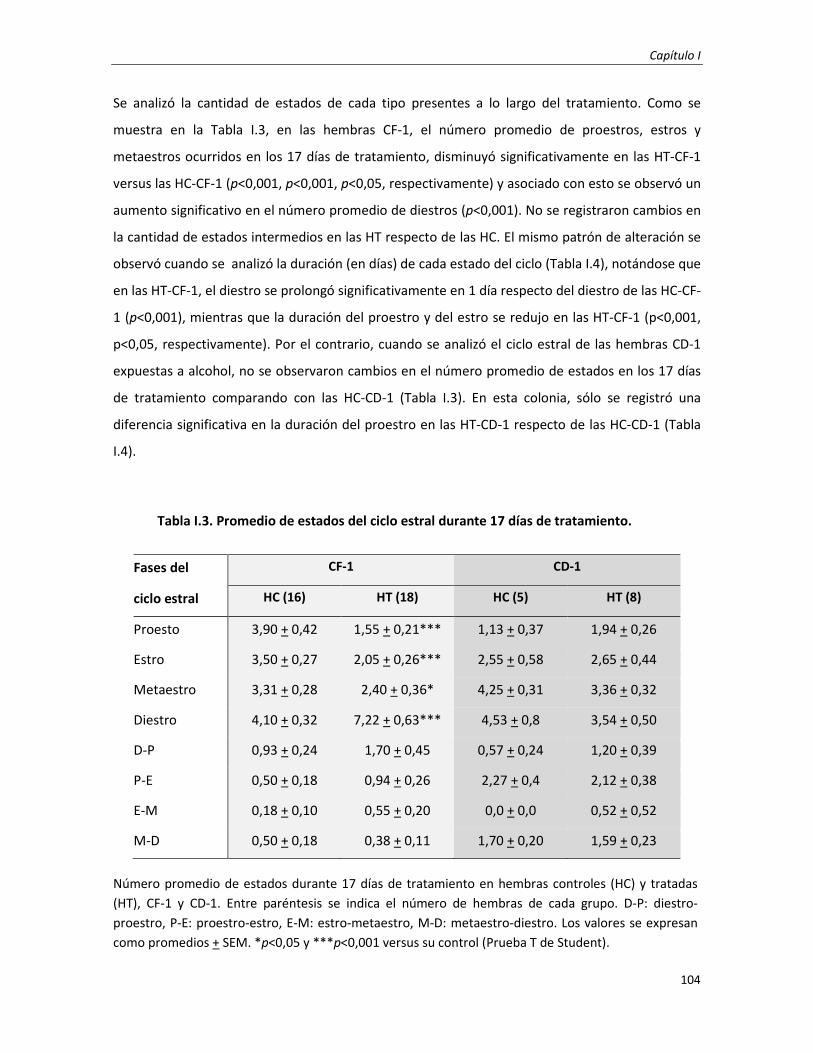
Capítulo I
104
�
Se analizó la cantidad de estados de cada tipo presentes a lo largo del tratamiento. Como se
muestra en la Tabla I.3, en las hembras CF-1, el número promedio de proestros, estros y
metaestros ocurridos en los 17 días de tratamiento, disminuyó significativamente en las HT-CF-1
versus las HC-CF-1 (p<0,001, p<0,001, p<0,05, respectivamente) y asociado con esto se observó un
aumento significativo en el número promedio de diestros (p<0,001). No se registraron cambios en
la cantidad de estados intermedios en las HT respecto de las HC. El mismo patrón de alteración se
observó cuando se analizó la duración (en días) de cada estado del ciclo (Tabla I.4), notándose que
en las HT-CF-1, el diestro se prolongó significativamente en 1 día respecto del diestro de las HC-CF-
1 (p<0,001), mientras que la duración del proestro y del estro se redujo en las HT-CF-1 (p<0,001,
p<0,05, respectivamente). Por el contrario, cuando se analizó el ciclo estral de las hembras CD-1
expuestas a alcohol, no se observaron cambios en el número promedio de estados en los 17 días
de tratamiento comparando con las HC-CD-1 (Tabla I.3). En esta colonia, sólo se registró una
diferencia significativa en la duración del proestro en las HT-CD-1 respecto de las HC-CD-1 (Tabla
I.4).
Tabla I.3. Promedio de estados del ciclo estral durante 17 días de tratamiento.
Fases del CF-1 CD-1
ciclo estral HC (16) HT (18) HC (5) HT (8)
Proesto 3,90 + 0,42 1,55 + 0,21*** 1,13 + 0,37 1,94 + 0,26
Estro 3,50 + 0,27 2,05 + 0,26*** 2,55 + 0,58 2,65 + 0,44
Metaestro 3,31 + 0,28 2,40 + 0,36* 4,25 + 0,31 3,36 + 0,32
Diestro 4,10 + 0,32 7,22 + 0,63*** 4,53 + 0,8 3,54 + 0,50
D-P 0,93 + 0,24 1,70 + 0,45 0,57 + 0,24 1,20 + 0,39
P-E 0,50 + 0,18 0,94 + 0,26 2,27 + 0,4 2,12 + 0,38
E-M 0,18 + 0,10 0,55 + 0,20 0,0 + 0,0 0,52 + 0,52
M-D 0,50 + 0,18 0,38 + 0,11 1,70 + 0,20 1,59 + 0,23
Número promedio de estados durante 17 días de tratamiento en hembras controles (HC) y tratadas
(HT), CF-1 y CD-1. Entre paréntesis se indica el número de hembras de cada grupo. D-P: diestro-
proestro, P-E: proestro-estro, E-M: estro-metaestro, M-D: metaestro-diestro. Los valores se expresan
como promedios + SEM. *p<0,05 y ***p<0,001 versus su control (Prueba T de Student).

Capítulo I
105
�
Tabla I.4. Duración promedio de cada estado del ciclo durante 17 días de tratamiento.
Fases del CF-1 CD-1
ciclo estral HC (16) HT (18) HC (5) HT (8)
Proesto 1,76 + 0,13 0,94 + 0,09*** 0,46 + 0,24 1,00 + 0,18*
Estro 1,36 + 0,10 1,07 + 0,08* 0,95 + 0,23 1,10 + 0,20
Metaestro 1,32 + 0,10 1,16 + 0,13 1,42 + 0,18 1,56 + 0,29
Diestro 1,47 + 0,08 2,56 + 0,27*** 1,71 + 0,76 1,69 + 0,49
Duración (en días) de cada estado del ciclo estral durante 17 días de tratamiento, en hembras
controles (HC) y tratadas (HT), CF-1 y CD-1. Entre paréntesis se indica el número de hembras de cada
grupo. Los valores se expresan como promedios + SEM. *p<0,05 y ***p<0,001 versus su control
(Prueba T de Student).

Capítulo I
106
�
I.3. CONCLUSIONES PARCIALES
El estudio de las variables biométricas es fundamental para la evaluación de la respuesta
fisiológica de las dos colonias murinas utilizadas frente al ingesta de alcohol, y así poder
monitorear la aparición de un posible factor de desnutrición asociado al consumo. Si así fuera,
dicho factor podría interferir con el/los efectos directos y propios del etanol sobre los parámetros
reproductivos en estudio. En tal sentido, se esperaba que el consumo pregestacional semicrónico
de etanol 10% durante un período de 17 días no modificara el consumo de calorías totales diarias
ni provocara alteraciones en el peso corporal de los animales tratados, descartándose entonces un
factor nutricional que pudiera interferir con los efectos de la exposición. A su vez, la aparición de
cambios o alteraciones en el ciclo estral luego del consumo de alcohol, podrían implicar un menor
rendimiento reproductivo, por un comportamiento poco receptivo de la hembra hacia el macho,
lo que llevaría a la disminución de la tasa de preñez. Según este resultado se decidió implementar
la estrategia reproductiva más adecuada para obtener preñez efectiva y sincronizada en las
hembras tratadas y controles.
1. En cuanto al efecto de la ingesta pregestacional de alcohol en variables biométricas de
hembras CF-1 y CD-1, se puede concluir que:
El consumo pregestacional de etanol 10%, si bien provocó una disminución en el grado de ingesta
líquida tanto en CF-1 como en CD-1, respecto de sus controles, no modificó la ingesta de calorías
totales ni los pesos corporales al final del tratamiento. En ambos grupos de hembras tratadas el
porcentaje de EDC fue similar y del orden de 18 y 14 % en CF-1 y CD-1, respectivamente.
Se puede confirmar que el tratamiento con alcohol no genera en las hembras tratadas un efecto
nutricional negativo respecto al peso de las hembras expuestas. Estos resultados, a su vez, avalan
la utilización de agua sola, sin suplemento glucosado, como bebida de las hembras controles.

Capítulo I
107
�
2. En cuanto a los cambios en el ciclo estral de hembras CF-1 y CD-1 durante el consumo
pregestacional de alcohol, se puede concluir que:
El ciclo estral de las hembras CF-1 tratadas presentó alteraciones en su ciclicidad, lo que fue
evidenciado por una alta proporción de diestros, registrados durante varios días consecutivos a lo
largo del período de tratamiento. Finalizado el período de consumo, se registró un 80% de
hembras con ciclo detenido en diestro desde la primera semana de tratamiento.
El ciclo estral de las hembras CD-1 tratadas, si bien presentó una variación en la duración del
proestro respecto de las controles, no se modificó ni la cantidad ni la duración de estros y diestros
a lo largo del tratamiento. Por otro lado, el porcentaje de hembras tratadas con alteraciones en el
ciclo no difirió significativamente del porcentaje de hembras controles.
Estos resultados fueron decisivos para establecer el esquema de apareamiento más adecuado a ser
aplicado, ya sea basado en el ciclo natural o en un ciclo inducido. Por lo tanto, dada la respuesta
diferencial de las colonias frente al tratamiento con alcohol, se decidió realizar dos diseños
experimentales diferentes para optimizar el rendimiento reproductivo en cada modelo. Para la
colonia CF-1, se optó por un esquema que involucró la intervención hormonal con gonadotrofinas
(modelo con estimulación ovárica), mientras que para la colonia CD-1, se optó por un esquema
basado en el ciclo natural (modelo sin estimulación ovárica).


Capítulo II�
108
�
Consumo perigestacional de alcohol: efectos en la morfogénesis y en el
crecimiento embrio-trofoblástico periimplantatorio murino.
II.1. MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo.
Se utilizaron hembras y machos adultos pertenecientes a las colonias CD-1 y CF-1. Según los
resultados obtenidos luego del tratamiento pregestacional (véase Capítulo I), para realizar el
tratamiento perigestacional, fueron planteados dos esquemas experimentales diferentes para
cada colonia (véase Materiales y métodos). Brevemente:
1.1. Tratamiento perigestacional con alcohol, sin régimen de estimulación ovárica.
Hembras CD-1 recibieron etanol 10% (m/v) en el agua de bebida 15 días previos al apareamiento y
hasta el día 4, 5 u 8 de gestación (hembras tratadas, HT). El grupo control (hembras controles, HC)
recibió sólo agua como bebida. Como no se detectaron alteraciones significativas en la regularidad
del ciclo estral (véase Capítulo I) que justificaran una intervención hormonal, como estrategia
reproductiva se optó por el ciclo natural. Las hembras fueron alojadas con machos para el
apareamiento entre los días 15 y 17 de tratamiento. Al día siguiente, la observación del tapón
vaginal indicó el día 1 de gestación. Las hembras con tapón vaginal continuaron recibiendo alcohol
(HT) o agua (HC) hasta el día 4, 5 u 8 de preñez. En la tarde del día 4 de gestación (entre las 13:00 y
16:00 hs), en la mañana (entre las 9:00 y 12:00 hs) del día 5 o del día 8 de gestación, las HC y HT
fueron sacrificadas por dislocación cervical para extraer los embriones o sitios de implantación
para realizar los estudios que se detallan a continuación (Figura II.1).�

Capítulo II�
109
�
Figura II.1. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, sin estimulación ovárica. Hembras CD-1 recibieron etanol 10% durante (hembras tratadas, HT)
15 días previos al apareamiento. Las hembras controles (HC) recibieron agua durante el mismo
período. Las hembras fueron puestas en contacto con el macho en días consecutivos y hasta 3 veces,
entre los días 15 y 17 de tratamiento. Cada mañana fue observada la presencia del tapón vaginal. Las
hembras con tapón (día 1 de gestación) continuaron recibiendo alcohol (HT) o agua (HC) hasta el día 4
(D4), 5 (D5) u 8 (D8) de gestación, días en que fueron sacrificadas para realizar los estudios indicados
en el esquema.
1.2. Tratamiento perigestacional con alcohol, con régimen de estimulación ovárica.
Hembras CF-1 recibieron etanol 10% (m/v) en el agua de bebida 17 días previos al apareamiento y
hasta el día 4 u 8 de gestación (hembras tratadas, HT). El grupo control (hembras controles, HC)
recibió sólo agua como bebida. Dado que la ingesta de alcohol durante 17 días indujo alteraciones

Capítulo II�
110
�
en el ciclo estral (en el 80% de las HT, véase Capítulo I), como estrategia reproductiva se recurrió a
un esquema de estimulación ovárica con eCG y hCG (aplicadas los días 15 y 17 de tratamiento,
respectivamente), para inducir la ovulación tanto en las HT como en las HC. Al día 17 de
tratamiento, las hembras fueron alojadas con machos para el apareamiento, y la observación del
tapón vaginal al día siguiente indicó el día 1 de gestación. Las hembras con tapón vaginal
continuaron recibiendo alcohol (HT) o agua (HC) hasta el día 4 u 8 de preñez. En la tarde del día 4
de gestación (entre las 13:00 y 16:00 hs, 96-99 hs post-hCG) o en la mañana del día 8 de gestación
(entre las 9:00 y 12:00 hs), las HC y HT fueron sacrificadas por dislocación cervical para extraer los
embriones o sitios de implantación para realizar los estudios que se detallan a continuación
(Figura II.2).
Figura II.2. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, con estimulación ovárica. Hembras CF-1 recibieron etanol 10% (hembras tratadas, HT)
durante 15 días previos al apareamiento. Las hembras controles (HC) recibieron agua el mismo
período. Al día 15 y 17 de tratamiento se indujo la ovulación con eCG y hCG, y luego de apareamiento,
la presencia de tapón vaginal indicó el día 1 de gestación (D1). Las hembras con tapón continuaron
recibiendo alcohol (HT) o agua (HC) hasta el día 4 (D4) u 8 (D8) de gestación, días en que fueron
sacrificadas para realizar los estudios que indicados en el esquema.

Capítulo II�
111
�
2. Metodologías empleadas.
2.1. Metodologías para el estudio del desarrollo embrionario de preimplantación:
Día 4 de gestación.
2.1.1. Recolección y procesamiento de embriones de preimplantación.
A la tarde del 4to. día de gestación, las hembras de las colonias CD-1 y CF-1 fueron sacrificadas, la
cavidad abdominal fue cuidadosamente abierta, y el útero rápidamente removido y colocado en
una placa de Petri con medio Krebs-Ringer. Los cuernos uterinos fueron separados de los
oviductos y del cérvix, y luego los embriones fueron extraídos por flushing del útero con medio
medio Krebs-Ringer - BSA 0,3% utilizando aguja de 25 G acoplada a una jeringa de 1 ml. Los
embriones fueron recolectados del líquido de flushing utilizando un microcapilar de vidrio de
diámetro adaptado al tamaño del embrión, acoplado a una pipeta bucal. Los embriones
recuperados fueron lavados sucesivas veces en microgotas de medio medio Krebs-Ringer - BSA
0,3%, fueron observados bajo microscopio invertido con contraste de fase y fotografiados para su
posterior clasificación según estadios embrionarios y análisis de la calidad morfológica.
Posteriormente, fueron procesados según la técnica descripta por Tarkowski para análisis de
crecimiento embrionario e índice mitótico.
2.1.2. Monitoreo de la preñez.
Con el fin de evaluar el desempeño reproductivo de HC y HT de ambas colonias al día 4 de
gestación, se registró el número total de embriones recuperados por hembra luego del flushing
uterino.
2.1.3. Determinación de la diferenciación embrionaria in-vivo.
Para evaluar los estadios de diferenciación en la fase de preimplantación, los embriones
recuperados de cada hembra fueron observados bajo microscopio invertido con contraste de fase
para ser clasificados en los estadios de diferenciación según la escala de Theiler (TS, Theiler Scale)

Capítulo II�
112
�
(Theiler, 1989; Kaufman y Bard, 1999). Los estadios embrionarios al día 4 de gestación incluyeron:
mórulas pre-compactadas y compactadas, blastocistos iniciales (con cavidad blastocélica en
formación ocupando menos del 50% del volumen del embrión), blastocistos expandidos (con
cavidad blastocélica completamente expandida) y blastocistos extruidos de la zona pelúcida
(Figura II.3, Tabla II.1). Se registró el número total de embriones en cada estadio en el total de
hembras utilizadas en cada grupo, y se calculó la frecuencia de cada tipo como porcentaje de cada
estadio sobre el total de embriones recuperados.
2.1.4. Determinación de la viabilidad y evaluación de la morfología embrionaria.
En primer lugar, se determinó la viabilidad de los embriones recuperados luego del flushing.
Aquellos embriones que presentaron un 50% o más del volumen embrionario con fragmentos y/o
signos de lisis celular fueron clasificados como inviables (Figura II.3). De esta manera, el número
total de embriones viables por hembra, así como, el número de embriones viables en el total de
hembras utilizadas fue contabilizado.
Aquellos embriones clasificados como viables, fueron observados en detalle para la evaluación de
la morfología embrionaria. Siempre que se observaron las siguientes características, los embriones
fueron clasificados como anormales: blastómeras fragmentadas y/o lisadas (ocupando entre un 10
y un 50% del volumen embrionario total), fragmentos citoplasmáticos en el espacio perivitelino,
vacuolas intracitoplasmáticas, blastómeras de tamaños variables, persistencia de blastómeras sin
compactar en mórulas compactadas, cavidad blastocélica anormal (por ejemplo: cavidades
tabicadas). El número de embriones morfológicamente anormales sobre el total de embriones
analizados fue registrado y las distintas anomalías embrionarias fueron fotografiadas en
microscopio invertido con contraste de fase.

Capítulo II�
113
�
Figura II.3. Diferenciación embrionaria in-vivo al día 4 de gestación. Micrografías representativas de
cada tipo embrionario. A, mórula pre-compactada; B, mórula compactada; C-D blastocistos iniciales
con distinto grado de expansión de la cavidad blastocélica; E, blastocisto expandido; F, blastocisto
extruido; G-H, embriones inviables: embrión fragmentado (G) y lisado (H). Aumento: 20x.
2.1.5. Determinación del grado de crecimiento embrionario e índice mitótico.
En las mórulas compactadas, blastocistos iniciales y blastocistos expandidos, el grado de
crecimiento embrionario, así como el índice mitótico fue determinado mediante la técnica de

Capítulo II�
114
�
Tarkowski (1966). Para ello, los embriones fueron incubados en gotas de una solución de citrato de
sodio 1% en agua destilada por 20 min (hasta evidenciar el hinchamiento de los embriones y/o
desaparición de la cavidad en el caso de los blastocistos). Luego fueron transferidos a portaobjetos
limpios y fueron fijados con metanol - ácido acético (3:1 v/v) mediante goteo directo sobre los
embriones, para provocar la ruptura de las membranas celulares, la dispersión del citoplasma, la
adhesión y fijación de los núcleos en el portaobjeto. Finalmente, luego del secado al aire, las
muestras fueron coloreadas con una solución de Giemsa en agua destilada (1:10) por 10 a 15 min,
lavadas en agua destilada y secadas a temperatura ambiente para su posterior observación en
microscopio óptico. El número de núcleos por tipo de embrión, indicador del grado de crecimiento
embrionario, fue cuantificado y expresado como número promedio de núcleos por embrión + SD.
Este recuento fue realizado en cada tipo embrionario previamente clasificado en cuanto a su
estadio, para determinar la correspondencia entre el grado de diferenciación embrionaria y el
rango numérico de núcleos por embrión, según las referencias que se indican en la Tabla II.1. El
índice mitótico (IM) fue calculado mediante la cuantificación del número de núcleos en división
mitótica (núcleos en profase, metafase, anafase y telofase) sobre el total de núcleos registrados en
el total de embriones de cada tipo.
Tabla II.1. Desarrollo embrionario de preimplantación. Estadios embrionarios.
Tipo de Embrión DG (hs post-hCG) Estadio según TS N° células/embr Ciclo celular
Embriones de 1 a 2 céls DG 1 (12-32 hs) TS 1-2 1 a 2 1er. Ciclo
Embriones de 2 a 4 céls DG 2 (48-50 hs) TS 2-3 2 a 4 2do. Ciclo
Embriones de 4 a 8 céls DG 3 (58-60 hs) TS 3 4 a 8 3er. Ciclo
Mórulas pre- y en
compactación DG 3 (60-72 hs) TS 3 8 a 16 céls 4to. Ciclo
Mórulas compactadas DG 3,5 (~84 hs) TS 3 16 a 32 céls 5to. Ciclo
Blastocistos iniciales DG 4 (90-98 hs) TS 4 32 a 64 céls 6to. Ciclo
Blastocistos expandidos DG 4 (98-110 hs) TS 4 64 a 128 céls 7mo. Ciclo
Blastocistos extruidos DG 5 (110-120 hs) TS 5 Aprox. 150 céls 8vo. ciclo

Capítulo II�
115
�
Según datos extraídos de literatura (Pratt, 1987; Theiler, 1989), se lista la secuencia del desarrollo
embrionario de preimplantación in-vivo (días de gestación -DG- siendo DG=1 el día de la observación
del tapón vaginal), según aspectos morfológicos y de crecimiento (en términos del número de células
por embrión y ciclo celular). Céls: células, hs post-hCG: hs de extracción embrionaria post aplicación de
hCG, N° células/embr: número de células por embrión, TS: escala de Theiler.
2.2. Metodologías para el estudio del desarrollo embrio-trofoblástico durante la implantación:
Día 5 de gestación.
2.2.1. Recolección y procesamiento de embriones implantativos.
A la mañana del 5to. día de gestación, las hembras de la colonia CD-1 fueron sacrificadas, la
cavidad abdominal fue cuidadosamente abierta, y el útero fue rápidamente removido y colocado
en una placa de Petri con buffer fosfato salino (PBS) estéril a 37°C. Los cuernos uterinos fueron
separados de su oviducto y del cérvix. Luego los embriones fueron extraídos por flushing del útero
con PBS suplementado con suero fetal bovino (SFB) 10% precalentado a 37°C utilizando aguja de
25 G acoplada a una jeringa de 1 ml. Los embriones fueron recolectados del líquido de flushing
usando un microcapilar de vidrio de diámetro adaptado al tamaño de los blastocistos, acoplado a
una pipeta bucal. Luego de sucesivos lavados mediante pasajes en microgotas de PBS-SFB estéril a
37°C, los embriones fueron transferidos a gotas de medio HTF (Human Tubal Fluid) estéril
suplementado con SFB 10%, cubiertas con aceite mineral. Bajo estereomicroscopio, rápidamente
fue registrado el número total de embriones y luego, bajo microscopio invertido con contraste de
fase, fueron observados y fotografiados para posterior evaluación del grado de diferenciación y
morfología. Los blastocistos implantativos (embriones extruidos de la zona pelúcida) fueron
transferidos, bajo flujo laminar, al sistema de cultivo para estudiar el proceso de implantación
(implantación in-vitro), como se detallará luego.
2.2.2. Monitoreo de la preñez.
Con el fin de evaluar el desempeño reproductivo de las HC y las HT al día 5 de gestación, se
registró el número total de embriones recuperados por hembra luego del flushing uterino.

Capítulo II�
116
�
2.2.3. Determinación de la diferenciación embrionaria implantativa in-vivo.
Los embriones implantativos fueron clasificados según su estadio de diferenciación en tres
categorías diferentes: embriones implantativos tipo 1, tipo 2 o tipo 3. Esta clasificación se basó en
el aspecto morfológico de los embriones, teniendo en cuenta la presencia o no de la cavidad
blastocélica, de procesos trofoectodérmicos (expansión inicial del trofoectodermo) y de células
gigantes trofoblásticas (CGTs) diferenciadas (Figura II.4; Tabla II.2). De acuerdo a esta clasificación,
el embrión más avanzado en su desarrollo corresponde al tipo 3. El número total de embriones de
cada tipo fue registrado y expresado como porcentaje del número total de embriones recuperados
sobre el total de hembras utilizadas en cada grupo.
Figura II.4. Diferenciación embrionaria in-vivo al día 5 de gestación. Micrografías representativas de
cada tipo embrionario. A, Embrión tipo 1: blastocisto implantativo temprano sin zona pelúcida, con
cavidad (*) y sin procesos trofoectodérmicos, donde se distingue claramente el MCI, el TP y el TM; B,
Embrión tipo 2: blastocisto implantativo con cavidad blastocélica reducida (*) e incipiente desarrollo de
procesos trofoblásticos (puntas de flechas). Aún se distingue el MCI; C, Embrión tipo 3: blastocisto
implantativo en estadio más avanzado, sin cavidad y desarrollo de procesos trofoblásticos (puntas de
flecha). MCI: macizo celular interno, TM: trofoectodermo mural, TP: trofoectodermo polar. Aumento:
20x.

Capítulo II�
117
�
Estudios in-vitro: capacidad implantativa.
2.2.4. Cultivo de embriones implantativos.
Por medio de la técnica de implantación in-vitro se evaluó la diferenciación embrionaria y el
crecimiento embrio-trofoblástico durante la implantación, así como la expansión del trofoblasto
para monitorear la capacidad implantativa y el grado de invasividad de las células trofoblásticas.
Los embriones implantativos extraídos por flushing uterino de 7 HC (n=50) y 8 HT (n=45) en la
mañana del día 5 de gestación, fueron colocados en cultivo. Para ello, los embriones fueron
cuidadosamente depositados sobre cubreobjetos de vidrio estériles ubicados en placas de 24
pocillos (3 a 5 embriones por pocillo) con 500 µl de medio D-MEM estéril (Dulbecco´s Modified
Eagle´s Medium), suplementado con SFB 10%, 100 UI/ml de penicilina, 100 µg/ml de
estreptomicina, 0,5% de aminoácidos no esenciales, 0,4 µg/ml de insulina, 520 µg/ml L(+) de
ácido láctico, 56 �g/ml de piruvato de sodio y 4 mg/ml de BSA. El cultivo de embriones fue
realizado a 36,5°C, con 5% CO2 en atmósfera húmeda, por 72 hs.
2.2.5. Determinación de la diferenciación embrionaria durante el cultivo.
Los embriones fueron fotografiados en microscopio con contraste de fase cada 24 hs, en el mismo
horario de la mañana y hasta cumplirse las 72 hs de cultivo, con el fin de analizar los estadios de
diferenciación embrionaria in-vitro de acuerdo con el aspecto morfológico de los embriones. Para
la clasificación de los tipos embrionarios se tomó en consideración el grado de desarrollo de las
células trofoblásticas y la presencia de CGTs, estableciéndose así, tres categorías de diferenciación
para las 24, 48 y 72 hs de cultivo: embriones tipo 1, tipo 2 y tipo 3 (Tabla II.2). De acuerdo a esta
clasificación, el embrión con desarrollo avanzado corresponde al tipo 3. El número total de
embriones de cada tipo fue registrado y expresado como porcentaje para cada tiempo de cultivo.
2.2.6. Evaluación de la morfología embrionaria implantativa.
Todos los embriones implantativos recuperados al día 5 de gestación (T0hs) y durante el cultivo
(T24 - 72hs), fueron observados para la evaluación de la morfología embrionaria. Siempre que se
observaron las siguientes características, los embriones fueron clasificados como anormales:

Capítulo II�
118
�
presencia de fragmentos citoplasmáticos, vacuolas intracitoplasmáticas, blastómeras con signos
de lisis, y en el caso de persistencia de la cavidad se observaron signos de cavitación anormal (dos
o más cavidades blastocélicas, tabiques). El número de embriones morfológicamente anormales
fue registrado y expresado como porcentaje respecto del total de los embriones analizados.
2.2.7. Determinación del grado de crecimiento de embriones implantativos.
Para analizar el crecimiento embrio-trofoblástico al día 5 de gestación, así como durante el cultivo
(grado de expansión embrio-trofoblástica), el área total del embrión (en µm2) fue medida
mediante el programa Image Pro Plus en micrografías obtenidas de microscopia invertida con
contraste de fase. Los datos de las áreas fueron agrupados en tres rangos numéricos y finalmente,
los embriones de cada tiempo de cultivo (D5 in-vivo o T0 hs de cultivo, T24 y T48 hs) fueron
clasificados dentro de estas tres categorías. Los tipos embrionarios fueron: para T0 hs: embriones
pequeños con áreas entre 4.500-7.400 µm2; embriones medianos con 7.401-10.300 µm2 y
embriones grandes con áreas entre 10.301-13.200 µm2. Para T24 hs: embriones pequeños con
áreas entre 5.400-22.930 µm2; embriones medianos con áreas entre 22.931-40.460 µm2 y
embriones grandes con 40.461-58.000 µm2 de área. Para T48 hs: embriones pequeños con 5.500-
26.600 µm2 de área, embriones medianos con áreas entre 26.601-47.700 µm2 y embriones grandes
con áreas entre 47.701-69.000 µm2. Se calculó el área promedio por embrión + SD para cada
categoría de tamaño embrionario.

Capítulo II�
119
�
Tabla II.2. Estadios de diferenciación según morfología embrionaria y por tiempo de cultivo.
Tiempo de
Cultivo (T) Embrión tipo 1 Embrión tipo 2 Embrión tipo 3
T0 hs (Día 5)
Con cavidad blastocélica
expandida
Sin desarrollo de
procesos trofoblásticos
Con cavidad
blastocélica reducida
Incipiente desarrollo
trofoblástico
Sin cavidad blastocélica
Incipiente desarrollo
trofoblástico
T24 hs
Sin/escaso desarrollo de
procesos trofoblásticos
Sin CGTs
Escaso desarrollo de
procesos trofoblásticos
Con CGTs
Moderado desarrollo
trofoblástico
Con CGTs
T48 hs
Sin/escaso desarrollo de
procesos trofoblásticos
Con CGTs
Escaso/moderado
desarrollo de procesos
trofoblásticos
Con CGTs
Moderado desarrollo
de procesos
trofoblásticos
Con CGTs
T72 hs
Moderado desarrollo de
procesos trofoblásticos
Con CGTs
Moderado/alto
desarrollo de procesos
trofoblásticos
Con CGTs
Alto desarrollo de
procesos trofoblásticos
Con CGTs
Los embriones observados al inicio del cultivo (T0 hs, correspondiente al día 5 de gestación) y cada 24
hs (T24 hs, T48 hs y T72 hs), fueron clasificados en embriones tipo 1, tipo 2, tipo 3 de acuerdo a la
presencia o no de cavidad blastocélica, procesos trofoblásticos y células gigantes del trofoblasto (CGTs)
diferenciadas.

Capítulo II�
120
�
2.3. Metodologías para el estudio del desarrollo embrio-trofoblástico durante la fase de
gastrulación: Día 8 de gestación.
2.3.1. Recolección y procesamiento de sitios de implantación.
Al 8vo. día de gestación, las hembras de las colonias CD-1 y CF-1 fueron sacrificadas y la cavidad
abdominal fue cuidadosamente abierta para extraer los cuernos uterinos. Con el fin de inhibir las
contracciones uterinas espontáneas del miometrio, que pudieran alterar la simetría y forma del
sitio de implantación, se colocó inmediatamente Papaverina (antiespasmódico de uso comercial)
sobre los cuernos uterinos, y luego de unos minutos los mismos fueron removidos de la cavidad
abdominal, lavados en medio Krebs-Ringer y transferidos a una placa de parafina donde, mediante
cortes con bisturí perpendiculares al eje longitudinal del útero, fueron delicadamente aislados los
sitios de implantación. Inmediatamente, los sitios fueron colocados en solución fijadora y,
posteriormente, procesados histológicamente para analizar el grado de diferenciación
embrionaria y realizar estudios histológicos, como se detallará a continuación.
2.3.2. Monitoreo de la preñez.
Con el fin de evaluar el desempeño reproductivo de las HC y las HT al día 8 de gestación, se
registró el número total de sitios de implantación y se calculó el número promedio por hembra.
2.3.3. Procesamiento histológico de los sitios de implantación.
Los sitios de implantación extraídos fueron fijados en paraformaldehído 4% por 18 hs a 4°C. Luego
de la fijación, fueron lavados en agua corriente y deshidratados mediante pasajes en alcoholes de
gradaciones crecientes. Posteriormente, fueron aclarados en xilol e incluidos en parafina. Con el
fin de obtener secciones transversales al eje longitudinal uterino, los sitios de implantación fueron
cuidadosamente orientados al momento de la realización de los tacos de parafina. Mediante corte
con micrótomo, se realizaron secciones de 5 µm de espesor y en forma seriada de todo el sitio de
implantación hasta llegar al embrión. Para seleccionar y conservar los cortes más centrales del
embrión, se realizó una coloración rápida de algunos cortes parafinados con azul de toluidina 1%

Capítulo II�
121
�
en agua destilada, los que fueron observados bajo microscopio óptico. De cada sitio de
implantación cortado, sólo fueron conservadas aquellas secciones correspondientes a la región
central del embrión, las que fueron montadas en portaobjetos limpios recubiertos con Poli-L-lisina
y secados por 30 min a 60°C. Posteriormente, se realizó una coloración topográfica con
hematoxilina-eosina, para lo cual los preparados fueron desparafinados en xilol, rehidratados
progresivamente en alcoholes de gradaciones decrecientes, lavados en agua destilada y
coloreados con hematoxilina de Mayer por 1 a 3 min. Durante 10 min se realizó el viraje en agua
corriente, y luego se continuó con eosina alcohólica por 30 seg. Después de los correspondientes
lavados fue realizado el montaje de los preparados en medio anhidro (DPX), previa deshidratación
de los cortes en alcoholes y diafanización en xilol. En todas las muestras histológicas de sitios de
implantación se analizó la diferenciación embrionaria y la calidad histológica del embrión, de los
tejidos extraembrionarios, de la interfase materno-trofoblástica y del cono ectoplacentario (Figura
II.5).
Figura II.5. Sitio de implantación al día 8 de gestación. Micrografía representativa de un sitio de
implantación incluido en parafina coloreado con hematoxilina-eosina. A, sitio de implantación; B,
detalle del embrión. CE: cono ectoplacentario, DAM: decidua antimesometrial, DM: decidua
mesometrial, E: embrión en gastrulación, InT-DA/DL: interfase trofoblástico-decidual
antimesometrial/lateral, ZE: zona de expansión trofoblástica del CE, ZP: zona de proliferación
trofoblástica del CE. A: aumento 2,5x; B: escala 100 µm.

Capítulo II�
122
�
2.3.4. Determinación del grado de diferenciación embrionaria en el sitio de implantación.
Con el fin de analizar el grado de diferenciación embrionaria luego de la exposición a alcohol hasta
el día 8 de gestación, en promedio dos sitios de implantación por hembra separados al azar fueron
histológicamente procesados. El estadio de diferenciación del concepto (embrión y membranas
extraembrionarias) fue determinado de acuerdo a la escala de Theiler (TS), según la presencia o no
de las cavidades embrionarias, el desarrollo de las capas y tejidos embrionarios (véase Figura 2.4
de la Introducción). Al día 8 de gestación en el ratón, los estadios embrionarios (E) encontrados,
fueron los que se describen, brevemente, a continuación:
» Embrión en TS9: embrión desarrollado al 6to. día y 15-20 hs de gestación. Embrión con canal
proamniótico y cono ectoplacentario. Los tejidos que constituyen el concepto en este estadio
son el ectodermo extraembrionario, el ectodermo embrionario, el endodermo visceral y parietal,
y la cavidad del saco vitelino (Figura II.6.A).
» Embrión en TS10: embrión en estadio de “Línea primitiva”.
En este estadio embrionario se presentan los siguientes subtipos de diferenciación:
» Embrión en TS10a: embrión desarrollado al 7mo. día y 3 hs de gestación.
Se caracteriza por la aparición del mesodermo, presencia de canal proamniótico, aún
continuo y adyacente al ectodermo extraembrionario y embrionario, y por la presencia de
cono ectoplacentario pequeño y poco extendido (Figura II.6.B).
» Embrión en TS10ab: embrión desarrollado al 7mo. día y 10 hs de gestación.
Se caracteriza por la aparición de los pliegues amnióticos (en el polo posterior del embrión
primero y luego en el polo anterior), la presencia del canal promaniótico e inicio de la
formación de la cavidad exocelómica y del cono ectoplacentario (Figura II.6.C).
» Embrión en TS10b: embrión desarrollado al 8vo. día de gestación.
Se caracteriza por presentar las tres cavidades bien definidas: la cavidad amniótica, la
exocelómica y la ectoplacentaria. Se distinguen las membranas del corion y del amnios. El
corion separa la cavidad ectoplacentaria de la exocelómica mientras que el amnios separa la
cavidad amniótica de la exocelómica (Figura II.6.D).
Los estadios embrionarios se determinaron y se cuantificó el número total de embriones de cada
tipo, número expresado como porcentaje del total de sitios analizados, en cada grupo
experimental.

Capítulo II�
123
�
Figura II.6. Diferenciación embrionaria al día 8 de gestación. Micrografías representativas de
embriones en gastrulación (cortes transversales de sitios de implantación incluidos en parafina). A,
embrión en estadio TS9, mostrando el canal proamniótico (*), el ectodermo extraembrionario (EctoEx)
y embrionario (EctoE), endodermo visceral (EndoV), endodermo parietal (EndoP), la cavidad del saco
vitelino (CSV) y células gigantes trofoblásticas primarias (CGT 1rias); B, embrión en estadio TS10a, donde
se observa la aparición del mesodermo (Me), y el canal proamniótico (*) continuo a lo largo del EctoEx
y ExtoE; C, embrión en estadio TS10ab, donde se observa la aparición del pliegue posterior (Pl amn P),
el exoceloma (Exocel) y la cavidad ectoplacentaria (Cav Ectopl); D, embrión en estadio TS10b, donde se
observan las tres cavidades bien definidas: Cav Ectopl, la cavidad exocelómica (Cav Exocel) y la cavidad
amniótica (CA); el corion y el amnios. En todos los casos se observa el cono ectoplacentario (CE) en el
lado mesometrial del sitio de implantación y CGTs 2rias. TS: Escala de Theiler. Aumento 10x.
2.3.5. Evaluación histológica embrio-trofoblástica al día 8 de gestación: estudio histopatológico.
Para analizar la calidad histo-morfológica de los tejidos embrionarios, extraembrionarios y
trofoblásticos en el sitio de implantación luego de la exposición a alcohol, en promedio, dos sitios
por hembra, elegidos al azar, fueron procesados histológicamente y analizados bajo microscopio
óptico. El estudio involucró la observación de las características descriptas a continuación:
» Cono ectoplacentario:
Se estudiaron las dos zonas que conforman el cono:
» Zona de proliferación trofoblástica (core proliferativo), se analizó:
.El grado de compactación del tejido. Patrón normal: tejido compacto.
.El grado de expansión. Patrón normal: en expansión moderada.

Capítulo II�
124
�
.La morfología de las células trofoblásticas. Patrón normal: células mononucleares
pequeñas, en estado proliferativo.
.La presencia o no de glóbulos rojos materno. Patrón normal: zona sin glóbulos rojos.
» Zona de expansión trofoblástica, se analizó:
.El grado de desarrollo o expansión. Patrón normal: en expansión hacia el lado
mesometrial, zona de gran dispersión trofoblástica (células en migración).
.La morfología de las células trofoblásticas. Patrón normal: trofoblastos mononucleares de
diferentes tamaños, según su grado de diferenciación.
.La presencia y abundancia de células gigantes trofoblásticas secundarias (CGTs 2rias).
Patrón normal: presencia de CGTs 2rias diferenciadas, principalmente en la región más
periférica de la zona de expansión.
.La presencia y abundancia de glóbulos rojos maternos. Patrón normal: zona con moderada
infiltración de glóbulos rojos.
» Interfase trofoblástico - decidual:
En el perímetro lateral y antimesometrial del embrión, se analizó:
.La presencia de células CGTs 1rias. Patrón normal: presencia de CGTs 1rias en todo el
perímetro de la interfase, siendo frecuentemente más abundantes en la zona de la cripta
antimesometrial.
.La remodelación del tejido antimesometrial (cripta antimesometrial). Patrón normal: zona
relativamente compacta, con fragmentación moderada, luz uterina reducida, picnosis
nuclear moderada a alta y escasa a moderada cantidad de eritrocitos maternos.
.La remodelación del tejido lateral trofoblástico-decidual. Patrón normal: CGTs 1rias y
células deciduales (de morfología poliédrica) en contacto, ausencia de edema y de lagunas
sanguíneas amplias, fragmentación celular escasa a moderada, glóbulos rojo maternos
escasos.
» Tejidos extraembrionarios y embrionarios:
En el ectodermo, mesodermo y endodermo embrionarios y extraembrionarios, se analizó:
.La morfología de las células. Patrón normal: varía de acuerdo al tejido (se detalla en
Resultados)

Capítulo II�
125
�
.El grado de organización celular. Patrón normal: tejidos con disposición celular ordenada
(se detalla en Resultados)
En primer lugar, se analizó el patrón histológico normal por zona y tejido en las muestras
controles, y luego, en comparación con este patrón se detectaron los cambios histológicos en las
muestras de las hembras tratadas. Las alteraciones histológicas encontradas en cada tejido (cono
ectoplacentario, interfase trofoblástico-decidual, tejidos embrionarios y extraembrionarios),
fueron registradas para calcular la frecuencia de anomalías en el total de sitios analizados para el
grupo control y el tratado.
Estudios in-vitro del desarrollo trofoblástico: capacidad invasiva.
2.3.6. Cultivo de conos ectoplacentarios.
Al 8vo. día de gestación, HC y HT de la colonia CD-1 fueron sacrificadas por dislocación cervical. Los
cuernos uterinos fueron rápidamente removidos y lavados en PBS estéril con SFB 10% a 37°C.
Luego fueron colocados sobre una placa de parafina y mediante un corte longitudinal desde un
extremo del cuerno hacia el otro extremo, el miometrio fue completamente aislado y descartado.
Cada decidua aislada fue transferida a una placa de Petri con PBS a 37°C. Bajo estereomicroscopio
y con ayuda de instrumental oftalmológico fue aislado de la decidua, cuidadosamente, el concepto
junto con el cono ectoplacentario. Finalmente, con bisturí oftalmológico, se separó el cono del
embrión (Figura II.7). Todos los conos, así disecados, fueron lavados varias veces en placas con
PBS-SFB estéril hasta lograr la remoción total de las células sanguíneas, siendo posteriormente
colocados en cultivo. Para ello, los conos ectoplacentarios fueron transferidos a placas de 24
pocillos con 500 µl de medio D-MEM estéril suplementado con SFB (10%), BSA (1%), insulina (0,4
µg/ml), ácido láctico (0,5 mg/ml), piruvato (0,5 mg/ml), Mitos+TM (0,2 ng/ml), nucleosidos (timidina
0,24 mg/ml, adenosina 0,8 mg/ml, guanosina 0,85 mg/ml, uridina 0,73 mg/ml) y antibióticos (50
UI/ml de penicilina G y 50 µg/ml estreptomicina), y colocados sobre portaobjetos circulares de
vidrio limpios y estériles (3 a 4 conos por pocillo). Los conos fueron incubados en estufa de cultivo
con atmósfera húmeda y con 5% de CO2 en aire, a 36,5°C por 72 hs. El medio de cultivo fue
parcialmente renovado cada 24 hs.

Capítulo II�
126
�
Figura II.7. Disección de cono ectoplacentario al día 8 de gestación. Micrografías de
estereomicroscopio representativas de sitios de implantación (A-B) y embriones disecados (C-D), de
hembras CD-1. A, decidua aislada; B, decidua alojando en su interior al concepto (embrión y sus tejidos
extraembrionarios, punta de flecha); C, embrión disecado de la decidua; D, cono ectoplacentario
separado del embrión. CE: cono ectoplacentario, DAM: decidua antimesometrial, DM: decidua
mesometrial, E: embrión. Aumentos: A y B: 10x; C y D: 20x.
2.3.7. Análisis de la viabilidad, desarrollo y capacidad invasiva de los conos ectoplacentarios en
cultivo.
Con el fin de evaluar la viabilidad, la capacidad de desarrollo in-vitro, así como, la capacidad
invasiva (en relación al crecimiento trofoblástico y diferenciación de CGTs) de los conos
provenientes de hembras expuestas a alcohol respecto de los controles, se realizaron los
siguientes análisis:
En primer lugar, la viabilidad trofoblástica in-vitro fue verificada, mediante la observación bajo
microscopio invertido con contraste de fase, de la adhesión o no de los conos ectoplacentarios a
las placas de cultivo, registrándose el número de conos adheridos sobre el total de conos
colocados en cultivo. Aquellos conos adheridos a las 24 hs fueron considerados como conos
viables. En los conos clasificados como viables fue analizada la capacidad de desarrollo

Capítulo II�
127
�
trofoblástico, en el lapso de 24 a 48 hs de cultivo, para lo que se cuantificó el número de conos
adheridos con o sin desarrollo trofoblástico a las 24 y 48 hs relativizando dicho valor al total de
conos adheridos a las 24 hs. Este procedimiento se llevó a cabo para cada cono ectoplacentario
colocado en cultivo, el que fue individualizado con el fin de realizar el seguimiento de su desarrollo
hasta las 48 hs.
Para el análisis del crecimiento o expansión trofoblástica in-vitro, cada cono adherido a la placa fue
fotografiado bajo microscopio con contraste de fase a las 24 y 48 hs. De acuerdo a los valores de
área trofoblástica total (µm2) obtenidos por análisis morfométrico de imágenes con el programa
Image Pro Plus, y debido a la gran variabilidad de tamaños, en cada tiempo de cultivo (T24 y T48
hs), los conos fueron categorizados a T24 y T48 hs en tres tipos: conos pequeños: con baja
expansión trofoblástica; conos medianos: con moderada expansión trofoblástica; y conos grandes:
con gran expansión trofoblástica. El número de conos pertenecientes a cada categoría fue
cuantificado y expresado como porcentaje sobre el número total de conos con desarrollo
trofoblástico. Para cada uno de los tipos de conos ectoplacentarios en cultivo, además de registrar
morfométricamente el área trofoblástica total, se registró el área correspondiente al core de
proliferación. Luego, el área de expansión trofoblástica se calculó como la diferencia entre el área
trofoblástica total y el área del core. Así, el área promedio + SD fue calculada para cada tamaño de
cono y tiempo de cultivo.
2.3.8. Análisis de la morfología de los conos ectoplacentarios en cultivo.
Para el análisis morfológico de los conos ectoplacentarios in-vitro a las 24 y 48 hs de cultivo fueron
observadas, en micrografías con contraste de fase, las características de las células trofoblásticas
centrales del core proliferativo y de las células trofoblásticas periféricas pertenecientes al área de
expansión trofoblástica del cono. En esta última zona se evaluó la presencia de proyecciones
citoplasmáticas trofoblásticas, características de células en migración (filopodios), y la
diferenciación a células gigantes trofoblásticas (células de grandes dimensiones y con numerosos
nucléolos).

Capítulo II�
128
�
2.3.9. Análisis de la diferenciación de las células trofoblásticas del cono ectoplacentario en
cultivo: Inmunohistoquímica para BrdU.
El nivel relativo de síntesis de ADN de las células trofoblásticas de los conos ectoplacentarios a las
48 hs de cultivo, estimado en función de la cantidad de células que incorporan bromodeoxiuridina
(BrdU) mediante inmunohistoquímica para BrdU, se utiliza para monitorear el proceso de
diferenciación de las células trofoblásticas (endorreduplicación del ADN, preponderante a T48 hs).
Los conos ectoplacentarios de las HC y HT cultivados hasta las 45 hs fueron incubados con BrdU
(0,3 µg/ml) por 3 hs en estufa de cultivo gaseada. Posteriormente, fueron lavados con PBS para
remoción del medio de marcación y fijados con una solución de paraformaldehído 2% por 20 min.
La localización del BrdU incorporado en las células fue realizada por una secuencia de
incubaciones intercaladas de lavados con PBS. Brevemente, las muestras fueron permeabilizadas
con acetona a -20°C por 10 min. Luego las peroxidasas endógenas fueron inhibidas con peróxido
de hidrógeno 3% en metanol por 10 min. Posteriormente, los conos fueron incubados con HCl 1N
con pepsina 0,1% 10 min a 60°C, HCl 1N con pepsina 0,1% 10 min a 37°C y con HCl 0,1N 20 min a
37°C, para favorecer los procesos de hidrólisis. El bloqueo de sitios inespecíficos fue realizado
mediante la incubación con PBS - BSA 1% - Saponina 0,05% - glicina 0,2%, 1h a 37°C, a lo que le
siguió la incubación con el anticuerpo primario anti BrdU biotinilado, overnight a 4°C. Al día
siguiente, fueron incubados con estreptoavidina - peroxidasa 40 min a 37°C y luego revelados con
diaminobenzidina (DAB; 0,5 mg/ml - peróxido de hidrógeno 3%). La contracoloración fue realizada
con hematoxilina de Mayer y finalmente el montaje fue realizado en óleo mineral. El índice de
incorporación de BrdU en los conos cultivados por 48 hs fue calculado a partir de la cuantificación
de células con marca positiva sobre un total de 500 células trofoblásticas del área de expansión.
Las células del core proliferativo, altamente positivas para BrdU, no fueron incluidas en el
recuento. Los datos fueron expresados como número total de células BrdU-positivas sobre el total
de conos en cultivo muestreados para HC y HT.
3. Análisis estadístico.
Los valores expresados como promedios + SD del grupo control y tratado fueron comparados
mediante prueba T de Student. Los valores de frecuencias, expresados como porcentajes, fueron
comparados mediante una prueba exacta de Fisher o Chi-cuadrado, según el caso. Para el análisis

Capítulo II�
129
�
estadístico fue utilizado el programa GraphPad Instat v2.05a. Un valor de p<0,05 fue considerado
estadísticamente significativo.

Capítulo II�
130
�
II.2. RESULTADOS
Dado que en nuestro laboratorio demostramos que la ingesta de alcohol, en hembras CF-1, desde
un período previo a la gestación (15 días) y hasta la etapa de organogénesis temprana (día 10 de
gestación) induce aumento de reabsorciones y retrasa la tasa de diferenciación embrionaria (Coll y
col., 2011), postulamos que estas alteraciones producidas por el consumo perigestacional tendrían
su origen en días previos, en días 7-8 de gestación (postimplantación temprana) o aún antes, en
día 4 (preimplantación), por inducción de anomalías tanto en linaje del propio embrión como en el
linaje trofoblástico, llevando a arresto del desarrollo embrionario en períodos periimplantatorios y
al día 10 de gestación. Para poner a prueba esta hipótesis, hembras murinas CD-1 y CF-1 fueron
tratadas con alcohol, desde antes de la gestación y hasta los días 4, 5 y 8 de preñez, momento en
los cuales se analizaron los efectos en el desarrollo y morfogénesis embrio-trofoblástica.
Al 4to. y 8vo. día de gestación, se evaluaron las variaciones en el consumo líquido y sólido, las
calorías ingeridas y los cambios en el peso corporal, de acuerdo a lo detallado en Materiales y
métodos. Si bien, se detectaron diferencias significativas en el consumo líquido en los grupos
tratados respecto de los controles, la cantidad total de calorías consumidas en ambos grupos no
varió significativamente entre los grupos. Tampoco se vio alterado el peso corporal de las HT en
comparación con el de las HC.
1. Efectos del consumo perigestacional de alcohol en embriones de preimplantación:
Día 4 de gestación.
Dado que se observó que en embriones en estadios preimplantorios (desde dos células) expuestos
in-vitro a alcohol impacta de formas diferentes en el desarrollo hasta blastocisto, dependiendo de
las condiciones de exposición (dosis y tiempos de exposición) (Stacheski y col., 1994a, 1994b), el
objetivo de esta sección fue evaluar los efectos de la administración de bebida alcohólica desde
antes de la preñez y hasta el día 4 de gestación, en el desarrollo in-vivo de los blastocistos en dos
modelos murinos (CD-1 y CF-1), determinando asimismo si el mismo esquema de exposición induce
patrones de desarrollo similares en ambas colonias murinas.

Capítulo II�
131
�
1.1. Efecto del consumo perigestacional de alcohol en el desempeño reproductivo.
Con el fin de evaluar el desempeño reproductivo tanto en las hembras CD-1 como CF-1, luego de
la ingesta de alcohol, se registró el número total de embriones recuperados por hembra al día 4 de
gestación. Asimismo, se determinó la proporción de embriones viables (sin fragmentaciones o con
menos del 50% del volumen embrionario fragmentado/lisado) y de inviables (más del 50% del
volumen embrionario fragmentado/lisado). Los resultados se muestran en la Tabla II.3. El número
de embriones recuperados y de embriones viables por hembra en ambas colonias murinas no
resultó alterado por el consumo de alcohol. Sin embargo, se observó que las HT-CF-1 presentaron
un aumento significativo del número total de embriones fragmentados y reducción del total de
embriones viables (p<0,01) comparando con las HC-CF-1. Estas alteraciones no fueron halladas en
las HT-CD-1.
Tabla II.3. Desempeño reproductivo al día 4 de gestación.
CD-1 CF-1
HC (5) HT (6) HC (10) HT (6)
N° total de embriones 56 64 172 130
N° embr totales/hembra + SD 11,2 + 3,1 10,5 + 2,2 17,2 + 8,5 21,6 + 9,6
N° embr viables/hembra + SD 11,2 + 3,1 10,5 + 2,2 16,4 + 7,3 18,8 + 10,6
N° embr viables/totales (%) 56/56 (100) 64/64 (100) 164/172 (95,4) 113/130 (86,9)**
N° embr fragm/totales (%) 0/56 (0) 0/64 (0) 8/172 (4,6) 17/130 (13,0)**
Monitoreo de la preñez y rendimiento gestacional al día 4 de gestación en hembras controles (HC) y
tratadas con alcohol (HT), CD-1 y CF-1. Entre paréntesis se indica el número de hembras de cada
grupo. embr: embriones, fragm: fragmentados. Los valores se expresan como promedios + SD y como
frecuencias (porcentajes, %). **p<0,01 versus su control (Prueba Chi-cuadrado).
1.2. Efecto del consumo perigestacional de alcohol en la diferenciación embrionaria.
El objetivo de esta sección fue analizar el desarrollo embrionario in-vivo, luego del consumo
materno de alcohol, en relación al grado de diferenciación embrionaria o “timing” de
embriogénesis. Para ello, los embriones recuperados de hembras HC y HT al día 4 de gestación se

Capítulo II�
132
�
clasificaron según su grado de diferenciación en: mórulas (pre- y compactadas; TS 3), blastocistos
iniciales (con cavidad blastocélica en formación; TS 4), blastocistos expandidos (con cavidad
blastocélica expandida; TS 4) y blastocistos extruidos (embrión implantativo, sin zona pelúcida, con
cavidad blastocélica visible o no; TS 5). Tanto en las HT-CD-1 como en las HT-CF-1 se evidenció
elevado número de mórulas (Figura II.8.B y D) comparando con el alto número de blastocistos
característico del tiempo gestacional (día 4) hallado en las HC de ambas colonias (Figura II.8.A y C).
Figura II.8. Diferenciación embrionaria in-vivo al día 4 de gestación. Micrografías con contraste de
fase representativas de embriones de preimplantación de hembras CD-1 (A, B) y CF-1 (C, D); controles
(A, C) y tratadas con alcohol (B, D) donde se muestran los distintos estadios embrionarios observados
al día 4 de gestación. Bl Inic: blastocisto inicial, Bl Exp, blastocisto expandido, M: mórula. Aumento:
10x.
El número de embriones de cada estadio fue expresado como porcentaje sobre el total de
embriones viables y se presenta en la Figura II.9. Al día 4 de gestación se esperaba que el
porcentaje mayoritario de embriones correspondiera a aquellos embriones en estadio de
blastocisto. Para las HC de ambas colonias lo esperado se condice con lo observado. Sin embrago,
cuando se analizó la distribución de tipos embrionarios en las HT, en ambas colonias, disminuyó
significativamente la proporción de blastocistos expandidos y de extruidos (p<0,05, Figura II.9) y
aumentó significativamente la cantidad de mórulas (HT-CD-1: p<0,01, HT-CF-1: p<0,05, Figura II.9)

Capítulo II�
133
�
lo que indicó un menor grado de diferenciación de la población embrionaria de CD-1 y CF-1
expuesta a alcohol.
Figura II.9. Diferenciación embrionaria in-vivo al día 4 de gestación. Los embriones de hembras
controles (HC) y tratadas con alcohol (HT), CD-1 (A) y CF-1 (B) fueron recolectados y clasificados según
cada tipo embrionario. Bl Inic: blastocistos iniciales, Bl Exp: blastocistos expandidos, Bl Extr:
blastocistos extruidos. Los valores se expresan como porcentajes del total de los embriones viables
recuperados.*p<0,05 y **p<0,01 versus su control (Prueba Chi-cuadrado).
1.3. Efecto del consumo perigestacional de alcohol en la morfología embrionaria.
Se analizó la calidad de los embriones de preimplantación recuperados al día 4 de gestación. Tanto
en las hembras tratadas de la colonia CD-1 como CF-1 se encontró una alta proporción de
embriones morfológicamente anormales (Figura II.10), caracterizados por la presencia de
fragmentaciones citoplasmáticas, blastómeras con signos de lisis, blastómeras de tamaño no
homogéneo, persistencia de blastómeras sin compactar en mórulas compactadas, siendo
principalmente observados signos de cavitación anormal en los blastocistos iniciales.

Capítulo II�
134
�
Figura II.10. Morfología embrionaria al día 4 de gestación. Micrografías con contraste de fase
representativas de embriones de preimplantación de hembras CD-1 (A) y CF-1 (B) tratadas con alcohol,
donde se muestran distintas anomalías morfológicas. 1: blastocisto inicial o expandido normal, 2:
blastocisto con cavidad blastocélica anormal (punta de flecha); 3: mórula compactada anormalmente
con blastómeras protruyendo (*); 4: embrión con espacio perivitelino aumentado; 5: embrión lisado
(inviable). Aumento: 10x.
En ambos grupos murinos expuestos a alcohol antes y hasta el día 4 de gestación, el número de
embriones morfológicamente anormales, calculado sobre el total de embriones viables, resultó
aumentado (p<0,05, Tabla II.4). Se destaca en las HT-CF-1 una mayor proporción de blastocistos
anormales respecto de los controles HC-CF-1 (p<0,05, Tabla II.4). En las hembras CD-1, si bien, no
se puedo establecer significancia estadística en la cantidad de mórulas y blastocistos con
morfología anormal, se indica la tendencia observada.
Tabla II.4. Morfología embrionaria al día 4 de gestación.
CD-1 CF-1
HC (5) HT (6) HC (10) HT (6)
N° total de embriones viables 56 64 164 113
N° embr anormales/totales (%) 1/56 (1,8) 8/64 (12,5)* 15/164 (9,1) 19/113 (16,8)*
N° M anormales/M totales (%) 0/4 (0) 4/18 (22,2) (*) 4/20 (20,0) 5/25 (20,0)
N° Bl anormales/Bl totales (%) 1/52 (1,9) 4/46 (8,7) (*) 11/144 (7,6) 14/88 (15,9)*
Cuantificación del número de embriones anormales al día 4 de gestación en hembras controles (HC) y
tratadas con alcohol (HT), CD-1 y CF-1. Para cada grupo, entre paréntesis se indica el número de
hembras utilizadas. Bl: blastocistos, embr: embriones, M: mórulas. Los valores se expresan como
frecuencias (porcentajes, %). *p<0,05 versus su control (Prueba Chi-cuadrado), (*): tendencia
observada de aumento.

Capítulo II�
135
�
1.4. Efecto del consumo perigestacional de alcohol en la tasa de crecimiento embrionario.
Con el fin de estudiar si el consumo materno de alcohol altera el crecimiento embrionario
comprometiendo así su potencial desarrollo, el número de núcleos por embrión fue cuantificado.
Dicho recuento se realizó tanto en mórulas como en blastocistos de hembras CD-1 y CF-1. A su
vez, el índice mitótico (IM) fue calculado. En la Figura II.11, se muestran fotos representativas de
núcleos de embriones obtenidos mediante la técnica de Tarkowski (1966).
El crecimiento embrionario (número de núcleos por embrión) sólo se vio significativamente
disminuido en los blastocistos iniciales provenientes de las HT tanto de CD-1 (p<0,05; Figura II.12A)
como de CF-1 (p<0,01; Figura II.12B).
Figura II.11. Crecimiento embrionario al día 4 de gestación. Micrografías representativas de núcleos
de embriones de preimplantación. A, mórula; B, blastocisto; C, detalle de núcleos en distintas fases del
ciclo celular: interfase (in), profase (pr), metafase (me) y telofase (te); D, detalle de dos núcleos en
metafase (me). Técnica de Tarkowski. Aumentos: A y B: 10x, C: 40x, D: 60x.

Capítulo II�
136
�
Figura II.12. Crecimiento embrionario al día 4 de gestación. Recuento del número de núcleos por tipo
embrionario en hembras controles (HC) y tratadas (HT), CD-1 (A) y CF-1 (B). Bl Inic: blastocistos
iniciales, Bl Exp: blastocistos expandidos, Bl Extr: blastocistos extruidos. Los valores se expresan como
promedios + SD. *p<0,05 y **p<0,01 versus su control (Prueba T de Student).
Para analizar la tasa de división celular en cada tipo embrionario, se calculó el índice mitótico (IM)
(Figura II.13). La proporción de núcleos en división resultó alterada en los blastocistos
provenientes de HT de ambas colonias. Los blastocistos iniciales de las HT-CD-1 presentaron IM
significativamente reducido (p<0,05), mientras que los blastocistos expandidos presentaron IM
significativamente aumentado (p<0,01), respecto de su control. Por otro lado, tanto las mórulas
como los blastocistos iniciales y expandidos de las HT-CF-1 presentaron IM significativamente
disminuido respecto de su control (p<0,05, p<0,01 y p<0,001, respectivamente).
Figura II.13. Índice mitótico por tipo embrionario al día 4 de gestación. N° de núcleos en división por
tipo embrionario en hembras controles (HC) y tratadas con alcohol (HT), CD-1 (A) y CF-1 (B). Bl Inic:
blastocistos iniciales, Bl Exp: bl. expandidos. Los valores expresan el índice mitótico (número de núcleos
en división sobre número total de núcleos contados, para cada tipo embrionario y en un total de 5
hembras para cada grupo). *p<0,05, **p<0,01 y ***p<0,001 versus su control (Prueba Chi-cuadrado).

Capítulo II�
137
�
2. Efectos del consumo materno perigestacional de alcohol en el desarrollo embrio-trofoblástico
durante la implantación: Día 5 de gestación.
Dado que la ingesta de alcohol hasta el día 4 de gestación afectó negativamente la tasa de
diferenciación, crecimiento y morfología de los blastocistos, el siguiente objetivo fue determinar si
la exposición a alcohol hasta el período implantatorio (día 5 de gestación) era capaz de inducir
alteraciones en el desarrollo de embriones durante las fases de la implantación (adhesión e
invasión del trofoblasto). Si así fuera, los efectos negativos embrio-trofoblásticos producidos por la
ingesta de alcohol podrían afectar la capacidad invasiva embrionaria y se manifestarían como
reabsorciones tempranas en estadios posteriores. Para dilucidar esta hipótesis, en primer lugar, se
monitoreó al día 5 de gestación el desempeño reproductivo, la tasa de diferenciación, la
morfología y el crecimiento de embriones en estadio implantativo. En segundo lugar, se analizó in-
vitro, si la diferenciación y el crecimiento embrio-trofoblástico durante la implantación resultaron
alterados por la exposición in-vivo a alcohol. Dado que la embriogénesis preimplantativa (día 4) en
los dos modelos utilizados, CD-1 y CF-1, resultó afectada negativamente, se decidió llevar a cabo el
estudio de este objetivo en la colonia CD-1.
2.1. Efecto del consumo perigestacional de alcohol hasta el día 5 de preñez en el desempeño
reproductivo, en la diferenciación, morfología y crecimiento embrionario implantativo (in-vivo).
Con el fin de monitorear la calidad de la preñez de las hembras CD-1 expuestas a alcohol, se
registró el número total de embriones recuperados al día 5 de gestación. El número de embriones
recuperados luego del flushing disminuyó significativamente en las hembras tratadas comparando
con el número embriones recuperado de las controles (p<0,05, Tabla II.5). Asimismo, el número de
embriones implantativos viables resultó disminuido en las hembras expuestas a alcohol (p<0,05,
Tabla II.5).

Capítulo II�
138
�
Tabla II.5. Desempeño reproductivo al día 5 de gestación.�
CD-1
HC (23) HT (23)
N° total de embriones 116 84
N° embr totales/hembra + SD 5,1 + 2,2 3,7 + 1,8*
N° embr viables/hembra + SD 4,9 + 1,2 3,5 + 1,9*
N° embr viables/totales (%) 114/116 (98,3) 81/84 (96,5)
N° embr fragm/totales (%) 2/116 (1,7) 3/84 (3,5)
Monitoreo de la preñez y rendimiento gestacional al día 5 de gestación en hembras controles (HC) y
tratadas con alcohol (HT), CD-1. Entre paréntesis se indica el número de hembras de cada grupo. Embr:
embriones, fragm: fragmentados. Los valores se expresan como promedios + SD o frecuencias
(porcentajes, %). *p<0,05 versus su control (Prueba T de Student).
Para analizar la diferenciación embrionaria al comienzo de la fase de implantación (mañana del día
5 de gestación), luego de la exposición a alcohol, los blastocistos implantativos (sin zona pelúcida)
recuperados fueron observados y clasificados en tres categorías de diferenciación, según su
aspecto morfológico: embriones implantativos tipo 1, tipo 2 y tipo 3 (véase Figura II.4). Para ello se
analizó: la presencia de cavidad blastocélica, el inicio de la expansión de las células
trofoectodérmicas (procesos trofoblásticos) y la presencia de CGTs diferenciadas (véase Tabla II.2).
El embrión implantativo tipo 1 presenta cavidad blastocélica visible y expandida; el embrión
implantativo tipo 2 presenta cavidad blastocélica reducida e incipiente desarrollo del trofoblasto,
procesos trofoblásticos, que protruye del embrión; y el embrión implantativo tipo 3 no presenta
cavidad blastocélica visible y el desarrollo trofoblástico ya es moderado, lo que coincide con la
preparación del embrión para la implantación. De acuerdo con esta clasificación, el embrión con
mayor grado de desarrollo corresponde al tipo 3. El número de embriones de cada tipo fue
registrado y expresado como porcentaje del total de embriones recuperados. Como se observa en
la Figura II.14, encontramos diferencias significativas entre las HC y las HT para los embriones tipo
2 y tipo 3. Mientras que las HC presentaron mayor porcentaje de embriones en el estadio más
avanzado (tipo 3), las HT tuvieron un incremento significativo de embriones tipo 2 (p<0,05) y un
reducido porcentaje de embriones tipo 3 (p<0,05), respecto del control, sugiriendo que la

Capítulo II�
139
�
población de embriones de las HT presenta retraso de la diferenciación al día 5 de gestación, al
inicio del período de implantación.
Figura II.14. Diferenciación embrionaria in-vivo al día 5 de gestación. Los embriones de hembras
controles (HC) y tratadas con alcohol (HT), CD-1, fueron recolectados y clasificados según cada tipo
embrionario. Embr: embriones. Los valores se expresan como porcentajes del total de los embriones
viables recuperados.*p<0,05 versus su control (Prueba Chi-cuadrado).
Se analizó la calidad morfológica de los embriones implantativos, para lo cual fueron consideradas
como anormales las siguientes características: signos de alteraciones de la cavitación (Figura
II.15.B), fragmentaciones en el trofoblasto y/o macizo celular interno (Figura II.15.C).
Figura II.15. Morfología embrionaria al día 5 de gestación. Micrografías con contraste de fase
representativas de embriones implantativos de hembras CD-1 tratadas con alcohol, donde se muestran
distintas anomalías morfológicas. A, blastocisto implantativo normal; B, blastocisto implantativo con
cavidad blastocélica anormal (punta de flecha); C, blastocisto implantativo anormal con expansión
trofoblástica con signos de fragmentación y lisis celular (punta de flecha). Aumento: 20x.

Capítulo II�
140
�
Tanto para las HC-CD-1 como para las HT-CD-1, el número de embriones morfológicamente
anormales fue registrado y expresado como porcentaje del total de los embriones analizados
Como se muestra en la Figura II.16, dicho valor se encontró significativamente aumentado en las
HT comparando con el valor de las HC (p<0,05).
Figura II.16. Morfología embrionaria al día 5 de gestación. Los embriones de hembras controles (HC) y
tratadas con alcohol (HT), CD-1, fueron recolectados, analizados según su aspecto morfológico y
clasificados en normales o anormales. Los valores se expresan como porcentajes del total de los
embriones viables recuperados.*p<0,05 versus control su control (Prueba Chi-cuadrado).
Con el fin de analizar el crecimiento embrionario, luego del consumo materno de alcohol hasta el
día 5 de gestación, se midió el área de los embriones recuperados (estudio morfométrico). Los
valores de área obtenidos se agruparon en tres categorías de crecimiento embrionario según los
rangos de valores (véase Métodos en este mismo Capítulo): embriones pequeños, medianos y
grandes. Para cada tipo embrionario fue calculada el área promedio por embrión, resultados que
se muestran en la Figura II.20. Al día 5 de gestación, el tamaño promedio de cada tipo embrionario
(pequeños, medianos y grandes) de las HT fue similar al tamaño de los embriones de las HC,
indicando que el crecimiento embrionario en términos de expansión embrio-trofoblástica, no fue
modificada por el consumo materno de alcohol.
2.2. Efecto del consumo perigestacional de alcohol hasta el día 5 de preñez sobre eventos de la
implantación (estudios in-vitro).
Luego de observar que la ingesta materna de alcohol retrasa la diferenciación, afecta el
crecimiento y aumenta la frecuencia de anomalías morfológicas de los embriones implantativos, el

Capítulo II�
141
�
paso siguiente planteado, fue determinar si los blastocistos expuestos in-vivo al alcohol eran
capaces de proseguir el desarrollo normal o si su capacidad implantativa e invasiva resultaba
afectada como consecuencia de la exposición. Para esto se utilizó el cultivo de embriones
implantativos hasta las 72 hs, momento en el que nuevamente se analizó la diferenciación, la
viabilidad y la morfología embrionaria, así como, la dinámica del crecimiento y la expansión
trofoblástica.
Se identificaron los estadios de diferenciación embrionaria según el aspecto morfológico del
embrión observado cada 24 hs y hasta las 72 hs de cultivo (considerando el grado de desarrollo de
las células trofoblásticas y la presencia de CGTs diferenciadas), estableciéndose tres tipos
embrionarios que expresan el grado de diferenciación a las 24, 48 y 72 hs de cultivo (véase Tabla
II.2). El embrión, durante 72 hs de cultivo, se va desarrollando como una monocapa de células
trofoblásticas que se expande gradualmente y que rodea a un agregado de células centrales aún
indiferenciadas que corresponde al macizo celular interno. La expansión de dicha monocapa de
células trofoblásticas es la consecuencia de tres procesos que ocurren de manera simultánea: la
proliferación, la migración y la diferenciación celular, siendo uno u otro más preponderante según
la etapa particular del cultivo (como ocurre in-vivo). Teniendo en cuenta la clasificación del
desarrollo embrionario in-vivo, según la escala de Theiler, las características del desarrollo
embrionario durante el tiempo de cultivo fueron las siguientes: a T0 hs (correspondiente a TS5 ó
estadio embrionario (E) 4 del desarrollo in-vivo) los blastocistos, sin zona pelúcida, presentan o no
cavidad blastocélica visible y tienen un incipiente desarrollo de procesos trofoblásticos; a T24 hs
de cultivo (correspondiente a TS6/E4,5 del desarrollo in-vivo), los blastocistos están estableciendo
contactos (adhesión) con el sustrato, presentan desarrollo de procesos trofoblásticos escaso a
moderado y escasas cantidad de CGTs; a T48 hs de cultivo (correspondiente a TS6/E5 del
desarrollo in-vivo), los embriones inician la migración/invasión en el sustrato, por lo tanto,
presentan mayor desarrollo trofoblástico (polarizado) y son evidentes CGTs diferenciadas. Con 72
hs de cultivo (correspondiente a TS7/E6 del desarrollo in-vivo, en fase de penetración), el embrión
muestra mayor expansión trofoblástica y con gran cantidad de CGTs diferenciadas hacia la
periferia. La Figura II.17 muestra estos tipos embrionarios desarrollados in-vitro hasta las 72 hs.
Una vez establecidos los estadios de diferenciación in-vitro, el siguiente paso fue analizar la
frecuencia de desarrollo de estos tipos durante las 72 hs de cultivo. Para ello, primero se

Capítulo II�
142
�
determinó la viabilidad in-vitro a través de evaluación morfológica, en relación al grado de
fragmentación y/o lisis celular. La viabilidad embrionaria resultó notablemente alterada en los
embriones provenientes de HT. La Figura II.18 muestra micrografías representativas de embriones
anormales encontrados a T24 hs, T48 hs y T72 hs. Tanto la lisis y como la fragmentación celular se
observó, en mayor grado, localizada en el aglomerado celular de la región central del embrión. Los
embriones con estas características no fueron considerados para la cuantificación de la frecuencia
de los tipos embrionarios (diferenciación embrionaria).
Figura II.17. Fases de desarrollo implantacional in-vitro desde el día 5 de gestación, durante 72 hs de
cultivo. Micrografías con contraste de fase representativas de embriones tipo 3, correspondiente al
estadio más avanzado para cada día de desarrollo in-vitro. A, T0 hs: embrión implantativo extruido de
la zona pelúcida, con incipiente desarrollo trofoblástico (punta de flecha); B, T24 hs: embrión con
moderado desarrollo trofoblástico (filopodios, punta de flecha) y algunas CGTs diferenciadas, con
núcleo de grandes dimensiones y varios nucléolos (*); C, T48 hs: embrión con moderada expansión
trofoblástica, ligeramente polarizada (filopodios, punta de flecha) y CGTs (*); D, T72 hs: embrión con
gran expansión trofoblástica (lamelipodios, puntas de flecha) y evidente desarrollo de CGTs en el frente
de invasión (*). Aumento: 20x.
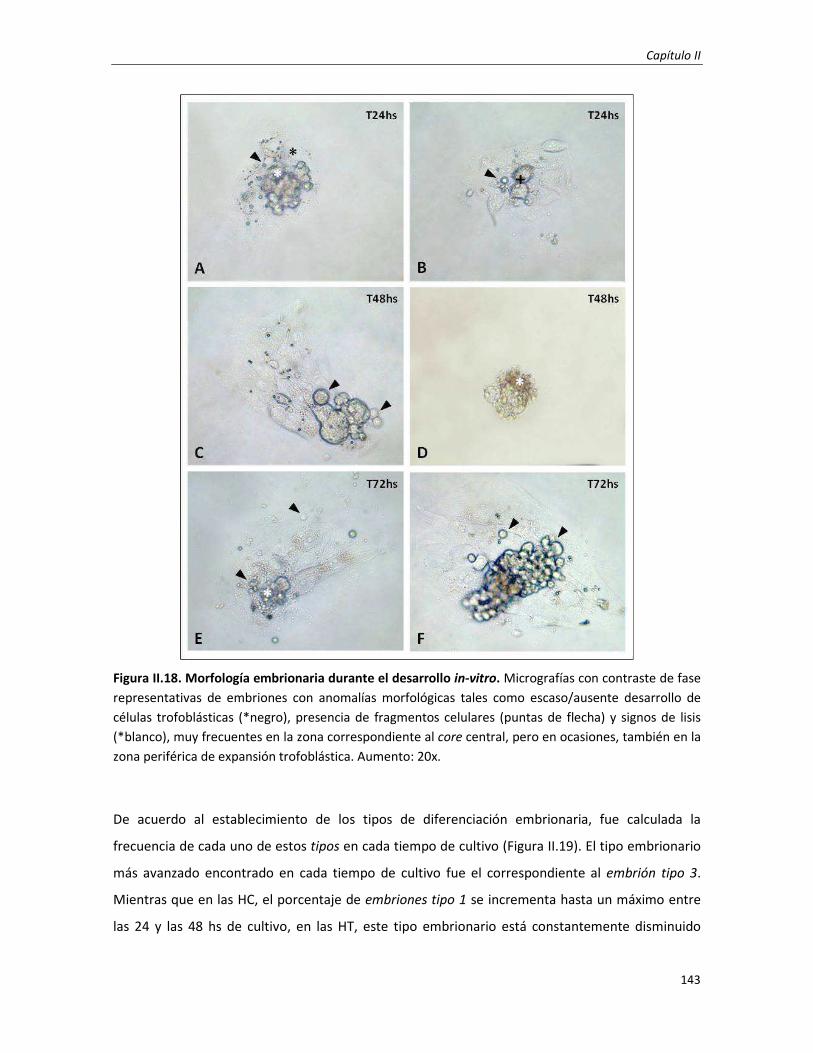
Capítulo II�
143
�
Figura II.18. Morfología embrionaria durante el desarrollo in-vitro. Micrografías con contraste de fase
representativas de embriones con anomalías morfológicas tales como escaso/ausente desarrollo de
células trofoblásticas (*negro), presencia de fragmentos celulares (puntas de flecha) y signos de lisis
(*blanco), muy frecuentes en la zona correspondiente al core central, pero en ocasiones, también en la
zona periférica de expansión trofoblástica. Aumento: 20x.
De acuerdo al establecimiento de los tipos de diferenciación embrionaria, fue calculada la
frecuencia de cada uno de estos tipos en cada tiempo de cultivo (Figura II.19). El tipo embrionario
más avanzado encontrado en cada tiempo de cultivo fue el correspondiente al embrión tipo 3.
Mientras que en las HC, el porcentaje de embriones tipo 1 se incrementa hasta un máximo entre
las 24 y las 48 hs de cultivo, en las HT, este tipo embrionario está constantemente disminuido

Capítulo II�
144
�
(p<0,05, respecto de su control) siendo esta reducción más pronunciada a las 72 hs (Figura II.19.A).
El porcentaje de embriones tipo 2 en las HC oscila en el rango de 13 y 40% en los tiempos de
cultivo. El porcentaje de embriones tipo 2 de HT sigue un patrón similar al control, pero a cada
tiempo de cultivo, el porcentaje resultó significativamente incrementado respecto del control
(Figura II.19.B). El patrón de desarrollo de los embriones tipo 3 de HC fue constante hasta las 24 hs,
luego, decae significativamente a las 48 hs (p<0,05) pero se incrementó significativamente a las 72
hs (p<0,05). El porcentaje de embriones tipo 3 de HT fue constante y similar hasta las 24 hs,
disminuyó significativamente a las 48 hs de cultivo (43,3 versus 18,1%, p<0,05), pero a las 72 hs,
este porcentaje no cambió significativamente respecto de su valor a las 48 hs, como sí ocurrió en
las HC, valor que incrementó. Por lo tanto, a las 72 hs de cultivo, el porcentaje de embriones tipo 3
de las HT resultó significativamente reducido respecto de su control (p<0,05) (Figura II.19.C).
Finalmente, se estudió el grado de crecimiento o expansión embrio-trofoblástica (área en µm2)
durante la implantación en el sistema in-vitro, basado en la hipótesis que el consumo materno de
alcohol afecta negativamente la dinámica del crecimiento y expansión embrio-trofoblástica,
siendo este parámetro un indicador de la capacidad implantativa/invasiva de las células
trofoblásticas. Para esto, los embriones desarrollados en cada tiempo de cultivo fueron agrupados
en tres categorías de tamaño según rangos de áreas de expansión: embriones pequeños, medianos
y grandes. El área promedio para cada tipo embrionario fue calculada y representada en la Figura
II.20. El área de los embriones pequeños de las HC y de las HT aumentó de manera similar desde T0
hs a T24 hs. Sin embargo, a las 48 hs de cultivo, los embriones pequeños de las HT tuvieron un
área significativamente mayor comparado con su control (p<0,05, Figura II.20.A). Este incremento
representó aproximadamente el 33,8% del valor del control. Los embriones medianos de las HC y
los de las HT presentaron similar incremento del área de expansión hasta T24 hs, pero este valor
de área se encontró incrementado en T48 hs en los embriones de HT (p<0,05). En ese momento,
los embriones derivados de HT, presentaron un incremento del orden del 12% sobre el valor
control (Figura II.20.B). Los embriones grandes de las HT a T24 hs presentaron un área
significativamente reducida (p<0,05), comparado con el área de embriones de HC, con un
descenso del 21,5% versus el control (Figura II.20.C).
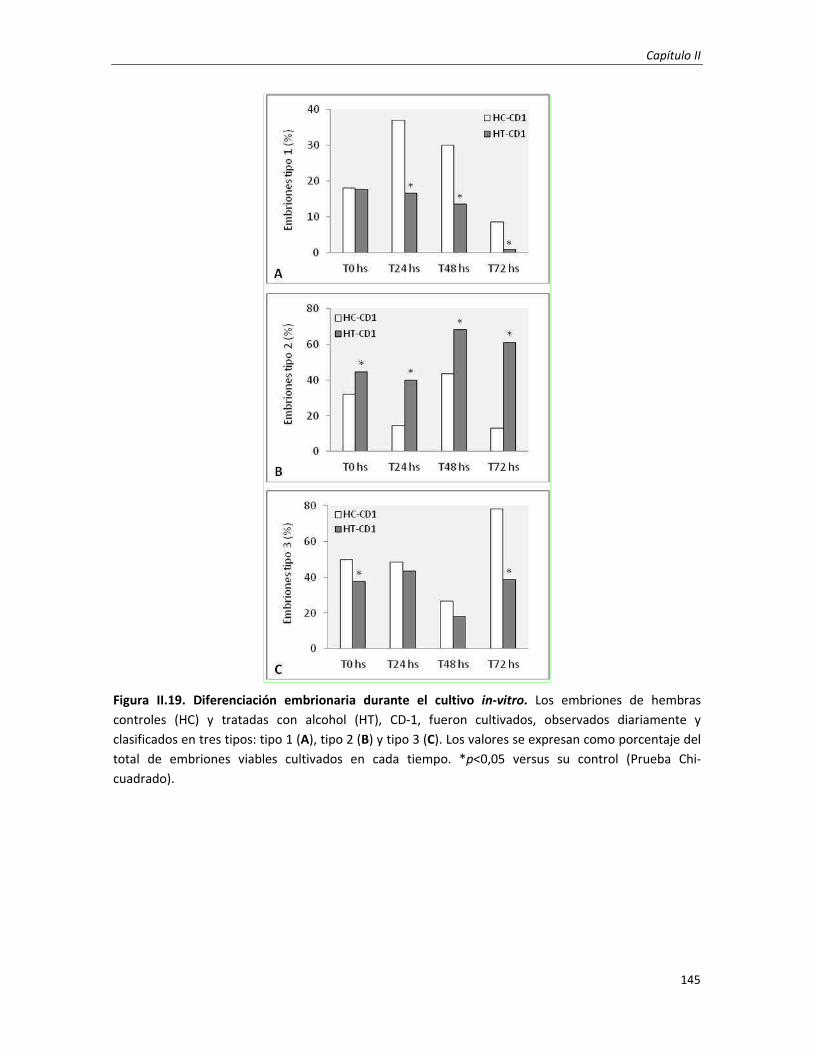
Capítulo II�
145
�
Figura II.19. Diferenciación embrionaria durante el cultivo in-vitro. Los embriones de hembras
controles (HC) y tratadas con alcohol (HT), CD-1, fueron cultivados, observados diariamente y
clasificados en tres tipos: tipo 1 (A), tipo 2 (B) y tipo 3 (C). Los valores se expresan como porcentaje del
total de embriones viables cultivados en cada tiempo. *p<0,05 versus su control (Prueba Chi-
cuadrado).

Capítulo II�
146
�
Figura II.20. Crecimiento embrio-trofoblástico durante el cultivo in-vitro. Los embriones de hembras
controles (HC) y tratadas (HT), CD-1, fueron cultivados y clasificados de acuerdo a tres intervalos de
áreas de expansión embrio-trofoblástica en: embriones pequeños (A), medianos (B) y grandes (C) en
cada tiempo de cultivo (T0 hs, T24 hs y T48 hs). Los valores se expresan como promedio de áreas de
expansión trofoblástica + SD. *p<0,05 versus su control (Prueba T de Student).

Capítulo II�
147
�
3. Efectos del consumo perigestacional de alcohol en el desarrollo embrio-trofoblástico en la fase
de gastrulación: Día 8 de gestación.
Hallazgos previos de nuestro laboratorio demuestran que la ingesta perigestacional de alcohol
hasta el día 10 de gestación, en hembras CF-1, produce efectos deletéreos relacionados con alta
tasa de pérdida gestacional por reabsorción y arresto embrionario en estadios de gastrulación (E7
y 8) (Coll y col., 2011). Por esto, postulamos que la exposición a alcohol, desde antes de la
gestación y hasta el día 8 de preñez, induce daño embrionario, llevando a arresto temprano del
desarrollo del sitio de implantación. Para evaluar esta hipótesis, se estudió en las colonias CD-1 y
CF-1, el desempeño reproductivo, la diferenciación y morfología de las estructuras embrio-
trofoblásticas, al día 8 de gestación.
3.1. Efecto del consumo perigestacional de alcohol en el desempeño reproductivo.
En primer lugar, se evaluó el desempeño reproductivo de las hembras expuestas a alcohol de
manera perigestacional y hasta el día 8 de preñez, en comparación con el rendimiento gestacional
de las controles. Para ello, el número total de sitios de implantación fue registrado en cada grupo
experimental (Tabla II.6). Como se observa, tanto en las HT-CD-1 como las HT-CF-1, el número de
sitios de implantación por hembra varió significativamente respecto de sus controles, resultando
aumentado en las HT-CD-1 y disminuido en las HT-CF-1 (p<0,05).
Tabla II.6. Desempeño reproductivo al día 8 de gestación.
CD-1 CF-1
HC (10) HT (10) HC (6) HT (6)
N° total sitios de implantación 95 115 152 122
N° total sitios de impl/hembra + SD 9,5 + 1,7 11,5 + 1,8* 26,5 + 1,7 20,3 + 1,6*
Monitoreo de la preñez y rendimiento gestacional al día 8 de gestación en hembras controles (HC) y
tratadas con alcohol (HT), CD-1 y CF-1. Para cada grupo, entre paréntesis se indica el número de
hembras utilizadas. Impl: implantación. Los valores se expresan como promedios + SD. *p<0,05 versus
su control (Prueba T de Student).

Capítulo II�
148
�
3.2. Efecto del consumo perigestacional de alcohol en la diferenciación embrionaria in-vivo.
Basados en la idea que la exposición a alcohol hasta el estadio de gastrulación (día 8) causa el
mismo patrón de retraso de desarrollo embrionario al observado cuando el consumo se extiende
hasta la fase de organogénesis (día 10), nos propusimos evaluar el grado de diferenciación
embrionaria en cortes de sitios de implantación colorados con hematoxilina-eosina. Se analizó el
grado de diferenciación embrionaria alcanzado de acuerdo a la escala de Theiler y considerando
los estadios de diferenciación que se encuentran al 8vo. día de preñez en el ratón (véase la Figura
II.6). Tanto en las HT-CD-1 como en HT-CF-1 se observó retraso en el desarrollo embrionario
(Figura II.21). Las HC-CD-1 (Figura II.21.A) presentaron la mayoría de los embriones en el estadio
más avanzado, TS10b, mientras que en las HT-CD-1 hubo una reducción significativa en la
frecuencia de estos embriones, en concordancia con un aumento de la frecuencia de embriones
en el estadio de desarrollo que le antecede (TS10ab, p<0,05). De la misma manera, en la colonia
CF-1, la mayor proporción de los embriones proveniente de HC pertenecieron al estadio más
avanzado de desarrollo, TS10b (Figura II.21.B), mientras que en las HT-CF-1 la mayoría de los
mismos presentaron un menor grado de diferenciación (TS10ab, p<0,05). A su vez, y a diferencia
de lo observado en la colonia CD-1, sólo en las HT-CF-1 encontramos un 17% de embriones en
estadio TS9. En conjunto, estos resultados indican que las hembras CD-1 y CF-1 expuestas a
alcohol resultaron afectadas en cuanto al grado de diferenciación embrionaria, con un retraso de
entre 8 y 24 hs, dependiendo de la colonia murina.
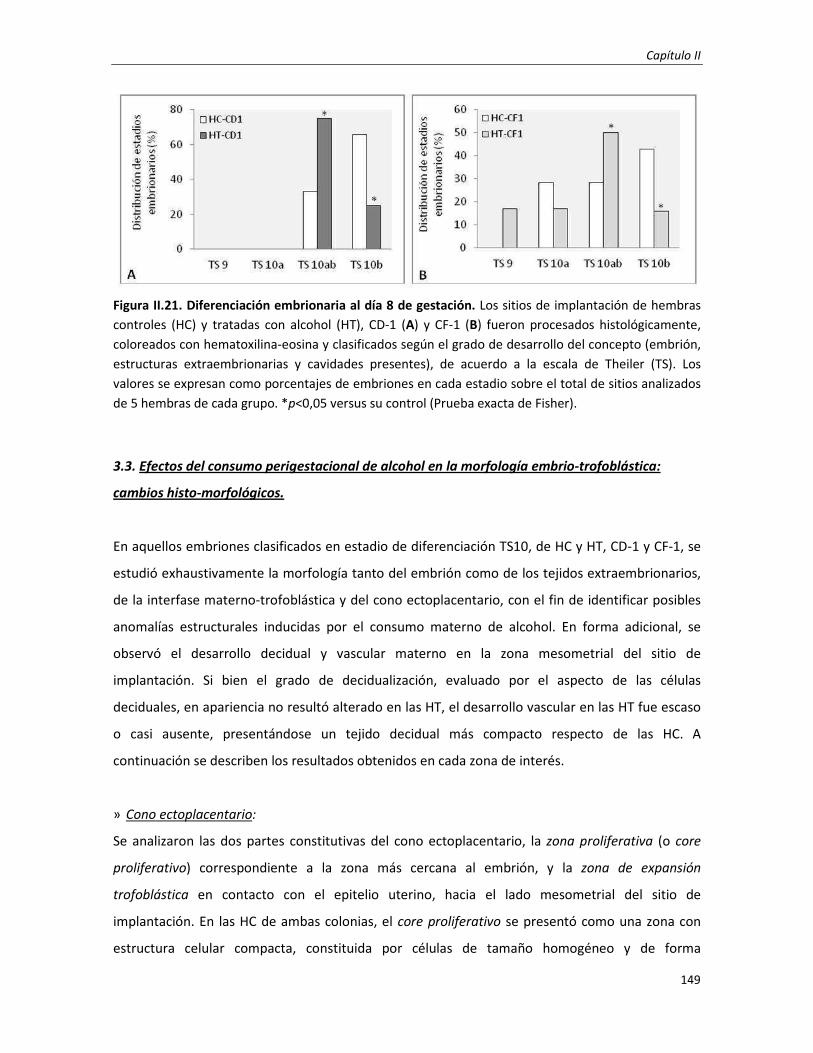
Capítulo II�
149
�
Figura II.21. Diferenciación embrionaria al día 8 de gestación. Los sitios de implantación de hembras
controles (HC) y tratadas con alcohol (HT), CD-1 (A) y CF-1 (B) fueron procesados histológicamente,
coloreados con hematoxilina-eosina y clasificados según el grado de desarrollo del concepto (embrión,
estructuras extraembrionarias y cavidades presentes), de acuerdo a la escala de Theiler (TS). Los
valores se expresan como porcentajes de embriones en cada estadio sobre el total de sitios analizados
de 5 hembras de cada grupo. *p<0,05 versus su control (Prueba exacta de Fisher).
3.3. Efectos del consumo perigestacional de alcohol en la morfología embrio-trofoblástica:
cambios histo-morfológicos.
En aquellos embriones clasificados en estadio de diferenciación TS10, de HC y HT, CD-1 y CF-1, se
estudió exhaustivamente la morfología tanto del embrión como de los tejidos extraembrionarios,
de la interfase materno-trofoblástica y del cono ectoplacentario, con el fin de identificar posibles
anomalías estructurales inducidas por el consumo materno de alcohol. En forma adicional, se
observó el desarrollo decidual y vascular materno en la zona mesometrial del sitio de
implantación. Si bien el grado de decidualización, evaluado por el aspecto de las células
deciduales, en apariencia no resultó alterado en las HT, el desarrollo vascular en las HT fue escaso
o casi ausente, presentándose un tejido decidual más compacto respecto de las HC. A
continuación se describen los resultados obtenidos en cada zona de interés.
» Cono ectoplacentario:
Se analizaron las dos partes constitutivas del cono ectoplacentario, la zona proliferativa (o core
proliferativo) correspondiente a la zona más cercana al embrión, y la zona de expansión
trofoblástica en contacto con el epitelio uterino, hacia el lado mesometrial del sitio de
implantación. En las HC de ambas colonias, el core proliferativo se presentó como una zona con
estructura celular compacta, constituida por células de tamaño homogéneo y de forma

Capítulo II�
150
�
redondeada, y sin glóbulos rojos maternos entre las mismas (Figura II.22.B). Por otro lado, la zona
de expansión trofoblástica se caracterizó por presentar células trofoblásticas dispersas, algunas de
las mismas, reconocidas por su mayor dimensión, en diferenciación o ya diferenciadas a CGTs 2rias.
Entre ellas, un moderado infiltrado de glóbulos rojos maternos fue frecuentemente observado
(Figura II.22.A). A diferencia de lo observado en las muestras controles, el core proliferativo de los
conos de las HT de ambas colonias, presentó un escaso desarrollo, siendo casi ausente en algunos
casos. En consonancia con esta observación, la zona de expansión trofoblástica también mostró un
menor desarrollo con un abundante infiltrado de glóbulos rojos maternos (Figura II.22.C).
Figura II.22. Morfología del cono ectoplacentario al día 8 de gestación. Micrografías representativas
de cortes transversales de sitios de implantación (hematoxilina-eosina) de hembra control (A y B) y
tratada con alcohol (C), CF-1. A, zona de expansión trofoblástica (ZE): células dispersas formando una
red entre los glóbulos rojos maternos; B, zona proliferativa (ZP): estructura celular compacta y sin
eritrocitos maternos; C, ZE con abundante infiltrado de glóbulos rojos (punta de flecha) y escasas
invasión trofoblástica, ZP con escaso desarrollo trofoblástico (*). Escala: 20 µm.
» Interfase trofoblástico-decidual:
Dada la relevancia que adquiere la interacción entre las células deciduales y las trofoblásticas para
establecer un adecuado diálogo materno-embrionario durante los estadios tempranos de invasión
trofoblástica (período postimplantatorio), se analizó la calidad morfológica de la interfase
trofoblástico-decidual a lo largo del perímetro lateral y antimesometrial del embrión. También se
evaluó la calidad histológica de la cripta antimesometrial de implantación, la abundancia de
glóbulos rojos en dicha zona, así como el grado de picnosis y fragmentación celular. En este
sentido, el 100% de los sitios de implantación de las HC de ambas colonias presentaron una cripta
antimesometrial con células poliédricas, moderada cantidad de fragmentos, escasa luz uterina,
moderada picnosis nuclear y escasa a moderada presencia de eritrocitos maternos; características
que también se hallaron en los laterales del embrión (Figura II.23.A). Este patrón morfológico

Capítulo II�
151
�
observado en las muestras analizadas de las HC, se tomó como criterio de una adecuada
remodelación de la interfase trofoblástico-decidual antimesometrial. Por el contrario,
aproximadamente, entre el 50 y 60% de las HT analizadas presentaron defectos en la
remodelación de la cripta de implantación, caracterizada por presentar excesiva picnosis nuclear,
abundante cantidad de fragmentos celulares, células de morfología irregular y gran cantidad de
glóbulos rojos en toda la interfase materno-embrionaria (Figura II.23.B). Tanto las anomalías
morfológicas como el número de muestras afectadas en las HT de ambas colonias, CD-1 y CF-1,
fueron similares.
Figura II.23. Morfología de la interfase trofoblástico-decidual al día 8 de gestación. Micrografías
representativas de cortes transversales de sitios de implantación (hematoxilina-eosina) de hembra
control (A) y tratada con alcohol (B), CF-1. A, zona antimesometrial de la interfase trofoblástico-
decidual (InT-DA) donde se observa escasa cantidad de glóbulos rojos (GR) entre el trofoblasto
embrionario y las células deciduales. B, en la misma zona se observa gran cantidad de glóbulos rojos
maternos (puntas de flecha). E: embrión, (*): CGT1ria. Escala: 20 µm.
» Tejidos extraembrionarios:
Entre el 100% (CD-1) y el 86% (CF-1) de las muestras controles analizadas se caracterizaron por
presentar un ectodermo extraembrionario conformado por una capa de tejido con células de
morfología cuboidal y columnar (Figura II.24.A y B). Por el contrario, entre el 75% (CD-1) y el 50%
(CF-1) de las muestras de HT analizadas, presentaron anomalías respecto del patrón descripto para
las HC. La capa se presentó desorganizada y con cambios morfológicos principalmente asociados a
alteraciones en la forma y tamaño de las células, las cuales, perdieron su característica forma
cuboidal para transformarse en células redondeadas con signos de poca adhesión celular (Figura
II.24.C y D). A su vez, se observó la presencia de fragmentos celulares proveniente de dicha capa,

Capítulo II�
152
�
hacia la cavidad exocelómica (Figura II.24.C). El mesodermo extraembrionario se caracterizó por
presentar, en el 100% de las muestras controles analizadas, células en disposición ordenada, con
formas poliédricas, y en algunos casos con leves prolongaciones citoplasmáticas que dan indicios
del proceso de migración de las mismas (Figura II.24.B). Sólo el 60% de las muestras analizadas de
las hembras tratadas mostraron dicho patrón. El 40% restante presentó alteraciones de forma y
tamaño, y disposición desorganizada (Figura II.24.D). El endodermo extraembrionario visceral, en
las muestras controles (85% en ambas colonias), se presentó células con morfología columnar y
vacuolas en la zona apical (Figura II.24.A). Sin embargo, el 100% (CD-1) y el 66% (CF-1) de las
muestras provenientes de HT, presentaron anomalías histológicas tales como una gran
desorganización, con núcleos deslocalizados (pérdida de su posición basal y localización central),
sumándose un aparente aumento en el número de vacuolas y/o aumento de tamaño de las
mismas (Figura II.24.D). En el endodermo parietal, a diferencia del visceral, caracterizado por
conformar una capa continúa de células planas, no se encontraron evidencias de alteraciones
significativas en las muestras de las HT analizadas respecto de las de las HC, en ninguna de las
colonias murinas.
» Embrión:
El ectodermo embrionario de las muestras de las hembras controles analizadas (100%, ambas
colonias) se caracterizó por presentar células columnares elongadas, con buena compactación y
disposición ordenadas (Figura II.25.A). Este patrón morfológico normal, sólo representó el 34% de
los sitios de implantación muestreados de las hembras tratadas. El porcentaje de sitios restante
presentó células con morfología alterada, con formas ahusadas y redondeadas, signos de adhesión
celular deficiente y distribución desorganizada (Figura II.25.B). El mesodermo embrionario de las
hembras controles (100% de las muestras analizadas) se presentó como una capa de células
poliédricas en migración, con procesos celulares (Figura II.25.A). En las muestras de las HT se
encontraron alteraciones respecto de dicho patrón morfológico, tales como, cambios en la forma y
tamaño celular (células ahusadas, pequeñas) conformando un tejido con desorganización y con
amplios espacios intercelulares (Figura II.25.B). El endodermo visceral de los embriones de las
controles se caracterizó por tener morfología celular cuboidal baja, sin vacuolas (100% de las
muestras controles de ambas colonias murinas). El 16% de las HT tuvo alteraciones de forma y
tamaño de las células (Figura II.25.B).

Capítulo II�
153
�
Figura II.24. Morfología de tejidos extraembrionarios al día 8 de gestación. Micrografías
representativas de cortes transversales de sitios de implantación (hematoxilina-eosina) de hembra
control (A-B) y tratada con alcohol (C-D), CF-1. Las HT presentaron alteraciones en la morfología y
organización histológica en las distintas capas extraembrionarias (puntas de flecha). EctoEx: ectodermo
extraembrionario, EndoP: endodermo parietal, EndoV: endodermo visceral, Me: mesodermo, Exocel:
exoceloma, Cav Ectopl: cavidad ectoplacentaria, Cav Exocel: cavidad exocelómica, CA: cavidad
amniótica, corion y amnios. Escala: 20 µm.
Figura II.25. Morfología del embrión en gastrulación al día 8 de gestación. Micrografías
representativas de cortes transversales de sitios de implantación (hematoxilina-eosina) de hembra
control (A) y tratada con alcohol (B), CF-1. Las HT presentaron alteraciones en la morfología y
organización celular en las distintas capas embrionarias (puntas de flecha). EctoE: ectodermo
embrionario, EndoP: endodermo parietal, EndoV: endodermo visceral, Me: mesodermo, CA: cavidad
amniótica. Escala: 20 µm.

Capítulo II�
154
�
3.4. Efecto del consumo perigestacional de alcohol hasta el día 8 de preñez en la viabilidad y
capacidad de desarrollo e invasión del cono ectoplacentario (estudio in-vitro).
Tanto la adecuada expansión como la diferenciación de las células trofoblásticas del cono
ectoplacentario son eventos claves para el desarrollo normal del embrión y de la placenta.
Alteraciones en la regulación de la dinámica de proliferación, migración y/o diferenciación de esta
estructura derivada del trofoectodermo polar del blastocisto implantado, determinarán cambios en
la tasa de implantación y, de esta manera, podrán constituirse en causas de pérdida gestacional
manifestada como reabsorción embrionaria temprana. En este marco, el objetivo siguiente fue
profundizar el análisis de la viabilidad, la capacidad de desarrollo, el grado de crecimiento, la
expansión media y el estado de diferenciación del cono ectoplacentario de las hembras tratadas en
comparación con las controles. Dado que encontramos anomalías histo-morfológicas en el cono
ectoplacentario de las hembras tratadas CD-1 y CF-1 de manera similar, para este objetivo, se
realizaron cultivos de explantes de conos ectoplacentarios utilizando el modelo murino de
exposición in-vivo a alcohol en CD-1.
3.4.1. Viabilidad y capacidad de desarrollo del cono ectoplacentario.
Para estudiar la viabilidad y capacidad de desarrollo de los conos ectoplacentarios de las hembras
controles y tratadas, se cuantificó el número de conos adheridos a la placa a las 24 hs de cultivo
(conos viables) sobre el total de conos colocados en cultivo a T0 hs, así como, el número de conos
que presentaron expansión trofoblástica entre las 24 y 48 hs (conos con capacidad de desarrollo)
sobre el total de conos adheridos al sustrato a T24 hs de cultivo (Tabla II.7). Como puede
observarse, el porcentaje de conos viables (adheridos a T24 hs) provenientes de hembras
expuestas a alcohol disminuye significativamente en comparación con el porcentaje de conos
adheridos provenientes de hembras controles (p<0,05).

Capítulo II�
155
�
Tabla II.7. Viabilidad y capacidad de desarrollo de conos ectoplacentarios en cultivo.
HC HT
N° total de conos ectoplacentarios en cultivo (T0 hs) 66 90
N° conos adheridos a las 24 hs de cultivo 59 70
N° conos adheridos a las 24 hs/conos totales (%) 59/66 (89,4) 70/90 (77,8)*
Monitoreo de la viabilidad y capacidad de desarrollo in-vitro de conos ectoplacentarios de hembras
controles (HC) y tratadas con alcohol (HT), CD-1. Los valores se expresan como frecuencias
(porcentajes, %). *p<0,05 versus su control (Prueba Chi-cuadrado).
Se evaluó la tasa de desarrollo entre las 24 y 48 hs de cultivo. En la Tabla II.8 se muestra el
recuento de los conos que presentaron o no desarrollo trofoblástico tanto a las 24 como a las
48 hs. Se observó que a las 24 hs la proporción de conos sin desarrollo trofoblástico resultó
mayor en las HT respecto de las HC (p<0,05). Sin embargo, aproximadamente el 50% de los
conos de las HC y de las HT, que a las 24 hs no mostraban desarrollo trofoblástico, a las 48 hs
de cultivo lograron desarrollarse (HC: 10/20; HT: 22/39). Con lo cual, se ve que una proporción
de los conos de las HT que no presentaban desarrollo trofoblástico a las 24 hs al cabo de las 48
hs lograron compensar la diferencia de las 24 hs y equiparar el porcentaje de conos con
desarrollo con los valores del control (Tabla II.8). De este análisis, a su vez, se desprende que
existe un porcentaje de conos sin capacidad de desarrollo trofoblástico entre las 24 y las 48 hs,
resultando dicho valor aumentado para las HT (HC: 16,9% (10/59) vs HT: 24% (17/70), p<0,05).
Tabla II.8. Desarrollo trofoblástico de conos ectoplacentarios a las 24 y 48 hs de cultivo.
24 hs 48 hs
HC HT HC HT
N°conos adheridos 59 70 59 70
N°conos sin desarrollo/conos adh (%) 20/59 (33,9) 39/70 (55,7)* 10/59 (16,9) 17/70 (24,3)
N°conos con desarrollo/conos adh (%) 39/59 (66,1) 31/70 (44,3)* 49/59 (83,0) 53/70 (75,7)
Monitoreo del desarrollo trofoblástico de cono ectoplacentario in-vitro de hembras controles (HC)
y tratadas con alcohol (HT), CD-1, cultivados por 48 hs. Adh: adheridos. Los valores se expresan
como frecuencias (porcentajes, %). *p<0,05 versus su control (Prueba Chi-cuadrado).

Capítulo II�
156
�
3.4.2. Crecimiento de los conos ectoplacentarios en cultivo.
Con el fin de estudiar el crecimiento de los conos ectoplacentarios, los conos viables (adheridos a
la placa de cultivo) y que presentaron desarrollo trofoblástico entre las 24 y 48 hs fueron
evaluados morfométricamente a través de la medición del área total de expansión trofoblástica.
Dada la gran variabilidad de tamaños observados, los conos fueron categorizados en: conos
pequeños, medianos y grandes, para cada tiempo de cultivo. La Tabla II.9 muestra la proporción
de conos en cada categoría, sobre el total de conos con desarrollo trofoblástico. La distribución de
los tipos de conos resultó similar entre el grupo de HC y HT para cada tiempo de cultivo.
Tabla II.9. Proporción de conos pequeños, medianos y grandes a las 24 y 48 hs de cultivo.
24 hs 48 hs
HC HT HC HT
N° conos con desarrollo trofoblástico 39 31 49 53
N° conos pequeños (%) 16/39 (41,0) 17/31 (54,8) 9/49 (18,3) 8/53 (15,1)
N° conos medianos (%) 16/39 (41,0) 10/31 (32,2) 31/49 (63,3) 37/53 (69,8)
N° conos grandes (%) 7/39 (17,9) 4/31 (12,9) 10/49 (20,4) 8/53 (15,1)
Distribución de los tipos de conos ectoplacentarios desarrollados in-vitro provenientes de hembras
controles (HC) y tratadas con alcohol (HT), CD-1, cultivados por 48 hs. Los conos fueron clasificados
de acuerdo a su tamaño en conos pequeños: con escasa expansión; conos medianos: con moderada
expansión; conos grandes: con gran expansión trofoblástica. Los valores se expresan como
frecuencias (porcentajes, %) (Prueba Chi-cuadrado).
En la Figura II.26 se muestran micrografías de los tipos de conos (según el área total de expansión
trofoblástica) en cada tiempo de cultivo. Tanto a T24 y T48 hs, pueden observarse conos
ectoplacentarios sin desarrollo trofoblástico (Figura II.26.A y B), conos pequeños, con incipiente
desarrollo trofoblástico (Figura II.26.C y D), conos medianos, con moderado desarrollo
trofoblástico (Figura II.26.E y F) y conos grandes, con gran desarrollo trofoblástico (Figura II.26.G y
H).

Capítulo II�
157
�
Figura II.26. Desarrollo de conos ectoplacentarios en cultivo. Micrografías con contraste de fase
representativas de conos cultivados por 24 hs (A, C, E, G) y por 48 hs (B, D, F, H), clasificados según el
grado de expansión del trofoblasto. Nótese la monocapa de células trofoblásticas (m) formando una
red alrededor del core proliferativo (c), los filopodios (puntas de flecha) y las CGT diferenciadas (*). A y
B, conos adheridos a la placa sin desarrollo trofoblástico; C y D, conos pequeños; E y F, conos medianos;
G y H, conos grandes. Aumento: 10x.

Capítulo II�
158
�
Se determinó el grado de expansión trofoblástica media de los tipos de conos por medición, en
micrografías con contraste de fase, del área del cono en su totalidad (área total) y del área de la
masa de células centrales (área del core proliferativo), y luego se calculó el área de expansión de la
monocapa de células trofoblástica como la diferencia entre el área total y el área del core. A su
vez, fue calculada la relación entre el área ocupada por el core proliferativo en función del área
total. Todos los valores de las áreas promedio fueron calculados para cada tipo de cono a las 24 y
48 hs de cultivo, datos que se muestran en la Tabla II.10.
Según el comportamiento observado en los conos de las HC, el aumento del área total de los
conos cultivados in-vitro es un proceso dinámico que ocurre a expensas tanto de las células del
core proliferativo como de la expansión de la monocapa de células trofoblásticas, involucrando
procesos de proliferación celular, migración o invasión trofoblástica, así como, la diferenciación de
dichas células en CGTs, las cuales, al aumentar notablemente sus dimensiones, contribuyen
también al incremento del área de expansión. Dichos procesos, si bien ocurren de manera
simultánea, generalmente existen preponderancias de unos sobre otros según el momento de
desarrollo. En los tres tipos de conos de las HC, se observó una disminución de la relación área
core/área total entre las 24 y 48 hs (disminución en función del tiempo de cultivo). Esto indica que
el core disminuye su tamaño a medida que aumenta el área total y el área de expansión
trofoblástica, y/o que el core permanece de igual tamaño pero que el área total aumenta. Sin
embargo, en los conos de las HT este comportamiento se observó parcialmente desregulado. A
continuación se describe, el comportamiento in-vitro para cada tipo de cono y según el tiempo de
cultivo (Tabla II.10).
Los conos pequeños de las HT a las 24 hs de cultivo presentan un área total, área del core y área de
expansión trofoblástica significativamente disminuida respecto del control. Es decir, todo el cono
en su conjunto es menor respecto de cono del control, y por tal motivo la relación de áreas
(core/total) se mantiene constante. Tanto el core de las HC como el de las HT, ocupa
aproximadamente un 30% del área total. Ya a las 48 hs, este patrón observado para los conos de
las HT, se invierte, presentando una área total y de expansión trofoblástica significativamente
mayor respecto a lo observado en los conos controles, sin embargo alcanzando un tamaño de core
similar al observado en los controles. Por este motivo en este caso la relación área core/área total
resulta significativamente disminuida (el core, ocupa una menor proporción del área total de
cono). En otras palabras, el cono pequeño de las HT, a las 48 hs estaría compensando el tamaño
disminuido de las 24 hs, exacerbando su crecimiento a expensas fundamentalmente del área de

Capítulo II�
159
�
expansión trofoblástica, superando así el tamaño total promedio de los conos controles.
Los conos medianos de las HT a las 24 hs de cultivo presentaron un área total y un área de
expansión trofoblástica aumentada significativamente respecto del control, sin diferencias
significativas en el tamaño alcanzando por el core, motivo por el cual, la relación área core/área
total resultó significativamente disminuida (el core ocupa una menor proporción del área total de
cono de HT). A las 48 hs, este patrón observado para los conos de las HT, como ocurrió para los
conos pequeños, se invierte respecto a lo observado a las 24 hs, presentando en este caso, un área
total y de expansión trofoblástica significativamente disminuidas, alcanzando sin embargo un
tamaño de core similar al observado en los controles, motivo por el cual la relación área core/área
total resulta, en este caso, significativamente aumentada (el core ocupa una mayor proporción del
área total de cono). Es decir, el cono mediano de las HT, a las 48 hs estaría compensando el
tamaño aumentado a las 24 hs frenando en cierta forma su crecimiento a expensas
fundamentalmente de área de expansión trofoblástica resultando, este área, disminuida respecto
del tamaño promedio observado en los conos controles.
A diferencia de las alteraciones observadas en la expansión in-vitro de los conos pequeños y
medianos de las HT en relación a los de las controles, los conos grandes de las HT, tanto a las
24 como a las 48 hs de cultivo, presentaron un comportamiento semejante al observado en los
controles. La única diferencia significativa surgió en la relación área core/área total, la cual
resultó disminuida a las 48 hs en los conos de HT. En otras palabras, la proporción ocupada por
el core (respecto del total) en los conos de las hembras tratadas es del 6%, proporción
ligeramente disminuida respecto del control (7%).
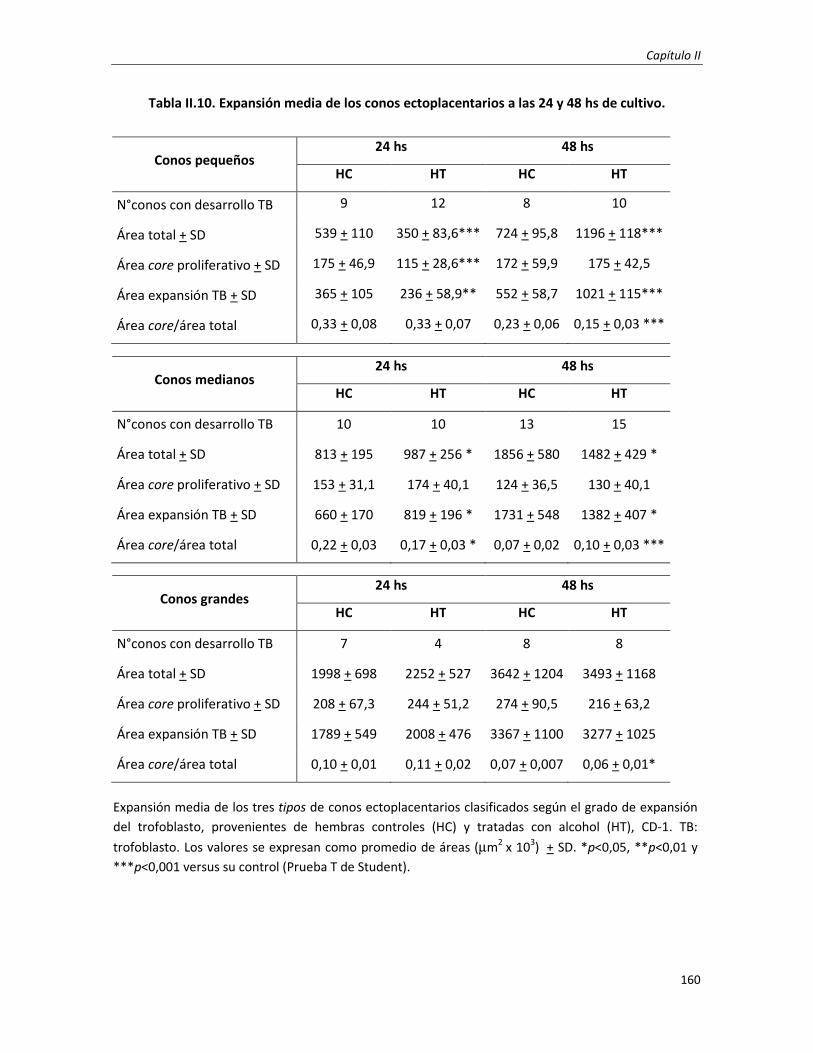
Capítulo II�
160
�
Tabla II.10. Expansión media de los conos ectoplacentarios a las 24 y 48 hs de cultivo.
Conos pequeños 24 hs 48 hs
HC HT HC HT
N°conos con desarrollo TB 9 12 8 10
Área total + SD 539 + 110 350 + 83,6*** 724 + 95,8 1196 + 118***
Área core proliferativo + SD 175 + 46,9 115 + 28,6*** 172 + 59,9 175 + 42,5
Área expansión TB + SD 365 + 105 236 + 58,9** 552 + 58,7 1021 + 115***
Área core/área total 0,33 + 0,08 0,33 + 0,07 0,23 + 0,06 0,15 + 0,03 ***
Conos medianos 24 hs 48 hs
HC HT HC HT
N°conos con desarrollo TB 10 10 13 15
Área total + SD 813 + 195 987 + 256 * 1856 + 580 1482 + 429 *
Área core proliferativo + SD 153 + 31,1 174 + 40,1 124 + 36,5 130 + 40,1
Área expansión TB + SD 660 + 170 819 + 196 * 1731 + 548 1382 + 407 *
Área core/área total 0,22 + 0,03 0,17 + 0,03 * 0,07 + 0,02 0,10 + 0,03 ***
Conos grandes 24 hs 48 hs
HC HT HC HT
N°conos con desarrollo TB 7 4 8 8
Área total + SD 1998 + 698 2252 + 527 3642 + 1204 3493 + 1168
Área core proliferativo + SD 208 + 67,3 244 + 51,2 274 + 90,5 216 + 63,2
Área expansión TB + SD 1789 + 549 2008 + 476 3367 + 1100 3277 + 1025
Área core/área total 0,10 + 0,01 0,11 + 0,02 0,07 + 0,007 0,06 + 0,01*
Expansión media de los tres tipos de conos ectoplacentarios clasificados según el grado de expansión
del trofoblasto, provenientes de hembras controles (HC) y tratadas con alcohol (HT), CD-1. TB:
trofoblasto. Los valores se expresan como promedio de áreas (µm2 x 103) + SD. *p<0,05, **p<0,01 y
***p<0,001 versus su control (Prueba T de Student).

Capítulo II�
161
�
3.4.3. Morfología de los conos ectoplacentarios en cultivo.
La morfología del trofoblasto invasivo (zona de expansión del cono ectoplacentario) fue analizado
durante el cultivo. A T24 hs, los conos adheridos a la placa evidenciaron desarrollo trofoblástico
periférico, alrededor de la masa de células centrales. Entre las 24 y las 48 hs, los conos formaron
una estructura que se organizó como una red de células trofoblásticas en forma de monocapa
expandida alrededor de un aglomerado de células generalmente centrales, de pequeñas
dimensiones y esféricas, superpuestas unas sobre otras. En comparación con las células centrales,
las células trofoblásticas en la zona de expansión exhibieron dimensiones celulares y nucleares
mayores, algunas de ellas presentaron las características típicas de las células trofoblásticas
gigantes, con 1 a 4 nucléolos de gran tamaño y proyecciones citoplasmáticas (filopodios) hacia la
zona externa del área de expansión. Generalmente, la expansión de esta monocapa de células
trofoblásticas ocurre de manera polarizada, es decir, las células avanzan en conjunto hacia un
mismo lado (Figura II.27). De acuerdo a estas observaciones, la morfología de los conos
ectoplacentarios en cultivo provenientes de HT no difirió de la morfología observada en los conos
de HC. Tampoco se registró lisis o fragmentación celular en las muestras de las HT, durante los
tiempos de cultivo estudiados.
3.4.4. Diferenciación celular en los conos ectoplacentarios en cultivo.
Dado que la expansión de la monocapa trofoblástica del cono ectoplacentario se vincula, en parte,
con la diferenciación trofoblástica (transformación a CGTs), se evaluó esta posibilidad a través de
la determinación de la incorporación de BrdU a las 48 hs de cultivo, como marcador de la
presencia de células trofoblásticas en diferenciación a CGT (que se encuentran en activa síntesis
de ADN: endorreduplicación) (Figura II.28). Este estudio reveló marcación positiva para BrdU tanto
en las células de pequeñas dimensiones presentes en la región central del cono (Figura II.28.A)
como en la monocapa de células trofoblásticas expandidas (Figura II.28.B). Como la marcación del
aglomerado central podría estar asociada también al proceso de proliferación celular, dicha región
no fue considerada en el análisis, el cual sólo se focalizó en la región periférica. En tal sentido,
luego de cuantificar y comparar el número de células BrdU-positivas en el área de expansión
trofoblástica entre los conos de HC y HT, no se hallaron diferencias significativas entre sí.

Capítulo II�
162
�
Figura II.27. Morfología de conos ectoplacentarios en cultivo. Micrografías con contraste de fase
representativas de conos cultivados por 24 hs (A, B, C) y por 48 hs (D, E, F). Notar la red de células
trofoblásticas expandida formando una monocapa (m) alrededor de un aglomerado central de células
pequeñas y superpuestas (core, c). En la periferia se observan células con filopodios (puntas de flecha),
células en migración (flechas) y células gigantes trofoblásticas (*). Aumento: 20x.
Figura II.28. Inmunolocalización de BrdU en conos ectoplacentarios en cultivo. Micrografías
representativas de conos cultivados por 48 hs y sometidos a inmunohistoquímica para BrdU. La
marcación positiva de las células trofoblásticas periféricas, es un indicador del proceso de
diferenciación a CGT. A y B, células centrales (*) y periféricas (puntas de flecha) exhiben reactividad
positiva para BrdU; C, control negativo en el cual se omitió el anticuerpo primario. Aumento: 20x.

Capítulo II�
163
�
II.3. CONCLUSIONES PARCIALES
1. En cuanto al efecto de la ingesta pregestacional de alcohol en el desarrollo in-vivo de
embriones de preimplantación (día 4 de gestación), en hembras CF-1 y CD-1, se puede concluir
que:
Si bien no se registró un impacto negativo en el número de embriones provenientes de hembras
tratadas de ambas colonias, las HT-CF-1 presentaron elevada tasa de fragmentación embrionaria.
Tanto las hembras CD-1 como CF-1, mostraron retraso de diferenciación de la población
embrionaria, aumento de blastocistos morfológicamente anormales y crecimiento de blastocistos
iniciales disminuido con desregulación de la división celular. Estos resultados fueron similares en
ambas colonias murinas.
2. En cuanto al efecto de la ingesta perigestacional de alcohol en el desarrollo embrio-
trofoblástico durante la implantación (día 5 de gestación) en hembras CD-1, se puede concluir
que:
El patrón de alteraciones en el desarrollo embrionario fue similar al encontrado en el día 4 de
gestación, aunque más pronunciadas, viéndose desregulación de la dinámica de diferenciación y
crecimiento de los embriones junto con anomalías en el grado de expansión/invasión trofoblástica
en la implantación.
Estudios in-vivo (inicio de la implantación):
Se evidenció un impacto negativo de la ingesta de alcohol por reducción del número de embriones
recuperados al día 5 de gestación, retraso en la diferenciación embrionaria, disminución de la
frecuencia de embriones en estadio implantativo más avanzado (tipo 3), y aumento del número de
embriones morfológicamente anormales.
Estudios in-vitro (capacidad implantativa):
Se evidenció retraso de diferenciación embrionaria durante la fase tardía de la implantación, por
reducción en el número de embriones tipo 3 (avanzados) a las 72 hs. A su vez, se observaron
alteraciones en invasividad trofoblástica durante la implantación, detectándose una mayor tasa de

Capítulo II�
164
�
expansión trofoblástica en la población de embriones pequeños (tipo 1) y embriones medianos
(tipo 2), respecto de los mismos tipos embrionarios provenientes de hembras controles,
sugiriendo esto, una posible estimulación de la invasividad trofoblástica inducida por la exposición
a alcohol.
3. En cuanto al efecto de la ingesta perigestacional de alcohol en el desarrollo embrio-
trofoblástico durante la fase de gastrulación (día 8 de gestación), en hembras CF-1 y CD-1, se
puede concluir que:
Estudios in-vivo:
Se evidenció retraso del desarrollo embrionario (diferenciación), entre 8 y 24 hs, similar al patrón
observado en estudios previos en la etapa de organogénesis (día 10). Las anomalías histo-
morfológicas en los tejidos embrio-trofoblásticos, incluyeron: conos ectoplacentarios con escaso o
ausente desarrollo de la zona proliferativa y con zona de expansión trofoblástica pequeña y con
abundante infiltrado de glóbulos rojos maternos; interfase trofoblástico-decidual con alteraciones
histológicas asociada con remodelación anormal de la cripta de implantación (excesiva picnosis
nuclear, abundante cantidad de fragmentos celulares, gran cantidad de glóbulos rojos); tejidos
extraembrionarios y embrionarios con anomalías histológicas (desorganización celular y cambios
morfológicos celulares).
Estudios in-vitro (capacidad invasiva):
Además de una reducida viabilidad, los conos de hembras tratadas presentaron una menor
capacidad de desarrollo entre las 24 y las 48 hs. La dinámica de expansión del trofoblasto de los
conos pequeños y medianos resultó variable y desregulada durante los tiempos del cultivo, con
patrones de desarrollo opuestos al de los conos de las hembras controles. Sin embargo, la
dinámica de expansión de conos grandes no fue afectada por la exposición a alcohol. La similar
tasa de incorporación de BrdU a T48 hs en las células trofoblásticas periféricas del cono de las
hembras tratadas respecto de las controles indicó que la diferenciación de estas células no fue
afectada por la ingesta de alcohol, por lo que este parámetro no estaría involucrado en el
crecimiento alterado observada en los conos de HT.

Capítulo II�
165
�
Por lo tanto, en conjunto se concluye que la administración por vía oral, de etanol 10%, en ratones
CD-1 y CF-1, desde antes de la gestación y hasta los días 4, 5 u 8 de gestación, induce anomalías
embrionarias y trofoblásticas que llevan a retraso de la diferenciación, dismorfogénesis
embrionaria y arresto del desarrollo periimplantatorio, alteraciones que en consecuencia, podrían
vincularse con la pérdida gestacional temprana observada en el período organogénico.


Capítulo III�
166
�
Consumo perigestacional de alcohol:
efectos en las uniones intercelulares, metaloproteasas y genotoxicidad en
el tejido embrio-trofoblástico periimplantatorio murino.
III.1. MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo.
Se utilizaron hembras y machos adultos pertenecientes a las colonias CD-1 y CF-1, en los que se
realizaron los esquemas de apareamiento correspondientes a cada colonia, como fue detallado en
Materiales y métodos. Brevemente, se describen a continuación.
1.1. Tratamiento perigestacional con alcohol, sin régimen de estimulación ovárica.
Hembras CD-1 recibieron etanol 10% (m/v) en el agua de bebida 15 días previos al apareamiento y
hasta el día 4 u 8 de gestación (hembras tratadas, HT). El grupo control (hembras controles, HC)
recibió sólo agua como bebida. Las hembras fueron alojadas con machos para el apareamiento
entre los días 15 y 17 de tratamiento. Al día siguiente, la observación del tapón vaginal indicó el
día 1 de gestación. Las hembras con tapón vaginal continuaron recibiendo alcohol (HT) o agua (HC)
hasta el día 4 u 8 de preñez. En la tarde del día 4 de gestación (entre las 13:00 y 16:00 hs) o a la
mañana (entre las 9:00 y 12:00 hs) del día 8 de gestación, las HC y HT fueron sacrificadas por
dislocación cervical para extraer los embriones o sitios de implantación para realizar los estudios
que se detallan a continuación (Figura III.1). Se utilizó un mínimo de 5 animales por grupo.

Capítulo III�
167
�
Figura III.1. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, sin estimulación ovárica. Hembras CD-1 recibieron etanol 10% (hembras tratadas, HT) durante
15 días previos al apareamiento. Las hembras controles (HC) recibieron agua durante el mismo
período. Las hembras fueron puestas en contacto con el macho en días consecutivos y hasta 3 veces,
entre los días 15 y 17 de tratamiento. Cada mañana fue observada la presencia del tapón vaginal. Las
hembras con tapón (día 1 de gestación) continuaron recibiendo alcohol (HT) o agua (HC) hasta el día 4
(D4) u 8 (D8) de gestación, días en que fueron sacrificadas para realizar los estudios indicados en el
esquema.
1.2. Tratamiento perigestacional con alcohol, con régimen de estimulación ovárica.
Hembras CF-1 recibieron etanol 10% (m/v) en el agua de bebida 17 días previos al apareamiento y
hasta el día 4 u 8 de gestación (hembras tratadas, HT). El grupo control (hembras controles, HC)
recibió sólo agua como bebida. Al día 15 y 17 de tratamiento con alcohol, tanto en las HT como en
las HC, se indujo la ovulación con gonadotrofinas (eCG/hCG). Al día 17 de tratamiento, las hembras
fueron alojadas con machos para el apareamiento, y la observación del tapón vaginal al día
siguiente indicó el día 1 de gestación. Las hembras con tapón vaginal continuaron recibiendo
alcohol (HT) o agua (HC) hasta el día 4 u 8 de preñez. En la tarde del día 4 de gestación (entre las

Capítulo III�
168
�
13:00 y 16:00 hs, 96-99 hs post-hCG) o en la mañana del día 8 de gestación (entre las 9:00 y 12:00
hs), las HC y HT fueron sacrificadas por dislocación cervical para extraer los embriones o sitios de
implantación para realizar los estudios que se detallan a continuación (Figura III.2).
Figura III.2. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, con estimulación ovárica. Hembras CF-1 recibieron etanol 10% (hembras tratadas, HT)
durante 15 días previos al apareamiento. Las hembras controles (HC) recibieron agua el mismo
período. Al día 15 y 17 de tratamiento se indujo la ovulación con eCG y hCG, y luego del apareamiento,
la presencia de tapón vaginal indicó el día 1 de gestación (D1). Las hembras con tapón continuaron
recibiendo alcohol (HT) o agua (HC) hasta el día 4 (D4) u 8 (D8) de gestación, días en que fueron
sacrificadas para realizar los estudios que indicados en el esquema.

Capítulo III�
169
�
2. Recolección y procesamiento de embriones, sitios de implantación, conos ectoplacentarios y
médula ósea materna.
2.1. Recolección y procesamiento de embriones de preimplantación (día 4 de gestación).
A la tarde del 4to. día de gestación, las hembras de la colonia CF-1 fueron sacrificadas y la cavidad
abdominal fue cuidadosamente abierta. El útero fue rápidamente removido y colocado en una
placa de Petri con medio Krebs-Ringer. Los cuernos uterinos fueron separados de los oviductos y
del cérvix, y luego los embriones fueron extraídos por flushing del útero con medio Krebs-Ringer -
BSA 0,3% utilizando aguja de 25 G acoplada a una jeringa de 1 ml. Los embriones se recolectaron
del líquido de flushing con un microcapilar de vidrio acoplado a una pipeta bucal. Los embriones
recuperados fueron lavados sucesivas veces en microgotas de medio Krebs-Ringer - BSA 0,3%.
Luego observados bajo microscopio invertido con contraste de fase, se clasificaron según estadios
embrionarios en mórulas, blastocistos iniciales, blastocistos expandidos o blastocistos extruidos de
la zona pelúcida. Un grupo de blastocistos iniciales y expandidos fue inmediatamente fijado en
paraformaldehído 2% por 20 min a temperatura ambiente (TA), lavado en PBS - BSA 0,3% y
almacenado a 4°C en microgotas de PBS cubierta con aceite mineral, hasta la realización de la
inmunohistoquímica (para el estudio de moléculas de adhesión) y el ensayo de TUNEL (para el
estudio de la muerte celular). Otro grupo de embriones (mórulas, blastocistos iniciales y
expandidos) provenientes de hembras CD-1 como CF-1, fue procesado según la técnica de
Tarkowski (véase Métodos del Capítulo II), para estudios de genotoxicidad embrionaria.
2.2. Recolección y procesamiento de sitios de implantación (día 8 de gestación).
Al 8vo. día de gestación, las hembras de las colonias CD-1 y CF-1 fueron sacrificadas y la cavidad
abdominal cuidadosamente abierta para extraer los cuernos uterinos, como fue descripto en el
Capítulo II (véase Métodos). Los sitios de implantación fueron fijados en paraformaldehído 4%,
deshidratados y embebidos en parafina para luego realizar cortes transversales al eje longitudinal
del útero de 5 µm de espesor. Luego de la obtención de cortes con las secciones embrionarias más
centrales, estos fueron adheridos a portaobjetos con Poli-L-lisina. Luego de colorear con
hematoxilina-eosina las secciones seriadas recolectadas, se seleccionaron sitios de implantación
con embrión desarrollado en los subestadios TS10a, TS10ab o TS10b (véase Métodos del Capítulo

Capítulo III�
170
�
II). Así, sólo aquellas secciones correspondientes al estadio embrionario esperado al día 8 de
gestación fueron utilizadas para los estudios de las moléculas de adhesión, inmunomarcación de
metaloproteasas, estudios nucleares y ensayo de TUNEL.
2.3. Recolección y procesamiento de conos ectoplacentarios (día 8 de gestación).
Al 8vo. día de gestación, HT y HC de la colonia CD-1 fueron sacrificadas por dislocación cervical. Los
cuernos uterinos fueron rápidamente removidos y lavados en PBS estéril con SFB 10%. Luego
fueron colocados sobre una placa de parafina y mediante un corte longitudinal desde un extremo
del cuerno hacia el otro extremo, el miometrio fue completamente aislado y descartado. Cada
decidua aislada fue transferida a una placa de Petri con PBS a 4°C. Bajo estereomicroscopio, y con
ayuda de instrumental oftalmológico fue aislado de la decidua, cuidadosamente, el concepto junto
con el cono ectoplacentario. Finalmente, con microbisturí se separó el cono del embrión (véase
Figura II.7 del Capítulo II). Todos los conos disecados fueron lavados en placas con PBS estéril a
4°C, tras lo cual fueron colectados en tubos Eppendorf con PBS estéril a 4°C y mantenidos en hielo
hasta comenzar la lisis del tejido. De cada hembra se colectaron tres tubos con tres conos
ectoplacentarios cada uno (aproximadamente 1500 células por tubo). Dichas muestras fueron
utilizadas en estudios de expresión génica de metaloproteasas, como se detallará más adelante.
2.4. Recolección y procesamiento de médula ósea.
Al 4to. día de gestación, luego del tratamiento perigestacional con alcohol, hembras HC y HT
preñadas de la colonia CF-1 fueron sacrificadas por dislocación cervical para disección de la
médula ósea. Para ello, ambos fémures fueron extraídos y luego de la remoción de tejidos
blandos, ambas apófisis fueron cortadas con bisturí. La médula ósea de ambos huesos
(proveniente de un mismo animal) fue colectada en un tubo cónico por inyección de medio F10-
SFB dentro de la cavidad ósea con jeringa de 1 ml. Luego de unos minutos, la suspensión de células
de médula ósea fue centrifugada a 1500g durante 5 min. El sobrenadante fue descartado y pellet
celular fue resuspendido en el líquido residual para realizar extendidos celulares en portaobjetos
(3 extendidos por hembra). Luego del secado al aire, las muestras fueron fijadas en alcohol 70%
durante 15 min y coloreadas con una solución de Giemsa en agua destilada (1:10) durante 10 min.
Dichas muestras fueron utilizadas para estudios de genotoxicidad materna.

Capítulo III�
171
�
3. Metodologías empleadas.
3.1. Metodologías para el estudio de moléculas de adhesión celular en embriones en el período
periimplantatorio: Día 4 y 8 de gestación.
3.1.1. Inmunofluorescencia para ZO-1 en blastocistos en suspensión.
Los blastocistos iniciales y expandidos fijados fueron permeabilizados por 30 min con una solución
de PBS - Tritón X-100 0,25% (PBST) e incubados por 30 min en solución de bloqueo PBST 0,25% -
BSA 5%. Luego de los lavados en PBST 0,25%, fueron incubados con anticuerpo primario anti ZO-1-
FITC diluido 1:80 en PBST 0,01% - BSA 5%, overnight a 4°C. Luego de los lavados con PBST 0,25%,
los embriones fueron incubados con anticuerpo secundario anti FITC biotinilado diluido 1:100 en
PBST 0,01% - BSA 5%, 1 h a TA. Posteriormente fueron lavados en PBST 0,25% e incubados con
neutral avidine FITC diluido 1:200 en PBST 0,01% - BSA 5%, 45 min a TA. Transcurrido ese tiempo,
los embriones fueron nuevamente lavados en PBST 0,25%, montados cuidadosamente en gotas de
glicerol - PBS (1:1) sobre portaobjeto y cubiertos con cubreobjetos con soportes de plastilina para
evitar la compresión de los blastocistos. Los embriones fueron observados y fotografiados bajo
microscopio de fluorescencia y microscopio confocal para la evaluación de la distribución y patrón
de expresión de la inmunomarca. El control de especificidad de la reacción fue realizado omitiendo
el anticuerpo primario.
3.1.2. Inmunofluorescencia para E-cadherina en blastocistos en suspensión.
Los blastocistos iniciales y expandidos fijados fueron permeabilizados y bloqueados como fue
explicado en el punto anterior. Luego de los lavados en PBST 0,25% fueron incubados con
anticuerpo primario mouse anti E-cadherina diluido 1:80 en PBST 0,01% - BSA, overnight a 4°C.
Luego de los lavados con PBST 0,25%, los embriones fueron incubados con anticuerpo secundario
donkey anti mouse Rodamina diluido 1:75 en PBST 0,01% - BSA 5%, 1 h a TA. Posteriormente
fueron lavados en PBST 0,25% y montados en gotas de glicerol - PBS (1:1), entre porta y
cubreobjestos como fue explicado en el punto anterior. Los blastocistos fueron observados y
fotografiados bajo microscopio de fluorescencia y confocal para la evaluación de la distribución y

Capítulo III�
172
�
patrón de expresión de la inmunomarca. El control de especificidad de la reacción fue realizado
omitiendo el anticuerpo primario.
3.1.3. Doble inmunofluorescencia para ZO-1 y E-cadherina en blastocistos en suspensión.
Los blastocistos iniciales y expandidos fijados fueron permeabilizados y bloqueados como fue
anteriormente explicado. Luego de los lavados en PBST 0,25% fueron incubados con anticuerpo
primario anti-ZO-1-FITC diluido 1:80 en PBST 0,01% - BSA 5%, overnight a 4°C. Al día siguiente,
luego de los lavados con PBST 0,25%, los embriones fueron incubados con anticuerpo secundario
anti FITC biotinilado diluido 1:100 en PBST 0,01% - BSA 5%, 1 h a TA. Posteriormente fueron
lavados en PBST 0,25% e incubados con neutral avidine FITC diluido 1:200 en PBST 0,01% - BSA 5%,
45 min a TA. Trascurrido ese tiempo, los embriones fueron lavados e incubados con anticuerpo
primario mouse anti E-cadherina diluido 1:80 en PBST 0,01% - BSA 5%, overnight a 4°C. Al día
siguiente, luego de los lavados con PBST 0,25%, los embriones fueron incubados con anticuerpo
secundario donkey anti mouse Rodamina 1:75 en PBST 0,01% - BSA 5%, 1 h a TA. Posteriormente
fueron lavados en PBST 0,25%, y montados en gotas de glicerol - PBS (1:1) entre porta y
cubreobjetos. Finalmente, lo embriones fueron observados y fotografiados bajo microscopio de
fluorescencia y confocal para la evaluación del patrón de co-localización de ambas moléculas. El
control de especificidad de la reacción fue realizado omitiendo el anticuerpo primario.
3.1.4a. Inmunofluorescencia para E-cadherina en sitios de implantación.
Los sitios de implantación procesados histológicamente fueron desparafinados en xilol y re-
hidratados progresivamente por pasaje en alcoholes de gradaciones decrecientes y lavados en PBS
(3 veces de 5 min cada una, 3 x 5 min). Luego los cortes fueron expuestos a una recuperación
antigénica en microondas a 400 w por 10 min y permeabilizados 20 min con PBST 0,25%.
Posteriormente, las secciones fueron lavadas en PBS (3 x 5 min) e incubadas con suero de cabra
1:40 en PBS, 1 h a TA. Luego de lo cual fueron incubadas con anticuerpo primario mouse anti E-
cadherina 1:5 en PBS overnight a 4°C en cámara húmeda. Al día siguiente, luego de los lavados en
PBS (3 x 5 min), los cortes fueron incubados con anticuerpo secundario goat anti mouse Alexa 488
diluido 1:25 en PBS, 1 h a TA en oscuridad. Posteriormente fueron lavados en PBS y montados en
glicerol - PBS (1:1) para ser observados en microscopio confocal. La especificidad de la reacción fue

Capítulo III�
173
�
determinada mediante la omisión del anticuerpo primario. El embrión fue fotografiado a un
aumento de 20x + zoom 2x para la evaluación de la distribución, patrón de expresión e intensidad
relativa de la inmunomarca en el ectodermo embrionario mediante el programa computacional
Scion Image.
3.1.4b. Análisis de imágenes y semicuantificación de la intensidad de inmunofluorescencia.
Para semicuantificar el nivel de inmunorreactividad para E-cadherina en la región ectodérmica de
embriones al día 8 de gestación, de HC y HT, se empleó el método descrito por Tolivia y
colaboradores (2006). Todos los ajustes y las condiciones de captura de imágenes digitales
realizadas en el microscopio confocal se mantuvieron constantes para todas las muestras. Las
micrografías adquiridas fueron guardadas en formato de imagen TIFF y procesadas usando Adobe
Photoshop CS2 para reconocer, seleccionar y separar las señales del cromógeno del fondo
inespecífico. De cada imagen a 20x se seleccionó y recortó la región correspondiente al ectodermo
embrionario, excluyendo los tejidos extraembrionarios y la cavidad exocelómica. Las imágenes
fueron luego convertidas a escala de grises. La iluminación global de cada pixel en las imágenes
convertidas corresponde a la intensidad de la fluorescencia. Cada micrografía convertida fue luego
invertida para que los pixeles grises y negros representen zonas de alta fluorescencia sobre un
fondo blanco y fueron guardadas como una nueva imagen TIFF para ser analizada con el programa
Scion Image. En cada micrografía (conteniendo exclusivamente el ectodermo embrionario) se
midió la intensidad media de la señal del cromógeno (densitometría promedio, DP) (Tolivia y col.,
2006). Los valores promedios de DP en pixeles para cada tratamiento fueron comparados
estadísticamente.

Capítulo III�
174
�
3.2. Metodologías para el estudio de la expresión proteica y génica de MMP-2 y MMP-9 en
embriones en el período postimplantatorio: Día 8 de gestación.
3.2.1a. Inmunohistoquímica para MMP-2 y MMP-9 en sitios de implantación.
Cortes transversales de sitios de implantación correspondientes al 8vo día de preñez fijados,
fueron desparafinados, hidratados y lavados en PBS (3 x 5 min). Luego fueron permeabilizados en
PBST 0,25% por 15 min a TA. Posteriormente, las secciones fueron lavadas en PBS y las
peroxidasas endógenas inhibidas con peróxido de hidrógeno (30v) al 3% en PBS durante 30 min a
TA. Luego de los lavados en PBS (3 x 5 min) los sitios de unión inespecíficos fueron bloqueados con
una solución de PBS - leche 3%, 1 h a TA. Transcurrido dicho período, las secciones fueron
incubadas overnight a 4°C en cámara húmeda con anticuerpo primario anti metaloproteasas: goat
anti MMP-2 diluido 1:50 en PBS ó con anticuerpo primario rabbit anti MMP-9 diluido 1:200 en
PBS. La especificidad de la reacción fue determinada mediante la omisión del anticuerpo primario.
Al día siguiente, luego de lavados en PBS (3 x 5 min), los cortes fueron incubados 1 h a TA en
cámara húmeda con anticuerpo secundario rabbit anti goat biotinilado 1:400 en PBS ó goat anti
rabbit biotinilado 1:400 en PBS. Luego de los lavados, en ambos casos, se prosiguió con un paso de
amplificación con el complejo Streptoavidin horseradish peroxidase (HRP) diluido 1:400 en PBS, 1 h
a TA en cámara húmeda. El revelado con 3-3`diaminobencidina (DAB) fue realizado utilizando un
kit comercial y posterior contracoloración con hematoxilina de Harris. Las muestras fueron
deshidratadas, aclaradas en xilol y montadas en medio de montaje anhidro (DPX). Se analizaron
dos sitios de implantación en estadio embrionario TS10 por animal de un total de 5 animales de
cada grupo experimental.
3.2.1b. Análisis de imágenes y semicuantificación de la inmunohistoquímica para MMPs.
Se adquirieron micrografías de cada zona del sitio de implantación inmunomarcado con
microscopio equipado con cámara de video conectada a un programa de análisis de imágenes
Nikon. En cada imagen (40x) se analizó la localización de la marca (intracelular y/o extracelular), el
patrón de marcación (granular u homogénea), la distribución (presente o ausente en el tejido
analizado), así como el nivel de intensidad relativo de la inmunomarca (en aquellos tejidos donde
se encontró marcación positiva). Para este último caso, se establecieron valores relativos de

Capítulo III�
175
�
intensidad (score) mediante la observación de la marca en cada zona de los sitios de implantación
de hembras controles. Los valores de intensidad establecidos fueron: “0”: sin marca; “1”:
marcación tenue; “2”: marcación moderada; “3”: marcación intensa. Luego, se analizaron todas las
imágenes de las zonas de los sitios de las HT a las que se les asignó un valor de intensidad, el cual
fue contrastado con los valores de las HC. Finalmente se realizó un promedio del nivel de
intensidad por zona para cada muestra de las HC y de las HT, para ser comparado
estadísticamente. Las zonas analizadas fueron: el cono ectoplacentario, incluyendo la zona de
proliferación, la zona de expansión trofoblástica y las células CGTs 2rias
; los tejidos embrionarios;
los tejidos extraembrionarios y la interfase trofoblástico-decidual (células CGTs 1rias
y tejido
decidual limitante).
3.2.2. PCR en Tiempo Real para MMP-2 y MMP-9 de cono ectoplacentario.
Para analizar los niveles de expresión génica de MMP-2 y MMP-9 en conos ectoplacentarios
colectados como fue indicado anteriormente, se utilizó el Kit Power SYBR® Green Cells-to-CtTM
el
cual incluye los reactivos necesarios para la retrotranscripción del ARNm seguida de la
amplificación por PCR en Tiempo Real del ADN copia (ADNc), directamente de muestras lisadas sin
previa extracción de RNA. Por lo tanto, luego de la obtención de los conos, se procedió de acuerdo
a lo indicado en el protocolo del kit comercial.
Para la lisis celular, los conos colectados fueron incubados por 5 min a 28°C con solución de Lisis
suplementada con Dnasa I para remover el ADN genómico. Las células lisadas durante este
procedimiento liberan el ARN a la solución de lisis la cual contiene los reactivos que inactivan
RNAsas endógenas. Una vez transcurridos los 5 min, se adicionó solución stop e incubó por 2 min a
TA. El lisado obtenido fue almacenado a -20°C hasta la síntesis del ADNc.
Para la síntesis de ADNc por retrotranscripción (RT) del ARN en el lisado celular se siguieron las
instrucciones del kit. Al lisado se adicionó una solución RT Master Mix compuesta por 2X SYBR® RT
buffer y 20X RT Enzyme Mix. Usando un termociclador Mastercycler ep, el programa utilizado para
la síntesis de ADNc fue 60 min a 37°C, seguido de 5 min a 95°C. Finalizado ese período, las
muestras de ADNc fueron almacenadas a -20°C hasta la realización de la PCR en Tiempo Real. Tres
muestras de ADNc fueron sintetizadas a partir de cada lisado conteniendo 3 conos
ectoplacentarios cada uno.

Capítulo III�
176
�
El ADNc sintetizado fue amplificado en un termociclador StepOnePlus™ Real-Time PCR Systems. La
PCR en Tiempo Real (RT-qPCR) fue realizada para la cuantificación relativa del ADNc generado por
transcripción reversa mediante la utilización de una solución de Power SYBR Green PCR Master
Mix con los primers de interés, conforme a lo indicado por el fabricante en placas multiwell
MicroAmp® Fast Optical 96-Well Reaction Plate (Applied Biosystems). Las placas fueron
centrifugadas (1 min a 800 rpm). Como control endógeno fue utilizado el gen Ywhaz (tyrosine 3-
monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide [Mus
musculus]). Como control negativo fue utilizada una solución Mix completa en la cual el ADNc fue
omitido. Los primers, sintetizados por Exxtend Biotecnologia Ltda., consistieron en las siguientes
secuencias: Mus musculus MMP-2 sense: 5´-GTTCAACGGTCGGGAATACA-3´, MMP-2 antisense: 5´-
GCCATACTTGCCATCCTTCT-3´ (103 pares de bases, pb. NCBI Ref. Seq. mRNA: NM_008610.2); Mus
musculus MMP-9 sense: 5´-CTGGAACTCACACGACATCTT-3´, MMP-9 antisense: 5´-
AAATGAGGTGAACAAGGTGGA -3´ (102 pb. NCBI Ref. Seq. mRNA: NM_013599.3); Mus musculus
Ywhaz sense: 5’-GAAGCCACAATGTTCTTGGCCCAT-3’, Ywhaz antisense: 5´-
AAACCAACAGAGACTTGGAAGCAC- 3’ (84 pb. NCBI Ref. Seq. mRNA: NM_001253805.1). Las
amplificaciones fueron realizadas utilizándose las siguientes condiciones de ciclado: 10 min a 95°C
para activación de la enzima, seguido de 40 ciclos de desnaturalización a 95°C por 15 seg,
annealing a 60°C por 30 seg y extensión a 72 °C por 30 seg. La curva de disociación (melting curve:
15 seg a 95°C, 60 seg a 60°C, 15 seg a 95°C) fue realizada con los productos de amplificación para
asegurar la especificidad de cada primer. Fue utilizado el método comparativo CT (threshold cycle)
para calcular el nivel de expresión relativa de los genes (StepOne™ Real-Time PCR Software v2.1,
Applied Biosystems). Los valores de CT obtenidos para Mmp-2 y Mmp-9 fueron normalizados al
control endógeno Ywhaz para la comparación del nivel de expresión relativa. Los valores de CT
normalizados fueron expresados como promedios + SEM.

Capítulo III�
177
�
3.3. Metodologías para el estudio de la genotoxicidad materna y embrionaria, y de la muerte
celular en embriones en el período periimplantatorio: Días 4 y 8 de gestación.
3.3.1. Genotoxicidad embrionaria preimplantatoria (día 4 de gestación).
Se utilizó la técnica de Tarkowski, descripta en el Capítulo II, para analizar la morfología de los
núcleos interfásicos en mórulas y blastocistos recuperados al día 4 de gestación. Los núcleos
embrionarios fijados en portaobjetos y coloreados con Giemsa, fueron observados al microscopio
óptico para evaluar la presencia o no de alteraciones nucleares, tales como: puentes cromatínicos,
gemaciones o lóbulos (núcleos preapoptóticos), picnosis (núcleos con cromatina altamente
condensada), y/o micronúcleos (MNs). La identificación de los MNs se basó en los siguientes
criterios: (1) estructuras pequeñas, redondas y con tono similar al nuclear luego de la coloración
con Giemsa; (2) generalmente, con un diámetro menor a un tercio del diámetro del núcleo
principal; (3) con localización dentro de los 3 diámetros del núcleo principal; (4) aquellas
blastómeras que presentaron uno o más MNs fueron contadas como una única célula
micronucleada. Imágenes de dichas alteraciones se muestran en la Figura III.3. En cada tipo
embrionario, se registró el número de núcleos con alteraciones y el resultado se expresó como
frecuencia sobre el número total de núcleos analizados.
3.3.2. Ensayo de TUNEL in-toto en embriones de preimplantación (día 4 de gestación).
Con fin de evaluar la muerte celular por apoptosis, los blastocistos fijados como fue descripto
anteriormente, fueron permeabilizados en una solución PBST 0,5% durante 10 min a TA. Luego de
varios lavados en microgotas de PBS - BSA 0,3%, las peroxidasas endógenas fueron inhibidas con
peróxido de hidrógenos (30v) 2% en PBS por 5 min a TA. Luego de lavados en PBS - BSA 0,3%, un
grupo de embriones fue incubado por 10 min a 37°C con DNAsa 0,5 µg/ml (control positivo). Luego
de los lavados, se procedió de acuerdo a lo establecido en el Kit comercial para la determinación
de la apoptosis (ApopTag® Plus Peroxidase In Situ Apoptosis Detection Kit). Los blastocistos fueron
incubados en microgotas de equilibration buffer por 5 min a TA, luego de lo cual fueron incubados
por 1h a 37°C con la enzima terminal deoxynucleotidyl transferase (TdT, 30%) preparada en
reaction buffer (70%) conteniendo los nucleótidos necesarios para la reacción, en las proporciones
indicadas��La especificidad de la reacción fue determinada mediante la omisión de la TdT en un

Capítulo III�
178
�
grupo de embriones (control negativo). Una vez transcurrido el tiempo de incubación, los
blastocistos fueron transferidos a microgotas con stop/wash buffer por 10 min a TA y luego fueron
lavados en PBS - BSA 0,3%. La siguiente incubación fue con el complejo anti digoxigenina
peroxidasa durante 30 min a TA. Luego de los lavados se prosiguió con el revelado con DAB
durante 6 min. Finalmente, los embriones fueron cuidadosamente montados en gotas glicerol -
PBS (1:1) sobre portaobjetos cubiertos con poli-L-lisina con cubreobjetos con soportes de
plastilina. Siempre que las incubaciones de los embriones fueron realizadas en la estufa de cultivo
a 37°C, las gotas fueron protegidas de la evaporación con aceite mineral. Bajo microscopio óptico
fue registrado el número de células con marca positiva (células TUNEL-positivas) por embrión,
distinguiendo en cada caso si dichas células fueran del macizo celular interno o del
trofoectodermo (polar o mural).

Capítulo III�
179
�
Figura III.3. Genotoxicidad embrionaria al día 4 de gestación. Micrografías representativas de núcleos
de embriones preimplantatorios procesados según la técnica de Tarkowski. A, blastocisto inicial (10x);
B, blastocisto expandido (10x); C, núcleos interfásicos (in) y metafásicos (me) normales (60x); D-H,
núcleos interfásicos anormales: D, núcleos interfásicos picnóticos (flechas, 40x); E, detalle de núcleos
picnóticos (pic), con cromatina condensada, intensamente coloreada y con tamaño nuclear reducido
(60x); F, núcleo interfásico con puente cromatínico (flecha, 60x); G, núcleos interfásicos con
gemaciones (flechas, 60x); H, núcleo interfásico con un MN (flecha, 60x).

Capítulo III�
180
�
3.3.3. Ensayo de TUNEL en sitios de implantación (día 8 de gestación).
El ensayo de TUNEL se llevó a cabo en sitios de implantación procesados histológicamente como
fue descripto anteriormente. Los cortes fueron desparafinados en xilol, hidratados
progresivamente en alcoholes y lavados en PBS (3 x 5 min). Luego fueron permeabilizados 10 min
con PBST 0,25%. Posteriormente, las secciones fueron lavadas en PBS (3 x 5 min) e incubadas con
proteinasa K 20 µg/ml en PBS durante 15 min a TA. Luego de los lavados en PBS (3 x 5 min) los
cortes fueron�post-fijados por 5 min con paraformaldehído 4% a TA. Las peroxidasas endógenas
fueron inhibidas con peróxido de hidrógeno (30v) 3% en PBS, 5 min a TA. Luego de los lavados, un
portaobjeto seleccionado como control positivo, fue incubado con DNAsa 0,5 µg/ml por 10 min a
37°C en cámara húmeda. Transcurrida dicha incubación se procedió de acuerdo a lo indicado por
el kit comercial (ApopTag® Plus Peroxidase In Situ Apoptosis Detection Kit). Todas las secciones
fueron incubadas con equilibration buffer, 5 min a TA a lo que le siguió la incubación con la enzima
TdT (30%) junto con reaction buffer (70%) en las proporciones indicadas, 1 h a 37°C en cámara
húmeda. La especificidad de la reacción fue determinada mediante la omisión de la TdT (control
negativo). Una vez concluido el tiempo de incubación, todas las secciones fueron incubadas con
stop/wash buffer, 10 min a TA, prosiguiendo con lavados en PBS (3 x 1 min). Inmediatamente,
fueron incubadas 30 min con el complejo anti digoxigenina peroxidasa a TA en cámara húmeda,
lavadas en PBS (3 x 5 min) y reveladas 5 min con DAB. Se utilizó Hematoxilina de Harry como
colorante de contraste. Luego las muestras fueron deshidratadas en alcoholes y aclaradas en xilol
para su montaje en DPX. Las muestras fueron observadas bajo microscopio óptico donde se
registró el número de células TUNEL-positivas en el cono ectoplacentario, en la interfase
trofoblástico-decidual, en el embrión y en los tejidos extraembrionarios. Los datos fueron
expresados como número promedio de células TUNEL-positivas por sitio de implantación. Las
imágenes fueron adquiridas con microscopio equipado con una cámara de video conectada a un
programa de análisis de imágenes Nikon.
3.3.4. Marcación nuclear con Hoechst 33342 en sitios de implantación (día 8 de gestación).
La marcación nuclear con Hoechst 33342 se llevó a cabo en sitios de implantación procesados
histológicamente como fue descripto. Los cortes fueron desparafinados en xilol, hidratados
progresivamente en alcoholes, lavados en PBS (3 x 5 min) e incubados, en oscuridad, con una

Capítulo III�
181
�
solución de Hoechst 33342 en PBS (0,5 mg/ml) por 5 min a TA. Luego de los lavados en PBS (3 x 5
min), las secciones fueron montadas con una solución de PBS - glicerol (1:1) y fotografiadas bajo
microscopio de fluorescencia. En la interfase trofoblástico-decidual y en el ectodermo embrionario
de cada muestra, se analizó la organización (disposición) nuclear en los tejidos, la forma y tamaño
de los núcleos así como el grado de condensación de la cromatina y la presencia de
fragmentaciones nucleares.
3.3.5. Test de micronúcleos para evaluación de genotoxicidad materna.
A partir de extendidos celulares de médula ósea, obtenidos como fue descripto anteriormente, se
procedió al estudio de la genotoxicidad materna, mediante la identificación y recuento de los MNs
en eritrocitos policromáticos (PCE). La identificación de los MNs se basó en los siguientes criterios:
(1) estructuras pequeñas y redondas (2) generalmente, con un diámetro menor a un tercio del
diámetro del núcleo principal; (3) con localización dentro de los 3 diámetros del núcleo principal;
(4) aquellos PCE que presentaron uno o más MNs fueron contados como una única célula
micronucleada (Figura III.4). Bajo microscopio óptico de campo claro, se registró el número de PCE
con MNs sobre un total de 1000 PCE contados por hembra. Los datos fueron expresados como
promedio de PCE con MN/1000 PCE.
Figura III.4. Test de micronúcleos para evaluación de la genotoxicidad. Micrografías representativas
de micronúcleos (MNs, flechas) en eritrocitos policromáticos (PCE) de médula ósea femoral. A, muestra
representativa de una hembra control con un PCE con MN; B, muestra representativa de una hembra
tratada con alcohol evidenciando tres PCE con MNs. Cebral y colaboradores (2011). Escala: 10 μm.

Capítulo III�
182
�
4. Análisis estadístico.
Los datos expresados como promedios de los valores ± SD o ± SEM de las muestras de las HC y HT
se compararon por Prueba T de Student o ANOVA no paramétrica y Test de Mann-Whitney. Los
datos expresados como proporciones o frecuencias numéricas se analizaron por prueba exacta de
Fisher o Chi-Cuadrado. Se utilizó el programa estadístico GraphPad Instat v2. Un p<0,05 fue
considerado estadísticamente significativo.

Capítulo III�
183
�
III.2. RESULTADOS
Dado que la ingesta perigestacional de alcohol hasta la fase pre- y postimplantatoria temprana
(gastrulación) en hembras CD-1 y CF-1, afectó negativamente la diferenciación, el crecimiento, la
morfología embrionaria y la invasividad del tejido trofoblástico del cono ectoplacentario desde la
fase de implantación, se postuló que estas anomalías podrían ser causadas, al menos en parte, por
la acción adversa del alcohol sobre algunas moléculas claves para el desarrollo embrio-placentario
temprano, alterando en particular la expresión de moléculas de adhesión intercelular embrionaria,
y la expresión de metaloproteasas embrio-trofoblásticas en la fase postimplantatoria temprana. En
paralelo con estas anomalías, los efectos negativos del consumo de alcohol podrían implicar
genotoxicidad embrionaria, así como un incremento de la muerte celular en los tejidos
embrionarios y placentarios en desarrollo. En este contexto, un aumento de la apoptosis
embrionaria y/o la aparición de anomalías nucleares en las células embrio-trofoblásticas, podría
ocurrir junto con genotoxicidad materna reforzando el efecto deletéreo del alcohol sobre el
desarrollo embrio-placentario temprano. Por lo tanto, para dilucidar estas hipótesis, se analizó el
patrón de inmunoexpresión de proteínas de las uniones adherentes y estrechas, la expresión génica
y proteica de la MMP-2 y MMP-9, las alteraciones núcleo-genotóxicas y apoptosis embrio-
trofoblásticas periimplantatorias, junto con la frecuencia de micronúcleos en eritrocitos de médula
ósea materna, luego del consumo perigestacional de alcohol.
1. Efectos del consumo perigestacional de alcohol en moléculas de adhesión celular en
embriones en el período periimplantatorio: Día 4 y 8 de gestación.
1.1. Estudio de las uniones intercelulares en embriones preimplantatorios: Día 4 de gestación.
Según nuestros hallazgos en los cuales las hembras CD-1 y CF-1 tratadas con alcohol hasta el día 4
de gestación presentaron una alta proporción de blastocistos morfológicamente anormales, con
alteraciones principalmente en la cavitación (véase Figura II.10 del Capítulo II), decidimos analizar
por inmunofluorescencia el patrón de inmunomarcación de la proteína ZO-1, importante
componente de las uniones estrechas del trofoectodermo para la formación de la cavidad durante
el desarrollo de los blastocistos. Dado que las alteraciones morfológicas de la cavidad en los

Capítulo III�
184
�
blastocistos de las HT-CF-1 en comparación con las HT-CD-1 fueron más pronunciadas, decidimos
abordar estos estudios en CF-1.
Se detectó expresión de ZO-1 en el trofoectodermo polar (TP) y mural (TM) de los blastocistos
iniciales y expandidos. En los blastocistos provenientes de las HC (Figura III.5.A y B; Figura III.6.A),
ZO-1 se localizó en la periferia del citoplasma, en los contactos celulares en forma
submembranosa. La marca se observó continua y definida. Por el contrario, en los embriones
provenientes de HT, si bien ZO-1 también fue detectada en los contactos celulares del TP y TM, se
evidenció deslocalización de la marca con desplazamiento hacia el centro (marca
intracitoplasmática perinuclear), donde se observó un patrón de marcación engrosada, granular e
intensa en el 90% de los embriones analizados. A su vez, en los contactos intercelulares, dicha
marca se presentó discontinua, delgada y/o tenue indicando, posiblemente una unión debilitada
(Figura III.5.C y D; Figura III.6.B y C).
�
�
Figura III.5. Inmunomarcación de ZO-1 en blastocistos iniciales al día 4 de gestación. Micrografías
representativas de embriones de preimplantación de hembras controles (A-B) y tratadas con alcohol
(C-D), CF-1. A-B, ZO-1 se localiza en los contactos celulares de TP y TM. La marca es continua y definida
(flechas); C-D, la marca para ZO-1 se evidencia engrosada, granular y deslocalizada hacia el núcleo. La
marca en los contactos celulares es discontinua y delgada/tenue (puntas de flecha). TM:
trofoectodermo mural, TP: trofoectodermo polar. Aumento: 40x.

Capítulo III�
185
�
Figura III.6. Inmunomarcación de ZO-1 en blastocistos expandidos al día 4 de gestación. Micrografías
representativas de embriones de preimplantación de hembras controles (A) y tratadas con alcohol (B-
C), CF-1. A, la ZO-1 se localiza en los contactos celulares del TP y TM. La marca es continua y definida
(flechas); B-C, la marca de ZO-1 en el TP se ve engrosada, granular e intensa (B: punta de flecha), en el
TM en se deslocaliza hacia el núcleo (*), siendo en los contactos intercelulares una marcación delgada
y/o tenue (C: puntas de flecha). TM: trofoectodermo mural, TP: trofoectodermo polar. Aumento: 40x.
Con el fin de determinar si la exposición a alcohol afectó las uniones adherentes de embriones
preimplantativos, tanto blastocistos iniciales como expandidos fueron inmunomarcados para
detectar E-cadherina. E-cadherina se localizó en las superficies de contacto (membrana) del TP y
TM de ambos tipos de blastocistos. Los blastocistos de las HC presentaron un patrón de marcación
continuo y definido en las superficies celulares (Figura III.7.A; Figura III.8.A). Sin embargo, los
embriones de las HT tuvieron patrones de expresión alterados tales como marcación engrosada y
discontinua con distorsiones y acumulaciones membranosas (Figura III.7.B, C y D; Figura III.8.B).
�

Capítulo III�
186
�
Figura III.7. Inmunomarcación de E-cadherina en blastocistos inicales al día 4 de gestación.
Micrografías representativas de embriones de preimplantación de hembras controles (A) y tratadas
con alcohol (B-D), CF-1. A, la E-cadherina se localiza en los contactos celulares del TP y TM. El patrón de
marcación es continuo y definido (flecha); B-D, la E-cadherina se inmunolocaliza en los contactos
celulares de TP y TM pero la marca se presenta engrosada (B: punta de flecha) y discontinua (C y D:
puntas de flecha). TP: trofoectodermo polar, TM: trofoectodermo mural. Aumento 40x.
Figura III.8. Inmunomarcación de E-cadherina en blastocistos expandidos al día 4 de gestación.
Micrografías representativas de embriones de preimplantación de hembras controles (A) y tratadas
con alcohol (B), CF-1. A, la E-cadherina se localiza en las superficies de contactos intercelulares del TP y
TM con un patrón continuo y definido (flechas); B, la E-cadherina se inmunolocaliza en los contactos
intercelulares pero se presenta engrosada y discontinua (punta de flecha). TM: trofoectodermo mural,
TP: trofoectodermo polar. Aumento: 40x.

Capítulo III�
187
�
Como es sabido, la biogénesis de las uniones estrechas es dependiente del previo y adecuado
establecimiento de uniones adherentes, procesos que se evidencian por un patrón de
localización/distribución diferencial de estas proteínas de unión, ya sea temporal como
espacialmente (membranosa y/o citoplasmática). Por ello, nuestro siguiente objetivo fue analizar
el grado de co-localización (doble inmunomarcación) entre la ZO-1 y la E-cadherina. Observamos
que el 100% de los embriones analizados provenientes de HC presentaron un alto grado de co-
localización (Figura III.9.C) de la ZO-1 (Figura III.9.A) con la E-cadherina (Figura III.9.B),
particularmente en el TP. En las HT, por el contrario, sólo el 50% de los embriones presentó co-
localización positiva, con menor grado de doble marcación para ambas proteínas (amarillo)
únicamente en TM y con un patrón puntiforme (Figura III.9.F), comparado con el patrón de
marcación de los embriones provenientes de HC (Figura III.9.C).
�

Capítulo III�
188
�
Figura III.9. Co-localización de ZO-1 y E-cadherina en blastocistos expandidos al día 4 de gestación.
Micrografías confocales representativas de embriones de preimplantación de hembras controles (A-C)
y tratadas con alcohol (D-F), CF-1. A, la ZO-1 (FITC, verde) se localiza en contactos celulares de TM y TP
de forma continua; B, la E-cadherina (Rodhamina, rojo) se localiza en la membrana de los contactos
celulares de TM y TP en forma continua; C, co-localización (amarillo) positiva y continua de ambas
proteínas en el TP; D, la ZO-1 (verde) se localiza en los contactos celulares en forma discontinua; E, la E-
cadherina (rojo) se presenta en los contactos celulares de la TM y el TP en forma membranosa pero
discontinua e irregular; F, la co-localización (amarillo) de la ZO-1 y la E-cadherina en embriones de HT
fue sólo levemente positiva. TM: trofoectodermo mural, TP: trofoectodermo polar. Escala: 50 µm

Capítulo III�
189
�
1.2. Estudio de las uniones adherentes en el ectodermo embrionario: Día 8 de gestación.
Basados en nuestras observaciones en cuanto a la histopatología de los embriones provenientes
de HT al 8vo. día de gestación, en los cuales se evidenció en un alto grado de desorganización
celular principalmente, del ectodermo embrionario (véase Figura II.25 del Capítulo II), se postuló
que alteraciones en el patrón de expresión de moléculas de las uniones adherentes,
particularmente de E-Cadherina, podrían estar involucradas en las anomalías histológicas
observadas en los embriones en gastrulación. El patrón de inmunomarcación para E-cadherina fue
analizado en embriones en estadio en TS10 de HC y HT, provenientes de la colonia CF-1 (Figura
III.10). Si bien no se encontraron diferencias en el patrón de marcación en el ectodermo
embrionario derivado de HT respecto del derivado de HC, el que se observó continuo y lineal entre
los contactos intercelulares de las células ectodérmicas, el análisis densitométrico de la
inmunoexpresión de dicha molécula mostró una reducción significativa de la marca de embriones
de HT (p<0,05, Figura III.10.B) respecto de la expresión de la E-cadherina de los embriones de HC
(Figura III.10.A).
Figura III.10. Inmunomarcación de E-cadherina en embriones en gastrulación al día 8 de gestación.
Micrografías representativas de embriones de postimplantación en estadio TS10 de hembras controles
(A) y tratadas con alcohol (B), CF-1. A, la inmunomarcación de E-cadherina (FITC, verde) se presentó
fuerte y definida en el EctoE (flecha); B, la intensidad de la inmunomarcación de E-cadherina en el
EctoE se redujo respecto de la marca de los embriones controles (punta de flecha). EctoE: ectodermo
embrionario. Aumento: 20x.

Capítulo III�
190
�
2. Efectos del consumo perigestacional de alcohol en la expresión proteica y génica de MMP-2 y
MMP-9 en embriones en el período postimplantatorio: Día 8 de gestación.
Según la hipótesis planteada, por la cual la ingesta perigestacional de alcohol hasta el día 8 de
gestación induce alteraciones en la expresión de MMPs en el tejido embrio-trofoblástico, los
objetivos fueron, en primer lugar, analizar tanto en CD-1 como CF-1, el patrón de
inmunomarcación de MMP-2 y MMP-9 en cuanto a la localización, distribución e intensidad
relativa en el tejido embrionario, extraembrionario, cono ectoplacentario y en la interfase
trofoblástico-decidual. En segundo lugar, el objetivo fue analizar la expresión génica de ambas
MMPs en el cono ectoplacentario aislado del sitio de implantación de día 8 de gestación tejido
involucrado, principalmente, en la invasión en el tejido decidual.
�
2.1. Estudio del patrón de inmunomarcación de MMP-2 y MMP-9 en el tejido embrio-
trofoblástico.
Se analizó la inmunoexpresión de MMP-2 y MMP-9 en sitios de implantación de HC y HT de las
colonias CD-1 y CF-1, particularmente observándose el cono ectoplacentario, la interfase
trofoblástico-decidual, los tejidos embrionarios y extraembrionarios. La inmunomarca de ambas
proteasas resultó en un patrón similar en las zonas mencionadas en las dos colonias, CD-1 y CF-1.
En ambas colonias, tanto para las muestras provenientes de HC como de HT, el patrón de
inmunoexpresión para MMP-2, cuando ésta fue positiva, fue principalmente extracelular y de
aspecto granular (Figura III.11.C y D). El cono ectoplacentario mostró reactividad positiva para
MMP-2 en la zona de proliferación (core) y en la de expansión trofoblástica, no así en las CGTs 2rias
,
tanto en muestras provenientes de HC como HT (Figura III.11.E y F, respectivamente). El análisis de
la intensidad relativa de la marca, según una escala numérica establecida previamente, mostró un
aumento significativo de la expresión de MMP-2 únicamente en la zona de proliferación
trofoblástica de las HT respecto de las HC (p<0,05, Figura III.13.A). El tejido ectodérmico
embrionario en las HT presentó mayor inmunomarcación que en las HC (Figura III.11.G y H; p<0,05,
Figura III.14.A). Lo mismo se observó para el mesodermo embrionario. El endodermo parietal y
visceral presentó mayor marcación de MMP-2 en las HT que el de las HC (p<0,05, Figura III.14.A).
Tanto en las HC como en las HT, el ectodermo extraembrionario no presentó marca positiva

Capítulo III�
191
�
(Figura III.14.A) así como tampoco las células CGTs 1rias
en la interfase trofoblástico-decidual
(Figura III.11.E-H).
El patrón de inmunomarcación de MMP-9, en ambas colonias CD-1 y CF-1, tanto en HC como HT,
fue continuo, homogéneo y extracelular. En el cono ectoplacentario de las muestras de HT, la
inmunomarca de MMP-9 se presentó significativamente disminuida tanto en la zona de
proliferación como en la zona de expansión respecto de las muestras controles (Figura III.12.C y D,
respectivamente; p<0,01, Figura III.13.A). De la misma manera, las CGTs 2rias
de las hembras
tratadas presentaron un nivel de marcación reducido respecto de las controles (Figura III.12.E y F,
respectivamente; p<0,01; Figura III.13.B). La inmunoexpresión de MMP-9 en el ectodermo
extraembrionario de las HT no presentó diferencias significativas respecto de las HC y su
marcación fue muy reducida (Figura III.12.H y G, respectivamente; Figura III.14.B). El endodermo
extraembrionario, por el contrario, presentó alta inmunomarcación, sin embargo, tampoco se
observaron diferencias significativas entre las HT respecto de las HC (Figura III.12.H y G,
respectivamente; Figura III.14.B). El tejido ectodérmico del embrión de las HT tuvo mayor
inmunoexpresión de MMP-9 que el de las HC (Figura III.12.J y I, respectivamente; p<0,05, Figura
III.14.B). En la interfase trofoblástico-decidual, las CGTs 1rias
mostraron marca positiva, la cual
disminuye significativamente en las células derivadas de HT respecto de las controles (Figura
III.12.K y L; valores promedio HC: 1,60 + 0,16 versus HT: 1,33 + 0,13; p<0,05).
�
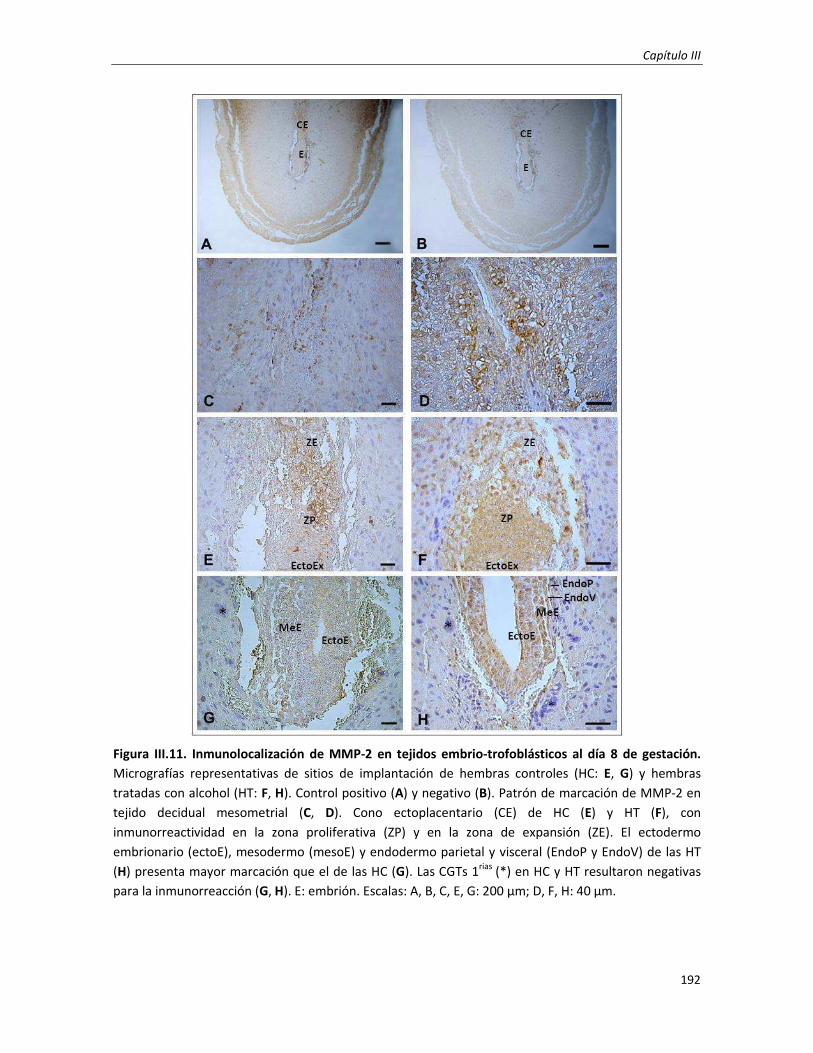
Capítulo III�
192
�
Figura III.11. Inmunolocalización de MMP-2 en tejidos embrio-trofoblásticos al día 8 de gestación.
Micrografías representativas de sitios de implantación de hembras controles (HC: E, G) y hembras
tratadas con alcohol (HT: F, H). Control positivo (A) y negativo (B). Patrón de marcación de MMP-2 en
tejido decidual mesometrial (C, D). Cono ectoplacentario (CE) de HC (E) y HT (F), con
inmunorreactividad en la zona proliferativa (ZP) y en la zona de expansión (ZE). El ectodermo
embrionario (ectoE), mesodermo (mesoE) y endodermo parietal y visceral (EndoP y EndoV) de las HT
(H) presenta mayor marcación que el de las HC (G). Las CGTs 1rias
(*) en HC y HT resultaron negativas
para la inmunorreacción (G, H). E: embrión. Escalas: A, B, C, E, G: 200 µm; D, F, H: 40 µm.

Capítulo III�
193
�

Capítulo III�
194
�
Figura III.12. Inmunolocalización de MMP-9 en tejidos embrio-trofoblásticos al día 8 de gestación.
Micrografías representativas de sitios de implantación de hembras controles (HC: C, E, G, I, K) y
hembras tratadas (HT: D, F, H, J, L). Control positivo (A) y negativo (B). Se observa inmunorreactividad
positiva en la zona proliferativa (ZP) y de expansión trofoblástica (ZE) del cono ectoplacentario (CE) de
HC (C) y HT (D). Las CGT 2rias
(*) de las HC (E) presentan alta inmunomarcación con respecto a las de HT
(F). El endodermo visceral (EndoV) de las HC (G) y HT (H) presenta alta marcación positiva. El
ectodermo extraembrionario (EctoEx) resultó con escasa marca en HC (G) y HT (H). El ectodermo
embrionario (EctoE) de las HT (I) presentó mayor marcación que el de las HC (J). Las CGTs 1rias
(*) de las
HC (K) presentan mayor marcación respecto a las de HT (L). E: embrión. Escalas: A y B: 100 µm; C y D:
20 µm; E-L: 10 µm.
Figura III.13. Inmunomarcación de MMP-2 y MMP-9 en cono ectoplacentario al día 8 de gestación. En
muestras provenientes de hembras CF-1 controles (HC) y tratadas con alcohol (HT), se analizó la
intensidad relativa de la inmunomarca de MMP-2 (A) y MMP-9 (B) según un score preestablecido. Zona
prolif: zona de trofoblasto proliferativo, zona expan: zona de expansión trofoblástica, CGT 2rias
: células
gigantes trofoblásticas secundarias. Los valores se expresan como promedio de la intensidad relativa +
SEM. *p<0,05 y **p<0,01 versus su control (Prueba T de Student).

Capítulo III�
195
�
Figura III.14. Inmunomarcación de MMP-2 y MMP-9 en tejidos embrionarios y extraembrionarios al
día 8 de gestación. En muestras provenientes de hembras CF-1 controles (HC) y tratadas con alcohol
(HT), se analizó la intensidad relativa de la inmunomarca de MMP-2 (A) y MMP-9 (B) según un score
preestablecido. EctoEx: ectodermo extraembrionario, EndoEx: endodermo extraembrionario, EctoE:
ectodermo embrionario. Los valores se expresan como promedio de la intensidad relativa + SEM.
*p<0,05 versus su control (Prueba T de Student).
2.2. Estudio de la expresión génica de MMP-2 y MMP-9 en el cono ectoplacentario.
Dada la importancia de la MMP-2 y MMP-9 en el proceso de invasión trofoblástica en el tejido
decidual, y de acuerdo a las diferencias encontradas en la inmunoexpresión de estas proteasas en
el cono ectoplacentario de las HT respecto del de las HC, se decidió analizar la expresión génica de
ambas metaloproteasas por RT-qPCR en conos ectoplacentarios aislados de HC y HT. Como el
patrón de expresión proteica para ambas metaloproteasas resultó similar en ambas colonias CD-1
y CF-1, el análisis del nivel de expresión génica fue realizado en muestras provenientes de hembras
CD-1. Como muestra la Figura III.15, el nivel de expresión génica de MMP-2 no mostró diferencias
significativas entre las HC y las HT, mientras que la expresión de MMP-9 sí resultó
significativamente disminuida en los conos ectoplacentarios provenientes de HT (p<0,05).

Capítulo III�
196
�
Figura III.15. Expresión génica de MMP-2 y MMP-9 en cono ectoplacentario al día 8 de gestación. En
muestras provenientes de hembras CD-1 controles (HC) y tratadas con alcohol (HT), se analizó la
expresión de MMP-2 (A) y MMP-9 (B) por qRT-PCR. Los valores fueron relativizados al gen endógeno
Ywhaz. Los valores se expresan como promedio de la cuantificación relativa + SEM. *p<0,05 versus su
control (Prueba T de Student).
3. Efectos del consumo perigestacional de alcohol en la inducción de genotoxicidad materna y/o
embrionaria y en la muerte celular embrio-trofoblástica: Días 4 y 8 de gestación.
Postulamos la existencia de un posible efecto genotóxico del etanol sobre embriones provenientes
de madres consumidoras de alcohol perigestacionalmente. Bajo esta hipótesis, fue evaluada la
morfología nuclear de embriones de preimplantación y postimplantación temprana. A su vez, con
el fin de detectar un posible daño genotóxico en la madre expuesta, se determinó la frecuencia de
micronúcleos en eritrocitos policromáticos de médula ósea materna. También postulamos que
tanto la potencial pérdida o alteración de las uniones de adhesión embrionaria, que llevarían a la
modificación de la expresión de E-cadherina y ZO-1, así como cambios en la expresión de MMPs en
los tejidos embrionarios y extraembrionarios, son factores que podrían inducir muerte celular por
apoptosis en los embriones expuestos. Por lo tanto, fue evaluado el grado de apoptosis en
embriones periimplantatorios luego de la exposición a alcohol.
3.1. Estudio genotóxico e incidencia de muerte celular programada en embriones de
preimplantación: Día 4 de gestación.
Se procedió al análisis genotóxico de mórulas y blastocistos mediante la observación de las
características de los núcleos interfásicos en extendidos nucleares (técnica de Tarkowski) de cada

Capítulo III�
197
�
tipo embrionario provenientes de HC y HT de las colonias CD-1 y CF-1. En cada embrión fue
evaluada y registrada la presencia de micronúcleos (MNs), puentes cromatínicos, lóbulos o
gemaciones, y picnosis nuclear. En las HT, tanto CD-1 como CF-1, la frecuencia de núcleos
embrionarios con MNs fue similar a la frecuencia encontrada en las HC de ambas colonias, siendo
dicho valor casi nulo. Tampoco se encontraron diferencias significativas en el número de núcleos
con puentes o gemaciones, entre las HC y las HT, el cual resultó cercano a cero. Sin embargo, se
encontraron núcleos picnóticos tanto en los embriones derivados de HC como en aquellos
derivados de HT. Como se observa en la Tabla III.1, los blastocistos iniciales de las HT-CD-1
presentaron un incremento significativo del número de núcleos con cromatina altamente
condensada comparado con el número de núcleos picnóticos del mismo tipo embrionario
proveniente de HC (p<0,05). En las HT-CF-1, también se encontró alta frecuencia de picnosis
nuclear, pero en este caso fue en blastocistos expandidos (p<0,001) (Tabla III.1).
Tabla III.1. Genotoxicidad embrionaria luego del consumo materno de alcohol.
CD-1 HC HT
Mórulas Bl Inic Bl Exp Mórulas Bl Inic Bl Exp
N° total de núcleos 104 575 1296 378 650 1650
N° núcleos picnóticos 4 4 1 15 15 0
N° núcl picn/núcl totales (%) 3,8 0,7 0,08 3,9 2,3* 0
CF-1 HC HT
Mórulas Bl Inic Bl Exp Mórulas Bl Inic Bl Exp
N° total de núcleos 191 738 754 198 662 627
N° núcl picnóticos 0 0 0 0 3 17
N° núcl picn/núcl totales (%) 0 0 0 0 0,45 2,7***
Embriones provenientes de las colonias CD-1 y CF-1, colectados al día 4 de gestación, fueron
clasificados en mórulas, blastocistos iniciales (Bl Inic) o blastocistos expandidos (Bl Exp) y sometidos a la
técnica de Tarkowski para evaluación de la calidad nuclear. Núcl: núcleos, picn: picnóticos. La cantidad
de núcleos picnóticos se expresa como porcentaje sobre el número total de núcleos contados. *p<0,05
y ***p<0,001 versus su control (Prueba Chi-cuadrado).

Capítulo III�
198
�
Dados estos resultados, se prosiguió con el ensayo de TUNEL para verificar la inducción de la
muerte celular mediada por apoptosis en los embriones de las HT.
Para ello, en primer lugar se analizó el nivel de apoptosis (como el número de células TUNEL-
positivas) en blastocistos iniciales (Figura III.16.A) y expandidos (Figura III.16.B-D) provenientes de
HC, comparando asimismo, la presencia de células TUNEL-positivas en el MCI, en el TP y en el TM,
en cada tipo embrionario (Figura III.17.A). Como puede observarse, los blastocistos iniciales de las
HC presentaron un mayor número de células apoptóticas por embrión (p<0,05) respecto de los
blastocistos expandidos, y dicha diferencia se manifestó principalmente a nivel del MCI (p<0,05).
Tanto en los blastocistos iniciales como expandidos, la cantidad de células apoptóticas en el TP y/o
TM fue reducida y no hubo diferencias entre ambos tipos embrionarios. Cuando se analizó el
índice de apoptosis en los blastocistos de las HT respecto de los embriones de las HC, se encontró
un mayor grado de apoptosis en los blastocistos iniciales (Figura III.16.E y F) y expandidos (Figura
III.16.G y H) de las HT versus los blastocistos iniciales (Figura III.16.A) y expandidos (Figura III.16.B,
C y D) de las HC. Estos datos se ven reflejados en la Figura III.17.B y Figura III.17.C en cuanto al
número total de células TUNEL-positivas por tipo embrionario. De la cuantificación del número de
células marcadas positivamente, se desprendió que el TP de los blastocistos iniciales de las HT
presentó significativamente mayor cantidad de células TUNEL-positivas que los de las HC (Figura
III.17.B). Por otra parte, en los blastocistos expandidos de las HT, las zonas con mayor incidencia
de apoptosis correspondieron tanto al MCI como al TM (Figura III.17.C).

Capítulo III�
199
�
Figura III.16. Ensayo de TUNEL in-toto en blastocistos al día 4 de gestación. Micrografías
representativas de embriones de preimplantación de hembras CF-1 controles (HC: A-D) y tratadas con
alcohol (HT: E-H). A, blastocisto inicial con 3 células TUNEL-positivas en el MCI (flecha); B, blastocisto
expandido con 2 células TUNEL-positivas en MCI (flecha) y 1 en el TM (punta de flecha); C, blastocisto
expandido con 1 célula TUNEL-positivas en TM (punta de flecha); D, blastocisto expandido con 1 célula
TUNEL-positiva en MCI (flecha); E-F, blastocistos iniciales con abundante células TUNEL-positiva en el
MCI (flechas) y en el TE (puntas de flecha); G-H, blastocistos expandidos con abundantes células
TUNEL-positivas en el MCI (flechas) y en el TE (puntas de flecha). I-J, blastocisto inicial y expandido,
respectivamente, incubados con DNAsa 0,5 µg/ml (controles positivos). K-L, blastocisto inicial y
expandido, respectivamente, donde se omitió la enzima TdT (controles negativos). MCI: macizo celular
interno, TdT: terminal deoxynucleotidyl transferase, TE: trofoectodermo, TM: trofoectodermo mural,
TP: trofoectodermo polar. Escala: 20 µm.
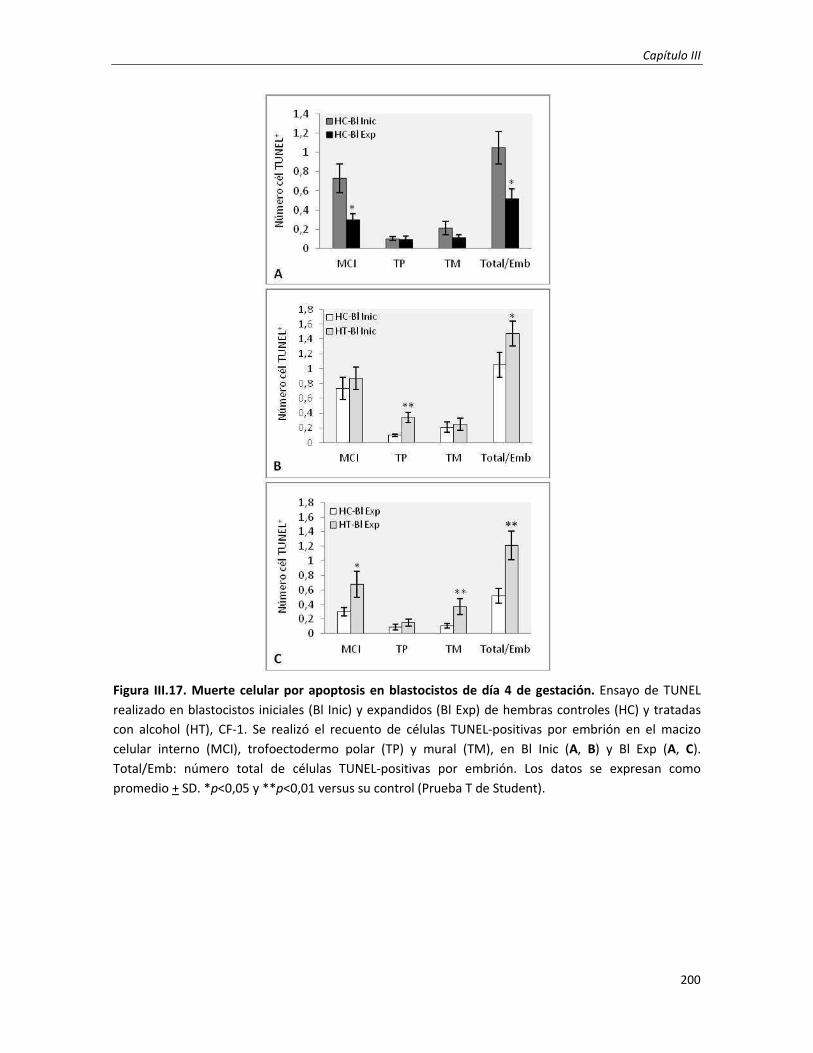
Capítulo III�
200
�
Figura III.17. Muerte celular por apoptosis en blastocistos de día 4 de gestación. Ensayo de TUNEL
realizado en blastocistos iniciales (Bl Inic) y expandidos (Bl Exp) de hembras controles (HC) y tratadas
con alcohol (HT), CF-1. Se realizó el recuento de células TUNEL-positivas por embrión en el macizo
celular interno (MCI), trofoectodermo polar (TP) y mural (TM), en Bl Inic (A, B) y Bl Exp (A, C).
Total/Emb: número total de células TUNEL-positivas por embrión. Los datos se expresan como
promedio + SD. *p<0,05 y **p<0,01 versus su control (Prueba T de Student).

Capítulo III�
201
�
3.2. Estudio genotóxico e incidencia de muerte celular programada en embriones
postimplantación temprana: Día 8 de gestación.
Dado que encontramos una elevada incidencia de picnosis nuclear y aumento de apoptosis en
blastocistos luego de la ingesta materna de alcohol hasta el día 4 de gestación, en forma más
acentuada en hembras de la colonia CF-1 que en CD-1, nos propusimos investigar si este efecto
adverso también se producía en los tejidos embrionarios, extraembrionarios y trofoblásticos, en el
período de gastrulación luego de la ingesta de alcohol hasta el día 8 de gestación. De esta manera,
se intentó determinar si los embriones de postimplantación en estadio más avanzado del
desarrollo resultan igualmente susceptibles a los efectos adversos de la exposición a alcohol que
los del estadio preimplantatorio, en cuanto a cambios en la morfología nuclear y grado de
apoptosis.
Mediante marcación nuclear con Hoechst 33342 de cortes histológicos de sitios de implantación,
se identificaron cambios en la morfología nuclear analizándose la organización, distribución
nuclear, la forma y tamaño relativo de los núcleos y la condensación de la cromatina, en los
embriones provenientes de hembras expuestas a alcohol y comparando con los controles. Las
alteraciones nucleares más evidentes se hallaron en la interfase trofoblástico-decidual y en el
ectodermo embrionario. La interfase materno-embrionaria lateral de los sitios de implantación de
las HT mostró desorganización nuclear (Figura III.18.B, C y D) respecto a lo observado en las
muestras controles (Figura III.18.A). A su vez, la zona antimesometrial de la interfase trofoblástico-
decidual de las HT se caracterizó por presentar alta fragmentación y núcleos con morfología
irregular, siendo algunos de ellos llamativamente de gran tamaño (Figura III.18.F, G y J) comparado
con el tamaño y forma homogénea observado en las muestras controles (Figura III.18.E). En los
embriones de las HT, el ectodermo también mostró alteraciones morfológicas nucleares (Figura
III.18.I y J) respecto de los embriones de las HC (Figura III.18.H). En términos generales, en todas
las muestras de HT se detectaron fragmentos nucleares y alta proporción de núcleos con
cromatina altamente condensada (con tamaño reducido y alta intensidad fluorescente; Figura
III.18).
�

Capítulo III�
202
�
Figura III.18. Marcación nuclear con Hoechst 33342 en sitios de implantación al día 8 de gestación.
Micrografías representativas de sitios de implantación de hembras CF-1 controles (HC: A, E, H) y
tratadas con alcohol (HT: B, C, D - F, G - I, J). A, E, H: núcleos de HC con morfología poliédrica y tamaño
homogéneo. B, C, D: interfase trofoblástico-decidual lateral (InT-DL) de las HT con gran desorganización
nuclear. F, G, J: interfase trofoblástico-decidual antimesometrial (InT-DA) con fragmentación nuclear
(flechas) y núcleos de tamaños heterogéneos (*). I, J: núcleos del ectodermo embrionario de las HT
con morfología irregular y fragmentación (flechas). EctoE: ectodermo embrionario, EndoP: endodermo
parietal, EndoV: endodermo visceral, SV: saco vitelino. Núcleos con cromatina altamente condensada:
puntas de flecha. Aumentos: A-G, J: 40x; H, I: 60x.
La muerte celular por apoptosis en tejido embrio-trofoblástico al día 8 de gestación fue verificada
realizando el ensayo de TUNEL. Los controles negativo y positivo, así como un detalle de la marca
positiva se presentan en la Figura III.19.A, B y C, respectivamente. Como se muestra en la Figura
III.20.A, la zona de expansión del cono ectoplacentario de los embriones provenientes de HC se

Capítulo III�
203
�
caracterizó por presentar gran cantidad de células con marcación positiva, al igual que en las HT
(Figura III.20.B). El número promedio de células apoptóticas, en dicha zona, no resultó
significativamente diferente entre las HC y HT (Figura III.21.A). La zona proliferativa del cono no
presentó células TUNEL-positivas ni en las muestras controles ni en las tratadas. La interfase
trofoblástico-decidual de las HT presentó mayor número de células apoptóticas respecto de las HC
(Figura III.21.B). Esto se observó, en las HT, tanto en la interfase lateral (respecto del embrión)
(Figura III.20.D) como en la zona de interfase antimesometrial (zona de la cripta de implantación)
(Figura III.20.F), mientras que las hembras controles presentaron escasas células apoptóticas en
dichas regiones (Figura III.20.C y E). Finalmente, se analizaron los tejidos embrionarios y
extraembrionarios. Se encontraron células apoptóticas principalmente en el embrión, no así en el
tejido extraembrionario. En particular, el ectodermo embrionario de las HT presentó un alto
número de células apoptóticas (Figura III.20.F; Figura III.21.C) respecto de las hembras controles
(Figura III.20.E; Figura III.21C). Los tejidos mesodérmicos como endodérmicos de las HT
presentaron un escaso número de células TUNEL-positivas, y este valor no resultó diferente entre
los tratamientos.
El índice de apoptosis en los tejidos analizados de los sitios de implantación de las HC y HT de la
colonia CF-1 resultó similar al índice encontrado para las HC y HT de la colonia CD-1.
Figura III.19. Ensayo de TUNEL en sitios de implantación al día 8 de gestación. Micrografías
representativas de un control negativo, donde se omitió la enzima TdT (A); un control positivo
incubado con DNAsa 0,5 µg/ml (B); detalle de células TUNEL-positiva (puntas de flecha, C). Dam:
decidua antimesometrial, De: decidua mesometrial, E: embrión. Escala: A y B: 200 µm, C: 20 µm.
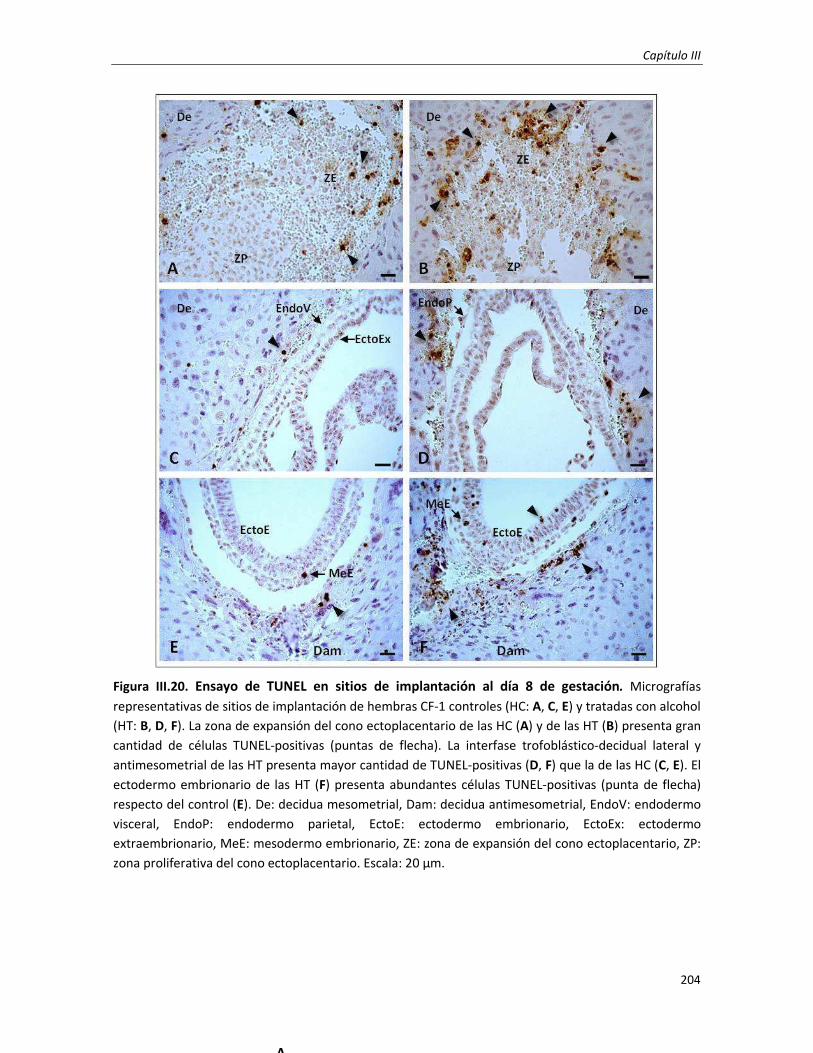
Capítulo III�
204
�
Figura III.20. Ensayo de TUNEL en sitios de implantación al día 8 de gestación. Micrografías
representativas de sitios de implantación de hembras CF-1 controles (HC: A, C, E) y tratadas con alcohol
(HT: B, D, F). La zona de expansión del cono ectoplacentario de las HC (A) y de las HT (B) presenta gran
cantidad de células TUNEL-positivas (puntas de flecha). La interfase trofoblástico-decidual lateral y
antimesometrial de las HT presenta mayor cantidad de TUNEL-positivas (D, F) que la de las HC (C, E). El
ectodermo embrionario de las HT (F) presenta abundantes células TUNEL-positivas (punta de flecha)
respecto del control (E). De: decidua mesometrial, Dam: decidua antimesometrial, EndoV: endodermo
visceral, EndoP: endodermo parietal, EctoE: ectodermo embrionario, EctoEx: ectodermo
extraembrionario, MeE: mesodermo embrionario, ZE: zona de expansión del cono ectoplacentario, ZP:
zona proliferativa del cono ectoplacentario. Escala: 20 µm.
A
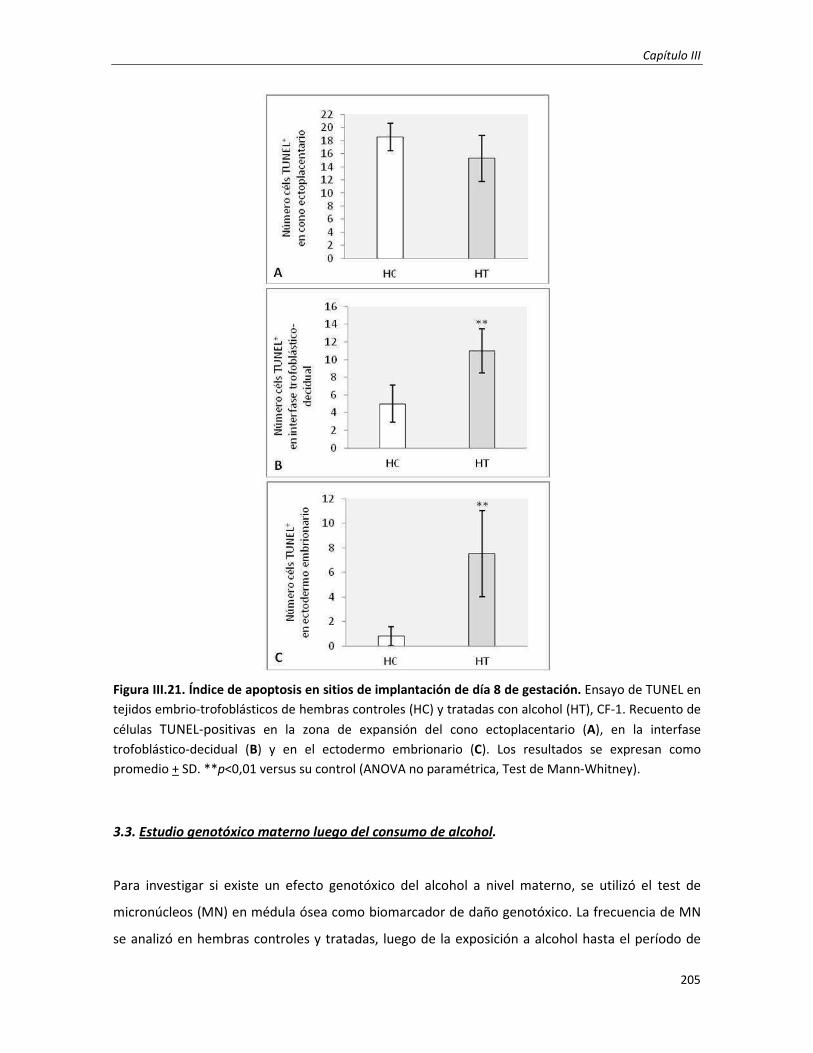
Capítulo III�
205
�
Figura III.21. Índice de apoptosis en sitios de implantación de día 8 de gestación. Ensayo de TUNEL en
tejidos embrio-trofoblásticos de hembras controles (HC) y tratadas con alcohol (HT), CF-1. Recuento de
células TUNEL-positivas en la zona de expansión del cono ectoplacentario (A), en la interfase
trofoblástico-decidual (B) y en el ectodermo embrionario (C). Los resultados se expresan como
promedio + SD. **p<0,01 versus su control (ANOVA no paramétrica, Test de Mann-Whitney).
3.3. Estudio genotóxico materno luego del consumo de alcohol.
Para investigar si existe un efecto genotóxico del alcohol a nivel materno, se utilizó el test de
micronúcleos (MN) en médula ósea como biomarcador de daño genotóxico. La frecuencia de MN
se analizó en hembras controles y tratadas, luego de la exposición a alcohol hasta el período de

Capítulo III�
206
�
preimplantación (día 4 de gestación). Para las HC el número de MN cada 1000 eritrocitos
policromáticos fue de 2,1 + 1,2, mientras que dicho valor se duplicó en las HT (4,07 + 1,5; p<0,01).
�

Capítulo III�
207
�
III.3. CONCLUSIONES PARCIALES
1. En cuanto al efecto de la ingesta perigestacional de etanol 10%, desde antes de la fecundación
y hasta la fase preimplantatoria (día 4 de gestación) o postimplantatoria temprana (día 8 de
gestación) sobre las moléculas de adhesión celular, se puede concluir que:
Tanto la localización/distribución como el patrón de expresión de proteínas de las uniones
adherentes (E-cadherina) y estrechas (ZO-1) en blastocistos iniciales y expandidos fueron
afectados negativamente; mientras que se indujo una reducción de la expresión de E-cadherina en
el ectodermo del embrión en gastrulación.
2. En cuanto al efecto de la ingesta perigestacional de etanol 10%, desde antes de la fecundación
y hasta la fase postimplantatoria temprana (día 8 de gestación) en la expresión proteica y
génica de MMP-2 y MMP-9, se puede concluir que:
La expresión génica de MMP-2 en el cono ectoplacentario, no fue modificada, pero sí aumentó la
expresión proteica en el trofoblasto proliferativo del cono ectoplacentario, en el endodermo
extraembrionario y en ectodermo embrionario.
La expresión génica de MMP-9 sí se redujo significativamente en el tejido trofoblástico del cono
ectoplacentario, lo que correlacionó con la reducción de la expresión proteica en la zona
proliferativa, en la zona de expansión y en las CGTs de la interfase trofoblástico-decidual, mientras
en el ectodermo embrionario la expresión proteica de la MMP-9 resultó incrementada
3. En cuanto al efecto del consumo perigestacional de etanol 10%, desde antes de la fecundación
y hasta la fase preimplantatoria (día 4 de gestación) o postimplantatoria temprana (día 8 de
gestación) a nivel genotóxico materno – embrionario, y en relación a la muerte celular embrio-
trofoblástica, se puede concluir que:
En blastocistos expuestos se indujo un aumento en la incidencia de picnosis nuclear coincidiendo
con el incremento en la muerte celular por apoptosis, tanto en el macizo celular interno como en
el trofoectodermo. En embriones en gastrulación y en la interfase trofoblástico-decidual se
produjeron tanto alteraciones morfológicas nucleares como apoptosis. Luego de la exposición a

Capítulo III�
208
�
alcohol hasta el día 4 de gestación, se incrementó la frecuencia de micronúcleos en eritrocitos de
médula ósea de las hembras expuestas y por lo tanto se indujo genotoxicidad materna.
Estos resultados sugieren que la exposición perigestacional a alcohol puede alterar la expresión de
proteínas de las uniones adherentes y estrechas, modificar los niveles de expresión proteica y
génica de MMPs, inducir daño genotóxico y apoptosis embrio-trofoblástica. Estas alteraciones
podrían ser causa de las anomalías morfológicas, del retraso de diferenciación y del crecimiento
embrionario, y de las alteraciones trofoblásticas durante la fase de invasión en el tejido materno
observadas luego de la ingesta de alcohol, entre los días 4 y 8 de gestación.


Capítulo IV�
209
�
Consumo perigestacional de alcohol:
efectos en el desarrollo feto-placentario en la gestación a término.
IV.1. MÉTODOS
1. Modelos animales y diseños experimentales: tratamientos in-vivo.
Se utilizaron hembras y machos adultos pertenecientes la colonia CD-1, en los que se llevó a cabo
el esquema de apareamiento correspondiente a la dicha colonia, como fue detallado en
Materiales y métodos. Brevemente, se describe a continuación.
1.1. Tratamiento peri-gestacional sin régimen de estimulación ovárica.
Hembras CD-1 recibieron etanol 10% (m/v) en el agua de bebida 15 días previos al apareamiento y
hasta el día 4, 8 o 10 de gestación (hembras tratadas, HT). El grupo control (hembras controles,
HC) recibió sólo agua como bebida durante todo el tratamiento. Las hembras fueron alojadas con
machos para el apareamiento entre los días 15 y 17 de tratamiento. Al día siguiente, la
observación del tapón vaginal indicó el día 1 de gestación. Las hembras con tapón vaginal
continuaron recibiendo alcohol (HT) hasta el día 4, 8 o 10 de preñez, luego de lo cual, el alcohol
fue reemplazado por agua hasta el día 18 de gestación (gestación a término). En la mañana del día
18 (entre las 9:00 y 12:00 hs), las HC y HT fueron sacrificadas por dislocación cervical para extraer
los fetos y placentas para realizar los estudios que se detallan a continuación (Figura IV.1). Se
utilizó un mínimo de 5 animales por grupo.

Capítulo IV�
210
�
Figura IV.1. Representación esquemática del diseño experimental. Tratamiento perigestacional con
alcohol, sin estimulación ovárica. Hembras CD-1 recibieron etanol 10% (hembras tratadas, HT) durante
15 días previos al apareamiento. Las hembras controles (HC) recibieron agua durante el mismo
período. Las hembras fueron puestas en contacto con el macho en días consecutivos y hasta 3 veces,
entre los días 15 y 17 de tratamiento. Cada mañana fue observada la presencia del tapón vaginal. Las
hembras con tapón (día 1 de gestación) continuaron recibiendo alcohol (HT) hasta el día 4 (D4), 8 (D8)
o 10 (D10) de gestación, a partir de los cuales el alcohol fue reemplazado por agua hasta el día 18 de
preñez, día en que fueron sacrificadas para realizar los estudios indicados en el esquema.
2. Recolección de fetos y placentas.
Al 18vo. día de gestación, las hembras de la colonia CD-1 fueron sacrificadas y la cavidad
abdominal fue cuidadosamente abierta para extraer los cuernos uterinos. Los mismos fueron
removidos de la cavidad abdominal, lavados en PBS y transferidos a una placa de parafina donde la
pared uterina y los sacos amnióticos fueron delicadamente diseccionados para liberar los fetos.
Las unidades feto-placentarias fueron separadas de la pared uterina y los fetos aislados de su
placenta a través de un corte en el cordón umbilical. Luego de registrar la viabilidad, el peso y largo
fetal, estos fueron sacrificados en cámara de CO2. Los fetos y las placentas fueron procesados para
los diferentes estudios como se detalla a continuación.

Capítulo IV�
211
�
3. Metodologías para el estudio del desarrollo feto-placentario a término: Día 18 de gestación.
3.1. Monitoreo de la preñez.
Con el fin de evaluar el desempeño reproductivo de las HC y las HT, en cada grupo experimental
fue contabilizado el número total de fetos por hembra (discriminando entre fetos vivos y muertos,
luego de aplicar un suave estímulo táctil directo sobre ellos), el número total de sitios de
implantación (SI) y el número total de reabsorciones, discriminando entre reabsorciones
tempranas y tardías según las características observadas, detalladas a continuación.
Para realizar el recuento del número total de SI, luego de haber retirado los fetos, las placentas y
las reabsorciones, los cuernos uterinos fueron incubados en una solución de sulfato de amonio
10% en agua destilada por 15 min (Salewski, 1964) el cual revela todos los sitios donde ocurrieron
implantaciones (marca regiones uterinas con elevada vascularización). Por lo tanto, el total de
sitios de implantación, evidenciados como puntos oscuros en el tejido uterino, fue cuantificado.
Las reabsorciones fueron clasificadas en tempranas o tardías, según el estadio gestacional en el
que aproximadamente se detuvo el desarrollo. Para ello, se tuvieron en cuenta los criterios que se
mencionan a continuación. Se consideró reabsorción tardía a aquellas unidades feto-placentarias
con signos de reabsorción, conteniendo embrión/feto muerto y en estado de degeneración,
desarrollado hasta los 10 a 16 días de gestación, y con un tamaño mayor a 0,5 cm. Se consideró
reabsorción temprana a embriones que detuvieron su desarrollo aproximadamente entre los 5,5 y
9 días de gestación. En esta categoría se incluyen las reabsorciones tempranas visibles y las no
detectables a simple vista. Las reabsorciones visibles (estadios de arresto del desarrollo: 7 a 9 días
de gestación, aproximadamente) se observaron como pequeños sitios de implantación oscuros en
degeneración, de tamaño entre 0,2 y 0,5 cm. Las reabsorciones no visibles (estadios de arresto del
desarrollo: 5,5 a 6,5 días de gestación, aproximadamente) se observaron como marcas uterinas (o
puntos oscuros) evidenciadas únicamente luego de la incubación con sulfato de amonio.
Por lo tanto, el número de sitios de implantación (SI), se estableció como:
N° total de SI = N° de fetos + N° de reabsorciones tardías + N° reabsorciones tempranas

Capítulo IV�
212
�
Donde: N° de reabsorciones tempranas = reabsorciones tempranas visibles + reabsorciones
tempranas no visibles (evidenciadas con sulfato de amonio, y calculadas como la diferencia entre el
total de SI marcados y el número de fetos, reabsorciones tardías y tempranas visibles).
Los datos fueron registrados como promedio y frecuencia en el total de hembras y en el total de
sitios de implantación utilizados en cada grupo.
3.2. Análisis del crecimiento fetal y placentario.
Para estudiar el crecimiento fetal y placentario, el peso de cada feto vivo y el largo corporal fue
registrado. De la misma manera, las placentas fueron pesadas individualmente. Los valores fueron
expresados como promedio para cada grupo experimental. A su vez, con el fin de analizar el
crecimiento del feto en relación al crecimiento de su placenta, fue determinado el índice
Feto/Placentario (IF/P), calculado como la relación entre el peso fetal y el placentario sobre el total
de las muestras analizadas para cada grupo.
3.3. Análisis de la morfología fetal externa.
Se determinó la incidencia de malformaciones fetales externas luego del consumo materno de
alcohol hasta distintas fases de la gestación. Para ello, luego del sacrificio, los fetos fueron fijados
por inmersión en una solución de formaldehído 5%, en la cual fueron mantenidos hasta su
evaluación. Los fetos fueron examinados bajo estereomicroscopio y de manera sistemática, en
cuanto a la morfología craneofacial, del cuerpo y de las extremidades, como se detalla en la Tabla
IV.1. Cada cambio o alteración morfológica estructural fue clasificada y registrada. Asimismo, fue
registrada la presencia de más de una malformación por feto (combinación) y las malformaciones
severas (fetos amorfos). Por lo tanto, la frecuencia de fetos malformados (expresada como
porcentaje), así como el tipo de anomalía encontrada en ellos, fue calculada sobre el número total
de fetos analizados en cada grupo experimental.

Capítulo IV�
213
�
Tabla IV.1. Tipos de malformaciones fetales asociadas al síndrome de alcoholismo fetal.
Craneofaciales Cuerpo Miembros
Microcefalia Fisura palpebral corta Onfalocele Cambios en el número
Anencefalia Baja inserción
de orejas Gastrosquisis Cambios en el tamaño
Acefalia Surco nasolabial liso Hidropesía Cambios en la posición
Acrania
Labio superior fino Cambios en la forma
Exencefalia Fisura labial (labio
leporino)
Cambios en el número
de dedos (ectrodactilia,
hipodactilia, polidactilia)
Microftalmia Fisura palatina Cambios en el
tamaño de dedos
Anoftalmia Macroglosia Fusión de dedos (Sindactilia)
Exoftalmia Micrognatia
Al día 18 de gestación los fetos fueron analizados en cuanto a la presencia de alteraciones estructurales
externas, frecuentemente asociadas al FAS. Microcefalia: cabeza de tamaño reducido según lo esperado
para la edad gestacional; anencefalia: ausencia parcial o total del cerebro y cráneo debido a defecto en el
cierre del tubo neural; acefalia: ausencia de la cabeza; acrania: ausencia de huesos de la bóveda craneana,
puede estar asociada a anencefalia o a exencefalia; exencefalia: cerebro situado fuera del cráneo;
microftalmia: ojos con escaso desarrollo respecto a lo esperado; anoftalmia: ausencia completa de uno o
ambos ojos; exoftalmia: exteriorización del globo ocular; fisura palpebral corta: acortamiento, respecto de
lo esperado, de la distancia entre la comisura interior y exterior del ojo generando ojos pequeños; anomalía
en la forma o posición de la oreja: como la baja inserción de oreja, con inserción anormal o pliegues del
pabellón auricular dismórficos, generalmente asociados con anomalías estructurales internas; surco
nasolabial liso: la pequeña depresión entre el labio superior de la boca y la nariz no está definida por las dos
crestas la piel resultando en un aspecto liso; fisura labial: hendidura en el labio superior, por una fusión
incompleta de los procesos maxilar y nasomedial, y se presenta frecuentemente con fisura palatina; fisura
palatina: el paladar presenta una grieta que comunica la boca con la cavidad nasal; macroglosia:
malformación de la lengua siendo ésta más grande de lo normal y que en posición de reposo, protruye hacia
afuera de la boca; micrognatia: la mandíbula es menor a lo esperado; onfalocele: malformación asociada
con defectos en el cierre de la pared abdominal en el que las vísceras, cubiertas por una membrana, se
presentan fuera del cuerpo fetal; gastrosquisis: malformación en la que los órganos de la cavidad abdominal
(generalmente intestino) quedan expuestos al exterior del cuerpo sin membranas envolventes, por fusión
incompleta de los pliegues laterales del cuerpo; hidropesía: presencia de edema debido a una cantidad
excesiva de líquido en la cavidad abdominal, alrededor del corazón o de los pulmones o edema generalizado
del cuerpo; ectrodactilia: ausencia congénita de todo o parte de uno o más dedos. Abarca una variedad de
estados desde la afalangia a la adactilia; hipodactilia/polidactilia: menor/mayor cantidad de dedos según lo
esperado; sindactilia: dedos fusionados.

Capítulo IV�
214
�
3.4. Análisis de la morfología fetal interna: sistema esquelético.
Con el fin de analizar la aparición de malformaciones óseas luego de la exposición materna a
alcohol, un mínimo de dos fetos por hembra separados al azar de cada tratamiento fue analizado.
Luego de la fijación con formaldehído al 5%, en cada feto se realizó un corte transversal debajo de
las costillas y dos cortes oblicuos, uno del lado derecho y otro del lado izquierdo, hasta la región
anal, formando un pequeño triangulo. Utilizando una pinza de puntas finas, las vísceras
abdominales, las torácicas y el diafragma fueron retirados de la cavidad abdominal. La
evisceración fue realizada con absoluta delicadeza evitando lesiones y fracturas del esqueleto.
Posteriormente, se prosiguió con la etapa de diafanización de los tejidos blandos y coloración del
esqueleto con alizarina red-S. Para ello, los fetos fueron colocados en una solución de hidróxido de
potasio (KOH) 1% en agua destilada por 96 hs hasta alcanzar el macerado/clareado total de la piel
y de los músculos, quedando los huesos visibles. Luego, la solución de diafanización fue
reemplazada por una solución colorante compuesta por alizarina red-S 0,25% en KOH 1% durante
72 hs, momento en el que se logró la coloración ósea completa. Dicha solución fue renovada cada
día. La alizarina, por su capacidad de unirse a sitios con depósitos de calcio, permite la
visualización del esqueleto. La intensidad de la coloración rojiza depende del grado de osificación
del hueso, por lo tanto, si la osificación fuera incipiente, la coloración roja resultante será tenue.
Finalmente, la solución de alizarina fue reemplazada por una solución clareadora constituida por
alcohol bencílico/glicerina/etanol absoluto (1:2:2 v/v), donde los fetos fueron mantenidos por 72
hs. Una vez aclarados, los fetos fueron conservados en glicerina pura hasta el análisis óseo.
Para el examen de las alteraciones del esqueleto, cada feto fue retirado de la solución de
almacenamiento, colocado en una placa de Petri y observado bajo estereomicroscopio. El
esqueleto fue examinado de manera sistemática de cabeza a cola, y las anomalías encontradas
fueron registradas.
Para el examen de las alteraciones óseas, los huesos del cráneo, del rostro y del cuerpo fueron
analizados en cuanto al tamaño, forma, posición, fusiones y número (adiciones y/o ausencias)
(Tabla IV.2, Figura IV.2). Cada alteración fue registrada y expresada como porcentaje de fetos con
alteraciones esqueléticas sobre el total de fetos analizados. A su vez, un análisis más detallado fue
realizado en los huesos craneofaciales, el cual involucró la evaluación del grado de osificación
(osificación completa o incompleta) y el grado de densidad ósea según la coloración de los huesos.
Para ello, se estableció un score basado en la intensidad de coloración con alizarina,

Capítulo IV�
215
�
correspondiendo un score de 3 a la coloración más intensa, un valor de 2 a una coloración media y
1 a una coloración tenue, como se ejemplifica en la Figura IV.3. El número de fetos con osificación
ósea incompleta, con densidad ósea disminuida (score menor o igual a 2) o con la combinación de
ambas anomalías, fue registrado y expresado como porcentaje respecto al número total de fetos
analizados. La intensidad de coloración con alizarina, como indicador de la densidad ósea, fue
expresada como promedio + SD.
Tabla IV.2. Análisis del sistema esquelético en fetos a término.
Principales piezas óseas
Craneofaciales Hueso nasal, frontal, parietal, intra-parietal, supra-occipital y exoccipital;
escamoso, cigomático, pre-maxilar, maxilar
Vértebras Un centro y dos arcos en cada vértebra; 7 cervicales, 13 torácicas, 6 lumbares, 5
sacras y caudales de número variable
Caja torácica 13 costillas pares; esternón conformado por 6 puntos de osificación: manubrio,
4 centros esternales y 1 apéndice xifoides; 2 clavículas; 2 escápulas
Miembros
anteriores
3 huesos largos: húmero, radio, cúbito;
4 metacarpos; 4 o 5 falanges
Pelvis 2 crestas ilíacas; isquion; pubis
Miembros
posteriores
3 huesos largos: tibia, fémur, fíbula (peroné)
5 metatarsos; 4 o 5 falanges
Al día 18 de gestación los fetos fueron analizados en cuanto a la presencia de alteraciones óseas en los
principales huesos del cráneo, rostro, columna, tórax, pelvis y miembros.

Capítulo IV�
216
�
Figura IV.2. Osificación fetal. Imágenes representativas de fetos coloreados con alizarina. El análisis de
la morfología esquelética fue realizado examinando los huesos señalados en la figura en cuanto a
tamaño, forma, posición, fusiones y número. A, 1-10 principales huesos craneofaciales. 1: nasal, 2:
frontal, 3: parietal, 4: intra-parietal, 5: supra-occipital, 6: exoccipital, 7: escamoso, 8: cigomático, 9: pre-
maxilar, 10: maxilar. 11: vértebras cervicales (7). 12: escápula (2). 13: húmero (2). B, 14: costillas (13
pares). C, 15: esternón, 16: clavícula (2). D, 17: vértebras torácicas (13), 18: vértebras lumbares (6), 19:
vértebras sacras (5), 20: crestas ilíacas, 21: pubis, 22: isquion. E, 23: fémur, 24: tibia, 25: fíbula, 26:
metatarsos (5), 27: falanges (4 o 5), 28: vértebras caudales (número variable). F, 29: radio, 30: cúbito,
31: metacarpos (4), 32: falanges (4 o 5).

Capítulo IV�
217
�
Figura IV.3. Densidad ósea. Imagen representativa de fetos con diferente densidad ósea manifestada
por el nivel de coloración con alizarina alcanzado luego de 72 hs de coloración. A la izquierda, feto con
densidad ósea adecuada para la edad gestacional, considerada como densidad ósea grado 3
(coloración intensa). A la derecha, feto con densidad ósea menor a la esperada para su edad
gestacional, clasificada como grado 1 (coloración tenue generalizada).
3.5a. Procesamiento histológico de las placentas.
Para el análisis histológico placentario fue empleada la técnica histoquímica basada en la reacción
de PAS (del inglés, Periodic Acid Schiff), la cual permite la detección de hidratos de carbono, libres
o conjugados, cuando están presentes en los tejidos en cantidades relativamente grandes,
generando un color magenta intenso. Para ello, las placentas fueron cortadas transversalmente en
dos partes iguales y fijadas en paraformaldehído 4% por 24 hs a 4°C. Transcurrido ese período
fueron lavadas en agua y conservadas en alcohol 70% a 4°C hasta la inclusión en parafina. Para
ello, las placentas fueron deshidratadas por pasajes en alcoholes de gradación creciente, aclaradas
en xilol y embebidas en parafina. Secciones transversales de 5 µm de espesor correspondientes a
la porción central de la placenta fueron obtenidas. Para llevar a cabo la reacción de PAS, las
muestras fueron desparafinadas en xilol, hidratadas progresivamente en alcoholes y colocadas en
ácido periódico 1% en etanol 40%, 10 min a TA. Luego, las muestras fueron colocadas en etanol
50% por 1 min seguido de la inmersión en reactivo de Schiff por 7 min. Posteriormente, fueron
lavadas en ácido sulfuroso (2 x 5 min) y se prosiguió con la coloración de contraste con
hematoxilina. Las muestras fueron fotografiadas en microscopio equipado con cámara de video
con un analizador de imágenes Nikon.

Capítulo IV�
218
�
3.5b. Análisis morfométrico y cualitativo de las placentas.
A partir de imágenes 2,5x digitalizadas de cortes placentarios, tratados según la técnica
histoquímica de PAS, fue realizado un análisis morfométrico con el programa Image Pro Plus de
las zonas constitutivas de la placenta: la zona de unión y zona laberíntica (Figura IV.4). El área de
cada zona fue relativizada al área placentaria total y la relación entre el área laberíntica y el área
de la zona de unión fue calculada. Un mínimo de dos placentas de cada hembra por grupo fue
analizado.
Asimismo, se realizó el análisis histopatológico examinando en detalle las características
morfológicas de las zonas de la placenta. En la zona de unión se observaron las células gigantes
trofoblásticas, las células glicogénicas PAS-positivas y las células espongiotrofoblásticas. En la zona
laberíntica se observó la red de vasos fetales y espacios sinusoides maternos (limitados por una
capa trilaminar de trofoblasto). Para el análisis se tuvo en cuenta: la organización/disposición
celular, integridad de los tejidos, proporción de tipos celulares en cada zona y la presencia o no de
infiltrado leucocitario. En la Figura IV.5, se muestran imágenes representativas de los principales
tipos celulares constitutivos de la placenta a término. Un mínimo de dos placentas de cada
hembra por grupo fue analizado.
Figura IV.4. Zonas de la placenta a término.
Micrografía representativa de placenta coloreada
según histoquímica PAS. Se observa la decidua
(De); la zona de unión (ZU), intensamente
coloreada por la presencia de células glicogénicas
y espongiotrofoblásticas; y la zona laberíntica (ZL),
red vascular de coloración más clara. El trazo
indica las áreas de la ZU y ZL consideradas para el
análisis morfométrico. Escala: 1 mm.

Capítulo IV�
219
�
Figura IV.5. Histología de la placenta a término. Tipos celulares presentes en la placenta al día 18 de
gestación. A, vista topográfica indicando las regiones detalladas en B, C y D; B, células deciduales (De) y
una sección de la zona de unión con células gigantes del trofoblasto (CGT) y células glicogénicas
intensamente marcadas (CG); C, zona de unión, donde se observan células espongiotrofoblásticas
basófilas (EPG) y CG con sus típicos gránulos de glicógeno de color magenta (flechas); D, zona
laberíntica, donde se observan espacios sanguíneos maternos (m) separados de los capilares fetales (cf)
por una fina estructura trilaminar conformada por sinciciotrofoblasto (st) y trofoblasto mononuclear
gigante (ct) y por el endotelio de los capilares fetales (ef). Escalas: A: 1 mm, B: 50 µm, C y D: 25 µm.
4. Análisis estadístico.
Los datos expresados como promedios de los valores ± SD de los grupos controles y grupos
tratados se compararon mediante ANOVA y prueba de Dunnett. Los datos expresados como
frecuencias (porcentajes) se compararon mediante prueba Chi-cuadrado. Se utilizó el programa
estadístico GraphPad Instat v2.05a. Un valor de p<0,05 fue considerado estadísticamente
significativo.

Capítulo IV�
220
�
IV.2. RESULTADOS
Es sabido que el consumo de altas cantidades de alcohol durante la gestación produce
malformaciones fetales, restricción del crecimiento, muerte intrauterina y alteraciones postnatales
a largo plazo. Dado que en nuestro modelo, la ingesta de alcohol desde antes de la gestación y
hasta períodos gestacionales tempranos induce anomalías embrionarias en estadios de
preimplantación, implantación y gastrulación, nos propusimos evaluar los efectos feto-placentarios
del consumo perigestacional de alcohol a largo plazo, en el desarrollo a término, aún cuando la
exposición a alcohol cesa en etapas tempranas/medias de la preñez (día 4, 8 o 10). Para ello, se
administró alcohol al 10% desde antes de la gestación, hasta el estadio de preimplantación, día 4
de gestación; hasta el estadio de gastrulación, día 8 de gestación; o hasta el estadio de
organogénesis, día 10 de gestación.
Efectos del consumo perigestacional de alcohol en el desarrollo feto-placentario a término:
Día 18 de gestación.
1.1. Estudio de los efectos nutricionales.
Con el fin de descartar efectos nutricionales en el desarrollo fetal y placentario, asociados al
consumo de alcohol, fue monitoreado el peso corporal de las hembras al inicio de los tratamientos
con alcohol, al último día de tratamiento (día 4, 8 o 10) y a término (día 18 de preñez), junto con el
consumo calórico final en cada grupo experimental. Como se muestras en la Tabla IV.3, al finalizar
el período de alcoholización (días 4, 8 y 10 de gestación), en ninguno de los casos fueron
encontradas diferencias significativas en el peso corporal entre los grupos controles y tratados. De
la misma manera, el peso de las hembras tratadas al día 18 de gestación no difirió
significativamente del peso de las controles. Tampoco fueron diferentes los patrones diarios de
ingesta calórica entre el grupo control y el tratado en cada esquema experimental al final del
período de exposición.
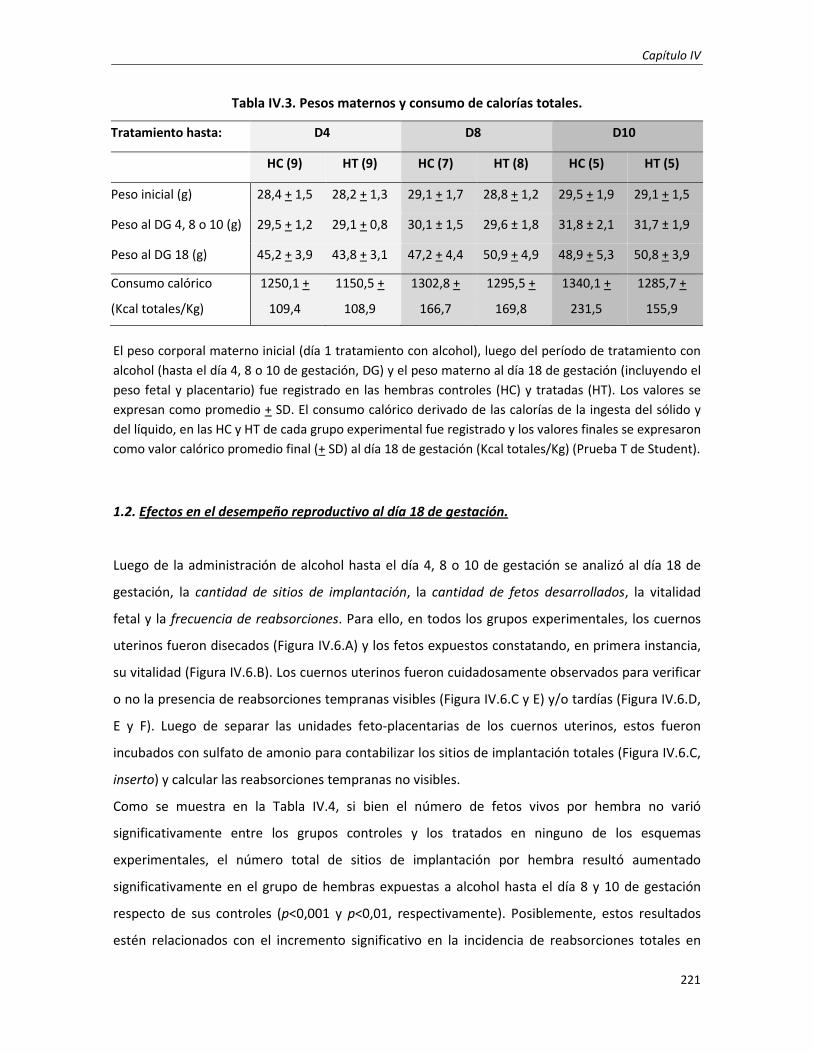
Capítulo IV�
221
�
Tabla IV.3. Pesos maternos y consumo de calorías totales.�
Tratamiento hasta: D4 D8 D10
HC (9) HT (9) HC (7) HT (8) HC (5) HT (5)
Peso inicial (g) 28,4 + 1,5 28,2 + 1,3 29,1 + 1,7 28,8 + 1,2 29,5 + 1,9 29,1 + 1,5
Peso al DG 4, 8 o 10 (g) 29,5 + 1,2 29,1 + 0,8 30,1 ± 1,5 29,6 ± 1,8 31,8 ± 2,1 31,7 ± 1,9
Peso al DG 18 (g) 45,2 + 3,9 43,8 + 3,1 47,2 + 4,4 50,9 + 4,9 48,9 + 5,3 50,8 + 3,9
Consumo calórico�
(Kcal totales/Kg)
1250,1 +
109,4
1150,5 +
108,9
1302,8 +
166,7
1295,5 +
169,8
1340,1 +
231,5
1285,7 +
155,9
El peso corporal materno inicial (día 1 tratamiento con alcohol), luego del período de tratamiento con
alcohol (hasta el día 4, 8 o 10 de gestación, DG) y el peso materno al día 18 de gestación (incluyendo el
peso fetal y placentario) fue registrado en las hembras controles (HC) y tratadas (HT). Los valores se
expresan como promedio + SD. El consumo calórico derivado de las calorías de la ingesta del sólido y
del líquido, en las HC y HT de cada grupo experimental fue registrado y los valores finales se expresaron
como valor calórico promedio final (+ SD) al día 18 de gestación (Kcal totales/Kg) (Prueba T de Student).
1.2. Efectos en el desempeño reproductivo al día 18 de gestación.
Luego de la administración de alcohol hasta el día 4, 8 o 10 de gestación se analizó al día 18 de
gestación, la cantidad de sitios de implantación, la cantidad de fetos desarrollados, la vitalidad
fetal y la frecuencia de reabsorciones. Para ello, en todos los grupos experimentales, los cuernos
uterinos fueron disecados (Figura IV.6.A) y los fetos expuestos constatando, en primera instancia,
su vitalidad (Figura IV.6.B). Los cuernos uterinos fueron cuidadosamente observados para verificar
o no la presencia de reabsorciones tempranas visibles (Figura IV.6.C y E) y/o tardías (Figura IV.6.D,
E y F). Luego de separar las unidades feto-placentarias de los cuernos uterinos, estos fueron
incubados con sulfato de amonio para contabilizar los sitios de implantación totales (Figura IV.6.C,
inserto) y calcular las reabsorciones tempranas no visibles.
Como se muestra en la Tabla IV.4, si bien el número de fetos vivos por hembra no varió
significativamente entre los grupos controles y los tratados en ninguno de los esquemas
experimentales, el número total de sitios de implantación por hembra resultó aumentado
significativamente en el grupo de hembras expuestas a alcohol hasta el día 8 y 10 de gestación
respecto de sus controles (p<0,001 y p<0,01, respectivamente). Posiblemente, estos resultados
estén relacionados con el incremento significativo en la incidencia de reabsorciones totales en

Capítulo IV�
222
�
dichos grupos, en comparación con los controles, alcanzando valores cercanos al 14% (p<0,01) en
el grupo HT-D4, 7% en el grupo HT-D8 y 11% en el grupo HT-D10 (p<0,05 versus sus respectivos
controles), como puede observarse en la Tabla IV.5. Las reabsorciones fueron clasificadas y
cuantificadas en términos del momento gestacional en que, aproximadamente, pudo haber
ocurrido el arresto en el desarrollo. Llamativamente, el número de reabsorciones tempranas sólo
resultó significativamente elevado, en el grupo tratado hasta el día 4 de gestación (p<0,05 versus
su control).
Figura IV.6. Desempeño reproductivo al día 18 de gestación. Imágenes representativas de las
unidades feto-placentarias (UFPs) disecadas al día 18 de gestación. A, disposición de las UFPs en los
cuernos uterinos; B, Recuento de fetos, evaluación de su vitalidad y clasificación de reabsorciones
visibles; C, útero con reabsorción temprana (Rte) visible (porción de tejido menor a 0,5 cm, punta de
flecha); C inserto, cuerno uterino luego de la incubación con sulfato de amonio. Se observan los tres
sitios de implantación correspondientes a las UFPs y la Rte mostrados en C; D, útero con una
reabsorción tardía (RTa, tejido mayor a 0,5 cm con placenta (Pl), punta de flecha); E, comparación de
tamaños entre feto normal, RTa (punta de flecha blanca) y RTes visibles (punta de flecha negra); F,
detalle de la RTa mostrada en E donde se observa desarrollo placentario y feto (F) detenido
aproximadamente en estadio medio de gestación. Escalas: B-E: 1 cm, F: 0,5 cm.

Capítulo IV�
223
�
Tabla IV.4. Desempeño reproductivo al día 18 de gestación.�
Tratamiento hasta: D4 D8 D10
HC HT HC HT HC HT
N° hembras 9 9 7 8 5 5
N° SI totales 97 104 79 111 57 72
N° SI/hembra 10,8 + 2,3 11,5 + 1,9 11,3 + 1,3 13,9+1,2*** 11,4 + 1,4 14,4+1,8**
N° fetos totales 96 90 77 103 56 64
N° fetos/hembra 10,7 + 2,3 10,0 + 2,4 11,2 + 1,8 12,9 + 2,3 11,2 + 1,2 12,8 + 2,2
N° fetos vivos (%) 96 (100) 88 (97,8) 77 (100) 101 (98,0) 56 (100) 62 (96,9)
N° fetos vivos/hembra 10,7 + 2,3 9,8 + 2,4 11,2 + 1,8 12,6 + 1,9 11,2 + 1,2 12,4 + 2,2
Monitoreo de la preñez al día 18 de gestación. El número de sitios de implantación (SI), de fetos totales
y vivos fue registrado sobre el total de hembras utilizadas en cada grupo (hembras tratadas hasta día 4,
8 o 10 de gestación, HT; y sus respectivos controles, HC). Los valores se expresan como promedios + SD
sobre el número total de hembras. El número de fetos vivos se expresa como porcentaje del total de
fetos. **p<0,01 y ***p<0,001 versus su control (Prueba T de Student y Chi-cuadrado, según
corresponda).

Capítulo IV�
224
�
Tabla IV.5. Frecuencia de reabsorciones al día 18 de gestación.�
Tratamiento hasta: D4 D8 D10
HC HT HC HT HC HT
N° hembras 9 9 7 8 5 5
N° SI totales 97 104 79 111 57 72
N° Rte/hembra 0,11+0,33 1,00+1,22* 0,14+0,36 0,63+1,06 0,0+0,0 1,40+1,14
N° Rta/hembra 0,0 + 0,0 0,55+1,01 0,0+0,0 0,37+0,52 0,20+0,42 0,20+0,45
N° reabs tot/hembra 0,11+0,33 1,55+1,66* 0,14+0,36 1,10+1,07* 0,20+0,42 1,60+1,14*
N° Rte (%) 1/97 (1,0) 9/104 (8,7)a 1/79 (1,3) 5/111 (4,5) 0/57 (0) 7/72 (9,7)
N° Rta (%) 0/97 (0,0) 5/104 (4,8) 0/79 (0,0) 3/111 (2,7) 1/57 (1,7) 1/72 (1,4)
N° reabs totales
(%)
1/97
(1,0)
14/104
(13,5)b
1/79
(1,3)
8/111
(7,2)a
1/57
(1,7)
8/72
(11,1)a
Monitoreo de la preñez al día 18 de gestación. El número de reabsorciones totales (reabs tot),
tempranas (RTe) y tardías (RTa) fue registrado sobre el total de hembras utilizadas en cada grupo
(hembras tratadas hasta día 4, 8 o 10 de gestación, HT; y sus respectivos controles, HC). Los resultados
se expresan como promedio + SD sobre el número total de hembras o como frecuencia (porcentaje, %).
*p<0,05 versus su control (Prueba T de Student); a: p<0,05 y b: p<0,01 respecto de su control (Prueba
Chi-cuadrado).
1.3. Desarrollo a término del feto y la placenta.
1.3 .1. Estudio del crecimiento fetal y placentario.
Dado que uno de los principales efectos del consumo gestacional de alcohol es la restricción del
crecimiento intrauterino, nos propusimos analizar el grado de crecimiento de los fetos vivos
conjuntamente con el crecimiento de su placenta. Como puede observarse en la Tabla IV.6, el peso
promedio de los fetos solamente resultó significativamente incrementado en el grupo que recibió
alcohol hasta el día 8 de gestación respecto del control (p<0,001), no encontrándose cambios ni en
el peso ni en el largo fetal en ninguno de los grupos analizados. Por otro lado, el peso placentario
sí mostró una disminución significativa en todos los grupos tratados, siendo más significativo para

Capítulo IV�
225
�
el grupo HT-D10. Esta disminución en el peso placentario conlleva una alteración en el índice de
peso feto/placentario, el cual resultó incrementado significativamente en todos los grupos
tratados respecto de los controles, indicando una desproporción entre el crecimiento del feto y la
placenta.
Tabla IV.6. Crecimiento fetal y placentario al día 18 de gestación.
Tratamiento hasta: D4 D8 D10
HC HT HC HT HC HT
N° fetos o placentas 96 88 77 101 56 62
Peso fetal (g) 1,3+0,13 1,3+0,16 1,4+0,16 1,5+0,12*** 1,4+0,12 1,3+0,21
Largo fetal (cm) 3,3+0,27 3,2+0,32 3,5+0,21 3,3+0,17 3,4+0,11 3,4+0,31
Peso placentario (g) 0,15+0,02 0,14+0,03* 0,16+0,01 0,15+0,02* 0,14+0,02 0,13+0,03***
Índice peso F/P 9,15+1,34 9,98+1,53** 9,11+1,50 9,88+1,41** 9,72+1,2 11,03+1,98***
El crecimiento de los fetos (F) y las placentas (P) fue evaluado a través de la medición de los pesos
(gramos, g) y tamaños (largo fetal, cm) en cada grupo experimental (hembras tratadas hasta día 4, 8 o
10 de gestación, HT; y sus respectivos controles, HC). Los resultados se expresan como promedio + SD
sobre el número total de fetos o placentas en cada grupo. El Índice peso F/P fue calculado como el
peso fetal sobre el peso placentario en el total de muestras.*p<0,05, **p<0,01 y ***p<0,001 versus su
control (Prueba T de Student).
1.3.2. Estudio de las anomalías morfológicas fetales externas.
Con el fin analizar posibles efectos teratogénicos de la exposición perigestacional a alcohol hasta
estadios tempranos (día 4 u 8) o medios (día 10) de la gestación, los fetos fueron evaluados en
cuanto la incidencia de malformaciones externas (véase Tabla IV.1 de Métodos). La Figura IV.7
muestra imágenes representativas de las alteraciones encontradas en todos los grupos tratados
con alcohol. Además de observarse fetos con una única malformación, en muchos casos dichas
alteraciones se encontraron combinadas (más de una por feto). Las malformaciones encontradas
con mayor frecuencia fueron las craneofaciales, las cuales incluyeron el labio leporino
(evidenciado por la protrusión de la lengua), exoftalmia, baja inserción de oreja, y la acrania con

Capítulo IV�
226
�
exencefalia, términos que hacen referencia a la exposición del cerebro por ausencia de huesos del
cráneo (Figura IV.7.B y C). La Figura IV.7.D, muestra un feto con edema generalizado (hidropesía) y
anoftalmia. En las hembras expuestas a alcohol, también fueron encontrados fetos con defectos
en el cierre de la pared abdominal, con exposición de vísceras, como el onfalocele (Figura IV.7.E) y
gastroquistis (Figura IV.7.C), en este último caso, asociado con alteraciones en las extremidades,
como la presencia de miembros rotados respecto de su posición normal (Figura IV.7.C). La
presencia de dedos supranumerarios (Figura IV.7.F) también fue observada. Además se
encontraron fetos con malformaciones múltiples (Figura IV.7.G) e inclusive fetos o masas de tejido
amorfo con sólo algunas estructuras diferenciadas (Figura IV.7.H).
En cada grupo tratado, la frecuencia de fetos con malformación fue calculada y comparada con sus
respectivos controles, resultado que se resume en la Tabla IV.7. Las hembras controles tuvieron un
bajo porcentaje de fetos con malformaciones siendo la mayoría de ellas de tipo craneofacial (baja
inserción de oreja). Sin embargo, en todos los grupos tratados se encontró un elevado porcentaje
de fetos con combinación de malformaciones. Sorprendentemente, no sólo el grupo que recibió
alcohol durante la organogénesis (HT-D10) mostró defectos teratogénicos, también se vio
incrementada significativamente la frecuencia de fetos con anomalías cuando el alcohol se ingirió
hasta la etapa de gastrulación (HT-D8) o hasta la etapa preimplantatoria (HT-D4), comparando con
sus controles. Así, el porcentaje total de fetos con malformaciones resultó incrementado en todos
los grupos tratados. Las alteraciones craneofaciales más severas, como la acrania y exencefalia,
fueron encontradas en los grupos tratados hasta el día 8 y día 10. A su vez, éste último grupo se
caracterizó por presentar una mayor proporción de fetos con múltiples malformaciones
(combinadas).
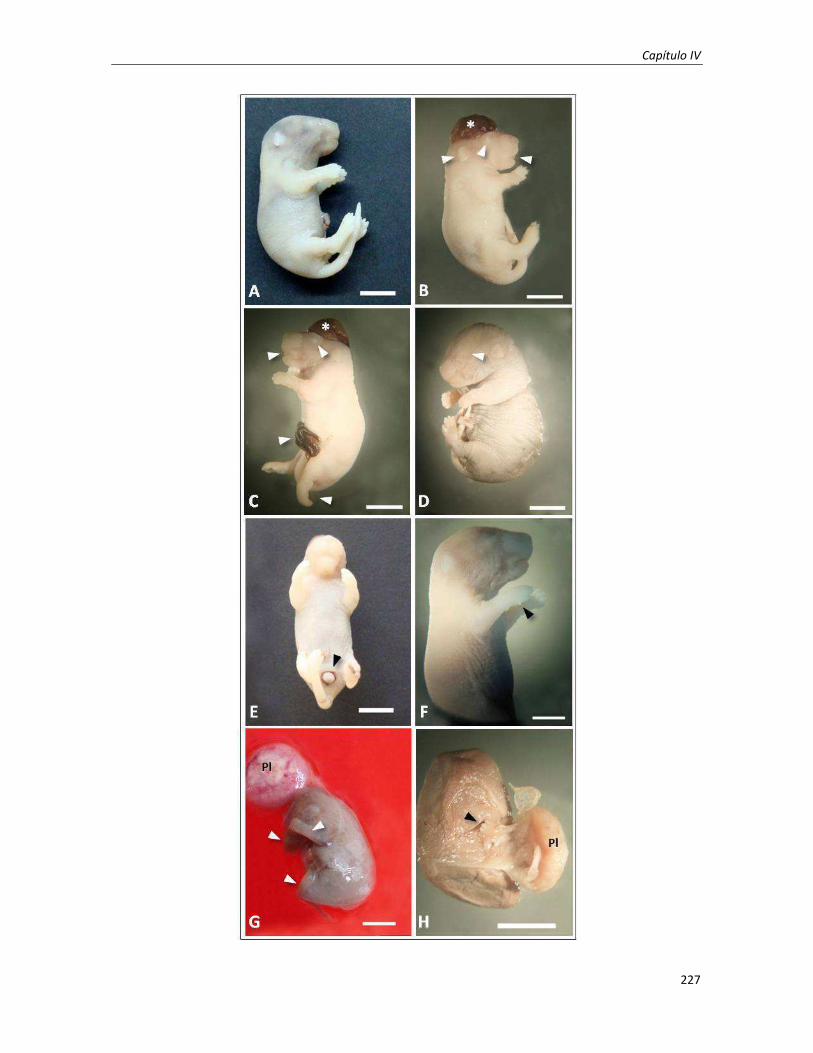
Capítulo IV�
227
�

Capítulo IV�
228
�
Figura IV.7. Malformaciones fetales externas al día 18 de gestación. Imágenes representativas de
fetos con malformaciones. A, feto normal; B, feto con anomalías craneofaciales: acrania con
exencefalia (*), exoftalmia, labio leporino (protrusión de la lengua) y baja inserción de oreja (puntas de
flecha); C, feto con las mismas anomalías craneofaciales que B, con gastrosquistis (con exposición de
intestino) y miembro posterior rotado (puntas de flecha); D, feto con hidropesía, tamaño corporal
disminuido y anoftalmia (punta de flecha); E, feto con exposición de vísceras recubiertas por una
membrana (onfalocele, punta de flecha); F, feto con polidactilia en miembro anterior (punta de flecha);
G, feto con desarrollo facial y corporal posterior incompleto, y con esbozos de miembros anteriores
(puntas de flecha); H, feto sin diferenciación, amorfo, sólo se distingue el desarrollo de los dedos de
una de las extremidades (punta de flecha). Pl: placenta. Escala: 0,5 cm.
Tabla IV.7. Frecuencia de malformaciones fetales externas.
Tratamiento hasta: D4 D8 D10
HC HT HC HT HC HT
N° fetos 96 90 77 103 56 64
MALFORMACIÓN
Sólo craneofacial (%) 12 (100) 32 (86,5) 8 (88,9) 53 (76,8) 7 (100) 34 (70,8)
Sólo miembros (%) 0 0 1 (11,1) 7 (10,1) 0 0
Sólo cuerpo (%) 0 0 0 0 0 4 (8,3)
Combinadas (%) 0 3 (8,1) 0 7 (10,1) 0 8 (16,7)
Severos/amorfos (%) 0 2 (5,4) 0 2 (2,9) 0 2 (4,2)
N° total de fetos
malformados (%)
12/96
(12,5)
37/90
(41,1)***
9/77
(11,7)
69/103
(66,9)***
7/56
(12,5)
48/64
(75,0)***
El número de fetos malformados y el tipo de anomalía fue registrado en cada grupo experimental
(hembras tratadas hasta día 4, 8 y 10 de gestación, HT; y sus respectivos controles, HC). Para cada tipo
de malformación, se cuantificó el número de fetos con alguna malformación sobre el total de fetos
malformados y el total de fetos malformados sobre el total de fetos analizados (%) en cada grupo
experimental. ***p<0,001 versus su control (Prueba de Chi-cuadrado).
1.3.3. Estudio de las anomalías morfológicas fetales internas: análisis del esqueleto.
Dado que el consumo materno de alcohol puede provocar alteraciones en el esqueleto fetal, nos
propusimos evaluar las anomalías esqueléticas mediante el recuento bajo estereomicroscopio del
número de huesos, y la observación del tamaño, la forma y la posición de los principales huesos

Capítulo IV�
229
�
del cráneo, rostro, tórax, pelvis, columna y miembros, considerando también el grado de
osificación y densidad ósea craneofacial.
La Figura IV.8.A, muestra una imagen representativa de un feto proveniente de un grupo control
con desarrollo esquelético normal, en cuanto al número de huesos, forma y tamaño, y osificación
craneofacial adecuada. Por el contrario, en los fetos de las hembras tratadas de los tres grupos
expuestos a alcohol, las alteraciones óseas fueron evidentes, encontrándose fetos con variaciones
en la densidad ósea (manifestada por el grado de coloración con alizarina) y en el grado de
osificación. Las Figuras IV.8.B, C y D muestran ejemplos de fetos con diversas anomalías, como
densidad ósea disminuida generalizada y curvatura anormal de la columna vertebral (Figura
IV.8.B); osificación incompleta del cráneo, presencia de costillas con desarrollo rudimentario,
ausencia de falanges (Figura IV.8.C). En la Figura IV.8.D, se muestra un feto con ausencia de huesos
de la bóveda craneana (acrania). Es importante destacar que, si bien las alteraciones esqueléticas
más significativas encontradas en los tres grupos tratados se observaron a nivel de craneofacial,
otras alteraciones tales como ausencia de costillas (Figura IV.9.A), alineamiento irregular de los
centros esternales (Figura IV.9.B), ausencia de falanges y de vértebras caudales (Figura IV.9.C)
fueron encontradas en los grupos tratados. Sin embargo, la frecuencia de fetos con alteraciones
de esta índole no resultó estadísticamente significativa respecto del porcentaje hallado en los
controles (datos no mostrados).
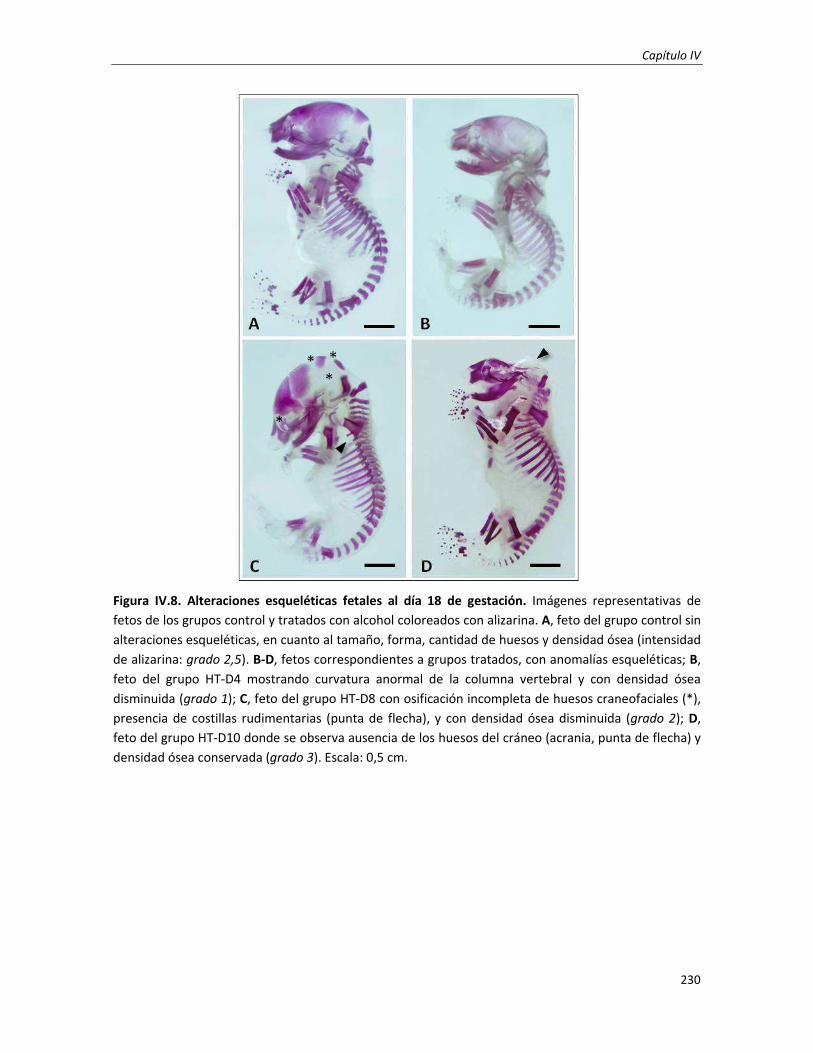
Capítulo IV�
230
�
Figura IV.8. Alteraciones esqueléticas fetales al día 18 de gestación. Imágenes representativas de
fetos de los grupos control y tratados con alcohol coloreados con alizarina. A, feto del grupo control sin
alteraciones esqueléticas, en cuanto al tamaño, forma, cantidad de huesos y densidad ósea (intensidad
de alizarina: grado 2,5). B-D, fetos correspondientes a grupos tratados, con anomalías esqueléticas; B,
feto del grupo HT-D4 mostrando curvatura anormal de la columna vertebral y con densidad ósea
disminuida (grado 1); C, feto del grupo HT-D8 con osificación incompleta de huesos craneofaciales (*),
presencia de costillas rudimentarias (punta de flecha), y con densidad ósea disminuida (grado 2); D,
feto del grupo HT-D10 donde se observa ausencia de los huesos del cráneo (acrania, punta de flecha) y
densidad ósea conservada (grado 3). Escala: 0,5 cm.

Capítulo IV�
231
�
Figura IV.9. Detalle de otras alteraciones esquelética. Imágenes representativas de fetos provenientes
de hembras tratadas con alcohol coloreados con alizarina donde se observan algunas alteraciones
esqueléticas y detalle a nivel del cuerpo (caja torácica, miembros y vértebras). A, ausencia de la 9na
costilla (punta de flecha); B, esternón con alineación anormal de los cuatro centros esternales (punta
de flecha); C, ausencia de falanges en miembros posteriores (puntas de flechas) y ausencia de
vértebras caudales (*).
Debido a la alta incidencia de alteraciones craneofaciales, se analizó el grado de osificación
craneana, observando el grado de fusión de dichos huesos (Figura IV.10). Fue considerada
osificación normal cuando la fusión fue casi completa (Figura IV.10.A, D y E) y osificación
incompleta (OI) cuando ésta fue baja (Figura IV.10.B y C). La densidad ósea fue determinada según
el grado de coloración con alizarina. Para ello se estableció un score de valoración de la intensidad
de color (véase Métodos). Así, se estableció como densidad ósea normal cuando la coloración
rojiza de la alizarina fue intensa (Figura IV.10.D). En contraste, coloraciones más pálidas como las
mostradas en la Figura IV.10.E y F, fueron consideradas indicadores de baja densidad ósea (BDO).

Capítulo IV�
232
�
Figura IV.10. Grado de osificación y densidad ósea craneofacial. Imágenes representativas de fetos
coloreados con alizarina donde se observan alteraciones esqueléticas de cráneo en detalle. A-C, vista
de la osificación del cráneo; A, osificación normal; B, osificación incompleta con ausencia del hueso
escamoso (*) y menor desarrollo del hueso parietal (punta de flecha); C, osificación incompleta del
hueso frontal (*). E-F, alteraciones en la densidad ósea según intensidad de coloración con alizarina; D,
feto del grupo control con valor de densidad ósea = 3; E, feto del grupo HT-D10 con valor de densidad
ósea = 2; F, feto del grupo HT-D4 con valor de densidad ósea = 1.
Así, el número de fetos con osificación incompleta (OI) craneana fue evaluado en el total de fetos
procesados por la técnica de alizarina. Los resultados fueron expresados como frecuencias sobre el
número total de fetos con anomalías craneanas. También se analizó el número de fetos con baja
densidad ósea craneana (BDO, valores entre 2,5 y 1) el que se expresó como frecuencia sobre el
total de fetos con anomalías craneanas. Se registró el número de fetos con combinación de ambas
anomalías (baja densidad ósea y osificación incompleta) y se realizó un promedio del valor de
densidad ósea en todos los fetos analizados. Como se observa en la Tabla IV.8, en todos los grupos
que recibieron alcohol hubo una alta incidencia de fetos con anomalías craneanas. Entre el 87% y
90% de los fetos de madres tratadas presentaron alguna alteración ósea craneofacial, siendo dicho
valor cercano al 30-35% en los controles (p<0,001 para HT-D4, p<0,01 para HT-D8 y HT-D10). La
cantidad de fetos con baja densidad ósea de los huesos del cráneo, con osificación incompleta o
combinación de ambas alteraciones, no resultó significativamente diferente entre los grupos
tratados y sus respectivos controles. Sin embargo, la valoración relativa de la intensidad de
coloración como indicador de la densidad ósea sí mostró diferencias significativas en todos los

Capítulo IV�
233
�
grupos tratados respecto de sus controles. Mientras que en los fetos controles, la intensidad de
alizarina resultó en un promedio cercano al valor 3 (establecido como máximo), en los fetos de los
grupos tratados dichos valores resultaron entre 1,8 y 2 (p<0,001 para HT-D4 y HT-D8, y p<0,01 para
HT-D10).
Tabla IV.8. Grado de osificación y densidad ósea craneofacial fetal.
Tratamiento hasta: D4 D8 D10
HC HT HC HT HC HT
N° fetos analizados 18 18 14 16 10 10
ANOMALÍA
Sólo BDO (%) 0 (0,0) 7 (43,7)
1 (20,0)
7 (50,0)
1 (33,3)
2 (22,2)
Intensidad alizarina + SD 2,75+0,45 1,81+0,53a 2,75+0,42 1,80+0,63
a 2,85+0,51 2,15+0,47
b
Sólo OI (%) 2 (33,3) 0 (0,0) 2 (40,0) 2 (14,3) 1 (33,3) 2 (22,2)
Combinación: BDO + OI (%) 4 (66,6) 9 (56,0) 2 (40,0) 3 (21,4) 1 (33,3) 1 (11,1)
Acrania (%) 0 (0,0) 0 (0,0) 0 (0,0) 2 (14,3) 0 (0,0) 4 (44,4)
N° total de fetos con
anomalías craneanas (%)
6/18
(33,3)
16/18
(88,8)***
5/14
(35,7)
14/16
(87,5)**
3/10
(30,0)
9/10
(90,0)**
El número de fetos con baja densidad ósea (-Sólo BDO-, considerando en tal categoría a fetos con
intensidad de alizarina con valores entre 2,5 y 1), el número de fetos con osificación incompleta (-Sólo
OI-), el número de fetos con ambas anomalías (-Combinación: BDO+OI-) y el número de fetos con
acrania fue registrado en cada grupo experimental y se expresa como porcentaje del número total de
fetos con anomalías craneanas. Los valores de intensidad de coloración (densidad ósea) se expresan
como promedio + SD de todos los fetos procesados. El número total de fetos con anomalías
esqueléticas craneofaciales fue registrado y se expresa como porcentaje del total de fetos analizados.
Un mínimo de dos fetos por hembra fue analizado. **p<0,01 y ***p<0,001 versus su control (Prueba
Chi-cuadrado); a: p<0,001 y b: p<0,01 versus su control (Prueba T de Student).
1.3.4. Estudio de la placenta a término.
Diversas sustancias pueden interferir y generar cambios en la estructura y función placentaria
conduciendo a pérdida gestacional y/o anomalías fetales. La evaluación histológica de la placenta,
luego de la exposición a alcohol durante su establecimiento y desarrollo temprano, es

Capítulo IV�
234
�
fundamental para analizar potenciales consecuencias adversas en el crecimiento del feto a
término que estuvieran relacionadas con alteración morfológica de la placenta. Con este objetivo,
las placentas de día 18 de gestación, luego de la ingesta perigestacional de alcohol hasta las fases
de periimplantación (días 4 y 8) o de organogénesis (día 10), fueron procesadas según la
histoquímica de PAS y analizadas, en primer lugar, en cuanto al desarrollo de las capas que forman
la cara fetal (la zona de unión, ZU; y la zona laberíntica, ZL) mediante evaluación morfométrica de
dichas capas. Luego fueron evaluadas en cuanto a las características histológicas de la ZU
(constituida por células gigantes del trofoblasto, CGTs; células glicogénicas, CGs; y células
espongiotrofoblásticas, EPGs); y de la ZL (constituida por vasos fetales, trofoblasto y espacios
sanguíneos maternos).
1.3.4a. Morfología de la cara fetal de la placenta: zona de unión y zona laberíntica.
Un minucioso examen histológico fue realizado en la cara fetal de la placenta observándose la ZU y
la ZL. La Figura IV.11 muestra imágenes topográficas (A-D) donde se evidencia la ZU de coloración
violeta intenso por contener CGs PAS-positivas y las células EPGs de coloración más basófila. En
contraste, la ZL, es de coloración clara. Se pueden observar cambios en las proporciones de la ZU
respecto de la ZL en las placentas de las HT (Figura IV.11.B-D) respecto de la ZU de las placentas de
las hembras controles (Figura IV.11.A). En todas las placentas provenientes de los grupos tratados,
se observaron extensiones celulares de la ZU hacia el laberinto, y en algunos casos, islas de este
tejido en el área laberíntica. En la Figura IV.11.B`-D` se muestran en detalle dichas proyecciones
celulares hacia el laberinto, correspondientes a CGs y a células EPGs, presentes en las placentas
provenientes de madres expuestas en comparación con las placentas de las controles (Figura
IV.11.A`).
1.3.4b. Áreas relativas de la zona de unión y zona laberíntica: análisis morfométrico.
La observación de las alteraciones en la relación de las áreas de la ZU y de la ZL, anteriormente
descriptas, llevó al análisis morfométrico de dichas áreas con el fin de corroborar las diferencias
observadas en las placentas de los grupos de HT versus las de las controles. Como se muestra en la
Tabla IV.9, en comparación con un valor representativo control de los promedios de las placentas
de controles (HC-D4, HC-D8 y HC-D10), en todas las placentas pertenecientes a los grupos tratados

Capítulo IV�
235
�
(HT-D4, HT-D8 y HT-D10) se observó una alteración en la proporción de la ZU respecto del área
total. Si bien, el área total no se modificó significativamente en los grupos tratados versus el
control, el área correspondiente a la ZU (constituida por CGTs, CGs y células EPGs) resultó
aumentada mientras que el área laberíntica se redujo significativamente comparando con el
control. Así, la relación entre el área de la zona del laberinto y la zona de unión, resultó disminuida
significativamente en las placentas de las HT respecto del control, indicando que, luego de los
diferentes períodos de exposición temprana a alcohol, las placentas d las HT desarrollaron área
laberíntica menor que el control.

Capítulo IV�
236
�

Capítulo IV�
237
�
Figura IV.11. Desarrollo placentario al día 18 de gestación. Micrografías representativas de cortes de
placentas de hembras controles (HC) y tratadas con alcohol (HT) (técnica de PAS), donde se observa la
decidua (De), la zona de unión (ZU) y la zona laberíntica (ZL). A-D, vistas topográficas (2,5x); A´-D´,
detalles (10x) de las zonas recuadradas en las imágenes topográficas. A-A´, placenta de HC. Se observa
integridad histológica en la ZU y la ZL, con un límite definido continuo y lineal entre ambas. B-B´,
placenta del grupo HT-D4. Puede observarse intromisión de la ZU en la ZL. C-C`, placenta perteneciente
al grupo HT-D8. Se observan extensiones irregulares de la ZU hacia el área laberíntica y una gran laguna
sanguínea cercana a la zona decidual (*). D-D`, placenta del grupo HT-D10. Se observa la misma
extensión irregular o protrusión de la ZU hacia la ZL, formándose islas de células pertenecientes a la ZU
en la zona vascular de la placenta. CG: células glicogénicas (PAS-positivas), EPG: células
espongiotrofoblásticas (basófilas).
Tabla IV.9. Morfometría de las placentas a término.
Tratamiento hasta: D4 D8 D10
HC HT HT HT
RELACIÓN
ZU/área total 0,42 + 0,05 0,55 + 0,03** 0,55 + 0,08** 0,51 + 0,04*
ZL/área total 0,57 + 0,05 0,45 + 0,03** 0,45 + 0,08** 0,48 + 0,04*
ZL/ZU 1,37 + 0,33 0,83 + 0,10** 0,83 + 0,16** 1,08 + 0,15*
El área total placentaria, el área de la zona de unión (ZU) y de la zona laberíntica (ZL) y la relación entre
el área de ZU y ZL fue calculada en cada grupo experimental (hembras tratadas hasta día 4, 8 o 10 de
gestación, HT) y comparada con el promedio representativo de las áreas de los controles (HC). Los
valores se expresan como promedio + SD. *p<0,05 y **p<0,01 (Test de Dunnett versus el control).
1.3.4c. Análisis histológico de la zona de unión.
El estudio de las características de la ZU involucró la observación de la región de la ZU limitante
con la decidua (posición mesometrial) y de la zona en contacto con el laberinto (lado
antimesometrial). La ZU del lado mesometrial, en contacto con la decidua, está conformada por
CGTs, de grandes dimensiones, que forman el límite entre la ZU y la decidua. En contacto con
éstas, hacia el centro de la ZU y en contacto con el laberinto (hacia el lado antimesometrial) se
encuentran las CGs (de menor dimensión, núcleos pequeños y con citoplasma con gránulos de
glicógeno), entre clusters de células EPGs (redondas y voluminosas). En las muestras controles, las

Capítulo IV�
238
�
células, en estas dos interfases de la ZU, se encuentran organizadas y ordenadas, donde las CGTs
forman una capa periférica limitante con la decidua, posición que hace referencia a una de sus
funciones como frente de invasión (Figura IV.12.A). Las CGs, que forman agrupaciones entre las
células EPGs, forman parte de la región central de la ZU y hacen contacto con el laberinto (Figura
IV.12.A`) proveyendo a este tejido de los nutrientes (CGs) y aporte hormonal (EPGs) necesarios
para el crecimiento fetal. Las placentas de las HT presentaron alteraciones en la organización y
distribución celular dentro de la ZU respecto de morfología de las placentas de las hembras HC. En
la interfase ZU-mesometrial y decidua, las CGTs se observaron entre las CGs, perdiendo su
característica localización periférica limitante. A su vez, próximo a la zona decidual, se evidenció
con frecuencia la presencia de lagunas sanguíneas y un llamativo aumento de CGs (Figuras IV.12.B,
C y D). En la interfase ZU-lado antimesometrial con el laberinto, de la misma manera, se detectó
un aumento en la cantidad de células PAS-positivas respecto de las EPGs, presentando estas
últimas, alteraciones morfológicas (células ahusadas), siendo esta observación más predominante
en el grupo tratado hasta el día 10 de gestación. En particular en este grupo tratado también fue
observado un elevado infiltrado leucocitario (Figuras IV.12. B`, C` y D`).

Capítulo IV�
239
�

Capítulo IV�
240
�
Figura IV.12. Zona de Unión de la placenta a término. Micrografías representativas de placentas de
hembras controles (HC) y tratadas con alcohol (HT) (técnica de PAS), donde se observa la Zona de
Unión (ZU) con sus células características: células gigantes trofoblásticas (CGTs), espongiotrofoblásticas
(EPG) y glicogénicas (CG, PAS-positivas). A-D, interfase ZU-lado mesometrial y decidua (De); A`-D`,
interfase ZU-lado antimesometrial y zona laberíntica (ZL). A-A`, placenta de HC; A, se observan las CGTs
en contacto con la De, las CG rodeadas por clusters de células EPG de características voluminosas; A`,
células EPG y células PAS-positivas en contacto con la ZL. B-B`, placenta del grupo HT-D4; B, se observa
una laguna de sangre (*) entre la De y las CGTs, abundante cantidad de CG y escasa cantidad de células
EPG; B`, las células EPG presentan morfología ahusada respeto del control. C-C`, placenta del grupo HT-
D8; C, se observa congestión vascular (*) entre la De y las CGTs, y abundancia de células PAS-positivas;
C`, hacia la zona laberíntica se observa el mismo patrón que en HT-D4, con alteración de la integridad
del tejido. D-D`, placenta del grupo HT-D10; D, se observa congestión vascular (*), deslocalización de
las CGTs y mayor cantidad de CG respecto de las EPG, como en B y C; D`, se observan las mismas
alteraciones descriptas, junto a un gran infiltrado leucocitario (>).
1.3.4d. Análisis histológico de la zona laberíntica.
El laberinto es un tejido especializado en el intercambio de nutrientes y gases entre la madre y el
feto. En el tejido normal (placenta de HC) se observa una red de espacios sanguíneos maternos
amplios separados de los capilares fetales por una delgada estructura trilaminar de trofoblasto,
conformada por dos capas de sinciciotrofoblasto (st), asociadas al endotelio de los vasos fetales, y
una capa de células trofoblásticas gigantes mononucleares o citotrofoblastos (ct), que delimitan
los espacios sinusoides materno. Si bien, al microscopio óptico no es posible identificar las dos
capas de st, en las muestras controles se percibe el delgado límite entre los espacios sanguíneos
fetales y los vasos maternos (Figura IV.13.A y A`).
La zona del laberinto de las muestras tratadas evidenció cambios significativos respecto de las
muestras controles. La integridad y organización de este tejido se vio claramente afectado. Se
observó un tejido más compacto con evidente engrosamiento de la capa trofoblástica que separa
los espacios sanguíneos (también llamada membrana interhemal), lo cual lleva a una reducción de
dichos espacios de intercambio respecto del control. Además se encontraron células PAS-positivas,
EPGs y leucocitos, a diferencia de las muestras controles donde las mismas no fueron observadas
(Figura IV.13.B y B`, C y C`, D y D`).

Capítulo IV�
241
�

Capítulo IV�
242
�
Figura IV.13. Zona Laberíntica de la placenta a término. Micrografías representativas de placentas de
grupos controles (HC) y tratados con alcohol (HT) (técnica de PAS), donde se observa la Zona
Laberíntica (ZL, A-D) y un detalle de la estructura histológica que separa los compartimientos de sangre
materna y fetal (A`-D`). A-A`, placenta de HC. Una fina capa de trofoblasto y endotelio fetal (<->) separa
los amplios espacios sanguíneos maternos (m) de los capilares fetales (cf). En los tres grupos tratados
(B-B`, HT-D4; C-C`, HT-D8 y D-D`, HT-D10), se observa el tejido más compacto, reducción de los
espacios sanguíneos maternos (m), engrosamiento de la capa trofoblástica (st, ct) y presencia de
células PAS-positivas, EPG y leucocitos (<). ct: citotrofoblasto gigante, ecf: núcleo del endotelio del cf, *:
glóbulos rojos fetales, ^: glóbulos rojos maternos, st: sinciciotrofoblasto.

Capítulo IV�
243
�
IV. 3. CONCLUSIONES PARCIALES
La ingesta perigestacional de alcohol al 10%, desde antes de la fecundación y hasta la fase
preimplantatoria (día 4 de gestación), postimplantatoria temprana (gastrulación, día 8 de
gestación) o postimplantatoria media (organogénesis, día 10 de gestación) produjo, en ratones
de la colonia CD-1, los siguientes efectos en la gestación a término (día 18 de gestación):
Un aumento en la incidencia de reabsorciones embrionarias en todos los grupos tratados.
El crecimiento fetal, en términos de peso corporal, sólo se vio afectado en el grupo tratado hasta
el día 8.
La frecuencia de fetos con malformaciones se incrementó en los tres grupos tratados con alcohol
tanto hasta la organogénesis (HT-D10) como en los grupos tratados hasta períodos gestacionales
más temprano (HT-D8 y HT-D4). También se indujo una alta incidencia de fetos con anomalías
óseas craneanas (baja densidad ósea y/u osificación incompleta) en todos los grupos de hembras
tratadas.
El crecimiento de la placenta (peso placentario) resultó disminuido en todos los grupos tratados,
siendo más pronunciado en el tratado hasta la organogénesis. La cara fetal (zona de unión y la
zona laberíntica) de las placentas provenientes de los grupos tratados fue afectada, observándose
grandes extensiones de la zona de unión hacia el laberinto junto con islas de células glucogénicas y
espongiotrofoblásticas entre la red vascular del laberinto. Esto concuerda con el aumento del área
ocupada por la zona de unión y la disminución del área laberíntica en las placentas de las hembras
tratadas. En la zona de unión se incrementaron las lagunas de sangre, se deslocalizaron las CGT y
aumentó la proporción de CGs respecto de las EPGs. La integridad y organización de la zona del
laberinto de las placentas de las hembras tratadas también se vio afectada, observándose
compactación del tejido, engrosamiento de la capa trofoblástica y consiguiente reducción de
espacios sanguíneos de intercambio.
La ingesta perigestacional de alcohol hasta estadios tempranos/medios del desarrollo embrio-
placentario en hembras murinas CD-1, tuvo consecuencias negativas en la gestación a término,
evidenciadas por desarrollo anómalo del feto, principalmente a nivel craneofacial ya sea por la alta
frecuencia de malformaciones externas como en el nivel de osificación, y alteraciones en el
crecimiento y estructura de la placenta.


Discusión
244
Discusión
Es bien conocido que el consumo gestacional de alcohol produce alteraciones embrionarias,
fetales y placentarias que pueden derivar en muerte embrionaria o fetal, o pueden tener un
impacto en la vida postnatal. Estas alteraciones se expresan en un alto porcentaje de los niños
nacidos de madres consumidoras manifestándose como el Síndrome de Alcoholismo Fetal
(FAS) (Jones y Smith, 1973). El impacto negativo de esta patología no sólo involucra al
individuo que lo padece, sino también a la familia y a la sociedad en su conjunto, razón por la
cual los trastornos asociados al FAS constituyen un importante problema de salud pública. Más
aún, el riesgo de desarrollo de FAS por la ingesta materna de bebidas alcohólicas es motivo de
preocupación internacional, ya que no es un problema de una etnia o grupo social
determinado, sino una problemática del estilo de vida de la sociedad moderna en su conjunto
(Riley y McGee, 2005). Mucho se ganaría si las patologías de programación intrauterina
asociadas al consumo materno de alcohol pudieran ser prevenidas. Tanto la profundización del
conocimiento general de dichas anomalías como la investigación de los mecanismos
moleculares implicados en la generación de las alteraciones embrio-placentarias tempranas,
son objetivos fundamentales para el avance en la resolución de dicha problemática social y de
salud pública. Es por ello, que en el presente trabajo se planteó como objetivo general estudiar
los efectos de la ingesta perigestacional de alcohol en el desarrollo del embrión y del tejido
trofoblástico que va a formar la placenta, en los estadios tempranos de la gestación murina, y
conjuntamente analizar los cambios en algunos mecanismos involucrados en la inducción de
anomalías embrionario-trofoblásticas. Finalmente, se evaluaron las consecuencias de los
efectos a largo plazo de la exposición temprana a alcohol, en el desarrollo fetal y placentario al
momento del nacimiento.
1. Efectos del consumo pregestacional de alcohol en las variaciones de parámetros
biométricos y en el ciclo estral en dos modelos murinos.
El tiempo y el momento de la exposición a alcohol, tanto en el período reproductivo como
durante la gestación, determinaran qué estructuras en desarrollo se verán afectadas y así
mismo, el modo de inducción de la anomalía. Esto ha sido demostrado en estudios
epidemiológicos en la población humana y en modelos animales, en los que se indica
claramente que la variabilidad clínica del FAS está relacionada con el estadio reproductivo y
gestacional en el que la droga actúa. En relación a ello, se pueden distinguir las siguientes

Discusión
245
ventanas de exposición: el período pregestacional (antes de la gestación) y el período
gestacional, incluyendo este último las fases de preimplantación, implantación,
postimplantación (gastrulación y organogénesis) y la fase de desarrollo fetal (Haycock y
Ramsay, 2009). El período perigestacional involucra los días previos al inicio de la gestación y
los días posteriores al inicio de la misma (postimpantación temprana). En cada período, tanto
factores genéticos maternos y embrionarios, como nutricionales, e incluso la edad materna,
juegan un papel importante en el tipo de efecto que se induce tras la exposición a alcohol.
Estos factores afectan el grado de metabolización de la droga y con ello, el nivel de alcohol en
sangre, influyendo en la sensibilidad o susceptibilidad del organismo expuesto. Por esto, no es
sorprendente que cantidades similares de alcohol administradas durante la gestación
produzcan efectos patológicos diferentes (Riley y McGee, 2005; Simpson y col., 2005).
Para estudiar los efectos de la ingesta perigestacional de alcohol, el primer objetivo
establecido en el presente trabajo fue analizar la respuesta animal frente al consumo
pregestacional de alcohol en cuanto al grado de ingesta sólida y líquida, las variaciones en el
peso corporal, los niveles de alcohol en sangre y el ciclo estral, en dos modelos exocriados de
hembras murinas.
En este estudio, se utilizaron ratones hembras sexualmente maduros de colonias exocriadas,
CF-1 y CD-1, como modelos experimentales con respuestas variables semejantes a las
encontradas luego de la ingesta de alcohol en humanos. En términos de la vía de
administración y dosis del etanol, se optó por el tratamiento oral de alcohol suministrado en el
agua de bebida, con lo cual se establece un método de exposición similar a la del consumo
humano y a una concentración equivalente a la del vino (10%). En estas condiciones, un
período corto pero continuo (diario y por 17 días) de ingesta de alcohol, previo a la gestación,
generó una concentración de alcohol en sangre baja, acorde a los valores mínimos de
alcoholemia que se producen por administración vía oral, en contraposición con las
concentraciones elevadas de etanol en sangre obtenidas por tratamiento intraperitoneal o
agudo utilizados por otros autores (Sulik y Johsnston, 1983; Sulik y col., 1984; Lieber y col.,
1989; Tabakoff y Hoffman, 2000). Los efectos de dosis moderadas a bajas de alcohol son
particularmente importantes dado que este es el patrón de consumo más habitual en la
población general, y al cual normalmente no se le atribuye un papel nocivo o perjudicial.
Aunque es conocido que la gama de efectos producidos por la exposición materna a alcohol se
relaciona con las distintas condiciones de consumo como el número de bebidas consumidas y
la concentración de alcohol, el parámetro más importante de referencia a tener en cuenta en

Discusión
246
términos comparativos es la cantidad de alcohol que alcanza al embrión en desarrollo dado
por la concentración de alcohol en sangre (BAC). Esta concentración de alcohol en sangre o
alcoholemia variará de acuerdo al grado de metabolización en cada individuo, siendo ésta el
resultado de la participación de las variables que se han mencionado como moduladoras de los
efectos (dosis, vía de administración, patrón de exposición crónico o agudo, factores genéticos
y nutricionales, entre otros).
En el presente estudio, ambas colonias murinas tratadas por un corto período de 17 días,
presentaron similares niveles de alcoholemia, entre 25 y 30 mg/dl, valores correspondientes a
los producidos por la ingesta de alcohol de tipo “social” en el humano (Izquierdo, 2002). En
relación con ello, valores de BAC entre 50 y 100 mg/dl (con un 36% de calorías derivadas del
alcohol, EDC) son considerados como propios del consumo excesivo de alcohol, mientras que
valores de BAC inferiores a 50 mg/dl (con un 25% de EDC) son propias de un consumo
moderado (Simpson y col., 2005). Los modelos animales de exposición aguda a etanol,
reportados en la literatura para el estudio de diversos aspectos a nivel embrionario o fetal,
también dependiendo del período de gestación de exposición, producen un valor de BAC en el
intervalo de 100 a 400 mg/dl (Kotch y Sulik, 1992a). En modelos de exposición crónica (durante
la gestación hasta estadios embrionarios específicos, y/o durante la lactancia) generalmente
producen valores de BAC inferiores, entre 65 y 220 mg/dl (Amini y col., 1996; Miñana y col.,
2000; Parnell y col., 2006; Boehm y col., 2009). Estos datos demuestran la gran variación en los
niveles de alcoholemia según los tratamientos, lo que indica que, para establecer
comparaciones sobre los efectos producidos, es necesario tomar el BAC como referencia.
Por otro lado, a pesar de la similitud de BAC entre ambas colonias, se pudo observar una leve
diferencia en el nivel de calorías derivadas del alcohol (%EDC), del orden del 17% en CF-1 y del
13% en CD-1. Estos valores posiblemente deriven de las diferencias encontradas en el volumen
de alcohol ingerido en cada colonia, aunque el total calórico para cada una de las colonias
tratadas fue similar a sus controles, donde no hubo diferencias significativas. En estos dos
modelos experimentales, estos parámetros biométricos fueron analizados particularmente con
el fin de monitorear la aparición de un posible factor de desnutrición asociado al consumo de
etanol, que pudiera interferir con el efecto per se del alcohol en la aparición de efectos
negativos. El tratamiento de etanol 10% en el agua de bebida en ambas colonias, no produjo
variaciones en el peso corporal final de las hembras expuestas respecto de sus controles,
hecho que estuvo en concordancia con la invariabilidad de los valores calóricos totales. Es
sabido que la pérdida o ganancia de peso corporal puede ser responsable de alteraciones de la
homeostasis interna. En particular, la desnutrición está comúnmente asociada a cuadros de

Discusión
247
alcoholismo severo, la que entonces sería en parte responsable de los defectos atribuidos a la
ingesta de alcohol.
Dado que la ingesta de etanol 10% podía interferir en el ciclo estral de las hembras expuestas
y, por lo tanto, afectar el comportamiento reproductivo y, en consecuencia la tasa de
apareamiento, se determinó el efecto de la exposición sobre el ciclo estral. Cuando se
administró diariamente etanol 10% en el agua de bebida a hembras de las dos colonias,
durante 17 días, el ciclo se alteró significativamente en la mayoría de las hembras CF-1
evidenciado esto por la aparición de una mayor proporción de diestros los que a su vez fueron
más prolongados. Finalizado el período de consumo, un 80% de hembras presentaron ciclo
detenido en diestro. Por el contrario, si bien en las hembras CD-1 se observaron algunas
irregularidades en el ciclo, éstas fueron menos marcadas que las halladas en CF-1, no
modificándose ni la cantidad ni la duración de estros ni de diestros a lo largo del tratamiento.
Si bien, las diferencias entre colonias no fueron objeto de análisis del presente estudio, éstas
demuestran una respuesta diferencial dependiente de la colonia murina, sugiriendo la
existencia de respuestas variables del ciclo estral de cada colonia frente a la ingesta de la
droga. Más allá de estas respuestas diferenciales, estos resultados demuestran que el
consumo de alcohol puede interferir en el normal funcionamiento del sistema reproductivo y
del ciclo estral, posiblemente por alteración hormonal, como fue visto por diversos autores
(Mello y col., 1983; Valimaki y col., 1984; Sanchis y col., 1985; Mello, 1988; Canteros y col,
1995). En concordancia con esto, previamente fue demostrado que el tratamiento de ratones
hembras híbridas (F1 (C57xCBA) con etanol 5 y 10 % en el agua de bebida por 30 días previos a
la gestación, produce disfunción reproductiva femenina (Cebral y col., 1997, 1998a, 1998b),
particularmente retraso de la ovulación (Cebral y col., 1998a) y alteración de la calidad
oocitaria expresada como un alto porcentaje de activación partenogenética de los oocitos
ovulados (Cebral y col., 1998b).
El resultado obtenido en cuanto a la variación del ciclo estral frente al consumo de alcohol en
ambas colonias fue fundamental para definir la estrategia reproductiva más conveniente a
seguir en cada una de ellas, con el fin de optimizar las posibilidades de preñez para los estudios
subsiguientes. Así, la ovulación de las hembras CF-1 fue inducida con gonadotrofinas exógenas,
mientras que en las hembras CD-1, la leve alteración del ciclo no justificó la intervención
hormonal para la puesta en apareo y obtención de preñez.
En síntesis, el estudio realizado en el período pregestacional indicó que las dos colonias
murinas poseen similares respuestas biométricas frente al consumo de alcohol, efectos
homologables a los que ocurren en el humano tras el consumo leve o bajo de alcohol; pero

Discusión
248
que la respuesta fisiológica reproductiva de ambas colonias presenta diferencias en cuanto a
su nivel de susceptibilidad frente al consumo de alcohol, y que por ello, el ulterior desarrollo
embrionario, bajo estas condiciones de exposición, también puede responder en forma
diferencial.
2. Efectos el consumo perigestacional de alcohol en la morfogénesis y en el crecimiento
embrio-trofoblástico periimplantatorio murino: días 4, 5 y 8 de gestación.
Estudios previos de nuestro laboratorio han evidenciado que la ingesta de alcohol, desde un
período corto previo a la gestación (15 días) y hasta la etapa de organogénesis temprana (día
10 de preñez) induce, en hembras CF-1, aumento de reabsorciones embrionarias en estadios
tempranos y genera retraso en el grado de diferenciación de los embriones (Coll y col., 2011).
En virtud de esos hallazgos, postulamos que dichas alteraciones, producidas por el consumo
perigestacional de etanol, tendrían origen en días previos al día 10 de gestación, posiblemente
entre los días 7 - 8 (postimplantación temprana) o inclusive antes, entre los días 4 - 5
(preimplantación), por inducción de anomalías tanto en estructuras del propio embrión como
en el linaje trofoblástico. Para dilucidar este interrogante, y habiendo establecido que la
administración de alcohol por 17 días en las colonias murinas utilizadas afecta
diferencialmente al ciclo estral, la administración previa de alcohol continuó durante la
gestación hasta los días 4, 5 y 8 de preñez, momento en los cuales se estudiaron los efectos en
el desarrollo y morfogénesis embrio-trofoblástica temprana.
El primer objetivo fue analizar el desarrollo periimplantativo in-vivo (días 4 y 5 de preñez) en
relación con la diferenciación y crecimiento embrio-trofoblástico.
En el ratón, el día 4 de gestación representa el momento del desarrollo en que predominan, en
la luz uterina, los blastocistos; mientras que el día 5, representa el momento en predominan
los embriones hatched (extruidos de la zona pelúcida) capacitados para iniciar la implantación,
a través del linaje celular extraembrionario ya diferenciado. El proceso implantativo es
dependiente de la formación de blastocistos competentes en la etapa final del desarrollo de
preimplantación (Hardy y col., 1989; Watson, 1992). Para ello, dos eventos morfogenéticos
importantes deben ocurrir de manera coordinada y regulada, la compactación y la cavitación
del embrión (Pratt y col., 1982; Watson, 1992; Motosugi y col., 2005; Sasaki, 2010). Por un
lado, al día 4 de gestación en ambas colonias, el consumo perigestacional de alcohol no tuvo
un impacto negativo en el desempeño reproductivo (número de embriones por hembra), ya
que el número de embriones recuperados no resultó estadísticamente diferente entre las
hembras tratadas y las controles. El elevado número de embriones recuperados de las

Discusión
249
hembras CF-1 respecto de CD-1, estuvo asociado al esquema de estimulación ovárica aplicado
en CF-1; mientras que el número de embriones recuperados de las hembras CD-1, fue el
esperado del apareamiento de hembras murinas con ciclo ovulatorio espontáneo o natural. La
viabilidad embrionaria (porcentaje de fragmentación) no resultó afectada en CD-1 por la
exposición a alcohol, contrariamente a lo esperado y a lo observado en CF-1. En la colonia CF-
1, la presencia de fragmentos celulares en los embriones recuperados pudo también estar
asociada a la manipulación hormonal exógena, mediante la cual se estimula la maduración y
ovulación de gran cantidad de oocitos por ciclo, pero con cierto detrimento en la calidad de la
gameta (Chappel y Howles, 1991). El hecho que los embriones altamente fragmentados tienen
un limitado potencial de desarrollo y raramente consiguen implantar adecuadamente (Plachot
y Mandelbaum, 1990; Erenus y col., 1991; Ebner y col., 2001; Chi y col., 2011), sugirió que el
ulterior desarrollo y la tasa implantativa embrionaria, al menos en CF-1, podrían estar
afectados.
Por otra parte, tanto las hembras CD-1 (Pérez-Tito y col., 2014) como las CF-1 tratadas con
alcohol, tuvieron retraso de la diferenciación de la población embrionaria, manifestado por la
alta proporción de mórulas encontradas y la diminución de blastocistos, según lo esperado al
día 4 de gestación murina, donde como lo marcan los controles, el estadio predominante es el
de blastocisto expandido. Como se mencionó, el proceso de diferenciación a blastocisto, en la
última fase del período de preimplantación, es dependiente de la adecuada compactación y
posterior cavitación de la mórula. Alteraciones en estos procesos, pueden generar anomalías
en la diferenciación de los blastocistos originando, en consecuencia, embriones poco
competentes para iniciar y/o mantener con éxito el proceso de implantación en el tejido
materno (Hardy y col., 1989).
Por otra parte, las hembras tratadas de CD-1 y CF-1 presentaron aumento de la frecuencia de
embriones morfológicamente anormales. De manera recurrente, los blastocistos de las
hembras expuestas presentaron alteración de la cavidad blastocélica, hecho que pone de
manifiesto la posibilidad que el alcohol interfiera en la cavitación, uno de los procesos
morfogenéticos fundamentales en la etapa de preimplantación. La transición de mórula a
blastocisto es dependiente, entre otros factores, del adecuado funcionamiento de la bomba
Na+/K+-ATPasa la que, con gasto de ATP, promueve el transporte de iones y de fluido,
contribuyendo al desarrollo de la cavidad blastocélica. La regulación de la actividad de la
bomba es multifactorial, incluyendo la expresión de las subunidades de la enzima (Watson,
1992) y la concentración intracelular de ATP (Leese, 1991, 1993), entre otros. En tal sentido, es
conocido el requerimiento de fuentes de energía exógena para el desarrollo del blastocisto, ya

Discusión
250
que la formación del blastocele se ha visto asociada a un incremento en el consumo y
metabolización de glucosa (Leese, 1991). Si la exposición perigestacional a etanol generara un
efecto perjudicial sobre el metabolismo de la glucosa, en consecuencia, un efecto negativo en
la actividad de Na+/K+-ATPasa es posible. Así, la reducción de los niveles celulares de ATP
podría conducir a una inhibición de la bomba, y por lo tanto, alterar la acumulación de líquido
en la cavidad naciente (Brison y Leese, 1994). En particular, se ha visto una inhibición de la
actividad de la Na+/K+-ATPasa placentaria, luego de la exposición directa a etanol (Swann,
1990). A su vez, el desarrollo de un epitelio trofoectodérmico (TE) polarizado es fundamental
para el mantenimiento de la localización de la bomba en la membrana basolateral. En este
aspecto, la estabilización del TE mediante uniones intercelulares estrechas, junto con la
presencia de E-cadherina, es crítica para la formación de la cavidad (Watson, 1992; Moriwaki y
col., 2007; Wang y col., 2008), aspecto que se discutirá más adelante. Además de los
mencionados, diversos factores tales como TGF-α, EGF, PARD6B, actúan de manera
coordinada para el proceso de cavitación (Watson, 1992; Alarcon, 2010), donde alteraciones
en los niveles de expresión podrían contribuir en la formación anómala del blastocele.
Investigaciones anteriores de nuestro grupo han demostrado que la administración crónica de
bajos niveles de etanol reduce la síntesis de la prostaglandina E en blastocistos desarrollados
in-vivo (Cebral y col., 1998b). Dado que las prostaglandinas están implicadas en la expansión
de la cavidad y en el hatching de los blastocisto (Kennedy, 1977), la deficiente diferenciación
embrionaria vista en las hembras murinas CD-1 y CF-1 tratadas, incluyendo la anormal
cavitación, podría estar relacionada con niveles alterados de prostanoides, tal como fue visto
anteriormente.
Por otro lado, el alto número de blastocistos fragmentados y con signos de lisis observados en
las hembras tratadas, pudo derivarse de una elevada frecuencia de oocitos activados
partenogenéticamente. Esta hipótesis es posible, ya que previamente nuestro grupo pudo
demostrar que una exposición a etanol 5% y 10 % durante un período prolongado (un mes)
antes de la gestación indujo, en ratones hembras híbridos (C57/B16 x CBA), alta tasa de
activación espontánea tanto en los oocitos ovulados (Cebral y col., 1998a) como luego de la
fecundación (Cebral y col., 1997). En relación con ello, se ha visto que los embriones
partenogenéticos, si bien pueden clivarse en las primeras etapas del desarrollo, generalmente,
cuando alcanzan la etapa de mórula o de blastocisto presentan una morfología alterada,
mostrando pequeños fragmentos citoplasmáticos extruidos en el espacio perivitelino, y no
logran proseguir su desarrollo más allá de esos estadios (Henery y Kaufman, 1992; Takase y
col., 1995). Este arresto embrionario en etapas tempranas del desarrollo preimplantativo,

Discusión
251
luego de la exposición pregestacional a alcohol, también fue demostrado previamente por
nuestro grupo (Cebral y col., 1999, 2000, 2001). Además de un evento partenogenético
periovulatorio inducido por la ingesta de alcohol, y dado que el consumo de la droga continúa
durante la gestación, otra posibilidad es la inducción de anomalías genéticas durante el
desarrollo de los embriones expuestos (Pellestor y col., 1994; Munné y Cohen, 1998) y la
muerte celular programada desencadenada para eliminar blastómeras con aberraciones
cromosómicas o como consecuencias de un entorno poco saludable para el embrión (Chen y
col., 2010; Chi y col., 2011).
Asumiendo que el número de núcleos es igual al número de células, a través del recuento de la
cantidad de núcleos fue posible estimar el crecimiento de los embriones. El número de núcleos
en las mórulas derivadas de hembras controles y tratadas de ambas colonias, no resultó
estadísticamente diferente entre sí. Sin embargo, el reducido número de núcleos en los
blastocistos iniciales derivados de las hembras CD-1 y CF-1 expuestas indicó la disminución del
grado de crecimiento embrionario. Esto podría estar asociado con una menor velocidad del
ciclo mitótico embrionario, evidenciado por la disminución del índice mitótico (IM), sugiriendo
una reducción en la proliferación celular embrionaria entre el 5to. y 6to. ciclo celular. Dado
que el número de células embrionarias es el resultado del balance entre la velocidad de clivaje
(proliferación celular del embrión) y la muerte celular, un reducido número de núcleos en los
blastocistos de las hembras tratadas bien podría ser el resultado del aumento de muerte
celular, manifestado por el incremento de núcleos picnóticos en dichos blastocistos
tempranos. Así, la ingesta de alcohol durante el período preimplantacional, podría inducir un
desequilibrio en la relación proliferación/apoptosis en una alta proporción de la población
embrionaria expuesta, llevando a disminución del grado de crecimiento por pérdida de células.
Esta anomalía en el crecimiento puede ser inclusive la consecuencia de mitosis anormales ya
en las primeras divisiones que, si generaran células con complementos cromosómicos
desbalanceados u otro tipo de daño y los mismos no fueran reparados, podrían derivar en
alteraciones en el ciclo celular (tiempo de clivaje, detenimiento en la mitosis, inducción de
apoptosis) (Musacchio y Hardwick, 2002). Por otro lado, la reducción del número de células
embrionarias aumentará la proporción de blastocisto defectuosos. Blastocistos con un número
de células reducido puede traer aparejado tanto anomalías en la implantación como en el
desarrollo de postimplantación, ya que ha sido reportado que un número crítico de células es
necesario para la formación normal del macizo celular interno (MCI), que permita al embrión
proseguir con su adecuado desarrollo (Gjørret y col., 2003).

Discusión
252
Por otro lado, los blastocistos expandidos de las hembras tratadas CD-1 y CF-1 no tuvieron
reducción del número de células. Sin embargo, en CD-1, los blastocistos expandidos expuestos
presentaron un IM incrementado, y al contrario, los blastocistos expandidos expuestos de las
hembras CF-1 presentaron un IM reducido. Estos resultados, indican que al momento de la
evaluación de los embriones, en el día 4 de gestación, la tasa de división de una proporción de
blastocistos expandidos de las hembras CD-1 expuestas es afectada en forma diferente que la
tasa de división de los blastocistos expandidos de las hembras CF-1. Así, mientras que el
aumento del IM de blastocistos de CD-1 podría implicar el desarrollo de embriones con un
mayor número de células en un estadio más avanzado, comparando con sus controles, en el
caso de los blastocistos de las hembras CF-1 expuestas, el menor IM llevaría a la formación de
embriones con un crecimiento deficiente por menor número de células por embrión. Sin
embargo, y en el caso de los blastocistos de CD-1, una alta tasa de proliferación celular,
indicada por el aumento del IM, podría estar operando en conjunto con una elevada apoptosis,
llevando entonces a la no modificación del número de núcleos en los blastocistos expandido,
resultados que se discutirán posteriormente. En conclusión, estos posibles desequilibrios entre
la proliferación y la apoptosis en los blastocistos expandidos, si bien, en esta fase del desarrollo
no resultan en cambios netos del número de células en cada embrión, sí podrían manifestarse
en etapas más avanzadas del desarrollo, como lo es en la etapa implantativa (al día 5 de
gestación) y, posiblemente, en las subsiguientes fases de postimplantación.
Visto que la ingesta de alcohol hasta el día 4 de gestación afectó negativamente la tasa de
diferenciación, el crecimiento y la morfología de los blastocistos de las hembras CD-1 y CF-1, el
desarrollo embrionario en el período próximo a la implantación, día 5 de gestación, también
podría resultar afectado de forma semejante. A diferencia de lo observado en el día 4, tanto en
las hembras tratadas CD-1 como CF-1, sí se registró una reducción en el número de embriones
recuperados. Dado que ya a la mañana del día 5 de gestación comenzó el primer evento de la
interacción del TE con el epitelio uterino, el proceso de attachment, y aunque por esto, la
recuperación uterina embrionaria fuera más dificultosa, la menor cantidad de embriones
obtenidos en hembras tratadas podría deberse a varias razones. Una de ellas implica la
posibilidad que embriones con anomalías morfológicas y/o con menor grado de diferenciación
al día 4, detuvieran su desarrollo dentro de las 24 hs posteriores, y por consiguiente se
produjera un arresto embrionario implicando la pérdida de los mismos en la cavidad uterina al
día 5 (momento de la recuperación). Otra posibilidad es que, en las hembras tratadas, una
cierta proporción de embriones haya establecido fuertes interacciones con el epitelio uterino

Discusión
253
al momento de la extracción, dando como resultado un proceso implantativo adelantado,
respecto del control, y como consecuencia el total de embriones desarrollados al día 5 no
pudieron ser recuperados. Estas posibilidades serán discutidas posteriormente en el contexto
de los resultados obtenidos al día 8 de gestación.
Así, como fue observado en el día 4 de gestación, también al inicio de la implantación (día 5) se
vio que, dentro de la población embrionaria derivada de las hembras CD-1 tratadas, la
frecuencia de embriones en estadio implantativo más avanzado, clasificados como embriones
tipo 3, fue disminuida, siendo ésta la población embrionaria predominante en las hembras
controles. Asimismo, se encontró un incremento de embriones morfológicamente anormales.
Por otra parte, con el fin de evaluar la población embrionaria en términos de crecimiento, los
embriones de las hembras controles y tratadas fueron agrupados en tres categorías, según el
rango del área de expansión trofoblástica: embriones pequeños, medianos y grandes, y dentro
de cada grupo fue calculada el área promedio por embrión y comparada con su respectivo
control. El tamaño promedio para cada tipo embrionario de las hembras tratadas no cambió
respecto de su control, indicando que el crecimiento embrionario, medido como expansión
trofoblástica, al día 5 de gestación, no fue alterado por el consumo materno de alcohol (Pérez-
Tito y col., 2014).
El siguiente objetivo fue analizar los efectos del consumo perigestacional de alcohol hasta el
día 5 de preñez en las fases de las de la implantación. La hipótesis planteada fue que el
desarrollo trofoblástico durante la implantación resultaría alterado por la exposición in-vivo a
alcohol, tanto por reducción o por aumento del área de expansión, y que en consecuencia una
alta proporción de blastocistos expuestos in-vivo a alcohol, que presentaban retraso de
diferenciación, tampoco proseguirían un desarrollo normal durante la fase de implantación.
Para ello, se utilizó un abordaje in-vitro el cual permitió determinar tanto la diferenciación,
viabilidad y morfología embrionaria, así como, la dinámica del crecimiento o expansión
trofoblástica como indicador de la capacidad implantativa del embrión.
El proceso de implantación ha sido ampliamente estudiado in-vitro. Blastocistos incubados
bajo condiciones adecuadas, son capaces de adherirse y expandirse en placas de Petri
reflejando, en consecuencia, que su capacidad invasiva correspondiente a las primeras etapas
de la implantación en el útero puede ser reproducida en cultivo (Yelian y col., 1995). El sistema
de implantación in-vitro es útil para evaluar los efectos producidos por la exposición in-vivo a
diferentes agentes, sobre procesos tales como la diferenciación celular, expansión y migración
del trofoblasto, procesos que in-vivo resultan ser de dificultoso seguimiento y estudio.

Discusión
254
En animales que desarrollan placentas de tipo hemocorial, como los roedores y primates, las
células trofoblásticas invaden el estroma endometrial hasta alcanzar la sangre materna, a
través de la cual sustentan el desarrollo embrionario. La actividad invasiva del trofoblasto, a su
vez, está garantizada por la habilidad de estas células de adherir y moverse en presencia de
determinadas moléculas de la matriz extracelular (MEC) (Armant y col., 1986; Sutherland y
col., 1988; Damsky y col., 1993; Schultz y Armant, 1995). Particularmente en ratones, las fases
iniciales de la implantación coinciden con la adquisición de relevantes propiedades por parte
del trofoblasto que, a grandes rasgos, incluyen la diferenciación de las células trofoblásticas en
células gigantes poliploides, las que como consecuencia, adquieren un fenotipo típicamente
invasivo asociado a la migración, capacidad fagocítica, síntesis de hormonas y de factores de
regulación (Bevilacqua y Abrahamsohn, 1988; Graham y col., 1993; Hohn y col., 1993).
Considerando estos aspectos relativos a la función trofoblástica durante la implantación, este
objetivo del trabajo fue realizado en el intento de evaluar el comportamiento de las células
trofoblásticas de blastocistos de ratones controles y tratados, cultivados por un período de 72
hs luego de la exposición in-vivo a etanol. Es importante destacar que el desarrollo
prácticamente exclusivo del linaje trofoblástico, y no de las células provenientes del MCI, fue
garantizado a través del adecuado manejo del medio cultivo. Varios estudios han mostrado
que determinadas variables del sistema de cultivo, como el contenido de aminoácidos (Spindle
y Pedersen, 1973), la fuente de suero y su concentración, entre otros (Spindle y Pedersen,
1973; McLaren y Hensleigh, 1975), pueden, aisladamente o en combinación, promover
diferentes grados de desarrollo de las células trofoblásticas o de las células del MCI. Así, por
ejemplo, condiciones específicas de cultivo utilizadas por Koi y colaboradores (1995),
resultaron en una intensa proliferación de las células del MCI, con consecuente diferenciación
de una capa germinativa, que sobrepuesta al cono ectoplacentario, se diferenció formando un
cilindro embrionario. A su vez, Spindle y Pedersen (1973), demostraron que el crecimiento de
las células trofoblásticas y, el crecimiento y diferenciación del MCI en endodermo y
ectodermo, podía ser modulado por la presencia o ausencia de determinados aminoácidos y
diferentes concentraciones de suero. De acuerdo con los autores, la adición de suero al 10% en
ausencia de aminoácidos esenciales en el medio de cultivo permite el crecimiento de las
células trofoblásticas y lleva a escaso desarrollo e inclusive a la pérdida del MCI. Basados en
estos antecedentes, se utilizó medio de cultivo sin aminoácidos esenciales y suplementado con
suero al 10%, además de fuentes de energía para sustentar el desarrollo de los embriones; de
manera de promover sólo el crecimiento de las células trofoblásticas. En este caso, por la
observación de la restricta dimensión de este aglomerado celular central, sumado a la falta de

Discusión
255
evidencias morfológicas sobre la presencia de células embrionarias diferenciadas (ectodermo,
endodermo, mesénquima) suponemos que las células del MCI no prosiguieron el proceso de
diferenciación y se mantuvieron en niveles basales de supervivencia.
Por otra parte, la capacidad de las células trofoblásticas in-vitro de adherirse y diferenciarse en
células gigantes, es atribuida a un comportamiento intrínseco de estas células (Gardner y col.,
1973; Gonçalves y col., 2003). Muchos autores sugieren que el comportamiento de las células
puede ser influenciado por las propiedades físicas de la MEC, la cual, ejerce resistencia física
sobre las células cuando estas extienden procesos citoplasmáticos para expandirse y migrar
por el sustrato (Harris y col., 1980). Sin embargo, basados en las investigaciones de Gonçalves
(1997), donde se sugiere que moléculas de la MEC como fibronectina (FN), laminina (LN) y
ácido hialurónico (AH), no estimulan el crecimiento celular de los blastocistos, y más aún, los
mayores índices encontrados fueron semejantes a valores controles en que ningún
componente de la MEC fue utilizado, sumado a que el área total de expansión de los
blastocistos cultivados sobre LN, FN, AH y sin sustrato fue semejante entre sí, por lo tanto,
optamos por realizar el cultivo de blastocistos hatched sin adición de sustrato.
El aumento del área de expansión trofoblástica es el resultado de variables que actúan en
simultáneo durante el desarrollo embrionario, tales como: el aumento del número de células
(proliferación), el aumento de las dimensiones de las células trofoblásticas a medida que se
diferencian y se tornan gigantes y también, debido a la capacidad migratoria del trofoblasto
sobre el sustrato.
Según la literatura, los blastocistos mantenidos en cultivo se organizan en una estructura
tridimensional, con células que se aplanan y se expanden sobre la superficie de la placa de
cultivo y con células que se aglomeran formando un macizo celular, generalmente central,
sobrepuesto a las células expandidas. Ansell y Snow (1975), McLaren y Hensleigh (1975) y
Sutherland y colaboradores (1988), caracterizaron como células trofoblásticas a las células que
se adhieren y expanden como una monocapa en la superficie de la placa de cultivo y como
células del MCI a las pertenecientes al macizo redondeado. Nuestras observaciones de la
morfología de los blastocistos en cultivo concuerdan con las características descriptas por
dichos autores. Extensas proyecciones citoplasmáticas, propias de la actividad migratoria de
las células trofoblásticas, fueron evidentes en la periferia de la monocapa, configurando un
aspecto de red citoplasmática a través de las cuales se mantienen los contactos celulares entre
sí. Este patrón de organización es semejante al reportado in-vivo, cuando las células
trofoblásticas están en franco proceso de invasión endometrial. Por otra parte, cambios en las
dimensiones celulares y nucleares, durante el cultivo, pusieron en evidencia el proceso de

Discusión
256
diferenciación de las células trofoblásticas más periféricas de la monocapa. Este proceso
también es comparable al que ocurre in-vivo. En ese sentido, in-vivo, el proceso de
diferenciación del trofoblasto comienza casi inmediatamente luego del hatching del
blastocisto y continúa durante la implantación (Potts, 1968; Reinius, 1967). En este proceso,
las células trofoblásticas murales se diferencian en células gigantes del trofoblasto primarias
(CGTs 1rias) (Dickson, 1963, 1966; Barlow y col., 1972; Bevilacqua y Abrahamsohn, 1988, 1989).
Estas células pierden la capacidad proliferativa, entran en un proceso de endorreduplicación
del ADN que, siendo progresivo a lo largo del período de implantación, lleva a la formación de
núcleos de grandes dimensiones y alto grado de poliploidía (Barlow y col., 1972; Gardner y col.,
1973; Ilgren, 1983).
Por lo tanto durante el cultivo in-vitro, dadas estas características el proceso de diferenciación
trofoblástica fue monitoreado morfológicamente, por observación de los cambios en las
dimensiones celulares y nucleares, esperando que a las 72 hs de cultivo se alcance un alto
porcentaje de CGTs en la monocapa embrionaria (Gillies y col., 1986, Gonçalves y col., 2003).
Estas observaciones son las obtenidas en nuestro sistema de cultivo, no encontrándose
diferencias en el grado de diferenciación trofoblástica entre los embriones de las hembras
controles y tratadas. Por otro lado, a través de observación de prolongaciones citoplasmáticas,
lamelipodios y filopodios en las células trofoblásticas periféricas fue monitoreada la
adquisición de un fenotipo invasivo. Estas características también fueron observadas en
embriones cultivados tanto de hembras controles como tratadas.
Si bien, los mencionados aspectos del desarrollo embrio-trofoblástico en cultivo fueron
observados en los embriones de ambos grupos, una elevada frecuencia de embriones de las
hembras expuestas presentaron alteraciones morfológicas, como ser alto grado de
fragmentación y lisis, principalmente en el aglomerado celular de la región central del
embrión, tanto a las 24, 48 como a las 72 hs de cultivo (Pérez-Tito y col., 2014).
En términos generales del desarrollo embrionario en cultivo, se evidenció un retraso en la
diferenciación durante el cultivo de los embriones derivados de las hembras tratadas,
manifestado principalmente en la fase tardía de la implantación (72 hs de cultivo). Allí, las
hembras tratadas presentaron menor proporción de embriones tipo 3 (avanzados) y un
predominio de embriones tipo 2, en comparación con la frecuencia de los controles (Pérez-Tito
y col., 2014). Este resultado indica que el retraso y/o desregulación de la diferenciación
embrionaria ocurrida al día 4 de gestación, luego de la exposición a alcohol, se extiende y
continúa durante la implantación, sugiriendo que las fases tempranas del desarrollo embrio-
trofoblástico postimplantatorio también podrían resultar afectadas.

Discusión
257
Al analizar las posibles alteraciones en la invasividad trofoblástica de las poblaciones
embrionaria en cultivo, a las 48 hs se detectó que los embriones pequeños (tipo 1) y medianos
(tipo 2) provenientes de hembras tratadas tuvieron una mayor tasa de expansión trofoblástica,
respecto de los mismos tipos embrionarios de hembras controles. Esta mayor invasividad,
posiblemente dada por un efecto estimulatorio del alcohol sobre las células trofoblásticas,
podría estar relacionada con alteraciones en tasa de proliferación, diferenciación y/o
migración. Estos resultados sugieren que la exposición perigestacional a alcohol, reduce la
cantidad de embriones en estadio de diferenciación avanzado (tipo 3) pero aumenta el
número de embriones tipo 1 y 2, y en ellos induce una expansión del trofoblasto
incrementando así su capacidad de invasión durante la implantación. Mientras, la reducida
población de embriones grandes (tipo 3) de las hembras tratadas, tuvo un área de expansión
trofoblástica disminuida a las 24 hs, aunque parece recuperarse a las 48 hs cuando alcanzan un
valor de área semejante a los valores controles (Pérez-Tito y col., 2014).
Al respecto, un efecto estimulatorio del etanol en el desarrollo in-vitro fue visto cuando se
incubaron embriones de preimplantación de ratón en presencia de etanol (Leach y col., 1993;
Stachecki y col. 1994a, 1994b). Así, el cultivo de embriones de 1 y 2-células por 24 hs en medio
con etanol 0,1% incrementó el porcentaje de formación de blastocistos, aceleró la mitosis y
estimuló la capacidad de liberación de la zona pelúcida con lo que se sugirió que se induciría
una precoz diferenciación trofoblástica (Leach y col., 1993). Cuando Stachecki y colaboradores
(1994a, 1994b) incubaron en las mismas condiciones (etanol 0,1%) mórulas, la cavitación
resultó acelerada. Inclusive, sólo 5 min de exposición fueron tan efectivos como 24 hs para
alterar la tasa de cavitación, donde el disparo de un pico transciente en la concentración de
calcio intracelular inducido por el etanol fue el mecanismo propuesto. Estos estudios sugieren
que, al menos en el desarrollo trofoblástico del grupo de embriones pequeños, la exposición
in-vivo a etanol hasta el día 5 de gestación, podría tener un efecto estimulatorio acelerando el
crecimiento o expansión del área trofoblástica. Tomando en conjunto los resultados aquí
presentados y los antecedentes de exposición directa a etanol, la respuesta diferencial del
etanol durante el desarrollo temprano que en primera instancia parecería arrojar resultados
contradictorios, está relacionada en parte con la forma en que el embrión de preimplantación
metaboliza el alcohol.
Las enzimas alcohol y aldehído deshidrogenasa (ADH y ALDH, respectivamente) son las
responsables de la metabolización de alcoholes y aldehídos. En el ratón, transcriptos de las
isoformas Clase I, III, y IV de la ADH han sido detectados el comienzo de la gastrulación (Ang y
col., 1996). Sin embargo, muchos de los ARNm examinados fueron detectados sólo en oocitos

Discusión
258
y cigotos, y resultaron ausentes durante la transcripción del genoma embrionario, sugiriendo
su origen materno. En particular, la ausencia de la ADH clase I en los embriones de
preimplantación, sugiere que dichos embriones se encuentran desprovistos de
deshidrogenasas que oxiden el etanol. Por lo tanto, la ausencia de la principal enzima
metabolizadora del etanol durante el desarrollo de preimplantación impediría la formación de
acetaldehído a nivel embrionario durante el consumo materno de alcohol. La baja toxicidad del
etanol observada en cultivos in-vitro de embriones (Leach y col., 1993; Stachecki y col, 1994a,
1994b) podría surgir de la incapacidad del embrión de acumular acetaldehído en él mismo
(Rout y Armant, 2002). En tal sentido, experimentalmente, se observó que el crecimiento de
mórulas y blastocistos se deteriora severamente luego de la exposición in-vitro a
concentraciones de acetaldehído 600 veces más bajas que los niveles tolerables de etanol
(Kalmus y Buckenmaier, 1989), y embriones de 2-células mostraron ser 16 veces más sensibles
a la toxicidad del acetaldehído que a la del etanol (Lau y col., 1991). Ya in-vivo, la situación es
diferente. El acetaldehído generado por la madre durante el consumo sería potencialmente
negativo para el desarrollo de preimplantación. Inicialmente, la ALDH mitocondrial de oocitos
y cigotos pueden oxidar el acetaldehído celular. Sin embargo, una vez que la embriogénesis
comenzó, la expresión de la enzima no se mantiene, representando esto un potencial daño
celular por parte de los aldehídos maternos generados durante el consumo de alcohol. La
ausencia de la enzima ADH clase I puede favorecer la tolerancia del embrión a la exposición a
etanol durante un consumo materno moderado. Sin embargo, la ausencia de una enzima
metabolizadora de acetaldehído materno en las etapas finales del desarrollo de implantación,
puede aumentar los efectos deletéreos de la acumulación sistémica de acetaldehído durante
el consumo de alcohol (Hard y col., 2001) produciendo entonces retraso en el crecimiento y
alteraciones morfológicas en gran parte de los embriones expuestos. De modo que los efectos
del etanol dependen también de la exposición directa (in-vitro) o indirecta (in-vivo), donde el
metabolismo materno participa activamente, ya se potenciando o atenuando los efectos del
etanol sobre el embrión.
En conclusión, los resultados obtenidos muestran que el consumo perigestacional de alcohol
hasta los días 4 y 5 de preñez desreguló la dinámica de la diferenciación y crecimiento
embrionario, además de provocar alteraciones morfológicas en gran parte de la población
expuesta. Estos resultados sugieren la participación de un entorno materno hostil para el
desarrollo embrionario (Chen y col., 2010; Chi y col., 2011), el que contribuirá, en parte, con la
generación de blastocistos poco competentes para iniciar el crucial proceso de implantación.

Discusión
259
Dado que en estudios previos demostramos que la ingesta perigestacional de alcohol hasta el
día 10 de preñez, en hembras CF-1, produjo alta tasa de reabsorción, determinada por arresto
embrionario en los estadios E7 y E8 (Coll y col., 2011), se quiso investigar si estos efectos se
manifestaban al día 8 de gestación momento en que cesa la ingesta de alcohol. Por otra parte,
según los hallazgos en el desarrollo en los días 4 y 5 de preñez, el objetivo fue determinar si, en
estas fases de preimplantación e implantación, el retraso del crecimiento, la reducción del
número de embriones totales en día 5 de gestación, la desregulación de la expansión
trofoblástica de un grupo de embriones de las hembras tratadas, se evidenciaba al día 8 de
gestación como pérdida embrionaria y/o retraso del desarrollo, de forma similar o con mayor
severidad que en los días previos.
El período de gastrulación embrionario es un momento crucial del desarrollo, en el que el
embrión está en proceso de formación de sus capas germinales mientras que el trofoblasto de
la región mesometrial, se encuentra en proceso de invasión y remodelación del tejido
materno. En este estadio gestacional, particularmente susceptible a agentes externos, se
estudiaron los efectos de la exposición materna a alcohol sobre la diferenciación y morfología
embrio-trofoblástica, en ambas colonias murinas CF-1 y CD-1.
En primera instancia, se evaluó el desempeño reproductivo analizando el número de sitios de
implantación por hembra. Se pudo constatar que en las hembras CF-1 (superovuladas), la
ingesta de alcohol indujo una disminución del número de sitios de implantación por hembra,
concordante con la reducción de la cantidad de sitios hallados en las hembras expuestas en el
día 10 de gestación (Coll y col., 2011). Por otro lado, en las hembras CD-1 los resultados
obtenidos refuerzan la idea que la menor tasa de recuperación de embriones al día 5 de
preñez, probablemente se debiera a que una población de embriones se implanta
precozmente y por ello se evidencia un mayor número de sitios de implantación al día 8. Esta
hipótesis no contradice los resultados obtenidos en cuanto a la alta proporción de embriones
con retraso de diferenciación hallada en día 4 y 5, ya que aún los embriones pequeños (día 5) o
con menor crecimiento (tamaño), siendo viables, presentaron mayor área de expansión
trofoblástica que los embriones pequeños de las hembras controles, lo que sugiere que
posiblemente este grupo embrionario expuesto pudo haber comenzado los eventos de la
implantación más rápidamente que sus pares controles, dando así una diferencia en la tasa de
recuperación tanto al día 5 (menor, que el control) como de implantación al día 8 (mayor, que
el control). A diferencia de lo que ocurre en la colonia CD-1, en las hembras CF-1,
probablemente la combinación de la estimulación ovárica con el alcohol, potencia sus efectos

Discusión
260
negativos llevando a la perdida embrionaria, que se manifiesta en un menor número de
embriones (sitios de implantación) en las hembras tratadas al día 8 de gestación.
Posteriormente, en preparados histológicos de sitios de implantación de las hembras CD-1 y
CF-1, se analizó el grado de diferenciación embrionaria luego de la exposición in-vivo a etanol,
evidenciándose, en ambas colonias, nuevamente un retraso del desarrollo. Según los estadios
de diferenciación hallados se pudo determinar que el retraso correspondía a un período
mínimo de 8 hs y máximo de 24 hs, respecto de los embriones derivados de las hembras
controles. Este patrón de diferenciación fue congruente con el observado al día 10 de
gestación, luego de la ingesta de alcohol en CF-1 (Coll y col., 2011). Aparte de estas
alteraciones en la población embrionaria de las hembras tratadas al día 8 de gestación,
quisimos investigar los efectos particulares de la exposición a alcohol sobre la histogénesis de
los embriones expuestos, analizando los potenciales cambios tanto en los tejidos embrionarios
como en la interfase materno-embrionaria, en los tejidos extraembrionarios y en el cono
ectoplacentario. Este estudio sólo fue realizado en los sitios de implantación clasificados en el
estadio E8 (TS10), mientras que los embriones o sitios con retraso no fueron incluidos en la
evaluación.
En relación con los tejidos embrionarios y extraembrionarios, en esta etapa del desarrollo, el
concepto (embrión y membranas extraembrionarias) se encuentra atravesando un activo e
intenso proceso de diferenciación celular, inducciones génicas y progresivos movimientos
morfogenéticos que llevarán a la formación de las tres capas germinales primarias: el
ectodermo, mesodermo y endodermo; y por lo tanto, al establecimiento del plan corporal
embrionario (Lewis y Tam, 2006). De modo que la gastrulación ha sido sugerida como una de
las etapas del desarrollo embrionario vulnerable a la acción de diversos teratógenos, y en
particular del etanol (Webster y col., 1980).
Tanto en los tejidos embrionarios como extraembrionarios, la mayoría de las muestras
derivadas de las hembras expuestas a alcohol, revelaron severas anomalías histológicas, como
ser irregularidad en la organización celular, alteraciones morfológicas según la capa germinal,
fragmentación y picnosis nuclear. Aunque fue claro que la exposición perigestacional a alcohol
hasta el día 8 de gestación, induce profundas consecuencias a nivel celular, no todas las capas
histológicas dentro del concepto fueron igualmente afectadas. Sin embargo, las anomalías
histológicas observadas en las muestras derivadas de hembras expuestas a etanol, fueron
recurrentes. Junto con la desorganización histológica, los cambios morfológicos celulares y la
baja adhesión celular, en algunos casos también se observó presencia de grandes espacios

Discusión
261
intercelulares. Estas anomalías sugieren la inducción de importantes cambios en los procesos
morfogenéticos del desarrollo del embrión durante la gastrulación. Nuestros resultados
concuerdan con estudios de otros autores, quienes demostraron que la exposición a alcohol
durante el período de gastrulación disminuye la tasa de división celular en embriones murinos
(Sulik, 1984) y altera significativamente el proceso de migración de células mesodérmicas hacia
la línea primitiva (Nakatsuji y Johnsson, 1984). Sulik y colaboradores (1984) vieron que las
alteraciones en las células mesodérmicas durante la gastrulación son responsables de
alteración de la diferenciación neuroepitelial, un efecto adverso que podría resultar en la
reducción del tamaño en la placa neural, particularmente de la región anterior del embrión. De
la misma manera, otros autores sugirieron que la exposición sostenida a etanol durante el
período de gastrulación, tendría un impacto negativo en el ectodermo neural y
consecuentemente en el desarrollo del cerebro embrionario (Rubert y col., 2006, Mooney y
Miller, 2007). Similares alteraciones en el tejido ectodérmico y mesodérmico fueron halladas
en nuestro laboratorio, en embriones al día 10 de gestación de hembras CF-1 expuestas a
alcohol en forma perigestacional (Coll y col., 2011).
Por otro lado, el desarrollo embrionario durante la gastrulación es fuertemente dependiente
de la invasión trofoblástica en el lado mesometrial del sitio de implantación. Así, luego de la
implantación, en la región mesometrial, el trofoblasto mantiene su capacidad proliferativa
para formar el cono ectoplacentario (CE) (Bevilacqua y Abrahamsohn, 1988), estructura que
adquiere gran relevancia entre los días 7 y 8 de gestación, y que luego dará lugar a la porción
fetal de la placenta (Simmons y col., 2007). En estos momentos, las células más periféricas del
CE se diferencian en gigantes poliploides (CGTs 2rias), más tardíamente que las de la región
mural y participan en la invasión decidual temprana de la región mesometrial (Snell y Stevens,
1966; Copp, 1979). Así, el CE en las etapas que siguen a la implantación y antes de iniciar el
proceso de placentación presenta células trofoblásticas proliferativas (que posteriormente se
diferenciarán en otros tipos celulares encontrados en la placenta) y CGTs poliploides. Las CGTs
no presentan actividad proliferativa, sino que como consecuencia de la poliploidización,
adquieren numerosas y complejas funciones relacionadas con la invasión, fagocitosis y
producción de hormonas (Zybina y Zibina, 1996). En el estroma uterino, la invasión
trofoblástica prosigue con la ruptura de los vasos endometriales y consecuente
extravasamiento de la sangre materna, que pasa a circular entre las células trofoblásticas
(Welsh y Enders, 1987; Bevilacqua y Abrahamsohn, 1989). Así, las células trofoblásticas que
envuelven al embrión durante toda la gestación se establecerán definitivamente en el estroma
uterino y mantendrán interacción directa con la sangre materna hasta el final de la gestación

Discusión
262
(Bevilacqua y Abrahamsohn, 1989). Es por este significativo papel del trofoblasto, tanto en la
implantación como en la placentación, que en nuestro estudio evaluamos el grado de
desarrollo del CE, sus alteraciones histomorfológicas junto con algunos factores moleculares
que participan en su función y actividad.
Al día 8 de gestación, la mayoría de las hembras CD-1 y CF-1 tratadas presentaron un escaso o
ausente desarrollo de la zona proliferativa del CE, mientras que también la zona de expansión
trofoblástica (región de CGTs en la periferia del CE) se encontró significativamente reducida
junto con una abundante cantidad de glóbulos rojos maternos. Asimismo, en la interfase
trofoblástico-decidual tanto de la cripta antimesometrial como de la interfase lateral del
embrión, se indujeron alteraciones histológicas, evidenciadas por una excesiva picnosis
nuclear, abundante fragmentación celular y elevada infiltración eritrocitaria. Estos resultados
se correlacionaron con otros estudios que asocian alteraciones de la invasión del trofoblasto
con graves complicaciones durante el embarazo temprano como los abortos espontáneos
(Khong y col., 1987), la preeclampsia y restricción del crecimiento intrauterino (Khong y col.,
1986). Así, nuestros resultados sugieren, acorde con otros autores (Jauniaux y col., 2003) que
una anormal invasión trofoblástica y remodelación tisular, ante la exposición a alcohol,
podrían causar no sólo disminución del flujo sanguíneo materno al embrión temprano, sino
que también constituirse en el origen de una placentación defectuosa en estadios
gestacionales posteriores.
Consecutivamente a los hallazgos mencionados, se quiso investigar la viabilidad, la capacidad
de desarrollo e invasión del CE mediante estudios in-vitro. La expansión del trofoblasto al día 8
de gestación es clave para la adquisición de una mayor superficie de diálogo e intercambio
entre el organismo materno y el embrión (Stroband y Van der Lende, 1990). El crecimiento o
expansión de las células trofoblásticas del CE en cultivo, fue utilizada como indicador de su
capacidad invasiva.
Además de una reducida viabilidad, los conos provenientes de las hembras tratadas
presentaron una menor capacidad de desarrollo entre las 24 y las 48 hs de cultivo.
Así como fue observado en cuanto al crecimiento embrio-trofoblástico al día 5 de gestación y
durante las fases de implantación (in-vitro), debido a la variabilidad de tamaños, los CE
cultivados se clasificaron en conos pequeños, medianos y grandes. Si bien, la distribución de los
tres tipos de conos de las hembras tratadas no fue diferente de la de las hembras controles,
durante los tiempos de cultivo; la dinámica de expansión del trofoblasto se vio alterada
respecto del control. Particularmente, en el caso de los conos pequeños, el grado de expansión
a las 24 hs fue reducido para luego incrementarse a las 48 hs; mientras que en el desarrollo de

Discusión
263
los conos medianos, se vio el patrón opuesto registrándose un incremento de su expansión a
las 24 hs para luego producirse una reducción a las 48 hs. Sin embargo, los conos grandes
presentaron una dinámica de expansión similar a la observada en los controles. Del mismo
modo a lo observado al día 5 de gestación y hacia el final del cultivo de los embriones
implantativos, nuevamente y en forma similar se indujo alteración de la dinámica de expansión
de conos pequeños y medianos, luego de la exposición a alcohol. Estos resultados sugieren que
la invasividad de conos con escaso desarrollo puede estar desregulada, con lo cual, tanto la
placentación como el desarrollo de esa población de embriones podrá estar afectada.
El área de expansión de los CE in-vitro resulta del balance de la proliferación, muerte celular,
diferenciación y migración. Al momento en el que el CE es aislado para ser colocado en cultivo,
las células trofoblásticas son predominantemente diploides (~90%), proporción que sin
embargo, disminuye partir del momento en que estas células son colocadas en cultivo (Ilgren,
1981; Rossant y Tamura-Lis, 1981), por el comienzo del proceso de endorreduplicación del
ADN, imitando el comportamiento de las células trofoblásticas in-vivo. Luego de 48 hs de
cultivo, la frecuencia de células diploides disminuye a medida que continúa, en forma lenta, el
aumento del proceso de poliploidización. Gonçalves y colaboradores (2003), correlacionaron
los valores de ploidía con valores de áreas nucleares, corroborando que los núcleos mayores
mostraban efectivamente mayor contenido de ADN, estableciendo así una forma sencilla, a
través del tamaño nuclear, de identificación de poblaciones celulares diferenciadas o no.
Complementariamente, la localización diferencial en el cono en cultivo de un tipo celular y
otro permite también su clara distinción: central, en el caso de las células diploides; y
periférica, en el caso de las células poliploides.
En condiciones de cultivo, Ilgren (1981) encontró el valor máximo de ploidía para las células
gigantes luego de 72 hs de cultivo. Hathaway y Babiarz (1988), por su parte, verificaron el
gradual aumento de ploidía en las células trofoblásticas entre las 48 y 96 hs de cultivo,
mientras que Gonçalves y colaboradores (2003) encontraron los niveles máximos a las 72 y 96
hs de cultivo. Para poner en evidencia la ploidía de las células trofoblásticas del CE, el índice de
incorporación de BrdU también puede ser utilizado. Los CE en cultivo presentan una tasa de
proliferación trofoblástica relevante principalmente en las primeras 24 hs de cultivo, luego de
lo cual, este proceso se detiene gradualmente (Gonçalves y col., 2003) sugiriendo que, aunque
las células centrales del cono sean potencialmente proliferativas, esta actividad se pierde por
su aislamiento de los demás tejidos embrionarios, donde por ausencia de señales
embrionarias se bloque la mitosis trofoblástica y se induce la diferenciación a CGTs (Gardner,
1972; Rossant y Offer, 1977; Ilgren, 1981; Rossant y Tamura-Lis, 1981; Rossant, 1986; Soares y

Discusión
264
col., 1993; Hamlin y Soares, 1995). En este contexto, es posible que el crecimiento celular
verificado en las primeras 24 hs de cultivo del CE, refleje todavía resquicios de una mutua
interacción embrio-trofoblástica. Así, durante el cultivo de las células trofoblásticas, hay una
desactivación de la proliferación celular y un proceso de diferenciación en CGTs,
contrariamente a lo que ocurre in-vivo, donde el cono mantiene los dos programas celulares
activos: proliferación y endorreduplicación. Estas dos actividades conjuntas le confieren al
trofoblasto la capacidad de expandir la población de células trofoblásticas precursoras para
posterior diferenciación en los diferentes tipos trofoblásticos de la placenta, al mismo tiempo
que la endorreduplicación prosigue para la formación de las CGTs.
Basados en estos antecedentes, en el presente trabajo se utilizó la marcación con BrdU para
verificar la síntesis de ADN, concordante con células metabólicamente activas y en proceso de
diferenciación. Dado que, como hemos visto, el alcohol interfiere con los programas de
diferenciación celular, nos preguntamos si podía estar alterado este proceso en las células
trofoblásticas en cultivo. El estudio se realizó a las 48 hs dado que, por lo anteriormente
expuesto, el proceso de diferenciación es preponderante a partir de ese momento. En
referencia a ello, no encontramos diferencias en el patrón de marcación ni en el número de
células BrdU-positivas en el área de expansión trofoblástica (zona de CGTs) entre los CE de las
hembras tratadas y controles. A partir de este hallazgo inferimos que el programa de
endorreduplicación del ADN, y por lo tanto, la diferenciación trofoblástica, no fueron afectados
por el consumo materno de etanol. Por lo tanto, las alteraciones en el área de expansión de los
CE derivados de hembras tratadas estarían relacionadas con otros eventos. Dado que la
morfología de los CE en cultivo, no fue modificada en las hembras tratadas, la que fue
semejante a lo reportado (Zybina y Zybina, 1996), la exposición a alcohol no afectó la
formación del patrón morfológico invasivo o migratorio de las células trofoblásticas.
Probablemente, dadas las alteraciones vistas en el ciclo celular de los embriones de etapas
más tempranas, la desregulación en el balance proliferación/muerte celular podría ser, en
parte, responsable de las alteraciones de la dinámica de crecimiento trofoblástico en la fase de
gastrulación.
3. Efectos del consumo perigestacional de alcohol sobre las uniones intercelulares,
metaloproteasas y genotoxicidad en el tejido embrio-trofoblástico periimplantatorio murino.
El desarrollo embrionario es dinámico y bien orquestado. Las células proliferan, se diferencian,
migran y mueren, y su toma de decisiones está en parte influenciada por su microambiente.

Discusión
265
En virtud de las alteraciones generadas por la ingesta perigestacional de alcohol en hembras
CD-1 y CF-1, hasta fase de preimplantación (días 4 y 5 de gestación) y hasta la de
postimplantación temprana (día 8), en relación con la diferenciación, el crecimiento, la
morfología embrionaria y los procesos morfogenéticos de los tejidos embrio-trofoblásticos,
postulados que estas anomalías podrían ser la resultante, al menos en parte, de la acción
adversa de la exposición a alcohol sobre moléculas claves para el desarrollo embrionario e
invasión trofoblástica en la fase de periiimplantación: las moléculas de unión intercelular y las
metaloproteasas (MMPs), MMP-2 y MMP-9. Por otra parte, pensamos que los efectos
negativos del alcohol podrían involucrar la generación de genotoxicidad embrionaria y
desbalance en los niveles de muerte celular en el tejido embrio-trofoblástico. En este contexto,
tampoco se descartó un posible efecto genotóxico del etanol a nivel materno en el período
temprano de la gestación murina.
- Efectos del consumo perigestacional de alcohol en moléculas de adhesión celular
embrionarias en el período periimplantatorio: días 4 y 8 de gestación.
Considerando que las uniones célula-célula están involucradas en la morfogénesis embrionaria
y en el establecimiento y mantenimiento de la arquitectura de los tejidos, la expresión
anómala de las proteínas que integran dichas uniones podría ser causa de anomalías
morfológicas del embrión temprano, inducidas por el consumo materno de alcohol.
Basados en nuestros resultados, en los cuales, las hembras de ambas colonias, CD-1 y CF-1,
que recibieron alcohol hasta el día 4 de gestación presentaron una alta proporción de
blastocistos con anomalías en la cavitación analizamos, por inmunofluorescencia, el patrón de
expresión de la proteína ZO-1 de las uniones estrechas (UE) y de la E-cadherina de las uniones
adherentes (UA), tanto en blastocistos iniciales como expandidos en el modelo murino CF-1.
El desarrollo normal del embrión de preimplantación es crítico para la embriogénesis de las
etapas posteriores, ya que defectos en la formación del blastocisto podrán dar lugar a pérdida
embrionaria temprana. Las moléculas de las uniones intercelulares, como las adherentes y las
estrechas, juegan un rol fundamental en la morfogénesis y diferenciación de los tejidos y del
embrión (Tunggal y col., 2005), siendo la regulación de sus niveles de expresión crucial para la
integridad de las uniones célula-célula y la diferenciación y remodelación celular.
La formación del blastocisto requiere la expresión de diversos productos génicos incluidos el
de la Na+/K+-ATPasa, la E-cadherina, y los componentes de las UE (Violette y col., 2006). En
particular, para que la cavitación ocurra, el embrión requiere el desarrollo de un epitelio

Discusión
266
polarizado y de un mecanismo para generar y/o transportar fluido a la cavidad naciente
(Watson, 1992). Este proceso se inicia con la polarización progresiva de las blastómeras
externas del embrión de 8 células durante la fase de compactación de la mórula (Johnson y
McConnell, 2004) donde las UA, mediadas por complejos cadherina-catenina, se establecen
como preludio a la formación de las UE (Fleming y col., 2001; Johnson y McConnell, 2004). El
establecimiento progresivo y secuencial de las UA y UE en la superficie basolateral y
apicolateral de las células, respectivamente (Ziomek y Johnson, 1980; Fleming y col., 2001)
contribuye a la formación del primer tipo celular diferenciado que surge durante la
embriogénesis, el TE. Así, la diferenciación del TE ocurre a medida que las UE se forman,
generando dos dominios (uno apical y otro basolateral). La formación y estabilidad de las UE
en el TE influencia la aparición y mantenimiento de la distribución polarizada de la bomba
Na+/K+-ATPasa, ya desde el estadio de mórula tardía (Fleming y col., 1989; Watson y col.,
1990b). Sin embargo, la formación adecuada del blastocisto sólo ocurre cuando se han
establecido UE funcionales y, en consecuencia, se desarrolla la barrera de permeabilidad que
restringe la fuga de líquido vía paracelular, asegurando la expansión de la cavidad (Biggers y
col., 1988; Sheth y col., 2000). El complejo de UE es, sin duda, uno de los componentes
celulares más importantes que participan en la cavitación (Watson, 1992). Alteraciones en la
regulación espacio-temporal de las moléculas de las uniones intercelulares en la embriogénesis
temprana son causas potenciales de anomalías embrionarias posteriores.
Las UE apicolaterales son complejos multiproteicos, donde ocludina y claudina son proteínas
integrales de membrana y los miembros de la familia ZO (ZO-1, ZO-2 y ZO-3) forman un
andamio submembranoso (Tsukita y col., 2001). La proteína ZO-1 se expresa por primera vez
durante la fase de compactación, en embriones de 8 células, por lo que se sugirió su
participación en la transición de mórula a blastocisto (Sheth y col., 1997, 2000). Wang y
colaboradores (2008) demostraron que la inactivación de ZO-1 con siRNA en embriones de
ratón, efectivamente, inhibía el pasaje de mórula a blastocisto, por lo que se pensó que ZO-1
sería necesaria para mantener la integridad de la capa de TE, esencial para la formación del
compartimento blastocélico (Wang y col., 2008). Además, en ausencia de ZO-1, las UA no
proporcionan suficiente cohesión a las células externas de la mórula para iniciar la
acumulación de líquido en el interior del embrión. Por otro lado, se vio que la inactivación de
ZO-1 reduce la expresión CDX2 involucrado en la diferenciación de los blastómeros polares
para formar TE (Niwa y col., 2005). Dado el importante papel de la ZO-1 en el proceso de
diferenciación de mórula a blastocisto, la inhibición o alteración de su patrón de expresión

Discusión
267
afectará notablemente el desarrollo del embrión. En el presente trabajo, tanto en los
blastocistos iniciales como expandidos de ambos grupos experimentales, se detectó ZO-1 en el
trofoectodermo polar (TP) y mural (TM). En los blastocistos de hembras controles, la ZO-1 se
localizó en los contactos celulares, en forma submembranosa, continua y definida. Sin
embargo, en los embriones de las hembras tratadas, si bien, la ZO-1 se localizó en los
contactos celulares del TE, la marca se presentó discontinua, delgada y débil, sugiriendo
debilitamiento de las uniones. Por otra parte, otros grupos de embriones tuvieron marca de
ZO-1 deslocalizada, engrosada, granular e intensa, que formó acúmulos citoplasmáticos
perinuclear. En conjunto, estos resultados y según los estudios previos sobre el papel
fundamental de la ZO-1 en el proceso de diferenciación de mórula a blastocisto, la interrupción
o alteración de la UE mediada por ZO-1 en la mayoría de los embriones de las hembras
alcohólicas, podría llevar a la imposibilidad de diferenciación de la mórula compactada, por lo
que en consecuencia se evidencia un retraso de crecimiento o desarrollo de blastocistos. Por
otra parte, algunas investigaciones demostraron que la distribución anómala, deslocalizada y
discontinua de la proteína ZO-1 en el TE, es indicador de rearreglos estructurales anormales de
las proteínas del complejo de la UE, alteraciones que pueden conducir a anomalías funcionales
de la bomba Na+/K+-ATPasa y consiguiente aumento de la permeabilidad intercelular e
inhibición del pasaje de mórula a blastocisto (Violette y col., 2006; Madan y col., 2007).
Mientras que el potencial de desarrollo de los embriones parece correlacionar con la
expresión/funcionalidad de ZO-1, embriones de preimplantación desprovistos de ZO-3 se
desarrollan normalmente hasta el estadio de blastocisto e inclusive evolucionan en ratones
fértiles (Adachi y col., 2006). Por otro lado, se ha demostrado que en embriones deficientes en
ZO-2, aunque retrasada, ocurrió cavitación. Llamativamente, en los embriones desprovistos de
ZO-2, se observó un mayor ensamblaje de ZO-1, posiblemente, como un mecanismo de
compensación a la ausencia de ZO-2, mientras que el doble knockdown de ZO-2 y ZO-1, generó
una inhibición severa de la formación de blastocele, indicando que la deficiencia de ZO-1, más
que la ausencia de ZO-2 sería la responsable de un fenotipo alterado más severo (Sheth y col.,
2008). Tomando en conjunto estos antecedentes y nuestros resultados, donde se ve en parte
un mayor ensamblaje de ZO-1, retraso en la diferenciación y alteración en la cavitación, podría
ser que la expresión de ZO-2 esté alterada por el consumo materno de alcohol.
La construcción de la capa epitelial trofoectodérmica con sus típicas UE requiere de la
formación previa de las UA basadas en E-cadherinas, siendo estas dos redes de unión las que
en conjunto regula la fuerza de adhesión entre las células para la formación del epitelio

Discusión
268
(Gumbiner y col., 1988; Gumbiner, 2000). Las cadherinas, componentes integrales de la
membrana de las UA, fueron originalmente identificadas en mórulas de ratón en proceso de
compactación (Yoshida y Takeichi, 1982), pero a diferencia de la ZO-1, que se expresa a partir
del estadio de 8 células, la proteína E-cadherina está presente en todos los estadios del
desarrollo embrionario de preimplantación (Wang y col., 2008). Ella provee la fuerza de
adhesión célula-célula en la compactación y es absolutamente necesaria para la diferenciación
del TE (Larue y col., 1994, 1996; Peinado y col., 2004; Eckert y col., 2005) en la transición de
mórula compactada a blastocisto, asegurando el correcto establecimiento de las uniones en el
TE (Larue y col., 1994, 1996; Kan y col., 2007). Esto ha sido demostrado mediante la inhibición
de su función. Así, los embriones knockout para E-cadherina no logran desarrollarse mas allá
de la fase de mórula, aunque se compacten (Larue y col., 1994; Ohsugi y col., 1997 ). También
Watson y colaboradores (1990b), demostraron que el tratamiento de embriones de 8 células
con anticuerpos anti-E-cadherina previno la formación de complejos de UE en el TE con la
consecuente imposibilidad de mantener la distribución polarizada de la Na+/K+-ATPasa, lo cual
alteró el comienzo y progresión de la cavitación. En nuestras observaciones un alto número de
blastocistos de las hembras tratadas poseían anomalías blastocélica, vistas como particiones
de la cavidad, varias cavidades o cavidades pequeñas. Estas anomalías, bien pudieron
relacionarse con las alteraciones de la expresión de E-cadherina, como fue sugerido en otros
estudios en que se vieron embriones con defectos en la expresión de esta molécula, con un
número variable de cavidades llenas de fluido (vesículas) (Manejwala y col., 1989; Watson y
col., 1990b), con deslocalización de la bomba Na+/K+-ATPasa en pequeñas vesículas. En el
presente trabajo, la E-cadherina se localizó en las superficies de contacto del TP y TM, tanto en
blastocistos iniciales como expandidos en ambos grupos experimentales. Pero, mientras los
blastocistos de las hembras controles presentaron un patrón de marcación continuo y definido
en las superficies blastoméricas, los embriones derivados de las hembras tratadas tuvieron
marcación engrosada y discontinua con acumulaciones membranosas.
La intensidad relativa de marca tanto para ZO-1 como para E-cadherina fue alta en los
embriones de hembras tratadas, sin embargo, la deslocalización de la marca en ambos casos
fue evidente, por lo tanto no descartamos la posibilidad que una falla en el ensamblaje de las
uniones pueda ser un disparador para sobreexpresar las moléculas involucradas en la unión,
como fue observado en investigaciones previas en embriones en estadio organogénicos donde
hallamos que la expresión proteica de N- y E-cadherina se encuentra incrementada en
embriones al día 10 de gestación luego del consumo murino periconcepcional de alcohol (Coll
y col., 2011). De modo que no descartamos la posibilidad que la exposición in-vivo a alcohol

Discusión
269
induzca una sobreexpresión de ZO-1 y E-cadherina como un mecanismo compensatorio a la
disrupción o debilitamiento de estas uniones intercelulares, como consecuencia de un
ensamblaje anómalo, en los embriones de preimplantación.
La importancia de los estudios de la expresión de ZO-1 y E-cadherina en embriones, radica en
la participación que tienen estas moléculas en el desarrollo de preimplantación. Fleming y
colaboradores (1989), demostraron la distribución membranosa al azar de la molécula ZO-1
luego del tratamiento con anticuerpos anti-E-cadherina, lo que altera la formación del epitelio
trofoectodérmico. Violette y colaboradores (2006) sugirieron que, por un lado la E-cadherina
es esencial para los eventos iniciales de la diferenciación, ya que dispara la translocación de
ZO-1 a la membrana plasmática. Por otro lado, basados en la inhibición experimental de la
bomba Na+/K+-ATPasa con ouabaina, también ésta sería crucial en la regulación de la
formación adecuada de la ZO-1 en la membrana, y en consecuencia, en la completa
diferenciación del TE. En nuestro estudio, no sólo la deslocalización de la marca para ZO-1 y E-
cadherina fue evidente, sino que también es posible que una falla en el ensamblaje de las
uniones, dada por la alteración en el patrón de expresión de ambas moléculas, pueda disparar
la sobreexpresión de estas moléculas, como fue mencionado. De modo que no descartamos la
posibilidad que la exposición in-vivo a alcohol induzca un mecanismo compensatorio en la
expresión de la ZO-1 y de E-cadherina ante un debilitamiento de estas uniones intercelulares
como consecuencia de un ensamblaje anómalo.
Los resultados de nuestro trabajo sugieren que la exposición perigestacional a alcohol induce
alteraciones en el programa de biogénesis de las UE y UA, aún desde el establecimiento de la
mórula. Este hecho es posible dado que mientras la colocalización de la ZO-1 y la E-cadherina,
estudiada por análisis de microscopía confocal, en todos los blastocistos de las hembras
controles, fue altamente positiva, sólo el 50% de los embriones de las hembras tratadas
presentó colocalización positiva, siendo la señal de la misma de carácter débil. Fue visto que la
diferenciación del trofoectodermo en el blastocisto, requiere en primer lugar de la formación
de UA (E-cadherina) para construir y establecer las UE (Gumbiner y col., 1988 ), por lo que las
primeras etapas en la biogénesis de las UE dependen de la expresión de E-cadherina. Esta, si es
alterada, provoca el montaje al azar de las UE en la membrana, alterando así la formación del
epitelio trofoectodérmico, mientras que la interrupción de la formación de la UE conduce al
detenimiento del desarrollo embrionario en la etapa de mórula. Los resultados del presente
trabajo sugieren que la exposición perigestacional a alcohol induce alteraciones en el
programa de biogénesis de las UE y UA en los blastocistos, donde se pierde la co-localización

Discusión
270
de la E-cadherina con la ZO-1 en la superficie de contactos trofoectodémicos, llevando a una
inadecuada diferenciación del blastocisto que se evidencia en menor proporción de
blastocistos expandidos, con anomalías morfológicas de la cavitación (Pérez-Tito y col., 2013).
Basados en la elevada desorganización histológica observada en los embriones provenientes
de hembras tratadas, al día 8 de gestación, se llevó a cabo el estudio de la inmunoexpresión de
E-cadherina particularmente en el ectodermo embrionario.
Los complejos de UA también juegan un papel crítico en una variedad de eventos de la
embriogénesis, no sólo en los iniciales (compactación y diferenciación del TE), sino también en
estadios más avanzados, donde predominan procesos de diferenciación celular, crecimiento y
proliferación, como ocurre en la gastrulación, neurulación, cardiogénesis y somitogénesis
(Peinado y col., 2004). Si bien, ha sido demostrado que la morfogénesis de los tejidos es
dependiente de la presencia de las cadherinas y que éstas han sido originalmente asociadas a
la adhesión intercelular (Yoshida y Takeichi, 1982), hoy en día es claro que el papel de las
cadherinas no sólo se limita a la unión mecánica entre las células. Las funciones de las
cadherinas abarcan múltiples aspectos en la génesis de los tejidos incluyendo la señalización
intracelular, la modulación de la proliferación, el reconocimiento celular y, la determinación y
el mantenimiento de la diferenciación, la coordinación del movimiento celular, así como la
inducción y mantenimiento de la arquitectura de tejidos epiteliales (Yoshida y Takeichi, 1982;
Takeichi, 1991; Halbleib y Nelson, 2006). Durante la gastrulación, cambios en la expresión de la
E-cadherina están involucrados en la transición epitelio-mesenquimática y la adquisición de un
fenotipo apto para la migración (Savagner, 2001). La pérdida de E-cadherina es una fuerza
motriz para la transformación de las células epiteliales en mesenquimáticas durante la
formación de mesodermo. Más tardíamente, durante la neurulación, la E-cadherina es
necesaria para la migración y los reordenamiento de los tejidos (Lien y col., 2006). Cambios de
expresión de E-cadherina a N-cadherina ocurren en la placa neural invaginada y son necesarios
para la formación de la placa neural (Hatta y Takeichi, 1986). Sin embargo, si bien el
mantenimiento de la integridad de los tejidos es una de las funciones críticas de las proteínas
involucradas en las UA, sus componentes también participan como mediadores en la
señalización intracelular, controlando aspectos como la proliferación y expresión génica. Así,
muchas anomalías en mutantes para caherinas o cateninas resultan no por defectos en la
adhesión, sino por defectos en la señalización interna, corroborando el papel de estas
proteínas en vías críticas del desarrollo del embrión. Mediante su participación en estas vías,
las proteínas de las UA pueden transmitir información, la cual es interpretada de manera

Discusión
271
célula-específica dependiendo del contexto intracelular, siendo las respuestas celulares de
muerte, de diferenciación o proliferación, entre otras (Lien y col., 2006). Embriones mutantes
para β-catenina, los cuales fallan en la formación de la línea primitiva y en la producción de
mesodermo y endodermo (Haegel y col., 1995), presentan defectos que no se asociaron a una
pérdida en la adhesión celular, la cual se mantuvo por la participación de otras moléculas, sino
que resultó por una falla en la vía de señalización (Fang y col., 2004). Asimismo, defectos
craneofaciales también pueden surgir como consecuencia de alteraciones en la señalización
intercelular, siendo la actividad señalizadora de β-catenina y p120-catenina requerida para la
migración y especificación del destino celular de las células de la cresta neural (Ciesiolka y col.,
2004).
En nuestros estudios, mientras que el patrón de marcación de la E-cadherina en los embriones
de las controles, fue continuo y lineal entre los contactos intercelulares de las células
ectodérmicas, la expresión de esta molécula en los embriones de hembras tratadas resultó
deficiente (Pérez-Tito y col., 2013), sugiriendo que, en parte, las alteraciones histológicas
observadas en estos tejidos, podrían vincularse con la pérdida de adhesión celular por
disrupción de la expresión de E-cadherina y/o con disrupciones en las vías de señalización en
las que dichas proteínas participan.
Cambios en el patrón espacio-temporal de la expresión de E-cadherina durante la
embriogénesis tiene un marcado impacto negativo en la mecánica de adhesión de las células
(Wheelock y col., 2008) y en la señalización interna. Las anomalías de las uniones celulares
constituyen importantes mecanismos inductores de defectos morfológicos del embrión en
desarrollo, como fue recientemente demostrado con la aparición de anomalías de cierre del
tubo neural, las alteraciones mesodérmicas y los cambios en la expresión de cadherinas de
embriones organogénicos luego de la exposición a alcohol (Coll y col., 2011). En este caso, al
día 10 de gestación, la expresión de E-cadherina embrionaria resultó incrementada,
comparando con los controles, como fue observado en gran parte de los blastocistos
expuestos. Sin embargo, el patrón de expresión de E-cadherina opuesto al día 8 de gestación,
no descarta la posibilidad que el efecto nocivo del alcohol sobre estas moléculas de unión sea
dependiente y especifico del tejido y del momento del desarrollo, ya que posiblemente
diferentes mecanismos compensatorios puedan ser disparados según los requerimientos
moleculares embrionarios para generar o reparar uniones de adhesión que estén afectadas.

Discusión
272
- Efectos del consumo perigestacional de alcohol en la expresión proteica y génica de MMP-2
y MMP-9 en embriones en el período postimplantatorio (día 8 de gestación).
Tanto el escaso desarrollo del cono ectoplacentario, como las alteraciones morfológicas y las
anomalías en su invasividad, observadas in-vitro al día 5 y 8 de gestación, luego de la ingesta
materna de alcohol, llevaron a hipotetizar que cambios en los niveles de MMPs en el tejido
embrio-trofoblástico afectarían la regulación del desarrollo y función trofoblástica temprana.
Las MMPs están involucradas tanto en procesos fisiológicos normales como patológicos. Estas
proteasas no son meras enzimas de degradación de componentes de la MEC, sino que
orquestan diversas vías de señalización a través del procesamiento proteolítico de receptores y
factores anclados en la MEC, desenmascarando y/o liberándolos de ella, para regular su
biodisponibilidad. Así, las MMPs están involucradas en el control de la división, proliferación
celular y homeostasis de los tejidos (Rodríguez y col. 2010). Participan en el control de la
apoptosis, induciendo o previniéndola según el procesamiento proteolítico de determinadas
moléculas (Tanaka y col., 1998). Así, las MMPs, a través de la interrupción de las uniones
célula-célula por escisión de cadherinas y otras moléculas de membrana, pueden inducir
procesos de apoptosis (Ilan y col., 2001). También están involucradas en procesos
angiogénicos y de neovascularización, regulando la biodisponibilidad de factores angiogénicos
secuestrados en la MEC, como es el caso de la liberación del factor de crecimiento del
endotelio vascular (VEGF) por la MMP-9 (Bergers y col., 2000; Lee y col., 2005), o la
degradación de la MEC y activación de factores por la MMP-2 (Dean y col., 2007).
Tradicionalmente, las MMPs han sido consideradas las principales facilitadores de la migración
celular a través de la ruptura de las barreras de la MEC, facilitando el avance celular y la
invasión de las células por proteólisis de componentes barrera de la MEC, de mediadores
extracelulares de adhesión célula-MEC o célula-célula y por escisión de receptores (Koshikawa
y col., 2000). La proteína transmembrana E-cadherina de las UA parece ser uno de los blancos
principales de las MMPs para la interrupción de los contactos célula-célula y facilitar la
motilidad de las células (Lochter y col, 1997; Noe y col., 2001). Las MMPs participan en la
implantación, decidualización, morfogénesis de los tejidos embrionarios/fetales y
placentación (Chen y col., 2007; Rodríguez y col., 2010) a través de la interacción con sus
inhibidores que se expresan en las células deciduales y trofoblásticas de la interfase materno-
embrionaria (Anacker y col., 2011). Específicamente la MMP-2 y, principalmente la MMP-9,
están implicadas en el comportamiento invasivo del trofoblasto (Alexander y col., 1996;
Sharkey y col., 1996; Peters y col., 1999; Chen y col., 2007).

Discusión
273
En base a estas importantes funciones, nos centramos en el estudio del patrón de expresión
proteica de MMP-2 y MMP-9 tanto en el CE, en la interfase trofoblástico-decidual como en los
tejidos embrionarios y extraembrionarios, luego de la exposición a alcohol hasta el día 8 de
gestación, siendo que en este momento de la gestación murina se produce la máxima
invasividad trofoblástica en el tejido materno.
Varios reportes describieron la expresión de dichas MMPs ya desde etapas tempranas del
desarrollo. Mensajeros y actividad de MMP-9 fue detectada en blastocistos implantativos de
ratón a las 24 hs de expansión in-vitro, e in-vivo en blastocistos implantativos y sitios de
implantación al día 5,5 (Sharkey y col., 1996; Whiteside y col., 2001; Kim y col., 2002; Chen y
col., 2007; Plaks y col., 2013), coincidiendo con el momento de expansión de las células
trofoblásticas. Wang y colaboradores (2003) demostraron expresión de ARNm de MMP-2 en
todos los estadios embrionarios de preimplantación. Sin embargo, Sharkey y colaboradores
(1996) mostraron expresión génica de MMP-2 sólo en blastocistos en expansión después de las
24 hs de cultivo, siendo su expresión menor respecto de los niveles de MMP-9, mientras que a
las 72 hs de cultivo, Chen y colaboradores (2007), detectaron ARNm de MMP-2 pero no
actividad. De modo que aún no está claro el perfil de expresión de MMP-2 en las etapas
iniciales del desarrollo. Al día 8 de gestación en el ratón, Bany y colaboradores (2000),
demostraron por RT-PCR la expresión de MMP-2 y MMP-9 tanto en embrión, CE y decidua. En
particular, diversos autores describieron un pico de expresión génica, proteica y actividad de la
MMP-9 en las CGTs entre E7,5 y E8,5, momento en que dichas células degradan la MEC, se
expanden e invaden la decidua (Soares y col., 1996; Peters y col., 1999). En roedores, la
adquisición de un fenotipo trofoblástico endocrino e invasivo es dependiente de la
diferenciación. Así, sólo aquellas células que se diferenciaron a CGT mostraron un fenotipo
migratorio asociado a la activación del promotor de MMP-9, mayor expresión génica y proteica
y mayor actividad proteolítica (Peters y col., 1999). Nuestros resultados concuerdan con estos
antecedentes. En el CE de las hembras controles y tratadas, la MMP-2 fue detectada, por
inmunohistoquímica, en la zona proliferativa y de expansión del CE, pero no en las CGT2rias,
mientras que la MMP-9 se observó en las tres zonas mencionadas.
Si bien, las alteraciones de la expresión y/o de la regulación de la actividad de las MMPs in-vivo
fueron analizadas en una amplia gama de patologías, los efectos de la ingesta materna de
alcohol sobre la expresión de las MMPs, embrionaria y trofoblástica temprana, no ha sido
previamente abordada.
Luego de la administración de alcohol hasta el día 8 de gestación, la inmunoexpresión de
MMP-9 en el sitio de implantación resultó significativamente disminuida, tanto en la zona de

Discusión
274
proliferación, en la de expansión del CE, así como en las CGTs 2rias y CGT 1rias. En concordancia
con este resultado, la expresión génica de la MMP-9 fue reducida en los CE aislados de las
hembras tratadas. Sin embargo, sólo el nivel de inmunoexpresión de MMP-2 resultó
incrementado en la zona de proliferación del CE, luego de la exposición a alcohol, ya que su
expresión génica, evaluada por RT-qPCR en CE aislados, no mostró diferencias significativas
con los CE de las hembras controles. Si bien, es importante para el éxito reproductivo murino
que ambas MMPs estén presentes, fue visto que la inhibición de las mismas, no inhibe
completamente la implantación, dado que dicho proceso ocurre aún en ratones deficientes
para MMP-2 o MMP-9 (Itoh y col., 1997; Plaks y col., 2013), donde otras proteasas podrían
compensar la ausencia de estas MMPs. Posiblemente éste sea el caso de las células
trofoblásticas de los embriones implantativos de las hembras tratadas, ya que el incremento
de la MMP-2 en la zona de proliferación del CE podría haber compensando la disminución de
la MMP-9 en dicha zona en el día 8 de gestación. Sin embargo, la expresión desregulada de
ambas MMPs ha sido asociada con importantes patologías gestacionales. Así, la ausencia o
baja expresión de MMP-9 se relaciona con preeclampsia en el humano (Plaks y col., 2013).
Plaks y colaboradores (2013), utilizando ratones deficientes (null) en MMP-9, mostraron sitios
de implantación en estadio E7,5 pequeños, junto con elevada tasa de reabsorciones, en
relación a las camadas wild type. Al examinar el CE de los ratones deficientes en MMP-9,
vieron que, en ausencia de MMP-9, la matriz que rodea a las células deciduales no es
eficientemente erosionada y fagocitada por las CGTs del CE, y por lo tanto se produjo una
invasión superficial del trofoblasto en la decidua materna, además de deficiencia en la
decidualización. Los autores observaron que el CE de los ratones deficientes en MMP-9
presentaba escaso desarrollo (área disminuida respecto de los wild type), estaban mal
orientados, mostraban acumulación de sangre en la periferia del CE y tapones de fragmentos
(debris) que bloquearían la invasión de las células trofoblásticas en el polo mesometrial de la
decidua. Nuestros resultados concuerdan con los obtenidos en el estudio mencionado. Los CE
de las hembras tratadas, expresando menor cantidad del mensajero como de la proteína de
MMP-9, fueron de menor tamaño y tuvieron exacerbada infiltración de glóbulos rojos
maternos, además de una reducida área de proliferación y expansión, tanto in-vivo como in-
vitro. Más aún, nosotros observamos alta fragmentación en la interfase entre las CGT del CE y
la decidua, del lado mesometrial, lo que probablemente, resulte de una incapacidad fagocítica
de las células trofoblásticas para eliminar las células deciduales muertas y abrir espacios para
el avance en el tejido materno. Así como fue sugerido por algunos autores (Plaks y col., 2013),
el trofoblasto del CE deficiente en MMP-9 tendrá un potencial de desarrollo restringido, y esta

Discusión
275
situación es la que probablemente contribuya a los defectos embrio-placentarios observados
en el estadio de organogénesis (día 10 de gestación), en nuestro modelo de ingesta de alcohol.
En el día 8 de gestación, la función placentaria es aún ejercida por los tejidos
extraembrionarios. Así, la placenta coriovitelina es la que lleva adelante el crecimiento del
embrión a través de la expansión e invasión de las células trofoblásticas de la interfase
materna, en todo el perímetro del embrión. El endodermo extraembrionario parietal, que
sintetiza y deposita los componentes de la membrana de Reichert, es una fuente de secreción
de MMP-9, siendo ésta un factor de regulación de la diferenciación y migración de las células
endodérmicas (Behrendtsen y Werb, 1997). Plaks y colaboradores (2013), hallaron que, en los
embriones deficientes para MMP-9, la migración y expansión de endodermo parietal y visceral
estaba deteriorada, con lo cual se podría ver obstaculizada la migración del trofoblasto
cercano al ectodermo extraembrionario, en la base del cono ectoplacentario, conduciendo a
un retraso en el desarrollo de los embriones deficientes en MMP9 en el estadio E7,5-8. Dado
que en nuestro trabajo, la inmunomarcación de MMP-9 en el endodermo y ectodermo
extraembrionario de las hembras tratadas no resultó diferente respecto del tejido de las
hembras controles, al menos las alteraciones morfológicas vistas en estos tejidos, luego de la
ingesta de alcohol, no pueden atribuirse a la disrupción de la expresión de la MMP-9. Sin
embargo, la inmunoexpresión de MMP-2 en el endodermo parietal sí fue incrementada,
sugiriendo un papel de esta proteasa tanto en las anomalías morfológicas observadas en este
tejido como en la función placentaria coriovitelina de los embriones de las hembras tratadas.
Debido a que observamos alteraciones histomorfológicas en el ectodermo embrionario, junto
con una reducida expresión de E-cadherina en los embriones de las hembras tratadas,
analizamos la inmunoexpresión de las MMPs en este tejido. Luego de la ingesta de alcohol
hasta el día 8, el ectodermo embrionario evidenció mayor inmunomarcación de MMP-2 y
MMP-9 que el mismo tejido en las hembras controles. Se ha visto que las MMPs pueden
producir una escisión excesiva de E-cadherina lo que se vinculó con la inducción de la
transición epitelio-mesenquimática exacerbada (Lochter y col., 1997). El incremento de las
MMPs en el ectodermo de las hembras expuestas, podría favorecer la ruptura de las
interacciones célula-célula y así relacionarse con la baja expresión de E-cadherina encontrada
en este mismo tejido.
Tomando como referencia otros modelos y patologías en relación al consumo de alcohol y su
incidencia en la expresión de MMPs, cabe mencionar, que el consumo de alcohol promueve
carcinogénesis mamaria por estimulación de la proliferación e invasión de células cancerosas a

Discusión
276
través del aumento en los niveles de MMP-9 y MMP-2 (Luo y Miler, 2000; Meng y col., 2000).
También otros autores (Aye y col., 2004), demostraron que el etanol puede activar la MMP-2
de fibroblastos expuestos a etanol, alterando significativamente el comportamiento invasivo
de las células tumorales. Mediante ensayos in-vitro en células HepG2, se demostró que el
acetaldehído, incrementa la expresión génica y la actividad de MMP-9, promoviendo la
invasión celular (Hsiang y col., 2007). También, en la fibrosis hepática asociada al abuso de
alcohol, la expresión de MMPs y sus inhibidores, los TIMPs, se encuentra alterada (Tsukada y
col., 2006; Friedman, 2008, Thompson y col., 2011).
En resumen, de la misma manera a lo observado por otros autores, sugerimos que las
alteraciones significativas en la expresión MMPs principalmente en el trofoblasto invasivo y en
el tejido embrionario, al día 8 de gestación podrían ser las que, también en nuestro modelo de
ingesta de alcohol, causen el importante retraso de diferenciación y reabsorción embrionaria
visto al día 10 de gestación (Coll y col., 2011).
- Efectos del consumo perigestacional de alcohol en la inducción de genotoxicidad materna
y/o embrionaria y en la muerte celular embrio-trofoblástica.
Bajo la hipótesis de un efecto genotóxico-apoptótico en los embriones periimplantativos,
inducido por la exposición materna a alcohol, que lleva a pérdida gestacional temprana y a
alteraciones morfológicas embrionarias, posiblemente disparado por la disrupción de las
uniones intercelulares embrionarias así como por la elevada síntesis de MMPs, se
monitorearon los cambios morfológicos nucleares, el índice de apoptosis y el efecto
genotóxico materno luego de la ingesta periconcepcional de alcohol hasta el día 4 y 8 de
gestación.
En primera instancia, cabe mencionar que una elevada fragmentación embrionaria también es
indicador de daño genotóxico. Como se describió anteriormente, las hembras CF-1, y no las
CD-1, tuvieron alto índice de embriones fragmentados luego de la ingesta de alcohol hasta el
día 4 de gestación.
Fragmentación celular es observada frecuentemente en embriones de preimplantación
(Antczak y Van Blerkom, 1999) y es diferente de la fragmentación del ADN asociada a la
apoptosis característica de estadios tardíos del desarrollo de preimplantación (Hardy y col.,
2001; Spanos y col., 2002). Tanto una como otra han sido correlacionadas negativamente con
el potencial de implantación (Alikani y col., 1999; Pelinck y col., 2010). En base a esto

Discusión
277
pensamos que, al menos en CF-1, existe una importante pérdida embrionaria periimplantativa
que conduce a la disminución del número de sitios de implantación observada en día 8 de
gestación luego del consumo de alcohol. Se ha sugerido que una gran proporción de los
embriones que exhiben fragmentación celular son aneuploides (Chavez y col., 2012), ya que de
estudios de colocalización de un marcador centromérico (CENP-A) y de un marcador de
envoltura nuclear (LAMIN-B1), se pudo corroborar que los fragmentos contenían cromosomas
envueltos por membrana nuclear. Esto también indicó la existencia de micronúcleos (MN)
embrionarios, estructuras que encapsulan cromosomas perdidos producto de ruptura del ADN
dentro de envoltura nuclear, formando un MN antes de ser secuestrados en un fragmento
citoplasmático. En base al timing de fragmentación, se sugirió que el embrión de
preimplantación puede inicialmente sufrir fragmentación, más que muerte celular, en
respuesta a una posible aneuploidía generada como consecuencia de inestabilidad genómica.
Sin embargo, aún no está claro si la generación de MN o la fragmentación celular representan
una forma de corregir una aneuploidía embrionaria o son el inicio de un daño en el desarrollo.
Así, se sugirió que a medida que avanza el desarrollo, los fragmentos citoplasmáticos pueden
permanecer como unidades separadas, que llevan ADN cromosómico y citoplasma, pero
alternativamente, estos fragmentos pueden ser reabsorbidos por la misma blastómera que los
generó o por una vecina, llevando a la formación de complejos patrones de aneuploidías
(mosaicismo), o entrar en apoptosis (Chavez y col., 2012).
En los embriones de preimplantación, luego de la ingesta de alcohol hasta el día 4 de
gestación, se realizó un análisis genotóxico mediante la evaluación de alteraciones de los
núcleos interfásicos, entre ellas la observación de la frecuencia de MN en el total de núcleos
interfásicos cuantificados en la población total embrionaria. En dichos embriones debido a la
división asincrónica que los caracteriza, son pocas las metafases que podrían obtenerse para
un análisis citogenético, por lo tanto, el ensayo de MN es utilizado para estudios de
genotoxicidad (Titenko-Holland y col., 1998). La frecuencia de MN en embriones de
preimplantación derivados de hembras tratadas con alcohol hasta el día 4 de gestación no
resultó diferente del valor observado en los embriones controles, siendo en ambos casos
prácticamente nulo. Sin embargo, una alta frecuencia de MN maternos fue detectada en las
hembras tratadas hasta el día 4 de gestación, respecto de la frecuencia de las hembras
controles. Estudios in-vitro e in-vivo, examinaron los intercambios de cromátides hermanas, la
formación de MN y las aberraciones cromosómicas, para corroborar la genotoxicidad del
etanol. Sin embargo, hoy en día, la información acerca de la capacidad del etanol para producir
cambios cromosómicos, continua siendo controvertida (Phillips y Jenkinson, 2001). Mientras

Discusión
278
que los estudios in-vitro, muestran un panorama más claro de los efectos genotóxicos del
alcohol, como lo indica el trabajo de Kayani y Parry (2010), quienes han demostrado que tanto
el etanol como el acetaldehído producen efectos genotóxicos, induciendo ambos la formación
de MN de manera dosis dependiente. Los estudios in-vivo continúan generando discusión.
Phillips y Jenkinson (2001) sostienen que los datos disponibles de los estudios sobre el etanol
como genotóxico, aún son contradictorios.
Varios autores demostraron que el alcoholismo crónico severo en humanos puede causar daño
en los cromosomas de linfocitos de sangre periférica (Burim y col., 2004), induciendo aumento
de MN (Maffei y col., 2000, Almeida y col., 2002). Sin embargo, muchos informes sobre las
aberraciones cromosómicas in-vivo, producto de la exposición a alcohol en los animales,
fueron negativos. El etanol no mostró ningún efecto sobre la incidencia de MN en médula ósea
de rata, cuando éste fue administrado en el agua de bebida al 5% durante 10 a 30 días
(Balansky y col., 1993) o al 10% y 20% durante 3 a 7 semanas (Tates y col., 1980). También, se
observó un aumento no significativo en la frecuencia de aberraciones cromosómicas cuando se
administró etanol al 20% a ratas durante 30 días (Tavares y col., 2001). Ellahueñe y
colaboradores (2012), en ninguna de las dosis de etanol ensayadas (5, 10, o 15% v/v)
encontraron un aumento en la frecuencia de MN en eritrocitos policromáticos (PCE) de
médula ósea de ratón respecto de los valores controles. Sin embargo, nuestro grupo demostró
que la ingesta de etanol 10% durante 27 días indujo la frecuencia de MN en médula ósea de
ratones machos y hembras (Cebral y col., 2011). Más aún, la exposición perigestacional a
etanol 10%, hasta el día 10 de preñez, en CF-1, aumentó los MN en la médula ósea materna.
En concordancia, Kotova y colaboradores (2013), también demostraron que el consumo de
alcohol 10% por 4 semanas en ratas resultó genotóxico. El incremento de MN observado en los
PCE de médula ósea de las hembras expuestas, en el presente trabajo, confirma nuestra
hipótesis de un efecto genotóxico materno y concuerda con los estudios previos realizados en
el modelo murino, expuesto hasta el día 10 de gestación (Cebral y col., 2011) y con los de
Kotova y colaboradores (2013).
Kotova y colaboradores (2013), indicaron al acetaldehído como el principal metabolito
inductor de inestabilidad genómica y por lo tanto, de genotoxicidad, que de manera dosis
dependiente y como consecuencia de las alteraciones cromosómicas, induce citotoxicidad,
ruptura en la doble cadena del ADN y un retraso en la progresión de la replicación celular.
Como posibles explicaciones a las diferencias encontradas en los reportes in-vivo, surgen
algunas hipótesis. Los niveles diferenciales de acetaldehído en suero de humanos, que inducen
genotoxicidad en forma variable, son dependientes de los polimorfismos de las enzimas que

Discusión
279
metabolizan el alcohol, lo que explica las diferencias encontradas en la susceptibilidad humana
a sufrir daño genotóxico. Particularmente, se ha reportado que individuos con genotipo
ALDH2*2 tienen un nivel de acetaldehído en suero más alto entre los bebedores de alcohol
(Neumark y col., 2004), y evidenciaron una mayor inducción de MN en PCE, que aquellos que
expresan la isoforma ALDH2*1. Otra situación que explicaría la ausencia de efecto genotóxico
luego de un tratamiento prolongado in-vivo con etanol en roedores, tiene que ver con que el
consumo crónico de etanol estimula al sistema MEOS, el que oxida rápidamente el alcohol,
generando un aumento de la tolerancia/adaptación al mismo (Lieber, 1997). Esta
adaptación/tolerancia a concentraciones de etanol sostenidas en el tiempo evita que el alcohol
en circulación alcance niveles excesivos (Tavares y col., 2001). Sin embargo, dado que el
sistema MEOS es un reconocido generador de radicales libres (Nordmann y col., 1992) y de
estrés oxidativo (Izzotti y col., 1998), la hipótesis de la adaptación al alcohol debe tomarse con
recaudo.
Del análisis de los núcleos interfásicos, encontramos un incremento de picnosis nuclear en los
blastocistos iniciales de hembras tratadas CD-1 y en blastocistos expandidos de tratadas CF-1.
Estos núcleos con cromatina altamente condensada se asocian a la muerte celular por
apoptosis, que refleja en muchos casos, la eliminación de células individuales sin impacto
negativo en la viabilidad y progreso del desarrollo del embrión, como probablemente ocurre
en los embriones controles. Sin embargo, si este proceso se viera exacerbado, esto podría
acarrar consecuencias negativas para el desarrollo embrionario subsiguiente. Por lo tanto,
dado que la calidad del blastocisto predice su capacidad para implantarse y su futuro
crecimiento, el siguiente paso fue esclarecer sobre la posibilidad que la alta picnosis se
asociara con un elevado índice de apoptosis en los mismos embriones de hembras tratadas. La
apoptosis, sin dudas, está relacionada con el control de calidad del embrión, ya que juega un
papel activo en el desarrollo del embrión a través de la eliminación de células genéticamente
anormales, brindando así una función protectora del período previo a la implantación (Hardy,
1999). Pero, en los blastocistos, la muerte celular programada, además de eliminar células
dañadas, está asociada con la diferenciación celular, porque la apoptosis ocurre
frecuentemente para eliminar aquellas células del MCI que a medida que pierden su
totipotencialidad retienen su capacidad de formar TE (Brison y Schultz, 1997). Así se logra
mantener la fidelidad del MCI librando al embrión de células aberrantes. Por lo tanto, un
patrón normal de apoptosis en los blastocistos de mamíferos, es crucial para el desarrollo, ya
que funciona como mecanismo controlador temprano de la salud de la futura descendencia

Discusión
280
(Byrne y col., 1999). Nosotros observamos estos patrones de apoptosis en los blastocistos
iniciales y expandidos de las hembras controles. Los resultados obtenidos destacan que los
blastocistos iniciales tienen mayor índice de apoptosis que los expandidos, en la situación
normal. Además, tanto en los blastocistos iniciales como en los expandidos, se evidenció, una
alta apoptosis en el MCI, mientras que fue escasa en el TE mural y polar. Al respecto cabe
destacar que los blastocistos iniciales tienen ambos linajes celulares en plena diferenciación, y
que por eso, el control de la “fidelidad” sería más importante que en el estadio ligeramente
más avanzado, en los blastocistos expandidos. Así como fue observado en el presente trabajo,
en la mayoría de los casos, la pérdida de células por apoptosis en los blastocistos ocurre
predominantemente a expensas del MCI y no del TE (Pampfer, 2000). Un incremento en la
relación TE/MCI puede conducir a un desbalance del tamaño placentario y fetal (Leese y col.,
1998). Experimentos más antiguos aún, demostraron que el MCI ejerce un cierto control en el
crecimiento del TE. Ansell y Snow (1975) mostraron que cuando la cantidad de células del MCI
se reduce significativamente en el blastocisto, la proliferación del trofoblasto se ve limitada y
la transformación a CGTs ocurre más rápidamente luego de la implantación. Esta alteración en
la etapa de periimplantación puede ser el presagio de anomalías ultraestructurales en etapas
más avanzadas (Ansell y Snow, 1975). Se ha planteado que el incremento de la apoptosis en las
células del MCI puede reflejar una mayor susceptibilidad intrínseca de estas células en relación
a las TE (Brison y Schultz, 1997). Las células del linaje del MCI y del TE divergen
tempranamente en respuesta a una combinación compleja de acontecimientos celulares y
moleculares. Así, las diferencias resultantes en los requerimientos metabólicos,
posicionamiento celular y microambiente se consideran algunas de las principales causas
subyacentes de la sensibilidad diferencial de las MCI y TE observada bajo ciertas circunstancias,
por ejemplo, ante agentes embriotóxicos. En la mayoría de los casos, las células del MCI
parecen menos resistentes a la alteración que las células del TE (Pampfer, 2000). La diferente
susceptibilidad del MCI vs el TE en la muerte celular inducida puede estar asociada con la
expresión diferencial de alguno de los principales genes involucrados en el control o ejecución
de la apoptosis. En relación a esto, se ha planteado la posibilidad que los dos linajes puedan
diferir en cuanto a los efectores pro- y anti-apoptóticos que expresan (Pampfer, 2000).
En función de su incidencia, el proceso apoptótico puede dejar de tener un efecto beneficioso,
y transformarse en un evento perjudicial o patológico (Jurisicova y col., 1996; Hardy, 1997).
Por lo tanto, la proporción de células apoptóticas se ha considerado uno de los parámetros
más importantes para la evaluación de la salud o calidad de los embriones. La frecuencia
umbral a partir de la cual la apoptosis es perjudicial en blastocistos de mamíferos aún no ha

Discusión
281
podido ser determinada (Fabian y col., 2007). En nuestro trabajo, como era esperado, tanto los
blastocistos iniciales como expandidos provenientes de las hembras tratadas con alcohol,
tuvieron un mayor grado de apoptosis que los controles. Particularmente, la ingesta materna
de alcohol indujo un alto número de células TUNEL-positivas en el TP de los blastocistos
iniciales, mientras que en los blastocistos expandidos, las zonas con mayor incidencia de
apoptosis correspondieron al MCI y al TM, lo que sugiere diferente sensibilidad de estas
poblaciones celulares de acuerdo al estadio de diferenciación del embrión preimplantativo.
Esta elevada incidencia de apoptosis en los blastocistos, como se explicó, puede tener claras
implicancias en la desregulación del proceso de implantación, el que está mediado por el TM,
aunque el TP participa en la invasión embrionaria en etapas más avanzadas, cuando éste
deriva en el cono ectoplacentario. Este resultado sugiere que el retraso embrionario, las
alteraciones en la cantidad de embriones que se diferencian adecuadamente durante la
implantación y las anomalías de la expansión trofoblástica del CE en las etapas de
postimplantación, observadas al día 8 de gestación, podrían relacionarse con el incremento de
muerte celular. Así, como las células del TE contribuyen a la placentación y son requeridas para
el desarrollo del concepto (Cross y col., 1994), una reducción en este linaje celular podría
reducir la implantación y la viabilidad embrionaria temprana o tardíamente (Pampfer y col.,
1990a; Kelly y col., 1992). Por otra parte, distorsiones del patrón de apoptosis en el MCI del
blastocisto pueden comprometer la maduración embrio-fetal a futuro, pudiendo conducir ya
sea a muerte embrionaria temprana o a la formación de anomalías en el feto (Brison y Schultz,
1997). Así, si la proporción de células apoptóticas es elevada, esto perjudicaría la homeostasis
del embrión, pudiendo no implantarse, detener su desarrollo o morir (Betts y King, 2001). Es
más probable que embriones con un gran número de células en relación con otros con menor
número, se implanten y den lugar a crías vivas (Van Soom y col., 1997). Byrne y colaboradores
(1999) muestran que los blastocistos con un pequeño número de células y, por lo tanto,
posiblemente con un potencial reducido de desarrollo, presentan una alta incidencia de
apoptosis. Otros estudios muestran que una reducción del 30% o más en el número de células
del MCI, por apoptosis excesiva, está asociada a un riesgo de pérdida embrionaria o a
desarrollo anormal, incluso cuando la tasa de implantación y el número de células TE es
normal (Tam, 1988). Por lo tanto, la cantidad de células del MCI es esencial para la adecuada
implantación del embrión, ya que una reducción en este linaje celular puede potencialmente
causar una disminución en la viabilidad (Pampfer y col., 1990a; Kelly y col., 1992), retraso y
malformaciones en el desarrollo embrionario de postimplantación (Pampfer, 2000).

Discusión
282
La alta incidencia de apoptosis embrionaria asociada al consumo de alcohol hasta la etapa de
preimplantación nos llevó a investigar si este efecto adverso, producto de la exposición
continua a alcohol, también se producía en los tejidos embrionarios, extraembrionarios y
trofoblásticos, en el período de gastrulación, al día 8 de gestación. De esta manera, se intentó
clarificar si los embriones en estadio más avanzado del desarrollo resultaban igualmente
susceptibles a los efectos adversos de la exposición a alcohol. Como primera aproximación, un
análisis de la organización y morfología nuclear, junto con el grado de compactación de la
cromatina fue realizado mediante marcación del ADN con Hoechst 33342. Las alteraciones
nucleares más evidentes se hallaron en la interfase trofoblástico-decidual y en el ectodermo
embrionario. La interfase materno-embrionaria lateral al embrión de los sitios de implantación
de las hembras tratadas mostró una gran desorganización nuclear respecto a lo observado en
las muestras controles. A su vez, la zona antimesometrial de la interfase trofoblástico-decidual
de las hembras tratadas tuvo alta fragmentación y gran heterogeneidad morfológica, sobre
todo relacionada con el tamaño nuclear, ya que se observaron núcleos llamativamente
grandes comparados con los núcleos de las muestras controles. El ectodermo de los embriones
de las hembras tratadas también mostró alteraciones morfológicas nucleares respecto del
control. En términos generales, luego de la ingesta de alcohol, en todas las muestras se
detectaron fragmentos nucleares y alta proporción de núcleos con cromatina altamente
condensada, resultados que llevaron a la evaluación de la apoptosis en dichos tejidos.
También en la etapa de postimplantación, la apoptosis participa de la eliminación de células
anormales, deslocalizadas, no funcionales o perjudiciales, controlando el número de células y
desempeñando así un papel clave en la adecuada formación de las estructuras
extraembrionarias y del propio embrión (Jacobson y col., 1997; Runic y col., 1998). Durante la
gestación normal, se ha identificado una determinada proporción de células apoptóticas tanto
en el compartimiento materno (células deciduales) como en el embrionario (trofoblasto),
dependiendo del estadio gestacional y la presencia de estas células apoptóticas en adecuada
proporción está relacionada con distintas etapa del desarrollo, incluyendo la adhesión y la
invasión del trofoblasto, la transformación de las arterias espiraladas, la diferenciación y
renovación del trofoblasto, y el parto. A su vez, también se ha demostrado que la apoptosis
desempeña un papel importante promoviendo la tolerancia inmunológica materna a los
antígenos paternos expresados por las células trofoblásticas (Huppertz y col., 2006). Dada
entonces la importancia de la apoptosis para el desarrollo normal del embrión y de la placenta,
alteraciones en la regulación de la muerte celular contribuyen a la inducción de múltiples
patologías de la gestación. Los abortos espontáneos, los partos prematuros, la preeclampsia y

Discusión
283
la restricción del crecimiento intrauterino, la mortalidad perinatal, entre otros (Huppertz y col.,
2006), han sido atribuidos al deterioro de la invasión trofoblástica por apoptosis exacerbada de
las células trofoblásticas. Si el aumento de la apoptosis del trofoblasto se produce al principio
de la gestación, puede verse limitada la invasión a las arterias uteroplacentarias, reduciéndose
su remodelación y en consecuencia, generándose una disminución en el flujo de sangre
materna a la placenta. En condiciones normales, las células trofoblásticas inducen apoptosis de
las células endoteliales y del músculo liso vascular. Esta remodelación arterial, asegura el
suministro adecuado de sangre al feto en desarrollo. Así, defectos en la funcionalidad invasiva
del trofoblasto y/o una falta de respuesta del endotelio o del músculo liso vascular a estímulos
proapoptóticos trofoblásticos, pueden explicar una escasa transformación vascular, como en el
caso de patologías como la preeclampsia (Pringle y Roberts, 2007). Por otra parte, los abortos
espontáneos también se asocian a una escasa o nula invasión trofoblástica debido a un
aumento de los niveles de apoptosis en el trofoblasto (Khong y col., 1987).
Según nuestros hallazgos en el día 8 de gestación, mientras la zona proliferativa del CE de las
muestras controles y de las tratadas no presentó células TUNEL-positivas, la zona de expansión
de los embriones de ambos grupos experimentales, sí tuvo abundante y similar cantidad de
células con marcación TUNEL-positiva. Estos elevados niveles de apoptosis son esperados en
dicha zona, ya que derivan de la diferenciación del TP de los blastocistos, donde se encontró
alta tasa de muerte celular. Sin embargo, como el trofoblasto de la zona de expansión está en
pleno proceso de invasión mesometrial, muchas de las células apoptóticas pueden
corresponder a células deciduales que, luego de su muerte, son fagocitadas por las células
trofoblásticas. Sin embargo, apoyándonos en las observaciones histológicas de los embriones
de las hembras tratadas, en que los mismos presentaron un escaso desarrollo del CE,
sugerimos que las células apoptóticas cuantificadas podrían corresponder a células
trofoblásticas del frente de invasión. En concordancia con esta idea, observamos que la
interfase trofoblástico-decidual lateral y antimesometrial de las muestras de las hembras
tratadas también presentó un mayor índice de apoptosis respecto de las muestras controles,
indicando que aún la invasión y expansión del lado antimesometrial del concepto, en el día 8
de gestación, se encuentra afectada por la exposición a etanol. Por lo tanto, una escasa
invasión trofoblástica, como consecuencia de una apoptosis exacerbada, podría ser
responsable de remodelación decidual anormal o deficiente y, en definitiva, ser causa de los
defectos placentarios tanto en fase de organogénesis como a largo plazo, en la gestación a
término.

Discusión
284
Mientras que en los tejidos extraembrionarios no fue detectada apoptosis, en el ectodermo
embrionario del embrión en gastrulación de las hembras tratadas se presentó un alto número
de células apoptóticas respecto de las hembras controles, resultado que se condice con
diversos estudios vinculados a la apoptosis y el FAS. La desregulación del patrón normal de
muerte de células embrionarias es uno de los eventos más importantes que preceden a la
aparición de anomalías estructurales independientemente de su naturaleza (Brill y col., 1999).
En particular, la apoptosis es muy frecuente en los embriones de postimplantación expuestos
al alcohol. Ha sido documentado que las células de embriones expuestas a alcohol presentan
ADN nuclear fragmentado y condensado en fragmentos picnóticos, signos característicos de la
apoptosis (Sulik y col, 1988; Cartwright y col., 1998).
El análisis de los defectos craneofaciales inducidos por alcohol observados en humanos reveló
ciertos elementos comunes entre las estructuras fetales afectadas. Gran parte de estas
estructuras derivan de un subconjunto de células de la cresta neural (CCN), las CCN craneana
(Noden, 1991). Estudios de Sulik y colaboradores (1988) en ratones destacaron que las
malformaciones craneofaciales (exencefalia, labio leporino, hipoplasia maxilar), observadas en
la etapa fetal, se debían a la exposición a alcohol de embriones murinos de 7, 8 o 9 días de
gestación, la que indujo apoptosis dentro de las 12 horas de la exposición. Así, la muerte
celular generalizada dentro de regiones embrionarias enriquecidas con CCN proporcionó
fuerte evidencia que dicha población es blanco directo del alcohol (Sulik y col., 1988; Sulik,
2005). Para investigar el mecanismo de la muerte de las CCN y sobre todo identificar la
ventana de sensibilidad crítica a la apoptosis inducida por alcohol, algunos investigadores
recurrieron al embrión de pollo. Como se vio en ratones, embriones de pollo expuestos a
niveles de alcohol superiores a 0,15% exhibieron muerte celular significativa en las CCN
craneana y en ciertas secciones del cerebro primitivo (Cartwright y col., 1998). Llamativamente
y al igual que en los embriones de ratón, escasa muerte celular fue observada otros
precursores/tejidos en desarrollo. Esta observación enfatizó la sospecha que la muerte celular
probablemente no era una consecuencia de la toxicidad generalizada, sino que poblaciones
particulares de células embrionarias eran susceptibles diferencialmente el efecto inducido por
alcohol. Los estudios en pollo tendientes a identificar la ventana de sensibilidad crítica
revelaron que la exposición a alcohol causó muerte celular sólo si el alcohol se administraba
antes de la migración de las CCN de su lugar de nacimiento en el neuroectodermo,
demostrándose que una vez que las células comenzaban a migrar hacia su destino, se
tornaban resistentes a la muerte inducida por el alcohol y, por lo tanto ya no se originaban

Discusión
285
defectos faciales. Así, se sugirió que la ventana de sensibilidad crítica para la producción de
malformaciones faciales es muy estrecha, extendiéndose sólo desde la gastrulación a la
migración de la CCN (Cartwright y Smith, 1995). Una ventana de sensibilidad idéntica para la
inducción de defectos craneofaciales fue encontrada en ratones, correspondiendo al corto
período de tiempo durante la formación de tres capas germinales (la gastrulación) y durante la
organización del sistema nervioso primitivo (la neurulación) (Webster y col., 1980). En
roedores, cuando el etanol se administró durante la gastrulación (justo antes y/o durante la
migración de las CCN a las regiones cervicales y craneofaciales) se generó un daño en la placa
neural anterior, que resultó en malformaciones craneofaciales (Sulik y col., 1988). Sulik y
colaboradores (1988) proporcionaron otros datos significativos respecto de la regulación de la
apoptosis. Reportaron que la muerte celular inducida por teratógenos es más frecuente en las
regiones embrionarias ya sometidas a algún grado de muerte celular endógena durante la
morfogénesis. Cartwright y Smith (1995) señalaron la misma observación en el modelo de
pollo. La coincidencia de la muerte inducida por el alcohol con la incidencia normal de ésta en
la cresta neural, llevó a la hipótesis que la exposición a alcohol activa de forma aberrante la
maquinaria apoptótica en las CCN craneanas (Smith, 1997).
Ya más recientemente, Dunty y colaboradores (2001), investigaron el patrón tanto de la
muerte celular programada normal como el resultante de la inducida por etanol, en el sistema
nervioso central en desarrollo y en la región craneofacial, a intervalos de 0,5 días, entre los
días 6,5 y 11 de gestación en el ratón. Una dosis aguda de etanol (2,9 g/kg) administrada por
vía intraperitoneal a ratones C57BL/6J preñados 12 hs antes del análisis de la apoptosis, dio
como resultado un aumento dramático en la muerte celular principalmente en regiones donde
normalmente este proceso ocurría como parte del proceso de morfogénesis. Las muestras de
embriones expuestos a etanol exhibieron muerte celular excesiva en distintas poblaciones de
células dependientes de la etapa del desarrollo en que se aplicara el etanol, en particular, se
vio afectado el tejido que daría origen al sistema nervioso central. Así, la apoptosis inducida
por etanol fue notable en el neuroectodermo al día 7,5 de gestación, en la placa neural y en la
línea primitiva al día 8 de gestación; en la cresta neural presuntiva de la parte posterior del
cerebro rostral al día 9 de gestación; en los arcos branquiales y rombomeros al día 9,5-10; y en
el diencéfalo, ganglios basales y en el cerebelo en desarrollo al día 11 de gestación.
Corroborando que poblaciones de células específicas de acuerdo a la etapa del desarrollo del
cerebro y región craneofacial, son vulnerables a la apoptosis inducida por etanol, clarificando
aún más la génesis de los defectos de nacimiento relacionados con la apoptosis y el alcohol.

Discusión
286
Así, en base a las alteraciones asociadas al FAS quedó plasmado en los reportes de Sulik y col.,
1988 (en ratones) y de Cartwright y col., 1998 (en pollos) que las CCN son particularmente
vulnerables a la muerte inducida por el alcohol, y si bien, los mecanismos de malformación
craneofacial son complejos, una característica común es la apoptosis excesiva que el embrión
puede ser incapaz de compensar (Sulik y col., 1988).
La comprensión de la regulación molecular de la apoptosis ayuda a explicar cómo el alcohol
inicia la vía apoptótica y da como resultado la muerte de algunas, aunque no todas, las
poblaciones de células. De modo que se plantearon algunas hipótesis sobre los mecanismos
que activarían esta vía después de la exposición al alcohol. Muy probablemente, el
desencadenante de la apoptosis inducida por alcohol se encuentra dentro de la misma
población de células blanco, debido a que muchas otras poblaciones de células no mueren a
pesar de exposiciones equivalentes a alcohol. Un de las hipótesis con la vía del ácido retinoico
(AR). El AR es esencial para la supervivencia de la cresta neural, es regulador clave de la
morfogénesis embrionaria y un potente mediador de la diferenciación en muchos linajes
celulares. La exposición al alcohol se asocia con deficiencias nutricionales de AR que afecta
potencialmente a los embriones en desarrollo (DeJonge y Zachman 1995; Deltour y col., 1996).
Debido a que los bajos niveles de AR resultan en la apoptosis de las poblaciones de la cresta
neural y su posterior eliminación del embrión (Maden y col., 1997), la deficiencia de AR podría
estar implicada en la apoptosis inducida por alcohol. Deltour y colaboradores (1996)
demostraron que las concentraciones de alcohol que se encuentra en las mujeres alcohólicas
están asociadas con la reducción de los niveles de AR. Otro mecanismo propuesto para la
inducción de apoptosis inducida por alcohol implica la capacidad de éste para perturbar la
función de las proteínas asociadas a la membrana de las células y así interferir en la
comunicación intercelular. En este proceso, quizás a través de interacciones directas proteína-
alcohol, se podría causar la activación aberrante o la inhibición de las vías de comunicación
vitales para la supervivencia celular. Utilizando diferentes líneas de células en cultivo, se ha
documentado la capacidad del alcohol para alterar la comunicación intracelular a través de
una variedad de mecanismos dependiendo de la célula blanco (Stachecki y col., 1994). Por lo
tanto, no se descarta la interferencia del alcohol con señales intracelulares críticas para la
supervivencia celular durante el desarrollo. Por último, una de las explicaciones más relevante
inherentes al mecanismo que subyace a la apoptosis se basa en la capacidad del alcohol para
causar lesión celular por el estrés oxidativo generado como consecuencia de su metabolización
(Kotch y col., 1995). La exposición a alcohol puede incrementar los niveles de moléculas
reactivas del oxígeno, llevando a las células a agotar potencialmente los compuestos de

Discusión
287
protección (antioxidantes). Bajo estas condiciones de estrés oxidativo, los radicales libres
interactúan con componentes vitales de la célula provocando severas lesiones. Uno de los
mecanismos mejor conocidos es la peroxidación lipídica (Aitken y col., 1989; Halliwell y Chirico,
1993). Por esta vía de estrés oxidativo, además del daño en los lípidos de la membrana, se
interfiere con los sistemas de regulación celular, con la comunicación intercelular, la síntesis de
proteínas y con la estructura y función del ADN lo que lleva a apoptosis (Halliwell y Aruoma,
1991; Wells y col., 2009). Aunque el embrión está equipado con mecanismos de defensa
antioxidantes enzimáticos, varios estudios han demostrado que la expresión de las enzimas
superóxido dismutasa, catalasa y glutatión peroxidasa es baja durante el desarrollo
embrionario temprano en comparación con los niveles maternos (Zaken y col., 2000), por lo
que el embrión es susceptible al daño inducido por estrés oxidativo generado por la exposición
a alcohol. En los embriones de preimplantación, las especies reactivas del oxígeno (ROS),
presentes en fluido blastocélico, inducen muerte de las células del MCI (Gramzinski y col.,
1990).
Mientras que algunos embriones no resultaron afectados, total o parcialmente, por la
exposición a alcohol, probablemente por la potencial capacidad que tienen para reparar el
daño ejercido por el alcohol y sus derivados, por la redundancia de señales celulares que
compensen la interferencia del alcohol con los mecanismos de señalización, otra importante
proporción de embriones, sufrió un significativo nivel de apoptosis. Ésta, probablemente,
tenga relación con deficiencia de AR, aumento de ROS, alteraciones en la señalización
intracelular, y como hemos visto, por inducción de alteraciones en las uniones intercelulares y
aumento de la expresión de MMPs, entre otros mecanismos.
Los resultados obtenidos en el presente trabajo, refuerzan la idea que el éxito de procesos
fundamentales para el desarrollo del embrión, como la implantación, la invasión trofoblástica y
la gastrulación, es la consecuencia de una compleja interacción entre diversos factores tanto
de origen materno como embrionario, que dirigen y regulan el timing apropiado de cada
evento. La ingesta de alcohol, desde antes de la gestación y hasta etapas tempranas del
desarrollo embrio-trofoblástico, inclusive en baja concentración, es suficiente como factor de
riesgo para la generación de daño en embriones tempranos, y para la salud del individuo en
desarrollo inclusive en la vida postnatal.
4. Consumo perigestacional de alcohol: efectos en el desarrollo feto-placentario en la
gestación a término.

Discusión
288
Dado que, en nuestro modelo, la ingesta de alcohol desde antes de la gestación y hasta
períodos gestacionales tempranos induce anomalías embrionarias en los estadios de
preimplantación, implantación y gastrulación, nos propusimos investigar si estas alteraciones
conducían a anomalía fetal y placentaria a término, cuando la exposición a alcohol cesa en
etapas tempranas de la preñez (días 4, 8 o 10 de gestación), siendo asimismo de interés
determinar si la exposición murina perigestacional a concentración moderada de alcohol es
capaz de producir patología perinatal típica del FAS.
El alcohol es uno de los teratógenos más comunes y su consumo no controlado en mujeres
gestantes puede originar un daño postnatal irreversible en el individuo en desarrollo. Así, la
exposición excesiva prenatal a alcohol puede afectar gravemente el desarrollo físico y
neuroconductual de un niño. El FAS es considerado un cuadro patológico que refleja las
manifestaciones máximas del abuso de alcohol durante la gestación, caracterizado por las
malformaciones congénitas, retraso mental, restricción del crecimiento, deficiencias
funcionales en general. Estos efectos teratogénicos han sido demostrados después de la
exposición tanto aguda (1 o 2 días) como crónica de etanol, empleando una variedad de vías
de administración en diversos modelos animales (Becker y col., 1996).
Dada la alta incidencia de FAS (entre el 9 al 10/1000 de los nacimientos vivos) (Guilliam y col.,
2011), es que resulta de interés su abordaje en modelos experimentales de investigación. Si
bien a partir del reconocimiento del embarazo, la mayoría de las mujeres en forma espontánea
reducen el consumo de alcohol, dado que muchas veces el reconocimiento del mismo no se
produce hasta 4ta o 5ta semanas de gestación, existe la posibilidad que hayan consumido
alcohol en las etapas más tempanas de la gestación o previo a la misma, situación que
representa un riesgo de anomalía congénita propia del FAS. Los niños que incluyen una historia
confirmada de exposición a alcohol por consumo materno antes de nacer, muchos de ellos
presentan todas las características típicas del FAS, otros, presentan un fenotipo parcial en
relación al síndrome (generalmente no presentan las características físicas del FAS pero sí los
déficits cognitivos y problemas conductuales), mientras que algunos, no muestran ningún
síntoma en absoluto, condición conocida como resistencia a etanol (Riley y McGee, 2005;
Gemma y col., 2006, 2007). La razón de las variaciones fenotípicas observadas está relacionada
con la existencia de factores que moderan la gama de resultados. Sin duda, uno de los
principales factores que influyen en los resultados es la cantidad de alcohol que alcanza el
embrión o el feto en desarrollo. Esto está, en parte, determinado por la dosis y el patrón de
exposición a alcohol, además de la capacidad individual de metabolización.

Discusión
289
En el presente trabajo, vimos que el mismo patrón de exposición a alcohol, desde antes de la
gestación y hasta los días 4, 8 o 10 de gestación, genera bajas alcoholemias en las hembras de
las dos colonias murinas, CD-1 y CF-1, utilizadas como modelos para explicar las respuestas
variables que se presentan en la población humana luego de la ingesta de alcohol. Al respecto,
los estudios en animales de experimentación, han proporcionado pruebas del potencial
teratogénico del etanol mediante el control de numerosos factores, sin que factores
“confundentes” asociados comúnmente al consumo crónico de alcohol en la población
humana, como la desnutrición, las enfermedades, el tabaquismo y el consumo de otras drogas,
puedan interferir en los resultados. Por otro lado, mediante el control del patrón de consumo
(dosis, vías de administración, tiempo y período). Además, el uso adecuado de modelos
animales permite evaluar el papel relativo de factores genéticos que influyen en la
susceptibilidad al etanol (Becker y col., 1996). Como fue mencionado, los roedores han sido la
especie más comúnmente utilizada para estudiar los efectos perjudiciales de la exposición
prenatal a etanol, en donde todas las características clínicas distintivas del FAS han sido
demostradas (Becker y col., 1989; Gilliam y col., 2011). Un número de cepas se han utilizado en
diversos estudios y la respuesta variable, es decir, la susceptibilidad a los efectos dañinos del
etanol sobre el crecimiento y el desarrollo fetal vistas entre las cepas apoyan la idea de que los
factores genéticos influyen, en parte, en la vulnerabilidad a etanol (Becker y col., 1996). En
particular, dichos estudios han puesto en evidencia la mayor sensibilidad de las cepas puras en
relación a los ratones consanguíneos (colonias) (Webster y col., 1980). Dado que en el
presente trabajo, el objetivo fue abordar las consecuencias de la ingesta materna
perigestacional de alcohol para explicar, al menos en parte, los efectos observados en el
humano, se utilizó el modelo exocriado.
El consumo prolongado de alcohol puede ser causa tanto de desnutrición primaria, por
generar un comportamiento de tipo anoréxico, como de desnutrición secundaria, por mala
absorción de nutrientes (Almeida Domingues y col., 2009). La posibilidad de que la
desnutrición materna induzca malformaciones fetales no ha reunido apoyo clínico o
experimental nacer (Stein y Susser, 1975; Lumey, 1992; Lumey y Stein, 1997). Sin embargo, si
la desnutrición conduce o se combina con alguna deficiencia nutricional específica, sí se han
reportado malformaciones bajo dichas circunstancias. Estas condiciones incluyen deficiencia
de folato (Czeizel, 2000), vitamina B12 (Ray y Blom, 2003), vitamina K (Menger y col., 1997), o
vitamina A. Por otra parte, el déficit alimentario materno incide frecuentemente en el peso al
nacer en la descendencia, por lo que ha sido asociado a la restricción de crecimiento
intrauterino (Wentzel y col., 2006). Por ello, en primer lugar evaluamos si la ingesta

Discusión
290
perigestacional de alcohol produce desnutrición en la madre expuesta. En este contexto,
nuestros resultados indicaron que el consumo calórico total al final de la gestación no fue
modificado en los grupos tratados hasta el día 4, 8 o 10 de gestación, lo que correlacionó con
la ausencia de cambios en los pesos corporales de las hembras tratadas a término, en relación
con los pesos de las hembras controles.
Por otra parte, es sabido que el momento gestacional de exposición a alcohol determinará, en
parte, el grado y tipo de anomalía feto-placentaria (Riley y McGee, 2005). Por eso, las
respuestas fetotóxicas al alcohol reflejan una etiología multifactorial (Riley y McGee, 2005). El
nivel mínimo de concentración de etanol que resulta en FAS todavía no fue establecido. Ya
desde hace algunos años varios estudios admiten que incluso la exposición prenatal a bajos
niveles de alcohol puede afectar negativamente el desarrollo fetal (Charness y col., 1994). En
esta condición de consumo se contextualiza el presente estudio. Sin embargo, y como se
mencionó, es el BAC producido el parámetro de referencia para establecer comparaciones. En
nuestros modelos murinos exocriados expuestos, el nivel de BAC producidos al final del
tratamiento, fueron bajos (cernos a 25-30 mg/dl). Mucho se ha debatido sobre el papel de los
niveles de BAC en la etiología del FAS y al respecto, parece que no existiera un consenso en la
literatura sobre la dosis teratogénica, principalmente en lo relativo a los valores moderados y
bajos de BAC, ya que los valores altos, sin lugar a duda, generan severas alteraciones
(malformaciones fetales, restricción del crecimiento, muerte intrauterina, alteraciones
postnatales a largo plazo). Generalmente, en los modelos de exposición aguda, por vía
intraperitoneal o por gavage, se prueba la incidencia de valores altos de BAC, entre 250 y 500
mg/dl, en días críticos del desarrollo (Webster y col., 1980). Sin bien, dichas concentraciones
pueden ser consideradas, en parte, poco representativas o realistas del consumo habitual en
humanos, la importancia de estos estudios radica en que ellos permiten determinar cuáles
serían las principales estructuras fetales afectadas, según el día de aplicación del alcohol. En
este contexto, los resultados obtenidos con el presente trabajo con un régimen de exposición
que genera valores bajos de BAC, contribuyeron al entendimiento del origen de las
alteraciones que se producen tempranamente y que se manifiestan en el desarrollo feto-
placentario a término. Más aún, lo interesante del régimen crónico es que permite el testeo de
dosis moderadas a bajas sostenidas en el tiempo y que abarquen diferentes períodos de
gestación.
Luego de la ingesta perigestacional de alcohol hasta fases tempranas del desarrollo, se analizó,
a término, la cantidad de sitios de implantación totales, el número de fetos desarrollados, la

Discusión
291
vitalidad fetal y la frecuencia de reabsorciones. Mientras que el número de sitios de
implantación en las hembras CD-1 que recibieron alcohol hasta el día 4 de gestación, no fue
diferente del control, en las hembras tratadas con alcohol hasta los días 8 y 10 de gestación, la
cantidad de sitios resultó aumentada, siendo este resultado similar a lo encontrado en el día 8
de gestación, luego de la ingesta de alcohol. La continua exposición a alcohol hasta la fase de
postimplantación temprana (días 8 y 10 de gestación) parece haber inducido en CD-1, por un
lado, el adelantamiento de la implantación embrionaria, además de la implantación de un
mayor número de embriones respecto al número de las controles, probablemente asociado a
una falla de los mecanismos que controlan el número de crías en la camada. Sin embargo, a
pesar de este resultado, no encontramos una diferencia significativa en la camada (cantidad de
fetos por hembra) en ninguno de los grupos tratados. Este mismo resultado también fue
observado por Oyedele y Kramer (2008), quienes luego de tratar hembras murinas C57BL/6J
bajo un régimen semicrónico de tres días consecutivos con etanol al 25% (v/v) mediante
gavage, en los días 7, 8 y 9, tampoco observaron una reducción en la número de fetos a
término respecto de los controles. Estos resultados están íntimamente relacionados con la
tasa de reabsorción que, si bien, en los tres grupos alcoholizados fue elevada en comparación
con los controles, alcanzando valores cercanos al 14% en el grupo tratado hasta el día 4, al 7%
en el grupo tratado hasta el día 8 y al 11% en el grupo tratado hasta día el 10 de gestación, no
fueron suficientes para generar una disminución en el tamaño de las camadas. Para su estudio,
las reabsorciones embrionarias encontradas al día 18 de gestación fueron clasificadas y
cuantificadas en términos del momento gestacional en pudieron originarse, identificándose
entonces, reabsorciones tanto tardías como tempranas. Si bien, todos los grupos tratados
presentaron alta tasa de reabsorción total, lo que pudo haber contribuido a la normalización
del número de fetos totales en los grupos alcoholizados, el número de reabsorciones
tempranas sólo resultó significativamente elevado en las hembras tratadas hasta el día 4 de
gestación. Esto demostró que la exposición a alcohol hasta el día 4 de gestación llevó a una
importante pérdida embrionaria inmediatamente después de la implantación, mientras que en
los grupos expuestos hasta los días 8 y 10 de gestación, este mecanismo temprano de control
que opera durante o luego de la implantación, no sería funcional, derivando en un número de
fetos aumentado al día 18 respecto de sus controles. Las reabsorciones en este caso, podrían
ser consecuencia de las alteraciones intrínsecas embrionarias observadas en las fases
tempranas por los efectos deletéreos del alcohol, ser producto de defectos en los mecanismos
maternos de control del número de embriones, o ambas opciones. Nuestros resultados

Discusión
292
concuerdan con otros trabajos publicados en los que se vio que la exposición a etanol tuvo un
efecto significativo en relación con las pérdidas embrionarias (Oyedele y Kramer, 2008).
Por otro lado, a diferencia de varios reportes donde se constata una reducción de la viabilidad
fetal (Gundogan y col., 2008; Oyedele y Kramer, 2008), en el presente trabajo, el etanol no
tuvo un efecto significativo en la mortalidad prenatal, dado que el número de fetos vivos por
hembra no varió significativamente entre los grupos controles y los tratados en ninguno de los
esquemas experimentales. La alta mortalidad fetal vista en dichos estudios, probablemente,
tenga estrecha relación con las dosis elevadas utilizadas.
Siendo la restricción del crecimiento intrauterino, uno de los principales efectos del consumo
gestacional de alcohol asociado al FAS, nos propusimos analizar el grado de crecimiento de los
fetos vivos, evaluando el peso y largo fetal. Los resultados no mostraron crecimiento fetal
retardado luego de la ingesta de etanol perigestacional. Más aún, mientras que el peso de los
fetos de los grupos tratados hasta los días 4 y 10 no varió respecto de sus controles, el peso
promedio de los fetos del grupo tratado hasta el día 8, resultó significativamente
incrementado respecto del control. El largo fetal no mostró variaciones en ninguno de los
grupos tratados. Este hallazgo es congruente con otros estudios en los cuales no han
reportado una relación entre el consumo de alcohol y la disminución de peso al nacer cuando
la exposición se lleva a cabo en los días 7, 8 y 9 de preñez (Oyedele y Kramer, 2008). Para
entender estos resultados debemos considerar algunas variables. El retraso del crecimiento,
como resultado de la exposición gestacional a etanol, ha sido demostrado en una serie de
estudios que emplearon una amplia variedad de cepas de ratón (Becker y Randall 1987, Becker
y col., 1996; Gundogan y col., 2008). La restricción del crecimiento neonatal ha sido asociada
con altos niveles de BACs (>100 mg/dl) (Samson, 1981; Gallo y Weinberg, 1986) pero sobre
todo, se relacionó con la exposición durante la última etapa de la preñez (días 14 a 19) la cual
parece ser más crítica para la inducción de esta anomalía (Wentzel y col., 2006; Gundogan y
col., 2008). Mientras que en varios estudios, el retraso del crecimiento fue evidente al
momento del nacimiento, en algunos otros, la disminución del peso corporal en las crías no se
hizo evidente sino hasta etapas postnatales avanzadas (Becker y col., 1996). Es decir, diversos
antecedentes demuestran que la exposición a etanol, principalmente, durante el período
postorganogénico sería la responsable de la restricción del crecimiento. Tras el período de
organogénesis, los sistemas de órganos formados entran en un período de crecimiento y
maduración funcional, siendo en esta etapa vulnerables a los efectos deletéreos del alcohol,
en relación a esas variables. En concordancia, nuestros resultados muestran que la exposición
a alcohol, no más allá del día 10 de gestación, indujo anomalías que no implicaron la

Discusión
293
disminución del tamaño y peso fetal. Como se mencionó anteriormente, otro factor
importante que ha sido asociado a la restricción del crecimiento intrauterino, es el estado
nutricional materno. Que el peso de los neonatos debe estar relacionado con el aumento del
peso de la madre, parece ser una suposición razonable y podría explicar la falta de diferencias
significativas entre los grupos de fetos de hembras tratadas y controles, lo que también fue
encontrado por Oyedele y Kramer (2008). Dichos autores observaron un patrón similar de
aumento de peso en ambos grupos de hembras (tratadas y no tratadas). De la misma manera,
Randall y Taylor (1979) no vieron diferencias significativas en el peso al nacimiento de crías
provenientes de ratones alimentados con etanol, donde las hembras tratadas no perdieron
peso sino que lo mantuvieron en un rango similar al de las controles. Esto mismo fue
constatado en la presente tesis, donde se descarta un posible factor de desnutrición materna,
asociado al consumo de alcohol, en el crecimiento fetal, ya que no se vieron diferencias
significativas en el peso corporal materno al inicio, en el último día de tratamiento (día 4, 8 o
10) y a término (día 18 de preñez) de los grupos de hembras tratadas. De la misma manera,
tampoco fueron diferentes los patrones diarios de ingesta calórica entre los grupos controles y
las tratados, en cada esquema experimental, al final del período de exposición. Por lo tanto, en
el presente trabajo, con un régimen crónico de exposición (previo a la gestación y hasta un
máximo de 10 días posteriores al inicio de la misma), al igual que Oyedele y Kramer (2008), con
una exposición corta (semicrónica, entre el día 6 a 8 de gestación), las madres tratadas con
etanol, no mostraron diferencias de pesos respecto de las controles a lo largo de la gestación.
La incidencia de malformaciones congénitas en la gestación a término fue analizada. Mientras
que en los grupos controles, la cantidad de fetos con malformaciones fue escasa
(aproximadamente del 12%), siendo éstas principalmente de tipo craneofacial (baja inserción
de oreja), en los tres grupos tratados con alcohol, una gran proporción de fetos presentó
malformaciones externas, como las típicas del FAS. En varios casos, un mismo feto resultó
portador de varias malformaciones en simultáneo, lo que fue observado fundamentalmente
en el grupo tratado hasta el día 10 de gestación. También en los grupos tratados, las
malformaciones encontradas con mayor frecuencia fueron las craneofaciales, las que
incluyeron labio leporino, baja inserción de oreja y acrania con exencefalia. También se vieron
fetos con defectos en los ojos (anoftalmia y exoftalmia), con fallas en el cierre de la pared
abdominal (onfalocele y gastroquistis), o alteraciones de las extremidades (miembros rotados
y dedos supernumerarios). Aunque menos frecuentes, se encontraron también fetos o masas
de tejido amorfos con sólo algunas pocas estructuras diferenciadas. Llamativamente, no sólo el
grupo que recibió etanol durante la gastrulación, así como, el grupo tratado hasta el día 10

Discusión
294
mostraron efectos teratogénicos, sino que además se vio incrementada significativamente la
frecuencia de fetos con anomalías cuando el alcohol se ingirió solamente hasta la etapa de
preimplantación. El hallazgo de anomalías morfológicas craneofaciales en fetos después del
consumo materno de etanol hasta el día 4 de preñez, resultó de sumo interés. Las
consecuencias teratogénicas de la exposición a etanol durante el período de preimplantación
no han recibido tanta atención, y en consecuencia son escasos los estudios. Esto posiblemente
aún se deba, en parte, a la creencia tradicional que suponía al embrión de mamíferos
refractario a estímulos adversos antes de la implantación. Sin embargo, el aumento de abortos
espontáneos luego de exposiciones durante el período de preimplantación, comenzó a
considerarse como un efecto deletéreo extremo de la exposición gestacional temprana a
etanol, ya que culmina en muerte embrionaria y en interrupción de la preñez (Becker y col.,
1996). Padmanabhan y Hameed (1988), investigaron los efectos sobre el feto de la
administración materna de dosis agudas de alcohol en estadios de preimplantación, aplicando
una dosis única de etanol 25% v/v (intraperitonealmente) en uno de los días 1 a 6 de la
gestación de ratones MF1. La exposición a etanol durante este período de preñez, produjo
malformaciones en el 80-100% de los fetos viables, siendo las mismas tanto craneofaciales
como sistémicas. Si bien, el porcentaje de fetos malformados encontrados fue mayor que los
vistos en nuestros resultados (posiblemente por la mayor dosis aplicada), dichos resultados
apoyan nuestros datos sobre la inducción de anomalías congénitas como consecuencia de una
exposición preimplantacional. El aumento significativo del número de fetos con anomalías
congénitas en el grupo alcoholizado hasta el día 4 de gestación respecto de su control, plantea
un verdadero desafío a la hora de hipotetizar sobre el origen de dichas alteraciones.
Probablemente, los efectos nocivos del etanol administrado a hembras murinas exocriadas,
repercuten en el MCI de los blastocistos, linaje del que derivan las células del propio feto. Esto,
en parte, fue demostrado en el presente trabajo, con la observación de la elevada tasa de
apoptosis, inducida por el etanol, en dicho estadio embrionario. Apoyados en el hecho que,
para una adecuada diferenciación embrionaria de postimplantación es necesario el desarrollo
de un cierto número umbral de células en el MCI, si el número de células del MCI de
blastocistos expuestos a etanol se reduce significativamente, pensamos que la diferenciación
de los embriones podría ser defectuosa. Por lo tanto, estas alteraciones del crecimiento
preimplantativo podrían contribuir al desarrollo de malformaciones y/o a muerte embrionaria.
Además, si ocurrieran mitosis aberrantes en las primeras divisiones embrionarias, podrían
generase células con complementos cromosómicos anormales que derivarían en anomalías
morfológicas en el embrión y que persistan en el feto (Musacchio y Hardwick, 2002).

Discusión
295
Las alteraciones craneofaciales más severas, como la acrania y la exencefalia, fueron
encontradas en los grupos tratados hasta el día 8 y día 10 de gestación. Por su parte, una alta
frecuencia de fetos de las hembras alcoholizadas hasta día 10 de preñez mostró un amplio
espectro de malformaciones aunque las craneofaciales tuvieron mayor prevalencia. Si bien,
por un lado, el etanol puede causar pérdida fetal de manera independiente del período de
administración, los efectos teratogénicos parecen ser estadio-dependiente. Así entonces, el
etanol administrado hasta el período de gastrulación (día 8) o hasta el de organogénesis (día
10), propició la aparición de una amplitud de malformaciones, con algunas diferencias, como
se indicó. Nuestros hallazgos demuestran así que los ratones exocriados expuestos a etanol
durante las fases tempranas de la gestación exhiben malformaciones craneofaciales como las
que se encuentran en los niños con FAS. Esto fue demostrado parcialmente en algunos
estudios con diferentes patrones de exposición a alcohol, coincidiendo en la mayoría de los
mismos en que las dosis testeadas fueron superiores a la utilizada en el presente trabajo. Por
lo tanto, es importante destacar que una concentración moderada, como etanol 10%, es
suficiente para recrear, en parte, el espectro de alteraciones asociadas al FAS. Oyedele y
Kramer (2008), con un régimen semicrónico de etanol al 25% (v/v) aplicado a ratones C57BL/6J
durante los días 7, 8 y 9 de gestación (período de formación de surco primitivo, pliegues
neurales y tubo neural), detectaron anoftalmia y agnatia en los grupos tratados. De la misma
manera, ratas tratadas con etanol 20% en el agua de bebida bajo un régimen crónico a lo largo
de la gestación, presentaron un aumento en la proporción de fetos malformados. Entre las
malformaciones encontradas se destacaron grandes hematomas, exencefalia y onfalocele
(Wentzel y col., 2006). Adicionalmente, los reconocidos trabajos de (Sulik y col., 1988; Dunty y
col., 2002), demostraron que los ratones expuestos a una dosis aguda de alcohol en el día 7 y 8
de gestación presentaron anomalías faciales al día 14 de gestación, análogas a las observadas
en el FAS.
Para entender los mecanismos de inducción de las alteraciones observadas, debemos
considerar que la morfogénesis craneofacial normal se desarrolla bajo la influencia de
múltiples interacciones entre tejidos embrionarios, tales como las CCN craneanas, el
mesodermo y el ectodermo, lo que requiere una regulación precisa de los movimientos
celulares, del crecimiento y del patrón de diferenciación de los tejidos involucrados (Sant'Anna
y Tosello, 2006). La exposición a alcohol durante el período de gastrulación de embriones de
ratón, reduce la tasa de división celular (Sulik, 1984) y el proceso de migración de las células
del mesodermo hacia la línea primitiva (Nakatsuji y Johnsson, 1984). Como se discutió
anteriormente, se ha demostrado que uno de los efectos deletéreos del alcohol en etapas

Discusión
296
específicas del desarrollo temprano es el desencadenamiento excesivo de apoptosis en células
destinadas a dar origen a estructuras craneofaciales, como las CCN craneana en períodos bien
definidos de vulnerabilidad, como lo es la gastrulación y la neurulación (Sulik, 1984; Sulik y col.,
1986; Sulik y col., 1988; Chen y Sulik, 1996; Cartwright y col., 1998).
La gastrulación y la organogénesis en el ratón abarcan desde el día 7 al 14 de gestación (que
corresponde a la 3ra y a la 8va semanas del embarazo humano). En la mayoría de los estudios,
la administración de etanol durante estos períodos de preñez resulta en un aumento, dosis-
dependiente, de la mortalidad prenatal (reabsorciones), de la incidencia de malformaciones de
órganos y del esqueleto. Dentro de estos períodos de desarrollo embrionario, la secuencia
temporal de acontecimientos del desarrollo, es única para cada sistema de órganos. Por lo
tanto, aunque la exposición a etanol durante estas fases resulta en el desarrollo de una
variedad de anomalías morfológicas, incluyendo craneofaciales, cerebrales, cardíacas,
urogenitales y defectos esqueléticos, el perfil específico de anomalías dependerá del momento
de la inducción del daño por el etanol (Becker y col., 1996). Así, la exposición aguda a etanol de
2,9 g/kg en ratones C57BW6J durante los días día 8 o 9 de gestación, produjo un aumento de
la mortalidad prenatal y una serie de anomalías craneofaciales (anoftalmia, microftalmia,
fisuras palpebrales cortas, hipoplasia de maxilar y mandibular, nariz pequeña, labio superior
fino, surco nasolabial liso, labio leporino y baja inserción de orejas) y del cerebro (microcefalia,
exencefalia). La exposición a etanol en el día 9 de gestación también originó malformaciones
cardíacas, mientras que la administración aguda al día 10 resultó en alteraciones urogenitales
y anomalías de las extremidades (polidactilia, sindactilia y adactilia) (Becker y col., 1996). Los
estudios dirigidos por Sulik (1988, 1992, 2005) muestran en un modelo de FAS en ratón, que
los rasgos craneofaciales similares a los de los seres humanos afectados, se producen con el
tratamiento materno agudo con una alta dosis de etanol en los días 7, 8 o 9 de gestación. Estas
alteraciones características del FAS, mostradas ampliamente en la literatura, se deben como
fue mencionado a defectos de las estructuras derivadas de las CCN craneana luego de la
exposición a alcohol (Sulik y col., 1988; Cartwright y col., 1998).
De manera similar a lo observado en los modelos de exposición aguda, los estudios de
exposición prenatal crónica a etanol en el período preorganogénico y organogénico, han
demostrado una gran variedad de defectos congénitos típicos del FAS, que en general, resultan
de la sumatoria de varios días de exposición (Becker y col., 1996). A diferencia de otros
órganos, el cerebro parece ser sensible a las acciones teratogénicas del etanol a lo largo de
todo su desarrollo ya que es uno de los primeros órganos en desarrollarse y el último en
madurar por completo, proceso que abarca todo el período gestacional y se extiende hasta la

Discusión
297
vida postnatal temprana, en roedores (West y Goodlett, 1990). Por lo tanto, las anomalías
cerebrales, en los modelos de ratón, han sido observadas después de la exposición a etanol
durante la mitad de la preñez, al final y durante toda la gestación. Estas anomalías incluyeron
exencefalia, hidrocefalia, microcefalia y defectos estructurales de diversas regiones cerebrales
(Becker y col., 1996).
Por otro lado, al igual que lo reportado por Wentzel y colaboradores (2006), cuando
analizamos de manera cualitativa la posición intrauterina de los fetos dismórficos y/o
reabsorciones en los grupos expuestos al etanol, encontramos que aquellos fetos con
malformaciones más severas, así como las reabsorciones, se hallaron principalmente en los
extremos de los cuernos uterinos. El hallazgo de esta distribución desigual de fetos anómalos y
de reabsorciones en los cuernos uterinos sugiere un gradiente intrauterino de nutrientes y de
etanol que influye en el riesgo de desarrollar embriopatía alcohólica. Dada esta observación,
según lo sugerido (Wentzel y col., 2006), con la exposición a altos niveles de etanol,
aumentaría el riesgo que los fetos posicionados más internamente resulten también afectados.
Menos estudiadas que las malformaciones congénitas, pero no por ello menos importantes,
son las alteraciones en el esqueleto fetal inducidas por el alcohol. Cada vez más evidencias
sugieren que el ambiente intrauterino puede influir en la salud ósea a largo plazo y en el riesgo
de desarrollar osteoporosis en la vida posterior. Si bien, el nivel de exposición a etanol
necesario para producir defectos esqueléticos es desconocido, se ha documentado que la
exposición intrauterina a alcohol genera alteraciones del desarrollo óseo fetal (Mishra y col.,
2003; Simpson y col., 2005; Snow y Keiver, 2007).
En virtud de estos antecedentes nos propusimos estudiar si la exposición prenatal a etanol
tiene efectos específicos en el desarrollo del esqueleto fetal. Evaluamos el número, tamaño,
forma y posición de los principales huesos del cráneo, rostro, tórax, pelvis, columna y
miembros, en los tres grupos expuestos a alcohol perigestacionalmente hasta el día 4, 8 o 10
de gestación. Contrariamente a lo observado en los controles, en los fetos provenientes de los
tres grupos experimentales expuestos a etanol, las alteraciones óseas fueron evidentes.
Fueron frecuentes fetos con densidad ósea disminuida en forma generalizada y con bajo grado
de osificación, ausencia de algunas piezas óseas, tales como, costillas y falanges y alineamiento
irregular de los centros esternales. Las alteraciones esqueléticas más significativas encontradas
en los tres grupos expuestos a etanol se observaron a nivel craneofacial, las cuales incluyeron
la ausencia de huesos de la bóveda craneana, baja densidad ósea y osificación incompleta.
Cerca del 90% de los fetos de madres tratadas con alcohol, independientemente del período

Discusión
298
de exposición, presentaron alguna alteración ósea craneofacial, porcentaje que fue
significativamente menor en las hembras controles. La cantidad de fetos con huesos craneanos
con baja densidad ósea o con osificación incompleta, o combinación de ambas alteraciones, no
resultó significativamente diferente entre los grupos tratados y con sus respectivos controles.
Sin embargo, la valoración relativa de la densidad ósea sí mostró diferencias estadísticamente
distintas en todos los grupos tratados respecto de sus controles. Mientras que para los
controles la intensidad de la coloración con alizarina, resultó en un valor relativo promedio
cercano a 3 (establecido arbitrariamente como máximo) para los fetos de los grupos tratados
dichos valor resultó entre 1,8 y 2, significativamente diferentes de sus controles. Por lo tanto,
este hallazgo podría sugerir que el etanol interfirió con procesos de crecimiento y expansión
del tejido óseo. Remarcando que estos efectos deletéreos sobre los huesos fetales se vieron
con la dosis de etanol 10%, considerada baja o moderada por la alcoholemia que produce,
consideramos que este régimen de alcoholización no fue suficiente para inducir cambios óseos
que afecten el peso y largo fetal al momento del nacimiento. Así, debido a la linealidad que
existe entre el tamaño corporal (peso y longitud) y la masa ósea, es difícil disociar un efecto
general en el tamaño corporal de un efecto específico en el desarrollo del esqueleto.
Actualmente no se conoce con certeza si el efecto en el esqueleto fetal, como resultado de la
exposición prenatal a etanol, es consecuencia de un retraso de crecimiento generalizado o
consecuencia de un efecto específico en el proceso de desarrollo óseo. Para discernir entre
ambas posibilidades debemos considerar que si el efecto del etanol en el desarrollo del
esqueleto fuera debido a un efecto general en el crecimiento, entonces la exposición a etanol
resultaría en un retraso uniforme en todos los parámetros asociados al crecimiento y muchos
de los huesos deberían mostrar un patrón similar de retraso. Alternativamente, si el etanol
posee efectos específicos sobre el desarrollo del esqueleto, entonces un retraso o alteración
diferencial podría surgir en los parámetros medidos. Nuestras evidencias sugieren que la
exposición prenatal a alcohol, hasta períodos tempranos de la gestación, produce efectos
adversos sobre el desarrollo del esqueleto fetal, independientes de los efectos en el
crecimiento general de los fetos. Así, la osificación fetal pareciera ser más sensible a los
efectos del etanol que el peso y largo corporal del feto. Inclusive se detectó una sensibilidad
diferencial según el hueso (Simpson y col., 2005). Por lo expuesto, los reportes son
coincidentes con nuestros resultados, ya que a pesar de no registrar efectos a nivel de
crecimiento global de los fetos expuestos, sí detectamos anomalías en el desarrollo óseo.
Simpson y colaboradores (2005), estudiaron en ratas los efectos de tres niveles de exposición
distintos de etanol, que se correspondieron con niveles bajos, moderados y altos a etanol

Discusión
299
(dietas líquidas con 15%, 25% o 36% de EDC), durante 6 semanas (3 antes del apareo y durante
21 días de gestación). La dosis más alta (36% EDC) generó una disminución del peso y largo
fetal y en la osificación esquelética. Sin embargo, efectos deletéreos en la osificación, pero no
en el peso corporal o en la longitud, fueron observados con la dosis moderada (25% EDC). Es
decir, se producen efectos nocivos a nivel óseo, inclusive, ante el consumo materno moderado
de etanol y en ausencia de alteración generalizada del crecimiento. Este hallazgo sugirió que
los defectos en la osificación del esqueleto fetal ocurrieron con niveles de etanol prenatal
menores a los requeridos para afectar el peso y largo fetal. Por lo tanto, y en consonancia con
nuestros resultados, los efectos del etanol en el desarrollo no se limitan a reflejar efectos en el
crecimiento general, sino que reflejan en un efecto específico. Más evidencias al respecto,
muestra que el número de huesos afectados y la gravedad de los defectos sobre la osificación
tendieron a aumentar con la dosis de etanol administrada (Simpson y col., 2005). Es
interesante resaltar que estos resultados sugieren una respuesta diferencial en el tamaño
corporal (peso y largo) y en la osificación del esqueleto con los diferentes niveles de exposición
a etanol. Otros resultados publicados, congruentes con los nuestros, también demostraron
que el consumo de alcohol durante la preñez afecta negativamente el desarrollo del esqueleto
fetal. Oyedele y Kramer (2008), también demostraron que fetos de hembras murinas C57BL/6J
expuestas a etanol 25% (v/v) mediante gavage esofágico en los días 7, 8 y 9 de gestación, que
sin presentar anomalías obvias a término, tuvieron defectos craneofaciales, tales como
ausencia de ciertos hueso (por ejemplo: premaxilar y mandíbula) y cierta alteración en el grado
de osificación.
Aún no son conocidas las razones de la susceptibilidad diferencial de los distintos huesos a las
acciones negativas del alcohol. Se ha sugerido que la sensibilidad al etanol sería mayor en
huesos que empiezan la osificación tempranamente en la gestación (Simpson y col., 2005).
Estudios in-vitro demostraron que el etanol inhibe la diferenciación de los osteoblastos, su
proliferación y función (Farley y col., 1985; Chavassieux y col., 1993; Klein, 1997; Gong y
Wezeman, 2004). Así, el etanol podría tener un efecto directo sobre las células involucradas en
la formación de hueso. Alternativamente, el etanol ejercería un efecto indirecto a través de
perturbaciones en la fisiología materna y fetal. Según diversos estudios, el consumo materno
de alcohol resulta en una hipocalcemia materna y fetal, y la severidad de la hipocalcemia varia
con el BAC (Keiver y col., 1996; Keiver y Weinberg, 2004). Esta disminución de calcio en sangre,
materna y fetal, está asociada con los efectos adversos en el desarrollo del esqueleto, incluida
una reducida mineralización (Chalon y Garel, 1985). Por lo tanto, una homeostasis alterada en
los niveles de calcio podría ser uno de los mecanismos por el cual el etanol afecta el desarrollo

Discusión
300
óseo fetal. Nuevamente, los efectos deletéreos del alcohol observados en el grupo
alcoholizado hasta el día 4 de gestación representan un desafío a la hora de justificar el origen
de los defectos óseos. Probablemente, la suma de las alteraciones inducidas en el MCI junto
con alteraciones fisiológicas de la madre, como consecuencia del consumo de alcohol,
contribuyan a la aparición de las alteraciones esqueléticas en este grupo.
Las alteraciones fetales, además de ser inducidas como efectos específico del alcohol y/o del
acetaldehído, puede ser generada indirectamente por lesión placentaria, lo que contribuye a
su mal funcionamiento y posible malnutrición fetal (Bosco, 2005; Bosco y Diaz, 2012).
El paso de los nutrientes de la sangre materna al feto está mediado por la placenta, un órgano
que establece conexiones funcionales críticas para la supervivencia fetal (Cross y col., 1994). La
placenta transporta oxígeno, nutrientes y anticuerpos para el feto, y elimina el dióxido de
carbono y desechos metabólicos; sirve como una barrera protectora contra los efectos
perjudiciales producidos por ciertos fármacos y microorganismos; actúa como una barrera
parcial entre la madre y el feto; y produce hormonas esenciales para el mantenimiento de la
preñez (Bosco, 1995, 2005). Es crítico entonces, para un metabolismo y crecimiento normal del
feto, que tenga lugar un adecuado intercambio a través de la placenta. Dado que el desarrollo
de la placenta es un proceso altamente regulado, este órgano presenta una elevada
susceptibilidad frente a agentes externos, los que pueden interferir con las funciones de la
placenta: la función inmunológica, la producción y liberación de hormonas y enzimas, el
transporte de nutrientes y productos de desecho, el crecimiento y maduración celular (Bosco y
Diaz, 2012).
Los efectos de la exposición a alcohol en la placentación, el crecimiento y función de la
placenta fueron revisados por Burd y colaboradores (2007). La exposición prenatal a alcohol
durante períodos variables de la preñez o durante la misma, originan una amplia gama de
efectos adversos tales como disfunción placentaria, disminución o aumento del tamaño de la
placenta, alteración del flujo sanguíneo y del transporte de nutrientes, cambios endocrinos,
restricción del crecimiento intrauterino, aumento de la tasa de muerte fetal y desprendimiento
de placenta. Una inadecuada función placentaria compromete el desarrollo embrionario y
fetal, e inclusive podría tener un impacto negativo en la vida postnatal (Barker, 2004). Sin
embargo, no se ha determinado si la exposición al alcohol de manera perigestacional y sólo
hasta las primeras etapas del desarrollo conduce a una placentación anormal y, en
consecuencia, a defectos en la placenta a término. En el presente trabajo, se analizaron los
efectos en el desarrollo general de la placenta luego que hembras murinas CD-1 fueron

Discusión
301
expuestas a etanol 10%, 17 días previos a la gestación y sólo hasta los tres períodos críticos del
desarrollo: preimplantación (día 4), postimplantación temprana-gastrulación (día 8) o
postimplantación media-organogénesis (día 10). Cumplidos dichos períodos, la exposición a
etanol cesó y continuaron recibiendo agua hasta la gestación a término (día 18).
En todos los grupos tratados con etanol fue significativa la reducción del peso placentario,
siendo más pronunciado en el grupo que recibió alcohol hasta el día 10 de gestación. Esta
disminución en el peso de la placenta, provocó una alteración en los índices de peso
feto/placentario, el cual resultó incrementado significativamente en todos los grupos tratados
respecto de los controles, sugiriendo un efecto especifico del etanol en el crecimiento de la
placenta. Una importante consecuencia de ello, es que, aunque al día 18 no se detectaron
cambios en el peso fetal de las hembras tratadas, el impacto negativo de la exposición en el
desarrollo de la placenta se ponga de manifiesto en la etapa postnatal, como fue sugerido por
algunos autores (Barker, 2004). Por otro lado, en particular, la relación entre la exposición a
etanol durante el período de preimplantación y el crecimiento de la placenta ha recibido
relativamente poca atención. En un estudio realizado por Padmanabhan y Hameed (1988), el
efecto del etanol sobre el peso de la placenta resultó ser altamente variable. La administración
de etanol 25%, ya sea al día 1,5 o 2,5 de gestación, resultó en un aumento del crecimiento de
la placenta al día 15. Sin embargo, las placentas recolectadas al día 18,5 no se vieron afectadas
después de la administración de etanol al 25% al día 2,5. Cuando el momento de la
administración fue el día 6,5 de gestación, los pesos placentarios registrados al día 18,5, se
redujeron en comparación con los controles (Padmanabhan y Hameed, 1988). Por lo tanto, los
efectos del etanol en relación al crecimiento de la placenta, ya sea que se administre durante
los períodos de preimplantación o postimplantación, parecen depender de la dosis y del
tiempo de exposición.
El análisis de la estructura placentaria demostró que la exposición perigestacional a etanol
afecta la morfogénesis de la placenta. En todos los grupos tratados, fue evidente el incremento
del área de la zona de unión en detrimento de la zona laberíntica, mientras que el área
placentaria total resultó inalterada respecto de los controles. Este hallazgo sugiere un cambio
en el balance entre la zona de unión (células gigantes trofoblásticas, CGTs; células
espongiotrofoblásticas, EPGs; células glicogénicas, CGs;) y el laberinto (zona vascular). Una
disminución en esta región, el laberinto, dará como resultado intercambios metabólicos
deficientes entre la madre y el feto, pudiendo esta situación ser inductora de restricción del
crecimiento fetal. Sin embargo, en ninguno de los grupos tratados el crecimiento fetal
intrauterino resulto disminuido.

Discusión
302
Las placentas de todos los grupos de hembras alcoholizadas mostraron alteraciones
histopatológicas tanto en la zona de unión como en la laberíntica. En la zona de unión se
observaron grandes lagunas de sangre y deslocalización de las CGTs, las cuales se alejaron de
su posición típicamente lindante con la decidua y se situaron más internamente, en la zona de
unión. También fueron observadas alteraciones en la morfología de las células EPGs, lo que
podría implicar alteraciones de su funcionalidad (síntesis y secreción de factores). Sin embargo,
el hallazgo más significativo fue el aumento de la cantidad de CGs, a lo largo de toda la zona de
unión, manifestado por la presencia de gran cantidad de células PAS-positivas. Por otro lado, el
laberinto, siendo un tejido especializado en el intercambio de nutrientes y gases entre la
madre y el feto, en las placentas de las hembras controles, se observó como una red de
espacios sanguíneos maternos amplios y distendidos, separados de los capilares fetales, por
una delgada estructura trilaminar de células trofoblásticas, conformada por dos capas de
sinciciotrofoblasto, que están asociadas al endotelio de los vasos fetales, y una capa de células
trofoblásticas gigantes mononucleares o citotrofoblastos sinusioidales, que delimitan los
espacios sinusoides maternos. Así, mientras que en las muestras controles se observó el
delgado límite entre los sinusoides maternos y los capilares fetales, en las muestras
provenientes de hembras tratadas se evidenciaron cambios significativos en la integridad y
organización de este tejido. Se observó un tejido más compacto con evidente engrosamiento
de la capa trofoblástica que separa los espacios sanguíneos, y por consiguiente, se apreció una
reducción de los espacios sanguíneos de intercambio. También se encontraron células PAS-
positivas, EPGs en la capa laberíntica y leucocitos en los espacios sanguíneos maternos
(principalmente al día 10 de gestación). Estos defectos placentarios a largo plazo, en el día 18
de gestación, estarían relacionados con los efectos adversos del etanol en las primeras etapas
de la placentación, siendo aún susceptibles las células embrionarias que darán origen a la
placenta, en el período de preimplantación.
La formación de la placenta hemocorial, depende de la normal actividad invasiva del
trofoblasto, desde el período implantativo, cuando se establecen íntimas interacciones
perfectamente sincronizadas entre el trofoblasto y la mucosa uterina. Que se alcance tal grado
de intimidad depende de una gran variedad de factores, entre los que se destaca la actividad
invasiva del trofoblasto (Finn y Martin, 1967). Luego de la implantación, ya en el estroma
endometrial, la invasión trofoblástica prosigue con la ruptura de los vasos endometriales con
el consecuente extravasamento de sangre materna (Bevilacqua y Abrahamsohn, 1989), lo que
permite el intercambio entre el trofoblasto y la sangre materna, condición que persiste
durante todo el desarrollo. Cuando la sangre materna circula directamente contra la superficie

Discusión
303
del trofoblasto, antes de la formación de la placenta corioalantoidea, el saco vitelino
desempeña un papel fundamental en la absorción y transporte de nutrientes de la madre al
embrión en desarrollo (Cross y col., 1994). Así, la nutrición histotrófica parece ser
especialmente importante en esta etapa. En el período organogénico, en ausencia de órganos
internos totalmente desarrollados, los vasos vitelinos pueden transportar los nutrientes al
embrión y eliminar los residuos de los mismos de manera más eficiente. El desarrollo de la
circulación vitelina permite que el embrión pase de la dependencia de la entrega de nutrientes
por difusión a un sistema más eficiente de conductos vasculares (Jollie, 1990). Nogales y
colaboradores (1993), observaron una asociación entre la incidencia de abortos espontáneos y
una reducción en el tamaño o la ausencia completa del saco vitelino, lo que sugiere una
relación entre las anomalías del saco vitelino y el desarrollo embrionario. Xu y colaboradores
(2005), demostraron que la exposición de embriones a etanol (1, 2 y 4 mg/ml) in-vitro por 48
hs a partir del día 9,5 de gestación (organogénesis) produjo efectos nocivos, concentración
dependiente, en la morfología y funcionalidad del saco vitelino. En este estudio, el desarrollo
de los vasos vitelinos fue notablemente afectado por el etanol, observándose una menor
cantidad, más delgados y menos ramificados respecto del control, mientras que una alta
concentración de alcohol inhibió por completo la formación vascular en este tejido. En
condiciones normales, las células endodérmicas del saco vitelino adquieren nutrientes por
pinocitosis, y los transfieren en forma de vesículas de almacenamiento. Después de la
exposición a etanol también se describieron alteraciones morfológicas de estas células (Xu y
col., 2005), mostrándose anomalías ultraestructurales, como una reducción de las
microvellosidades, disminución en la cantidad de invaginaciones pinocíticas y de vesículas de
almacenamiento, lo que sugirió una actividad pinocítica reducida o suprimida. Estas
alteraciones morfológicas y de la función de nutrición histotrófica, probablemente, fueron las
que condujeron a crecimiento retardado y malformaciones del embrión expuesto, que
correspondieron principalmente a fallas en el cierre del tubo neural y anormalidades del
sistema cardiovascular embrionario.
El desarrollo de la placenta murina ha sido descripto en detalle por diversos autores (Soares y
col., 1996; Rossant y Cross, 2001; Georgiades y col., 2002; Malassiné y col., 2003). Brevemente,
la placenta corioalantoidea madura de ratón comienza a establecerse a partir del día 10,5-11
de gestación, y hacia el día 12,5 de gestación la placenta definitiva está claramente subdividida
en dos compartimentos, uno materno y otro fetal. El compartimiento materno,
correspondiente a la decidua, está compuesto por células deciduales, endoteliales y

Discusión
304
trofoblasto que ha migrado de la placenta fetal. El compartimiento fetal está compuesto por
dos zonas morfológica y funcionalmente distintivas: (i) la zona de unión, compuesta por una
capa externa y distal al feto de CGTs lindantes con la decidua, y por una capa media de células
EPGs y CGs; y (ii) la zona laberíntica (interna y proximal al feto) compuesta por trofoblasto,
vasos fetales y sangre materna.
La exposición a etanol en el momento del establecimiento de la placenta madura o en días
previos, podía producir daño a largo plazo, hipótesis que podía estar relacionada con efectos
negativos sobre la tasa de invasividad del trofoblasto, insuficiente remodelación arterial,
alteraciones en los factores reguladores de la placentación, entre otros. Los resultados
obtenidos de los estudios al día 4, 8 y 10 de gestación, luego de la exposición materna alcohol,
en relación con las anomalías del desarrollo trofoblástico, convalidan la hipótesis planteada.
Si bien, muchos estudios han utilizado con frecuencia sólo el peso y/o el volumen de la
placenta como un indicador del desarrollo de dicho órgano, ellos sólo proporcionan una
estimación del desarrollo de la placenta, pero no permiten discriminar diferencias en el
crecimiento de los compartimentos individuales que la componen. Por lo tanto, no pueden
reflejar con precisión la capacidad funcional del órgano. Las diferencias en el crecimiento de
los compartimentos individuales de la placenta pueden ser analizadas aplicando metodologías
histológicas. Un reflejo de esto es el hecho que mientras que el peso y volumen total de la
placenta alcanzan un máximo en el estadio E16,5, la ampliación del volumen del laberinto es
muy significativa más allá de ese estadio (siendo activa entre E12,5 y 18,5). Además, durante el
desarrollo de la placenta hasta su crecimiento a término, existen importantes cambios en el
número y en la morfología de las células que conforman cada capa, cambios que conllevan
importantes implicancias a nivel funcional. Inicialmente, en el estadio E12,5, las CGTs aparecen
como una capa continua lindante con la decidua, pero esta se hace discontinua más tarde en el
desarrollo (Cross y col., 1994). Estas células continúan representando el componente fetal más
distal de la placenta del ratón a lo largo de toda la preñez. Con respecto a la zona de unión,
Coan y colaboradores (2004), demostraron que el volumen de la misma a lo largo de la
gestación presenta un comportamiento bifásico, expandiéndose notablemente hasta E16,5, y
luego reduciéndose en E18,5, siendo las alteraciones en el tamaño celular, la proliferación y la
migración, los principales responsables de estos cambios dinámicos en la zona de unión de la
placenta. Mientras el volumen de las células EPGs se duplica y el número de las mismas
aumenta de manera constante desde E12,5 a E16,5, un evento proliferativo dramático en ese
mismo período produce un aumento explosivo de aproximadamente 80 veces en el número de
GCs, con un pico en E16,5. Este hallazgo sugiere que el principal contribuyente a la expansión

Discusión
305
de la zona de unión serían las CGs, que junto con el aumento en el número y tamaño celular
del EPG en E16,5, marcan el punto máximo de tamaño celular y número de células en la zona
de unión durante la gestación. El aumento de las CGs en este estadio, es seguido de una
disminución significativa de estas células hasta el día E18,5 ya que una parte de estas células
abandonan la zona de unión e invaden la decidua. La disminución de CGs al día 18, no sólo
tiene que ver con la migración de las células a la decidua sino con la pérdida de CGs por otro
mecanismo (Coan y col., 2004, 2006). Así, Waddell y colaboradores (2000) sugirieron que la
muerte de CGs por apoptosis también podría desempeñar un papel en la reducción del
volumen de la zona de unión. Las evidencias morfométricas e inmunohistoquímicas, indican
que, luego que la zona de unión ha alcanzado su nivel máximo de expansión en E16,5, la
proliferación celular en dicha zona cesaría, en contraste con la zona del laberinto, que continúa
proliferando, creciendo y expandiéndose hasta el desarrollo a término (Coan y col., 2004).
En cuanto a la estructura, tanto las células EPGs como las CGs se agrupan, en la placenta a
término, conformando grupos discretos o formaciones de tipo “cordones” de células, que
abarcan la zona de unión desde el laberinto hasta las CGTs (próximas a la decidua). Muy
ocasionalmente se han descripto CGs profundamente dentro de la zona laberíntica, aunque la
importancia funcional de esta localización aún no se ha determinado (Coan y col., 2006). En
nuestro modelo, las placentas de las hembras tratadas presentaron alteraciones en la
organización y distribución celular dentro de la ZU, y en la interfase de ésta con la decidua, en
el lado mesometrial, las CGTs se observaron entre las CGs, perdiendo su característica
localización periférica limitante. Uno de los resultados más llamativos es la evidencia de un
aumento en la cantidad de CGs PAS-positivas en la ZU respecto de las EPGs. Las EPGs sufrieron
cambios morfológicos (células ahusadas), lo que fue más predominante en el grupo tratado
hasta el día 10 de gestación. Estos cambios morfológicos celulares podrían indican anomalías
funcionales relacionadas con la producción hormonal y demás factores vinculados con la
regulación de la función placentaria (Coan y col., 2005). Particularmente, en el caso de las CGs,
aparte de su comportamiento transitorio y de la capacidad de acumular glucógeno, no se
conoce mucho más acerca de su función. Así, varios son los interrogantes en torno a estas
células, entre ellos, ¿Qué proporción de la composición celular de la zona de unión consiste en
CGs durante la gestación?, ¿realmente puede el tamaño y la migración de las CGs explicar los
cambios en la estructura y el volumen de la zona de unión en la gestación tardía? Además de
acumular glucógeno, las CGs han sido asociadas con la expresión de glucagon en la zona de
unión (Coan y col., 2006). En términos generales, el glucagon se une a receptores en células
que almacenan glucógeno tales como hepatocitos, estimulando la producción de glucosa a

Discusión
306
través de la glucogenolisis del glucógeneo almacenado (Lodish y col., 1998). Coan y col (2006)
sugirieron que el glucagon presente en las CGs en la fase final de la gestación sería clave
cuando un suministro de energía adicional podría ser requerido por el feto, la placenta o la
madre para completar con éxito la gestación. La inmunomarcación positiva para glucagon en la
gestación tardía, que podría atribuirse exclusivamente a las GCs, y por lo tanto, dado que hacia
el final de la preñez aumenta la demanda del feto sobre los recursos maternos, la reserva de
glucógeno de las CGs sería convertida en glucosa, liberándose directamente a la sangre
materna (Coan y col., 2006). La glucosa circularía al feto a través del laberinto. Apoyando esta
idea, López y colaboradores (1996) demostraron bioquímicamente que el glucógeno
placentario total disminuyó entre E15 y E18 en la placenta normal.
Durante la embriogénesis, la formación temprana de un sistema vascular funcional altamente
integrado en la zona más cercana al feto, es fundamental. Así, la capa laberíntica es quizás el
sitio de interacción materno-fetal más importante de la placenta, ya que contiene sangre fetal
y sangre materna circulando independientemente, pero en estrecha asociación. La sangre
materna (que fluye por espacios irregulares o sinusoides maternos), está separada de la
sangre fetal (que circula dentro de los capilares fetales) por una capa de células trofoblásticas,
que algunos autores han denominado “membrana interhemal” la que, en ratones, comprende
tres capas de células trofoblásticas (Georgiades y col., 2002; Coan y col., 2004). Desde el
estadio E10,5 hasta el desarrollo a término, la placenta sufre un gran crecimiento y expansión
de la capa laberíntica. En parte, el éxito de la placentación depende de la remodelación de los
vasos maternos dentro del laberinto, la cual ocurre por dos rutas. Una ruta “intersticial” donde
los vasos maternos pierden su revestimiento de células endoteliales, después de haber sido
erosionados por el trofoblasto fetal invasivo formando los espacios de sangre materna. Otra,
involucra al trofoblasto endovascular que migra dentro de las arterias uterinas espiraladas
provocando su remodelación dado que reemplazan el endotelio vascular (Pijnenborg y col.,
1991). Si bien, el número de arterias espiraladas no cambia, el diámetro se incrementa
notablemente entre E10,5 y E14,5. El reemplazo de la capa muscular de las dichas arterias por
parte del trofoblasto endovascular conduce a un alto flujo de sangre y a una circulación de
baja resistencia en el espacio laberíntico (Adamson y col., 2002). A la remodelación de las
arterias uterinas también contribuyen las CGs, a través de la expresión de MMP-9 y por ende
degradación de la matriz extracelular que rodea las arterias maternas facilitando así su
dilatación (Adamson y col., 2002). Como mencionamos anteriormente, si bien, el volumen
máximo de la placenta se alcanza en el estadio E16,5, la proporción de tejido laberíntico sigue

Discusión
307
aumentando, aparentemente, a expensas de la zona de unión y de la decidua. La expansión del
laberinto se asocia a un aumento en el volumen del trofoblasto y en el volumen ocupado por
los capilares fetales y los espacios sanguíneos maternos (Coan y col., 2004). Dentro de la zona
del laberinto, el volumen y el área superficial total de los sinusoides maternos se expande
rápidamente entre E14,5 y E16,5, precedido y sucedido por períodos relativamente
quiescentes de expansión (no hay diferencia en volumen y superficie entre E12,5 y E14,5 ni
entre E16,5 y E18,5). Así, el volumen de sangre materna que puede circular a través de la
placenta, alcanza su máximo en E16,5. En contraste, la dinámica del desarrollo de los capilares
fetales en el laberinto es diferente a la de los sinusoides maternos. La expansión de los
capilares fetales en el laberinto es continua en el tiempo, desde E12,5 hasta 18,5, lo que
sugiere una continua necesidad de mayores volúmenes de sangre fetal para intercambiar con
la circulación materna y así obtener suficiente nutrición y oxígeno para el crecimiento fetal
(Malassiné y col., 2003; Coan y col., 2004). El desarrollo de la membrana interhemal, que
separa las circulaciones materna y fetal en el laberinto, aumenta a medida que se desarrolla la
placenta, particularmente entre E12,5 y E16,5, donde este incremento es significativo. A partir
de E16,5, cuando el volumen total de placenta deja de ampliarse, el volumen de la membrana
también permanece inalterado. Sin embargo, previo a la expansión en volumen, la membrana
interhemal sufre un significativo adelgazamiento. Uno de los parámetros más importante para
la fisiología de la placenta es el espesor de dicha membrana. Así, la presencia de zonas
delgadas en la barrera es de importancia clave para la difusión efectiva de los solutos. Entre
E12,5 y E16,5, ocurren reducciones significativas del espesor. Ya en E16,5, se alcanza el espesor
final, de aproximadamente 4,4 µm, y no se producen más reducciones (Coan y col., 2004). Este
proceso en el desarrollo de la placenta determina la formación de una barrera fina con gran
superficie que minimiza la distancia de difusión y maximiza el área de intercambio. Por lo
tanto, alcanzar el espesor promedio “armónico” es uno de los determinantes más críticos de la
capacidad de difusión de la placenta. Ligeros descensos en el espesor de la membrana pueden
tener un efecto positivo (Mayhew y col., 1986), pero, por el contrario, si la membrana
interhemal se mantiene gruesa, esto podría perturbar la difusión de sustancias al feto (Coan y
col., 2004). En las placentas de todos los grupos de hembras tratadas, encontramos que la
zona del laberinto tuvo importantes cambios en la integridad y organización del tejido. La capa
trofoblástica que separa los espacios sanguíneos (membrana interhemal) presentó un espesor
engrosado, junto con una mayor compactación del tejido laberíntico en comparación con las
placentas controles, lo que llevó a una reducción de los espacios de intercambio sanguíneo.

Discusión
308
El crecimiento del feto depende del intercambio transplacentario de gases respiratorios,
nutrientes y desechos entre los sistemas circulatorios materno y fetal del laberinto, por lo cual
el deterioro del desarrollo vascular placentario o la remodelación arterial deficiente pudo
conducir a mortalidad fetal temprana (Gundogan y col., 2008, 2010). Sin embargo, nosotros no
encontramos mortalidad fetal elevada en el día 18 de gestación, en las hembras expuestas a
alcohol hasta los días 4, 8 o 10 de gestación. En diversos modelos experimentales se ha
probado el deterioro de la placentación asociado a la exposición crónica a etanol. La isquemia
o infarto observado en estas placentas expuestas reduce el espesor del órgano en su conjunto
por aumento de la necrosis celular, lo que pone en peligro el suministro de nutrientes para el
feto y resultando en restricción del crecimiento. Así, Wentzel y colaboradores (2006)
mostraron que ratas expuestas a un régimen crónico de etanol 20% vía oral hasta la gestación
a término, indujo la disminución del peso corporal materno y fetal, aunque la exposición
produjo un aumento en el peso placentario, efecto análogo al generado por la diabetes
materna (Eriksson y Jansson, 1984). Por su parte, Gundogan y colaboradores (2008) vieron que
ratas expuestas de manera crónica a etanol al 37% desde el día 6 de gestación y hasta el día
16, presentaron alta reabsorción fetal y deterioro del desarrollo de la placenta, visto como una
reducción del peso placentario al día 16. Los estudios histológicos demostraron que la capa
laberíntica es particularmente afectada por la exposición a alcohol, debido a la inducción de
isquemia que genera trombosis en el parénquima de las capas laberínticas. En el presente
trabajo de tesis, en los tres grupos tratados también fue observada una reducción de los
espacios sanguíneos maternos y alta cantidad de leucocitos, lo que sugiere un efecto
inflamatorio resultante de la injuria del tejido. Estos efectos podrían también estar asociados
con anomalía de la remodelación vascular, tal como fue propuesto por Gundogan y col. (2008,
2010). Los autores describieron que las alteraciones placentarias vistas luego de la exposición
gestacional crónica a etanol relacionadas con la escasa remodelación de los vasos sanguíneos
del útero, comprometen el flujo sanguíneo placentario y el intercambio de nutrientes. Esta
alteración del flujo sanguíneo útero-placentario se ha relacionado con abortos tempranos y
con restricción del crecimiento intrauterino. Cabe destacar que la disminución del crecimiento
fetal, en estos modelos animales, se produjo por la exposición a etanol durante toda la
gestación, desde el inicio de la placentación hasta el desarrollo a término (Wentzel y col.,
2006; Rutland y col., 2007; Gundogan y col., 2008, 2010), casos en que los efectos del etanol
sobre la placenta y/o el feto, son de tipo directos. En nuestro modelo de ingesta temprana de
alcohol, cuando su administración cesó al día 4, 8 o 10 de gestación, los efectos adversos del
etanol sobre el feto y la placenta fueron indirectos y que, por lo tanto, un efecto hipóxico e

Discusión
309
ineficiente transporte de nutrientes inducido por defectos vasculares dados por las
alteraciones en el laberinto, pudo haberse desencadenado tardíamente. Así, el deterioro en la
expansión del laberinto, la vasculopatía, sumado al aumento del espesor de la membrana
interhemal, observado en el presente trabajo, serían algunos de los mecanismos morfológicos
más importantes por los cuales la exposición materna a etanol durante la preñez temprana
resulta en alteración placentaria a término.
Aunque el desarrollo de la placenta fue afectado por el alcohol, lo que induciría crecimiento y
desarrollo intrauterino fetal retrasado, como fue sugerido por otros autores (Reynolds y
Redmer, 1995; Rutland y col., 2007), la deficiente expansión de la zona laberíntica no
repercutió en el peso fetal de los tres grupos tratados, y por lo tanto la exposición temprana a
alcohol no llevó a restricción del crecimiento fetal. Una hipótesis que explicaría este resultado,
está relacionada con los cambios observados en el espesor de la zona de unión y abundancia
de CGs. Planteamos que la acción negativa del etanol sobre el desarrollo del laberinto induciría
una consecuente reacción compensatoria en la zona de unión, ya sea aumentando la cantidad
de CGs o impidiendo su reducción hacia el final de la gestación, incrementando así las reservas
energéticas para el aporte nutricional necesario en las últimas etapas de la gestación donde la
demanda fetal es alta. Sugerimos que el incremento de esta reserva energética evitaría, al
menos en parte, la restricción del crecimiento fetal. Sin embargo, aunque no se detectó
restricción del crecimiento intrauterino de acuerdo al análisis de los parámetros relacionados
estudiados, no podemos descartar la posibilidad de la existencia de una deficiencia funcional
del laberinto en todos los grupos de hembras tratadas. Esta hipótesis se apoya en los
resultados vistos por otros autores. De la misma manera, las placentas de ratas diabéticas
presentan una zona de unión de mayor espesor que el control, mientras que el laberinto, tuvo
membranas trofoblásticas interhemales significativamente más gruesas. Los autores, ante
estas observaciones, propusieron que el aumento del número de CGs en las placentas de la
ratas diabéticas, podría ser un mecanismo de compensación para aliviar sus deficiencias
nutricionales (Padmanabhan y Shafiullah, 2001). Más aún, en los últimos años, varios grupos
de investigación han informado casos de desarrollo placentario alterado, donde las
observaciones cualitativas indican un cambio en el equilibrio entre las células EPGS y CGs, y
que la ampliación de la zona de unión está relacionada al aumento del número de CGs (Yu y
col., 2008).
Sin duda, para explicar en forma completa las alteraciones placentarias observadas a largo
plazo en los grupos tratados con etanol hasta los días 8 y 10 de gestación, y sobre todo en el

Discusión
310
grupo expuesto hasta el día 4, se debe considerar que, probablemente, las alteraciones en el
desarrollo temprano del linaje trofoblástico, en la regulación de la invasión del trofoblasto
durante la implantación y decidualización deficiente, entre otras causas podrían justificar las
anomalías feto-placentarias observadas a largo plazo en el presente trabajo.
En síntesis, el consumo de alcohol desde antes de la preñez, en baja concentración, y durante
períodos gestacionales tempranos y medios, es inductor de anomalías embrio-feto-
placentarias, las que podrían implicar un riesgo para la vida postnatal futura. Tanto la pérdida
gestacional temprana, la inducción de malformaciones fetales, las anomalías en el esqueleto
fetal así como las alteraciones placentarias a término, todas características del FAS, son
demostradas por primera vez, con el presente trabajo, en el modelo murino exocriado
expuesto a bajas dosis de alcohol perigestacionalmente hasta la preñez temprana.
Consideraciones finales
A pesar de los conocimientos con que se cuentan hoy en día acerca del papel teratogénico del
alcohol, la incidencia del Síndrome de Alcoholismo Fetal en la población humana continúa
siendo relevante a nivel médico-social. Por esto, las investigaciones que profundizan el
conocimiento en relación a los efectos y mecanismos de acción del consumo prenatal de
etanol son de suma importancia para enfatizar sobre la prevención de la ingesta materna de
alcohol, esclarecer acerca de los umbrales de concentración nocivos y ahondar en el desarrollo
de estrategias para mitigar los daños por exposición a etanol durante la gestación.
Según diversos trabajos en animales de experimentales, el tratamiento con alcohol en dosis o
concentraciones clasificadas como altas, valores numéricos que aún no han sido
uniformemente definidos, sin dudas produce efectos deletéreos en los procesos del desarrollo
embrio-placentario estudiados en el presente trabajo. Sin embargo, dados ciertos
interrogantes acerca de potenciales efectos nocivos por exposiciones a dosis bajas y
moderadas de etanol durante la preñez temprana, nuestra investigación se focalizó en
dilucidar estos efectos en el desarrollo y crecimiento embrio-trofoblástico en fases
periimplantatorias de la gestación del ratón.
En base a los resultados aportados por el presente estudio, podemos sugerir la inducción de
efectos deletéreos significativos a nivel embrionario, fetal y placentario, en diferentes etapas
claves de la gestación, luego de la administración de etanol al 10% durante un período
perigestacional y no más allá de la mitad de la gestación murina.

Discusión
311
Los abordajes in-vivo permitieron inferir conclusiones en el desarrollo temprano respecto del
efecto del etanol sin perder de vista el entorno materno. Los abordajes in-vitro, por su parte,
permitieron el estudio de eventos específicos del desarrollo y discernir sobre la inducción de
anomalías específicamente embrionarias sin la influencia de mecanismos regulatorios
maternos, y permitió el seguimiento de procesos que in-vivo no es posible. La utilización de
dos colonias exocriadas diferentes, enriqueció notablemente el conocimiento actual sobre la
temática de estudio y el comportamiento del modelo animal. Los resultados obtenidos en las
colonias CF-1 y CD-1 permitieron, además de la evaluación de sus respuestas frente a la
exposición a alcohol, reflexionar acerca de la introducción de protocolos hormonales para la
estimulación ovárica y su posible influencia en los efectos reproductivos-gestacionales
generados específicamente por el alcohol. Por otra parte, la diferente respuesta reproductiva
frente a la ingesta de alcohol de la colonia CD-1 permitió en particular realizar estudios a largo
plazo, para evaluar a nivel fetal y placentario los efectos de la exposición materna temprana a
alcohol que, sin duda, en el modelo murino superovulado, no hubiera sido posible realizar.
En conclusión, la ingesta perigestacional de etanol 10% desde antes y hasta la gestación
temprana/media, produjo:
-Anomalías embrionarias y trofoblásticas en los procesos del desarrollo temprano, relativas a la
inducción, al día 4, 5 y 8 de gestación, de retraso de la diferenciación y del crecimiento
embrionario, dismorfogénesis, desregulación de la capacidad invasiva trofoblástica temprana y
arresto del desarrollo periimplantatoria.
-Disrupción de la expresión de moléculas de adhesión intercelular en blastocistos (ZO-1 y E-
cadherina) y en ectodermo de embriones en grastrulación (E-cadherina); alteración de los
niveles de expresión de MMP-2 y MMP-9 en tejido embrio-trofoblástico de embriones en
gastrulación; genotoxicidad materna y embrionaria junto con alta apoptosis en embriones de
preimplantación y en gastrulación.
-Malformaciones craneofaciales, alteraciones esqueléticas fetales y anomalías estructurales y
del crecimiento placentario, en la gestación a término.


Bibligrafía
312
-Abrahamsohn P. A. (1983). Ultrastructural study of the mouse antimesometrial decidua. Anat Embryol 166
(2), 263-74.
-Abrahamsohn P. A. y Zorn T. M. T. (1993). Implantation and decidualization in rodents. J Exp Zool 266 (6),
603-628.
-Adachi M., Inoko A., Hata M., Furuse K., Umeda K., Itoh M. y Tsukita S. (2006). Normal establishment of
epithelial tight junctions in mice and cultured cells lacking expression of ZO-3, a tight-junction MAGUK
protein. Mol Cell Biol 26, 9003–9015.
-Adamson S. L., Lu .Y, Whiteley K. J., Holmyard D., Hemberger M., Pfarrer C. y Cross J. C. (2002).
Interactions between trophoblast cells and the maternal and fetal circulation in the mouse placenta. Dev
Biol 250 (2), 358–373.
-Ain R., Canham L. N. y Soares M. J. (2003). Gestation stage-dependent intrauterine trophoblast cell
invasion in the rat and mouse: novel endocrine phenotype and regulation. Dev Biol 260 (1), 176-190.
-Aitken R. J., Clarkson J. S. y Fishel S. (1989). Generation of reactive oxygen species, lipid peroxidation, and
human sperm function. Biol Reprod 41 (1), 183-197.
-Alarcon V. B. (2010). Cell polarity regulator PARD6B is essential for trophectoderm formation in the
preimplantation mouse embryo. Biol Reprod 83 (3), 347–358.
-Alexander C. M., Hansell E. J., Behrendtsen O., Flannery M. L., Kishnani N. S, Hawkes S. P. y Werb Z.
(1996). Expression and function of matrix metalloproteinases and their inhibitors at the maternal-embryonic
boundary during mouse embryo implantation. Development 122 (6), 1723-1736.
-Alikani M. , Cohen J., Tomkin G., Garrisi G. J., Mack C., Scott R. T. (1999). Human embryo fragmentation in
vitro and its implications for pregnancy and implantation. Fertil Steril 71 (5), 836–842.
-Alireza Shirpoor, Syranush Minassian, Siamak Salami, Mohammad Hassan Khadem-Ansari y Marine
Yeghiazaryan (2009). Vitamin E protects developing rat hippocampus and cerebellum against ethanol-
induced oxidative stress and apoptosis. Food Chemistry 113 (1), 115–120.
-Almeida Domingues J., Tancredo Toledo M. y Guimarães Moraes S. (2009). Maternal and fetal liver
histomorphological analysis of malnourished pregnant rats submitted to exposure to ethanol. Rev Fac Ciênc
Méd Sorocaba 11 (3), 9-17.
-Amini S. A., Dunkley P. R. y Murdoch R. N. (1996). Teratogenic effects of etanol in the Quackenbush Special
mouse. Drug Alcohol Depend 41 (1), 61-69.
-Amoroso E. C. (1952). Placentation. En: Parkes A. S. Marshall´s Physiology of Reproduction. Longman,
London pp 127-311.
-Anacker J., Segerer S. E., Hagemann C., Feix S., Kapp M., Bausch R. y Kämmerer U. (2011). Human decidua
and invasive trophoblasts are rich sources of nearly all human matrix metalloproteinases. Mol Hum Reprod
17 (10), 637–652.
-Anderson R. A. Jr, Willis B. R., Phillips J. F., Oswald C. y Zaneveld L. J. (1987). Delayed pubertal
development of the male reproductive tract associated with chronic ethanol ingestion. Biochem Pharmacol
36 (13), 2157-2167.
-Ang H. L., Deltour L., Hayamizu T. F., Zgombić-Knight M., Duester G. (1996). Retinoic acid synthesis in
mouse embryos during gastrulation and craniofacial development linked to class IV alcohol dehydrogenase
gene expression. J Biol Chem 271 (16), 9526-34.
-Ansell J. D. y Snow M. H. (1975). The development of trophoblast in vitro from blastocysts varying amounts
of inner cell mass. J Embryol Exp Morphol 33, 177-185.
-Antczak M. y Van Blerkom J. (1999). Temporal and spatial aspects of fragmentation in early human
embryos: possible effects on developmental competence and association with the differential elimination of
regulatory proteins from polarized domains. Hum Reprod 14 (2), 429–447.
-Anthony B., Zhou F. C., Ogawa T., Goodlett C. R. y Ruiz J. (2008). Alcohol exposure alters cell cycle and
apoptotic events during early neurulation. Alcohol Alcohol 43 (3), 261–273.
-Aragón A. S., Kalberg W. O., Buckley D., Barela-Scott L., Tabachnick B. G y May P. A. (2008).
Neuropsychological study of FASD in a sample of American Indian children: processing simple versus
complex information. Alcohol Clin Exp Res 32 (12), 2136-2148.
-Arends M.J. y Wyllie A.H. (1991). Apoptosis: mechanisms and roles in pathology. Int Rev Exp Pathol 32,
223-254.

Bibligrafía
313
-Armant D. R., Kaplan H. A. y Lennarz W. I. (1986). Fibronectin and laminin promete in vitro attachment and
outgrowth of mouse blastocysts. Dev Biol 116 (2), 519-523.
-Aye M. M., Ma C., Lin H., Bower K. A., Wiggins R. C. y Luo J. (2004). Ethanol-induced in vitro invasion of
breast cancer cells: the contribution of MMP-2 by fibroblasts. Int J Cancer 112 (5), 738–746.
-Balansky R. M., Blagoeva P. M., Mircheva Z. I. y Flora S. (1993). Coclastogenicity of ethanol with cigarette
smoke in rat erythroblasts and anticlastogenicity in alveolar macrophages. Cancer Lett 72 (3), 183-189.
-Balda M. S. y Matter K. (2000). The tight junction protein ZO-1 and an interacting transcription factor
regulate ErbB-2 expression. The EMBO J 19 (9), 2024-2033.
-Bannigan J. y Cotteli D. (1984). Ethanol teratogenicity in mice: an electron microscopic study. Teratology 30
(2), 281-290.
-Bany B.M., Harvey M. B. y Schultz G. A. (2000). Expression of matrix metalloproteinases 2 and 9 in the
mouse uterus during implantation and oil-induced decidualization. J Reprod Fertil 120 (1), 125–134.
-Barash V. y Shafrir E. (1990). Mobilization of placental glycogen in diabetic rats. Placenta 11 (6), 515-521.
-Barker D. J. (2004). The developmental origins of adult disease. J Am Coll Nutr 23 (6 Suppl), 588S-595S.
-Barlow P., Owen D. A. J. y Graham C. (1972). DNA synthesis in the preimplantation mouse embryo. J
Embryol Exp Morphol 27 (2), 431-445.
-Bazer F. W., Spencer T. E., Johnson G. A., Burghardt R. C. y Wu G. (2009). Comparative aspects of
implantation. Reproduction 138 (2), 195-209.
-Becker H. C. y Randall C. L. (1987). Two generations of maternal alcohol consumption in mice: Effect on
pregnancy outcome. Alcohol Clin Exp Res 11 (3), 240-242.
-Becker H. C., Randall C. L. y Middaugh L. D. (1989). Behavioral teratogenie effects of ethanol in mice. Ann
N Y Acad Sci 562, 340-341.
-Becker H. C., Diaz-Granados J. L. y Randall C. L. (1996). Teratogenic actions of ethanol in the mouse: A
Minireview. Pharmacol Biochem Behav 55 (4), 501-513.
-Behrendtsen O., Alexander C. M. y Werb Z. (1992). Metalloproteinases mediate extracellular matrix
degradation by cells from mouse blastocyst outgrowth. Development 114 (2), 447-56.
-Behrendtsen O. y Werb Z. (1997). Metalloproteinases regulate parietal endoderm differentiating and
migrating in cultured mouse embryos. Dev Dyn 208 (2), 255–265.
-Benavides F. J. y Guénet J. L . (2003). Las líneas genéticamente estandarizadas y los controles de la pureza
genética. En: Manual de Genética de Roedores de Laboratorio: Principios Básicos y Aplicaciones. Publicación
Madrid: Universidad De Alcalá, 105-136.
-Bergers G., Brekken R., McMahon G., Vu T.H., Itoh T., Tamaki K., Tanzawa K., Thorpe P., Itohara S., Werb
Z. y Hanahan D. (2000). Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogénesis.
Nat Cell Biol 2, 737–744.
-Bertrand J., Floyd R. L., Weber M. K., O’Connor M., Riley E. P., Johnson K. A. y Cohen D. E. (2004). National
Task Force on FAS/FAE: Guidelines for Referral and Diagnosis. Atlanta: Centers for Disease Control and
Prevention, pp1-50.
-Betts D. H. y King W. A. (2001). Genetic regulation of embryo death and senescence. Theriogenology 55
(1), 171-191.
-Bevilacqua E. (1984). Estudo ultra-estrutural da interação materno-embrionária durante o inicio da
gestação de camundongo. Tesis de Doctorado. Instituto de Ciências Biomédicas, Universidade de São Paulo,
Brasil.
-Bevilacqua E. M. y Abrahamsohn P. A. (1988). Ultrastructure of trophoblast giant cell transformation
during the invasive stage of implantation of the mouse embryo. J Morphol 198 (3), 341-351.
-Bevilacqua E. y Abrahamsohn P. A. (1989).Trophoblast invasion during implantation of the mouse embryo.
Arch Biol Med Exp. 22, 107-118.
-Bevilacqua E. M., Faria M. R. y Abrahamsohn P. A. (1991). Growth of mouse ectoplacental cone cells in
subcutaneous tissues. Development of placental-like cells. Am J Anat. 192 (4), 382-399.
-Bevilacqua E., Hoshida M. S., Amarante-Paffaro A., Albieri-Borges A. y Zago Gomes S. (2010). Trophoblast
phagocytic program: roles in different placental systems. Int J Dev Biol 54 (2-3), 495-505.
-Biggers J. D., Bell J. E. y Benos D. J. (1988). Mammalian blastocyst: transport functions in a developing
epithelium. A J Physiol 255 (4 Pt 1), 419-32.

Bibligrafía
314
-Biggers J. D., Baltz J. M. y Lechene C. (1991). Ions and preimplantation development. En: Animal
Applications of Research in Mammalian Development. Cold Spring Harbor, New York: Cold Spring Harbor
Laboratory Press, pp 121-146.
-Billington W. D. (1971). Biology of the trophoblast. Adv Reprod Physiol 5, 27-66.
-Bischof P. y Martelli M. (1992). Current topic: proteolysis in the penetration phase of the implantation
process. Placenta 13 (1), 17-24.
-Bischof P., Martelli M. y Campana A. (1994). Controlled extracellular-matrix degradation: A fundamental
mechanism in the implantation process. En: Glasser S. R., Mulholand J. y Psychoyos A. (Eds). Endocrinology
of Embryo-Endometrium Interactions. New York, Plenum pp 379-390.
-Bischof P., Meisser A. y Campana A. (2000). Paracrine and autocrine regulators of trofoblast invasion.
Placenta 21, Supl A S55-60.
-Blakley P. M. (1988). Expperimental teratology of ethanol. En: Kalter H. (Ed) Issues and Reviews in
Teratoloty, vol 4 New York, Plenum pp 237-283.
-Blomberg L., Hashizume K. y Viebahn C. (2008). Blastocyst elongation, trophoblastic differentiation, and
embryonic pattern formation. Reproduction 135 (2), 181-195.
-Bloor D. J., Metcalfe A. D., Rutherford A., Brison D. R. y Kimber S. J. (2002). Expression of cell adhesion
molecules during human preimplantation embryo development. Mol Hum Reprod 8 (3), 237-245.
-Boehm S. L., Moore E. M., Walsh C. D., Gross C. D., Cavelli A. M., Gigante E., Linsenbardt D. N. (2008).
Using drinking in the dark to model prenatal binge-like exposure to ethanol in C57BL/6J mice. Dev
Psychobiol 50(6), 566-578.
-Boller K. y Schmid W. (1970). Chemical mutagenesis in mammals. The Chinese hamster bone marrow as an
in vivo test system. Hematological findings after treatment with trenimon. Human genetik 11 (1), 35-54.�
-Borland R. M. (1977). Transport processes in the mamalian blastocyst. En: Johnson M. (Ed) Development in
Mamamls. New York, North Holland Pusblishing Co., pp 31-68.
-Borson W. F. y Li T. K. (1986). Genetic polymorphism of human liver alcohol and aldehyde dehydrogenases,
and their relationship to alcohol metabolism and alcoholism. Hepatology 6 (3), 502-510.
-Bosco C. (2005). Alcohol and xenobiotics in placenta damage. In Preedy V. R. y Watson R. R. (Eds).
Comprehensive Handbook of Alcohol Related Pathology, 2. London: Elsevier Science, 921-935.
-Bosco C. y Diaz E. (2012). Placental hypoxia and foetal development versus alcohol exposure in pregnancy.
Alcohol Alcohol 47 (2), 109–117.
-Brannigan J. y Burke P. (1982). Ethanol teratogenicity in mice: a light microscope study. Teratology 26 (3),
247-254.
-Brembeck F. H., Rosario M. y Birchmeier W. (2006). Balancing cell adhesion and Wnt signaling, the key role
of beta-catenin. Curr Opin Genet Dev 16 (1), 51-59.
-Brill A., Torchinsky A., Carp H. y Toder V. (1999). The role of apoptosis in normal and abnormal embryonic
development. J Assist Reprod Genet 16 (10), 512-519.
-Brison D. R. y Leese H. J. (1994). The role of exogenous energy sbstrate in blastocele fluid accumulation in
the rat. Zigote 2 (1), 69-77.
-Brison D. R. y Shultz R. M. (1997). Apoptosis during mouse blastocyst formation: evidence for a role for
survival factors including Transforming Growth Factor a. Biol Reprod 56 (5), 1088-1096.
-Burd L., Roberts D., Olson M., y Odendaal H. (2007). Ethanol and the placenta: a review. J Matern Fetal
Neonatal Med 20 (5), 361–375.
-Burdsal C. A., Damsky C. H. y Pedersen R. A. (1993). The role of E-cadherin and integrins in mesoderm
differentiation and migration at the mammalian primitive streak. Development 118 (3), 829-844.
-Burim R. V., Canalle R., Takahashi C. S., Tavares D. C., Martinelli Ad Ade L. y Sakamoto-Hojo E. T. (2004).
Clastogenic effect of ethanol in chronic and abstinent alcoholics. Mutat Res 560(2), 187-198.
-Byrne A. T., Southgate J., Brison D. R. y Leese H. J. (1999). Analysis of apoptosis in the preimplantation
bovine embryo using TUNEL. J Reprod Fertil 117 (1), 97-105.
-Byrne A. T., Southgate J., Brison D. R. y Leese H. J. (2002). Regulation of apoptosis in the bovine blastocyst
by insulin and the insulin-like growth factor (IGF) superfamily. Mol Reprod Dev 62 (4), 489-495.
-Cahill P. A. y Redmond E. M. (2012). Alcohol and cardiovascular disease-modulation of vascular cell
function. Nutrients 4, 297-318.

Bibligrafía
315
-Canteros G., Rettori V., Franchi A., Genaro A., Cebral E., Faletti A., Gimeno M. y McCann S. M. (1995).
Ethanol inhibits luteinizing hormone-releaseing hormone (LHRH) secretion by blocking the response of LHRH
neuronal terminals to nitric oxide. Proc Natl Acad Sci USA 92 (8), 3416-3420.
-Carmona J. C. H. (2008). Metabolismo del etanol: comparación del efecto de las grasas saturadas e
insaturadas en la reducción de problemas hepáticos inducidos por el alcohol. Archivos de Medicina
(Universidad de Manizales) 8 (1), 55-62.
-Carneige J. A. y Cabaca (1993). Extracellular matrix composition and resilience: two parameters that
influence the in vitro migration and morphology of rat inner cell mass-derived cells. Biol Reprod 48, 287-299.
-Cartwright M. M., Tessmer L. L. y Smith S. M. (1998). Ethanol-induced neural crest apoptosis is coincident
with their endogenous death, but is mechanistically distinct. Alcohol Clin Exp Res 22 (1), 142-149.
-Cartwright M. M. y Smith S. M. (1995). Stage dependent effects of ethanol on cranial neural crest cell
development: Partial basis for the phenotypic variations observed in fetal alcohol syndrome. Alcohol Clin
Exp Res 19 (6), 1454-1462.
-Casarett L. J. y Doull´s J. (2001). Toxicology, the basis of the science of poisons, 6a Klaassen C. D., Amdus M.
O. Y Doull´s J. (Eds). Macmillan Publishing Company.
-Cebral E., Lasserre A., Rettori V. y Gimeno M. A. F. (1997). Impaired mouse fertilization by low chronic
alcohol treatment. Alcohol Alcohol 32 (5), 563-572.
-Cebral E., Lasserre A., Faletti A. B. y Gimeno M. A. F. (1998a). Response to ovulatory induction following
moderate chronic ethanol administration in mice. Med Sci Res 26, 29–31.
-Cebral E., Lasserre A., Motta A. y Gimeno M. A. F. (1998b). Mouse oocyte quality and prostaglandin synthesis
by cumulus oocyte complex after moderate chronic ethanol intake. Prostaglandins Leukot Essent Fatty Acids 58
(5), 381-387.
-Cebral E., Lasserre A., Rettori V. y De Gimeno M. A. (1999). Deleterious effects of chronic moderate alcohol
intake by female mice on preimplantation embryo growth in vitro. Alcohol Alcohol 34 (4), 551-558.
-Cebral E., Lasserre A., Rettori V. y de Gimeno M. A. (2000). Alterations in preimplantation in vivo
development after preconceptional chronic moderate alcohol consumption in female mice. Alcohol Alcohol
35 (4), 336-343.
-Cebral E., Rettori V., de Gimeno M. A. (2001). Impact of chronic low-dose ethanol ingestion during sexual
maturation of female mice on in-vitro and in-vivo embryo development. Reprod Toxicol 15 (2), 123-129.
-Cebral E., Abrevaya X. C. y Mudry M. D. (2011). Male and female reproductive toxicity induced by sub-
chronic ethanol exposure in CF-1 mice. Cell Biol Toxicol 27 (4), 237-248.
-Ceccanti M., Spagnolo A. P., Tarani L., Attilia M. L., Chessa L., Mancinelli R., Stegagno M., Sasso F.
G., Romeo M., Jones K. L., Robinson L. K., Del Campo M., Gossage P. J., May P.A., Hoyme E. H. (2007).
Clinical delineation of fetal alcohol spectrum disorders (FASD) in Italian children: comparison and contrast
with other racial/ethnic groups and implications for diagnosis and prevention. Neurosci Biobehav Rev 31 (2),
270-277.
-Chalon S. y Garel J. M. (1985). Plasma calcium control in the rat fetus. I. Influence of maternal hormones.
Biol Neonate 48 (6), 313-322.
-Chappel S. C. y Howles C. (1991). Re-evaluation of the roles of luteinizing hormone and follicle-stimulating
hormone in the ovulatory process. Hum Reprod 6, 1206-1212.
-Charness M. E., Safran R. M. y Perides G. (1994). Ethanol inhibits neural cell-cell adhesion. J Biol Chem 269
(12), 9304-9309.
-Chavassieux P., Serre C. M., Vergnaud P., Delmas P. D. y Meunier P. J. (1993). In vitro evaluation of dose-
effects of ethanol on human osteoblastic cells. Bone Miner 22 (2), 95-103.
-Chavez S. L., Loewke K. E., Han J., Moussavi F., Colls P., Munne S., Behr B. y Reijo Pera R. A. (2012).
Dynamic blastomere behaviour reflects human embryo ploidy by the four-cell stage. Nat Commun 3, 1251.
-Checiu M. y Sandor S. (1986). The effect of ethanol upon early development in mice and rats. IX. Late effect
of acute preimplantation intoxication in mice. Morphol Embryol 32 (1), 5-11.
-Chen L., Nakai M., Belton R. J. Jr y Nowak R. A. (2007). Expression of extracellular matrix metalloproteinase
inducer and matrix metalloproteinases during mouse embryonic development. Reproduction 133 (2), 405–
414.

Bibligrafía
316
-Chen C. C., Hsieh M. S., Hsuuw Y. D., Huang F. J. y Chan W. H. (2010). Hazardous effects of curcumin on
mouse embryonic development through a mitochondria-dependent apoptotic signaling pathway. Int J Mol
Sci 11 (8), 2839-2855.
-Chernoff G. F. (1980). The fetal alcohol syndrome in mice: maternal variables. Teratology 22 (1), 71-75.
Chi H. J., Koo J. J., Choi S. Y., Jeong H. J. y Roh S. I. (2011). Fragmentation of embryos is associated with both
necrosis and apoptosis. Fertil Steril 96 (1), 187-192.
-Ciesiolka M., Delvaeye M., Van Imschoot G., Verschuere V., McCrea P., van Roy F. y Vleminckx K. (2004).
p120 catenin is required for morphogenetic movements involved in the formation of the eyes and the
craniofacial skeleton in Xenopus. J Cell Sci 117 (Pt 18), 4325-4339.
-Clarren S. K. y Smith D. W. (1978). The fetal alcohol syndrome. N Engl J Med 298 (19), 1063-1067.
-Clugston R. D. y Blaner W. S. (2012). The Adverse Effects of Alcohol on Vitamin A Metabolism. Nutrients 4
(5), 356-371.
-Coan P. M., Ferguson-Smith A. C. y Burton G. J. (2004). Developmental Dynamics of the Definitive Mouse
Placenta Assessed by Stereology. Biol Reprod 70 (6), 1806–1813.
-Coan P. M., Ferguson-Smith A. C., Burton G. J. (2005). Ultrastructural changes in the interhaemal
membrane and junctional zone of the murine chorioallantoic placenta across gestation. J Anat 207 (6), 783-
796.
-Coan P. M., Conroy N., Burton G. J. y Ferguson-Smith A. C. (2006). Origin and Characteristics of glycogen
cells in the developing murine placenta. Dev Dyn 235 (12), 3280-3294.
-Cohen M., Meisser A. y Bischof A. (2006). Metalloproteinases and human placental invasiveness. Placenta
27 (8), 783-793.
-Coll T. A., Pérez-Tito L., Sobarzo C. M. A. y Cebral E. (2011). Embryo developmental disruption during
organogenesis produced by CF-1 murine periconceptional alcohol consumption. Birth Defects Res B 92 (6),
560-574.
-Collins J.A., Schandl C.A., Young K.K., Vesely J. y Willingham M. C. (1997). Major DNA fragmentation is a
late event in apoptosis. J Histochem Cytochem 45 (7), 923-934.
-Cook J. D. (2003). Biochemical markers of alcohol use in pregnant woman. Clin Biochem 36 (1), 9-19.
-Copp A. J. (1978). Interaction between inner cell mass and trophectoderm of the mouse blastocyst. A study
of cellular proliferation. J Embryol Exp Morphol 48, 109-125.
-Copp A. J. (1979). Interaction between inner cell mass and trophectoderm. The fate of the polar
trophectoderm. J Embryol Exp Morphol 51, 109-120.
-Copp A. J., Greene N. D. y Murdoch J. N. (2003). The genetic basis of mammalian neurulation. Nat Rev
Genet 4 (10), 784–793.
-Coucouvanis E. y Martin G. R. (1995). Signals for death and survival: a two-step mechanism for cavitation in
the vertebrate embryo. Cell 83(2), 279–287.
-Crabb D. W., Bosron W.F y Li T. K. (1987). Ethanol metabolism. Pharmac Ther 34 (1), 59-73.
-Cross J. C., Werb Z. y Fisher S. J. (1994). Implantation and the placenta: key piceces of the development
puzzle. Science 266 (5190), 1508-1518.
-Croy B. A., Ashkar A. A., Foster R. A., DiSanto J. P., Magram J., Carson D., Gendler S. J., Grusby M. J.,
Wagner N., Muller W. y Guimond M. J. (1997). Histological studies of gene-ablated mice support important
functional roles for natural killer cells in the uterus during pregnancy. J Reprod Immunol 35 (2), 111-133.
-Cross J. C. (2000). Genetic insights into trophoblast differentiation and placental morphogenesis. Semin Cell
Dev Biol 11 (2), 105-113.
-Cudd T. (2005). Animal model systems for the study of alcohol teratology. Exp Biol Med 230 (6), 389-393.
-Cunningham C. C., Coleman W. B. y Spach P. I. (1990). The effects of chronic ethanol consumption on
hepatic mitocondrial energy metabolism. Alcohol Alcohol 25 (2-3), 127–136.
-Cuthbertson K. S. R. (1983). Parthenogenetic activation of mouse oocyte in vivo with ethanol and benzil
alcohol. J Exp Zool 226 (2), 311-314.
-Czechanski A., Byers C., Greenstein I., Schrode N., Donahue L. R., Hadjantonakis A. K. y Reinholdt L. G.
(2014). Derivation and characterization of mouse embryonic stem cells from permissive and nonpermissive
strains. Nat Protoc 9 (3), 559–574.

Bibligrafía
317
-Czeizel A. E. (2000). Primary prevention of neural-tube defects and some other major congenital
abnormalities: recommendations for the appropriate use of folic acid during pregnancy. Paediatr Drugs 2
(6), 437-449.
-D`Amato F. (1989). Polyploidy in cell differentiation. Caryologia 42, 183-211.
-Damsky C. H., Sutherland A. y Fisher S. (1993). Extracellular matrix 5: adhesive interactions in early
mammalian embryogenesis, implantation and placentation. FASEB J 7 (14), 1320-1329.
-Darzynkiewicz Z., Bedner E. y Traganos F. (2001). Difficulties and pitfalls in analysis of apoptosis. Methods
Cell Biol 63, 527–546.
-Das S. K., Yano S., Wang J., Edwards D. R., Nagase H. y Dey S. K. (1997). Expression of matrix
metalloproteinases and tissue inhibitors of metalloproteinases in the mouse uterus during the peri-
implantation period. Dev Genet 21 (1), 44-54.
-Davies J. y Glasser S. R. (1968). Histological and fine structural observation on the placenta of the rat. Part I.
Organization of the normal placenta. Acta Anat 69 (4), 542-608.
-Day N. L. y Richardson G. A. (1991). Prenatal alcohol exposure: a continuum of effects. Semin Perinatol 15
(4), 271-279.
-De Almeida Reis Sr, Sadigursky M., Andrade M. G. S., Soares L. P., Do Espirito Santo A. R. y Vilas Boas D. S.
(2002). Efeito genotoxico do etanol em células da mucosa bucal. Pesqui Odontol Bras 16(3), 221-225.
-De Vries W. N., Evsikov A. V., Haac B. E., Fancher K. S., Holbrook A. E., Kemler R., Solter D. y Knowles B. B.
(2004). Maternal beta-catenin and E-cadherin in mouse development. Development 131 (18), 4435-4445.
-Dean R.A., Butler G.S., Hamma-Kourbali Y., Delbe J., Brigstock D.R., Courty J. y Overall C.M. (2007).
Identification of candidate angiogenic inhibitors processed by matrix metalloproteinase 2 (MMP-2) in cell-
based proteomic screens: disruption of vascular endothelial growth factor (VEGF)/heparin affin regulatory
peptide (pleiotrophin) and VEGF/Connective tissue growth factor angiogenic inhibitory complexes by MMP-
2 proteolysis. Mol Cell Biol 27, 8454-8465.
-DeJonge M. H. y Zachman R. D. (1995).The effect of maternal ethanol ingestion on fetal rat heart vitamin
A: A model for fetal alcohol syndrome. Pediatr Res 37 (4), 418–423.
-Deltour L., Ang H. L. y Duester G. (1996). Ethanol inhibition of retinoic acid synthesis as a potential
mechanism for fetal alcohol syndrome. FASEB J 10 (9), 1050-1057.
-Deng X. y Deitrich R. A. (2007). Ethanol metabolism and effects: nitric oxide and its interaction. Curr Clin
Pharmacol 2 (2), 145–153.
-Dickson A. D. (1963). Trophoblastic giant cell transformation of mouse blastocysts. J Reprod Fertil 6, 465-
466.
-Dive C. y col. (1992). Analysis and discrimination of necrosis and apoptosis (programmed cell death) by
multiparameter flow cytometry. Biochim Biophys Acta 1133 (3), 275-285.
-Downs K. M. (2011). Lineage commitments: emphasis on embryonic-extraembryonic interfaces. EMBO rep
12 (10), 987-990.
-Downs K. M. y Davies T. (1993). Staging of gastrulating mouse embryos by morphological landmarks in the
dissecting microscope. Development 118 (4), 1255-1266.
-Driscoll C. D., Streissguth A. P., Riley E. P. (1990). Prenatal alcohol exposure: comparability of effects in
humans and animal models. Neurotoxicol Teratol 12 (3), 231-237.
-Duester G., Ang H. L., Deltour L., Foglio M. H., Hayamizu T. F. y Zgombic-Knight M. (1995). Class I and class
IV alcohol dehydrogenase (retinol dehydrogenase) gene expression in mouse embryos. Adv Exp Med Biol
372, 301-313.
-Duester G. (2000). Families of retinoid dehydrogenases regulating vitamin A function: production of visual
pigment and retinoic acid. Eur J Biochem 267 (14), 4315-4324.
-Dunty W. C. Jr, Chen S. Y., Zucker R. M., Dehart D. B. y Sulik K. K. (2001). Selective Vulnerability of
Embryonic Cell Populations to Ethanol-Induced Apoptosis: Implications for Alcohol-Related Birth Defects and
Neurodevelopmental Disorder. Alcohol Clin Exp Res 25(10), 1523-1535.
-Ebner T., Yaman C., Moser M., Sommergruber M., Pölz W. y Tews G. (2001). Embryo fragmentation in vitro
and its impact on treatment and pregnancy outcome. Fertil Steril 76 (2), 281–285.
-Eckert J. J. y Fleming T. P. (2008). Tight junction biogenesis during early development. Biochim Biophys Acta
1778 (3), 717–728.

Bibligrafía
318
-Eckert J. J., McCallum A., Mears A., Rumsby M. G, Cameron I. T. y Fleming T. P. (2005). Relative
contribution of cell contact pattern, specific PKC isoforms and gap junctional communication in tight
junction assembly in the mouse early embryo. Dev Biol 288 (1), 234-247.
-Egeblad M. y Werb Z. (2002). New function for the matrix metalloproteinases in cancer progression. Nat
(3) Rev Cancer 2, 161-174.
-Eijnde S. M., Luijsterburg A. J., Boshart L., De Zeeuw C. I., van Dierendonck J. H., Reutelingsperger C. P. y
Vermeij-Keers C. (1997). In situ detection of apoptosis during embryogenesis with annexin V: from whole
mount to ultrastructure. Cytometry 29 (4), 313-320.
-Ellahueñe M. F., Pérez-Alzola L. P. y Olmedo M. I. (2012). Chronic ethanol consumption in mice does not
induce DNA damage in somatic or germ cells, evaluated by the bone marrow micronucleous assay and the
dominant lethal mutation assay. Biol Res 45 (1), 27-31.
-El-Shershaby A. M. y Hinchliffe J. R. (1974). Cell redundancy in the zona intact preimplantation mouse
blastocyst: a light and electron microscopic study of dead cells and their fate. J Embryol Exp Morphol 31 (3),
643-654.
-Enari M., Sakahira H., Yokoyama H., Okawa K., Iwamatsu A. y Nagata S. (1998). A caspase-activated
DNAse that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 391 (6662), 43-50.
-Enders A. C. y Schlafke S. (1967). A morphological analysis of the early implantation stages in the rat. Am J
Anat 120, 185-226.
-Enders A. C. y Schlafke S. (1981). Differentiation of the blastocyst of the rhesus monkey. Am J Anat 162 (1),
1–21.
-Erenus M., Zouves C., Rajamahendran P. et al. (1991). The effect of embryo quality on subsequent
pregnancy rates after in vitro fertilization. Fertil Steril 56, 707–710.
-Eriksson U. J. y Jansson L. (1984). Diabetes in pregnancy: decreased placental blood flow and disturbed
fetal development in the rat. Pediatr Res 18 (8), 735–738.
-Exley G. E., Tang C., McElhinny A. S. y Warner C. M. (1999). Expression of caspase and BCL-2 apoptotic
family members in mouse preimplantation embryos. Biol Reprod 61 (1), 231-239.
-Fabian D., Il’ková G., Rehák P., Czikková S., Baran V., Koppel J. (2004). Inhibitory effect of IGF-I on induced
apoptosis in mouse preimplantation embryos cultured in vitro. Theriogenology 61 (4), 745-755.
-Fabian D., Koppel J. y Maddox-Hyttel P. (2005). Apoptotic processes during mammalian preimplantation
development. Theriogenology 64 (2), 221–231.
-Fabian D., Makarevich A. V, Chrenek P., Bukovská A. y Koppel J. (2007). Chronological appearance of
spontaneous and induced apoptosis during preimplantation development of rabbit and mouse embryos.
Theriogenology 68 (9), 1271-1281.
-Fang X., Ji H., Kim S. W., Park J. I., Vaught T. G., Anastasiadis P. Z., Ciesiolka M. y McCrea P. D. (2004).
Vertebrate development requires ARVCF and p120 catenins and their interplay with RhoA and Rac. J Cell Biol
165 (1), 87-98.
-Farley J. R., Fitzsimmons R., Taylor A. K., Jorch U. M. y Lau K. H. (1985).Direct effects of ethanol on bone
resorption and formation in vitro. Arch Biochem Biophys 238 (1), 305-314.
-Fenech M. y Morley A. A. (1985). Measurement of micronuclei in lymphocytes. Mutat Res 147 (1-2), 29-36.
-Fenech M., Kirsch-Volders M., Natarajan A. T., Surralles J., Crott J. W., Parry J., Norppa H., Eastmond D.
A., Tucker J. D. y Thomas P. (2011). Molecular mechanisms of micronucleus, nucleoplasmic bridge and
nuclear bud formation in mammalian and human cells. Mutagenesis 26 (1), 125-132.
-Festing MF. (1979). Inbred strains in biomedical research. Macmillan Press, London: Oxford University
Press, New York.
-Finn C. A. y Martin L. (1967). Patterns of cells during early pregnancy. J Endocr 39 (4), 393-397.
-Fleming T. P., McConnell J., Johnson M. H. y Stevenson B. R. (1989). Development of tight junctions de
novo in the mouse early embryo: Control of assembly of the tight junction-specific protein, ZO-1. J Cell Biol
108 (4), 1407-1418.
-Fleming T. P., Sheth B. y Fesenko I. (2001). Cell adhesion in the preimplantation mammalian embryo and its
role in trophectoderm differentiation and blastocyst morphogenesis. Front Biosci 6, 1000-1007.
-Folgueras A. R., Pendas A. M., Sanchez L. M. y Lopez-Otin C. (2004). Matrix metalloproteinases in cancer:
from new functions to improved inhibition strategies. Int J Dev Biol 48 (5-6), 411-424.

Bibligrafía
319
-Fowden A. (2001). Growth and metabolism. En: Fetal Growth and Development, Harding R and Bocking A.
D. (Eds). Cambridge: Cambridge University Press, pp. 44-69.
-Friedman S. L. (2008). Mechanisms of hepatic fibrogenesis. Gastroenterology 134 (6), 1655-1669.
-Fromenty B., Vadrot N., Massart J., Turlin B., Barri-Ova N., Lettéron P., Fautrel A. y Robin M. A. (2009).
Chronic ethanol consumption lessens the gain of body weight, liver triglycerides, and diabetes in obese
ob/ob mice. J Pharmacol Exp Ther 331 (1), 23-34.
-Froster U.G. y Baird P. A. (1992). Congenital defects of the limbs and alcohol exposure in pregnancy: data
from a population based study. Am J Med Genet 44 (6), 782-785.
-Gallo P. V. y Weinberg J. (1986). Organ growth and cellular development in ethanol-exposed rats. Alcohol 3
(4), 261-267.
-Gardiner C. S., Williams J. S. y Menino A. R. Jr. (1990a). Sodium/potassium adenosine triphosphatase a-
and b-subunit and a-subunit mRNA levels during mouse embryo develompent in vitro. Biol Reprod 43 (5),
788-794.
-Gardner R. L. (1972). An investigation of inner cell mass and trophoblast tissue following their isolation
from the mouse blastocyst. J Embryol Exp Morphol 28, 279–312.
-Gardner R. L., Papaionnou V. E. y Barton S. C. (1973). Origin of the ectoplacental cone and secundary giant
cells in the mouse blastocysts reconstituted from isolated trophoblast and inner cell mass. J Embryol
Morphol 30 (3), 561-572.
-Gavrieli Y., Sherman Y. y Ben-Sasson S. A. (1992). Identification of programmed cell death in situ via
specific labeling of nuclear DNA fragmentation. J Cell Biol 119 (3), 493–501.
-Gemma S., Vichi S. y Testai E. (2006). Individual susceptibility and alcohol effects: biochemical and genetic
aspects. Ann Ist Super Sanita 42 (1), 8–16.
-Gemma S., Vichi S. y Testai E. (2007). Metabolic and genetic factors contributing to alcohol induced effects
and fetal alcohol syndrome. Neurosci Biobehav Rev 31 (2), 221-229.
-Georgiades P., Ferguson-Smith A. C. y Burton G. J. (2002). Comparative developmental anatomy of the
murine and human definitive placentae. Placenta 23 (1), 3-19.
-Gilliam D., Valdez N., Branson S., Dixon A. y Downing C. (2011). Maternal effects on ethanol teratogenesis
in a cross between A/J and C57BL/6J mice. Alcohol 45 (5), 441-449.
-Gillies R. J., Didier N. y Denton M. (1986). Determination of cell number in monolayer cultures. Anal
Biochem 159 (1), 109-113.
-Giorgetti C., Terriou P., Auquier P., Hans E., Spach J. L., Salzmann J. y Roulier R. (1995). Embryo score to
predict implantation after in vitro fertilization: based on 957 single embryo transfers. Hum Reprod 10 (9),
2427-2431.
-Gjørret J. O., Knijn H. M., Dieleman S. J., Avery B., Larsson L. I. y Maddox-Hyttel P. (2003). Chronology of
apoptosis in bovine embryos produced in vivo and in vitro. Biol Reprod 69 (4), 1193–1200.
-Godin E. A., O’Leary-Moore S. K., Khan A. A., Parnell S. E., Ament J. J., Dehart D. B., Johnson B. W., Allan
Johnson G., Styner M. A. y Sulik K. K. (2010). Magnetic resonance microscopy defines ethanol-induced brain
abnormalities in prenatal mice: effects of acute insult on gestational day 7. Alcohol Clin Exp Res 34 (1), 98-
111.
-Gonçalvez C. R. (1997). Comporatmento das células trofoblásticas de camundongo na presença de
componentes da matriz extracelular. Estudo in vitro. Tesis de Maestría. Instituto de Ciências Biomédicas,
Universidade de São Paulo, Brasil.
-Gonçalves C. R., Antonini S., Vianna-Morgante A. M., Machado-Santelli G. M. y Bevilacqua E. (2003).
Developmental changes in the ploidy of mouse implanting trophoblast cells in vitro. Histochem Cell Biol 119
(3), 189-198.
-Gong Z. y Wezeman F. H. (2004). Inhibitory effect of alcohol on osteogenic differentiation in human bone
marrow-derived mesenchymal stem cells. Alcohol Clin Exp Res 28 (3), 468-479.
-González-Mariscal L., Tapia R. y Chamorro D. (2008). Crosstalk of tight junction components with signaling
pathways. Biochim Biophys Acta 1778 (3), 729-756.
-Graham C. H., McCrae K. R. y Lala P. K. (1993). Molecular mechanisms controlling trophoblast invasion the
uterus. Trophoblast Res 7, 237-250.
-Gramzinski R. A., Parchment R. E. y Pierce G. B. (1990). Evidence linking programmed cell death in the
blastocyst to polyamine oxidation. Differentiation 43 (1), 59-65.

Bibligrafía
320
-Grifo, J. A., Tang, Y. X., Munne S. y Krey L. (1996). Update in preimplantation genetic diagnosis: successes,
advances, and problems. Curr Opin Obstet Gynecol 8 (2), 135–138.
-Grimberg T., Henly D. C. y Worrall S. (1998). The effect of ethanol and cytokines on hepatic metabolism in
rats. Alcohol Clin Exp Res 22, 170A.
-Guide for the care and use of laboratory animals (2010). 8a Ed. The Natinal Academies Press, Washington,
D.C.
-Gumbiner B. (1987). Structure, biochemistry and assembly of epithelial tight junctions. Am J Physiol 253 (6
Pt 1) , 749-758.
-Gumbiner B., Stevenson B. y Grimaldi A. (1988). The role of the cell adhesion molecule uvomorulin in the
formation and maintenance of the epithelial junctional complex. J Cell Biol 107 (4), 1575-1587.
-Gumbiner B., Lowekopf, T. y Apatira D. (1991). Identification of a 160KDa polypeptidethat-binds to the
tight junction protein ZO-1. Proc Natl Acad Sci U S A 88 (8), 3460-3464.
-Gumbiner B. M. (2000). Regulation of cadherin adhesive activity. J Cell Biol 148 (3), 399-403.
-Gundogan F., Elwood G., Longato L., Tong M., Feijoo A., Carlson R. I., Wands J. R., De la Monte S. M.
(2008). Impaired placentation in fetal alcohol syndrome. Placenta 29 (2), 148-157.
-Gundogan F., Elwood G., Mark P., Feijoo A., Longato L., Tong M., de la Monte S. M. (2010). Ethanol-
induced oxidative stress and mitochondrial dysfunction in rat placenta: relevance to pregnancy loss. Alcohol
Clin Exp Res 34 (3), 415–423.
-Gupta M., Gulamhusein A. P. y Beck F. (1982). Morphometric analysis of the visceral yolk sac endoderm in
the rat in vivo and in vitro. J Reprod Fertil 65 (1), 239-245.
-Gutierrez-Adan A., Rizos D., Fair T., Moreira P. N., Pintado B., de la Fuente J., et al. (2004). Effect of speed
of development on mRNA expression pattern in early bovine embryos cultured in vivo or in vitro. Mol
Reprod Dev 68 (4), 441–448.
-Haegel H., Larue L., Ohsugi M., Fedorov L., Herrenknecht K. y Kemler R. (1995). Lack of β-catenin affects
mouse development at gastrulation. Development 121 (11), 3529-3537.
-Hahn-Dantona E., Ramos-DeSimone N., Sipley J., Nagase H., French D. L. y Quigley J. P. (1999).
Activation of ProMMP-9 by a Plasmin/MMP-3 Cascade in a Tumor Cell Model: Regulation by Tissue
Inhibitors of Metalloproteinases. Ann N Y Acad Sci 878, 372-87.
-Halbleib J. M. y Nelson W. J. (2006). Cadherins in development: cell adhesion, sorting, and tissue
morphogenesis. Genes Dev 20 (23), 3199-3214.
-Halliwell B. y Aruoma O. I. (1991). DNA damage by oxygen-derived species. Its mechanism and
measurement in mammalian systems. FEBS Lett 281 (1-2), 9-19.
-Halliwell B. y Chirico S. (1993). Lipid peroxidation: its mechanism, measurement, and significance. Am J Clin
Nutr 57 (5 Suppl), 715S-724S.
-Halsted C.H. (2004). Nutrition and alcoholic liver disease. Semin Liver Dis 24 (3), 289–304.
-Hamlin G. P., Lu X. J., Roby K. F. y Soares M. J. (1994). Recapitulation of the pathway for trophoblast giant
cell differentiation in vitro: stage-specific expression of members of the prolactin gene family. Endocrinology
134, 2390-2396.
-Hamlin G. P. y Soares M. J. (1995). Regulation of deoxyribonucleic acid synthesis in proliferating and
differentiating trophoblast cells: involvement of transferrin, transforming growth factor-b, and tyrosine
kinase. Endocrinology 136, 322-331.
-Handler J. A. y Thurman R. G. (1988). Catalase-dependent ethanol oxidation in perfused rat liver.
Requirement for fatty-acid-stimulated H2O2 production by peroxisomes. Eur J Biochem 176 (2), 477-484.
-Handyside A. H. y Hunter S. (1986). Cell division and death in the mouse blastocyst before implantation
Roux’s. Archives of Developmental Biology 195, 519–526.
-Hard M. L., Einarson T. R. y Koren G. (2001). The role of acetaldehyde in pregnancy outcome after prenatal
alcohol exposure. Ther Drug Monit 23 (4), 427-434.
-Hard M. L., Abdolell M., Robinson B. H. y Koren G. (2005). Gene-expression analysis after alcohol exposure
in the developing mouse. J Lab Clin Med 145 (1), 47–54.
-Hardy K., Handyside A. H. y Winston R. M. L. (1989). The human blastocyst: cell number, death and
allocation during late preimplantation development in vitro. Development 107 (3), 597–604.
-Hardy K. (1993). Development of human blastocysts in vitro. En: Preimplantation Embryo Development,
Bavister B. D. (Ed). Springer-Verlag, New York, pp 184-199.

Bibligrafía
321
-Hardy K. y Handyside A.H. (1996). Metabolism and cell allocation during parthenogenetic mouse
preimplantation development. Mol Reprod Dev 43 (3), 313-22.
-Hardy K. (1997). Cell death in the mammalian blastocyst. Mol Hum Reprod 3 (10), 919-925.
-Hardy K. (1999). Apoptosis in the human embryo. Rev Reprod 4 (3), 125-134.
-Hardy K., Spanos S., Becker D., Iannelli P., Winston R. M., Stark J. (2001). From cell death to embryo arrest:
mathematical models of human preimplantation embryo development. Proc Natl Acad Sci U S A 98 (4),
1655-1660.
-Hardy K. y Spanos S. (2002). Growth factor expression and function in the human and mouse
preimplantation embryo. J Endocrinol 172 (2), 221-236.
-Harris S. I., Balaban R. S. y Mandel L. J. (1980). Oxygen consumption and cellular ion transport: evidence
for adenosine triphosphate to O2 ratio near 6 in intac cell. Science 208 (4448), 1148-1150.
-Harvey M. B., Leco K. J., Arcellana-Panlilio M. Y., Zhang X., Edwards D. R. y Schultz G. A. (1995).
Proteinase expression in early mouse embryos is regulated by leukemia inhibitory factor and epidermal
growth factor. Development 121 (4), 1005-1014.
-Hatta K. y Takeichi M. (1986). Expression of N-cadherin adhesion molecules associated with early
morphogenetic events in chick development. Nature 320 (6061), 447-9.
-Hay E. D. (1990). Role of cell-matrix contacts in cell migration and epithelial-mesenchymal transformation.
Cell Differ Dev 32 (3), 367-375.
-Hay E. D. (1991). Introductory remarks. En: Cell biology of extracellular matrix. Hay E. D. (Ed). 2a ed New
York, Pleum Press, pp 1-4.
-Haycock P. C. (2009). Fetal alcohol spectrum disorders: the epigenetic perspective. Biol Reprod 81 (4), 607-
617.
-Heddle J. A., Hite M., Kirkhart B., Mavournin K., MacGregor J. T., Newell G. W. Salamone M. F. (1983).
The induction of micronuclei as a measure of genotoxicity. A report of the U. S. Environmental Protection
Agency Gene-Tox Program. Mutat Res 123 (1), 61-118.
-Helige C., Ahammer H., Hammer A., Huppertz B., Frank H. G. y Dohr G. (2008). Trophoblastic invasion in
vitro and in vivo: similarities and differences. Hum Reprod 23 (10), 2282-2291.
-Henery C. C. y Kaufman M.H. (1992). Cleavage rate of haploid and diploid parthenogenetic mouse embryos
during the preimplantation period. Mol Reprod Dev 31 (4), 258–263.
-Hogan B., Constantini F. y Lacy E. (1986). Manipulating the mouse embryo: A laboratory manual. Cold
Spring Harbor, New York, Cold Spring Harbor Laboratory Press.
-Hohn H. P., Boots L. R. y Denker H. W. (1993). Differentiation of human trophoblast cells in vitro stimulated
by extracellular matrix. Trophoblast Res 7, 181-200.
-Holmes R. S., Duley J. A., Algar E. M., Mather P. B. y Rout U. K. (1986). Biochemical and genetic studies on
enzymes of alcohol metabolism: the mouse as a model organism for human studies. Alcohol Alcohol 21 (1),
41-56.
-Hoover K. B, Liao S-Y y Bryant P. J. (1998). Loss of the Tight Junction MAGUK ZO-1 in Breast Cancer:
Relationship to Glandular Differentiation and Loss of Heterozygosity. Am J Pathol 153 (6), 1767-1773.
-Hoshida M. S., Gorjão R., Lima C., Daher S., Curi R. y Bevilacqua E. (2007). Regulation of Gene Expression in
Mouse Trophoblast Cells by Interferon-gamma. Placenta 28, 1059-1072.
-Houghton F. D. (2005). Role of gap junctions during early embryo development. Reproduction 129 (2), 129–
135.
-Hoyme H. E., May P. A., Kalberg W., Kodituwakku P., Gossage J. P., Trujillo P. M., Buckley D., Miller J. H.,
Aragon A. S., Khaole N., Viljoen D. L., Jones K. L. y Robinson L. K. (2005). A practical clinical approach to
diagnosis of fetal alcohol spectrum disorders: clarification of the 1996 Institute of Medicine criteria.
Pediatrics 115 (1), 39-47.
-Hsiang C. Y., Wu S. L., Chen J. C, Lo H. Y., Li C. C, Chiang S. Y., Wu H. C. y Ho T. Y. (2007). Acetaldehyde
induces matrix metalloproteinase-9 gene expression via nuclear factor-κB and activator protein 1 signaling
pathways in human hepatocellular carcinoma cells: Association with the invasive potential. Toxicol Lett 171
(1-2), 78-86.
-Hsuuw Y.D., Chang C.K., Chan W.H. y Yu J.S. (2005). Curcumin prevents methylglyoxal-induced oxidative
stress and apoptosis in mouse embryonic stem cells and blastocysts. J Cell Physiol 205 (3), 379-386.

Bibligrafía
322
-Hu D. y Cross J. C. (2010). Development and function of trophoblast giant cells in the rodent placenta. Int J
Dev Biol 54 (2-3), 341-354.
-Huang F.J., Shen C.C., Chang S.Y., Wu T.C. y Hsuuw Y.D. (2003). Retinoic acid decreases the viability of
mouse blastocysts in vitro. Hum Reprod 18 (1), 130-136.
-Hugues J. N., Cofte T., Perret G., Jayle M. S., Sebaoun J. y Modigliani E. (1980). Hypothalamo-pituitary ovarian
function in 31 women with chronic alcoholism. Clin Endocrinol 12 (6), 543-551.
-Huppertz B., Kadyrov M. y Kingdom J. C. (2006). Apoptosis and its role in the trophoblast. Am J Obstet
Gynecol 195 (1), 29-39.
-Hurle J. M., Ros M. A., Climent V. y Garcia-Martinez V. (1996). Morphology and significance of
programmed cell death in the developing limb bud of the vertebrate embryo. Microsc Res Tech 34 (3),236-
246.
-Hyafil F., Morello D., Babinet C. y Jacob F. (1980). A cell surface glycoprotein involved in the compaction of
embryo carcinoma cells and cleavage stage embryos. Cell 21 (3), 927-934.
-Ikonomidou C., Bittigau P., Ishimaru M. J., Wozniak D. F., Koch C., Genz K., Price M. T., Stefovska V.,
Horster F., Tenkova T., Dikranian K., Olney J. W. (2000). Ethanol-induced apoptotic neurodegeneration and
fetal alcohol syndrome. Science 287 (5455), 1056-1060.
-Ilan N., Mohsenin A., Cheung L. y Madri J.A. (2001). PECAM-1 shedding during apoptosis generates a
membrane-anchored truncated molecule with unique signaling characteristics. FASEB J 15 (2), 362-372.
-Ilgren E. B. (1981). On the control of trophoblastic giant-cell transformation in the mouse: homotypic
cellular interaction and polyploid. J Embryol Exp Morphol 62, 183-202.
-Ilgren E. B. (1983). Control of trophoblast growth. Placenta 4 (3), 307-328.
-Islas S., Vega J., Ponce L. y Gonzalez-Mariscal L. (2002). Nuclear localization of the tight junction protein
ZO-2 in epithelial cells. Exp Cell Res 274 (1), 138-148.
-Ismail S., Buckley S., Budacki R., Jabbar A. y Gallicano G. I. (2010). Screening, diagnosing and prevention of
fetal alcohol syndrome: is this syndrome treatable? Dev Neurosci 32 (2), 91–100.
-Itoh T., Ikeda T., Gomi H., Nakao S., Suzuki T. y Itohara S. (1997). Unaltered secretion of b-amyloid
precursor protein in gelatinase A (matrix metalloproteinase 2)-deficient mice. J Biol Chem 272 (32), 22389-
22392.
-Izquierdo, M. (2002). Intoxicación alcohólica aguda. Adicciones 14 (SUPL 1), 175-193.
-Izzotti A., Balansky R. M., Blagoeva P. M., Mircheva Z. I., Tulimiero L., Cartaglia C. y De Flora S. (1998).
DNA altarations in rat organs after chornic exposuer to cigarette smoke and / or etanol ingestion. FASEB J
12, 753-758.
-Jacobson M. D., Weil M. y Raff M. C. (1997). Programmed cell death in animal development. Cell 88 (3),
347-354.
-Jaffe, J. H. (1985). Drug addiction and drug abuse. En: The Pharmacological Basis of Therapeutics. Gilman A.
G., Goodman L. S y Rall T. W (Eds). 7a Ed, MacMillan, New York, pp. 522–573.
-Jamieson M. E., Coutts J. R. T. y Connor J. M. (1994). The chromosome constitution of human
preimplantation embryos fertilized in vitro. Hum Reprod 9 (4), 709-715.
-Jauniaux E., Gulbis B. y Burton G. J. (2003). Physiological implications of the materno-fetal oxygen gradient
in human early pregnancy. Reprod Biomed Online 7 (2), 250-253.
-Javed Q., Fleming T.P., Hay M. y Citi S. (1993). Tight junction protein cingulin is expressed by maternal and
embryonic genomes during early mouse development. Development 117 (3), 1145.
-Johnson M. H. y McConnell J. M. L. (2004). Lineage allocation and cell polarity during mouse
embryogenesis. Semin Cell Dev Biol 15 (5), 583-597.
-Jollie W. P. (1990). Development, morphology, and function of the yolk-sac placenta of laboratory rodents.
Teratology 41 (4), 361-381.
-Jones K. L. y Smith D. W. (1973). Recognition of the fetal alcohol syndrome in early infancy. Lancet 302
(7836), 999-1001.
-Jones K. L., Smith D. W., Ulleland C. N. y Streissguth A. P. (1974). Pattern of malformation in offspring of
chronic alcoholic mothers. Lancet 1 (7815), 1267-71.
-Jurisicova A., Varmuza S. y Casper R. F. (1995). Involvement of programmed cell death in preimplantation
embryo demise. Hum Reprod Update 1 (6), 558-566.

Bibligrafía
323
-Jurisicova A., Varmuza S. y Casper R.F. (1996). Programmed cell death and human embryo fragmentation.
Mol Hum Reprod 2 (2), 93-98.
-Jurisicova A, Rogers I., Fasciani A., Casper R.F. y Varmuza S. (1998a). Effects of maternal age and
conditions of fertilization on programmed cell death during murine preimplantation embryo development.
Mol Hum Reprod 4, 139-145.
-Jurisicova A., Latham E., Casper R. F. y Varmuza S.L. (1998b). Expression and regulation of genes associated
with cell death during murine preimplantation embryo development. Mol Reprod Dev 51 (3), 243–253.
-Jurisicova A. y Acton B. M. (2004). Deadly decisions: the role of genes regulating programmed cell death in
human preimplantation embryo development. Reproduction 128 (3), 281–291.
-Kalmus G. W. y Buckenmaier CC. (1989). Effects of ethanol and acetaldehyde on cultured pre-implantation
mouse embryos. Experientia 45 (5), 484-487.
-Kan N. G., Stemmler M. P., Junghans D., Kanzler B, de Vries W. N., Dominis M y Kemler R. (2007). Gene
replacement reveals a specific role for E-cadherin in the formation of a functional trophectoderm.
Development 134 (1), 31-41.
-Kato S., Alderman J. y Lieber C. S. (1988). In vivo role of the microsomal ethanol-oxidizing
system in ethanol metabolism by deermice lacking alcohol dehydrogenase. Biochem Pharmacol 37 (13),
2706-2708.
-Katz S. y Abrahamshon P. A. (1987). Involution of the antimesometrial decidua in the mouse. An
ultrastructural study. Anat Embryol 176 (2), 251-258.
-Kaufman M. H. (1983a). Ethanol-induced chromosomal abnormalities at conception. Nature 302 (5205),
258-260.
-Kaufman M. H. (1983b). The origin, properties and fate of trophoblast in the mouse. En: Biology of
Trophoblast. Loke Y. W. y Whyte A. (Eds). Amsterdam: Elsevier, pp 401-467.
-Kaufman M. H. y Bain I. M. (1984a). Influence of ethanol on chromosome segregation during the first and
second meiotic divisions in the mouse egg. J Exp Zool 230 (2), 315-320.
-Kaufman M. H. y Bain I. M. (1984b). The development potential of ethanol-induced monosomic and
trisomic conceptus in the mouse. J Exp Zool 231 (1), 149-155.
-Kaufman M. H. (1985). An hypothesis regarding the origin of aneuploidy in man: indirect evidence from an
experimental model. J Med Genet 22 (3), 171-178.
-Kaufman M. H. (1997). The teratogenic effects of alcohol following exposure during pregnancy and its
influence on the chromosome constitution of the pre-ovulatory egg. Alcohol Alcohol 32 (2), 113-128.
-Kaufman M. H. y Bard J. B. L. (1999). The anatomical basis of mouse development. Edinburgh, United
Kingdom, Academic Press.
-Kayani M. A. y Parry J. M. (2010). The in vitro genotoxicity of ethanol and acetaldehyde. Toxicol in Vitro 24
(1), 56-60.
-Kaye P. L. (1997). Preimplantation growth factor physiology. Rev Reprod 2 (2), 121–127.
-Keiver K., Herbert L. y Weinberg J. (1996). Effect of maternal ethanol consumption on maternal and fetal
calcium metabolism. Alcohol Clin Exp Res 20 (7), 1305-1312.
-Keiver K y Weinberg J. (2004). Effect of duration of maternal alcohol consumption on calcium metabolism
and bone in the fetal rat. Alcohol Clin Exp Res 28 (3), 456-467.
-Kelly S. J. (1975). Studies of potency of the early cleavage blastomeres of the mouse. En: The early
development of mammals. Balls M. y Wild A. E. (Eds). Cambridge: Cambridge University Press, pp 97-107.
-Kelly S. J. (1979). Studies of the developmental potential of 4- and 8-cell stage mouse blastomeres. J. Exp
Zool 200 (3), 365-376.
-Kelly S. M., Robaire B. y Hales B. F. (1992). Paternal cyclophosphamide treatment causes postimplantation
loss via inner cell mass-specific cell death. Teratology 45 (3), 313-318.
-Kemler R., Babinet C., Eisen H. y Jacob, F. (1977). Surface antigen in early differentiation. Proc Natl Acad Sci
U S A 74 (10), 4449-4452.
-Kennedy T. G. (1977). Evidence for a role for prostaglandins in the initiation of blastocyst implantation in
the rat. Biol Reprod 16, 286–291.
-Kenneth R., Warren y Richard J. B. (1988). Alcohol-Related Birth Defects: an Update 103 (6), 638-642.
-Kerr J. R. F., Wyllie A. H. y Currie A. R. (1972). Apoptosis: a basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer 26 (4), 239-257.

Bibligrafía
324
-Khong T. Y., De Wolf F., Robertson W. B. y Brosens I. (1986). Inadequate maternal vascular response to
placentation in pregnancies complicated by pre-eclampsia and by small-for-gestational age infants. Br J
Obstet Gynaec 93, 1049–1059.
-Khong T. Y., Liddell H. S. y Robertson W. B. (1987). Defective haemochorial placentation as a cause of
miscarriage: a preliminary study. Br J Obstet Gynaecol 94, 649–655.
-Kim S. W., Park J. I., Spring C. M., Sater A. K., Ji H., Otchere A. A., Daniel J. M. y McCrea P. D. (2004). Non-
canonical Wnt signals are modulated by the Kaiso transcriptional repressor and p120-catenin. Nat Cell Biol 6
(12), 1212-1220.
-Kirby D. R. S. (1960). Development of mouse eggs beneath the kidney capsule. Nature 187, 707-708.
Kiso Y., Yamashiro S., McBey B. A. y Croy B. A. (1992). Tissue-specific differentiation of a natural killer cell
subset in ectopically grafted murine uterine tissue. Transplantation 54 (1), 185-187.
-Klein R. F. (1997). Alcohol-induced bone disease: impact of ethanol on osteoblast proliferation. Alcohol Clin
Exp Res 21 (3), 392-399.
-Knipp G. T., Audus K. L. y Soares M. J. (1999). Nutrient transport across the placenta. Adv Drug Deliv Rev 38
(1), 41-58.
-Koi H., Tachi C., Tojo H., Kubota T. y Aso T. (1995). Effects of matrix proteins and heparin-binding
components in fetal bovine serum upon the proliferation of ectoplacental cone cells in mouse blastocysts
cultured in vitro. Biol Reprod 52 (4), 759-770.
-Koshikawa N., Giannelli G., Cirulli V., Miyazaki K. y Quaranta V. (2000). Role of cell surface
metalloprotease MT1-MMP in epithelial cell migration over laminin-5. J Cell Biol 148 (3), 615-624.
-Kotch L. E. y Sulik K. K. (1992). Experimental fetal alcohol syndrome: proposed pathogenic basis for a
variety of associated facial and brain anomalies. Am J Med Genet 44 (2), 168-176.
-Kotch L. E., Chen S. Y. y Sulik K. K. (1995). Ethanol-induced teratogenesis: free radical damage as a possible
mechanism. Teratology 52 (3), 128-136.
-Kotova N., Vare D., Schultz N., Meesters D. G., StepnikM., Grawé J., Helleday T., Jenseen D. (2013).
Genotoxicity of alcohol is linked to DNA replication-associated damage and homologos recombination
repair. Carcinogenesis 34 (2), 325-330.
-Kowalczyk C. L., Stachecki J. J., Shultz J. F., Leach R. E. y Randall C. A. (1996). Effects of alcohol on murine
preimplantation development: relationship to relative membrane disordering potency. Alcohol Clin Exp Res
20 (3), 566-571.
-Krueger A. W., Bo W. J. y Rudeen P. K. (1982). Female reproduction during chronic ethanol consumption in
rats. Pharmacol Biochem Behav 17 (4), 629-631.
-Kucheira K., Taneja N. y Mohan D. (1986). Chromosomal aberrations and sister chromatid exchanges in
chronic male alcoholics. Indian J Med Res 83, 417-421.
-Gonzalez-Mariscal A., Betanzos A. y Avila-Flores. (2000). MAGUK proteins: structure and role in the tight
junction. Semin Cell Dev Biol 11 (4), 315-324.
-Lane M. y Gardner D. K. (1997). Differential regulation of mouse embryo development and viability by
amino acids. J Reprod Fetil 109 (1), 153-164.
-Larsen W. J. (1993). Human Embryology. Singapore, Churchill Livingstone.
-Larue L., Ohsugi M., Hirchenhain J. y Kemler R. (1994). E-cadherin null mutant embryos fail to form a
trophectoderm epithelium. Proc Natl Acad Sci U S A. 91 (17), 8263-8267.
-Larue L., Antos C., Butz S., Huber O., Delmas V., Dominis M. y Kemler R. (1996). A role for cadherins in
tissue formation. Development 122 (10), 3185-3194.
-Lau C. F., Vogel R., Obe G. y Spielmann H. (1991). Embryologic and cytogenetic effects of ethanol on
preimplantation mouse embryos in vitro. Reprod Toxicol 5 (5), 405-510.
-Lawson K. A., Meneses J. J. y Pedersen R. A. (1991). Clonal analysis of epiblast fate during germ layer
formation in the mouse embryo. Development 113 (3), 891-911.
-Leach R. E., Stachecki J. J. y Armant D. R. (1993). Development of in vitro fertilized mouse embryos
exposed to ethanol during the preimplantation period: accelerated embryogenesis at subtoxic levels.
Teratology 47 (1), 57-64.
-Lee M. y Wakabayashi K. (1985). Hormonal changes in rats consuming alcohol prior to and during
gestation. Alcohol Clin Exp Res 9 (5), 417-420.

Bibligrafía
325
-Lee D. M., Osathanondh R. y Yeh J. (1988). Localization of Bcl-2 in the human fetal mullerian tract. Fertil
Steril 70 (1), 135-140.
-Lee S., Jilani S.M., Nikolova G.V., Carpizo D. y Iruela-Arispe M.L. (2005). Processing of VEGF-A by matrix
metalloproteinases regulates bioavailability and vascular patterning in tumors. J Cell Biol 169, 681-691.
-Leese H. J. (1991). Metabolism of the preimplantation mammalian embryo. Oxf Rev Reprod Biol 13, 35-72.
-Leese H. J., Donnay I. y Thompson J. G. (1998). Human assisted conception: a cautionary tale. Lessons from
domestic animals. Hum Reprod 13 Suppl 4, 184-202.
-Leese H. J. (1993). Energy metabolism in preimplantation development. En: Preimplantation Embryo
Development, Bavister B.D. (ed), Oxford: Oxford University Press pp 35-72.
-Lemoine P., Harousseau H., Borteyru J. P., Menuet J. C. (1968). Children of alcoholic parents: Anomalies
observed in 127 cases. Quest Medicale 25, 476–82.
-Levine E., Lee C. H., Kintner C. y Gumbiner B. M. (1994). Selective disruption of E-cadherin function in early
Xenopus embryos by a dominant negative mutant. Development 120 (4), 901-909.
-Lewis S. L. y Tam P. P. L. (2006). Definitive endoderm of the mouse embryo: formation, cell fates, and
morphogenetic function. Dev Dyn 235 (9), 2315-2329.
-Lieber C. (1985). Alcohol and the liver metabolism of ethanol, metabolic effects and pathogenesis of injury.
Acta Med Scand Suppl 703, 1-55.
-Lieber C.S., Decarli L. M. y Sorrell M. F. (1989). Experimental methods of ethanol administration.
Hepatology 10 (4), 501–510.
-Lieber C. S. y Decarli L. M. (1991). Hepatotoxicity of ethanol. J Hepatol 12 (3), 394–401.
-Lieber C. S. (1997). Ethanol metabolism, cirrosis and alcoholism.Clin Chim Acta 257 (1), 59-84.
-Lieber C.S. (2003). Relationships between nutrition, alcohol use, and liver disease. Alcohol Res Health 27
(3), 220-231.
-Lien W.H., Klezovitch O., Vasioukhin V. (2006). Cadherin-catenin proteins in vertebrate development. Curr
Opin Cell Biol 18 (5), 499-506.
-Lim H. J. y Wang H. (2010). Uterine disorders and pregnancy complications: insights from mouse models. J
Clin Invest 120 (4), 1004-1015.
-Lin H., Mossmann T. R., Guilbert L., Tuntipopipat S. y Wegnann T. G. (1993). Synthesis of T-helper-2-type
cytokines at the maternal-fetal interface. J Immunol 151 (9), 4562-4573.
-Liu L., Trimarchi J. R. y Keefe D. L. (1999). Thiol oxidation-induced embryonic cell death in mice is
prevented by the antioxidant dithiothreitol. Biol Reprod 61 (4), 1162-1169.
-Liu L., Trimarchi J. R. y Keefe L. D. (2000). Involvement of Mitochondria in Oxidative Stress-Induced Cell
Death in Mouse Zygotes. Biol Reprod 62 (6), 1745-53.
-Liu Y., Balaraman Y., Wang G., Nephew K. P. y Zhou F. C. (2009). Alcohol exposure alters DNA methylation
profiles in mouse embryos at early neurulation. Epigenetics 4 (7), 500–511.
-Lochter A., Galosy S., Muschler J., Freedman N., Werb Z. y Bissell M.J. (1997). Matrix metalloproteinase
stromelysin-1 triggers a cascade of molecular alterations that leads to stable epithelial-to mesenchymal
conversion and a premalignant phenotype in mammary epithelial cells. J Cell Biol 139 (7), 1861-1872.
-Lodish H., Baltimore D., Berk A., Zipursky S. L., Matsudaira P. y Darnell J. (1998). Molecular cell biology.
New York: Scientific American Books, Inc.
-Lois M., Brown L. A., Moss I. M., Roman J., Guidot D.M. (1999). Ethanol ingestion increases activation of
matrix metalloproteinases in rat lungs during acute endotoxemia. Am J Respir Crit Care Med 160 (4), 1354-
1360.
-Lopata A., Kohlman D. J. y Kellow G. N. (1982). The fine structure of human blastocysts developed in
culture. En: Embryonic Development, Part B: Cellular Aspects. Alan R Liss (Ed). New York, pp 69-85.
-López M. C., Roubicek M. y Arzeno M. (2001). Chronic alcohol ingestion and chromosomal aberrations. A
popular study in Mar del Plata, Argentina. J. Basic Applied Genetics 14 (1), 1-4.
-Lopez M. F., Dikkes P., Zurakowski D. y Villa-Komaroff L. (1996). Insulin-like growth factor II affects the
appearance and glycogen content of glycogen cells in the murine placenta. Endocrinology 137 (5), 2100-
2108.
-Lorenti Garcia C., Mechilli M., Proietti De Santis L., Schinoppi A., Kobos K. y Palitti F. (2009). Relationship
between DNA lesions, DNA repair and chromosomal damage induced by acetaldehyde. Mut Res 662 (1-2), 3-
9.

Bibligrafía
326
-Lumey L. H. (1992). Decreased birthweights in infants after maternal in utero exposure to the Dutch famine
of 1944–1945. Paediatr Perinat Epidemiol 6 (2), 240-253.
-Lumey L. H. y Stein A. D. (1997). Offspring birth weights after maternal intrauterine undernutrition: a
comparison within sibships. Am J Epidemiol 146 (10), 810-819.
-Luo J., y Miller M.W. (2000). Ethanol enhances erbB-mediated migration of human breast cancer cells in
culture. Breast Cancer Res Treat 63 (1), 61-69.
-Luzhna L., Kathiria P., Kovalchuk O. (2013). Micronuclei in genotoxicity assessment: from genetics to
epigenetics and beyond. Front Genet 4, 1-17.
-MacAuley A., Cross J. C. y Werk Z. (1998). Reprogramming the cell cycle for endoreduplication in rodent
trophoblast cells. Mol Biol Cell 9, 795-807.
-Madan P., Rose K. y Watson A. J. (2007). Na/K-ATPase beta1 subunit expression is required for blastocyst
formation and normal assembly of trophectoderm tight junction-associated proteins. J Biol Chem 282 (16),
12127-12134.
-Maden M., Graham A., Gale E., Rollinson C. y Zile M. (1997). Positional apoptosis during vertebrate CNS
development in the absence of endogenous retinoids. Development 124 (14), 2799-805.
-Maffei F., Fimognari C., Castelli E., Stefanini G. F., Forti G. C. y Hrelia P. (2000). Increased cytogenetic
damage detected by FISH analysis on micronuclei in peripheral lymphocytes from alcoholics. Mutagenesis 15
(6), 517-523.
-Majno G. y Joris I. (1995). Apoptosis, oncosis, and necrosis. An overview of cell death. Am J Pathol 146 (1),
3–15.
-Malassiné A., Frendo J. L. y Evain-Brion D. (2003). A comparison of placental development and endocrine
functions between the human and mouse model. Hum Reprod Update. 9 (6), 531-539.
-Manaseki S. y Searle R.F. (1989). Natural killer (NK) cell activity of first trimester human decidua. Cell
Immunol 121 (1), 166-173.
-Mancinelli R., Ceccanti M., Laviola G. (2007). Fetal alcohol spectrum disorders (FASD): from experimental
biology to the search for treatment. Neurosci Biobehav Rev 31 (2), 165-167.
-Manejwala F. M., Cragoe E. J. Jr y Schultz R. M. (1989). Blastocoel expansion in the preimplantation mouse
embryo: role of extracellular sodium and chloride and possible apical routes of their entry. Dev Biol 133 (1),
210-220.
-Martin L. y Finn C. A. (1968). Hormonal regulation of cell division in epithelial and connective tissue of
mouse uterus. J Endocrinol 41 (3), 363-371.
-Martin S. J., Reutelingsperger C. P. M., McGahon A. J., Rader J. A., van Schie R. C. A. A., LaFace D. M. y
Green D. R. (1995). Early redistribution of plasma membrane phosphatidylserine is a general feature of
apoptosis regardless of the initiating stimulus: inhibition by overexpression of Bcl-2 and Abl. J Exp Med 182
(5), 1545-1556.
-Marzusch K., Buchholz F., Ruck P., Handgretinger R., Geiselhart A., Engelmann L. y Dietl J. (1997).
Interleukin-12- and interleukin-2-stimulated release of interferon-gamma by uterine CD56++ large granular
lymphocytes is amplified by decidual macrophages. Hum Reprod. 12 (5), 921-924.
-May P. A., Fiorentino D., Phillip G. J., Kalberg W. O., Eugene H. H., Robinson L. K. y col. (2006).
Epidemiology of FASD in a province in Italy: prevalence and characteristics of children in a random sample of
schools. Alcohol Clin Exp Res 30 (9),1562-1575.
-Mayhew T. M., Jackson M. R. y Haas J. D. (1986). Placental morphology and oxygen diffusion. Placenta 7
(2), 121-131.
-McLaren A. y Hensleigh H. C. (1975). Culture of mammalian embryos over the implantation period. En: The
early development of mammals. Balls M. y Wild A. E. (Eds). Cambridge, Cambridge Univ Press, pp 45-60.
-Mello N. K., Bree M. P., Mendelson J. H., Ellingobe J., King N. W. y Sehgal P. (1983). Alcohol self-
administration disrupts reproductive function in female macaques monkeys. Sciences 221 (4611), 677-679.
-Mello N. K. (1988). Effects of alcohol abuse on reproductive function in women. Recent Dev Alcohol 6, 253-
276.
-Mendelson J. H. y Mello N. K. (1988). Chronic alcohol effects on anterior pituitary and ovarian hormones in
healthy women. J. Pharmacol Exp Ther 245 (2), 407-412.
-Meng Q., Gao B., Goldberg I. D., Rosen E. M. y Fan S. (2000). Stimulation of cell invasion and migration by
alcohol in breast cancer cells. Biochem Biophys Res Commun 273 (2), 448-453.

Bibligrafía
327
-Menger H., Lin A. E., Toriello H. V., Bernert G. y Spranger J. W. (1997). Vitamin K deficiency embryopathy:
a phenocopy of the warfarin embryopathy due to a disorder of embryonic vitamin K metabolism. Am J Med
Genet 72 (2), 129-134.
-Mirkes P. E., Wilson K. L. y Cornel L. M. (2000). Teratogen-induced activation of ERK, JNK, and p38 MAP
kinases in early postimplantation murine embryos. Teratology 62 (1), 14-25.
-Mirkes P. E. (2002). 2001 Warkany lecture: to die or not to die, the role of apoptosis in normal and
abnormal mammalian development. Teratology 65 (5), 228-239.
-Mishra S. R., Sahai A., Srivastava A. K., Agrawal A. K., Singh P. J. y Mishra R. K. (2003). Skeletal anomalies
in fetal alcohol syndrome: a study on developing mice embryos. J Anat Soc India 52 (1), 51-44.
-Mohr L.R. y Trounson A. O. (1982). Comparative ultrastructure of hatched human, mouse and bovine
blastocysts. J Reprod Fertil 66 (2), 499-504.
-Mooney S. M. y Miller M. W. (2007). Time-specific effects of ethanol exposure on cranial nerve nuclei:
gastrulation and neurogenesis. Exp Neurol 205, 56–63.
-Mori C., Nakamura N., Kimura S., Irie H., Takigawa T. y Shiota K. (1995). Programmed cell death in the
interdigital tissue of the fetal mouse limb is apoptosis with DNA fragmentation. Anat Rec 242 (1), 103-110.
-Moriwaki K., Tsukita S. y Furuse M. (2007). Tight junctions containing claudin 4 and 6 are essential for
blastocyst formation in preimplantation mouse embryos. Dev Biol 312 (2), 509–522.
-Mossman H. W. (1987). Vertebrate fetal membranes. New Brusnswich N. J. Rutgers University Press.
-Motosugi N., Bauer T., Polanski Z., Solter D. y Hiiragi T. (2005). Polarity of the mouse embryo is established
at blastocyst and is not prepatterned. Genes Dev 19 (9), 1801-1892.
-Mudry M. D. y Abrevaya X. (2006). En: Genética toxicológica. Mudry M. D. y Carballo M. A. Buenos Aires:
De los Cuatro Vientos, pp 277-298.
-Mudry M. D. y Greizerstein (2006). En: Genética toxicológica. Mudry M. D. y Carballo M. A. Buenos Aires:
De los Cuatro Vientos, pp 57-81.
-Müntener M. y Hsu Y. C. (1977). Development of trophoblast and placenta of the mouse. Acta Anat 98 (3),
241-252.
-Munné S. y Cohen J. (1998). Chromosome abnormalities in human embryos. Hum Reprod Update 4 (6),
842-855.
-Musacchio A., Hardwick K. G. (2002). The spindle checkpoint: structural insights into dynamic signaling. Nat
Rev Mol Cell Biol 3 (10), 731-741.
-Nakatsuji N. y Johnsson K. E. (1984). Effects of ethanol on the primitive streak stage mouse embryo.
Teratology 29 (3), 369-375.
-Narayanan V. (1997). Apoptosis in development and disease of the nervous system. Naturally occurring cell
death in the developing nervous system. Pediatr Neurol 16 (1), 9-13.
-Neuber E., Rinaudo P., Trimarchi J. R. y Sakkas D. (2003). Sequential assessment of individually cultured
human embryos as an indicator of subsequent good quality blastocyst development. Hum Reprod 18 (6),
1307-1312.
-Neumark Y. D., Friedlander Y., Durst R., Leitersdorf E., Jaffe D., Ramchandani V. A., O' Connor S., Carr L. G.
y Li T. K. (2004). Alcohol dehydrogenase polymorphisms influence alcohol-elimination rates in
a male Jewish population. Alcohol Clin Exp Res 28 (1), 10-14.
-Niwa H., Toyooka Y., Shimosato D., Strumpf D., Takahashi K., Yagi R. y Rossant J. (2005). Interaction
between Oct3/4 and Cdx2 determines trophectoderm differentiation. Cell 123, 917-929.
-Noden D. M. (1991). Vertebrate craniofacial development: The relation between ontogenetic process and
morphological outcome. Brain Behav Evol 38 (4-5), 190–225.
-Noe V., Fingleton B., Jacobs K., Crawford H.C., Vermeulen S., Steelant W., Bruyneel E., Matrisian L.M. y
Mareel M. (2001). Release of an invasion promoter E-cadherin fragment by matrilysin and stromelysin-1. J
Cell Sci 114 (Pt 1), 111-118.
-Nogales F.F., Beltran E. y Gonzalez F. (1993). Morphological changes of the secondary human yolk sac in
early pregnancy wastage. En: The Human Yolk Sac and Yolk Sac Tumors. Nogales F. F. (Ed.). Springer-Verlag,
Heidelberg, Germany, pp 174-194.
-Nordmann R., Ribière C. y Rouach H. (1992). Implication of free radical mechanisms in ethanol-induced
cellular injury. Free Radic Biol Med 12 (3), 219-240.
-Obe G. y Anderson D. (1987). Genetic effects of ethanol. Mutat Res 186 (3), 177-200.

Bibligrafía
328
-Ohsugi M., Larue L., Schwarz H. y Kemler R. (1997). Cell-junctional and cytoskeletal organization in mouse
blastocysts lacking E-cadherin. Dev Biol 185 (2), 261-271.
-Opas M. (1989). Expression of differentiated phenotype by epithelial cells in vitro is regulated by both
biochemistry of the substratum. Dev Biol 131, 281-293.
-Ornoy A. y Ergaz Z. (2010). Alcohol Abuse in Pregnant Women: Effects on the fetus and newborn, mode of
action and maternal treatment. Int J Environ Res Public Health 7 (2), 364-379.
-Oyedele O. O. y Kramer B. (2008). Acute ethanol administration causes malformations but does not affect
cranial morphometry in neonatal mice. Alcohol 42 (1), 21-27
-Pacchierotti F., Tiveron C., D’Archovio M., Bassani B., Cordelli E., Leter G. Y Spano M. (1994). Acrylamide-
induced chromosomal damage in male mouse germ cells detected by cytogenetic analysis of one-cell
zygotes. Mutat Res 309, 273-284.
-Padmanabhan R. y Hameed M.S. (1988). Effects of acute doses of ethanol administered at pre-
implantation stages on fetal development in the mouse. Drug Alcohol Depend 22 (1-2), 91-100.
-Padmanabhan R. y Shafiullah M. (2001). Intrauterine growth retardation in experimental diabetes: possible
role of the placenta. Arch Physiol Biochem 109 (3), 260-271.
-Pampfer S., De Hertogh R., Vanderheyden I., Michiels B. y Vercheval M. (1990a). Decreased inner cell
mass proportion in blastocysts from diabetic rats. Diabetes 39 (4), 471-476.
-Pampfer S., Vanderheyden I., Michiels B. y De Hertogh R. (1990b). Cell allocation to the inner cell mass and
the trophectoderm in rat embryos during in vivo preimplantation development. Roux´s Arch Dev Biol 198,
257-263.
-Pampfer S. (2000). Apoptosis in rodent peri-implantation embryos: differential susceptibility of inner cell
mass and trophectoderm cell lineages-a review. Placenta 21 (Suppl A), S3-10.
-Pardi G., Marconi A. M. y Cetin I. (2002). Placental-fetal interrelationship in IUGR fetuses-a review.
Placenta 23 (Suppl A), S136–S141.
-Paris L., Tonutti L., Vannini C. y Bazzoni. (2008). Structural organization of the tight junctions. Biochim
Biophys Acta 1778 (3), 646-659.
-Parnell S. E., Dehart D. B., Wills T. A., Chen S. Y., Hodge C. W., Besheer J., Waage-Baudet H. G., Charness
M. E. y Sulik K. K. (2006). Maternal oral intake mouse model for fetal alcohol spectrum disorders: ocular
defects as a measure of effect. Alcohol Clin Exp Res 30 (10), 1791-1798.
-Parnell S. E., O'Leary-Moore S. K., Godin E. A., Dehart D. B., Johnson B. W., Allan Johnson G., Styner M. A.
y Sulik K. K. (2009). Magnetic resonance microscopy defines ethanol-induced brain abnormalities in prenatal
mice: effects of acute insult on gestational day 8. Alcohol Clin Exp Res 33 (6), 1001-1011.
-Parr E. L., Tung H. N. y Parr M. B. (1987). Apoptosis as the mode of uterine epithelial cell death during
embryo implantation in mice and rats. Biol Reprod 36 (1), 211-225.
-Parr E. L. y Parr M. B. (1989a). Epithelial cell death during rodent embryo implantation. En: Blastocyst
implantation. Yoshinaga K. (Ed). Boston, Adams, pp 105-115.
-Parr E. L. y Parr M. B. (1989b). The implantation reaction. En: Biology of the Uterus. Wynn R. N. y Jollie W.
P. (Eds). New York, Plenum, pp 233-277.
-Partridge C. R., Sampson H. W. y Forough R. (1999). Long-term alcohol consumption increases matrix
metalloproteinase-2 activity in rat aorta. Life Sci 65 (13), 1395-13402.
-Payne D., Flaherty S. P., Barry M. F. y Matthews C. D. (1997). Preliminary observations on polar body
extrusion and pronuclear formation in human oocytes using time-lapse video cinematography. Hum
Reprod 12 (3): 532-541.
-Peel S. y Bulmer D. (1977). Proliferation an differentiation of trophoblast in the establishment of the rat
chorio-allantoic placenta. J Anat 124 (3), 675-687.
-Peinado H., Portillo F. y Cano A. (2004). Transcriptional regulation of cadherins during development and
carcinogenesis. Int J Dev Biol 48 (5-6), 365-375.
-Pelinck M. J., Hoek A., Simons A. H., Heineman M. J., van Echten-Arends J. y Arts E. G. (2010).
Embryo quality and impact of specific embryo characteristics on ongoing implantation in unselected embryo
sderived from modified natural cycle in vitro fertilization. Fertil Steril 94 (2), 527-534.
-Pellestor F., Girardet A., Andréo B., Arnal F. y Humeau C. (1994). Relationship between morphology and
chromosomal constitution in human preimplantation embryo. Mol Reprod Dev 39 (4), 141-146.

Bibligrafía
329
-Penaloza C., Lin L., Lockshin R. A. y Zakeri Z. (2006). Cell death in development: shaping the embryo.
Histochem Cell Biol 126 (2), 149–158.
-Pereira P. N. G., Dobreva M. P., Graham L., Huylebroeck D., Lawson K. A. y Zwijsen A.N. (2011). Amnion
formation in the mouse embryo: the single amniochorionic fold model. BMC Dev Biol 11, 48-60.
-Perez M. J., Velasco E., Monte M. J., Gonzalez-Buitrago J. M. y Marin J. J. (2006). Maternal ethanol
consumption during pregnancy enhances bile acid-induced oxidative stress and apoptosis in fetal rat liver.
Toxicology 225(2-3), 183-194.
-Pérez-Tito L., Bevilaqcua E. y Cebral E. (2014). Peri-implantational in vivo and in vitro embryo-trophoblast
development after perigestational alcohol exposure in the CD-1 mouse. Drug Chem Toxicol 37 (2), 184-197.
-Pérez-Tito L., Sobarzo C., Coll T. A., Ventureira M. R. y Cebral E. (2013). La ingesta periconcepcional
moderada de alcohol altera las uniones intercelulares de embriones peri-implantatorios murinos. Cs Morfol
15 (2), 1-16.
-Peters T. J., Albieri A., Bevilacqua E., Chapman B. M., Crane L. H., Hamlin G. P., Seiki M. y Soares M. J.
(1999). Differentiation-dependent expression of gelatinase B/matrix metalloproteinase-9 in trophoblast
cells. Cell Tissue Res 295 (2), 287-296.
-Phillips B. j. y Jenkinson P. (2001). Is ethanol genotoxic? A review of the published data. Mutagenesis 16
(2), 91-101.
-Pierce G. B., Lewellyn A. L. y Parchment R. E. (1989). Mechanism of programmed cell death in the
blastocyst. Proc Natl Acad Sci U S A 86 (10), 3654-3658.
-Pijnenborg R., Anthony J., Davey D. A., Rees A., Tiltman A., Vercruysse L. y van Assche A. (1991). Placental
bed spiral arteries in the hypertensive disorders of pregnancy. Br J Obstet Gynaecol 98 (7), 648-655.
-Pijnenborg R., Robertson W. B. y Brosens I. (1985). Morphological aspects of placental ontogeny and
phylogeny. Placenta 6 (2), 155-162.
-Plachot M. y Mandelbaum J. (1990). Oocyte maturation, fertilization and embryonic growth in vitro. Br
Med Bull 46 (3), 675–694.
-Plaks V., Rinkenberger J., Dai J., Flannery M., Sund M., Kanasaki K., Ni W., Kalluri R., y Werb Z. (2013).
Matrix metalloproteinase-9 deficiency phenocopies features of preeclampsia and intrauterine growth
restriction. PNAS 110 (27), 11109–11114.
-Potts D. M. (1966). The attachment phase of ovoimplantation. Am J Obstetr Gynecol 96 (8), 1122-1128.
-Potts D. M. (1968). The ultrastructure of implantation in the mouse. J Anat 103, 77-90.
-Potts D. M. (1969). The ultrastructure of egg implantation. Adv Reprod Physiol 4, 241-267.
-Prater M. R., Laudermilch C.L., Liang C. y Holladay S.D. (2008). Placental oxidative stress alters expression
of murine osteogenic genes and impairs fetal skeletal formation. Placenta 29 (9), 802–808.
-Pratt H. P. M. (1987). Isolation, culture and manipulation of pre-implantation mouse embryos. En:
Mammalian development a practical aproach. Monk M. (Ed). Oxford, IRL Press, pp 13-42.
-Pratt H. P. M., Ziomek C. A., Reeve W. J. D. y Johnson M. H. (1982). Compaction of the mouse embryo: An
analysis of its components. J Embryol Exp Morphol 70, 113-132.
-Pringle K. G. y Roberts C. T. (2007). New light on early post-implantation pregnancy in the mouse: roles for
insulin-like growth factor-II (IGF-II)? Placenta 28 (4), 286-297.
-Qian X., Karpova T., Sheppard A. M., McNally J. y Lowy D. R. (2004). E-cadherin-mediated adhesion inhibits
ligand-dependent activation of diverse receptor tyrosine kinases. EMBO J 23 (8), 1739-1748.
-Ralston A. y Rossant J. (2008). Cdx2 acts downstream of cell polarization to cell-autonomously promote
trophectoderm fate in the early mouse embryo. Dev Biol 313 (3), 614-629.
-Randall C. L. y Taylor W. J. (1979). Prenatal ethanol exposure in mice: teratogenic effects. Teratology 19 (3),
305-311.
-Randall C. L. (1987). Alcohol as a teratogen: A decade of research in review. Alcohol Alcohol Suppl 1, 125-
132.
-Ray J. G. y Blom H. J. (2003). Vitamin B12 insufficiency and the risk of fetal neural tube defects. Q J Med 96
(4), 289-295.
-Redline R. W., Chernicky C. L., Tan H-Q., Ilan J. y Ilan J. (1993). Differential expression of insulin-like growth
factor-II in specific regions of the late (post 9.5) murine placenta. Mol Reprod Dev 36 (2), 121-129.
-Reinius S. (1965). Morphology of the mouse embryo from the time of implantation to mesoderm
formation. Z. Zellforsch Mikrosk Anat 68 (5), 711-723.

Bibligrafía
330
-Reinius S. (1967). Ultrastructure of blastocyst attachment in the mouse. Z Zellforsch Mikrosk Anat 77 (2),
257-266.
-Reynolds L. P. y Redmer D. A. (1995). Utero-placental vascular development and placental function. J Anim
Sci 73(6), 1839-1851.
-Rhodes J. S., Best K., Belknap J. K., Finn D. A. y Crabbe J. C. (2005). Evaluation of a simple model of ethanol
drinking to intoxication in C57BL/6J mice. Physiology and Behavior 84 (1), 53-63.
-Riley E. P. y McGee C. L. (2005). Fetal Alcohol Spectrum Disorders: An Overview with Emphasis on Changes
in Brain and Behavior. Exp Biol Med 230 (6), 357–365.
-Rodríguez D., Morrison C. J. y Overal C. M. (2010). Matrix metalloproteinases: what do they not do? New
substrates and biological roles identified by murine models and proteomics. Biochim Biophys Acta 1803 (1),
39–54.
-Rodriguez T. A., Sparrow D. B., Scott A. N., Withington S. L., Preis J. I., Michalicek J., Clements M., Tsang T.
E., Shioda T., Beddington R. S. P. y Dunwoodie S. L. (2004). Cited1 is required in trophoblasts for placental
development and for embryo growth and survival. Mol Cell Biol 24 (1), 228-244.
-Rogers P. A. W. y Murphy C. R. (1989). Uterine receptivity for implantation: human studies. En: Blastocyst
Implantation. Yoshinaga K. (Ed). Boston, Adams, pp 231-238.
-Rossett H. L., Weiner L. y Edelin K. C. (1981). Strategies for prevention of fetal alcohol effects. Obstet
Gynecol 57 (1), 1-7.
-Rosett H. L. (1980). Clinical perspective of tbe fetal alcohol syndrome. Alcohol Clin Exp Res 4 (2), 119-22.
-Ross M.H. y Pawlina W. (2007). Histología. Texto y atlas color con Biología Celular y Molecular. 5a ed. Ed
Médica Panamericana, pp 108-145.
-Rossant J. y Offer L. (1977). Properties of extra-embryonic ectoderm isolated from post-implantation
mouse embryos. J Embryol Exp Morphol 39, 183–194.
-Rossant J. y Tamura-Lis W. (1981). Effect of culture conditions on diploid to giant-cell transformation in
postimplantion mouse trophoblast. J Embryol Exp Morph 62, 217-227.
-Rossant J. (1986). Development of extraembryonic cell lineages in the mouse embryo. En: Experimental
approaches to mammalian embryonic development. Rossant J. y Pedersen R. A. (Eds). New York, Cambridge
University Press, pp 97-120.
-Rossant J. y Cross J. C. (2001). Placental development: lessons from mouse mutants. Nat Rev Genet 2 (7),
538-548.
-Rout U. K. y Armant D. R. (2002). Expression of genes for alcohol and aldehyde metabolizing enzymes in
mouse oocytes and preimplantation embryos. Reprod Toxicol 16, 253–258.
-Rout U. K. y Holmes R. S. (1996). Alcohol dehydrogenases and aldehyde dehydrogenases among inbred
strains of mice: multiplicity, development, genetic studies and metabolic roles. Addict Biol 1 (4), 349-62.
-Rubert G., Miñana R., Pascual M. y Guerri C. (2006). Ethanol exposure during embryogenesis decreases the
radial glial progenitorpool and affects the generation of neurons and astrocytes. J Neurosci Res 84 (3), 483-
496.
-Runic R., Lockwood C. J., LaChapelle L., Dipasquale B., Demopoulos R. I., Kumar A. y Guller S. (1998).
Apoptosis and Fas expression in human fetal membranes. J Clin Endocrinol Metab 83 (2), 660-666.
-Rutland C. S., Latunde-Dada A. O., Thorpe A., Plant R., Langley-Evans S. y Leach L. (2007). Effect of
gestational nutrition on vascular integrity in the murine placenta. Placenta 28 (7), 734-742.
-Sadler T.W. (1995). Langman Embriología médica. 7a Ed. Médica Panamericana y Williams y Wilkins Press,
Maryland, USA pp 115-133.
-Saitou M. y Yamaji M. (2010). Germ cell specification in mice: signaling, transcription regulation, and
epigenetic consequences. Reproduction 139 (6), 931-942.
-Saiz N. y Plusa B. (2013). Early cell fate decisions in the mouse embryo. Reproduction 145 (3), 65-80.
-Salewski E (1964). Farbemethode zum makrospischen Nachweis von Implantationsstellen am Uterus der
Ratte. Arch Exp Path Pharm 247, 367.
-Sampson P. D., Streissguth A. P., Bookstein F. L., Little R. E., Clarren S. K., Dehaene P., Hanson J. W. y
Graham J. M. (1997). Incidence of fetal alcohol syndrome and prevalence of alcohol-related
neurodevelopmental disorder. Teratology 56 (5), 317-326.
-Sampson P. D., Streissguth A. P., Bookstein F. L. y Barr H. M. (2000). On categorization in analyses of
alcohol teratogenesis. Environ Health Perspect 3, 421-28.

Bibligrafía
331
-Samson H. H. (1981). Maternal ethanol consumption and fetal development in the rat: a comparison of
ethanol exposure techniques. Alcohol Clin Exp Res 5 (1), 67-74.
-Sanchis R., Esquifino A. y Guerri C. (1985). Chronic ethanol intake modifies estrous cyclicity and alters
prolactin and LH levels. Pharmacol Biochem Behav 23 (2), 221-224.
-Sant´Anna L. B. y Tosello D. O. (2006). Fetal alcohol syndrome and developing craniofacial and dental
structures-a review. Orthod Craniofac Res 9 (4), 172–185.
-Santos N. S. S, Biscaro M. D. A, Beatriz Christina Lorenzetti Santos B. C. L. y Guimarães Moraes S. (2009).
Congenital malformations in embryos of female mice exposed to alcohol and nicotinamide. Einstein 7 (1 Pt
1), 52-57.
-Sasaki H. (2010). Mechanisms of trophectoderm fate specification in preimplantation mouse development.
Dev Growth Differ 52 (3), 263-273.
-Sathananthan A. H., Wood C. y Leeton J. F. (1982). Ultrastructural evaluation of 8–16 cell human embryos
cultured in vitro. Micron 13, 193–203.
-Savagner P. (2001). Leaving the neighborhood: molecular mechanisms involved during epithelial-
mesenchymal transition. Bioessays 23 (10), 912-923.
-Schuckit M. A. (2006). Drug and alcohol abuse: a clinical guide to diagnosis and treatment. Plenum Press 6a
Ed, New York.
-Schultz G. A. (1986). Utilization of genetic information in the preimplantation mouse embryo. En:
Experimental aproaches to mammalian preimplantation development. J Rossant R. A. Pedersen (Eds). New
York, Cambridge University Press, pp 239-266.
-Schultz R. M. (1993). Regulation of zygotic gene activation in the mouse. Bioessays 15 (8), 531-538.
-Schultz J. F. y Armant D. R. (1995). Beta1- y Beta3-class integrins mediate fibronectin binding activity at the
surface of developing mouse peri-implantation blatocysts. J Biol Chem 270 (19), 1522-1531.
-Sedelnikova O. A., Nakamura A., Kovalchuk O., Koturbash I., Mitchell S. A., Marino S.A. Brenner D. J.
y Bonner W. M. (2007). DNA double-strand breaks form in bystander cells after microbeam irradiation of
three-dimensional human tissue models. Cancer Res 67 (9), 4295-4302.
-Sharkey M. E., Adler R. R., Nieder G. L. y Brenner C. A. (1996). Matrix metalloproteinase expression during
mouse peri-implantation development. Am J Reprod Immunol 36 (2), 72-80.
-Sherman M. I. (1983). Endocrinology of rodent trophoblast cells. En: Biology of Trophoblast. Loke Y. W. y
Whyte A (Eds). Amsterdam, Elisevier, pp 401-467.
-Sheth B., Fesenko I., Collins J. E., Moran B., Wild A. E., Anderson J. M. y Fleming T. P. (1997). Tight junction
assembly during mouse blastocyst formation is regulated by late expression of ZO-1 alpha + isoform.
Development 124 (10), 2027-2037.
-Sheth B., Fontaine J. J., Ponza E., McCallum A., Page A., Citi S., Louvard D., Zahraoui A. y Fleming T.P.
(2000). Differentiation of the epithelial apical junctional complex during mouse preimplantation
development: a role for rab13 in the early maturation of the tight junction. Mech Dev 97 (1-2), 93-104.
-Sheth B., Nowak R. L., Anderson R., Kwong W. Y., Papenbrock T. y Fleming T. P. (2008). Tight junction
protein ZO-2 expression and relative function of ZO-1 and ZO-2 during mouse blastocyst formation. Exp Cell
Res 314 (18), 3356-3368.
-Shiina Y., Kaneda M., Matsuyama K., Tanaka K., Hiroi M. y Doi K. (1993). Role of the extracellular Ca2+ on
the intracellular Ca2+ changes in fertilized and activated mouse oocytes. J Reprod Fertil 97 (1), 143-150.
-Simmons D. G. y Cross J. C. (2005). Determinants of trophoblast lineage and cell subtype specification in
the mouse placenta. Dev. Biol. 284 (1), 12-24.
-Simmons D. G. Fortier A. L. y Cross, J. C. (2007). Diverse subtypes and developmental origins of trophoblast
giant cells in the mouse placenta. Dev Biol 304 (2), 567-578.
-Simmons D. G., Natale D. R. C., Begay V., Hughes M., Leutz A. y Cross J. C. (2008). Early patterning of the
chorion leads to the trilaminar trophoblast cell structure in the placental labyrinth. Development 135 (12),
2083-2091.
-Simpson M. E., Duggal S. y Keiver K. (2005). Prenatal ethanol exposure has differential effects on fetal
growth and skeletal ossification. Bone 36 (3), 521-532.
-Smith S. M. (1997). Alcohol-induced cell death in the embryo. Alcohol Health Res World 21 (4), 287-297.
-Snell G. D. y Stevens L. C. (1966). Early embryology. En: Biology of the Laboratory mouse. Snell G. D. (Ed).
New York, McGraw-Hill, pp 205-245.

Bibligrafía
332
-Snow M. E. y Keiver K. (2007). Prenatal ethanol exposure disrupts the histological stages of fetal bone
development. Bone 41 (2), 181-187.
-Soares M. J., Faria T. N., Roby K. F. y Deb S. (1991). Pregnancy and the prolactin family of hormones:
coordination of anterior pituitary uterine and placental expression. Endocr Rev 12 (4), 402-423.
-Soares M. J., Faria T. N., Hamlin G. P., Lu X. J. y Deb S. (1993). Trophoblast cell differentiation: expression
of the placental prolactin family. En: Trophoblast Cells: Pathways for maternal-embryonic communication.
Soares M. J., Handwerger S. y Talamantes F. (Eds). New York, Spring-Verlag, pp 43-65.
-Soares M. J., Chapman B. M., Easmussen C. A., Dai G., Kamei T. y Orwig K. E. (1996). Differentiation of
trophoblast endocrine cells. Placenta 17 (5-6), 277-289.
-Soares M. J. (2004). The prolactin and growth hormone families: pregnancy-specific hormones/cytokines at
the maternal-fetal interface. Reprod Biol Endocrinol 2, 51.
-Soltes B. A., Anderson R., Radwanska E. (1996). Morphologic changes in offspring of female mice exposed
to ethanol before conception. Am J Obstet Gynecol 175 (5): 1158-1162.
-Spadoni A. D., McGee C. L., Fryer S. L. y Riley E. P. (2007). Neuroimaging and fetal alcohol spectrum
disorders. Neurosci Biobehav Rev 31 (2), 239-245.
-Spanos S., Rice S., Karagiannis P., Taylor D., Becker D. L., Winston R. M., Hardy K. (2002). Caspase activity
and expression of cell death genes during development of human preimplantation embryos. Reproduction
124 (3), 353-363.
-Spindle A. I. y Pedersen R. A. (1973). Hatching, attachment, and outgrowth of mouse blatocysts in vitro:
fixed nitrogen requirements. J Exp Zool 186, 305-318.
-Stachecki J. J., Yelian F. D., Leach R. E. y Armant D. R. (1994a). Mouse blastocyst outgrowth and
implantation rates following exposure to ethanol or A23187 during culture in vitro. J Reprod Fetil 101 (3),
611-617.
-Stachecki J. J., Yelian F. D., Schultz J. F., Leach R. E. y Armant D. R. (1994b). Blastocyst cavitation is
accelerated by ethanol- or ionophore-induced elevation of intracellular calcium. Biol Reprod 50 (1), 1-9.
-Staun-Ram E. y Shalev E. (2005). Human trofoblast function during the implantation process. Reprod Biol
Endocrinol 3, 56-68.
-Stein Z. y Susser M. (1975). The Dutch famine, 1944–1945, and the reproductive process. I. Effects or six
indices at birth. Pediatr Res 9 (2), 70-76.
-Stevenson B. R., Siliciano J. D., Mooseker M. S. y Goodenough D. A. (1986). Identification of ZO-1: A high
molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J
Cell Biol 103 (3), 755-766.
-Straszewski-chavez S. L., Abrahams V. M. y Mor G. (2005). The role of apoptosis in
the regulation of trophoblast survival and differentiation during pregnancy. Endocr Rev 26 (7), 877-897.
-Streissguth A.P., Landesman-Dweyr S., Martin J. C. y Smith D. W. (1980). Teratogenic effects of alcohol in
humans and laboratory animals. Science 209 (4454), 353-361.
-Streissguth A. P., Barr H. M., Sampson P. D. y Bookstein F. L. (1994). Prenatal alcohol and offspring
development: the first fourteen years. Drug Alcohol Depend 36 (2), 89-99.
-Sulik K. K., Johnston M. C. y Webb M. A. (1981). Fetal alcohol syndrome: embryogenesis in a mouse model.
Science 214 (4523), 936-938.
-Stroband H. W. y Van der Lende T. (1990). Embryonic and uterine development during early pregnancy in
pigs. J Reprod Fertil Suppl. 40, 261-277.
-Sulik K. K. y Johnston M. C. (1983). Sequence of developmental alterations following acute ethanol
exposure in mice: craniofacial features of the fetal alcohol syndrome. Am J Anat 166 (3), 257-269.
-Sulik K. K. (1984). Critical periods for alcohol teratogenesis in mice, with special reference to the
gastrulation stage of embryogenesis. Ciba Found Symp 105, 124-141.
-Sulik K. K., Lauder J. M. y Dehart D. B. (1984). Brain malformations in prenatal mice following acute
maternal ethanol administration. Int J Dev Neurosci 2 (3), 203-214.
-Sulik K. K., Johnston M. C., Daft P. A., Russell W. E. y Dehart D.B. (1986). Fetal alcohol syndrome and
DiGeorge anomaly: critical ethanol exposure periods for craniofacial malformations as illustrated in an
animal model. Am J Med Genet Suppl 2, 97-112.
-Sulik K. K., Cook C. S. y Webster W. S. (1988).Teratogens and craniofacial malformations: Relationships to
cell death. Development 103 (Suppl.), 213–231.

Bibligrafía
333
-Sutherland A. E., Calarco P. G. y Damsky C. H. (1988). Expression and function of cell surface extracellular
matrix receptors in mouse blastocyst attachment and outgrowth. J Cell Biol 106 (4), 1331-1348.
-Swann A. C. (1990). Ethanol and (Naþ,Kþ)-ATPase: alteration of Naþ-Kþ selectivity. Alcohol Clin Exp Res 14
(6), 922–927.
-Tabakoff B. y Hoffman P. L. (2000). Animal Models in Alcohol Research. Alcohol Research and Health 24 (2),
77-84.
-Takase K., Ishikawa M. y Hoshiai H. (1995). Apoptosis on degeneration process of unfertilized mouse ova.
Tohoku J Exp Med 175, 69–76.
-Takeichi M. (1988). The cadherins: cell-cell adhesion molecules controlling animal morphogenesis.
Development 102, 639-655.
-Takeichi M. (1991). Cadherin cell adhesion receptors as a morphogenetic regulator. Science 251, 1451-55.
-Takeichi M. (1995). Morphogenetic roles of classic cadherins. Curr Opin Cell Biol 7, 619-627.
-Tam P.P. (1988). Postimplantation development of mitomycin C-treated mouse blastocysts. Teratology 37
(3), 205-212.
-Tanaka M., Itai T., Adachi M. y Nagata S. (1998). Downregulation of Fas ligand by shedding. Nat Med 4 (1),
31-36.
-Tarkowski A. K. (1966). An air drying method for chromosome preparations from mouse egg. Cytogenetics
(Basel) 5, 394-400.
-Tates A. D., Vogel N. y Neuteboom I. (1980). Cytogenetic effects in hepatocytes, bone marrow cells and
blood lymphocytes of rats exposed to ethanol in the drinking water. Mutat Res 70 (3), 285-288.
-Tavares D. C., Cecchi A. O., Jordao A. A. Jr, Vannucchi H. y Takahashi C. S, (2001). Cytogenetic study of
chronic ethanol consumption in rats. Teratog Carcinog Mutagen 21 (5), 361-368.
-Tebbs R. S., Flannery M. L., Meneses J. J., Hartmann A., Tucker J. D., Thompson L. H., Cleaver J. E. y
Pedersen R. A. (1999). Requirement for the Xrcc1 DNA base excision repair gene during early mouse
development. Dev Biol 208 (2), 513-529.
-Teodoro W. P. R. (1995). Quantificacao e caraterizacao bioquímica e imunoquímica do colágeno no
endométrio de camundongas virgens e grávidas. Tesis de Maestría, Instituto de Ciências Biomédicas,
Universidade de Sao Paulo, Brasil.
-Theiler K. (1989). The house mouse: Atlas of Embryonic Development. New York, Spring-Verlag.
-Thompson K. J., McKillop I. H. y Schrum L. W. (2011). Targeting collagen expression in alcoholic liver
disease. World J Gastroenterol 17(20), 2473-2481.
-Titenko-Holland N., Ahlborn T., Lowe X., Shang N., Smith M. T. y Wyrobek A. J. (1998).
Micronuclei and developmental abnormalities in 4-day mouse embryos after paternal treatment
with acrylamide. Environ Mol Mutagen 31 (3), 206-217.
-Tolivia J., Navarro A., del Valle E., Perez C., Ordoñez C. y Martínez E. (2006). Application of Photoshop and
Scion Image analysis to quantification of signals in histochemistry, immunocytochemistry and
hybridocytochemistry. Anal Quant Cytol Histol 28 (1), 43-53.
-Tsukada S., Parsons C. J. y Rippe R. A. (2006). Mechanisms of liver fibrosis. Clin Chim Acta 364 (1-2), 33-60.
Tsukamoto H., Towner S. J., Ciofalo L. M., y French S. W. (1986). Ethanol-induced liver fibrosis in rats fed high
fat diet. Hepatology 6 (5), 814-822.
-Tunggal J.A., Helfrich I., Schmitz A., Schwarz H., Günzel D., Fromm M., Kemler R., Krieg T. y Niessen C. M.
(2005). E-cadherin is essential for in vivo epidermal barrier function by regulating tight junctions. EMBO J 24
(6), 1146-1156.
-Underwood J.L., Murphy C.G., Chen J., Franse-Carman L., Wood I., EpsteinD.L. y Alvarado J. A. (1999).
Glucocorticoids regulate transendothelial fluid flow resistance and formation of intercellular junctions. Am J
Physiol 277 (2 Pt 1), 330-342.
-Valimaki M., Pelkonen R., Salaspuro M., Harkonen M., Hirvonen E. y Ylikahri R. (1984). Sex hormone in
amenorrheic women with alcoholic liver disease. J Clin Endocrinol Metab 59 (1), 133-138.
-Van Soom A., Ysebaert M.T y de Kruif A. (1997). Relationship between timing of development, morula
morphology, and cell allocation to inner cell mass and trophectoderm in in vitro-produced bovine embryos.
Mol Reprod Dev 47 (1), 47-56.
-Vaux D. L. (2002). Apoptosis and toxicology - what relevance? Toxicology 181-182, 3-7.

Bibligrafía
334
-Viebahn C. (1995). Epithelio-mesenchymal transformation during formation of the mesoderm in the
mammalian embryo.Acta Anat. 154 (1), 79-97.
-Violette M. I., Madan P. y Watson A. J. (2006). Na+/K
+ -ATPase regulates tight junction formation and
function during mouse preimplantation development. Dev Biol 289 (2), 406-419.
-Vu T. H., Shipley J. M., Bergers G., Berger J.E., Helms J. A., Hanahan D., Shapiro S. D., Senior R. M. y Werb
Z. (1998). MM-9/gelatinase B is a key regulator of growth plate angiogenesis and apoptosis of hypertrophic
chondrocytes. Cell 93 (3), 411-422.
-Waddell B. J., Hisheh S., Dharmarajam A. M. y Burton P. J. (2000). Apoptosis in rat placenta is zone-
dependent and stimulated by glucocorticoids. Biol Reprod 63 (6), 1913–1917.
-Walker S. K., Hartwich K. M. y Robinson J. S. (2000). Long-term effects on offspring of exposure of oocytes
and embryos to chemical and physical agents. Hum Reprod Update 6 (6), 564-577.
-Wang G. y Bieberich E. (2010). Prenatal alcohol exposure triggers ceramide-induced apoptosis in neural
crest-derived tissues concurrent with defective cranial development. Cell Death and Disease 1, e46.
-Wang H., Ding T., Brown N., Yamamoto Y., Prince L. S., Reese J. y Paria B.C. (2008). Zonula occludens-1
(ZO-1) is involved in morula to blastocyst transformation in the mouse. Dev Biol 318 (1), 112–125.
-Wang H. y Dey S. K. (2006). Roadmap to embryo implantation: clues from mouse models. Nat Rev Genet 7
(2), 185-199.
-Wang X., Thomas P., Xue J. y Fenech M. (2004). Folate deficiency induces aneuploidy in human
lymphocytes in vitro-evidence using cytokinesis-blocked cells and probes specific for chromosomes 17 and
21. Mutat Res 551 (1-2), 167-180.
-Waring A. J., Rottenberg H., Ohnishi T. y Rubin E. (1982). The effect of chronic ethanol consumption on
temperature-dependent physical properties of liver mitochondrial membranes. Arch Biochem Biophys 216
(1), 51-61.
-Warner C. M., Cao W,, Exley G. E., McElhinny A. S., Alikani M., Cohen J., Scott R.T. y Brenner C.A. (1998a).
Genetic regulation of egg and embryo survival. Hum Reprod 13 (Supl 3), 178–190.
-Warner C. M., Exley G. E., McElhinny A. S. y Tang C. (1998b). Genetic regulation of preimplantation mouse
embryo survival. J Exp Zool 282 (2), 272-279.
-Watson A. J., Damsky C. H. y Kidder G. M. (1990b). Differentiation of an epithelium: Factors affecting the
polarized distribution of Na/K-ATPase in mouse trophectoderm. Dev Biol 141 (1), 104-114.
-Watson A. (1992). The cell biology of blastocyst development. Mol Reprod Dev 33 (4), 492-505.
-Watson E. D. y Cross J. C. (2005). Development of structures and transport functions in the mouse
placenta. Physiology 20, 180-193.
-Webster W. S., Walsh D. A., Lipson A. H. y McEwen S. E. (1980). Teratogenesis after acute alcohol exposure
in inbred and outbred mice. Neurobehavior and Toxicology 2, 227–234.
-Webster W. S., Lipson A. H. y Sulik K. K. (1988). Interference with gastrulation during the third week of
pregnancy as a cause of some facial abnormalities and CNS defects. Am J Med Genet 31 (3), 505-512.
-Webster W. S. y Ritchie H. E. (1991). Teratogenic effects of alcohol and isotretinoin on craniofacial
development: an analysis of animal models. J Craniofac Genet Dev Biol 11 (4), 296-302.
-Wells P. G., McCallum G. P., Chen C. S., Henderson J. T., Lee C. J., Perstin J., Preston T. J., Wiley M. J. y
Wong A. W. (2009). Oxidative stress in developmental origins of disease: teratogenesis,
neurodevelopmental deficits, and cancer. Toxicol Sci 108 (1), 4-18.
-Welsh A. O. y Enders A. C. (1987). Trophoblast-decidual cell interaction and establishment of maternal
blood circulation in the parietal yolk sac placental of the rat. Anat Rec 217 (2), 213-219.
-Wentzel P., Rydberg U. y Eriksson U. J. (2006). Antioxidative treatment diminishes ethanol-induced
congenital malformations in the rat. Alcohol Clin Exp Res 30 (10), 1752-1760.
-West J. R. y Goodlett C. R. (1990). Teratognic effects of alcohol on brain development. Ann Med 22 (5),
319-325.
-Wewer V. M., Damjanov A., Weiss J., Liotta L. A. y Damjanov I. (1986). Mouse endometrial stromal cells
produce basement-membrane components. Differentiation 32 (1), 49-58.
-Wheelock M. J., Shintani Y., Maeda M., Fukumoto Y. y Johnson K. R. (2008). Cadherin switching. J Cell Sci
121 (Pt 6), 727-735.
-White E. (1996). Life, death, and the pursuit of apoptosis. Genes Dev 10 (1), 1–15.

Bibligrafía
335
-Whiteside E. J., Jackson M. M., Herington A. C., Edwards D. R. y Harvey M. B. (2001). Matrix
metalloproteinase-9 and tissue inhibitor of metalloproteinase-3 are key regulators of extracellular matrix
degradation by mouse embryos. Biol Reprod 64 (5), 1331-1337.
-Wiebold J. L. y Becker W. C. (1987). In-vivo and in-vitro effects of ethanol on mouse preimplantation
embryos. J Reprod Fertil 80 (1), 49-57.
-Wiley L. M. (1988). Trophectoderm. The first epithelium to develop in the mammalian embryo. Scanning
Microsc 2 (1), 417-426.
-Winston N., Johnson M., Pickering S. y Braude P. (1991). Partenogenetic activation and development of
fresh and aged human oocytes. Fertil Steril 56 (5), 904-912.
-Wu D. y Cederbaum A. I. (2003). Alcohol, oxidative stress, and free radical damage. Alcohol Res Health 7(4),
277-284.
-Wyllie A. H., Kerr J. F. R. y Currie A. R. (1980). Cell death: the significance of apoptosis. Int Rev Cytol 68,
251-306.
-Wyllie A. H. (1997). Apoptosis: an overview. Br Med Bull 53 (3), 451-465.
-Xu Y., Xiao R. y Li Y. (2005). Effect of ethanol on the development of visceral yolk sac. Hum Reprod 20 (9),
2509-2516.
-Yalcin B., Nicod J., Bhomra A., Davidson S., Cleak J., Farinelli L., Østeras M., Whitley A., Yuan W., Gan X.,
Goodson M., Klenerman P., Satpathy A., Mathis D., Benoist C., Adams D. J., Mott R. y Flint J. (2010).
Commercially Available Outbred Mice for Genome-Wide Association Studies. PLos Genetics 6 (9), 1-15.
-Yang H. W., Hwang K. J., Kwon H. C., Kim H. S., Choi K. W. y Oh K. S. (1998). Detection of reactive oxygen
species (ROS) and apoptosis in human fragmented embryos. Hum Reprod 13 (4), 998–1002.
-Yelian F. D., Yang Y., Hirata J. D., Schultz J. F. y Armant D. R. (1995). Molecular interactions between
fibronectin and integrins during mouse blastocyst outgrowth. Mol Reprod Dev 41 (4), 435-448.
-Yoshida C. y Takeichi M. (1982). Teratocarcinoma cell adhesion: identification of a cell-surface protein
involved in calcium-dependent cell aggregation. Cell 28 (2), 217-224.
-Young T. A., Bailey S. M., Van Horn C. G. y Cunningham C. C. (2006). Chronic ethanol consumption
decreases mitochondrial and glycolytic production of ATP in liver. Alcohol Alcohol 41 (3), 254-260.
-Yu Y., Singh U., Shi W., Konno T., Soares M. J., Geyer R. y Fundele R. (2008). Influence of murine maternal
diabetes on placental morphology, gene expression, and function. Arch Physiol Biochem 114 (2), 99-110.
-Yung J. y Russfield A. B. (1972). Prolactin cells in the hypophysis of cirrhotic patients. Arch Pathol 94 (3), 265-69.
Zaken V., Kohen R. y Ornoy A. (2000). The development of antioxidant defense mechanism in young rat
embryos in vivo and in vitro. Early Pregnancy 4 (2), 110-123.
-Zakeri Z., Quagliano D. y Ahuja H.S. (1994). Apoptotic cell death in the mouse limb and its suppression in
the hammertoe mutant. Dev Biol 165 (1), 294–297.
-Zakeri Z. F. y Ahuja H. S. (1997). Cell death/apoptosis: Normal, chemically induced and teratogenic effect.
Mutat Res 396 (1-2), 149-161.
-Zhang X., Wang H. M., Lin H. Y., Liu G. Y., Li Q. L. y Zhu C. (2004). Regulation of matrix metalloproteinases
(MMPs) and their inhibitors (TIMPs) during mouse peri-implantation: Role of nitric oxide. Placenta 25 (4),
243-252.
-Zhang Y., Yang Z. y Wu J. (2007). Signaling pathways and preimplantation development of mammalian
embryos. FEBS J 274 (17), 4349–4359.
-Ziomek C. A. y Johnson M. H. (1980). Cell surface interactions induce polarization of mouse 8-cell
blastomeres at compaction. Cell 21 (3), 935–942.
-Zorn T. M. T., Bevilacqua E. M. A. F y Abrahamsohn P. A. (1986). Collagen remodeling during
decidualization in the mouse. Cell Tissue Res 244 (2), 443-448.
-Zorn T. M. T., Bijovsky A. T., Bevilacqua E. M. A. F. y Abrahamsohn P. A. (1989). Phagocytosis of collagen
by mouse decidual cells. Anat Rec 225, 96-100.
-Zorn T. M. T., Pinhal M. A. S., Carvalho J. J., Nader H. B., Abrahamsohn P. A. y Dietrich C. P. (1995).
Biosynthesis of glycosaminoglycans in the endometrium during the initial stages of pregnancy of the mouse.
Cell Mol Biol 41 (1), 97- 106.
-Zucoloto S., Rossi M. A., Wright N. A. (1984). Experimental models of chronic alcoholism, solid diet versus
liquid diet. Int J Vitam Nutr Res 54 (4), 387-391.
-Zybina E. V. y Zybina T. G. (1996). Polytene chromosomes in mammalian cells. Int Rev Cytol 165, 53-119.