

CITOQUININAS - franciscosaborio.files.wordpress.com€“Activa crecimiento de yemas laterales ......
42
CITOQUININAS
Transcript of CITOQUININAS - franciscosaborio.files.wordpress.com€“Activa crecimiento de yemas laterales ......

CITOQUININAS

1892 Weisner (Observaciones y Teorías) Sugiere existencia de compuesto que regula la división celular.
1913/1921 Haberlandt Extracto de floema induce división celular en parénquima de tubérculos de papa y heridas Si herida es lavada con agua, no se forma callo
– XLT: compuesto soluble
Define término: totipotencia
1950 Skoog (U. Wisconsin)
Estudiaba brotación de novo en tallos de tabaco cultivados in vitro
Respuesta muy variable > in vitro shoots en medio de cultivo
Prueba agua de coco – Steward at Cornell investiga componentes- que no interfiera
Skoog 1500 m en Suecia 1932 contactó depto de Bioq. 1952 reportan fracción de
agua de coco 4.000 x mas activa, estable a temp, no volatil, orgánico
1951 Carlos Miller
Beca en grupo de Skoog. Busca en extracto de levadura actividad promotora de brotación.
Encontró cultivo (botella) con alta actividad promotora en tallos de tabaco, no las otras,
Observó que actividad precipitaba con AgNO3, igual q purinas y pirimidinas y probó estas
individualmente pero no hubo resp. pero la duda persistió x reporte de skoog de auxinas +
adenina promovían división. Probó diferentes fuentes de ADN entre ella Herring Sperm
(arenque) y tenía alta actividad y precipitaba con AgNO3!
HISTORIA

Se compró HS pero no tenía actividad. Cuando HS se puso “viejo” empezó a mostrar activdad. Probó si autoclavando favorecía el envejecimiento y así fue. El compuesto se logró purificar
1955 Miller et al – Describen actividad en artículo. Y proponen ue se denomine kinetina.
– El grupo de Steward encontró en coco a difenilurea

Historia cont.
1957 Skoog y Miller
Proponen teoría del crecimiento y desarrollo: este es controlado por la relación de auxina y citoquininas (kinetina). Callos de tabaco.
1956 Skoog y Liberman Observan que kinetina aumenta el tamaño de las hojas
1958 Wickson y Thimann
Observan que kinetina rompe dominancia apical en Pisum sativum y rompe letargo de semillas de lechuga, tabaco, trévol otros.
1962 Oberbeek and Loeffler
Kinetina alarga vida de vegetales al reducir tasa de descomposición de proteínas
1963 Letham Australia y Miller en US
primeros en aislar citoquinina de plantas, ambos en granos de maíz.
ZEATINA

Definición
Sustancia que en presencia de una concentración óptima de auxina, induce división celular en cultivos in vitro de médula de tabaco

Bioensayos
Inducción de división en células de médula de tabaco
Inducción de división en células de callo soya
Expansión de células de cotiledones de rábano
Inhibición de la senescencia medido por reducción de degradación de la clorofila.

Citoquininas naturales
Adenina
Zeatina (trans y cis)
Dihidro-zeatina
Dimetilalil adenina (DMAA)
Isopentenil adenina
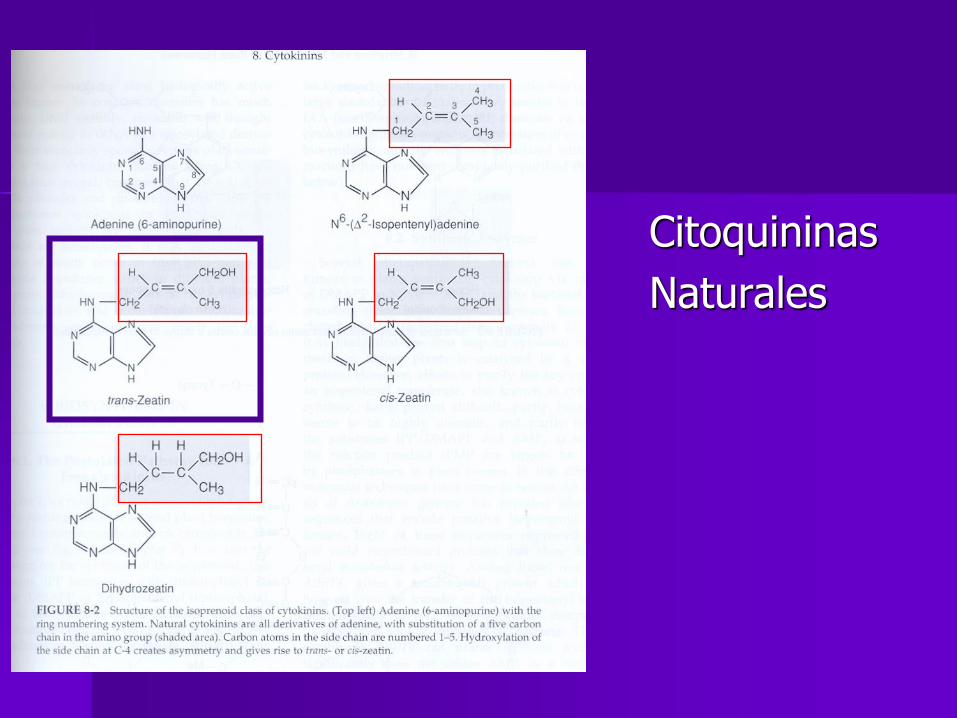
Citoquininas
Naturales

Ribosido-Z Ribotido-Z
tRNA

Citoquinas sintéticas
Kinetina
Benzyl-adenina
Tetrahydropyranyl-benzyl-adenina

Anticitoquininas
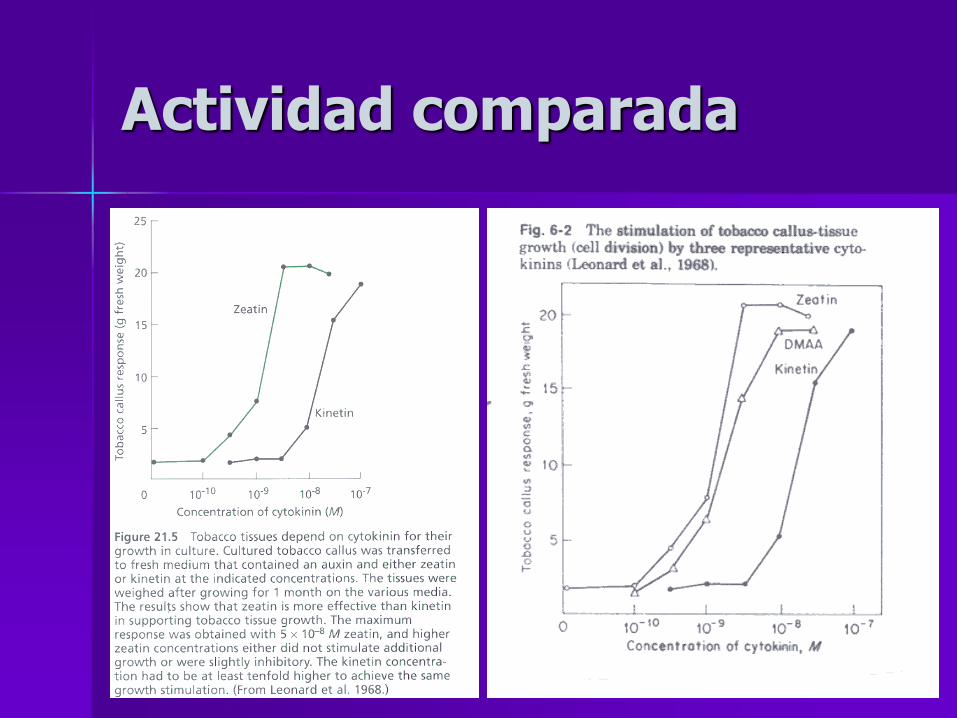
Actividad comparada

Factores de Sensibilidad
RECEPTIVIDAD cambio en número de receptores AFINIDAD cambio en receptores por modificaciones covalentes o cambios alostéricos causados por unión de una molécula al receptor CAPACIDAD DE RESPUESTA cambios en la cadena de eventos posteriores a la unión del RC al receptor EFICIENCIA DE ABSORCION cambios en el sistema de absorción de RC METABOLISMO degradación endógena

Sitios de síntesis
Raíces Meristemos de raíz y tallo Cambium Tejidos de almacenamiento (conjugados) Tejidos en crecimiento
– Semillas – Frutos – Raíces – Yemas laterales – Endospermo líquido

Biosíntesis

Ru
ta d
el
Acid
o M
eva
lón
ico


Síntesis a partir de tRNA
1. En tRNA predomina isómero cis y en la planta predomina trans
2. En callos de tabaco que no necesitan ck para crecer la tasa de degradación de tRNA-z no es suficiente para mantener el nivel de ck libre observado

Conjugación
Combinación reversible de citoquininas con diferentes compuestos y se usan en momentos específicos
Se almacenan en vacuolas y/o ret. endopl.
Compuestos son transportables
Tipo de conjugación depende de especie y de etapa del desarrollo
Se puede combinar con: Glucosa, ribonucleósidos y ribonucleótidos

Conjugación
Intermediarios en síntesis?

Catabolismo
Remoción irreversible de citoquininas
Vía Citoquinina oxidasa
Isopentenil adenina adenina 3 metyl-2-butenal + Citoquinina oxidasa

N-conjugación

UREA
ZEATINA

Ribonucleótido de zeatina
– Forma más común de transporte
Bidireccional
Transporte

Homeostasis
Auxina
Auxina
Conjug. de Auxina Metabolitos Inactivos
Adenina y derivados de adenina
Ck oxidasa
Precursores de citoquinina
AMP Isopentenylpirofosfato
Citoquininas activas Conjugados inactivos
Zeatina, ribosido de Z Isopentenyladenina Isopentenyladenosina
7 y 9 N-glucósidos
O-glucósidos
B-glucosidasa

Efectos Fisiológicos
En combinación con auxinas regulan la relación parte aérea:raiz
Regula dominancia apical – Activa crecimiento de yemas laterales
Induce división celular
Induce formación de órganos in vitro
Retarda senescencia de hojas
En combinación con etileno y luz regula el crecimiento de dicots en la oscuridad

División Celular
Auxina
Agostino y Kiever, 1999
Auxinas
Citoquininas
Sacarosa
Señales específicas
Regulación metabólica

División celular y giberelinas
•mRNA de cdc activada por Giberelinas
•mRNA de ciclina •Síntesis de DNA •Acumulación de células en G2 • Alista células para entrar a M
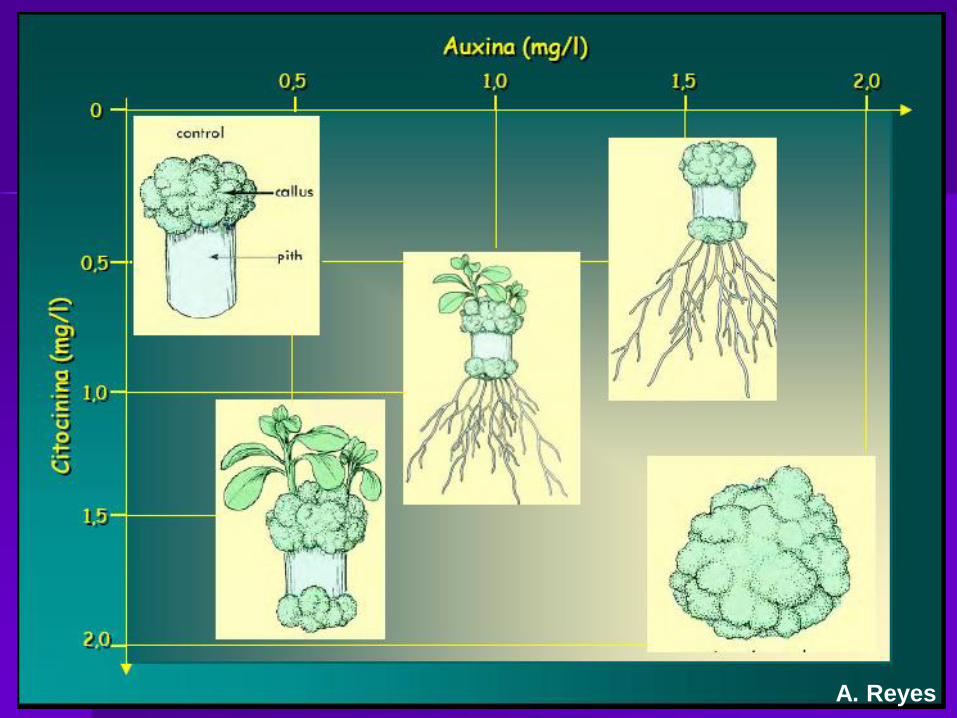
Citoquininas y morfogénesis
A. Reyes


Agrobacterium tumefaciens

Otros efectos fisiológicos
Estimula pérdida de agua por transpiración
Elimina dormancia en algunas semillas
Estimula formación de tubérculos
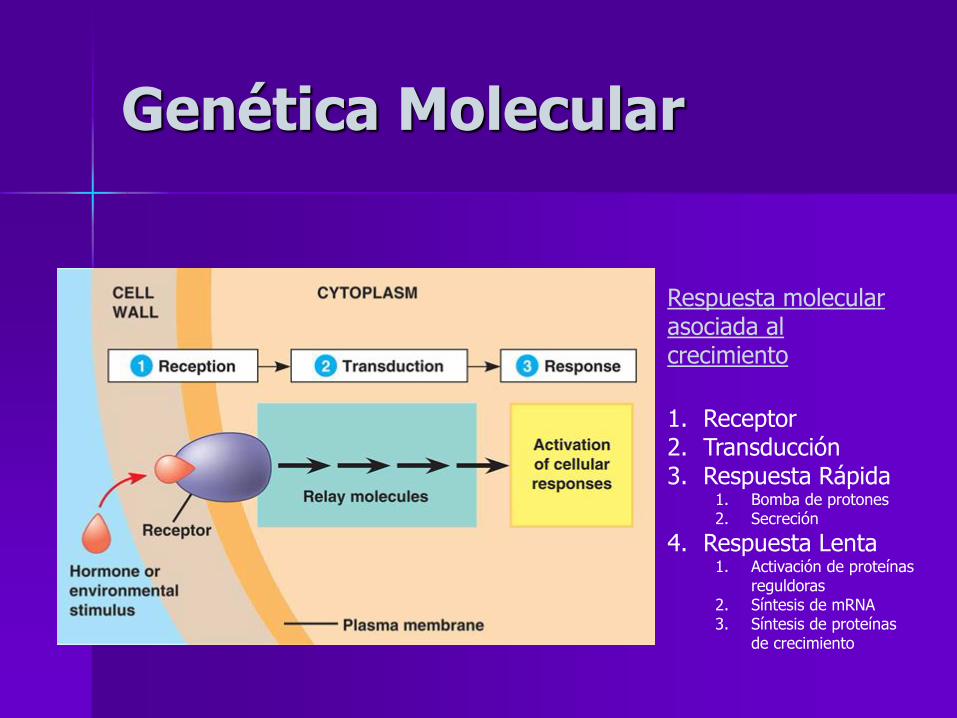
Genética Molecular
Respuesta molecular asociada al crecimiento
1. Receptor 2. Transducción 3. Respuesta Rápida
1. Bomba de protones 2. Secreción
4. Respuesta Lenta 1. Activación de proteínas
reguldoras 2. Síntesis de mRNA 3. Síntesis de proteínas
de crecimiento

Sistema de dos componentes
Kakimoto, 2003

Genes asociados a auxinas
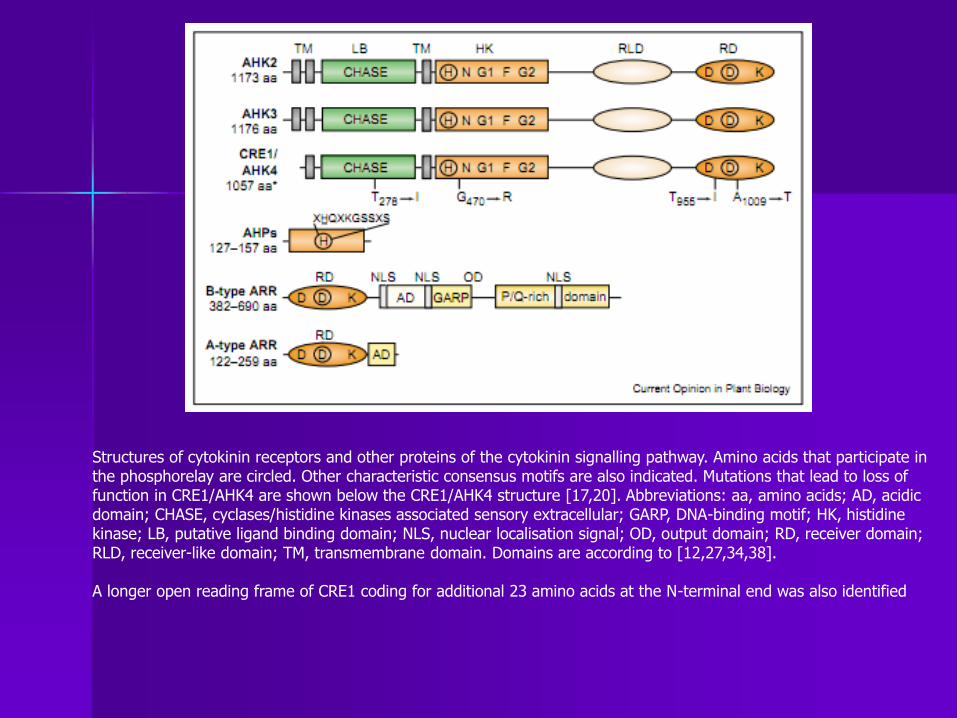
Structures of cytokinin receptors and other proteins of the cytokinin signalling pathway. Amino acids that participate in the phosphorelay are circled. Other characteristic consensus motifs are also indicated. Mutations that lead to loss of function in CRE1/AHK4 are shown below the CRE1/AHK4 structure [17,20]. Abbreviations: aa, amino acids; AD, acidic domain; CHASE, cyclases/histidine kinases associated sensory extracellular; GARP, DNA-binding motif; HK, histidine kinase; LB, putative ligand binding domain; NLS, nuclear localisation signal; OD, output domain; RD, receiver domain; RLD, receiver-like domain; TM, transmembrane domain. Domains are according to [12,27,34,38]. A longer open reading frame of CRE1 coding for additional 23 amino acids at the N-terminal end was also identified

Kakimoto, 2003
Tipo B: activador de TC no influenciado por ck Tipo A: represor de TC, influenciado por ck Type A no fosforilado inhibe la TC inducida por Tipo B, su fosforilación permite TC de Tipo B. Tipo B sintetiza Tipo A (autoregulación)
ARR: arabidopsis response regulator

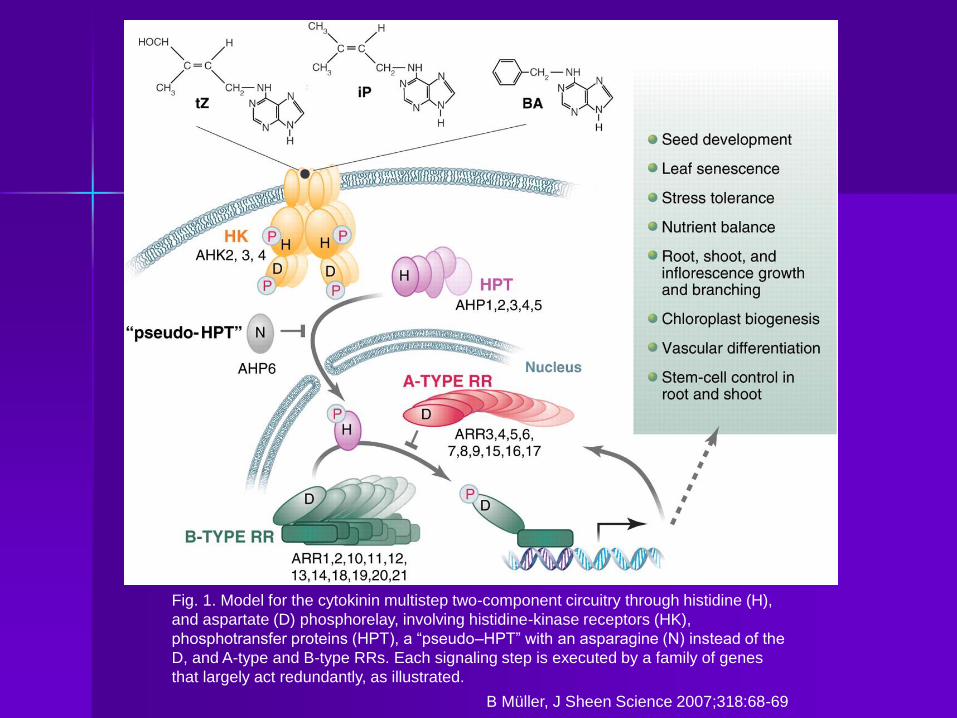
Fig. 1. Model for the cytokinin multistep two-component circuitry through histidine (H),
and aspartate (D) phosphorelay, involving histidine-kinase receptors (HK),
phosphotransfer proteins (HPT), a “pseudo–HPT” with an asparagine (N) instead of the
D, and A-type and B-type RRs. Each signaling step is executed by a family of genes
that largely act redundantly, as illustrated.
B Müller, J Sheen Science 2007;318:68-69




















