







Idiomas
Páginas
Jurídico


PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE Facultad de Ciencias Biológicas
Programa de Doctorado en Ciencias Biológicas Mención Ciencias Fisiológicas
Tesis Doctoral:
PARTICIPACIÓN DE LOS HEMICANALES FORMADOS POR CONEXINA EN LA ATROFIA MUSCULAR INDUCIDA POR CÁNCER Y SEPSIS
Por
Aníbal Antonio Vargas Ríos
Abril 2018

II
PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE Facultad de Ciencias Biológicas
Programa de Doctorado en Ciencias Biológicas Mención Ciencias Fisiológicas
PARTICIPACIÓN DE LOS HEMICANALES FORMADOS POR CONEXINAS EN LA ATROFIA MUSCULAR INDUCIDA POR CÁNCER O SEPSIS
Tesis entregada a la Pontificia Universidad Católica de Chile en cumplimiento parcial de los requisitos para optar al Grado de Doctor en Ciencias
con mención en Ciencias Fisiológicas
Por
Aníbal Antonio Vargas Ríos
Director de Tesis:
Juan Carlos Sáez C.
Abril 2018

III
Esta Tesis está dedicada a Carol José, José, Petty, Andrés y Myriam por todo el apoyo y
soporte que me han dado durante estos años.

IV
Agradecimientos
Esta Tesis fue concebida gracias a la guía de mi Director Juan Carlos Sáez junto con las
observaciones y aportes de todos los integrantes de mi comisión, de los cuales estoy
agradecido por el tiempo dedicado para sacar adelante esta Tesis. También mi gratitud al Dr.
Pérez-Acle y a su laboratorio, en especial a Sebastián y Martín por su valiosa ayuda en el
ámbito bioinformático y a Alex Vielma por todo el apoyo en el campo de la electrofisiología.
Finalmente, del laboratorio de JCS, para Fujiko, Elisa, Elsa, Ximena, Camila, Rosalba, Tere,
Paola y a los que ya dejaron de ser parte de él, pero siempre estuvieron presentes,
especialmente a Bruno, Mario, Paloma, Carlos, Adam, Kenji, Helmut, Pablo y Vania mis
agradecimientos por todo el apoyo profesional, intelectual, técnico y humano durante el
desarrollo de esta Tesis.

V
Financiamientos
1. Proyectos de Investigación
Fondecyt Regular 1141092: Connexin hemichannels as mediators of
sarcolemmal and mitochondrial dysfunction in respiratory and peripheral
muscle in the pathogenesis of sepsis associated-Intensive Care Unit Acquired
Weakness (ICUAW)
Iniciativa Científica Mileno, Centro Interdisciplinario de Neurociencias de
Valparaíso. P09-022-F.
2. Becas de Estudio
Beca Conicyt para estudios de Doctorado en Chile
Beca de Doctorado del Centro Interdisciplinario de Neurociencias de
Valparaíso, a través de la Iniciativa Científica Milenio.

VI
Índice
1. Resumen: .................................................................................................................... 1
1. Abstract....................................................................................................................... 4
1. Introducción ................................................................................................................ 7
1.1. La pérdida de masa muscular asociado a la inflamación sistémica ......................... 7
1.2. Modelos animales de caquexia ................................................................................ 9
1.3. Disfunción muscular en respuesta al aumento de la permeabilidad de la membrana celular ............................................................................................................. 10
1.4. Aumento de permeabilidad del sarcolema se relaciona con la expresión de HCs-Cxs en músculos esqueléticos atróficos ............................................................................ 11
1.5. Las conexinas (Cxs): expresión y función ............................................................... 12
1.6. Función y regulación de HCs-Cxs ............................................................................ 13
1.7. La Cx39 y su función indeterminada como HC ....................................................... 14
1.8. Expresión de Cx39 en células musculares .............................................................. 15
Hipótesis: ........................................................................................................................... 17
Objetivo general: ............................................................................................................... 17
1.9. Objetivos específicos: ............................................................................................. 17
4. Materiales y métodos ................................................................................................ 18
5. Resultados................................................................................................................. 30
5.1.1. Células de melanoma B16F0 inoculadas vía venosa, colonizan el pulmón e inducen el aumento de citoquinas pro-inflamatorias circulantes en ratones. ................. 30
5.1.2. La atrofia muscular esquelética inducida por cáncer ocurre sin participación de las Cxs 43 y 45. ................................................................................................................... 33
5.1.3. El sarcolema de fibras musculares aisladas desde ratones con cáncer no presenta permeabilidad a colorantes. .............................................................................. 36
5.1.4. El PMR de las miofibras de los músculos esqueléticos de ratones tratados con células B16F0 no se altera. ................................................................................................ 39
5.1.5. Las Cxs 39, 43 y 45 no cambian su expresión en miofibras de músculos esqueléticos de ratones inoculados con células de melanoma B16F0. ............................ 42
5.1.6. Los ratones que no expresan Cxs 43 y 45 en miofibras exhiben significativamente menor atrofia muscular esquelética inducida por endotoxemia ....... 44

VII
5.1.7. Las miofibras de músculo TA de ratones sometidos con endotoxemia expresan Cx39, Cx43 y Cx45 .............................................................................................................. 47
5.1.8. Las miofibras de ratones con endotoxemia presentan menor PMR y aumento en la captación de colorantes. ........................................................................................... 49
5.1.9. La expresión y actividad de HCs-Cxs disminuye el PMR de células HeLa. .......... 52
5.2. Caracterización de Cx39 y su función como hemicanal en células HeLa. ..................... 57
5.2.1. Las células Hela-Cx39 no forman CUH funcionales. ........................................... 57
5.2.2. Un aumento de actividad de los HCs-Cx39 incrementa la corriente macroscópica de la membrana en células HeLa-Cx39. ............................................................................ 60
5.2.3. Los HC-Cx39 aumentan su actividad en ausencia de cationes divalentes extracelulares y presentan conductancia unitaria característica. .................................... 63
5.2.4. Los HCs-Cx39 permiten el paso de un grupo selectivo de moléculas con diferentes propiedades fisicoquímicas .............................................................................. 65
5.2.5. Electronegatividad, potencial de ionización, flexibilidad, valencia, tamaño, polarizabilidad y geometría son características claves para definir el paso de moléculas a través de los HCs-Cx39 ...................................................................................................... 68
5.2.6. Los HCs-Cx39 abiertos no permiten el ingreso del ion Ca2+ ............................... 71
5.2.6.1. El pH alcalino induce aumento de la captación de colorante pero no de Ca2+
en células HeLa-Cx39 ......................................................................................................... 71
5.2.6.2. El estrés mecánico (EM) induce aumento de la captación de colorante pero no de Ca2+ en células HeLa-Cx39 ....................................................................................... 73
5.2.7. La captación de colorante en células HeLa-Cx39 es inhibida por algunos pero no por otros bloqueadores de HCs-Cxs. ................................................................................. 75
5.2.8. El modelamiento comparativo de los HCs-Cx39 revela constricciones en el poro que no se visualizan en los HCs-Cx43 y en los HCs-Cx26................................................... 78
5.2.9. La estructura del poro de los HC-Cx39 posee una distribución de cargas eléctricas singular en comparación a los HCs-Cx26 y HCs-Cx43 ....................................... 81
6. Discusión ................................................................................................................... 83
7. Conclusiones ............................................................................................................ 104
8. Bibliografía ............................................................................................................... 106
9. Anexo I. Certificado de Bioética ............................................................................... 117
9.1. Anexo II. Portada de publicación relacionado con esta Tesis ..................................... 118

VIII
Lista de abreviaturas
7-ADD: 7-aminoactinomicina D
ACMA: 9-amino-6-cloro-2-metoxiacridina
AE˗4: Azul de Evans
AM+1: Azul de metileno
AST: Área de sección transversal
ATP: Adenosina tri-fosfato
BBG-: Brilliant blue G
BSA: Albumina de suero de bovino (Bovine serum albumine en inglés)
Ca2+: Ion calcio
CBX: Carbenoxolona
CUH: Canal de unión en hendidura
Cx26: Conexina 26
Cx39: Conexina 39
Cx43: Conexina 43
Cx45: Conexina 45
DAPI+2: 4´,6´-diamidino-2-fenilindol
EM: Estrés mecánico
Etd+: Bromuro de etidio
FDB: Flexor Digitalis Brevis
GA: Algoritmo genético (genetic algoritm en inglés)

IX
gj: Conductancia intercelular
HCs-Cxs: Hemicanales formados por conexinas
HeLa-P: Células HeLa parentales
H-etd+4: Homodímero de etidio
I: Corriente
Ij: Corriente intercelular
IL1-β: Interleukina 1-β
IL-6: Interleukina-6
INF-γ: Interferon-γ
IP2+: Ioduro de propidio
KCL: Cloruro de potasio
KNN: Vecinos K más cercanos (K-nearest neighbors en inglés)
La3+: Ion lantano
LCP: Ligación cecal y punción
LPS: Lipopolisacarido
LY-2: Amarillo Lucifer (Lucifer yellow, en inglés)
SLCD: Solución Libre de Cationes Divalentes
Na+: Ion sodio
NAD+: Nicotinamida adenina dinucleótido
NO: Óxido nítrico (“nitric oxide” en inglés)
Ole: Oleamida
PBS: Tampón fosfato salino (phosphate buffer saline, en inglés)

X
PGE2: Prostaglandina E2
PMR: Potencial de membrana en reposo
SFB: Suero Fetal Bovino
TA: Tibialis Anterior
TNF-α: Factor de necrosis tumoral alfa (tumor necrosis factor alpha en inglés)
V: Voltaje
VI-: Verde de indocianina
β-Ga: Ácido 18-β-glicirritínico

1
1. Resumen:
La pérdida de masa corporal se asocia directamente a la atrofia muscular en pacientes
con ciertas enfermedades inflamatorias sistémicas crónicas y agudas como el cáncer y la
sepsis, respectivamente. Es por ello que caracterizar los eventos que participan en la atrofia
muscular ha sido de gran interés. En otras condiciones asociadas a la pérdida de masa
muscular, como la denervación y la administración prolongada de glucocorticoides, se ha
demostrado que una parte importante es consecuencia de la expresión de conexinas (Cxs)
39, 43 y 45, y su actividad como hemicanales (HCs-Cxs), aunque se desconoce si Cx39 forma
HCs-Cxs funcionales. La dependencia de la atrofia muscular a la expresión y función de HCs-
Cxs, se ha establecido con el uso de ratones KO-Cx43/Cx45 miofibra específico (las cuales no
expresan Cx43 ni Cx45, pero siguen expresando Cx39, en músculos diferenciados atrofiados).
En estas condiciones, disminuye el PMR y aumenta de permeabilidad del sarcolema a
colorantes, como etidio y azul de Evans, y al Ca2+, lo que precede a la atrofia muscular. El
aumento de permeabilidad a colorantes y baja del PMR también se han observado en atrofia
muscular inducida por sepsis. Sin embargo, no ha sido dilucidado si estos cambios dependen
de la expresión de HCs-Cxs. Además, se desconoce si los HCs-Cxs participan en la atrofia
muscular inducida por cáncer.

2
En esta tesis se estudiaron: 1) la posible participación de los HCs-Cxs en la atrofia
muscular inducida por sepsis y cáncer y 2) se caracterizaron propiedades de los canales
formados por la Cx39. Se propuso la siguiente hipótesis: “El aumento de la expresión y
actividad de los HCs-Cxs induce atrofia y disfunción de los músculos esqueléticos en
condiciones inflamatorias asociadas al cáncer o sepsis.”
Objetivos generales: 1) Determinar si la atrofia muscular y el aumento de la permeabilidad
del sarcolema, dependen de la expresión y estado funcional de los HCs-Cxs en un modelo de
cáncer y un modelo de sepsis. 2) Determinar si la Cx39 forma canales funcionales y de ser así
determinar sus propiedades biofísicas y sensibilidades a inhibidores.
En el modelo de cáncer generado con por la inyección endovenosa de células de
melanoma de ratón (células B16F0), se produjo una atrofia muscular cercana al 25%, tanto
en ratones controles como en ratones que no expresan Cx43/Cx45 en miofibra. Además, no
se evidenciaron cambios en la permeabilidad del sarcolema de las miofibras de ratones con
cáncer comparados con ratones controles y el PMR se mantuvo en valores normales en
ambos casos. Por otro lado, en ratones con sepsis inducida por ligación cecal y punción
(LCP), se observó el aumento de permeabilidad del sarcolema y atrofia muscular, que fueron
dependientes de la expresión de Cx43 y Cx45. La atrofia muscular se redujo a cerca de un
30% en ratones con miofibras deficientes en la expresión de Cx43/Cx45, a pesar de mantener
la expresión de Cx39.
La Cx39 y su función como HCs-Cxs fue se caracterizaron en células HeLa transfectadas
establemente con el cDNA de la Cx39 ratón (la cual es ortóloga de la Cx40.1 humana), en

3
donde se encontró HCs-Cxs funcionales con una conductancia unitaria de 75 pS, permeables
a diversos colorantes, y la cual aumenta su probabilidad de apertura en soluciones libres de
cationes divalentes [SLCD], pH extracelular alcalino, estrés mecánico y polarización de la
membrana celular a potenciales positivos. Sin embargo, Ca2+ no pasa a través de los
HCs˗Cx39 abiertos.
Los resultados de este trabajo indican que la atrofia muscular inducida por cáncer sería
independiente de la expresión y actividad de HCs-Cxs, sin embargo la atrofia inducida por
sepsis es significativamente dependiente de la expresión de HCs-Cxs. Ya que la Cx39 forma
HCs funcionales muy poco permeables o impermeables a Ca2+, parece no favorecer la
inducción de atrofia por sepsis.

4
1. Abstract
Loss of body mass is directly associated with muscle atrophy in patients with certain
systemic chronic inflammatory diseases such as cancer and acute as sepsis. In sick humans
and animal models, muscle atrophy is directly related to survival, and permits to estimate the
time and quality of rehabilitation of survivors to severe sepsis as well. It is for this reason that
characterizing the events that participate in muscular atrophy in both pathologies has been
relevant. Previously, the dependence of muscular atrophy on the expression and activity of
hemichannels formed by Cxs 43, 45 or 39 has been described in other models of muscle mass
loss, such as denervation- and glucocorticoid-induced atrophy, in which the phenomena of
increase in permeability of the sarcolemma to dyes and Ca2+, and drop in resting membrane
potential (RMP) precede muscle atrophy. The increase in sarcolemma permeability is not
observed in muscle-specific KO-Cx43/Cx45 mice, which are also, significantly resistant to
atrophy induced by these conditions. However, the fraction of atrophy response observed in
KO-Cx43/Cx45 mice, might still be mediated by Cx39 of which neither its function as HC nor
permeability properties have been determined.
As observed in denervation-induced muscle atrophy, in the early stages of muscle
atrophy induced by sepsis, loss of membrane integrity of myofibers has been reported based
on increased dye uptake, as well as decrease in RMP. However, it has not been established
whether these changes and muscular atrophy depend on Cx HCs. It is also unknown whether
in cancer-induced muscle atrophy these phenomena exist and whether they are dependent
on Cx HCs.

5
In this thesis, (1) the participation of Cx HCs in muscle loss induced by sepsis and
cancer was determined. Moreover, (2) properties of Cx39-based channels were studied. The
following hypothesis was proposed: "The increase in the expression and activity of
hemichannels formed by Cxs induces atrophy and dysfunction of the skeletal muscles in
inflammatory conditions induced by cancer and sepsis."
General objectives:
1. To determine if muscle atrophy, as well as previous events including increase in
sarcolemma permeability, depend on the expression and function of Cx HCs in cancer
and sepsis models.
2. To determine whether Cx39 forms functional channels and if so determining their
biophysical properties and sensitivity to known gap junction and hemichannel
blockers.
Either control or Cx43/Cx45 skeletal muscle deficient mice inoculated with mouse
melanoma cells (B16F0 cells) showed about 25% muscle atrophy. In addition, no change in
sarcolemma permeability of myofibers from mice with cancer was observed since the RMP
and dye uptake of the myofibers were comparable to that of myofibers from control mice.
At the same time, mice subjected to acute sepsis by cecal ligation and puncture (LCP),
showed increase in dye permeability and drop in RMP of myofibers dependent on the

6
expression of Cx43 and Cx45. In addition, close to 50% muscle atrophy was observed in
surviving mice, in which about 30% of this atrophy was Cx43/Cx45 expression dependent.
Cx39 and hemichannel function were characterized in HeLa cells stably transfected with
this protein. Functional hemichannels were found in cells exposed to conditions known to
increase the open probability of other Cx HCs (e.g., divalent cation free media (SLCD),
extracellular alkaline pH, mechanical stress and positive membrane potentials). Under
conditions that Cx39 HCs increased their activity in the presence of extracellular Ca2+, it was
observed that this ion does not pass through open Cx39 HCs. Under these conditions unitary
events of 75 pS were frequent in HeLa-Cx39 cells and absent in parental cells.
Given the above results, it was determined that cancer-induced muscle atrophy is
independent of Cx HC expression and activity. However, muscle atrophy induce by sepsis is
partially due to Cx HC expression. In addition, it was shown that Cx39 forms functional HCs
however they are not permeable to Ca2+ and thus this feature does not contribute to the
sepsis-induced muscle atrophy.

7
1. Introducción
1.1. La pérdida de masa muscular asociado a la inflamación sistémica
Caquexia es un síndrome asociado a enfermedades crónicas, como el cáncer o el sida y
también agudas, como el shock séptico, caracterizada por la pérdida de masa corporal
superior al 5% en un rango menor a 12 meses, asociado a pérdida de fuerza y atrofia
muscular, fatiga, anorexia, inflamación entre otros síntomas. En pacientes con este síndrome,
disminuye rápidamente la calidad de vida y aumenta la morbilidad y mortalidad. La atrofia
muscular esquelética asociada a caquexia, tiene una prevalencia de entre el 60 al 80% en
enfermos de cáncer, en donde un 30% muere directamente por este síndrome y el 50%
restante muere con algún grado de atrofia muscular (von Haehling, S y Anker, SD 2010). A su
vez, en sepsis grave, existe una prevalencia de entre el 20-60%, que en el corto plazo
desarrollan neuropatía (Tennila, A et al. 2000), cuya gravedad está estrechamente
relacionada con la menor sobrevida de los internados en unidades de cuidados intensivos y el
aumento del tiempo de recuperación de los sobrevivientes (Visser, LH 2006, Callahan, LA y
Supinski, GS 2009). La pérdida masa muscular en ambos casos se ha tratado de estabilizar
con aproximaciones hormonales para inhibir la anorexia y estimular la hipertrofia muscular
pero sus resultados son dispares, ya que no se ha observado un aumento significativo de la
sobrevida, a la vez que los tratamientos crónicos con agonistas adrenérgicos no entregan
resultados favorables ya que a corto plazo provocan alteraciones vasculares y cardíacos
(Amitani, M et al. 2013, Joassard, OR et al. 2013, Kalantar-Zadeh, K et al. 2013). Es por ello
que los estudios actuales, se enfocan en el catabolismo del músculo inducido por un estado

8
inflamatorio. Estudios, tanto in vitro como in vivo de modelos animales de cáncer, sepsis y
otras patologías asociadas que desarrollan atrofia muscular, han revelado que el aumento
del catabolismo proteico es mayor que el aumento del anabolismo y el cual es consecuencia
de un estado inflamatorio exacerbado, mediado por citoquinas pro-inflamatorias como TNF-
α, IL-1β, IL-6 e INF-γ (Bonaldo, P y Sandri, M 2013) y glucocorticoides (Schakman, O et al.
2013). Estas citoquinas en el músculo esquelético activan factores de transcripción de vías
proinflamatorias, entre ellas Nf˗κB, el cual induce la expresión de proteínas catalíticas de la
vía ubiquitin/ligasa- proteosoma, MURF-1 y Atrogina (Argiles, JM et al. 2005, Baltgalvis, KA et
al. 2008, Sishi, BJ y Engelbrecht, AM 2011). Por otro lado, se ha evidenciado una
dependencia importante de la atrofia muscular esquelética a la expresión de Cx39, Cx43 y
Cx45 en las miofibras de un modelo de atrofia muscular inducida por denervación (Cea, LA et
al. 2013, Cisterna, BA et al. 2016) o glucocorticoides (Cea, LA et al. 2016). Ante la nula
expresión de Cxs43/45 en los músculos esqueléticos, la atrofia muscular es drásticamente
reducida (~75% de la atrofia inducida por denervación) o totalmente prevenida (atrofia
inducida por glucocorticoides). Esto último indica que, independiente del estímulo,
compartirían el mismo mecanismo de atrofia muscular dependiente de la expresión de Cxs,
por lo que, eventualmente, sería un blanco a considerar en el tratamiento de la atrofia
inducida por cáncer y/o sepsis.

9
1.2. Modelos animales de caquexia
Sepsis: La septicemia es una condición inflamatoria sistémica aguda provocada
principalmente por el sistema inmune innato en respuesta a la presencia de agentes
patógenos. En humanos se reconocen dos fases, la primera presenta un alto gasto cardiaco
con una baja resistencia vascular y la segunda presenta bajo gasto cardiaco, la cual es la fase
del shock séptico. La alta incidencia de muerte por shock séptico se debe a la falla
multiorgánica a corto plazo. Los diversos modelos de condiciones asociadas al shock séptico
como la endotoxemia con endotoxina bacteriana (LPS), y de sepsis propiamente tal con
bacterias o ligación cecal y punción (LCP) manifiestan un aumento de citoquinas
proinflamatorias y glucocorticoides plasmáticos (Vary, TC 1998). El modelo de LCP reproduce
el aumento de citoquinas proinflamatorias y los eventos hemodinámicos característicos de la
sepsis en humanos. Las ventajas de este modelo, es que desarrolla una sepsis polimicrobiana
endógena y reproduce de mejor manera lo que se observa en humanos al romperse el tubo
intestinal en casos de apendicitis/peritonitis (Holecek, M 2012). Este es un modelo de
caquexia aguda, en donde en ratas y después de 6 horas de la intervención aumenta los
niveles de citoquinas proinflamatorias y comienza la proteólisis en los músculos esqueléticos
como EDL y soleus (Holecek, M 2012).
Cáncer: es el crecimiento celular descontrolado que se inicia por la desregulación del
ciclo celular en cualquier tejido. Clínicamente se reconocen cuatro fases de progresión
crónica del cáncer que dependen del crecimiento del tumor, colonización de nódulos

10
linfáticos y de otros órganos (metástasis) por estas células. Para su estudio se usan
principalmente tres modelos de cáncer en ratas y ratones que corresponden a: 1) trasplante
de tumores (líneas celulares derivados de tumores desarrollados en la misma especie), 2)
mutantes (inmunosuprimidos o con regulación celular alterada) que requieren de la
inyección de un carcinógeno o por inducción espontanea en los tejidos y 3) por inyección
directa de un tumor humano. De estos modelos, el trasplante de tumores de la misma
especie, resulta en un modelo ideal si se requiere conocer la respuesta inflamatoria del
cáncer (generado en la misma especie) en otros tejidos, con su sistema inmunológico intacto,
y sin la intervención de cancerígenos que pueden inducir una inflamación aguda local. Este
modelo entrega la posibilidad de modular la respuesta de otros tejidos, como el muscular, en
un sistema inflamado inducido por cáncer. De las líneas celulares disponibles para generar
cáncer en ratón C57/BL6, se encuentran las células B16F0 derivadas de melanoma de ratón
C57BL/6J, que se caracterizan por generar una pobre respuesta inmunogénica, lo cual, al
inyectarlas por vía subcutánea o venosa, no genera respuesta de rechazo por parte de los
ratones (lo que no incluye inducción de una respuesta inflamatoria aguda a la inoculación de
estas células). Por lo tanto, las respuestas inflamatorias observadas en este modelo de cáncer
se deben a la proliferación y desarrollo del tumor y la metástasis dentro del animal y no al
efecto agudo de la detección de las células cancerígenas por el sistema inmune (Dranoff, G
2012).
1.3. Disfunción muscular en respuesta al aumento de la permeabilidad de la
membrana celular

11
En sepsis, junto con la atrofia, también se ha descrito disfunción muscular, reflejada en el
aumento de la permeabilidad del sarcolema a colorantes (Lin, MC et al. 1998) y la
disminución del PMR de las miofibras (Liu, SH et al. 1997). En este sentido, TNF-α induce una
disminución del PMR de las miofibras de manera concentración-dependiente en músculos de
rata ex-vivo e in vivo en las primeras dos horas de incubación o post-administración (Tracey,
KJ et al. 1986). A su vez, en modelo de endotoxemia inducido por inyección de
lipopolisacarido bacteriano (LPS) en ratones, se produce la pérdida del 8% del potencial de
membrana en el músculo diafragma en un rango de tiempo similar (Liu, SH et al. 1997). A
pesar de que se ha establecido que condiciones inflamatorias inducen la caída del PMR,
registros de este fenómeno junto cambios en la permeabilidad a colorantes en modelo de
atrofia muscular inducido por cáncer no han sido estudiados en detalle.
1.4. Aumento de permeabilidad del sarcolema se relaciona con la expresión de HCs-
Cxs en músculos esqueléticos atróficos
La caída del PMR y la entrada de colorantes a las fibras musculares en modelos de
sepsis en ratas, sugiere que las propiedades de permeabilidad del sarcolema están alteradas.
En atrofias musculares dependientes de HCs-Cxs, se ha observado aumento de la
permeabilidad del sarcolema a colorantes, tanto in vitro (fibras aisladas) como in vivo (Cea,
LA et al. 2013), como también la acumulación neta de [Ca2+]i proveniente del espacio
extracelular (Cisterna, BA et al. 2016). Junto con lo anterior, se ha observado que la caída de
PMR (Cea, LA et al. 2016) y el ingreso neto de Na+ (Cisterna, BA et al. 2016) son inhibidos por

12
bloqueadores de HCs como el La3+ y CBX y no ocurre en ratones KO músculo-específico
Cx43fl/flCx45fl/fl:Myo-Cre (Cea, LA et al. 2012, Cea, LA et al. 2013, Cisterna, BA et al. 2016). Sin
embargo, en atrofia muscular inducida por denervación en esta cepa de ratones KO, persiste
la expresión de Cx39, por lo que es posible que, la fracción de atrofia remanente, podría
estar asociada a la expresión de HCs-Cx39. Se desconoce si los efectos descritos arriba son
compartidos por modelos inflamatorios como el cáncer o la sepsis.
1.5. Las conexinas (Cxs): expresión y función
Las Cxs son proteínas integrales de membrana expresadas en la mayoría de los tejidos
de los vertebrados, y en humanos son codificadas por 21 genes diferentes (von Maltzahn, J et
al. 2004). Seis subunidades de Cxs compatibles forman HCs homoméricos y dos HCs-Cxs
compatibles forman un CUH homotípico. Los HCs comunican los medios intra y extracelular
mientras que los CUH comunican los citoplasmas de dos células adyacentes (Sáez, JC et al.
2003). Por otro lado, las Cxs también cumplen funciones que no corresponden a CUH o HCs
(Jiang, JX y Gu, S 2005), como por ejemplo, siendo parte en transducción de señales
mediadas por receptores purinérgicos P2Y (Scemes, E 2008), en mecanismos de adhesión
celular (Elias, LA et al. 2007), regulación del citoesqueleto (Olk, S et al. 2010), de expresión
génica (Iacobas, DA et al. 2004), función de exosomas (Soares, AR et al. 2015) y función
mitocondrial (Rodriguez-Sinovas, A et al. 2017), ya que se pueden localizar en todos los
subcompartimentos celulares mencionados arriba.

13
1.6. Función y regulación de HCs-Cxs
Los HCs-Cxs están involucrados en la señalización celular autocrina y paracrina, ya que
son permeables a diferentes moléculas pequeñas involucradas en señalización celular,
incluyendo NAD+, ATP, glutamato, NO y PGE2 y de sustratos metabólicos como glucosa y
glutatión (Bennett, MV et al. 2003, Figueroa, XF et al. 2013, Orellana, JA y Stehberg, J 2014).
Además, son canales de baja selectividad a iones, siendo incluso permeables a Ca2+ (Sánchez,
HA et al. 2009, Sánchez, HA et al. 2010, Schalper, KA et al. 2010). Es por esto, que los HCs-
Cxs constituyen una vía alternativa de ingreso de este ión al citoplasma y por tanto favorecen
la activación de vías de degradación de proteínas dependientes de Ca2+ como las calpaínas
(Smith, IJ et al. 2008). La actividad de los HCs puede ser regulada por fosforilación (Bao, X et
al. 2007, Puebla, C et al. 2016), potencial redox (Contreras, JE et al. 2002, Retamal, MA et al.
2006, Retamal, MA et al. 2007), pH (Beahm, DL y Hall, JE 2002, Schalper, KA et al. 2010,
D'Hondt, C et al. 2013) entre otras condiciones (Sáez, JC et al. 2005). En adición, la
probabilidad de apertura de diferentes HCs-Cxs incrementa en células expuestas a bajas
concentraciones de Ca2+ extracelular (Sáez, JC et al. 2005), potenciales de membrana
positivos (Sáez, JC et al. 2005) o estrés mecánico (D'Hondt, C et al. 2013), y es drásticamente
reducido por La3+ y bloqueadores de CUH como CBX, β-Ga y octanol entre otros (D'Hondt, C
et al. 2013).

14
1.7. La Cx39 y su función indeterminada como HC
A pesar de conocerse algunos mecanismos que regulan los HCs-Cx43 y HCs-Cx45, que
se expresan en músculo en ciertas condiciones de atrofia, aún se desconoce si las
condiciones anteriormente descritas afectan el estado funcional delos HCs-Cx39 y si ésta
podría participar en la atrofia del músculo de ratones que expresan esta Cx.
Se ha postulado que los HCs-Cxs presentan baja probabilidad de apertura en
condiciones basales in vitro, particularmente en sistemas de expresión exógenos (Trexler, EB
et al. 1996, Kondo, RP et al. 2000, Contreras, JE et al. 2003, Sáez, JC et al. 2005). El rango de
conductancia unitaria de HCs-Cxs varía entre 17 pS en HCs-Cx32 (Gómez-Hernández, JM et al.
2003, Oh, S et al. 2004) hasta 352 pS en HCs-Cx52 (Valiunas, V y Weingart, R 2000). En
general, la conductancia unitaria de HCs-Cxs es más alta que la de CUH-Cxs homotípicos
(Sáez, JC et al. 2005), específicamente la conductancia unitaria de HCs compuestos por Cx30,
Cx43 o Cx45 es cercano al doble de la conductancia de CUH homotípicos formados por las
mismas Cxs (Valiunas, V y Weingart, R 2000, Valiunas, V 2002, Contreras, JE et al. 2003).
Debido a que la conductancia unitaria de CUH-Cx39 no ha sido descrita, no es predecible la
conductancia unitaria de los HCs-Cx39.
La permeabilidad de los HCs-Cxs a moléculas ha sido frecuentemente estudiada por
medio de la captación de sondas fluorescentes (Schalper, KA et al. 2008), con diferentes
tamaños, formas y cargas netas como bromuro de etidio (Etd+), 4´,6´-diamidino-2-fenilindol
(DAPI2+) y amarillo Lucifer (LY2-) entre otros (Sáez, JC et al. 2005). Con esta herramienta, los
HCs-Cx26 (Figueroa, VA et al. 2014), HCs-Cx32 (Sánchez, HA et al. 2009) y HCs-Cx43

15
(Contreras, JE et al. 2003, Retamal, MA et al. 2007, Orellana, JA et al. 2011), así como los HCs
formados por otras Cxs descritas hasta ahora y activados con diferentes estímulos, presentan
características de permeabilidad similares. Hasta ahora, los HCs-Cx39 expresados en células
HeLa han mostrado ser permeables a azul de Evans (AE4-) (Cea, LA et al. 2013), pero sus
características de permeabilidad permanecen mayormente sin determinar, por lo que su
caracterización es relevante para entender su posible participación tanto en fenómenos
fisiológicos como fisiopatológicos incluyendo la atrofia muscular.
1.8. Expresión de Cx39 en células musculares
La Cx39 codificada en el genoma de ratón comparte cerca del 61% de identidad de
secuencia con su ortólogo humano, la Cx40.1 (von Maltzahn, J et al. 2004). La Cx39 se
expresa transitoriamente en mioblasto de ratón durante el desarrollo embrionario tardío
(von Maltzahn, J et al. 2004), después de la denervación en miofibras de ratones adultos
(Cea, LA et al. 2013) como también en miofibras de ratones mdx, un modelo de la
enfermedad de distrofia muscular de Duchenne (Cea, LA et al. 2016). Por otro lado, los
ratones deficientes en Cx39 muestran una miogénesis y regeneración acelerada (von
Maltzahn, J et al. 2011). En sistemas de expresión exógena, la Cx39 no forma CUH
(estudiados usando un número pequeño de sondas permeables) (von Maltzahn, J et al.
2004). Sin embargo, la formación de CUH-Cx39 no ha sido demostrada con técnicas más
sensibles, como las mediciones de acoplamiento eléctrico. Se podría especular que la Cx39
podría estar relacionada con una actividad independiente de HCs o que su actividad como

16
CUH pueda estar altamente regulada en el músculo, por lo que es necesaria su
caracterización en un sistema aislado en células HeLa transfectadas con Cx39.
Al haberse determinado que distintas condiciones que inducen atrofia muscular (denervación
o glucocorticoides), tienen la misma dependencia de expresión de Cxs y considerando que,
en ambos casos, el bloqueo farmacológico de los HCs-Cxs puede evitar de forma importante
dicho proceso, hace interesante conocer si las atrofias musculares derivada de cáncer o
sepsis, las cuales son refractarias a tratamientos actuales de aumento de masa muscular (ej.:
tratamientos hormonales y/o cambios de dieta), podrían ser también dependientes del
aumento de permeabilidad del sarcolema, debido al incremento de la expresión y la
actividad de HCs-Cxs. Esta posible dependencia de la atrofia muscular esquelética, en
condiciones de inflamación sistémica crónica y aguda, a la expresión de las Cxs aún no ha
sido estudiada.

17
Hipótesis: el aumento de la expresión y actividad de HCs-Cxs en miofibras induce atrofia
y disfunción de los músculos esqueléticos en condiciones inflamatorias sistémicas inducidas
por caquexia aguda o crónica.
Objetivo general: determinar si la atrofia y disfunción muscular inducida por caquexia de
forma crónica por cáncer, y aguda por sepsis depende de la actividad de HCs formados por
Cx39, Cx43 yCx45
1.9. Objetivos específicos:
1. Determinar si la caquexia crónica inducida por cáncer afecta la expresión y actividad
in vitro de los HCs formados por Cx39, Cx43 y Cx45
2. Determinar si la caquexia aguda inducida por LCP induce la expresión y actividad in
vitro de HCs formados por Cx39, Cx43 y Cx45
3. Determinar si la expresión y actividad de HCs-Cxs inducen atrofia y disfunción
muscular en ambos modelos de caquexia.
4. Caracterizar los canales formados por la Cx39

18
4. Materiales y métodos
Reactivos: Octanol, heptanol, CBX, oleamida, LaCl3, Etd+, procion orange MX2R (PO, 3-),
Brilliant Blue G (BBG, 1-), Azul de Evans (AE, ˗4) y ioduro de propidio (IP, +2) fueron
comprados de Sigma-Aldrich (Saint Louis, Missouri, EUA). 9-amino-6-cloro-2-metoxiacridina
(ACMA, 1+),4´,6´-diamidino-2-fenilindol (DAPI, 2+), ioduro de hexidio (IH, 1+), amarillo
nuclear (AN, 1+), 1-ioduro BOBO (BOBO, 4+), 7-aminoactinomicina D (7-ADD, 0), 1-ioduro
YOYO (YOYO, 4+), 1-ioduro TOTO (TOTO, 4+), y homodímero de etidio (H-etd, 4+) fueron
obtenidos de Invitrogen (Massachusetts, EUA), verde de indocianina (VI, 1+), de Pulsion
Medical Systems (Feldkirchen, Alemania) azul de metileno (AM, 1+), de Euromed (Santiago,
Chile).
Ratones: El uso de animales en esta tesis tuvo la autorización del comité de Bioética de la
Pontificia Universidad Católica de Chile, rol CBB 233/2013 y N° 150323001. Se utilizaron
ratones C57bl/6 machos de dos meses y ratones doble mutantes para Cxs 43/45 músculo
específicos, C57bl/6 Cx43fl/fl-Cx45fl/fl:Myo-Cre con su respectiva línea de origen como control.
Todos con un régimen alimenticio ad libitum antes y durante el ensayo. Todos los animales
fueron sacrificados bajo anestesia y dislocación cervical.
Ensayo de metástasis y caquexia: Se indujo colonización y crecimiento de células
tumorales en ratones utilizando células de melanoma B16F0, las cuales previamente fueron
cultivadas en placas Nunc™ (Thermo Fisher, Dinamarca) y alimentadas con medio DMEM alto

19
en glucosa, suplementado con 10% de suero fetal bovino (SFB) y 50 U/ml de penicilina y
estreptomicina, a pH 7.4 y mantenidos a 37°C en atmósfera de 5% CO2 / 95% aire y
alimentadas cada dos días. Las células al 60-70% de confluencia se disgregaron con tripsina
(3 min, 37oC) bajo agitación. Luego de la neutralización con medio completo que contenía
SFB, las células fueron lavadas para eliminar la tripsina, se re-suspendieron en PBS para la
posterior inyección en ratones C57/BL6 y Cx43fl/fl-Cx45fl/fl:Myo-Cre (5x107 células, iv). Los
ratones, fueron eutanasiados a la semana 1 o 2 post-inoculación. Cada día se les controló
peso y signos de dolor según escala de observación y evaluación de gestos y conducta en
ratones.
Ensayo de LCP: El modelo de sepsis grave fue realizado por LCP como se ha descrito
anteriormente (Holecek, M 2012). En resumen, los ratones fueron anestesiados usando
isofluorano (Baxter Healthcare, Puerto Rico), luego el abdomen fue afeitado y se realizó una
incisión en la línea media abdominal bajo el diafragma. El ciego fue aislado, ligado con sutura
quirúrgica de nylon (Tagumedica, Chile) y punzado con aguja 21-G. Posteriormente, una
pequeña cantidad de material fecal fue extruida. El ciego fue retornado a la cavidad
abdominal y se agregó 200 µl de PBS estéril para asegurar sepsis. La incisión fue cerrada con
sutura quirúrgica de nylon. Después de la cirugía a todos los ratones se le administró
tramadol (30 mg/kg) cada 24 h, para disminuir el dolor y se verificó estado general y peso.
Ratones controles Sham, fueron sometidos al mismo protocolo de operación, pero sin ligar ni
punzar el ciego.

20
Captación in vitro de fibras musculares aisladas en tiempo real: Para ensayos de
permeabilidad se utilizaron fibras aisladas de músculo Flexor Digitalis Brevis (FDB), las cuales
se obtuvieron mediante digestión con colagenasa (1 mM, disuelta en DMEM F12, 10% FBS,
3h a 37oC) y disgregadas mecánicamente por trituración a través de un pipeta Pasteur de
boca ancha, y luego se permitió su adherencia sobre placas de plástico por 10 minutos. La
solución de registro Krebs que contenía (en mM) NaCl (145), KCl (5), CaCl2 (3), MgCl2 (1),
glucosa (5), HEPES (10), Tris (5) (pH 7.4), se le agregó Etd+ (MW 394,3 g/mol, W: 620 nm) a
una concentración final de 5 µM. El registro de captación se realizó en microscopio de
fluorescencia invertido Nikon Ti, tomando fotografías cada 30 segundos con software de
control NIS Elements (Nikon, Japón), analizando posteriormente la intensidad de
fluorescencia de los núcleos por un tiempo de 5 min con procesador de imágenes ImageJ V
1.48. Se compararon las tasas de captación entre las distintas condiciones.
Inmunofluorescencia: Se utilizaron cortes transversales de músculo Tibialis Anterior
(TA) fijados en paraformaldehído al 3%. Las muestras se incubaron con solución de bloqueo
(50 mM NH4Cl, 0,025%Tritón, 1% BSA en PBS) durante toda la noche a 4oC para disminuir la
unión no específica del anticuerpo primario. Luego, las muestras se incubaron con anticuerpo
policlonal contra Cx39, Cx43 o Cx45, por 12 h. Posteriormente, las muestras se lavaron 3
veces con PBS, y luego fueron incubadas por 30 minutos con anti-IgG de conejo acoplado con
Cy3, y posteriormente fueron lavadas 3 veces con PBS. Las muestras fueron finalmente
montadas en un portaobjeto con fluoromont G y observadas en microscopio de fluorescencia
invertido Nikon Ti, obteniendo las imágenes con software NIS Elements (Nikon, Japón).

21
Medición de potencial de membrana (PMR): Se utilizó músculo FDB completo
sumergido en solución Krebs (pH 7.4) para mediciones de PMR en configuración “current
clamp” se utilizó un amplificador Axopatch 1D (Molecular Devices, CA, EUA). El electrodo de
referencia fue insertado en un puente de agarosa 3M KCl y sumergido en solución salina a pH
7.4 y la pipeta de registro (25-30mΩ) fue fabricada a partir de un capilar de vidrio (A-M
System, Inc. Cat # 596800) con estirador horizontal P97 (Sutter instruments, CA, EUA) la cual
fue llenada con una solución 3 M KCl.
Medición de citoquinas séricas: Se obtuvo muestras de sangre mediante un corte y
recolección de sangre desde la cola de los ratones, obteniendo un volumen aproximado de
100 µL. Se permitió la formación del coágulo a temperatura ambiente por 1 h y luego se
obtuvo el suero por centrifugación (2.000 RPM, 4oC, 5 min). La muestra se guardó a -80oC
hasta el ensayo de medición de citoquinas con kit Milliplex map (Millipore). En resumen, el
kit presenta 96 platos de muestras con “beads” magnéticos de inmunodetección para
citoquinas IFN-, IL-1b, IL-6, TNF-, mediante fluorescencia que se detectó con Luminex 200 y
fue analizada con software xPONENT (Millipore Corp.)
Medición de área transversal de fibras musculares: Se utilizaron cortes transversales
de músculo TA fijados en paraformaldehído al 3%, teñidos en eosina/hematoxilina. El área
de sección transversal (AST) fue determinada mediante el marcaje del contorno de cada fibra
en el corte, con dibujo libre de ROI de medición, utilizando el programa ImageJ v.148 para
finalmente calcular el área dentro del contorno demarcado, usando el mismo programa.

22
Líneas celulares: Las células HeLa-p y células HeLa transfectadas con Cxs fueron
cultivadas en medio DMEM suplementado con 10% de SFB, 50 U/ml de penicilina y
estreptomicina, a pH 7.4 y mantenidos a 37°C en atmósfera de 5% CO2/95% aire. Las células
transfectadas HeLa-Cx39, HeLa-Cx45 y Hela Cx43 fueron seleccionadas con 0.5 mg/ml
puromicina y HeLa-Cx43-EGFP con 0.3 mg/ml geneticina (Invitrogen, MA, EUA), como se
describió anteriormente (Jordan, K et al. 1999, von Maltzahn, J et al. 2004). Células HeLa-
Cx39 fueron amablemente facilitadas por Dr. Klaus Willecke (Instituto Lymes, Universidad de
Bonn, Bonn Alemania). Para cada experimento, las células fueron sembradas sobre
cubreobjetos de vidrio, hasta que la confluencia apropiada fue alcanzada. Las células fueron
alimentadas un día antes del experimento.
La expresión de la Cx39 en células HeLa fue confirmada por inmunofluorescencia. En
resumen, células sembradas sobre cubreobjetos de vidrio y fueron lavadas con PBS, fijadas
con formaldehído al 3% e incubadas con solución de bloqueo (1% BSA, 0.025% triton X-100 y
50 mM NH4Cl, en PBS a pH 7.4) por 12 h. Luego, las células fueron incubadas con anticuerpo
de conejo anti-Cx39 (Invitrogen, MA, EUA) por 6 h a temperatura ambiente para
posteriormente lavarlas con PBS e incubarlas 30 min con anticuerpo secundario anti-IgGs de
conejo (Santa Cruz, Texas, EUA) conjugado con Cy2. Las preparaciones fueron lavadas con
PBS y los cubre objetos fueron montados con Fluoromont-G sin DAPI (Electron Microscopy
Science, PA, EUA).

23
Electrofisiología: El estado funcional de los HCs fue evaluado en experimentos de
fijación de voltaje en modalidad de célula completa como ha sido previamente descrito
(Contreras, JE et al. 2003, Sánchez, HA et al. 2009). La solución de baño contenía: (en mM)
140 NaCl, 5.4 CsCl, 1 MgCl2, 1.8 CaCl2, 2 BaCl2, y 10 HEPES, pH 7.4. La solución de la pipeta
contenía (en mM): 130 CsCl, 10 AspNa, 0.26 CaCl2, 1 MgCl2, 2 EGTA, 7 TEA-Cl, and 5 HEPES, pH
7.2. La resistencia de la pipeta en el baño fue de 10-15 MΩ. Todos los registros fueron
realizados a temperatura ambiente (21-23oC). Las corrientes macroscópicas y microscópicas
fueron obtenidos con protocolos de pulsos rectangulares de voltaje (0 mV a +60 mV por 40 s)
y protocolos de rampas de voltaje (-80 mV a +80 mV, por 9 s). Las conductancias fueron
calculadas siguiendo la ley de Ohm: 𝑔𝑚 =(𝐼𝑚𝑏
−𝐼𝑚𝑎)
𝑉𝑚 donde (Imb – Ima) es la diferencia de
corriente, entre el estado abierto y cerrado del canal, respectivamente, y Vm el voltaje de
membrana fijado en el protocolo, con el cual se observa las transiciones de corrientes
medidas.
Las mediciones de PMR fueron realizadas con microelectrodos bajo configuración de
fijación de corriente en célula completa a 37oC en células individuales. La pipeta de registro
fue llenada con solución de KCl 3 M con una resistencia de 25-30 MΩ y la solución del baño
fue la misma que la utilizada en la medición de potenciales de membrana de las fibras
musculares de los ratones tratados con cáncer y sepsis. El valor registrado corresponde al
PMR medido cuando la pipeta cruza la membrana celular y se mantuvo estable por 5 s desde
el acceso.

24
Para evaluar el acoplamiento eléctrico, cultivos de alta densidad celular (60 a 70%)
fueron bañados con una solución salina que contenía (en mM): 150 NaCl, 4 KCl, 1 MgCl2, 1.2
CaCl2, y HEPES a pH 7.4. La solución de pipeta fue la misma utilizada en experimentos con
HCs suplementado con 18 µM DAPI2+ en unas de las pipetas para identificar las células
registradas para la posterior detección de la Cx39 por inmunofluorescencia indirecta. Las
corrientes de canales intercelulares fueron registradas en células adyacentes usando
protocolos de pulsos de voltajes rectangulares incrementados cada 20 mV desde -100 a 0 mV
y con una duración de 200 ms cada uno. El potencial de membrana fijada para la célula no
estimulada fue de -60 mV y la conductancia de CUH (gj) fue calculada desde 𝑔𝑗 =
−𝐼𝑏 (𝑉𝑎 − 𝑉𝑏)⁄ , como se describe previamente, siendo Ib la corriente de unión intercelular (Ij)
registrada en la célula no estimulada (Van Rijen, HV et al. 1998). Todos los experimentos
fueron realizados usando un microscopio invertido Olympus IX51 con amplificador
Axopatch1-D, digitalizador Digidata 1322 y programa de adquisición de datos Clampex 9.1
con escala digitalizada x 10. Los datos fueron analizados con programa Clampfit 2.1
(Molecular Devices, Ca, EUA).
Captación de colorante en tiempo real de células HeLa: Las células HeLa fueron
bañadas en la misma solución salina utilizada en la captación de colorante de fibras
musculares in vitro, suplementada con 5 µM de cada colorante, que normalmente no
ingresan a la célula a través de la membrana celular. Las células fueron registradas por 5 min
en solución control, y fueron lavadas 5 veces con solución control sin Ca2+ ni Mg2+ (SLCD) para
inducir la apertura de HCs, como se ha mostrado para otras Cxs (Cea, LA et al. 2013). Para

25
inducir apertura de los HCs-Cxs por estrés mecánico (EM), las células fueron registradas en
medio salino normal por 5 min, posteriormente se aplicó el estímulo que consistió en un
goteo continuo de 1 ml, de la misma solución, lanzado con una pipeta desde 15 cm de altura
impactando directamente en la muestra. Las imágenes fueron registradas cada 30 s por 20
min con microscopio invertido Eclipse Ti y programa NIS-Elements (Nikon, Tokio, Japón); los
análisis fueron realizados con programa ImageJ 1.42q (NIH, MD, EUA).
Señal de Ca2+: La señal de Ca2+ intracelular correspondiente a la razón de emisiones
F340/F380 (nombrada desde ahora como señal de Ca2+), fue registrada como se describió
anteriormente (Harcha, PA et al. 2015). En resumen, células sembradas sobre cubreobjetos
de vidrio fueron cargadas por 30 minutos a 37oC con 5 µM FURA-2AM (Invitrogen, MA, EUA)
en la misma solución salina que la utilizada en ensayos de captación de colorante y fueron
lavadas con la misma solución sin FURA-2AM. Para las mediciones de la señal de Ca2+, la
intensidad de fluorescencia fue capturada cada 3 s. La cuantificación de la razón de la
intensidad de fluorescencia (Señal de Ca2+ = F340/F380) e imágenes fue realizada en
microscopio invertido Nikon Eclipse Ti implementado con el programa NIS Elements (Nikon,
Tokio, Japón).
Modelamiento molecular: Modelos moleculares de HCs y CUH fueron producidos
usando la estructura cristalizada de Cx26 humana como molde (Maeda, S et al. 2009). Se
computaron alineamientos secuenciales para producir los modelos moleculares al introducir
la secuencia aminoacídica en el molde. Luego, múltiples alineamientos de secuencias fueron

26
recomputadas por Mafft v7 (Araya-Secchi, R et al. 2012) usando la matriz de substitución
PAM200 y la estrategia lenta con todos los pares de alineamientos locales. La identidad de
secuencia versus la estructura molde resultó en 31.3% y 42.3% para Cx39 y Cx43,
respectivamente. Constricciones espaciales necesarias para producir los modelos
moleculares de Cx39 y Cx43 fueron compilados desde la estructura molde utilizando
MODELLER v.9.14 (Eswar, N et al. 2006). En total, 1.000 moldes fueron producidos por cada
secuencia objetivo, mientras que la calidad estructural fue determinada utilizando MAIDEN
(Postic, G et al. 2015). MAIDEN evalúa los ajustes de las distancias interatómicas entre los
residuos localizados en las hélices transmembrana en la bicapa lipídica usando la función de
energía empírica. El modelo con puntaje máximo para cada proteína fue elegido para análisis
posteriores.
Radio del poro: Perfiles de radio del poro para los modelos moleculares y la
estructura cristalizada fueron computados con la aplicación detectora de cavidades Epock
(Laurent, B et al. 2015), implementado en VMD (Humphrey, W et al. 1996). Para ello, un eje
prospectivo de 120Å y 200Å de largo para HCs y CUH, respectivamente, fue trazado a lo largo
del eje Z para cada modelo molecular. Un radio prospectivo de 16Å fue seleccionado,
mientras el volumen disponible fue computado usando una grilla de 0,5Å de espacio.
Evaluación del potencial electrostático: El potencial electrostático de nuestros
modelos moleculares fue determinado usando PDB2QPR implementado en la herramienta
APBS 2.1 de PyMOL (Lerner, M y Carlson, H 2006). Esta herramienta evalúa átomos de

27
hidrógeno y cargas para modelos moleculares acorde al campo de fuerza CHARMM. Luego,
el potencial electrostático fue computado utilizando la ecuación linearizada de Poisson-
Boltzmann y parámetros por defecto. Las figuras fueron producidas utilizando las
capacidades de dibujo en 3D de PyMOL.
Modelamiento in silico de colorantes y caracterización: Todos los colorantes
permeables fueron modelados usando el software de diseño molecular Avogadro v1.0.3
(Hanwell, MD et al. 2012). La codificación molecular se realizó utilizando los códigos SMILES
para cada molécula obtenidos desde el sitio PuBChem de NCBI (Bolton, EE et al. 2008).
Cuando los códigos SMILES no estaban disponibles, las moléculas fueron dibujadas a mano
con herramienta de dibujo del software Avogadro. Todas las moléculas fueron sometidas a
optimización geométrica, realizando un procedimiento de minimización de energía utilizando
los algoritmos de gradientes conjugados y campos de fuerzas MMFF94 (Halgren, TA 1996) en
software Avogadro. Luego, un total de 1.875 descriptores moleculares fueron calculados
usando el programa descriptor molecular PaDEL v2.21 (Yap, CW 2011). Descriptores que no
pudieron ser computados para más de una molécula, o que eran equivalentes a 0 para todas
las moléculas, pero menos de uno, fueron descartados como no informativos. Para
seleccionar los descriptores que mejor representan al grupo de moléculas, se utilizó una
aproximación basada en clasificadores de vecinos K más cercanos (KNN, K-nearest neighbors)
combinados con un algoritmo genético (GA, genetic algorithm). Cada combinación GA
generada de los descriptores fue utilizada para clasificar cada molécula dentro de dos clases
basadas en su capacidad de pasar a través de HCs-Cx39. Esta clasificación fue realizada con

28
un clasificador KNN y un procedimiento “dejar uno afuera”. El GA usado, trabaja con lo
siguiente: una población de 5.000 diferentes sub-sets (ej. Genomas o soluciones) de
descriptores moleculares generados en la primera iteración. Cada uno de estos genomas
tuvo un vector numérico de largo equivalente al total del número de descriptores, en el que
los elementos vectoriales asignados a ser utilizados se establecieron en 1, mientras que los
descriptores no utilizados se establecieron en 0. En la población inicial el primer sub-set
incluyó todos los descriptores que podrían ser producidos por todas las moléculas (1.532
descriptores, todos los elementos establecidos en 1). Los siguientes sub-sets fueron
realizados por cada descriptor, y los sub-sets restantes fueron elegidos al azar como una
combinación de descriptores. Subsecuentemente, los 1.000 sub-sets que correctamente
clasificaron un gran número de moléculas, pasaron directamente (y sin alterar) a la próxima
generación, y fueron usados para crear las 4.000 soluciones restantes. Estas 4.000 soluciones
fueron creadas por entrecruzamiento al azar de mil sub-sets, seguidos por mutaciones
puntuales (un cambio del valor en el elemento vector al otro valor posible) que ocurre con
una probabilidad de 10-3. Durante el proceso de entrecruzamiento, una posición vectorial fue
azarosamente elegida para combinar dos de 1.000 genomas, creando dos genomas hijos.
Este protocolo asegura rápida convergencia de sub-set de descriptores que clasificaron
correctamente todas las moléculas usando el KNN.
Análisis estadístico: Todos los resultados son presentados como el promedio ± EE. Y
las diferencias significativas entre grupos fueron determinados usando ANOVA de una vía
seguido por comparaciones múltiples a posteriori con test de Tukey. Las diferencias fueron

29
consideradas significativas con p<0,05. Los análisis estadísticos fueron realizados utilizando
Excel 2013 (Microsoft, WA, EUA) y GraphPad Prism 6 (GraphPad, CA, EUA).

30
5. Resultados
5.1.1. Células de melanoma B16F0 inoculadas vía venosa, colonizan el pulmón e
inducen el aumento de citoquinas pro-inflamatorias circulantes en ratones.
Se estableció el modelo de cáncer en ratón induciéndolo mediante inoculación de células de
melanoma B16F0 (5X105) vía intravenosa (Timmons, JJ et al. 2016)y el progreso de la
metástasis se evaluó a una y dos semanas post-inoculación de las células (Fig. 1A). Estos
tiempos de evaluación fueron seleccionados en base a la progresión de atrofia muscular
dependiente HCs-Cxs, observada en otros modelos como el inducido por denervación, en el
cual se evidencia atrofia significativa a partir de la primera semana de producido el daño
(Cea, LA et al. 2013, Cisterna, BA et al. 2016). El peso y signos de dolor (según lo observado
en comportamiento y gesticulación facial, comparados con tabla de bienestar y dolor
animal), fueron evaluados diariamente sin encontrarse variaciones significativas en el peso
durante los periodos señalados (Fig. 1A), ni dolor evidente, entre ratones tratados con células
de melanoma como comparados con ratones control. A pesar de no encontrarse diferencias
evidentes, tanto conductuales como de peso, entre los ratones controles e inyectados, la
invasión de las células B16F0 en el segundo grupo se manifestó en forma de colonias de color
negro (por la expresión de melanina), distribuidas en el tejido pulmonar a los 7 días post-
inyección, las cuales crecieron y se fusionaron con otras colonias, como se observó a los 14
días post-tratamiento (Fig. 1B). Como ha sido reportado anteriormente (Gil-Ad, I et al. 2006)
desde el día 14, y hasta el día 20, decae rápidamente el estado general de los ratones, con
signos evidentes de dolor e hiperventilación, debido a la colonización y crecimiento masivo

31
de las células de melanoma, por lo que se determinó finalizar el experimento a las dos
semanas post-inyección.
Como se ha establecido anteriormente en estos modelos de cáncer (Argiles, JM y
Lopez-Soriano, FJ 1999), la concentración sérica de citoquinas proinflamatorias aumentó
significativamente desde la primera semana en ratones inyectados con células B16F0, en
comparación con el suero obtenido de ratones inyectados sólo con PBS, desde la primera
semana de tratamiento: IL1-β (346 ± 29 pg/ml vs. 39,05 ± 5,80 pg/ml, n=4, p<0,0001), IL˗6,
(346,0 ± 29,2 pg/ml vs. 43,8 ± 8,7 pg/ml n=4 p<0,0001), TNF-α (39,8 ± 2,9 pg/ml vs. 4,4 ± 0,4
pg/ml, n=4 p<0,0001) e IFN-γ (1,3 ± 0,1 pg/ml vs. 0,06 ± 0,01 pg/ml n=4, p<0,0001). Estas
diferencias se mantuvieron hasta el día 14, IL1-β (179,3 ± 7,0 pg/ml vs. 15,9 ± 2,1 pg/ml, n=4,
p=0,0003), IL˗6, (249,3 ± 28,8 pg/ml vs. 18,5 ± 6,0 pg/ml n=4 p=0,0003), TNF-α (35,2 ± 3,0
pg/ml vs. 3,3 ± 0,6 pg/ml, n=4 p<0,0001) e IFN-γ (0,3 ± 0,05 pg/ml vs. 0,04 ± 0,001 pg/ml n=4,
p=0,04)(Fig. 1C), por lo que se establece que, a la primera semana post-administración de las
células B16F0, se alcanza un estado inflamatorio sostenido, y por lo tanto se decidió evaluar
la atrofia muscular esquelética y la dependencia de esta condición a la expresión de los HCs-
Cxs.

32
Fig. 1: Las células B16F0 i.v. colonizan el pulmón e inducen un estado inflamatorio general A. Modelo de cáncer en ratón inducido con células de melanoma B16F0 (5 x 10^5 células/ ratón i.v.). El peso de los ratones tratados hasta el final del protocolo no reveló diferencias significativas. B. El pulmón fue colonizado por las células B16F0, lo que se evidenció como manchas negras (debido a la expresión de melanina) que formaron colonias en la superficie. En el día 14 el tamaño de las manchas fue mayor que a los 7 días posiblemente debido al crecimiento y fusión de las colonias. Además, el tamaño del pulmón aumentó debido al menos en parte a la invasión y el crecimiento de las colonias de melanoma que además inducen congestión pulmonar. C. Concentración de citoquinas pro-inflamatorias plasmáticas. La concentración de TNF-α, IL-1β, IL-6 e INF-γ aumentó en el suero ratón inyectado con células de melanoma, desde la primera semana y se mantuvo elevado hasta la segunda semana de evaluación.(* p<0,05; *** p<0,005; **** p<0,0001).
B
C
A

33
5.1.2. La atrofia muscular esquelética inducida por cáncer ocurre sin participación de
las Cxs 43 y 45.
Una vez establecido el modelo de inflamación crónica, inducido por la colonización de células
de melanoma B16F0, se evaluó si este estado inflamatorio se asociaba con atrofia muscular
esquelética y si esta depende de los HCs-Cxs, mediante la medición del AST de las miofibras
del músculo TA. A 7 días post-inyección de las células B16F0, el AST de las miofibras de
ratones Cx43fl/fl-Cx45fl/fl, los cuales normalmente expresan Cx39, Cx43 y Cx45 en los músculos
esqueléticos en desarrollo y en condiciones de daño, denervación o inflamación, disminuyó
significativamente (alrededor de un 25%) con respecto a los ratones tratados sólo con
vehículo. Esta atrofia muscular, inducida por las de células cancerosas, se mantuvo sin
variaciones hasta el día 14 (Fig. 2).
Para determinar si la expresión de Cx43 y Cx45 participan en la atrofia muscular
inducida por cáncer, se utilizaron ratones Cx43fl/flCx45fl/fl:Myo-Cre que no expresan
Cx43/Cx45 de forma músculo-específica, pero que en condiciones de denervación aún
expresan Cx39. Tanto en el día 7 como el día 14, la atrofia muscular esquelética inducida por
cáncer, fue similar a lo observada en ratones Cx43fl/fl-Cx45fl/fl que si expresan Cxs (Fig. 2), por
lo que las Cx43 y Cx45 no participarían en la atrofia muscular en el modelo de cáncer
estudiado. En paralelo, se utilizó un control positivo de atrofia muscular esquelética
dependiente de HCs-Cxs inducido por denervación al seccionar el nervio ciático en ratones
Cx43fl/flCx45fl/fl y Cx43fl/flCx45fl/fl:Myo-Cre, como se ha descrito anteriormente (Cea, LA et al.

34
2013, Cisterna, BA et al. 2016). Tanto en el día 7 como en el día 14, la atrofia muscular
esquelética fue de 40% en ratones Cx43fl/flCx45fl/fl mientras que ratones Cx43fl/flCx45fl/fl:Myo-
Cre alcanzó una atrofia total de sólo un 25% (Fig. 2).

35
Fig.2: La atrofia muscular esquelética inducida por colonización de células B16F0 en el pulmón también ocurre en ratones que no expresan Cx43-45 en los músculos esqueléticos. Cortes de músculo TA en ratones WT y KO-músculo específico de Cxs43-45 a los 7 y 14 días post i.v. de células B16F0, comparados con ratones denervados en el mismo tiempo. B. Porcentajes promedios AST de las miofibras presentes en cortes transversales de músculos TA, con respecto a las de ratones Sham. Cada uno de los tratamientos con células B16F0 indujo la caída del tamaño de las fibras cercana al 25%. Barras blancas: WT; barras negras KO-Cx43-45. n=4 por grupo, 600 a 900 áreas de fibras medidas por corte, 3 cortes por n *p<0,05, y **p<0,001. Barra de cal. 50 µm.

36
5.1.3. El sarcolema de fibras musculares aisladas desde ratones con cáncer no presenta
permeabilidad a colorantes.
A pesar de observarse que la presencia de células de melanoma inducen la misma atrofia en
los ratones Cx43fl/flCx45fl/fl:Myo-Cre como en ratones Cx43fl/flCx45fl/fl, se evaluó si las fibras
musculares aisladas de ratones Cx43fl/flCx45fl/fl presentan permeabilidad a colorantes que
pasan a través de HCs-Cxs, en respuesta al cáncer, como se ha observado en un modelo de
atrofia muscular inducido por denervación, en el cual el aumento de la permeabilidad del
sarcolema de las fibras musculares precede a la atrofia muscular (Cisterna, BA et al. 2016).
Para esto, se utilizó como control positivo el incremento de permeabilidad de la membrana
celular a colorantes, de las fibras musculares aisladas de ratones denervados Cx43fl/flCx45fl/fl y
Cx43fl/flCx45fl/fl:Myo-Cre (Cea, LA et al. 2013, Cisterna, BA et al. 2016). A los 7 y 14 días post-
inoculación de células B16F0 se aislaron miofibras de músculo FDB, y se evaluó la
permeabilidad del sarcolema mediante la captación de Etd+. Las tasas de captación de Etd+
fueron similares a las mostradas por las miofibras de ratones tratados con vehículo (0,15 ±
0,12 UA/min y 0,15 ± 0,03 UA/min vs. 0,16 ± 0,03, n=5, p>0,98 y p>0,99, respectivamente). A
su vez, la captación basal de Etd+ de miofibras de ratones tratados con células B16F0 fue
inhibida por La3+, tanto a los 7 como a los 14 días post-inoculación (0,14 ± 0,01 vs. 0,05 ± 0,01
UA/min y 0,15 ± 0,03 vs. 0,06 ± 0,02 UA/min, n=5, p<0,05, respectivamente) (Fig.3A). Sin
embargo, la tasa de captación de Etd+ aumentó significativamente en fibras aisladas de
ratones denervados (7 días post-denervación), comparado con ratones controles (0,78 ± 0,05

37
AU/min vs. 0,17 ± 0,03 AU/min, n=6, p<0,0001) y ratones con cáncer (0,78 ± 0,05 AU/min vs.
0,15 ± 0,12 AU/min). Este aumento de captación fue significativamente menor en miofibras
de ratones Cx43fl/flCx45fl/fl:Myo-Cre comparado con las miofibras de músculo denervados de
ratones Cx43fl/flCx45fl/fl (0,15 ± 0,03 AU/min vs. 0,78 ± 0,05 AU/min, respectivamente, n=6,
p<0,0001), los cuales presentan tasas de captación similares a las fibras controles, como se
ha visto anteriormente (Cisterna, BA et al. 2016). La tasa de captación de colorante de
miofibras de ratones denervados Cx43fl/flCx45fl/fl:Myo-Cre presentaron valores similares a la
de miofibras de ratones con cáncer (0,15 ± 0,03 AU/min vs. 0,15 ± 0,12 AU/min, n=6, p=0,99)
(Fig. 3D).

38
Fig.3: La inoculación con células B16F0 no aumenta de captación de Etd+ en fibras aisladas de músculo esquelético. A. Curvas representativas de captación de Etd+ de fibras aisladas de músculo FDB de ratones inoculados con células de melanoma y controles. Ambas curvas tienen pendientes sin diferencias significativas. La captación de Etd+ de ambos tipos de miofibras fue inhibida por La3+. B. Razones de captaciones de Etd+ de miofibras aisladas de ratones post inoculación de células B16F0 7d y 14d. C. Curvas de captación en tiempo real de Etd+ en miofibras aisladas de ratones denervados Cx43fl/fl Cx45fl/fl y Cx43fl/fl Cx45fl/fl:MC, KO músculo-específico comparados con ratones Cx43fl/fl Cx45fl/fl inyectados con células de melanoma. Se observa que sólo aumenta la permeabilidad del sarcolema en las fibras denervadas de ratones que expresan Cxs. D. Tasas de captaciones calculadas de las curvas representadas en C, la tasa de captación de colorante solo aumentó en ratones Cx43fl/fl Cx45fl/fl. (**** p<0,0001, n = 5).

39
5.1.4. El PMR de las miofibras de los músculos esqueléticos de ratones tratados con
células B16F0 no se altera.
A pesar de no observarse un aumento de la permeabilidad a Etd+ en las fibras musculares
aisladas de ratones tratados con células B16F0, se evaluó el PMR como una estrategia de
medición de permeabilidad más sensible que la captación de colorantes, por depender del
flujo de iones. Al estar la membrana permeable de manera inespecífica, como sería en el
caso de presentar HCs-Cxs activos, los iones difundirían a través de la membrana a favor de
su gradiente electroquímico, alterando el equilibrio iónico final, llegando a un nuevo estado
estable, con un nuevo PMR. Además, anteriormente se ha observado que la acumulación
neta de [Na+]i depende de la actividad de HCs-Cxs en fibras de ratones denervados y precede
a la atrofia muscular (Cisterna, BA et al. 2016), por lo que el PMR del músculo debería estar
alterado. Para resolver este objetivo, se realizaron registros de PMR con microelectrodos en
músculo FDB in vivo de ratones tratados con células B16F0 a los 7 y 14 días post-inyección de
las células. En estos ensayos nuevamente se utilizó, como control positivo (datos no
publicados), ratones denervados Cx43fl/flCx45fl/fl y Cx43fl/flCx45fl/fl:Myo-Cre, a los cuales se les
midió el PMR in vivo. El PMR de miofibras de ratones inyectado con células B16F0 a los 7 y
14 días post-inyección fue similar al de miofibras de ratones controles (˗77,1 ± 1,2 mV y ˗75,0
± 1,1 mV, respectivamente vs. ˗75,5 ± 0,6 mV, n=3 ratones, 30 registros por n, p=0,99),
mientras que las miofibras de ratones que fueron denervados y registrados a 7 y 14 días
denervación, mostraron caídas significativas en el PMR comparados con el valor registrado

40
en miofibras de ratones controles (˗55,7 ± 0,7 mV y ˗55,8 ± 0,6 mV vs. ˗75,5 ± 0,6 mV, n=3
ratones, 30 registros por n, p<0,0001). Esta caída de PMR no se observó en miofibras de
ratones Cx43fl/flCx45fl/fl:Myo-Cre denervados, evaluados en ambos periodos de tiempo (˗55,7
± 0,7 mV y ˗55,8 ± 0,6 mV, respectivamente vs. ˗75,9 ± 0,6 mV y 75,8 ± 0,6 mV,
respectivamente, n=3 ratones, 30 registros por n, p<0,0001) registrando valores similares de
PMR en miofibras de ratones controles y tratados con células B16F0 (Fig. 4).

41
Fig.4: El PMR de miofibras esqueléticas in vivo no se altera en ratones inoculados con células de melanoma B16F0. A. Valores representativos de potenciales de membrana en reposo (PMR) en los tres tratamientos registrados mediante microelectrodo en músculo FDB. Se utilizaron ratones denervados para la comparación de caída de PMR en músculo atrofiado que expresan Cxs. B. PMR. (n= 5, 30 registros por experimento o animal ****p<0,0001). Ratones Cx43fl/flCx45fl/fl :MC no expresan Cx43/Cx45 en las miofibras esqueléticas, mientras que las miofibras de ratones Cx43fl/flCx45fl/fl denervados si expresan y fueron utilizadas como controles de la línea mutante.
A B
4 0
6 0
8 0
1 0 0
D e n e rv a c ió n
7 1 4 7 1 4
B 1 6 F 0
7 1 40D ía s
C x 4 3fl/fl
C x 4 5fl/fl
:M C
C x 4 3fl/fl
C x 4 5fl/fl
PM
R (
-mV
)
4 0
6 0
8 0
1 0 0
PM
R (
- m
V)
D e n e rv a c ió n
7 1 4 7 1 4
B 1 6 F 0
7 1 40D ía s
* * * *
C x 4 3f l/ f l
C x 4 5f l/ f l
C x 4 3f l/ f l
C x 4 5f l/ f l
:M C

42
5.1.5. Las Cxs 39, 43 y 45 no cambian su expresión en miofibras de músculos
esqueléticos de ratones inoculados con células de melanoma B16F0.
Ante la ausencia de efecto de la inoculación de células de melanoma B16F0 sobre la
permeabilidad de los fibras musculares, evidenciado con los registros de captación de
colorante y PMR, y ante la falta de participación de las Cxs en la atrofia muscular evidenciada
en ratones Cx43fl/flCx45fl/fl:Myo-Cre, que no expresan Cx43 y Cx45, aún podría ser posible que
Cx43, Cx45 y Cx39 se expresaran de igual forma bajo estas condiciones inflamatorias, tal
como se ha observado anteriormente en otras condiciones como denervación o en respuesta
a glucocorticoides, pero que en los tiempos evaluados todavía no estén funcionales como
HCs-Cxs o su actividad estuviera fuertemente inhibida en ratones con cáncer. Sin embargo,
inmunofluorescencias realizadas en cortes transversales de músculo TA tanto para Cx39,
Cx43 como Cx45 obtenidos de ratones inoculados con células B6F0 durante 7 y 14 días, no
mostraron diferencias significativas con respecto a condiciones control. Por lo tanto, la
ausencia de cambios de permeabilidad del sarcolema y la atrofia de los músculos de ratones
KO para Cx43/45 revelan que en la atrofia muscular inducida por cáncer causado por las
células B16F0 (Fig.5) no se explica por cambios en la expresión de Cxs.

43
Fig.5: La inoculación de células B16F0 no induce la expresión de las Cxs 39, 43 y 45 en músculo TA. Detección por inmunofluorescencia de Cx39, Cx43 y Cx45 en cortes de músculo TA de ratones a los 7 y 14 días post inoculación con células B16F0 (n=5). Cortes de TA de ratones a los 7 días post-denervación (der.) como control positivo. Azul: DAPI. Barra: 50 µM.

44
5.1.6. Los ratones que no expresan Cxs 43 y 45 en miofibras exhiben
significativamente menor atrofia muscular esquelética inducida por
endotoxemia
Otra condición de atrofia muscular esquelética asociada a un cuadro inflamatorio agudo es
sepsis. En este caso, el modelo elegido fue ratones sometidos a LCP, condición que asemeja a
la condición de humanos con peritonitis, causada por infección polimicrobiana en el espacio
peritoneal, la cual es una condición grave que lleva en poco tiempo a shock séptico (Siempos,
II et al. 2014). Como se ha descrito anteriormente para ratones WT sometidos a LCP
(Gosemann, JH et al. 2010, Kaynar, AM et al. 2014), los ratones, tanto Cx43fl/flCx45fl/fl como
Cx43fl/flCx45fl/fl:Myo-Cre, sometidos a LCP, presentaron una rápida y significativa diferencia
de baja de peso (Δg) hasta el día dos post-operación, comparado con el peso inicial a tiempo
0 (˗2,9 ± 0,3 g y ˗2,5 ± 0,4 g, respectivamente, n=8) sin observarse diferencias significativas en
el Δg entre estas dos cepas de ratones, mientras que los ratones Sham, registrados en el
mismo tiempo, tuvieron un Δg significativamente menor (˗0,87 ± 0,2 g y ˗0,96 ± 0,3,
respectivamente, n=8), sin observarse también diferencias ente las cepas de ratón. El Δg final
alcanzado en el día 2 por los ratones con LCP se mantuvo hasta el día 7, sin diferencias
significativas entre ratones Cx43fl/flCx45fl/fl y Cx43fl/flCx45fl/fl:Myo-Cre pero siendo
significativamente mayor que el Δg de ratones Sham (Fig. 6A).
Para estudiar la atrofia muscular en ratones sometidos a LCP, se realizaron análisis de
AST de fibras de músculo TA, al igual que se realizaron en músculos de ratones tratados con

45
células B16F0 (Fig. 6B). Las mediciones de AST revelaron una atrofia muscular significativa, de
aproximadamente 50%, en ratones Cx43fl/flCx45fl/fl sometidos a LCP comparados con ratones
Sham, mientras que en ratones Cx43fl/flCx45fl/fl:Myo-Cre, la atrofia muscular fue de un total
de 20% aprox., la cual es significativamente menor que la observada en ratones
Cx43fl/flCx45fl/fl, lo que representa un 30% aprox. menos de disminución de AST de las fibras,
por lo que ese porcentaje de atrofia está directamente relacionado con la expresión
muscular de Cxs (Fig. 6C).

46
Fig.6. Los ratones KO de Cx43 y Cx45 en miofibras esqueléticas manifiestan menor atrofia muscular esquelética inducida por sepsis. A. Curvas de peso corporal promedio bruto (izq.) y neto (der.) de ratones Cx43fl/fl Cx45fl/fl:MC y Cx43fl/fl Cx45fl/fl sometidos a ligación y punción cecal (sepsis) hasta el día 7 post-cirugía. La LCP causó una pérdida de aprox. un 20% del peso en ambos tipos de ratones. B Cortes transversales de músculo TA en ratones Cx43fl/fl Cx45fl/fl:MC y Cx43fl/fl Cx45fl/fl teñidos con hematoxilina/eosina. El gráfico ilustra la disminución del AST de las fibras de los ratones sometidos a LCP (izq.) en relación al valor de las miofibras de músculos de animales controles. En ratones que expresan Cx43 y Cx45, LCP disminuyó aproximadamente un 50% con respecto a su control. Miofibras de ratones Cx43fl/fl Cx45fl/fl:MC presentaron una disminución de aprox. un 20% lo que evidencia un 30% aprox. de protección contra la atrofia muscular inducida por LCP. Barra: 50 µM.

47
5.1.7. Las miofibras de músculo TA de ratones sometidos con endotoxemia expresan
Cx39, Cx43 y Cx45
Los resultados anteriores sugieren fuertemente que la atrofia del músculo TA inducida por
LCP depende de la expresión de Cxs, al igual que en modelos de atrofia muscular inducida
por denervación (Cea, LA et al. 2013) o glucocorticoides (Cea, LA et al. 2016), por lo que se
realizaron ensayos de inmunofluorescencia para detectar la presencia de Cxs en cortes
transversales de músculos de ratones Cx43fl/flCx45fl/fl y Cx43fl/flCx45fl/fl:Myo-Cre bajo
tratamiento con LCP. En músculo TA de ratones Cx43fl/flCx45fl/fl a 7 días post-LCP, se detectó
marca de inmunofluorescencia positiva para Cx39, Cx43 y Cx45. Como se esperaba, la marca
de inmunofluorescencia para Cx43 y Cx45 no se observó en ratones Cx43fl/flCx45fl/fl:Myo-Cre,
tanto en condiciones controles como a los 7 días post-LCP (Fig. 7). Sin embargo, la marca
para Cx39 fue positiva en ratones Cx43fl/flCx45fl/fl:Myo-Cre con LCP al igual que en ratones
Cx43fl/flCx45fl/fl bajo el mismo tratamiento (Fig. 7).

48
Fig.7. LCP induce la expresión de Cx39, Cx43 y Cx45 en miofibras de músculo TA de ratones Cx43fl/fl Cx45fl/fl. Panel de inmunofluorescencias en cortes transversales de TA para Cx43, Cx45 y CX39 en ratones Cx43fl/fl Cx45fl/fl y Cx43fl/fl Cx45fl/fl:MC, se puede observar que en miofibras de ratones Cx43fl/fl Cx45fl/fl:MC sólo se expresa Cx39. En azul se marcan los núcleos con DAPI. Barra: 20 µM.

49
5.1.8. Las miofibras de ratones con endotoxemia presentan menor PMR y aumento en
la captación de colorantes.
Ya que se observó la expresión de Cx39, Cx43 y Cx45 en miofibras de músculos de ratones
con LCP, queda por determinar si la pérdida de PMR y aumento de permeabilidad observada
anteriormente (Liu, SH et al. 1997, Lin, MC et al. 1998) depende de la expresión y función de
HCs-Cxs, al igual a lo observado en ratones denervados (Cisterna, BA et al. 2016) o sometidos
a glucocorticoides (Cea, LA et al. 2016), en donde la caída del potencial de membrana y el
aumento de permeabilidad de las miofibras es dependiente de la expresión y función de HCs-
Cxs. Como se esperaba, ratones WT sometidos a LCP tuvieron una baja significativa cercano
al 10% de PMR 24 h post-LCP, comparado con ratones sham (˗78,0 ± 1,5 mV vs. ˗71,0 ± 0,8
mV, respectivamente, n=3, 30 fibras por n, p<0,001). Del mismo modo las miofibras de
músculos de ratones Cx43fl/flCx45fl/fl sometidos a LCP presentaron una baja de PMR similar y
significativa con respecto a su control (˗80,0 ± 1,8 mV vs. ˗63,0 ± 1,7 mV, respectivamente,
n=3, 30 fibras por n p<0,001). Sin embargo, en miofibras de ratones Cx43fl/flCx45fl/fl:Myo-Cre
sometidos a LCP no se observó la baja de PMR en el mismo período de tiempo,
manteniéndose sin diferencias significativas comparado con el valor registrado en miofibras
de ratones controles. Así mismo, se observó un aumento significativo de la captación de Etd+
en fibras aisladas de ratones WT sometidos a LCP en comparación a ratones control (0,15 ±
0,03 vs. 0,39 ± 0,06 UA/min, respectivamente, n=3, p<0,001) la cual es inhibida
significativamente por La3+ (200µM) aplicado de forma aguda (0,39 ± 0,06 vs. 0,06 ± 0,04

50
UA/min, respectivamente, n=3, p<0,001). Además, no se observó este aumento de captación
de Etd+ en miofibras de músculos de ratones Cx43fl/flCx45fl/fl:Myo-Cre, los cuales presentaron
una captación similar a la de miofibras de los ratones WT en condiciones basales (0,14 ± 0,04
vs. 0,15 ± 0,03, respectivamente, n=3, p>0,9).

51
Fig.8 LCP induce caída del PMR y aumento de la permeabilidad del sarcolema de miofibras del músculo FDB controles, pero no en músculos deficientes en la expresión de Cx43/Cx45. A. (Arriba a la izquierda) Registros representativos de PMR obtenidos con microelectrodos en miofibras de músculo FDB de ratones a las 24 h post-LCP, en donde cada punto es una medición individual. (Abajo a la izquierda) PMR bajo las mismas condiciones detalladas en el gráfico anterior (n=3, 30 mediciones por grupo). B. (Arriba a la derecha).Registro representativo de captación de Etd+ en tiempo real de fibras aisladas de músculo FDB, de ratones tratados con LCP y controles, solo se ve el aumento significativo en la captación de Etd+ en ratones WT bajo LCP, la cual es inhibida por La3+, mientras que en ratones Cx43fl/flCx45fl/fl:Myo-Cre no se observa el aumento de la captación de Etd+.(Abajo a la derecha) Tasas de captación de Etd+ de fibras aisladas de ratones 24 h post-LCP (n=3). ***p<0,001; ****p<0,0001.

52
5.1.9. La expresión y actividad de HCs-Cxs disminuye el PMR de células HeLa.
Para determinar si específicamente la expresión HCs-Cxs funcionales puede alterar el PMR de
las fibras musculares esqueléticas en condiciones de inflamación o desuso, como en el caso
los ratones denervados, se realizaron registros de PMR con microelectrodos en cultivos de
células aisladas HeLa-P y transfectadas con Cx39, Cx43 o Cx45, las cuales se expresan en el
músculo atrófico post denervación. De nuestro conocimiento, estudios similares no se han
realizado utilizando otros HCs-Cxs, con la excepción de lo reportado sobre la débil influencia
que tiene la expresión de Cx43 en células HeLa sobre el PMR (Contreras, JE et al. 2003). El
PMR registrado en células HeLa-P mantenidas en solución salina normal (-24 ± 1 mV, n=40) se
mantuvo sin diferencias significativas en condiciones que podrían alterar la función de los
HCs-Cxs como la incubación con SLCD (-21,6 ± 1,5 mV, n=40, p>0,9), SLCD + La3+ (-22,5 ± 0,4
mV, n=30, p>0,9) y La3+ (-24 ± 1 mV, n=25, p> 0,9).
En la mayoría de las células, la bomba Na+/K+ ATPasa contribuye a la mantención del
PMR, y su inhibición causa despolarización de la membrana celular (Redmann, K y Walliser, S
1981). De acuerdo con el rol de la bomba Na+/K+ ATPasa sobre el PMR, en células HeLa-P el
tratamiento de 30 min con ouabaina (0,1 μM), un inhibidor de la bomba Na+/K+ ATPasa
(Clausen, T et al. 1983), redujo el PMR de forma significativa tanto en solución extracelular
control (˗13,0 ± 0,3 mV, n=40, p<0.0001) como en SLCD + ouabaina (˗10,7 ± 0,6 mV, n=25,
p<0,0001).
A diferencia de las células HeLa-P que no expresan Cxs, células que expresan Cxs
como las células HeLa-Cx43 y HeLa-Cx45, bañadas en solución salina normal presentan PMR

53
significativamente más bajos que HeLa-P bajo las mismas condiciones (˗15,0 ± 0,4 mV, n=53 y
˗18,0 ± 0,4 mV, n=90 vs. -24 ± 1 mV, n=40, respectivamente, p<0,0001), lo que indica que la
expresión por si sola de las Cxs estudiadas y su actividad basal como HCs podría ser suficiente
para lograr un nuevo estado estacionario iónico. Sin embargo, el PMR células HeLa-Cx39 no
presentó diferencias significativas con respecto al de células HeLa-P mantenidas en
condiciones controles (˗19,7 ± 1,7 mV, n=30 vs. -24 ± 1 mV, n=40, p>0,09). La incubación por
30 min con SLCD, condición conocida por aumentar la probabilidad de apertura de los HCs-
Cxs, en células HeLa-Cxs el PMR disminuyó significativamente comparado con el registrado
en condiciones controles, tanto en células HeLa-Cx39 (˗19,7 ± 1,7 mV, n=30 vs. ˗9,0 ± 1,1 mV,
n=25, p<0,0001), HeLa-Cx43 (˗15,0 ± 0,4 mV, n=53 vs. ˗8,5 ± 0,3 mV n=60, p<0,0001) como en
células HeLa˗Cx45 (˗18,0 ± 0,4 mV, n=90 vs. ˗11,0 ± 0,4 mV, n=25, p<0,0001), indicando que
los HCs-Cx39 son inhibidos por cationes divalentes como el Ca2+, como ha sido demostrado
anteriormente para otros HCs-Cxs (DeVries, SH y Schwartz, EA 1992, Pfahnl, A y Dahl, G 1999,
Kondo, RP et al. 2000, Contreras, JE et al. 2003, Gómez-Hernández, JM et al. 2003, Sáez, JC et
al. 2005, Lopez, W et al. 2014, Zonta, F et al. 2014). La disminución del PMR inducido por
SLCD en células HeLa transfectadas con Cxs fue evitado mediante la co-incubación de
SLCD+La3+(200 µM, 30 min), un conocido bloqueador de HCs-Cxs (Sáez, JC et al. 2005,
D'Hondt, C et al. 2009), quedando los valores de PMR similares y sin diferencias significativas
con respecto a condiciones controles tanto en células HeLa-Cx39 (˗21,0 ± 0,9 mV, n= 30 vs.
˗19,7 ± 1,7 mV, n=30 p>0,9), HeLa-Cx43 (˗18,3 ± 1,0 mV, n=30 vs. ˗15,0 ± 0,4 mV, n=53, p>
0,96) como HeLa-Cx45 (˗21,0 ± 0,4 mV, n=40 vs. ˗18,0 ± 0,4 mV, n=90, p>0,99). Por otro lado,
en comparación con condiciones controles, la incubación con La3+ (200µM, 30 min) indujo

54
hiperpolarización de la membrana celular: el PMR en células HeLa-Cx39 (˗19,7 ± 1,7 mV, n=30
vs. ˗27,0 ± 1,8 mV, n=30, p<0,0001), HeLa-Cx43 (˗15,0 ± 0,4 mV, n=53 vs. ˗28,5 ± 2,1 n=25,
p<0,0001) y HeLa-Cx45 (˗18,0 ± 0,4 mV, n=90 vs. ˗25,4 ± 1,4 n=25, p<0,0001) fue afectado de
modo similar. Sin embargo, la hiperpolarización de la membrana inducida por La3+ en células
HeLa transfectadas con Cxs, no difirieron significativamente de los valores de PMR
registrados en células HeLa-P, tanto incubadas con La3+ como medio salino normal. Por otro
lado, la incubación con ouabaina (0,1 µM, por 30 min) indujo una despolarización
significativa comparada con las condiciones controles en células HeLa-Cx39 (˗8,6 ± 0,6 mV
n=25 vs. ˗19,7 ± 1,7 mV, n=30 p<0,0001), HeLa-Cx43 (˗11,0 ± 0,5 mV n=30 vs. ˗15,0 ± 0,4 mV,
n=53, p=0,03) y HeLa-Cx45 (˗11,1 ± 0,6 mV n=30 vs. ˗18,0 ± 0,4 mV, n=90, p<0,0001), sin
embargo esta despolarización no es significativa si se compara con la despolarización
inducida por SLCD en los mismos tipos celulares. Finalmente, se indujo la despolarización por
la incubación de SLCD + ouabaina para observar si el efecto de estas condiciones se potencia.
Sólo en células HeLa-Cx45 se registró una despolarización significativa en respuesta al efecto
de la aplicación simultanea de SLCD + ouabaina comparado con el efecto de la aplicación de
sólo SLCD (˗5,3 ± 0,3 mV, n=40 vs. ˗11,0 ± 0,4 mV, n=25, p=0,0004). La disminución del PMR
en HeLa-Cx43 y HeLa-Cx39 tratadas con SLCD + ouabaina no mostró diferencia significativa
comparado con la caída de PMR inducida por SLCD (Fig. 9).
En resumen, LCP induce la expresión de Cx39 en músculo esquelético de ratones
Cx43fl/flCx45fl/fl:Myo-Cre (Fig. 7), sin embargo, en estos ratones no se observa el componente
dependiente de Cxs sobre la atrofia muscular, o el aumento de captación de colorante y

55
disminución del PMR (Fig. 8), a pesar de la continua la expresión de Cx39 (Fig.6). Este
fenómeno también se observa en fibras de músculos denervados en el mismo tipo de ratón,
considerando también que en ellos el Ca2+ y Na+ no se acumulan dentro de la miofibra por
efecto de la ausencia de la expresión de Cx43 y Cx45 en el músculo de ratones KO-músculo-
específico (Cisterna et al 2016), pero aún mantienen la expresión de Cx39. Hasta la fecha, no
se conoce como podría participar la Cx39 en la atrofia muscular por desconocimiento de la
función que tendría Cx39 como hemicanal y de sus propiedades de permeabilidad, por lo que
fue necesario caracterizarlo en un sistema aislado y reconstruido establemente en células
HeLa.

56
Fig.9: La SLCD induce caída del PMR en células HeLa que expresan Cx43, Cx45 o Cx39, respuesta que es prevenida con La3+. Se utilizó células HeLa sembradas subconfluentemente y aisladas unas de otras, y el PMR se midió con microelectrodos. Ouabaina (0,1 µM) se utilizó como control para evidenciar la caída de PMR en las HeLa-P por inhibición de la bomba Na+/K+ ATPasa. (Exp. realizado para esta tesis y gráfico modificado del presentado por Saavedra F. en tesis de pregrado bioquímica Univ. de Valparaíso)

57
5.2. Caracterización de Cx39 y su función como hemicanal en células HeLa.
5.2.1. Las células Hela-Cx39 no forman CUH funcionales.
Todas las células HeLa-Cx39 muestran inmunoreactividad positiva principalmente como una
marca intracelular difusa, en pequeños parches en la periferia celular y en aparentes
interfaces célula-célula (Fig.10 der. marcada por flechas), por lo que no se descarta la
posibilidad que formen uniones funcionales. A pesar de que ya se ha descrito la ausencia de
transferencia de colorantes, como LY-2, DAPI2+, IP2+, Etd+ o Neurobiotina en células HeLa-Cx39
(von Maltzahn, J et al. 2004) quedaba por determinar si células que expresan Cx39,
establecen acoplamiento eléctrico, parámetro mucho más sensible para detectar la
comunicación a través de canales intercelulares que pudiera formar la Cx39, y que se puede
evidenciar registrando el traspaso de iones a través de estos canales. La aplicación de pulsos
rectangulares de voltaje de 20 mV (desde -100 a 0 mV con 200 ms de duración) en células
HeLa-Cx43-EGFP generan una evidente Ij la cual no es detectada entre células HeLa-Cx39
(n=9) (Fig. 10). Los valores de Ij registrados en HeLa-Cx39 fueron cero con todos los voltajes
aplicados. De acuerdo con lo previamente descrito (von Maltzahn, J et al. 2004) la
transferencia intercelular de DAPI2+ fue negativa (Fig. 10A der). En células HeLa-Cx43-EGFP,
el cálculo de gj (conductancia macroscópica) de los canales en hendidura, desde el valor de Ij
registrado en la célula no estimulada a diferentes voltajes, produjo valores similares dentro
del rango de voltaje ilustrado (-40 mV a +40 mV), indicando la ausencia de dependencia de

58
voltaje. Por otro lado, el gj entre células HeLa-Cx39 a cualquier voltaje aplicado fue cero (Fig.
10C), revelando la ausencia de flujo de iones entre células HeLa-Cx39 vecinas.

59
Fig.10. Las células HeLa-Cx39 no forman uniones de hendidura funcionales. A y B, contraste de fase (izquierda) e inmunofluorescencia de Cx39 (A, derecha) en HeLa-Cx39; y fluorescencia Cx43-EGFP en HeLa-Cx43EGFP (B, derecha). Barra escala: 20 μm. C Ij a diferentes voltajes (izquierda) y gj normalizada a diferentes voltajes (derecha) utilizando mediciones en “dual voltage-clamp” en células HeLa-Cx39 y HeLa-Cx43EGFP. Los valores de Ij para las células HeLa-Cx39 fueron cero a todos los voltajes estudiados. Registros de la transferencia de la corriente intercelular de célula1 (C1) a célula2 (C2) y viceversa en células HeLa-Cx39 (debajo de las células mostradas en A) y en HeLa-Cx43EGFP (bajo las células mostradas en B) bajo pulsos de voltaje (aumentando en 20 mV de -100 a 0 mV con una duración de 200 ms, trazos de fondo izquierdo) aplicadas a C1 y C2, respectivamente (N = 9).

60
5.2.2. Un aumento de actividad de los HCs-Cx39 incrementa la corriente macroscópica
de la membrana en células HeLa-Cx39.
Para probar que la SLCD incrementa la probabilidad de apertura de HCs-Cx39, se
registraron las corrientes de membrana, macroscópicas y microscópicas, de células HeLa-
Cx39 utilizando configuración de fijación de voltaje en célula completa. La corriente de
membrana registrada (Fig. 11B) y la pendiente de corriente (tasa de corriente) calculada
desde la curva corriente vs. tiempo (Fig. 11B inserto izq.) bajo condiciones controles fueron
bajos y similares a los valores de células HeLa-P (0,016 ± 0,008 pA/ms vs. 0,013 ± 0,005
pA/ms, respectivamente p > 0,05 n= 10). Sin embargo, la tasa de corriente de membrana de
células HeLa-Cx39 se incrementó significativamente bajo la exposición de SLCD,
particularmente a potenciales de membrana positivos (> +25 mV) y disminuyó drásticamente
bajo el tratamiento con La3+, (0,080 ± 0,006 pA/ms vs. 0,020 ± 0,001 pA/ms,
respectivamente, n= 5 p < 0,0001). Cabe señalar que la curva I/V registrada en condiciones
controles fue comparable con la registrada en SLCD + La3+ (Fig. 11B), sugiriendo que la
mayoría si no todo, el incremento de la corriente de membrana inducida por la SLCD fue
debido a la apertura de HCs-Cx39. En todas las condiciones, las corrientes macroscópicas a 0
mV, I=0, indicando que los HCs-Cx39 no tienen selectividad iónica, como los HCs-Cx43
(Kondo, RP et al. 2000, Contreras, JE et al. 2003).
Bajo la aplicación de rampas de voltaje (-80 mV a +80 mV), las curvas de corriente,
mostraron eventos unitarios que fueron frecuentemente observadas a potenciales positivos

61
en células HeLa-Cx39 expuestas a SLCD (Fig. 11B). Los valores de transiciones discretas de
corriente fueron convertidas punto por punto a valores de conductancia, revelando valores
de conductancia unitarios de ~75 pS (Fig. 11B, inserto der.). En SLCD la probabilidad de
apertura de los eventos unitarios de corriente se incrementaron con respecto a las
condiciones controles. En células HeLa-P, la aplicación de SLCD no indujo la aparición de
eventos unitarios discretos de corriente como se ha reportado previamente (Contreras, JE et
al. 2003, Retamal, MA et al. 2007, Sánchez, HA et al. 2009)

62
Fig.11. La SLCD incrementa la corriente macroscópica de membrana en células HeLaCx39. Curva representativa I/V de membrana obtenida bajo la aplicación de una rampa de voltaje desde -80 a +80 mV en células HeLa-Cx39 bajo condiciones controles (Control), SLCD o SLCD-La3+ (200 µM), un bloqueador de HCs-Cxs. La corriente macroscópica a V=0 mV fue cero en todas las condiciones. Inserto a la derecha, muestra una corriente unitaria (rectángulo de líneas discontinuas) que se traduce en una conductancia unitaria de aprox. 75 pS, registrado en la membrana de células HeLa-Cx39. Inserto a la izquierda, muestra la curva de corrientes macroscópicas bajo diferentes condiciones indicadas en la figura (n=10 ****p<0,0001).

63
5.2.3. Los HC-Cx39 aumentan su actividad en ausencia de cationes divalentes
extracelulares y presentan conductancia unitaria característica.
Los eventos de corriente unitarios también fueron evaluados usando pulsos de voltaje
rectangulares. Células HeLa-P que fueron fijadas a -60 mV en solución salina control y en
SLCD no mostraron eventos de corriente unitaria (Fig. 12A). En contraste, en células HeLa-
Cx39 los eventos unitarios con una probabilidad de apertura bastante baja (Po = 0.3), fueron
evidentes después de fijar el voltaje de membrana a +60 mV por cerca de 20 s. Sin embargo,
en células expuestas a SLCD, eventos discretos de corriente unitaria mostraron una alta
probabilidad de apertura (Po = 0.9) y fueron completamente bloqueados por La3+ (Fig. 12A).
La mayoría de los eventos discretos registrados en células HeLa-Cx39 presentaron valores de
eventos unitarios de ~75 ± 5 pS (n = 40) en condiciones controles como en SLCD (Fig. 12A,B) y
los tiempos de transición entre el estado completamente cerrado a completamente abierto,
y vice-versa, de los HCs-Cx39 no fueron significativamente diferentes (21,0 ± 0,5 ms vs. 20,4 ±
0,5 ms, P > 0,05, n = 30) (Fig. 12C), pero ambos valores fueron más bajos que los registrados
en HeLa-Cx43-EGFP (full-closed to full-open: 27,2 ± 1,9 ms vs. 32,6 ± 2,8 ms, P < 0,05, n = 30
and full-open to full-closed: 20,4 ± 0,5 ms vs. 32,6 ± 2,8 ms, P < 0,0001, n = 30) (Fig. 12C).

64
Fig12. La SLCD incrementa la probabilidad de apertura de los HCs-Cx39. A. Eventos de corriente unitaria registrados bajo voltage-clamp a +60 mV en células HeLa-P (dos trazos superiores) o células HeLa-Cx39 bajo las condiciones indicadas. La frecuencia de eventos registrada bajo cada condición se muestra a la derecha de cada trazo. B. Gráfico de frecuencia de conductancias obtenidas en células HeLa-Cx39 en SLCD. EL evento de conductancia unitaria más frecuente registrado fue 75 ± 5 pS en células bañadas con SLCD (n=40 células), y fue bloqueado por La3+ (n=10). C. Tiempo de retardo de apertura y cierre de los HCs-Cx39 and -Cx43-EGFP (*** P<0,0005, n=30).

65
5.2.4. Los HCs-Cx39 permiten el paso de un grupo selectivo de moléculas con
diferentes propiedades fisicoquímicas
Diferentes colorantes (ej.: DAPI2+, Etd+ y IP2+) han sido usados anteriormente para
observar cambios en la permeabilidad de membrana debido a la presencia de HCs-Cxs
(Orellana, JA et al. 2011, Hansen, DB et al. 2014). Para identificar las características de
permeabilidad de los HCs-Cx39, se realizaron ensayos de captación de colorante en tiempo
real, usando moléculas con diferentes masas y cargas netas que fluorescen al intercalarse
con ácidos nucleicos intracelulares (Tabla 1 and Fig. 13). Las células Hela-P no mostraron
cambios en la captación de Etd+ bajo la exposición a SLCD (Fig. 13, inserto). Similarmente, las
células HeLa-Cx39 en presencia de cationes divalentes mostraron baja captación de Etd +y
IP2+. Las células HeLa-Cx39 expuestas a SLCD, mostraron un drástico aumento de captación
de Etd+ que fue completamente bloqueado por La3+ (200 μM) (Fig. 13, inserto). De hecho, la
tasa de captación de colorante de todos los trazadores usados en células HeLa-Cx39 fue baja
en condiciones control, pero la tasa de captación de varios colorantes incrementó
significativamente bajo la exposición a SLCD; Etd+, DAPI+2, AE˗4 hexidio, AM+1 y procion
orange MX2R, que también fue bloqueada por La3+ (Fig. 13). En contraste, la tasa de
captación de IP2+, mostró una tendencia de aumento menor en SLCD, que fue reducida por
La3+ (no significativo), mientras que la tasa de captación de H-etd4+ y BBG- no fue
significativamente afectada por la SLCD.

66
Fig. 13. La captación de colorante es incrementada por SLCD. Tasa de captación de diferentes moléculas con diferente masa y carga neta (lista inferior) de células HeLa-Cx39 bajo cada condición indicada (n=6 para cada colorante). Inserto muestra captación representativa de Etd+ e IP2+, en células HeLa-Cx39, respectivamente, y Etd+ en células HeLa-P. Después de 5 min en condición control, la solución extracelular fue rápidamente cambiada por SLCD y la captación de colorante fue registrada por 8 min seguido por adicionales 5 min de registro en presencia de 200 µM La3+. (****P<0,0001 y ***p<0,0005 comparado con los valores basales en cada caso; n=4, con 30 células registradas en cada caso). UA: unidades arbitrarias.

67
Tabla1. Captación de colorante de células HeLaCx39 vía HCs-Cx39
Nombre comercial P.M. Carga neta
Captación
Inhibido por
bloqueador de HCs
ACMA (9-Amino-6-Chloro-2-
Methoxyacridine) 258,7 + - n/a
Azul de metileno 319,8 + + +
DAPI 350,2 2+ + +
Bromuro de etidio 394,3 + + +
Ioduro de hexidio 497,4 + + +
Amarillo nuclear 651,0 + - n/a
Ioduro de propidio 668,3 2+ - n/a
Procion orange MX2R 715,5 3- + +
Verde de indocianina 774,9 - + +
BBG 854,0 - - n/a
Homodímero de etidio 856,7 4+ - n/a
Azul de Evans 960,8 4- + +
BOBO™-1 Ioduro 1.202,6 4+ - n/a
7-AAD (7-Aminoactinomicina D) 1.270,4 0 + +
YOYO®-1 Ioduro 1.270,7 4+ + +
TOTO®-1 Ioduro 1.302,8 4+ + +
n/a: No aplica

68
5.2.5. Electronegatividad, potencial de ionización, flexibilidad, valencia, tamaño,
polarizabilidad y geometría son características claves para definir el paso de
moléculas a través de los HCs-Cx39
Las moléculas anteriormente evaluadas para conocer la selectividad de los HCs-Cx39
difieren notoriamente en carga neta, tamaño o forma. Sin embargo, estos criterios de
selección no responden la pregunta de por qué pasan a través de los HCs-Cx39. Para superar
esta limitación, se usó una aproximación bioinformática para explorar las propiedades físico-
químicas de los diferentes trazadores de permeabilidad. Se identificaron 11 de 1.000
descriptores moleculares (Tabla 2) que clasifican los trazadores en términos de su habilidad
para pasar a través de los HCs-Cx39 (Tabla1) entre las propiedades fisicoquímicas
seleccionadas, las más frecuentes pertenecen a seis categorías diferentes: (1)
electronegatividad, (2) potencial de ionización, (3) polarizabilidad, (4) tamaño y geometría,
(5) flexibilidad y (6) valencia. Mientras que los valores de algunos descriptores moleculares
fueron altos, como los de electronegatividad (TDB6e, ATSC5s y GATS5s), de potencial de
ionización (TDB8iy TDB10i), tamaño y geometría (TDB6v yTDB5u), otros fueron más
pequeños (RotBtFrac y VP-6) para moléculas que pasan a través de HCs-Cx39 comparado a
moléculas que no pasan (Tabla1). Ya que, algunos valores de varios descriptores son
característica común para todas las moléculas que pasan a través de los HCs-Cx39, mientras
que otras son características de las que no pasan, el valor de un solo descriptor no es
suficiente para establecer si una molécula en particular pase a través de los HCs-Cx39. Por lo

69
tanto, el paso de los colorantes a través de los HCs-Cx39 de los colorantes podría ser una
propiedad emergente obtenida desde una combinación no-lineal de varias propiedades
fisicoquímicas, enumeradas anteriormente.…………………………………………………………………………….
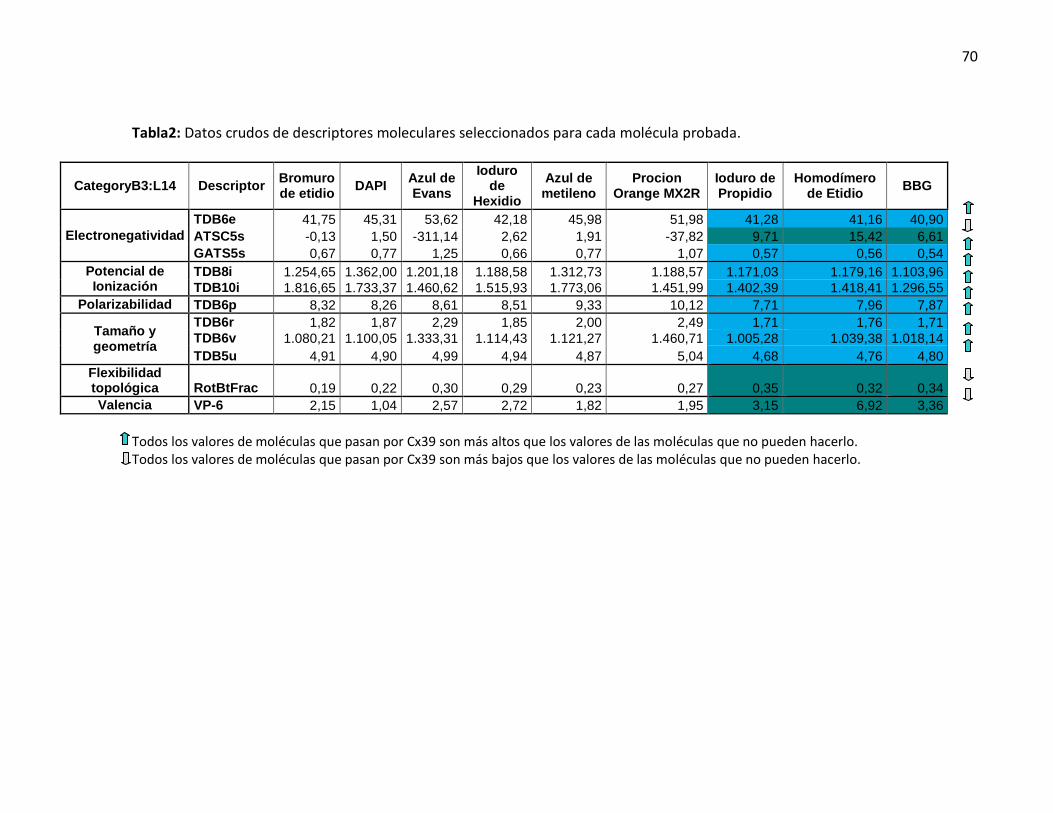
70
Tabla2: Datos crudos de descriptores moleculares seleccionados para cada molécula probada.
CategoryB3:L14 Descriptor Bromuro de etidio
DAPI Azul de Evans
Ioduro de
Hexidio
Azul de metileno
Procion Orange MX2R
Ioduro de Propidio
Homodímero de Etidio
BBG
Electronegatividad
TDB6e 41,75 45,31 53,62 42,18 45,98 51,98 41,28 41,16 40,90
ATSC5s -0,13 1,50 -311,14 2,62 1,91 -37,82 9,71 15,42 6,61
GATS5s 0,67 0,77 1,25 0,66 0,77 1,07 0,57 0,56 0,54
Potencial de Ionización
TDB8i 1.254,65 1.362,00 1.201,18 1.188,58 1.312,73 1.188,57 1.171,03 1.179,16 1.103,96
TDB10i 1.816,65 1.733,37 1.460,62 1.515,93 1.773,06 1.451,99 1.402,39 1.418,41 1.296,55
Polarizabilidad TDB6p 8,32 8,26 8,61 8,51 9,33 10,12 7,71 7,96 7,87
Tamaño y geometría
TDB6r 1,82 1,87 2,29 1,85 2,00 2,49 1,71 1,76 1,71
TDB6v 1.080,21 1.100,05 1.333,31 1.114,43 1.121,27 1.460,71 1.005,28 1.039,38 1.018,14
TDB5u 4,91 4,90 4,99 4,94 4,87 5,04 4,68 4,76 4,80
Flexibilidad topológica RotBtFrac 0,19 0,22 0,30 0,29 0,23 0,27 0,35 0,32 0,34
Valencia VP-6 2,15 1,04 2,57 2,72 1,82 1,95 3,15 6,92 3,36
Todos los valores de moléculas que pasan por Cx39 son más altos que los valores de las moléculas que no pueden hacerlo. Todos los valores de moléculas que pasan por Cx39 son más bajos que los valores de las moléculas que no pueden hacerlo.

71
5.2.6. Los HCs-Cx39 abiertos no permiten el ingreso del ion Ca2+
5.2.6.1. El pH alcalino induce aumento de la captación de colorante pero no de Ca2+
en células HeLa-Cx39
Para evaluar la captación de Ca2+, se indujo la apertura de los HCs-Cxs en presencia de Ca2+
extracelular, como ocurre bajo la exposición de pH alcalino (Schalper, KA et al. 2010). Así, se
expusieron las células a una solución salina con pH 8.5 y la captación de Etd+ y Ca2+ fueron
registrados simultáneamente. Como previamente se demostró (Schalper, KA et al. 2010), las
células HeLa-Cx43 mostraron un incremento significativo en la captación de Etd+ como
también de la señal de Ca2+ (Fig.14 A, B). El mismo tratamiento indujo captación de Etd+ en
células HeLa-Cx39 (9,3 ± 1,3 AU/min vs. 2,4 ± 0,4 AU/min, respectivamente p < 0,0001 n = 4)
(Fig. 14 A), pero la señal de Ca2+ no aumentó significativamente (Fig. 14B, inserto). Por otro
lado, las células HeLa-P expuestas a pH alcalino no mostraron cambios en la captación de
Etd+ ni en la señal de Ca2+ (Fig. 14 A, B).

72
Fig. 14. El pH alcalino abre los HCs-Cx39, los cuales son permeables al Etd+ pero no al Ca2+. Células HeLa-P, HeLa-Cx39 o HeLa-Cx43 fueron expuestas a solución salina extracelular con pH 7.4 o pH 8.5. A and B. Tasa de captación de Etd+ y Ca2+, respectivamente, fueron evaluados a tiempo real. B Inserto. Incremento en tiempo real de la señal de Ca2+ en células HeLa-P, HeLa-Cx39 y HeLa-Cx43, bañados con solución alcalina después del registro en condiciones control. ****p<0,0001 y ***p<0,001, respectivamente, n=3 experimentos y 20 células fueron registradas en cada experimento. UA: Unidades arbitrarias.

73
5.2.6.2. El estrés mecánico (EM) induce aumento de la captación de colorante pero
no de Ca2+ en células HeLa-Cx39
Para estudiar más a fondo la posible permeabilidad de los HCs-Cx39 al Ca2+, se indujo la
apertura de estos mediante la aplicación de EM, un estímulo que induce la apertura de HCs
constituido por otras Cxs y en otros tipo celulares (Stout, CE et al. 2002, Batra, N et al. 2012).
Se encontró que hay una relación directa entre la intensidad del estímulo de EM y se midió la
tasa de captación de Etd+ en células HeLa-Cx39 (Fig. 15A). Mientras que, la aplicación de 2
estímulos seriales, goteo de 1 ml desde 15 cm de altura cada uno, no incrementó
significativamente la tasa captación de colorante, la aplicación de 4 estímulos causó un
incremento similar en la tasa de captación de colorante, al inducido por 6 o por 8 estímulos.
(Fig. 15A). Estos incrementos en la tasa de captación fueron completamente suprimidos por
La3+ aplicado durante la medición en tiempo real (Fig. 15A), indicando que la captación del
trazador ocurre a través de HCs-Cx39. Por otro lado, la aplicación de 8 estímulos sobre
células HeLa-P no incrementó la tasa de captación de colorante (Fig. 15A), lo cual es
consistente con la ausencia de HCs-Cx39. Una vez que se calibró la condición que indujo la
mayor respuesta en la captación del trazador, se procedió a realizar los ensayos para evaluar
el posible flujo de Ca2+ en células HeLa-Cx39. A pesar que se procedió con EM con el máximo
de 8 estímulos sobre células HeLa-Cx39, la señal de Ca2+ intracelular no aumentó (Fig. 15B).
Sin embargo, la aplicación de ionóforo de Ca2+ A23187 (2,5 μM) rápidamente incrementó la
señal de Ca2+, que después decreció progresivamente (Fig. 15B)

74
Fig.15. Los HCs-Cx39 de células expuestas a EM son permeables a Etd+ pero no a Ca2+. A. tasa de captación de Etd+ de células HeLa-P y HeLa-Cx39 mecánicamente estimuladas con diferentes ciclos de aplicación de 1 ml de solución salina desde 15 cm de altura y bajo condiciones controles. Las células HeLa-Cx39 fueron mecánicamente estimuladas con 8 ml en presencia de 200 µM La3+ (8+La3+). Las células HeLa-P también fueron estimuladas con 8 ml de solución salina (****p<0,0001 n=5 con 30 células registradas en cada caso) B. Señal intracelular de Ca2+ de células HeLa-Cx39 bajo condiciones control (primeros 2,5 min), después de estímulo mecánico (8 ml desde 15 cm de altura) y después de la adición del ionóforo de Ca2+ A23187 (2,5 µM) como control de ingreso de Ca2+extracelular. EM: Estrés mecánico.

75
5.2.7. La captación de colorante en células HeLa-Cx39 es inhibida por algunos pero no
por otros bloqueadores de HCs-Cxs.
Varios bloqueadores de CUH-Cxs y HCs-Cxs y sus propiedades han sido descritas
anteriormente (Bodendiek, SB y Raman, G 2010, Verselis, VK y Srinivas, M 2013). A pesar de
que los HCs-Cx39 (Fig. 13 B, inserto) y HCs-Cx43 (Contreras, JE et al. 2003) son rápidamente
bloqueadas por La3+ en células expuestas a SLCD, la aplicación de octanol (310 μM), heptanol
(350 μM), β-Ga (50 μM), Ole (100 μM) o CBX (50 μM) no bloqueó los HCs-Cxs abiertos por
SLCD. Esto podría sugerir que estos bloqueadores no bloquean los HCs-Cx43 o de HCs-Cx39
abiertos. Sin embargo, la co-aplicación de ácido β-Ga (50 μM), Ole (100 μM) o CBX (50 μM)
con SLCD a células HeLa-Cx39 o células HeLa-Cx43-EGFP evitó significativamente el
incremento de la captación de colorante inducida por SLCD en ambos tipos celulares,
sugiriendo que los bloqueadores actúan cuando HCs-Cx están en estado cerrado. En
contraste, heptanol u octanol no previnieron el incremento de captación de colorante
inducido por SLCD en células HeLa-Cx39 (Fig. 16). Sin embargo, estos dos alcoholes
efectivamente previnieron la apertura inducida por SLCD en células HeLa-Cx43 (Fig. 16).
Como CBX (50 μM) es un bloqueador conocido de HCs-Cxs y HCs-Panx1, se evaluó la
posibilidad que los HCs-Cx39 presentaran sensibilidad farmacológica similar a la de los HCs-
Panx1. Con este fin, se probó si una baja concentración de CBX (5 μM), cual es un
bloqueador clásico de HCs-Panx1 (Bruzzone, R et al. 2005, Ma, W et al. 2009), afecta el
estado funcional de HCs-Cx39. La apertura de los HCs-Panx1 fue inducida por EM en células

76
HeLa-Panx1 usadas como control (Bao, L et al. 2004, Locovei, S et al. 2006, Taylor, KA et al.
2015) y 5 μM CBX drásticamente redujo la captación de colorante. En contraste la misma
concentración de CBX no afectó la captación de colorante inducida por EM en células HeLa-
Cx39 (Fig. 16).

77
Fig.16. Los HCs-Cx39 son inhibidos por algunos bloqueadores convencionales. El gráfico muestra los datos normalizados, con respecto a las condiciones basales, de la tasa de captación de Etd+ inducida por SLCD en células HeLa-Cx43 EGFP y HeLa Cx39. Heptanol (350
µM), octanol (310 µM), β-Ga(50 µM, -GA), Ole (100 µM) o CBX (5 o 50 µM) fueron aplicado junto SLCD (n=10, &&&&p<0,0001, &&&p<0,0005 and ****p<0,0001 comparado solo con células HeLa-Cx39 y HeLa-Cx43 EGFP, respectivamente, bajo el incremento de la captación de colorante inducido por SLCD vs. todos los bloqueadores de captación; ####p<0,0001 comparado solo entre células HeLa-Panx1).

78
5.2.8. El modelamiento comparativo de los HCs-Cx39 revela constricciones en el poro
que no se visualizan en los HCs-Cx43 y en los HCs-Cx26.
Para conocer mejor las diferencias estructurales y/o funcionales entre los HCs-Cx43 y los
HCs-Cx39, lo cual podría ayudar a explicar porque Cx39 no forma CUH funcionales en células
HeLa, se desarrolló un set de modelos moleculares para ambos HCs-Cxs y CUH basados en la
estructura cristalizada de los HCs-Cx26 como un molde (Maeda, S et al. 2009) (Fig. 17A).
Como se esperaba, e independiente de las variaciones de secuencia individual, todos los
modelos asemejan a la estructura general de un CUH-Cx26. Sin embargo, se pueden
encontrar algunas diferencias en la estructura del poro a lo largo del canal (Fig. 17, fila
superior). Para determinar si estas diferencias estructurales pueden influenciar en la función
del canal, el radio del poro de todos los HCs-Cxs y CUH-Cxs fue analizado. El radio del poro de
CUH (Fig. 17B izq.) y HC (Fig. 17B der.) formado por Cx26 muestra una estrecha constricción
en la región helicoidal N-terminal (NTH), que alcanza los 4.5 Å, mientras que la parte más
ancha se encuentra en la parte extracelular en la región para-helicoidal (PH), con un radio de
poro cercano a 14 Å. El radio del poro calculado para canales formados por Cx39 y Cx43
sigue una tendencia similar. Mostrando una parte angosta del poro en la región NTH,
alcanzando un radio de menos de 2Å. Ambos canales muestran la parte más ancha del poro
en el lado extracelular de la región PH, similar a los canales formados por Cx26, alcanzando
un radio cercano a 12Å. Sin embargo, el radio del poro de los CUH (Fig. 17B izq.) y HCs (Fig.
17B der.) formados por Cx39 muestran una notoria constricción, la cual está ausente en las

79
estructuras formadas por las otras Cxs modeladas: un radio de poro cercano a 4Å en el lado
intercelular de la región PH.

80
Fig.17. La mCx39 se asemeja a un canal de una Cx, pero muestra diferencias estructurales en la región PH en comparación con mCx43 y hCx26. A. Fila superior, representaciones estructurales de la estructura cristalizada de hCx26-HC utilizada como plantilla, y los modelos moleculares de mCx39 y mCx43. Cada HC muestra dos monómeros que están representados por cintas y codificados por colores para representar diferentes regiones de la proteína (NTH = magenta, TM1 = rojo, PH = cian, TM2 = naranja, TM3 = amarillo y TM4 = verde). Otros dos monómeros se representan en una superficie sólida gris, mientras que los otros dos en la parte delantera se omitieron por claridad. A. Fila inferior, representaciones estructurales del CUH-hCx26utilizado como plantilla, y los modelos moleculares de mCx39 y mCx43. Cada HC muestra tres monómeros representados por dibujos animados (cilindros = alfa hélices, flechas = hojas beta, tubos = bobinas y bucles) y codificados por colores para diferenciarse entre sí. B. Radio del poro a lo largo del eje z de los canales de Cxs. A la izquierda, el radio de poro a lo largo del eje z calculado para la estructura cristalina hCx26 (negro) de los CUH y el modelo molecular de mCx39 (rojo) y de Cx43 (azul). Derecha, radio de poro a lo largo del eje z calculado para la estructura cristalina de hCx26 (negro) de los modelos moleculares de los HCs-mCx39 (rojo) y HCs-Cx43 (azul). La región del NTH se representa en magenta mientras que las regiones del PH aparecen en cian.

81
5.2.9. La estructura del poro de los HC-Cx39 posee una distribución de cargas eléctricas
singular en comparación a los HCs-Cx26 y HCs-Cx43
Para profundizar las implicaciones de estas diferencias estructurales entre la Cx39 y Cx43,
considerando las variaciones en su secuencia, se sometió a evaluación bioinformática el
potencial electrostático de los modelos moleculares obtenidos. El potencial electrostático de
ambos HCs formados por Cx43 y Cx26 exhibe una tendencia general. Las porciones
intracelulares muestran una confluencia positiva, mientras que lados extracelulares tienden a
ser electronegativos, mostrando un patrón difuso en cual las cargas negativas están
mezcladas con regiones neutrales y, en menor extensión, con regiones positivas.
Interesantemente, los HCs-Cx39 escapan a esta tendencia, exhibiendo un lado intracelular
más neutral, mostrando un patrón difuso de cargas positivas, y una marcada confluencia
electronegativa al lado extracelular flanqueado por dos regiones neutrales localizadas a
ambos lados de esta acumulación de cargas negativas. Además este parche electronegativo
co-localiza con la constricción única encontrada en los HCs-Cx39 en la región intracelular (Fig.
18).

82
Fig.18. La Cx39 posee fuertes cargas electronegativas en una constricción particular al interior del poro de los CUH y HCs, en comparación a los poros de CUH y HCs formados por Cx26 y Cx43. Modelamiento de la superficie molecular a lo largo del poro de los canales basados en Cx coloreados por el potencial electrostático. Fila superior, el potencial electrostático “mapeado” sobre la superficie molecular de la estructura cristalina de los HCs-hCx26 utilizados como molde, y los modelos moleculares de mCx39 y mCx43. Fila inferior, el potencial electrostático mapeado sobre la superficie molecular de la estructura cristalina CUH-hCx26 utilizado como molde, y los modelos moleculares de la mCx39 y mCx43. Todas las representaciones estructurales representan 4 monómeros por HC mientras que los otros dos en el frente (en negro) se omitieron por claridad. La codificación del color corresponde al valor del potencial electrostático según el espectro mostrado a la derecha (en unidades kT/e).

83
6. Discusión
El trasplante de tumores utilizando líneas celulares como las B16, como modelo de estudio,
es uno de los métodos más simples y rápidos para formar grandes tumores y observar sus
consecuencias, en este caso, en músculo esquelético. Una de las grandes ventajas al utilizar
este modelo, es la baja respuesta inmunogénica de rechazo al tumor. Sin embargo, por su
eficacia, los ratones deben ser sacrificados a las pocas semanas por la agresiva invasión. Por
otro lado, una de las debilidades de este modelo es la velocidad de crecimiento y
reproducibilidad de los tumores. Además, la inoculación de varios miles de células y el daño
al tejido blanco, puede generar respuestas inflamatorias diversas (Dranoff, G 2011).
La técnica de inoculación i.v. de células B16F0, tiene la desventaja que para evaluar la
progresión del tumor, en este caso, las colonias que se forman en el pulmón, se tiene que
necesariamente sacrificar al animal, por lo que no se conoce, por ningún patrón observable
in vivo, si los ratones poseen o no el tumor y que tan reproducibles estuvieron las
inyecciones dentro de los grupos tratados. En este caso, al no evidenciarse variaciones
negativas en el peso de los ratones tratados con células de melanoma, tampoco se pudo
predecir cuales eran los que estaban comprometidos en formación de tumores. Aunque en
la literatura se ha evidenciado que hay bajas de peso significativas, con respecto al peso
inicial antes del tratamiento, en modelos de trasplante de tumor (Gil-Ad, I et al. 2006,
Dranoff, G 2011, Graulich, T et al. 2014), se debe considerar algunas variables: los tipos
celulares y la cantidad de células inoculadas para generar tumores como por ejemplo células
de cáncer de Lewis, CT26 de colón o B16 de melanoma; el lugar de colonización y crecimiento

84
tumoral, como subcutánea, peritoneal, pulmonar o intramuscular; y, finalmente, el tiempo
evaluado. Sin embargo, en la literatura se ha informado ambos casos, tanto la disminución de
más de 2 gramos en ratones de 21 días post-tratamiento subcutáneo con B16 y carcinoma de
Lewis (Graulich, T et al. 2014), como también las fluctuaciones de peso sin diferencias
significativas de ratones control comparados con ratones inoculados con células B16 i.v.
durante los primeros 20 días de tratamiento (Gil-Ad, I et al. 2006).
Los ratones inoculados con células B16F0 i.v., decaen en su estado de salud general
rápidamente. La inoculación i.v. de 5x106 células induce un rápido deterioro del estado
general entre los 15 a 17 días post tratamiento, al igual como se ha descrito anteriormente
en ratones inoculados con 2x106 células, en donde el deterioro abrupto del estado general se
retarda hasta la cuarta semana (Gil-Ad, I et al. 2006).
El estado inflamatorio de los ratones que poseían tumores en desarrollo, resultó
evidente con el análisis del perfil de citoquinas pro-inflamatorias que, como se ha descrito,
las concentraciones resultaron significativamente más altas que en condiciones controles
(Thejass, P y Kuttan, G 2007, Das, SK et al. 2011, Pratheeshkumar, P y Kuttan, G 2011, Zhang,
L et al. 2015), las cuales se mantuvieron altas durante todo el protocolo temporal,
verificándose el estado inflamatorio crónico, con excepción de INF-γ, el cual al día 14 decae
significativamente. Esto puede deberse a que la cantidad de INF˗γ registrado durante la
primera semana sea producto de una respuesta inflamatoria aguda del tejido huésped de las
células tumorales, en este caso el pulmón, pues se ha registrado que este órgano responde
con la producción de INF˗γ liberado por las células NK, como parte de una resistencia natural
del tejido pulmonar a la metástasis por células B16 (Takeda, K et al. 2011). Estas citoquinas

85
pro-inflamatorias están identificadas por ser pro-caquécticas (Argiles, JM y Lopez-Soriano, FJ
1999), lo que quedó demostrado con la medición del AST de las fibras tanto en ratones
Cx43fl/fl-Cx45fl/fl como en Cx43fl/flCx45fl/fl:Myo-Cre, la cual resultó ser de similar magnitud a lo
observado en otros modelos de atrofia inducida por células tumorales inoculadas (Cai, D et
al. 2004).
La atrofia muscular inducida por cáncer en ratones que no expresan Cx43/Cx45 indica
que este componente de atrofia no depende de la expresión de Cxs. Cabe señalar que, en
comparación a la atrofia dependiente de HCs-Cxs observada en ratones denervados (Cea, LA
et al. 2013), el componente de atrofia muscular inducido por este modelo de cáncer es de
menor magnitud y similar a la atrofia muscular residual observada en ratones
Cx43fl/flCx45fl/fl:Myo-Cre denervados, por lo que estas dos atrofias compartirían una vía en
común, siendo un componente de disminución del tamaño de la fibra independiente de la
expresión y actividad de HCs-Cxs. Este componente de atrofia en cáncer se puede atribuir a
la acción directa de las citoquinas proinflamatorias circulantes sobre la miofibra, las cuales
activan las vías de degradación vía ubitiquin-ligasa y autofagia, donde a su vez también,
inducen la inhibición de las vías pro-anabólicas, dependientes de la vía mTORC1 y mTORC2
(Bonaldo, P y Sandri, M 2013), induciendo así el desbalance neto hacia la degradación
proteica. La participación de los HCs-Cxs, en el segundo componente de la atrofia muscular,
sería inducida por un daño agudo, como la denervación, o bajo otro perfil de concentración
de moléculas pro˗inflamatorias circulantes, que podrían inducir una inflamación aguda y una
rápida perdida de función en otros sitios, como ocurre con la unión neuromuscular (UNM) en
condiciones de endotoxemia inducido por LCP (Liu, L et al. 2016). En este mismo sentido,

86
según datos de laboratorio, existiría un rol protector del componente de atrofia muscular
HCs-Cxs-dependiente, inducida por la inervación de la miofibra, en donde se ha observado
que la estimulación constante de fibras aisladas con un análogo de acetilcolina evitaría la
expresión de HCs-Cxs y la consecuente pérdida de integridad de permeabilidad de
membrana, evidenciando que la mantención del estímulo nervioso de la UNM evita la
expresión y actividad de HCs-Cxs. Acorde con esto último y asociado al cáncer, se ha
identificado en humanos que las patologías neuromusculares desarrolladas en cualquiera de
los segmentos del nervio motor, son causadas por infiltración de tumores, daño mecánico
local por compresión de tejidos y por repuesta autoinmune de los receptores de acetilcolina
(rACh) y otros componentes de la UNM en ciertos casos de timomas (Grisold, W et al. 2016).
Sin embargo, estos daños neuromusculares debido a tumores son escasos en comparación a
la incidencia de caquexia en pacientes con cáncer (Grisold, W et al. 2016), por lo que es poco
probable que exista un déficit de neurotransmisión en este modelo de cáncer, en donde el
estado inflamatorio inducido no alcanzaría a sobrepasar los mecanismos que aún operan en
el músculo esquelético y que evitarían el componente de atrofia dependiente de Cxs. Sin
embargo tanto la conducción nerviosa como la integridad de la UNM en cáncer quedarían
por ser descritas ya que no se han reportado datos sobre estos parámetros en modelos
animales de cáncer.
De igual forma, así como en la atrofia inducida por cáncer no estaría involucrada la
expresión de HCs-Cxs, la integridad de la membrana no se vio afectada, a diferencia de como
se ha observado en la evaluación de permeabilidad a Etd+ y la caída del PMR en fibras
aisladas de ratones que expresan HCs-Cxs en otras condiciones que inducen atrofia (Cea, LA

87
et al. 2016). No obstante, está descrito que TNF-α exógeno induce disminución de PMR de
forma concentración-dependiente en músculo esquelético de rata tanto in vivo como ex-vivo
(Tracey, KJ et al. 1986). Así es como se ha observado la disminución del PMR, utilizando
concentraciones letales de TNF-α (sobre 2500 pg/ml) in vivo (Tracey, KJ et al. 1986). Pero, en
comparación al perfil de citoquinas obtenido de los ratones tratados con células B16, los
ensayos descritos de dependencia del PMR al TNF-α exógeno en músculo, fueron realizados a
concentraciones 6 veces más altas que el TNF-α sérico registrado en los ratones inyectados
con células B16. Sin embargo, esas concentraciones séricas de esta citoquina, sí se podría
encontrar en un ratón con inflamación sistémica como en sepsis. Aun conociendo que el
PMR del músculo esquelético depende de la concentración de TNF-α, es necesario evaluar el
PMR del músculo en un modelo de liberación endógena de TNF-α como en sepsis.
Caso contrario al del cáncer, la atrofia muscular inducida por LCP si fue prevenida en
ratones Cx43fl/flCx45fl/fl:Myo-Cre. En este caso, se evidenció una prevención parcial de la
atrofia muscular debido a la ausencia de la expresión de HCs-Cxs cercano a la misma
magnitud que muestra la prevención de la atrofia muscular los ratones Cx43fl/flCx45fl/fl:Myo-
Cre sometidos a denervación (Cea, LA et al. 2013, Cisterna, BA et al. 2016). A diferencia de la
atrofia muscular inducida por cáncer, en sepsis se ha observado anteriormente una pérdida
similar de la integridad de la unión neuromuscular (UNM) en condiciones de LCP, que en
comparación a ratas denervadas, se desarrolla en los mismos tiempos, pero con una potencia
inferior a lo observado en denervación. Sin embargo, este efecto es suficiente para que a las
24h los potenciales de acción en la miofibra sean significativamente de menor amplitud y de
mayor duración, evidenciando la pérdida de estímulo nervioso sobre la miofibra (Liu, L et al.

88
2016). Además, se ha establecido que ratas sometidas a LCP, expresan receptores TLR-4 en
forma tiempo-dependiente en el cordón espinal, (con un máximo a las 24 h), inflamación
espinal (infiltración de microglia/macrófagos TLR-4 positivos), desmielinizacion del nervio
ciático y disfunción neuromuscular (observado en el aumento de expresión de otros subtipos
de rACH y pérdida de excitación eléctrica de la miofibra)(Xie, F et al. 2017). Así mismo, se ha
descrito que la disminución de la actividad de la UNM está asociado a la expresión y actividad
de los HCs-Cxs (Tesis Doctoral de Bruno Cisterna, 2016), por lo que la atrofia muscular
inducida por LCP en ratones estaría asociada a un daño tipo-denervación, que induce, al igual
que en el modelo de denervación, la expresión y actividad de HCs-Cxs y la atrofia
HCs˗Cxs˗dependiente. Por lo anterior, se podría establecer que la baja actividad de la UNM
en ratones inducida por sepsis, es un evento previo a la expresión y actividad de HCs-Cxs, lo
cual llevaría al segundo componente de atrofia. Este daño tipo-denervación observado en
sepsis ha sido retratado anteriormente en la literatura (Tsukagoshi, H et al. 1999, Nayci, A et
al. 2005, Cankayali, I et al. 2007), sin embargo la identidad de los efectores o el mecanismo
de señalización de la pérdida de la UNM en la miofibra en estas condiciones aún no se ha
establecido.
La prevención parcial, pero significativa, en los ratones KO-músculo específico para
Cxs43/45, nos muestra que la atrofia observada puede poseer dos componentes que se
potencian: 1) un primer componente podría ser independientes de expresión y función HCs-
Cxs, como por ejemplo por citoquinas pro-inflamatorias que señalicen directamente sobre
rutas catabólicas y anabólicas, como sería la atrofia observada en el modelo de cáncer;

89
aunque también es posible 2) que la Cx39 participe en la atrofia muscular de los ratones KO
para Cx43 y Cx45 pudiendo condicionar la resistencia a la prevención total.
Al igual como se ha observado en atrofia inducida por denervación (Cisterna, BA et al.
2016), se ha establecido que la [Ca2+]i aumenta en condiciones de LCP tanto en ratas como en
ratones (Benson, DW et al. 1989, Fischer, DR et al. 2001, Balboa, E et al. 2017). En el contexto
que la atrofia por sepsis inducida por LCP genera una atrofia muscular tipo-denervación, el
segundo componente de atrofia observado, que es evitado en ratones que no expresan
Cx43/Cx45 en miofibras, estaría dado por la acumulación de Ca2+ en el sarcolema, el cual
induciría la activación de calpaínas las cuales degradan miofibras, lo que posteriormente
aumentaría la vía de degradación proteica dependiente del sistema ubiquitin-proteosoma
(Smith, IJ et al. 2008) y la inhibición de la señalización vía Akt (Smith, IJ y Dodd, SL 2007), la
cual induce finalmente la disminución de las vías anabólicas dependientes de mTOR
(Egerman, MA y Glass, DJ 2014), la cual también es inhibida vía citoquinas proinflamatorias y
glucocorticoides que están presentes en el modelo de LCP (Bonaldo, P y Sandri, M 2013),
resultando finalmente en una ganancia neta hacia la degradación proteica en el balance
síntesis-degradación.
A pesar de que ratones Cx43fl/flCx45fl/fl:Myo-Cre evitan de forma parcial, pero
significativa, la atrofia muscular, debida al tratamiento con LCP, esta no se refleja en una
prevención a la pérdida de peso. Esto puede deberse a que, tanto los ratones
Cx43fl/flCx45fl/fl:Myo-Cre como los Cx43fl/flCx45fl/fl sometidos a LCP, poseen síntomas de
anorexia inducido por citoquinas pro-inflamatorias lo que podría inducir una disminución de
ingesta de comida debido al dolor e inflamación que provocaría finalmente en ambos ratones

90
la baja de peso (Bernstein, IL et al. 1991). Probablemente uno de los componentes de atrofia
muscular de los ratones con LCP sea por condición nutricional, la cual no sería evitada en
ratones Cx43fl/flCx45fl/fl:Myo-Cre.
Tanto la baja del PMR (Liu, SH et al. 1997) como el aumento de permeabilidad del
sarcolema a colorantes (Lin, MC et al. 1998) ya se habían observado anteriormente en
músculo de diafragma de ratas bajo estas condiciones. A pesar de la baja de peso corporal de
los ratones Cx43fl/flCx45fl/fl:Myo-Cre en condiciones de LCP, es evidente la ausencia del
incremento en la tasa de captación de Etd+ y la mantención del PMR en niveles normales, al
contrario de lo observado en las miofibras de ratones Cx43fl/flCx45fl/fl sometidos a la misma
condición; esto nos hace concluir que los fenómenos de permeabilidad descritos para el
modelo de LCP, dependerían de la actividad de HCs-Cxs, así como se ha observado en otros
modelos de atrofia muscular (Cea, LA et al. 2013, Cea, LA et al. 2016, Cea, LA et al. 2016, Cea,
LA et al. 2016, Cisterna, BA et al. 2016). Sin embargo, esta ausencia del aumento de la
permeabilidad de las miofibras se mantiene aún en presencia de la expresión de la Cx39. Por
lo que la actividad de esta Cx como HC podría estar regulada o ser insuficiente para inducir
cambios evidentes en la permeabilidad del sarcolema de las miofibras.
Como es ampliamente conocido, el PMR en fibras esqueléticas depende de las
conductancias de K+ y Cl- de la membrana en estado de reposo (Bretag, AH 1987). De esta
manera, la pérdida de PMR en condiciones de LCP, se ha descrito como una consecuencia de
la pérdida de conductancia de Cl-, directamente a través de la disminución de la actividad de
canales de Cl- (Liu, SH et al. 1997), del cual depende más del 80% del de la conductancia
iónica de membrana en reposo (Palade, PT y Barchi, RL 1977), por lo que el rol que tienen los

91
HCs-Cxs activos en la baja de PMR en ratones sometidos a LCP, como HCs transmembrana de
poca selectividad iónica que permiten el paso de los iones según su gradiente
electroquímico, podría estar sobre interpretada. Sin embargo, la disminución de la
conductancia de ion Cl-, dependiente de canales de cloro CLC-1 en músculo esquelético de
ratas, han mostrado ser dependiente del aumento de la [Ca2+]i y de la actividad de la proteína
kinasa Ca2+˗dependiente (PKC) (Liantonio, A et al. 2007). De esta manera, el aumento de
[Ca2+]i (proveniente de una liberación de Ca2+ mitocondrial al citoplasma, inducido por ácido
niflúmico) en las miofibras, provoca finalmente la activación de PKC que fosforila a CLC-1
disminuyendo su actividad, reduciendo posteriormente la conductancia de Cl-, que se
traduce en la baja del PMR. En este sentido, se ha descrito que el aumento de actividad de
los HCs-Cx43 y HCs-Cx45 permite el paso de Ca2+ desde el espacio extracelular, además se
induce el aumento de la actividad de los HCs por la acumulación de este ion en el espacio
intracelular (Schalper, KA et al. 2008), por lo que se podría generar un ingreso reverberante y
una acumulación neta de Ca2+ dentro de las miofibras. Como se ha demostrado en
condiciones de denervación, el Ca2+ se acumula en las miofibras de ratones que expresan
Cx43/45, pero no en miofibras de ratones KO˗músculo específicos de Cx43/45 (Cisterna, BA
et al. 2016), en los cuales tampoco se observa en absoluto una baja del PMR inducida por
denervación, lo que se explicaría por la incapacidad de acumular Ca2+ intracelular debido a la
nula expresión de HCs formados Cx43 y Cx45 en la miofibra. A su vez, también se ha
observado el aumento de un 25% de los niveles basales de la [Ca2+]i en fibras musculares
aisladas de ratones sometidos a LCP, comparados con fibras musculares aisladas de ratones

92
control (Balboa, E et al. 2017). Este aumento también podría contribuir a la baja de PMR
dependiente de Ca2+ en estas condiciones.
A pesar de que en músculo esquelético la pérdida de PMR seria Ca2+˗dependiente en
condiciones de denervación y sepsis inducida por LCP, no se ha descrito la dependencia de
PMR y la expresión/función de HCs-Cxs, con excepción de la débil influencia que tiene Cx43
sobre el PMR en células HeLa en condiciones normales (Contreras, JE et al. 2003), que se
podría sumar al fenómeno anteriormente descrito. En este caso, la reducción de PMR en
células HeLa expuestas a SLCD es probablemente mediada por la apertura de HCs-Cxs que
permiten el paso transmembrana de iones (como el Na+ o el K+). En esta misma línea, ha sido
descrito que HCs-Cx43, permite el paso de Na+ (Li, F et al. 2001) y K+ (Miro-Casas, E et al.
2009), y al igual que estos últimos, los HCs-Cx39 también son canales no-selectivos como se
describe más adelante. Es por ello que, los gradientes transmembrana de Na+ y K+ impulsan
el flujo de iones según su gradiente electroquímico en células HeLa-Cx39, reduciendo el PMR
a un nuevo valor en estado estable. Consistente con esta interpretación es que la exposición
a SLCD no afecta el PMR en células HeLa-P y que La3+, un bloqueador de HCs-Cxs, evite la
reducción del RMP inducido por SLCD en HeLa-Cx39, HeLa-Cx43 y HeLa-Cx45. Además, el
PMR en células HeLa transfectadas con Cx39, Cx43 y Cx45, inmersas en solución salina
control fue incrementado con La3+, lo cual podría ser explicado por la inhibición de actividad
basal de los HCs. Otros HCs-Cxs (Kondo, RP et al. 2000, Contreras, JE et al. 2003) son
inhibidas por cationes divalentes extracelulares como el Ca2+, y la ausencia de este ion en el
medio extracelular incrementa la probabilidad de apertura, lo cual induciría la apertura
también de HCs-Cx39 induciendo la baja del PMR.

93
En la mayoría de las células, la bomba Na+/K+ ATPasa contribuye a la mantención del
PMR, y su inhibición causa la despolarización de la membrana celular (Redmann, K y Walliser,
S 1981). De acuerdo con el efecto de ouabaina sobre la fracción de PMR dependiente de la
Na+/K+ ATPasa, células HeLa-P expuestas a ouabaina mostraron una reducción similar de PMR
como las registradas en células HeLa transfectadas. Según estas observaciones, la
disminución del PMR en las fibras musculares en condiciones inflamatorias también puede
depender de una alta actividad de HCs-Cxs por sí sola, como lo visto en células HeLa-Cxs
bañadas con SLCD (sin Ca2+ ni Mg2+) (Fig. 9). En resumen, la Cx39 sigue siendo expresada en
ratones Cx43fl/flCx45fl/fl:Myo-Cre, sometidos a LCP, sin embargo, la atrofia muscular
esquelética, el PMR y la captación de colorante es prevenida por la ausencia de Cx43/Cx45,
aún en presencia de la Cx39. En modelos de denervación de músculo en ratones
Cx43fl/flCx45fl/fl:Myo-Cre, se ha observado una protección significativa del aumento de
captación de Etd+ observado en fibras aisladas (Cisterna, BA et al. 2016) y del PMR, el cual no
disminuye en presencia de la expresión de Cx39. Esto puede implicar que la fibra muscular
posee mecanismos compensatorios que no han sido sobrepasados por la expresión de Cx39,
como por ejemplo la actividad de la bomba Na+/K+ ATPasa, los cuales mantienen el mismo
estado estable iónico que lo observado en condiciones normales. En este contexto, se ha
observado anteriormente que la bomba Na+/K+ ATPasa aumenta su actividad en un 46% en el
músculo esquelético de ratas sometidas a LCP, comparado con condiciones basales, sin verse
afectados los niveles del mensajero ni proteico de esta bomba (O'Brien, WJ et al. 1996). Esta
compensación, de aumento de la actividad de la bomba, puede ser en respuesta al nuevo
flujo iónico que se genera en las miofibras por el aumento de la permeabilidad del sarcolema

94
dependiente de los HCs-Cxs, lo cual lleva a un nuevo estado estable, que sería la sumatoria
del aumento de flujo iónico, con el aumento de la actividad de la bomba Na+/K+ ATPasa,
dando como resultado la observada baja neta del PMR. Por el contrario, la falta de expresión
y actividad de los HCs-Cx43 o HCs-Cx45 es suficiente para que la expresión de Cx39 no pueda
ejercer una influencia importante en el flujo iónico que afecte finalmente el PMR de las
miofibras de ratones sometidos a LCP.
Ante esto último, se puede hipotetizar de que los HCs-Cx39 podrían estar altamente
regulados, tener una permeabilidad restringida o conductancia menor, en comparación a
HCs-Cx43 o HCs-Cx45, expresados en el músculo esquelético y que los mecanismos celulares
de compensación no son sobrepasados. En esta misma línea, sólo hay referencia de un dato
que sugiere el funcionamiento de la Cx39 como HCs con captación positiva de azul de Evans,
en células HeLa (Cea, LA et al. 2013). Así también, en comparación a fibras musculares
obtenidas de ratones Cx43fl/flCx45fl/fl denervados, se observó que en ratones
Cx43fl/flCx45fl/fl:Myo-Cre denervados, el Ca2+ no ingresa a la fibra aun en presencia de la
expresión de Cx39 (Cisterna, BA et al. 2016). Por lo que se hizo necesaria la caracterización
de Cx39 y su capacidad de formar HCs funcionales en un sistema aislado en células HeLa.
La Cx39 de roedores es ortólogo de la Cx40.1 expresada en humanos, y se encuentra
sólo en músculo esquelético de ratón en etapas tempranas del desarrollo (von Maltzahn, J et
al. 2004) y la falta de expresión acelera la diferenciación y regeneración de las miofibras (von
Maltzahn, J et al. 2011). Sobre la Cx39 se desconocían las propiedades funcionales como CUH
o HCs, por lo que no podíamos esclarecer su posible participación en la atrofia muscular.

95
Se había establecido que células HeLa-Cx39 no presentan acoplamiento a colorantes
(von Maltzahn, J et al. 2004), lo cual se confirmó, en esta tesis, a través de una técnica mucho
más sensible como el acoplamiento eléctrico. Existe precedentes de otras Cxs que no forman
CUH funcionales como mCx29 (Altevogt, BM et al. 2002)y mCx33 (Chang, M et al. 1996) en
las cuales no se ha observado acoplamiento positivo tanto transfectadas en células de
mamífero como en ovocitos de xenopus. Sin embargo, y en comparación a lo sucedido en
células HeLa-Cx39, al realizarse ensayos de acoplamiento con Alexa488 en diafragma aislado
de neonatos de ratón, que no expresan otra CUH-Cxs diferente a Cx39, se pudo observar el
traspaso de colorante a fibras vecinas a diferencia de rodamina dextrano (10 kDa), que no
difunde a células vecinas (von Maltzahn, J et al. 2004), por lo que el acoplamiento con Alexa
488 podría ocurrir a través de canales de membrana de CUH-Cx39 funcionales con
propiedades de permeabilidad limitada. Una posibilidad ante este fenómeno es que uno o
más factores para la formación de CUHs-Cx39, presentes en las fibras musculares, se pierdan
en células HeLa. Además, y según el contexto en el cual se expresa Cx39 en el músculo
esquelético, esto es, junto a Cx43 y Cx45, queda por determinar la compatibilidad y posible
formación de canales heterotípicos entre canales homoméricos de Cx39 con otros canales
homoméricos formados por Cx43 y Cx45. Por otro lado, es posible que el acoplamiento a
colorante observado in situ por von Maltzahn y col (2004) sea un artefacto causado por
liberación de colorante via HCs-Cx39 y/o HCs-Panx1 seguido de captación vía hemicanales en
células contiguas como lo describió Cea y col. (2013).
La diminución en PMR mostrado en células HeLa-Cx39 inducido por SLCD, coincide
con un incremento en las corrientes macroscópicas, como se determinó por técnica de

96
fijación de voltaje en célula completa, balo la misma condición. El incremento de las
corrientes macroscópicas inducido por SLCD fue revertido por La3+ a valores basales. En todas
las condiciones a 0 mV, I = 0, indicando que los HCs-Cx39 no tienen selectividad iónica al igual
que los HCs-Cx43 (Kondo, RP et al. 2000, Contreras, JE et al. 2003). La presencia de eventos
unitarios de alrededor de 75 pS fue observada en registros de corrientes macroscópicas con
protocolos de rampas en SLCD a potenciales negativos, el cual fue confirmado con protocolos
de pasos rectangulares a +60 mV bajo las mismas condiciones, indicando que la conductancia
unitaria es de 75 ± 5 pS. Interesantemente, en comparación con los HCs-Cx43-EGFP, el
tiempo de transición de apertura y cierre de los HCs-Cx39 fueron más rápidos, pero los HCs-
Cx39 tienen el mismo tiempo de transición desde cerrado a abierto y vice-versa. Valores
similares de tiempo de transición han sido observado previamente para los HCs-Cx43
(Contreras, JE et al. 2003) y HCs-Cx56 (Ebihara, L et al. 1999).
El uso de diferentes colorantes como DAPI, Etd+, propidio y otros han sido reportados
antes para observar la actividad de permeabilidad a través de HCs-Cxs (Orellana, JA et al.
2011, Hansen, DB et al. 2014). Sin embargo, las propiedades de selectividad del poro de la Cx
no son fácilmente visualizadas con una lista corta de colorantes permeables. El incremento
en las corrientes de membrana en células HeLa-Cx39 en SLCD coincide con un notorio
incremento en la permeabilidad a diferentes colorantes en la misma solución, con excepción
de H-etd+4 (MW: 875) y BBG- (MW: 854), mientras que pasa muy poco IP+2 (MW: 668). La
captación de todos estos colorantes fue inhibida por La3+ en condiciones de SLCD. La
permeabilidad de los HeLa-Cx39 a moléculas varía entre moléculas grandes con alta carga
neta, como el colorante TOTO® (MW: 1300, +4), YOYO® (MW: 1200, +4) o AE˗4 (MW: 960), a

97
moléculas pequeñas con baja carga neta, los cuales no pasan a través de los HCs-Cx39, como
ACMA® (MW: 258, +1). Debido a que los colorantes permeables difieren en términos de su
naturaleza molecular (tabla 1), explicar por qué un colorante pasa a través de los Cx39˗HCs
debe considerar la naturaleza del poro de los HCs-Cx39 y las características moleculares del
colorante.
Medir la permeabilidad de los HCs-Cx39 con varios colorantes podría proveer
información útil sobre la naturaleza del poro y su afinidad a ciertas moléculas, acorde a la
carga neta, forma y tamaño. Sin embargo, estos criterios de selección parecen ser
insuficientes para determinar cuáles son las características básicas de las moléculas que
cruzan la membrana a través de los HCs-Cx39. Para explorar estas propiedades
fisicoquímicas que podrían explicar la permeabilidad de HCs-Cx39, el uso de una
aproximación de aprendizaje-mecanizado para clasificar permitió descubrir propiedades
específicas de los diferentes colorantes que favorecen o impiden el paso de estos, resultando
ser: electronegatividad, potencial de ionización, polarizabilidad y valencia. Otras
propiedades representadas fueron relacionadas con el tamaño, geometría y flexibilidad de
las moléculas. Como la electronegatividad es proporcional a la valencia, es de esperar que
los descriptores TDB6e, ATSC5s, GATS5s & VP-6 podrían mostrar tendencias similares (por
ejemplo parches electronegativos se extienden y se separan y átomos de baja valencia son
cercanos entre sí). Procion orange MX2R y azul de Evans tienen varios grupos SO3-distribuido
a través de su estructura, y pasan a través de los HCs-Cx39. BBG también tiene una pareja de
grupo SO3-, pero como se discute más adelante, los descriptores relacionados con el tamaño
lo clasifican como favorable para cruzar los HCs-Cx39. Potenciales de ionización y

98
polarizabilidad también muestran tendencias similares, principalmente debido al hecho que
la mayoría de los colorantes probados tienen estructuras resonantes que permiten la
deslocalización del electrón (por ejemplo la polarización se reparte por toda la molécula). En
el caso del IP2+, a pesar de que tiene estructuras resonantes similares a Etd+ y hexidio, este
posee dos cargas positivas cercanas una de otra. Esto es contrario a lo que pasa para el
DAPI2+, que tiene dos cargas positivas lejanas entre sí. Interesantemente, la flexibilidad
molecular tiende a relacionarse negativamente con la capacidad de difundir a través de los
HCs-Cx39: colorantes que tienen baja o nula tasa de captación (propidio, homodímero de
Etd+, BBG-) tienen altos valores para el descriptor RotBtFrac. Esta noción es reforzada debido
a que BBG- y H˗etd4+ presentan una estructura más abultada que el resto de las moléculas
(según los descriptores de tamaño y geometría), a pesar de que tienen pesos moleculares
similares y volúmenes de van der Waals al AE4˗. Además DAPI2+, AE4˗, y AM+, tienen una
estructura lineal general, mientras que Etd+ y hexidio, son relativamente pequeños, lo cual
podría favorecer la difusión a través de los HCs-Cx39. En el caso de procion orange MX2R, a
pesar de ser una molécula cargada como el AE4˗, este posee dos átomos de cloro que podrían
dificultar su difusión. El radio del poro también podría ser un factor de impedimento para
que estas moléculas abultadas puedan difundir a través de los HCs-Cx39. En contraste con
otros HCs-Cxs, los HCs˗Cx39 exhiben una constricción en la región PH que está ausente en los
HCs-Cx26 y HCs˗Cx43. Por lo tanto, es evidente que una estructura abultada, como BBG- y H-
etd+4, tendrá más dificultad en cruzar la barrera formada por la constricción de la región PH
de los HC˗Cx39. En cuanto al IP2+, aunque su estructura es similar al Etd+ y al hexidio, este

99
posee un grupo amonio cuaternario, el cual puede rotar libremente, posiblemente
impidiendo su difusión a través de los HCs-Cx39.
Se ha descrito anteriormente, que el Ca2+ aumenta en las miofibras en condiciones
inflamatorias como denervación, y que esta entrada de Ca2+ depende de los HCs-Cxs
(Cisterna, BA et al. 2016). Sin embargo, este ingreso de Ca2+ no se observa en ratones
denervados que no expresan Cx43/Cx45, específicamente en el músculo esquelético
(Cisterna, BA et al. 2016). Ante esto, hay dos posibilidades por las que la señal de Ca2+ puede
no haber sido detectada en esos ensayos: una entrada significativamente menor de Ca2+ a
través de los HCs-Cx39, que la fibra puede evacuar rápidamente del citoplasma (a diferencia
de la entrada constante y masiva de Ca2+ que se generaría al tener activos los HCs-43 y
HCs˗Cx45), o que los HCs-Cx39 no permitan el paso de Ca2+, por lo que fue necesario conocer
si Ca2+ pasa a través de los HCs-Cx39. Ambos estímulos, la alcalinización del medio
extracelular (pH: 8.5)(Schalper, KA et al. 2010) y el EM (Bao, L et al. 2004, D'Hondt, C et al.
2009) aplicados a células transfectadas con Cxs, indujeron el aumento de la tasa de captación
de Etd+, no se detectó señal de Ca2+ intracelular en células HeLa-Cx39, indicando que los HCs-
Cx39 son impermeables a este ion bajo estas condiciones. Interesantemente los HCs-Cx39
son los primeros HCs que observa que son impermeables a Ca2+, sugiriendo que su apertura
podría ser menos dañina para las células bajo condiciones patológicas como denervación del
músculo esquelético (Cea, LA et al. 2013, Cisterna, BA et al. 2016) y en distrofia muscular de
Duchenne (Cea, LA et al. 2016). Sin embargo aún es posible que los HCs-Cx39 permitan la
liberación de ATP u otras moléculas de señalización que participen en le atrofia muscular.

100
Anteriormente se han identificado bloqueadores de los HCs y CUH formados por Cxs,
y se han descrito sus propiedades (Herve, JC y Sarrouilhe, D 2005, Bodendiek, SB y Raman, G
2010, Verselis, VK y Srinivas, M 2013). Los experimentos de captación de colorante en
células HeLa-Cx39 sugieren que alcoholes de cadenas largas, derivados de ácido glicirritínico
y ácidos grasos no bloquean completamente los HCs-Cx39 o HCs-Cx43 después de la
exposición a SLCD y que su bloqueo es preventivo. Por otro lado, octanol y heptanol no
previenen el incremento de captación de colorante en HeLa-Cx39 inmersas en SLCD. La
acción de alcoholes de cadena larga como heptanol ha sido explicada por sus efectos sobre la
fluidez de la membrana celular (Bastiaanse, EM et al. 1993), mientras que una interacción
específica de alta afinidad con los HCs-Cx50 ha sido reportado para octanol (Eskandari, S et
al. 2002). Oleamida, una amida primaria de ácido graso, la que posee propiedades
anestésicas, no muestra efecto sobre la trasmisión de ondas de calcio estimuladas
mecánicamente (Guan, X et al. 1997, Boger, DL et al. 1998). Finalmente, los bloqueadores
clásicos relacionados estructuralmente, β-Ga y CBX, parcialmente y completamente
bloquean la actividad de los HCs-Cx39, respectivamente. Estos derivados parecen bloquear
diferentes de CUH-Cxs sin ser específicos para ningún subtipo de Cxs y el mecanismo de
acción propuesto podría involucrar la inserción de los derivados del ácido glicirritínico en la
membrana plasmática, por lo tanto uniendo a las subunidades de Cxs de los CUH e
induciendo un cambio conformacional que conduce al cerrado del canal (Davidson, JS et al.
1986).
El estudio de la relación estructura-función que podría explicar por qué las células
HeLa que expresan la Cx39 no forman CUH funcionales pero si forman HCs con modelos

101
moleculares, reveló características de interés interpretativo que en comparación poseen
otros HCs-Cxs o CUH-Cxs formados por Cx26 o Cx43. A pesar de la similar estructura general
exhibida por los modelos moleculares, algunas diferencias notables aparecen en un análisis
detallado. Como se notó, ambos HCs-Cx43 y HCs-Cx39 muestran zonas de constricciones
notorias en el NTH que, debido a restricciones estéricas, podrían evitar el paso de la mayoría
de las moléculas a través del canal, produciendo canales inactivos o constitutivamente
cerrados. Sin embargo, no es el caso para los HCs-Cx43 que forman ambos HCs y CUH
funcionales. Por lo tanto, se hipotetizó que esta constricción podría ser el resultado de la
aplicación de una restricción espacial obtenida desde la estructura cristalizada de la Cx26
durante el procedimiento del modelamiento comparativo. Según se describió anteriormente,
la estructura cristalizada de los HCs-Cx26, fue obtenida en estado abierto (Maeda, S et al.
2009). Sin embargo, dinámicas de simulación molecular realizadas anteriormente (Kwon, T
et al. 2011) usando la estructura cristalizada de los HC-Cx26 concluyeron que esta estructura
improbablemente represente a un canal abierto. Además, más tarde (Zonta, F et al. 2014) se
alcanzó la misma conclusión: la estructura cristalizada de canales formados por la Cx26
debería estar en formación cerrada. A pesar de esta controversia, evidencia experimental
disponible sugiere que la región NTH está relacionada a la apertura rápida dependiente de
voltaje del canal (Oh, S y Bargiello, TA 2015). Por lo tanto, se hipotetizó que cualquier
restricción presente en esta región podría ser afectada por la relocalización del NTH en
presencia de diferencias de potencial transmembrana. Esta noción es apoyada por los
trabajos previos que sugieren que el sensor de voltaje puede ser una parte integral del NTH
(Ek-Vitorin, JF y Burt, JM 2013, Araya-Secchi, R et al. 2014)

102
Por otro lado, los canales formados por la Cx39 exhiben una constricción singular en
el lado intracelular de la región PH que está ausente en el modelo formado por la Cx43 y por
la estructura molde de la Cx26. Similar a NTH, esta región también ha sido relacionada a
regulación voltaje dependiente de canales basados por Cxs, es decir, la regulación lenta (Oh,
S y Bargiello, TA 2015). Como se mencionó antes, células HeLa-Cx39 no forman canales de
unión en hendidura. Por lo tanto, se hipotetizó que la constricción presente en los canales
formados por la Cx39 en la región PH podrían responder a una diferencia de voltaje
transmembrana. Si este fuera el caso, aparte de la restricción estérica, la región PH podría
también presentar otras características que podrían explicar la pérdida de funcionalidad de
CUH. En este sentido, el potencial electrostático reveló, que en comparación a los HCs y CUH
formados por Cx43 y Cx26, los HCs y CUH formados por Cx39 poseen cargas fuertemente
negativas justamente en la constricción particular que posee en la región PH, lo cual revelaría
que no solo existiría una restricción espacial-estérica, sino que también eléctrica.
Considerando la importancia de la localización de las cargas para la selectividad iónica de los
canales formados por Cxs (Ek-Vitorin, JF y Burt, JM 2013, Escalona, Y et al. 2016), es de
esperar que cualquier alteración en este patrón pudiera impedir corrientes iónicas o censar
el voltaje, por lo tanto afectando la función del canal.
Interesantemente, la reciente liberación de la estructura cristalizada de canal
formado por Cx26 obtenido en presencia de Ca2+ extracelular, sugiere que a pesar de que
ambas regiones PH y NTH puedan responder a diferencias de voltaje a través de la
membrana celular, el bloqueo de los canales inducida por Ca2+ podría ser el resultado de una

103
obstrucción electrostática en lugar de un conjunto de cambios conformacionales (Bennett,
BC et al. 2016)

104
7. Conclusiones
Cx39 no forma canales de unión en hendidura en células HeLa-Cx39
Cx39 forma HCs funcionales en células HeLa y su conductancia unitaria es de 75 pS
Ca2+ no pasa a través de HCs-Cx39, por lo tanto los HC-Cx39 no serían una vía de
ingreso de Ca2+ en músculo sometido a condiciones que inducen atrofia muscular
dependiente de HCs-Cxs.
HC-Cx39 presenta permeabilidad a colorantes de diverso tamaño, lo cual indica que
podría estar activo como HCs en fibras musculares atróficas.
La atrofia muscular esquelética inducida por el modelo de cáncer con células de
melanoma no depende de la expresión ni actividad de HCs-Cxs.
LCP induce atrofia muscular esquelética y depende parcialmente de la expresión y
función de HCs-Cxs (Fig. 19).
Cx39 forma HCs funcionales en HeLa-Cx39 pero su participación en el músculo
atrófico queda por ser determinado.

105
Figura 19. La atrofia muscular inducida por sepsis depende de la expresión y función de HCs-Cxs. 1: Los HCs-Cxs 43,45 y 39 se expresan en el músculo atrófico de ratones sometidos a LCP, pero no en músculos atrofiados de ratones inyectados con células de melanoma B16F0 2: Los HCs-Cxs se abren y se mantienen abiertos en las fibras musculares por un mecanismo aún no determinado. 3: La permeabilidad del sarcolema aumenta, dependiente de la actividad de HCs-Cx. El Ca2+
no pasa a través de HCs-Cx39. 4: Como consecuencia del aumento de la permeabilidad, el PMR de las miofibras disminuye por el nuevo estado estable alcanzado por el flujo iónico a través de los HCs-Cxs.

106
8. Bibliografía
1 Altevogt, BM, Kleopa, KA, Postma, FR, Scherer, SS and Paul, DL (2002). "Connexin29 is uniquely distributed within myelinating glial cells of the central and peripheral nervous systems." J Neurosci 22(15): 6458-6470. 2 Amitani, M, Asakawa, A, Amitani, H and Inui, A (2013). "Control of food intake and muscle wasting in cachexia." Int J Biochem Cell Biol 45(10): 2179-2185. 3 Araya-Secchi, R, Lagos, CF, Abarca, F, Martinez, AD and Perez-Acle, T (2012). "Molecular Dynamics Simulations of Cx26-Wt and Deafness Related Mutants M34A, A40G and V37I." Biophys J 102(3): 106a-107a. 4 Araya-Secchi, R, Perez-Acle, T, Kang, SG, Huynh, T, Bernardin, A, Escalona, Y, Garate, JA, Martinez, AD, Garcia, IE, Sáez, JC and Zhou, R (2014). "Characterization of a novel water pocket inside the human Cx26 hemichannel structure." Biophys J 107(3): 599-612. 5 Argiles, JM, Busquets, S and Lopez-Soriano, FJ (2005). "The pivotal role of cytokines in muscle wasting during cancer." Int J Biochem Cell Biol 37(10): 2036-2046. 6 Argiles, JM and Lopez-Soriano, FJ (1999). "The role of cytokines in cancer cachexia." Med Res Rev 19(3): 223-248. 7 Balboa, E, Saavedra, F, Cea, LA, Vargas, AA, Ramírez, V, Escamilla, R, Sáez, JC and Regueira, T (2017). "Sepsis-Induced Channelopathy in Skeletal Muscles is Associated with Expression of Non-Selective Channels." Shock. 8 Baltgalvis, KA, Berger, FG, Pena, MM, Davis, JM, Muga, SJ and Carson, JA (2008). "Interleukin-6 and cachexia in ApcMin/+ mice." Am J Physiol Regul Integr Comp Physiol 294(2): R393-401. 9 Bao, L, Locovei, S and Dahl, G (2004). "Pannexin membrane channels are mechanosensitive conduits for ATP." FEBS Lett 572(1-3): 65-68. 10 Bao, L, Sachs, F and Dahl, G (2004). "Connexins are mechanosensitive." Am J Physiol Cell Physiol 287(5): C1389-1395. 11 Bao, X, Lee, SC, Reuss, L and Altenberg, GA (2007). "Change in permeant size selectivity by phosphorylation of connexin 43 gap-junctional hemichannels by PKC." Proc Natl Acad Sci U S A 104(12): 4919-4924. 12 Bastiaanse, EM, Jongsma, HJ, van der Laarse, A and Takens-Kwak, BR (1993). "Heptanol-induced decrease in cardiac gap junctional conductance is mediated by a decrease in the fluidity of membranous cholesterol-rich domains." J Membr Biol 136(2): 135-145.

107
13 Batra, N, Burra, S, Siller-Jackson, AJ, Gu, S, Xia, X, Weber, GF, DeSimone, D, Bonewald, LF, Lafer, EM, Sprague, E, Schwartz, MA and Jiang, JX (2012). "Mechanical stress-activated integrin alpha5beta1 induces opening of connexin 43 hemichannels." Proc Natl Acad Sci U S A 109(9): 3359-3364. 14 Beahm, DL and Hall, JE (2002). "Hemichannel and junctional properties of connexin 50." Biophys J 82(4): 2016-2031. 15 Bennett, BC, Purdy, MD, Baker, KA, Acharya, C, McIntire, WE, Stevens, RC, Zhang, Q, Harris, AL, Abagyan, R and Yeager, M (2016) An electrostatic mechanism for Ca(2+)-mediated regulation of gap junction channels. Nat Commun 7, 8770 DOI: 10.1038/ncomms9770 16 Bennett, MV, Contreras, JE, Bukauskas, FF and Sáez, JC (2003). "New roles for astrocytes: gap junction hemichannels have something to communicate." Trends Neurosci 26(11): 610-617. 17 Benson, DW, Hasselgren, PO, Hiyama, DT, James, JH, Li, S, Rigel, DF and Fischer, JE (1989). "Effect of sepsis on calcium uptake and content in skeletal muscle and regulation in vitro by calcium of total and myofibrillar protein breakdown in control and septic muscle: results from a preliminary study." Surgery 106(1): 87-93. 18 Bernstein, IL, Taylor, EM and Bentson, KL (1991). "TNF-induced anorexia and learned food aversions are attenuated by area postrema lesions." Am J Physiol 260(5 Pt 2): R906-910. 19 Bodendiek, SB and Raman, G (2010). "Connexin modulators and their potential targets under the magnifying glass." Curr Med Chem 17(34): 4191-4230. 20 Boger, DL, Patterson, JE, Guan, X, Cravatt, BF, Lerner, RA and Gilula, NB (1998). "Chemical requirements for inhibition of gap junction communication by the biologically active lipid oleamide." Proc Natl Acad Sci U S A 95(9): 4810-4815. 21 Bolton, EE, Wang, Y, Thiessen, PA and Bryant, SH (2008). Chapter 12 - PubChem: Integrated Platform of Small Molecules and Biological Activities. Annual Reports in Computational Chemistry. A. W. Ralphand C. S. David, Elsevier. Volume 4: 217-241. 22 Bonaldo, P and Sandri, M (2013). "Cellular and molecular mechanisms of muscle atrophy." Dis Model Mech 6(1): 25-39. 23 Bretag, AH (1987). "Muscle chloride channels." Physiol Rev 67(2): 618-724. 24 Bruzzone, R, Barbe, MT, Jakob, NJ and Monyer, H (2005). "Pharmacological properties of homomeric and heteromeric pannexin hemichannels expressed in Xenopus oocytes." J Neurochem 92(5): 1033-1043. 25 Cai, D, Frantz, JD, Tawa, NE, Jr., Melendez, PA, Oh, BC, Lidov, HG, Hasselgren, PO, Frontera, WR, Lee, J, Glass, DJ and Shoelson, SE (2004). "IKKbeta/NF-kappaB activation causes severe muscle wasting in mice." Cell 119(2): 285-298.

108
26 Callahan, LA and Supinski, GS (2009). "Sepsis-induced myopathy." Crit Care Med 37(10 Suppl): S354-367. 27 Cankayali, I, Dogan, YH, Solak, I, Demirag, K, Eris, O, Demirgoren, S and Moral, AR (2007). "Neuromuscular deterioration in the early stage of sepsis in rats." Crit Care 11(1): R1. 28 Cea, LA, Balboa, E, Puebla, C, Vargas, AA, Cisterna, BA, Escamilla, R, Regueira, T and Sáez, JC (2016). "Dexamethasone-induced muscular atrophy is mediated by functional expression of connexin-based hemichannels." Biochim Biophys Acta 1862(10): 1891-1899. 29 Cea, LA, Bevilacqua, JA, Arriagada, C, Cardenas, AM, Bigot, A, Mouly, V, Sáez, JC and Caviedes, P (2016). "The absence of dysferlin induces the expression of functional connexin-based hemichannels in human myotubes." BMC Cell Biol 17 Suppl 1: 15. 30 Cea, LA, Cisterna, BA, Puebla, C, Frank, M, Figueroa, XF, Cardozo, C, Willecke, K, Latorre, R and Sáez, JC (2013). "De novo expression of connexin hemichannels in denervated fast skeletal muscles leads to atrophy." Proc Natl Acad Sci U S A 110(40): 16229-16234. 31 Cea, LA, Puebla, C, Cisterna, BA, Escamilla, R, Vargas, AA, Frank, M, Martínez-Montero, P, Prior, C, Molano, J, Esteban-Rodriguez, I, Pascual, I, Gallano, P, Lorenzo, G, Pian, H, Barrio, LC, Willecke, K and Sáez, JC (2016). "Fast skeletal myofibers of mdx mouse, model of Duchenne muscular dystrophy, express connexin hemichannels that lead to apoptosis." Cell Mol Life Sci 73(13): 2583-2599. 32 Cea, LA, Riquelme, MA, Cisterna, BA, Puebla, C, Vega, JL, Rovegno, M and Sáez, JC (2012). "Connexin- and pannexin-based channels in normal skeletal muscles and their possible role in muscle atrophy." J Membr Biol 245(8): 423-436. 33 Cisterna, BA, Vargas, AA, Puebla, C and Sáez, JC (2016). "Connexin hemichannels explain the ionic imbalance and lead to atrophy in denervated skeletal muscles." Biochim Biophys Acta 1862(11): 2168-2176. 34 Clausen, T, Kjeldsen, K and Norgaard, A (1983). "Effects of denervation on sodium, potassium and [3H]ouabain binding in muscles of normal and potassium-depleted rats." J Physiol 345: 123-134. 35 Contreras, JE, Sáez, JC, Bukauskas, FF and Bennett, MV (2003). "Gating and regulation of connexin 43 (Cx43) hemichannels." Proc Natl Acad Sci U S A 100(20): 11388-11393. 36 Contreras, JE, Sanchez, HA, Eugenin, EA, Speidel, D, Theis, M, Willecke, K, Bukauskas, FF, Bennett, MV and Sáez, JC (2002). "Metabolic inhibition induces opening of unapposed connexin 43 gap junction hemichannels and reduces gap junctional communication in cortical astrocytes in culture." Proc Natl Acad Sci U S A 99(1): 495-500. 37 Chang, M, Werner, R and Dahl, G (1996). "A role for an inhibitory connexin in testis?" Dev Biol 175(1): 50-56.

109
38 D'Hondt, C, Iyyathurai, J, Vinken, M, Rogiers, V, Leybaert, L, Himpens, B and Bultynck, G (2013). "Regulation of connexin- and pannexin-based channels by post-translational modifications." Biol Cell 105(9): 373-398. 39 D'Hondt, C, Ponsaerts, R, De Smedt, H, Bultynck, G and Himpens, B (2009). "Pannexins, distant relatives of the connexin family with specific cellular functions?" Bioessays 31(9): 953-974. 40 Das, SK, Eder, S, Schauer, S, Diwoky, C, Temmel, H, Guertl, B, Gorkiewicz, G, Tamilarasan, KP, Kumari, P, Trauner, M, Zimmermann, R, Vesely, P, Haemmerle, G, Zechner, R and Hoefler, G (2011). "Adipose triglyceride lipase contributes to cancer-associated cachexia." Science 333(6039): 233-238. 41 Davidson, JS, Baumgarten, IM and Harley, EH (1986). "Reversible inhibition of intercellular junctional communication by glycyrrhetinic acid." Biochem Biophys Res Commun 134(1): 29-36. 42 DeVries, SH and Schwartz, EA (1992). "Hemi-gap-junction channels in solitary horizontal cells of the catfish retina." J Physiol 445: 201-230. 43 Dranoff, G (2011). "Experimental mouse tumour models: what can be learnt about human cancer immunology?" Nat Rev Immunol 12(1): 61-66. 44 Dranoff, G (2012). "Experimental mouse tumour models: what can be learnt about human cancer immunology?" Nat Rev Immunol 12(1): 61-66. 45 Ebihara, L, Xu, X, Oberti, C, Beyer, EC and Berthoud, VM (1999). "Co-expression of lens fiber connexins modifies hemi-gap-junctional channel behavior." Biophys J 76(1 Pt 1): 198-206. 46 Egerman, MA and Glass, DJ (2014). "Signaling pathways controlling skeletal muscle mass." Crit Rev Biochem Mol Biol 49(1): 59-68. 47 Ek-Vitorin, JF and Burt, JM (2013). "Structural basis for the selective permeability of channels made of communicating junction proteins." Biochim Biophys Acta 1828(1): 51-68. 48 Elias, LA, Wang, DD and Kriegstein, AR (2007). "Gap junction adhesion is necessary for radial migration in the neocortex." Nature 448(7156): 901-907. 49 Escalona, Y, Garate, JA, Araya-Secchi, R, Huynh, T, Zhou, R and Perez-Acle, T (2016). "Exploring the Membrane Potential of Simple Dual-Membrane Systems as Models for Gap-Junction Channels." Biophys J 110(12): 2678-2688. 50 Eskandari, S, Zampighi, GA, Leung, DW, Wright, EM and Loo, DD (2002). "Inhibition of gap junction hemichannels by chloride channel blockers." J Membr Biol 185(2): 93-102. 51 Eswar, N, Webb, B, Marti-Renom, MA, Madhusudhan, MS, Eramian, D, Shen, MY, Pieper, U and Sali, A (2006). "Comparative protein structure modeling using Modeller." Curr Protoc Bioinformatics Chapter 5: Unit 5 6.

110
52 Figueroa, VA, Retamal, MA, Cea, LA, Salas, JD, Vargas, AA, Verdugo, CA, Jara, O, Martínez, AD and Sáez, JC (2014). "Extracellular gentamicin reduces the activity of connexin hemichannels and interferes with purinergic Ca(2+) signaling in HeLa cells." Front Cell Neurosci 8: 265. 53 Figueroa, XF, Lillo, MA, Gaete, PS, Riquelme, MA and Sáez, JC (2013). "Diffusion of nitric oxide across cell membranes of the vascular wall requires specific connexin-based channels." Neuropharmacology 75: 471-478. 54 Fischer, DR, Sun, X, Williams, AB, Gang, G, Pritts, TA, James, JH, Molloy, M, Fischer, JE, Paul, RJ and Hasselgren, PO (2001). "Dantrolene reduces serum TNFalpha and corticosterone levels and muscle calcium, calpain gene expression, and protein breakdown in septic rats." Shock 15(3): 200-207. 55 Gil-Ad, I, Shtaif, B, Levkovitz, Y, Nordenberg, J, Taler, M, Korov, I and Weizman, A (2006). "Phenothiazines induce apoptosis in a B16 mouse melanoma cell line and attenuate in vivo melanoma tumor growth." Oncol Rep 15(1): 107-112. 56 Gómez-Hernández, JM, de Miguel, M, Larrosa, B, Gonzalez, D and Barrio, LC (2003). "Molecular basis of calcium regulation in connexin-32 hemichannels." Proc Natl Acad Sci U S A 100(26): 16030-16035. 57 Gosemann, JH, van Griensven, M, Barkhausen, T, Kobbe, P, Thobe, BM, Haasper, C, Pape, HC, Krettek, C, Hildebrand, F and Frink, M (2010). "TLR4 influences the humoral and cellular immune response during polymicrobial sepsis." Injury 41(10): 1060-1067. 58 Graulich, T, Das, SK, Wessels, L, Kummer, W, Hoefler, G and Muhlfeld, C (2014). "Effects of Lewis lung carcinoma and B16 melanoma on the innervation of the mouse trachea." Auton Neurosci 183: 106-110. 59 Grisold, W, Grisold, A and Loscher, WN (2016). "Neuromuscular complications in cancer." J Neurol Sci 367: 184-202. 60 Guan, X, Cravatt, BF, Ehring, GR, Hall, JE, Boger, DL, Lerner, RA and Gilula, NB (1997). "The sleep-inducing lipid oleamide deconvolutes gap junction communication and calcium wave transmission in glial cells." J Cell Biol 139(7): 1785-1792. 61 Halgren, TA (1996). "Merck molecular force field. I. Basis, form, scope, parameterization, and performance of MMFF94." Journal of Computational Chemistry 17(5-6): 490-519. 62 Hansen, DB, Braunstein, TH, Nielsen, MS and MacAulay, N (2014). "Distinct permeation profiles of the connexin 30 and 43 hemichannels." FEBS Lett 588(8): 1446-1457. 63 Hanwell, MD, Curtis, DE, Lonie, DC, Vandermeersch, T, Zurek, E and Hutchison, GR (2012). "Avogadro: an advanced semantic chemical editor, visualization, and analysis platform." J Cheminform 4(1): 17.

111
64 Harcha, PA, Vargas, A, Yi, C, Koulakoff, AA, Giaume, C and Sáez, JC (2015). "Hemichannels Are Required for Amyloid beta-Peptide-Induced Degranulation and Are Activated in Brain Mast Cells of APPswe/PS1dE9 Mice." J Neurosci 35(25): 9526-9538. 65 Herve, JC and Sarrouilhe, D (2005). "Connexin-made channels as pharmacological targets." Curr Pharm Des 11(15): 1941-1958. 66 Holecek, M (2012). "Muscle wasting in animal models of severe illness." Int J Exp Pathol 93(3): 157-171. 67 Humphrey, W, Dalke, A and Schulten, K (1996). "VMD: visual molecular dynamics." J Mol Graph 14(1): 33-38, 27-38. 68 Iacobas, DA, Scemes, E and Spray, DC (2004). "Gene expression alterations in connexin null mice extend beyond the gap junction." Neurochem Int 45(2-3): 243-250. 69 Jiang, JX and Gu, S (2005). "Gap junction- and hemichannel-independent actions of connexins." Biochim Biophys Acta 1711(2): 208-214. 70 Joassard, OR, Durieux, AC and Freyssenet, DG (2013). "beta2-Adrenergic agonists and the treatment of skeletal muscle wasting disorders." Int J Biochem Cell Biol 45(10): 2309-2321. 71 Jordan, K, Solan, JL, Dominguez, M, Sia, M, Hand, A, Lampe, PD and Laird, DW (1999). "Trafficking, assembly, and function of a connexin43-green fluorescent protein chimera in live mammalian cells." Mol Biol Cell 10(6): 2033-2050. 72 Kalantar-Zadeh, K, Rhee, C, Sim, JJ, Stenvinkel, P, Anker, SD and Kovesdy, CP (2013). "Why cachexia kills: examining the causality of poor outcomes in wasting conditions." J Cachexia Sarcopenia Muscle 4(2): 89-94. 73 Kaynar, AM, Yende, S, Zhu, L, Frederick, DR, Chambers, R, Burton, CL, Carter, M, Stolz, DB, Agostini, B, Gregory, AD, Nagarajan, S, Shapiro, SD and Angus, DC (2014). "Effects of intra-abdominal sepsis on atherosclerosis in mice." Crit Care 18(5): 469. 74 Kondo, RP, Wang, SY, John, SA, Weiss, JN and Goldhaber, JI (2000). "Metabolic inhibition activates a non-selective current through connexin hemichannels in isolated ventricular myocytes." J Mol Cell Cardiol 32(10): 1859-1872. 75 Kwon, T, Harris, AL, Rossi, A and Bargiello, TA (2011). "Molecular dynamics simulations of the Cx26 hemichannel: evaluation of structural models with Brownian dynamics." J Gen Physiol 138(5): 475-493. 76 Laurent, B, Chavent, M, Cragnolini, T, Dahl, AC, Pasquali, S, Derreumaux, P, Sansom, MS and Baaden, M (2015). "Epock: rapid analysis of protein pocket dynamics." Bioinformatics 31(9): 1478-1480. 77 Lerner, M and Carlson, H (2006). "APBS plugin for PyMOL." Ann Arbor: University of Michigan.

112
78 Li, F, Sugishita, K, Su, Z, Ueda, I and Barry, WH (2001). "Activation of connexin-43 hemichannels can elevate [Ca(2+)]i and [Na(+)]i in rabbit ventricular myocytes during metabolic inhibition." J Mol Cell Cardiol 33(12): 2145-2155. 79 Liantonio, A, Giannuzzi, V, Picollo, A, Babini, E, Pusch, M and Conte Camerino, D (2007). "Niflumic acid inhibits chloride conductance of rat skeletal muscle by directly inhibiting the CLC-1 channel and by increasing intracellular calcium." Br J Pharmacol 150(2): 235-247. 80 Lin, MC, Ebihara, S, El Dwairi, Q, Hussain, SN, Yang, L, Gottfried, SB, Comtois, A and Petrof, BJ (1998). "Diaphragm sarcolemmal injury is induced by sepsis and alleviated by nitric oxide synthase inhibition." Am J Respir Crit Care Med 158(5 Pt 1): 1656-1663. 81 Liu, L, Xie, F, Wei, K, Hao, XC, Li, P, Cao, J and Min, S (2016). "Sepsis induced denervation-like changes at the neuromuscular junction." J Surg Res 200(2): 523-532. 82 Liu, SH, Lai, JL, Lin, RH, Lin, MJ and Lin-Shiau, SY (1997). "Involvement of nitric oxide in the in vivo effects of lipopolysaccharide on the contractile and electrical properties of mouse diaphragm." Naunyn Schmiedebergs Arch Pharmacol 356(4): 500-504. 83 Locovei, S, Bao, L and Dahl, G (2006). "Pannexin 1 in erythrocytes: function without a gap." Proc Natl Acad Sci U S A 103(20): 7655-7659. 84 Lopez, W, Liu, Y, Harris, AL and Contreras, JE (2014). "Divalent regulation and intersubunit interactions of human connexin26 (Cx26) hemichannels." Channels (Austin) 8(1): 1-4. 85 Ma, W, Hui, H, Pelegrin, P and Surprenant, A (2009). "Pharmacological characterization of pannexin-1 currents expressed in mammalian cells." J Pharmacol Exp Ther 328(2): 409-418. 86 Maeda, S, Nakagawa, S, Suga, M, Yamashita, E, Oshima, A, Fujiyoshi, Y and Tsukihara, T (2009). "Structure of the connexin 26 gap junction channel at 3.5 A resolution." Nature 458(7238): 597-602. 87 Miro-Casas, E, Ruiz-Meana, M, Agullo, E, Stahlhofen, S, Rodriguez-Sinovas, A, Cabestrero, A, Jorge, I, Torre, I, Vazquez, J, Boengler, K, Schulz, R, Heusch, G and Garcia-Dorado, D (2009). "Connexin43 in cardiomyocyte mitochondria contributes to mitochondrial potassium uptake." Cardiovasc Res 83(4): 747-756. 88 Nayci, A, Atis, S, Comelekoglu, U, Ozge, A, Ogenler, O, Coskun, B and Zorludemir, S (2005). "Sepsis induces early phrenic nerve neuropathy in rats." Eur Respir J 26(4): 686-692. 89 O'Brien, WJ, Lingrel, JB, Fischer, JE and Hasselgren, PO (1996). "Sepsis increases skeletal muscle sodium, potassium-adenosinetriphosphatase activity without affecting messenger RNA or protein levels." J Am Coll Surg 183(5): 471-479. 90 Oh, S and Bargiello, TA (2015). "Voltage regulation of connexin channel conductance." Yonsei Med J 56(1): 1-15.

113
91 Oh, S, Rivkin, S, Tang, Q, Verselis, VK and Bargiello, TA (2004). "Determinants of gating polarity of a connexin 32 hemichannel." Biophys J 87(2): 912-928. 92 Olk, S, Turchinovich, A, Grzendowski, M, Stuhler, K, Meyer, HE, Zoidl, G and Dermietzel, R (2010). "Proteomic analysis of astroglial connexin43 silencing uncovers a cytoskeletal platform involved in process formation and migration." Glia 58(4): 494-505. 93 Orellana, JA, Diaz, E, Schalper, KA, Vargas, AA, Bennett, MV and Sáez, JC (2011). "Cation permeation through connexin 43 hemichannels is cooperative, competitive and saturable with parameters depending on the permeant species." Biochem Biophys Res Commun 409(4): 603-609. 94 Orellana, JA and Stehberg, J (2014). "Hemichannels: new roles in astroglial function." Front Physiol 5: 193. 95 Palade, PT and Barchi, RL (1977). "Characteristics of the chloride conductance in muscle fibers of the rat diaphragm." J Gen Physiol 69(3): 325-342. 96 Pfahnl, A and Dahl, G (1999). "Gating of cx46 gap junction hemichannels by calcium and voltage." Pflugers Arch 437(3): 345-353. 97 Postic, G, Ghouzam, Y and Gelly, JC (2015). "An empirical energy function for structural assessment of protein transmembrane domains." Biochimie 115: 155-161. 98 Pratheeshkumar, P and Kuttan, G (2011). "Vernonia cinerea Less. inhibits tumor cell invasion and pulmonary metastasis in C57BL/6 mice." Integr Cancer Ther 10(2): 178-191. 99 Puebla, C, Cisterna, BA, Salas, DP, Delgado-Lopez, F, Lampe, PD and Sáez, JC (2016). "Linoleic acid permeabilizes gastric epithelial cells by increasing connexin 43 levels in the cell membrane via a GPR40- and Akt-dependent mechanism." Biochim Biophys Acta. 100 Redmann, K and Walliser, S (1981). "Different changes in transmembrane potential of cultured cells after ouabain-inhibited active Na+/K+-transport." Arch Geschwulstforsch 51(1): 96-102. 101 Retamal, MA, Cortes, CJ, Reuss, L, Bennett, MV and Sáez, JC (2006). "S-nitrosylation and permeation through connexin 43 hemichannels in astrocytes: induction by oxidant stress and reversal by reducing agents." Proc Natl Acad Sci U S A 103(12): 4475-4480. 102 Retamal, MA, Schalper, KA, Shoji, KF, Bennett, MV and Sáez, JC (2007). "Opening of connexin 43 hemichannels is increased by lowering intracellular redox potential." Proc Natl Acad Sci U S A 104(20): 8322-8327. 103 Rodriguez-Sinovas, A, Ruiz-Meana, M, Denuc, A and Garcia-Dorado, D (2017). "Mitochondrial Cx43, an important component of cardiac preconditioning." Biochim Biophys Acta. 104 Sáez, JC, Berthoud, VM, Branes, MC, Martinez, AD and Beyer, EC (2003). "Plasma membrane channels formed by connexins: their regulation and functions." Physiol Rev 83(4): 1359-1400.

114
105 Sáez, JC, Retamal, MA, Basilio, D, Bukauskas, FF and Bennett, MV (2005). "Connexin-based gap junction hemichannels: gating mechanisms." Biochim Biophys Acta 1711(2): 215-224. 106 Sánchez, HA, Mese, G, Srinivas, M, White, TW and Verselis, VK (2010). "Differentially altered Ca2+ regulation and Ca2+ permeability in Cx26 hemichannels formed by the A40V and G45E mutations that cause keratitis ichthyosis deafness syndrome." J Gen Physiol 136(1): 47-62. 107 Sánchez, HA, Orellana, JA, Verselis, VK and Sáez, JC (2009). "Metabolic inhibition increases activity of connexin-32 hemichannels permeable to Ca2+ in transfected HeLa cells." Am J Physiol Cell Physiol 297(3): C665-678. 108 Scemes, E (2008). "Modulation of astrocyte P2Y1 receptors by the carboxyl terminal domain of the gap junction protein Cx43." Glia 56(2): 145-153. 109 Schakman, O, Kalista, S, Barbe, C, Loumaye, A and Thissen, JP (2013). "Glucocorticoid-induced skeletal muscle atrophy." Int J Biochem Cell Biol 45(10): 2163-2172. 110 Schalper, KA, Palacios-Prado, N, Orellana, JA and Sáez, JC (2008). "Currently used methods for identification and characterization of hemichannels." Cell Commun Adhes 15(1): 207-218. 111 Schalper, KA, Palacios-Prado, N, Retamal, MA, Shoji, KF, Martínez, AD and Sáez, JC (2008). "Connexin hemichannel composition determines the FGF-1-induced membrane permeability and free [Ca2+]i responses." Mol Biol Cell 19(8): 3501-3513. 112 Schalper, KA, Sánchez, HA, Lee, SC, Altenberg, GA, Nathanson, MH and Sáez, JC (2010). "Connexin 43 hemichannels mediate the Ca2+ influx induced by extracellular alkalinization." Am J Physiol Cell Physiol 299(6): C1504-1515. 113 Siempos, II, Lam, HC, Ding, Y, Choi, ME, Choi, AM and Ryter, SW (2014). "Cecal ligation and puncture-induced sepsis as a model to study autophagy in mice." J Vis Exp(84): e51066. 114 Sishi, BJ and Engelbrecht, AM (2011). "Tumor necrosis factor alpha (TNF-alpha) inactivates the PI3-kinase/PKB pathway and induces atrophy and apoptosis in L6 myotubes." Cytokine 54(2): 173-184. 115 Smith, IJ and Dodd, SL (2007). "Calpain activation causes a proteasome-dependent increase in protein degradation and inhibits the Akt signalling pathway in rat diaphragm muscle." Exp Physiol 92(3): 561-573. 116 Smith, IJ, Lecker, SH and Hasselgren, PO (2008). "Calpain activity and muscle wasting in sepsis." Am J Physiol Endocrinol Metab 295(4): E762-771. 117 Soares, AR, Martins-Marques, T, Ribeiro-Rodrigues, T, Ferreira, JV, Catarino, S, Pinho, MJ, Zuzarte, M, Isabel Anjo, S, Manadas, B, J, PGS, Pereira, P and Girao, H (2015). "Gap junctional protein Cx43 is involved in the communication between extracellular vesicles and mammalian cells." Sci Rep 5: 13243.

115
118 Stout, CE, Costantin, JL, Naus, CC and Charles, AC (2002). "Intercellular calcium signaling in astrocytes via ATP release through connexin hemichannels." J Biol Chem 277(12): 10482-10488. 119 Takeda, K, Nakayama, M, Sakaki, M, Hayakawa, Y, Imawari, M, Ogasawara, K, Okumura, K and Smyth, MJ (2011). "IFN-gamma production by lung NK cells is critical for the natural resistance to pulmonary metastasis of B16 melanoma in mice." J Leukoc Biol 90(4): 777-785. 120 Taylor, KA, Wright, JR and Mahaut-Smith, MP (2015). "Regulation of Pannexin-1 channel activity." Biochem Soc Trans 43(3): 502-507. 121 Tennila, A, Salmi, T, Pettila, V, Roine, RO, Varpula, T and Takkunen, O (2000). "Early signs of critical illness polyneuropathy in ICU patients with systemic inflammatory response syndrome or sepsis." Intensive Care Med 26(9): 1360-1363. 122 Thejass, P and Kuttan, G (2007). "Modulation of cell-mediated immune response in B16F-10 melanoma-induced metastatic tumor-bearing C57BL/6 mice by sulforaphane." Immunopharmacol Immunotoxicol 29(2): 173-186. 123 Timmons, JJ, Cohessy, S and Wong, ET (2016). "Injection of Syngeneic Murine Melanoma Cells to Determine Their Metastatic Potential in the Lungs." J Vis Exp(111). 124 Tracey, KJ, Lowry, SF, Beutler, B, Cerami, A, Albert, JD and Shires, GT (1986). "Cachectin/tumor necrosis factor mediates changes of skeletal muscle plasma membrane potential." J Exp Med 164(4): 1368-1373. 125 Trexler, EB, Bennett, MV, Bargiello, TA and Verselis, VK (1996). "Voltage gating and permeation in a gap junction hemichannel." Proc Natl Acad Sci U S A 93(12): 5836-5841. 126 Tsukagoshi, H, Morita, T, Takahashi, K, Kunimoto, F and Goto, F (1999). "Cecal ligation and puncture peritonitis model shows decreased nicotinic acetylcholine receptor numbers in rat muscle: immunopathologic mechanisms?" Anesthesiology 91(2): 448-460. 127 Valiunas, V (2002). "Biophysical properties of connexin-45 gap junction hemichannels studied in vertebrate cells." J Gen Physiol 119(2): 147-164. 128 Valiunas, V and Weingart, R (2000). "Electrical properties of gap junction hemichannels identified in transfected HeLa cells." Pflugers Arch 440(3): 366-379. 129 Van Rijen, HV, Wilders, R, Van Ginneken, AC and Jongsma, HJ (1998). "Quantitative analysis of dual whole-cell voltage-clamp determination of gap junctional conductance." Pflugers Arch 436(1): 141-151. 130 Vary, TC (1998). "Regulation of skeletal muscle protein turnover during sepsis." Curr Opin Clin Nutr Metab Care 1(2): 217-224.

116
131 Verselis, VK and Srinivas, M (2013). "Connexin channel modulators and their mechanisms of action." Neuropharmacology 75: 517-524. 132 Visser, LH (2006). "Critical illness polyneuropathy and myopathy: clinical features, risk factors and prognosis." Eur J Neurol 13(11): 1203-1212. 133 von Haehling, S and Anker, SD (2010). "Cachexia as a major underestimated and unmet medical need: facts and numbers." J Cachexia Sarcopenia Muscle 1(1): 1-5. 134 von Maltzahn, J, Euwens, C, Willecke, K and Sohl, G (2004). "The novel mouse connexin39 gene is expressed in developing striated muscle fibers." J Cell Sci 117(Pt 22): 5381-5392. 135 von Maltzahn, J, Wulf, V, Matern, G and Willecke, K (2011). "Connexin39 deficient mice display accelerated myogenesis and regeneration of skeletal muscle." Exp Cell Res 317(8): 1169-1178. 136 Xie, F, Min, S, Chen, J, Yang, J and Wang, X (2017). "Ulinastatin Inhibited Sepsis-induced Spinal Inflammation to Alleviate Peripheral Neuromuscular Dysfunction in an Experimental Rat Model of Neuromyopathy." J Neurochem. 137 Yap, CW (2011). "PaDEL-descriptor: an open source software to calculate molecular descriptors and fingerprints." J Comput Chem 32(7): 1466-1474. 138 Zhang, L, Tao, L, Shi, T, Zhang, F, Sheng, X, Cao, Y, Zheng, S, Wang, A, Qian, W, Jiang, L and Lu, Y (2015). "Paeonol inhibits B16F10 melanoma metastasis in vitro and in vivo via disrupting proinflammatory cytokines-mediated NF-kappaB and STAT3 pathways." IUBMB Life 67(10): 778-788. 139 Zonta, F, Mammano, F, Torsello, M, Fortunati, N, Orian, L and Polimeno, A (2014). "Role of gamma carboxylated Glu47 in connexin 26 hemichannel regulation by extracellular Ca(2)(+): insight from a local quantum chemistry study." Biochem Biophys Res Commun 445(1): 10-15.

117
9. Anexo I. Certificado de Bioética

118
9.1. Anexo II. Portada de publicación relacionado con esta Tesis
Top Related