
XVII SIMPOSIO DE ZOOLOGÍA - cezug.org.mxcezug.org.mx/czug/Documentos_files/memorias XVII...
58
-
Upload
trinhquynh -
Category
Documents
-
view
223 -
download
0
Transcript of XVII SIMPOSIO DE ZOOLOGÍA - cezug.org.mxcezug.org.mx/czug/Documentos_files/memorias XVII...


XVII SIMPOSIO DE ZOOLOGÍA DIRECTORIO
Universidad de Guadalajara RECTOR GENERAL
Mtro. Izcóatl Tonatiuh Bravo Padilla VICERRECTOR
Dr. Miguel Ángel Navarro Navarro SECRETARIO GENERAL
Mtro. José Alfredo Peña Ramos
Centro Universitario de Ciencias Biológicas y Agropecuarias RECTOR
Dr. Salvador Mena Munguía SECRETARIO ACADÉMICO Mtro. Salvador González Luna
SECRETARIO ADMINISTRATIVO Mtro. José Rizo Ayala
División de Ciencias Biológicas y Ambientales
DIRECTOR Dr. Carlos Beas Zárate
SECRETARIO Dr. Guillermo Barba Calvillo
Departamento de Botánica y Zoología
JEFE DE DEPARTAMENTO Dr. Ramón Rodríguez Macias
Centro de Estudios en Zoología
JEFE DE CENTRO Dr. Sergio Guerrero Vázquez
Cuerpo Académico de Zoología UdeG-CA-51
Responsable Dr. José Luis Navarrete Heredia
COMITÉ ORGANIZADOR DEL XVII SIMPOSIO DE ZOOLOGÍA
Georgina Adriana Quiroz Rocha
Miguel Vásquez Bolaños Margarito Mora Núñez
José Luis Navarrete Heredia Sergio Guerrero Vázquez
Silvia S. Zalapa Hernández Agustín Camacho Rodríguez
Gustavo Moya Raygoza Ana Laura González Hernández
Martha Elizabeth Anguiano Santana Pablo Antonio Martínez Rodríguez

COMITÉ EDITORIAL DE MEMORIAS XVII SIMPOSIO DE ZOOLOGÍA
Georgina Adriana Quiroz Rocha
Miguel Vásquez Bolaños

XVII SIMPOSIO DE ZOOLOGÍA INSTITUCIONES PARTICIPANTES
SCRIPPS INSTITUTION OF OCEANOGRAPHY-US SAN DIEGO
UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO Instituto de Biología
Colección Nacional de Arácnidos (CNAN)
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR Departamento de Biología Marina
Programa de Investigación para la Conservación de la Fauna Arrecifal
UNIVERSIDAD AUTÓNOMA DE NUEVO LEÓN
Facultad de Ciencias Biológicas Laboratorio de Ornitología
UNIVERSIDAD JUÁREZ DEL ESTADO DE DURANGO Facultad de Ciencias Forestales
INSTITUTO POLITÉCNICO NACIONAL CIIDIR - IPN Unidad Durango
INSTITUTO DE ECOLOGÍA, A. C.
INSTITUTO NACIONAL DE LA PESCA Centro Regional de Investigaciones Pesqueras- Manzanillo
INSTITUTO DE CIENCIAS DEL MAR Y LIMNOLOGÍA Unidad Mazatlán
EL COLEGIO DE LA FORNTERA SUR
Unidad San Cristóbal de las Casas, Chiapas
INSTITUTO TECNOLÓGICO DE BAHÍA DE BANDERAS

UNIVERSIDAD DE GUADALAJARA
Coordinación de Bibliotecas
Centro Universitario de Ciencias Económico Administrativas Departamento de Estudios Regionales/ DER-INESER
Centro Universitario de la Costa Sur Departamento de Estudios Sustentables para el Desarrollo de Zonas Costeras
Licenciatura en Biología Marina
Ingeniería en Recursos Naturales y Agropecuarios
Instituto Manantlán de Ecología y Conservación de la Biodiversidad
Centro Universitario de Ciencias Biológicas y Agropecuarias
División de Ciencias Veterinarias Departamento de Salud Pública
División de Ciencias Biológicas y Ambientales
Instituto de Limnología
Departamento de Ecología Laboratorio de Ecosistemas Marinos y Acuicultura (LEMA)
Departamento de Ciencias Ambientales
Departamento de Biología Celular y Molecular
Departamento de Botánica y Zoología
Centro de Estudios en Zoología (CZUG) Entomología
Vertebrados

PROGRAMA XVII SIMPOSIO DE ZOOLOGÍA 19-23 de octubre de 2015
Homenaje al Dr. Miguel Ángel Morón Ríos
Lunes 19 de octubre. Auditorio de usos múltiples CUCBA. 9:00-9:15 Inscripciones Pag. 9:15-9:45 Inauguración XVII SIMPOSIO DE ZOOLOGÍA 9:45-10:00
Alimentación del bobo café, Sula leucogaster (Aves: Suliformes) de una colonia de anidación en la isla Peña blanca, Colima. Luis Abraham Aguilar Nuño, Salvador Hernández Vazquez, Humberto Almanza Rodríguez, Ana María Almaguer Hernández, José Angel Hinojosa Larios, Bernabé Aguilar Palomino, José Manuel Bojórquez Castro y Miriam Daniela Rodríguez Torres.
29
10:00-10:15
Abundancia y alimentación del cuervo común Corvus corax clarionensis (Aves: Corvidae) en la Isla Clarión del Archipiélago Revillagigedo, México. Humberto Almanza Rodríguez, Luis Abraham Aguilar Nuño, Salvador Hernández Vazquez, Diego Adolfo González Zamora y Luis Felipe Angulo Castellanos.
31
10:15-10:30
Especies de langostino (Palaemonidae: Macrobrachium) en el Arroyo Seco, Cihuatlán, Jalisco. Rodolfo Castro Barbosa, Arnulfo H. Díaz, Carlos Alberto Amezcua Gómez y Jesús Eduardo Sainz Urzua.
19
10:30-10:45
Distribución y abundancia del langostino crecedor Macrobrachium americanum (Invertebrata: Decapoda) en el Río Marabasco, Jalisco-Colima. Jesús Eduardo Sainz Urzúa, Arnulfo H. Díaz, Carlos Alberto Amezcua Gómez y Rodolfo Castro Barbosa.
21
10:45-11:00
Receso
11:00-12:00
Conferencia Magistral. "Experiencias de nuestro laboratorio con diferentes herramientas para abordar los misterios de las tortugas marinas: genética, modelaje de deriva oceánica, telemetría, embriología". F. Alberto Abreu Grobois. Instituto de Ciencias del Mar y Limnología, Unidad Mazatlán.
12:00-12:15
Receso
12:15-12:30
Isópodos marinos (Crustacea: Isopoda) de Bahía de Chamela, Jalisco: Listado faunístico y nuevos registros. Diego Alfredo Gutiérrez Anzaldo, Manuel Ayón Parente, Eduardo Ríos Jara y Cristian Moisés Galván Villa.
22
12:30-12:45
Situación actual de las poblaciones de Octopus hubbsorum (Molusca: Octopoda) y algunas alternativas de manejo del recurso en la Costa de Jalisco. Carlos Alberto Amezcua Gómez, Rodolfo Castro Barbosa y Jesús Eduardo Sainz Urzua.
23
12:45-13:00
Estado del conocimiento de la Manta birostris (Batoidea; Mobulidae): Una revisión. Miguel de Jesús Gómez García, Daniel Enrique Godínez Siordia y Carlos Alberto Amezcua Gómez.
24
13:00-13:15
Foto identificación de la manta gigante Manta birostris (Vertebrata: Elasmobranchii) en Bahía de Banderas, Jalisco-Nayarit. Ramiro I. Gallardo-Hernández, Adrián Maldonado-Gasca, Javier Tovar-Ávila y Joshua Stewart.
25

13:15-13:30
Poliaminas Intestinales en tilapia (Oreochromis niloticus) y bagre (Ictalurus punctatus). Ramón Guillermo Ortiz-García, Anne Santerre, María del Rosario Huizar-López, Martha Cecilia Téllez-Bañuelos, Josefina Casas-Solís, Patricia Castro-Félix y Eduardo Juárez-Carrillo.
27
13:30-13:45
Condición poblacional y parasitosis aproximada del Tiro Goodea atripinnis (Goodeidae: Actinopterigy) en la Laguna de Cajititlán, Jalisco, durante la alta mortandad de peces (2014-2015). Agustín Camacho Rodríguez, Eduardo Juárez Carrillo, Martha Alicia Lara González y Manuel Guzmán Arroyo.
26
13:45-14:00
Receso
14:00-15:00
Foro “Políticas públicas en el medio ambiente ¿qué pretende el gobierno federal?”. Participantes: Arturo Curiel Ballesteros, Sergio Graf Montero, Enrique Jardel, Miguel Enrique Magaña Virgen, Óscar Francisco Reina Bustos.
Martes 20 de octubre. Sala de tesis, División de Ciencias Biológicas y Ambientales, CUCBA.
10:00-10:15
Inscripciones
10:15-10:30
Composición del ensamble de aves (Chordata: Aves) en cuatro sitios del Área de Protección de Flora y Fauna Maderas del Carmen, Coahuila, México. Nadia Martínez Gallegos, Juan Antonio García Salas y Sergio Guerrero Vázquez.
28
10:30-10:45
Comparación de dos métodos bioacústicos para el análisis de la riqueza de murciélagos insectívoros (Mammalia: Chiroptera) en un bosque templado de Jalisco, México. Carolina Orendain García, Silvia Socorro Zalapa Hernández y Sergio Guerrero Vázquez.
32
10:45-11:00
Biología poblacional de Artibeus hirsutus (Chiroptera: Phyllostomidae), en Isla Grande Atoyac, Jalisco. Rogelio García Sandoval, Silvia Socorro Zalapa Hernández y Sergio Guerrero Vázquez.
33
11:00-11:15
Estructura genética de Artibeus lituratus (Chiroptera, Phyllostomidae), en Jalisco. Andres Eduardo Gonzalez Yepez, Patricia Castro Félix, Silvia S. Zalapa Hernández y Felipe J. López Chávez.
34
11:15-11:30
Evaluación de gremios tróficos de murciélagos (Mammalia: Chiroptera) como bioindicadores potenciales de genotóxicos en el ambiente. Jorge Elías De la Torre Miguel, Silvia Socorro Zalapa Hernández, María Luisa Ramos Ibarra y Sergio Guerrero Vázquez.
35
11:30-11:45
Descripción del esqueleto postcraneal de los murciélagos (Mammalia:Chiroptera) Tadarida brasiliensis y Macrotus waterhousii. Jorge Manuel Vargas Aldaco y Silvia Socorro Zalapa Hernández.
36
12:45-12:00
Receso
12:00-12:15
¡Estamos extintos, Mexicanos!, Aspectos de Paleontología y Educación en México. Roberto Ramírez Espitia.
37

12:15-12:30
Colección Paleontológica del Centro de Estudios en Zoología de la Universidad de Guadalajara. Margarito Mora-Núñez y Diego Balderas Noriega.
37
12:30-12:45
Uso de técnicas de escalada y espeleología en la exploración de campo para monitoreo y muestreo en actividades de investigación zoológica y botánica. Rodrigo Andrade Villegas y Héctor Leonel Ayala-Téllez.
38
12:45-13:00
Receso
13:00-14:00
Conferencia Magistral. Biología Subterránea: un mundo por descubrir. Héctor Leonel Ayala Téllez. CUCBA, U. de G.
17
Miércoles 21 de octubre. Auditorio de usos múltiples CUCBA. 9:00-9:15
Diversidad de corales blandos (Cnidaria: Octocorallia: Alcyonacea) de Bahía de Chamela. Victoria Belén Muñoz Jiménez, Cristian Moisés Galván Villa, Eduardo Ríos Jara y Carlos Armando Sánchez Ortiz.
19
9:15-9:30
El género Vaejovis C. L. Koch, 1836 (Scorpiones: Vaejovidae) en Jalisco. Gerardo Adalberto Contreras Félix y Oscar Francke.
39
9:30-9:45
Selección de las presas por parte del depredador generalista Geocoris punctipes (Hemiptera: Lygaeidea): análisis de movilidad, estadío y coloración. Walter Ernesto Printzen Cruz y Ricardo Ramírez Romero.
40
9:45-10:00
Descripción de la estructura de las redes mutualistas planta-abeja (Hymenoptera: Apoidea: Anthophila) en dos sitios con diferente recurrencia de incendios en Bosque Escuela de la Sierra La Primavera. Christian Fernando Barajas-Daniel, Alejandro Muñoz-Urías, Osiris Yanid Serrano-Pineda, Hugo Eduardo Fierros-López y Claudia Aurora Uribe-Mú.
40
10:00-10:15
Hormigas (Hymenoptera: Formicidae) del Jardín Botánico Vallarta, Cabo Corrientes, Jalisco. Anahí Cisneros Caballero y Miguel Vásquez Bolaños.
42
10:15-10:30
Las sociedades de hormigas (Hymenoptera: Formicidae) como modelo en el estudio de sistemas complejos. Luigui Michel Gallardo-Becerra y Christian Fernando Barajas-Daniel.
43
10:30-10:45
Uso médico de la grana cochinilla en la época colonial mexicana. Fernando Rafael Villaseñor Ulloa.
44
10:45-11:00
Receso
11:00-12:00
Homenaje Dr. Miguel Ángel Morón Ríos
12:00-13:00
Conferencia Magistral. Un panorama sobre el estudio de los escarabajos melolóntidos y cetónidos en América Latina. Miguel Ángel Morón Ríos. Instituto de Ecología A. C., Xalapa, Veracruz, México.
3
13:00-13:15
Receso
13:15- Diversidad de melolóntidos nocturnos (Insecta: Coleoptera) en un 45

13:30 bosque tropical caducifolio de la Sierra del Águila, Ameca, Jalisco, México. José Ernesto Colima-Lara, Luis Eugenio Rivera-Cervantes, Adriana Elena Castro-Ramírez y Edith García-Real.
13:30-14:00
30 años conociendo la biodiversidad de los escarabajos (Coleoptera: Scarabaeoidea), de los sistemas montañosos del sur oeste de Jalisco, México. Luis Eugenio Rivera-Cervantes y Edith García-Real
46
14:00-14:15
Insectos descortezadores de los géneros Ips y Pseudips en Pinus cooperi B. y P. durangensis Mtz. en Pueblo Nuevo, Durango. Yolanda Ontiveros Moreno, Rebeca Álvarez Zagoya y José Javier Corral Rivas.
46
Jueves 22 y viernes 23 de octubre. Aula Posgrado CUCBA. 9:00-14:00 Taller “Escarabajos melolóntidos con importancia agrícola”.
Imparte Dr. Miguel Ángel Morón Ríos. INECOL, Xalapa, Veracruz.

RESUMEN CURRICULAR
Dr. Miguel Ángel Morón Ríos
Posición actual: Investigador Titular "D", Red de Biodiversidad y Sistemática, Instituto de Ecología, A. C. (Sector CONACYT) Xalapa, Veracruz, México.
Campo de Investigación: Taxonomía, Sistemática, Ecología, Zoogeografía, Evolución e importancia agropecuaria y forestal de los adultos y larvas de insectos Coleópteros Lamelicornios americanos, con énfasis en la Zona de Transición Mexicana.
Estudios: Licenciatura en Biología (1975), Maestría en Ciencias (Biología) (1977) y Doctorado en Ciencias (Biología) (1985). Facultad de Ciencias, UNAM, México, D.F.
Experiencia en investigación: Instituto de Biología, UNAM (1971-1976); UAM-Xochimilco (1975-1981); Instituto de Ecología, A. C. (1981 a la fecha). Responsable de 16 proyectos apoyados por el CONACYT (1983-2005) y CONABIO (1993-2003). 31 estancias de investigación en instituciones de Inglaterra, Francia, Alemania, España, Holanda, Canadá, EUA, Colombia, Nicaragua y Costa Rica. Profesor-Investigador Titular "C" Invitado en período sabático, Departamento de Investigación en Ciencias Agrícolas (1996), Centro de Agroecología (2014) Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla, Puebla, México.
Publicaciones: 256 artículos de investigación en revistas con arbitraje de México, EUA, Brasil, Canadá, Costa Rica, Colombia, Venezuela, Emiratos Árabes, España, Gran Bretaña, Italia, India, Japón, Nueva Zelandia; 12 libros, 66 capítulos de libro; y 67 artículos de divulgación científica, ensayos, notas, artículos de síntesis y trabajos en extenso en memorias de reuniones; 4 catálogos y 104 resúmenes en memorias de congresos.
Impacto de las publicaciones: 3,161 citas entre 1984 y 2013.
Participación en reuniones científicas: 95 congresos, simposia y mesas redondas en México, Alemania, España, Portugal, EUA, Venezuela, Colombia, Bolivia, Brasil, Ecuador, Chile, Costa Rica y Guatemala.
Cargos desempeñados: Director del Área Ecología y Biosistemática de Animales, INECOL (1989-91); Jefe de la División de Ecología y Biosistemática de Animales, INECOL (1992-93); Jefe del Departamento de Biosistemática de Insectos, INECOL (1992-1995); Jefe del Proyecto Biosistemática, INECOL (1987-1991); Encargado del Insectario, DPAA, DCBS, UAM-X (1977-1981).
Experiencia en docencia: 58 cursos de licenciatura, 20 cursos de posgrado y 7 de especialización, en Facultad de Ciencias, UNAM; UAM-Xochimilco (1974-1989); Instituto de Ecología (1993-2010); CATIE, Costa Rica (1989 y 1994); Univ. Nal. Colombia, Medellín (1994 y 2007); Univ. Michoacana San Nicolás de Hidalgo (1995); CORPOICA, Bogotá, Colombia (1995); EMBRAPA-Trigo, Passo Fundo, Brasil (1999);

Benemérita Universidad Autónoma de Puebla (1996-2002, 2013-2014); CITRO-Universidad Veracruzana (2009-2011); Universidad Nacional de Río Cuarto, Córdoba, Argentina (2006).
Tesis dirigidas: 28 de licenciatura en Biología, 15 de Maestría en Ciencias (Biología) y seis de Doctorado en Ciencias (Biología), todas sobre aspectos entomológicos.
Distinciones Académicas: Candidato a Investigador Nacional SNI (1985-1988);
Premio "Weizmann 1987" área Ciencias Naturales, Fundación Weizmann y AIC, México, por la tesis doctoral titulada "Revisión del género Phyllophaga Harris en México (Insecta: Coleoptera, Melolonthidae)"; Investigador Nacional nivel II, SNI (1988-1991); Miembro Regular de la Academia Mexicana de Ciencias, A.C. México (desde 1989); Investigador Nacional nivel III, SNI (1991-2017); Premio de Investigación Científica 1992, área de Ciencias Naturales. AIC, México; Cátedra Patrimonial de Excelencia Nivel II, CONACYT (1997-1999).
Otros logros académicos: Autor o coautor de 12 géneros y 250 especies
americanas de coleópteros lamelicornios nuevos para la ciencia, sobre todo de la fauna mexicana. Asesor científico y gráfico de la Exposición Itinerante "Escarabajos, 200 Millones de Años de Evolución" que recorrió México (1983- 1992) y España (1993-1996).
Cargos Honoríficos: Presidente, Vicepresidente y Secretario de la Sociedad Mexicana de Entomología, A.C. (1984-1990); Editor de la revista científica Folia Entomológica Mexicana (1981-1992); Miembro de los Consejos Editoriales de 5 revistas mexicanas de Zoología (1984-2008). Curador General de la Colección Entomológica del Instituto de Ecología, A.C. (IEXA) (2002-2012).

1
RESUMEN CURRICULAR
Dr. Miguel Ángel Morón Ríos
Posición actual: Investigador Titular "D", Red de Biodiversidad y Sistemática, Instituto de Ecología, A. C. (Sector CONACYT) Xalapa, Veracruz, México.
Campo de Investigación: Taxonomía, Sistemática, Ecología, Zoogeografía, Evolución e importancia agropecuaria y forestal de los adultos y larvas de insectos Coleópteros Lamelicornios americanos, con énfasis en la Zona de Transición Mexicana.
Estudios: Licenciatura en Biología (1975), Maestría en Ciencias (Biología) (1977) y Doctorado en Ciencias (Biología) (1985). Facultad de Ciencias, UNAM, México, D.F.
Experiencia en investigación: Instituto de Biología, UNAM (1971-1976); UAM-Xochimilco (1975-1981); Instituto de Ecología, A. C. (1981 a la fecha). Responsable de 16 proyectos apoyados por el CONACYT (1983-2005) y CONABIO (1993-2003). 31 estancias de investigación en instituciones de Inglaterra, Francia, Alemania, España, Holanda, Canadá, EUA, Colombia, Nicaragua y Costa Rica. Profesor-Investigador Titular "C" Invitado en período sabático, Departamento de Investigación en Ciencias Agrícolas (1996), Centro de Agroecología (2014) Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla, Puebla, México.
Publicaciones: 256 artículos de investigación en revistas con arbitraje de México, EUA, Brasil, Canadá, Costa Rica, Colombia, Venezuela, Emiratos Árabes, España, Gran Bretaña, Italia, India, Japón, Nueva Zelandia; 12 libros, 66 capítulos de libro; y 67 artículos de divulgación científica, ensayos, notas, artículos de síntesis y trabajos en extenso en memorias de reuniones; 4 catálogos y 104 resúmenes en memorias de congresos.
Impacto de las publicaciones: 3,161 citas entre 1984 y 2013.
Participación en reuniones científicas: 95 congresos, simposia y mesas redondas en México, Alemania, España, Portugal, EUA, Venezuela, Colombia, Bolivia, Brasil, Ecuador, Chile, Costa Rica y Guatemala.
Cargos desempeñados: Director del Área Ecología y Biosistemática de Animales, INECOL (1989-91); Jefe de la División de Ecología y Biosistemática de Animales, INECOL (1992-93); Jefe del Departamento de Biosistemática de Insectos, INECOL (1992-1995); Jefe del Proyecto Biosistemática, INECOL (1987-1991); Encargado del Insectario, DPAA, DCBS, UAM-X (1977-1981).
Experiencia en docencia: 58 cursos de licenciatura, 20 cursos de posgrado y 7 de especialización, en Facultad de Ciencias, UNAM; UAM-Xochimilco (1974-1989); Instituto de Ecología (1993-2010); CATIE, Costa Rica (1989 y 1994); Univ. Nal. Colombia, Medellín (1994 y 2007); Univ. Michoacana San Nicolás de Hidalgo (1995); CORPOICA, Bogotá, Colombia (1995); EMBRAPA-Trigo, Passo Fundo, Brasil (1999); Benemérita Universidad Autónoma de Puebla (1996-2002, 2013-2014); CITRO-

2
Universidad Veracruzana (2009-2011); Universidad Nacional de Río Cuarto, Córdoba, Argentina (2006).
Tesis dirigidas: 28 de licenciatura en Biología, 15 de Maestría en Ciencias (Biología) y seis de Doctorado en Ciencias (Biología), todas sobre aspectos entomológicos.
Distinciones Académicas: Candidato a Investigador Nacional SNI (1985-1988);
Premio "Weizmann 1987" área Ciencias Naturales, Fundación Weizmann y AIC, México, por la tesis doctoral titulada "Revisión del género Phyllophaga Harris en México (Insecta: Coleoptera, Melolonthidae)"; Investigador Nacional nivel II, SNI (1988-1991); Miembro Regular de la Academia Mexicana de Ciencias, A.C. México (desde 1989); Investigador Nacional nivel III, SNI (1991-2017); Premio de Investigación Científica 1992, área de Ciencias Naturales. AIC, México; Cátedra Patrimonial de Excelencia Nivel II, CONACYT (1997-1999).
Otros logros académicos: Autor o coautor de 12 géneros y 250 especies
americanas de coleópteros lamelicornios nuevos para la ciencia, sobre todo de la fauna mexicana. Asesor científico y gráfico de la Exposición Itinerante "Escarabajos, 200 Millones de Años de Evolución" que recorrió México (1983- 1992) y España (1993-1996).
Cargos Honoríficos: Presidente, Vicepresidente y Secretario de la Sociedad Mexicana de Entomología, A.C. (1984-1990); Editor de la revista científica Folia Entomológica Mexicana (1981-1992); Miembro de los Consejos Editoriales de 5 revistas mexicanas de Zoología (1984-2008). Curador General de la Colección Entomológica del Instituto de Ecología, A.C. (IEXA) (2002-2012).

3
Conferencias magistrales Un panorama sobre el estudio de los escarabajos melolóntidos y cetónidos en América
Latina Miguel Ángel Morón
Instituto de Ecología, A. C. Xalapa, Veracruz, México. [email protected]
Aunque no disponemos de un inventario preciso o actualizado, se estima que entre México, las Antillas, Argentina y Chile, existen poco más de 5000 especies de escarabajos melolóntidos y cetónidos, cifra que representa el 90% de las especies referidas para América y el 25% de las especies enlistadas para el mundo (Krajcik 2012). Antes de continuar es necesario aclarar el criterio taxonómico empleado en este ensayo. La clasificación a nivel familia atiende a la propuesta de Endrödi (1966) con las modificaciones sugeridas por Morón (1984, 1997a, 2003, 2010, 2013), Cherman y Morón (2014), para los agrupamientos de especies fito-saprófagas, de manera que en los territorios de América Latina la familia Melolonthidae queda formada por las subfamilias Melolonthinae, Hopliinae, Rutelinae, Dynastinae y Orphninae; en tanto que la familia Cetoniidae está representada por especies de las subfamilias Cetoniinae, Trichiinae y Valginae (Cuadro 1). Consideramos fito-saprófagos a los escarabajos cuyos adultos se alimentan con tejidos vegetales, secreciones azucaradas o polen, mientras que sus larvas se alimentan con raíces, tallos subterráneos, madera en descomposición, hojarasca o humus (Morón et al. 2010). Si las especies de estos escarabajos estuvieran uniformemente repartidas en los 137 millones de kilómetros cuadrados de la superficie emergida del planeta (excluyendo la Antártida, donde no se han registrado escarabajos) en todo el continente americano deberían existir cuando menos 6200 especies de melolóntidos y cetónidos, de forma que sólo nos faltaría describir y catalogar alrededor de 550 especies. Pero existen varios factores que complican estos cálculos, como por ejemplo: a) las especies usualmente son más diversas y están concentradas entre los trópicos de Cáncer y Capricornio; b) cuando menos un tercio del territorio intertropical americano es poco accesible para la exploración biológica, y no se cuenta con muestras representativas de numerosos biomas; c) más de la mitad de la misma área se encuentra sujeta a una presión antrópica continua que afecta directamente las comunidades silvestres y restringe o elimina especies de melolóntidos y cetónidos con hábitos especializados y distribución restringida, y favorece el establecimiento de especies nativas oportunistas. Además debemos considerar la escasez de estudiosos de los melolóntidos y cetónidos, quienes difícilmente hemos logrado avanzar en la catalogación de esta riqueza biológica durante los últimos cien años. Las mariposas diurnas (Papilionoidea), las hormigas (Formicidae) y los escarabajos lamelicornios y pectinicornios (Scarabaeoidea) se han citado en varias ocasiones como ejemplos de los grupos de insectos mejor conocidos a nivel mundial, sin embargo esto puede corresponder a un efecto engañoso de proporción, ya que la mayor parte de los grupos de insectos y otros artrópodos han recibido mucha menos atención a pesar de que entre todos constituyen el 80% de la riqueza de especies animales efectivamente catalogada en el mundo hasta principios del siglo XXI (Morón 1984, Bellés 1995, Zhang 2011).

4
El decir que los escarabajos figuran entre los grupos “mejor conocidos” usualmente significa que con cierta facilidad es posible asignarle un binomio específico a un ejemplar, pero en realidad no sabemos casi nada sobre sus hábitos, desarrollo, valencia en un ecosistema, origen, etc. En realidad contamos con los expedientes biológicos medianamente completos de unas 50 especies americanas de escarabajos melolóntidos y cetónidos (equivalentes al 1% de los taxones catalogados), sobre todo debido a que varias de ellas son abundantes, tienen distribución geográfica y ecológica amplia, o se les ha considerado perjudiciales en alguna región. Sobre las otras 4955 especies referidas para América Latina solo tenemos la descripción taxonómica de los adultos, algunos registros de distribución, datos esporádicos sobre sus preferencias alimentarias o forma de captura. En cuanto a los estados inmaduros se han descrito los caracteres larvales de cerca de 150 especies y las pupas de 53 especies de Melolonthinae, Rutelinae, Dynastinae, Orphninae, Cetoniinae y Trichiinae. El primer estudio integral sobre la fauna de escarabajos lamelicornios fitófagos de México fue publicado por Henry W. Bates como parte de la magna obra Biologia Centrali-Americana (1887-1889), la cual contiene 1398 registros de distribución para 416 especies, y las descripciones de 202 especies nuevas de Melolonthidae y Cetoniidae de México. Dicha información abarcó localidades en 26 estados y los alrededores de la ciudad de México (Cuadro 2). Entre 1890 y 1943 varios autores norteamericanos y europeos publicaron descripciones nuevas o registros precisos para 242 especies de estos grupos, las cuales junto con las referidas por Bates fueron compiladas por Blackwelder (1944) en un listado con referencia solo a países latinoamericanos que incluyó 658 especies para México (Cuadro 3). Como un punto de referencia para medir los avances en el estudio e inventario de estos coleópteros a continuación nos referimos a las cifras de nuevas especies en cuya descripción ha participado el autor de estas líneas, así como a las larvas y pupas cuyas características diagnósticas se han dado a conocer por dicho autor y sus colaboradores (Cuadro 4). Sumando las aportaciones de una docena de autores, en un período de 70 años a partir del catálogo de Blackwelder (1944) se ha logrado incrementar en un 38% la lista de especies de Melolonthidae y Cetoniidae distribuídas entre México, las Antillas y el Cono Sur. En lo correspondiente a México, el incremento es equivalente a un 79% del cuál un 38% se originó en las publicaciones de Morón y colaboradores (199 especies nuevas para la Ciencia). Pero el impulso al estudio de los estados inmaduros es notablemente mayor, porque en el mismo lapso de tiempo el número de larvas descritas aumentó hasta ocho veces, y prácticamente se dio inicio al estudio comparativo de las pupas de estos grupos. Durante el mismo período las aportaciones fueron mucho menos numerosas para el catálogo centroamericano, y poco significativas para Sudamérica, donde residen el 68% de las especies de nuestro interés, aunque el avance en las descripciones de larvas y pupas fue relevante. Además de contribuir a complementar y actualizar los catálogos, es pertinente recordar que la identificación específica precisa es fundamental para otorgar el debido valor a las investigaciones sobre biología, fisiología, comportamiento, ecología, zoogeografía y manejo de plagas. Pero ello no debe restringir las actividades de un investigador a los aspectos taxonómicos o descriptivos, sino al contrario, es muy conveniente relacionar un tema con otros para ampliar y profundizar la visión sobre el grupo, permitiendo aproximarse a la comprensión del sitio que ocupan los escarabajos en el ambiente, como sus actividades pueden favorecernos o afectarnos, y como nuestras acciones los

5
impactan. Para aproximarnos a esta propuesta, desde 1985 M. A. Morón definió la línea de investigación titulada “Taxonomía, Sistemática, Ecología, Zoogeografía, Importancia Agropecuaria y Forestal de adultos e inmaduros de coleópteros lamelicornios en América Latina”, dividida en cinco enfoques, relacionados entre sí en diversos grados, en los cuales se fueron enmarcando sus publicaciones y colaboraciones. 1.- Estudios faunísticos. Es uno de los enfoques más completos para ubicarse en la línea de investigación citada, porque el primer objetivo consiste en colectar lamelicornios empleando todos los medios posibles, durante seis meses a dos años, en una región definida por la confluencia de dos o más tipos de vegetación o usos de suelo. Para cumplir este objetivo se requiere aplicar una buena diversidad de conocimientos y técnicas, no solo de tipo entomológico, porque es necesario conocer suficiente de la vegetación, el suelo, el clima, la historia geológica, los antecedentes agropecuarios y forestales de la región. Después de las capturas siguen los procedimientos para la conservación e identificación de los ejemplares, hasta obtener una lista de binomios específicos organizados en un sistema de clasificación, y una colección de referencia bien etiquetada. Con esta información se construye una base de datos, gráficas y cuadros comparativos que nos ayudarán a analizar la distribución espacial y temporal de las especies representadas en la muestra local, a concentrar las observaciones sobre sus hábitos y preferencias, hasta lograr gráficos que sinteticen los resultados para apoyar las comparaciones dentro de la región estudiada y las relaciones con otras localidades donde se hayan registrado las mismas especies. Durante el proceso se evalúa la abundancia y el predominio de las especies, y se pondera su valor como indicador ecológico o biogeográfico, así como su potencial como integrante de ambientes agrícolas, pecuarios, forestales o urbanos. Gran parte del ejercicio descrito lo realizó el autor de estas líneas durante 1973-1975 como tesis de licenciatura en Biología en Villa de Allende, localidad situada cerca del centro del Eje Neovolcánico Transmexicano y, valorando su alcance como recurso formativo que permite al estudiante integrar muchos de los contenidos del plan de estudios con la realidad nacional, se propuso como alternativa de tesis en varias regiones del país, con cierto grado de éxito, ya que en un lapso de 26 años 14 estudiantes evaluaron la fauna de lamelicornios en 14 localidades de siete estados de la federación (Cuadro 5). Mediante esa introducción al ejercicio profesional, el 50% de esos biólogos han continuado investigando aspectos de la taxonomía, biología, ecología o importancia de estos coleópteros y el 25% de ellos han asesorado a otros jóvenes sobre la misma línea de trabajo. Como aportación al inventario faunístico nacional, durante los estudios faunísticos desarrollados en las tesis de Escoto-Rocha (1984), Deloya-López (1987), Delgado-Castillo (1989), Carrillo-Ruiz (2000), Percino-Figueroa (2002), Muñoz- Hernández (2003), Reyes-Novelo (2003), Chacón-Aguayo (2005), Ramírez-Ponce (2005), García de Jesús (2006), Salamanca-Calixto (2006), Delgado-Solano (2007), Cuate-Mozo (2008) y Minor-Montiel (2010) se obtuvieron 508 registros precisos para 55 géneros y 267 especies de Melolonthidae y Cetoniidae de México, y ejemplares que representaron a 32 especies nuevas para la Ciencia. 2.- Taxonomía y Sistemática. Es otro de los enfoques de la línea de investigación antes dicha que abarca las revisiones de subtribus, grupos de especies, subgéneros o géneros y las descripciones aisladas de nuevos taxones, así como la descripción de estados

6
inmaduros, todo lo cual con frecuencia ha incluido claves dicotómicas. Estas actividades requieren de un apoyo gráfico que usualmente consiste en ilustraciones esquemáticas o realzadas con sombreado, cuya realización no se reconoce durante las evaluaciones académicas de investigación, tal vez porque se considera una labor técnica similar a la preparación de ilustraciones basadas en fotografías. Sin embargo es frecuente que los investigadores dediquen bastante tiempo para realizar personalmente estas actividades complementarias del trabajo taxonómico. En este caso, el suscrito ha incorporado 3440 ilustraciones a tinta en 124 artículos de investigación y otras 410 ilustraciones en siete libros, aparte de mapas, gráficas, esquemas y fotografías a color o gama de grises, que representan especies de México, América Central y en menor proporción, de Sudamérica. Además de los métodos taxonómicos convencionales algunos trabajos de investigación en posgrado han incorporado técnicas apropiadas para los análisis filogenéticos basados en caracteres morfológicos (Carrillo-Ruiz 2006, Ramírez-Ponce 2007, 2012, Rivera-Gasperín 2012) algunos de ellos producto de la exploración con microscopía electrónica de barrido que, entre otros caracteres interesantes, ha permitido descubrir una gran diversidad estructural en los órganos sensoriales cuticulares de estos escarabajos, como los receptores antenales y la microestructura de las sedas piliformes y escamiformes (Romero-López et al. 2013). 3.- Ecología y Zoogeografía. Desde el proyecto original sobre la fauna de Villa de Allende fue evidente la importancia de relacionar la presencia de las especies de Melolonthidae con factores ambientales, como la marcha anual de temperaturas y precipitaciones mensuales, el predominio de las especies arbóreas y la existencia de cultivos. También se aceptó el valor de la fenología para interpretar la sucesión y el predominio de cada especie a lo largo del ciclo anual. Al paso de los años, con los estudios faunísticos llevados a cabo aparte de los trabajos de tesis, se fueron afinando los conceptos sobre riqueza específica, ensambles, complejos, gremios, diversidad, composición taxonómica, similitud faunística, endemismo, centros de origen, corredores y barreras, aplicables a los escarabajos de la Zona de Transición Mexicana y la región Neotropical, hasta iniciar la fundamentación de las propuestas para considerar a los Melolonthidae y Cetoniidae como grupos bioindicadores ecológicos y biogeográficos (Morón 1991, 1994, 1997b, 2014, Lobo y Morón 1993). Durante los trabajos de campo se evaluó la participación de las larvas de Melolonthidae en las comunidades edafícolas (Morón 1981) y se dedicó atención a los ensambles de escarabajos asociados con el arbolado muerto, para evaluar la composición de especies de Melolonthidae con larvas saproxilófagas, y su coexistencia con adultos e inmaduros de otras familias de coleópteros (Morón 1985, Morón y Terrón 1986, Morón et al. 1988). Al finalizar la primera etapa del estudio de la fauna de lamelicornios de la estación de Biología Tropical de la UNAM en Los Tuxtlas, Veracruz (Morón 1979) se inició la evaluación de la composición de géneros y especies registradas en cada localidad explorada, al comparar Villa de Allende con Los Tuxtlas. En el primer estudio faunístico de Melolonthidae en la Reserva de la Biosfera La Michilía, Durango (Morón 1981), se evaluó la biomasa de las especies atraídas por la trampa de luz y su frecuencia por hora, y se aplicaron los índices de predominio específico (c) de Simpson y de diversidad (d) de Menhinick para discutir los resultados sobre la actividad de las especies locales. La fauna de melolóntidos de La Michilía se comparó con las faunas de

7
Villa de Allende, las montañas de Hidalgo (Zacualtipán y Molango) y el bosque tropical de Los Tuxtlas, empleando el índice de similitud (QS) de Sørenson, registrando los valores a nivel específico y genérico en una matriz para las cinco localidades. A partir del trabajo efectuado en Cuetzalan, Puebla (Carrillo-Ruiz 2000) en los estudios faunísticos se utilizó el análisis de parsimonia de endemismos para obtener hipótesis de las relaciones entre las localidades mediante la similitud de su fauna de melolóntidos a nivel de especies. Buena parte de la información reunida sobre la distribución de especies de Melolonthidae se discutió en un ensayo basado en análisis de trazos individuales y en las provincias biogeográficas delineadas para México (Morón 2006). Algunas investigaciones realizadas como parte de los programas de posgrado han incluido análisis de agrupamientos faunísticos de Melolonthidae con la estimación de la riqueza específica por tipo de vegetación empleando el índice (H’) de Shannon-Wiener, un modelo de dominancia/diversidad mediante el logaritmo decimal de la proporción de individuos de cada especie para visualizar gráficamente la estructura de la comunidad de escarabajos, así como la estimación de la eficiencia de captura y predicción de la riqueza total mediante curvas de acumulación con el programa EstimateS 8.0, y el índice de complementariedad de Coldwell-Coddington para determinar la diferencia en la composición de especies en diferentes localidades, entre otros recursos estadísticos aplicables a las muestras disponibles (García de Jesús 2011, Delgado-Solano 2011). Otros trabajos de posgrado se realizaron con la premisa de considerar a los melolóntidos como bioindicadores ecológicos, empleando agrupamientos de larvas edafícolas (Tapia-Rojas 2005) o muestras de adultos obtenidos con trampas atrayentes y larvas edafícolas, que permitieron calcular las diversidades alfa, beta, gamma y la dominancia (Yanes-Gómez 2007). 4.- Contribución al control de plagas. La identificación precisa y el conocimiento paulatino sobre las preferencias alimentarias de las larvas de Melolonthidae y Cetoniidae, así como su abundancia y heterogeneidad, han permitido generar hipótesis sobre las alternativas para propiciar el equilibrio de las poblaciones de “gallina ciega” que pueden ocasionar pérdidas importantes en la producción agrícola o forestal, o cuya actividad mejora las condiciones de los suelos cultivables. En el transcurso de 40 años se han tenido pocas oportunidades para diseñar trabajo de campo encaminado a evaluar las poblaciones edafícolas de estos escarabajos, sin embargo ahora se cuenta con información confiable básica para aplicarla en distintos ambientes, aunque una de las premisas fundamentales descubiertas es que cada localidad tiene características únicas que obligan a tratamientos específicos. La experiencia más amplia y detallada, que incluye la mayoría de los factores determinantes del problema “gallina ciega” se encuentra ejemplificada en los trabajos realizados en la región cañera de Tepic, Nayarit (Morón et al. 1996, 1998). Cabe destacar los estudios desarrollados en cultivos de maíz y sorgo en Tamaulipas (Rodríguez del Bosque 1993), en cultivos de maíz en Jalisco y Michoacán (Nájera-Rincón 1993, 1998) y en Los Altos de Chiapas (Ramírez-Salinas et al. 2000, Castro-Ramírez et al. 2005), así como el trabajo reciente en plantaciones de arbolitos de Navidad en Veracruz (Morón et al. 2014). En algunos trabajos de posgrado se han logrado avances regionales sobre los aspectos fundamentales de las comunidades subterráneas de melolóntidos, por ejemplo en la región central del estado de Puebla (Aragón 1997, 2005) y en el norte de Sinaloa (Lugo-García 2010). 5.- Divulgación. En 1983 se reunió un equipo de diseñadores, museográfos, artistas y especialistas, para organizar un programa de divulgación sobre la importancia de los escarabajos, conformado por una exposición itinerante, folletos, conferencias y un libro

8
con nivel de divulgación universitario (Morón 1984). Durante casi diez años la exposición recorrió parte del país acompañada de algunas conferencias y folletos sencillos, generando un buen índice de interés en el público, atrayendo a estudiantes y coleccionistas, y el libro tuvo dos ediciones y una versión digitalizada recientemente. Panorama futuro en México. Aun cuando se ha logrado un buen nivel de conocimiento sobre la distribución de la mayor parte de los grupos específicos de Melolonthidae y Cetoniidae en México, persiste la necesidad de continuar con el enfoque faunístico de la línea de investigación, principalmente en las regiones menos conocidas del país (como los núcleos de las Sierras Madre Occidental y del Sur) o en los estados donde se han obtenido menos datos, como Aguascalientes, Campeche, Guanajuato, Querétaro y Zacatecas (Morón et al. 2014b). También es importante registrar y evaluar los cambios ocurridos después de varios años en las faunas de melolóntidos en localidades ya estudiadas, para conocer más sobre la dinámica de los ensambles o gremios de especies (Lobo y Morón 1993) y estimar con detalle el impacto que la expansión de las fronteras agropecuaria y urbana tienen en las comunidades de estos insectos. En el enfoque sistemático es necesario completar las revisiones de los géneros con mayor número de especies en el país, como Phyllophaga (+390), Diplotaxis (+180) y Paranomala (+80), así como estudiar la filogenia de los agrupamientos genéricos originados o diversificados en la Zona de Transición Mexicana, empleando caracteres morfológicos y moleculares. También se requiere profundizar en la taxonomía de los estados inmaduros de un gran número de especies y géneros característicos de México, para disponer de descripciones y claves fundamentales para otro tipo de estudios. En forma paralela, es importante obtener información sobre los hábitos de alimentación y reproducción de un mayor número de especies representativas de los linajes de melolóntidos y cetónidos en el país, a fin de contar con un soporte adecuado para regular las especies dañinas y proteger a las especies benéficas o a las endémicas amenazadas. Las actividades de divulgación deben intensificarse a fin de reclutar más interesados en el tema de investigación, que continúen y profundicen el estudio de estos grupos, además de promover el desarrollo de líneas similares en otras familias de insectos aún poco conocidas. Visión retrospectiva y panorama futuro en Centro y Sudamérica. A pesar de la importancia de las cifras de riqueza específica (Cuadro 3) todos los aspectos hasta aquí citados para el estudio de los melolóntidos han recibido menos atención al sur de México (Cuadro 4), por la escasez de especialistas, y la ausencia de una obra de compilación equivalente a la Biologia Centrali Americana. El autor ha promovido la línea de investigación en colaboración con profesores y estudiantes de instituciones centro y sudamericanas, con éxito relativo a causa de la gran desproporción entre las dimensiones de la problemática del inventario en el enorme y diverso territorio ubicado al sur de Panamá, y la disponibilidad de recursos humanos y materiales. Con enfoque faunístico se han llevado a cabo algunas investigaciones en Colombia (Pardo-Locarno 1993, Restrepo-Giraldo 1998, Restrepo-Giraldo et al. 2003, Neita-Moreno 2004), en Ecuador (Paucar-Cabrera 2005) y en Brasil (Morón 2004). Los

9
estudios sobre taxonomía y sistemática de los grupos neotropicales típicos realizados por latinoamericanos han sido escasos (Cano y Morón 1998, Solís 2004, Escalona y Joly 2006, Abadie et al. 2008, Cherman 2015). Algunos aspectos de la ecología, los estados inmaduros y la importancia agrícola de melolontinos, dinastinos, rutelinos y cetoninos se han documentado en localidades de Colombia (Vallejo 1997, Neita-Moreno 2006, Pardo-Locarno 2008), Uruguay (Alzugaray et al. 1998), Brasil (Cherman 2010, Oliveira et al. 2014), Bolivia (Copa-Bazán y Morón 2014). Uno de los primeros objetivos para el futuro inmediato de este tipo de estudios en América Central y Sudamérica debería consistir en completar inventarios nacionales de melolóntidos y cetónidos. En forma simultánea sería importante caracterizar la fauna de las principales formaciones vegetales originales entre Guatemala y Chile-Argentina, así como iniciar las revisiones taxonómicas de los géneros menos estudiados y con mayor número de especies en América del Sur, como Plectris (+350), Leucothyreus (+160), Isonychus (+135), Ceraspis (+90), Geniates (+40), y Gymnetis (+30). La descripción de larvas de tercer estadio para los principales géneros y grupos específicos en cada país sería un complemento indispensable para proseguir los estudios sobre ciclos vitales y hábitos de las especies que han demostrado capacidad para producir daños en plantas cultivadas. La coordinación de programas de divulgación con el apoyo a programas de estudios universitarios y de posgrado para reclutar estudiantes y facilitar su formación y especialización en esta disciplina debería sustentarse en programas nacionales o regionales adecuados para promover la investigación sobre todos los aspectos básicos y aplicados relacionados con la biodiversidad del neotrópico. Referencias Abadie, E. I., P. C. Grossi and P. S. Wagner. 2008. A field guide of the Dynastidae family of the south of South America. Publicado por los autores. Argentina. Alzugaray, R., A. Ribeiro, M. S. Zerbino, E. Morelli y E. Castiglioni. 1998. Situación de los insectos del suelo en Uruguay. Pp. 151-164. In: Morón, M. A. y A. Aragón (eds.). .) Avances en el estudio de la diversidad, importancia y manejo de los coleópteros edafícolas americanos. Publicación Especial Benemérita Universidad Autónoma de Puebla y Sociedad Mexicana de Entomología. Puebla, México. Aragón-García, A. 1997. Estudio de los Coleópteros Melolontidos asociados a la rizosfera de la caña de azúcar, en la zona de Atencingo, Puebla. Tesis de Maestría en Ciencias inédita. División de Estudios de Postgrado, Facultad de Ciencias, UNAM. Aragón-García, A. 2005. Biología, comportamiento e importancia de cinco especies del género Phyllophaga (Coleoptera: Melolonthidae) en agrosistemas del estado de Puebla. Tesis doctoral inédita. Posgrado en Ciencias Ambientales, Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla. Bates, H. W. 1886-1890. Biologia Centrali Americana. Insecta, Coleoptera, Volume II, Part 2, Pectinicornia and Lamellicornia. Taylor and Francis, London. 432 pp. Bellés, X. 1995. Una estirpe de rancio abolengo. El mundo de los insectos. Investigación y Ciencia, Temas 2: 2-8. Blackwelder, R. E. 1944. Checklist of the coleopterous insects of Mexico, Central America, the West Indies, and South America. Part 2. Bulletin United States National Museum 185: 189-341. Cano, E. B. y M. A. Morón. 1998. Phyllophaga (Coleoptera: Melolonthinae) de Guatemala. Sistemática, diversidad, biología y biogeografía. Informe de investigación inédito. Universidad del Valle de Guatemala y Secretaria Nacional de Ciencia y Tecnología, Guatemala.

10
Carrillo-Ruiz, H. 2000. Fauna de Coleoptera Lamellicornia de Cuetzalan del Progreso, Puebla, México. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Carrillo-Ruiz, H. 2006. Análisis filogenético de la tribu Hoplini (Coleoptera: Melolonthidae). Tesis doctoral inédita. División de Posgrado, Instituto de Ecología, A.C. Xalapa. Castro-Ramírez, A. E., H. Delfín-González, V. Parra-Tabla y M. A. Morón, 2005. Fauna de Melolóntidos (Coleoptera: Scarabaeoidea) asociados al maíz (Zea mays L.) en Los Altos de Chiapas, México. Folia Entomológica Mexicana 44 (3): 339-365. Chacón-Aguayo, A. L. 2005. Fauna de Coleoptera Lamellicornia de Teotlalzingo, Puebla, México. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Cherman, M. A. 2010. Análise populacional de Melolonthidae (Coleoptera) da regiao do Plan Alto do Rio Grande do Sul. Tesis Maestria em Ciências (inédita). Centro de Ciências Rurais, Universidade Federal de Santa Maria, RS, Brasil. 139 pp. Cherman, M. A. 2015. Revisão e análise cladística de Liogenys Guérin-Meneville (Coleoptera, Melolonthidae). Tesis doctoral inédita. Programa de Posgraduados, Universidade Federal do Parana, Curitiba, Brasil. Cherman, M. A. y M. A. Morón. 2014. Validación de la familia Melolonthidae Leach, 1819 (Coleoptera: Scarabaeoidea). Acta Zoológica Mexicana (nueva serie) 30(1): 201-220. Copa-Bazan, F. y M. A. Morón. 2014. Importancia del gusano blanco (Coleoptera: Melolonthidae: Dynastinae) en cultivos de caña de azúcar en Santa Cruz, Bolivia. pp. 51-64. In: Aragón-García, A. (ed.) Diversidad e importancia agrícola de Coleópteros edafícolas. Benemérita Universidad Autónoma de Puebla. México. Cuate-Mozo, V. A. 2008. Fauna de Coleópteros Lamellicornia del municipio de Chiautla de Tapia, Puebla. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Delgado-Castillo, L. L. 1989. Fauna de Coleópteros Lamelicornios de Acahuizotla, Guerrero, México". Tesis profesional inédita. Facultad de Ciencias, UNAM. México, D. F. Delgado-Solano, J. M. 2007. Fauna de Coleoptera Lamellicornia del municipio de Chignahuapan, Puebla, México”. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Delgado-Solano, J. M. 2011. Diversidad de Scarabaeoidea (Coleoptera) en los principales tipos de vegetación de Montebello, Chiapas, México. Tesis de Maestría en Recursos Naturales y Desarrollo Rural (inédita). El Colegio de la Frontera Sur (ECOSUR). San Cristóbal de Las Casas, Chiapas, México. 134 pp. Deloya-López, A.C. 1987. Fauna de Coleópteros Lamelicornios del Sur de Morelos, México. Tesis profesional inédita. Facultad de Ciencias, UNAM. México, D.F. Endrödi, S. 1966. Monographie der Dynastinae (Coleoptera: Lamellicornia) I Teil. Entomologische Abhandlungen Museum Tierkunde Dresden 33: 1-457. Escalona, H. E. y L. J. Joly. 2006. El género Ligyrus Burmeister 1847 (Coleoptera: Scarabaeidae: Dynastinae, Pentodontini) en Venezuela. Boletín Sociedad Entomológica Aragonesa 39: 111-137. Escoto-Rocha, J. 1984. Análisis de la fauna de coleópteros Scarabaeidae y Melolonthidae de Calvillo, Aguascalientes. Tesis profesional inédita. Escuela de Biología. Universidad Autónoma de Aguascalientes, Aguascalientes.

11
García de Jesús, S. 2006. Estudio faunístico de Coleoptera Lamellicornia de la región La Malinche, Tlaxcala, México. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. García de Jesús, S. 2011. Fauna de Coleoptera Scarabaeoidea de los bosques de coniferas de los municipios de Emiliano Zapata y Terrenate, Tlaxcala, México. Tesis de Maestría en Ciencias, inédita. Universidad Autónoma de Tlaxcala. Krajcik, M. 2012. Checklist of the World Scarabaeoidea. Supplement 5. Animma.X, Plzen, Czech Republic. 278 pp. Lobo, J. y M.A. Morón. 1993. La modificación de las comunidades de coleópteros Melolonthidae y Scarabaeidae en dos áreas protegidas mexicanas. Giornale Italiano di Entomologia 6: 391-406. Lugo-García, G. A. 2010. Diversidad, daños y manejo de Scarabaeoidea en el cultivo de maíz en el norte de Sinaloa, México. Tesis doctoral inédita. Instituto de Fitosanidad, Colegio de Posgraduados, Montecillo, México. Minor-Montiel, P. 2010. Coleópteros lamelicornios de la barranca de Huehuetitla, Tlaxcala, México. Tesis profesional inédita. Escuela de Biología, Universidad Autónoma de Tlaxcala. Morón, M. A. 1975. Coleópteros Lamelicornios de Villa de Allende, Estado de México. Tesis profesional inédita. Facultad de Ciencias, Universidad Nacional Autónoma de México. Morón, M. A. 1979. Fauna de coleópteros Lamelicornios de la Estación de Biología Tropical U.N.A.M. "Los Tuxtlas", Ver. México. Anales Instituto Biología UNAM serie Zoología 50 (1): 375-454. Morón, M.A. 1981. Fauna de coleópteros Melolonthidae de la Reserva de la Biósfera "La Michilía", Durango. México. Folia Entomológica Mexicana 50: 3-69. Morón, M. A. 1984. Escarabajos, 200 millones de años de evolución. Instituto de Ecología, A. C. México. 111 pp. Morón, M.A. 1985. Observaciones sobre la biología de dos especies de rutelinos saproxilófagos en la Sierra de Hidalgo, México. (Col. Melolonthidae, Rutelinae). Folia Entomológica Mexicana 64:41-53. Morón, M. A. 1991. Los escarabajos fitófagos, un ejemplo de la riqueza biótica de Mesoamérica (Coleoptera: Scarabaeoidea). Giornale Italiano di Entomologia Vol. 5: 209-218. Morón, M. A. 1994. La diversidad genérica de los Coleopteros Melolonthidae en México. Acta Zoológica Mexicana (n.s.) 61: 7-19. Morón, M. A. 1997a. Antecedentes. Pp. 1-8. In: Morón, M. A., B. C. Ratcliffe y C. Deloya. Atlas de los escarabajos de México. Coleoptera Lamellicornia. Vol. I. Familia Melolonthidae. CONABIO y Sociedad Mexicana de Entomología, A. C. México. Morón, M. A. 1997b. Inventarios faunísticos de los Coleoptera Melolonthidae Neotropicales con potencial como bioindicadores. Giornale Italiano di Entomologia, 8: 265-274 Morón, M. A. 2003. Antecedentes. Pp. 11-18. In: Morón, M. A. (ed.). Atlas de los escarabajos de México. Coleoptera Lamellicornia. Vol. II. Familias Scarabaeidae, Trogidae, Passalidae y Lucanidae. Argania editio, Barcelona. Morón, M.A. 2005. Melolontídeos edafícolas. Pp. 133-166. In: Salvadori, J. R., J. C. Avila y M. T. B. Silva, (Eds.). Pragas de Solo en Brasil. Empresa Brasileira de Pesquisa Agropecuária, Passo Fundo, RS. Morón, M. A. 2006. Patrones de distribución de la familia Melolonthidae (Coleoptera). Pp. 295-331. In: Morrone, J. J., y J. Llorente-Bousquets (eds.). Componentes bióticos

12
principales de la entomofauna mexicana. Volumen 1. Facultad de Ciencias, UNAM, México, D. F. Morón, M. A. 2010. Diversidad y distribución del complejo “gallina ciega” (Coleoptera: Scarabaeoidea). Pp. 41-63. In: Rodríguez del Bosque, L. A. y M. A. Morón (eds.). Plagas del suelo. Mundi-Prensa, México. Morón, M. A. 2013. Introducción al conocimiento de los escarabajos de Puebla. Pp. 1-27. In: Morón, M. A., A. Aragón-García y H. Carrillo-Ruiz (eds.). Fauna de escarabajos del estado de Puebla. Publicado por Miguel Ángel Morón, Coatepec, Veracruz, México. Morón, M. A. 2014. Los escarabajos lamelicornios como indicadores ecológicos y biogeográficos. Pp. 307-324. In: González-Zuarth, C. A., A. Vallarino, J. C. Pérez-Jiménez y A. M. Low-Pfeng (eds.). Bioindicadores: guardianes de nuestro futuro ambiental. Instituto Nacional de Ecología y Cambio Climático (INECC) – Colegio de la Frontera Sur (ECOSUR). México. Morón, M. A., S. Hernández-Rodríguez y A. Ramírez, 1996. El complejo “gallina ciega” (Coleoptera: Melolonthidae) asociado con caña de azúcar en Tepic, Nayarit, México. Folia Entomológica Mexicana 98: 1-44. Morón, M.A., S. Hernández-Rodríguez y A. Ramírez-Campos, 1998. Las especies de Phyllophaga (Coleoptera: Melolonthidae) con importancia agrícola en Nayarit, México. pp. 79-98 In: Morón, M. A. y A. Aragón (Eds.) Avances en el estudio de la diversidad, importancia y manejo de los coleópteros edafícolas americanos. Publicación Especial Benemérita Universidad Autónoma de Puebla y Sociedad Mexicana de Entomología. Puebla, México. Morón, M. A., L. A. Rodríguez del Bosque, A. Aragón y C. Ramírez-Salinas. 2010. Biología y hábitos de coleópteros escarabaeoideos. Pp. 65-82. In: Rodríguez del Bosque, L. A. y M. A. Morón (eds.). Plagas del suelo. Mundi-Prensa, México. Morón, M. A., C. V. Rojas-Gómez y R. Arce-Pérez. 2014a. Fluctuación de poblaciones de “gallina ciega” en una plantación de Pseudotsuga macrolepis (Pinaceae) en Las Vigas, Veracruz, México. pp. 15-35. In: Aragón-García, A. (ed.) Diversidad e importancia agrícola de Coleópteros edafícolas. Benemérita Universidad Autónoma de Puebla. México. Morón, M. A., G. Nogueira, C. V. Rojas-Gómez & R. Arce-Pérez. 2014b. Biodiversidad de Melolonthidae (Coleoptera) en México. Revista Mexicana de Biodiversidad 85, Suplemento: 298-302. Morón, M.A. y R. Terrón. 1986. Coleópteros xilófagos asociados con Liquidambar styraciflua (L.) en la Sierra de Hidalgo, México. Folia Entomológica Mexicana 67:25-35. Morón, M.A., J. Valenzuela y R. Terrón. 1988. La macro-coleopterofauna saproxilófila del Soconusco, Chiapas, México. Folia Entomológica Mexicana 74:145-158. Muñoz-Hernández, A. 2003. Estudio de los Coleópteros Lamelicornios de Teziutlán, Puebla. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Nájera-Rincón, M. B. 1993. Coleópteros rizófagos asociados al maíz de temporal en el centro del estado de Jalisco, México. Identificación, ecología y control. Pp. 143-154. In: Morón, M. A. (ed.) Diversidad y manejo de plagas subterráneas. Sociedad Mexicana de Entomología e Instituto de Ecología, Xalapa, México. Nájera-Rincón, M. B. 1998. Diversidad y abundancia del complejo “gallina ciega” (Coleoptera: Melolonthidae) en agroecosistemas de maíz en la región templada de Michoacán. Pp. 99-106. In: Morón, M. A. y A. Aragón (eds.) Avances en el estado de la diversidad, importancia y manejo de los coleópteros edafícolas americanos.

13
Benemérita Universidad Autónoma de Puebla y Sociedad Mexicana de Entomología, Puebla, México. Neita-Moreno, J. C. 2004. Escarabajos (Scarabaeidae: Pleurosticti) de la selva baja del bosque pluvial tropical, Chocó, Colombia. Tesis de Ingeniero Forestal (inédita). Universidad Tecnológica del Chocó, Quibdó, Colombia. Neita-Moreno, J. C. 2006. Taxonomía y distribución de chisas (Coleoptera: Melolonthidae) en cinco cultivos de importancia agrícola en el Departamento de Cundinamarca, Colombia. Tesis de Maestría en Ciencias Agrarias con énfasis en Entomología, inédita. Escuela de Posgrados. Facultad de Agronomía, Universidad Nacional de Colombia-Bogotá. Oliveira, C. M., M. A. Morón y M. R. Frizzas. 2014. Phyllophaga nitididorsis Frey (Coleoptera: Melolonthidae) nova praga da soja em Cerrado no Minas Gerais, Brasil. pp. 233-247. In: Aragón-García, A. (ed.) Diversidad e importancia agrícola de Coleópteros edafícolas. Benemérita Universidad Autónoma de Puebla. México. Pardo-Locarno, L. C. 1993. Estudio preliminar de las especies de Melolonthidae del Valle del Cauca, Colombia con énfasis en la cuenca Calima-San Juan (Valle-Chocó). Pp. 83-90. In: Morón, M. A. (ed.) Diversidad y manejo de plagas subterráneas. Sociedad Mexicana de Entomologia e Instituto de Ecología, Xalapa. México. Pardo-Locarno, L. C. 2008. Macroinvertebrados edafícolas en agroecosistemas del municipio de El Cerrito (Valle) con énfasis en la comunidad de escarabajos Melolonthidae (Coleoptera: Scarabaeoidea). Tesis Doctorado en Ciencias-Biología (inédita). Facultad de Ciencias Naturales y Exactas, Universidad del Valle, Santiago de Cali. Colombia. 228 pp. Paucar-Cabrera, A. 2005. A catalog and distributional analysis of the Rutelinae (Coleoptera: Scarabaeidae) of Ecuador. Zootaxa 948: 1-92. Percino-Figueroa, S. M. 2002. Fauna de Coleóptera Lamelicornia del municipio de Zacatlán, Puebla, México. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Ramírez-Ponce, A. 2005. Fauna de Coleópteros Lamelicornios de Santiago Xiacui, Sierra Norte, Oaxaca, México. Tesis profesional inédita. Instituto Tecnológico Agropecuario de Oaxaca no. 23, Oaxaca. SEP-SEIT. Ramírez-Ponce, A. 2007. Relaciones filogenéticas del género Anomala Samouelle (Coleoptera: Melolonthidae; Rutelinae)” Tesis de Maestría en Ciencias, inédita. Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México. Ramírez-Ponce, A. 2012. Sistemática supraespecífica del género Paranomala Casey en Norte y Centroamérica (Coleoptera: Melolonthidae; Rutelinae: Anomalini)” Tesis doctoral inédita. Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México. Ramírez-Salinas, C., M.A. Morón y A. Castro-Ramírez, 2000. Descripción de los estados inmaduros de seis especies de Phyllophaga (Coleoptera: Melolonthidae, Melolonthinae) de la región Altos de Chiapas, México. Folia Entomológica Mexicana 109: 73-106. Restrepo-Giraldo, H. 1998. Aproximación al estudio de los escarabajos fitófagos (Coleoptera: Melolonthidae) en Colombia. Tesis profesional inédita. Facultad de Ciencias, Universidad Nacional de Colombia, sede Santafé de Bogotá. Restrepo-Giraldo, H., M. A. Morón, F. Vallejo, L.C. Pardo-Locarno y A. López-Avila, 2003. Catálogo de Coleoptera Melolonthidae (Scarabaeidae Pleurosticti) de Colombia. Folia Entomológica Mexicana 42 (2): 239-263

14
Reyes-Novelo, E. A. 2003. Fauna de las familias Melolonthidae y Passalidae (Coleoptera: Scarabaeoidea) en dos zonas de Yucatán, México. Tesis profesional inédita. Instituto Tecnológico Agropecuario No. 2, Conkal, Yucatán, SEIT-SEP. Rivera-Gasperín, S. L. 2012. Relaciones filogenéticas de la sutribu Rhizotrogina Burmeister, 1855 (Coleoptera: Melolonthidae: Melolonthinae) basada en caracteres morfológicos. Tesis de Maestría en Ciencias, inédita. División de Posgrado, Instituto de Ecología, A.C. Xalapa. Rodríguez del Bosque, L. A. 1993. Abundancia estacional y ecología de coleópteros rizófagos: un estudio durante 15 años en agroecosistemas del norte de Tamaulipas. Pp.7-15. In: Morón, M. A. (ed.). Diversidad y manejo de plagas subterráneas. Sociedad Mexicana de Entomología e Instituto de Ecología, Xalapa, México. Romero-López, A. A., H. Carrillo-Ruiz y M. A. Morón. 2013. Morphological diversity of antennal sensilla in Hopliinae (Coleoptera: Scarabaeoidea: Melolonthidae). Academic Journal of Entomology 6(1): 20-26. Salamanca-Calixto, C. 2006. Fauna de Coleoptera Lamellicornia en la región de Tlachichuca y Ahuatepec del Camino (Ciudad Serdán), Puebla, México. Tesis profesional inédita. Escuela de Biología, Benemérita Universidad Autónoma de Puebla. Solís, A. 2004. Escarabajos fruteros de Costa Rica. Instituto Nacional de Biodiversidad. Santo Domingo de Heredia. Costa Rica. Tapia-Rojas, A. M. 2005. Estudio de las larvas de coleópteros melolóntidos edafícolas en ambientes forestales de Puebla: su posible uso como bioindicadores. Tesis de Maestría en Ciencias, inédita. Posgrado en Ciencias Ambientales, Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla. Vallejo, F. 1997. Contribución al conocimiento de las plagas subterráneas-chisas (Coleoptera: Melolonthidae) del Oriente de Antioquia. Tesis de Maestría en Entomología (inédita). Facultad de Ciencias, Universidad Nacional de Colombia-Medellín. Yanes-Gómez, G. 2007. Coleópteros Scarabaeoidea de Santo Domingo Huehuetlan, Puebla, y su posible uso como indicadores ecológicos. Tesis de Maestría en Ciencias, inédita. Posgrado en Ciencias Ambientales, Instituto de Ciencias, Benemérita Universidad Autónoma de Puebla. Zhang, Z.Q. 2011. Animal biodiversity. An introduction to higher-level classification and taxonomic richness. Zootaxa 3148: 7-12.

15
Cuadro 1. Clasificación supragenérica de los Scarabaeoidea fito-saprófagos en América Latina, y riqueza específica por tribus (abreviaturas: Mex= México; Ant.= Antillas;
CA= Centroamérica; SA= Sudamérica). Datos inéditos de M. A. Morón. Familias Subfamilias Tribus Núm aprox spp. Distribución Melolonthidae Melolonthinae Melolonthini 844 Mex, Ant, CA, SA
Diplotaxini 281 Mex, CA, SA Liparetrini 20 SA Sericini 172 Mex, CA, SA Sericoidini 53 SA Podolasini 11 Mex Oncerini 1 Mex Macrodactylini 939 Mex, CA, SA Pachydemini 29 Mex, Ant, SA
Hopliinae Hopliini 25 Mex, CA Rutelinae Rutelini 740 Mex, Ant, CA, SA
Anomalini 215 Mex, Ant, CA, SA Anoplognathini 87 Mex, CA, SA Geniatini 327 Mex, Ant, CA, SA Spodochlamyini 17 CA, SA
Dynastinae Dynastini 50 Mex. Ant, CA, SA Cyclocephalini 456 Mex, Ant, CA, SA Pentodontini 118 Mex, Ant, CA, SA Oryctini 132 Mex, Ant, CA, SA Agaocephalini 43 Mex, Ant, CA, SA Phileurini 162 Mex, Ant, CA, SA
Orphninae Aegidini 7 Mex, Ant, CA, SA Cetoniidae
Cetoniinae Cetoniini 53 Mex, CA, SA Gymnetini 148 Mex, Ant, CA, SA Goliathini 4 Mex Cremastocheilini 29 Mex, CA, SA
Trichiinae Trichiini 28 Mex, CA, SA Incaini 12 Mex, CA, SA
Valginae Valgini 1 Mex Totales 8 29 5004 Cuadro 2. Registros de especies de Melolonthidae y Cetoniidae por entidad en México
(Bates 1888-89). Entidad registros Entidad registros Entidad registros Aguascalientes 5 Guerrero 96 Puebla 65 Campeche 5 Hidalgo 17 Querétaro 2 Chiapas 31 Jalisco 14 San Luis Potosí 14 Chihuahua 20 México 24 Sinaloa 11 Coahuila 10 Michoacán 8 Sonora 22 Colima 22 Morelos 33 Tabasco 22 Cd. de México 38 Nayarit 3 Tamaulipas 1 Durango 77 Nuevo León 2 Veracruz 451 Guanajuato 34 Oaxaca 217 Yucatán 31
Cuadro 3. Especies de Melolonthidae y Cetoniidae por regiones americanas
(Blackwelder 1944). subfamilias México Centroamérica Sudamérica Antillas Totales Melolonthinae 291 118 1011 90 1510 Rutelinae 170 149 791 10 1120 Dynastinae 110 57 381 40 588 Cetoniinae 76 28 92 4 200 Trichiinae 10 3 9 0 22
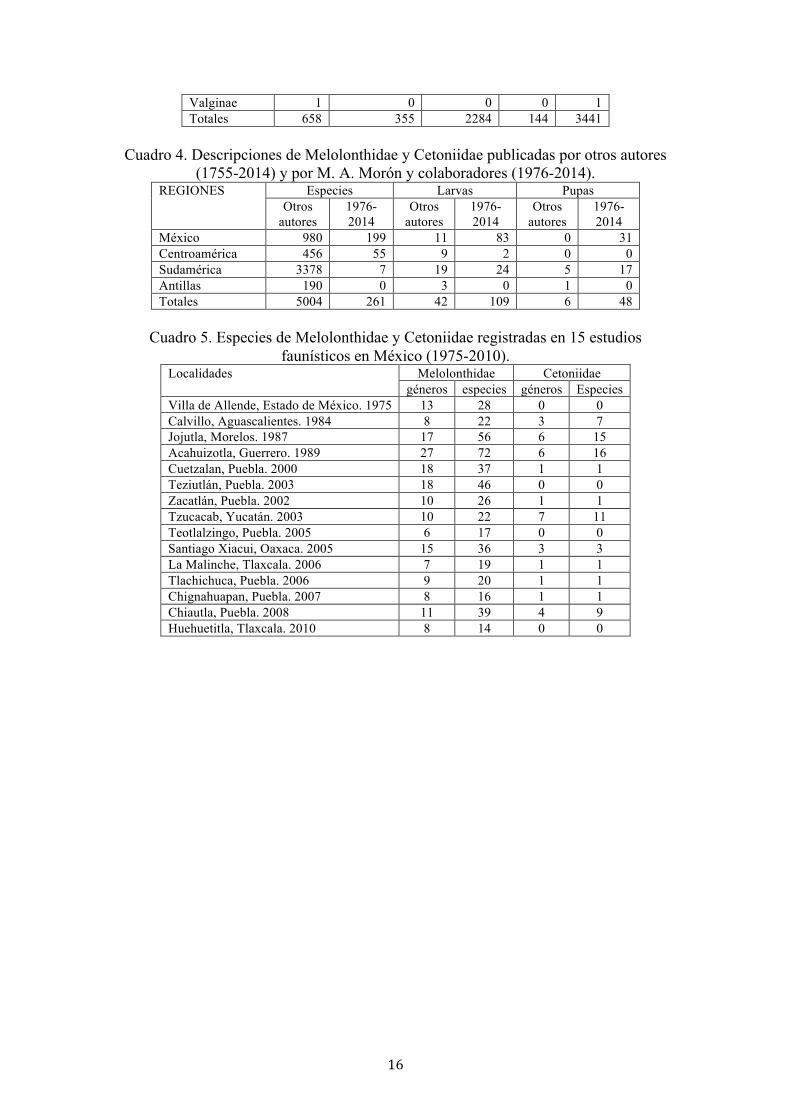
16
Valginae 1 0 0 0 1 Totales 658 355 2284 144 3441
Cuadro 4. Descripciones de Melolonthidae y Cetoniidae publicadas por otros autores
(1755-2014) y por M. A. Morón y colaboradores (1976-2014). REGIONES Especies Larvas Pupas
Otros autores
1976- 2014
Otros autores
1976- 2014
Otros autores
1976-2014
México 980 199 11 83 0 31 Centroamérica 456 55 9 2 0 0 Sudamérica 3378 7 19 24 5 17 Antillas 190 0 3 0 1 0 Totales 5004 261 42 109 6 48
Cuadro 5. Especies de Melolonthidae y Cetoniidae registradas en 15 estudios
faunísticos en México (1975-2010). Localidades Melolonthidae Cetoniidae
géneros especies géneros Especies Villa de Allende, Estado de México. 1975 13 28 0 0 Calvillo, Aguascalientes. 1984 8 22 3 7 Jojutla, Morelos. 1987 17 56 6 15 Acahuizotla, Guerrero. 1989 27 72 6 16 Cuetzalan, Puebla. 2000 18 37 1 1 Teziutlán, Puebla. 2003 18 46 0 0 Zacatlán, Puebla. 2002 10 26 1 1 Tzucacab, Yucatán. 2003 10 22 7 11 Teotlalzingo, Puebla. 2005 6 17 0 0 Santiago Xiacui, Oaxaca. 2005 15 36 3 3 La Malinche, Tlaxcala. 2006 7 19 1 1 Tlachichuca, Puebla. 2006 9 20 1 1 Chignahuapan, Puebla. 2007 8 16 1 1 Chiautla, Puebla. 2008 11 39 4 9 Huehuetitla, Tlaxcala. 2010 8 14 0 0

17
Biología Subterránea: un mundo por descubrir Héctor Leonel Ayala Téllez
Departamento de Ciencias Ambientales, Centro Universitario de Ciencias Biológicas y Agropecuarias. Km. 15.5 Carretera Guadalajara-Nogales, Predio las Agujas, Nextipac,
Zapopan, Jalisco. [email protected]
La biología de los hábitats subterráneos (conocida como biología subterránea, espeleobiología, bioespeleología, o biología de cuevas) es un campo de investigación que incorpora todos los aspectos del estudio biológico de la biota de los hábitats subterráneos y/o el estudio de los hábitats subterráneos por sí mismos. La biología subterránea no solo estudia los animales y otros grupos bióticos, sino que toma en cuenta el ambiente en que se encuentran y como éste los afecta. Más allá de la entrada, donde hay un área de penumbra, la característica más obvia de las cuevas es su obscuridad total. Cierta similitud existe con la obscuridad que rodea a miles de especies que viven en los abismos de los océanos o en las bacterias y parásitos que viven dentro de los animales. Otra particularidad de las cuevas es que su clima es prácticamente uniforme, un factor importante para las especies que no pueden regular la temperatura de sus cuerpos. En estos ecosistemas, sobre todo en las partes más profundas, están representados algunos ejemplares de pocas especies, aunque con atributos que los hacen únicos. Los animales que habitan las cuevas exhiben diferentes grados de adaptación a este ambiente, debido ante todo, a las singulares condiciones de obscuridad, silencio, humedad relativa y temperatura, que convierten al sistema subterráneo en un entorno terrestre excepcional (Cano y Martínez 1999; Culver y Pipan, 2009). El clima subterráneo condiciona la estabilidad de las reacciones químicas y los procesos físicos que tienen lugar en el interior de las cuevas, controla el crecimiento y desarrollo de los espeleotemas y soporta la diversidad biológica subterránea; de todos estos elementos, la temperatura del aire subterráneo es quizá el más importante (Hoffmann et al., 1986; Cano y Martínez, 1999). El primer organismo de ambientes hipogeos en ser estudiado fue Proteus anguinus, un anfibio descrito por Laurenti en 1768. En el siglo XIX, naturalistas como J.C. Schiödte, A. Schmidl y E. Simon en Europa y T. Tellkrampf, F. W. Putnam y A. S. Packard Jr. en Norteamérica, se interesaron en la fauna habitante de las cuevas. Sin embargo, el comienzo de la investigación sistemática de la biología de los organismos cavernícolas data de principios del siglo XX, cuando E. G. Racovitza y R. Jeannel dieron realce a la bioespeleología en Europa. A mediados del siglo XX se lograron progresos notables en el conocimiento de la taxonomía, ecología y fisiología de la fauna de cuevas estudiadas en Europa y Norteamérica. Este conocimiento se integró de manera destacada en dos monografías: una en 1964, de A. Vandel, titulada "Bioespeleología" y otra de T. C. Barr, titulada "Ecología de las cavernas y evolución de los troglóbios", publicada en 1968. De ahí en adelante se emprendió la investigación en diferentes áreas que involucran el conocimiento de los organismos y su grado de evolución. En años recientes se han descubierto cuevas con gran riqueza faunística, con numerosos hábitats subterráneos que presentan complejos mecanismos de colonización (Hoffmann et al.,1986; Christiansen, 1995; Cano y Martínez, 1999). Entre las aportaciones más grandes que hacen las cuevas al mundo, se halla la posibilidad de que desempeñen un papel importante como salvaguardas de especies animales únicas, incrementando así la biodiversidad de las regiones del planeta donde éstas se encuentran. La diversidad de animales que habitan las cuevas, motivó a muchos biólogos a iniciar su investigación, es así como nació la bioespeleología (del griego bios, vida, spelaion, cueva, y logos, tratado), definida como la ciencia que estudia a los

18
organismos que viven en las cuevas. Dentro de las cuevas, se pueden encontrar especies de animales pertenecientes a diferentes grupos o Phyla; México ofrece mucho en el campo de la investigación subterránea, la particularidad faunística con más de 2100 especies, entre mamíferos, aves, reptiles, anfibios, peces, gasterópodos, insectos, crustáceos, arácnidos y muchas otras formas de vida, propias de las cavernas. Muchos investigadores de expediciones extranjeras y mexicanas, coinciden en que México tiene una de las faunas cavernícolas más rica y diversa que cualquier región del mundo (Reddell, 1981, 1982; Lazcano, 1983, Hoffmann et al., 1986); así, la Biología Subterránea o Bioespeleología para nuestra nación, se convierte en una disciplina científica de frontera, un mundo por descubrir.

19
Ponencias
Diversidad de corales blandos (Cnidaria: Octocorallia: Alcyonacea) de Bahía de Chamela
Victoria Belén Muñoz Jiménez1, Cristian Moisés Galván Villa2, Eduardo Ríos Jara3 y Carlos Armando Sánchez Ortiz4
1,2,3Centro Universitario de Ciencias Biológicas y Agropecuarias, Depto. de Ecología Aplicada, Laboratorio de Ecosistemas Marinos y Acuicultura, Camino Ramón Padilla Sánchez 2100, Nextipac, 44600, Zapopan, Jalisco, México, 4Universidad Autónoma de Baja California Sur, Depto. Biología Marina, Programa de Investigación para la Conservación de la Fauna Arrecifal, Carretera al Sur Km 4.5, 23080 La Paz, B.C.S.
[email protected], [email protected], [email protected], [email protected]
Las gorgonias o abanicos de mar son uno de los principales componentes de las comunidades bentónicas en arrecifes rocosos del Pacífico Oriental (Gamero, 2015). Se caracterizan por ser organismos sedentarios, coloniales y con pólipos que presentan ocho tentáculos pinnados y ocho septos en el mesenterio (Brusca y Brusca, 2003). En el Pacífico Americano se encuentra una gran diversidad de estos organismos, sin embargo muy poco se conoce sobre el número de especies, su distribucióny biología en general (Hernández, 2010, Muñoz et al., 2014). Se realizó un inventario de la diversidad de corales blandos (orden Alcyonacea) del Santuario Islas e Islotes de Bahía Chamela, localizado en el Pacífico Central Mexicano. Los registros se obtuvieron de un total de diez localidades dentro de la bahía y la colecta de organismos se realizó mediante buceo SCUBA entre los años 2007 y 2015. A la fecha se han determinado 11 especies nominales pertenecientes a cuatro géneros (Heterogorgia, Leptogorgia, Muricea, Pacifigorgia) y dos familias (Gorgoniidae y Plexauriidae). Las colonias que no se han podido determinar se encuentran clasificadas en tres grupos: Muricea sp. 1, Pacifigorgia sp. 1, Pacifigorgia sp. 2. El sitio con mayor riqueza fue Isla Cocinas con seis especies, seguido de Isla Pajarera con cinco especies. Comparando los resultados obtenidos hasta el momento con la literatura existente para el Pacífico Oriental Tropical se ha encontrado un número considerable de especies para la bahía; sin embargo, se espera que con futuros muestreos y mayor revisión de los ejemplares colectados aumente el número de especies y géneros. Especies de langostino (Palaemonidae: Macrobrachium) en el Arroyo Seco, Cihuatlán,
Jalisco Rodolfo Castro Barbosa1, Arnulfo H. Díaz1, Carlos Alberto Amezcua Gómez2 y Jesús
Eduardo Sainz Urzua1 1Centro Universitario de la Costa Sur, Departamento de Estudios Sustentables para el Desarrollo de Zonas Costeras, Licenciatura en Biología Marina, Gómez Farías # 82, C. P. 48980. San Patricio-Melaque, Cihuatlán, Jalisco. 2Instituto Nacional de la Pesca, Centro Regional de Investigaciones Pesqueras- Manzanillo, Playa Ventanas s/n, Col. Campos, C. P. 28860 Manzanillo, Colima.
[email protected], [email protected], [email protected], [email protected]
La familia Palaemonidae son algunos de los langostinos más abundantes y diversos, que habitan en el mar y agua dulce en aguas tropicales y templadas (Wicksten, 1983). El

20
género Macrobrachium es conocido por el abundante número de especies, y su amplia distribución geográfica, así como la importancia comercial (Holthuis, 1952; Villalobos, 1982). Se distinguen de otros géneros por los siguientes rasgos morfológicos; Caparazón con una tribuna proyectada, primer par de pereiópodos delgados y largos como el caparazón, el segundo par de quelas, a menudo es más largo que el cuerpo en los machos, tres pares posteriores de pereiópodos simples, telson triangular, que termina en una sóla punta (Bate, 1868; Holthuis, 1952; Mossolini & Bueno, 2003). También tienen espinas hepáticas y antenal y dos pares de espinas en la superficie dorsal del telson (Hedgpeth, 1949; Holthuis, 1952). Existen algunos trabajos tendientes a enumerar e identificar las especies nativas de ciertas regiones, tal es el caso de Holthuis (1952), el cual habla de 26 especies del genero Macrobrachium spp., de agua dulce para todo el continente Americano y de ellas 7 se localizan en la vertiente del Pacífico mexicano (Villalobos-Hiriart et al., 1993; Román et al., 2000; Wicksten & Hendrickx, 2003 Macrobrachium amazonicum, M. panamesis, M. heterochirus, M. carnicus M. americanum. M. tenellum, y M. acanthurus), de ellas sólo siete tienen importancia comercial y/o de cultivo (Holthuis, 1952). El objetivo de este estudio fue identificar el número de especies del género Macrobrachium en el Río Arroyo Seco, en Cihuatlán, Jalisco. Con el fin de generar un catálogo fotográfico para la identificación de las especies presentes. El trabajo en campo se realizó por medio de muestreos semanales por tres meses (época de secas), la captura de los organismos fueron por medio de “Chacaleras” y buceo libre, fueron medidos en Laboratorio Húmedo del DESZC con un ictiometro y se pesaron con una balanza Ohaus con .01 gr precisión. Se identificó el sexo por medio de la nomenclatura anatómica según de acuerdo McLaughlin (1980). En el estudio se encontró dos especies Macrobranchium americanum y Macrobranchium tenellum, de los cuales fueron un total de 15 organismos, 6 organismos fueron M. americanum y 9 M. tenellum, de lo cual del M. americanum fueron 4 machos y 2 hembras, y del M. tenellum los 9 fueron machos, la talla mínima del M. americanum del peso fue 12.5 (cm), y máxima 14.8 (cm), promedio es de 13.73 (cm), y del peso fue; máxima 21.75 (gr), mínima 18.09 (gr), y promedio fue de 19.79 (gr), de tal manera se hizo el mismos procedimiento a la otra especie la cual fue M. tenellum, sus tallas fueron: talla máxima 12.2 (cm), mínima 9.2 (cm), promedio 11.11 (cm), y de peso fueron; la máxima 16.24 (gr), mínima 12.66 (gr), promedio 15.19 (gr). Cuando se hicieron los primeros muestreos fueron bajas las capturas, ya que en la zona más alta del río Arroyo Seco capturamos algunos organismos con mayor frecuencia, mas sin embargo se observó que el M. americanum fue más abundante en zona más alta, y del M. tenellum, fueron en zonas de fosas (charcos), donde la visibilidad del agua era muy diferente al otro sitio (zona alta) e incluso la temperatura, En el primer punto de muestreo capturamos el M. americanum el agua muy transparente y fondo arenoso, con muy poco fluido de agua, con profundidades máxima de 1.5 mts, y como característica algunas rocas donde se refugiaban los organismos, y de la especie de M. tenellum sus condiciones de hábitat es sustrato lodoso, y su captura fue en fosas (charcos) y con muy baja visibilidad. En la región se reconocen otras especies pero que sólo se suelen capturar en temporada de lluvias, M. occidentale, M. digueti, M.olfesii, M. hobbsi y M. michoacanus; pero entre las siete especies que se consideran para la cuenca del Pacífico (Villalobos-Hiriart et al., 1993; Romaán et al., 2000; Wicksten & Hendrickx, 2003) se ha propuesto un rearreglo taxonómico en dos subgéneros Macrobrachium y Allobrachium (Jayachandran, 2001; Hernandez et al., 2007).

21
Distribución y abundancia del langostino crecedor Macrobrachium americanum (Invertebrata: Decapoda) en el Río Marabasco, Jalisco-Colima
Jesús Eduardo Sainz Urzúa1, Arnulfo H. Díaz1, Carlos Alberto Amezcua Gómez2 y Rodolfo Castro Barbosa1
1Centro Universitario de la Costa Sur, Departamento de Estudios Sustentables para el Desarrollo de Zonas Costeras, Licenciatura en Biología Marina, Gómez Farías # 82, C.P. 48980. San Patricio-Melaque, Cihuatlán, Jalisco. 2Instituto Nacional de la Pesca, Centro Regional de Investigaciones Pesqueras- Manzanillo, Playa Ventanas s/n, Col. Campos, C.P. 28860 Manzanillo, Colima.
[email protected], [email protected], [email protected], [email protected]
El langostino Macrobrachium americanum es una especie con un gran potencial para cultivo comercial debido a su tamaño, mercado y valor económico. (Holthuis, 1952) señala que M. americanum es uno de los langostinos más grandes del género, llegando a medir hasta 25 cm en machos, 19 cm en hembras. El ciclo biológico es relativamente corto, de hábitos nocturnos y de alimentación omnívora, es depredador activo de otros crustáceos y larvas de insectos acuáticos. M. americanum es apreciado por su rápido crecimiento, su alto contenido proteico y exquisito sabor. La distribución de los organismos pertenecientes a la familia Palemonidae del género Macrobrachium, se extiende desde Baja California hasta Perú e Islas Galápagos (Holthuis 1952). Existen 29 especies del género Macrobrachium en América, de los cuales siete tienen importancia comercial y de cultivo: M. americanum M. tenellum M. carcinus M. acanthurus M. panamense M. heterochirus y M. amzonicum. En nuestro país existen cuatro especies de importancia comercial las cuales son M. acanthurus M. carcinus para el golfo de México y el Caribe, M. americanum y M. tenellum para el Pacífico. Estas se encuentran dentro de los climas tropicales y semitropicales con una temperatura ambiental media anual de 16 °C como una mínima y de 32 °C como una máxima. El objetivo de este estudio es determinar la distribución y abundancia del langostino Macrobrachium americanum en el Río de Marabasco, Jalisco-Colima, México. El área de estudio se realizó en el Río Marabasco, en el límite geográfico entre los estados de Jalisco y Colima, Los muestreos fueron nocturnos entre 10 pm a 2 am y comenzaron en la parte baja del río hacia la parte alta de la sierra, como arte de pesca se utilizó una atarraya de 1.5 m y una luz de malla de 1.5 cm, se utilizó pulpa de coco como alimento para ayudar a la captura ya que es muy conocido la preferencia de estos organismos por ese alimento, se tomó la temperatura del agua para saber su ventana óptima de temperatura de estos, se auxilió de una lámpara para poder ver a los langostinos durante la noche y guiarnos. Los muestreos fueron semanales y se realizaron durante cinco semanas, empezando desde el 21 de abril de 2014 y finalizando el 19 de mayo de 2014. Se muestrearon cinco puntos con una diferencia de 500 m aproximadamente, los organismos capturados fueron transportados al Departamento de Estudios Sustentables de Zonas Costeras, fueron medidos con un ictiómetro desde la parte del telson hasta el rostro, fueron pesados con una balanza de capacidad de 600 g y fotografiados. El sexo se determinó de acuerdo a características propuestas por Ling et al. (1976), ya que el macho posee quelas más largas y gruesas que en la hembra. En el color del macho es de café rojizo y el exoesqueleto es poco brillante; en la hembra es café grisáceo y casi opaco. Como resultado se capturaron 50 organismos donde se encontró una mayor abundancia de machos que hembras, con un total de 36 machos y 14 hembras, la talla promedio de los machos fue de 17. 4 cm y el peso promedio de 143.36 gr, la talla promedio de las hembras fue de 14.29 cm y un peso promedio de 52.79 gr, en el primer

22
muestreo se capturaron 8 organismos machos y 2 hembras, en el segundo muestreo se capturaron 9 machos y 3 hembras, en el tercer muestreo se capturaron 6 machos y 3 hembras, en el cuarto muestreo sólo se capturaron 6 organismos machos y en el quinto muestreo se capturaron 7 machos y 6 hembras. La temperatura osciló entre los 28 y 29 °C. Todos los organismos se capturaron en un tipo de sustrato roco y arenoso. De acuerdo a los resultados se encontró que en épocas de secas podemos encontrar más organismos machos que hembras, el organismo más grande fue un macho con una talla de 22 cm con un peso de 264.7 gr y el de menor tamaño fue una hembra de 12.2 cm con un peso de 48.6 gr. A manera de conclusión en época de secas se pueden encontrar pocos organismos debido a que no es época de reproducción, de tal manera que es menos probable poder capturarlos ya que se encuentran encuevados, en piedras, raíces o troncos de árboles, etc. En época de secas la forma más conveniente de poder capturarlos es por la noche, siempre y cuando no haya luna nueva para poder verlos mejor. Isópodos marinos (Crustacea: Isopoda) de Bahía de Chamela, Jalisco: Listado faunístico
y nuevos registros Diego Alfredo Gutiérrez Anzaldo1, Manuel Ayón Parente2, Eduardo Ríos Jara3 y
Cristian Moisés Galván Villa4. 1,2Centro Universitario de Ciencias Biológicas y Agropecuarias, Km15.5 Carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco.
[email protected], [email protected], [email protected], [email protected]
En la actualidad existen grupos de crustáceos poco conocidos que tienen un papel muy importante en la conservación del equilibrio ecológico en el sistema marino (Barnes, 1986). La mayoría de estos (~50%) lo conforman especies del superorden Peracarida, el cual incluye al orden Isopoda. Actualmente los isópodos están representados por alrededor de 10, 297 especies, de las cuales poco más de 9500 son de hábitats acuáticos (Boyko et al., 2008; Espinosa-Pérez et al., 2009). Los isópodos marinos tienen gran importancia ecológica ya que actúan como reductores de materia orgánica compactada en materia particulada, papel importante en el flujo energético del ecosistema marino (Schultz, 1969). También son de importancia económica, ya que algunas especies del género Limnoria tienen hábitos excavadores de estructuras de madera (muelles, embarcaciones, etc) utilizadas por el hombre, otras más (e.g., Bopyridea, Cymothodea) son parásitos de peces y crustáceos de interés comercial (Markham, 1985). Ocupan una diversidad de hábitats, van desde las zonas intermareales hasta mar profundo; habitan entre otros sustratos, raíces de mangle (Ellison y Farnsworth, 1990), fondos blandos (Taylor y Moore, 1995), pastos marinos (Kang y Yun, 1988), comunidades de macroalgas (Arrontes y Anadón, 1990), corales (Delaney, 1984), rocas o sustratos arenosos (Dexter, 1976). Dada la importancia ya mencionada y la carencia de información de este grupo para las costas de Jalisco, el presente estudio tiene como objetivo elaborar un listado faunístico de los isópodos de Bahía Chamela, Jalisco. Los especímenes fueron obtenidos en muestreos realizados durante 2013 y 2014 en el intermareal y submareal somero de Bahía Chamela. Las colectas se efectuaron mediante buceo, snorkel y utilizando una pequeña draga biológica. En total se recolectaron 225 organismos correspondientes a 18 especies. Los Cymothoidea con 10 especies y seis géneros fueron los mejor representados, seguidos por Sphaeromatidea con tres 3 géneros y tres especies, el resto de las familias contó con un género y una especie. El

23
88% de las especies representan nuevos registros para la Bahía, sólo Ligia occidentalis y Tylos punctatus habían sido registradas para la localidad. Tres de las especies colectadas son parásitas de peces (Cymothoa exigua, Rocinela angustata y R. sigmata), el resto son de vida libre. Para cada especie se presentan mapas de distribución.
Situación actual de las poblaciones de Octopus hubbsorum (Molusca: Octopoda) y algunas alternativas de manejo del recurso en la Costa de Jalisco
Carlos Alberto Amezcua Gómez1,2, Rodolfo Castro Barbosa2 y Jesús Eduardo Sainz Urzua2
1Instituto Nacional de la Pesca, Centro Regional de Investigaciones Pesqueras- Manzanillo, Playa Ventanas s/n, Col. Campos, C.P. 28860 Manzanillo, Colima. 2Centro Universitario de la Costa Sur, Departamento de Estudios Sustentables para el Desarrollo de Zonas Costeras, Licenciatura en Biología Marina, Gómez Farías # 82, C.P. 48980. San Patricio-Melaque, Cihuatlán, Jalisco. En México puntualmente en el Golfo de México se tiene un manejo de la pesquería del recurso pulpo, de tal manera que parte importante de este manejo es una veda que ayuda a la recuperación de los stock de este recurso y la operación de trampas cuyas capturas son selectivas y de acuerdo a la Normas establecidas, al contrario del Océano Pacífico el recurso pulpo se mantiene sin ningún manejo solo las recomendaciones que emite la Carta Nacional Pesquera, a lo largo de la Costa de Pacífico el pulpo se captura por medio de gancho y ayudado con equipo tipo hooka, el gancho no es selectivo, es agresivo y pueden estar susceptibles a la captura hembras ovígeras y juveniles, además que la baja calidad de producto capturado por el daño con el gancho, el objetivo de este trabajo es conocer el estatus en las poblaciones de pulpo en la Costa de Jalisco en el año 2014. Para conocer la situación actual de la población del pulpo se realizaron muestreos mensuales en 4 cooperativas dedicadas a la captura de pulpo, un día por cada cooperativa, ahí; se midieron los organismos con un ictiometro, las medidas que se obtuvieron fueron longitud de manto y longitud total, así mismo se pesaron con una báscula Ohaus con capacidad de 3 kilos y precisión .01 g , a los organismos se sexaron revisando el tercer brazo para ver si tiene el hectocotilo para los machos y la ausencia de este para las hembras, así mismo se hizo un reconocimiento del estado de madurez de manera visual. Los organismos fueron capturados con la colaboración de los buzos de cada región debido a que estos cuentan con mayor experiencia en la pesca del pulpo, fue por medio de buceo con gancho, en las inmersiones se utilizó equipo semiautónomo, el cual consiste en aletas, visor, cinturón de plomo, computadora de buceo, regulador de aire y boquillas así como un compresor a gasolina tipo “hooka” de 2 mangueras (100 m de largo) y una bolsa de colecta de malla de seda, conocida como “chango”. Las localidades muestreadas fueron San Patricio-Melaque, La Manzanilla, Chamela, Punta Perula. El presente estudio se inició a partir del mes de enero del 2014, en su totalidad se muestreo un total de 986 organismos, la abundancia del recurso empezó a partir de abril hasta el mes de julio, esta fluctuación se considera anómala ya que en otros años los meses de mayo hasta agosto esta anomalía se presume a que fue debido a la presencia de “El Niño”, de acuerdo a la bitácoras de los pescadores a partir del mes de agosto ya no hubo organismos por lo que los usuarios optaron por ir a pescar recurso escama o dorado. La estructura de tallas para el año 2014 se presentó: Longitud total entre 27 a 99 cm con una moda de 48 cm y talla promedio de 51.8 cm, la estructura por tallas (longitud del manto, LDM) del pulpo en base a la medición de los individuos, se presentó entre 6.0 y 19.9.0 cm, con una moda de 10.0 mm y talla

24
promedio de 10.6 cm, la estructura por peso del pulpo, se presentó entre 103 g a 1,992 g con un peso promedio de 503.56 g y con una moda de 390 g, la cual acumula el 65% de la población estudiada. De acuerdo a López-Uriate, 2006 para esta especie, las hembras son aptas para reproducirse a partir de los 758 g, por lo que la captura se está centrando a organismos juveniles. También se muestra que los meses de Mayo y Junio encontramos los organismos con mayor peso, por lo que es de conjeturar que estos organismos pertenecen a un stock diferente ya que a partir del mes de agosto hasta finales de año fue muy poca la captura de organismos, durante el periodo de estudio se observó una proporción sexual de 1.14 hembras por 1 macho. A lo largo de todo el año la captura solo se incrementó en el mes de julio y los meses donde regularmente no había captura (enero, febrero) si la hubo, es de resaltar que el mes donde inicia la temporada (mayo) se presentó una captura muy baja y ese misma observación se mantiene en todos los meses a excepción de julio. Como recomendaciones es necesario que se regule la captura de los organismos que aún no están en edad de madurez, es necesario que se prohíba la captura, venta y se respete a las hembras que están encuevadas ya que al sacarlas del nidos los huevos están a expensas de hongos o depredadores, por su potencial de crecimiento es necesario que se proyecten alternativas de cultivo para pulpo en México. Por parte del INAPESCA se está proyectando la engorda de juveniles en maricultivo y a su vez promoviendo nuevos métodos de captura que sean más sustentables para el recurso pulpo.
Estado del conocimiento de la Manta birostris (Batoidea: Mobulidae): Una revisión Miguel de Jesús Gómez García1, Daniel Enrique Godínez Siordia2 y Carlos Alberto
Amezcua Gómez3 1,2 Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras; Centro Universitario de la Costa Sur, Universidad de Guadalajara. Gómez Farías N° 82, San Patricio-Melaque, Jalisco, C. P. 48980. 3Instituto Nacional de la Pesca, Carretera a Campos, Playa Ventanas S/N, Manzanillo, Colima, C. P. 28860.
[email protected], [email protected], [email protected]
La mantarraya o manta gigante Manta birostris es un elasmobranquio del orden de los myliobatiformes, sub familia mobulinae, puede tener una longitud de los 6.7 a los 9 m y un peso de hasta 2,400 kg (Bigelow y Schroeder, 1953; Last y Stevens, 1994). Se encuentra en mares tropicales y sub tropicales de todo el mundo, alimentándose mediante filtración, principalmente del zooplancton, aunque también consume pequeños crustáceos y peces (FAO, 2000; Osada, 2010). Su amplia distribución y hábitos migratorios han hecho difícil su estudio (Marshall, 2008; Couturier, 2012). Se reproducen de manera vivípara aplacentaria (Tomita et al., 2012) teniendo de una a dos crías cada 3 años, su carácter sociable y baja fecundidad las ponen en una posición vulnerable para su conservación (Marshal et al., 2011). Este organismo se encuentra clasificado en la lista roja de la IUCN (International Union for Conservation of Nature) como especie bajo amenaza (Marshall et al., 2006). A pesar que en algunos países han establecido normas que prohíben su pesca (CITES, 2013) las mantarrayas continúan siendo amenazadas ya que sus branquias deshidratadas se emplean en la medicina tradicional, llegando a alcanzar un precio por libra de $140 dólares (Dewar, 2002). El estudio de las mantarrayas se ha visto favorecido en años recientes debido a su gran importancia turística, llegando a generar hasta $8 millones de dólares por año en algunas zonas. (Anderson et al., 2010.; Osada, 2010). La historia evolutiva de la manta

25
gigante es prácticamente desconocida, los datos más antiguos provienen de fósiles a inicios del período pleistoceno encontrados en Norte América (Snyder et al., 1983; Bourdon 1999), mientras que los estudios más recientes de filogenética son los realizados por Kashiwagi et al. (2012). El género manta se dividió recientemente en dos especies, la manta gigante M. birostris y la de arrecife M. alfredi (Marshall et al., 2009) con lo que se ha descubierto que la mayor parte de la información obtenida hasta el momento pertenece a M. alfredi, más asociada con arrecifes y de distribución mucho menor (Couturier et al., 2012) estando ausente en el Este del Pacífico y el Oeste del Atlántico, sin embargo existe un número creciente de estudios acerca de M. birostris, algunos con marcas satelitales (Graham et al., 2012) con el fin de elucidar su distribución. Ha sido reportada con frecuencia en las islas Revillagigedo a las afueras de Jalisco (Marshall et al., 2009; Kashawagi et al., 2011), y fue con datos de esta misma zona que Hinojosa-Alvares et al. (2014) describieron el genoma mitocondrial de la especie. Ari y Correia (2008) estudiaron en el Acuario de Lisboa en Portugal, el comportamiento alimenticio de un ejemplar hembra durante tres años, y descubrieron que las mantas gigantes buscan su alimento usando tanto el olfato como la vista, mientras que Ari (2011) examinó su morfología cerebral, descubriendo que M. birostris posee el cerebro más grande en relación al cuerpo de todos los peces, aunque no se sabe si esta característica se deba a su estilo de vida activo, sus hábitos migratorios (memoria), o a su desarrollado sentido de la vista y olfato. En los últimos años han surgido una serie de estudios acerca de la ocurrencia de esta especie (Mejía-Fala et al., 2014; Couturier et al., 2015; DeBoer et al., 2015; Girondot et al., 2015; Medeiros et al., 2015) La mayoría de estos se basan en los patrones de pigmentación únicos de la especie como medio para su identificación, existiendo bases de datos internacionales utilizadas con este fin (Marshal et al., 2006; Deakos et al., 2011) sin embargo estudios recientes en especímenes en cautiverio sugieren que estos patrones pueden no ser tan confiables, ya que existen cambios a corto (Ari, 2014) y largo plazo en las tonalidades y patrones de la piel de los organismos (Ari, 2015). Descubrimientos como este resaltan la importancia de dar continuidad a los estudios acerca de esta especie, ya que siguen sin conocerse aspectos biológicos y ecológicos básicos, como su distribución, periodos reproductivos e incluso hábitos migratorios. Al igual que con muchas especies de carácter pelágico importantes para el ecosistema, aun hace falta mucho por aprender si se quiere tener un buen manejo y conservación de la especie. Foto identificación de la manta gigante Manta birostris (Vertebrata: Elasmobranchii) en
Bahía de Banderas, Jalisco-Nayarit Ramiro I. Gallardo-Hernández1, Adrián Maldonado-Gasca2, Javier Tovar-Ávila3 y
Joshua Stewart4 1, 2 Instituto Tecnológico de Bahía de Banderas. Crucero a Punta de Mita S/N. Bahía de Banderas, Nayarit. C.P. 63734 México. 3Centro Regional de Investigación Pesquera, Tortuga No 1, Bahía de Banderas, Nayarit. C. P. 24280, México. 4 Scripps Institution of Oceanography-US San Diego, La Jolla, California, C. P. 92093. USA.
[email protected], [email protected], [email protected], [email protected]. Correspondencia: [email protected]
La manta gigante (Manta birostris) es uno de los animales de mayor tamaño en el océano, está en la Lista Roja de la IUCN como especie Vulnerable a nivel mundial, con tendencias que apuntan a la reducción de sus poblaciones y en México se encuentra protegida en la NOM-029-PESC-2006. Con el objetivo de estimar la abundancia,

26
patrones de coloración, proporción sexual y distribución espacio-temporal de las mantas oceánicas en la Bahía de Banderas, Jalisco-Nayarit, se llevaron a cabo muestreos de campo entre Julio de 2013 y Noviembre de 2014. Se tomaron fotografías utilizando una cámara GoPro 3+ Black y equipo de buceo libre. Las fotografías de cada individuo fueron digitalizadas y analizadas con el programa I3S-Spot. Se registraron 35 individuos, de los cuales 24 presentan una coloración chevron y 11 una coloración melánica (negras en la región dorsal). Del total de las mantas identificadas, 15 fueron machos, nueve hembras y a 11 no se pudo identificar el sexo. Cuatro mantas presentaron algún daño corporal (11.4%), como: mutilación de uno de los lóbulos cefálicos, mutilación de la punta en la aleta pectoral, y heridas a causa de artes de pesca. La región donde hubo mayor incidencia fue en Yelapa y los meses con mayor número de registros fueron septiembre 2013, agosto y noviembre de 2014. Se considera que la zona sur de Bahía de Banderas es un área de alimentación de las mantas, por la alta productividad primaria y de zooplancton generada por las surgencias estacionales. Asi mismo se han observado sitios que pueden ser utilizados por las mantas como estaciones de limpieza y posiblemente también sea una zona de reproducción.
Condición poblacional y parasitosis aproximada del Tiro Goodea atripinnis (Goodeidae: Actinopterigy) en la Laguna de Cajititlán, Jalisco, durante la alta
mortandad de peces (2014-2015) Agustín Camacho Rodríguez1, Eduardo Juárez Carrillo2, Martha Alicia Lara González2
y Manuel Guzmán Arroyo3 1Centro Universitario de Tonalá. Av. Nuevo Periférico No. 555, Ejido San José Tatepozco, 48525 Tonalá, Jal. 2Laboratorio de Ecosistemas Marinos y Acuicultura, CA-UDEG 582 Hidrobiología, Centro Universitario de Ciencias Biológicas y Agropecuarias. Km 15.5 Caretera Guadalajara-Nogales, Predio Las Agujas, Nextipac,Zapopan, Jalisco. 3Instituto de Limnologia, Universidad de Guadalajara, A.P. 310, 045900, Ajijic, Jalisco.
[email protected], [email protected], [email protected]
La laguna de Cajititlán es un cuerpo de agua natural de agua dulce que se encuentra en una cuenca endorreica en el Municipio de Cajititlán aproximadamente a 30 Km de la Ciudad de Guadalajara, localizada en las coordenadas geográficas 20º 24´20” y 20º 24´50” latitud norte y 103º 16´50” 103º 23´32” longitud oeste (Cruz-Solís, M.E., 1993). Es una laguna eutrófica, monomictica tropical con una profundidad media de 2.5 m y una profundidad máxima de 5.00 m es un cuerpo de agua semicerrado debido a que se alimenta por un canal artificial (canal de Cedros) y el agua, actualmente sale por rebosamiento por el antiguo canal que alimenta la presa de la Calera-Puertas de Juanacatlán. Tiene una superficie aproximada de 2000 ha. Desde el mes de Agosto del año 2013 al de 2015 se han registrado altas mortandades de peces, hasta 365 toneladas, en el vaso lacustre. Si bien todas las especies presentes en la laguna son afectadas la que mayor mortandad presenta es Goodea atripinnis Jordan (Tiros). Goodea atripinnis se distribuye en la vertiente del Pacífico, cuenca de los ríos Lerma-Santiago y partes antiguas (pleistocénicas) de este sistema. En las cuencas de los ríos Balsas, Armería y tributarios independientes en el Pacífico de Nayarit, aguas adyacentes en Jalisco y Michoacán. Vertiente del Atlántico, ríos Santa María, San Juan del Río y afluentes del alto río Pánuco en Querétaro y San Luis Potosí. Lo encontramos en lagos, estanques, manantiales y efluentes y arroyos. Agua clara, turbia o lodosa; vegetación rara vez rala,

27
típicamente algas verdes, además de chara jacinto de agua, Potamogeton, Lemna y Nasturtium; corriente nula, a veces moderadamente fuerte; sustrato de lodo, arcilla, arena, grava y rocas. El alimento consiste en algas verdes filamentosas, microcrustáceos y moluscos. Como lo indica su largo intestino convoluto, abundantes branquia espinas y dientes mandibulares externos bífidos, se alimenta por ramoneo y filtración. La presencia en la laguna de Cajititlán de juveniles entre enero y julio indica una temporada reproductiva larga. Esta especie generalmente nada entre la mitad de la columna de agua y el fondo, alimentándose durante el día del perifiton. Forma cardúmenes de peces casi inmóviles justo por encima del fondo. Longitud máxima es de 200 milímetros (Miller, 2009). A fin de conocer la condición poblacional Goodea atripinnis se han realizado muestreos desde el mes de septiembre de 2014 al de Agosto de 2015, utilizando para la captura de los peces una red atarraya de 3m de radio con una bolsa conformada por tirantes de 30 cm y luz de maya de 2.5 cm, operada de manera individual desde una lancha de 18 pies de eslora con motor fuera de borda. Los sitios de muestreo se han elegido atendiendo al criterio de accesibilidad y mejor representatividad de las condiciones hidrológicas promedio de la laguna, estos sitios se han denominado: Cajititlán, Cuexcomatitlán, San Juan Evangelista, San Lucas y Centro de la laguna. Se han capturado 3,436 peces de los cuales 3,150 son Goodea atripinnis, 134 Chapalichthyes encaustus, 41 Poecilia mexicana, cuatro Menidia sp., dos Ameca splendens, y 105 Oreochromis niloticus. Se han analizado por disección a 62 especímenes de la especie Goodea atripinis y sin importar el sexo la incidencia parasitaria es de 42%, esto es 29 de ellos poseen alguna especie de parasito, destaca por su tamaño el llamado gran gusano rojo. Eustrongylides tubifex, que afecta principalmente la cavidad célomica en donde se le observa en forma de adultos enquistados principalmente en el mesenterio. Se han observado numerosos quistes de trematodos alojados principalmente en hígado, éste órgano se manifiesta en la totalidad de los peces analizados con un evidente daño en forma de manchas de colores pálidos, petequias rojizas, una condición deleznable y muy fácilmente rompible, su anatomía es anormal manifiesta en la irregularidad de su superficie y en su irregularidad anatómica entre los individuos. La proporción sexual para Goodea atripinnis en la Laguna de Cajititlan guarda una relación de 0.4838 (machos/hembras) o de 1.066 hembras por cada macho. La talla promedio de una muestra compuesta por 31 ejemplares capturados el 6 de Octubre de 2014 es 31.64 mm con un error típico de de 0.793 mm. Poliaminas intestinales en tilapia (Oreochromis niloticus) y bagre (Ictalurus punctatus)
Ramón Guillermo Ortiz-García1, Anne Santerre2, María del Rosario Huizar-López3, Martha Cecilia Téllez-Bañuelos4, Josefina Casas-Solís5, Patricia Castro-Félix6 y
Eduardo Juárez-Carrillo7 Departamento de Biología Celular y Molecular y Departamento de Ecología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Jalisco, México.
[email protected], [email protected], [email protected], [email protected], [email protected],
[email protected], [email protected] La acuicultura ofrece una alternativa alimenticia de gran valor nutricional y representa el sector de producción de alimento de mayor crecimiento, con una expansión a nivel mundial de 5% a 10% anual en los últimos 10 años. La tilapia (Oreochromis sp.) y el bagre (Ictalurus sp.) son las especies más populares en la acuicultura de peces de agua

28
dulce de México. Actualmente, la alimentación de los organismos en el sector acuicultura depende de dietas formuladas con alimentos industriales y representa del 50 al 60 % del costo de producción de las especies acuícolas (Watanabe, 2002). Una producción acuícola sustentable debe considerar un cambio en la formulación del alimento comercial de los peces de granja a través del reemplazo, parcial o total, de los aceites de pescado por aceites de origen vegetal. Una elección equivocada del tipo y proporción de aceite, podría producir una baja ganancia de peso, prolongar el tiempo para llegar a una talla comercial y afectar el rendimiento de los sistemas de producción intensiva (García & Villarroel, 2009). Los objetivos de este trabajo fueron investigar el efecto de cuatro dietas sobre los niveles de poliaminas en intestino y el crecimiento de tilapia del Nilo (Oreochromis niloticus Linnaeus, 1758) y bagre del canal (Ictalurus punctatus Rafinesque, 1818) juveniles. La dieta control (C) contiene 2% de aceite de pescado y 6% de aceite de soya. La dieta alternativa (S) está formulada con 8% de aceite de soya y no contiene aceite de pescado. Las dietas CP y SP corresponden a las dietas C y S, suplementadas con un probiótico formulado para el sector acuícola. Las poliaminas biogénicas: putrescina (Pu), espermidina (Spd) y espermina (Spm) fueron evaluadas por HPLC durante un periodo experimental de 120 días. Los niveles promedios de las poliaminas en intestino de tilapia fueron Pu 0.0194 ± 0.0048, Spd 0.0949 ± 0.0266, Spm 0.0561 ± 0.0196 nmol µL-1 y en bagre Pu 0.0144 ± 0.0025; Spd 0.1054 ± 0.0142, Spm 0.1036 ± 0.0143 nmol µL-1. Los datos experimentales indican que las dietas S, CP y SP no afectaron el crecimiento y los niveles de poliaminas de los peces en comparación con la dieta C, sin embargo, se observaron cambios en las poliaminas intestinales a través del tiempo de experimentación. Los resultados sugieren que es factible el reemplazo total del aceite de pescado por el aceite de soya en la formulación de dietas prácticas en tilapia y bagre. Estos datos complementan los de otros autores que proponen sustituir un porcentaje del aceite de pescado por aceites de origen vegetal, como se realizó en el salmón del Atlántico (Salmo salar) donde el reemplazo del 50 y 80% del aceite de pescado por diferentes aceites vegetales no comprometió el crecimiento, la sobrevivencia y la utilización alimenticia (Rosenlund et al., 2001; Caballero et al., 2002), o en la especie marina Sciaenops ocellatus donde la sustitución de 70 al 80% de aceites de pescado por aceites vegetales no redujo el crecimiento (Lochmann & Gatlin, 1993; Tucker et al., 1997).
Composición del ensamble de aves (Chordata: Aves) en cuatro sitios del Área de Protección de Flora y Fauna Maderas del Carmen, Coahuila, México
Nadia Martínez Gallegos1, Juan Antonio García Salas2 y Sergio Guerrero Vázquez3
1,3Centro Universitario de Ciencias Biológicas y Agropecuarias, Camino Ing. Ramón Padilla Sánchez #2100, Las Agujas, Nextipac, Zapopan, Jalisco, CUCBA, Universidad de Guadalajara. 2Laboratorio de Ornitología, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Ciudad Universitaria, C. P. 66451, San Nicolás de los Garza, Nuevo León.
[email protected] El Área de Protección de Flora y Fauna Maderas del Carmen (APFFMC) fue decretada el 7 de noviembre de 1994; es considerada un importante corredor biológico entre el sur de Estados Unidos y noreste de México (CONANP, 2008), también es relevante como área de anidamiento y hábitat durante la migración de la avifauna (CONANP & SEMARNAT, 2008). Se ubica en el Desierto Chihuahuense en el extremo noroeste del Estado de Coahuila; situándose dentro de tres municipios: Múzquiz, Ocampo y Acuña.

29
Cubre una superficie total de 208,381 ha; con un rango altitudinal desde los 500 msnm a la orilla del río Bravo hasta los 2,720 msnm en los picos más altos. El presente trabajo tiene como objetivo evaluar la estructura del ensamble de aves en cuatro diferentes sitios (El Jardín, Jaboncillos, Pilares y San Francisco) dentro del APFFMC, cada uno definido por tener una asociación vegetal predominante y diferente de los otros tres. Para implementar el monitoreo se realizaron 14 visitas al área durante los años 2012 y 2013, utilizando el método de transectos en línea con puntos de conteo (Ralph et al., 1996; Bibby et al., 1992); cada transecto tenía 10 puntos de conteo y una longitud total de 3000 metros. El muestreo iniciaba al amanecer; en cada punto se identificaron y registraron todos los individuos captados visual y auditivamente en un lapso estricto de 10 minutos. Se registraron un total de 193 especies; el sitio con mayor riqueza fue San Francisco (143), seguido de Pilares (135), Jaboncillos (85) y El Jardín (53). La mayor abundancia se registró en San Francisco con 1961 individuos, mientras que la menor corresponde a El Jardín, con 356. El Orden mejor representado fue Passeriformes con 114 especies y 4142 individuos; dentro del cual, la Familia con mayor número de especies fue Emberizidae. Con respecto a la permanencia estacional y temporal (Howell & Webb, 1995; Garza de León et al., 2007), se registraron 66 especies residentes, 64 invernales, 25 transitorias, 24 veraniegas, 9 casuales y 2 accidentales; las aves invernales, residentes, transitorias y veraniegas se observaron normalmente en los cuatro sitios, mientras que las accidentales y casuales solo se pudieron observar en tres de ellos. Se identificaron 10 gremios alimenticios (Ehrlich, Dobkin & Wheye, 1988); los insectívoros fueron los más abundantes con 121 especies y distribuyéndose en los cuatro sitios, seguidos de los granívoros con 22 y los carnívoros con 13; el gremio menos representado fue el de ictiófagos con 1 sola especie, la cuál fue observada en un solo sitio. Tomando en consideración los criterios establecidos por la NOM-059-SEMARNAT- 2010, se encontraron al menos 12 especies con alguna categoría de riesgo, entre ellas: Aquila chrysaetos (amenazada), Cyrtonyx montezumae (protegida) y Falco mexicanus (amenazada); por su parte, la IUCN cataloga a Colinus virginianus, Passerina ciris y Vireo bellii como especies casi amenazadas (NT) , mientras que a Vireo atricapilla como especie vulnerable (VU). Aunque la avifauna del APFFMC ha sido estudiada anteriormente en listados (CONANP, 2010; Panjabi, Youngberg & Levandoski, 2010; Cantú de Leija, 2013; Castillo- Muñóz, 2014), planes de manejo del área (CONANP, 2010; SEMARNAT & CONANP, 2013) y estudios sobre el efecto de la variabilidad climática en la avifauna (Castillo- Muñóz, 2014); no se ha profundizado en cuanto a su variación en relación con los diferentes hábitat. Es por lo anterior que se realizó el trabajo anterior, para contribuir en el conocimiento de la composición del ensamble de aves en esta zona, así como evaluar sus cambios en tiempo y en espacio.
Alimentación del bobo café, Sula leucogaster (Aves: Suliformes) de una colonia de anidación en la isla Peña blanca, Colima
Luis Abraham Aguilar Nuño1, Salvador Hernández Vazquez2, Humberto Almanza Rodríguez3, Ana María Almaguer Hernández, José Angel Hinojosa Larios, Bernabé
Aguilar Palomino, José Manuel Bojórquez Castro y Miriam Daniela Rodríguez Torres Universidad de Guadalajara, Centro Universitario de la Costas Sur. Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras. Gómez Farías #82, San Patricio Melaque, Cihuatlán, C.P. 48980, Jalisco.

30
Las aves marinas son un grupo muy diverso de especies adaptadas para explotar eficientemente los recursos marinos, entre los que figuran como principales se encuentran los peces pelágicos, crustáceos y calamares (Furness y Monaghan, 1987; Branco et al., 2005). El estudio del papel de las aves en el flujo de energía en los ecosistemas marinos así como la rapidez de respuesta a los cambios en la disponibilidad de las presas ha demostrado su importancia como depredadores tope en la cadena, por lo tanto este grupo puede ser empleado como indicador para el monitoreo de los cambios en la diversidad de los ecosistemas (Bost y Maho, 1993; Montevecchi y Myers, 1995, 1996; Ceyca y Mellink, 2009). El bobo café (Sula leucogaster) es una especie común en aguas tropicales de todo el mundo (Schreiber y Norton, 2002; Ceyca y Mellink, 2009), en el Pacifico mexicano es una especie ampliamente distribuida y numerosa, se han registrado colonias desde el norte del golfo de California hasta Guerrero (Anderson, 1983; Mellink y Riojas-López, 2005; Ceyca y Mellink, 2009). S. leucogaster se reproduce en colonias las cuales pueden ser de tamaños y densidades variables, ambos padres participan en la construcción del nido, incubación y alimentación de los pollos, (Dorward, 1962; Nelson, 1978, Simmons, 1967; de Campos y Martins, 2003), esta especie presenta dimorfismo sexual en la coloración del pico, patas y la mancha ocular, además de que las hembras son más grandes que los machos (Sick, 1997; de Campos y Martins, 2003). Los estudios sobre alimentación en aves marinas generalmente se realizan durante la temporada de anidación (Stratoudakis, 1999; Branco et al., 2005). Entre sus presas más comunes se encuentran los peces voladores y calamares (Dorward, 1962; Nelson, 1978; Harrison et al., 1983; Mellink, 2001), aunque no exclusivamente, existe variación en cuanto a sitios además de que hacen un uso importante de los recursos temporales (Cramp y Simmons, 1977, Nelson 1978; Mellink 2001). Ángeles-Pérez y colaboradores (1991) encontraron que no existe una diferencia en cuanto a la composición de las presas, respecto a machos y hembras, aunque si existe una diferencia en cuanto al peso, mostrando un peso mayor los regurgitados de las hembras. Este trabajo pretende hacer una descripción de los componentes tróficos del Sula leucogaster durante la temporada de anidación en una isla del Pacífico central mexicano. La isla Peña Blanca se encuentra al norte del puerto de Manzanillo, Colima a 1.8 km de la línea de costa frente a una playa desierta. Se hicieron visitas periódicas del 15 de enero del 2014 al 17 de julio del 2014, durante la temporada de anidación, en total se recolectaron 90 regurgitados, posteriormente en laboratorio fueron identificadas las presas de cada regurgitado hasta su máximo nivel taxonómico posible, las presas digeridas que no se pudieron identificar se clasificaron como materia digerida. Se encontró un total de 32 componentes en los regurgitados, de los cuales se obtuvo el número de presas, el peso, la frecuencia de aparición y se calculó el índice de importancia relativa. Los componentes con las mayores apariciones fueron Cheilopogon heterurus hubbsi, Fistularia corneta y Oxiphoranfus micropterus, con un total de 80, 50 y 29 individuos respectivamente. Los componentes que representaron el peso mayor fueron Cheilopogon heterurus hubbsi, Coryphaena hippurus y Selar crumenophalmus con un peso de 2230.5, 980 y 693 gr. respectivamente. Los componentes más frecuentes fueron Cheilopogon heterurus hubbsi, Coryphaena hippurus, Selar crumenophalmus y Oxiphoranfus micropterus apareciendo en un total de 26, 13, 13, y 12 regurgitados respectivamente. El índice de importancia relativa (IIR) Pinkas et al. (1971) se calculó mediante la fórmula IIR = (W+N) *F. Donde W es el porcentaje del peso, N es el porcentaje del número de organismos y F es el porcentaje de la frecuencia. Las especies más importantes fueron Cheilopogon heterurus hubbsi, Coryphaena hippurus, Oxiphoranfus micropterus y Selar crumenophalmus. Se observó que la dieta S. leucogaster es muy diversa aunque Cheilopogon heterurus hubbsi tiene

31
casi el 60% de importancia relativa, esto podría deberse a que S. leucogaaster es un buceador pasivo que captura a sus presas por medio de un clavado (Lewis et al., 2004) y C. heterurus es un pez volador, lo que aumenta su detectabilidad y su capturabilidad, también podría deberse a su disponibilidad en esa temporada. Las especies presa de S. leucogaster son distintas a las reportadas en otros trabajos del Pacífico mexicano lo que demuestra su capacidad de respuesta ante un cambio de composición en la diversidad del sitio, en este caso espacial.
Abundancia y alimentación del cuervo común Corvus corax clarionensis (Aves: Corvidae) en la Isla Clarión del Archipiélago Revillagigedo, México
Humberto Almanza Rodríguez1,Luis Abraham Aguilar Nuño2, Salvador Hernández Vazquez3, Diego Adolfo González Zamora4 y Luis Felipe Angulo Castellanos5
1,2,3,4. Universidad de Guadalajara, Centro Universitario de la Costas Sur. Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras. Gómez Farías #82, San Patricio Melaque, Cihuatlán, C.P. 48980, Jalisco. 5Cemtro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA), Universidad de Guadalajara. Apartado Postal 2100. Zapopan, Jalisco, C.P. 45220, México.
[email protected], [email protected], [email protected], [email protected]
El cuervo común (Corvus corax) es un ave de la familia Corvidae que se distribuye en gran parte de México, excepto en la vertiente del golfo, la franja costera del Pacífico central y la península de Yucatán (Navarro & Peterson, 2007). Su alimentación es muy variada, son depredadores (Ratcliffe, 1998; Amar et al., 2010) aunque tienden a ser más carroñeros (Marquis et al., 1978; Amar et al., 2010). Los métodos empleados para conseguir su alimento son muy diversos (Heinrich, 1989, 1999; Ratcliffe, 1997; Boarman & Heinrich, 1999), han sido registrados depredando huevos, pollos e incluso juveniles de algunas aves (Marquiss et al., 1978; Ewins et al., 1986; Marquiss & Booth 1986; Amar et al., 2010). Los recursos provenientes de asentamientos humanos pueden aumentar la densidad de depredadores y afecta a las poblaciones de sus presas (Chapman et al., 1996; Holt, 1984; Schneider, 2001; Boarman et al., 2006). En el desierto de Mojave se cree que los residuos humanos fueron responsables de aumentos (>1,000 % en más de 24 años) en las poblaciones de cuervo en el desierto (Boarman y Berry, 1995; Boarman et al., 2006). Con base a lo anterior el objetivo de este estudio es evaluar la abundancia temporal y el grado de depredación de Corvux corax en la isla Clarión, Archipiélago de Revillagigo. El estudio se llevó a cabo en la isla Clarión, en el Archipiélago de Revillagigedo. La isla se localiza a 710 km al suroeste de Cabo San Lucas, B.C. y 1100 km al oeste del Puerto de Manzanillo, Colima. Mide 8.5 km de longitud y 3.6 km de ancho, con una superficie de 1,980 ha. Esta isla es un sitio importante para la anidación de aves marinas y aves terrestre (Wanless et al., 2009).Los datos fueron obtenidos del 19 de marzo al 30 de junio del 2015. La abundancia de C. corax fue estimada atreves de censos visuales realizados semanalmente en cuatro puntos estratégicos distribuidos a lo largo de la isla (a, b, c y d). La depredación de C. corax fue evaluado en las colonias reproductivas de Sula sula y Sula dactylatra. En el caso de la primer especie se realizaron observaciones semanales en tres parcelas de de 20 x 20 m cada una, y distribuidas en toda la colonia reproductiva (“1”, “2” y “3”). En el caso de Sula dactylatra no se establecieron parcelas debido al tamaño reducido de su colonia y las observaciones se realizaron semanalmente en todos los nidos registrados. Con base a la información generada en los puntos de observación. En el punto A, las abundancias

32
fueron mayores el mes de marzo (entre 23 a 36 organismos) y abril (de 18 a 21 organismos) posteriormente hubo una ligera reducción en mayo y junio manteniéndose una población de entre 6 y 16 cuervos. En marzo en el punto B ocurrió la mayor de las abundancias en todos los puntos observados (69 organismos); para este punto el número de organismos disminuyó a partir de la segunda semana del mes de mayo (12 – 16 organismos) y en una de las semanas de abril (16 organismos); durante los meses de marzo y junio las abundancias fueron superiores a los 25 individuos. Para el punto C hubo una disminución de 50 organismos hasta llegar a 7, en este punto no se muestreo la semana 12 y 13 correspondientes al mes de junio. El punto D hubo una gran variación en las abundancias, ya que se registró la abundancia más baja; en este sitio no se muestreo la semana 13. En la parcela 1 fueron registrados nueve nidos de S. sula, de los cuales fueron depredado un huevo y seis pollos. En la parcela 2 se registraron 13 nidos; solo un huevo y un pollo fueron depredados. En la parcela 3 se registraron 10 nidos y 2 huevos y 4 pollos depredados. Para el bobo enmascarado (S. dactylatra) se registraron 30 nidos y 34 huevos. De los cuales 12 huevos fueron destruidos por C. corax. Para esta especie no se registraron ataques hacia los pollos. Aparte de los ataques a las colonias de anidación de estas dos especies, también se observó depredando aves terrestres, conejos y tortugas marinas que acababan de salir del nido. Este trabajo reporta por primera vez datos de la abundancia y depredación de C. corax en las colonias reproductivas de S. sula y S. dactylatra en la isla Clarión. Sin embargo se requieren estudios que permitan evaluar mas a detalle aspectos de su ecología trófica y reproductiva con el fin de generar estrategias de manejo adecuadas para la conservación de las especies que habitan esta isla. Comparación de dos métodos bioacústicos para el análisis de la riqueza de murciélagos
insectívoros (Mammalia: Chiroptera) en un bosque templado de Jalisco, México Carolina Orendain García1, Silvia Socorro Zalapa Hernández2 y Sergio Guerrero
Vázquez3 1,2,3 Centro de Estudios en Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Km. 15.5 Carretera Guadalajara-Nogales, predio Las Agujas, Nextipac, Zapopan, Jalisco, C.P. 45100. [email protected], [email protected], [email protected] Los murciélagos perciben el mundo a través de los pulsos que ellos emiten y los ecos que reciben (Neuweiler, 2000; Hans-Ulrich y Kalko, 2001). Los llamados de ecolocalización (pulsos) en murciélagos cumplen varias funciones incluyendo orientación y búsqueda de alimento, en ésta última se pueden identificar las fase de búsqueda, acercamiento y captura de presas (Rivera y Burneo, 2013). Los llamados de la fase de búsqueda se sabe que son característicos de familias, géneros y especies, son denominados como firmas vocales las cuales ayudan a identificar especies en vuelo libre (Trejo, 2011). La forma de obtener estos llamados de ecolocalización es mediante detectores y micrófonos ultrasónicos (Pech-Canche et al., 2010), que incorporan distintos mecanismos para transformar los ultrasonidos en frecuencias audibles para el humano (Fenton, 2002) y para extraer el contenido espectral de los mismos (Parsons et al., 2000), lo que permite entre otras cosas, la distinción de especies (Adams et al., 2012). El objetivo de este trabajo es comparar dos métodos bioacústicos: Estación fija vs Transecto móvil, con la intención de aportar información que ayude a lograr mejores inventarios, reduciendo el tiempo y esfuerzo invertido en trabajo de campo, así como a la homogeneización de métodos de campo que permita la comparación entre diferentes

33
estudios. El trabajo se lleva a cabo en el Área de Protección de Flora y Fauna La Primavera, donde se eligieron tres zonas de muestreo: Las Ánimas, Cerritos Colorados y Los Guayabos. Los muestreos se llevarán a cabo de marzo a octubre de 2015 una noche por sitio por mes. En cada sitio se implementan los dos métodos bioacústicos de muestreo durante la misma noche. El método bioacústico Estación fija, que consiste en: la utilización de un detector ultrasónico modelo SM2BAT marca Wildlife Acoustics, activo durante 4 horas a partir de la puesta del sol. El detector se coloca cerca de un cuerpo de agua, en el mismo sitio durante todos los meses de muestreo. Se usa un micrófono SMX2 omnidireccional Wildlife Acustics, elevado aproximadamente a 5 metros de altura. Y el método bioacústico Transecto móvil, que consiste en elegir ocho estaciones de grabación con una separación de 500 m en línea recta entre ellas, las cuales están ubicadas a través de caminos existentes en el área, cada transecto inicia o pasa junto a la Estación fija de cada zona de muestreo. En cada estación se llevan a cabo grabaciones de 20 minutos continuos, iniciando 30 minutos después de la puesta del sol. Se utiliza un detector ultrasónico modelo SM2BAT marca Wildlife Acoustics, y un micrófono modelo SMX1, elevado a aproximadamente 3 metros de altura. Después de cada muestreo, las grabaciones obtenidas son procesadas para obtener los pases que serán analizados, generando una base de datos en Excel, estos pases son visualizados en el programa SonoBat. La determinación de ellos se realizará a nivel más específico posible comparando con bases de referencia existentes y grabaciones del área de estudio. Se obtendrá la riqueza por sitio por mes y por método bioacústico, se elaborarán curvas de acumulación de especies para compara la eficiencia de los métodos bioacústicos. Presentamos los resultados obtenidos hasta mayo de 2015. En total se tiene 5,045 pases de murciélagos; para Cerritos Colorados 2,301 de estos 2,169 para el método bioacústico Estación fija y 132 para el método bioacústico Transecto móvil; en Los Guayabos 937, 849 para la Estación fija y 88 para el Transecto móvil; mientras que en Las Ánimas 1,807, 1,746 en la Estación fija y 61 en el Transecto móvil. Se cuenta con el análisis a nivel especie solo para el mes de marzo, en el que se registraron más especies en el método bioacústico Estación fija que en el método bioacústico Transecto móvil, en todos los sitios. En Las Ánimas se registraron 17 especies en la Estación fija y solo 2 en el Transecto móvil; en Los Guayabos 11 especies en la Estación fija y 2 en el Transecto móvil; mientras que en Cerritos Colorados, 9 especies en la Estación fija y 5 en el Transecto móvil. En total se han registrado 19 especies, 16 pertenecen a la familia Vespertilionidae, 2 para Molossidae y 1 para Mormoopidae. Biología poblacional de Artibeus hirsutus (Chiroptera: Phyllostomidae), en Isla Grande
Atoyac, Jalisco Rogelio García Sandoval1, Silvia Socorro Zalapa Hernández2 y Sergio Guerrero
Vázquez3 1,2,3 Centro de Estudios en Zoología de la Universidad de Guadalajara. Centro Universitario de Ciencias Biológicas y Agropecuarias, Km. 15.5 Carretera Guadalajara-Nogales, predio Las Agujas, Nextipac, Zapopan, Jalisco.
[email protected], [email protected], [email protected] El aporte ecológico que tienen los murciélagos para un ecosistema es fundamental, al influir en la diversidad y dinámica de las comunidades vegetales, así como en el mantenimiento y regeneración de los bosques, al ser de los principales agentes dispersores de semillas, polinizadores y controladores de plagas, entre otros beneficios

34
(Fleming, 1981; Medellín y Gaona, 1999). Sin embargo, para conocer su función ecológica en los ambientes, es necesario contar con información básica de su biología y ecología poblacional. Artibeus hirsutus es un murciélago frugívoro de la Subfamilia Stenodermatinae, es endémico de México y se distribuye en la Vertiente del Pacífico se encuentra en selvas tropicales y bosques templados desde el nivel del mar hasta los 2,575 m de elevación (Webster y Jones, 1983). Es una especie con la cual se ha trabajado poco, solo se cuenta con algunos trabajos que abordan aspectos de su sistema de apareamiento y variabilidad genética (Ortega et al., 2009) y sobre la composición de su dieta (Lumbreras, 2012), así que no se cuenta con información adecuada para determinar su situación actual. De tal manera que el objetivo de este estudio es conocer algunos aspectos de la biología poblacional de Artibeus hirsutus presente en Isla Grande, Atoyac, Jalisco, Isla Grande que es una UMA ecoturistica, ubicada dentro de la cuenca endorreica Zacoalco-Sayula, perteneciente en su mayoría al municipio de Atoyac, en el estado de Jalisco. El trabajo se llevará a cabo de enero a diciembre 2015 visitando la isla 1 día de cada mes. La captura de los murciélagos se realiza en dos cuevas existentes en el área, con la ayuda una trampa-cubeta propuesta por Martínez-Medina (2010). Una vez capturados los murciélagos se colocan en bolsas de manta individuales para su procesamiento. A todos los individuos se les toman los datos convencionales, se marcaran con anillos de metal tamaños 4, 5 y 6 de National Band and Tag Co., y se liberaran dentro de la cueva de la cual fueron capturados. Hasta el momento se han realizado 8 muestreos (enero a agosto), en los cuales se han capturado 119 individuos en total, 80 hembras y 39 machos, siendo febrero y junio los meses con mayor capturas 28 y 22 individuos respectivamente, de los cuales se cuenta 27 individuos recapturados. La proporción de sexos ha sido de 1:1 en los meses de enero, marzo, junio y agosto, mientras que estuvo sesgada hacia las hembras en febrero, abril y julio, y hacia los machos en mayo. Se han evidenciado 2 picos de actividad reproductiva, en hembras los meses de febrero y julio, y en machos enero y junio. La mayoría de los individuos son adultos con apenas el 10% de crías y juveniles.
Estructura genética de Artibeus lituratus (Chiroptera: Phyllostomidae), en Jalisco 1Andres Eduardo Gonzalez Yepez, 2Patricia Castro Félix, 3Silvia S. Zalapa Hernández y
4Felipe J. López Chávez 1Centro Universitario de Ciencias Biologicas y Agropecuarias, Km. 15.5 Carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco. [email protected], [email protected], [email protected],
[email protected] El género Artibeus está distribuido a lo largo de América tropical desde México hasta Argentina incluyendo las Antillas en el Caribe describen 11 especies para el género Artibeus. En México están presentes 3 especies, una de estas es Artibeus lituratus, especie de amplia distribución y abundancia en ambientes tropicales (Merchan-Rivadeneira et al., 2010). Si bien A. lituratus está ampliamente distribuida es evidente que existen barreras geográficas a lo largo de su distribución. En el estado de Jalisco encontramos colonias de esta especie a lo largo de la costa así como en la parte central del estado, situación que nos lleva a pensar que las colonias de la costa y el centro del estado, al estar separadas por las sierra madre del sur, existe una diferenciación genética. ¿Cuáles son los patrones de movimiento entre poblaciones? ¿Las colonias del centro del estado se mueven hacia la costa o viceversa? Estas son preguntas que es difícil de contestar utilizando los estudios tradicionales de ecología, como el de marcaje

35
y recaptura. Para contestas a las preguntas anteriores podemos hacer uso de la genética de poblaciones en este caso con los marcadores de microsatélites. Hasta el momento llevamos probados ocho loci de microsatélites, de los cuales 6 han dado resultados favorables para el estudio.
Evaluación de gremios tróficos de murciélagos (Mammalia: Chiroptera) como bioindicadores potenciales de genotóxicos en el ambiente
Jorge Elías De la torre Miguel1, Silvia Socorro Zalapa Hernández2, María Luisa Ramos Ibarra3 y Sergio Guerrero Vázquez4
1, 2,4 Centro de Estudios en Zoología, 3Departamento de Salud Pública, Centro Universitario de Ciencias Biológicas y Agropecuarias, Km. 15.5 Carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco.
[email protected], [email protected], [email protected], [email protected]
Los quirópteros ocupan una amplia variedad de nichos ecológicos, se desempeñan como controladores de poblaciones de insectos, como polinizadores, dispersores de semillas, depredan y a su vez son depredados por otros organismos. Debido a que sus fuentes de alimentación son tan variadas, se ven expuestos a alimentarse de recursos que podrían estar contaminados, dado que los compuestos usados en la agroindustria como lo son fertilizantes, fungicidas e insecticidas contienen compuestos y sustancias que resultan toxicas como los son los metales pesados que terminan impactando su hábitat (Zucal et al., 2015). Los insectos, frutos y flores de las cuales se alimentan los murciélagos contendiendo estos compuestos gradualmente van generando una acumulación de contaminantes en ellos, que resulta en daños a mediano y largo plazo e inclusive la muerte de los organismos por las altas concentraciones de esto compuestos (Ramos-Ibarra, 2002; Zucal et al., 2015). Otros trabajos ya relacionan directamente a cinco especies de murciélagos frugívoros como bioindicadores de genotóxicos en el ambiente (Núñez, 2015). Fue a partir de este último que surgió la pregunta ¿el tipo de alimento que consumen los murciélagos influye en su nivel de daño genético (considerado como frecuencia de micronúcleos) y con esto algunos puedan ser mejores bioindicadores que otros? Por lo que el objetivo de este trabajo es evaluar a murciélagos de diferentes gremios tróficos como potenciales bioindicadores de genotóxicos en el ambiente. El daño genético se evaluará mediante la prueba de micronúcelos en sangre periférica, los micronúcleos son fragmentos de material genético perdidos durante la mitosis celular, la prueba consiste en la toma una muestra se sangre y su posterior tinción usando un pigmento especial para ácidos nucleicos (Naranja de Acridina), se requiere de un microscopio de fluorescencia para realizar los conteos; las ventajas que tiene el uso de esta prueba sobre otras es que no requiere el sacrificio de los organismos por lo que se pueden realizar estudios a largo plazo además de ser confiable y relativamente económica (Mojica De Leon, 2013; Núñez, 2015). Una de sus desventajas es que no permite la identificación del agente que causa el daño. Se realizarán muestreos en cuatro locaciones en el Área de Protección de Flora y Fauna La Primavera. Se usarán técnicas estándar para la captura y manejo de los organismos. A cada individuo se le tomarán los siguientes datos: largo de antebrazo, peso, sexo, edad, condición reproductiva y se determinarán a nivel especie, del mismo modo a cada organismo se le tomará una muestra de sangre de la vena cefálica con una lanceta estéril y se realizarán dos frotis delgados sobre portaobjetos nuevos y limpios, una vez se verifique el organismo está en buenas condiciones y sin sangrar se procederá a su liberación. Posteriormente los frotis

36
serán fijados en el laboratorio con etanol al 100% durante 10 minutos. Para su análisis deberán ser teñidos con naranja de acridina (Mojica De Leon, 2013), Para su análisis se usará un microscopio de fluorescencia marca zeiss con objetivo de 100X, se realizará el conteo de eritrocitos micronuleados (EMN) en cada 10,000 eritrocitos totales (ET), eritrocitos policromáticos con micronúcleos (EPCMN) en cada 10,000 eritrocitos policromáticos (EPC) y EPC/1,000 ET. Se obtendrán las medias y las desviaciones estándar para cada gremio trófico, se probaran diferencia estadísticas mediante las pruebas paramétricas t de Student y ANOVA, o en su caso no paramétricas U de Mann-Whitney y Kruscal-Wallis (Triola, 2009).
Descripción del esqueleto postcraneal de los murciélagos (Mammalia: Chiroptera) Tadarida brasiliensis y Macrotus waterhousii
Jorge Manuel Vargas Aldaco1 y Silvia Socorro Zalapa Hernández2 1,2 Centro de Estudios en Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Km. 15.5 Carretera Guadalajara-Nogales, predio Las Agujas, Nextipac, Zapopan, Jalisco, C.P. 45100.
[email protected], [email protected] De manera general las especies de murciélagos se agrupan en 6 gremios tróficos: frugívoros, nectarívoros, insectívoros, omnívoros, carnívoros y hematófagos, se ha considerado que existe una relación entre las habilidades de vuelo que es uno de sus atributos característicos y su modo de forrajeo. Por otro lado, el tamaño y características del esqueleto postcraneal que estos presentan, si tiene una relación directa con sus habilidades de vuelo (Wimsatt, 1970). Los murciélagos poseen uno de los esqueletos con mayor diferenciación en comparación con los demás mamíferos, principalmente por el considerable alargamiento de las extremidades anteriores (humero, radio, metacarpos y falanges), aunque también la arquitectura de otras estructuras se han visto modificadas considerablemente, como la columna vertebral, cintura escapular, esternón y la primera costilla (Wimsatt, 1970). Sin embargo, a pesar de su relevancia se le ha dado un menor énfasis a las características postcraneales en relación a las craneales (López-Gonzales y Polanco, 2001). Dentro de los pocos trabajos esta el de Wimsatt (1970) que describe y compara de manera general el esqueleto postcraneal de algunas especies de murciélagos; mientras que, descripciones detalladas a nivel especie para México solo se cuenta con el trabajo de López-Gonzales y Polanco (2001), quién describen el esqueleto postcraneal de Glossophaga morenoi. Por lo que el objetivo de este trabajo es describir el esqueleto postcraneal de 2 especies de murciélagos, Tadarida brasiliensis y Macrotus waterhousii, con la intención de elaborar material didáctico que apoye las materias de Mamíferos y Grupos Zoológicos (Chiropteros) y Anatomía. El material fue donado a la Colección de Vertebrados del Centro de Estudios en Zoología. Lo ejemplares se entregaron muertos y en congelación por lo que se procedió a su limpieza mediante remojo en agua y jabón, hasta obtener el esqueleto limpio, se tomaron fotografías del esqueleto postcraneal por regiones así como del esqueleto completo con una cámara digital. Las imágenes obtenidas sirvieron para medir las estructuras óseas de los murciélagos con el programa: Image-Pro Plus v6, por último los esqueletos fueron armados y montados para su exhibición. Se encontró que en M. waterhouisii los huesos que componen las extremidades anteriores son más grandes, tanto la suma de sus partes como en la comparación individual de cada pieza, presentan una mayor numero de vertebras torácicas y costillas; mientras que T. brasiliensis los huesos que componen las extremidades anteriores son más pequeños, tanto en la suma de sus partes como en la

37
comparación individual de cada pieza y el numero de vertebras torácicas y costillas es menor. El humero que particularmente en los murciélagos tiene un valor taxonómico, en estas 2 especies presenta una extensión similar.
¡Estamos extintos, Mexicanos!, Aspectos de Paleontología y Educación en México Roberto Ramírez Espitia
Centro Universitario de Ciencias Económico Administrativas, Periférico norte y Parres Arias, c.p. 45180, Zapopan, Jalisco, México. Departamento de Estudios Regionales/ DER-INESER
[email protected] ¿Cuántos dinosaurios mexicanos conoces?. México es uno de los países mas importantes a nivel mundial en restos de dinosaurios, los registros más abundantes pertenecen al grupo de los llamados “picos de pato” de la familia Hadrosauridae. También se han encontrado Ceratópsidos, que se caracterizaban por tener cuatro patas y cuernos en el rostro y los Terópodos, que fueron carnívoros de dos patas (Hernández, 2009). El potencial paleontológico de México es abundante y diverso, ya que es posible encontrar evidencias de los diferentes grupos biológicos que han estado presentes en el planeta desde hace 2700 millones de años. Los fósiles forman parte del patrimonio nacional como lo marca la Ley Federal de Monumentos y Zonas Arqueológicas, Artísticos e Históricos. Desafortunadamente existe una compleja problemática que impide que este conocimiento llegue a la población, como ejemplo, no existen programas oficiales de educación básica ni media superior, que acerquen a los estudiantes a estos conocimientos y a nivel profesional es poca la oferta educativa, por lo que parece importante proponer que se incorpore al plan de estudio en educación básica o media superior, con el fin de que esté al alcance de la población estudiantil de México.
Colección Paleontológica del Centro de Estudios en Zoología de la Universidad de Guadalajara
Margarito Mora-Núñez1 y Diego Balderas Noriega2 1Centro de Estudios en Zoología, Departamento de Botánica y Zoología, C.U.C.B.A., Universidad de Guadalajara, México. 2 Estudiante de Licenciatura en Biología, C.U.C.B.A., Universidad de Guadalajara, México.
[email protected] Las colecciones científicas tienen una gran importancia, tanto en la investigación como en la conservación de material que representa el patrimonio natural de determinada región. Las colecciones paleontológicas, en específico, resguardan acervos de material fósil que sirve de base a múltiples investigaciones de diversa índole. La organización de fondo de las colecciones, así como la cantidad y calidad de datos de los ejemplares son los elementos que les dan un valor real a las mismas. La Colección Paleontológica del Centro de Estudios en Zoología comenzó su formación en el año 1999 con la incorporación y sistematización de algunas colectas de fósiles hechas por estudiantes dentro de la asignatura de Paleobiología entre los años 1994 a 1998. Desde esa fecha se ha ido incorporando material obtenido por salidas de prospección y colecta por parte de personal del CEZUG, así como material donado por particulares y estudiantes. La elaboración de la base de datos y el registro de ejemplares está actualmente en proceso.

38
Actualmente, la colección cuenta con ejemplares principalmente de Jalisco, pero también existe material de Coahuila, Puebla, Hidalgo y Zacatecas. Además, existe material de invertebrados cretácicos donados, procedentes del estado de Texas, en Estados Unidos. Los municipios jaliscienses representados en la colección son principalmente: Chapala, Ameca, Amatitán, Zacoalco de Torres y Zapopan. Los ejemplares son principalmente piezas permineralizadas e impresiones. Los grupos biológicos representados son básicamente mamíferos, actinopterigios, crustáceos, gasterópodos, bivalvos, coníferas, angiospermas, y, en menor proporción (representados por piezas aisladas) existen aves, reptiles y anfibios. El material jalisciense procede básicamente de estratos del Cretácico inferior, Mioceno, Plioceno y Pleistoceno. El trabajo de sistematización de la colección comenzó en 2007 con apoyo de estudiantes en su servicio social; tal actividad continúa actualmente y se encuentra en proyecto la elaboración de la base de datos, el catálogo de localidades y la formación de una colección osteológica de referencia. Posteriormente se solicitará al INAH su registro formal. Es fundamental que la carrera de Biología y en general la Universidad de Guadalajara cuente con una colección de fósiles que represente al extenso patrimonio paleontológico del Estado de Jalisco, y con esto contribuir al conocimiento de la historia natural de la región occidente del país. Uso de técnicas de escalada y espeleología en la exploración de campo para monitoreo
y muestreo en actividades de investigación zoológica y botánica Rodrigo Andrade Villegas1 y Héctor Leonel Ayala-Téllez2
1Estudiante de Lic. en Biología, 2Departamento de Ciencias Ambientales, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Km. 15.5 Carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco.
[email protected], [email protected] El hombre ha sido explorador por naturaleza, ha modificado lo que tiene a la mano para crear herramientas, las ha utilizado para conquistar todo terreno, ahora el hombre va en busca de conocimiento, explora para buscar respuestas, ha ido a la cima más alta, a los polos, al fondo marino y para ello requiere de conocimiento especializado de técnicas que le ayuden a lograrlo. Las practicas espeleológicas y de escalada son actividades especiales, cada una con su técnica, equipo y herramientas propias a utilizar. Estas actividades nos permiten realizar recorridos con fines deportivos, sin embargo se pueden utilizar en el campo educativo, ilustrativo, investigativo y centrándonos en nuestro trabajo a la identificación tanto de fauna y flora (Alvarez-Garay, 2014). Nosotros como investigadores vamos al campo, para ello, se necesita un conocimiento mínimo de progresión de acuerdo con la dificultad del terreno, se podrá utilizar solo nuestro cuerpo o se deberá adquirir conocimiento más especializado, ya sea la aplicación de las técnicas de ascenso y descenso o el uso de materiales especiales para ser capaz de progresar por cualquier tipo de terreno agreste de forma segura. Con ayuda de estas técnicas, los investigadores han alcanzado lugares difíciles, como la copa de una secuoya, el fondo marino, la sima de una cueva o los glaciares de las altas montañas. Para aumentar el conocimiento en las investigaciones de campo dentro de la Biología, es importante estar complementado los protocolos de investigación, cuando así se requieran, con técnicas especializadas que ayuden a cubrir e incrementar sitios y rangos nuevos que por su difícil acceso, quedan sin estudiar ni conocer (barrancos, acantilados, cuevas, etc.). Moreno, (2007) menciona que ningún método es capaz de

39
registrar todos los componentes de la fauna para realizar un inventario completo y es necesario combinar todos los métodos y técnicas posibles para aumentar los esfuerzos de muestreo en el campo, aunque estas sean más costosas y requiera de la participación de personal capacitado. Por este motivo se pretende hacer mención de técnicas especializadas que en un principio nacen con otros fines, pero en el ámbito de la investigación zoológica y botánica pudieran contribuir a alcanzar sitios inexplorados o que por su naturaleza topográfica tales como acantilados, barrancos, copas de árboles y cuevas resultan más difíciles de acceder e investigar.
El género Vaejovis C. L. Koch, 1836 (Scorpiones: Vaejovidae) en Jalisco Gerardo Adalberto Contreras Félix y Oscar Francke
Colección Nacional de Arácnidos (CNAN), Instituto de Biología, Universidad Nacional Autónoma de México, 3er. Circuito Exterior, Cd. Universitaria. México D. F., C. P. 04510.
[email protected] En el mundo se conocen cerca de 1900 especies de alacranes (Francke, 2014). Para México, la familia Vaejovidae, presenta la mayor diversidad de especies y géneros en México y toda Norte América; dentro de esta familia, el género Vaejovis, es el que presenta la mayor diversidad con 57 especies (Soleglad y Fet, 2008). Este género ha sido tradicionalmente dividido en varios grupos de especies (Hoffmann, 1931; Soleglad, 1973; Williams, 1980; Sissom y Francke, 1985; Soleglad y Fet, 2008; Graham y Bryson, 2010; Santibáñez, López y Francke, 2010), lametablemente, las diagnosis y límites que comprenden a estos grupos son aún confusas (Sissom, 2000; Prendini y Wheleer, 2005; Soleglad y Fet, 2008). En Jalisco, con una superficie de 78’588 Km2 (Inegi, 2012), la diversidad de escorpiones es baja en relación a su tamaño, con solo 6 géneros de los cuales Vaejovidae presenta la mayor diversidad con 4; de los cuales solo 3 especies pertenecen al Vaejovis (Francke y Santibáñez-López, 2014; Contreras-Félix et al., 2015). Aunque por trabajos en curso es probable que aumente la diversidad de este género a mas de 5 especies (Contreras-Félix y Francke, en prensa). En Jalisco se presentan solo dos grupos del género Vaejovis: el denominado grupo “mexicanus” y el grupo “nigrescens” (o “nitidulus”, dependiendo el autor que se decida seguir). El grupo “mexicanus” se reconoce por la presencia de las quillas ventral medianas de los segmentos I-IV del metasoma granulares, además de la ausencia de un tapón genital en el espermatóforo; mientras que el grupo “nigrescens” se reconoce por presentar las quillas ventral medianas de los segmentos I-IV del metasoma lisas y la presencia de un tapón genital. Hay una considerable disparidad entre los dos grupos presentes en Jalisco, esto en gran medida por la cantidad personas trabajando con cada uno de esos grupos de especies; mientras que para el grupo “nigrescens” se han descrito 2 especies en los últimos 10 años, para el grupo “mexicanus” se han descrito mas de 9 en los últimos 10 años, mas otras 5 especies de este grupo que están en proceso de ser descritas (Francke, com. pers.). Con este precedente se espera que el numero de Vaejovis en Jalisco aumente considerablemente en los próximos años, pero es necesario más trabajo de campo en zonas donde existan pocos registros para llegar a un mejor entendimiento de la distribución de las especies.

40
Selección de las presas por parte del depredador generalista Geocoris punctipes (Hemiptera: Lygaeidae): análisis de movilidad, estadío y coloración
Walter Ernesto Printzen Cruz¹ y Ricardo Ramírez Romero² ¹,²Centro Universitario de Ciencias Biológicas y Agropecuarias, Km. 15.5 Carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco.
¹[email protected], ²[email protected] Geocoris punctipes Say (Hemiptera: Lygaeidae) es un insecto depredador y se encuentra entre los enemigos naturales más eficaces de plagas de insectos que infestan los cultivos de campo e invernadero (Crocker & Whitcomb, 1980; Boyd & Boethel, 1998). Myzus persicae Sulzer (Hemiptera: Aphididae) es una especie polífaga asociado a hortalizas, siendo plaga para diversos cultivos como es el chile serrano (Capsicum annuum). El manejo de esta plaga ha sido básicamente con insecticidas sintéticos, provocando problemas ambientales y en su control (Ortega, 2008). Una alternativa es el control biológico, para regular la plaga mediante agentes biológicos, como parasitoides y depredadores (Sánchez-Ruiz, 1997). Sin embargo es importante conocer la ecología del depredador, para optimizar su empleo como agente de control biológico. El objetivo del presente estudio fue determinar si el depredador G. punctipes tiene preferencia por ciertas presas en función de diferentes factores como la movilidad, el estadío y la coloración. Para ello se realizaron dos bioensayos en una arena de observación referida como microcosmo, donde se determinó la tasa de depredación bajo diferentes tratamientos. El primer bioensayo fue sin opción y se establecieron los siguientes tratamientos: 1) control, 2) presas inmóviles, 3) presas móviles, 4) presas de primer estadío, 5) presas de cuarto estadío, 6) presas adultas, 7) presas oscuras y 8) presas claras. El segundo bioensayo fue con opción y se probaron los siguientes tratamientos: 1) presas móviles vs. inmóviles, 2) presas del primer estadío vs. presas adultas y 3) presas oscuras vs. claras. Estos bioensayos se realizaron bajo condiciones de laboratorio a una temperatura de 24±3°C y humedad relativa de 50±10%. Análisis estadísticos preliminares en ambos bioensayos indican que G. punctipes, no presenta diferencias significativas en la elección de sus presas bajo los factores probados. Sin embargo se observan ciertas tendencias: 1) la tasa de depredación tiende a ser mayor en presas móviles, 2) la tasa de depredación tiende a ser mayor en presas de primer estadío y 3) la tasa de depredación tiende a ser mayor en presas claras. Será necesario realizar más replicas para aumentar el poder del experimento. Sin embargo es posible que estas tendencias se observen porque las presas móviles son más visibles, las presas pequeñas posiblemente son más fáciles de ser sometidas y las presas claras indiquen un estado de buena salud.
Descripción de la estructura de las redes mutualistas planta-abeja (Hymenoptera: Apoidea: Anthophila) en dos sitios con diferente recurrencia de incendios en Bosque
Escuela de la Sierra La Primavera Christian Fernando Barajas-Daniel1, Alejandro Muñoz-Urías2, Osiris Yanid Serrano-
Pineda3, Hugo Eduardo Fierros-López4 y Claudia Aurora Uribe-Mú5 Centro Universitario de Ciencias Biológicas y Agropecuarias, Camino Ramón Padilla Sánchez 2100, Nextipac, 44600, Zapopan, Jalisco, México.
[email protected], [email protected], [email protected], [email protected], [email protected]

41
El análisis de las interacciones a través de la teoría de redes se considera una herramienta útil en el monitoreo del funcionamiento de los ecosistemas (Hegland et al., 2010), y se han utilizado para evaluar el cambio en las interacciones por tipo de vegetación (Razo-León, 2015), por escala temporal (Díaz-Castelazo et al., 2010) y por grados de perturbación (Vázquez & Simberloff, 2003). Se conoce el efecto que tienen los incendios sobre la composición de las comunidades vegetales (Cárdenas-Arévalo & Vargas-Ríos, 2008), sin embargo, son escasos los estudios acerca de su efecto sobre las comunidades de antófilos y sus interacciones, por lo cual el presente trabajo tiene como objetivo describir el efecto de la recurrencia de incendios sobre las redes mutualistas planta-abeja. El trabajo de campo se llevó a cabo dentro del Área de Protección de Flora y Fauna Bosque La Primavera en el municipio de Tala, Jalisco (entre los meridianos 103°37’15’’ y 103°40’08’’ Oeste, y los paralelos 20°36’26’’ y 20°34’34’’ N), que cuenta con un registro de 104 especies de abejas (Fierros-López et al., 2014). En el área de estudio se han registrado hasta 60 incendios forestales en los últimos 15 años (Huerta-Martínez & Ibarra-Montoya, 2014). Se definieron dos sitios de muestreo de 30 x 100 m; uno de ellos se incendió en dos periodos “+if”, mientras que el otro no cuenta con registros como evidencia de incendios “-if” desde 1995 (Huerta-Martínez & Ibarra-Montoya, 2014). En ambos sitios se hicieron recorridos de aproximadamente dos horas, donde se permaneció alrededor de 10 minutos en cada planta que se encontraba en floración (recurso disponible). Los muestreos se llevaron a cabo durante los meses de febrero a diciembre de 2012 y 2013. Con los datos obtenidos en campo se estimaron los siguientes parámetros: Riqueza específica; número de enlaces o links; conectancia; asimetría de la red; anidamiento; proporción de especies que contribuyen al patrón de anidamiento de la red e índice de similitud de Jaccard para la composición de interacciones de la red, así como la composición específica. Se utilizó la librería bipartite del software R 3.1.2 (R Core team, 2014) y el software ANINHADO (Guimarães & Guimarães, 2006). En el sitio “-if”, se registraron 76 especies que participan en la red planta-antófilo, de estas, 46 de abejas y 30 de plantas con un total de 81 enlaces, mientras que para la red del sitio “+if”, se registraron 59 especies en total, 31 de abejas y 28 de plantas con un total de 54 enlaces. Cuando se analizó el número medio de enlaces, se registraron valores de 1.88 en “-if” y 1.74 “+if” para abejas, mientras que los valores fueron de 2.7 en “-if” y 1.92 en “+if” para plantas. El valor de conectancia en ambas redes fue de 0.06, la cual representa el 6% del número total de interacciones posibles. En el sitio “-if” se registró una asimetría de 0.18, mientras que en “+if” fue de 0.05. Ambas redes presentaron patrón de anidamiento (p < 0.0001), y ambos sitios no mostraron diferencias estadísticamente significativas entre sus temperaturas (TM) (X2=0.0927, p= 0.7608). No hubo diferencias significativas entre los sitios "+if" y "-if” en el análisis del porcentaje de especies que contribuyen al anidamiento, donde se registraron valores entre 34.88% y 38.71% para las abejas, mientras que los porcentajes para especies de plantas oscilaron entre 26.67% y 32.14%. El índice de Jaccard para evaluar el recambió de las especies que integran la red, mostró valores entre 21% y 30%, mientras que la similitud de interacciones de manera general, fue de 6.3%, y tomando únicamente en cuenta las especies compartidas, se registró que sólo el 22.86% de las interacciones fueron las mismas en ambos sitios, sugiriendo que existen cambios en la dinámica mutualista. Dado que en este estudio registramos que la asimetría entre redes es distinta, y que existe una elevada tasa de recambio de especies e interacciones, estos resultados sugieren que la recurrencia de incendios forestales no solamente tiene un efecto sobre la estructura de la comunidad de plantas que integran la red, sino que también influye en las interacciones que forman la red mutualista.

42
Hormigas (Hymenoptera: Formicidae) del Jardín Botánico Vallarta, Cabo Corrientes, Jalisco, México
Anahí Cisneros-Caballero1 y Miguel Vásquez-Bolaños2 Entomología, Centro de Estudios en Zoología, Departamento de Botánica y Zoolgía, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Camino Ing. Ramón Padilla Sánchez #2100, Las Agujas, Nextipac, Zapopan, Jalisco, México.
[email protected], [email protected] Las especies de hormigas se agrupan en la familia Formicidae, pertenecen al orden Hymenoptera (junto con abejas, avispas y similares). Las colonias de hormigas tradicionalmente se componen por castas: una reina, obreras estériles y machos (Fernández, 2003). Las hormigas están presentes en la tierra desde hace aproximadamente 120 millones de años, su éxito se debe a la presencia de la glándula metapleural que les permitió explotar y colonizar el ambiente terrestre al secretar sustancias antibióticas para combatir los microorganismos presentes en este ecosistema. Son un grupo diverso y abundante que habita en casi todos los ambientes terrestres del planeta, desde los bosques tropicales hasta los pastizales templados, zonas áridas y semiáridas, así como en los agroecosistemas y zonas urbanas. Se distribuyen desde el nivel del mar hasta los 4,000 metros, la mayoría de las especies se concentran en los trópicos y entre los 800 y 1,600 msnm, están ausentes en los polos y en las altas montañas. Presentan hábitats muy variados las hay con nidos subterráneos, en hojarasca, en la madera en descomposición, sobre el suelo, en las plantas o dentro de ellas (Vásquez-Bolaños y Quiroz-Rocha, 2013; Alatorre-Bracamontes y Vásquez-Bolaños, 2010). Para México se han registrado 927 especies, pertenecientes a 93 géneros y once subfamilias (Vásquez-Bolaños, 2015). El objetivo del presente trabajo es realizar un inventario de las especies de hormigas presentes en el Jardín Botánico Vallarta. El cual se encuentra en el Km. 24 de la carretera Puerto Vallarta-Barra de Navidad, Las Juntas y Los Veranos, Cabo Corrientes, Jalisco, México. Tiene una extensión de ocho hectáreas, limita al norte con el cauce del río Los Horcones, cuenta con un gradiente altitudinal va de los 300 a los 400 msnm. El tipo de vegetación predominante es el Bosque Tropical Subcaducifolio pero también presenta áreas perturbadas y se cultivan especies vegetales nativas y exóticas. Se realizaron colectas de manera directa en tres fechas: noviembre 2013, julio 2014 y agosto 2014, en diferentes puntos del jardín para tratar de abarcarlo en su totalidad. Las hormigas colectadas se conservaron en alcohol diluido al 70%, se etiquetaron con la información correspondiente al sitio de colecta. Se transportaron al Laboratorio de Entomología del Centro de Estudios en Zoología de la Universidad de Guadalajara, en donde se encuentra depositado el material. Se separaron, determinaron con las claves para el grupo a nivel especie y etiquetaron; algunos ejemplares se montaron. Hasta el momento se tienen determinadas 50 morfoespecies pertenecientes a 23 géneros y siete subfamilias: Dolichoderinae.- Azteca sp., Dorymyrmex sp., Tapinoma meanocephalum; Dorylinae.- Eciton mexicanum, Eciton burchellii parvispinum, Nomamyrmex esenbeckii mordax; Ectatomminae.- Ectatomma tuberculatum, Gnamptogenys striatula; Formicinae.- Anoplolepis sp., Brachymyrmex sp. 1, Brachymyrmex sp. 2, Camponotus sericeiventris, Camponotus sansabeanus, Camponotus lespesii melancholicus, Camponotus sp., Nylanderia steinheili; Myrmicinae.- Atta mexicana, Cephalotes maculatus, Cephalotes goniodontus, Crematogaster sp., Monomorium sp., Mycocepurus smithii, Pheidole sp. 1, Pheidole sp. 2, Pheidole sp. 3, Pheidole sp. 4, Pheidole sp. 5, Pheidole sp. 6, Rogeria sp., Solenopsis geminata, Solenopsis sp. 1,

43
Solenopsis sp. 2, Solenopsis sp. 3, Trachymyrmex sp., Wasmannia auropunctata; Ponerinae.- Odontomachus sp., Neoponera verenae; Pseudomyrmecinae.- Pseudomyrmex gracilis, Psedomyrmex ferrugineus. Los géneros más abundantes son: Solenopsis con 122 individuos, Pheidole con 82, Wasmannia con 49 y Eciton con 48. Los menos abundantes son: Odontomachus, Anoplolepis, Nylanderia y Ectatomma con tres individuos cada uno, Rogeria con dos, Crematogaster y Neoponera que sólo cuentan con un individuo cada uno. Los géneros con mayor diversidad son Pheidole con ocho morfoespecies y Solenopsis con seis. Es probable que el número de especies sea mayor al encontrado en este trabajo, debido a las condiciones ambientales y a la complejidad de la vegetación. Se requiere mayor esfuerzo en el trabajo de campo para conocer la diversidad de hormigas en el jardín, así como para determinar si se tratan de especies nativas o introducidas. Las sociedades de hormigas (Hymenoptera: Formicidae) como modelo en el estudio de
sistemas complejos Luigui Michel Gallardo-Becerra1 y Christian Fernando Barajas-Daniel2
Centro Universitario de Ciencias Biológicas y Agropecuarias, Camino Ramón Padilla Sánchez 2100, Nextipac, 44600, Zapopan, Jalisco, México.
[email protected], [email protected] Los sistemas complejos se definen como un conjunto de unidades similares que interaccionan y generan propiedades emergentes. Dichas propiedades no pueden ser explicadas por la unidad (Konieczny et al., 2014). Actualmente se reconoce que el abordaje de los sistemas biológicos a través de las ciencias de la complejidad, significan estudios de frontera. Esto se refleja en múltiples avances realizados recientemente en distintas áreas de la biología (e.g. genómica, biología de sistemas y ecología), que han utilizado dichas herramientas (Kauffman, 2014). Las hormigas (Hymenoptera: Formicidae), al estar representadas por casi 13,000 especies (Agosti & Johnson, 2005) y ser todas eusociales (Plowes, 2010), han servido de inspiración para otras áreas del conocimiento además de la biología, entre ellas la computación y la robótica. Un ejemplo son los algoritmos de optimización, como el caso de Linepithema humile en el experimento del puente doble, donde la ruta más utilizada del hormiguero al alimento, depende de las marcas de feromonas y la tasa de evaporación de estas; o el de Leptothorax unifasciatus, donde se utiliza su sistema de clasificación de crías en analogía con la clasificación de datos (Bonabeau, 2000). Como todos los sistemas complejos, las sociedades de hormigas se caracterizan por la inexistencia de un control central. El comportamiento colectivo de la colonia se rige por las interacciones locales entre sus miembros, y se ha registrado recientemente en Camponotus fellah, que el aislamiento de un individuo tiene como consecuencia una reducción en su esperanza de vida en comparación con aquellos que se encuentran en grupo (Koto et al., 2015). En 1980, Douglas Hofstadter sugirió en su libro “Gödel, Escher, Bach”, que pensáramos en una colonia de hormigas como si fuera un sistema de procesamiento distribuido, donde un conjunto de computadoras se conectan a la red, dándole al usuario la sensación de ser una sola (Hernández-Vázquez, 2005). Sin embargo, Deborah Gordon en su “Ant Encounters” (2010), afirma que dicha analogía no hace justicia a las hormigas, dado que las colonias de estos insectos demuestran con mayor frecuencia un elemento muy importante: las propiedades emergentes. Por ejemplo, el comportamiento individual de Pogonomyrmex barbatus es caótico, además de no realizar actividades complejas (e.g. forrajeo y construcción de nido), sin embargo continúa alimentándose y desplazándose

44
(Gordon, 2010). La existencia de una colonia propicia la interacción entre los individuos y asignación de tareas. Sin embargo, no todas las interacciones entre los individuos son posibles al mismo tiempo; éstos “huecos” generados en el posible universo de interacciones facilitan la división del trabajo. Esta emergencia surge en un punto medio entre el orden y el desorden, que algunos autores lo definen como “borde del caos” (Miramontes, 2000). Por último, la auto-organización (generación aparentemente espontánea de patrones), puede ser observada en una colonia en respuesta a condiciones dadas en un momento determinado. Muchos son los ejemplos de hormigas que ensamblan sus cuerpos para desarrollar una actividad (Anderson et al., 2002). Tal es el caso de Solenopsis invicta, las cuales construyen con sus cuerpos una estructura que simula a una balsa cuando son sumergidas en agua. No obstante, sólo recientemente se desarrollaron experimentos que revelan las propiedades subyacentes de dicha estructura, que pudieran ser de utilidad en la ingeniería (Foster et al., 2014). Debido a que es común observar distintas castas de hormigas que desempeñan diversas tareas, además de que interactúan con mecanismos químicos y mecánicos para mantener la colonia, es posible que estos insectos nos sigan proporcionando herramientas que contribuyan al conocimiento de aquellos sistemas en los que las relaciones causa-efecto dependen de múltiples variables. Uso médico de la grana cochinilla en la época colonial mexicana Fernando Rafael Villaseñor Ulloa Coordinación de Bibliotecas. Universidad de Guadalajara, Av. Hidalgo #935, Colonia Centro, Guadalajara, Jalisco. C.P. 44100.
[email protected] Uno de los más importantes descubrimientos comerciales que se realizaron al inicio de la época colonial mexicana fue sin duda la grana cochinilla (Dactylopius coccus Costa), pequeño insecto parásito del nopal cuya fama e importancia comercial procedió de su capacidad para proporcionar un colorante productor de diversas tonalidades rojas. Sin embargo, es importante señalar, que aunque existía evidencia de su uso médico por parte de los pueblos prehispánicos, lo cierto es que fue documentado de forma muy pobre. Aunque con el nacimiento de la vida colonial, la grana cochinilla se fue ganando una fuerte reputación como productora de riqueza (18% de los ingresos por exportación de Nueva España provenían de su comercio), y por lo tanto, todas sus virtudes se volvieron convenientemente fuentes de estudio. La medicina encontró entonces un elemento con múltiples posibilidades de aplicación para la salud de seres humanos, las cuales, si bien nos podrían parecer a éstas alturas de la civilización algo descabellado, en su momento, fueron el reflejo de la más pura ciencia de la época. Si bien y a diferencia de otros usos para el insecto, importantes exploradores del siglo XVIII como Humboldt, Clavijero y Menonville, desconocen su aplicación médica, otros autores menos conocidos y algunas fuentes especializadas como diccionarios, farmacopeas, compendios de cirugía y crisoles médicos si dan cuenta de esa utilidad suya. Entre los más notables aspectos que eran tratados por medio de la aplicación de alguno de los componentes de la grana cochinilla podemos encontrar: su uso como sedante, su capacidad de restitución de la salud cardiaca, su uso como sudorípara para restaurar de fiebres de larga duración, su capacidad para restaurar articulaciones lastimadas o desgastadas por la edad, liberador del estómago, remedio para la sarna humana y animal, agente de limpieza dental, reconstituyente sexual, excelente auxiliar para evitar

45
abortos y evitar males de piedra en el riñón. Mostrar éstos descubrimientos y los textos de donde provienen son los objetivos del presente.
Diversidad de melolóntidos nocturnos (Insecta: Coleoptera) en un bosque tropical caducifolio de la Sierra del Águila, Ameca, Jalisco, México
José Ernesto Colima-Lara1, Luis Eugenio Rivera-Cervantes2, Adriana Elena Castro-Ramírez3 y Edith García-Real2
1Ingeniería en Recursos Naturales y Agropecuarios, Centro Universitario de la Costa Sur, Universidad de Guadalajara, Avenida Independencia Nacional 151, Autlán de Navarro, Jalisco, C.P. 48900, México. 2Instituto Manantlán de Ecología y Recursos Naturales, Centro Universitario de la Costa Sur. 3El Colegio de la Frontera Sur, Carretera Panamericana y Periférico s/n, María Auxiliadora, San Cristóbal de las Casas, Chiapas, C.P. 29290, México.
[email protected], [email protected], [email protected], [email protected]
La Sierra del Águila es un área natural protegida (21,109.3019 ha) decretada en 2010 para la protección hidrológica de la región Valles. Ubicada entre los municipios de Ahualulco de Mercado, Ameca, Etzatlán y San Juanito de Escobedo, Ameca ocupa el segundo lugar en extensión (6,816.7109 ha, 32.29%). Nosotros analizamos la diversidad de los Melolonthidae adultos de hábitos nocturnos, presentes en un bosque tropical caducifolio del municipio de Ameca, Jalisco; con el objetivo de determinar la composición específica y la abundancia estacional, así como sus patrones fenológicos y comprobar mediante índices la diversidad y la relación fenológica de los escarabajos con la precipitación registrada. Las recolectas se realizaron por un año de manera sistemática, de julio 2013 ha junio 2014. Los muestreos se realizaron tres noches por mes siguiendo el periodo de luna nueva (máxima oscuridad) entre las 19:30 y las 22:30, utilizando una trampa de luz tipo pantalla. En total se recolecto 2,297 adultos de 3 sufamilias, 12 géneros y 35 morfoespecies. Los géneros más abundantes fueron; Cyclocephala con 1050 individuos (46%), Diplotaxis con 605 (26%), Phyllophaga con 410 (18%) y Paranomala con 173 (7%). Los géneros más diversos fueron Phyllophaga con 13 morfoespecies, Paranomala con 6 y Diplotaxis y Isonychus con 3. La proporción de machos y hembras, está varió según el género, en Cyclocephala y Diplotaxis existió una relación 1:1, en Phyllophaga los machos fueron más abundantes en una proporción 1.5:1, en cambio en Paranomala las hembras fueron más abundantes en una proporción de 1.75:1. La mayor riqueza y abundancia se presentó en los meses de junio y julio (inicio del periodo de lluvias). En cuanto a la actividad de los melolónthidos adultos, está fue distinta y abarco 8 meses. A nivel específico Cyclocephala forcipulata fue dominante en relación con las otras registradas. Los periodos de actividad de vuelo para las cuatro especies más abundantes (C. forcipulata, Phyllophaga chiapensis, Diplotaxis sp. 1 y Paranomal aff. inconstans) va de las 20:00 a las 22:30, presentando su pico más elevado entre las 21:00 y las 22:30. La diversidad alfa fue analizada mediante los índices de Shannon-Weiner y Brillouin dando como resultado H´= 1.89 y HB= 1.86 respectivamente, por otro lado Spearman mostró una relación positiva entre la riqueza y precipitación (0.54, p<0.05), mientras que para la abundancia y precipitación no existió alguna (0.47, p<0.05). Phyllophaga fue el más rico en especies, sin embargo el más abundante fue Cyclocephala¸ comportamiento que solo se ha visto muy pocas veces de acuerdo a estudios realizados para el país. Por

46
último, se pudo observar que existe una relación estrecha entre la fenología de los escarabajos nocturnos con la precipitación. 30 años conociendo la biodiversidad de los escarabajos (Coleoptera: Scarabaeoidea), de
los sistemas montañosos del sur oeste de Jalisco, México Luis Eugenio Rivera-Cervantes y Edith García-Real
Instituto Manantlán de Ecología y Conservación de la Biodiversidad, Centro Universitario de la Costa Sur, Universidad de Guadalajara. Avenida Independencia Nacional 151, Autlán de Navarro, Jalisco, C.P. 48900, México.
[email protected]; [email protected] A mediados de la década de los 80´s del pasado S. XX, la Universidad de Guadalajara creó el ahora Instituto Manantlán de Ecología y Conservación de la Biodiversidad (IMECBIO), para entre otros objetivos, generara información biológica (flora y fauna) de la actual Reserva de la Biosfera Sierra de Manantlán. De esta manera es que a partir de 1986 personal del laboratorio de zoología del IMECBIO inician los inventarios entomológicos en esta área natural protegida del occidente de México, en particular la biodiversidad de escarabajos en los diferentes ecosistemas forestales de esta sierra: bosque mesófilo de montaña, bosque de oyamel, bosque de pino-encino, bosque de encino subperennifolio y caducifolio, y el bosque tropical caducifolio y subcaducifolio. En la última década del S. XX y los 15 años que llevamos de esté S. XXI, los inventarios de escarabajos se han extendidos a otros sistemas montañosos del sur de Jalisco como son: Sierra de Tapalpa, Sierra de Quila, Sierra de Cacoma y Sierra Occidental (Mascota). Los trabajos de inventario se han realizado de manera sistemática o esporádica, dependiendo del apoyo del que se cuente o de la situación de inseguridad que se presente en la región. Las técnicas de colecta han incluido: trampas de luz fluorescente, trampas cebadas para la captura de escarabajos necrófagos, coprófagos o carpófagos, y colectas directas con redes entomológicas y hachas (escarabajos xilófagos). A la fecha se han capturado más de 45,000 especímenes, los que se encuentran montados o preservados en alcohol al 70%, todos ellos depositados en la colección entomológica del IMECBIO, en Autlán, Jalisco. En estos 30 años de inventario de los escarabajos del suroeste de Jalisco, nos han permitido descubrir más de 25 especies nuevas para la ciencia de las familias Scarabaeidae, Melolonthidae y Passalidae, más de 10 nuevos registros de distribución, la realización de seis tesis de licenciatura y tres de maestría. La posición geográfica del estado de Jalisco, su gradiente altitudinal, su historia geológica y volcánica, hacen que confluyan especies de los dos reinos biogeográficos del continente, así como endemismos propios, lo cual ha permitido que nuestro estado se ubique entre los primeros cinco con mayor riqueza de escarabajos en México. Sin embargo, falta mucho por conocer aún, existen muchos municipios y sistemas montañosos pobremente estudiados.
Insectos descortezadores de los géneros Ips y Pseudips en Pinus cooperi B. y P. durangensis Mtz. en Pueblo Nuevo, Durango
1Yolanda Ontiveros Moreno, 2Rebeca Álvarez Zagoya y 3José Javier Corral Rivas 1,3Universidad Juárez del Estado de Durango, Facultad de Ciencias Forestales, Río Papaloapan y Boulevard Durango S/N, Col. Valle del Sur, Durango, Dgo. C.P. 34120. 2CIIDIR - IPN Unidad Durango, Sigma No.119, Fraccionamiento 20 de Noviembre II, Durango, Dgo. C.P.34220.

47
[email protected], 2 [email protected], [email protected] En el estado de Durango se presentó una plaga de insectos descortezadores de los géneros Dendroctonus e Ips, teniendo una pérdida anual del 25 % en el área forestal, este fenómeno se atribuyó por el calentamiento global, cambios de temperatura, incendios forestales y sequías que ocurrieron en el año 2010-2012 (Aguilar et al., 2013). Con la presencia de estos descortezadores se emplearon varios tipos de saneamiento, y a su vez, estudios e investigaciones realizados en estas zonas afectadas, siendo una de ellas el Paraje Santa Bárbara, Municipio Pueblo Nuevo, Durango, con la finalidad de conocer y evaluar el impacto de estos. Se realizaron tres muestreos en dos especies arbóreas de importancia, Pinus cooperi y Pinus durangensis, con derribo, desramado y descortezado del tronco total de cada árbol, donde se tomaron muestras en secciones de 50 cm cada una, siendo un total de 260 secciones revisadas bajo estereomicroscopio, en 3 árboles por fecha de muestreo. Cada sección de corteza se mantuvo en bolsas plásticas, en refrigeración para la revisión de los insectos (descortezadores y otros), se separaron e identificaron a nivel de especie, y su separación en hembras y machos. Los datos de insectos en cada sección, por fecha y especie arbórea, fueron analizados estadísticamente con los índices de Margalef y Shannon (Indice de diversidad y de abundancia respectivamente). Los resultados muestran la abundancia de los descortezadores a lo largo del fuste, de acuerdo a la especie, sexo y fecha, para tres especies de plagas primarias, Ips pini, Ips lecontei y Pseudips mexicanus, de las cuales el descortezador Ips pini presentó una abundancia de 1188 individuos en ambos pinos ya mencionados anteriormente, seguido de Pseudips mexicanus y por ultimo Ips lecontei, siendo estos dos últimos no competitivos para Ips pini, por la presencia baja de individuos. De las dos especies de pinos, la más susceptible en este trabajo fue Pinus durangensis, ya que presentó las tres especies de descortezadores y mayor abundancia que Pinus cooperi.


















