Tema XI. ANABOLISMO · anabolismo heterótrofo de los glúcidos puede iniciarse con moléculas más...
12
1 Tema XI. ANABOLISMO Concepto de anabolismo Panorama general de anabolismo autótrofo Panorama general de anabolismo heterótrofo Anabolismo de los glúcidos - Gluconeogénesis Anabolismo de los lípidos Anabolismo de las proteínas. Anabolismo autótrofo: la fotosíntesis Fase luminosa (procesos primarios) - Naturaleza de la luz - Absorción de la energía luminosa - Formación de energía química libre y poder reductor - Fotofosforilación cíclica: Fase oscura (procesos secundarios). - El ciclo de Calvin: - Reducción y fijación de nitrógeno y azufre - La fotorrespiración Plantas C 3 y C 4 Factores que influyen en la fotosíntesis Anabolismo autótrofo: la quimiosíntesis CONCEPTO DE ANABOLISMO La reacción esquemática de todo fenómeno anabólico es la siguiente: A + B + Energía AB en la cual A y B son sustancias pobres en energía, mientras que el compuesto AB es rico en ella. De acuerdo con esto, en todo proceso anabólico o de síntesis, hay una incorporación de energía que puede tener tres orígenes: Energía luminosa del sol. Los únicos seres que pueden incorporarla son las plantas verdes y algunas bacterias (bacterias verdes y purpúreas). Energía procedente de reacciones químicas exotérmicas que se desarrollan en el medio ambiente. Solamente algunas bacterias (nitrificantes, sulfobacterias y ferrobacterias) pueden utilizar este tipo de energía. Energía química almacenada en determinados compuestos orgánicos fabricados por cualquiera de los seres vivos que pueden captar la energía libre de los dos casos anteriores. Los animales y las plantas no verdes son los organismos que utilizan esta fuente de energía. Los dos primeros tipos de incorporación de energía caracterizan la síntesis o anabolismo autótrofo; el último es propio del anabolismo heterótrofo. PANORÁMICA GENERAL DEL ANABOLISMO AUTÓTROFO La síntesis autótrofa se caracteriza porque partiendo de sustancias inorgánicas e incorporando a las mismas energía libre, son elevadas a la categoría de materia orgánica. Teniendo en cuenta que los componentes básicos de la materia orgánica son el C, O, H, y N, serán los compuestos inorgánicos sencillos de estos elementos los que constituirán el punto de partida del anabolismo autótrofo. Tales compuestos son: para el C, el anhídrido carbónico (CO 2 ); para el Nitrógeno, los nitratos (ion NO 3- ), y para el H y O, el agua. Como consecuencia de ello, dentro del anabolismo autótrofo se pueden distinguir dos tipos fundamentales: el del Carbono (en forma de CO 2 ) y el del Nitrógeno (en forma de NO 3- ). El del Hidrógeno y Oxígeno en forma de agua es simultáneo con el del Carbono.
Transcript of Tema XI. ANABOLISMO · anabolismo heterótrofo de los glúcidos puede iniciarse con moléculas más...

122
1
Tema XI. ANABOLISMO
Concepto de anabolismo
Panorama general de anabolismo autótrofo
Panorama general de anabolismo heterótrofo
Anabolismo de los glúcidos - Gluconeogénesis
Anabolismo de los lípidos
Anabolismo de las proteínas.
Anabolismo autótrofo: la fotosíntesis
Fase luminosa (procesos primarios) - Naturaleza de la luz - Absorción de la energía luminosa - Formación de energía química libre y poder reductor - Fotofosforilación cíclica:
Fase oscura (procesos secundarios). - El ciclo de Calvin: - Reducción y fijación de nitrógeno y azufre - La fotorrespiración
Plantas C3 y C4
Factores que influyen en la fotosíntesis
Anabolismo autótrofo: la quimiosíntesis
CONCEPTO DE ANABOLISMO
La reacción esquemática de todo fenómeno anabólico es la siguiente:
A + B + Energía AB
en la cual A y B son sustancias pobres en energía, mientras que el compuesto AB es rico en ella. De acuerdo con esto, en todo proceso anabólico o de síntesis, hay una incorporación de energía que puede tener tres orígenes:
Energía luminosa del sol. Los únicos seres que pueden incorporarla son las plantas verdes y algunas bacterias (bacterias verdes y purpúreas).
Energía procedente de reacciones químicas exotérmicas que se desarrollan en el medio ambiente. Solamente algunas bacterias (nitrificantes, sulfobacterias y ferrobacterias) pueden utilizar este tipo de energía.
Energía química almacenada en determinados compuestos orgánicos fabricados por cualquiera de los seres vivos que pueden captar la energía libre de los dos casos anteriores. Los animales y las plantas no verdes son los organismos que utilizan esta fuente de energía.
Los dos primeros tipos de incorporación de energía caracterizan la síntesis o anabolismo autótrofo; el último es propio del anabolismo heterótrofo.
PANORÁMICA GENERAL DEL ANABOLISMO AUTÓTROFO
La síntesis autótrofa se caracteriza porque partiendo de sustancias inorgánicas e incorporando a las mismas energía libre, son elevadas a la categoría de materia orgánica.
Teniendo en cuenta que los componentes básicos de la materia orgánica son el C, O, H, y N, serán los compuestos inorgánicos sencillos de estos elementos los que constituirán el punto de partida del anabolismo autótrofo. Tales compuestos son: para el C, el anhídrido carbónico (CO2); para el Nitrógeno, los nitratos (ion NO
3-), y para el H y O, el agua.
Como consecuencia de ello, dentro del anabolismo autótrofo se pueden distinguir dos tipos fundamentales: el del Carbono (en forma de CO2) y el del Nitrógeno (en forma de NO
3-). El del Hidrógeno y
Oxígeno en forma de agua es simultáneo con el del Carbono.

122
2
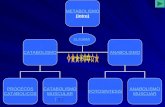
Con el anabolismo del Carbono (al que acompaña siempre el del H y O en forma de agua) se forman monosacáridos, glicerina y ácidos grasos. Con el del Nitrógeno se forma amoniaco, que como radical amino (-NH2) se combina con alguno de los compuestos orgánicos de carácter ácido obtenidos en el anabolismo del Carbono, y se forman aminoácidos. Todas estas sustancias orgánicas sencillas sirven luego para formar principios inmediatos más complejos (polisacáridos, grasas, proteínas, etc.) por mecanismos químicos que son ya muy semejantes a los del anabolismo heterótrofo, pues como en éste, se parte ya de moléculas orgánicas.
Si dentro del anabolismo autótrofo, tenemos en cuenta la fuente que proporciona la energía para transformar las sustancias inorgánicas en orgánicas, podemos distinguir dos tipos de anabolismo autótrofo:
Fotosíntesis, cuando dicha energía procede de la luz solar; puede ser fotosíntesis del Carbono o fotosíntesis del Nitrógeno.
Quimiosíntesis, cuando procede de reacciones químicas exotérmicas; de la misma forma, ésta puede ser del Carbono o del Nitrógeno.
El anabolismo autótrofo es un proceso típicamente reductor. Esta reducción se consigue
porque mediante la energía luminosa (fotosíntesis) o mediante reacciones químicas exotérmicas (quimiosíntesis) se liberan protones (H
+) y electrones (e
-) que transportados por algún enzima se fijan
sobre compuestos inorgánicos diversos a los que reducen, logrando así su incorporación a la materia orgánica. También en todos los casos la energía (lumínica o calórica) es transferida como energía química al ATP para ser de esta forma utilizada en el proceso.
PANORÁMICA GENERAL DEL ANABOLISMO HETERÓTROFO El anabolismo heterótrofo, a diferencia del autótrofo, parte ya de sustancias orgánicas
sencillas, limitándose con ellas a elaborar otras progresivamente más complejas. En los vegetales, el anabolismo heterótrofo viene a ser como la segunda parte de su
anabolismo total, pues como consecuencia de la síntesis autótrofa el vegetal dispone ya de sustancias orgánicas sencillas, con las cuales puede elaborar compuestos orgánicos más complejos por un mecanismo paralelo al que se desarrolla en los seres heterótrofos (animales y plantas sin clorofila), los cuales ya inician su anabolismo con productos orgánicos sencillos, dada su incapacidad para formarlos a partir de la materia inorgánica.
Por tanto, todo lo que se diga sobre el anabolismo heterótrofo es válido, en líneas generales, para ambos grupos de seres vivientes.
ANABOLISMO DE LOS GLÚCIDOS: Los monosacáridos procedentes de la digestión, representados fundamentalmente por
glucosa, son transportados al hígado por la sangre, donde dicho azúcar se transforma en glucógeno que es almacenado. Para llevar a cabo esta transformación, la glucosa se fosforila (es decir, se le une una molécula de ácido fosfórico al carbono 6, formando glucosa 6-fosfato), ya que solamente las moléculas de glucosa fosforilada son capaces de unirse para formar disacáridos y luego el polisacárido glucógeno. Este proceso se denomina glucogenogénesis.
A medida que las células del organismo van necesitando glucosa, el glucógeno del hígado se vuelve a convertir en glucosa 6-fosfato y ésta a su vez en glucosa, que es vertida en la sangre (donde gracias a la insulina se mantiene en una proporción constante del 1 por 1.000) que la hace llegar a todas las células del organismo. El proceso de obtención de moléculas de glucosa a partir del glucógeno se denomina glucogénesis
Estas dos funciones que realiza el hígado de elaborar glucógeno y liberar glucosa, constituyen la llamada función glucogénico-glucémica. Esta doble función está regulada por dos hormonas de acción antagónica: la adrenalina, que incrementa la liberación de glucosa (acción hiperglucemiante), y la insulina, que actúa frenando dicha liberación (acción hipoglucemiante).
Otras partes del organismo, especialmente los músculos, pueden almacenar también glucosa en forma de glucógeno, pero este glucógeno sólo es utilizado para liberar energía en el lugar donde se encuentra sin que pueda ser movilizado hacia otras células.
Si en el organismo ingresan una excesiva cantidad de glúcidos, de tal manera que se satura la capacidad de almacenamiento en el hígado y en los músculos, entonces el exceso de glucosa es transformado en grasa que se almacena en los tejidos, especialmente el tejido adiposo.

122
3
En los vegetales, los monosacáridos obtenidos por fotosíntesis son también anabolizados, formando con ellos reservas de almidón (equivalente al glucógeno animal) que almacenan en forma de gránulos en el interior de sus células.
La glucosa y demás monosacáridos con los que los organismos anabolizan polisacáridos, pueden proceder de los alimentos (animales) o de la fotosíntesis (vegetales), pero en ocasiones el anabolismo heterótrofo de los glúcidos puede iniciarse con moléculas más sencillas que los monosacáridos, concretamente a partir del ácido pirúvico. Este proceso se conoce con el nombre de gluconeogénesis. En líneas generales la obtención de glucosa a partir del ácido pirúvico tiene lugar por una cadena metabólica inversa a la de la glucolisis, en la cual la glucosa se transforma en dos moléculas de ácido pirúvico.
Gluconeogénesis (Biosíntesis de glucosa “de novo”)
La gluconeogénesis es la ruta por la cual los precursores no azúcares (lactato, piruvato, propionato, glicerol y aminoácidos) se convierten en glucosa. Puede llevarse a cabo en el citoplasma o en la mitocondria, dependiendo del lugar en el que se encuentre el precursor utilizado. No debemos de confundirlo con la glucogenolisis, ya que en ésta se forma glucosa pero a partir del glucógeno no es una síntesis “de novo”. Como sabemos, la glucosa ocupa un papel central en el metabolismo, como combustible y como precursor de otros carbohidratos y biomoléculas. El cerebro (una persona normal consume unos 160±20 g de glucosa por día, un 75% por el cerebro) y los eritrocitos dependen de la glucosa como fuente energética (otros son la lente, la córnea, los testículos, la médula del riñón). Sin embargo el hígado solamente puede almacenar glucosa en forma de glucógeno para proporcionar al cerebro glucosa durante solo medio día en condiciones de ayuno (entre 180 y 200 g de glucógeno se almacenan en un individuo normal y los fluidos corporales tienen unos 20 g de glucosa libre). Por lo tanto, durante condiciones de ayuno o metabólicamente parecidas la mayor parte de la glucosa se debe de formar por la gluconeogénesis (nueva síntesis de glucosa) a partir de precursores que no son carbohidratos. Mediante estudios de marcaje, la fuente de la glucosa en condiciones de ayuno por la gluconeogénesis es de cerca del 64% del total de la glucosa presente en sangre (durante las primeras 22 h) y prácticamente toda la glucosa a las 48 h. Por lo tanto, incluso durante unas horas sin alimentarse, la gluconeogénesis es la ruta por la cual la mayor parte de la glucosa se forma en los animales. Tiene lugar principalmente en el hígado (90%) y menos en el riñón (10%).
Los precursores que se pueden convertir en glucosa son: lactato y piruvato, producidos en la glucolisis, intermediarios del ciclo de los ácidos tricarboxílicos y los esqueletos carbonados de la mayoría de los aminoácidos.
La gluconeogénesis es, en esencia, el proceso inverso a la glucólisis; pero conviene aclarar que, si bien todos lo metabolitos intermedios son idénticos en ambos procesos, no ocurre lo mismo con los sistemas enzimáticos que intervienen como catalizadores.
De los nueve enzimas, seis catalizan reversiblemente en sentido glucolítico o gluconeogénico (pasos 2, 4, 5, 6 y 7); pero hay tres que son específicos y catalizan irreversiblemente en sentido glucolítico (1, 3, y 8) o gluconeogénico (1’, 3’ y 8’), de esta forma se hace posible la regulación independiente de los dos procesos, actuando

122
4
(mediante acción hormonal, por ejemplo) sobre la actividad o la síntesis de algunos de estos enzimas. Mediante la gluconeogénesis se sintetiza glucosa a expensas del ácido pirúvico o de cualquier
metabolito que pueda transformarse en dicho ácido. Las reacciones específicamente glucolíticas (1, 3 y 8) son energéticamente favorables y
transcurren irreversiblemente, impulsando “cuesta abajo” la degradación de la glucosa, que suministra un total de 2 ATP. Sin embargo, las reacciones inversas de la gluconeogénesis (1’, 3’ y 8’) son energéticamente desfavorables y requieren un aporte de energía para impulsar “cuesta arriba” el proceso gluconeogénico, que consume 4 ATP + 2 GTP, lo que equivale en total a 6 ATP por cada molécula de glucosa sintetizada (conviene observar que en las reacciones 1’ y 3’ de la gluconeogénesis la hidrólisis del grupo fosfato no se acopla con la síntesis de ATP).
La reacción 8’ es en realidad un conjunto de transformaciones que convierten el ácido piríuvico en forfoenolpirúvico, a través de unas etapas intermedias que transcurren en parte de la mitocondria y en parte en el citosol.
ANABOLISMO DE LOS LÍPIDOS El anabolismo de las grasas puede desarrollarse de dos maneras diferentes: a partir de las
que aportan los alimentos y a partir de los glúcidos. En los vegetales sólo es posible la segunda vía, ya que no son capaces de absorber grasas en su nutrición.
Las grasas procedentes de los alimentos desdobladas durante la digestión en glicerina y ácidos grasos, son reconstruidas nuevamente apenas absorbidas por el intestino. Una pequeña cantidad ingresa en el hígado, donde es almacenada, pero la mayor parte va a través del sistema linfático a almacenarse en los tejidos del cuerpo. Aunque todas las células pueden almacenar algo de grasa, esta función se desarrolla especialmente por células especializadas para este fin, que constituyen los tejidos grasos o adiposos (debajo de la piel, alrededor del corazón y riñones, etc.).
Las grasas pueden formarse en el organismo a expensas de los glúcidos. Este proceso metabólico exige la formación por una parte de glicerina, y por otra de ácidos grasos. La glicerina se forma a partir del 3-fosfogliceraldehido, sustancia que las plantas consiguen en el proceso de la fotosíntesis, y los animales a partir de la glucosa. Los ácidos grasos se forman a partir del acetil-CoA, obtenido a su vez del ácido pirúvico durante el catabolismo de la glucosa, por un proceso metabólico inverso al de la b-oxidación.
ANABOLISMO DE LAS PROTEÍNAS Los vegetales son capaces de fabricar los aminoácidos mediante la fotosíntesis del Nitrógeno,
cosa que no pueden hacer los animales que los absorben tras la digestión de las proteínas. En ambos casos, los aminoácidos deben unirse para formar cadenas de péptidos que
después se unen para formar proteínas. La síntesis de proteínas a partir de aminoácidos constituye un complicado proceso en el que intervienen de una manera directa los ácidos nucleicos y que se desarrolla en dos importantes fases conocidas como transcripción y traducción genética.
ANABOLISMO AUTÓTROFO: LA FOTOSÍNTESIS Puede definirse como un proceso anabólico mediante el cual muchos seres vivos son
capaces de utilizar la energía luminosa para sintetizar materia orgánica a partir de agua, dióxido de carbono, nitratos, sulfatos y fosfatos.
La Fotosíntesis la realizan los siguientes seres vivos: bacterias fotosintéticas, algunos organismos unicelulares (Diatomeas y Dinoflagelados), algas, musgos y vegetales superiores.
La fotosíntesis del carbono es la principal y la mejor conocida, pero no es la única ruta
fotosintética. Fue la primera que se descubrió y su importancia radica en que es el proceso que suministra hidratos de carbono a las partes no fotosintéticas de la planta y a otros organismos. Es, por tanto, el origen de todo lo vivo que hay en la Tierra ya que la energía solar es utilizada para sintetizar los compuestos orgánicos que después serán ingeridos y degradados por los seres vivos no fotosintéticos. Gracias a la fotosíntesis, la energía solar es el origen de la energía química de la célula.
La importancia de la fotosíntesis del carbono también radica en su economía, ya que utiliza
como fuente de energía el sol, como fuente de carbono el CO2 y para reducirlo utiliza agua.

122
5
La reacción general de la fotosíntesis para sintetizar una molécula de glucosa sería
6CO2 + 12H2O + LUZ C6H12O6 + 6O2 + 6H2O
Se trata por tanto de un proceso inverso al catabolismo aerobio de la glucosa que se podría resumir de la siguiente manera: Para sintetizar una molécula de glucosa se precisa una fuente de carbono que es el CO2 cuyos carbonos es preciso reducir combinándolos con hidrógenos, por lo cual se necesita un dador de hidrógenos que es el H2O, la cual al perder sus hidrógenos liberará O2. Todo ello no se realiza espontáneamente sino que requiere un aporte de energía que proviene de la luz solar.
El proceso requiere de una primera etapa en la que se absorbe la energía solar y se transforma en energía química libre ATP y en poder reductor (NADPH), y una segunda etapa donde se utilizan esos productos para la reducción del CO2. Son las denominadas FASE LUMINOSA (procesos primarios) y FASE OSCURA (procesos secundarios).
FASE LUMINOSA O PROCESOS PRIMARIOS: Se produce la captación de energía luminosa y su transformación en energía química libre (ATP) y poder reductor (NADP reducido). Para que esto se produzca es necesaria la presencia de luz.
FASE OSCURA O PROCESOS SECUNDARIOS. Se produce la fijación del CO2 en una molécula aceptora y su posterior reducción para formar hidratos de carbono para lo cual se utilizan los productos obtenidos en la fase anterior. El ATP y el NADPH se utilizan para reducir los compuestos en un alto grado de oxidación (CO2, NO3
-, etc), uno como fuente de energía y el
otro como reductor. No es necesaria la presencia de luz.
FASE LUMINOSA (PROCESOS PRIMARIOS) NATURALEZA DE LA LUZ: La luz visible es una radiación electromagnética con longitudes
de onda entre 400nm (violeta) y 700nm (rojo). Según la teoría cuántica, la luz se propaga de modo discontinuo en forma de fotones que representan paquetes de energía sin masa. El contenido de energía de un fotón es q =h·ν donde h es la constante de Planck y ν la frecuencia, es decir, una variable del movimiento ondulatorio que es inversamente proporcional a la longitud de onda. Por ello, a menor longitud de onda, mayor energía lleva asociada esa radiación.
Las sustancias absorben parte de la luz que reciben y emiten otra parte que se percibe como el color. De la que absorben reciben también la energía de los fotones lo que hace que se produzca una excitación en sus moléculas y un electrón salte a una órbita de mayor energía para volver inmediatamente a su estado fundamental emitiendo la energía absorbida en forma de luz, de calor o ambas cosas a la vez.

122
6
ABSORCIÓN DE LA ENERGÍA LUMINOSA: Las células fotosintéticas poseen una serie de pigmentos localizados en los tilacoides de los cloroplastos. Los más importantes son las clorofilas, aunque existe también una cierta cantidad de pigmentos accesorios (carotenos y xantofilas).
Estos pigmentos se encuentran en la membrana tilacoidal asociados en grupos o conjuntos funcionales que constituyen unidades fotosintéticas llamadas fotosistemas. Cada uno está compuesto por cientos de moléculas de clorofila y carotenoides que actúan como moléculas colectoras que absorben la luz, el llamado complejo antena, y la transmiten como en un embudo hacia una molécula de clorofila especializada que forma el llamado centro de reacción y que al recibir la excitación por la energía transmitida es capaz de perder un electrón excitado que pasa a un nivel energético superior y que será captado por otra molécula cercana. Este hecho supone la transformación de la energía luminosa en energía química.
En las células eucarióticas fotosintéticas y en las cianobacterias hay dos tipos de fotosistemas que se diferencian en el máximo de absorción de la clorofila que forma el centro de reacción. El fotosistema I tiene un máximo de absorción a 700nm y su clorofila se llama P700 (pigmento 700nm), mientras que el fotosistema II tiene el máximo a 680nm (P680). El resto de bacterias fotosintéticas sólo poseen el fotosistema I.
FORMACIÓN DE ENERGÍA QUÍMICA LIBRE Y PODER REDUCTOR: Cuando inciden dos
fotones sobre el fotosistema I, el P700 se excita y emite dos electrones a una órbita de elevada energía que son inmediatamente captados por una proteína ferrosulfurada que lo cede a la ferredoxina. Como el potencial de reducción de esta molécula es muy alto, el NADP
+ acepta los electrones y se reduce a
NADPH.

122
7
¿Cómo se rellena el hueco dejado por los electrones en el P700? Para ello actúa el fotosistema II. Al ser iluminado por dos fotones el P680 se excita y emite dos electrones que son captados por una cadena transportadora de electrones formada por la feofitina (Feo), la plastoquinona (Pq), el complejo de los citocromos b-f y la plastocianina (Pc). El complejo citocromo b-f actúa además como una bomba de protones, utilizando parte de la energía en impulsar protones al espacio tilacoidal. De la plastocianina los electrones pasan ya al P700 que recupera los que justo antes había perdido.
Ahora es el P680 el que presenta un hueco electrónico. Éste es rellenado por los electrones procedentes del agua como consecuencia de la ruptura de la molécula de agua (fotolisis del agua), proceso en el que interviene un enzima con manganeso no muy bien conocido y mediante el cual una molécula de agua se escinde en dos protones, dos electrones y un átomo de oxígeno.
Los protones que libera la fotolisis del agua en el espacio tilacoidal se suman a los que bombea el complejo citocromo b-f con lo que se genera un gradiente electroquímico. Ahora, al igual que ocurre en las mitocondrias según la hipótesis quimiosmótica, los protones son impulsados por el gradiente de pH a través del ATP-sintetasa que se encuentra en la membrana tilacoidal, con lo que se sintetiza ATP a partir de ADP + Pi. Esta síntesis de ATP recibe el nombre de fotofosforilación no cíclica debido a que el flujo de electrones comienza en la molécula de agua y termina en el NADP, es decir, no es un flujo cíclico.
En conjunto, en la transferencia no cíclica, a partir de una molécula de agua y 4 fotones de luz, se forman una molécula de NADPH, una de ATP y ½O2.

122
8
FOTOFOSFORILACIÓN CÍCLICA o ANOXIGÉNICA: Además del flujo no cíclico de electrones, en casi todas las especies tiene lugar también un flujo cíclico en el que interviene sólo el fotosistema I. Se dice que es un proceso cíclico porque los electrones perdidos por el P700 regresan de nuevo a dicho pigmento. La finalidad del proceso cíclico es generar ATP para subsanar el déficit que, en caso contrario, se produciría ya que en la fase oscura se necesitan 3 moléculas de ATP por cada 2 de NADPH.
Al igual que en el proceso no cíclico, al incidir los fotones sobre el fotosistema I, los electrones adquieren la energía necesaria para ser capturados por la ferredoxina, pero ahora, en vez de continuar hacia el NADP, son desviados hacia la cadena de transporte de electrones que conecta los fotosistemas I y II, concretamente al complejo citocromo b-f que bombea protones al espacio tilacoidal para que luego se sintetice ATP al pasar por el canal de la ATP-sintetasa. Se llama ahora fotofosforilación cíclica ya que el flujo de electrones es cíclico, son los mismos que perdió el P700 los que vuelven cíclicamente a él.
Así pues, en este caso no se forma NADPH, no interviene el agua ni se libera O2.
El flujo cíclico de electrones es también característico de las bacterias fotosintéticas que no desprenden oxígeno (bacterias del azufre y purpúreas) ya que no tienen el fotosistema II.
FASE OSCURA (PROCESOS SECUNDARIOS). En la fase oscura se
utilizan el poder energético (ATP) y el poder reductor (NADPH) obtenidos durante la fase luminosa para impulsar la transformación de sustancias inorgánicas en materia orgánica. Esta transformación es muy desfavorable energéticamente y por ello se acopla a las reacciones de la fase luminosa. El proceso fundamental de la fase oscura es la fijación reductora del C a partir del CO2 formándose primero glúcidos sencillos de los que derivarán el resto de compuestos orgánicos (Ciclo de Calvin). Pero además existen otras vías para aquellos compuestos orgánicos que contienen nitrógeno o azufre en las que las plantas utilizan como fuente de estos elementos nitratos (NO3
-) o sulfatos (SO4
2-)
a los que hay que reducir.

122
9
EL CICLO DE CALVIN: La fijación reductora del CO2 ocurre en la mayoría de las plantas a través de una secuencia cíclica de reacciones conocida como ciclo de Calvin o ciclo C3 debido a que la mayoría de los metabolitos intermediarios tienen 3 carbonos. El ciclo se inicia a partir de un enzima de elevado peso molecular, la ribulosa difosfato carboxilasa (RuBisCo), que cataliza la incorporación al ciclo del CO2 atmosférico o disuelto en agua si se trata de plantas acuáticas. Este enzima se encuentra en grandes cantidades en los cloroplastos y se considera que es la proteína más abundante de la Biosfera.
Se distinguen 3 etapas en el ciclo de Calvin:
Fijación: Mediante el enzima RuBisCo el CO2 se fija a un azúcar preexistente, la ribulosa-1,5-difosfato, formándose un compuesto muy inestable de 6 carbonos que se rompe inmediatamente en 2 moléculas de 3-P-glicerato.
Reducción: El 3-P-glicerato se reduce a 3-P-gliceraldehido consumiéndose el NADPH y el ATP que se obtuvieron en la fase luminosa.
Regeneración del aceptor: De las 6 moléculas de 3-P-gliceraldehido que se forman en 3 vueltas del Ciclo de Calvin, una se considera el rendimiento neto de la fotosíntesis porque en cada vuelta se fija solamente un carbono al captar una molécula de CO2. Deben fijarse 3 carbonos para sintetizar una nueva molécula de 3-P-gliceraldehido. Las otras 5 sufren una serie de transformaciones consecutivas en las que también se consume ATP para regenerar la ribulosa 1,5-difosfato, con la que se cierra el ciclo.
REDUCCIÓN Y FIJACIÓN DE NITRÓGENO Y AZUFRE. A partir de sustancias presentes en el ciclo de Calvin se pueden formar ácidos grasos, aminoácidos y nucleótidos.
Los vegetales obtienen el nitrógeno en forma de nitratos (NO3) disueltos en el agua, en lo que se luele llamar sales minerales; su reducción necesaria utiliza la energía obtenida en las reacciones luminosas de la fotosíntesis. El NO3 se reduce primero a nitrito (NO2
-) y luego a amoniaco (NH3). El
amoniaco se incorpora como grupo amino (NH2) para formar aminoácidos y nucleótidos, necesarios para sintetizar proteínas y ácidos nucleícos.
El azufre es captado como sulfato (SO42-), y reducido, primero a sulfito (SO3
2-) y luego a
sulfuro de hidrógeno (H2S), el cual se incorpora como grupo tiol (-SH) a la cisteína.
CICLO DE CALVIN

122
10
LA FOTORRESPIRACIÓN. En el ciclo de Calvin intervienen diversas enzimas, cada paso
requiere la acción de alguna enzima; la más conocida es la llamada Rubisco (Ribulosa 1-5 difosfato carboxilasa-oxidasa), que puede actuar como carboxilasa o como oxidasa según la concentración de CO2.
- Cuando la concentración de CO2 es mayor que la de O2 el enzima actúa como carboxilasa, fija una molécula de CO2 en la ribulosa 1- 5 difosfato y da lugar a dos moléculas de ácido 3 fosfoglicérico.
- Cuando la concentración de CO2 es menor que la de O2, el enzima actúa como oxigenasa y da lugar a la rotura de la molécula de ribulosa 1-5 difosfato, que se transforma en una molécula de ácido 3-fosfoglicérico incorporada al Ciclo de Calvin, y otra de ácido fofoglicólico. Este último compuesto se transforma en los peroxisomas mediante un proceso (vía glicólica) en el que intervienen las mitocondrias y mediante el cual, parte de los átomos de carbono que contiene se libera a la atmósfera en forma de
CO2, lo que supone una pérdida de eficacia del proceso fotosintético. En condiciones
atmosféricas normales, hasta el 50% del carbono fijado durante la
fotosíntesis puede ser reoxidado a CO2 durante la fotorrespiración. Así, la
fotorrespiración reduce en gran medida la eficiencia fotosintética de algunas
plantas.
PLANTAS C3 Y C4 La mayoría de las plantas son C3, son las más primitivas y comprenden al grupo más numeroso,
pues incluyen a la mayoría de las plantas y a todas las algas. Se llaman así porque el CO2 penetra en el estroma de los cloroplastos y se combina con un compuesto de 5 C, la ribulosa 1-5 difosfato, para dar un compuesto intermedio de 6 C, muy inestable, que se escinde inmediatamente en dos moléculas de ácido 3-fosfoglicérico. Estas moléculas tienen 3 C y son los primeros compuestos estables que se detectan inmediatamente después de la fijación del CO2.
Las plantas C4 incluyen en numerosas especies adaptadas a climas secos y calurosos, como las cañas, papiros, el maíz, el mijo, etc.; así como las plantas crasas y cactus propias de climas desérticos.
La actividad oxigenasa de la ribulosa 1-5 difosfato carboxilasa aumenta más rápidamente con la temperatura que la actividad carboxilasa. ¿Cómo evitan las plantas tropicales la elevada velocidad de fotorrespiración inútil?
Su solución es conseguir una elevada concentración de CO2 localizada en el sitio en el transcurre el ciclo de Calvin en sus células fotosintéticas. Para eso, el CO2 atmosférico se difunde al interior de las hojas y penetra, en primer lugar, en los cloroplastos del mesófilo. Aquí comienza el ciclo de Hatch-Slack, de manera que el CO2 se combina con el ácido fosfoenol pirúvico para formar oxalacético (el enzima que cataliza este proceso es mucho más activo que la Rubisco); esta es la molécula C4 (que da nombre a estas plantas) que se desplaza a las células de la vaina perivascular, donde sufre descarboxilación, para regenerar de nuevo ácido fosfoenol pirúvico (vuelve a las células de mesófilo para cerrar el ciclo), y el CO2 liberado es captado por la enzima Rubisco para incorporarlo al Ciclo de Calvin como las plantas C3.

122
11
La estructura anatómica de las hojas de las plantas C4 presenta una distribución característica adaptada a la función que desempeña; alrededor de los vasos transportadores de savia se disponen dos coronas de células concéntricas: las células del mesófilo (donde se forma el compuesto C4) y la vaina perivascular o células envolventes del haz (donde tiene lugar el Ciclo de Clavin). Las interrelaciones metabólicas de las dos coronas permiten que la fijación del CO2 transcurra en dos etapas sucesivas. Primero en el ciclo de Hatch-Slack y, posteriormente en el Ciclo de Calvin.
Gracias al dispositivo de las plantas C4, la concentración de CO2 en las células de la vaina perivascular (el único lugar donde se produce el Ciclo de Calvin) es unas cincuenta veces superior a la que existe en las plantas C3, con lo cual aumenta la eficacia de la fotosíntesis al eliminarse casi por completo la actividad fotorrespiratoria de la Rubisco. Podría considerarse que el ciclo de Hatch-Slack se comporta “como una activa nodriza que suministra grandes cantidades de CO2 a la perezosa Rubisco, con el fin de disminuir al máximo sus caprichos fotorrespiratorios”.
Observese que, cuando la vía C4 libera el CO2 al ciclo de Calvin, por cada hexosa formada se consumen 30 ATPs, a diferencia de los 18 ATPs que se consumían en ausencia de la vía C4. La alta concentración de CO2 en las células de la vaina del haz de las plantas C4, debida al consumo de 12 ATPs de más, es crítica para conseguir una rápida velocidad fotosintética, porque, cuando la luz es abundante, el CO2 es precisamente el factor limitante de esta velocidad. Una concentración elevada de CO2 tambien reduce al mínimo la perdida de energía debida a fotorrespiración.
Las plantas C4 tienen ventaja en un ambiente cálido y muy iluminado, lo que justifica su prevalencia en los trópicos. Las plantas C3, que consumen solamente 18 ATPs por hexosa, formada en ausencia de fotorrespiración (comparado con los 30 ATPs de las plantas C4), son más eficaces a temperaturas inferiores a 28ºC, y por tanto predominan en los ambientes menos cálidos.
La Rubisco, surgió pronto en la evolución, cuando la atmósfera era rica en CO2 y casi libre de O2. Inicialmente, el enzima no se seleccionó para trabajar en un ambiente como el actual, rico en O2. En esencia, la via C4 crea un microcosmos en el que abunda el CO2, lo que para la fotosíntesis significa un retorno al paraíso.

122
12
FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
Intensidad luminosa: La actividad fotosintética aumenta con la intensidad luminosa hasta alcanzar un límite máximo característico de cada especie. Para una misma intensidad luminosa, el rendimiento fotosintético es superior en las plantas adaptadas a climas secos y calurosos.
Concentración de CO2: La actividad fotosintética aumenta conforme va creciendo la concentración de CO2, hasta alcanzar un límite en el que se estabiliza.
Temperatura: Como toda actividad enzimática, la fotosíntesis aumenta con la temperatura hasta alcanzar un límite máximo (variable según las especies de climas cálidos, templados o fríos), por encima del cual se produce la desnaturalización de los enzimas.
Fotoperíodo: El rendimiento está en relación directa a las horas de exposición a la luz que tenga la planta.
Humedad ambiental: Cuando hay escasez de agua, los estomas (aberturas de la epidermis de las zonas verdes de las plantas superiores) se cierran para evitar pérdidas de agua por transpiración, lo cual dificulta el paso de CO2 y la actividad fotosintética disminuye.
Concentración de O2: Al aumentar baja el rendimiento fotosintético debido a las pérdidas por fotorrespiración.
Color de la luz: Si se ilumina la planta con luz roja, con longitud de onda superior a 680 nm, no actúa el fotosistema II, sólo se estimula el fotosistema I y se produce fotofosforilación cíclica, con lo que el rendimiento fotosintético es mucho menor. Este hecho se denomina declive del rojo.
ANABOLISMO AUTÓTROFO: LA QUIMIOSÍNTESIS. OTRA FORMA DE NUTRICIÓN AUTÓTROFA
La quimiosíntesis es también una forma de nutrición autótrofa en la que, a diferencia de la fotosíntesis, la energía y los electrones (ATP y NADPH) necesarios para los procesos de anabolismo van a proceder de la oxidación de sustancias inorgánicas.
Se trata de una forma de nutrición típicamente bacteriana. En la que las diferentes especies se han especializado en la oxidación de distintos substratos. Según el substrato oxidado tendremos:
• Bacterias quimiosintéticas nitrificantes. Están muy ampliamente distribuida en suelo y aguas.
Unas (Ej.: Nitrosomonas) oxidan amoniaco a nitrito (NO2-) y otras (Nitrobacter) prosiguen la
oxidación hasta nitrato (NO3-). Ambos géneros se complementan y comparten el mismo hábitat.
Por ello, de ambas depende, en parte, que se cierre el ciclo del nitrógeno y contribuyen a que los suelos sean ricos en nitrato, compuesto que las plantas asimilan para sintetizar aminoácidos.
• Bacterias quimiosintéticas del azufre. Estas bacterias oxidan azufre a sulfitos o a sulfatos. Se usan para desalcalinizar suelos muy calcáreos. Se añade azufre , y las bacterias producirán sulfúrico que transforma el carbonato cálcico en sulfato cálcico (yeso) que se elimina lavando las tierras.
• Bacterias del hierro. Oxidan los compuestos ferrosos a férricos.
Estos dos últimos tipos de bacterias crecen, sobre todo en los yacimientos de
azufre y hierro de origen volcánico y en particular en los llamados humeros negros. Es de destacar, que las bacterias quimiosintéticas son los únicos seres vivos no dependientes, ni
directa ni indirectamente, de la luz solar.