TEMA: NUTRICIÓN CELULAR I CATABOLISMO · La respiración es un tipo de catabolismo total, en el...
26
1 TEMA: NUTRICIÓN CELULAR I CATABOLISMO INTRODUCCIÓN El metabolismo es el conjunto de reacciones químicas que permiten la transformación de los nutrientes (para producir energía, para producir materia propia para crecimiento o renovación o para producir funciones como contracción muscular...) en el interior de la célula. Los nutrientes son los elementos necesarios para realizar las funciones vitales de la célula a través de un proceso metabólico. Son nutrientes los bioelementos, el agua, las sales minerales, glúcidos, lípidos, el oxígeno molecular..., es decir, todos aquellos átomos o moléculas necesarias para el funcionamiento del organismo. No son siempre los mismos en todas las células u organismos, por ejemplo para las células fotosintéticas el CO 2 es un nutriente neccesario para el proceso metabólico de la fotosíntesis, mientras que para una célula heterótrofa el CO 2 es un desecho producido en un proceso metabólico como la respiración celular o en la fermentación alcohólica. Incluso en la misma célula fotosintética cuando no realiza la fotosíntesis (por la noche) produce CO 2 en sus mitocondrias que al no usarse para la fotosíntesis el CO 2 es un desecho en este caso y no un nutriente. Otro ejemplo sería el O 2 que es un nutriente para organismos aerobios que hacen la respiración celular, mientras que resulta tóxico para microorganismos anaerobios estrictos. CONCEPTO DE NUTRICIÓN CELULAR Todos los seres vivos necesitan materia y energía para vivir: El objeto de la nutrición celular es: Incorporar Materia y Energía Transformarla en el interior mediante una serie de reacciones químicas Eliminar los residuos que se producen en las reacciones anteriores. . De este modo, las células, y en general los organismos, consiguen obtener materia propia para crecer o renovarse y la energía necesaria para realizar cualquier actividad. 1. TIPOS DE NUTRICIÓN SEGÚN LA FUENTE DE MATERIA Y ENERGÍA Según la fuente de materia tenemos la nutrición autótrofa y la nutrición heterótrofa. En la nutrición autótrofa, la fuente de carbono es el CO 2 y en la heterótrofa la fuente de carbono son moléculas orgánicas. Nutrición autótrofa : Las sustancias inorgánicas como CO 2 , H 2 O y sales minerales son nutrientes a partir de los cuales formarán moléculas orgánicas; para lo que se necesita un aporte de energía. Materia y energía se incorporan por separado. ENERGÍA Sustancias inorgánicas (H 2 O, CO 2 , Sustancias orgánicas sencillas (glicerol, aa, glucosa, ac. Grasos…..)
Transcript of TEMA: NUTRICIÓN CELULAR I CATABOLISMO · La respiración es un tipo de catabolismo total, en el...

1
TEMA: NUTRICIÓN CELULAR I
CATABOLISMO INTRODUCCIÓN
El metabolismo es el conjunto de reacciones químicas que permiten la transformación de los nutrientes (para producir energía, para producir materia propia para crecimiento o renovación o para producir funciones como contracción muscular...) en el interior de la célula.
Los nutrientes son los elementos necesarios para realizar las funciones vitales de la célula a través de un proceso metabólico. Son nutrientes los bioelementos, el agua, las sales minerales, glúcidos, lípidos, el oxígeno molecular..., es decir, todos aquellos átomos o moléculas necesarias para el funcionamiento del organismo. No son siempre los mismos en todas las células u organismos, por ejemplo para las células fotosintéticas el CO2 es un nutriente neccesario para el proceso metabólico de la fotosíntesis, mientras que para una célula heterótrofa el CO2 es un desecho producido en un proceso metabólico como la respiración celular o en la fermentación alcohólica. Incluso en la misma célula fotosintética cuando no realiza la fotosíntesis (por la noche) produce CO2 en sus mitocondrias que al no usarse para la fotosíntesis el CO2 es un desecho en este caso y no un nutriente. Otro ejemplo sería el O2 que es un nutriente para organismos aerobios que hacen la respiración celular, mientras que resulta tóxico para microorganismos anaerobios estrictos.
CONCEPTO DE NUTRICIÓN CELULAR Todos los seres vivos necesitan materia y energía para vivir:
El objeto de la nutrición celular es:
Incorporar Materia y Energía
Transformarla en el interior mediante una serie de reacciones químicas
Eliminar los residuos que se producen en las reacciones anteriores.
. De este modo, las células, y en general los organismos, consiguen obtener materia propia para crecer o renovarse y la energía necesaria para realizar cualquier actividad.
1. TIPOS DE NUTRICIÓN SEGÚN LA FUENTE DE MATERIA Y ENERGÍA
Según la fuente de materia tenemos la nutrición autótrofa y la nutrición heterótrofa. En la
nutrición autótrofa, la fuente de carbono es el CO2 y en la heterótrofa la fuente de carbono son moléculas orgánicas.
Nutrición autótrofa: Las sustancias inorgánicas como CO2, H2O y sales minerales son nutrientes a partir de los cuales formarán moléculas orgánicas; para lo que se necesita un aporte de energía. Materia y energía se incorporan por separado. ENERGÍA
Sustancias inorgánicas
(H2O, CO2,
Sustancias orgánicas sencillas (glicerol,
aa, glucosa, ac. Grasos…..)

2
Según la fuente de esta energía se distinguen dos tipos de nutrición autótrofa: fotosíntesis y quimiosíntesis.
Fotosíntesis: es el tipo de nutrición autótrofa en el que las sustancias inorgánicas (CO2, H2O y sales minerales) son transformadas en orgánicas (glúcidos, proteínas...) mediante el aporte de energía lumínica.
Células en las que aparecen: células con pigmentos Organismos formados por est tipo de células: Protoctistas (algas), Metafitas (plantas), Bacterias.
Quimiosíntesis: es el tipo de nutrición autótrofa en el que las sustancias inorgánicas (CO2, H2O y sales minerales) son transformadas en orgánicas (glúcidos, proteínas...) mediante el aporte de energía química procedente de la oxidación de moléculas químicas inorgánicas. Organismos quimioautótrofos son únicamente algunos tipos de bacterias, por ejemplo las bacterias del metano que oxidan el metano (CH4) a CO2 obteniendo ATP.
Los organismos autótrofos tienen un gran interés biológico dado el papel de los autótrofos en los ciclos biogeoquímicos, pues mantienen el ciclo de la materia en los ecosistemas, ya que son los productores que están en la base de todas las relaciones tróficas aportando materia y energía para que pueda ser usada por los siguientes niveles tróficos.
Nutrición heterótrofa: en la nutrición heterótrofa, la fuente de carbono son moléculas orgánicas más o menos complejas que contienen ya energía en sus enlaces químicos, por lo que la materia y la energía se incorporan juntas. También usan sustancias inorgánicas.
Organismos heterótrofos animales, hongos, protozoos y muchas bacterias (todas las bacterias menos las fotosintéticas).
2. ETAPAS DE LA NUTRICIÓN CELULAR
A. INCORPORACIÓN DE NUTRIENTES B. TRANSFORMACIÓN DE NUTRIENTES (METABOLISMO) C. ELIMINACIÓN DE PRODUCTOS DE DESECHO
A. INCORPORACIÓN DE NUTRIENTES
Se realiza por diferentes mecanismos según el tamaño y la naturaleza de las sustancias (hidrofílicas o hidrofóbicas). En células autótrofas: Agua: osmosis Gases : CO2 y O2 por difusión simple Iones: (sales minerales) : Difusión facilitada (permeasas y canales iónicos)
-Transporte pasivo (sin gasto energético) a favor de gradiente -Transporte activo ( con gasto energético) en contra de gradiente
En células heterótrofas: Además de sustancias inorgánicas como las anteriores incorporan materia orgánica. Sustancias orgánicas sencillas: (glucosa, aminoácidos……) Mediante transporte mediado por receptor, en el caso de moléculas hidrofílicas Sustancias orgánicas complejas: (polisacáridos, proteínas, grandes partículas, fragmentos celulares………) Mediante procesos de endocitosis (fagocitosis, pinocitosis, endocitosis mediada por receptor)

3
B. TRANSFORMACIÓN DE NUTRIENTES ( METABOLISMO) Metabolismo: Conjunto de reacciones químicas que permiten la transformación de los nutrientes en el interior de la célula, para producir energía o para formar materia propia para el crecimiento o renovación, o para realizar otras funciones por ejemplo para la contracción muscular, transmisión del impulso nervioso... Reacciones para formar materia propia: Anabolismo Reacciones para obtener energía: Catabolismo
METABOLISMO: CONCEPTOS, TIPOS, CARACTERÍSTICAS Y FUNCIONES
Rutas metabólicas: Secuencias de reacciones químicas que están ligadas entre sí, siendo
el producto de una reacción, el sustrato de la siguiente.
Metabolitos: cada una de las moléculas intermediarias que aparecen en las reacciones
metabólicas.
El qué las reacciones ocurran en un determinado orden es porque son procesos controlados
por:
Enzimas (proteínas catalizadoras)
Acoplamiento de reacciones: Las reacciones endergónicas, absorben la energía
producida en las reacciones exergónicas
Transportadores energéticos que llevan energía de las reacciones exergónicas a las
endergónicas (ATP)
Coenzimas de oxido- reducción (NAD2H, FAD2H…….)
TIPOS DE RUTAS METABÓLICAS:
Rutas catabólicas=Catabolismo: Degradación enzimática de moléculas orgánicas
complejas aa moléculas orgánicas sencillas. Son reacciones exergónicas (se desprende
energía= ATP). Son reacciones de oxidación. (perder H o perder electrones).
Rutas anabólicas=Anabolismo: Síntesis enzimática de moléculas orgánicas complejas a
partir de moléculas sencillas.(orgánicas, en anabolismo heterótrofo. Inorgánicas en
anabolismo autótrofo). Son reacciones energónicas o endotérmicas (Absorben energía
=ATP). Reacciones de reducción. (ganancia de H o electrones). El anabolismo y el catabolismo están interconectados y se necesitan el uno al otro, ya que si se
producen oxidaciones en unas moléculas ( NAD+ o FAD+ NADH+H+ o FADH2 . Se obtienen
coenzimas reducidos)para reducir sustancias en el anabolismo (el anabolismo regenera coenzimas coenzimas oxidados) y si unos procesos necesitan energía (anabolismo), otros deben proporcionar esa energía (catabolismo).
H2 -
H2-
X+Y XY
XY X+Y
ADP+P ATP NADH+H NAD

4
CATABOLISMO ANABOLISMO
Son reacciones de DEGRADACIÓN, es decir, de moléculas más grandes se obtienen moléculas más pequeñas.
Son reacciones de SÍNTESIS, es decir, de moléculas más pequeñas se obtienen moléculas más grandes.
Es un proceso DE OXIDACCIÓN que produce energía (ATP).
Es un proceso DE REDUCCIÓN que necesita energía (ATP).
Son rutas metabólicas convergentes, es decir, partiendo de sustancias muy diferentes acaban produciendo las mismas sustancias. Ejemplo: CO2 y H2O
Son rutas metabólicas divergentes, es decir, partiendo de las mismas sustancias acaban produciendo sustancias muy diferentes.
Ejemplo de ruta catabólica es la respiración: materia orgánica + O2 → CO2 + H2O + energía (ATP)
Ejemplo de ruta anabólica es la fotosíntesis: CO2 + H2O + energía luminosa → materia orgánica + O2
CARACTERÍSTICAS DE LAS REACCIONES METABÓLICAS:
Las reacciones metabólicas están acopladas energéticamente mediante el ATP pues las reacciones catabólicas liberan energía mientras que las anabólicas requieren energía. Esta energía se transfiere del catabolismo al anabolismo almacenada en la molécula de ATP.
ATP ↔ ADP + Pi + Energía (7,3 KCAL) Hay dos formas de obtener ATP:
1- Fosforilación a nivel de sustrato: consiste en que una molécula que tiene unido un grupo fosfato se lo cede al ADP formando ATP.
X-P + ADP → X + ATP 2- Fosforilación oxidativa en este caso el ADP se une a un fosfato inorgánico (que no
estaba unido a ninguna molécula orgánica), esto sucede en la membrana mitocondrial interna, a nivel de las ATP sintasas de las crestas mitocondriales, aprovechando la energia liberada de las reacciones de oxidorreducción.
3- Fotofosforilación: el ADP se une a un fosfato inorgánico,para formar ATP, en la
membrana de los tilacoides donde hay ATPasas o partículas F. La Energía procede del sol que a su vez produce reacciones de oxidorreducción.
Las reacciones metabólicas son reacciones de oxido-reducción catabolismo es un proceso de oxidación (libera H+ o electrones) anabolismo es un proceso de reducción (absorbe H+ (o electrones)) Se puede decir por tanto que el catabolismo es un proceso de deshidrogenación y el anabolismo lo es de hidrogenación.

5
Las sustancias que transportan los hidrógenos son coenzimas como NAD+, FAD, FMN, NADP... formando NADH+ H+, FADH2, FMNH2, NADPH2... Oxido-reducción: Transferencia de electrones entre dos sustancias. Oxidación: perdida de electrones, aumento del número de oxidación. perdida de hidrógeno o incorporación de oxígeno. Reducción: ganancia de electrones disminución del número de oxidación, ganancia de hidrógeno o perdida de oxígeno. Un potencial redox muy electronegativo significa que tiene mucha facilidad para oxidarse es decir para oxidarse. Cuando los electrones son cedidos de una molécula con un potencial mas electronegativo a otra con un potencial redox menos electronegativo se libera energía, en el caso de las células se forma ATP. oxidación Fe2+ ⎯⎯⎯→ Fe3+ + e-
reducción Cl + e- ⎯⎯⎯⎯→ Cl-
Las reacciones metabólicas tienen una secuencia encadenada y catalizada por enzimas: Una vía o ruta metabólica consiste en una sucesión de reacciones encadenadas en las que se generan intermediarios metabólicos o metabolitos. Cada reacción está catalizada por una enzima diferente. Las rutas metabólicas pueden ser lineales (como la glucólisis) o circulares (como el ciclo de Krebs. Las lineales a su vez pueden ser convergentes como en el catabolismo o divergentes como en el anabolismo.
Existen mecanismos de autorregulación de estas rutas como inhibición por producto final o retroalimentación.
Las reacciones metabólicas están compartimentalizadas pues las distintas vías metabólicas se sitúan en zonas celulares diferentes, debido a que las enzimas que necesitan están en diferentes lugares de la célula. En el metabolismo tienen lugar muchas reacciones (muchas vías o rutas), que tienen lugar de forma simultánea, y para evitar interferencias entre ellas (por ejemplo que interfieran enzimas que hacen reacciones opuestas) cada una ocurre en un compartimento celular (en un orgánulo), es decir, las rutas están compartimentalizadas, y con ello la eficacia enzimática es mayor. Por ejemplo:
o Citoplasma: Glucólisis, gluconeogénesis, glucogenogenesis, síntesis de triglicéridos y de proteínas (traducción).
o Mitocondria: Ciclo de krebs, -oxidación, fosforilación oxidativa. o Retículo endoplasmático: síntesis de lípidos y de proteínas. o Núcleo: duplicación y transcripción.

6
C. ELIMINACIÓN DE PRODUCTOS DE DESECHO Moléculas que son o que pueden resultar tóxicas. Los residuos no digeridos salen desde el endosoma por exocitosis, en células animales se le denomina defecación celular. En células vegetales, las sustancias tóxicas, reaccionan entre sí y se acumulan en el interior de las vacuolas en forma de nicotina cafeína(Sustancias que resultan menos tóxicas) Las células vegetales eliminan agua, dióxido de carbono y amoniaco Las células animales eliminan agua, dióxido de carbono, amoniaco y productos de defecación celular.
3. TIPOS DE CATABOLISMO: RESPIRACIÓN CELULAR AERÓBICA Y FERMENTACIONES
La respiración es un tipo de catabolismo total, en el que la materia orgánica se descompone totalmente hasta moléculas inorgánicas liberando gran cantidad de energía (ATP), ya que la oxidación es completa. El más conocido es la respiración celular aerobia o aeróbica donde el oxígeno es el que recoge los H+ y electrones liberados en la oxidación formándose agua (½O2 + 2H+ + 2e- → H2O ). La respiración incluye una cadena de transporte de electrones y el proceso de fosforilación oxidativa.
En algunas bacterias existe la respiración anaeróbica donde no usan oxígeno sino que el aceptor final de los H+ y electrones liberados en las oxidaciones es otra molécula inorgánica distinta al oxígeno, por ejemplo algunas bacterias usan el azufre: S + 2H+ + 2e- → H2S.
Producen moléculas inórganicas distintas del agua (CO 2 y CH4)
La fermentación es un tipo de catabolismo parcial, en el que la materia orgánica solo se degrada y oxida parcialmente dando otras moléculas orgánicasmás sencillas. Como es una oxidación parcial se obtiene poca cantidad de energía (poco ATP) y los H+ y electrones liberados en la oxidación los acepta la propia molécula orgánica resultante de la oxidación. Además en la fermentación no hay cadena de transporte de electrones ni fosforilación oxidativa.

7
RESPIRACIÓN FERMENTACIÓN
Catabolismo completo (oxidación total) Catabolismo parcial (oxidación incompleta)
Se obtiene mucho ATP (38) Se obtiene poco ATP(2)
El aceptor final de los hidrógenos (H+ y e-) es el oxígeno.
El aceptor final de los hidrógenos (H+ y e-) es la propia molécula orgánica resultante de la oxidación
Posee cadena de transporte de electrones y fosforilación oxidativa
No posee cadena de transporte de electrones ni fosforilación oxidativa
Sucede en la mitocondria (en bacterias con respiración celular la cadena de transporte de electrones y la fosforilacion oxidativa sucede en la membrana plasmática.(mesosomas))
Sucede en el citoplasma
Con Ciclo de Krebs en la matriz mitocondrial (en bacterias con respiración celular el ciclo de Krebs es en el citoplasma)
Sin Ciclo de Krebs
Células eucariotas y procariotas En procariotas, algunas eucariotas cómo células músculares y levaduras.
Clasificación de las células según su catabolismo
Según el tipo de catabolismo que realizan las células, estas pueden clasificarse en células
aerobias, anaerobias estrictas y anaerobias facultativas. Las células anaerobias estrictas no solo no usan el oxígeno en su catabolismo, sino que además les resulta tóxico, estas células realizan el proceso catabólico de la fermentación; las células aerobias necesitan el oxigeno porque realizan el proceso catabólico de la respiración celular aerobia y las células anaerobias facultativas pueden vivir con o sin oxígeno ya que pueden hacer tanto fermentaciones como respiración celular aerobia, según la disponibilidad de oxígeno en el medio (les interesa más hacer la respiración porque obtiene mucho más ATP, pero si no hay oxígeno realizará fermentaciones).

8
4. RESPIRACIÓN AEROBIA
Resumen de la respiración celular aerobia
El catabolismo de las diferentes macromoléculas (glúcidos, lípidos y proteínas) converge en la
formación de Acetil-CoA en la matriz mitocondrial. El grupo acetil del Acetil-CoA entra en el ciclo de Krebs donde se oxida totalmente hasta CO2. Todos estos procesos catabólicos han producido coenzimas reducidos (NADH+ H+ y FADH2) que aportan los H+ y electrones a la cadena de transporte de electrones, la cadena de transporte de electrones bombea los H+ al espacio intermembrana produciendo un gradiente quimioosmótico de H+ que permitirá la fosforilación oxidativa (formación de ATP a partir de ADP + Pi) en las partículas F de la membrana mitocondrial
interna. El último aceptor de los H+ y electrones es el oxígeno formándose agua: 2H+ + 2e- + O2 H2O.
Materia orgánica + O2 → CO2 + H2O + Energía (ATP)
Las céluas usan cómo moléculas energética principalmente glúcidos y lípidos. La ruta central del catabolismo aerobio es el catabolismo de glúcidos.
5. CATABOLISMO DE GLÚCIDOS
A. ETAPA PRELIMINAR (DEGRADACIÓN DE POLISACÁRIDOS PARA PRODUCIR GLUCOSA) (CITOPLASMA)
B. GLUCOLISIS ( DEGRADACIÓN DE GLUCOSA A PIRÚVICO) (CITOPLASMA) C. ETAPA INTERMEDIA (TRANSFORMACIÓN DE PIRÚVICO A ACETIL COA)(MATRIZ
MITOCONDRIAL) D. CICLO DE KREBS (MATRIZ MITOCONDRIAL) E. FOSFORILACIÓN OXIDATIVAA O CADENA DE TRANSPORTE ELECTRÓNICO
(MEMBRANA INTERNA MITOCONDRIAL)

9
A. ETAPA PRELIMINAR A partir de de polisacáridos formar glucosa que intervenga en la siguiente fase
Vegetales:
Por descomposición de almidón en maltosa, dextrinas, glucosa, por acción de la - amilasa
(ruptura enlaces a (1-4)) . A continuación actuarían la (1-6) glucooxidasa para las ramificaciones y la la maltasa para romper la matosa. Otras fuentes de glucosa en vegetales:
- Fotosíntesis - A partir de lípidos transformados en glucosa mediante el ciclo del ácido glioxílico en
los glioxisomas
Animales:
Puede provenir de la digestión en el tubo digestivo por acción de la -amilasa y las mismas enzimas anteriores. Si el aporte de glucosa en la dieta no es suficiente, los animales recurren a sus reservas de glucógeno que hay en músculo e hígado. La función del glucógeno muscular y hepático es diferente. El glucógeno muscular se usa a corto plazo para la contracción muscular y se consume rápidamente. El hepático, para suministrar glucosa a otros tejidos. El proceso de degradación del glucógeno es mediante GLUCÓGENOLISIS OTRAS FUENTES DE GLUCOSA SON DISACÁRIDOS INGERIDOS EN LOS ALIMENTOS (LACTOSA, SACAROSA….)
GLUCÓGENO (n- glucosas)
Glucógeno
Fosforilasa
GLUCÓGENO(n-1 glucosas) + GLUCOSA-1P
(1-6)
Glucooxidasa
GLUCOSA
GLUCOSA +P
GLUCOSA-6P
CONTRACCIÓN
MUSCULAR
SANGRE
Hígado
Músculo
10
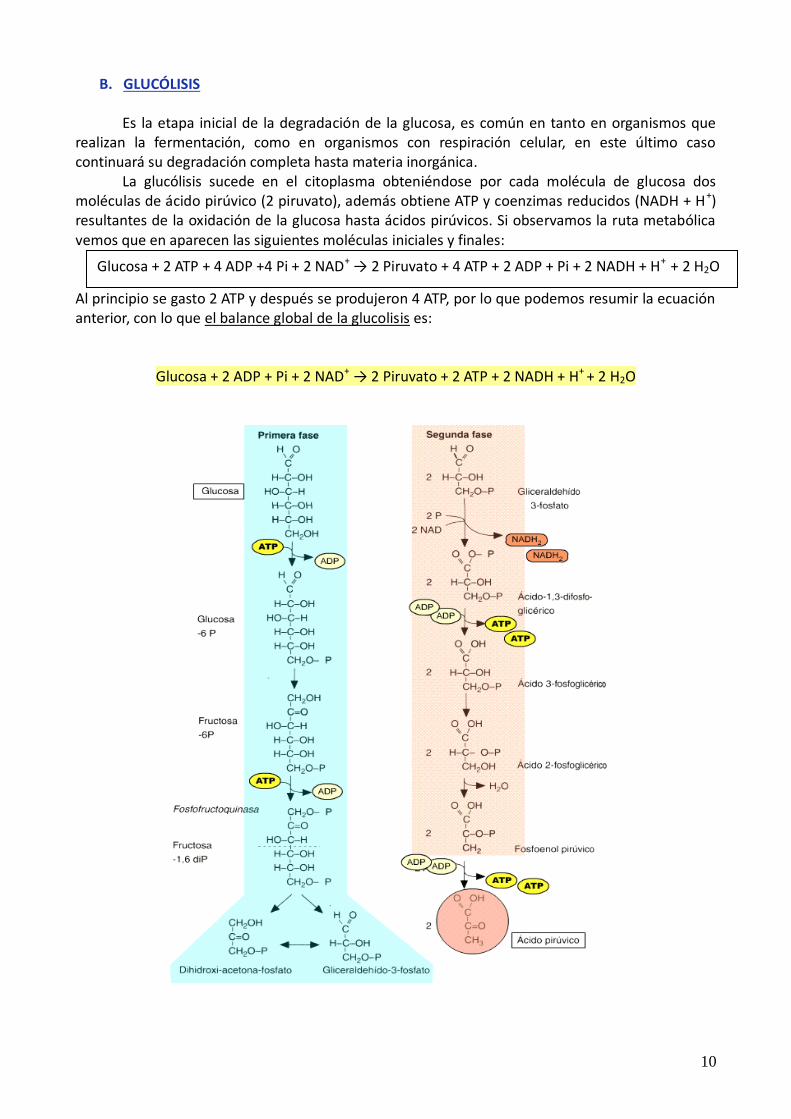
B. GLUCÓLISIS Es la etapa inicial de la degradación de la glucosa, es común en tanto en organismos que realizan la fermentación, como en organismos con respiración celular, en este último caso continuará su degradación completa hasta materia inorgánica. La glucólisis sucede en el citoplasma obteniéndose por cada molécula de glucosa dos moléculas de ácido pirúvico (2 piruvato), además obtiene ATP y coenzimas reducidos (NADH + H+) resultantes de la oxidación de la glucosa hasta ácidos pirúvicos. Si observamos la ruta metabólica vemos que en aparecen las siguientes moléculas iniciales y finales: Al principio se gasto 2 ATP y después se produjeron 4 ATP, por lo que podemos resumir la ecuación anterior, con lo que el balance global de la glucolisis es:
Glucosa + 2 ADP + Pi + 2 NAD+ → 2 Piruvato + 2 ATP + 2 NADH + H+ + 2 H2O
Glucosa + 2 ATP + 4 ADP +4 Pi + 2 NAD+ → 2 Piruvato + 4 ATP + 2 ADP + Pi + 2 NADH + H+ + 2 H2O

11
Las etapas de control de la glucolisis son:
De Glucosa a Glucosa6P: La hexoquinasa es inhibida por el producto de la reacción, la G-6-
P y activada por Pi. La isoenzima de la hexoquinasa en hígado se llama glucoquinasa y
tiene menor afinidad por la glucosa que la HK, luego tendrá una KM más alta
Fructosa6P a Fructosa 1,6DP: es la enzima clave en el control de la glucolisis; es una
enzima alostérica y está regulada por metabolitos activadores (F-2,6-BP, AMP) y otros
inhibidores (ATP, citrato, H+ ).
Fosfoenol pirúvico a ácido pirúvico : La piruvato quinasa es inhibida por el ATP, ALA,
Acetil-CoA y los ácidos grasos de cadena larga. Los últimos pueden proporcionar ATP a
través del Ciclo de Krebs. Es activada por F1,6-BP. En hígado es inhibida por fosforilación.
Metabolismo de Fructosa-2,6-bisfosfato Las tres etapas son irreversibles, pero las dos primeras sólo usan otra enzima para reversivilizar la reacción mientras que la tercera tiene que ir por una via alternativa.

12
Incorporación de otros glúcidos a la glucolisis o vías alimentadoras de la glucolisis
C. ETAPA INTERMEDIA
VÍAS ALTERNATIVAS DEL ÁCIDO PIRÚVICO
La glucólisis es común en la fermentación y en la respiración celular, por lo que hay dos vías o rutas metabólicas posibles para los ácidos pirúvicos obtenidos en la glucólisis: la formación de Acetil-CoA y la fermentación. El primer caso sucede en organismos capaces de realizar la respiración celular en presencia de oxígeno (organismos aerobios y anaerobios facultativos en presencia de oxígeno) y el segundo caso sucede en ausencia de oxígeno (organismos anaerobios).
- Formación de Acetil-CoA o etapa intermedia: El pirúvico atraviesa la membrana interna y externa de la membrana mitocondrial. Esta etapa se produce en la matriz mitocondrial y consiste en una descarboxilación oxidativa
del ácido pirúvico, es decir, pierde un C en forma de CO2, al mismo tiempo que se oxida perdiendo hidrógenos obteniendo NADH + H+. Por último, el coenzima A (HS-CoA ) se une a la molécula de 2 carbonos (grupo acetilo) resultante, dando Acetil-CoA que irá al ciclo de Krebs. Por cada ácido pirúvico se obtiene la siguiente ecuación:
Piruvato + NAD+ + HS-CoA → Acetil-CoA + CO2 + NADH + H+
La enzima que cataliza el proceso es la piruvato deshidrogenasa complejo enzimático de gran tamaño que realiza las tres funciones: descarboxilación, oxidación y unión del grupo acetilo al coenzima A Esta reacción es irreversible, cuando la cantidad de ATP en la célula es alta, se detiene la actuación del sistema, ya que su función es proporcionar combustible al ciclo de Krebs para obtener energía. El coA-SH= molécula de compuesta por: nucleótido de adenosina, molécula de ácido pantoténico y una etilamina unida al grupo tiol (SH)
EL Acetil-CoA tenemos que multiplicarlo todo por 2 porque de la glucólisis se obtienen 2 ácidos pirúvicos por cada glucosa.
En organismos anaerobios, al no haber respiración celular, los ácidos pirúvicos siguen una vía
de fermentación necesaria para reciclar el NAD+.

13
D. CICLO DE KREBS
También llamado : CICLO DE LOS ÁCIDOS TRICARBOXÍLICOS O CICLO DEL ÁCIDO CÍTRICO. Es la vía para la oxidación completa de glúcidos, ácidos grasos y proteínas.
Sucede en la matriz mitocondrial. Comienza con la unión del Acetil-CoA con el ácido oxalacético que posee 4 carbonos, dando lugar a una molécula de 6 carbonos y el coenzima A vuelve a quedar libre en la matriz (para poder seguir formando Acetil-CoA).
Cosiste en la oxidación de los grupos metilo del acetil CoA a dos moléculas de CO2 y la transferencia de los hidrógenos a los coenzimas NAD+ y FAD+
El ciclo se cierra regenerando el ácido oxalacético . En resumen, por cada Acetil- CoA que entra en el ciclo de Krebs se obtienen 3 NADH + H+, 1 FADH2, 1 GTP (ATP) y 2CO2 (el GTP equivale a un ATP porque se forma ATP a partir de él).
Acetil-CoA → 3 NADH + H+ + 1 FADH2 + 1 GTP (ATP) + 2CO2
1. citrato sitetasa: condesa el acetilCoa con oxalacético para formar citrato
2. Citrato isomerasa: el citato se isomeriza a isocítrico
3. IsocitratoDH Descarboxilación oxidativa del isocitrato
4. -cetoglutarato DH: descarboxilación oxidativa del
- cetoglutarato 5. SuccinilCoA sintetasa:
formación de GTP, fosforilación a nivel de sustrato.
6. SuccinatoDH: Oxidación del Succinico
7. Fumarasa:formación de malato.
8. MalatoDH: oxidación del malato para regenerar oxalacetato.
De estas reacciones 2 son descarboxilaciones y 4 oxidaciones. Durante el ciclo se liberan 4 pares de átomos de hidrógeno de los metabolitos, 3 de los cuales son captados por NAD+ , y 1 captado por FAD+ . Se origina GTP por fosforilación a nivel de sustrato que se puede convertir enzimáticamente en ATP
GTP+ADP GDP+ATP

14
Debido al pH fisiólogico del interior de la matriz mitocondrial la mayor parte de los grupos ácidos se encuentran ionizados, por eso hablamos de : Fumarato, succinato……..
La velocidad del ciclo está controlada por:
- ADP= Es un modulador positivo; Aumenta su velocidad - ATP y NADH+H+=Moduladores negativos; Bajan la velocidad del ciclo
En el ciclo de Krebs no se consume O2, pero es necesaria su disponibilidad para que los coenzimas (NAD2H y FAD2H) puedan ceder sus H y así regenerarse. Este ciclo es el centro del metabolismo aerobio, aquí confluyen la mayor parte de los procesos catabólicos (glucosa, ácidos grasos, proteínas), pero también producen intermediarios de algunas rutas anabólicas como el oxalcetato que se utiliza para la síntesis
del ácido aspártico (aa), succinilcoA (síntesis del grupo hemo) y cetoglutarato (síntesis ácido glutámico = aa) Por eso el ciclo de Krebs es un ciclo anfibólico, es decir áctua cómo ruta anabólica y catabólica. Cómo por cada glucosa producimos 2 acetilCoA, serán necesarias 2 vueltas de ciclo. Balance hasta ahora:
glucolisis glucosa 2 pirúvicos+ 2ATP+2NAD2H
Intermedia 2 pirúvicos 2AcetilCoA+2NAD2H+2CO2
Ciclo de Krebs 2AcetilCoA 2(3NAD2H+FAD2H+GTP)
Global
Glucosa 10 NAD2H + 2FAD2H +4 ATP
Todos los coenzimas reducidos producidos en el ciclo de Krebs se encuentran en la matriz mitocondrial y serán utilizados por la cadena respiratoria. Hay que recordar que en la glucolisis también se produjeron coenzimas reducidos que están en el citoplasma. La membrana interna mitocondrial es impermeable a este coenzima y para poder llegar a la matriz mitocondrial va utilizar un sistema de trnasporte que se denominan lanzaderas de sustrato.
C4
C6
C5
C4
C4
C4
CO2
NAD2H
GTP CO2
FAD2H
C2
NAD2H
NAD2H

15
LANZADERAS Existen 2 tipos de lanzaderas: Lanzadera glicerol- fosfato: en células de músculo y cerebro Lanzadera malato- aspartato: en células de hígado y corazón LANZADERA GLICEROL- FOSFATO: Los electrones del NADH+H+ son cedidos al FAD2H de la matriz mitocondrial
LANZADERA MALATO- ASPARTATO
DHAP
DHAP
G3P
G3P H2
22
H2
22
Membrana interna
mitocondrial
citoplasma
NAD+
NAD2H
FAD FAD2H
Matriz mitocondrial
MALATO
OO ASPARTATO.
AT H2
22
H2
22
Membrana interna
mitocondrial
citoplasma
NAD+
NAD2H
NAD2H NAD2H
Matriz mitocondrial
ASPARTATO.
AT MALATO
OO

16
E. CADENA DE TRANSPORTE DE ELECTRONES Y FOSFORILACIÓN OXIDATIVA. TEORÍA QUIMIOOSMÓTICA
La cadena de transporte de electrones y la fosforilación oxidativa suceden en la membrana mitocondrial interna si son células eucariotas y en la membrana celular en procariotas (MESOSOMAS). La cadena de transporte de electrones es un conjunto de moléculas que se encuentran en la membrana interna de la mitocondria que permiten mediante una serie de reacciones de oxidación-reducción transportar los electrones desde los coenzimas reducidos hasta el oxígeno. La finalidad es la síntesis de ATP. En la membrana interna mitocondrial existen una serie de transportadores de electrones, que los transfieren de acuerdo a su potencial redox. Los electrones pasan del transportador electrónico que tiene el potencial redox más electronegativo (mayor energía) hacia el que tiene un potencial redox más positivo (menos energía). Las moléculas transportadoras de la cadena de transporte de electrones son principalmente proteínas que se encuentran ordenadas en posiciones fijas; son 6 componentes, 4 de ellos son grandes complejos protéicos (se nombran en números romanos: I, II, III y IV), otro es una pequeña molécula lipídica: la ubiquinona Q que transporta los electrones de los complejos I y II al III, y por último, una pequeña proteína: el citocromo C que transporta electrones del complejo III al IV.
El hecho de que cada molécula acepte electrones de la molécula anterior es debido a que en cada molécula los electrones van ocupando posiciones de menor energía, por lo que van liberando energía cada vez que pasan a la molécula siguiente.

17
El NADH + H+ cede sus hidrógenos al complejo I ( Complejo NADH- deshidrogenasa) situado en un nivel energético mayor que el resto de los complejos, este se los cede a la ubiquinona
(coenzima Q) que es una flavoproteína. La diferencia de potencial existente en esta oxidorreducción es suficiente para bombear protones desde la matriz al espacio intermembrana.
Complejo III (complejo citocromo b-c) Acepta los electrones de la ubiquinona y los tras lada hasta el citocromo c que actúa como intermediario. (ya no se aceptan protones sólo electrones, los protones quedan fuera de la cadena) . También se crea la energía suficiente para bombear protones al espacia perimitocondrial.
Complejo IV (complejo citocromo oxidasa) Toma los electrones del citocromo c y los cede al
oxígeno que actúa como último aceptor de electrones. La energía liberada también permite transportar protones en contra de gradiente.
Cuando ½ O2 capta los dos electrones se obtiene oxígeno ionizado O2- que tiene gran apetencia por los H+libres en el medio y forma H2O.
Cómo resultado de la fosforilación oxidativa del NAD2H se forman 3 ATP. ½ O2 +NAD2H H2O + NAD++ 3 ATP El FADH2 cede los hidrógenos al complejo II (complejo succinato deshidrogenasa) y este a la
ubiquinona, situado en el nivel energético inferior, cómo los dos son flavoproteínas (sus potenciales redox son parecidos), no existe la diferencia de potencial suficiente cómo para transportar protones al espacio perimitocondrial. El resto del transporte es igual que para el NAD2H. (complejo III y complejo IV)
½ O2 +FAD2H H2O + FAD++ 2ATP La teoría quimioosmótica afirma que la energía liberada en la cadena de transporte de
electrones (al ir los electrones ocupando posiciones de menor energía) es utilizada por los complejos I, III y IV para bombear protones (H+) al espacio intermembrana, formando un gradiente electroquímico entre el espacio intermembrana con gran cantidad de H+ y la matriz con poca cantidad de H+. Además del bombeo de protones por los complejos I, III y IV, otro factor que disminuye los H+ en la matriz y que por tanto, aumenta este gradiente electroquímico, es la retirada de H+ de la matriz por el oxígeno para formar agua.
Este gradiente electroquímico generado posee energía potencial acumulada que se utiliza para formar ATP, ya que la membrana mitocondrial interna es impermeable a los H+ y estos sólo pueden volver a la matriz a través del canal de las partículas F o ATPsintetasas; esto produce un movimiento de rotación y el giro del tallo dentro del rosetón provoca un cambio conformacional en sus unidades en la ATPsintetasa que permite la formación de ATP a partir de ADP y Pi (la parte que gira es la F1 y es la energía cinética del giro la que produce la formación de ATP). A esto se le conoce como fosforilación oxidativa. ( trnasformación de energía potencial en energía química).

18
La ATP sintetasa esta formada por 9 cadenas proteicas: - 3 hidrófobas, incluidas en la membrana=F 0 - 6 Hidrófilas que forman una cabeza esférica que sale de la membrana mitocondrial hacía
la matriz = F 1 Se calcula que por cada NADH + H+ que entra en el transporte de electrones se obtienen 3 ATP (en los libros más actuales de bioquímica afirman que son 2,5 ATP). Como el FADH2 se incorpora a la cadena de transporte de electrones en un nivel energético más bajo se obtienen sólo 2 ATP (en los libros de bioquímica 1,5 ATP) .
BALANCE ENERGÉTICO DEL CATABOLISMO DE LA GLUCOSA ( USANDO LANZADERA GLICEROL
FOSFATO) Glucosa + O2 → CO2 + H2O + Energía (38 ATP)
Lugar Ruta metabólica
Número de coenzimas reducidos
ATP obtenidos por fosforilación oxidativa
ATP obtenidos por fosforilación a nivel de sustrato
ATP total
Citoplasma Glucólisis 2 NAD2H
2 ATP 2ATP
Matriz Mitocondrial
2 ác. Pirúvicos a Acetil Co-A ( etapa intermedia)
2NAD2H
Matriz Mitocondrial
2 Acetil Co-A al ciclo de Krebs
6NAD2H 2FAD2H
2 GTP 2 GTP
Crestas mitocondriales
Cadena de transporte electrónico
3 ATP por NAD2H 2 ATP por FAD2H
Total
10 NAD2H 2 FAD2H
30 ATP 4 ATP
4ATP 38
Si el proceso ocurre en hígado o corazón se usa la lanzadera malato- aspartato y se obtienen
36 ATP Hidrólisis de 1 mól de glucosa(180 g) genera 277 Kcal ( 38 ATP por 7,3 Kcal) En laboratorio se ha medido que la combustión total de la glucosa es de 686 Kcal. El rendimiento sería de un 40%.

19
Regulación de la fosforilación oxidativa: Se basa en la relación ADP/ATP; NAD/NAD2H y presión parcial de oxígeno (pO2)
Si la relación ADP/ATP es alta aumenta la velocidad del proceso; eso indica que la relación NAD/NAD2H aumenta Si la relación ADP/ATP es baja la velocidad del proceso; eso indica que la relación NAD/NAD2H disminuye Una alta presión parcial de oxígeno hace que la velocidad de proceso aumente. Factores inhibidores y desacopladores: Inhibidores: Bloquean algún lugar específico de la cadena de transporte impidiendo la transferencia de electrones y la fosforilación oxidativa Desacopladores: impiden el transporte de protones, lo que hace que se genere menos ATP pero aumenta la velocidad de la cadena y la energía del transporte de electrones se va en forma de calor. En algunas células se produce desacoplamiento, para producir calor (tejido adiposos de los animales que hibernan). Las células de grasa parda producen calor en vez de ATP cuando oxidan sus reservas de grasa . Este calor les ayuda a despertar. RUTA DEL FOSFOGLUCANATO O DE DESVIACIÓN DE LAS HEXOSAS Ruta de degradación con función de biosíntesis: proporciona NADP2H y ribosa-5-fosfato para reacciones de biosíntesis, pero también puede degradar glucosa, o pentosas de los nucleótidos procedentes de la hidrólisis de los ácidos nucleicos de la dieta, hasta CO2 y agua. Tiene dos fases: La fase oxidativa genera por cada molécula de glucosa; 2 moléculas de NADPH, 1molécula de
ribulosa-5-fosfato y una molécula de CO2.
La fase no oxidativa convierte 3 azúcares fosfato de 5 carbonos en 2 azúcares fosfato de 6
carbonos y 1 azúcar fosfato de 3 carbonos
Balance global
3 glucosa 6-P + 6 NADP+ + 3 H2O → 2 fructosa 6-P + gliceraldehído 3-P + 6 NADPH + 6 H+ + 3
CO2
La principal función de esta vía no es degradar glucosa sino generar NADP2H necesario para la
biosíntesis de ácidos grasos y colesterol en hígado. En músculo no existe esta via porque no generan
ácidos grasos.
Esta vía también es utilizada para convertir hexosas en pentosas necesarias para la síntesis de
nucleótidos.
En eritrocitos el NADP2H se utiliza para reducir el agua oxigenada y evitar su acción oxidante
sobre lípidos y proteínas y asi conseguir que no se rompa la membrana.

20
6. FERMENTACIONES
El objetivo principal de las fermentaciones es eliminar los hidrógenos del NADH + H+ para
tener NAD+ con el que poder seguir haciendo la glucólisis y así obtener el ATP que necesitan (en la respiración celular los NAD+ se reciclan al cederse los hidrógenos del NADH + H+a la cadena de transporte de electrones).
Son procesos catabólicos anaerobios de degradación de glucosa. El aceptor final de protones es una molécula orgánica, diferente según el tipo de
fermentación. Es un catabolismo parcial de la materia orgánica, los productos finales aun contienen cierta
cantidad de energía en sus enlaces. Tienen un rendimiento energético escaso. Se obtiene energía por fosforilación a nivel de sustrato. En la industria hay veces que se usa el término fermentación para cualquier proceso que tiene
lugar en tanques. Hablando de fermentación anoxidativa cuando no requieren aireación y oxidativa cuando requieren aireación.
Las fermentaciones oxidativas no son auténticas fermentaciones ya que interviene el oxígeno.
Fermentación acética para producción de vinagre.
En la fermentación láctica el NADH + H+ cede los hidrógenos directamente al ácido pirúvico
obteniendo ácido láctico; esta fermentación la realizan las bacterias del yogur como Streptococcus lactis o Lactobacillus sp. O Leuconostoc. Se emplea en la obtención de derivados lacteos como: Mantequilla, yogur, cuajada, queso……
Nuestras células musculares, en caso de sobreesfuerzo en los que falta oxígeno pueden
realizar la fermentación láctica. En condiciones anaerobias el piruvato se acumula y la velocidad de reacción de la glucolisis tiende a cero, para que la glucolisis pueda continuar e necesario eliminar
el pirúvico y para eso se utiliza la fermentación láctica.
El rendimiento es mucho menor pero por lo menos se puede seguir produciendo energía a partir de la glucosa.
El aumento de concentración del ácido láctico produce una bajada de pH, lo que ocasiona fatiga muscular. El ácido láctico puede llegar a cristalizar y producir micro roturas en el tejido muscular (agujetas).
Después de esta situación de ejercicio, cuando hay aporte de oxígeno suficiente parte del lactato puede reciclarse a piruvato, el resto del láctico va a al hígado donde se convierte en glucosa mediante
gluconeogénesis. La glucosa puede regresar al músculo vía sanguínea.

21
Fermentación alcohólica el ácido pirúvico se descarboxila (pierde un CO2) dando una molécula de dos carbonos que es la que recibe los hidrógenos del NADH + H+ formándose etanol;
la fermentación alcohólica la realiza la levadura Saccharomyces cerevisiae que son los microorganismos que se utilizan para fabricar las bebidas alcohólicas (cerveza, vino, sidra...) y los productos de repostería (bizcochos, galletas, rollos...).
Actualmente la industria química utiliza para la repostería industrial levadura química que les sale más barato aunque nutricionalmente es peor para el consumidor (entre otras cosas por la mayor cantidad de azúcares presentes por no haber microorganismos que los fermenten).
Otras fermentaciones: Fermentación propiónica: para la producción de algunos quesos. Prpionibacterium Fermentación fórmica: Enterobacterias- E. coli Fermentación butírica: sepuede producir durante l proceso de ensilado. Clostridium. Fermentación pútrida: Descomposición de cadáveres y restos vegetales. Proteínas y aa CO2 +NH3 +sustancias orgánicas degradadas. En general las fermentaciones tienen importancia en los ecosistemas, ya que realizan procesos
de descomposición. El olor y el sabor de algunos quesos y vinos se producen de esta manera.
7. CATABOLISMO DE LÍPIDOS
El proceso de saponificación para degradar las grasas ocurre en el tubo digestivo y en el citoplasma. Triglicéridos Glicerol + 3 ac. grasos El glicerol y los ácidos grasos que forman las grasas, una vez separados, siguen caminos distintos en su degradación:
- Catabolismo de la glicerina: la glicerina en el citoplasma se transforma fácilmente en un intermediario de la glucólisis (gliceraldehído 3P o dihidroxiacetona fosfato) al oxidarse y añadirse
un Pi, con lo cual se incorpora a la glucólisis.
lipasas

22
- Catabolismo de los ácidos grasos o β-oxidación de los ácidos grasos: los ácidos grasos se degradan en la matriz mitocondrial mediante la llamada β-oxidación de los ácidos grasos, pero primero, antes de entrar en la mitocondria, el ácido graso debe activarse.
La activación del ácido graso consiste en añadirle un CoA (HS-CoA) con gasto de 2 ATP (en realidad se usa solo un ATP pero en lugar de dar ADP + Pi da AMP + 2Pi por lo que equivale al gasto de 2 ATP) formando un acil-CoA (ácido graso con un CoA).Esto ocurre en el citoplasma.
Una vez activado atraviesa la membrana mitocondrial unido a un transportador: la carnitina. Una vez en la matriz se inicia la β-oxidación del ácido graso.
La β-oxidación de los ácidos grasos consiste en oxidar el carbono β (es el carbono 3) de -CH2 a
-C=O, los hidrógenos de esta oxidación son recogidos por los coenzimas NAD+ y el FAD, al carbono β posteriormente se le une un CoA, lo que ocasionará por un lado la liberación de un acetil-CoA (irá al ciclo de Krebs) y por otro lado la formación de un acil-CoA con dos carbonos menos.
Este proceso se repite hasta que el ácido graso se quede sin carbonos, al repetirse el proceso es como una hélice (al tener en cada vuelta dos carbonos menos se va estrechando), por eso recibe el nombre de hélice de Lynen en honor a uno de sus descubridores.
El balance total es de un NADH + H+
, un FADH2 y un acetil-CoA por cada vuelta en la hélice de Lynen, es decir, por cada dos carbonos que tenga el ácido graso excepto los dos últimos carbonos (dan acetil-CoA pero no coenzimas reducidos al no tener carbono β) y menos 2 ATP gastados en la activación del ácido graso.
Los coenzimas reducidos van a la cadena de transporte de electrones de la respiración celular de la mitocondria y el acetil-CoA va al ciclo de Krebs
Hay que tener en cuenta que en la activación del ácido graso usamos dos CoASH, el primero sería un CoASH citoplásmico y el 2º un CoASH mitocondrial ya que la membrana interna mitocondrial es impermeable al CoASH.

23
Etapas de la oxidación: 1- El acilCoA sufre una oxidación
a cargo de FAD.(Formación molécula doble enlace.
2- Hidratación (OH y H se incorporan a ambos lados del doble enlace)
3- 2ª oxidación a cargo NAD ( se oxida el grupo alcohol a grupo cetónico)
4- Hidrólisis: Formación de acetilCoA que va al ciclo de Krebs y un ácido graso con (n-2) atomos de carbono, que se vuelve a incorporar a otra hélice de oxidación. Hasta que queden sólo un acilCoA de 2 átomos de carbono (acetilCoA)
La oxidación en mamíferos puede ocurrir en mitocondrias y en peroxisomas, mientras que en levaduras sólo ocurre en peroxisomas. Los ácidos grasos de número impar se degradan de la misma forma pero el último resto es de propionilCoA que se transforma en ácido succínico, el cual se incorpora al ciclo de Krebs. En situación de ayuno, o hipoglucemias, se acumulan muchas moléculas de acetilCoA provenientes
de la oxidación de ácidos grasos( en células del hígado), estos acetilCoA se unen entre sí formando los cuerpos cetónicos que salen a la sangre y son utilizados como combustible en los tejidos periféricos(Corazón y cerebro) RENDIMIENTO ENERGÉTICO DE UN ÁCIDO GRASO EJ: Acido Capróico (6 átomos de carbono)
Lugar Ruta metabólica
REACCIONES
citoplasma Activación ácido graso
CH3-(CH2)4-COOH+2ATP CH3-(CH2)4-CO-S-CoA
Matriz mitocondrial
-oxidación (2 )
CH3-(CH2)4-CO-S-CoA 3CH3-CO-SCoA+ 2FAD2H+2NAD2H
Matriz mitocondrial
CICLO DE kREBS
3CH3-CO-SCoA 9NAD2H+3FAD2H+6CO2+3GTP
TOTAL
CH3-(CH2)4-COOH 11NAD2H+5FAD2H+6CO2+1ATP
Lugar Ruta metabólica
REACCIONES
CRESTAS MITOCONDRI-ALES
TRANSPORTE ELECTRÓNICO FOSFORILACION OXIDATIVA
11NAD2H+11/2 O2 33ATP+ H2O 5 FAD2H+5/2 O2 10ATP+ H2O

24
Ejercicio: Calcula el ATP obtenido (indirectamente) al degradar el ácido palmítico que es un ácido graso de 16C. ¿Cuál sería el rendimiento energético de una molécula de triestearina( triglicérido con 3 moléculas de ácido esteárico (18 C))? Conclusiones finales: El rendimiento energético de un ácido graso de 6 átomos de carbono es mayor que el de la glucosa que tiene l mismo número de átomos de carbono. Las grasas tienen un gran valor como combustibles metabólicos especialmente en células animales, son la principal reserva energética(Triglicéridos), se acumulan en los adipocitos del tejido adiposo. ¿Porqué se almacena más grasas que glucógeno? El almacenar más gras ue glucógeno se debe a que a igualdad de peso las grasas, proporcionan mucha más energía que el glucógeno. Además el glucógeno es una molécula hidrofílica por lo que pesará más que un lípido que es hidrofóbico. Así si nuestra reserva mayoritaria fuese glucógeno en vez de lípidos incrementaríamos nuestro peso en aproximadamente 30 Kg. Necesitaríamos mayor cantidad de tejido esquelético y más energía para moverlo. En plantas como no se mueven no importa tanto el aumento de peso por lo que acumulan principalmente almidón. En el caso de las semillas que han de ser dispersadas, supone una ventaja acumula lípidos. Por eso muchas especies, ricino, girasol……. Almacenan aceites. En condiciones normales un adulto acumula glucógeno para un dia mientras que las grasas suministran energía para más de un mes de actividad normal. En situación de ayuno casi todo el acetilCoA proviene de las grasas, mientras que el de después de comer proviene de la glucosa. Los azúcares se transforman en grasas por tanto un aumento de azúcares implicaría un aumento de grasas. Las células prefieren los azúcares a las grasas para catabolizar por varias razones:
- La movilización de grasas es más lenta. - En condiciones anaerobias las grasas no pueden metabolizarse mientras que los
azúcares si. - Los ácidos grasos no pueden transformarse en grasas al revés si. - Los ácidos grasos necesitan mecanismos especiales de transporte en sangre mientras
que la glucosa.
8. CATABOLISMO DE PROTEÍNAS Las proteínas no se suelen degradar para producir energía, salvo en caso de ayuno prolongado
o por un exceso de proteínas en la dieta. Aspectos generales La digestión de
proteínas de la dieta comienza en el estómago. La ingesta de proteínas estimula la síntesis de la hormona gastrina que estimula la liberación de pepsinógeno, forma inactiva de la pépsina. El proceso continua en el intestino delgado por acción de las peptidasas pancreáticas: quimotripsina y tripsina y otras enteropeptidasas.

25
Los aa se absorben en los enterocitos del intestino y se distribuyen por la sangre hasta los órganos y tejidos. La degradación de aa excedentes supone que el grupo α-amino se convierta en urea para su excreción, mientras que los esqueletos carbonados se transformen a Acetil-CoA, piruvato o intermediarios del ciclo del Ac. Cítrico y la energía consiguiente de su oxidación. La cadena carbonada de los AA podrá transformarse finalmente en cuerpos cetónicos (AA cetogénicos) o glucosa (AA glucogénicos).
Los aminoácidos deben seguir dos pasos para su degradación: primero se les debe quitar el grupo amino (-NH2) y segundo el resto carbonado que queda sin el grupo -NH2 se transforma según el aminoácido (ver imagen) del que procede en ácido pirúvico o en acetil-CoA o en un intermediario del ciclo de Krebs, incorporándose a las rutas catabólicas que correspondan (formación de acetil-CoA o ciclo de Krebs).
REACCIONES GENERALES DE AMINOACIDOS: transaminacion, desaminacion y
descarboxilacion La pérdida del grupo -NH2 se realiza por transaminación y desaminación. Transaminación es
que el grupo -NH2 pasa a otra molécula (dando glutamato) que posteriormente eliminará el -NH2 en el hígado por una reacción llamada desaminación formando amoniaco (NH3) o amonio (NH4
+). El NH3 o NH4
+ (en nosotros) va al ciclo de la urea transformándose en urea (en animales uricotélicos se transforma en ácido úrico y en animales amoniotélicos el NH3 o NH4
+ se elimina directamente al exterior).
El tercer tipo de reacciones que se pueden producir son de descarboxilación: R-CH-COOH R-CH2-NH2+CO2 NH2
Ciclo de la urea Ocurre parte en el citoplasma y parte en la mitocondria.
descarboxilasa

26
9.CATABOLISMO DE ÁCIDOS NUCLÉICOS No tiene finalidad energética. Por acción de las nucleasas (se encuentran en el jugo intestinal) se rompen los enlaces fosdiester entre nucleótidos y estos pueden reutilizarse en síntesis de ARN y ADN.
VISIÓN GLOBAL DEL CATABOLISMO: El catabolismo glúcidico es la vía central, ya que aquí confluyen el de proteínas, ácidos grasos y ácidos nucléicos. La mitocondria es la central energética de la célula ya que es a nivel de la fosforilación oxidativa cuando se sintetiza más ATP. El ATP sintetizado en la mitocondria sale hacia el citoplasma y otros orgánulos para ser utilizado en reacciones de síntesis, transporte a nivel de membrana, movimiento, ciclosis…….. Los procesos catabólicos tienen consecuencias nocivas para la célula, ya que se forman radicales libres (moléculas muy oxigenadas y muy reactivas que resultan agresivas y tóxicas), que rompen membranas y son la causa del envejecimiento celular. Por eso el efecto saludable de antioxidantes como la vitamina C.
Nucleótidos
ácido fosfórico
reutilización formación ATP
eliminación por l orina
pentosa metabolismo de glúcidos ruta del fosfoglucanato
bases nitrogenadas
púricas
pirimidínicas
Fomación urea
Ácido úrico