Tema 8 el catabolismo
142
El catabolismo utrición celular: Metabolismo 1
-
Upload
rosa-berros-canuria -
Category
Education
-
view
1.337 -
download
1
Transcript of Tema 8 el catabolismo

El catabolismo
Nutrición celular: Metabolismo 1

Características

El metabolismo celularEs el conjunto de procesos que tienen lugar en la célula.Mediante ellos, unas moléculas se transforman en otras.El objetivo es obtener materia y energía para llevar a cabo las funciones de nutrición, relación y reproducción.Así se mantiene diferente al entorno.Si pierde la capacidad para realizar estas reacciones, se iguala al entorno, es decir: muere.

Materia y energíaLa materia se utiliza para:
crecer (incluido el desarrollo embrionario). renovar las estructuras (renovar células muertas, tejidos dañados, etc.).
La energía se almacena en los enlaces de la materia orgánica compleja y se usa:
moverse. mantener la temperatura. realizar reacciones endotérmicas (que requieren energía).

Reacciones metabólicasLas reacciones del metabolismo se denominan rutas metabólicas.Las moléculas que intervienen se llaman metabolitos.La sustancia inicial se llama sustrato.Las sustancias finales de una ruta se llaman productos. Las vías metabólicas pequeñas, que enlazan otras grandes se denominan metabolismo intermedio.

Las reacciones metabólicas pueden ser de dos tipos:
Exergónicas o exotérmicas. Se realizan de forma espontánea. Son reacciones de degradación Desprenden energía que se almacena en los enlaces del ATP. Los productos tienen menos energía que los reactivos. Estas reacciones se engloban en la parte del metabolismo llamada Catabolismo.
Reacciones metabólicas

Endergónicas o endotérmicas. No se realizan de forma espontánea, sino con gasto de energía. Son reacciones de síntesis. Requieren un aporte de energía, que se obtiene rompiendo los enlaces del ATP. Los productos tienen más energía que los reactivos. Estas reacciones se engloban en la parte del metabolismo llamada Anabolismo.
Reacciones metabólicas

CATABOLISMO
ANABOLISMO· Son reacciones de
degradación.· Son reacciones de
oxidación.· Desprenden energía.· A partir de muchos
sustratos diferentes se forman casi siempre los mismos productos, principalmente dióxido de carbono, ácido pirúvico y etanol.
· Es un conjunto de vías metabólicas convergentes.
· Son reacciones de síntesis.· Son reacciones de
reducción.· Precisan energía.· A partir de unos pocos
sustratos se pueden formar muchos productos, diferentes.
· Es un conjunto de vías metabólicas divergentes.
Etapas del metabolismo

Nutrición celular Según la materia y energía utilizada, se distinguen distintos tipos de metabolismo
Materia
Energía
Inorgánica(autótrofos)
Orgánica(heterótrofos)
Solar(fotótrofos) fotoautótrofos fotoheterótrofos
Química(quimiótrofos) quimioautótrof
osquimioheterótrof
os

Diferencias entre células autótrofas y heterótrofas
Las células autótrofas presentan dos secuencias de reacciones anabólicas:
en primer lugar, anabolismo autótrofo: paso de materia inorgánica a materia orgánica simple (fotosíntesis o quimiosíntesis) en segundo lugar, anabolismo heterótrofo: síntesis de moléculas orgánicas complejas a partir de moléculas orgánicas sencillas.
Las células heterótrofas sólo tienen anabolismo heterótrofo.El catabolismo es idéntico en ambos tipos de células.

CARACTERÍSTICAS DE LAS
REACCIONES METABÓLICAS

Características 1- Son reacciones de oxidación-
reducción (redox) lo que supone: Están acopladas energéticamente a través del ATP. Requieren transportadores de electrones (NAD, NADH, FAD, etc).
2- Están encadenadas3- Están compartimentadas4- Están catalizadas por enzimas

1-OXIDACIÓN REDUCCIÓN

Son reacciones de oxidorreducción Una molécula se oxida si pierde electrones, al tiempo que otra molécula gana esos electrones y se reduce.La oxidación y reducción se pueden llevar a cabo por: OXIDACIÓN REDUCCIÓN
Pérdida de e-, conlleva aumento del número de oxidación: Cu+ Cu2+
Ganancia de e-, conlleva disminución del número de oxidación: Cu2+ Cu+
Pérdida de hidrógeno: R-H RLos e- van acompañados de H+
Incorporación de hidrógeno: R R-HLos e- van acompañados de H+
Incorporación de oxígeno: R R-O
El Oxígeno atrae hacia sí los e-
Pérdida de oxígeno: R-O REl Oxígeno atrae hacia sí los e-

OxidaciónLas moléculas que pierden electrones quedan oxidadas.Se le llama dador de electrones, porque se lo cede a otra molécula. También se le llama agente reductor porque la molécula a la que cede los electrones se reduce. Cuando una molécula se oxida, reduce a otra. OJO!!! La molécula oxidada es el agente reductor.

Las moléculas que ganan electrones quedan reducidas.Se le llama aceptor de electrones, porque lo acepta de otra molécula. También se le llama agente oxidante porque la molécula de la que los acepta se oxida. Cuando una molécula se reduce, oxida a otra. OJO!!! La molécula reducida es el agente oxidante.
Reducción

Transportadores intermedios
En las reacciones de oxidación se libera energía y electrones (a veces acompañados de H+, formando Hidrógeno) Se precisan moléculas que almacenen y/o lleven los electrones (o el Hidrógeno) y la energía desde las reacciones catabólicas hasta las anabólicas donde se necesitan Esto es así porque ambos tipos de reacciones se dan en distintos lugares de la célula y en diferentes momentos.

Transportadores de energía
Se utilizan nucleótidos trifosfato de Adenina (ATP), Guanina (GTP), Citosina (CTP) y Uracilo (UTP), aunque el más utilizado con mucha diferencia es el ATP.Los enlaces que unen entre sí los grupos fosfato son muy energéticos y cuando se rompen [al pasar de trifosfato (ATP) a difosfato (ADP) y de difosfato a monofosfato (AMP)], liberan la energía almacenada. La energía se utiliza para todas las necesidades.

El ATP se puede sintetizar de dos formas: fosforilación a nivel de sustrato:
Para transferir un grupo fosfato al ADP y obtener ATP, se utiliza la energía liberada en la hidrólisis de compuestos fosforilados muy ricos en energía. Se les quita el grupo fosfato y se usa dicho fosfato y la energía liberada al desprenderlo.
X-P + ADP X + ATP Glucosa-6-P + ADP Glucosa + ATP
Requieren la intervención del ATP

Por ejemplo con Glucosa-6-fosfato
Glucosa 6-P Glucosa
ADP + P ATP
Transportadores de energía

Fosforilación acoplada al transporte de membranas.
Se realiza por sucesivos pasos de e- de unas sustancias a otras de manera que en cada paso los electrones pierden energía.Esta energía se utiliza por una enzima llamada ATP-sintetasa para sintetizar ATP a partir de ADP + Pi (se llama así, pero es fosfato)Todas las sustancias por las que pasan los electrones, y la ATP-sintetasa se encuentran formando parte de membranas
Transportadores de energía

Dependiendo de qué membranas se trate se habla de:
fosforilación oxidativa: membrana interna de mitocondrias–Se realiza como parte del catabolismo.
fotofosforilación: transformación de la energía lumínica en las membranas de los tilacoides de cloroplastos.–Se realiza como parte del anabolismo.
Transportadores de energía

ADENOSÍN-TRIFOSFATO (ATP)
OHO — P — O — P — O — P — O
H
CH2
OH OH
H
Adenina
HH
OHOH OH
OO O
ATP
ATP + H2O → ADP + Pi + energía (7,3 kcal/mol)
ADP + H2O → AMP + Pi + energía (7,3 kcal/mol)
ADP
AMP
OHO — P — O — P — O
H
CH2
OH OH
H
Adenina
HH
OH OH
O O
O HO — P — O
H
CH2
OH OH
H
Adenina
HH
OH
O
La célula y la energía
Transportadores de energía

Transportadores de electrones Los electrones y/o hidrógenos que se desprenden en la oxidación, deben ser llevados hasta los lugares de la reducción. Para ello se usan intermediarios que:
se reducen al captar electrones (o hidrógenos) de la sustancia que se ha oxidado. los transportan. los ceden (se oxidan) reduciendo a otra sustancia.

Transportadores de electrones
Las sustancias que transportan hidrógeno o electrones más H+ son, principalmente:
Nucleótidos de Piridina oxidados: NAD, NADP reducidos: NADH, NADPH
Nucleótidos de Flavina oxidados: FAD, FMN reducidos: FADH2, FMNH2

NAD, NADP El NAD (Nicotinamin Adenin Dinucleótido) es, como su nombre indica, la unión de dos nucleótidos, uno cuya base nitrogenada es Adenina y el otro con base nitrogenada nicotinamida. El NADP (Nicotinamin Adenin Dinucleótido Fosfato) es igual, pero tiene un fosfato en el carbono 2 de la ribosa del nucleótido de Adenina.

NAD, NADP
Adenina
Nicotinamida
NAD NADP
Lugar de la reducción

NADH NADPH
NADH, NADPH Cuando captan H, en los lugares indicados, se transforman en NADH y NADPH
H H

FAD, FMN El FAD (Flavin Adenin Dinucleótido) es también la unión de dos nucleótidos, uno con Adenina y otro con Flavina como base nitrogenada. El FMN (Flavin MonoNucleótido) es un único nucleótido con Flavina como base nitrogenada.
Adenin

Se pueden reducir: una vez (H naranja en 5 para dar FADH, FMNH) dos veces (H verde en 1 además del naranja, para dar FADH2 y FMNH2)
En cualquier caso, los hidrógenos se unen siempre a la Flavina en los lugares indicados en rojo.
FAD, FMN

Son coenzimas de deshidrogenasas (catalizan reacciones redox)
FMNH2
FADH2
FADH, FADH2, FMNH, FMNH2
H
H

Transportadores de electrones

Otras características2- Están encadenadas:
El producto de una reacción es el sustrato de la siguiente.Los productos intermedios se denominan metabolitos.Las vías pueden ser lineales, ramificadas o cíclicas.
3- Están compartimentadasDistintas reacciones se llevan a cabo en orgánulos diferentes.
4- Están catalizadas por enzimasLas enzimas que actúan son específicas y las condiciones pueden ser diferentes en los distintos compartimentos.

CATABOLISMO

Características y tipos

CaracterísticasConjunto de reacciones de transformación de moléculas orgánicas en otras moléculas orgánicas más simples o en moléculas inorgánicas. Los productos finales constituyen los productos de excreción.Son reacciones de oxidación y exergónicas.

La energía liberada se almacena en los enlaces del ATP y se utiliza para las actividades celulares o para fabricar moléculas más complejas en reacciones de reducción que requieren energía (anabolismo).Los electrones (o H) liberados se almacenan en sustancias transportadoras (NAD, FAD, etc.) para fabricar moléculas más complejas en reacciones de reducción que requieren electrones (anabolismo).
Características

Catabolismo y Oxígeno.En función de su relación con el Oxígeno, las células pueden clasificarse en
Aerobias: son la mayoría y utilizan el O2 como aceptor final de e-, obteniendo mucha energía.Anaerobias facultativas: Levaduras o muchas células de organismos superiores (musculares de animales). Normalmente son aerobias, pero si falta el O2 siguen una vía anaerobia.Anaerobias estrictas: Tienen que vivir en ausencia de O2 ya que les es nocivo. Son algunos microorganismos (bacterias de suelos profundos o fondos oceánicos).

TiposRespiración: da lugar a sustancias inorgánicas.
Respiración aerobia: El aceptor final de todos los e- (en forma de H2) que se desprenden en las oxidaciones es el O2 que se reduce a H2O.
La realizan las células eucariotas y muchas procariotas.
Respiración anaerobia: presenta como último aceptor de e- sustancias inorgánicas diferentes del O2
Exclusivo de algunas células procariotas.

TiposFermentación: El último aceptor de e- es una molécula orgánica.
Se considera un catabolismo parcial ya que las sustancias finales podrían oxidarse más.Los productos finales son sustancias aún orgánicas (aunque más oxidadas que las iniciales). Son reacciones anaerobias y liberan poca energía. La realizan células procariotas y algunas eucariotas como levaduras y células musculares de animales.

Tipos de anaerobiosis Hay dos tipos de catabolismo anaerobio, como se ha visto:
fermentación: el aceptor final es una sustancia orgánica y los productos finales son orgánicos.
No hay cadena de transporte (se verá más adelante)
respiración anaerobia: aunque es anaerobia, no se debe confundir con la fermentación.
En ésta el aceptor final es una molécula inorgánica.

La respiración anaerobiaEs un proceso redox con cadena de transporte electrónico en la que el aceptor final de electrones es una molécula inorgánica distinta del oxígeno (más raramente, una molécula orgánica muy sencilla como metano). No se usa oxígeno, sino otra sustancia oxidante, como el sulfato o el nitrato.En la siguiente tabla se muestran distintos aceptores de electrones, sus productos y algunos ejemplos de microorganismos que realizan tales procesos:

La respiración anaerobia

Rutas catabólicas de los distintos nutrientes

Rutas catabólicas de los distintos nutrientes
Las rutas catabólicas más importantes de los distintos nutrientes son:
Glúcidos: glucolisis.Lípidos: b-Oxidación de ácidos grasos.Prótidos: transaminación y desaminación.
En general, todas estas rutas convergen en una ruta central de todo el catabolismo llamada Ciclo de Krebs ya que terminan en un complejo llamado Acetil CoA (que inicia el ciclo) o en algún otro compuesto que forma parte del ciclo en distintos lugares.

CATABOLISMO DE LA
GLUCOSA

Respiración aerobiaEs el catabolismo total de cualquier forma de molécula orgánica que se oxida hasta CO2, libera energía e hidrógenos que son captados por el O2 para formar H2O.Generalmente los sustratos son glúcidos y lípidos.La glucosa es la molécula más utilizada y la reacción global sería:C6H12O6 (glucosa) + 6O2 6CO2 + 6H2O +
energía (36-38 ATP)
Esta reacción no se lleva a cabo directamente, pues se perdería la mayor parte de la energía en forma de calor.

Los organismos heterótrofos la incorporan tras convertir los alimentos en nutrientes (polisacáridos en monosacárido glucosa)Los autótrofos la sintetizan en la fotosíntesis a partir de materia inorgánica.Puede también obtenerse:
por transformación de otras moléculas orgánicas (gluconeogénesis en anabolismo) por la hidrólisis de polisacáridos de reserva (glucogenolisis)
Incorporación de glucosa

La enzima glucógeno fosforilasa (o almidón-fosforilasa en vegetales) cataliza la rotura del enlace a(1-4) de la glucosa de un extremoA la vez, le transfiere un fosfato, liberándose Glucosa-1-fosfato y quedando el glucógeno (o el almidón con una glucosa menos)Así sucesivamente, se van obteniendo G-1-PLas roturas de las ramificaciones, con enlace a(1-6), son más complicadas y dependen de otra enzima, pero igual se van desprendiendo.Posteriormente, la G-1-P se transforma en G-6-P que es la que se va a oxidar, definitivamente.
Resumen de glucogenolisis

Resumen de glucogenolisis
Polisacáridos Glucosa 1- PGlucógenolisis
Glucógeno/Almidón(n glucosas)
Glucosa 1-P +
Glucógeno/almidón (n-1) glucosas
Glucógeno fosforilasa
Pi
Glucosa-1-Fosfato Glucosa-6-Fosfatofosfogluco
mutasa

Vía fermentativa en ausencia de
oxígeno
Oxidación vía mitocondrial en
presencia de oxígeno
Destino de la glucosa
glucosa
Piruvato
Glucógeno, almidón,
Almacenamiento (anabolismo)
Oxidación vía glucólisis no es dependiente de oxígeno (catabolismo)
GLUCOLISIS

Catabolismo de la GlucosaLa glucosa es el monosacárido más abundante y sirve de ejemplo del catabolismo de los glúcidos. Su oxidación no se realiza de golpe (la energía se liberaría de forma súbita y se perdería en gran parte)Se va realizando poco a poco, en una serie de reacciones que desprenden la energía en pequeñas dosis fácilmente almacenables en moléculas de ATP.

El proceso consta de dos fases: 1ª fase: la glucosa se oxida en una serie de reacciones en una ruta metabólica llamada glucolisis de la que se desprende ATP y dos moléculas de ácido pirúvico. 2ª fase. El ácido pirúvico se oxida y puede ser:
De forma aerobia, mediante la respiración celular que lo degrada hasta CO2 y H2O, actuando el O2 como aceptor final de e-. De forma anaerobia, mediante fermentación, convirtiéndose en alguna molécula orgánica más sencilla. El aceptor de e- es otra molécula orgánica.
Catabolismo de la Glucosa

Glucolisis

GlucolisisConsta de dos fases:
Glucolisis I o de consumo de energía. Por cada Glucosa se consumen dos ATP y tras una serie de reacciones, se obtienen dos moléculas de Gliceraldehido-3-fosfato. Glucolisis II o de producción de energía. Por cada gliceraldehido-3-fosfato se forman dos ATP (4 en total) y se genera un ácido pirúvico.
En resumen: por cada glucosa obtenemos dos ATP (hay que restar los dos gastados) y dos de ácido pirúvico.

Glucolisis ILa glucosa se rompe en dos moléculas de 3 Carbonos: gliceraldehido-3-P y dihidroxiacetona fosfato. Para ello ha debido haber dos fosforilaciones y se han gastado 2 moléculas de ATP. La dihidroxiacetona, pasa a gliceraldehido. Al final, tenemos dos de gliceraldehido-P

C1
C6
C2C3
C4
C5 O
GLUCOSA
P
C1
C6
C2C3
C4
C5 O
P P P ATP P P ADP
GLUCOSA 6 FOSFATO
MEMBRANA CELULAR
1
Mg2
HEXOCINASA
Fase de inversión de energía, gasta 1 ATP

P
C1
C6
C2C3
C4
C5 O
C1C6
C2
C3C4
C5O
P
2 GLUCOSA 6 FOSFATO - FRUCTOSA 6 FOSTATO
GLUCOSA 6 FOSFATO
FRUCTOSA 6 FOSTATO
FOSFOHEXOSA
ISOMERASA

C1C6
C2
C3C4
C5O
P P
C1C6
C2
C3C4
C5O
P
FRUCTOSA 6 FOSFATO
P P P ATP P P ADP
FRUCTOSA 1-6 DIFOSFATO
3
Mg2
FOSFOFRUCTO
QUINASA
Fase de inversión de energía, gasta 1 ATP

C1C6
C2
C3C4
C5O
P P
C1
C2
C3
PO
Gliceraldehído 3 Fosfato Dihidroxiacetona - Fosfato
4
5
C1
C2
C3P
=
OH
ALDOLASA
TRIOSAFOSTATO
ISOMERASA
FRUCTOSA 1- 6 DIFOSFATO

RESUMEN
Fosforilaciónhexocinasa
Isomerizaciónfosfohexosaisomerasa
Fosforilaciónfosfofructocinasa
Rupturaaldolasa
IsomerizaciónFosfotriosa isomerasa
1
2
3
4
5

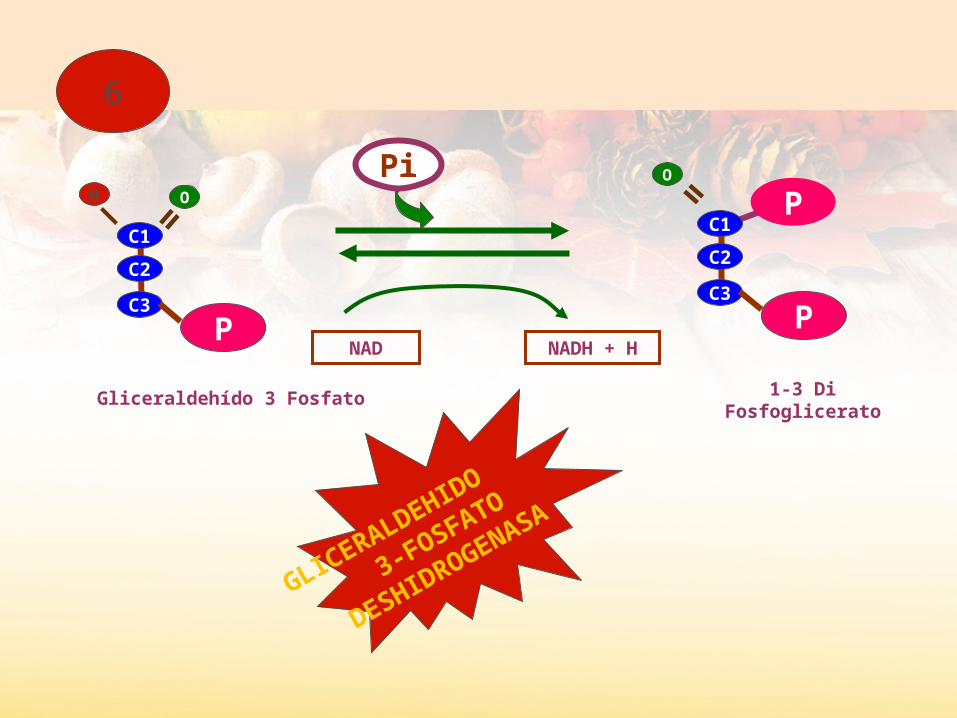
A partir de aquí, todo hay que multiplicarlo por dos.El gliceraldehido-3-P sufre una fosforilación y una oxidación simultáneas.
Fosforilación en el carbono 1. No se gasta ATP porque se obtiene el Pi tal cual.Oxidación del grupo aldehído a grupo ácido
Obtenemos al final una molécula de ácido 1-3-difosfoglicérico
Glucolisis II

Glucolisis IIEl Ácido 1-3-fosfoglicérico, se oxida hasta ácido pirúvico.Se desprenden 2 Pi de cada molécula obteniéndose ATP por fosforilación a nivel de sustrato. En total, se obtienen 4 ATP (restando los dos empleados en la etapa anterior, sacamos un balance positivo de 2 ATP)
C1
C2
C3P
P
NAD NADH + H
1-3 Di Fosfoglicerato
6
C1
C2
C3P
=OH
Gliceraldehído 3 Fosfato
GLICERALDEHIDO
3-FOSFATO
DESHIDROGENASA
Pi O =

C1
C2
C3P
P
P P P ATPP P ADP
C1
C2
C3P
1-3 Di Fosfoglicerato 3 Fosfoglicerato
P7
FOSFORILACION A NIVEL DE SUSTRATO
FOSFOGLICERATO
QUINASA
O = O = O

8
3 Fosfoglicerato 2 Fosfoglicerato
FOSFOGLICERATO
MUTASA
C1
C2
C3P
=OO
C1
C2
C3P
=
OO
Mg2

Fosfoenolpiruvato
P P P ATP
P P ADP
PIRUVATO
P
H20
9
10
C1
C2
CH2
P
OO=
H
HO
C2
CH2
P
OO
=C1
ENOLASA
2 Fosfoglicerato
C
C = OCH3
OO
=
PIRUVATOCINASA

RESUMENOxidación y
FosforilaciónGliceraldehido
3- fosfato deshidrogenasa
Fosforilación a nivel del sustrato
Fosfoglicerato cinasa
Isomerización
fosfogliceratomutasa
DeshidrataciónEnolasa
Fosforilación a nivel del sustrato
Piruvato cinasa
6
7
8
9
10

Glucolisis: balance
Glucosa 2 piruvatos
2 ATP +2 NADH
Utilizados: Ganados: 2 ADP 4 ATP
2 Pi 2 NADH2 NAD 2 H20
2 ATPGanancia neta: 2 ATP y 2 NADH

Glucolisis: balance
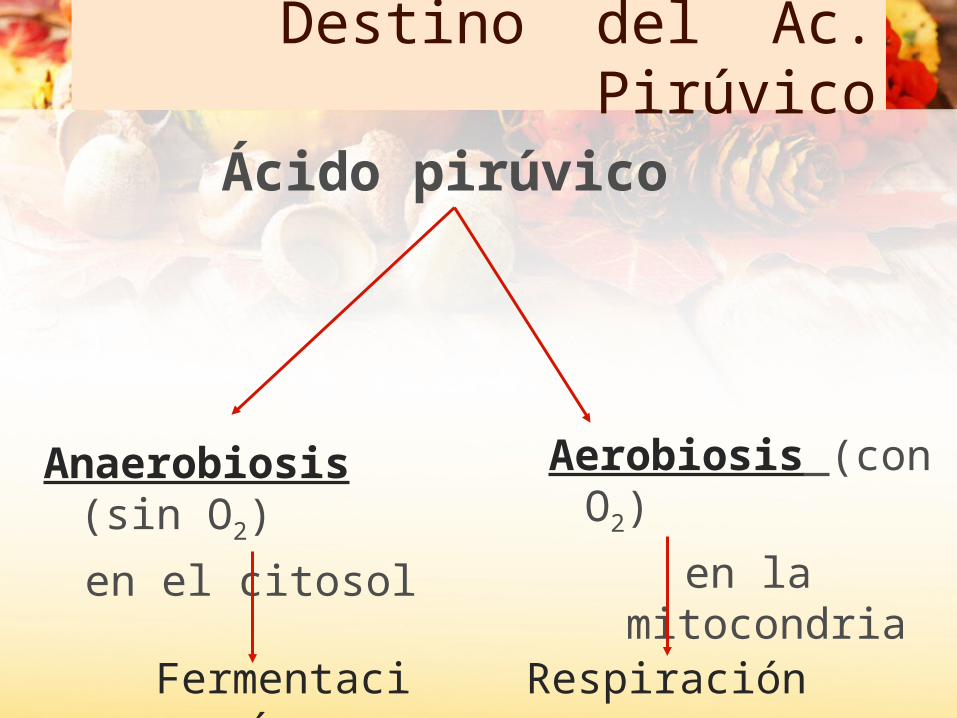
Destino del Ác. Pirúvico
Anaerobiosis (sin O2)
en el citosol
Aerobiosis (con O2)
en la mitocondria
Fermentación Respiración
Ácido pirúvico

glucosa
2 piruvatos
2 etanol + 2 CO2 2 lactato2 acetil-CoA
4 CO2 + 4 H2O
2 CO2
Anaerobias Anaerobia
s Aerobia
s Fermentación alcohólica en
levaduras
Fermentación láctica en músculo,
eritrocitos y bacterias
anaeróbicas
glucólisis
Ciclo de
Krebs
Células animales, vegetales y
microorganismos
independiente de O2

Piruvato: sin oxígeno

Fermentación del piruvato No requiere O2 como último aceptor de e-
El aceptor final es una molécula orgánica. Es un catabolismo parcial.Tiene lugar en el citosol.El ATP se obtiene por fosforilación a nivel de sustrato (se obtienen solo los dos ATP de la glucolisis)Rendimiento energético escaso.

Fosforilación a nivel de sustrato

Fermentación del piruvatoLa ventaja de este proceso es que el NAD se recicla una y otra vez, con lo que no es necesario que entre en una cadena transportadora (imposible sin O2) Cada vez que entra un Piruvato se oxida cediendo los Hidrógenos al mismo NAD que pasa a NADH.El NADH cede los electrones al aceptor final (distinto según el tipo de fermentación) y vuelve a ser NAD, listo para volver a actuar.

La fermentacion alcohólicaGlucosa + 2 (ADP + Pi) 2 Etanol + 2 CO2 + 2 ATPAceptor final: acetaldehído para convertirse en etanol (reciclado de NAD+)Importancia: cerveza, vino, pan (Saccharomyces)
Glucolisis
Fosforilación a nivel
de sustrato
Reciclado del NAD

La fermentacion lácticaGlucosa + 2 (ADP + Pi) 2 Ácido láctico (C3) + 2 ATPAceptor final: Piruvato que se convierte en lactato (reciclado de NAD)Importancia: prod. lacteos: mantequilla, queso, yogur, “músculo” “agujetas”
Glucolisis
Fosforilación a nivel
de sustrato
Reciclado del NAD

Piruvato: con oxígeno

Respiración celularLa respiración celular o aerobia es el proceso que sigue el ácido pirúvico en presencia de oxígeno.Por este proceso, cada glucosa se oxida totalmente hasta CO2 y H2O.En este proceso, el piruvato de la glucolisis (dos, en realidad) sufre tres procesos:
Descarboxilación oxidativa.Ciclo de KrebsTransporte de electrones con fosforilación oxidativa.

Descarboxilación oxidativa del piruvato
Consiste en la transformación del piruvato en Acetil Co-APara que esto suceda, el piruvato debe ingresar en la matriz mitocondrial.
ácido pirúvico
NAD
CoA-SH NADH2CO2
Piruvato deshidrogena
sa
acetil CoA

Descarboxilación oxidativa del piruvato
Consiste en: 2 Piruvato + 2 NAD+ + 2 CoA-SH 2 Acetil CoA + 2 CO2 + 2
NADHCada piruvato se transforma en acetato perdiendo un CO2 (2 en total)El acetato de une a la CoA por el grupo tiol (SH), formando Acetil Coenzima A.Se pierde el H del tiol que reduce NAD a NADH (2, uno por cada piruvato)Se forma Acetil CoAContinúa en el ciclo de
Ciclo de Krebs
“Acetil CoA Punto de encuentro de las
encrucijadas metabólicas”

El ciclo de Krebs

El acetato se desprende del CoA y se une a un ácido de 4 carbonos (oxalacetato) dando un ácido de 6 C (citrato). A partir de aquí tienen lugar un serie de reacciones tendentes a recuperar el oxalacetato y reiniciar el ciclo al entrar otro Acetil-CoA. A lo largo de esas reacciones:
Se desprenden 2 de CO2 por ciclo (4 por glucosa)Se desprenden 3 de NADH (6 por glucosa).Se desprende un FADH (2 por glucosa).Se desprende 1 GTP (2 por glucosa) con un rendimiento energético equivalente al ATP.
El ciclo de Krebs

El ciclo de Krebs

Ciclo de Krebs

Rendimiento del Ciclo de Krebs (hay que multiplicar
x2)Acetil CoA3 H2O3 NAD1 FAD1 GDP + Pi
2 CO2
1 H2O3 NADH2
1 FADH2
1 GTPSe ha transformado la materia orgánica en inorgánica.
Los electrones han sido recogidos como (H2) por coenzimas transportadoras de electrones (NAD y FAD).
Se ha sintetizado una molécula de GTP, análoga al ATP.
ENTRA SALE
12 ATP

Cadena de transporte electrónico: fosforilación
oxidativa

Cadena transportadora de e-
Se realiza con todas las moléculas reducidas de NADH y FADH, desprendidas en los procesos anteriores.Estas moléculas se oxidan al ceder los e- a una serie de sustancias (la cadena de transporte) que se los van cediendo unas a otras, en una cadena redox hasta cedérselos (junto a H+) al O2 para reducirlo y convertirlo en H2O.Todas las sustancias de la cadena están formando parte de la membrana de las crestas mitocondriales.

La cadena está constituida por: Cuatro complejos (I, II, III y IV) que son agrupaciones de proteínas transportadoras y que están fijos en la membrana de las crestas mitocondriales. Ubiquinona (Q) capaz de trasladarse por la membrana llevando e- del complejo I y II al complejo III. Citocromo c, también móvil, conecta el complejo III con el IV
Cadena transportadora de e-

Cadena transportadora de e-

Fosforilación oxidativa: Teoría quimiosmótica
La transferencia de electrones de unos complejos a otros va acompañada de un transporte de protones de la matriz al espacio intermembrana (teoría quimiosmótica de Mitchell)

Se produce una diferencia de potencial entre ambos espacios (potencial electroquímico).Para compensar dicho potencial, los protones son devueltos a la matriz por la ATP-sintetasaEn ese paso de protones de vuelta a la matriz, se fabrica ATP, por fosforilación oxidativa. Si los e- se incorporan al complejo I, salen protones suficientes para fabricar 3 ATP.Si los e- se incorporan al complejo II, solo salen protones suficientes para fabricar 2 ATP.
Fosforilación oxidativa: Teoría quimiosmótica

Fosforilación oxidativa: Teoría quimiosmótica
Los NADH y los FADH, obtenidos en las etapas anteriores, son los que ceden los electrones a uno de los complejos enzimáticos. Los NADH ceden electrones al complejo I por lo que hacen un recorrido completo obteniéndose 3 ATP .Los FADH ceden electrones al complejo II por lo que harán un recorrido menor y se obtienen solo 2 ATP.El último aceptor de electrones de la cadena es el O2 que se reduce a agua.

TRANSPORTE DE ELECTRONES
Fosforilación oxidativa: Teoría quimiosmótica

LanzaderasLos 2 NADH formados durante la glucolisis, en el citosol tienen que entrar en la mitocondria para incorporarse a la cadena de transporte. La membrana interna de las mitocondrias es impermeable al NADH por lo que en dicha membrana hay unos sistemas llamados lanzaderas.Las lanzaderas introducen los electrones de estos NADH, bien al complejo I o al complejo III, variando, de esta manera el balance final de ATP entre 36 y 38.

LANZADERASLanzadera malato-
aspartatoLa lanzadera malato aspartato, transfiere el NADH al interior de la mitocondria a nivel del complejo I, por lo que, en total, se generan 3 ATP. Esta lanzadera está presente en las mitocondrias de las células de corazón, riñón e hígado.Si los dos NADH del citoplasma entran por esta lanzadera, el balance del catabolismo total de la glucosa son 38 ATP

Lanzadera glicerol-fosfatoLa lanzadera del glicerolfosfato, transfiere el NADH al interior de la mitocondria a nivel del complejo III. En total se originan 2 moléculas de ATP. Esta lanzadera está en mitocondrias de células musculares y del cerebro.Si los dos NADH del citoplasma entran por esta lanzadera, el balance del catabolismo total de la glucosa son 36 ATP.

Lanzadera malato-aspartato (3ATP)
Lanzadera glicerol fosfato (2 ATP)
LA RESPIRACIÓN
CELULAR AEROBIA

Rendimiento máximo

Balance energético: 36/38 atp
PROCESO GLOBAL: Glucosa + 6 O2 6 CO2 + 6H2O + 38 ATP (máximo)que pueden ser 36, dependiendo del tipo de lanzadera que utilice la célula-
2
2
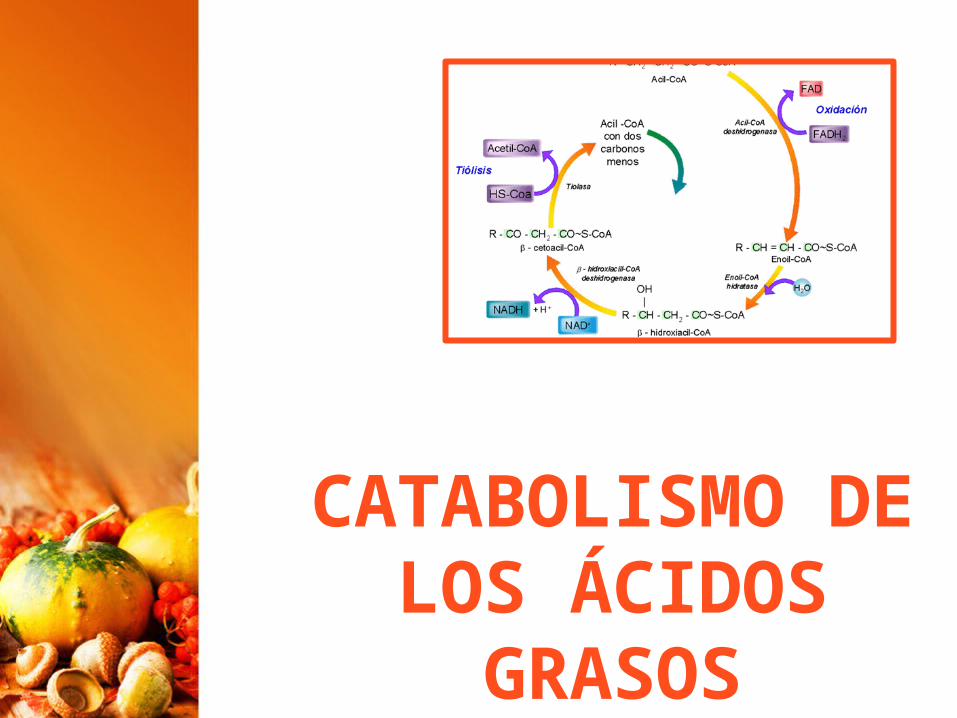
CATABOLISMO DE LOS ÁCIDOS
GRASOS

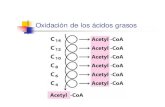
Catabolismo de los lípidosEn los seres vivos, sobre todo en animales, las grasas tienen una enorme importancia como combustible por su alto valor energético.La oxidación de un gramo de grasa proporciona 9,5 Kcal, mientras que un gramo de glúcidos o de proteínas proporciona unas 4,2 Kcal.La vía metabólica principal en el catabolismo de las grasas es la oxidación de los ácidos grasos que provienen de su hidrólisis.Los ácidos grasos también pueden provenir, aunque, en menor proporción, de los fosfolípidos.

Catabolismo de lípidos: Ácidos grasos
Triglicérido
glicerol
Gliceraldehido-P
NADPiNADH
a la glucolisis, ciclo de Krebs, transporte de
electrones, etc.
Ácidos grasos
a la β-oxidación o hélice de Lynen en la
mitocondria

La oxidación de ácidos grasos se produce en la matriz mitocondrial.La ruta se llama b-oxidación o hélice de Lynen.Consiste en una secuencia repetida de reacciones, de forma que, en cada una, se desprenden dos Carbonos del ácido graso en forma de Acetil-Co A.La pérdida de los dos Carbonos comienza por el extremo carboxilo.Como los ácidos grasos no pueden traspasar la membrana mitocondrial, deben activarse con lo que se gasta ATP (el equivalente a 2 ATP)
ß-oxidación o hélice de Lynen

Coenzima A
Si los ácidos grasos tienen número par de átomos de C darán solo Acetil-Co A
Si los ácidos grasos tienen número impar de átomos de C
darán además una molécula de Propionil-Co A
ß-oxidación o hélice de Lynen

Primera fase: Activación del ácido graso.El ácido graso, en el citosol, se une al CoA formando un compuesto llamado acil-CoA con gasto del equivalente de dos moléculas de ATP
(ATP AMP + 2 Pi).El acil-CoA tiene que entrar a la mitocondria para lo que, en el espacio intermembranoso, se une a la carnitina (y pierde el Co-A que favorecerá el paso, formando Acil-carnitina.Una vez en el interior de la mitocondria, la carnitina se separa quedando libre el acil-CoA (se une a otro CoA) y pasando a la segunda fase: la b-oxidación.
Activación del ácido graso

Activación del ácido graso
Hacia la hélice de Lynen

ß-oxidación o hélice de Lynen
Ácido Palmítico: 16 Carbonos

R-CH2-CH2-COOHácido graso
ATP AMP+2Pi
Ciclo de krebs
ß-oxidación o hélice de Lynen

ß-oxidación o hélice de Lynen
Acetil CoA
Acil CoA con 2 C menos
Al ciclo de
Krebs
A la cadena de
transporte

Segunda fase: hélice de Lynen. Los ácidos grasos se van oxidando, perdiendo los carbonos de dos en dos. Cada par de carbonos se acaba convirtiendo en Acetil-CoA que ingresa al ciclo de Krebs.
ÁCIDO GRASO DE n CARBONOS
n/2 CICLOS DE Krebs
ß-oxidación o hélice de Lynen

En cada vuelta, para perder dos carbonos en forma de acetil-Co-A, se desprende un FADH y un NADH que tendrán su rendimiento en la fosforilación oxidativa (cadena respiratoria o de transporte electrónico).
ÁCIDO GRASO DE n CARBONOS
(n/2)-1 VUELTAS (FADH y NADH)
ß-oxidación o hélice de Lynen

Rendimiento de la β-oxidación de ácidos grasos Ej. ácido caproico 6C
Nº de vueltas
Nº de ciclos de krebs
Moléculas reducidas por vuelta
Rendimiento del Ciclo de Krebs
Rendimiento de la fosforilación oxidativa
Ácido graso de N carbonos
N/2 - 1 N/2 1 NADH 1 FADHTOTAL(N/2-1) NADH (N/2-1) FADH
Cada acetil CoA rinde 3 NADH1 FADH 1 GTP
1 NADH rinde 3 ATP y 1 FADH rinde 2 ATP
Ácido caproico(6C, igual que la glucosa)
2 32 NADH2 FADH
9 NADH3 FADH3 GTP
33 ATP10 ATP 3ATP
TOTAL menos 2 ATP gastados para entrar = 44 ATP

Cada ciclo de krebs = 12 ATPCada vuelta = 5 ATP (3 de un NADH y 2 de un FADH)
Ácido graso de n Carbonosn/2 x 12 ATP + [(n/2) – 1] x 5 ATP – 2 ATP
Ácido graso de 20 Carbonos(10 x 12) + (9 x 5) – 2 = 163 ATP
Balance general: n carbonos
Ciclos de krebs = n/2
21 Nº de vueltas n
Ciclo de Krebbs Vueltas Gastados

Glucosa 38 ATP, Ácido caproico 44 ATP ¿Por qué se prefieren los glúcidos?
Movilización más rápida de glúcidos.Algunos tejidos sólo pueden utilizar glucosa (nervioso, eritrocitos...)Los ácidos grasos son insolubles y necesitan moléculas transportadoras.En anaerobiosis no funciona el ciclo de Krebs. Se hace imprescindible la glucolisis.En animales, los ácidos grasos no se transforman en glúcidos (al revés, sí)
Catabolismo de las
proteínas

Las proteínas que ya no se necesitan (orgánulos que se destruyen por autofagia, enzimas inservibles, etc.) son hidrolizadas para obtener aminoácidos, los cuáles serán empleados:
En la síntesis de nuevas proteínas, tal cual están.En la síntesis de nuevas proteínas después de haberlos transformado en otros por procesos de transaminación.Destruidos por no ser necesarios en un proceso catabólico. Para ello el grupo amina y el esqueleto carbonado son separados y degradados por rutas metabólicas interconectadas.
Catabolismo de proteínas

aa
a-cetoácido 2
aa
a-cetoácido 1
Transaminación: el grupo amino se puede transferir: el aminoácido que pierde el grupo amina se transforma en un a-cetoácido. Al transferir el grupo amina a otro a-cetoácido, se obtiene otro aminoácido.
Transaminación
Aminoácido 1 Aminoácido 2

El grupo amina se excretará de diferente manera según los organismos:
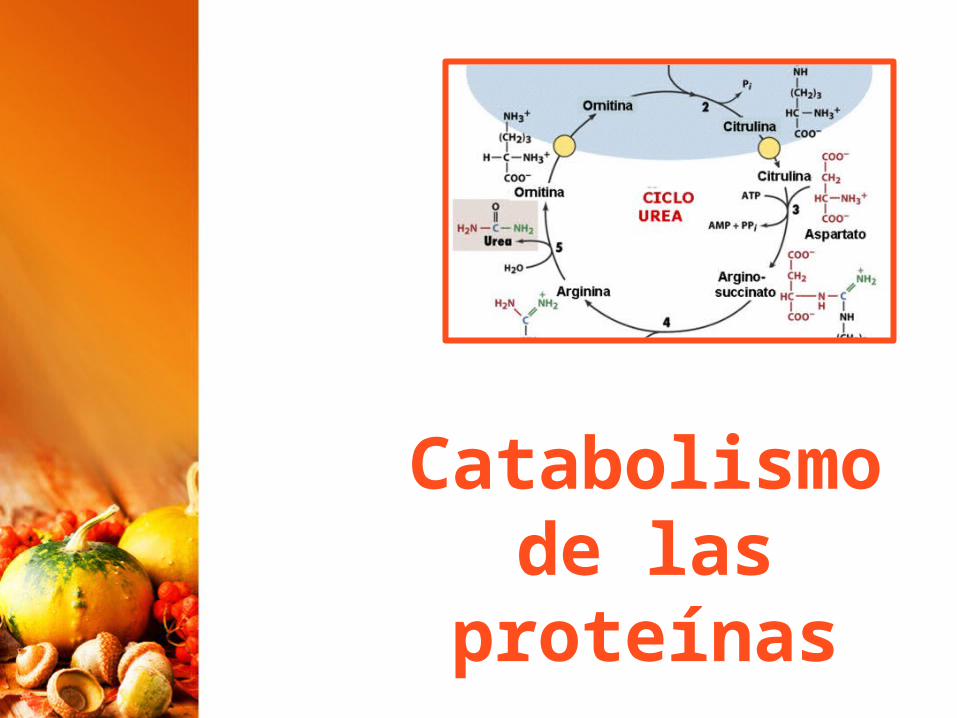
Los peces óseos, excretan amoníaco por lo que se les llama animales amoniotélicos.Las aves y también los reptiles son uricotélicos y lo excretan en forma de ácido úrico.La mayoría de animales terrestres son ureotélicos, por lo que lo excretan en forma de urea; En este caso, el amoníaco depositado en las mitocondrias de los hepatocitos se convierte en urea mediante el ciclo de la urea
Catabolismo de proteínas

CICiclo de la Urea

El resto de la cadena hidrocarbonada del aminoácido, una vez desaminado, se va a convertir en alguna sustancia (ácido pirúvico, acetil CoA, etc.) del ciclo de krebs y, por lo tanto, se introducirá en dicho ciclo, siendo totalmente degradada. No obstante, hay que recordar que las proteínas no tienen como función servir para proporcionar energía.Su catabolismo solo se lleva a cabo para eliminar de la célula estructuras u otras sustancias que ya no sirven.
Catabolismo de proteínas

CATABOLISMO DE PROTEÍNASCatabolismo de proteínas

Catabolismo de ácidos nucléicos

Catabolismo de los ácidos nucleicos
Los nucleótidos procedentes de la hidrólisis de los ácidos nucléicos se separan en la pentosa, grupos fosfato y la base nitrogenada:
La pentosa sigue las vías de los glúcidos en su degradación incorporándose, antes o después al ciclo de Krebs.Las bases nitrogenadas se utilizan para fabricar nuevos nucleótidos o se degradan hasta el producto nitrogenado de excreción del organismo (amoniaco, urea o ácido úrico)Los grupos fosfato se pueden excretar como tales disueltos en orina o ser utilizados para fabricar ATP, nuevos nucleótidos o para cualquier fosforilación.

Metabolismo y nutrición

Anaya

Anaya

Continúa en la siguiente
Anaya

Anaya

Anaya

Anaya
Continúa en la siguiente

Anaya

PAU Cantabria

PAU CantabriaDefine el ciclo de Krebs ¿Qué papel juega el ciclo de Krebs en el metabolismo aeróbico? ¿Qué tipo de biomoléculas utilizan este ciclo en su catabolismo?Describe, mediante un dibujo o esquema, el mecanismo que tienen las células eucarióticas para obtener ATP en presencia de oxígeno. ¿Puede haber respiración en ausencia de oxígeno? Razona tu respuesta y pon un ejemplo. ¿Dónde se produce ATP en una célula? ¿En qué parte/s de la misma tiene lugar este proceso? Indica un proceso celular en el que se consuma ATP.

Escribe un texto coherente, de no más de diez líneas, en el que se relacionen los siguientes conceptos referentes a un determinado fenómeno biológico: glucosa, fermentación láctica, glicolisis, anaerobiosis.Comente brevemente el destino catabólico de: proteínas, almidón y triglicéridos, indicando, mediante un esquema, las rutas metabólicas más relevantes (incluyendo las que son comunes a todos ellos) implicadas en los diferentes procesos, así como los metabolitos finales de triglicéridos y almidón. Considerar condiciones aerobias.
PAU Cantabria

Identifica las rutas metabólicas a, b, c, d y f que aparecen en el esquema de la figura, e indica la finalidad metabólica de cada una de ellas. Señala las que funcionarían en presencia de oxígeno. Razona la respuesta.
PAU Cantabria

En presencia de O2: ¿en qué etapa del metabolismo coinciden los procesos catabólicos de triglicéridos y glúcidos?¿Cuáles son los productos finales de sus respectivos catabolismos? ¿Cuál de los dos tipos de biomoléculas proporciona un mayor rendimiento calórico por molécula degradada en las citadas circunstancias? ¿En qué compartimento/orgánulo celular ocurre dicha etapa?
PAU Cantabria

Dentro de un esquema general del catabolismo aeróbico, comenta el destino de proteínas, polisacáridos y triglicéridos, indicando los productos finales de su degradación, así como las rutas metabólicas centrales en las que convergen dichos procesos.Explica el fundamento del ensayo de la presencia de catalasa en una muestra de tejido o células. Cita dos tejidos o muestras biológicas donde esperarías encontrar actividad catalasa.
PAU Cantabria

Completa la siguiente reacción de la degradación de la glucosa en la célula eucariota hasta sus productos finales.
Glucosa + O2
Indica, por orden de intervención, las rutas metabólicas que intervienen en dicho proceso y comenta la finalidad de cada una de ellas.
Haz un breve comentario sobre el catabolismo aerobio de los glúcidos y triglicéridos, indicando las rutas metabólicas comunes a ambos procesos, así como los rendimientos energéticos y los productos finales de degradación en cada caso.
PAU Cantabria

PAU CantabriaComenta con un esquema el papel del ciclo de Krebs en el metabolismo de los ácidos grasos. (Considera condiciones aerobias).Metabolismo fermentativo (fermentación) concepto, mecanismo, principales diferencias con el no fermentativo. Pon dos ejemplos de productos de metabolismo fermentativo que tengan interés industrial.Comenta y razona el concepto y funciones del metabolismo en seres vivos, poniendo ejemplos concretos en cada caso. ¿Puede haber seres vivos sin metabolismo? Razona la respuesta.

Fin