TB04 - Membranas Biológicas (1)

of 55
-
Upload
tayna-alejandra-hernandez -
Category
Documents
-
view
255 -
download
0
description
Membranas Biológica
Transcript of TB04 - Membranas Biológicas (1)
-
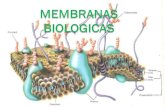
Membrana biolgica, composicin, estructura y funcin.
Transporte, tipos. Transporte pasivo, smosis, difusin simple y facilitada.
Transporte activo, bombas.
-
La antigua medicina china consideraba 404 causas para las enfermedades:
101 causas para enfermedades por la tierra (significaba huesos y msculo)101 por el agua (sangre)101 por el fuego (fiebre)101 por el aire (sistema respiratorio)
En la actualidad se describen mucho ms factores, virus bacterias hongos, alteraciones genticas, dietas inapropiadas, agentes contaminantes txicos, desechos radioactivos y hasta factores psicolgicos.
-
Las diferentes causas de las enfermedades tienen un factor comn, Todas causan dao celular, cuando las clulas mueren, los tejidos y rganos mueren y finalmente el organismo muere.
Todas las clulas estn rodeadas por una membrana celular y las funciones celulares son dependientes de un sistema de membranas altamente organizado, tanto membranas de superficie como intracelulares.los roles de los sistemas de membrana son fundamentales para cualquier organismo.
La desorganizacin de los sistemas de membrana pueden ser las causas o resultado de un proceso de enfermedad. En cada caso: La destruccin de los sistemas de membrana es sinnimo de muerte celular.
-
En 1925 E. Gorter y Grendel extrajeron lpidos de un determinado nmero de membranas de eritrocitos, correspondiente a una rea de superfice conocida.
Ellos determinaron el rea de superficie ocupada por una monocapa de los lpidos extrados, extendidos en la interfase aire agua, el area de superficie de la monocapa era el doble de la que ocupaban en la membrana del eritrocito, concluyendo que las membranas consistian de bicapas lipdicas
-
Las membranas separan la clula del medio exterior. Tambin separan compartimentos dentro de la clula donde se realizan procesos especializados.
Membranas celulares cumplen diversas funciones en diferentes regiones y organelas de una clula.
A nivel del microscopio electronico, las diferentes membranas comparten una estructura comn , la "Unidad de membrana" parecida a una lnea de ferrocarril, con dos lneas densas separadas por un espacio claro.
-
Esta imagen resulta de la unin de los metales pesados densos en electrones usados como tincin en la microscopa electrnica de transmisin a la cabeza polar de los fosfolpidos, las lneas densas estan separadas por la porcin interior de la membrana ligeramente teida, que contiene las cadenas hidrofbicas de los cidos grasos
-
Una perspectiva histrica
Danielli y Davson (1930) estudiaron los triglicridos de las bicapas lipdicas sobre una superficie de agua. Ellos encontraron que estas molculas se arreglaban con las cabezas polares de cara al exterior.
Sin embargo, ellos siempre formaban gotitas (el aceite en el agua) y la tensin superficial era mucho ms alta que en las de clulas, pero si se aaden protenas, se reduce la tensin superficial y las membranas se aplanan hacia fuera.
-
Robertson (1950) no vio ningn espacio para poros en las microfotografas electrnicas. Y supuso que el aspecto de la lnea de ferrocarril provena de unin del tetraoxido de osmio (usado en la ME) a protenas y los grupos polares de lpidos.
-
Lenard y Singer (1966 ) observaron que ms del 30 % de protenas de la membrana se encontraban como alfa hlice. Esto debido a que probablemente que haba muchas protenas esfricas.
-
Singer estudi los fosfolipidos de las bicapas y encontr que pueden formar una superficie aplanada sobre el agua, sin la exigencia de una cubierta de protenas. El punto decisivo en el modelo se produjo con el advenimiento de tcnicas de criofractura . La figura muestra el interior de una membrana y las prominencias, surcos, arrugas ". ms tarde se encontro que eran protenas.
-
La Arquitectura Bsica de la membrana
Hasta aqu hemos visto la unidad bsica de membrana. Todas las membranas contienen protenas y lpidos. Sin embargo, la proporcin de cada uno vara dependiendo la membrana. Por ejemplo:
La mielina, que asla fibras de nervio, contiene18 % de protenas y 76 % de lpidos.
-
La membrana mitocondrial interna contiene 76 % de protenas y slo del 24 %. de lpidos.
Las membranas celulares de glbulos rojos humanos y el hgado de ratn contienen las cantidades casi iguales (44, el 49 % respectivamente) de protenas y (43, el 52 % respectivamente) de lpidos.
-
La arquitectura de la membrana es la de una bicapa lipdica los lpidos son anfipticos, por lo que sus cabezas polares se orientan al exterior y la porcin hidrofbica al interior.
Las protenas estn embebidas en la bicapa, pasan a travs de de la bicapa (como protenas transmembrana, o pueden estar insertadas en la superficie citoplasmtica o en la externa.
-
En un corte transversal de la bicapa se puede observar las molculas de lpidos con una cabeza globular (polar) y una regin recta (no polar).
Cada fila de lpidos forma una capa. Por lo tanto, la membrana celular consiste de una bicapa con las regiones no polares dirigidas hacia adentro.
-
Los fosfolpidos de membrana
Uno de los tipos principales de lpidos en la membrana son los fosfolpidos Estos tienen una cabeza con un grupo polar y dos colas hidrocarbonadas.
La regin superior que comienza con el NH3 es el grupo polar. Esta unido por glicerol a dos colas de cidos grasos. Una de las colas es un cido graso de cadena recta (saturado). El otro tiene un doblez en la cola debido al doble enlace cis insaturado
-
Componentes lipdicos de la membrana plasmtica, la capa externa contiene predominantemente fosfatidilcolina, esfingomielina, y glicolipidos, mientras que la capa interna contiene fosfatidiletanolamina, fosfatidilserina y fosfatidil inositol (-). El colesterol se distribuye en ambas capas.Las cargas negativas de las cabezad de los gupos de fosfatidil serina y fosfatidiil inositol se encuentran marcadas grupos polares Los fosfolpidos se distribuyen asimetricamente en las mono capas
-
Dos roles criticos de la bicapa
La estructura de los fosfolipidos es responsable de la funcin bsica de la membrana como barrera entre dos compartimientos acuosos. Ya que el interior de la bicapa est ocupado por las cadenas hidrfbicas de los cidos grasos, la membrana es impermeable a molculas solubles en agua incluyendo iones y la mayora de molculas biolgicas .
Las bicapas son fludos viscosos no solidos la mayora de cidos grasos tienen uno o dos dobles enlaces que producen dobleces en la cadenas hidrocarbonadas y no les permite empacarse, las cadenas se mueven libremente en el interior de la membrana que se presentan como una estructura suave y flexible, adems los fosfolpidos y protenas son libres de difundir lateralmente en la membrana una propiedad crtica para muchas funciones.
-
La bicapa lipdica proporciona a la membrana sus caracterstica de fluidez.
-
Colesterol en la Membrana
La cantidad de colesterol vara con el tipo de membrana , las membranas plasmticas tienen casi una molcula de colesterol por molcula de fosfolpido, otras membranas (bacterias) no tienen colesterol .
-
Las molculas de colesterol no forman membranas por si mismos, se insertan en la membrana con la misma orientacin que las molculas de fosfolpidos
-
Debido a la rigidez de su estructura el anillo de colesterol juega un rol distinto en la membrana,
Dependiendo de la temperatura el colesterol tiene distintos efectos en la fluidez:
A elevadas temperaturas el colesterol interfiere con el movimiento de las cadenas de los cidos grasos de los fosfolpidos, produciendo menor fluidez en la parte externa de la membrana y reduce su permeabilidad a pequeas molculas
A bajas temperaturas, el colesterol tiene efecto opuesto por que interfiere con interacciones entre las cadenas de los cidos grasos,evitando el congelamiento y mantieneniendo la fluidez de la membrana
Aunque el colesterol no esta presente en las bacterias, es un componente esencial de las membranas plasmaticas celulares, las plantas carecen de colesterol pero contienen esteroles relacionados con funcin similar .
-
Los glicolpidos se ubican exclusivamente en la capa exterior con sus porciones de carbohidratos expuestas en la superficie celular, son los componentes menores de la membrana, 2% de todos los lpidos
Ellos pueden microagregarse en la membrana, para proteger, aislar, o servir de sitios receptores de ligandos. Entre las molculas que se unen a travs de glicoesfingolpidos se incluyen venenos, toxinas del colera y tetano.
-
Protenas de Membrana
Jonathan Singer and Garth Nicolson 1972, proponen el modelo de mosaico fluido paradigma bsico de la organizacin de todas las membranas biolgicas. En el la membrana se ve como dos fluidos dimensionales en el cual las protenas se insertan en las bicapas.
-
Mdodelo de mosaico fluido, las protenas estan insertadas en la bicapa, las protenas perifericas se unen por interacciones protena-protena, la mayora de protenas integrales de membrana son protenas transmembrana con porciones que se exiben a ambos lados de la bicapa. La porcin extracelular de estas protenas por lo general estn glicosiladas, bilayer.
-
Formacion de "Microdominios
No todos los lpidos difunden libremente, determinados dominios de membrana ricos en colesterol y esfingolpidos (esfingolmielina y glicolpidos), funcionan como balsas o plataformas a la que se adhieren protenas especficas, conforme la membrana se mueve alrededor de la clula y tambin durante las seales de transduccin, sealamiento celular y en la captacin de molculas extracelulares por endocitosis.
-
La funcin de las diferentes organelas , es dependiente de las protenas de sus membranas . Las protenas transmembrana son anfipticas, presentan regines hidrofbicas e hidrofilicas que estn orientadas en en las mismas regiones de la bicapa lipdica. Tambien se les conoce como protenas integrales
Otras de protenas pueden ligarse solo a la superficie citoplasmtica (por union a un cido graso) o a la superficie externa unida a un oligosacarido. O estas protenas que no son transmembrana pueden estar unidas a otras protenas de membrana. Estas se denominan protenas perifericas.
Protenas que se insertan una vez a travs de la membrana son llamadas protenas transmembrana de simple paso. Aquellas que atraviesan varias veces la membrana son llamadas protenas transmembrana multipaso, estas forman asas por fuera de la membrana.
-
Asociacin del citoesqueleto cortical del eritrocito con la membrana plasmatica . La membrana esta asociada con una red de tetrameros de espectrina ligados por entrcuzamieno por cortos filamentos de actina en asociacin con la protena 4.1. La red de espectrian actina est ligada a la membrana por ankirina que une tanto a espectrina como y banda 3 otra unin adicional la proporciona la union de la protena 4.1 a glicoforina.
-
Banda 3 es un transportador de Bicarbonato y cloruro, 929 AA y tiene 14 regiones de alfa helice que atravieza la membrana, dimeros de banda 3 forman una estructura globular con canales internos por los cuales los iones atraviezan la bicapa.
-
Un centro de reaccin fotosinttico de Rhodoseudomonas con tres protenas transmembranas L (rojo), M (amarillo), y H (verde) Las subunidades L y M tienen 5 helices transmembranas , mientras que H solo una. La cuarta subunidad es un citocromo (blanco) que es una protena periferica..
-
Membrana externa bacterial. La membrana plasmtica de algunas bacterias est rodeada por una pared celular y a una membrana externa diferente. La membrana externa contiene porinas (Barriles en Hojas beta)que forma canales cauosos abiertos que permitene el paso de iones y pequeas molculas.
-
una variedad de protenas integrales de membrana estn acncladas en la membrana por uniones coovalentes con glicosilfosfatidilinositol (GPI)
-
Mobilidad de las protenas de membrana
-
Celulas y Difusin
El agua, el dixido de carbono, y el oxgeno estn entre las pocas molculas simples que pueden cruzar la membrana de clula por la difusin (o un tipo de difusin conocida como smosis).
La difusin es la principal forma de movimiento de sustancias dentro de clulas, as como para el paso de pequeas molculas esenciales a travs la membrana celular.
El intercambio de gas en agallas y pulmones funciona por este proceso. El dixido de carbono es producido por todas las clulas como consecuencia de procesos celulares metablicos. Ya que la fuente de produccin esta dentro de la clula, el gradiente de concentracin se mantiene constantemente, as el flujo neto de CO2 es hacia fuera de la clula.
Procesos metablicos en animales y plantas por lo general requieren el oxgeno, que est en la concentracin inferior dentro de la clula, as el flujo neto de oxgeno es hacia el interior de la clula.
-
La smosis es la difusin del agua a travs una membrana semipermeable (o diferencialmente permeable o selectivamente permeable).
En la membrana de clula, as como en los tubos de dilisis, cubiertas de acetato de celulosa. La presencia de un soluto disminuye el potencial de agua de una sustancia. As hay ms agua por unidad de volumen en un cristal de agua dulce que en un volumen equivalente de agua de mar. En una clula, que tiene muchas organelas y otras molculas grandes, el flujo de agua es generalmente hacia la clula.
-
Soluciones hipertonicas son aquellas en las cuales el soluto esta presente en mayor concentracin respecto a la celula (menor potencial del agua)
Soluciones hipotonicas son aquellas en las cuales el soluto esta presente en menor concentracin respecto a la celula (mayor potencial del agua)
Soluciones isotonicas tienen igual concentracion, los potenciales del agua son iguales aunque hay desplazamiento de agua al interior y al exterior, el flujo neto es cero
Relaciones del agua y la forma de los eritrocitos
-
Relaciones del agua con una clula vegetal
-
Una de las funciones principales de sangre en animales es mantener un medio ambiente isotonico interno. Esto evita los problemas asociados con la prdida o el incremento de agua dentro o fuera de las clula (homeostasis).
Paramecium y otros organismos unicelulares de agua dulce tienen la dificultad ya que ellos son por lo general hipertnicos en relacin con su ambiente exterior. As el agua tender a fluir a travs de la membrana de la clula, hinchando la clula y eventualmente reventndola. La Vacuola contrctil del Paramecium responde a este problema. El de bombeo del agua fuera de la clula por este mtodo requiere energa ya que el agua se mueve contra gradiente de concentracin.
Ya que los ciliados (y muchos protozoarios de agua dulce) son hipotonicos, la remocin del agua a travs de la membrana de clula por la smosis es un problema significativo. Un mecanismo comnmente empleado es vacuola contrctil. El agua es recogida en el anillo central de la vacuola y activamente transportada de la clula.
-
El funcionamiento de una vacuola contrctil en Paramecium
-
Transporte Pasivo y ActivoEl transporte pasivo no requiere energa de la clula, ejemplos: difusin de oxgeno y dioxido de carbono, osmosis del agua y difusin facilitada
Tipos de transporte pasivo
-
Tipos de transporte activo El transporte activo requiere el aporte de energa por la clula, usualmente en forma de ATP. Ejemplos: el transporte de grandes molculas (no solubles en lpidos) y la bomba de sodio-potasio
-
Transporte mediado a travs de un transportador
Las protenas de transporte integradas en la membrana de clula son muy selectivas con las sustancias que van a transportar. Algunas de estas protenas pueden mover materiales a travs de la membrana slo cuando son ayudadas por un gradiente de concentracin, un tipo de transporte mediado es conocido como difusin facilitada. La difusin y la difusin facilitada se producen por las diferencias de energa potencial de un gradiente de concentracin.
La glucosa ingresa en la mayora de clulas por difusin facilitada. Al parecer hay un nmero limitado de protenas que transportan glucosa. La degradacin rpida de glucosa en la clula (proceso conocido como gluclisis) mantiene el gradiente de concentracin. Sin embargo, cuando la concentracin externa de glucosa se incrementa, el transporte de glucosa no excede de una cierta velocidad, lo cual sugiere una limitacin en el transporte.
-
En el caso de transporte activo, las protenas tienen que mover los solutos contra un gradiente de concentracin. Por ejemplo con la bomba de sodio-potasio de neuronas. Na+ se mantiene a concentraciones bajas dentro de la clula y el K+ a concentraciones ms altas. Contrario al exterior de la clula.
Cuando un mensaje de nervio es propagado, los iones pasan a travs de la membrana, propagando el mensaje. Despus que el mensaje ha pasado, los iones deben ser transportados activamente retornando a sus " posiciones de partida " a travs de la membrana. Cerca de 1/3 del ATP es usado por un animal en estado de reposo por la bomba de Na-K.
-
Tipos de molculas de transporte
Uniport transportan un soluto en un tiempo
Symport transporta un soluto y otro soluto es cotransportado a la vez en la misma direccin.
Antiport transporta el soluto al interior (o hacia fuera) y otro soluto es co-transportado en direccin opuesta. Uno ingresa y el otro va hacia fuera o viceversa.
-
Las vesiculas y vacuolas que se pueden fusionar con la membrana celular son utilizadas para liberar o transportar sustancias qumicas al exterior de la clula; o para permitir el ingreso de ellas. Exocitosis es el termino aplicado cuando el transporte es al exterior de la clula.
-
Endocitosis es el caso cuando una molcula hace que la membrana celular forme una concavidad hacia adentro, formndose una vescula.
La fagocitosis es el tipo de endocitosis en el cual una clula entera es ingresada.
Pinocitosis se refiere al ingreso de un fluido externo.
La endocitosis mediada por un receptor ocurre cuando el material para ser transportado se une a ciertas molculas especficas de la membrana. Ejemplos: el transporte de insulina y colesterol en clulas de animal.
-
Endocitosis y exocitosis
-
El glicocalix es una cubierta de carbohidratos formada por oligosacaridos, glicolpidos y glicoprotenas transmembrana protejen a la superficie celular y sirven como marcadores de una serie de interacciones clula-clula.
Un ejemplo de estas interacciones es la adhesion de leucocitos a celulas endoteliales de vasos sanguineos que media la respuesta inflamatoria, en este proceso intervienen las protenas selectinas que reconocen especficamente carbohidratos de la superficie celular.
-
Celula del epitelio intestina l polarizada; la superficie apical contiene microvellosidades especialzada en absorcin de nutrientes, del intestino al lumen , la superficie basolateral esta especializada en la transferencia de nutrientes absorvidos al tejido conectivo adyacente que contiene capilares sanguineos . Uniones estrechas separan los dominios apicales y basolaterales , las protenas de membrana son libres de difundir en cada dominio pero no cruzan de un dominio a otro.
-
La pared celular es una estructura rgida de prokaryotes, contiene peptidoglicanos, da la forma a la clula y rodea la membrana citoplasmatica. Proteje a la clula del ambiente.
las bacterias gram negativas tienen membrana externa, similar a la membrana celular, pero menos permeable y compuesta de lipopolisaccharides (LPS). LPS es una sustancia clasificada como un endotoxin.
-
Gracias por la atencion prestada