T.7 Intro Al Metabolismo
29
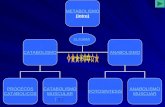
TEMA V II. I ntro duc ción al metabolismo Factores de supervivencia son senyales para que una célula se mantenga viva. Hay algunas células que no pueden fallar, morir o funcionar mal, para evitar esto estan todo el rato canviando su material para evitar que envejezcan y se oxiden. Una célula va a ploriferar, crecer si le interesa al organismo. La mayoria de las enfermedades neurodegenretiva s y los cáncers dependen de las proteïnas. Transformación oncogenica: canvió de una proteïna que puede provocar un inició de cáncer. CONCEPTOS BÁSICOS Metabolismo: actividad eficazmente coordinada en la que muchos sistemas multiezimáticos cooperan para: • Obtener energía química captando energía solar o degradando nutrientes. • Convertir los nutrientes en moléculas característica de la célula • Sintetizar macromoléculas a partir de precursores monoméricos • Sintetizar y degradar las biomoléculas que realizan funciones celulares específicas. Vía metabólica: conjunto de reacciones enzimáticas encadenadas mediante las cuales un precursor se convierte en producto a través de una serie de intermediarios metabólicos o metabolitos. Como una cadena de montaje, el producto de la primera es el sustrato de la siguiente. No hay ninguna máquina que su rendimiento supere el 12%. En el nuestro cuerpo el rendimiento es del 40-60 % hasta el 80%!! Ya que hacemos muchos pasos de oxidación. Metabolismo intermediario: vías que interconvierten presursores metabolitos y productos de bajo peso molecular ( <1.000 Da). Fuentes de energía y materia para los seres vivos • Ciclos del CO 2 y del O 2 entre organismos autótrofos y heterótrofos. Los autótrofos sintetizan la glucosa y todos sus demás compuestos orgánicos a partir del carbono inorgánico obtenido en forma de CO 2 , produciendo además O 2 . Los heterótrofos pueden sintetizar sus metabolitos orgánicos únicamente a partir de compuestos orgánicos por lo tanto han de consumir y también necesitan de O 2 y pruducen CO 2 . Se recambian unas 4 x 10 11 toneladas anuales de carbono en la biosfera. 1
Transcript of T.7 Intro Al Metabolismo

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 1/29
TEMA V II. Introducción al metabolismo
Factores de supervivencia son senyales para que una célula se mantenga viva.Hay algunas células que no pueden fallar, morir o funcionar mal, para evitar esto estantodo el rato canviando su material para evitar que envejezcan y se oxiden.Una célula va a ploriferar, crecer si le interesa al organismo.La mayoria de las enfermedades neurodegenretivas y los cáncers dependen de lasproteïnas.
Transformación oncogenica: canvió de una proteïna que puede provocar un inició decáncer.
CONCEPTOS BÁSICOS
Metabolismo: actividad eficazmente coordinada en la que muchos sistemasmultiezimáticos cooperan para:
• Obtener energía química captando energía solar o degradando nutrientes.• Convertir los nutrientes en moléculas característica de la célula
• Sintetizar macromoléculas a partir de precursores monoméricos
• Sintetizar y degradar las biomoléculas que realizan funciones celularesespecíficas.
Vía metabólica: conjunto de reacciones enzimáticas encadenadas mediante las cualesun precursor se convierte en producto a través de una serie de intermediariosmetabólicos o metabolitos.Como una cadena de montaje, el producto de la primera es el sustrato de la siguiente.
No hay ninguna máquina que su rendimiento supere el 12%. En el nuestro cuerpo elrendimiento es del 40-60 % hasta el 80%!! Ya que hacemos muchos pasos deoxidación.
Metabolismo intermediario: vías que interconvierten presursores metabolitos yproductos de bajo peso molecular ( <1.000 Da).
Fuentes de energía y materia para los seres vivos
• Ciclos del CO2 y del O2 entre organismos autótrofos y heterótrofos.
Los autótrofos sintetizan la glucosa y todos sus demás compuestos orgánicos a partirdel carbono inorgánico obtenido en forma de CO2 , produciendo además O2.Los heterótrofos pueden sintetizar sus metabolitos orgánicos únicamente a partir decompuestos orgánicos por lo tanto han de consumir y también necesitan de O2 ypruducen CO2.
Se recambian unas 4 x 1011 toneladas anuales de carbono en la biosfera.
1

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 2/29
• Ciclo del nitrógeno en la biosfera.La fuente más abundante de nitrógeno es el aire, que en sus cuatro quintas partes esnitrógeno molecular (N2). Sin embrago, relativamente pocas especies pueden convertir
el nitrógeno atmosférico en formas utilizables por los organismos vivos.El primer paso del ciclo del nitrógeno consiste en la fijación (reducción) del nitrógenoatmosférico por parte de las bacterias fijadoras de nitrógeno para proporcionaramoníaco (NH3 o NH+
4) Las bacterias del suelo son tan abundantes y activas queprácticamente todo el amoníaco que llega al suelo se oxida a nitratos, el amoníaco lopasan a nitrito ( NO-
2) y finalmente a nitrato (NO-3). Este proceso se conoce como
nitrificación. Las plantas y muchas bacterias pueden incorporar y reducir fácilmentenitrato i nitrito por acción de nitrato y nitrito reductasa. El amoníaco así formado esincorporado a los aminoácidos por las plantas. Los animales a su vez utilizan las plantascomo fuente de aminoácidos, tanto esenciales como no esenciales, para fabricar sus
proteínas. Lo expulsan en forma de amoníaco.El equilibrio entre el nitrógeno fijado y atmosférico es mantenido por las bacterias queconvierten el nitrato en N2 en condiciones anaeróbicas proceso denominadodesnitrificación. También pueden dar amoníaco.
2

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 3/29
• Flujo de energía en la biosfera.
RELACIONES ENERGÉTICAS ENTRE VÍAS ANABÓLICAS Y CATABÓLICAS
A partir de los nutrientes los oxidamos y formamos productos oxidados y hacemosATP, NADH y FADH2 (son básicamente e-).
Cuantos más electrones tenga una molécula mejor combustible será ya que nuestrometabolismo se basa en el intercambio de e-. Estos e- son llevados a la cadena de e-
que lleva a ATP.La filosofía es coger e- de donde sea y obtener ATP.
3

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 4/29
ENERGÍA LIBRE DE OXIDACIÓN DE COMPUESTOS MONOCARBONADOS
OH son descompensados ya que al ser el O y el N muy electronegativos el H estaráNH muy cerca y los electrones se quedaran porque no los podemos arrancar.
QUANTOS MÁS H HAYA HAY MÁS E- PAR ARRANCAR. A más H mejor combustible.
PRINCIPALES COMBUSTIBLES
Las grasas son mejores fuentes de enregía porque están más reducidas.Fatty acid: Tienen muchos H, por lo que los e- se pueden arrancar facilmente.
Glucosa: hay tambien H pero la mayoria unida a O y no se puede coger.
Comamos lo que comamos una vez llenados los depositos básicos los excedentes sevan a almacenar en forma de grasa, formando lipidos porque están más reducidos.Aunque uno podria vivir mucho tiempo solo de grasa (ayunas) moriría por falta deglucosa, necesaria por el cerebro.
TRANSFERENCIAS DE ELECTRONES EN LAS CÉLULAS
4

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 5/29
Las células vivas tienen un “circuito” biológico análogo, con un compuestorelativamente reducido, tal como la glucosa como fuente de electrones. A medida quela glucosa se oxida enzimáticamente, los electrones liberados fluyen espontáneamentea través de una serie de transportadores de electrones intermedios a otra especiequímica, tal como el O2. Este flujo de electrones es exergónico, porque el O2 tiene unaafinidad por los electrones mayor que la que tienen los transportadores de electronesintermedios.Los electrones, en la célula, se transfieren de una molécula (dador de electrones) a
otra (aceptor de electrones) de cuatro formas diferentes, todas ellas catalizadas por lafamilia de enzimas OXIDOREDUCTASAS:
1. Directamente como electrones. Por ejemplo, el par redox Fe2+/Fe3+ puede transferirun electrón al par redox Cu+/Cu2+ (OXIDADAD, REDUCTASAS):
2. En forma de átomos de hidrógeno. Recuérdese que un átomo de hidrógeno consisteen un protón (H+) y un único electron (e-). En este caso podemos escribir la ecuacióngeneral catalizada por DESHIDROGENASAS:
en donde AH2 es el dador de hidrógeno/electrón. (No confundir la reacción anteriorcon una disociación de ácido en la que interviene un proton pero no un electron). AH2y A constituyen conjuntamente un par redox conjugado (A/AH2), que puede reducirotro compuesto B (o un par redox B/BH2) por transferencia de átomos de hidrógeno.
3. En forma de ión hidruro (:H-), que tiene dos electrones. Esto sucede en el caso de lasdeshidrogenasas ligadas a NAD.
4. A través de una combinación directa con oxígeno. En este caso, el oxígeno secombina con un reductor orgánico y se incorpora de forma covalente en el producto,tal como sucede en la oxidación de un hidrocarburo a alcohol:
FORMAS OXIDADA Y REDUCIDA DE LOS NUCLEÓTIDOS DE FLAVINA
Las flavoproteínas son enzimas que catalizan reacciones de oxidación-reducciónutilizando flavina mononucleótido (FMN) o flavina adenina dinucleótido (FAD) comocoenzima. Estos coenzimas, los nucleótidos de flavina, provienen de la vitaminariboflavina. La estructura de anillos fusionados de los nucleóridos de flavina ( anilloisoaloxazina) se reduce de manera reversible, aceptando uno o dos electrones en
forma de uno o dos átomos de hidrógeno (cada átomo consiste en un electron más unproton) desde un sustrato reducido.
5

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 6/29
Las formas completamente reducidas se abrevian FMNH2 y FADH2. Cuando unnucleótido de flavina totalmente oxidado acepta solo un electron (un átomo dehidrógeno) se produce la forma semiquinona del anillo de isoaloxazina, abreviada
FADH· y FMNH
·. Debido a que los nucleótidos de flavina tienen una especialidad
química algo diferente de la de los coenzimas nicotinamida (la capacidad de participartanto en transferencias de uno como de dos electrones), las flavopreteínas intervienenen una mayor variedad de reacciones que las deshidrogenasas ligadas a NAD(P).
FORMAS OXIDADA Y REDUCIDA DE LOS DINUCLEOTIDOS DE NICOTINAMIDA YADENINA
La nicotinamide adenina dinucleótido (NAD;NAD+ en su foram oxidada) y su análogopróxima, el nicotinmaida adenina dinucleótido fosfato (NADP; NADP+ en su formaoxidada) están compuestos por dos nucleótidos unidos a través de sus grupos fosfatopor un enlace fosfoanhídrido. Debido a que el anillo de nicotinamida se parece a lapriridina, a estos compuestos se los denomina a veces nucleótidos de pirindina. Lavitamina niacina (PP factor o preventivo de pelagra) es la encargada de aportar laporción nicotinamida de lso nucleótidos de nicotinamida.Ambos coenzimas experimentan reducción reversible del anillo nicotinamida. Mientrauna molñecula de sustrato es oxidada (deshidrogenasión, cediendo dos átomos de
hidrógeno, la forma oxidada del nucleótido NAD+
o NADP+
) aceptan un ión hidruro( :H-, el equivalente de un protón y dos electrones) y se transforma en la fromareducida (NADH o NADHP). El segundo protón eliminado dels sustrato se libera aldisolvente acuoso. Las semirreacciones para estos cofactores nucleotídicos son:
6

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 7/29
CARACTERÍSTICAS GENERALES DE LAS VÍAS CATABÓLICA/ANABÓLICAS
• Irreversibles, muy exergónicas. Liberan e- que producen procesos de oxidaciónliberando eenrgia que es utilizada.
- Reacciones enzimáticas diferentes ( en los pasos claves de las vías para regularla vía y que no sean reversibles)- Regulación independiente
• Tienen almenos una etapa obligada.
- Reacción irreversible, generalmente al principio - Etapa limitante de la velocidad de la vía
7

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 8/29
• Tienen lugar en compartimientos específicos
• Catabolismo y anabolismo del mismo tipo de biomóleculas no tienen lugarsimultáneamente en la misma célula.Cuando hacemos catabolismo oxidamos y cuando hacemos anabolismoreducimos. Si se activa la glucolisi se inactiva la gluconeogenesi.
¿las vías anabólicas son exergónicas?
Una reacción de sintesi sera siempe endergónica, pero la vía es exergónica ya que elATP que gastamos es más del que necesitamos y el balance total es exergónico y haceque la vía funcione.
REGULACIÓN DE LA ACTIVIDAD DE ENZIMAS QUE CATALIZAN ETAPAS LIMITANTES
Generalmente al principio.
• Alosterismo. Respuesta inmediata a cambios en la concentración demetabolitos.
• Adaptacion gènica: por los factores de transcripción• Alteraciones en la concentración de enzima. Regulación a largo plazo por
cambios en la velocidad de síntesis o de degradación. Puede producirse enrespuesta a hormonar
• Modulación covalente. Fosforilación de proteínas.
Es una reacción catalizada por quinasas: catalizan la transferencia de un fosfato ά(último) del ATP a residuos de Ter o Tyr.Enzimas fosfatasas: puede canviar el estado de actividad.Cuando fosforilamos una molécula tambien biene O con dos cargas negativas.
3. Lo que nos hace más complejos capacidades de interacción entre proteínas, unaunion de diferentes proteínas puede dar funciones diferentes.4. Puede producir ubiquitinación.
8

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 9/29
CONTROL DEL METABOLISMO POR LA PROTEÍNA KINASA ACTIVADA POR AMP(AMPk)
Hipoxia: utilizando O2 para oxidar.+ ATP más estado energético, ATP+ADP+AMP siempre constante.
TRANSMISIÓN INTERCELULAR DE INFORMACIÓN
• Tipos de mensajeros o señales
- Hormonas- Neurotransmisores- Factores de crecimiento- Ferormonas- Olores- Sabores- Luz- Componentes de la matriz extracelular- Antígenos
• Acciones a nivel molecular- Alteración en el flujo de iones- Regulación de la actividad de enzimas- Inducción de la síntesi de proteínas
9

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 10/29
- Activación de sistemas de transporte
TIPOS DE SEÑALIZACIÓN INTERCELULAR
- Señales endocrinas: Las hormonas son producidas por células del sistema endocrino
y circulan por el torrente sanguíneo hasta alcanzar todos los lugares del cuerpo. Es de
respuesta lenta, inespecífica, larga duración y actúa a distancia.
- Señales paracrinas: Sólo actúan sobre células diana que se encuentran en la vecindad
de las células emisoras, como por ejemplo los neurotransmisores.
- Señales autocrinas: Afectan sólo a las células que son del mismo tipo celular como las
células emisoras. Un ejemplo de señales autocrinas se encuentra en las células del
sistema inmune.
- Señales yuxtacrinas: Son transmitidas a lo largo de la membrana celular a través de
proteínas o lípidos que integran la membrana celular y son capaces de afectar tanto a
la célula emisora como a las células inmediatamente adyacentes.
RECEPTORES: Moléculas proteicas a las que se asocian los mensajeros intercelulares
de forma reversible y específica.
• Receptores nucleares- Hormonas esteroideas, tiroideas, retinoides- Complejo hormona-receptor altera la expresión génica- Respuestas fisiólogicas lentas
• Receptores de membrana- Proteínas, péptidos, aminas, eicosanoides, olores, sabores, luz,...- Tipo de receptor:
Receptores-canal iónico
Receptores acoplados a proteínas GReceptores con actividad enzimáticaReceptores asociados a tirosina quinasas
• Respuestas bioquímicas y fisiológicas rápidas y lentas
10

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 11/29
DIFERENCIAS BÁSICAS EN LA SEÑALIZACIÓN A TRAVÉS DE RECEPTORESINTRACELULARES Y DE MEMBRANA
Receptores intracelularesAlgunas hormonas son liposolubles y atraviesan la membrana plasmática por difusiónsimple para unirse a receptores intracelulares. Estos son capaces de entrar en el
núcleo y alterar la transcripción de genes según lo que le transmita la hormona.
Receptores de membranaLos receptores transmembrana son proteínas que se extienden por todo el espesor dela membrana plasmática de la célula, con un extremo del receptor fuera de la célula(dominio extracelular) y otro extremo del receptor dentro (dominio intracelular).Cuando el dominio extracelular reconoce a una hormona, la totalidad del receptorsufre un cambio en su conformación estructural que afecta al dominio intracelular,
confiriéndole una nueva acción. En este caso, la hormona (u otro ligando) no atraviesala membrana plasmática para penetrar en la célula. Aunque un receptor sencillo puedetransducir alguna señal tras la unión del ligando, lo más frecuente es que la unión delligando provoque la asociación de varias moléculas receptoras.
11

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 12/29
PRINCIPALES TIPOS DE RECEPTORES DE MEMBRANA
a) Canal ionico de entrada regulada: se abre o se cierra en respuesta a laconcentración del ligando señal o al potencial de membrana.
b) Receptores ligados a enzimas: Los receptores de membrana ligados a enzimas
tienen la particularidad de tener actividad de receptor en el dominioextracelular de su molécula, y actividad enzimática en el dominio intracelular ocitosólico. Presentan dos estados conformacionales; en ausencia de moléculade señalización, el enzima está inactivo, en el otro estado, la unión al mediadorproduce un cambio conformacional en el receptor que activa a la enzima.
c) Receptores asociados a tirosina quinasa: la unión del ligando activa la actividadtirosina quinasa por autofosforilación. Son también enzimas, catalizan lafosforilación de diversas proteínas. Insulina o factor de crecimiento epidérmico(EGF-R)
d) Receptores acoplados a proteínas G: La unión de un ligando externo al
receptor activa una proteína de unión de GTP, (G), que regula un enzima quegenera un segundo mensajero intracelular. Es un tipo de receptor como β-adrenérgico qu detecta adrenalina (epinefrina)
12

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 13/29
EL MISMO MENSAJERO INTRACELULAR PUEDE EJERCER DIFERENTES ACCIONES ENDISTINTOS TEJIDOS ACTUANDO A TRAVÉS DE RECEPTORES DIFERENTES
El efecto del mensajero dependera de la unión con el receptor, un mismo mensajerohace diferentes funciones si se une con otro receptor diferente.
MECANISMO GENERAL DE LA TRANSDUCCIÓN DE SEÑALES
Tres factores explican la extraordinaria sensibilidad de lso transductores de señal: laelevada afinidad de los receptores para las moléculas señal, la cooperatividad (amenudo pero no siempre) en la interacción ligando y receptor y la amplificación deseñal por cascadas enzimáticas. La información que lleva la hormona va a acativardiferentes enzimas y estos a otros, en pocos segundos E1 puede activar a muchos
millones de enzimas.
13

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 14/29
TRANSDUCCIÓN DE SEÑALES A TRAVÉS DE RECEPTORES ACOPLADOS A PROTEÍNAS G
La transducción de señal, a através de receptores acoplados a proteínas G (GPRC) estádefinido por tres componentes esenciales: un receptor de membrana plasmática consiete segmentos helicoidales transmembrana, un enzima efector en la membranaplasmática que genera un segundo mensajero intracelular y una proteína fijadora de
un nucleótido de guanosia (proteína G) que activa el enzima efector.
La unión de la hormona al receptor provoca canvios conformacionales al interior de lacélula. Se van activar unas proteínas (G) estas activan a unos enzimas (efectores)una vez activados se va a producir unas moléculas (segundos mensajeros vidamedia muy corta) que tienen de función activar otras proteínas, la mayoria decasos proteínas quinasas.
Las quinasas fosforilan proteínas, canvaindo la estructura de la proteína canviando sufunción.Al final se produce la respuesta celular, es un procéso específico.Hay dos tipos:
- Proteínas mismo receptor tiene actividad quinasa- receptores tienen proteínas G.
14

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 15/29
Estructura de los principales mensajeros secundarios
AMP cíclico: rompe enlaces fosfodiésteres: actividad fosfodiesterasa.Diacilglicerol= 2ácidos grasos + glicerol
Fosfatidilinositol: segundo mensajero de la insulina.
Estructura de receptor acoplado a proteína G
El recepetor β-adrenégenico es una proteína integrals con siete regiones hidrofóbicasde 20 a 28 residuos de aminoácidos que serpentean de un lado a otro de la membranaplásmatica, tienen tambien un dominio N-terminal extracelular dónde se une lahormona y un domino C-terminal dónde se acopla la proteína G.
15

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 16/29
PROTEÍNAS G
Son proteínas periféricas de membrana,se activan si forman GTP, formadas porhetertrímeros αβγ que unen GDP/GTP en la subunidad alfa que además tiene lapropiedad de romper el GTP a GDP (actividad hidrolasa o GTPasa) para inactivarse.- Formas activas: subunidad α con GTP o subunidades βγ libres.
Como consecuencia de que el receptoru una una hormona se produce un canvioconformacional, que provoca la union de GTP en la subunidad α de la proteína G está
subunidad se separa de βγ y puede desplazarse por la membrana y activar otraproteína. Entonces la propia actividad hidrolasa hdroliza el GTP a GDP y vuelve areunirse con el receptro y las subunidades.Extisten tambien factores que regulan la actividad de las proteínas G:- Factores intercanviadores de nucleótidos de guanina (Guanine-nucleotide exhangefactor GEF) ayudan a que se activen más rapidamente.- Factores activadores de actividad GTPsas (GTPase-activating proteins GAP) ayudan ahidrolizar GTP a GDP.
16

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 17/29
1, 2, 3 β.Sistema receptor/proteína G/adenilil ciclasa
Activamos el adenilil ciclasa , la unión de la subunidad α provoca un canvió
conformacional al adenilil ciclasa y la activa.
La adenilil ciclasa es una proteína integral de membrana plasmática con sus sitio activoen la cara citósilica. La asociación de Gsα activa con la adenilil ciclasa estimula a laciclasa a catalizar la sintesi de cAMP a partir del ATP.El AMPciclico es capaz de activar a una proteína quinasa.Proteína quinasa A o PKA: - Dos subunidades que catalizan
- Dos subunidades reguladorasEl AMPciclico se une a estas ultimas y las subunidades catalizadoras son liberadas y yapueden fosforilar otras proteínas.
17

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 18/29
Proteína fosfatasa: proteína antagónica a la quinasa, ya que desfosforila.
Hay células que tienen los dos receptores ( activadore y inhibidores )
18

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 19/29
Acción del AMPcíclico vía PKA sobre la expresión génica
Las proteínas quinasas cuando llevan cierto tiempo activas van a parar al nucleo, qllí pueden canviar la conformación de los factores de transcripción (por fosforilación) ycanviar la expresión de genes.A corto plazo fosforilación de proteínas, a largo plazo canvió de expresión de genes.
SISTEMA PROTEÍNA G-FOSFOLIPASA C: SEGUNDOS MENSAJEROS DERIVADOS DEFOSFOINOSÍTIDOS. RECEPTOR α1.
Una segunda y amplia clase de GPCR están acoplados a través de una proteína G a unafosfolipasa C (PLC) de la membrana plasmática que es específica para el lípido demembrana fosfatidinositol 4,5-bifosfato, o PIP2. Cuando una hormona que actúa segúneste mecanismo se une a su receptor, el complejo hormona-receptor cataliza elintercanvio GTP-GDP en una proteína G asociada, Gq activandola. La Gq activada a suvez activa una PLC específica para PIP2 que cataliza de dos potentes segundosmensajeros diacilglicerol e inostol 1,4,5-trifosfato o IP3.
19

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 20/29
El inostol trifosfato IP3, hidrosoluble, difunde desde la membrana plásmatica alreticulo endoplásmatico (RE), donde se une a receptores específicos de canales de Ca2+
regulados por IP3 provocando su apertura (bomba SERCA), de manera que cuando seabren estos canales reguladores de Ca2+ y estos fluyen hacia el citosol.El diacilglicerol coopera con el Ca2+ en la activación de la proteína quinasa C (PKC) porlo que tambien actua como segundo mensajero.
EL ÓXIDO NÍTRICO, UN MENSAJERO INTERCELULAR ATÍPICO CON IMPORTANTESACCIONES EN EL TEJIDO VASCULAR, NERVIOSO E INMUNE
El gas óxido nitroso (NO) es un radical libre que difunde a través de la membranaplasmática y afecta a las células vecinas. El NO se forma a partir de la arginina y eloxígeno por la enzima óxido nitroso sintetasa, con citrulina como sustrato.
20

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 21/29
PAPEL DEL NO EN EL CONTROL DEL TONO VASCULAR
El NO funciona principalmente a través de receptores diana, la enzima solubleguanilato ciclasa, que cuando se activa produce el segundo mensajeroguanosinmonofosfato cíclico (GMPc). Que activa a la proteína kinasa G y induce a la
relajación del músculo.
En altas concentraciones el NO es tóxico, y se piensa que es el responsable de algunaslesiones después de un infarto. El NO realiza tres funciones principales:
• Relajación de los vasos sanguíneos.
• Regulación de la exocitosis de neurotransmisores.
• Respuesta celular inmune.
21

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 22/29
RECEPTORES CON ACTIVIDAD GUANILIL CICLASA
Las guanilil ciclasas son receptores enzimáticos que cuando se activan convierten elGTP en el segundo mensajero guanosina 3’,5’- monofosfato cíclico (GMP cíclico ocGMP).Muchas de las acciones del cGMP en animales se realizan mediante la proteína quinasa
dependiente de cGMP (PKG). Al activarse por acción del cGMP, la PKG fosforilaresiudos Ser y Thr en proteínas diana.
RECEPTORES CON ACTIVIDAD ENZIMÁTICA
• Tirosina quinasa (insulina, PDGF, EGF and FGF)
• Tirosina fosfatasa (CD45)
• Guanilil ciclasa (péptidos natriurétricos)
• Serina-Treonina quinasa (TGF-β)
RECEPTORES TIROSINA QUINASA
EJ: Receptor del factor de crecimiento epidérmico (EGF)
Son proteínas de membrana que tienen un dominió de unión en la cra extracelular yun sitio activo enzimático en el lado citósilico, con dos dominios conectados por unúnico segmento transmembrana. El dominio citoplásmatico es una proteína quinasaque fosforila a residuos de Tyr de proteínas diana específicasEn este caso son dímeros, el factor de crecimiento une ambos dimeros y se favoreceque se active el dominió tirosina quinasa.La fosforilación hace que vengan más proteínas y estas tienen otro dominio al que sepueden unir otras proteínas ( y estas permiten activar una vía).Todo se puede inhibir con un inhibidor de tyrosina quinasa
22

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 23/29
Papel de la proteína G pequeña RAS en la transducción de senyales mediante unreceptor con actividad tirosina quinasa
Una vez el recepter se autofosforila las tyrosinas fosfatadas son reconocidas por eldomini SH2 de unas proteínas llamadas Grb2 es una proteína adaptadora sin funciónenzimática su función es poner en contacto dos proteínas (en este caso EGF con Sos)que han de ineraccionar para la transducción de señal, esta se une a Sos mediante su
dominio SH3 que reconoce regiones ricas en resiuduos de Pro de Sos.Sos actua como factor de intercambio de nucleótidos de guanosina, GEF, catalizando lasustitución del GDP por GTP en RAS, una proteína G pequeña ya que es monomérica.
Papel de la cascada de las proteínas quinasas activadas por mitógenos (MAPKs)(ERK)en la transducción de señales de receptores con actividad tirosina quinasa
RAS va a activar a RAF (MAP-kinasa-kinasa-kinasa), no a fosforilarla, ya que esta seautofosforila pero en condiciones normales no puede porque tiene un dominioplegada en si mismo cerrando el centro activo e impidiendo que se una el ATP, quando
interactuen Raf va a poder abrir el dominio autoinhibidor y se va a autofosforilar.En este momento que RAS ya a echo su función GAP la va a desfosforilar y desactivarrapidamente.
23

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 24/29
RAF o MAP-kinasa-kinasa-kinasa va a fosforilar a MAP-kinasa-kinasa y esta va afosforilar a MAP-kinasa que puede translocar al nucleo y puede controlar la actividadde las ciclinas y la ploriferación celular.Las RAF y MEK van a ser desfosforiladas por proteínas fosfatasas.
RECEPTOR DE INSULINA
Los receptores de insulina ya son un dimero, la insulina provoca un canvioconformacional a α que va a pasar tambien a β que va a provocar una autofosforilaciónen los dominios de tyrosina quinasa.Las fosfotirosinas son reconocidas por diferentes proteínas (proteínas señalizadoras).
A diferencia de los receptores de los factores de crecimiento, el receptor tambienpuede fosforilar proteínas se llaman substartos del receptor de la insulina IRS.
24

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 25/29
Señalización a través del IRS
• INSULINA: evita que la célula sesuicide, actica la glucolisi, sintesi
de lipidos, inhibe traducción.
Receptor activado, autofosforiladoIRS: Sustrato del receptor deinsulina.
SEÑALIZACIÓN A TRAVÉS DEL IRS : Fosfatidilinistol 3-quinasa (PI3-K)
La ruta de señalización de la insulina se ramifica en IRS-1. Grb2 no es la únicaproteína que se asocia con IRS-1 fosforilado. El enzima fosfoinostol 3-quinasa (PI-3K) se une a IRS-1 a través del dominio SH2 de PI-3K. Activada de esta manera PI-3K convierte el lípido de membrana fosfatidilinostiol 4,5-biifosfato (PIP2) enfosfatidilinostiol 3,4,5-trifosfato (PIP3). El grupo de cabeza con múltiples cargas delPIP3, que sobresale en el lado citoplásmatico de la membrana plasmática, es elpunto de unión de una segunda rama de señalización en la que interviene otracascada de proteínas quinasas. Cuando esta unida a PIP3, la proteína quinasa B(PKB; Akt) es fosforilada y activada todavía por otra proteína quinasa PDK1. La PKBactivada fosforila entonces residuos de Ser o Thr de sus proteínas diana, una de lascuales es la glucógeno sintetasa quinasa 3 (GSK3).
25

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 26/29
Existe un mecanismo para terminar la actividad de la ruta PI-3K-PKB. Una fosfatasaespecífica de PIP3 (PTEN) elimina el grupo fosforilo en la posición 3 de PIP3 paradar PIP2, el cual ya no sirve como sitio de unión de PKB, con lo que se trunca la rutade señalización.En diversos tipos de cáncer a menudo se observa que el gen PTEN haexperimentado mutación. Fenómeno que da lugar a un circuito de regulacióndefectuoso y concentraciones anormalmente elevadas de PIP3 y actividad PKB. Elresultado parece ser una señal continua para la división celular y ,en consecuencia,el característico crecimiento tumoral.
REGULACIÓN DE LA EXPRESIÓN GÉNICA POR INSULINA. TRANSMISIÓN DE LAINFORMACIÓN AL NÚCLEO.
1. El receptor de insulina se une a insulina y experimenta autofosforilación enresiduos de Tyr carboxilo-terminales.
2. El receptor de insulina fosforila IRS-1 en sus residuos de Tyr.3. El dominio SH2 de Grb2 se une a la P-tyr de IRS-1. Sos de una a Grb2 y luego a
Ras, provocando la liberación de GDP y la unión de GTP a Ras.4. Ras activado se une y activa Raf-1.5. Raf-1 fosforila MEK en dos resiudos de SER, activándola. Mek fosforila ERK en
un resiudo de Thr y uno Tyr, activándola.6. ERK se meuev hacia dentro del núcleo y fosforila factores de transcripción
nucleares tales como Elk1, activándolos.
26

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 27/29
7. Elk1 fosforilada se une a SFR para estimular la transcripción y traducción degenes necesarios pra la división celular.
¿ Cómo la activación de un único receptor puede llevar a la activación dediferentes vías de señalización intracelulares?
Mecanismos de las proteínas señalizadoras
PROTEÍNAS “MODULARES”. DOMINIOS MODULARES
• Proteínas modulares: tambien llamasa prteínas armazón
• Desde los RTK conducen la señal con ayuda de interacciones específicasproteína-proteína
• Dominios específicos
• No hay patrón en cuanto a número o localización dentro de la proteína
• No están presentes en procariotas
27

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 28/29
MECANISMOS DE ACCIÓN DE HORMONAS CON RECEPTORES NUCLEARES
Las hormonas esteroideas (estrógeno, cortisol y progesterona..), demasiadohidrófobicas para disolverse fácilmente la sangre, se transportan por proteínas
específicas transportadoras hasta un punto de liberación hasta sus tejidos diana. EN lascélulas diana estas hormonas pasan la membrana plásmatica por fifusión simple uy seunen a proteínas receptoras especificas en el núcleo. Los receptores sin ligandonormalmente (aporreceptores a menúdo actuan suprimiendo la trasncripción de genesdiana. La unión de la hormona desencadena cambios de conformación de una proteínareceptora y de esta forma es capaz de interactuar con secuencias reguladoras de ADNdenomidanos elemetos de respuesto de ehormonal (HRE), alterando así la expresióngénica.
28

5/10/2018 T.7 Intro Al Metabolismo - slidepdf.com
http://slidepdf.com/reader/full/t7-intro-al-metabolismo 29/29
DOMINIO DE UNIÓN AL DNA CONSERVADO EN LOS RECEPTORES NUCLEARES
29