
Proyecto de aula de biologia
77
UNIVERSIDAD TÉCNICA DE MACHALA Calidad, Pertinencia y Calidez VICERRECTORADO ACADÉMICO CURSO DE NIVELACIÓN DE CARRERA ÁREA: SALUD CURSO DE NIVELACIÓN DE CARRERA SEGUNDO SEMESTRE 2013 ASIGNATURA: BIOLOGÍA TUTOR: BIOQ. CARLOS GARCIA MSC. PROYECTO DE AULA TEMA: PROYECTO DE ELABORACIÓN DE UN MANUAL PARA SEMBRAR UNA PLANTA DENTRO DE OTRA PLANTA MADREEN EL ÁREA DE LA SALUD DEL CURSO DE NIVELACIÓN DE UTMACH DE ENERO A FEBRERO DEL 2014. ESTUDIANTE: ROCIO DEL PILAR AÑAZCO VÉLEZ. 2013– 2014
-
Upload
rocio-anazco -
Category
Documents
-
view
325 -
download
1
Transcript of Proyecto de aula de biologia

UNIVERSIDAD TÉCNICA DE MACHALA Calidad, Pertinencia y Calidez
VICERRECTORADO ACADÉMICO CURSO DE NIVELACIÓN DE CARRERA
ÁREA: SALUD
CURSO DE NIVELACIÓN DE CARRERA
SEGUNDO SEMESTRE 2013
ASIGNATURA: BIOLOGÍA
TUTOR: BIOQ. CARLOS GARCIA MSC.
PROYECTO DE AULA
TEMA:
PROYECTO DE ELABORACIÓN DE UN MANUAL PARA
SEMBRAR UNA PLANTA DENTRO DE OTRA PLANTA
MADREEN EL ÁREA DE LA SALUD DEL CURSO DE
NIVELACIÓN DE UTMACH DE ENERO A FEBRERO DEL
2014.
ESTUDIANTE:
ROCIO DEL PILAR AÑAZCO VÉLEZ.
2013– 2014

DEDICATORIA
El presente trabajo realizado me permito dedicar de
todo corazón primeramente a Dios por que Él está
siempre en mi mente y corazón, a mis padres, a mis
hermanos y hermana política, al Bioq. Carlos García
Docente del módulo de Biología, dedico también a mis
mejores amigas; y, finalmente a mis compañeras de
curso.
RocioAñazco

AGRADECIMIENTO
De todo corazón me permito expresar mis sinceros
agradecimientos primeramente a Dios por permitirme
seguir adelante en este curso de nivelación y admisión,
y por la perseverancia que puso en mi corazón en la
realización de este trabajo, a mis padres por su apoyo
económico y moral, a mi profesor Bioq. Carlos
García,quién nos impartió sus conocimientos en éste
módulo interesante e importante como es Biología.
RocioAñazco

DATOS PERSONALES
DATOS PERSONALES:
NOMBRES: ROCIO DEL PILAR
APELLIDOS: AÑAZCO VÉLEZ
FECHA DE NACIMIENTO: 22 / DICIEMBRE / 1995
TIPO DE SANGRE: 0+ POSITIVO
ESTADO CIVIL:SOLTERA
NACIONALIDAD: ECUATORIANA
EDAD: 18 AÑOS
DIRECCION DOMICILIARIA: STA. ROSA (CDLA. JOSE MARIA OLLAGUE)
TELEFONO: 2 – 945 - 527
EMAIL:[email protected]
GMAIL: [email protected]
DATOS ACADÉMICOS:
PRIMARIOS: C.E.P.W.O.L. “ALTAMIRA”
SECUNDARIOS: COL. NACIONAL “ZOILA UGARTE DE LANDÍVAR”
TITULO: BACHILLER EN CIENCIAS (FÍSICO - MATEMÁTICO)

PRESENTACIÓN
AUTORETRATO
Mi nombre es Rocio del Pilar Añazco Vélez, nací en el cantón Santa Rosa
Provincia de El Oro, el 22 de Diciembre de 1995, soy hija de padres legalmente
casados; IltoAñazco Córdova y Odalia Vélez Murillo, soy la última de 5
hermanos,la mayor parte de mi infancia la pasé con mi madre y mis hermanos, ya
que mi padre se encontraba lejos cumpliendo con sus servicios a la ciudadanía, ya
que es parte de la Policía Nacional, mis estudios de Educ. Básica los inicié en el
―Complejo Educativo Particular Dr. Wenceslao Ollague Loayza‖ (actual
C.E.P.W.O.L. ALTAMIRA) hasta 7mo año de Educ. Básica, desde 8vo año de
Educ. Básica hasta 3er Año de bachillerato estudié en la ―Unidad Educativa ―Zoila
Ugarte de Landívar‖ de allí salí graduada de ―Físico-Matemático‖.
Soy la mimada de mis padres al igual que de mi hermana mayor, ellos son los que
más me han inculcado a los valores éticos y morales, tengo errores como toda
persona, pero de mis virtudes las que más me gustan y práctico son: la
responsabilidad, ser amistosa y ayudar en lo que pueda a los que me necesiten,

aunque algunas veces soy rencorosa, pero cuando me hacen algo y veo que me
afecto mucho soy vengativa, mi lema es ―no me creo más ni menos que otro y
demuestro siempre ser cada vez una mejor persona‖.
Para llegar a obtener este cupo e ingresar a la Universidad no se me hizo fácil,
pase y estoy pasando por muchas situaciones en con mi familia y con un gran
esfuerzo y sacrificio estoy aquí…

RESUMEN
El injerto es una técnica conocida desde la Antigüedad. Está documentado en
China desde comienzos del I milenio a. C., y en Occidente ya la Grecia clásica lo
conocía. Aristóteles describe con bastante detalle las técnicas empleadas en su
época, y los escritores agrícolas romanos las documentan así mismo. Aunque el
interés en la práctica continuó y se estimuló en el Renacimiento, no sería hasta el
siglo XVII en que Henri Louis Duhamel estudiase la función de los tejidos en el
proceso de injerto, investigaciones continuadas por HermannVochting que
sentaron las bases de los conocimientos modernos sobre injerto. A partir de los
años 1920 se cuenta con descripciones científicas del injerto en púa, y a partir de
los '50 se popularizó en cucurbitáceas y solanáceas. La extensión del cambium en
éstas es la que parece favorecer su aplicación.
La finalidad principal del injerto es el método de prevención de
enfermedades,evitando el contactó de una planta sensible con el suelo,agua o
restos de cultivo anterior infestados El injerto sobre una planta resiste a la
enfermedad que se desea prevenir, el portainjertos resistente permanece sano y
asegura,a partir del suelo una alimentación normal de la planta, a la que aísla del
parásito.

INTRODUCCIÓN
El injerto es una técnica de reproducción asexual que permite la formación de una
nueva planta mediante el trasplante de una yema o púa de la planta madre
seleccionada a un patrón.
Con esta técnica de multiplicación se unen porciones distintas de tal manera que
hay soldadura y paso de savia, constituyéndose un único individuo capaz de
crecer y desarrollarse.
Es muy importante conocer y desarrollar esta técnica de propagación. En
jardinería y agricultura, los injertos son considerados sumamente importantes
porque de ésta manera pueden multiplicarse y conservarse, sobre patrones de
buena vitalidad, las razas que no dan semillas como en los frutales, la vid, los
rosales, etc.

INTRODUCCIÓN
ANTECEDENTES DE LA PROBLEMÁTICA
Conceptualización de la problemática en relación con su historia.
El injerto es una técnica conocida desde la Antigüedad. Está documentado en
China desde comienzos del I milenio a. C., y en Occidente ya la Grecia clásica lo
conocía. Aristóteles describe con bastante detalle las técnicas empleadas en su
época, y los escritores agrícolas romanos las documentan asimismo. Aunque el
interés en la práctica continuó y se estimuló en el Renacimiento, no sería hasta el
siglo XVII en que Henri Louis Duhamel estudiase la función de los tejidos en el
proceso de injerto, investigaciones continuadas por HermannVochting que
sentaron las bases de los conocimientos modernos sobre injerto. A partir de los
años 1920 se cuenta con descripciones científicas del injerto en púa, y a partir de
los '50 se popularizó en cucurbitáceas y solanáceas. La extensión del cambium en
éstas es la que parece favorecer su aplicación.
Importancia social sobre la problemática.
Cuando se realiza un injerto se obtienen beneficios con algunos patrones ya que
transfieren a esta unión caracteres de rusticidad o resistencia frente a algún factor
desfavorable. Se perpetúan clones de frutales que únicamente son producibles
bajo este método. Se puede acelerar la madurez reproductiva de las plantas para
lograr rápida fructificación. Cambiar cultivares de plantas antiguas por variedades
nuevas que se estén apreciando. Obtener formas deseadas de crecimiento como
el achaparramiento en cítricos. Repara partes dañadas de árboles. Sirve de fuente
de estudio de enfermedades porque sirve de detector de la presencia de algún
agente patógeno.
Referente nacional y local sobre la problemática y su relación con el Plan de Buen Vivir.

El Buen Vivir se funda en una relación armónicaentre el hombre y la naturaleza; ya
no podemos centrar la vida solo en el hombre como lo hacía lavisión
antropocéntrica del pasado, hoy es imperativo entender que la vida del hombre no
seguirá siendo posible si la separamos del destino de la naturaleza, de sus
recursos vitales, del agua, de las tierrasfértiles, de los frutos que tan
generosamente nos permiten reproducir la vida humana y la vida animal; por eso
es importante inculcar a que las personas realicemos algún tipo de injerto ya que
eso nos ayudará a crear nuevas especies y da la oportunidad de que siga
teniendo vida.
Si este foro tiene como gran cobertor conceptual y práctico el nuevo humanismo,
nosotros creemos queel nuevo humanismo ha de entenderse en esta relación
armónica y respetuosa: el hombre depende nosolo del hombre, sino de su
entorno, de la materialidad natural que por años ha sido explotada más porlas
ambiciones de un sistema económico que prioriza las industrias agrediendo sin
cautelas al ambiente,que por un sistema que respeta el entorno y desarrolla una
ética sobre los alcances de una economía quedebe tomar solo que se necesita
para vivir, sin una permanente expoliación de los recursos naturales yla
―inundación‖ de productos industriales muchas de las veces innecesarios para la
vida humana.
El nuevo humanismo no es propiedad de ninguna cultura nipuede ser cristalizado
en tiempo histórico alguno. El humanismo está presente en todas las culturasbajo
distintas denominaciones y formas, y fluye hacia renovados significados del mismo
modo comotransita la especie humana, de una manera dinámica desde la
determinación hacia la libertad‖.
SITUACION PROBLEMICA
Mediante las encuesta realizadas al curso de nivelación en el paralelo V01 de la
UTMACH en el área salud el 80% respondió negativamente, podemos observar
que hay una gran demanda de personas que no conocen lo que es un injerto, ni

para qué sirve y por qué es necesario, lo cual me ha permitido realizar con ayuda
del TUTOR BIOQ.CARLOS GARCÍA un manual sobre uninjerto; sembrando una
planta de orquídea en una palmera.
1. Enunciación del problema
Proyecto de elaboración de un manual para sembrar una planta dentro de otra
planta madreen el área de la salud del curso de nivelación de utmach de enero a
febrero del 2014.
2. Justificación
La presente investigación la realizaré para analizar las condiciones de vida de
injerto, ya que es un proceso hortícola donde se inserta el tejido de una planta en
los tejidos de otra para formar una unión simbiótica.
Este proyecto de investigación tiene el propósito de realizar un manual de esta
situación cultural conla fijación de una orquídea (orchideceae)Phalaenopsis,
también llamado orquídeas mariposa porque se parecen a una polilla en vuelo, a
una palmera (Arecaceae, sinónimoPalmae), el injerto es más un proceso de unión
para que la orquídea pueda establecer su sistema de raíces y vivir en conjunción
con el árbol.
En la realización de este manual determinaré una propuesta de manera especial
hacia todos los colectivos y personas sensibilizados en temas de conocimiento y
conservación del medio natural, desde colectivos conservacionistas a grupos de
investigación o personas individuales comprometidas con el conocimiento de su
entorno más cercano.
3. Problematización

3.1 Problema Central
Proyecto de elaboración de un manual para sembrar una planta dentro de otra
planta madre.
3.2 Problema Complementario
El injerto es un método de multiplicación que admite aprovechar las buenas
características que aportan los patrones, permite tener en una misma planta de
flores de distintas variedades o varios tipos de frutas, provee una producción
temprana, tiene efecto enanizante, abastece un sistema de raíces adecuado,
daresultados beneficiosos del injerto, logra salvar la vida de una planta, mejora la
adaptabilidad del suelo, cambio de una copa no deseada, propaga plantas que no
puede propagarse ni por esquejes ni por acodos, tiene una buena resistencia a
plagas y enfermedades.
Los injertos en plantas monocotiledóneas son difíciles por la forma en que están
arreglado los haces vasculares, en algunos casos hay incompatibilidad de
especies, pero hay resultados beneficiosos del injerto
4.1. Objetivos generales
Obtener un injerto que conserve los caracteres de aquella que se desea y Conocer
su forma de aplicación, ventajas, desventajas y como actúa un injerto.
4.2. Objetivos específicos
Suministrar una idea de las diferentes relaciones y procesos que ocurren en los
tejidos meristemáticos de la planta en el proceso de desarrollo de un injerto.
Determinar la importancia que tienen los injertos, como una herramienta de
propagación de plantas y una forma de mejorar la calidad y eficiencia de
algunos cultivares.

Utilizar la información adquirida y llevar a práctica los métodos aprendidos en
el desempeño de nuestra carrera profesional
PLANTEAMINETO DE CONJETURA E HIPOTESIS
Proyecto de elaboración de un manual para sembrar una planta dentro de otra
planta madreen el área de la salud del curso de nivelación de UTMACH de enero
a febrero del 2014.
5. Variables y características
Variable Dependiente: Injerto (Orquídea)
Variable Independiente:Palmera (Aracaceae-Palmae)
Marco Referencial
Marco Teórico
EL INJERTO
El injerto viene a ser el proceso por el cual dos porciones de tejido vegetal viviente son
unidos entre sí, con la finalidad que luego ambos se desarrollen como si se tratara de una
sola planta. El injerto es considerado una práctica de propagación asexual que permite el
desarrollo de una variedad vegetal interrelacionada con otra y que cuyo producto de
dicha interrelación brinde un producto óptimo y deseado.
Patrón:
El patrón viene a ser la planta que alberga la unión con la púa y cuyos procesos
metabólicos le sirve a la púa que actúa solo como una parte de la planta. Es así que el
patrón es el responsable de la nutrición del injerto por cual la nutrición mineral de este es
de suma importancia para la obtención de buenos injertos.
Clases de patrones:

Los patrones pueden dividirse en dos grupos:
Patrones de plántula: Son aquellos que se desarrollan a partir de la semilla es decir son
cultivados en almácigos pasan por los procesos de trasplante y traslado a plántulas. Estos
patrones cuentan con ciertas ventajas debido a que su propagación es relativamente fácil
y económica además de poder adaptarse fácilmente a los métodos de propagación en
masa.
Además por tratarse de una reproducción por semilla se elimina la transmisión de
enfermedades causadas por virus, salvo rarísimas excepciones. En algunas especies
además se puede contar con un mejor enraizamiento en comparación con los patrones
obtenidos a partir de estacas.
La desventaja del uso de estos patrones es la presencia de variación genética, que se
muestra en una variación en el crecimiento y comportamiento de la púa que se injerte ya
que en dicho proceso se puede tener una planta heterocigota que puede responder
diferente que una planta genéticamente pura.
Patrones clonales: Muy utilizados en países europeos, incluye dentro de este grupo a los
patrones obtenidos por estacas enraizadas y acodos de banquillo. La ventaja de este tipo
de patrones es que por ser un tipo de propagación asexual se tiene la seguridad de que
todos los plantones sean genéticamente iguales y se puede esperar que en un ambiente
dado tengan características idénticas de desarrollo no solo de la misma planta sino
también de la púa con la que va a ser unido. Obviamente los plantones a clonar tienen
características óptimas como resistencia a enfermedades o a cualquier factor
desfavorable y se conoce su desempeño cuando se le someta al proceso de injerto.
El problema en la utilización de estos patrones es el cuidado que se debe de tener en la
propagación de enfermedades bajo este sentido se tiene que considerar el uso de
patrones y púas libres de organismos patógenos.
Injertos dobles:
Es la combinación posible que se puede realizar entre dos clones de plantas en un arreglo
o disposición vertical. Esto supone la inserción de un clon entre patrón y el injerto. Esta

inserción permitirá el desarrollo de una planta con tres tipos de especies vegetales, a esta
porción entre injerto y patrón se le conoce como patrón intermedio.
El uso de este patrón intermedio es importante para la solución de ciertos problemas de
incompatibilidad, es decir, el patrón intermedio sirve de conector entre especies vegetales
que normalmente son incompatibles.
Además el uso de este patrón intermedio también es asociado con ciertas características
deseadas con las que pueda aportar y que se encuentren ausentes tanto en la púa como
en el patrón. También se ha comprobado que el patrón intermedio influye sobre el
crecimiento de los árboles aportando también sus características.
Motivos por el cual se Injerta:
A) Obtención de Beneficios con algunos patrones:
Algunas especies presentan por su misma naturaleza un carácter de rusticidad frente a
condiciones adversas ya sea del tipo sustrato, del tipo climático o efectos producidos por
alguna plaga o enfermedad. Dicha rusticidad o resistencia a estas condiciones no la
comparten otras especies vegetales que son útiles para el hombre y gracias al injerto que
une estas características de los dos tipos de plantas, logra que estas especies
susceptibles pueden gozar de los beneficios de las otras especies resistentes creando así
una especie vegetal útil y a la vez resistente.
B) Perpetuación de Clones únicamente producidos a partir de injertos:
Un gran número de especies utilizables por el hombre tales como frutales y otras plantas
leñosas no se pueden reproducir por métodos asexuales como estacas o acodos por lo
que necesitan de otra especie que sí lo pueda lograr de manera efectiva. Además las
plantas obtenidas son heterocigotas por lo que su propagación por semillas no podría
mantener el clon deseado por lo que también se tendría que recurrir nuevamente al
injerto.
C) Acelerar la madurez reproductiva:
Las especies como frutales y otras necesitan de un tiempo o periodo vegetativo muy largo
para empezar el proceso de fructificación, por lo que el uso del injerto se haría para
disminuir el tiempo de espera para la obtención de los productos cosechables; esto se

logra injertando sobre plantas o patrones ya establecidos, es decir, sobre plantas ya
maduras.
D) Cambiar los cultivares de algunas plantas:
Un árbol puede producir un cultivar que por diversas razones ya no brinde los beneficios
que nosotros deseamos, por lo que desearíamos poder cambiar por otro tipo de cultivar,
el injerto hace posible este cambio.
Además puede resultar de interés para un agricultor el tener un solo árbol con varios
cultivares diferentes siempre y cuando sea cada una de estas variedades compatibles con
el patrón.
E) Obtener formas especiales de crecimiento:
El injerto también permite cambiar las formas de crecimiento no deseado de una especie
vegetal por otra que nos brinde mayores beneficios
F) Reparar partes dañadas de los árboles:
Por algún agente extraño una planta puede ser dañada y el injerto puede asegurar la
rápida recuperación salvando la planta.
G) Sirve de fuente de estudio de enfermedades:
Las enfermedades son transmisibles por medio del injerto, entonces al realizar esta
práctica se puede detectar la presencia de dichos virus en plantas que no mostraban
síntomas de enfermedad.
Procesos que ocurren en la unión de la púa con el Patrón:
1- Establecimiento de un contacto íntimo de una extensión considerable de región
cambial del patrón y de la púa en condiciones ambientales favorables: La temperatura
óptima para permitir la actividad celular abarca entre los 12,8 a 32 0C dependiendo
de la especie. Estas condiciones mayormente las encontramos en primavera, para el
caso de invernaderos las temperaturas son controladas.
Luego se forma un nuevo tejido de callo originado en la región cambial formado por
células de pared delgada turgentes que necesitan las condiciones de humedad
necesaria para seguir viviendo.

2- Producción y entrelazamiento de células de parénquima por el patrón y la púa: Este
mecanismo es utilizado por la planta debido a que las células vivas de los dos
componentes del injerto han quedado semi aislados por una capa de células muertas
y suberizadas. Las células vivas son capaces de producir una unión debido a su
rápida proliferación de dictiosomas que segregan sustancias entre los espacios de las
paredes celulares que sirven de guía de migración de vesículas al plasmalema,
produciendo una rápida adhesión entre las células parenquimáticas.
3- Producción de nuevo cámbium en el puente del callo: En un tiempo de 2 a 3 semanas
en los bordes del callo las células parenquimáticas que tocan las células cambiales
del patrón y de la púa se diferencian en unas nuevas células de cámbium. Este
cámbium avanza por el cámbium original del patrón y de la púa y a través del puente
de callo, hasta que se forme una conexión cambial continúa entre el patrón y la púa.
4- Formación de nuevo xilema y floema: Se produce a partir del nuevo cambium vascular
producido en el puente del callo el cual comienza su actividad cambial normal de
acumular xilema hacia el interior y floema hacia el exterior.
6.1.2 Proceso de cicatrización en el injerto de yema en T
En este método de injertar se tiene que la yema extraída por ejemplo de la Rosa contiene
epidermis, capa de corcho, floema, cambium y con frecuencia tejido del xilema que
dificulta mucho en el proceso de cicatrización.
Alrededor de los tres días de injertar las yemas terminales de los radios del xilema rotos y
las derivadas cambiales adyacentes de la superficie expuesta del patrón empieza a crecer
y dividirse, conduciendo a la producción de filamentos de callo. Del mismo modo
originándose en las células terminales de radios de floema rotos y de células jóvenes
adyacentes del floema secundario de la superficie cortada del escudete se forman
filamentos de callo.
En 14 días el espacio entre el patrón y el injerto se llena por completo de callo, el cual se
ha desarrollado principalmente de la proliferación del xilema secundario inmaduro del
patrón y del floema secundario también inmaduro del injerto.

Durante la segunda semana en este tejido de callo de nuevo desarrollo aparecen
pequeñas áreas de células cambiales. Hacia el décimo día se extiende sobre la cara del
patrón una faja completa de tejido de cámbium y se une con el cámbium no lesionado de
ambos lados del escudete.
Una vez que se completa la continuidad cambial, pronto se establece la conexión de tejido
vascular entre la yema y el patrón. En el injerto de yema la unión primaria se establece
entre la superficie del floema de la cara interna del escudete y la superficie meristemática
del xilema del patrón. Sin embargo, puede ocurrir un tipo secundario de unión en los
bordes del escudete.
Factores que influyen en la cicatrización de un injerto:
Clase de planta: Algunas plantas son mucho más dificiles de injertar que otras aunque nio
intervenga en ello la incompatibilidad. Algunas plantas que se injertan con facilidad forman
una goma de herida que tapa los elementos de xilema expuestos después de la operación
de injerto evitando con ello la desecación excesiva y la muerte de tejido.
Condiciones de temperatura humedad y oxígeno durante y después del injerto: Los
rangos óptimos para el normal desarrollo de un injerto depende también del tipo de planta
sin embargo existen rangos de temperaturas máximas y mínimas para que se produzca la
formación de callo, estos rangos oscilan entre 0 y 40 0C para la vid la temperatura optima
esta entre 24 a 27 0C por eso se recomienda la práctica de injertos en épocas de
primavera donde la temperatura se encuentra entre los rangos establecidos.
Actividad del crecimiento del patrón: El éxito de algunas tipos de injerto dependen mucho
de que las células del cámbium estén en división activa. La iniciación de la actividad
cambial se da en la primavera que es cuando las yemas empiezan a crecer. Este estímulo
se debe a la producción de auxinas y giberelinas que se originan en las yemas en
expansión.
Técnicas de propagación: Lastécnicas o tipos de injertos tienen que ser los adecuados de
acuerdo a la especie vegetal y depende mucho de la forma y calidad del injerto el éxito
de dicha práctica.

Contaminación con algún agente patógeno: Es importante el aislamiento de las especies
con las que se va a trabajar ya que con el injerto se pueden transmitir numerosas
enfermedades, además en el caso que se tratara de plantas sanas cabe la posibilidad de
contagio por virus existentes en el medio ambiente ya que el injerto deja una herida que
puede ser utilizada como vía de ingreso a cualquier patógeno.
Relación de sustancias de crecimiento: Se ha comprobado que el uso de sustancias
reguladoras del crecimiento como la auxina para la cicatrización no brinda resultados
esperados, sin embargo, la auxina y la Kineetina son muy favorables para la formación de
callo, además existen otras sustancias promotoras del desarrollo de callo como es el caso
de ácido absícico sobre todo en combinación con los anteriores reguladores de
crecimiento.
Relaciones entre el Injerto y el Patrón:
La combinación de dos plantas con genotipos diferentes produce a veces patrones de
crecimiento que difieren de aquellos que hubieran tenido las partes componentes
cultivadas por separado. Las características alteradas pueden resultar de reacciones de
incompatibilidad o del hecho de que una de las partes del injerto tenga uno o más
caracteres específicos que no se encuentran en la otra como ciertas resistencias o debido
a interacciones específicas entre el patrón y la púa que alteren ciertas características
como tamaño, desarrollo, productividad, calidad de fruto u otro atributo.
6.1.3 Efectos del Patrón sobre la púa:
El más saltante de ellos es el control del tamaño o a un cambio en la forma del árbol. Esto
se produce debido al cambio de vigor que se logra de la unión. Estas relaciones ayudan al
agricultor a la obtención de variedades enanas que le permitan facilidades en el manejo
por ejemplo de cítricos.
Para el caso de fructificación el patrón puede influir en la precocidad de la fructificación,
cuajado de frutos y formación de yemas y en general en el rendimiento de un árbol, estas
características vienen asociadas a los patrones que producen achaparramiento ya que
esto permite plantas más pequeñas y fructíferas que al ocupar menor espacio producen
mayor cantidad de rendimientos por hectárea. La influencia del patrón puede variar mucho

en las diferentes clases de plantas hay especies que al unirse permiten un aumento en la
producción debido a la mayor formación de flores y frutos, sin embargo, en otras especies
la mezcla puede influir de manera negativa para la producción disminuyendo
enormemente la fructificación.
En algunas especies vegetales como la vid el rendimiento depende mucho del vigor del
crecimiento del año en que se cosecha la utilización de patrones resistentes o vigorosos
es de suma importancia en estos casos.
Es importante recalcar que en la unión de patrones hay características que no se mezclan
y que no son apreciables en los frutos, es decir, los frutos del injerto no adquieren las
características de los frutos de los patrones, sin embargo ciertos patrones pueden afectar
la calidad de la variedad que se injerte sobre ellos un ejemplo son los cítricos, donde los
patrones producen efectos muy notables en las características de los frutos.
Otro efecto del patrón sobre la púa lo podemos encontrar en la resistencia que ofrecen
algunos patrones a efectos climáticos adversos, hay que considerar también que ciertos
comportamientos de los patrones son respuesta a diferentes tipos de fertilización del
suelo. Además hay casos de sobre fertilización donde las cantidades de algún
micronutriente se convierta en tóxico para la planta, pueda ser controlado por la
resistencia de algunos patrones a ciertas sustancias tóxicas.
Efectos de la cultivar sobre el patrón:
Se produce también la influencia de la púa sobre el patrón para el caso de
achaparramiento, es decir, el efecto de achaparramiento también puede ser influenciado
por la púa.
Además para el caso de vigor se tiene que especies de cultivares resistentes a ciertas
condiciones adversas al servir como púas también producen cierta resistencia y vigor al
patrón que las está soportando, un ejemplo claro lo tenemos en cítricos donde púas de
especies más vigorosas actúan sobre patrones menos vigorosos es la púa la que empieza
a controlar o a influenciar sobre la rapidez de crecimiento y tamaño final del árbol.
También esto se observa en el manzano.

La resistencia a condiciones climáticas adversas también son modificadas por ciertos
tipos de cultivares sobre patrones menos resistentes no necesariamente por el hecho
simple de que el cultivar imparta esa resistencia al patrón sino porque ciertas especies
tienden a prolongar el crecimiento de las raíces del patrón por bastante tiempo durante el
otoño de modo que los tejidos de la raíz no han madurado lo suficiente para cuando
ocurre las heladas fuertes de invierno.
Los efectos de algunos patrones intermedios también son de gran utilidad en la actividad
agrícola, uno de los efectos más importantes es el producido en los manzanos donde el
achaparramiento empleando un patrón intermedio achaparrante tiene la ventaja de
permitir el empleo como patrón de una plántula bien anclada, vigorosa en lugar de un clon
achaparrante quebradizo y mal anclado. Sin embargo el patrón intermedio puede
influenciar en una producción excesiva de brotes en las raíces. Existen datos que los
efectos observados del patrón intermedio se debe directamente a la influencia de una
porción del patrón intermedio más bien que a anormalidades en la unión de injerto.
Mecanismos de la influencia Recíproca entre patrón e injerto:
Estos mecanismos de relaciones recíprocas son procesos muy complejos y a veces son
atribuibles a otras causas por esto dichas explicaciones propuestas de los efectos
observados son especulativas por eso se consideran como posibles.
Uno de estos posibles mecanismos es la influencia del patrón como resultado de efectos
de translocación más bien que de la capacidad de absorción del sistema radicular. El
hecho de que cuando menos en cierto grado, la porción de tallo de un árbol tiene una
influencia definida se ha demostrado en experimentos donde patrones se han usado
como patrones intermedios entre un sistema radicular vigoroso y un cultivar que sirvió de
injerto. Los efectos esperados se manifestaron aunque en menor grado, no obstante que
los materiales se emplearon como patrón intermedio.
6.1.4 Incompatibilidad del injerto
Viene a ser un proceso por el cual la unión de las dos plantas no se produce en forma
exitosa y por ende tampoco el desarrollo satisfactorio como una sola planta. La diferencia

de una incompatibilidad es sumamente notoria ya que simplemente una unión
incompatible se trasluce en un crecimiento anormal.
Tipos de incompatibilidad:
La incompatibilidad de injerto es principalmente de dos tipos: Incompatibilidad localizada
e incompatibilidad translocada.
Incompatibilidad localizada: En la combinación incompatible generalmente la estructura de
la unión es mecánicamente débil presentando interrumpida la continuidad de los tejidos
vasculares y el cambium. Con frecuencia los síntomas se presentan en forma lenta
proporcionalmente con el grado de disturbio fisiológico presente en la unión del injerto,
provocando una defectuosa translocación entre injerto y el patrón produce un agotamiento
de las raíces.
Este tipo de incompatibilidad es notorio debido a la formación de tejido parenquimático en
la zona de unión envés de encontrar un tipo de tejido diferenciado. A veces los vasos
conductores establecen conexiones alrededor de masas de tejidos de callo, mientras que
otras, los extremos de los vasos están separados por masas de parénquima. En casos
extremos puede llegarse hasta un tipo de deformación del tejido vascular. El cambium de
ambas partes ha establecido límites y no ha permitido la traslocación entre ambos
elementos.
Un ejemplo de incompatibilidad localizada es la de un tipo de Peral Bartlett sobre un
membrillero sin embargo esta unión se puede lograr con el uso de un patrón intermedio y
la combinación de tres partes es completamente compatible.
Incompatibilidad translocada: Incluye los casos donde la incompatibilidad no se puede
solucionar con el uso de un patrón intermedio. Este tipo de incompatibilidad implica una
degeneración en el floema y se puede reconocer por el desarrollo en la corteza de una
línea de color pardo o una zona necrótica. En este tipo de incompatibilidad no se produce
los movimientos de carbohidratos. En diversa modalidades que se presentan en esta
categoría, la extensión de la descomposición del tejido cortical puede abarcar desde
prácticamente la falta de formación de unión mecánica débil con tejidos deformados hasta
una unión fuerte con tejidos conectados normalmente. Un ejemplo de esta categoría la

ofrece el durazno de la variedad Hale Early injertado en patrón de ciruelo Myrobalan que
forma una unión débil con tejidos deformados y acumula cantidades anormales de
carbohidratos en la base de la púa del durazno. Si se utiliza un patrón intermedio entre el
durazno y el ciruelo los síntomas de incompatibilidad todavía permanecen.
Otro ejemplo lo tiene el durazno injertado en patrón Marianna el cual forma una unión
incompatible, aunque los escudetes de durazno se unen con facilidad a este patrón de
ciruelo y crecen de manera satisfactoria durante la primera estación, después aparece un
engrosamiento arriba de la unión de injerto seguido por el marchitamiento de las hojas de
durazno y la muerte del árbol.
Los casos de incompatibilidad también se desarrollan en un tiempo determinado, es decir,
hay casos en que una unión compatible puede desempeñarse en forma normal duran te
20 o 30 años y luego para que se sigan obteniendo buenos resultados se necesita del uso
de un patrón intermedio que devuelva la compatibilidad a esa unión.
Causas de la incompatibilidad
La incompatibilidad está relacionada principalmente con las diferencias genéticas entre el
patrón y la púa que contiene materiales vegetales genéticamente diferentes que a su vez
producen o son responsables de un sistema fisiológico, anatómico y bioquímico
totalmente diferentes y con muchas interacciones posibles.
Un ejemplo es la diferencia morfológica y fisiológica entre ciertas variedades de peral
sobre membrilleros en la cual se nota que en la translocación de compuestos como
glucósido cianogénico propio del membrillero al pasar al peral este lo descompone en la
zona de unión produciendo el ácido cianhídrico como producto de dicha descomposición
no permitiendo así relaciones entre el membrillero y el peral, además este ácido produce
una falta de actividad cambial causando marcados desarreglos anatómicos dando paso a
una destrucción de tejidos del xilema y del floema los cuales son vitales para la planta por
ser los transportadores de agua y materiales este deterioro afecta al membrillero ya que
dicho problema llega a las raíces donde las grandes cantidades de ácido cianhídrico
matan grandes áreas del floema.

En las variedades de peral que son compatibles con el membrillero existe un inhibidor
soluble de agua que le permite al membrillero descomponer el ácido glicósido en forma
óptima no presentándose los problemas anteriormente mencionados.
Consideraciones del Injerto:
Para obtener resultados exitosos en la práctica de los injertos es necesario considerar
condiciones que tienen que ver mucho con el aspecto fisiológico de las plantas que
intervienen en esta unión.
Una de ellas, la más importante es que la púa y el patrón deben ser compatibles es decir
tengan la capacidad de unirse en la práctica se pueden utilizar plantas estrechamente
emparentadas y no plantas con relación distante por la diferencia en caracteres
morfológicos y fisiológicos de cada especie vegetal.
Otra consideración es la apropiada región para la colocación del injerto además del intimo
contacto que tienen que tener la púa con el patrón esto se logra con métodos aplicativos o
técnicas para injertar, además estos métodos deben asegurar una rápida cicatrización
para que la púa pueda ser provista de agua y nutrientes por el patrón para cuando
empiecen a abrirse las yemas.
El tiempo en que se realiza la operación es también un factor importante que determina el
éxito de esta práctica, se tiene que considerar el estado o edad fisiológica adecuada tanto
de la púa como del patrón para realizar la unión y que los procesos tales como la
producción de callo puedan efectuarse.
Dicha práctica debe de protegerse de efectos externos adversos para la unión, tales como
la desecación de la herida o el ingreso de algún patógeno por ser esta una ventana para
el ingreso de dichos elementos, se debe cubrir o proteger las heridas con algún material y
además se le debe de suministrar de suficiente humedad para que el injerto no se
deseque.
Posteriormente a la realización de dicha práctica se debe de tener ciertos cuidados para
evitar problemas como el brotamiento del patrón cerca de la púa pudiéndola ahogar, o en
ciertos casos crecen del injerto ramas tan rápido que es posible que puedan romper la
unión.

6.1.5 Métodos de injerto
Entre los principales podemos considerar:
Injerto Inglés:
Este método se utiliza en materiales de tamaño pequeño, es ventajoso debido a que el
área de contacto entre púa y patrón es grande, la única consideración a seguir es que el
diámetro de ambos debe ser semejante. Es recomendable que la púa tenga dos o tres
yemas, haciendo el injerto en la zona internodal lisa que quede abajo de la yema inferior.
La unión de las partes de cámbiumdeben de coincidir una con otra para permitir una
correcta formación del callo.
Injerto de empalme:
Tiene mucho parecido con el método de lengüeta, es de fácil ejecución y de mucha
utilidad para injertar plantas que tienen un tallo con mucha médula o en las cuales la
madera no es lo suficiente flexible para permitir un ensamble apretado.
Injerto de hendidura:
Muy utilizado para injertar la copa de los árboles ya sea en el tronco de un árbol pequeño
o en las ramas principales de un árbol más grande. Los beneficios de este método radican
en que se pueden realizar en cualquier época del año. Es importante considerar que los
cámbium tienen que coincidir y tratar en lo posible de la coincidencia de la corteza de la
púa con la del patrón.
Injerto de cuña:
Para su realización se realiza una muesca en V en el patrón y de igual forma se deja una
estructura similar en la púa para que ocupe ese espacio. Al igual que el injerto de
hendidura este método es realizable también en invierno de preferencia a finales o al
inicio de la primavera. Es de fácil ejecución y muy práctico para muchas especies
vegetales
Injerto de corteza:

Es muy usado en los árboles de madera suave, a los cuales es fácil desprenderlos de una
parte de la corteza de igual tamaño para ambos componentes para luego unirlos y
sellarlos con cera o con algún otro aislante. El método de corteza es realizable en árboles
de madera gruesa, si se hace uso de una cuña que permita ajustar apretadamente en la
ranura de la corteza, acomodándose con el corte principal y haciendo que el borde de la
cuña quede debajo de la porción suelta de corteza remanente esto se realiza para no
tener que insertar la púa debajo de la corteza.
6.1.6 Antes de seguir los pasos para sembrar un injerto se deben tomar en
cuenta los siguientes puntos:
-Proceso de unión del injerto:
El injerto completo las dos partes se comportan como una unidad, no solo el flujo
de agua de la planta, sino el envío de señales y coordinación.
El desarrollo del injerto comprende tres procesos: cohesión del patrón;
proliferación del callo y diferenciación vascular.La cohesión se produce como
resultado de la deposición y subsiguiente polimerización de materiales de las
membranas celulares,debida a la herida del injerto.
Cuando se injertan plantas de distintas especies aparecen nuevas
proteínas,mientras que algunas no permiten el intercambio. En las uniones poco
compatibles a 4 semanas el injerto disminuye completamente incompatible.
La fuerza de la unión del injerto evoluciona en tres fases; 1 y 4 esta fuerza
aumenta lentamente.4-8 lo hace rápidamente8-11 a veces vuelve a crecer más
lentamente.El fortalecimiento se retrasa al desviar las partes de su posición
habitual.
-Factores que influyen en la unión:
De las plantas que se unen, algunos factores ambientales facilitan o dificultan el
proceso, los siguientes:

-Temperatura:
Efecto sobre la formación de tejido de callo, el injerto cucurbitáceas una
temperatura de 25 a 28º en la fase de unión. Buenos resultados cuando la
mínimas no bajan de 15 ºC ni máximas exceden los 33 ºC.
-Humedad:
Deben mantenerse en condiciones de humedad elevada,pues en caso
contrario,las probabilidades de una buena unión son reducirlas. Las partes
expuestas a baja humedad se suberifican.Una alta humedad relativa y baja
luminosidad dificultan la marchites.La tasa de supervivientes y calidad del injerto
mejoran con una HR del 80-90 %.Si baja del 70% se ven perjudicadas.Las
instalaciones se colocan las plantas recién injertadas 97% un ligero movimiento
del aire puede ser conveniente.
-Superficie de contacto:
Un contacto eficaz depende del número y disposición de los conductores de las
dos plantas que se injertan.
-Contaminación por patógenos:
En las heridas producidas al injerto,bacterias u hongos que causan la pérdida del
injerto.
-Incompatibilidad:
No hay reglas para predecir el resultado de un injerto aunque, en termino
generales cuantas más afinidad botánica haya entre las plantas más
probabilidades de éxito en el injerto.La incompatibilidad se manifiesta en
-Aparición de miriñaque, un abultamiento de la zona superior al injerto
-Enrollamiento y amarillamiento de las hojas.-Muerte prematura de plantas. La

compatibilidad del injerto está determinada por un sistema de reconocimiento
.Según la disolución de las membranas celulares y subsiguiente contacto de jugos
celulares,hace que las proteínas liberadas se combinen que inicia un proceso que
culminan en un injerto perfecto.Los factores implicados son constituyentes
normales de los tejidos internodales que se han injertado; son componentes de la
paredes celulares y desprenden del contacto entre patrón y variedad.
No pueden ser transferidos de un injerto intermedio, permite el injerto compatible
entre dos especies poco afines.Las pectinas internodales reducen la capacidad de
formar conexiones vasculares en injertos.Algunas proteínas,podrían ser
responsables de la incompatibilidad entre calabazas.
La variedad con mayores niveles de la proteína son las menos afinadas tienen con
el melón, la compatibilidad puede explicarse sin un sistema de reconocimiento
celular la herida del injerto provoca la proliferación del callo.El destino de sus
células depende de sus proximidades, reciben un flujo de auxinas que producen
una diferenciación.La incompatibilidad no se haya formado sustancias que
promueven la formación de la unión, sino que estas sustancias están inhibidas por
la presencia de toxinas, que hacen a las células del callo incapaces de responder.
Los injertos de plantas genéticamente próximas son generalmente compatibles
hay una similitud bioquímicas entre ambas,las sustancias elaboradas por una de
ella no son toxicas para la otra.
-Interacción patrón - Variedad:
Las modificaciones del comportamiento de la variedad por efecto del patrón,
pueden ser producidas por:
-Reacción de incompatibilidad.
-Resistencia a enfermedades.
-Tolerancia a ciertas características del suelo.

-Interacciones específicas entre patrón y variedad, produzcan alteraciones del
desarrollo de la planta,tamaño del fruto, calidad etc.
La resistencia a enfermedades de suelo esta con el conjunto de raíz – hipocotilo al
tallo y hojas de la planta, cuando hay en el suelo un patógeno vascular puede
producir la infección de la planta a través de raíces en sandias injertadas con las
raíces del patrón. Encima del injerto los vasos conductores procedentes de la raíz
se necrosan de los del patrón es suficientes un suministro de agua y nutrientes
para la necesidades de la planta.La raíz del patrón induzca resistencia a la
variedad injertadas, impidiendo la propagación del patógeno.Cuando el patógeno
es el virus en melón o en pepino,tomase te transmite a través de raíces.
La resistencia a Ralstonia está relacionada con la difusión bacteriana en la parte
baja del tallo.Vigor de la planta injertada es intermedio de porta injertos y el de la
variedad, la variedad tiene también influencias sobre el tamaño de la raíz del
patrón.
El suplemento de vigor proporciona el patrón,y permite menos número de plantas
por unidad de superficie, las raíces vigorosas del patrón son capaces de absorber
agua y nutrientes más eficazmente.El flujo de savia procede de la raíz del patrón
contienen altas sustancias orgánicas y fitohormonas.La mayor duración del ciclo
de las plantas injertadas, debe solo a su tolerancia a enfermedades al mejor
suministro de agua y minerales.
El clorofila de las hojas jóvenes es mayor en las plantas injertadas el contenido en
la savia del xilema de iones minerales especialmente nitrato y fosfato y de
citoquinimas es mayor de melón injertadas.Cuando tienen fruto las plantas
injertadas son activas en el transporte de savia que las no injertadas.
Algunos portainjertos tienen capacidad de extraer agua del suelo, es posible que
mediante el injerto sobre patrones reducir el estrés iónico y mejorar la tolerancia.

El melón injertado sobre Cucúrbita es más tolerante al Boro.
Los porta injertos utilizados en tomate y berenjena,melón, sandía y pepino
funcionan estas mismas especies sin injertar,aunque la actividad fotosintética
contenido en clorofila y proteínas solubles disminuye con bajas temperaturas.
El impacto de altas temperaturas es menor en plantas de tomate injertadas, el
tamaño del fruto sobre patrones vigorosos aumenta de manera notable. El injerto
puede influir sobre la forma y color, grosor de la corteza textura y en el contenido
en sólidos solubres.El componente de los azucares en la carne de la sandía,
injertada o no es la sacarosa y en el mesocarpio glucosa y fructosa, es menor que
el de las no injertadas.
El injerto disminuye la actividad de la invertasa en los tejidos del fruto inmaduro y
aumenta la actividad.Tanto el injerto de sandía sobre Cucúrbita como el cuaje de
frutos mediante la aplicación de CPPU, ocasionan un retraso en la acumulación de
azúcar.
Las plantas de tomate injertadas sobre patrones híbridos intereses pacíficos tienen
menor en materia seca de plantas sin injertar. El contenido en sólidos
solubles,azucares totales y ácido ascórbico es ligeramente menor en los de
plantas de tomate injertadas, tienen mejor color y más licopeno.Hay
diferencias,dependiendo del patrón, en los contenidos de N, P, K, Ca y Mg entre
ambos. No hay diferencias entre plantas de melón injertadas o no, en vitamina C,
pH,firmeza y características sensoriales.La raíz puede controlar la inducción floral.
6.1.7 Pasos a seguir para sembrar un injerto
Para realizar un injerto, los tejidos de ambas plantas deben entrar en contacto. En
ningún momento las células se mezclan; los tejidos por encima y debajo del punto
de injerto permanecen por completo diferentes. La unión viene dada por la
formación de un callo parenquimático a raíz del corte. Algunas de las células del
callo se transforman, en condiciones adecuadas, en meristema cambial que puede

volver a producir tejido vascular, estableciendo así la comunicación simplástica
entre ambas partes. La temperatura y humedad del injerto debe mantenerse
controlada para favorecer esta unión.Pasos:
1.- conseguir una planta (orquídea) y una palmera.
2.- Hacer un orificio al costado de la palmera
3.- Colocar abono.
4.- Transplantar la orquídea.
5.- Regarle un poco de agua,
6.-Finalmente observar su crecimiento y cuidar que no se marchite.
6.2 Marco Conceptual
6.2.1 PALMERA
Las arecáceas (nombre científicoArecaceae, sinónimoPalmae), son una familia
de plantas monocotiledóneas, la única familia del ordenArecales (sinónimo
Principales). Normalmente se las conoce como palmeras o palmas. Esta
importante familia es fácil de reconocer. Son plantas leñosas (pero sin crecimiento
secundario del tronco, sólo primario). A pesar de ser monocotiledóneas muchas de

ellas son arborescentes, con grandes hojas en corona al final del tallo,
generalmente pinnadas (pinnatisectas) o palmadas (palmatisectas). Sus flores
poseen 3 sépalos y 3 pétalos, y se disponen en inflorescencias provistas de una o
varias espatas. El fruto es carnoso: una baya o una drupa. Están ampliamente
distribuidas en regiones tropicales a templadas, pero principalmente en regiones
cálidas.
La familia fue reconocida por sistemas de clasificación modernos como el sistema
de clasificación APG III (2009 ) y el APWeb (2001 en adelante ), y
tradicionalmente también fue reconocida en otros sistemas de clasificación debido
a sus caracteres morfológicos distintivos. En estos sistemas de clasificación, se
ubican en su propio orden monotípicoArecales, en la subclase Commelinidae.
Entre las palmeras se encuentran especies de importancia económica y especies
de valor ornamental, además de otras como el cocotero, la palmera datilera, el
palmito, el ratán, la cera de carnaúba, la rafia, entre otras.En el mundo crecen
como especies propias de zonas tropicales, existen concentraciones de ellas en
países como Madagascar. Colombia es el país con mayor número de variedades y
una de ellas es el árbol nacional. Adicionalmente hay varios jardines botánicos
especializados en las palmeras y son a menudo llamados palmetum. Entre
algunas de estas colecciones se pueden citar el Palmetum de Santa Cruz de
Tenerife y el Palmeral de Elche, en España, el Parque Nacional El Palmar en
Entre Ríos, Argentina, y en menor medida, el Jardín Botánico de Caracas, en
Venezuela, y el Jardín Botánico Nacional de La Habana, en Cuba.
Descripción
Árboles o arbustos con troncos sin ramificar o raramente ramificados,
ocasionalmente hierbas largamente rizomatosas, o lianas (Calamusspp.). El sexo
de la planta es variable. El crecimiento secundario está ausente. Ápice del tallo
con un meristema apical grande, las hojas se desarrollan helicoidalmente. Taninos

y polifenoles muchas veces presentes. Pelos variados, y plantas a veces
espinosas debido a segmentos de hojas modificados, fibras expuestas, raíces
puntiagudas, o crecimientos del pecíolo.
Las raíces son micorrícicas, sin pelos.
El tallo es usualmente arborescente con un tronco único sin ramificar (ramificado
dicotómicamente en Hyphaene) o en un grupo cespitoso de tallos erectos, o en un
rizoma erecto ramificado dicotómicamente (Nypa), o en una liana elongada con
largos internodos (ratanes).
Hojas típicas son bastante grandes, alternas y espirales (raramente dísticas o
trísticas), muchas veces agrupadas en una corona terminal (acrocaulis), pero a
veces bien separadas, enteras, envainadoras en la base, con un elongado, erecto
pecíolo (a veces referido como pseudopecíolo) entre la base envainadora y la
lámina. En los taxones arborescentes las bases envainadoras de las hojas
adyacentes pueden solaparse una a la otra, formando un capitel en el ápice del
tronco. Las hojas pueden ser simples, usualmente divididas en forma pinada o
palmada a medida que la hoja se expande, y en la madurez pareciendo
palmadamente lobadas (con segmentos irradiando de un solo punto),
costapalmadamente lobadas (con segmentos más o menos palmados divergiendo
de un eje central corto, o "costa"), pinadamente lobadas o compuestas (con un eje
central bien desarrollado portando segmentos pinados), o raramente dos veces
pinadamente compuestas. A veces bífidas. Con foliolos convertidos en espinas
presentes en algunos taxones. Lámina "plicada", y los segmentos o bien
induplicados (con forma de V en la sección transversal), o bien reduplicados (con
forma de Λ en sección transversal), cada segmento con venas más o menos
paralelas a divergentes. El pecíolo, en los taxones con hojas palmadas, con una
solapa ("hastula"), entre el pecíolo y la lámina. Con tejidos blandos muchas veces
decayendo hasta revelar variados patrones de fibras. Sin estípulas. Hojas

típicamente liguladas, con el apéndice, la lígula, en la unión interna entre la lámina
y el pecíolo. La venación es pinada -o palmado-paralela.
Ejemplares de Livistonachinensis.
Inflorescencias determinadas o indeterminadas, panículas o espigas de flores
solitarias o de unidades cimosas, típicamente axilares o también terminales, con
brácteas de persistentes pequeñas a grandes y deciduas. Las inflorescencias
emergen de debajo (infrafoliares) o entre (interfoliares) o por encima de
(suprafoliares) las hojas. El pedúnculo tiene por debajo un profilo muchas veces
grande con 1 a numerosas espatas.
Flores bisexuales o unisexuales (y entonces plantas monoicas a dioicas), radiales,
usualmente sésiles, con perianto usualmente diferenciado en cáliz y corola,
hipóginas.
Usualmente 3 sépalos, separados a connados, usualmente imbricados.
Usualmente 3 pétalos usualmente, separados a connados, imbricados a valvados.
Estambres 3 o 6 a numerosos, filamentos separados a connados, libres o adnatos
a los pétalos. Estaminodios presentes en algunas especies. Anteras
longitudinales, raramente poricidas en dehiscencia.

Polen usualmente monosulcado.
Carpelos usualmente 3, pero ocasionalmente tantos como 10, a veces pareciendo
que hay uno solo, separados a connados. Ovario súpero, usualmente con
placentación axilar, pero placentación variable.
Estilos, si presentes, separados o connados, estigmas sésiles o en la punta de los
estilos, estigma variado. Óvulos 1 por lóculo, anátropo a ortótropo, bitégmico.
Nectarios en los septos del ovario o sin nectarios.
El fruto es una drupa, usualmente de una sola semilla, muchas veces fibrosa, o
raramente una baya. Raramente dehiscente. Algunos con escamas externas
(Calamoideae), pelos, aguijones, u otras cosas.
Semillas usualmente 1 por fruto y con endosperma con aceites o carbohidratos
(hemicelulosas), a veces ruminado. Almidón ausente.
6.2.2Las Orquídeas
Las orquídeas u orquidáceas (nombre científicoOrchidaceae) son una familia de
plantas monocotiledóneas que se distinguen por la complejidad de sus flores y por
sus interacciones ecológicas con los agentes polinizadores y con los hongos con
los que forman micorrizas.

La familia comprende aproximadamente 25.000 (algunas fuentes informan de
30.000) especies, y quizá otros 60.000 híbridos y variedades producidas por los
horticultores, por lo que resulta ser una de las familias con mayor riqueza de
especies entre las angiospermas. Pueden ser reconocidas por sus flores de
simetría fuertemente bilateral, en las que la pieza media del verticilo interno de
tépalos —llamada labelo— está profundamente modificada, y el o los estambres
están fusionados al estilo, al menos en la base.
Las orquídeas constituyen un grupo extremadamente diverso de plantas, que
pueden tener desde unos pocos milímetros de longitud (ciertas especies de los
géneros Bulbophyllum y Platystele) hasta constituir gigantescas agregaciones de
varios cientos de kilogramos de peso (algunas especies de Grammatophyllum) o
presentar longitudes de hasta 13,4 m, como es el caso de Sobraliaaltissima, una
orquídea recién descrita en 1999 en el Perú. Del mismo modo, las flores de las
orquídeas varían en tamaño desde menos de 1 mm y difícilmente visibles a simple
vista (Platystele) pasando por las grandes flores de 15 a 20 cm de diámetro en
muchas especies de los géneros Paphiopedilum, Phragmipedium y Cattleya hasta
los 76 cm de las flores de Phragmipediumcaudatum. La fragancia de sus flores no
es menos variable, desde el delicado aroma de Cattleya hasta el repulsivo hedor
de las flores de ciertas especies de Bulbophyllum .
Se encuentran en la mayor parte del mundo, si bien son especialmente
abundantes en los trópicos. No obstante, su capacidad de adaptación les ha
permitido conquistar un sinnúmero de nichos ecológicos, desde los más secos y
calientes del planeta hasta los más húmedos y fríos ya que, literalmente, se
distribuyen desde las regiones polares hasta el ecuador.
La familia ha sido reconocida por los sistemas clásicos de clasificación de plantas,
como el sistema de Cronquist, así como por los más modernos, como el sistema
de clasificación APG II y el sistema de clasificación APG III.

DESCRIPCIÓN
Hábito
Las orquídeas son plantas herbáceas, perennes (raramente anuales), terrestres o
epífitas, ocasionalmente trepadoras, algunas veces saprófitas o, raramente,
micoheterotróficas. Con respecto a las orquídeas epífitas, se dice que pueden
llegar a ser eternas. De hecho, en la naturaleza, su supervivencia está ligada a la
vida del árbol que las sostiene. Se conocen plantas recolectadas a mediados del
siglo XIX que todavía están creciendo y floreciendo en muchas colecciones.
Los tallos son rizomas o cormos en las especies terrestres. En las especies
epífitas, en cambio, las hojas se hallan engrosadas en la base formando
pseudobulbos que sirven para almacenar agua y nutrientes y que, por lo general,
están recubiertos por las vainas foliares membranosas que se secan con la edad.
Existen dos tipos básicos de crecimiento dentro de la familia: el tipo simpodial, que
origina tallos múltiples, y el tipo monopodial, que origina un solo tallo. El tipo
simpodial de crecimiento es el más común dentro de la familia. La mayoría de
estas orquídeas presentan pseudobulbos que funcionan como reservorios de agua
y nutrientes. La planta sostiene los pseudobulbos casi verticalmente y el
crecimiento y desarrollo posterior de nuevos tallos se produce horizontalmente,
entre los pseudobulbos preexistentes. Cada nuevo pseudobulbo se origina en la
base de los anteriores y, con su crecimiento, origina nuevas hojas y raíces. Las
hojas originadas en cada pseudobulbo pueden durar muchos años, proveyendo
nutrientes para toda la planta, hasta que se tornan marrones y mueren. Aún sin
hojas, cada pseudobulbo continúa sosteniendo el crecimiento y suministrando la
energía necesaria para el crecimiento del resto de la planta y para la floración.
Algunos ejemplos de orquídeas con este tipo de crecimiento son los géneros
Cattleya, Dendrobium y Oncidium. Las orquídeas con crecimiento monopodial, a
diferencia de las anteriores, presentan un solo tallo principal que crece erecto e

indefinidamente desde el centro de la planta. Normalmente, el tallo va creciendo
hacia arriba y se originan raíces en los nudos, las cuales crecen hacia abajo. La
planta, conforme va creciendo, pierde las hojas inferiores a medida que se forman
nuevas hojas en el extremo superior. Algunas especies de orquídeas con este tipo
de crecimiento son aquellas pertenecientes a los géneros Ascocentrum,
Phalaenopsis y Vanda.
Raíz
Las orquídeas terrestres a veces presentan raíces tuberosas. En las orquídeas
epífitas, en cambio, las raíces son aéreas y están muy desarrolladas, cuelgan de
los árboles y son verdes y gruesas. Las raíces de las epífitas tienen una doble
función, son las estructuras que se encargan de captar los nutrientes que la planta
necesita y funcionan, además, como elementos de fijación. Las raíces en este tipo
de orquídeas típicamente poseen una epidermis esponjosa, formado por muchas
capas de células muertas a la madurez y con paredes celulares engrosadas,
llamada velamen. El velamen constituye una vaina esponjosa y blanquecina que
rodea por completo a la raíz. Si el tiempo está seco, sus células están llenas de
aire; pero cuando llueve se llenan de agua.Según algunos autores el velamen es
un tejido que absorbe agua, según otros nunca se ha observado el paso de agua
del velamen al córtex de la raíz. Su función principal parece ser la de protección
mecánica, además de impedir la excesiva pérdida de agua de la raíz en períodos
de deficiencia hídrica. Además, cuando el velamen se llena de agua se vuelve
transparente permitiendo a la luz alcanzar el tejido verde de las raíces y, por ende,
facilita la fotosíntesis.
Hojas
Del rizoma o de los tallos aéreos nacen las hojas, las cuales son simples y de
margen entero, generalmente alternas, espiraladas, dísticas o verticiladas, muchas
veces plicadas, basales o a lo largo del tallo, a veces reducidas a vainas o a

escamas, usualmente con venación paralela y envainadoras en la base. Pueden
presentar pecíolo o ser sésiles y no presentan estípulas. Las especies adaptadas
a períodos de sequía tienen hojas carnosas que cumplen la función de reserva de
agua en épocas de escasez.
Flor
Ninguna familia de plantas tiene una gama de flores tan variada. Las flores están
altamente especializadas en relación a sus polinizadores. Son bien conocidas las
variaciones estructurales que facilitan la polinización por una determinada especie
de insecto, pájaro o murciélago.
Disposición de las piezas del perianto en las orquídeas, se observan los pétalos
(P), los sépalos (S) y el labelo (L).
Flor de Cattleyalabiata, obsérvese la disposición de las piezas del perianto en
relación al esquema superior.

Las flores pueden surgir, dependiendo del género y la especie, de la base de la
hoja, del rizoma o de algún entrenudo del pseudobulbo. Son hermafroditas
(raramente unisexuales), en general zigomorfas (de simetría bilateral), usualmente
resupinadas (es decir, las partes florales giran 180° durante el desarrollo), muchas
veces conspicuas y epíginas (es decir, las piezas del perianto se disponen por
encima del ovario).
En la gran mayoría de los géneros, las flores están formadas por tres piezas
externas llamadas sépalos, dos laterales y uno dorsal, y tres elementos internos
llamados pétalos, el inferior modificado en un labio o labelo de tamaño mayor y
color más intenso que los demás. Algunos autores interpretan al perianto de las
orquídeas como un perigonio, formado por seis tépalos dispuestos en dos
verticilos. Las diferentes piezas del perianto pueden estar separadas entre sí o
fusionadas en la base.
Los sépalos, o tépalos externos, son usualmente petaloideos (similares a pétalos),
imbricados. A veces los dos sépalos laterales se encuentran fusionados en un solo
elemento llamado «sinsépalo». Los pétalos, o tépalos internos, están siempre
separados, a veces presentan puntos, manchas y colores muy variados. El
llamado «labelo» es el pétalo medio, es de tamaño mayor que los dos pétalos
laterales y su forma es extremadamente variable. Es la pieza más compleja y, en
cierto modo, un órgano característico de las orquídeas. Puede ser lobulado, y
entonces se dice que existe un lóbulo central y dos laterales (Orchis,
Dactylorhiza). En otras oportunidades, como en Epipactis, se diferencian
transversalmente dos partes que se denominan «hipoquilo» la basal y «epiquilo»
la distal. Puede tener áreas brillantes, crestas, quillas u otras protuberancias que
se suelen denominar como «callo» o «callus». También es frecuente que
desarrolle un espolón dirigido hacia atrás o hacia abajo en donde se aloja un
nectario. Este espolón puede ser largo y fino (Gymnadenia, Orchis), o como un
saco redondeado (Coeloglossumviride). También hay especies en que el espolón
no tiene néctar o puede haber nectarios no incluidos en el espolón.

El androceo está usualmente formado por uno o dos estambres (a veces tres), si
es uno solo deriva del estambre medio del verticilo externo ancestral y usualmente
con dos estaminodios vestigiales derivados de los estambres laterales de un
verticilo interno ancestral. En algunas subfamilias, como en Apostasioideae o
Cypripedioideae, hay dos o tres estambres fértiles. Cuando son dos, han derivado
de los dos estambres laterales del verticilo interno ancestral, y cuando son tres, se
han originado de los dos laterales del verticilo interno y del estambre medio del
verticilo externo. El androceo se halla fusionado al estilo y al estigma, los cuales
se hallan altamente modificados, formando una estructura conocida como
«columna», «ginostemo» o «ginostegio». Las tecas de las anteras se disponen en
la porción del ginostemo denominada «clinandro» o «androclino». El polen es
granular, en tétradas o aglutinado en grupos de dos a ocho masas suaves o duras
llamadas polinias. Estas polinias presentan un apéndice filiforme —llamado
«caudícula»— que se une con una masa pegajosa —«retináculo» o «viscidium»—
sobre el «rostelo», estructura derivada del estigma con forma de lóbulo alargado y
que se sitúa sobre la porción receptiva del estigma. El conjunto de polinios,
caudículas y retináculos se denomina «polinario», el cual es la unidad de
transporte del polen durante la polinización. Las anteras son de dehiscencia
longitudinal y su conectivo muchas veces se haya modificado en un «opérculo»
que cubre la antera hasta la polinización.
El gineceo está formado por tres carpelos fusionados entre sí, con el ovario ínfero,
que puede presentar un lóculo o tres, y numerosos óvulos (hasta millones) de
placentación usualmente parietal, pero ocasionalmente de placentación axilar.
Las orquídeas son, en general, productoras de néctar, sustancia que utilizan como
recompensa a los polinizadores. Los nectarios son variables en posición y tipo.
Por ejemplo, se encuentran en el espolón del labelo, o en los ápices de los
sépalos, o en las paredes internas del gineceo. Las especies que no producen
néctar son autógamas o apomícticas, es decir, no necesitan de polinizadores para
producir semillas.18

Inflorescencia
Las orquídeas llevan sus flores de diversos modos. Aun dentro del mismo género,
las diferentes especies pueden tener distintos modos de disponer las flores en
inflorescencias, las cuales son indeterminadas y, a veces, reducidas a una única
flor, terminal o axilar. La mayoría de las orquídeas tienen inflorescencias que
llevan dos o más flores, las que usualmente nacen de un eje floral más o menos
alargado que comprende un tallo denominado pedúnculo y una porción que lleva
las flores, llamada raquis. En la mayoría de las especies las flores se disponen en
un racimo erecto y alargado, con las flores arregladas en una espiral laxa
alrededor del raquis (como, por ejemplo, en Cymbidium). En esos racimos las
flores individuales se enlazan con el eje floral a través de un corto tallito llamado
pedicelo. Puede ser que las flores se articulen con el raquis directamente, sin
pedicelo, y —en ese caso— la inflorescencia se denomina espiga, como puede
observarse en los géneros Peristylus y Neuwiedia. Un grupo de orquídeas
pertenecientes al género Bulbophyllum, bastante espectacular por su floración,
presenta el raquis tan contraído que todas las flores parecen salir del mismo
punto, como en una umbela. Algunas otras orquídeas (Oncidium, Renanthera),
finalmente, presentan inflorescencias ramificadas que se denominan panículas.
Fruto y semilla
El fruto es una cápsulaloculícida, que se abre mediante tres o seis ranuras
longitudinales (a veces una sola); en raras ocasiones, el fruto de las orquídeas es
una baya.
Las semillas son diminutas y numerosas. El tegumento es crustoso o
membranoso, sin fitomelaninas, con sólo la capa externa persistente y los tejidos
internos colapsados. Las semillas son muchas veces membranosas y aladas, los
que les permite ser dispersadas por el viento. El embrión es muy pequeño y no se
halla acompañado por endosperma, ya que este tejido aborta muy temprano en el
desarrollo embrionario.

Emitología e historia
La palabra orquídea deriva del griegoορχις (orchis = testículo), vocablo que se
encontró por primera vez en los manuscritos de la obra De causisplantarum del
filósofo griego Teofrasto y que datan aproximadamente del año 375 antes de
Cristo. Tal vocablo hace referencia a la forma de los tubérculos de las especies del
género Orchis, orquídeas de hábito terrestre cuyos tubérculos dobles parecen
testículos, como puede apreciarse en la imagen de la derecha.
Fueron conocidas y apreciadas por los seres humanos desde la Antigüedad.
Existen escritos chinos de 1.500 años de antigüedad donde se hace referencia al
cultivo de las orquídeas. En la antigua Grecia se le atribuían propiedades curativas
y afrodisíacas. Los aztecas utilizaban una orquídea —la vainilla— para enriquecer
una bebida espesa hecha a base de cacao, destinada a los nobles y a los
guerreros y era conocida con el nombre de xocoatl.
En Europa, el interés por ellas se despertó hacia 1731 cuando floreció la primera
orquídea tropical del Nuevo Mundo, Bletia purpurea (sin.Bletia verecunda), en la
colección del almirante inglés Charles Wager quien la obtuvo del Jardín botánico
de Chelsea. Desde ese momento, se suscitó un interés sin igual por la adquisición
y cultivo de orquídeas exóticas, en particular por los miembros de las clases
sociales más acomodadas, quienes debían construir un orquideario como una
obligación acorde con su estatus. De hecho, cuando una orquídea florecía en tales
colecciones, el evento daba lugar a grandes fiestas y la noticia cubría las primeras
planas de la prensa. Para satisfacer este consumo de orquídeas raras y exóticas,
durante muchos años los recolectores profesionales provenientes en su mayoría
de Francia e Inglaterra se dedicaron a saquear sin misericordia los bosques
americanos, poniendo a muchas especies en peligro de extinción. A principios del
siglo XX, no obstante, la era de la denominada «orquideomanía» llegaba a su fin.
El costo para calefaccionar los invernaderos en los que se debían cultivar estas
plantas era extremadamente alto y la carestía energética —agudizada por la

primera guerra mundial— dificultó el mantenimiento de los orquidarios privados.
Con la depresión de 1929, el cultivo de orquídeas a gran escala definitivamente
pasó a manos de empresarios comerciales.
Distribución y hábitat
Las orquídeas conforman la familia más grande de las plantas con flores, con
alrededor de 20.000 especies divididas en unos 800 géneros distribuidos por todo
el mundo. Son una familia cosmopolita, que se halla distribuida desde dentro del
Círculo polar ártico hasta Tierra del Fuego y las islas al sur de Australia. Se hallan
ausentes solamente en los desiertos verdaderos y en los polos. Son más diversas
en las regiones tropicales, donde frecuentemente son epifitas. No obstante, la
mayoría de las especies se encuentran en los trópicos y subtrópicos, desde el
nivel del mar hasta los 5000 msnm, en casi todos los ambientes. En algunos
ecosistemas son el elemento dominante, particularmente en hábitats deficientes
en nutrientes. Solamente existen dos ambientes en la tierra donde no prosperan
estas plantas, los polos y los desiertos de arena. Son más diversas en las
regiones tropicales, donde frecuentemente son epifitas. La mayor cantidad de
especies se distribuyen en las regiones tropicales, particularmente en áreas
montañosas, las cuales representan barreras naturales y aíslan a las diversas
poblaciones de plantas, lo que ocasiona la formación de un elevado número de
endemismos. Algunas áreas con una marcada predominancia de orquídeas son
las islas y el área continental del sudeste asiático y la región montañosa de
Colombia y Ecuador. El tercer sitio con un gran número de especies es la masa
atlántica brasilera con, aproximadamente, 1.500 especies descritas. Otras áreas
importantes son las montañas del sur del Himalaya en la India y China, las
montañas de América Central y el sudeste africano, notablemente la isla de
Madagascar.
Ecuador es el país que presenta la mayor riqueza de especies, llegando a totalizar
3549 taxones de orquídeas registrados, seguido por Colombia con 2723, Nueva

Guinea con 2717 y Brasil con 2590. Borneo, Sumatra, Madagascar, Venezuela y
Costa Rica son también países con un elevado número de especies.
Su capacidad para adaptarse es notable, ya que pueden crecer tanto a nivel del
mar como en los páramos elevados. Muchas viven sobre los árboles (epifitas),
otras lo hacen sobre las rocas (litófitas), otras sobre la tierra y algunas especies se
desarrollan incluso en ambientes subterráneos. A pesar de lo que mucha gente
cree, no son parásitas, ya que no se alimentan del árbol donde viven, sino que lo
usan como medio de soporte y como vehículo para alcanzar la luz del sol. Algunas
sólo miden unos pocos centímetros y otras pueden tener el porte de un árbol. Sus
flores pueden ser tan diminutas que resulta imposible observarlas a simple vista,
mientras que otras llaman poderosamente la atención.
Biología de la polinización
Por lo general, las orquídeas florecen una sola vez al año, siempre por la misma
época, la cual está determinada por factores ambientales tales como la
disminución o elevación de la temperatura, el incremento de las horas de luz, los
cambios de estación y las variaciones en la humedad ambiental. Las flores pueden
permanecer abiertas desde un día (el caso de Sobralia) hasta más de tres meses
(como en Paphiopedilum y Phalaenopsis). Los híbridos artificiales pueden florecer
dos o más veces al año.
El 97% de las especies de orquídeas necesitan de un polinizador para que se lleve
a cabo la transferencia de los granos de polen de una planta a los pistilos de otro
individuo y, por ende, para que se produzca la fecundación y la formación de las
semillas. Se debe tener en cuenta que el polen de las orquídeas se halla agrupado
en masas compactas llamadas polinias (singular: polinario), de tal modo que por sí
solo, o por acción del viento, el polen no se puede dispersar de una flor a otra por
lo que los polinizadores son imprescindibles para asegurar su reproducción
sexual. Estos polinizadores son muy variados y, según cuál sea la especie en

cuestión, pueden ser moscas, mosquitos, abejas, avispas, coleópteros y aves
(especialmente colibríes).
La zoofilia que caracteriza a las orquídeas presupone que los animales
polinizadores visiten las flores de manera regular y se detengan en ellas el tiempo
suficiente; que las anteras y el estigma sean rozados o tocados con cierta
frecuencia y que el primero quede adherido a los visitantes de modo tan perfecto
que pueda llegar con la debida seguridad a los estigmas de otras flores. El
resultado de la zoofilia depende esencialmente de que los animales puedan
reconocer las flores desde una cierta distancia y de que se vean compelidos a
visitar durante un cierto tiempo las flores de la misma especie. Las flores zoófilas,
entonces, deben poseer productos atractivos (cebos, como el polen y el néctar),
medios de reclamo (tales como olores y colores) y, además, polen viscoso o
adherente.
Muchas especies de orquídeas recompensan a los polinizadores con alimento
(como por ejemplo, néctar, pelos alimenticios o aceites) y otros compuestos, tales
como ceras, resinas y fragancias. Estas recompensas, a su vez, refuerzan la
conducta de los polinizadores. No obstante, la especialización en un solo tipo de
polinizador para asegurar una transferencia más eficiente de polen, determinó una
creciente especialización morfológica y estructural en las flores de las orquídeas
para garantizar la atracción de una sola especie de insecto.
Por esa razón, las flores de las orquídeas son de formas extremadamente
variadas y pueden atraer a una amplia variedad de insectos (abejas, avispas,
moscas, mariposas, polillas) así como a pájaros, murciélagos o sapos para la
polinización. Algunas atraen visitantes generalistas, pero muchas están bastante
especializadas, atrayendo sólo a una o unas pocas especies como polinizadores.
Polen, néctar, o fragancias florales pueden ser empleadas como
recompensadores de la polinización, mientras que algunas flores (por ejemplo
Cypripedium) manipulan a sus polinizadores y no proveen ninguna recompensa, y

algunas especies de Ophrys y Cryptostylis mimetizan la forma y el olor de las
hembras de abejas, avispas, o moscas, y son polinizadas cuando los machos
tratan de aparearse con la flor (fenómeno llamado pseudocopulación).
Generalmente, el labelo funciona como una plataforma de aterrizaje y provee
señales visuales o táctiles que orientan al polinizador. La polinia se adjunta al
cuerpo del polinizador, y muchas veces es depositada en el estigma (usualmente
una depresión en la parte de abajo de la columna) de la siguiente flor visitada. El
género Coryanthes tiene un labelo como un bolsillo que se llena con un fluido
secretado por la columna, una abeja que cae en este fluido debe viajar a través de
un túnel, forzando la deposición del polinario en su cuerpo. La transferencia de
polen dentro de la polinia es una aparente adaptación para asegurar la fertilización
de muchos del tremendo número de óvulos. En algunas especies la polinización
es un evento bastante poco común, y las flores pueden permanecer funcionales y
vistosas por muchos días. El marchitamiento del perianto ocurre rápidamente solo
después de la fertilización. Angraecumsesquipedale, una orquídea de
Madagascar, también es conocida por su biología de la polinización. Esta especie
presenta un espolón de 20 a 35 cm de largo, y es polinizada por una mariposa
esfíngida, Xanthopanmorganiipraedicta con una probóscide de esa longitud, un
hecho que Charles Darwin había predicho antes del descubrimiento de tal
polinizador.
Biología de la germinación
En la mayoría de las especies las pequeñas semillas, que son como polvo, son
dispersadas por el viento y requieren nutrientes provistos por un hongo micorrícico
para poder germinar. Algunos miembros de Cypripedioideae y Vanilloideae
poseen frutos carnosos que fermentan in situ liberando compuestos fragantes (por
ejemplo la vainillina) que atraen a pájaros y mamíferos, que actúan como agentes
de dispersión.

Estas semillas están formadas por un embrión constituido por pocas células (entre
100 y 200), cubiertas por una testa muy dura. El número de semillas puede variar
de 13.000 a 4.000.000 por cápsula. El rango de peso de una semilla de orquídea
varía de 0,3 a 14 µg y miden de 0,25 a 1,2 mm de largo y 0,009 a 0,27 mm de
ancho. Estas semillas no poseen endosperma y consisten de un pequeño embrión
suspendido dentro de una membrana, comúnmente transparente, aunque en
ocasiones pigmentada. Las formas de las semillas pueden ser muy variables,
existiendo elípticas, filiformes, fusiformes, redondas, globulares o
prominentemente aladas. Todas estas características aparentemente maximizan
la fecundidad y la efectividad en la dispersión por el viento de las semillas de
orquídeas. La germinación de estas semillas tiene lugar por medio de un proceso
que es diferente al de la mayor parte de las angiospermas, porque los embriones
de orquídeas son, desde un punto de vista anatómico y estructural,
extremadamente reducidos y simples. Los embriones de orquídeas germinan y
crecen hasta producir una masa de células llamada «protocormo». Estos
protocormos, con sus «rizoides» (estructuras en forma de raíces) pueden o no de
inmediato comenzar a fotosintetizar. No obstante, para que el protocormo
sobreviva, se desarrolle y se transforme en retoño, primero debe establecer una
relación simbiótica con un hongo.
El papel que desempeña el hongo es el de suministrar azúcares al protocormo
(especialmente a aquellos que no poseen clorofila). El hongo obtiene el azúcar de
secciones del substrato (suelo u otro objeto sólido que sirva de organismo
huésped a la planta) de la orquídea, es decir, la corteza de un árbol o del suelo. El
protocormo, a su vez, provee al hongo con ciertas vitaminas y un hábitat donde
vivir. El hongo vive en área del protocormo y del substrato. Con el tiempo, el joven
retoño comenzará a producir sus propios nutrientes y la simbiosis no será más
necesaria.
Evolución y filogenia

Se han sugerido muchos orígenes posibles para las orquídeas, no obstante, la
familia de las hipoxidáceas (o plantas similares a ellas, ya extintas) parecen ser
sus progenitores más probables. A pesar de ser la familia de angiospermas más
diversa sobre la tierra, las orquídeas no poseen un registro fósil adecuado por lo
que muchos aspectos de su historia evolutiva permanecen oscuros. No obstante,
en 2007 se ha informado el hallazgo en la República Dominicana del polinario de
una orquídea (la que fue denominada Meliorchiscaribea) preservado en ámbar y
adherido al mesoescutelo de una especie extinta de abeja(Proplebeia
dominicana). Ese fósil proviene del Mioceno, aproximadamente 15 a 20 millones
de años atrás. Este descubrimiento constituye no sólo el primer fósil de orquídea
descubierto sino también el primer antecedente fósil de las interacciones entre las
plantas y sus polinizadores. Asimismo, este descubrimiento sumado a análisis
cladísticos sobre datos de la morfología, indican que el ancestro más reciente de
las orquídeas existentes vivió en el Cretácico superior, entre 76 a 84 millones de
años atrás. La monofilia de las orquídeas está sustentada tanto por la morfología
como por los análisis de secuencias de ADN (Dressler 1981, 1993, Dressler y
Chase 1995, Burns-Balogh y Funk 1986, Juddet ál. 1993, Chase et ál. 2000, Fayet
ál. 2000, Freudensteinet ál. 2004 ). Asimismo, las relaciones filogenéticas dentro
de la familia, motivo de activas investigaciones en las últimas décadas que todavía
continúan, han sido dilucidadas mediante análisis cladísticos de la morfología y de
secuencias conservadas del ADN (Dressler 1983, 1993, Chase 1986, 1988, Chase
y Hills 1992, Chase y Palmer 1992, Cameron et ál. 1999, Koreset ál. 2000,
Whittenet ál. 2000, Salazar et ál. 2003, Burns-Balogh y Funk 1986, Cameron 2006,
Dressler 1986, 1993, Juddet ál. 1993, Dressler y Chase 1995, Cameron et ál.
1999, Freudenstein y Rasmussen 1999, Cameron y Chase 2000, Freudensteinet
ál. 2000, 2004, Molvrayet ál. 2000, van den Berget ál. 2005).
La filogenia de las orquídeas se halla suficientemente aclarada en la actualidad, lo
que permite la reconstrucción de los estados ancestrales y el análisis de la
evolución de varios caracteres adaptativos novedosos, los que incluyen los

síndromes de polinización altamente especializados, la colonización de hábitats
epifíticos y la presencia de metabolismo ácido de las crasuláceas. El metabolismo
ácido de las crasuláceas (conocido como CAM, acrónimo inglés para
CrassulaceanAcidMetabolism) es una ruta fotosintética, muy distribuida
taxonómicamente, la cual ha evolucionado en plantas que habitan ambientes con
limitaciones de agua y de dióxido de carbono, los cuales incluyen el dosel de los
bosques tropicales con disponiblidad hídrica estacional o intermitente. La
fotosíntesis C3 es el estado ancestral de la familia y el metabolismo CAM ha
evolucionado al menos diez veces de modo independiente, con varias reversiones
al estado original. Un gran evento de radiación adaptativa hacia el metabolismo
CAM ocurrió dentro de Epidendroideae, el clado con mayor riqueza de especies
epífitas de todos los grupos de plantas conocidos, el cual estuvo ligado a la rápida
evolución de una gran cantidad de especies durante el período Terciario. De
hecho, se ha demostrado que existe una asociación muy estrecha entre el
metabolismo CAM y el hábito epifítico, caracteres que habrían evolucionado
paralelamente hace unos 65 millones de años, en correspondencia con la
progresiva aridificación y reducción de la concentración de dióxido de carbono en
la atmósfera durante el Terciario. Aparentemente, la riqueza de especies en las
orquídeas está ligada a la colonización de hábitats epifíticos, y la capacidad para
hacerlo estuvo relacionada con la adquisición del metabolismo CAM, como un
medio de llevar a cabo la fotosíntesis con bajo consumo de agua. No obstante, el
cambio del hábito de crecimiento terrestre al hábito epifítico no solo requiere del
metabolismo CAM, sino también de la presencia de varios otros atributos
adaptativos. Así la presencia de raíces finas, pequeñas y trepadoras es esencial
para la adhesión a un sustrato con escasa estabilidad, como las ramas de los
árboles. La suculencia y las raíces con velamen se requieren para soportar los
períodos prolongados de deficiencia hídrica al estar fijadas a sustratos con escasa
o nula capacidad de retención de agua. Finalmente, la densidad poblacional de los
organimosepifiticos es en general muy baja, por lo que requieren de sistemas de
polinización altamente especializados para lograr la transferencia eficiente del

polen entre plantas diferentes. Todas estas adaptaciones morfológicas y
fisiológicas se han desarrollado en las orquídeas en múltiples ocasiones, lo que ha
determinado la gran riqueza de especies de esta familia y que casi el 70% de las
mismas sean epífitas.
Los géneros Apostasía y Neuwiedia (pertenecientes a la subfamilia
Apostasioideae) son considerados hermanas del resto de los miembros de la
familia (Dressler 1993, Juddet ál. 1993, Dressler y Chase 1995, Neyland y
Urbatsch 1996, Cameron et ál. 1999, Freudenstein y Rasmussen 1999 ). Estos
dos géneros, y especialmente Neuwiedia, han retenido muchos caracteres
ancestrales, como por ejemplo la presencia de dos (Apostasia) o tres (Neuwiedia)
estambres en sus flores, los que están sólo ligeramente fusionados al estilo y los
granos de polen, independientes entre sí y no pegajosos. Las restantes orquídeas,
en cambio, presentan polen pegajoso o con los granos de polen fusionados entre
sí en el momento en el que son liberados de los estambres. Dentro del grupo de
orquídeas con polen pegajoso, las subfamilias Cypripedioideae y Vanilloideae son
los clados basales. La primera de ellas (que incluye, por ejemplo, a Cypripedium y
Paphiopedilum) es claramente monofilética y sus miembros comparten algunos
caracteres exclusivos, como su labelo en forma de saco (con la forma de una
zapatilla) y la antera media modificada en un estaminodio con forma de escudo y
dos estambres funcionales. Vanilloideae (que contiene por ejemplo a Vanilla,
Pogonia y Cleistes) se distingue porque sus flores tienen sólo un estambre
funcional y tampoco presentan polinias.
Las restantes subfamilias poseen un solo estambre funcional, los granos de polen
se encuentran agrupados formando polinias y el filamento del estambre se
encuentra completamente fusionado con el estilo (Dahlgrenet ál. 1985, Burns-
Balogh y Funk 1986, Juddet ál.1993 ). Este clado, que incluye a todas las especies
con polen agrupado en polinias, tiene flores con sólo un estambre funcional (se
dicen monandras) y los dos estambres laterales están transformados en esbeltos
estaminodios o faltan completamente).

Los análisis de morfología (Freudenstein y Rasmussen1999 ) y algunos análisis
moleculares (Cameron et ál. 1999, Molvrayet ál. 2000 ) indican que las orquídeas
monandras son monofiléticas, No obstante, otros análisis moleculares (Cameron
2006, Cameron y Chase 2000, Freudensteinet ál. 2004 ), sostienen que la
reducción del número de estambres funcionales ha ocurrido en dos oportunidades
durante la evolución de la familia. Entre las orquídeas monandras con polinia se
reconocen dos grandes subfamilias, Epidendroideae y Orchidoideae (incluyendo
Spiranthoideae). Los miembros de Epidendroideae comparten la apomorfía de
presentar una antera picuda e incumbente (es decir, la antera se halla curvada
sobre el ápice de la columna), mientras que los integrantes de Orchidoideae
comparten las apomorfías de un ápice de la antera agudo, tallos suaves y hojas
convolutas. El árbol filogenético de las subfamilias es el que sigue (Juddet ál.
2007, modificado de Cameron et ál. 1999, Kocyanet ál.2004 ):
Orchidaceae Apostasioideae
polen pegajoso Cypripedioideae
Vanilloideae
polinia Orchidoideae (incluyendo
Spiranthoideae)
Epidendroideae
Apostasioideae
Las orquídeas apostasioides se consideran el grupo de orquídeas más primitivo.
Presentan dos o tres estambres en sus flores, las cuales son «regulares» y se
parecen a las del género Hypoxis (de la familia Hypoxidaceae). Las hojas se
disponen en forma espiralada en los tallos, son plegadas, resupinadas (salvo en
Apostasia). El saco embrionario es bispórico, del tipo Allium. El número
cromosómico básico es x= 24. Incluye solo dos géneros (Apostasia y Neuwiedia) y
aproximadamente 16 especies.

Sinonimia: ApostasiaceaeLindley, NeuwiediaceaeReveal&Hoogland.
Cypripedioideae
Este segundo grupo de orquídeas representan un linaje independiente, con
categoría taxonómica de subfamilia: las cipripedióideas. También retienen
características primitivas, tales como la presencia de dos estambres en las flores.
Comprende cinco géneros: Cypripedium, Mexipedium, Paphiopedilum,
Phragmipedium y Selenipedium y cerca de 150 especies, las cuales se distribuyen
en cinco tribus monotípicas. Están ampliamente distribuidas en Eurasia y a través
de América.
Conocidas popularmente como «zapatillas de dama» debido a la abultada forma
de zapatilla de su labelo que funciona como atrapa insectos, ya que el insecto es
forzado a pasar con la espalda por el estaminodio, con lo que se recolectan o
depositan los polinia. En estas orquídeas, dos anteras fértiles se disponen a cada
lado de la columna. El estambre central es estéril y está curiosamente modificado
como un escudo que impide el acceso directo de los polinizadores desde del
frente de la flor a la parte central. Los otros dos estambres están escondidos
detrás de este estaminodio. El labelo saculiforme ha evolucionado como una
trampa para los polinizadores. Las paredes internas del labelo son muy resbalosas
pero una escalera de pelos yace en el interior de la pared dorsal. Este conduce
bajo el estigma ventral a una de las dos salidas en la base del labelo a cada lado
de la columna.
Sinonimia: CypripediaceaeLindley.
Vanilloideae
Las orquídeas vanilóideas son un pequeño grupo que incluye a Vanilla, un género
de aproximadamente 70 especies de lianas. Comprende 15 géneros y

180 especies que se distribuyen en la franja tropical y subtropical húmeda del
globo y en los Estados Unidos de América.
Orchidoideae
Esta subfamilia incluye en su mayoría orquídeas terrestres con tubérculos o
rizomas carnosos. El género tipo Orchis y las "orquídeas abeja" (Ophrys, que se
denominan así porque su labelo parece el abdomen de una abeja) pertenecen a
este grupo. Comprende 208 géneros y 3.630 especies distribuidas en todo el
mundo, excepto en los desiertos más secos, en el círculo polar Ártico y en la
Antártida. Los miembros representativos de Orchidoideae incluyen a Cynorkis,
Diuris, Goodyera, Habenaria, Orchis, Platanthera, Spiranthes, y Zeuxine.
Epidendroideae
Más de 500 géneros y cerca de 20.000 especies distribuidas en las mismas
regiones de Orchidoideae, si bien incluyen algunas especies subterráneas del
desierto australiano. Epidendroideae contiene numerosas epífitas tropicales, entre
los géneros representativos se incluyen Bulbophyllum, Catasetum, Dendrobium,
Epidendrum, Encyclia, Maxillaria, Oncidium, Pleurothallis y Vanda. La delimitación
de los géneros en este grupo es notoriamente problemática, y los géneros más
numerosos no son monofiléticos. La mayoría son epífitas tropicales (normalmente
con pseudobulbos), pero algunas son terrestres e incluso unas pocas saprofitas.
6.1.3.12 Importancia económica
Tradicionalmente las orquídeas han sido utilizadas por distintos pueblos con fines
ornamentales y medicinales. Los chinos fueron los primeros en cultivarlas desde,
aproximadamente, el año 500 a. C. Más tarde, en el siglo V, los griegos las
empleaban como plantas medicinales. En América, los aztecas las utilizaban como
plantas medicinales, especias, alimenticias y ornamentales. Una de las orquídeas
empleadas por este pueblo fue la popular vainilla («tlilxóchitl» en náhuatl, nombre

científico, Vanillaplanifolia), usada para aromatizar el chocolate,y llevada a Europa
por los conquistadores españoles a principios del siglo XVI y desde ahí a regiones
tropicales como Madagascar. Este país se ha convertido en el primer productor del
mundo de esta especia, utilizada como saborizante y aromatizante en todo el
mundo.88
A pesar de la gran diversidad de la familia, pocas orquídeas son cultivadas por
otra razón que no sea la belleza de sus flores. Además del ya mencionado cultivo
de Vanilla para producir vanillina, algunas pocas especies se utilizan para la
producción de aromatizantes del té (Jumellea) o del tabaco (Vanilla). En Turquía
se utilizan los tubérculos de Anacamptismorio para la preparación de una bebida
típica caliente que se bebe en los días fríos del invierno conocida como salep.
El cultivo de las orquídeas por la belleza de sus flores evolucionó lentamente
desde un simple pasatiempo hasta la explotación comercial. Las primeras
orquídeas ornamentales llegaron a Europa, procedentes del Nuevo Mundo, en
1731. Sin embargo, no fue sino hasta 1821 cuando se inició su cultivo comercial
en invernaderos cerca de Londres. Para 1913 se inauguró en Singapur la
compañía «SunKee» para producir y comercializar flores cortadas de orquídeas.
Actualmente, en Estados Unidos, Inglaterra, Francia, Taiwán, Japón, China,
Tailandia, Australia, Hawái y Singapur se ha profundizado el interés por el cultivo y
la explotación de orquídeas, con dos objetivos definidos. El primero es el de la
producción de flor cortada para abastecer el mercado internacional de floricultura.
El segundo objetivo es el de producir y comercializar plantas de diferentes
tamaños, en particular las que se hallan cerca de la floración, para abastecer de
plantas ornamentales el mercado interno de cada país.Tailandia es uno de los
países más especializados en la producción de flores de orquídeas para abastecer
la demanda de las principales ciudades alrededor del mundo, con un monto de
exportaciones de 40 millones de dólares para el año 2001.

Entre los géneros de orquídeas más comúnmente cultivados para flor de corte o
como plantas ornamentales se destacan Cattleya, Dendrobium, Epidendrum,
Paphiopedilum, Phalaenopsis, Vanda, Brassia, Cymbidium, Laelia, Miltonia,
Oncidium, Encyclia y Coelogyne. No obstante, la mayor proporción de cultivares
actuales de orquídeas (los que se cuentan por más de 100.000) han surgido a
través de hibridaciones artificiales entre dos o más especies, muchas veces de
distintos géneros.
Multiplicación
El método más simple de multiplicación, a menudo utilizado por los coleccionistas
y por los comerciantes de pequeña escala, es la división del tallo. En varias
especies de orquídeas, como las pertenecientes al género Dendrobium, el
pseudobulbo es largo y articulado, está formado por muchos nudos en los cuales
se desarrollan hijuelos. Desde la base de estos hijuelos se desarrollan raíces.
Para multiplicar este tipo de orquídeas, entonces, solo se deben cortar los hijuelos
enraizados, separarlos de la planta madre y trasplantarlos a otro recipiente. Las
especies de orquídeas de mayor importancia comercial, tales como Cattleya,
Laelia, Miltonia y Odontoglossum, pueden propagarse por división del rizoma en
secciones, las que deben llevar de tres a cuatro pseudobulbos. Los denominados
«bulbos traseros», aquellos que ya han perdido el follaje, se usan comúnmente
para propagar clones de Cymbidium. Estos bulbos se remueven de la planta y se
colocan en otro recipiente con un sustrato adecuado para que formen raíces.
Cuidados
Luz sí, pero no sol directo, la temperatura entre 18 y 25 °C., nunca inferior a 16 °C.
Pues moriría.
El riego, una vez a la semana se coloca la maceta en un plato y se llena el plato
de agua, al rato quitar el plato o las raíces se pudrirán. Se abona una vez al mes,

siguiendo las mismas instrucciones que para el riego, es decir, diluyes el abono en
el agua del plato y al rato cuando haya absorbido toda la humedad que necesita,
retiras el plato.
Para conseguir una mayor y mejor floración, cuando las flores se secan, cortar por
encima de la tercera yema y en pocos meses volverá a brotar con más vigor. La
maceta, como en todas las plantas, se cambia cuando empiezan a asomar por
debajo raíces, esa es señal de que ya no le queda tierra, ni sitio para las raíces.
Propagación a gran escala
Fruto abierto mostrando las semillas de Cattleyawalkeriana, especie nativa del
centro de Brasil.
Debido a la gran cantidad de semillas que se producen en cada fruto y a la
posibilidad de cultivar los meristemasin vitro, las orquídeas también pueden ser
multiplicadas a gran escala.
Las semillas de las orquídeas son muy pequeñas y las que se hallan en un solo
fruto pueden generar miles de nuevas plantas, cada una con características
diferente de la otra. No obstante, las semillas contienen escasas reservas y no
pueden germinar con sus propios recursos. De hecho, en la naturaleza deben
asociarse a un hongo durante la germinación, el cual le provee los nutrientes que
requiere para su crecimiento y desarrollo. Por esta razón, la manera más simple —
si bien la menos eficiente— de multiplicar orquídeas a través de semillas consiste

en esparcir las semillas sobre y alrededor de las raíces de orquídeas cultivadas en
maceta y asegurarse de que tengan humedad constante en el sustrato. Las
semillas germinan en unas semanas y crecen muy lentamente, de manera que
una planta obtenida de este modo florece por primera vez cuando alcanza de
cinco a diez años. Este proceso se denomina «germinación simbiótica» y, hasta
1922, era el único método de propagación de orquídeas a través de semillas. En
ese año Lewis Knudson de la Universidad Cornell publicó un trabajo en el que
describía un método artificial para hacer germinar a las orquídeas sin la
participación de un hongo. Este método, llamado de «germinación asimbiótica»,
hace uso de técnicas de micropropagación para lograr la germinación y
establecimiento de las plántulas en un medio de cultivo artificial y bajo condiciones
estériles.
La reproducción a través del cultivo in vitro de meristemas, o clonación, es más
eficiente y consiste en quitar la punta de la raíz o el extremo de un brote, situarlo
en un medio de cultivo adecuado bajo condiciones estériles. Bajo la influencia de
los fitohormona los meristemas se convierten en una masa de tejido
indiferenciado, capaces de dar lugar a nuevas plántulas. Las plántulas así
obtenidas se separan unas de otras y se cultivan en tubos de ensayo
independientes. Las plantas son clones perfectos de la planta original, por lo que
éste es el método más aplicable a la propagación masiva de una variedad
particular o de híbridos estériles.
Inflorescencia de Campylocentrumgrisebachii, una de las orquídeas más
pequeñas.

Orquídeas emblemáticas o extremas
Bulbophyllumminutissimum, originaria de Australia, es la orquídea más
pequeña, con solo 3 a 4 mm de altura.
Cattleyamossiae es la flor nacional de Venezuela.
Cattleyatrianae es la flor nacional de Colombia
Guariantheskinneri es la flor nacional de Costa Rica
Lycastevirginalis es la flor nacional de Guatemala
Peristeria elata es la flor nacional de Panamá.
Rhyncholaeliadigbyana es la flor nacional de Honduras.
Sobraliaaltissima, nativa del Perú, es la orquídea más alta del mundo.
6.2.3TAXONOMÍA
Algunos autores, como por ejemplo Dahlgrenet ál. (1985), reconocieron tres
familias de orquídeas basándose en el número de anteras del androceo:
Apostasiaceae, con dos o tres anteras sólo parcialmente fusionadas al gineceo;
Cypripediaceae, con dos anteras fusionadas al gineceo y Orchidaceae, la familia
más rica en cantidad de especies, las cuales presentan una única antera
fusionada al gineceo. No obstante, todas las evidencias moleculares hasta la
fecha muestran que Orchidaceae definida o circunscripta de ese modo sería
polifilética. Los datos morfológicos y moleculares indican, en cambio, que
Orchidaceae debería definirse en forma amplia, por lo que una nueva clasificación
de la familia que se corresponde con la filogenia conocida fue publicada por Chase
y colaboradores en 2003.
6.3 Marco Contextual
Datos informativos.
Lugar de investigación: Universidad Técnica de Machala
Ciudad: Machala

Dirección y croquis de ubicación: Se encuentra localizada en el kilómetro 5 1/2 vía
pasaje ciudadela universitaria.
Para llegar a la Universidad técnica de Machala, poniendo como punto de inicio al centro
de la ciudad de Machala (parque Juan Montalvo) se deberá movilizar por la vía
Rocafuerte y se procederá a cambiar de ruta en la intersección 10 de agosto y av. 25 de
junio, procede su recorrido por dicha avenida, hasta llegar al puente de retorno a la ciudad
el cual también le sirve de ingreso a los recintos de la Universidad.
Si viene de otra ciudad, ingresar por la ―Y del Cambio‖ dirección a la ciudad de Machala,
la cual pasa por la universidad.
Teléfono: (593) 07 2983-368 Ext. 140.
Representantes:

Rector: Ing. Cesar Javier Quezada Abad Mba.
Vicerrectora académica: Ing. Laura Amarilis Borja Herrera Mg.
Vicerrector administrativo: Soc. Jorge
Ramiro Ordoñez Morejón Mg.sc.
Directors DNNA:
Director institucional de nivelación y admisión UTMACH: Ing. Iván Villacrés Mieles

Director académico de nivelación y admisión UTMACH y coordinador académico:
Dr. Rubén Lema.
Talento humano de oficina:
Secretaría: Srta. Diana Muñoz.
Asistente de dirección: Lcdo. Iván Álvarez.

Año de creación del objeto de investigación: 14 de abril de 1969.
2.1.2 Caracterización la Universidad Técnica de Máchala.
Reseña histórica de la universidad técnica de Machala.
Grandes jornadas tuvo que cumplir la comunidad Orense para lograr la fundación de la
universidad, desde las luchas en las calles que costó la vida de hombres de nuestro
pueblo, hasta las polémicas parlamentarias, como producto de los intereses que se
reflejan al interior de la sociedad.
Después de una serie de gestiones y trámites, Universidad Técnica de Machala, se creó
por la resolución del honorable Congreso Nacional de la República del Ecuador, por
decreto ley No. 69-04, del 14 de abril de 1969, publicada en el Registro Oficial No. 161,
del 18 del mismo mes y año. Habiéndose iniciado con la Facultad de Agronomía y
Veterinaria.
Por resolución oficial se encargó a la Casa de la Cultura Núcleo de El Oro, presidida por
el Lcdo. Diego Minuche Garrido, la organización de la universidad, con la Asesoría de la
Comisión de Coordinación Académica del Consejo Nacional de Educación Superior, hasta
que se designe el rector.
El 23 de julio de 1969, el señor Presidente de la República Dr. José María Velasco Ibarra,
declaró solemnemente inaugurada la Universidad Técnica de Machala en visita a la
provincia de El Oro.
Estas autoridades efectuaron programaciones y obras que reclamaban las propias
exigencias del crecimiento de la población universitaria y el desarrollo del medio. Dieron
prioritaria atención a la adecuada marcha académico-administrativa de la Universidad, a
la iniciación de la construcción del Campus Universitario y el Complejo Deportivo y a la
elevada formación científico-técnica de los estudiantes.

En lo que respecta a la construcción de la Ciudadela Universitaria se dotó de un complejo
arquitectónico a la Facultad de Agronomía y Veterinaria; y se iniciaron las obras de los
edificios de las Facultades de Sociología, Ingeniería Civil y Ciencias Químicas. Durante
esta administración se creó el Departamento de Planificación y tres nuevas carreras:
Acuacultura, Educación Parvularia, y Enfermería.
Misión y Visión.
Misión: La Universidad Técnica de Machala es una Institución reconocida en su área de
influencias formadoras de profesionales, con capacidades científico-técnicas, éticas,
solidarias, con identidad nacional, que aporta, creativamente, a través de la docencia,
investigación, vinculación y gestión, a la solución de los problemas del desarrollo
sostenible y sustentable.
Visión: La Universidad Técnica de Machala para el año 2013 es una institución
acreditada, lidera el desarrollo territorial, forma y perfecciona profesionales competentes,
emprendedores, innovadores, críticos y humanistas.

Organigrama.
6.4Metodología
En la elaboración de este proyecto un papel muy importante fue la investigación, la
lectura, y las comparaciones, siendo un tema de gran interés se encontraron mucha
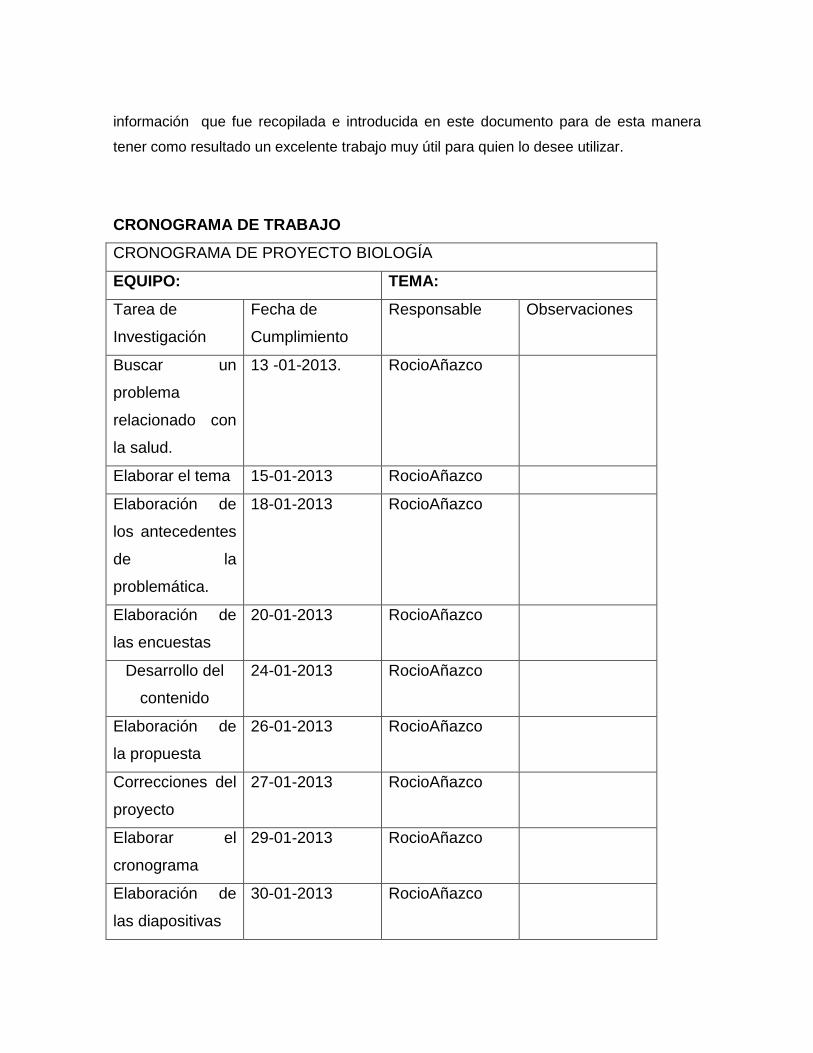
información que fue recopilada e introducida en este documento para de esta manera
tener como resultado un excelente trabajo muy útil para quien lo desee utilizar.
CRONOGRAMA DE TRABAJO
CRONOGRAMA DE PROYECTO BIOLOGÍA
EQUIPO: TEMA:
Tarea de
Investigación
Fecha de
Cumplimiento
Responsable Observaciones
Buscar un
problema
relacionado con
la salud.
13 -01-2013. RocioAñazco
Elaborar el tema 15-01-2013 RocioAñazco
Elaboración de
los antecedentes
de la
problemática.
18-01-2013 RocioAñazco
Elaboración de
las encuestas
20-01-2013 RocioAñazco
Desarrollo del
contenido
24-01-2013 RocioAñazco
Elaboración de
la propuesta
26-01-2013 RocioAñazco
Correcciones del
proyecto
27-01-2013 RocioAñazco
Elaborar el
cronograma
29-01-2013 RocioAñazco
Elaboración de
las diapositivas
30-01-2013 RocioAñazco

Defensa final del
proyecto
05-02-2013 RocioAñazco
PRESUPUESTO DEL PROYECTO (para elaborar el producto)
MATERIAL CANTIDAD VALOR
Palmera 1 0.00
Orquídea 1 $10.00
Abono 1 porción $1.00
TOTAL 11.00
PRESUPUESTO DEL PROYECTO (para la realización de este documento)
MATERIAL CANTIDAD VALOR
Investigación (Internet) Casa $0,00
Imprimir documentación 71 hojas $8.50
Imágenes 5 $0,75
Anillado 1.25 $1,25
TOTAL $10.50
PROPUESTA
Realizar campañas, para realizar injertos y mantener la variedad en la flora.
Repartir volantes para que asistan personas, a alguna conferencia y hacerles
conocer sobre los injertos y sus distintas clases para cultivar variedades con
requerimientos relativamente estrictos en materia de nutrición.

Conclusión
El injerto es el proceso por el cual dos porciones de tejido meristemático de dos
plantas son unidas, con la finalidad de que se desarrollen como si fuera una sola
planta.
Recomendación
Deshacer suavemente el cepellón antes de plantar, distribuyendo sus raíces en el hoyo de
plantación. Cortar y sanear aquellas raíces rotas, sobre todo si son grandes, puesto que
son la entrada para diversas enfermedades de hongos.
A la vez que se va plantando debemos ir regando, el primer riego debe de ser abundante.
Nunca esperar al día siguiente, a que nos den el riego o nos acaben de instalar el gotero,
etc.
Web-grafía
http://es.wikipedia.org/wiki/Arecaceae
http://www.palmerasonline.com/
http://proyectoorquidea-extremadura.blogspot.com/
http://exotic-plants-es.v145.com/como-un-injerto-phalaenopsis-orqu%C3%ADdea-
a-un-%C3%A1rbol.html
http://exotic-plants-es.v145.com/como-un-injerto-phalaenopsis-orqu%C3%ADdea-
a-un-%C3%A1rbol.html
http://es.wikipedia.org/wiki/Injerto#Historia

Glosario:
Púa: Es una porción de tejido meristemático, (yema) escogido porque en él se encuentra
una mayor actividad celular y hormonal.
Patrón: Es la parte encargada de llevar o acoger a esta yema, es decir, servir de soporte
y proveedor de nutrientes a la púa. Los patrones pueden ser de plántula si provienen de
semilla óclonales si provienen de estacas o acodos.


ENCUESTA DIRIGIDA A LOS ESTUDIANTES DE LA UNIVERSIDAD
TÉCNICA DE MACHALA.
TEMA: PROYECTO DE ELABORACIÓN DE UN MANUAL PARA
SEMBRAR UNA PLANTA DENTRO DE OTRA PLANTA MADREEN
EL ÁREA DE LA SALUD DEL CURSO DE NIVELACIÓN DE
UTMACH DE ENERO A FEBRERO DEL 2014.
OBJETIVO DE LA ENCUESTA:
Utilizar la información adquirida y llevar a práctica los pasos y métodos aprendidos
en el desempeño de este proyecto de aula.
INSTRUCCIONES:
1. Si desea guardar el anonimato, no registre nombre.
2. Los datos serán utilizados exclusivamente para el trabajo de investigación.
3. Lea detenidamente cada aspecto, antes de escribir la respuesta en cada
pregunta señale el número de opciones que se solicita.
4. No deje ninguna pregunta sin responder, marcando una (x) en el.

SISTEMA NACIONAL DE NIVELACIÓN Y ADMISIÓN
ATENCIÓN: Se le solicita que responda de acuerdo con su forma habitual de
trabajo.
DATOS GENERALES
NOMBRE DEL ENCUESTADO…………………………………
SEXO …………………………. EDAD…………..
1.- ¿Sabe usted en qué consiste un injerto?
2.- ¿Sabe usted como realizar un injerto?
SI
NO
SI
NO

3.- ¿Desea conocer las ventajas de un injerto?
4.- ¿Conoce usted a las plantas que se le pueden hacer algún injerto?
5.- ¿Desea conocer los diferentes tipos de injerto?
SI
NO
SI
NO
SI
NO

Uno de los momentos en que le ponía agua a la planta de orquídea.
Junto a la orquídea que sembré.

TEXTO CIENTÍFICO
TOMATES Y PAPAS EN UNA MISMA PLANTA
No es la primera vez que se crea una planta de este tipo, pero según sus productores, de
la marca Thomson and Morgan, es la primera vez que se hace a escala comercial."Por
encima de la tierra, los horticultores pueden recoger más de 500 tomates cherry", asegura
la compañía.Pero además, bajo tierra podrán encontrarse con una buena cosecha de
papas blancas.Según la firma, la idea ha estado desarrollándose durante los últimos 15
años.
La planta dura una temporada. "Los tomates pertenecen a la familia de las papas
(Solanaceae) y por lo tanto son naturalmente compatibles"."Cada planta se injerta
de forma manual para crear esta característica única de doble cosecha. No hay
modificación genética, es un proceso completamente natural", dice la empresa.
Una temporada
De acuerdo a GuyBarter, representante de la Real Sociedad de Horticultura de
Reino Unido (RHS) la flamante planta es de gran interés.Barter dice que ya se
habían creado muchos de estos vegetales –con la técnica de injerto– pero en

general todos tenían problemas de sabor."La estamos observando con interés
porque Thomson y Morgan es una firma muy reputada que tiene mucho que
perder y no podría descartar que sea una planta muy valiosa", dice el experto."En
el pasado no teníamos mucha fe en estas plantas, no han sido muy buenas, pero
la técnica de injerto ha mejorado mucho en años recientes"."Mucha gente no tiene
tanto espacio en sus jardines e imagino que este tipo de producto puede ser
atractivo", añade Barter."Es una operación que requiere mucha habilidad. Hemos
visto cosas similares, pero cuando se las observa en detalle, es una papa plantada
en la misma maceta que el tomate. En este caso es una sola planta que no
produce ramas de papa".La firma dice que el TomTato dura una temporada:
cuando los tomates están maduros, se pueden extraer las papas.También agrega
la empresa que se hicieron pruebas en los dos extremos de la planta para
certificar que está libre de alfa-selamina, un tóxico natural que puede producirse
dependiendo de las condiciones de crecimiento y almacenamiento.Un producto
similar, apodado en este caso Potato Tom, se presentó la semana pasada en
Nueva Zelanda.
Ejercicios aplicados con las asignaturas aprendidas anteriormente.
1.- Uno de los habitantes de la ciudadela ―ANA BOLENA‖ realizó un injerto 250
orquídeas en una planta madre, con el paso del tiempo se fueron muriendo 60
orquídeas. ¿Cuántas orquídeas le quedaron?
¿De qué trata del problema?
Del total de orquídeas
Aplica la estrategia de solución del problema.
250
60
190
Formula la respuesta del problema.
Al habitante le quedan 190 orquídeas.

2.- Analogías.
Semilla es a planta, como árbol es a fruto.
Patrón es a injerto, como rama es a hoja.


















