
Microbiota) FERMENTACIÓN) Ciego Intes-no)grueso)) · SISTEMAENDOCRINO’ENTÉRICO’(SEE)’...
20
Estómago Intes-no delgado Preestómago(s) Ciego Intes-no grueso Colon Recto Esófago Boca Microbiota FERMENTACIÓN DIGESTIÓN ABSORCIÓN La integración de estas tres funciones (diges-ón, absorción y fermentacion) con las secreciones y la mo-lidad se consigue gracias a la interacción de los sistemas nervioso (SNE) y endocrino (SEE) entéricos y el sistema nervioso autónomo (SNA, simpá-co y parasimpá-co)
-
Upload
truongkhanh -
Category
Documents
-
view
218 -
download
0
Transcript of Microbiota) FERMENTACIÓN) Ciego Intes-no)grueso)) · SISTEMAENDOCRINO’ENTÉRICO’(SEE)’...

Estómago
Intes-no delgado
Pre-‐estómago(s)
Ciego
Intes-no grueso Colon
Recto
Esófago
Boca
Microbiota FERMENTACIÓN
DIGESTIÓN ABSORCIÓN
La integración de estas tres funciones (diges-ón, absorción y fermentacion) con las secreciones y la mo-lidad se consigue gracias a la interacción de los sistemas nervioso (SNE) y endocrino (SEE) entéricos y el sistema nervioso autónomo (SNA, simpá-co y parasimpá-co)

SISTEMA NERVIOSO ENTERICO (SNE) Red de neuronas que forman circuitos complejos en la pared del tubo digestivo; tienen sus somas en los plexos submucoso (PS) y mioentérico (PM).
Las neuronas motoras del SNE que liberan acetilcolina tienen, en general, función excitatoria: contracción del músculo longitudinal, secreción en el epitelio.
Existen también neuronas motoras que liberan neurotransmisores inhibitorios, como el NO y el VIP: se conocen como neuronas inhibitorias no adrenérgicas no colinérgicas (NANC).
Las neuronas sensoriales extienden fibras hasta la mucosa, submucosa y el músculo. Son sensibles a cambios físicos o químicos en la luz y secreciones de células entero-endocrinas. Llevan la información a interneuronas o neuronas de los plexos PS y PM.
Chapter 42: Gastrointestinal Motility 471
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
including distension (stretch receptors), pH of the luminal con-tents, osmolarity, and even the presence of certain toxins. These sensory neurons can then relay that information to other neu-rons within the submucosal or myenteric plexus which may in turn activate efferent neurons within the submucosal and myen-teric nerve plexuses to respond to the detected change. The efferent neurons of the ENS can secrete a wide variety of neuro-transmitters to interact with receptors on their target cells. These include acetylcholine, norepinephrine (discussed in the section on neurotransmitters of the autonomic nervous system), dopamine, serotonin, and at least 30 other neurotransmitters and bioactive substances such as gastrointestinal peptide, vaso-active intestinal peptide (VIP), and calcitonin gene‐related pep-tide which have very specific actions within the gastrointestinal tract. Some of these actions are stimulatory and some inhibi-tory. Responses modulated by these widely varied transmitters might include contraction of the muscle layers in response to distension, secretion of fluids to neutralize acidity, and secretion of mucus to flush toxins away from an area. Interneurons can convey information between the submucosal and myenteric plexuses, so they are always in communication with one another. The ENS can control many of the actions of the gastrointestinal tract autonomously. However, its actions tend to be rather
localized and for coordinated actions over long sections of the gut, input from the autonomic nervous system is required.
Autonomic nervous system and the gastrointestinal tract
The efferent parasympathetic system is the predominant player when considering the effects of the autonomic nervous system on the gastrointestinal tract (Figure 42.5). The efferent sympathetic nervous system is often referred to as the “fight or flight” system for its action on heart and respiratory function. The parasympathetic system in contrast is referred to as the “rest and digest” system.
Mucous membrane
Muscularis mucosae
Submucous layerand plexus
Circular musclecoat
Myenteric plexus
Longitudinal musclecoat
Serous layer
Sympatheticpostganglionic !bers
Spinal afferent !bers
Sympatheticpreganglionic !bers
Vagal afferentand efferent!bers
Centripetal!bers
Figure 42.4 Scheme of the extrinsic and intrinsic innervation of the intestine. From Reece, W.O. (ed.) (2004) Dukes’ Physiology of Domestic Animals, 12th edn. Cornell University Press, Ithaca, NY. Reproduced with permission from Cornell University Press.
1 Describe the location of the presynaptic parasympathetic neurons that affect the intestinal tract.
2 How does the parasympathetic nervous system interact with the enteric nervous system to effect coordinated actions within the intestinal tract?
3 What are the major actions of the sympathetic nervous system on the intestinal tract?
Fibras centrípetas
Aferentes vagales
Eferentes vagales
Membrana mucosa
Muscular de la mucosa
Submucosa plexo submucoso
Músculo circular
Plexo mientérico
Músculo longitudinal
Capa serosa
Fibras postganglionares simpá-cas
Aferentes espinales
Fibras simpá-cas preganglionares

SISTEMA ENDOCRINO ENTÉRICO (SEE)
Células enteroendocrinas ubicadas en el eptelio de diferentes comparAmentos del tubo digesAvo
En respuesta a cambios químicos en la luz del tubos digesAvo (pH, osmolaridad, nutrientes específicos), las células del SEE secretan hormonas polipepKdicas con acción:
1) local (paracrina) sobre células vecinas o terminales eferentes del SNE o
2) a distancia (endocrina), vía circulación sanguínea, sobre otras células del sistema digesAvo o del otros sistemas corporales.
Las hormonas con efectos mejor conocidos son la gastrina, somatostaAna, colecistoquinina (CCK ), secreAna, pépAdo YY, enteroglucagon y moAlina.

BOCA
SELECCIÓN Y TOMA (O PRENSIÓN) DE ALIMENTOS Y AGUA MASTICACIÓN (TRITURADO) E INSALIVACIÓN DEL ALIMENTO
labios dientes
almohadilla dental (rumiantes), lengua (papilas gustaAvas),
carrillos, paladar duro
glándulas salivales

CONDUCTA INGESTIVA
Caballos (herbívoros perisodácAlos) Alimento: Usan los labios (muy móviles) como herramienta de selección y prensión del forraje a escala de bocado. Aprisionan el forraje entre los incisivos superiores e inferiores y cortan el bocado con un movimiento de la cabeza (predomina el movimiento lateral) Agua: Sumergen labios y parte de los carrillos en la aguada y toman agua por succión
Rumiantes (herbívoros arAodácAlos) Alimento: Usan la lengua (larga, áspera y móvil) como herramienta de prensión y selección del forraje a escala de bocado. Aprisionan el forraje entre los incisivos inferiores y la almohadilla dental y cortan el bocado con movimiento de la cabeza (hacia atrás o hacia adelante) Agua: Sumergen labios y parte de los carrillos en la aguada y toman agua por succión
Cerdos (omnívoros) Alimento: En condiciones de campo, cavan con el hocico en el suelo, y usan el labio inferior como principal instrumento de prensión de los alimentos. Agua: Sumergen labios y parte de los carrillos en la aguada y toman agua por succión
Felinos (carnívoros) y perros (carnívoros +) Alimento: Capturan la presa con patas anteriores y con la boca. Si es pequeña, pueden introducirla en la boca con movimientos de cabeza hacia arriba y atrás. Si es grande la cortan o desgarran con los molares. Agua: Toman agua con rápidos movimientos de la lengua, cuyo extremo libre curvan en forma de cuenco

MASTICACIÓN
Es la ruptura mecánica de los alimentos en la boca por accion de los dientes. Funciones: 1) Reduce el tamaño de parKculas aumentando la superficie de contacto entre las parAculas de alimento y la enzimas y acelerando la digesAón. 2) EsAmula la secreción de saliva y favorece el mezclado de las parAculas de alimento y la saliva.
CARNÍVOROS
En los carnívoros la masAcación es pobre. Los dientes Aenen superficies acuminadas y bordes filosos; se usan principalmente para cortar, arrancar y desgarrar el alimento que tragan rápidamente, casi sin masAcar. En carnívoros y omnívoros, los movimientos masAcatorios se realizan en el plano verAcal únicamente.

En los hervíboros, la masAcación es extensa en comparación con los carnívoros. Los molares Aenen superficies compuestas por cúspides y surcos, que trituran el forraje gracias a una combinación de movimientos de las mandíbulas en planos verAcal y lateral. La masAcación durante la prensión (y la rumia, en los rumiantes) es fundamental para mezclar el forraje con la saliva, conseguir una primera ruptura de las estructuras de sostén y conducción de la planta y de las paredes celulares de las células vegetales (consAtuidas por hidratos de carbono estructurales complejos con uniones glucídicas ß 1-‐4), y aumentar la superficie de acción de enzimas digesAvas o los siAos de adherencia de la microbiota en las cámaras de fermentación anteriores.
HERVÍBOROS MASTICACIÓN

470 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
The medulla coordinates the rest of the swallowing reflex. Respiration efforts are inhibited by the medulla, reducing the danger of food inhalation. Efferent motor neurons carried by cranial nerves VII, IX, X and XII carry out the following steps. The back of the tongue and floor of mouth elevate to drive the bolus to the caudal pharynx. The soft palate is elevated dorsally to close off the nasopharynx, preventing food from exiting via the nose (Figure 42.3). The hyoid apparatus elevates and the epi-glottis moves downward to cover the opening to the glottis. Laryngeal muscles tighten around the glottis as well to prevent food from entering the trachea. At this point the upper esopha-geal sphincter relaxes allowing the bolus to enter the esophagus. Snakes have an interesting adaptation that allows them to take minutes to hours to swallow their meal. In addition to unhing-ing their jaw to swallow objects many times wider than they are, snakes can extend their glottis and trachea forward and out the side of the mouth. This allows them to take in air during the entire swallowing process.
Difficulties with swallowing can signal a variety of problems in animals. Failure to close the glottis during the swallowing reflex can lead to aspiration pneumonia. Anesthetized or very weak animals can inhale vomitus or saliva because reflex centers
are depressed and do not respond to pharyngeal receptor stim-ulation that initiates the reflex. In horses, guttural pouch infec-tions can damage cranial nerves IX, X and XII interfering with the swallowing reflex. Animals born with cleft palates are unable to close off the nasopharynx and milk will be seen coming out of the nose during suckling. In all species, problems with swallow-ing can signal tumors within the medulla. As veterinarians, we must always consider rabies and other neurological diseases as a possible cause of swallowing difficulty in animals.
MOVEMENT OF INGESTA THROUGH THE GASTROINTESTINAL TRACT
Once the bolus of food has entered the esophagus the bolus takes a path through a long tube that varies in width and functions, but which essentially consists of at least two muscle layers that will act to propel the bolus down the tract. In some areas these muscles contract to prevent movement of material, forming sphincters or valves that open only occasionally. The contraction of these muscles is controlled by a novel and unique collection of neurons that form the enteric nervous system, the so‐called “brain of the gut.” This system can auton-omously control many actions within the gut. However, its actions are often coordinated over long distances in the gastro-intestinal tract by input from the autonomic nervous system and by input from endocrine or paracrine hormones produced within the intestinal tract.
The enteric nervous system
The enteric nervous system (ENS) functions from esophagus to anus. It consists of two layers of nerve cell bodies named on the basis of their location (Figure 42.4). The cell bodies of the sub-mucosal plexus (Meissner’s plexus) lie within the submucosa below the tunica mucosa. The cell bodies of the myenteric plexus (Auerbach’s plexus) lie between the inner circular smooth muscle layer that stretches around the circumference of the intestine and the outer longitudinal smooth muscle cells that runs parallel the length of the intestine. These nerve cell bodies extend sensory fibers to the secretory, absorptive, and enteroen-docrine cells lining the lumen of the gut, as well as sensory fibers within the lamina propria, submucosa, and muscle layers. These sensory neurons can detect a variety of changes within the gut,
1 Describe the location of the myenteric and submucosal plexuses.
2 What kinds of neurotransmitters are made by neurons of the enteric nervous system?
3 Describe how stretch of a segment of intestine can result in a localized segmental contraction without any input from nerves outside the intestine.
Clinical correlations
Whenever administering a pill to an animal, one must also succeed in getting the pill to the back of the tongue if it is to be actually swallowed by the patient.
Oropharynx
Nasopharynx
Esophagus
Glottis
Air
Trachea
Trachea
Oropharynx
Nasopharynx
Esophagus
Glottis
Bolus
Soft palate
Soft palate
Figure 42.3 Swallowing reflex. (Top) Normally the glottis is open and the soft palate is down to allow air to move through the nasopharynx and oropharynx into the trachea. The upper esophageal sphincter is kept closed to reduce air entry. Once food has moved to the back of the oropharynx by voluntary muscles, an involuntary reflex causes the glottis to close over the trachea and the dorsal soft palate to be elevated to close off the nasopharynx. The hyoid apparatus lifts and the upper esophageal sphincter relaxes and the bolus enters the esophagus.
Inspiración
Deglución
Orofaringe
Nasofaringe
Paladar blando
Paladar blando
Orofaringe
Nasofaringe Aire Esófago
Epiglo-s
Esófago
Bolo
Epiglo-s
Control VOLUNTARIO de la primera fase de la deglución: movimiento del bolo hacia atrás hasta la base de la lengua
Base de la lengua
Control INVOLUNTARIO de la segunda fase de la deglución: elevación del paladar blando, avance del hiodes, cierre de la tráquea por la epiglo-s, apertura del esZnter esofágico anterior
DEGLUCIÓN

ESÓFAGO
Es un órgano tubular que lleva el alimento desde la orofaringe hasta el estómago. La pared está formada por la membrana mucosa (con epitelio pano estra-ficado), la submucosa, las túnicas musculares (circular y longitudinal) y la túnica adven-cia. La luz del esófago es virtual: está ocluida en reposo y se abre para dejar pasar el alimento luego de la deglución.
EsZnter esofágico anterior (superior). Banda de músculo cricofaríngeo (estriado): cierre intenso para evitar reflujo de contenidos esofágico hacia la faringe
Presión nega-va leve: ayuda al ingreso de alimento al esófago en la deglución
EsZnter esofágico posterior (inferior). Banda gruesa de músculo liso circular: cierre intenso para evitar reflujo del contenido gástrico hacia el esófago

MÚSCULO ESTRIADO (LÍNEA PUNTEADA) Y LISO (LÍNEA CONTINUA) EN EL ESÓFAGO DE DIFERENTES ESPECIES

distensión movimientos peristál-cos propulsan el bolo hacia el estómago
VIP
SNE
relajación del esZnter esofágico posterior
-‐
Distensión
gastrina
+ contracción del esZter esofágico posterior
tono vagal (ace-lcolina)
+
PROPULSIÓN DEL BOLO DE ALIMENTO EN EL ESOFAGO

Músculos lisos del tubo digesAvo Luego de la deglución, el bolo de alimento entra en un largo sistema de tubos de diámetro variable, que forma diferentes compar-miento separados por esZnteres o válvulas.
9
b.- Cuando nos refiramos a órganos del sistema digestivo dentro de la cavidad abdominal, el siguiente esquema nos servirá como base para su análisis. Indica cuál es la estructura general de los órganos tubulares. 4.- Organización del esófago.
Túnica submucosa
Túnica muscular
Túnica serosa
Plexo mientérico
Capa muscular de la mucosa
Capa lámina propia
Capa epitelial
Túnica mucosa
a.- Elabora las referencias para las siguientes representaciones.
b.- Describe las representaciones.
c.- Formula al menos 5 preguntas sobre la organización histológica del esófago y sus componentes.
d.- Elabora un cuadro sinóptico con la histoarquitectura general del esófago.
Por fuera de la submucosa se encuentran 2 capas de músculo: la capa circular, con fibras perpendiculares al eje del tubo diges-vo, y b) la capa longitudinal, cuyas fibras siguen la misma dirección que el tubo diges-vo. El estómago -ene una cuarta capa de músculo, con fibras de dirección oblicua a las otras dos, ubicada entre la capa circular y la submucosa.
En estas túnicas se reconocen tres capas de músculo liso. La más cercana a la luz del tubo es la muscular de la mucosa: es una capa delgada ubicada en la túnica mucosa.
La pared de este tubo está cons-tuída por 4 túnicas o capas: mucosa, una submucosa, una muscular y una serosa o adven-cia.
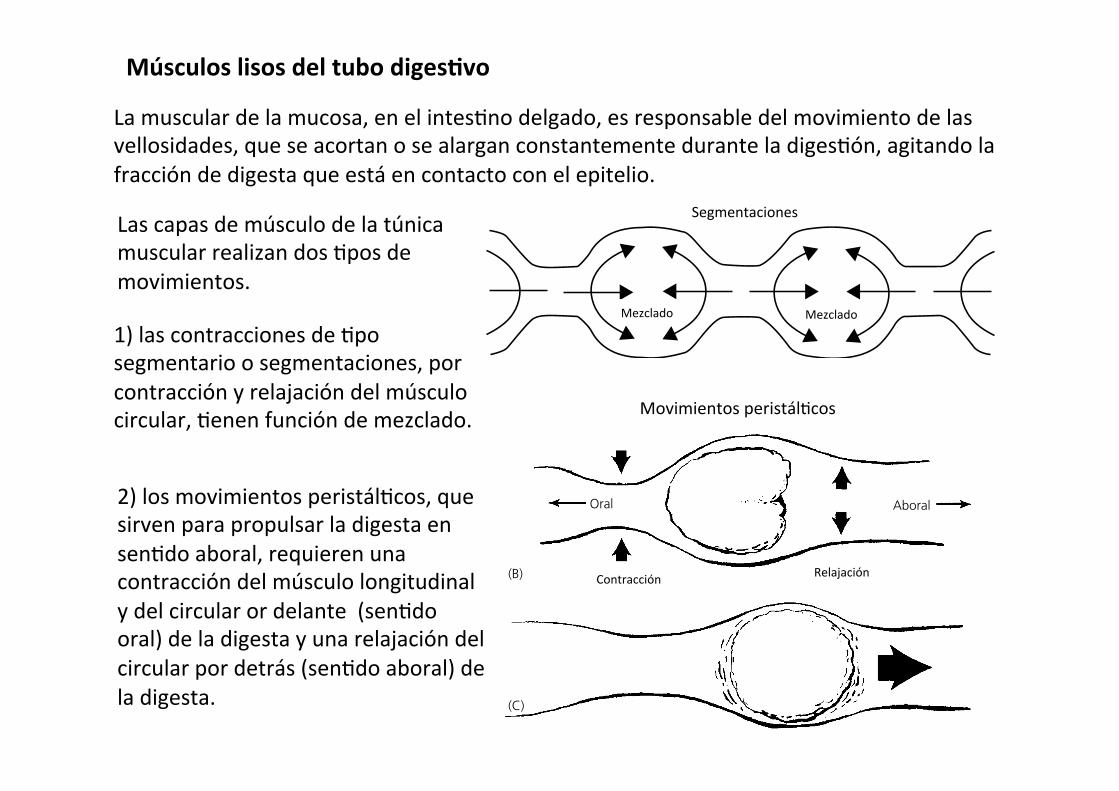
Músculos lisos del tubo digesAvo
La muscular de la mucosa, en el intes-no delgado, es responsable del movimiento de las vellosidades, que se acortan o se alargan constantemente durante la diges-ón, agitando la fracción de digesta que está en contacto con el epitelio.
2) los movimientos peristál-cos, que sirven para propulsar la digesta en sen-do aboral, requieren una contracción del músculo longitudinal y del circular or delante (sen-do oral) de la digesta y una relajación del circular por detrás (sen-do aboral) de la digesta.
Las capas de músculo de la túnica muscular realizan dos -pos de movimientos.
1) las contracciones de -po segmentario o segmentaciones, por contracción y relajación del músculo circular, -enen función de mezclado.
Chapter 42: Gastrointestinal Motility 479
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
stomach does not distend very much and the full stomach can distend to the point of initiating a vomition reflex. The stomach will try to contract (causing colic pain) and the abdominal mus-cles will contract but stomach contents cannot pass the lower esophageal sphincter. The horse’s abdominal muscles are so strong that prolonged attempts to vomit can cause the stomach wall to rupture. Interestingly, the veterinarian can pass a nasogas-tric tube through the lower esophageal sphincter into the stomach and can remove stomach contents to relieve colic symptoms.
Movement of the bolus through the small intestine
The primary stimulus for contraction in the small intestine is distension of the small intestine. Shortly after meals, mixing or segmental contractions predominate within the small intestine (Figure 42.9). Movement of ingesta through the small intestine involves peristaltic contractions of the circular and longitudinal muscles. This can be mediated over short distances (a few centi-meters) by ENS afferent and efferent neurons alone. Peristaltic contractions that move boluses of food much longer distances require coordination by the vagal sensory afferents and parasympathetic efferent fibers (Figure 42.10). Peristaltic con-tractions convey material distances of 10–20 cm in the dog as it digests its meal. Between meals, once digestion is complete it is not uncommon for peristaltic waves to propel boluses of material the entire length of the small intestine. It is believed
that this action sweeps the small intestine clean of undigestible material, protecting the small intestine from becoming popu-lated with high numbers of bacteria and reducing the risk of toxin production within the lumen.
The hormone gastrin, secreted by enteroendocrine cells of the pyloric stomach when the stomach is full, can stimulate small intestinal motility, presumably to provide more room for stomach emptying. CCK, secreted by enteroendocrine cells in the duodenum on sensing fat or amino acids in the lumen, can also stimulate contractions within the small intestine. Secretin, a hormone produced by duodenal enteroendocrine cells in response to decreased pH, slows intestinal motility. Severe distension of the intestine can sometimes occur due to blockage or necrosis of a segment of intestine. Pain from this distension elicits sympathetic discharge and release of epi-nephrine from the adrenal medulla, which causes the cessation of intestinal motility (ileus) and sweating.
Movement of the bolus through the large intestine
For material to enter the large colon it must move from the ileum to the colon (ruminants, cat and dog) through the ileoco-lic sphincter (valve). In some species the ileum empties into the cecum through the ileocecal sphincter (horse, pig, rabbit,
1 What is the difference between a segmental and a peristaltic contraction of the intestine?
Same length of intestine later in time
Mixing
Mixing Mixing
Mixing Mixing
Bidirectionalpropulsion
Propulsivesegment
Propulsivesegment
Propulsivesegment
Receivingsegment
Receivingsegment
Receivingsegment
Figure 42.9 Segmental contractions of the small intestine. Movement of chyme into the receiving (relaxed) segment by the propulsive (contracting) segment results in mixing. The receiving segment then becomes the propulsive segment, and mixing continues. From Rhoades, R.A. and Tanner, G.A. (2003) Medical Physiology, 2nd edn. Lippincott Williams & Wilkins, Baltimore. Reproduced with permission from Lippincott Williams & Wilkins.
1 Which segments of the colon are innervated by sacral spinal parasympathetic !bers?
2 What is the path of a bolus of food through the horse cecum and colon? Determine the sites most likely to suffer impaction causing colic.
Segmentaciones
Mezclado Mezclado
480 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
elephant and rat). These sphincters are normally kept closed. Distension in the ileum, and the hormone gastrin produced in response to a full stomach, cause the sphincter to relax. Distension of the colon or cecum will cause these sphincter muscles to contract more tightly to stop further entry of ingesta into the large intestine. Within the ascending colon of monogas-tric and ruminant species the contractions can be segmental mixing contractions or peristaltic contractions. The peristaltic contractions can be both antegrade (moving material toward the anus) and retrograde, which often moves material into the cecum or allows it to remain longer in the colon for extraction of more water and electrolytes. Cecal contractions can be both segmental and peristaltic and the peristaltic contractions gener-ally move material out of the cecum. Segmental mixing contrac-tions predominate in the transverse colon and both segmental and antegrade peristaltic contractions occur in the descending colon. In some species the segmented contractions of the descending colon become exaggerated, forming haustra. This allows more resident time in the descending colon to remove even more water from the ingesta before excretion. These species excrete pelleted feces. In the dog and cat (and human), between meals there can be mass aboral movement of material beginning near the ileocolic sphincter to the end of the descending colon.
As with the other segments of the gastrointestinal tract the ENS plays a role in segmental and peristaltic contractions and sensory afferent and parasympathetic input from the vagus (ascending colon) and sacral nerves (transverse and descending colon) control motility in the colon.
Megacolon may develop whenever the nerves conducting sensory afferent and parasympathetic fibers are compromised. In dogs and cats this may be due to damage to the pelvic splanchnic nerves following a traumatic experience (vehicle accident) that breaks the pelvic or sacral bones. The colon loses its parasympathetic tone and becomes greatly distended.
Horse cecum and colonThe horse is a hindgut fermenter with a cecum and ascending colon specially adapted to allow fermentation of plant cellulose and hemicellulose, providing the horse with energy in the form of volatile fatty acids. The hindgut strategy for obtaining energy from plant structural carbohydrates is also found in the rabbit, chinchilla, koala, elephant, and rhinoceros.
The cecum and ascending large colon of the horse are com-posed of three physiologically separate fermentation vats: the cecum, the ventral colon, and the dorsal colon (Figure 42.11). The dorsal colon empties into the transverse colon and then on
(A)
(B)
(C)
(D)
Distension
Intestine
Relaxation
AboralOral
Contraction
Bolus
Figure 42.10 Intestinal peristalsis and movement of contents. (A) Original distension. (B) Contraction occurs cranial to the distension and relaxation caudal to the distension. (C) Contraction and relaxation followed by movement of contents in an aboral direction. (D) A new distension point initiates a new locus of contraction and relaxation, which continues aborally as a wave.
Contracción Relajación
Movimientos peristál-cos

Músculo liso 1) reacciona más lentamente que el estriado 2) Aene filamentos de acAna más largos -‐> se contrae una distancia 3 o 4 veces mayor que una fibra estriada 3) su contracción depende del ingreso de Ca++ desde el líquido extracelular (LEC) -‐> muy suscepAble a la caída de la concentración de Ca++ en el LEC (hipocalcemia o paresa puerperal de la vaca lechera) 4) las fibras se contraen en forma simultánea con otras fibras adyacentes, con las que forman un sinciAo. Están conectadas mediante microtúbos proteicos que forman una vía de baja resistencia al pasaje de iones de una célula a otra. Los potenciales de acción se transmiten de una célula a otra sin que sea necesario que una neurona inerve individualmente a cada célula del músculo, como ocurre en el músculo estriado. 5) están en contacto, mediantes uniones gap (o de hendidura) con las Células IntersAciales de Cajal, que son el marcapasos de la acAvidad eléctrica del tubo digesAvo.
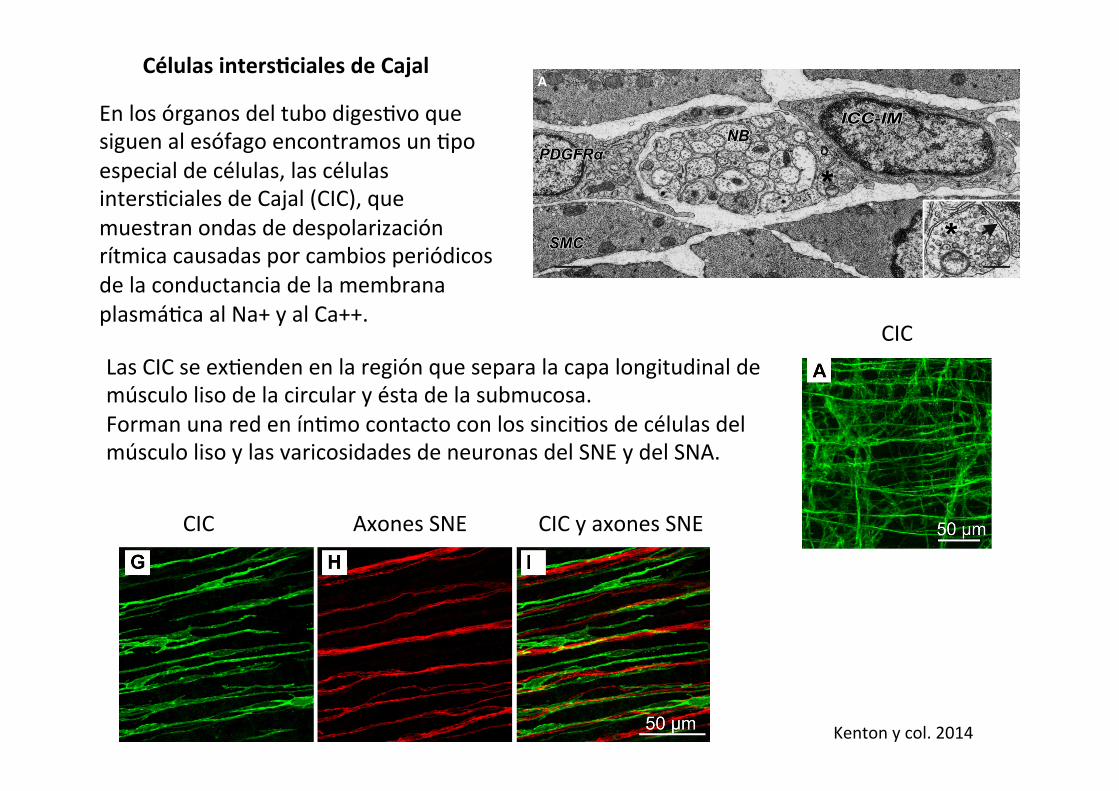
Células intersAciales de Cajal
En los órganos del tubo diges-vo que siguen al esófago encontramos un -po especial de células, las células inters-ciales de Cajal (CIC), que muestran ondas de despolarización rítmica causadas por cambios periódicos de la conductancia de la membrana plasmá-ca al Na+ y al Ca++.
stitial cells nor displayed functional defects. Thus progress inunderstanding the functions of interstitial cells in non-GI mus-cles has been slower. The discussion in this review will beginby reviewing highlights of research on ICC in the GI tract andthen focus on progress made on this class of cells in othersmooth muscle organs. Recent progress on a second class ofinterstitial cells, known for decades in the morphological liter-ature as “fibroblast-like cells,” will also be discussed. Thesecells are labeled specifically in several smooth muscles by an-tibodies against platelet-derived growth factor receptor !(PDGFR!) (174, 207), and this has provided an importantmeans of accessing these cells in physiological and genomicinvestigations.
II. ICC IN THE GASTROINTESTINAL TRACT
A. Structural Features of ICC and PDGFR!"
Cells and Networks
An important feature of GI interstitial cells (ICC andPDGFR!! cells; see FIGURE 1 and TABLE 1 for clarification ofthe terminologies used to describe these cells and their specificanatomical localizations) is electrical coupling to smooth mus-
cle cells. ICC and smooth muscle cells express a variety of gapjunction proteins (68, 136, 337), but little is known about thenature and regulation of the gap junctions between interstitialcells and smooth muscle cells. Gap junctions facilitate commu-nication between interstitial cells and smooth muscle cells(FIGURE 2). Electrical connectivity between interstitial cellsand smooth muscle cells in GI muscles causes the tunica mus-cularis to behave, in fact, as a multicellular electrical syncy-tium. We have referred to this functional structure as thesmooth muscle cell/ICC/PDGFR!! cell (SIP) syncytium (209,330; see FIGURE 3). Changes in electrical conductances in onetype of SIP cell affect the electrical excitability of the othercoupled cells. Spontaneous pacemaker activity generated byICC conducts to smooth muscle cells and drives electrical slowwaves and phasic contractions, and neural inputs to ICC andPDGFR!! cells can conduct to smooth muscle cells and mod-ulate contractile behavior. The integrated behavior of SIP cellsdefines what has been referred to classically as “myogenic”regulation of GI motor function. However, with currentknowledge, the term myogenic is too limited to define a majordivision of GI regulation, and we suggest replacement of thisterm with SIPgenic to refer to the complex and integratedregulatory processes operating beneath the level of neural andhormonal regulation. It is now recognized that remodeling or
FIGURE 1. A–C: c-Kit! ICC-MY in mouse,monkey, and human gastric antrums, re-spectively. Note similarities in the struc-tural organization of ICC in all three spe-cies. D–F: c-Kit! ICC-MY (D, green) andPDGFR!!-MY (fibroblast-like) cells (E, red)are distinct populations of cells. These pan-els show cells from murine colon. Mergedimages from D and E are shown in F. G–I:intramuscular ICC (ICC-IM; G, c-Kit is green)in the monkey gastric fundus. ICC-IM formclose associations with enteric motor nerveprocesses (nNOS! motor neuron processesare shown in H, red). I: merged image of Gand H. Scale bar in F applies to D–F, andscale bar in I applies to G–I. (Authors aregrateful to Dr. Masaaki Kurahashi for im-ages in D–F and Dr. Peter Blair for images inB and G–I.)
SANDERS ET AL.
860 Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from
stitial cells nor displayed functional defects. Thus progress inunderstanding the functions of interstitial cells in non-GI mus-cles has been slower. The discussion in this review will beginby reviewing highlights of research on ICC in the GI tract andthen focus on progress made on this class of cells in othersmooth muscle organs. Recent progress on a second class ofinterstitial cells, known for decades in the morphological liter-ature as “fibroblast-like cells,” will also be discussed. Thesecells are labeled specifically in several smooth muscles by an-tibodies against platelet-derived growth factor receptor !(PDGFR!) (174, 207), and this has provided an importantmeans of accessing these cells in physiological and genomicinvestigations.
II. ICC IN THE GASTROINTESTINAL TRACT
A. Structural Features of ICC and PDGFR!"
Cells and Networks
An important feature of GI interstitial cells (ICC andPDGFR!! cells; see FIGURE 1 and TABLE 1 for clarification ofthe terminologies used to describe these cells and their specificanatomical localizations) is electrical coupling to smooth mus-
cle cells. ICC and smooth muscle cells express a variety of gapjunction proteins (68, 136, 337), but little is known about thenature and regulation of the gap junctions between interstitialcells and smooth muscle cells. Gap junctions facilitate commu-nication between interstitial cells and smooth muscle cells(FIGURE 2). Electrical connectivity between interstitial cellsand smooth muscle cells in GI muscles causes the tunica mus-cularis to behave, in fact, as a multicellular electrical syncy-tium. We have referred to this functional structure as thesmooth muscle cell/ICC/PDGFR!! cell (SIP) syncytium (209,330; see FIGURE 3). Changes in electrical conductances in onetype of SIP cell affect the electrical excitability of the othercoupled cells. Spontaneous pacemaker activity generated byICC conducts to smooth muscle cells and drives electrical slowwaves and phasic contractions, and neural inputs to ICC andPDGFR!! cells can conduct to smooth muscle cells and mod-ulate contractile behavior. The integrated behavior of SIP cellsdefines what has been referred to classically as “myogenic”regulation of GI motor function. However, with currentknowledge, the term myogenic is too limited to define a majordivision of GI regulation, and we suggest replacement of thisterm with SIPgenic to refer to the complex and integratedregulatory processes operating beneath the level of neural andhormonal regulation. It is now recognized that remodeling or
FIGURE 1. A–C: c-Kit! ICC-MY in mouse,monkey, and human gastric antrums, re-spectively. Note similarities in the struc-tural organization of ICC in all three spe-cies. D–F: c-Kit! ICC-MY (D, green) andPDGFR!!-MY (fibroblast-like) cells (E, red)are distinct populations of cells. These pan-els show cells from murine colon. Mergedimages from D and E are shown in F. G–I:intramuscular ICC (ICC-IM; G, c-Kit is green)in the monkey gastric fundus. ICC-IM formclose associations with enteric motor nerveprocesses (nNOS! motor neuron processesare shown in H, red). I: merged image of Gand H. Scale bar in F applies to D–F, andscale bar in I applies to G–I. (Authors aregrateful to Dr. Masaaki Kurahashi for im-ages in D–F and Dr. Peter Blair for images inB and G–I.)
SANDERS ET AL.
860 Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from
CIC Axones SNE
CIC
CIC y axones SNE
Kenton y col. 2014
Las CIC se ex-enden en la región que separa la capa longitudinal de músculo liso de la circular y ésta de la submucosa. Forman una red en ín-mo contacto con los sinci-os de células del músculo liso y las varicosidades de neuronas del SNE y del SNA.
fibroblast-like (66, 113, 212, 309, 314). Cells more likesmooth muscle have more complete basal laminae andprominent caveolae; these features are more sparse or lack-ing in cells with a fibroblast-like appearance.
Most studies of ICC have been performed on mammalianspecies and limited primarily to common laboratory ani-mals and humans. Recent, morphological reports describeICC in the GI tract of zebrafish (14). The morphologicalfeatures of these cells are similar in description to theirmammalian counterpart; however, caveolae were not ob-served in either ICC or smooth muscle cells. ICC in ze-brafish can also be identified by antibodies for c-Kit.
Ultrastructural characteristics are useful for identificationof ICC and PDGFR!! cells, but the technical demands oftransmission electron microscopy (TEM) make this tech-nique suboptimal for efficient characterizations of intersti-tial cell distributions and clinical assessments of the state ofinterstitial cells in human muscles (where frequently ade-quate fixation to preserve identifying features is difficult toachieve).
1. Histological and immunomarkers for ICC
For many years interstitial cells were difficult to investigatebecause these cells represent only a fraction of the total cells
B
A
Innervation of SIP syncytium
Interstitial cells
Enteric motor neuron
Gap junctions
Kit+ (ICC-IM)PDGFR!+-IM Varicosity
SMC
PDGFR!PDGFR!PDGFR!NBNBNB
ICC-IMICC-IMICC-IM
SMCSMCSMC
****
FIGURE 3. Structures of the SIP syncytium. A: electron micrograph (originally provided by Professor Teru-masa Komuro) showing three types of cells in close proximity to nerve bundles (NB) and varicosities of entericmotor neurons (*): smooth muscle cells, ICC, and PDGFR!! cells. These cells are electrically coupled (gapjunctions not shown in this image), forming an electrical syncytium known as the SIP syncytium. In some cases,very close contacts ("20 nm; black arrow in inset) can be found between nerve varicosities and ICC or smoothmuscle cells. Scale bars are 0.5 "m in main image and 0.2 "m in inset. B: depiction of SIP syncytium (withdotted line showing approximate cut of a thin section such as shown in A). Smooth muscle cells, ICC, andPDGFR!! cells are arranged around projections of excitatory and inhibitory enteric motor neurons. [Redrawnfrom Sanders et al. (329). Copyright 2010 The Authors. Journal compilation Copyright 2010 The PhysiologicalSociety.]
INTERSTITIAL CELLS IN SMOOTH MUSCLES
863Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from

ies. It is also important to note that ICC in the region of thedeep muscular plexus (ICC-DMP) appeared normal inW/WV and Sl/Sld mice. Intestinal motility was altered inc-Kit and stem cell factor mutants, but responses to stimu-lation of intrinsic enteric motor neurons were retained andsmooth muscle tissues responded to neurotransmitters andgenerated Ca2! action potentials (80, 251, 403). Completeloss of motor activity would prohibit intestinal transit,amounting to a fatal pseudo-obstruction of the bowel.However, W/WV mice live into adulthood without obviousloss of nutritional assimilation (56). A normal pattern ofsegmentation and peristaltic waves, observed in the smallintestines of wild-type mice, was not present in W/WV mice.Intestinal muscles of these animals displayed action poten-tials and contractions, but these events were more randomand less coordinated, leading to weakened propulsive activ-ity (80).
Survival and apparent nutritional sufficiency in W/WV miceraises the question of the importance of ICC and slowwaves. Loss of ICC-MY in W/WV mice resulted in a 10- to17-mV depolarization in the smooth muscle syncytium(378, 403, 404). Depolarization alone was not responsiblefor loss of slow waves because this level of depolarization,as tested with elevated external K!, did not block slowwaves (378). A second function of ICC-MY, therefore, ap-pears to be setting membrane potential to more negativelevels than would be accomplished by the intrinsic ionicconductances of smooth muscle cells. Thus ICC-MY in thesmall intestine shift the membrane potential of the inte-grated SIP syncytium from a range in which smooth musclecells generate spontaneous action potentials to more nega-tive potentials in which the open probability of Ca2! chan-
nels is reduced, and the intrinsic excitability of smooth mus-cle cells is therefore suppressed between slow waves. Slowwaves, superimposed upon the influence of ICC-MY onmembrane potential, organize the generation of muscle ac-tion potentials and phasic contractions into the phasic pat-tern characteristic of the small intestine. The conductancesresponsible for the negative membrane potentials of ICChave not yet been determined.
2. ICC as gastric pacemakers
ICC-MY are present in the gastric corpus and antrum ofW/WV mice, but with application of neutralizing antibod-ies, lesions in gastric ICC-MY networks developed after9–40 days in organotypic cultures (279). Slow waves werenot recorded in regions of muscle where ICC-MY were lostbut persisted in areas where ICC networks remained. Elec-trophysiological recording from guinea pig stomach alsosupported the hypothesis that ICC-MY are pacemakers inthe stomach (82). When intracellular impalements weremade, slow wave activity was recorded from circularsmooth muscle cells, positively identified by dye filling dur-ing recording. More rarely, cells were impaled with muchfaster and larger amplitude slow wave events (termed driverpotentials by these authors). The cells exhibiting driver po-tentials were between the muscle layers and had morphol-ogies consistent with cells labeled with c-Kit antibodies.Simultaneous recording from ICC and nearby smooth mus-cle cells showed that the slow waves in ICC preceded theevents in smooth muscle cells (FIGURE 6). These investiga-tors also recorded from longitudinal muscle cells, and theslow waves in these cells were similar in waveform to eventsin ICC but of reduced amplitude. The slow waves recorded
Enteric motor neuron
ICC-MY
ICC-IM
CM bundle
LM
20 mv
1
2
3 sec
1
2
FIGURE 6. Propagation of slow waves from ICC-MY to circular smooth muscle cells in the guinea pig stomach(trace was provided by Professor David Hirst). In the experiment illustrated, an ICC-MY was impaled (electrode1) and a circular smooth muscle cell was impaled simultaneously. Traces above show slow wave events of largeamplitude in ICC-MY and attenuated amplitude in smooth muscle cell. Note slight delay in activation of slowwave in smooth muscle cell.
SANDERS ET AL.
870 Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from
fibroblast-like (66, 113, 212, 309, 314). Cells more likesmooth muscle have more complete basal laminae andprominent caveolae; these features are more sparse or lack-ing in cells with a fibroblast-like appearance.
Most studies of ICC have been performed on mammalianspecies and limited primarily to common laboratory ani-mals and humans. Recent, morphological reports describeICC in the GI tract of zebrafish (14). The morphologicalfeatures of these cells are similar in description to theirmammalian counterpart; however, caveolae were not ob-served in either ICC or smooth muscle cells. ICC in ze-brafish can also be identified by antibodies for c-Kit.
Ultrastructural characteristics are useful for identificationof ICC and PDGFR!! cells, but the technical demands oftransmission electron microscopy (TEM) make this tech-nique suboptimal for efficient characterizations of intersti-tial cell distributions and clinical assessments of the state ofinterstitial cells in human muscles (where frequently ade-quate fixation to preserve identifying features is difficult toachieve).
1. Histological and immunomarkers for ICC
For many years interstitial cells were difficult to investigatebecause these cells represent only a fraction of the total cells
B
A
Innervation of SIP syncytium
Interstitial cells
Enteric motor neuron
Gap junctions
Kit+ (ICC-IM)PDGFR!+-IM Varicosity
SMC
PDGFR!PDGFR!PDGFR!NBNBNB
ICC-IMICC-IMICC-IM
SMCSMCSMC
****
FIGURE 3. Structures of the SIP syncytium. A: electron micrograph (originally provided by Professor Teru-masa Komuro) showing three types of cells in close proximity to nerve bundles (NB) and varicosities of entericmotor neurons (*): smooth muscle cells, ICC, and PDGFR!! cells. These cells are electrically coupled (gapjunctions not shown in this image), forming an electrical syncytium known as the SIP syncytium. In some cases,very close contacts ("20 nm; black arrow in inset) can be found between nerve varicosities and ICC or smoothmuscle cells. Scale bars are 0.5 "m in main image and 0.2 "m in inset. B: depiction of SIP syncytium (withdotted line showing approximate cut of a thin section such as shown in A). Smooth muscle cells, ICC, andPDGFR!! cells are arranged around projections of excitatory and inhibitory enteric motor neurons. [Redrawnfrom Sanders et al. (329). Copyright 2010 The Authors. Journal compilation Copyright 2010 The PhysiologicalSociety.]
INTERSTITIAL CELLS IN SMOOTH MUSCLES
863Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from
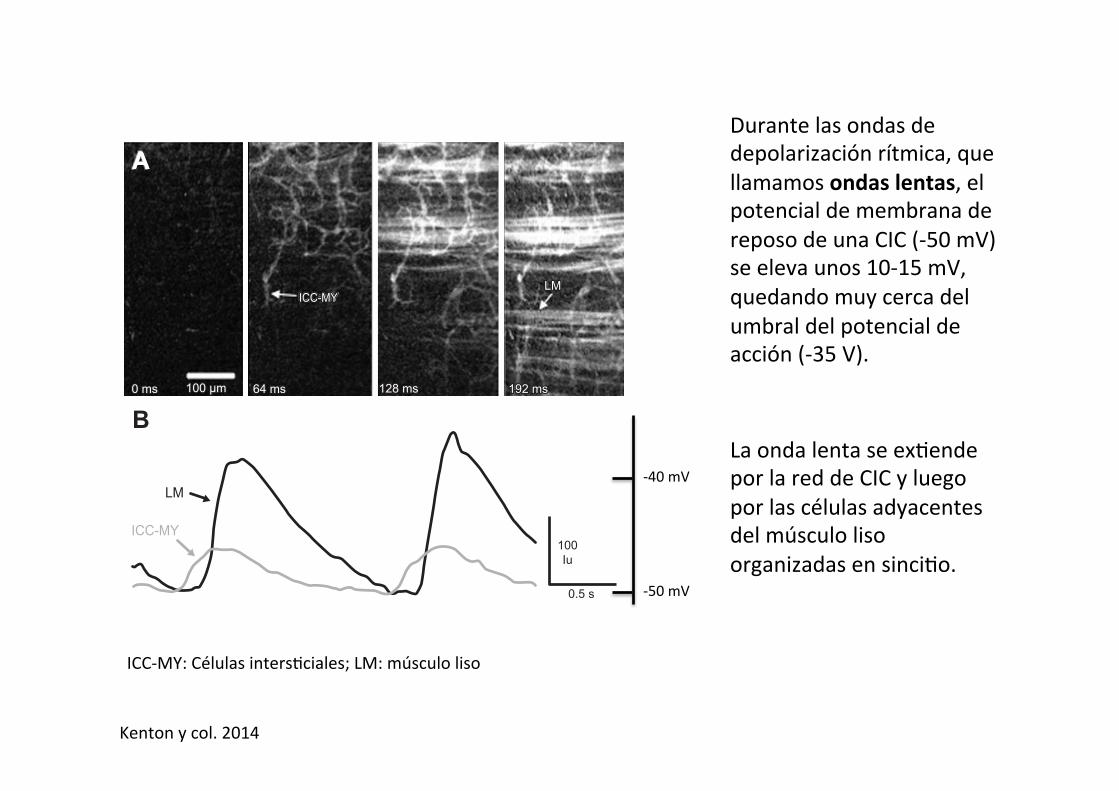
Durante las ondas de depolarización rítmica, que llamamos ondas lentas, el potencial de membrana de reposo de una CIC (-‐50 mV) se eleva unos 10-‐15 mV, quedando muy cerca del umbral del potencial de acción (-‐35 V).
La onda lenta se ex-ende por la red de CIC y luego por las células adyacentes del músculo liso organizadas en sinci-o.
ICC-‐MY: Células inters-ciales; LM: músculo liso
imaging also reinforced the pacemaker role for ICC-MYbecause Ca2! transients in closely spaced ICC-MY andsmooth muscle cells occur sequentially (FIGURE 9), as wouldbe expected if slow waves, generated in ICC-MY, initiateddepolarization and activation of voltage-dependent Ca2!
channels in neighboring smooth muscle cells (146, 147,283). Average propagation velocities of wave fronts in ICCnetworks in the mouse were between 2–6 mm/s, and there-fore consistent with the velocities of slow wave propagationin murine muscles. The propagation velocity of Ca2! tran-sients associated with slow waves greatly exceeds the ratesof Ca2! waves that result only from diffusion (10–50 !m/s)(367), supporting the idea that slow wave propagation de-pends on a voltage-dependent mechanism.
The mechanism of propagation was investigated by imag-ing of Ca2! dynamics in ICC networks, and these studies
showed that electrical coupling between cells was funda-mental to the organization of propagating Ca2! transients(283). Treatment of ICC-MY networks with 18"-glycyr-rhetinic acid, a gap junction uncoupler, did not block Ca2!
transients in individual cells, but blocked the spread of tran-sients through the network. After 18"-glycyrrhetinic acid,individual cells paced at their intrinsic frequencies, whichvaried from cell to cell, and failed to be entrained by adominant pacemaker frequency in the network. By tabulat-ing the time course of Ca2! transients in multiple cellswithin a network, it was possible to characterize the cellularsequence of activation and the time course of cellular tran-sients event to event. This analysis showed that the activa-tion sequence (i.e., spread of slow waves through a net-work) varies even when the direction of propagation re-mains constant. The time lag in activation between adjacentcells also varied from cycle to cycle, likely reflecting thedynamics of refractoriness and excitability of individualcells. These properties are consistent with the cablelikeproperties of a complex two- or three-dimensional networkof excitable cells.
Imaging studies also support the hypothesis that the volt-age-dependent mechanism for propagation is Ca2! entrythrough voltage-dependent, possibly T-type Ca2! channels(283). Nicardipine did not affect Ca2! transients in ICC-MY, but greatly reduced or blocked Ca2! transients insmooth muscle cells. Ni2! (100 !M) severely disruptedCa2! transients, and mibefradil blocked Ca2! transientspropagating through ICC networks. Drugs, such as cyclo-piazonic acid (10 !M) and 2-APB (50 !M), which affectintracellular Ca2! handling mechanisms, reduced the fre-quency of ICC-MY Ca2! transients, and ryanodine (10!M) did not affect frequency but reduced the amplitude ofthese events. These data are consistent with the idea thatCa2! entry entrains the discharge of localized Ca2! releaseevents, generates whole cell Ca2! transients, and facilitatespropagation of slow wave currents in ICC networks.
Imaging has also been used to study pacemaker activity andpropagation in larger animals and human muscles, andthese studies have allowed generalizations to be made aboutthe role of ICC in pacemaker activity and the mechanismfor pacemaking. For example, studies on muscles of thehuman jejunum showed Ca2! transients in ICC-MY withcharacteristics similar to the events in murine muscles(241). The Ca2! transients in human ICC-MY were bipha-sic in nature, and the initial component was sensitive toNi2! and mibefradil. Ca2! transients in human jejunalICC-MY were not affected by dihydropyridines, but werereduced or blocked by cyclopiazonic acid and 2-APB. Be-cause slow waves spread passively (i.e., with decrement)into smooth muscle bundles, electrical events cannot pene-trate very far into the depth of thicker muscles of largeranimals and humans (412). However, ICC lying within thesepta (ICC-SEP) between muscle bundles in these tissues
0 ms0 ms0 ms 64 ms64 ms64 ms 128 ms128 ms128 ms 192 ms192 ms192 ms
LMLMLMICC-MYICC-MYICC-MY
100 µm100 µm100 µm
AA
CaCa2+2+ Transients TransientsCa2+ Transients
see frames in Asee frames in Asee frames in A
Circ
umfe
rent
ial
Circ
umfe
rent
ial
Circ
umfe
rent
ial
1 s1 s1 s
100100µmµm100µm
100Iu
0.5 s
B
CC
LM
ICC-MY
FIGURE 9. Spread of Ca2! transients in the ICC network of murinesmall intestine. ICC were loaded with fluo 4. Sequence of videoimages shows development of Ca2! transient in ICC-MY and spreadof wave front from upper left quadrant to whole image. Streaks offluorescence are longitudinal smooth muscle cells (LM) that areabove the ICC-MY network. Note that ICC fire Ca2! transients priorto events in LM. Ca2! transients in regions of interest in ICC-MY andLM cells are shown as traces as a function of time in the bottomright of figure. Spatiotemporal maps were constructed from repet-itive Ca2! transients sweeping through the ICC-MY network. [Re-drawn from Hennig et al. (147). Copyright 2010 Blackwell Publish-ing Ltd.]
SANDERS ET AL.
878 Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from
-‐40 mV
-‐50 mV
Kenton y col. 2014

quired for slow wave generation and propagation. In fact,ion channel recovery from inactivation is categoricallymuch quicker than the long refractory period of slowwaves, suggesting that other mechanisms, such as reloadingof intracellular Ca2! stores, is likely to be the rate-limitingstep in slow wave frequency and responsible for long peri-ods of refractoriness.
ICC have a critical role in slow wave propagation becausethese cells provide the pathway for active propagation in GImuscles. Smooth muscle cells lack the ionic apparatus nec-essary to regenerate slow waves. Thus a coupled network ofICC is required for coherent propagation of slow waves andcoordination of contractions (FIGURE 8). This point wasdemonstrated by dissection experiments in which bits oftissue containing pacemaker cells were removed from mus-cles of the canine colon. In intact muscles, slow waves ofuniform amplitude were recorded from cells along the sub-
mucosal surface of the circular muscle layer, and activepropagation of slow waves occurred in this region. Slowwave activity was lost if a thin band of tissue along thesubmucosal surface (containing pacemaker ICC-SM) wasremoved, and slow waves decayed within a few millimetersfrom a region with the pacemaker cell network intact (332).A similar phenomenon was observed in gastric musclestreated with neutralizing c-Kit antibodies, slow waves weregenerated in areas where clusters of ICC were retained, butthese events did not propagate to regions devoid of ICC(279). Furthermore, after loss of ICC, gastric muscles couldnot be paced by application of current pulses.
Recordings of slow waves in ICC and from nearby smoothmuscle cells demonstrate how slow waves conduct via elec-trical coupling into smooth muscle cells. Large-amplitudeslow waves were recorded in ICC, and much attenuatedslow waves were recorded from nearby smooth muscle cells
1.
2.
3.
Single ICC
ICC network
ICC networkcoupled to SMCs
FIGURE 8. Active propagation of slowwaves. 1: The initiating step occurs by lo-calized Ca2! transients (elevated Ca2! de-picted by green color in all cells; arrow),initiating STICs in an ICC. 2: Depolarizationcaused by STICs activates Ca2! entry andCa2!-induced Ca2! release raising Ca2!
throughout the ICC and activating whole cellslow wave current. 3: Depolarization causesactive propagation of slow waves through theICC network (horizontal black arrow showsdirection of propagation), and slow wave cur-rents are activated cell to cell, as the wave-front spreads. 4: As slow waves propagatethrough the ICC network, they conduct pas-sively into electrically coupled smooth musclecells (SMCs). Slow waves depolarize SMCsand activate L-type Ca2! channels. Ca2! en-try (green) triggers SMC contraction. Spreadof slow waves in the ICC network leadsspread of contractions necessary for seg-mental and peristaltic contractions.
SANDERS ET AL.
876 Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from
Tiempo
1. Inicio de una onda lenta de despolarización en una sola CIC (área verde)
2. Despolarización completa de una sola CIC
3. La onda lenta se exAende por toda la red de CIC (verde). Las CIC del segmento de intesAno adyacnte (rosa), no se han despolarizado aún.
4. La onda lenta pasa al sinciAo de células de músculo liso (verde). Las células CIC y las células del sinciAo de músculos liso del segmento de intesAno adyacente (rosa), no se han despolarizado aún.
4.
Kenton y col. 2014

Las ondas lentas no causan contracción del músculo liso por sí mismas: solo acercan el potencial de membrana al umbral del potencial de acción. Cuando por acción del sistema autónomo, o del SNE o del las hormonas del SEE el potencial de reposo es algo más posi-vo (-‐40 mV), el potencial de membrana durante la onda lenta supera el umbral de -‐35 mV. Se pueden generar, entonces potenciales, de acción que se trasladan a los sinci-os del músculo liso , provocan la apertura de los canales de Ca++ y la contracción del músculo.
Chapter 42: Gastrointestinal Motility 475
Sect
ion
VII:
Dig
esti
on
, A
bso
rpti
on
, an
d M
etab
olis
mare pumped out of the cell, reestablishing the resting membrane potential of –50 mV. If the threshold potential to initiate an action potential is –35 mV, this rhythmic depolarization does not result in any muscular contractions. However, the resting membrane potential of these Cajal cells can be altered by ner-vous, hormonal, and mechanical influences. If a segment of
intestine is stretched by the presence of a bolus of food, or if muscarinic receptors on these cells have been activated by parasympathetic stimulation, the resting membrane potential may be reduced to just –40 mV. Now as the slow wave of depo-larization proceeds and sodium and calcium leak into the cell altering the potential difference by 10 mV, the membrane
Submucosalgland
Cells of Auerbachnerve plexus
Circularsmooth muscle
Submucosa
Muscularismucosae
Mucosa
Lumen
Mucosa
Submucosa
Circular smooth muscleLongitudinal smooth muscle
Serosa
Mesentery
Muscularis mucosae
Villi
Epithelial cells
SerosaLongitudinal smooth muscle
(B)
(A)
Figure 42.6 Schematic representation of the general organizational features of the mammalian gastrointestinal tract. (A) Cross‐section of small intestine with its mesenteric suspension that envelops the intestine as its serosa. (B) Boxed section from (A) to show greater detail. The myenteric or Auerbach nerve plexus controls gastrointestinal movements. The submucosal or Meissner plexus (not shown) is in the submucosa and controls secretions and blood flow. The muscularis mucosae produces folds in the mucosa for amplification of surface area. From Reece, W.O. (2009) Functional Anatomy and Physiology of Domestic Animals, 4th edn. Wiley‐Blackwell, Ames, IA. Reproduced with permission from Wiley.
Spikes
Slow wavesDepolarization
Hyperpolarization
Stimulation by1) Norepinephrine2) SympatheticsStimulation by
1) Stretch2) Acetylcholine3) Parasympathetics
Resting
Seconds
Mem
bran
e po
tent
ial (
mV
)
0
0 6 12 18 24 30 36 42 48 54
–10
–20
–30
–40
–50
–60
–70Figure 42.7 Membrane potentials in mammalian intestinal smooth muscle. Note the slow waves, spike potentials, and directions of depolarization and hyperpolarization. From Guyton, A.C. and Hall, J.E. (2000) Textbook of Medical Physiology, 10th edn. W.B. Saunders, Philadelphia. With permission from Elsevier.
Ondas lentas
Potencial de acción
Despolarización
Reposo
Es-mulación por 1) Norepinefrina 2) Sistema simpá-co
Hiperpolarización
Es-mulación por 1) Distensión 2) Ace-lcolina 3) Sistema parasimpá-co
Potencial de mem
brana (m
V)
Segundos
Inversamente, los eshmulos inhibitorios reducen el potencial de membrana de reposo provocando una hiperpolarización (-‐60 o -‐70 mV) de la membrana. El cambio de potencial durante la onda lenta queda más alejado del umbral, y los potenciales de acción -enen menos chances de ocurrir. Se consigue un efecto inhibitrorio, o de relajación del músculo. Kenton y col. 2014

Las ondas lentas son el origen (marcapaso) de toda la ac-vidad motora del tubo diges-vo.
intrinsic to ganglia or the tertiary plexus. Therefore, a labelimplying that they are part of the myenteric plexus is mor-phologically incorrect. Cells within muscle bundles havebeen referred to as intramuscular ICC or ICC-IM. Someauthors have used specific terms for ICC-IM within thecircular muscle (ICC-CM) or longitudinal muscle (ICC-LM); however, no distinctive morphological or functionaldata exist to suggest differences between these ICC. A spe-cialization of ICC-IM is found in the small intestine inwhich ICC are densely distributed and intimately associatedwith neurons of the deep muscular plexus, and thereforethese cells are referred to as ICC-DMP. Cells at the inneraspect of the circular muscle layer in the colon and moresparsely in this location in the stomach are in contact withthe submucosa and therefore called ICC-SM. Cells alongthe submucosal surface of the circular muscle layer aresometimes referred to as ICC-SMP, but these cells are dis-tinct and well removed from the submucous plexus, so thisdesignation is misleading. Cells within the septa that sepa-rate muscle bundles are referred to as ICC-SEP. TABLE 1provides localizations of the different classes of ICCthroughout the GI tract and attempts to also summarize thealternative terminologies that have been used in the litera-ture.
When it became possible to immunolabel fibroblast-likecells with PDGFR! antibodies (174), it was recognized thatthis population of interstitial cells occupies most of the sameanatomical niches as ICC, and therefore, similar terminol-ogy was proposed to designate these cells: 1) PDGFR!!
cells were utilized because identification by chemical codingis more specific than the vague term “fibroblast-like” cells,2) PDGFR!!-IM was applied to cells within muscle bun-dles, and 3) PDGFR!!-MY was applied to cells in the planeof the myenteric plexus between the circular and longitudi-nal muscle layers (FIGURE 1, D–I; TABLE 1).
C. Role of Interstitial Cells in Pacemakingand Generation of Electrical Rhythmicity
In many, but not all, regions of the GI tract, motor patternsare intrinsically phasic in nature. Spontaneous pacemakeractivity (termed, slow waves; FIGURE 4) drives and times thephasic contractile behavior. Slow waves occur in continu-ous rhythmic depolarization/repolarization cycles, and dif-ferent regions of the bowel display distinct intrinsic slowwave frequencies (FIGURE 4, A–C). Slow waves also vary induration from one to several seconds. Interslow wave trans-
HUMAN MURINE CANINE
mV
mV
mV
1 min
10 s
20 s
0
-55
-30
-70
0
-80
10 s 1 s
A Gastric antrum
B Small intestine
C Colon
FIGURE 4. Electrical slow waves recorded fromsmooth muscle cells in various regions of the gas-trointestinal (GI) tracts of 3 species. Slow wavesconsist of a relatively abrupt upstroke depolarizationfollowed by a stable or slowly changing plateauphase. Although frequencies and amplitudes of slowwaves vary in different tissues, note the regularity ofduration and frequency in muscles from each regionand species. The activity shown is integrated electri-cal behavior: a summation of slow waves generatedby ICC, conducted to smooth muscle cells, and re-sponses of smooth muscle cells. Depolarization ofsmooth muscle cells can activate voltage-dependentCa2! channels and generate Ca2! action potentials.Ca2! action potentials are elicited in muscles of thesmall intestine and colon during periods of enhancedexcitation. Action potentials are not typical in gastricmuscles, except in the terminal portion of the an-trum and pylorus. Note differences in resting mem-brane potentials (most negative potentials betweenslow waves) in different regions. Regulation of mem-brane potential sets the excitability of these musclesand the response elicited by slow wave activity. Dot-ted lines were drawn at "40 mV, the approximatethreshold for appreciable activation of L-type Ca2!
currents and excitation-contraction coupling. Notethat slow waves are membrane potential fluctua-tions from negative potentials where open probabilityof voltage-dependent Ca2! channels is very low topotentials where open probabilities increase andgenerate Ca2! entry. Thus slow waves are an intrin-sic mechanism organizing the motor output of GImuscles into phasic contractions. Propagation ofslow waves in ICC networks generates segmentaland peristaltic contractions. [From Sanders et al.(331).]
INTERSTITIAL CELLS IN SMOOTH MUSCLES
867Physiol Rev • VOL 94 • JULY 2014 • www.prv.org
by 10.220.32.247 on June 14, 2017http://physrev.physiology.org/
Dow
nloaded from Están presentes en todos los segmentos del tubo diges-vo, desde el estómago al colon.
Su frecuencia y velocidad varía en los diferentes compar-mentos
3
Table 1. Frequency and propagation velocity of contraction waves in stomach and small intestine in the dog, human, pig, sheep and rabbit. Maximal Velocity - frequency
[contractions/min] [cm/sec]
Stomach Dog 5,2 0.8-1.1 Rabbit 4.6 0.4-0.5 Pig 3.3 – Sheep 5.4 – Human 3 Small intestine Dog Duodenum 15.8–17.8 7-12 Jejunum 17–17.7 4.7 Ileum 13.3–13.8 0.7-0.8 Pig Duodenum 17–18 8 Jejunum 15 5.6 Ileum 11 0.5 Sheep Ileum 14.4–14.8 0.4 Human Duodenum 11 –
1.2 Function of the gastric pump
The main feature of the gastric pump is the peristaltic wave. It originates at the proximal stomach and propagates to the pylorus. The peristaltic waves are based on electrical waves originating in the gastric wall. In the wall of both the stomach and small intestine, there is a network of interstitial cells – called interstitial cells of Cajal (ICC). These interstitial cells produce electrical pacesetter potentials due to oscillations in their membrane potential. The pacesetter potential of the ICCs drive electrical events in the smooth muscle cells where they are reflected by slow waves. The pacesetter potentials and slow waves start in the proximal stomach and move aborally along the syncitium of the smooth muscle cells. The frequency of the pacesetter potentials differs among species. The pacesetter potentials determine the maximal frequency and the propagation velocity of the peristaltic waves (Table 1). However, the pacesetter potentials do not cause contractions by themselves: they are always present even when the stomach lacks any contractile activity. Contractions only occur, when excitatory neurotransmitters, one of the most prominent being acetylcholine, are released. Acetylcholine opens calcium channels during the maximum of the pacesetter potentials, so that influx of calcium into the smooth muscle cells occurs. The influx of calcium induces the electro-mechanical coupling. It is associated with the occurrence of spike potentials. The release of acetylcholine and thus the stimulation of gastric motility occurs by cephalic and gastric reflexes: they are elicited by mechano-receptors of the mouth during the ingestion of food and by mechano- and/or chemoreceptors receptors in the stomach. In the region of the gastric corpus the peristaltic waves are shallow; they represent – as mentioned above – the pump of the gastric reservoir. When the peristaltic wave reaches the antrum, the circular constriction becomes deeper. The emptying mechanism of the antral pump can be divided into three phases: 1) a phase of propulsion, 2) a phase of emptying and mixing, and 3) a phase of retropulsion and grinding (Fig. 4). Due to the regularly occurring pacesetter potentials these phases occur cyclically. When the peristaltic wave moves over the proximal antrum the previously contracting terminal antrum relaxes. Therefore chyme is propelled into the distal (or terminal) antrum (phase of propulsion).
Canine: canino; Murine: murino; Gastric antrum: antro gástrico; Small intes-ne: intes-no delgado; Maximal frequency: frecuencia máxima; Velocity. velocidad; Dog: perro; rabik: conejo; Pig: cerdo; Sheep: oveja; duodenum: duodeno; Jejunum: yeyuno; Ileum: íleon
Kenton y col. 2014 Ehrlein y Schemann, XXXX




![Semana 2. Microbiota[1]](https://static.fdocuments.ec/doc/165x107/5695d4251a28ab9b02a07179/semana-2-microbiota1.jpg)













