
Los Dinosaurios de Sangre Caliente - Adrian Desmond
1441
description
teoría sobre los dinosaurios como animales de sangre caliente
Transcript of Los Dinosaurios de Sangre Caliente - Adrian Desmond


El paleontólogo británico AdrianDesmond presenta en esta obrauna profusa recopilación de losargumentos en favor de que losdinosaurios eran de sangre calientey, al mismo tiempo, recrea de formaamena la aventura vital de laespecie que domino la Tierradurante más de 140 millones deaños. Después de leer este libro,que Isaac Asimov consideraba comoel mejor que se ha escrito sobreeste tema, se entiende por qué loscientíficos consideran ahora a losdinosaurios como animales activos,inteligentes, especializados y con

estructuras sociales desarrolladas,más cercanos a los mamíferos y lasaves que a los reptiles y lagartos.

Adrian Desmond
Los dinosauriosde sangrecaliente
ePub r1.0

Readman 07.05.15

Título original: The Hot-Blooded
Dinosaurs
Adrian Desmond, 1976Traducción: Rosana Tulla AttmanDirección científica: Jaume josa LlorcaAutores de la biografía y la presentación:Pilar Zueras y Néstor NavarreteDiseño de cubierta: Readman
Editor digital: ReadmanePub base r1.2

Los dinosaurios desangre caliente
Aun cuando los primeros restosfósiles de dinosaurios sedescubrieron en el siglo XVIII,
pasaron casi cien años antes de quelos paleontólogos tuvieran una ideaaproximada de la clase de animalesa que pertenecían tan colosalesrestos. A medida que elmonumental rompecabezas

empezaba a tomar forma, surgíannuevos interrogantes cuyasrespuestas provisionales chocabanunas veces contra las ideas de laciencia establecida y otras contralas nuevas teorías científicas que seabrían paso con fuerza, como elevolucionismo darwinista. Ya desdeun principio quedó claro que sehabían requerido dilatadísimosperíodos de tiempo para que losdinosaurios se desarrollaran yextinguieran sin que el hombrehubiera tenido posibilidad algunade conocerlos directamente. Porotra parte, el enorme tamaño demuchas especies y su súbita

desaparición sin dejar
aparentemente continuadoresplanteaban serios problemas desdeel punto de vista de la evolución. Laacumulación de hallazgos, lassucesivas mejoras en los sistemasde datación de los yacimientos enlos que se encontraban los fósiles yel espectacular avance de la
Anatomía comparada hicieronposible que hacia finales delsiglo XIX ya se pudieran hacer
reconstrucciones aceptables y setuviera una idea de la enormeantigüedad de los restos, así comode su parentesco con los reptilesmodernos.

Esta última cuestión, que se tuvocomo irrebatible durante variasdécadas, presentaba, por cierto, nopocos problemas a losinvestigadores más rigurosos. Losreptiles son animales demetabolismo exotérmico, es decir,dependen del calor externo paracalentar su propio cuerpo, por loque habitan generalmente en laszonas tropicales o templadas delplaneta y no pueden exceder ciertamasa corporal, que se consideracrítica. ¿Cómo se las arreglaban,pues, ciertas especies dedinosaurios, que sobrepasaban lasdiez, veinte y hasta treinta

toneladas, para calentar al sol unamasa tan enorme?
Las medias verdades
De haber llevado una existenciasemejante a la de los reptiles, losgrandes dinosaurios no solamentehabrían tenido que habitarexclusivamente en las partes mástórridas del planeta, sino que, aunen ese caso, les habría sidonecesario pasar más del 90% de sutiempo reposando al sol con el finde adquirir la energía que les eraimprescindible; cualquier esfuerzo

violento y continuado les habríaquedado vedado por carecer de lasuficiente energía para realizarlo, ymuchas especies habrían tenidoque habitar en zonas cálidas ypantanosas, las
únicas capaces de producirnutrientes en la cantidad suficientepara ellas. Sin embargo, muchoshechos no concordaban con esteesquema: se encontraron fósiles degrandes dinosaurios en lugares delplaneta que tenían clima frío en laépoca en que aquéllos lashabitaron, y la realización deavanzados estudios anatómicos

vino a demostrar que muchosdinosaurios eran animales veloces yágiles, con grados de actividadimpensables en un reptil.
La respuesta
Adrián Desmond recoge las dudasmanifestadas por diferentesinvestigadores a lo largo del tiempoen lo que concierne a la fisiologíade los dinosaurios para construiruna hipótesis rigurosa, y a untiempo extraordinariamenteamena: la de que los dinosauriostuvieron que ser, necesariamente,

animales de metabolismoendotérmico —de sangre caliente—,más cercanos a las aves y a losmamíferos que a los reptiles ylagartos. A partir de la década delos sesenta, el número depaleontólogos que defendía lahipótesis de los dinosaurios desangre caliente se fue haciendocada vez mayor y las pruebasacumuladas en su favoraumentaron hasta hacerseprácticamente irrebatibles veinteaños después. Hoy en día, a pesarde que las ideas populares sobreestos extraordinarios seres no sehan desprendido aún de conceptos

ya anticuados, son muchos losexpertos que están de acuerdo enconsiderar a los dinosaurios comoanimales de sangre caliente, conespecialidades complejas y conniveles avanzados de organizaciónsocial.
El largo adiós
Si la vida de los dinosaurios se haprestado a inacabablesinterpretaciones y controversias, lashipótesis sobre su desaparición nohan sido menos polémicas. AdrianDesmond recoge algunas de ellas,

como por ejemplo la que sostieneque la extinción de los dinosauriosfue producto de un envejecimientogenético. Otra explicación, muchomás pintoresca, se apoya en unapretendida «agresión química»originada tras la ingestión dealcaloides tóxicos aparecidos en lasprimeras plantas con flores, ytodavía hay especialistas quemantienen la teoría catastrofista,basada en un supuesto cataclismode proporciones cósmicas. Aunqueresulta mucho más plausible lahipótesis que contempla comogradual la extinción de esosanimales fabulosos que se

enseñorearon de la Tierra en elJurásico.
Otros libros de la colecciónrelacionados con el tema
El enigma de los dinosaurios deJohn N. Wilford
Fósiles y hombre de Eric Buffetaut

Adrian Desmond
1947 - Nace el 6 de octubre enGuilford, en el condado británico deSurrey.
1969 - Se gradúa en Ciencia yecnología por el Chelsea College
de la Universidad de Londres.
1971 - Cursa estudios sobre Historiay Filosofía de la Ciencia en laUniversidad de Londres.

1973 - Se especializa enPaleontología de los vertebradospor la Universidad de Londres yrealiza estudios complementariosen la norteamericana Universidadde Harvard.
1974 - En Natural History, publica«Central Park’s fragile dinosaurs»;se trata de un trabajo en el que elautor, combinando de modocaracterístico el rigor de lainvestigación histórica con laamenidad literaria, reconstruye elinsólito proyecto de fundación de unmuseo paleontológico en el granparque neoyorquino, acometido en

el pasado siglo; el museo debíaalbergar fantásticas reproduccionesde dinosaurios, pero éstas fuerondemolidas a martillazos en un actode caciquismo municipal.
1975 - Publica en The Times elartículo titulado «The puzzle overthe pterosaur: how did the world’sbiggest bird (!) ever manage tofly?».
1976 - Publica The hot-bloodeddinosaurs: A revolution inPaleontology, obra traducida anumerosos idiomas y que no tardaen convertirse en un bestsellermundial de la literatura de

divulgación científica.
1979 - En Ape’s reflexion, trata delos experimentos efectuados entorno al lenguaje de los signos queemplean los chimpancés.
1982 - En el marco de susinvestigaciones sobre la historia dela ciencia, publica Archetypes andancestors, un estudio sobre lasituación de la Paleontología en elLondres Victoriano.
1989 - Publica The politics oevolution, una aproximación a lasideas del evolucionismopredarwiniano.

1991 - En colaboración con JamesR. Moore, publica Darwin.

AGRADECIMIENTO
Quisiera dar las gracias a John H.Ostrom que leyó el manuscrito completo añadió numerosos comentarios y
críticas valiosas. Frances Mussetcorrigió los capítulos 4 y 7, yR. B. Freeman y Dale A. Rissell,revisaron los capítulos 1 y 8respectivamente. Las siguientes personas también contribuyeron a lacreación de esta obra ya sea por la víade la conversión o por la de lacorrespondencia: Donald Baird,Heinrich K. Erben, Kenneth A. Sellers yTony Swian. Barbara Desmond y

J. A. Cowie leyeron algunos capítulos.Es un placer para mí darle las gracias aPamela Gilí que no solamente revisó losnumerosos borradores del manuscritosino que también contribuyó de manerainmensurable al texto con ideas ysugerencias.
Jack Saxon, Leo Desmond yGeraldine Stanley ayudaron con lastraducciones al ruso, italiano y alemán.Jessica Gwynne fue la responsable delas restauraciones del pteranodón yKeith Penny hizo las fotografías de lasreconstrucciones del Crystal Palace.Eva Crawley fotografió las ilustracionesde los textos.
La investigación se efectuó en la

biblioteca de ciencias naturalesD. M. S. Watson del University Collegede Londres usando las extensascolecciones de reimpresión de losdifuntos D. M. S. Watson y Arthur SmitWoodward. Se reunió material adicionalen las bibliotecas del British Museu(historia natural), Senate House(Universidad de Londres) y del Museode Zoología Comparada de la HarvardUniversity. Le estoy muy agradecido alos bibliotecarios del UniversityCollege de Londres por tolerar misincesantes peticiones. Doy las gracias aAnthony Blond y Desmond Briggs y aJames O’Shea Wade por infundirmeconstantemente ánimo. Finalmente

quisiera expresar mi gratitud a EverettMendelson, Stephen Jay Gould y a laFacultad del Departamento de Historiade la Ciencia, Harvard University, por concederme un período de excedenciaen 1974-75, durante el cual concebí yescribí gran parte de este libro.

I. LA CUMBRE DELA CREACIÓN
En el tercer año de la RepúblicaFrancesa, el general Pichegru se hallabaserenamente cerca de las puertas deMaastricht, preparado para sitiarla. Elejército revolucionario, en su marchahacia el norte para liberar a Holanda e1795, fue frenado por la fortalezasituada en la colina de San Pedro cercade la pequeña ciudad holandesa.Próximo a la plaza fuerte se hallaba u pequeño castillo, la residencia de ueclesiástico local, el canónigo Godin, y

como tal era un objetivo digno de lashostilidades republicanas. La artilleríafrancesa bombardeó la guarnición perocuriosamente dejó intacta la residencia.El edificio también era el santuariodonde se guardaba una inapreciablereliquia y las órdenes de Pichegru erade preservarla a toda costa. Después dela capitulación, el general ordenó a sustropas que registraran la casa y seapoderaran de la reliquia para laRepública. Sin embargo, llegó tarde yaque durante la noche de la batalla elastuto canónigo se había llevado estavaliosa antigüedad a la seguridad de laciudad.
El tesoro que detuvo el avance de

los franceses era una pareja demaxilares fosilizados de aspectoimpresionante que constituían los restosde una bestia enorme que había cazadoen el mundo preadanita. Las noticias delcráneo, con colmillos como dagas en susmaxilares de doce metros de longitud, sehabían difundido rápidamente por todaEuropa, alcanzando el fósil ciertanotoriedad por las extraordinariascontroversias que lo rodeaban alládonde fuera. Nadie era capaz de afirmar con certeza de qué tipo de monstruoantediluviano se trataba. La aparición dela criatura había estado precedida por una amarga disputa y una costosaquerella legal para determinar al

legítimo propietario. Por lo tanto no eraninguna sorpresa que su presencia lefuera conocida al general francés. Laimportancia del cráneo fosilizado, siembargo, no residía en su dramáticaentrada en una Europa revolucionariasino en la profunda influencia queejerció sobre los paleontólogos duranteel siguiente medio siglo. Tambiéayudaría a efectuar una revolución en el pensamiento del hombre acerca de lavida primitiva de su planeta: haríaverosímil la idea de la extinción. Por otro lado, durante muchas décadasllevaría a conclusiones erróneas a loscientíficos que intentaban comprender lanaturaleza de los enormes saurios que

vivieron en la época mesozoica.Durante el siglo XVIII, las canteras
calizas de Maastricht fueron famosas por su riqueza en conchas fósiles, particularmente en curiosidades talescomo amonitas, belemnitas y erizos demar fosilizados. La colina de San Pedroera una zona de prospeccióespecialmente rica; se habían labradouna serie de canteras calizasdirectamente en el corazón de lamontaña y posteriormente ampliadocreando vastas galerías con techossostenidos por grandes pilares a modode las criptas románicas. Con laapertura de nuevas cámaras y laampliación de las antiguas, toda la

montaña quedó surcada por cavernassubterráneas. La extracción de talcantidad de creta provocóinevitablemente el desenterramiento demuchos fósiles inusuales y por lasgalerías subterráneas se pasearon loscoleccionistas con antorchas en busca deestos restos antediluvianos. Fue en 1770cuando la montaña ofreció su posesiómás espectacular: los trabajadores quese hallaban a gran profundidad de lacantera encontraron a 455 metros dedistancia de la entrada principal losmaxilares de un animal verdaderamentemonstruoso incrustado en roca sólida.Uno de los coleccionistas locales, el Dr.Hoffmann, un cirujano militar alemá

retirado, que había trabajado durantemuchos años en la región parasuministrar material al Museo Teylor deHaarlem, fue convidadoapresuradamente para que diera sopinión como experto. Deseandoadquirir los impresionantes maxilares para sí mismo, Hoffmann recompensógenerosamente a los trabajadores queservicialmente extrajeron el resto deeste valioso trofeo. El bloque fueexcavado intacto y Hoffmann volvió acasa con él. El anatomista holandésPieter Camper fue requerido para laidentificación de los enigmáticosmaxilares. Éste supuso que eran losrestos de una vieja ballena, una

suposición natural teniendo en cuenta eltamaño de los maxilares y las conchasmarinas encontradas en las rocascircundantes. Su hijo, Adrien Camper,sin embargo, declaró sorprendentementeque la bestia no era un mamífero sino ulagarto marino monstruoso. Siembargo, debido a que el mundo actualno es habitado por lagartos de estetamaño, otros científicos pensaron queera más probable que los maxilares pertenecieran a un cocodrilo prehistórico.
Mientras tanto, la controversia querodeaba a este inmenso cráneo se habíaconvertido en el tema principal dediscusión en la ciudad y finalmente llegó

a oídos del canónigo Godin, que era el propietario de los prados situados por encima de la cantera. Habiéndose dadocuenta de que poseía una valiosaventaja, solicitó el prestigioso fósilinvocando sus derechos feudales en uintento de reclamar la reliquia preadanita.
Godin llevó a Hoffmann ante lostribunales y ganó el juicio, entrando asíen posesión de los maxilaresfosilizados, mientras que Hoffmann tuvoque pagar las costas judiciales. Elcanónigo Godin colocó la reliquia euna urna de cristal y la expuso en scasa de campo, cerca de la montaña, para que pudieran venir a verla los

curiosos.Tuvieron que pasar varios años
hasta que finalmente se hizo justicia ycuando esto sucedió, Hoffmann ya habíamuerto. Cuando Maestricht cayó emanos de los republicanos, el canónigoescondió su tesoro en la ciudad.Pichegru se tomó la revancha a lamañana siguiente ofreciendo 6000 botellas del mejor vino a la persona quele trajera el fósil. Las tropas registrarola ciudad y una docena de granaderosllevaron triunfalmente los maxilaresfosilizados al general. Desde aquí, la bestia antediluviana fue llevada como botín a París donde se instaló en elJardin des Plantes[1]. Aquí, el anatomista

francés más importante del momento,Georges Cuvier, estableciódefinitivamente su valor científico.
En la época durante la cual se habíaapoderado de la capital el reino delTerror, el joven Cuvier se hallaba en elcampo, en Normandía, coleccionandomoluscos y estudiando la fauna local,mientras se ganaba la vida con utrabajo menor como oficinista. Cuvier era, sobre todo, un excelenteadministrador. Su talento y susinvestigaciones no pasaron inadvertidos en 1795, cuando sólo contaba 26 años
de edad, fue llamado a París paratrabajar junto al evolucionista Jean-Baptiste Lamarck en el Jardin des

Plantes. En París inmediatamente se hizo patente el don de Cuvier para laorganización. El trabajo de su vidaestuvo dominado por el deseo de poner todo en orden. La zoología encontró asía un sistematizador de fósiles y
apoleón a su vicepresidente delMinisterio del Interior, cuya función fuela reinstauración de un estricto ordesocial. Las reglas de la ley eran uimperativo político bajo el nuevorégimen, al igual que el poderoso ordenatural era un imperativo zoológico,siendo ambos perfectamentecompatibles bajo la competencia deCuvier. Como resultado de su exhaustivoestudio sobre los fósiles y animales

vivos, especialmente de los grandescuadrúpedos, Cuvier finalmente estuvoconvencido de que habían desaparecidorazas enteras de animales de lasuperficie de la Tierra. La extinción eraun viejo dilema. Existían razonesmetafísicas profundamente enraizadas para descartarla y siempre había faltadola prueba concluyente para lademostración inequívoca. A finales delsiglo XVII, el naturalista inglés John Rayhabía jugado con la idea de la extinció pero, al final, había vuelto a la opinióconvencional de que, criaturas como lasamonitas de concha en espiral, podíaestar viviendo en lugares remotos del planeta. (El descubrimiento del

emparentado nautilo en el sur delPacífico justificó parcialmente esteenfoque; el nautilo es el únicosuperviviente de una raza de moluscoscon tentáculos, parecidos a loscalamares, que viven en conchas deespiral compartimentadas yfrecuentemente hermosas). El predecesor de Cuvier en el siglo XVIII,el conde de Euffon, pensó en u principio que los restos de mastodontehallados en los bancos de arena del ríoOhio podrían haber pertenecido a unaespecie extinguida. Él también volvió ala explicación más tradicional,sugiriendo que el cráneo del mastodonteamericano no era otra cosa que una

mezcla de colmillos de elefante ydientes de hipopótamo. Así, la cuestiósiguió estando inconclusa hasta la épocade Cuvier, y resulta irónico que elderrumbamiento de una de las ideas másmimadas desde la época de Aristótelesse debiera a un incondicional y, segútodas las informaciones, bastante pomposo defensor de la ley y del orden.
Fue solamente cuestión de mesesdespués de la llegada como trofeo deguerra a París de los maxilaresfosilizados de Godin, que Cuvier finalizara su trabajo sobre fósiles yelefantes aún vivos y anunciara, deforma bastante categórica, que algunostipos de elefantes en efecto se había

extinguido[2]. La extinción se hizorealidad. El instrumento de Cuvier parademostrar su teoría era el rigurosoestudio comparativo de todos losespecímenes disponibles. Su técnicaconsistía en comparar lascorrespondientes estructuras dediferentes animales y correlacionar funciones y, en último término, con losdiferentes modos de vida. Declaró queel mastodonte americano y el mamutsiberiano no eran idénticos a loselefantes que aún vivían y que, debido aque era improbable que éstos siguieraviviendo en la Tierra, sólo se podíaconcluir que se habían extinguido. Lasimplicidad del razonamiento es

engañosa ya que existía un poderoso,aunque rara vez explícitamenteformulado, argumento contrario a éste, basado en gran medida en la fe. Estaidea inhibitoria era el concepto del
lenum: es decir, que un Dios generososólo podía haber poblado el mundo cocualquier tipo imaginable de organismocomo señal de su omnipotencia. Estosorganismos podían ser ordenados en unacadena de infinitas graduaciones, queabarcarían desde la criaturamicroscópica más pequeña hasta elhombre. La extinción rompería la cadenadebido a la introducción de saltos,situación considerada por los devotoscomo reflejo adverso de la fuerza del

creador.La prueba objetiva y detallada de
Cuvier, que apoyaba la extinción de losmamuts, destruyó finalmente la fe delhombre en el plenum y eliminó el primer eslabón de la cadena. También preparóel terreno para la aceptación de otrosmonstruos prehistóricos que no habíalogrado sobrevivir. Mientras que elmamut había vivido en un pasadoreciente, el «gran animal de Maestricht»,tal como era llamado, había vivido eépocas muy remotas y se parecía aúmenos a las criaturas de nuestros días.Cuvier concluyó que cuanto más hacia el pasado se miraba, menos se parecían lascriaturas a las existentes en la

actualidad. El reconocimiento de estateoría se produjo forzosamente debidoal descubrimiento en Baviera de u pterodáctilo, del tamaño de un pichón,en 1784, identificado por Cuvier como bestia con alas pero también comoreptil. Durante la época mesozoica, laTierra estaba poblada por seres exóticos extraños que hoy en día resultaría
bastante desconocidos. Cuvier razonóque fuera cual fuera el tipo de catástrofeque había exterminado a los mamuts eun reciente pasado, ésta también debíade haber causado estragos en épocasmás remotas de la historia de la Tierra,teniendo como víctimas a los extrañosreptiles mesozoicos.

Cuvier examinó el cráneo deMaestricht y confirmó que era, tal comohabía sugerido Adrien Camper, ulagarto emparentado con los monitoresque aún vivían en los trópicos, si bien setrataba de un ejemplar de proporcionesasombrosas. Mientras tanto, eInglaterra, el reverendo WilliaConybeare, en ausencia de otrassugerencias, había bautizado a lacriatura: la llamó Mosasaurus o«lagarto del Mosa» de acuerdo con laregión de donde procedía[3]. Cuvier hizouna interpretación completamentecorrecta. Pero a diferencia de losmonitores aún vivos, el mosasauriohabía sufrido modificaciones destinadas

a una existencia marina, se alimentabade peces, sus miembros se habíaconvertido en aletas y presentaba unafuerte cola para desplazarse. Elreverendo William Buckland especulóque, en base a la longitud del cráneo,«este monitor monstruoso de las remotas profundidades mediría entre 1,5 y 6metros de longitud[4]», aun cuando elmás grande de sus parientes entre loslagartos aún vivos apenas alcanzaba lamitad de este tamaño.
Cuando se desenterraron los primeros huesos de lo que hoy en díaconocemos como dinosaurios,comprensiblemente se supuso que éstos procedían de los lagartos gigantes

similares. «Puede parecer extraño — dijo Buckland acerca del mosasaurio deMaestricht— observar que susdimensiones superan a las de cualquier lagarto aún vivo, o encontrar génerosmarinos en el orden de los saurios, yaque actualmente no existe ningunaespecie en este orden capaz de vivir eel mar. Sin embargo, resulta apenasmenos extraño que las desviacionesanálogas encontradas en el
egalosaurus e Iguanodon, que proporcionan ejemplos de una expansióaún mayor, como las del monitor o de laiguana, hacia formas colosalesadaptadas para desplazarse haciatierra[5]». Fue el propio Buckland quien,

en 1824, describió el primero de estosgigantes terrestres. Buckland era profesor de mineralogía y geología en laUniversidad de Oxford por lo que eldescubrimiento de grandes huesos en lascanteras de pizarra de Stonesfield, asólo unos pocos kilómetros de distancia,atrajo naturalmente su atención y losfósiles rápidamente encontraron elcamino hacia el Museo de Oxford.
Aunque ninguno de los huesos fuedesenterrado en contacto con otros, aexcepción de algunas vértebras,Buckland se mostró bastante firme corespecto a la posición de la criatura eel esquema zoológico de las cosas, talcomo sugiere el título de su artículo

«Notice on the Megalosaurus or greatFossil Lizard of Stonesfield[6]». (Notasobre el megalosaurio o gran lagartofosilizado de Stonesfield). Influenciado por la convincente demostración deCuvier, de que los grandes maxilares deMaestricht habían pertenecido en otrostiempos a un monitor mesozoico,Buckland supuso naturalmente lo mismo para las enormes vértebras y huesos delos miembros que se hallaban en smuseo. De hecho, el nombre que seinventaron él y su cómplice en la causacatastrofista, el reverendo WilliaConybeare, delata las supuestasafinidades del monstruo: Megalosaurus
significa «lagarto gigante». Su tamaño

superaba incluso las estimaciones deBuckland para el mosasaurio. El fémur más grande del museo medíaescasamente 91 centímetros de longitud tenía una circunferencia de 20
centímetros en su punto más estrecho.Cuando se le informó sobre la existenciade este hueso, el barón Cuvier estimóque «si la criatura tenía las proporciones de un lagarto», probablemente habría tenido unalongitud superior a los 12 metros y elvolumen de un elefante.
Al igual que Cuvier, Buckland se basó en estos restos de reptil como prueba de que la Tierra había sidodevastada por una catástrofe. Si

embargo, Buckland añadió una nuevacontroversia teológica. Cuvier, ausiendo un devoto cristiano, había sido prudente al no mezclar la Biblia y lageología y había evitado hacer especulaciones acerca de la naturalezadel agente desencadenante de lasrevoluciones terrenales. (Nunca puso eduda de que éstas tuvieran unaexplicación natural). También cuestionóla extensión global de las revoluciones;la inundación de algunos continentesaislados habría sido suficiente. Perocuando su Preliminary Discourse, quetrataba sobre estas revoluciones(publicado por primera vez en 1812),cayó en manos de los traductores

ingleses en 1813, su tono fue alteradosubrepticiamente y el Diluvio Universalsustituyó a la última revolució(proporcionándole así a la Biblia ciertarespetabilidad científica). El lector inglés tenía la impresión como si el propio Cuvier estuviera criticando eldiluvio. Buckland fue un heredero deesta tradición inglesa. El reverendoBuckland, que más tarde se convertiríaen el Deán de Westminster, era uteólogo, que no compartía lasreticencias de Cuvier de utilizar a laciencia para apoyar a la RevelacióDivina, modificó drásticamente elsistema cuveriano y, por lo tanto, lo que para Cuvier había sido una reciente

incursión marina local se convirtió euna gran oleada enviada por elTodopoderoso para ahogar el mundo.Buckland buscó en todos los lugares delmundo —entre los trofeos de antiguasexpediciones al Himalaya y en lascuevas de Yorkshire— pruebas queconfirmaran que la inundación habíasido universal y catastrófica. Además,como las extinciones más recientes sehabían producido después de unainundación de inimaginable fuerza, losreptiles gigantescos debían de haber encontrado su fin en una inundacióanterior. El Todopoderoso, en ucaprichoso acto de «interferenciacreativa», había anulado el antiguo

orden; los huesos fósiles de Stonesfielderan el testimonio de la periodicidad delos diluvios universales enviado por Dios.
Gideon Mantell, un médico ycoleccionista de fósiles que vivía eLewes, no lejos de Brighton en la costasur de Inglaterra, había encontradohuesos gigantes similares en el bosquede Tilgate. Mantell también poseía elfémur de un Megalosaurus, si bien éstetenía una circunferencia de 50centímetros. Una vez más, tomandocomo base la analogía de los lagartos,se pensó que este fósil procedía de unacriatura de unos 18 o 21 metros delongitud. El bosque de Tilgate era una

buena zona de prospección para sauriosgigantes y ya había proporcionado aMantell restos de cocodrilos gigantes y plesiosaurios. También se habíaencontrado dientes enormes, diferentes atodos los observados hasta entonces. Lacorona de cada diente estaba desgastadaformando una superficie oblicua y suave la forma del diente revelaba que s
propietario se había alimentado de plantas. La cuestión de si los dientes podían proceder de un gran reptil eradudosa, debido a que actualmenteexisten muy pocos reptiles que seacapaces de masticar materia vegetal.Debido a que Charles Lyell estaba a punto de visitar París, Mantell le dio u

diente para que se lo mostrara al baróCuvier. Éste inmediatamente lo catalogócomo el incisivo superior de urinoceronte. Mantell no pudo aceptar que los rinocerontes hubieran vividounto a reptiles gigantes ni que los
dientes procedieran de estratossuperiores de la tierra, tal como habíasugerido Cuvier, de forma que llevó losfósiles al Museo Hunterian del ColegioReal de Cirujanos de Londres. Fue allídonde vio los dientes de las iguanas deSudamérica; el parecido era obvio einequívoco. Los pequeños dientes de lasiguanas se parecían tanto a los dientesgigantescos encontrados en las rocasmesozoicas, que Mantell incluso jugó

con la idea de darles a ambos el mismonombre. Sin embargo, a propuesta deConybeare, decidió darle a este precursor mesozoico el comprometidonombre de Iguanodon o «diente deiguana[7]». Ésta era la pruebaindependiente, en caso de ser necesaria,de que los saurios mesozoicosgigantescos eran simplemente lagartosglorificados. Mantell pensó que si unaiguana poseía dientes de este tamaño, lacriatura completa debía de medir más de16 metros.
Cuvier, Buckland y Mantell habíautilizado a los monitores o a las iguanascomo modelos para estimar el tamañode los supuestos antepasados de los

lagartos en el mundo antiguo. Éstosaumentaban el lagarto a escala, de formaque su fémur, diente o cualquier partedel esqueleto tuviera el mismo tamañoque el del fósil y, a continuación,calculaban el tamaño global del animalfosilizado. Los huesos fosilizados,especialmente los fémures, teníafrecuentemente una longitud quince oveinte veces superior a la de un lagarto,lo cual llevaba a la estimación dedimensiones fantásticas para los sauriosextinguidos. La descripción que hizoBuckland del dedo de 15,24 centímetrosde longitud de una de estas primitivas bestias hallada en la isla de Wight, nodeja lugar a dudas respecto al temor que

estos monstruos inspiraban. «Éste es — dijo— el hueso metacarpiano más largodescubierto hasta el momento; y si nosreferimos al animal extinguido del cual procedía, la escala según la cual las personas de la antigüedad medían aHércules…, debemos concluir que elsujeto, de cuyo cuerpo formaba parte,era el cuadrúpedo más enorme queamás haya pisado la superficie de
nuestro planeta[8]». El mosasaurio deCuvier, el primero de los leviatanesmesozoicos en ser desenterrado,difícilmente podría haber ejercido másinfluencia sobre los primeros paleontólogos. Muchos de loscoleccionistas de fósiles del siglo XIX

eran aficionados y sin sus incansablestrabajos de excavación, los geólogos-clérigos profesionales como Bucklandhabrían sufrido en gran medida unaescasez de materia prima con la cualtrabajar. Fueron los aficionadosentregados y perseverantes comoThomas Hawkins, Gideon Mantell yMary Anning, que vivían cerca de losacantilados mesozoicos ricos en fósilesde la costa sur de Inglaterra, los que por primera vez desenterraron muchos delos imponentes saurios gigantes que eotros tiempos habían dominado laTierra. Fueron estos mismos visionarioslos que trabajando la piedra calizaolítica inglesa (que data del período

urásico medio) y las arenas y arcillasdel cretáceo inferior (de los condados boscosos de Kent y Sussex) estimularola imaginación pública con cuentosacerca de monstruos marinos, dragonesvoladores y enormes lagartos terrestresque habitaban la Tierra en las épocasanteriores a Noé. Muchos de los primeros pioneros eran hombres ymujeres devotos, y algunos parecían ser eclesiásticos fanáticos, incluso para losniveles de aquella época. De esta forma,la investigación profunda sobre loshabitantes arcaicos del globo terráqueono solamente proporcionó una nuevavisión sobre las obras de laProvidencia, sino que también ofreció

un punto de vista distinto de la versió bíblica.
Debido a que Cuvier interpretó queel primer saurio había sido como ulagarto gigante (lo que en realidad fue),el Iguanodon y el Megalosaurus
recibieron un estatus similar. Cuvier siguió siendo reverenciado: la última palabra anatómica la tenía él. Econsecuencia, los lagartos constituyeroel estándar según el cual se comprendía —y medían— los extinguidos saurios, procedimiento que llevó a tamañosaparentemente absurdos para estosreptiles arcaicos.
Fue Richard Owen, profesor delColegio Real de Cirujanos de Londres,

quien finalmente rompió con latradición. El 2 de agosto de 1841, en lareunión de la Asociación Británicacelebrada aquel año en Plymouth, Owe pronunció un discurso de dos horas ymedia de duración titulado «Informesobre los reptiles fósiles en GraBretaña». Con 38 años de edad, Oweaun siendo relativamente joven, era laindiscutible estrella en ascensión de la paleontología, y era llamado por todosel Cuvier inglés. Sus ampliosconocimientos sobre los reptilesfosilizados mesozoicos no erasuperados por nadie.
De entre los saurios de la épocamesozoica, Owen distinguió tres

especies que eran algo diferentes alresto. Anunció que el Iguanodon, el
egalosaurus y el Hylaeosaurus (o«Lagarto de Wealden») de Mantell, u«lagarto fosilizado» enorme cubierto deun caparazón de placas escamosas,encontrado por Mantell en el bosque deTilgate en 1832, fueran agrupados por scaracterística común de poseer cincovértebras fundidas en el cinturó pélvico. A diferencia de otros sauriosgigantes, éstos eran animales terrestres
en lugar de acuáticos y sus enormesmiembros y sus cortos dedos del pie les proporcionaban un parecido más quesuperficial con los mamíferos paquidermos pesados[9]. Estas

características, unidas al enorme tamañode las bestias, le «ofrecían suficientesmotivos para establecer una tribu osuborden distinto de reptiles saurios — proclamó Owen—, para los cuales propongo el nombre de dinosaurios[10]».Dinosaurio significa literalmente«lagarto terrible», nombre evocador otorgado en reconocimiento de lasdimensiones de la criatura y de sindudable ferocidad. Pero Owen teníarazones adicionales, más importantesque las meramente necesarias para elestudio formal de los restosfragmentarios, para investigar a estosreptiles gigantes.
Desafortunadamente, la historia sólo

honra a aquellos que luchan al lado delos ganadores; Richard Owen fue partidario de la causa antidarwiniana ela discusión evolucionista, lo cualhistóricamente nunca le fue perdonado.Owen se hallaba en uno de los puntoscruciales de la historia intelectual. Eraun anatomista práctico brillante, peroaun así era hijo del orden establecido yno fue capaz de aceptar los cambios quese avecinarían. Owen entró en un mundoen el cual el «progresismo» de Cuvier era científicamente respetable: laevidencia de que se habían extinguidorazas enteras de animales en distintosintervalos del tiempo parecía exigir laexistencia de catástrofes periódicas. Por

otro lado, en esa época, la doctrina de laevolución era una herejía romántica,objetivamente insostenible, que carecíade cualquier mecanismo riguroso. Owedejó un mundo, en el cual el hecho de laevolución era aceptado universalmente,en el cual las pruebas habían comenzadoa desviarse a favor de ésta y donde elcatastrofismo era considerado unaquimera de los creacionistas. Nunca fuecapaz de adaptarse a estos cambios.Cuando, después de la publicación delOrigen de las especies de Darwin e1859, la opinión cambió, Owen asumióel papel de principal portavoz de lafracción antidarwinista, basándose en s prestigio más que en argumentos

razonados para ganar el caso.Sufrió humillantes derrotas una y
otra vez, lo cual provocó quedespreciara a los evolucionistas, entrelos cuales el joven e impetuosoT. H. Huxley era el más estruendoso.Owen —el Cuvier inglés— se llevó scausa a la tumba. La historianaturalmente ha sido injusta. En la épocade juventud de Owen realmente existían
razones para creer que la evolución,especialmente tal como la exponían susexponentes predarwinistas, no podíarecibir apoyo alguno en base a losestudios de los fósiles. Los dinosauriosse convertirían en un eslabón crucial ela línea de razonamiento de Owen.

Owen se mostraba escépticorespecto al método de Buckland para elcálculo de las dimensiones de lossaurios usando los lagartos comoestándar. Los parecidos entre el guanodon y la iguana, dijo con aire deautoridad, «han llevado a los paleontólogos que hasta ahora ha publicado los resultados de sus cálculosdel tamaño del Iguanodon aconclusiones erróneas; y por lo tanto, lasdimensiones de 30 metros de longitudobtenidas en base a la comparación delos dientes y de la clavícula del guanodon con los de la Iguana, de 22metros en base a una comparaciósimilar de sus fémures y de 24 metros e

base a los huesos de los dedos, tomandocomo modelo el espécimen más grandede Horsham en lugar del utilizado por eldoctor Mantell para su comparación,obtendríamos un resultado superior a los60 metros de longitud total para el guanodon[11]». Estas dimensiones erasimplemente absurdas. Debido a queeste procedimiento era obviamentedudoso, Owen propuso un nuevométodo. Midió las vértebras y estimó snúmero desde la cabeza hasta la cola,llegando de esta forma a una longitudmucho más razonable de 8,5 metros parael Iguanodon y de 9,1 metros para el
egalosaurus. Estos antiguos sauriosobviamente no eran tan delgados como

los lagartos. Owen pensaba que, adiferencia de los ágiles y ligeroslagartos, el Megalosaurus era unacriatura maciza, compacta y pesada, coextremidades largas que soportaban uvoluminoso cuerpo.
De la forma y el tamaño delas costillas (continuó Owen) sededuce que su tronco era másancho y grueso en sus proporciones que el de lossaurios modernos y, sin duda, sealzaba del suelo apoyado enextremidades proporcionalmentemás anchas y en especial máslargas, de modo que el aspecto

general del Megalosaurus envida debía de parecerse proporcionalmente a los grandesmamíferos cuadrúpedos queahora pueblan la Tierra, y quedebieron de ocupar en la eraolítica el lugar que hastaentonces tenían los grandesreptiles del extinguido orden delos dinosaurios[12].
En 1841, cuando finalmente losdinosaurios fueron reconocidos por Owen, los otros grupos importantes desaurios extinguidos ya eran conocidosdesde hacía algunas décadas. Muchos

tenían un tamaño de dimensionescolosales similares, lo cual en gramedida hizo disminuir el impacto de losdinosaurios. Mary Anning y ThomasHawkins habían rescatado esqueletosenteros de ictiosaurios («lagartos- peces», reptiles marinos aerodinámicos,con forma parecida a la de los delfines, pero que no presentaban el cerebroagrandado de los delfines) y plesiosaurios (literalmente «casilagartos»), reptiles nadadores que parasu desplazamiento usaban aletas, en losesquistos azules de los acantilados deLyme Regis del jurásico inferior en lacosta de Dorset, y Gideon Mantell habíaexcavado los huesos de cocodrilos

gigantes y mosasaurios en Sussex. Así, a pesar de que los dinosaurios resultabaenormes en proporción con los lagartos,en opinión de Owen eran, en realidad,apenas un poco más largos que los otrosleviatanes de la Tierra arcaica. Owecalculó que los dinosaurios eran apenasmás grandes que los elefantes y losrinocerontes, con los que probablementetendrían un cierto parecido físico. Eran,literalmente, lagartos elefantinos.
Si, consiguientemente, el tamaño eraun criterio menor (aunquefrecuentemente se refirió a éste con elfin de proporcionar a este grupo ciertogrado de impacto dramático), ¿por quécreó Owen los dinosaurios? ¿Se trataba

simplemente de que quería dar sreconocimiento a algunos caracteres maldefinidos o existía algún otro motivo?
La comparación que hizo Owen deestos reptiles gigantes con los elefantes rinocerontes proporciona un indicio
decisivo para sus motivaciones. Owecreía que los dinosaurios eran losreptiles más evolucionados que jamáshabían existido. «Los megalosaurios eiguanodones —exclamó, regocijándosede sus innegables modificaciones casi perfectas de tipo reptil—, alcanzaron smáximo volumen y seguramentedesempeñaron los papeles mássobresalientes en sus respectivoscaracteres como devoradores de

animales y herbívoros que jamás haya presenciado la Tierra, en criaturasovíparas (que ponen huevos) y de sangrefría[13]». Estos animales fueron lacumbre de la creación reptil, metáforaque caracterizaba las opinionesimperantes de Owen con extraordinariaexactitud. Sus investigaciones sobre losreptiles fósiles habían proporcionado,finalmente, la prueba que haríainsostenibles las especulacionesevolucionistas de Jean-BaptisteLamarck.
Lamarck había sido el colega deCuvier en el Museo de París, aunqueestos dos eminentes hombres siempremantuvieron opiniones diametralmente

opuestas sobre la historia de la vida.Mientras que Cuvier concebía la idea derazas enteras de animales que habíasido aniquiladas por misteriosascatástrofes y de la posterior aparicióde faunas completamente nuevas parasustituirlas, Lamarck creía en lasucesión gradual de la vida queavanzaba a través de cambiosinfinitamente pequeños. (La extinción, yno digamos las aniquilaciones en masa,no tenían espacio en el esquema de lascosas de Lamarck). Las teoríasevolucionistas de Lamarck habían sidoinspiradas por el aparente incremento dela complejidad de los animales en sascenso en la escala de la Naturaleza.

«Por lo tanto, se hizo importante saber —afirmó Lamarck en su Zoological
hilosophy en 1809, explicando cómollegó a sus teorías sobre la evolución—,cómo esta organización (de una criaturasimple), a través de algún tipo decambio, había dado lugar a otras menossimples, y de hecho, a la gradualmentecreciente complejidad observada a lolargo de toda la escala animal[14]».Postulaba que existía una tendenciainnata a la complejidad, de forma que ecada generación se debía producir uincremento infinitamente pequeño;cuanto más tiempo pasaba, máscomplejos eran los animales que sedesarrollaban a partir de otros más

simples. Esta tendencia intrínseca erafomentada por el mayor uso que hacía elanimal de determinadas estructuras; esteuso desarrollaba la estructura ecuestión (como, por ejemplo, el cuellode la jirafa) y provocaba su crecimiento.Lamarck estaba pisando terreno peligroso y Owen emprendió la lucha para socavar el conjunto de esta teoría.
Como paleontólogo, Owen vio el punto débil en los argumentos deLamarck: si podía demostrar que elincremento de la complejidad era unailusión, todo el edificio se derrumbaría.El aumento de la complejidad era u punto en el cual insistían la mayoría delos primeros evolucionistas que

pensaban que era crucial para su teoría.«Debido a que al ascender en la escalaanimal —insistió Lamarck—,comenzando por los animales másimperfectos, la organización incrementagradualmente su complejidad de formaextremadamente notable». Éste fue el punto débil escogido por Owen para elataque. Los primeros evolucionistas,antes de la aparición del Origen de las
especies en 1859 de Darwin, al exigir una tendencia de las especies alascenso, desde las formas más bajas alas más altas, forzosamente tuvieron queencontrarse con la resistencia del lobby paleontológico. Owen preguntó si lasespeculaciones de los evolucionistas

habían obtenido algún punto de apoyo «a partir de los hechos ya determinados eel departamento de reptiles de la paleontología[15]». Owen arguyó que silas actuales especies de animales habíasido el resultado del progresivodesarrollo de especies más antiguas,«cada categoría debería presentar ahorasus caracteres típicos en las condicionesmás organizadas de la organización». Élexamen superficial de los fósilesregistrados demostraba que éste no erael caso. Los lagartos y cocodrilosactuales eran, en opinión de Owen,creaciones inferiores que difícilmente podían compartir la magnificencia delos dinosaurios mesozoicos. Éstos

ocupaban un lugar más bajo en la escalaen una época mucho más reciente, hechoque apenas era compatible con elascenso lineal de las especies. Lasuperioridad de los dinosaurios quevivieron en una gloriosa «Era de losreptiles», fue un acto directo de laCreación Divina. Las especies no setransmutaban de unas a otras sino queeran colocadas sobre la Tierra por designio divino y, si parecía queformaban una sucesión, ello se debía ala planificación divina más que a laevolución.
La evidencia… no permiteotra conclusión que pensar que

las diferentes especies dereptiles fueron introducidasrepentinamente en la superficiede la Tierra, aunque con unacierta regularidad sistemática enel orden de su aparición[16].
A pesar de la regularidad, según lacual la era de los peces dio paso a la erade los reptiles y ésta finalmente a la delos mamíferos, no existe una ascensióinfalible sino un plan de organizacióestática. Los reptiles no terminan hoy edía en sus representantes másevolucionados, es decir, «en el orden delos dinosaurios, de los cuales sabemos

que el tipo de estructura reptil seaproximó al máximo a la de losmamíferos». Por lo tanto, el motivo deOwen para aislar a los dinosaurios fueel de expresar su inherente superioridadentre los reptiles. Al proceder de estaforma, también pudo disponer de uinstrumento para atacar a losevolucionistas. Si hubiese habido algútipo de ascenso, tal como exigían losevolucionistas predarwinistas, losdinosaurios paquidermos —la apoteosisdel orden reptil de sangre fría— estaríavivos actualmente. En lugar de ellovivieron en la primitiva Tierra en elmomento que el Creador consideróidóneo para ellos.

La razón, según Owen, por la cual elCreador escogió el período medio de lahistoria de la Tierra, época en la cual pobló abundantemente el globoterráqueo con reptiles gigantes de todotipo, radicó en que la atmósfera eradensa y con escaso oxígeno. Los reptilesson criaturas de sangre fría; necesitan elcalor del Sol para calentarse como preludio a la actividad. Scomportamiento está destinado aabsorber la energía solar en lugar degenerarla internamente mediante laactividad muscular, como los mamíferoso las aves. Por consiguiente, umamífero puede permanecer activo,independientemente de la temperatura

ambiental, pero necesita un consumomucho mayor de oxígeno para liberar laenergía a fin de mantener este período prolongado de actividad. El corazón ylos pulmones menos eficientes de losreptiles los obligan a sobrevivir comenos energía. Owen pensó que elextraordinario predominio de criaturasgigantes de sangre fría en la eramesozoica sólo podía significar quehabía una menor cantidad de oxígenodisponible. Y, debido a la existencia dereptiles como los pterodáctilos quevolaban, probablemente la atmósfera eramucho más densa en aquella época, deforma que los animales seguramenteflotarían en lugar de mantenerse en el

aire mediante el movimiento de sus alas.(Owen dudaba seriamente si la bajafisiología de los pterodáctilos podríahaber soportado los enérgicos aletazos).
El mundo mesozoico diseñado por laProvidencia se adecuabaadmirablemente a los reptiles de sangrefría que lo habitaban, pero eratotalmente inadecuado para las criaturas pertenecientes a una organizaciósuperior. Sólo cuando el oxígeno delaire aumentó y la presión descendió pudo producirse «la bella adaptación dela estructura de las aves a un medio quede esta forma se volvió más ligero yvigorizante». El descubrimiento dehuesos, aparentemente de aves, en los

estratos de Wealden del cretáceo (erealidad los restos de pterosaurios),indica el inicio del proceso de«vigorización» en aquella época. Scontinuación dejó al mundo inhabitable para los inmensos saurios que murieroen masa al final del cretáceo. Pero elos Wealden también se encontraron el guanodon y Hylaeosaurus de formaque, según Owen, difícilmentesorprendería que representaran lamáxima aproximación de un reptil a losanimales de sangre caliente. Owesupuso que, al igual que los cocodrilos,los dinosaurios poseían una buenacirculación para hacer frente al climamás vigorizante en el que se

encontraban. Necesitaban un buecorazón de cuatro cámaras, con unacompleta división entre la sangre venosaconsumida y la sangre arterial rica eoxígeno. En los lagartos inferiores, lasangre venosa se mezcla con la sangrearterial en el corazón. Comoconsecuencia de ello, los tejidos recibesangre sólo parcialmente renovada ytienen que realizar sus actividades comucho menos oxígeno.
Se podía concluir que losdinosaurios, que tenían la mismaestructura torácica que loscocodrilos, debieron de tener uncorazón de cuatro cámaras; y que

debido a su superior adaptacióna la vida terrestre habíandisfrutado de la función de unórgano central de la circulaciónaltamente organizado, en ungrado bastante aproximado alque ahora caracteriza a losvertebrados de sangrecaliente[17].
Así fue como Richard Oweconcluyó su informe —el primer informe — sobre los dinosaurios. Estasafirmaciones eran audaces conjeturas desu parte. Fue quizás el prestigio ydominio indiscutido de Owen en este

tema lo que hizo que pudiera sacar adelante estas ideas. «Un observador más precavido, tal vez se hubieraretraído ante tales especulaciones»reconoció Owen. Sin embargo, elrazonamiento de Owen no resiste deningún modo un examen. ¿Por qué uclima más «vigorizante» debería haber vuelto el mundo inhabitable para losdinosaurios? ¿Por qué sobrevivieron loslagartos? Owen evidentemente pensóque era necesario llegar a estosextremos para contrarrestar las teoríasde los evolucionistas. Debido a queéstos exigían un ascenso ininterrumpido,Owen los combatía en base a las arenasmovedizas de absolutos cualitativos

como «superior» e «inferior». Comoconsecuencia de ello, sus argumentoseran igual de frágiles que los de losevolucionistas, presentando manifiestasincongruencias.
¿Cómo debemos valorar lasopiniones de Owen? No debemosconfundirnos pensando que, de algunaforma, anticipó ideas modernas, basadasen estudios anatómicos y ecológicosdetallados. Con el fin de combatir lasespeculaciones heréticas de losevolucionista, se había visto impulsadoa crear un orden totalmente nuevo dereptiles a partir de sólo tres especiesconocidas y, posteriormente,reconocerlas como la cumbre de la

creación reptil. Los dinosaurios pareceque no fueron creados por Dios sino más bien por la ingenuidad humana. Oweliteralmente pobló su tierra mesozoicacon dinosaurios como estrategia contralos evolucionistas. Su sucesión deteorías fue formulada como reacción.
ecesitaba una raza de superreptiles deforma que creó una.
Aun a pesar de que los dinosaurioscompartían determinadas característicascon los mamíferos, Owen ciertamentenunca se apartó de su teoría de que estos«gigantescos lagartos-cocodrilo» noeran, en realidad, otra cosa que animalesde sangre fría. Fue este modelo dedinosaurio —un lagarto gigante—, más

que sus fantásticas especulaciones (querápidamente fueron olvidadas cuandolos evolucionistas dejaron de aferrarse ala simplista idea de la progresión linealascendente de la vida), el que tuvo unainfluencia duradera sobre los paleontólogos. El dinosaurio eradescrito como un lagarto monstruoso, parecido a un rinoceronte escamoso.
El legado de Owen quedóestablecido en términos más concretoscuando, trece años más tarde, tuvo laoportunidad de construir modelos etamaño natural de sus dinosaurios. Laocasión se presentó cuando el edificiocentral que albergaba a la GraExposición de 1851, el Crystal Palace

de Joseph Paxton, la edificacióacristalada con estructura metálica másgrande jamás construida, fue trasladadade Hyde Park a un nuevo lugar en lossuburbios de Sydenham. La exposicióde Hyde Park se había hecho paracelebrar los frutos de laindustrialización y fue diseñada comouna feria de comercio internacional paraexhibir las obras tecnológicas. Cuandoen 1852 fue cerrada, el palacio fuedesmantelado pieza por pieza y vuelto aedificar en Sydenham donde debíaformar un centro de exposició permanente para las artes y las ciencias.
Parece ser que fue el consorte de lareina Victoria, el príncipe Alberto,

quien sugirió por primera vez que la planta baja del nuevo parque podría ser adornado con las bestias restauradas deépocas pasadas. La tarea dereconstrucción fue encomendada a u pintor y escultor de la vida salvaje,Benjamin Waterhouse Hawkins, quehabía actuado en función de supervisor en la Gran Exposición. Hawkins recibiócarta blanca y escogió construir grandes mamíferos como el mastodonte.Posteriormente dio con las monografíasde Owen sobre los reptiles gigantes delmesozoico y decidió devolverle a estascriaturas su antigua gloria. Para la tareahercúlea de «revivir el mundo antiguo»,Hawkins trabajó en estrecha

colaboración con Owen desde la planificación hasta el mismomodelado[18]. A partir de modelos aescala en miniatura, de sólo unos pocoscentímetros de longitud, Hawkinsconstruía moldes de arcilla de tamañonatural, de los cuales el más grande pesaba treinta toneladas y, a partir deéstos, obtenía sus figuras. Seconstruyeron tres islas para albergar alos monstruos en un lago de 242 áreas,creado artificialmente en la planta bajadel palacio. Una vegetación real proporcionaba a las islas un marcotropical exuberante, escenario que seveía realzado por la construcción deréplicas de un tipo de helechos del

cretáceo, las cicádidas en forma de palmeras, con troncos que presentabaun dibujo en forma de hojas que lesdaban el aspecto de piñas petrificadas.Con el objetivo de aumentar la ilusióde una playa tropical (se suponía que lascriaturas habían sido habitantes de lacosta), se dispuso que el nivel de aguadel lago subiera y bajara unos cuantoscentímetros, como en las mareasnaturales, quedando las criaturas parcialmente sumergidas a intervalos[19].Sigue siendo un enigma cómo se pudolograr todo esto en un lago de 242 áreas.
Las criaturas representaban la vidareptil específica de Inglaterra en laépoca mesozoica, aunque tambié

existían algunos mamíferos terciarios eel escenario como el alce gigante.Hawkins describió su vocación comouna convocatoria «desde los abismosdel tiempo y desde las profundidades dela Tierra, de las vastas formas ygigantescas bestias diseñadasconvenientemente por el TodopoderosoCreador para poblar y precedemos eesta parte de la Tierra llamada GraBretaña». Los reptiles de las tres islasforman una secuencia, estando lascriaturas arcaicas de la era triásicasituadas en un extremo y los dinosauriosde la cumbre en el extremo cretáceo másreciente. Debido a que Hawkins sólodisponía de los cráneos de las bestias

del triásico, se vio obligado a recurrir acierto grado de ingenuidad para añadir los cuerpos. Sólo disponía de losreptiles modernos para hacer de guía deforma que, naturalmente, éstos fuerosus modelos. El Dicynodon, o «animalde dos colmillos», era un primo deaquellos reptiles que evolucionarohacia los mamíferos. Debido a que scráneo tenía un hocico escamoso, parecido al de las tortugas, Hawkins lorestauró en forma de tortuga enorme cocuerno. El Labyrinthodon, que recibíasu nombre por los pliegues laberínticosdel esmalte de sus dientes, era uanfibio primitivo acorazado, de aspectono muy diferente al de un cocodrilo

vivo. Hawkins restauró tres de estosanimales dándoles el aspecto de ranas ysapos gigantes con colas cortas.
Los dinosaurios estabarepresentados por las tres especies deOwen (éstas eran los únicos dinosauriosreconocidos oficialmente en 1854), el guanodon e Hylaeosaurus cretáceos yel Megalosaurus jurásico más antiguo.Los animales andaban a cuatro patas,tenían el aspecto de rinocerontes(parecido que aún asombraba más por elcuerno que el Iguanodon llevaba en elhocico, y que en realidad eran los restosde un dedo). Owen había derrotadofinalmente a los evolucionistas: estosdinosaurios eran los homólogos

concretos de los mamíferos rinocerinos difícilmente podían pertenecer al
mismo grupo como insignificanteslagartos. El mosasaurio fue restauradocomo si estuviese escondido en el agua,sólo la cabeza sobresalía del agua. Dehecho, sólo se esculpió la cabeza ya queel cuerpo aún era desconocido a pesar de que Mantell había encontrado algunosfragmentos en la creta inglesa. Dos pterodáctilos gigantes contemplabamajestuosamente toda la escena desdeun punto sobresaliente de un acantilado,mientras que en las orillas del lago loscocodrilos marinos de hocico largo,ictiosaurios y plesiosaurios tomaban elsol[20].

Una vez finalizado el proyecto,Hawkins y Owen lo celebraron dandouna cena en el interior del Iguanodon
en la víspera de año nuevo en 1853.Veintiún dignatarios ocuparon elestómago del reptil cargado socialmente el profesor Owen se sentó en la cabeza
(literalmente). El grupo festejó demanera suntuosa y el jolgorio continuóhasta entrada la noche con una sucesióde brindis de felicitación. Cuando, a primeras horas de la mañana, el banquete terminó, se pudo ver cómo elconjunto de eminentes científicos salíadel dinosaurio y cruzaban el parquealabando ruidosamente los méritos delas restauraciones de Hawkins[21]. El

incrédulo Quaterly Review informó:«Saurios y pterodáctilos, ¿pudisteissoñar alguna vez, en vuestras antiguasfiestas, que una raza por venir que seaposentaría sobre vuestras tumbas…cenaría en honor de vuestros fantasmas,evocados de las profundidades por sus brujos[22]?».
La reina Victoria y el príncipeAlberto volvieron a abrir el palacio el10 de junio de 1854 en presencia decuarenta mil espectadores. El consensode la opinión pública era claramente quese trataba de monstruos antediluvianos, bestias que habían sido destruidas por elDiluvio Universal, aun a pesar de queesta teoría había estado fuera de boga e

los círculos científicos durante muchosaños. Incluso las revistas literarias, quedeliberaron sobre este «mausoleo de lamemoria de los mundos perdidos», talcomo describía el Quaterly Review lacolección, discutieron los diversosméritos de los logros pasados delCreador. Los espectadores acudían etropel al palacio para contemplar losmonstruos que poblaron la Tierra antesde Noé, dejando tras de sí una estela dedevastación en su búsqueda derecuerdos.
El edificio del Crystal Palace fuecompletamente destruido por uincendio en 1936, lo cual probablementefue la causa de que los dinosaurios

cayeran en el olvido. Hoy en día, losreptiles se pueden seguir contemplandoen sus islas prehistóricas, siendo eltestimonio de la concepción victorianade los dinosaurios, concepción quedesde entonces ha dominado tanto lasideas del público como las de loscientíficos. Charles Dickens las resumióen los primeros párrafos de Bleak
ouse, escritos en el año en el cualHawkins comenzó las reconstruccionesdel palacio. «Implacable tiempo denoviembre —escribió Dickens—. Tanto barro en las calles como si las aguasacabaran de retirarse momentos antes dela faz de la Tierra, ¿no hubiera sidomaravilloso dar con un Megalosaurus,

de doce metros de largo, más o menos,marchando pesadamente, como uelefante, en dirección a la colina deHolborn[23]?».
De esta forma, los Victorianos noslegaron el dinosaurio como un lagartomonstruoso: una criatura con todas lasconnotaciones que se suponen en los«reptiles». Las restauraciones delCrystal Palace han sobrevivido hasta elsiglo XX como singular pieza victoriana,monumentos a una creencia de que losdinosaurios eran, según palabras deDickens, «lagartos elefantinos». Esterecuerdo visual, se ha visto acompañado

de la idea de que los dinosaurios eraesencialmente criaturas de sangre fría.Cuvier, al reconocer correctamente al
osasaurus como lagarto,inconscientemente incluyó a losdinosaurios contemporáneos en umismo grupo. Owen podría haber emancipado a los dinosauriosexcluyéndolos de la categoría de loslagartos, pero fue incapaz (si es quealguna vez realmente quiso) de descartar que los dinosaurios tuviesen unafisiología similar a la de los lagartos.Durante más de un siglo, esta visión deldinosaurio permaneció incuestionable.La situación, tal como aún se presentabaen 1968, fue descrita por Robert T.

Bakker, que por aquel entonces estabaen la Universidad de Yale:
Generalmente, los paleontólogos aceptaron que, entodos los detalles de su vida, losdinosaurios eran simplementecocodrilos, lagartos, o caimanesque habían crecido más de lonormal. Los cocodrilos y loslagártidos pasan gran parte de sutiempo inactivos, tomando el solechados en la roca o el troncomás conveniente, y comparadoscon otros animales modernos, lamayor parte de los reptilesmodernos son lentos y

perezosos. De aquí viene que lareconstrucción de un dinosauriocomo el Brontosaurus, nos lo presente como una montaña decarne fláccida que se mueve a sualrededor sólo de forma lenta y poco frecuente[24].
Sin embargo, existen profundasdiferencias entre los dinosaurios y loslagartos, diferencias que nos han llevadoa pensar que nuestro modelo de losdinosaurios no era del todo correcto.Incluso los «lagartos elefantinos» desangre fría, por la naturaleza de stamaño, serían bestias radicalmente

diferentes a los pequeños lagartosexistentes en la actualidad.
Hemos estado hablando sobre loslagartos, refiriéndonos a ellos como aanimales de «sangre fría». Sin embargo,éste es un término inapropiado ya que ssangre no es, en absoluto, fría. Lascaracterísticas que distinguen a losreptiles de los mamíferos y de las avesson que los primeros no dependen delcalor producido interiormente. Losmúsculos y órganos corporales de lascriaturas de «sangre caliente» produceel suficiente calor como para mantener el cuerpo a una temperatura alta, estable, proceso que es controlado por elhipotálamo en el cerebro. Los lagartos,

sin embargo, dependen del calor del Sol del aire circundante para calentar sus
cuerpos. Por esta razón es más correctollamarlos ectotermos: dependen defuentes de calor externas para elevar stemperatura corporal en contraste colas aves y los mamíferos endotérmicos.
Antes de que los lagartos fueraestudiados en su medio ambiente natural,se pensaba que su temperatura variabadirectamente con la del aire. A finalesdel siglo, los fisiólogos quemonitorizaban los lagartos bajocondiciones de laboratorio, naturalmenteencontraron que esto en realidad eracierto. Sin embargo, los lagartos queviven en plena Naturaleza son capaces

de elevar su temperatura por encima dela del aire y luego mantenerla dentro deunos estrictos límites, a pesar de las pequeñas fluctuaciones que se pueda producir en la temperatura ambiental.Los grandes lagartos sin orejas delsudoeste de Estados Unidos, por ejemplo, pueden mantener unatemperatura media que va de los 17 alos 39o durante el 75% de su vidaactiva[25]. La mayoría de lagartos, dehecho, poseen una temperatura alta,óptima, que se mantiene estrictamentedurante el tiempo que están despiertos.
¿Por qué estaban tan equivocadas lasobservaciones del laboratorio?«Seguramente por la misma razón por la

cual un hombre que tiene encadenadauna pierna a una pesada bola de hierrono puede demostrar lo rápido que puedecorrer», reflexionó Charles M. Bogertdel Museo Americano de Historia
atural[26]. El lagarto es capaz demantener una elevada temperaturainterna mediante respuestas delcomportamiento que se orientan por elsol; confinarlo a una jaula solamentesirve para inhibir su margen deactividades. A la mayoría de loslagartos les gusta «tomar el sol» yexplotar al máximo los rayos solares,adoptando una serie de posturas yorientaciones apropiadas. De estaforma, el lagarto puede variar la

absorción de calor con bastante precisión. Si, por ejemplo, stemperatura corporal desciende por debajo del punto óptimo, el animal secolocará en ángulo recto respecto a losrayos solares para maximizar laexposición. De forma similar, a primeras horas de la mañana o últimashoras de la tarde, cuando el sol está bajo, el lagarto se pegará a lassuperficies y pendientes rocosas a fin deque su cuerpo reciba los rayos solares perpendicularmente a la longitud de scuerpo, para recibir la máxima energía posible. Cuando el lagarto alcanzatemperaturas demasiado altas mientras busca alimento durante el día, se da la

vuelta y se pone de cara al sol paraminimizar el área de exposición.
De esta manera, algunos lagartos socapaces de mantener una temperatura taelevada como la del hombre. De hecho,algunos pueden estabilizarse etemperaturas mucho más altas: el lagartoamericano de cola de látigo mantiene stemperatura corporal entre los 42 oC ylos 45 oC. Se requiere una elevadatemperatura interna para elfuncionamiento óptimo de los procesos bioquímicos del cuerpo; cuando latemperatura desciende, las reaccionesquímicas se suceden con más lentitud ylos reptiles se vuelven perezosos. Cadaespecie tiene su propia temperatura

óptima, de forma que dos especies quevivan juntas en la Naturaleza (lagartosque estén a una misma temperaturaambiental controlada de laboratorio), pueden presentar diferencias de hasta 12oC debido a que tienen diferentesrespuestas al sol.
Teóricamente, cuanto mayor sea lamasa del animal, tanto mayor será svolumen en relación al área desuperficie. Consiguientemente, tambiénecesitará un tiempo de exposición alsol más prolongado para calentarse.Esto, de todos modos, es el caso de losobjetos inanimados; pero éstos nuncaserán tan versátiles como los lagartos.Esta relación no es válida cuando se

trata de pequeños reptiles ectotérmicos, precisamente debido a sus maravillosasreacciones heliotrópicas. Los lagartosmás grandes, teóricamente deberíacalentarse y enfriarse más lentamenteque sus primos más pequeños, debido aque su área de superficie esrelativamente más pequeña a pesar de smayor masa. Pero, en realidad, existe pocas diferencias debido a que adoptatodo tipo de posturas para maximizar sexposición al sol, mientras que al mismotiempo oscurecen el color de su piel para aumentar su capacidad deabsorción.
Todo esto está muy bien cuando setrata de pequeñas criaturas, que

necesitan sólo un corto espacio detiempo para alcanzar su temperaturaóptima. Sin embargo, existe un límitesuperior de tamaño, más allá del cualestas respuestas del comportamiento ycambios de color ya no les permiten aestas criaturas competir con sus parientes más pequeños. A un caimán leresultaría completamente imposiblecalentarse con la misma rapidez que a u pequeño lagarto. En un intento deconocer un poco más acerca de losdinosaurios (tanto acerca del tamañocomo de la relación), Bogert unió susfuerzas a las del experto en dinosaurios,Edwin H. Colbert, también del MuseoAmericano de Historia Natural, y

Raymond B. Cowles de la Universidadde California y, en 1946, escogieron alcaimán como animal de ensayo. En laestación biológica de Archbold, cercade Lake Placid en Florida, el caimáamericano podía ser estudiado en smedio ambiente natural, constituido por palmeras, cicádidas y coniferas que posiblemente era bastante parecido almundo cretáceo de los dinosaurios. Elequipo trabajó con trece caimanes quemedían entre 30 cm y casi dos metros delongitud, y pesaban desde unos pocosgramos a 226 kg. Los animales eraatados a postes bajo el sol y seregistraban las temperaturas eintervalos de diez minutos. En los

experimentos iniciales —tal como seesperaba—, inmediatamente se hizo patente que, bajo la luz solar directa, elgrado de incremento de la temperaturade los caimanes variaba inversamente asu tamaño. El caimán más pequeño sólonecesitaba 90 segundos para incrementar su temperatura en 1 grado, mientras quelos ejemplares de 136 kg necesitaban utiempo cinco veces superior.«Prosiguiendo con esta línea derazonamiento —dijeron Colbert, Cowles Bogert—, podría parecer probable que
en un dinosaurio adulto de dieztoneladas de peso, es decir, un animalcon un peso corporal deaproximadamente 9 millones de gramos,

el aumento de la temperatura fueramucho más lento que en un caimágrande. En efecto, si al dinosaurio (uanimal con una masa corporal 700 vecessuperior a la de un caimán grande), sele aplicara la misma diferencia deincremento de temperatura existenteentre un caimán pequeño y uno grande,se podría estimar que, para incrementar en 1 oC la temperatura corporal de losgigantes adultos extinguidos se tardaríamás de 86 horas[27]». ¡En otras palabras,si la temperatura del dinosauriodisminuyera sólo un grado por debajo desu umbral inferior de actividad, elanimal tendría que tomar el sol durante
más de tres días para volver a recuperar

su temperatura normal! (Naturalmente,suponiendo que no hubiera espaciosnocturnos interpuestos). Esto es absurdo.Lo que realmente significa es que,
suponiendo que existía una temperaturamedioambiental alta y constante, si lainmensa masa del dinosaurio era capazde retener el calor durante la noche, éstecomenzaría el nuevo día con apenas unadisminución de sus reservas caloríficas.Si los cálculos de Colbert eracorrectos, los dinosaurios podrían haber mantenido su temperatura corporalabsolutamente estable (dentro de umargen de variación de unos pocosgrados para la temperatura óptima)durante toda su vida adulta. Es decir,

suponiendo que realmente hubieravivido en regiones con temperaturasaltas estables. Aunque eran animales de«sangre fría», nunca se enfriaban. Estetrabajo elemental sobre los caimanesestableció que la naturaleza elefantinade los «lagartos» les otorgaba un patróde temperatura similar al de losmamíferos o de las aves, aunque se pensaba que su fisiología y metabolismoeran totalmente distintos.
Los colaboradores en el temahicieron numerosas objeciones a estecálculo excesivo de 86 horas para uincremento de la temperatura de ugrado, de forma que, al año siguiente,Colbert, Cowles y Bogert ya había

reducido sus estimaciones a «variashoras» arguyendo que, a menos que seutilizaran cocodrilos mucho más grandescomo animales de ensayo, nunca seconocería el grado exacto decalentamiento y enfriamiento de udinosaurio[28]. En la época en la queColbert escribió su libro Dinosaurs, e1962, había reducido las estimaciones ados o tres horas para el incremento de 1grado celsius en un Brontosaurus detreinta toneladas de peso[29]. Bogertredujo esta cifra aún más: sostuvo queun dinosaurio de diez toneladas de pesonecesitaría tomar el sol abrasador durante poco más de una hora paraelevar en dos grados su temperatura

corporal[30]. La reducción de lasestimaciones cambió drásticamente lasituación. Si aceptamos la cifrarevisada, el dinosaurio exotérmico
odría haber perdido una notablecantidad de calor durante la noche, talcomo ya previo Bogert:
Imaginemos que nuestrohipotético dinosaurio fueraactivo durante el día y sometidoa un enfriamiento durante lanoche, tal como sucedería encualquier región desierta ennuestros días. Si la temperaturadescendiera cuatro o cinco

grados por debajo de su umbralde actividad, el dinosaurionecesitaría tomar el sol durantegran parte del día siguiente a finde volver a recuperar latemperatura umbral deactividad[31].
Tomar el sol durante una serieininterrumpida de horas habría tenidograves consecuencias para losdinosaurios. El estudio de Colbert sobrelos caimanes previo estas consecuenciasde forma bastante dramática. Colbertmidió la temperatura interna de suscaimanes, pero cuando ésta se aproximó

al grado óptimo, dos de sus animalesmurieron repentinamente. La razón deello era que el sol estaba abrasando la parte externa de los cuerpos de losanimales y el calor debía ser llevado alos tejidos y órganos a través de laconducción y transporte mediante lasangre. Debido a esto se había creado ugradiente, de forma que, cuando latemperatura había alcanzado sólo 38 oC,internamente la superficie escamosa sehabía calentado tanto que los animales perecieron. Colbert había extrapolado eltiempo requerido de calentamiento delos dinosaurios a partir de los valoresdeterminados en los caimanes, pero sehabía descuidado de extrapolar el

gradiente de calor existente entre lasuperficie interna y externa. Losdinosaurios gigantes se caracterizaba por tener una inmensa masa pero un áreade superficie relativamente pequeña, yla única forma de que el sol pudieracalentar esta masa interna de carne eracalentando mucho la superficie dorsalexpuesta al sol. Los dinosaurios, al igualque los lagartos o los mamíferos,funcionan con una temperatura óptima(cualquiera que haya sido ésta). Paraincrementar la temperatura en las zonasmás profundas del cuerpo, la pieltendría que haber sufrido un calor abrasador que habría idodesapareciendo hasta que internamente

se hubiera alcanzado el grado óptimo. Sien los caimanes del Mississippi deColbert, el gradiente era considerable,en los dinosaurios de cincuentatoneladas de peso, éste debía ser sencillamente enorme. Si la sangre nohabía sido capaz de disipar rápidamenteel calor de la superficie(transportándolo a otro lugar) en loscaimanes atados, seguramente tampocolo podría haber hecho en un dinosaurio.Por esta razón, parece muy improbableque un dinosaurio de cincuenta toneladas pudiera haber alcanzado una temperaturaóptima uniforme tomando el sol comolos lagartos.
Esto es simplificar un poco

excesivamente el problema. Pero ilustracómo este estudio, practicado amediados de la década de 1940, hizosurgir muchas más preguntas de las que podía contestar. Los caimanes deColbert fueron atados a postes bajo elsol. Si hubiesen estado descansando aorillas de un río, se habrían vistoobligados a meterse en el agua o a lasombra mucho antes de alcanzar uestado crítico. De esta forma, udinosaurio ectotérmico no atado tendríaque haber estado todo el tiempocambiando de posición, alternando entreel sol y la sombra, intentando alcanzar un equilibrio entre sus temperaturasinternas y de superficie. Todo ello

suponiendo que no tenía que mantenersefirme bajo el sol abrasador durante todoel día, a fin de recuperar el calor perdido durante la noche. Debido a quese desconocen la cantidad de calor perdido durante la noche y el tiempo deexposición necesario para recuperarlo,es difícil saber si un dinosaurio desangre fría podría haber funcionado. Lasconsiderables divergencias de opinióacerca de cuánto tiempo de exposicióal sol era necesario para incrementar latemperatura del animal en un solo grado,ilustran el carácter especulativo de todoeste ejercicio. Faltaban datos precisosde forma que, en la década de 1940, fue posible concluir que estos leviatanes

reptiles eran ectotérmicos (lo cual nuncase cuestionó), pero que habían vivido eclimas tropicales donde nunca se producían descensos importantes de latemperatura durante la noche. Debido aque los dinosaurios se desplazaron haciaregiones situadas más al norte y alejadasde los actuales trópicos, se dio por sentado que, en la era mesozoica, laTierra era uniformemente cálida en unagran parte de su superficie. Noconstante, resulta difícil imaginarse auna criatura de sangre fría, cuyo tonelajesuperaba al de una gran familia deelefantes, manteniendo una temperaturaalta estable tomando baños de sol: lascontradicciones inherentes a u

ectotermo de cincuenta toneladastomando el sol parecen insuperables.Las preguntas surgidas a raíz del estudiode Colbert nunca fueron contestadas; lavasta cantidad de factores desconocidoshizo que el problema fuera insoluble.Cuando éste fue abordado de nuevo dosdécadas más tarde, se hizo desde u punto de vista completamente diferente ylos resultados de este nuevo enfoquemostraron por qué se habían encontradotantas dificultades utilizando al lagartocomo sustituto del dinosaurio.

II. EL TIRANOENCUENTRA SUS
PIES
Lejos del emplazamiento urbano delas festividades del Crystal Palace del príncipe Alberto, el Nuevo Mundoestaba presenciando los primerosdescubrimientos de dinosaurios. Seríaaquí donde el dinosaurio emergería deuna curiosa manera totalmenteinimaginable para Owen. En 1854, unaexpedición exploradora de losingenieros topográficos de EstadosUnidos se dirigió hacia el Oeste para

alcanzar el río Judith cerca de su uniócon el poderoso Missouri en lo que eaquel entonces era territorio de
ebraska (ahora Montana). Tratándosede una expedición de reconocimientocientífico oficial, el gobierno habíaenviado a Ferdinand Vandiveer Haydecomo geólogo del equipo, con elencargo de cartografiar la región yrecoger muestras. Hayden, que sólocontaba 26 años de edad, era uveterano de una anterior expedición e busca de fósiles cuando emprendió elcamino hacia el oeste desde Nueva York a las tierras de baldío del Missourisuperior. Hayden estaba familiarizadocon el terreno de las tierras de baldío y

recorrió los afloramientos desérticos yaccidentados del Judith. El terrenoexpuesto y desgastado por el tiempo,recorrido por profundos barrancos ycañones y carente de vegetacióarbustiva, hizo fácil la recolección demuestras y Hayden volvió al Este con ualijo de grandes dientes encontrados elas arenas y arcillas cretáceas[1]. A principios de 1858 donó estos hallazgosgeológicos a la Academia de Ciencias
aturales de Filadelfia, cuyo director era Joseph Leidy, que al mismo tiempoera también profesor de anatomía de laUniversidad de Pennsylvania. Lareacción inmediata de Leidy fueanunciar que Europa no había sido la

única que había estado poblada dedinosaurios en el mesozoico, y que el primitivo paisaje americanoseguramente también había presenciadoel ascenso y la caída de estas criaturasgigantes. Algunas de las coronas nuevasde los dientes del Judith eran cónicas yligeramente curvadas, con un bordecortante característico de las criaturasherbívoras: por lo tanto, Leidy llamó aeste saurio herbívoro Trachodon o«diente rudo». Al igual que Mantellhabía descubierto que el megalosaurio,un «contemporáneo carnívoro», habíasido un depredador del Iguanodon, en elOeste de Norteamérica, en el cretáceo,había existido un enorme depredador

que cazaba el Trachodon. La expedicióde Hayden regresó con docenas decolmillos de este carnívoro. Los largosdientes, curvados y aserrados debíahaber pertenecido a un reptilespantosamente armado, al que Leidydio el sugestivo nombre de Deinodon
horridus o «el más horrible de losdientes terroríficos». Leidy observó queel patrón de desgaste de los dientes dabala impresión de que «el maxilar superior e inferior se cerraban como las hojas deunas tijeras, de forma que estaban bieadaptados para penetrar, despedazar ycortar los alimentos animales[2]».Mientras los apacibles herbívoros, el guanodon y el Trachodon, pastaba

tranquilamente a lo largo de las orillasde los cursos de agua de las regionesinteriores del cretáceo, el
egalosaurus y el Deinodon infestabalas aguas como los cocodrilos, preparados para cazar a suscontemporáneos. La lucha por lasupervivencia en el remoto pasado, tavigorosamente descrita por Leidy, coherbívoros gigantes tenidos a raya por carnívoros gigantes, era una escena tantomás notable cuanto se basabaúnicamente en unos cuantos dientesrotos. Sin embargo, alrededor de 1850,Leidy pudo inspirarse también en lasexperiencias de los coleccionistas defósiles ingleses. Al igual que e

Inglaterra, los dientes de los dinosaurioshabían aparecido antes que el esqueleto,dando una visión atormentada de la vidaen el lejano mesozoico. Eldescubrimiento de los restos delesqueleto de estos gigantes fue esperadocon gran expectación, pero, cuandofinalmente se produjo, proporcionó uduro golpe a las ideas convencionales.Cuando Leidy vio por primera vez elesqueleto de uno de sus dinosauriosamericanos, éste no coincidía niremotamente con la concepción deRichard Owen que ya se había hechomundialmente famoso por las efigies delCrystal Palace.
Leidy no tuvo que esperar la llegada

de la siguiente expedición al Oeste paraobtener su esqueleto. William Parker Foulke, un miembro de la Academia deCiencias Naturales de Filadelfia, ycolaborador habitual en temasgeológicos, había pasado el verano yotoño de 1858 en Haddonfield, NewJersey. Se enteró que un granjero vecinohabía desenterrado unos grandes huesosmientras cavaba marga (un tipo de piedra caliza lodosa) en su granja, hacíaunos veinte años aproximadamente.Estas vértebras habían llamado mucho laatención por su tamaño, y la gente se lashabía llevado como recuerdo. Debido aque no se habían desenterrado aún de lamarga el cráneo y los miembros, Foulke

se dispuso a encontrar la antigua canteraen las tierras del granjero. Haddonfieldse hallaba sobre roca cretácea, por locual Foulke tenía la esperanza deencontrar los restos de saurios, sobretodo debido a que en la vecindad sehabían encontrado fragmentos demosasaurio. Finalmente la cantera demarga fue descubierta de nuevo por unode los antiguos trabajadores en el fondode un barranco, y Foulke contrató aexcavadores experimentados paracomenzar de nuevo las operaciones. Unavez apartadas las capas superficiales detierra, encontraron una delgada capa deconchas de moluscos fosilizados ydemenuzados a una profundidad de 2,43

metros. Cuando excavaron a mayor profundidad, a unos 3 metros, dieron couna acumulación de huesos de saurio decolor negro ébano. Éstos resultaron ser extremadamente friables y Foulke prodigó muchos cuidados para sacarlosindividualmente, usando sólo una paleta un cuchillo. Cuando al extraer los
huesos alguno de ellos se rompía, seconfeccionaba un dibujo in situ delhueso, de forma que más tarde pudieraser recompuesto.
Se informó a Leidy, en la Academiade Ciencias Naturales, y ésteinmediatamente fue a visitar la escenade la agitación ayudando a extraer losúltimos huesos que quedaban. Tras ser

informado de este tesoro en el mismolugar del descubrimiento, Leidy visitótodas las canteras de marga de la zona buscando restos de saurios gigantes.¡Pero a pesar de que muchostrabajadores habían visto y tocado loshuesos, éstos siempre habían sidotirados y se habían perdido! Leidy semarchó con algunos fragmentos decocodrilos y mosasaurios pero nada que pudiera competir con el impresionanteesqueleto de Foulke. Foulke le presentóa la Academia de Ciencias Naturales loshuesos fósiles el 14 de diciembre de1858 y Leidy comenzó su descripcióformal. Los restos pertenecían a un graherbívoro emparentado con el

guanodon inglés y Leidy les dio elnombre de Hadrosaurus foulkii paraconmemorar al descubridor de lacriatura. Buscando en la marga, Foulkehabía recogido además nueve dientes eforma de escudo de, aproximadamente, 5centímetros de largo procedentes delmaxilar inferior, dientes que claramenterelacionaban al hadrosaurio con el guanodon y Trachodon. Comoresultado de los laboriosos cuidados deFoulke, los frágiles huesos fuerorecuperados de la roca sin sufrir dañoalguno. Aparte de las vértebras, Leidyestaba también en posesión de loshúmeros, radios y cúbitos de lasextremidades anteriores y de los

correspondientes huesos de lasextremidades posteriores. Debido a queéstos fueron hallados juntos, era seguroque provenían de un solo animal.
Para sorpresa de todos existía una peculiar desproporción entre lasextremidades anteriores y posteriores.El fémur estaba completo y medía 1metro y, en cambio, el húmero del brazosuperior medía sólo 58 centímetros.Leidy admitió abiertamente que si nohubiese encontrado juntos los huesos,habría compuesto dos animales bastantediferentes a partir de ellos: unoconocido solamente a partir de lasextremidades posteriores y el otro a partir de las extremidades anteriores.

Estimando que el número de vértebrashabía sido de 80 aproximadamente,Leidy supuso que la longitud total del
adrosaurus había sido de 7,62 metros.Debido a que los miembros posterioreselevaban el trasero del reptil unos 1,8metros por encima del suelo, mientrasque los miembros anteriores medíamenos de 1,2 metros de largo, el saurioen vida no podía haber tenido ningú parecido con el modelo de Iguanodon
expuesto en Londres. Y, aún así, los dosestaban estrechamente emparentados.Owen había hecho más suposiciones delo que se pensaba.
La suplantación del Iguanodon
rinocerino de Owen trajo consigo sus

propios problemas: ¿Con qué podía ser remplazado? ¿Cuál era el aspecto evida del enigmático dinosaurio? Existemuy pocos reptiles modernos cuyasextremidades posteriores tengan utamaño muy superior al de lasanteriores. La primera impresión deLeidy fue que el Hadrosaurus semantenía erecto y caminaba como ucanguro, aunque éste fuera un mamífero no un reptil. «La gran desproporció
de tamaño existente entre las partesdelanteras y traseras del esqueleto del
adrosaurus, me llevó a sospechar queeste gran lagarto herbívoro extinguidodebía de tascar y ramonear manteniéndose erecto, como el canguro,

sosteniéndose sobre sus extremidades posteriores y su rabo[3]». Sin embargo,el Hadrosaurus era un reptil y no umamífero, hecho que claramenteinquietaba a Leidy que estaba buscandoun modelo de reptil que le permitieramantener a su hadrosaurio en una postura postrada. La solución vino dada por otra de las restauraciones de Owedel Crystal Palace, el Labyrinthodon.Hawkins se había encargado derestaurar el Labyrinthodon, dándole elaspecto de una rana de tamaño humano yLeidy se aferró a esto como la únicaalternativa viable al canguro en elámbito de los animales de sangre fría.«No es improbable —concluyó

cubriendo de esta manera todas lasopciones posibles— que el
adrosaurus adoptara normalmente una posición postrada, evolucionando posteriormente de la forma que se creehizo el Labysinthodon, un batracioextinguido (anfibio) en un períodoanterior [4]». Los anfibios del triásico, dehecho, no presentaban ninguna de lasespecializaciones peculiares para saltar que observamos en las ranas. Cuandoapareció el resto del cuerpo, el
abyrinthodon resultó ser una bestiacon forma de cocodrilo que probablemente vivió de forma similar elos ríos y pantanos del triásico. Leidy pronto abandonó la rana y volvió a

recurrir a la analogía del canguro.La sospecha de Leidy, de que el
voluminoso hadrosaurio había tenidouna locomoción similar a la del canguro,fue respaldada totalmente cuando en lascanteras de marga de New Jerseyapareció otro saurio, encontrado por unode sus antiguos estudiantes, EdwardDunker Cope. Cope había sido un niño precoz cuyo interés permanente habíaestado dedicado a los reptiles; a la edadde diez años hacía unos dibujos a escalaexactos de los complejos huesoscraneales de los ictiosaurios expuestosen la Academia de Ciencias Naturalesde Filadelfia. No obstante, su conductaen la escuela era considerablemente

peor de la esperada en un cuáquero. Stendencia pendenciera, tanto física comointelectual resurgiría años más tarde coconsecuencias dramáticas cuando se vioinvolucrado en su infame lucha contra el paleontólogo de Yale, O. C. Marsh.Cope resultaría ser la fuerza impulsora para la búsqueda de saurios mesozoicosdurante tres décadas en el NuevoMundo: «impulsor» es la palabra clavedebido a que su carácter era deurgencia, energía y ambición, mezcladocon orgullo y envidia: una combinacióexplosiva. Poseía un temperamentofiero, carácter combativo y unadesmedida capacidad de investigación.En 1858, a los 18 años de edad, Cope

desafió por primera vez a los eminentesacadémicos de Filadelfia, remitiéndolea la Academia de Ciencias Naturales uartículo sobre los aspectos másdelicados de la clasificación de lassalamandras que inmediatamente fuereconocido y publicado. En sólo tresaños aparecieron treinta artículosadicionales sobre la clasificación de lasserpientes, lagartos y anfibios. Al finalde esta época, Cope comenzó seducación zoológica formal asistiendoal curso de Joseph Leidy de anatomíacomparada en la Universidad dePennsylvania. (Cope publicó de forma prolífica —1400 artículos, libros ymonografías durante su vida—,

produciendo los artículos con tantarapidez que cometió numerosos erroresaunque su orgullo nunca le permitióadmitirlos). La habilidad de Cope fuereconocida rápidamente por loseminentes científicos de la Academia deCiencias Naturales y, en 1861, fueelegido como miembro; cuatro años mástarde se convirtió en su director. Con elcomienzo de la guerra civilnorteamericana, Cope, a pesar del votode pacifismo de los cuáqueros, se vio profundamente comprometido, haciendoque su padre lo enviara de viaje aEuropa para que no se uniera al Ejércitode la Unión. Cuando, en 1864, regresóde su viaje a Europa, se convirtió e

profesor de zoología en el HaverfortCollege cuando sólo contaba 24 años deedad. El carácter nervioso de Copehacía que éste no se adaptara a laexistencia universitaria sedentaria. Scarácter extremadamente individualistatampoco hubiese permitido que launiversidad le quitara todo su tiempo; laenseñanza había acaparado muchotiempo de Leidy, quedándole a éste muy pocas horas para la investigación, yCope había aprendido la lección.Además, las conferencias de Leidysobre los saurios gigantes extinguidoscomo el Hadrosaurus habían estimuladola imaginación de Cope, de forma queéste ansiaba ir a vivir al distrito de

Haddonfield donde estaban enterradoslos monstruos. Hizo frecuentes viajes alas canteras de marga de New Jerseyque contenían los restos de losdinosaurios y, en 1866, se compró unacasa en la región. Su recolección defósiles tuvo tanto éxito que dos años mástarde dimitió de su puesto en la facultad,vendió la granja que su padre le habíaregalado (con la vana esperanza de quesu hijo le tomara afición a la vida en elcampo) y «se retiró» a Haddonfield paradedicarse por completo a la búsquedade nuevos saurios.
En el verano de 1866, en una de sus periódicas visitas a las canteras de WestJersey, encontró su primer dinosaurio,

un fémur de hadrosaurio deaproximadamente 76 centímetros delongitud. Deliberadamente procuróconvertirse en un personaje conocido etoda la región de las canteras paraasegurarse de que le fueran comunicadostodos los hallazgos de fósiles en cuantoéstos se produjeran. Su sistemafuncionó. A finales del mismo verano,cuando una de las canteras estaba produciendo una gran cantidad de restosde saurios, Cope fue llamado precipitadamente. Los huesos había producido un estado de excitación yCope escribió a su padre describiéndolela vista que se le había ofrecido desdeel fondo de la cantera:

Hallé los restos que tenían uninterés mucho mayor del quehabía esperado —se trataba,nada más ni nada menos, ¡que deun dinosaurio carnívorogigantesco totalmente nuevo, probablemente del género Megalosaurus de Buckland!, quefue el devorador y destructor del
Hadrosaurus de Leidy y de todolo que podía atrapar—. Se haconservado una garra o falangede los dedos que como mínimomide 20 centímetros de longitud;su forma es como la garra de unáguila y como la zarpa de unleón[5].

Este descubrimiento reforzaría lareputación internacional de Cope. El
adrosaurus cretáceo de New Jerseyhabía finalmente encontrado su pareja.«El descubrimiento de este animal llenóel vacío en la fauna cretácea —escribióCope, adoptando la ética gladiatorial predominante—, revelando al enemigocarnívoro del gran Hadrosaurus
herbívoro[6]». El adversario delhadrosaurio no era el megalosaurioinglés, sino un protagonista mucho másformidable. Se trataba, dijo Cope,observando más detenidamente losdientes en forma de daga y las garras,del carnívoro terrestre más rapaz que elmundo haya conocido. Cuando exhibió

el dinosaurio ante la Academia deCiencias Naturales, en agosto de 1866,escogió para la bestia depredadora elnombre de Lealaps aquilunguis. (En lamitología griega, Laelapo, era un veloz perro que la diosa Diana había regaladoal joven Céfalo, amante de la caza. Udía durante la caza, Laelapo fueconvertido en piedra por los dioses,siendo su postura tan natural que parecíaestar a punto de saltar).
Afortunadamente, el esqueleto deCope incluía los huesos de lasextremidades anteriores y posteriores yla discrepancia de tamaño era inclusomás marcada que en el saurio de Leidy.El fémur de 78 cm de longitud hacía

parecer más pequeño al correspondientehueso del brazo que, restaurado, habríaalcanzado una longitud de 30centímetros. (El hueso de Cope estabaincompleto). Este hallazgo apoyó lateoría de Leidy de que estos animaleshabían sido canguros reptiles gigantes.El escenario cretáceo quedó así biedefinido: los Hadrosaurus y Laelaps
saltaban como canguros monstruosos a pesar de que tenían una longitudsuperior a los 6 metros y se perseguíamutuamente de esta manera. Lasextremidades anteriores ridículamente pequeñas del Laelaps no servían comoarmas de ataque; esta función quedabarelegada a las extremidades posteriores

con aspecto de garras de águila. Elaelaps saltaba persiguiendo al
hadrosaurio mientras quesimultáneamente lo hería con sus garras.O al menos esto era lo que pensabaCope.
Los descubrimientos de Cope fuerohechos en un momento oportuno. Susdinosaurios resucitados pronto seríadevueltos a su supuesto aspecto natural,al igual que lo habían sido suscontemporáneos en Londres unas cuantasdécadas antes. La fama de lasrestauraciones del Crystal Palace sehabía extendido hasta el otro lado delAtlántico seguida de cerca de sinventor Waterhouse Hawkins. Los

miembros de la comisión encargada delCentral Park en Nueva York se daba perfecta cuenta del valor educativo de laaventura de Londres. Andrew Green,que era el administrador de la comisión,observó, en 1868, que «aunque el parque de Sydenham está situado avarios kilómetros de distancia de lametrópoli, y a pesar de que hay que pagar una entrada, miles de personas lohan visitado anualmente en los últimosquince años, siendo las restauracionesde los animales un elemento permanentede atracción y una fuente de valiosainstrucción para multitud de personasque de ninguna otra manera hubiese podido adquirir este tipo de

conocimiento[7]». Al igual que Hawkins,Green creía en la educación a través dela «enseñanza objetiva» y fue la fuerzaimpulsora para la fundación del MuseoAmericano de Historia Natural en 1869.En mayo de 1868, Green se dirigió aHawkins, en Nueva York, con la petición «de realizar la resurrección deun grupo de animales de los períodosantiguos del continente americano».Hawkins no se pudo resistir a la oferta yrápidamente empezó a trabajar visitandolos museos de Washington, NewBruswick, Albany o New Haven e busca de sujetos dramáticos adecuados.En Filadelfia se encontró con Cope yLeidy en la Academia de Ciencias

aturales y su búsqueda acabó cuandovio los saurios gigantes expuestos en elmuseo de la Academia[8]. La insistenciade Leidy en la lucha por lasupervivencia con dientes y garras elos tiempos prehistóricos, visiómagnificada y descrita de forma aún mássanguinaria por Cope, evidentementetuvo una gran influencia sobre Hawkinsque se expresó a través de las posturasque éste tenía planeadas para suscriaturas.

a. Megalosaurus, el dinosaurio carnívoro jurásico, tal como se lo imaginaba Owen en1854. Éste contrastaba con la idea que teníaCope del carnívoro Laelaps menos de dosdécadas más tarde. Los paleontólogos quetrabajaban en América fueron los primerosen darse cuenta de que los carnívorosandaban sobre las patas traseras.
Desde setiembre hasta diciembre de

1868 estuvo haciendo moldes de losfósiles del Hadrosaurus y Laelaps delmuseo, rellenando los huecos dejados por los huesos que faltaban con réplicasde yeso. Mientras Hawkins estaba en elmuseo, los recolectores de Cope eKansas enviaron a Filadelfia u plesiosaurio de 12 metros de longitudcon cuello de serpiente encontrado en lacreta y, aparentemente, fue el propioHawkins quien durante un mes se dedicóa «sacarlo de la matriz» para Cope,antes de hacer los moldes para su propiomuseo propuesto en Central Park [9].
Este plesiosaurio de cuello largo,descrito por Cope en 1868 y llamado
lasmosaurus (o «reptil de cinta»),

desencadenaría la rivalidad paleontológica más explosiva de todoslos tiempos, enemistad que marcaría a la paleontología americana durante lossiguientes veinte años. Cope prosperócon la controversia pero fue el amargo perdedor. Así, su amistad con OthnielCharles Marsh, profesor de paleontología en el recientementeinaugurado Museo Peabody de Historia
atural, en la Universidad de Yale, seconvirtió repentinamente en antagonismo finalmente en amargura, cuando Mars
osó criticar la restauración delelasmosaurio de Cope.
A diferencia de sus contemporáneoscon talento y prolíficos, Marsh esperó

treinta años antes de publicar uartículo. En su infancia había sidotímido y arisco, e incluso a los 21 añosde edad aún se imaginaba un futuroseguro, o incluso trivial, en una profesión comercial mecánica. No eraun comienzo muy propicio para un futurolíder de la ciencia norteamericana. Elcatalizador que inició el tardío despeguefue el dinero. Marsh era el sobrino delmillonario George Peabody, a quien losnegocios prósperos en la Banca deLondres y tendencias filantrópicasconvirtieron en el objetivo natural de losnecesitados familiares. Peabody,consciente de su propia falta deeducación sólida, donó gran parte de s

fortuna a instituciones educativas. Marsdedujo sagazmente que Peabody nuncale daría dinero a un sobrino pocoambicioso. Así, en un intento deimpresionar a su tío, Marsh —que habíaingresado en la Academia Phillips deAndover, una escuela de preparación alos estudios universitarios, a los 21 añosde edad, donde adoptó el apodo de«Daddy»—, comenzó a reunir distinciones, honores y buenascalificaciones. Envió éstos a sdistinguido tío junto con una prudente petición de dinero que le permitiesecontinuar su educación en launiversidad. Peabody respondió a lainiciativa y envió al ya adulto Mars

(que entonces tenía 25 años) a laUniversidad de Yale.
Marsh pasó los años que duró laguerra civil de Estados Unidos eEuropa, examinando las coleccionesfósiles y calculando su siguiente jugada.Cope, en su gira electoral por lascapitales europeas, explorando lossótanos de todos los museos einformándose sobre cualquier personade importancia, visitó a Marsh en laUniversidad de Berlín en 1863. Éste fuesu primer encuentro. Marsh tenía 32años y Cope era nueve años más joven.Marsh, con sus comienzos tardíos,acababa de comenzar a contribuir a laciencia mientras que Cope ya había

publicado multitud de artículoscientíficos. Nada concreto los dividía,aunque Marsh pensó que «lasexcentricidades de la conducta» deCope eran difíciles de tolerar. En años posteriores, Marsh declaró que yaentonces dudó de la cordura de Cope, urecuerdo nublado por una vida de mutuoodio. Fue el clásico choque de personalidades: el impetuoso, enérgicoCope, imaginativo y excitable, rápido eformarse opiniones en contraposiciócon el impasible Marsh. Marsh, dehecho, se pasó los años en el extranjeroideando una manera de obtener aún másdinero de su tío. Esperaba poder utilizar su influencia para persuadir a Peabody

que fundara un museo de historia naturalen Yale, consiguiendo él de paso u puesto como profesor. La tácticafuncionó. Marsh asombró a sus profesores de Yale con el anuncio deque su tío iba a donar 150 000 dólares para la fundación de un museo y unacátedra de paleontología. A modo degratitud, las autoridades con muchogusto le concedieron a Marsh unacátedra. Teniendo ahora garantizados por Peabody los ingresos de por vida,Marsh podía libremente investigar yexplorar. Desde esta posición ventajosafue escalando puestos, cogiéndole elgusto al prestigio adquirido que sólo podía ser satisfecho con nuevos

nombramientos. Fue designado paleontólogo especializado evertebrados del Geological Survey deEstados Unidos, lo cual hizo que susfondos aumentaran aún más. Mientrasque las menguantes finanzas de Cope eaños posteriores provocaron su retiro alos confines de su casa, que servía almismo tiempo de museo y laboratorio,Marsh disfrutaba de todos los recursosdel espacioso Museo Peabody, delacceso al Survey (que patrocinaba susexploraciones en el campo y pagaba alequipo de Yale) y del lujo de unamansión de 18 habitaciones en NewHaven. Mientras proliferaban losartículos de Cope, el prestigio de Mars

iba aumentando y ambos hombres seobservaban recelosamente. Marsh,cuyos artículos científicos siempreestaban bien planeados y ejecutados,trabajaba metódicamente eindividualmente de forma que svolumen de producción sólo constituíauna fracción del publicado por Cope.Cope, siempre oportunista, pasaba de utema a otro, siendo incapaz de ignorar material alguno que se le presentase. Susartículos trataban sobre cualquier tema,desde los fósiles de tiburones hasta s propia teoría de los mecanismos de laevolución. Marsh, que desgraciadamenteno era un teórico, se limitaba al sólidotrabajo descriptivo, describiendo sus

resultados en la prosa paleontológicamás pesada. Marsh, de hecho, podía permitirse el lujo de desdeñar loserrores. Cope, que frecuentemente se precipitaba para obtener la prioridad,metió la pata más de una vez. Fue eeste punto cuando finalmente estos dosvoluntariosos protagonistas seenfrentaron abiertamente.
Las descripciones publicadas por Cope acerca del «reptil de cinta»hicieron que pareciese la criatura másnotable de todos los tiempos, taradicalmente diferente a todas las demásque necesitaba el reconocimiento de uorden de reptiles totalmente nuevo: losestreptosaurios o «reptiles retorcidos»,

llamados así por su largo y flexiblecuello. El boceto de la restauración deCope de esta «milagrosa creación», talcomo la denominaba sarcásticamenteMarsh, apareció en la revistaTransactions de la Sociedad FilosóficaAmericana. El dibujo se basaba en elesqueleto reconstruido del elasmosaurioen el museo de la Academia de Ciencias
aturales, exposición que Copeorgullosamente mostró a Marsh durantela visita que este último hizo aFiladelfia en 1870. Cope no pudodisfrutar mucho tiempo de su preciada posesión. Cuando Marsh vio elesqueleto y Cope le explicó sus peculiaridades, inmediatamente se dio

cuenta de cuál era la razón de lasingularidad de la criatura. «Observéque las articulaciones de las vértebrasestaban puestas al revés —explicóMarsh más tarde— y le sugerísuavemente que había confundido la parte delantera y trasera del animal[10]».Cope estaba furioso y se lo hizo saber aMarsh en términos no precisamentevagos. No había estado estudiando elanimal durante tantos años para nisiquiera poder distinguir un extremo deotro.
Parece que efectivamente sí se habíaequivocado ya que el profesor Leidycogió tranquilamente la última vértebrade la «cola» y la adaptó al cráneo.

Cope, en efecto, había colocado lacabeza en el extremo de la cola, creandoasí la maravillosa criatura a partir desus restauraciones. Cuando Marsh sacóa relucir las dudas de Leidy, «la vanidadherida de Cope recibió un golpe del cualnunca más se recuperó y desde entoncesha sido mi implacable enemigo», dijoMarsh más tarde. El desconcertadoCope rápidamente procuró comprar, cosu propio dinero, todos aquellosejemplares de Transactions quecontenían su restauración. Marsnaturalmente se guardó dos copias. A partir de este momento, Marsh y Copefueron implacables adversarios,compitiendo en cualquier aventura y

luchando continuamente por la primiciade las publicaciones. Durante dosdécadas, la paleontología americanaestuvo dominada por esta enemistad, yla motivación que incitaba a amboshombres a buscar fósiles, debe ser considerada un producto de la misma.La guerra, en realidad, no sedesencadenaría hasta 1877. En este año,Marsh anunció al mundo eldescubrimiento de un dinosauriorealmente gigante, el «reptil titánico» oTitanosaurus. En el mismo artículo queapareció en julio, aprovechó paraobservar que la bestia especial de Cope,el Laelaps, llevaba el mismo nombreque al menos otras dos criaturas. Debido

a que éstas habían recibido su nombreunos treinta años antes tenían prioridad.De acuerdo con las reglas que rigen ladesignación de nombres, «Laelaps»debería ser cambiado y el propio Marsle dio el nuevo nombre de
ryptosaurus[11]. Cope no le prestóninguna atención a la corrección ycontinuó utilizando el nombre de
aelapos hasta su muerte. ¡Sin embargo,a los pocos días de haberse publicado elartículo de Marsh, Cope se desquitódeclarando que Titanosaurus era unombre que ya pertenecía a otro animal que sería mejor que Marsh buscara
otro[12]! El «debate» continuó en estetono histérico mientras que los otros

paleontólogos, procurando no ser arrastrados hacia esta lucha, tomaron urumbo más sensato.
Entretanto, el culpable original, ellasmosaurus, al igual que los otros
saurios de la Academia de Cienciasaturales, había sido dibujado por el
infatigable Hawkins que también hizolos moldes. La Academia de Ciencias
aturales de Filadelfia le proporcionólas instalaciones y los equipos y, acambio, recibió la primera restauraciódel esqueleto en tamaño natural de u
adrosaurus de 11,9 metros delongitud, que se apoyaba sobre lasextremidades posteriores agarrado a uárbol con los brazos y ramoneando. Este

modelo permaneció expuesto en laAcademia durante muchos años antes deser desmantelado. En diciembre de1868, Hawkins volvió a Nueva York ycomenzó el trabajo de restauración en utaller paleontológico especialmenteinstalado en la parte sudoccidental delCentral Park. Allí empezó a reconstruir dos modelos de Hadrosaurus, unoerguido como el espécimen de laAcademia de Ciencias Naturales y otroacostado en una postura parecida a la delas esfinges, con miembros posterioresenormes y pies de ave.
Los reptiles gigantes debían ser alojados en el Museo Paleozoico planeado para el parque. Al igual que s

homólogo de Londres, el edificio estabadiseñado como un edificio enorme decristal con una estructura de hierro y utecho abovedado. Mucho antes de quecomenzaran los trabajos del museo,Hawkins ya había hecho dibujos de sinterior y, a partir de éstos, nos podemoshacer una idea de las posturas que tenía previstas. El Hadrosaurus más grandedel dibujo era atacado por uno de los
aelaps más pequeños pero feroces,mientras que otros dos Laelaps se peleaban encima de un enorme cadáver.A sus pies estaban escondidos en el agua plesiosaurios gigantes y mosasaurios.También estaba previsto realizar modelos de los animales

contemporáneos más espectaculares delhombre primitivo como los mamuts,alces gigantes, armadillos gigantes y perezosos gigantes. El objetivo era,según el informe anual de la comisión,representar «una historia visualcompleta del continente americanodesde los inicios de la creación hastanuestros días». Aun no alcanzando esteobjetivo, el museo hubiese ilustradográficamente aquellos períodos de lahistoria de la Tierra en los cualesverdaderamente reinaban los gigantes.
Los cimientos fueron echados por el principal jardinero paisajista de
orteamérica, Frederick Law Olmstead,en la acera opuesta de la Calle 63, y

antes de su debido tiempo aparecieromapas del parque que ya ilustraban eledificio del museo[13]. Los sucesos quese produjeron a continuación sodifíciles de esclarecer debido a que loshechos que conocemos son de carácter muy fragmentario. Durante varios años,William Marcy Tweed había estadoconstruyéndose un imperio privado e
ueva York. El círculo de Tweed habíalogrado, mediante dinero o amenazas, elcontrol completo sobre todos los bastiones de poder de la ciudad, desdelos cuarteles generales del PartidoDemócrata hasta la administración deusticia. Los secuaces de Tweed fuero
introducidos en todos los puestos

ejecutivos clave. El historiadorA. B. Callow dijo recientemente:«Desde 1866 a 1871, la organización deTweed saqueó la ciudad de Nueva York con tal precisión que ha recibido lasingular distinción de ser etiquetadacomo el modelo de corrupción cívica ela historia municipal de EstadosUnidos[14]». El saqueo mediante lacorrupción y robo del tesoro hicieron de«Boss» Tweed un hombre inmensamenterico, lo cual a su vez hizo que aumentarasu dominio completo sobre la ciudad yse convirtiera en el tercer propietariomás rico de bienes raíces en NuevaYork. Tweed conservaba su poder político mediante el patrocinio,

proporcionando puestos de trabajo acambio de un apoyo continuo. Para poder acomodar a sus partidariosnecesitaba tener el control de grandeszonas de la ciudad y no constituyeninguna sorpresa que pronto echara elojo a la dirección del Central Park. E1870 forzó a la asamblea legislativa aque aboliera la comisión de AndrewGreen y colocó a sus propios hombresen los puestos de control. Green vioimpotente cómo la mano derecha deTweed, Peter Brains Sweeny, tomaba elcontrol sobre el parque. El New York
Times, una de las pocas institucionesque fueron capaces de resistirse algrupo, profetizó la explotació

sistemática y el consiguiente ocaso del parque.
La nueva administración de Tweed,que se hizo cargo de la dirección el 1 demayo de 1870, desde el principio estuvoen contra del Museo Paleozoico.Aparentemente, el trabajo fueinterrumpido porque estaba costándoledemasiado dinero a los contribuyentes.El coste final estimado del edificio erade 300 000 dólares, lo cual «parecía unasuma demasiado grande como para ser gastada para un edificio dedicadototalmente a la paleontología, unaciencia que, pese a ser interesante, es ela actualidad tan imperfectamenteconocida como para no justificar u

gasto público tan grande parailustrarla». Resulta irónico a la luz deesta declaración que el grupo de Tweedse gastara 8 millones de dólares en los primeros dieciocho meses patrocinandoempleos a cuatro mil trabajadores en el parque, para el cual Green sólo habíanecesitado un cuarto de millón dedólares al año. Algunos meses mástarde, un portavoz del Liceo de Historia
atural del Museo Americano, uno delcada vez más creciente número deadversarios de Tweed, satirizó la posición de Tweed: «El profesor Hawkins —dijo imitando a Tweed— haestado estudiando libros y huesos; ¿quées lo que sabe acerca de la dirección de

Central Park? De todas formas, sólo eraun inglés y la idea de intentar obtener umuseo en esta ciudad sin tener un pla para dividir los beneficios eraabsurda[15]». El grupo de Tweed, al ver que el museo no le producía beneficioalguno, decidió demolerlo hasta loscimientos.
A principios de 1871 los modelos deHawkins estaban a punto de quedar terminados en su taller. Una ilustracióen la revista Repart de la comisiómuestra que, en 1868, había completadoel Hadrosaurus más pequeño yrealizado el molde del esqueleto de otromás grande. Probablemente tambiéhabía acabado el elasmosaurio co

cuello de serpiente. Un inventario del parque, realizado por el grupo de Tweedal día siguiente de la toma de posesión,muestra que había construido el rapaz
aelaps y «siete moldes de yeso» detipo no especificado. Pero éstos nosobrevivirían. Un día de primavera de1871, por orden de los funcionarios deTweed y con la bendición de Sweeny,unos vándalos entraron en el taller ydestrozaron los dinosaurios amartillazos enterrando posteriormentelos trozos en el parque. Los moldes ymaquetas fueron destruidos en un acto de puro vandalismo caprichoso, mientrasque el conmocionado Hawkins erainsultado diciéndole que «no se

preocupara de los animales muertoshabiendo tantos animales vivos quecuidar [16]». No existe motivación alguna para esta destrucción. Boss Tweed podría haberse forrado los bolsillosvendiendo los modelos a la SmithsoniaInstitution de Washington que con gustolos habría aceptado. El desprecio deTweed por el proyecto podría haber surgido a raíz de un prejuicio religioso profundamente arraigado. Ciertamente parecía ser escéptico respecto a laantigüedad de los dinosaurios,refiriéndose a los modelos de Hawkinscomo «especímenes de animales que sedice precedentes del período preadanita[17]». Unos meses más tarde se

produjo la caída de Tweed inducida por Louis Jenning, un editor ingléscontratado especialmente para este fi por el New York Times. Las invectivas yexposición de los fraudes realizados por el Times provocaron la pérdida de poder político del grupo de Tweed y el procesamiento de sus líderes. Tweed ySweeny huyeron al extranjero y lacomisión del parque volvió a manos delcompetente Andrew Green. Peroentonces el daño ya estaba hecho.
Hawkins, abrumado, regresó a laEscuela de Nueva Jersey (Universidadde Princeton) y, después de un retiroacadémico de varios años, volvió a sInglaterra nativa en 1877. La animosidad

de Tweed hacia las creaciones impíasde Hawkins no podía haber tenido unainfluencia duradera ya que, en 1875,Hawkins volvió a trabajar de nuevo y eel espacio de sólo dos años acabó (ocasi acabó) una serie de diecisietegrandes pinturas para la universidad.Las escenas prehistóricas, que se basaban en gran medida en susinvestigaciones colectivas del CrystalPalace y Central Park, son asombrosas por su intento de infundir algo de vidaen los dinosaurios resucitados[18].Incluso hizo un esfuerzo final parareanimar al Hadrosaurus dejado a ssuerte, interrumpiendo sus pinturas e1878 para construir una efigie de tamaño

natural para el centenario de laDeclaración de la Independencia,celebrado con una exposicióinternacional en Filadelfia. Debido aque el modelo fue encargado por el
ational Museum en Washington, mástarde fue devuelto a la capital ycolocado al aire libre enfrente delmuseo. Sin embargo, este modelotampoco estaba destinado a sobrevivir.Parece ser que Hawkins lo fabricó deeso y a consecuencia de estar expuesto
a la intemperie, simplemente se fuedegradando con el tiempo.
Una consecuencia deldescubrimiento de dinosaurios coaspecto de canguros era que muchas

personas miraban despectivamente a losmodelos del Crystal Palace, que parecían rinocerontes reptiles. Marsera un crítico especialmente duro. Noera capaz de ver las restauracionesdesde un punto de vista histórico, como producto del limitado conocimiento delque disponían Owen y Hawkins. Trasvisitar el palacio, en 1895, hizo unoscomentarios despectivos referentes a losmodelos en una convención de laAsociación Británica:
Los dinosaurios parecen…haber sufrido mucho tanto por sus enemigos como por susamigos. Muchos de ellos fueron

destruidos y desmembrados hacetiempo por sus enemigosnaturales, pero, incluso másrecientemente, sus amigos leshan hecho injusticia reuniendosus restos esparcidos yrestaurándolos con sus formas devida supuestas… Por lo que yo puedo juzgar, no existe nada parecido a ellos en el cielo, o enla Tierra, o en los mares o bajola Tierra. Nosotros ahorasabemos de buena fuente que el
Megalosaurus y el Iguanodon
eran bípedos, y representarlosarrastrándose, excepto en su mástierna juventud, sería casi tan

absurdo como hacer lo mismo enel género Homo[19].
Los monstruos podían haber parecido unas curiosas reliquias en laépoca de Marsh, pero sus críticas erainjustas: en 1854 no había ninguna razó para sospechar que muchos dinosauriostenían el aspecto de canguros gigantes.
De hecho, los paleontólogos ingleseshabían respondido rápidamente a losdescubrimientos americanos y no tardómucho en producirse la aceptación total.Thomas Henry Huxley, profesor extraordinario de paleontología en laEscuela de Ingeniería de Jermyn Street

(que más tarde se convertiría en elLondon University’s Imperial College),celebró el hallazgo de Leidy y dijo quelos dinosaurios ingleses como el guanodon también debían de haberseapoyado sobre sus patas traseras.Además, las huellas de los pies con tresdedos halladas en los Weald (hechas por una gran criatura de zancada larga: probablemente el mismo Iguanodon) lehicieron pensar que esta criaturacaminaba erguida sobre sus patastraseras. Huxley, sin embargo, comoardiente evolucionista, tenía otrosmotivos para aceptar las teorías deLeidy. Apenas se había publicado elOrigen de las especies, en 1859, cuando

la tenaz defensa de Huxley de sus principios ante los paleontólogos y elclero por igual le valieron el apodo de«bulldog de Darwin». Una vez Darwihubo preparado el trabajo preliminar yvisto publicado su libro, el Origen de
las especies, se retiró de la arena y dejóque Huxley combatiera por él. Por ello,cualquier cosa que apoyara la causadarwiniana sería aprovechada por Huxley. Allá donde Darwin no osabaentrar, Huxley se lanzaba a batallar.Huxley no tenía escrúpulos en sacar alhombre de su inamovible posición en elUniverso. El origen evolucionista delhombre era un tema incendiario, que cotoda garantía provocaría gritos de

indignación del clero y Huxleydisfrutaba acosando a la oposición. Laimagen victoriana barbuda, severa yceñuda de su retrato no muestra elencanto de este atractivo provocador
que se escondía en su interior. Nadie selibraba de los ataques de Huxley; primeros ministros, científicos yclérigos, todos eran tratados con elmismo entusiasmo. «Los extinguidosteólogos —se mofó en 1860— estáesparcidos alrededor de las cunas decada ciencia sin hacer nada, como lasserpientes estranguladas junto a la deHércules». En posteriores años dedicócada vez más tiempo a los trabajos polémicos, llevando la bandera

evolucionista al campo de losadversarios, con tal éxito que él mismose otorgó el título de Episcophagous o«tragaobispos». Fue el primero ecomprender completamente lasimplicaciones de la evolución para elhombre, y fue el primer agnóstico. Eldeshacerse hábilmente de la oposicióclerical, sin embargo, sólo era una víade escape para las inclinacionesmaliciosas de Huxley; fue en su batallacontra sus superiores académicos, yespecialmente en su utilización de losfósiles como prueba de la evolución,donde Huxley prestó su más valiososervicio a la causa evolucionista.
De esta forma ocurrió que, en una de

sus conferencias extremadamente populares en la Royal Institution e1868, Huxley comenzó su violentoataque contra los principales problemas paleontológicos con los que seenfrentaba la doctrina de la evolución.¿Cómo es que, arguyó en nombre de sus protagonistas, si todos los animales hasurgido de un mismo tronco común,existan esos grandes saltos que se daentre peces y reptiles, reptiles ymamíferos, etc.? Nosotros, los quecreemos en la evolución, replicamos queen otros tiempos existieron formas querevolucionaban unas especies con otras, pero que ahora están extinguidas. Por ejemplo, parece que existe un gran salto

entre los reptiles y las aves. Siembargo, el recientemente descubierto
rchaeopteryx, o «pluma antigua» deSolnhofen en Baviera era un perfectosemireptil, semiave que vivió en laépoca jurásica: tenía plumas pero almismo tiempo también dientes, una largacola ósea y dedos largos. Las avessaltan o corren con las patas traseras, deforma que Huxley investigó si existíaalgún reptil que hiciera lo mismo y que pudiera estar a medio camino de lasaves. El Iguanodon, con sus piestraseros con tres dedos, era la criaturaidónea. El Iguanodon erguido y susaliados estaban dotados de cuartostraseros «sorprendentemente parecidos»

a los de las aves, proporcionándole aHuxley el primer indicio de laantigüedad de las aves. El pequeñodinosaurio con el nombre deCompsognathus, un contemporáneo delhíbrido Archaeopteryx, hallado en losdepósitos de pizarra de Baviera, teníaun aspecto aún más parecido al de uave. Debido a que sólo medía 60,8centímetros de longitud, también se parecía más a las aves primitivas por stamaño y, por lo tanto, hacía que losargumentos de los adversarios,referentes al tamaño gigantesco del guanodon, fueran menos molestos.Unos miembros traseros largos ydelicados proporcionaban al

Compsognathus cierto parecido con lasaves. «Es imposible mirar laconformación de este extraño reptil — dijo Huxley durante una conferencia ela Royal Institution— y dudar de queandaba o saltaba en una postura erguidao semierguida, más o menos como uave, con las que debía de tener uextraordinario parecido gracias a scuello largo, su diminuta cabeza y sus pequeños miembros anteriores[20]».
El absoluto compromiso de Huxleycon la causa evolucionista le llevó aapreciar rápidamente la importancia delos descubrimientos de Leidy y acelerar su reconocimiento en Inglaterra. Huxleyestaba entonces en una posición que le

permitía reinterpretar las observacionesde Edward Hitchcock acerca de lashuellas de «ave» de la antigua NuevaPiedra Arcillosa Roja del triásico deConnecticut y Massachusetts. Hitchcock era el presidente del Amherst College y profesor de teología natural y geología
, gracias a su intervención, la escuelafue la primera en obtener algunas de lasextraordinarias huellas fósiles de tresdedos en 1835. Hitchcock las guardó esu «armario», nombre bajo el cual seconocía la colección de fósiles. Lashuellas habían sido descubiertas en elvalle cruzado por el río Connecticut yhabían sido llevadas a Hitchcock parasu identificación. Éste se dio cuenta de

su valor y estuvo trabajando durante tresdécadas, recorriendo los 128 kilómetrosde longitud del valle y descubriendo 21lugares con huellas de pies. En 1848 poseía las huellas características de 49especies, de las cuales 32 eran criaturaserguidas, bípedas con sólo tres o cuatrodedos. Debido a que las huellas se parecían a las de aves caminando por elagua, Hitchcock les dio el nombre de«huellas de ave pétreas». Por sugerenciade James Dana, que más tarde seconvertiría en el partidarioincondicional de Darwin en laUniversidad de Yale, Hitchcock asignónombres a todas sus huellas para evitar la confusión, dividiéndolas de acuerdo

con características tales como número ygrosor de los dedos, presencia de talón,etc. Algunos de sus colegas objetaroque no era una práctica común darlenombres a criaturas de las cuales no sehabían descubierto los restosverdaderos. Nunca se hallaron losesqueletos, pero Hitchcock argumentó persuasivamente que unos pies particulares debían caracterizar a unosanimales particulares: no existiríaconfusión alguna entre avestruces yáguilas o carnívoros y rumiantes aunquesólo se conocieran sus huellas. Erealidad, la anatomía comparada deCuvier había hecho que este enfoquefuera respetable. A partir de un único

hueso, el barón francés habíareconstruido un animal entero y de formasimilar, a partir de una sola huella de u pie podía evocar a su artífice.«Cualquiera —decía Cuvier— queobserve la huella de una pezuña podráconcluir que ésta ha sido hecha por uanimal rumiante, y considerar que estasconclusiones son tan ciertas como lo puedan ser otras en física o ética». A partir de una huella, la técnicacomparativa de Cuvier le permitíaevocar intensamente la criatura hasta el punto de poder describir sus dientes.Con este impresionante precedente,Hitchcock comenzó a trabajar describiendo las criaturas que, en la

época del triásico, habían pululado por el valle del Connecticut dejando sólosus huellas como recuerdo. Se pensabaque muchas de las huellas decuadrúpedos habían sido dejadas por el
abyrinthodon al pasar, pero, egeneral, las huellas eran de tres dedos ymuy grandes. El «animal gigante» deHitchcock, el Brontozoum Giganteum,era conocido sólo por sus huellas y era,con mucho, la criatura dominante en el paisaje tanto por su número como por stamaño: sus pies llegaban a medir 45centímetros de longitud y la zancada erade aproximadamente 1,8 metros. Estosgigantes, dijo Hitchcock en 1848, eraobviamente aves zancudas con u

tamaño cuatro o cinco veces superior alde las avestruces. «Debieron de ser losgobernantes gigantes de aquel valle — pensó Hitchcock, conclusióaparentemente confirmada por snúmero, ya que creía que en la regióhabían vivido muchas bandadas—. Scarácter gregario se deduce del hechoque, en algunas localidades, hallamoshileras paralelas de huellas a pocoscentímetros de distancia unas de otras».Al igual que muchas aves zancudas, las bandadas de estas criaturas se había pavoneado por las orillas del antiguolago.
Cualquiera que sea la duda

que podamos tener acerca delemplazamiento exacto de estosanimales en la escala zoológica, podemos tener la seguridad deque muchos de ellos eranextraños y gigantescos, y heexperimentado la excitación deun romance ya que he regresadoa aquellas épocas inmensamenteremotas y observado aquellasorillas donde se paseaban estosenormes y heteróclitos seres.Ahora he visto, como científico,un ave áptero, de unos 3,65 a 4,5metros de altura —mejor dicho,grandes bandadas de ellos—,caminando por la superficie

lodosa, seguidos de otrosmuchos de carácter análogo perode tamaño más pequeño. Acontinuación un animal bípedo,un ave quizá, con pies y talonesde casi 61 centímetros delongitud. Luego una multitud de bípedos más pequeños, con lamisma forma general; y entreellos algunos moviéndoseerguidos, otros arrastrándose por la superficie con miembrosextendidos… Pero el mayor milagro viene en forma de batracio bípedo, con unos piesde 50 centímetros de longitud.Hemos oído acerca del

Labyrinthodon en Europa —unarana tan larga como un buey; pero sus pies sólo medían 15 o20 centímetros de longitud—, un pigmeo comparado con elOtozoum de Nueva Inglaterra.Detrás de éste camina, sobre patas desproporcionadas, ungrupo de pequeños lagartos ySalamandridae, con patastrífidas o cuadrífidas. Más allá,medio escondidos en laoscuridad, se mueven animalestan extraños que difícilmente pueden ser incluidos en los tiposde organización existente. Estacolección de fieras de la remota

era de la arenisca es realmenteextraña[21].
El zoo prehistórico de Hitchcock estaba modelado según los tiposexistentes y, comprensiblemente, s primera y última impresión era que eltriásico había estado gobernado por avestruces zancudas monstruosamentegrandes. «Sin embargo —dijo una vezhaciendo sin querer un juego de palabras —, los geólogos deberían ser losúltimos hombres en confiar en sus primeras impresiones[22]». Hitchcock siguió obstinadamente fiel a susavestruces del triásico incluso tras el

descubrimiento de Leidy deladrosaurus con aspecto de canguro.
En Inglaterra, el joven y aventureroHuxley no tuvo los mismos escrúpulosen repoblar el valle del Connecticut.
Huxley se hallaba en una posiciómucho más fuerte. La maravillosaevidencia de la existencia de vida en laépoca triásica, proporcionada por estasareniscas a pesar de la falta de huesos o plumas, le sugerían numerosas ideasinstructivas acerca de las criaturas queen otra época habían caminado por estasarenosas orillas. «La importante verdadque revelan esas huellas —afirmóHuxley— es que, al comienzo de laépoca mesozoica, existían animales

bípedos que tenían pies de ave yandaban con la misma postura erguida osemierguida». Huxley supuso que lashuellas más pequeñas fueron dejadas por aves, pero que muchas de lasrestantes (especialmente las huellasgigantes con tres dedos) pertenecían adinosaurios. De esta forma, Huxleyestaba dispuesto a admitir que losgrandes dinosaurios bípedos habíavivido incluso en los remotos días del
triásico.De hecho, todas las huellas podía
ser atribuidas a reptiles y anfibiosarcaicos, las aves aún no habían hechosu aparición en el cielo aunque niHuxley ni Hitchcock lo sabían. No

obstante, el enfoque de Huxley estaba bien escogido: la confusión entre lashuellas de aves y dinosaurios bípedossirvió únicamente para destacar las
similitudes entre estos dos grandesgrupos. Si averiguando la ascendenciade las aves, Huxley podía demostrar queéstas se habían vuelto tan parecidas alos reptiles que ya no podían ser distinguidas de los dinosaurios coaspecto de ave, habría demostrado queno existe «nada salvaje o ilegítimo» ela hipótesis de que las aves habíatenido sus orígenes ancestrales en losdinosaurios. Éste fue probablemente el primer éxito importante de Huxley, yaque consiguió que la paleontología

estuviese al servicio de la evolución;este logro fue rápidamente superado por la descripción de Huxley, en 1870, delos pasos en la evolución de un caballode un dedo a partir de su ancestroarcaico de cuatro o cinco dedos quevivía en el eoceno.
La fe de Huxley en la bipedalidaddel Iguanodon estaba totalmenteustificada, en 1878, cuando en una mina
de carbón en Bélgica se descubrió ugrupo entero de iguanodontes. Losdinosaurios estaban enterrados a más de304 metros de profundidad eBernissart, cerca de Mons, no en elcarbón sino en una fisura que habíaquedado cubierta de tierra cretácea.

Desde el período arcaico carbonífero delos pantanos que formaron el carbóhasta el mundo del Iguanodon deWealden habían transcurrido 180millones de años. Sin embargo, en laépoca cretácea, una profunda barrancaen los depósitos de carbón habíaactuado como una trampa mortal paralos dinosaurios y en ella cayeron erápida sucesión no menos de 31 guanodones adultos. Los mineros queestaban abriendo una nueva galería en lamina Fosse Sainte-Barbe habían hechoun túnel que llegaba hasta esta barracacretácea obstruida, destruyendocompletamente un esqueleto antes dedarse cuenta. La enormidad del hallazgo

se hizo manifiesta cuando los expertosdel Museo Real de Historia Natural, eBruselas, intentaron construir una nuevagalería 30 metros por debajo de laoriginal para sacar los fósiles desdeabajo y encontraron una continuacióvertical del cementerio. La barranca devarios metros de longitud estabaliteralmente repleta de Iguanodones quehabían quedado atrapados en ella no pudiendo escapar y quedando enterradosen el fango cuando la fisura se llenó coagua de las inundaciones. Con laautorización del culto director de lamina, las galerías fueron requisadasdurante tres años para que el equipo delaboratorio y los ayudantes pudiera

bajar bajo tierra y sacar bloques enterosde huesos envueltos por la marga yenviarlos al museo. A Louis Dollo, aúveinteañero, le fue encomendada la tareade preparación y descripción de losesqueletos a la cual se dedicó durante elresto de su vida. En 1883 había montadosu primer esqueleto y a finales del siglose habían añadido cuatro esqueletoserectos más; las criaturas se erguíasobre sus patas traseras y tenían los brazos delanteros libres para agarrarse alos árboles. Actualmente, el Museo Realalberga 31 esqueletos completos y parciales extraídos de la mina queforman una de las exposiciones de grupomás sensacionales del mundo. Esta

increíble colección de bestias recreadareunida en un lugar permitía un gracampo para el estudio comparativo delas variaciones dentro de una misma
especie de dinosaurio. Dollo, erealidad, pensaba que tenía dos especiesque se distinguían entre sí por el tamaño,sin embargo, estas diferencias probablemente se deban a diferenciassexuales. Curiosamente, los individuosque cayeron en el barranco eran adultoscompletamente desarrollados, y no habíadinosaurios jóvenes junto a sus padres.La fecunda imaginación de Dollo evocóel cuadro de un cementerio deiguanodones, hacia el cual se dirigiríalos animales decrépitos y seniles para

morir, al igual que nuestro míticocementerio de elefantes[23].
Los Iguanodones erainnegablemente unos de los dinosauriosmás afortunados en la historia de laespecie. Todos eran grandes, peroalgunos alcanzaban proporcionesrealmente gigantescas. La expedicióSinclair del Museo Americano a losdepósitos de carbón de Mesaverde deWyoming y Colorado, en 1937 volviócon huellas enormes de iguanodontessacadas del carbón. Las huellas, cadauna de poco menos de 90 centímetros delongitud, aparecían en espacios de 4,5metros, indicando que se trataba de unacriatura de pasos enormes. En base a

estas cifras se pensó que la misteriosacriatura erecta alcanzaría una altura de10 metros. Sin embargo, este dinosauriosigue siendo un enigma: su esqueletonunca fue encontrado[24].
b. El Iguanodon es uno de los dinosauriosmás conocidos, en gran parte debido a los

estudios realizados por Louis Dollo de losespecímenes de Bernissart.
No todos los herbívoros alcanzaba proporciones gigantes. Uno de losdinosaurios británicos más fascinanteses también uno de los más pequeños.Cuando por primera vez se encontrarofragmentos en los Wealden Beds en laisla de Wight, Mantell pensó que lacriatura debía de haber sido un animaloven, muy pequeño de Iguanodon.
Huxley, que estaba trabajando con mejor material lo identificó como udinosaurio adulto en miniatura por derecho propio y lo llamó ypsilophodon. Esta bestia sólo medía

entre 1,2 y 1,5 metros de longitud y, por esta razón, parecía aún más pequeño allado de sus contemporáneos como el guanodon. Sus grandes ojos y el primer dedo del pie que se podía oponer alresto de dedos hicieron pensar a OthenioAbel, en Viena en 1912, que este ágilherbívoro había saltado de rama erama, posándose en un árbol para comer frutas u hojas que cogía con las pequeñas patas delanteras. Debido a queel Hypsilophodon era un dinosaurio primitivo, con manos con cinco dedos y pies con cuatro dedos muy largos paraagarrarlas, Abel especuló que losorígenes de los dinosaurios se debíaencontrar entre los primitivos reptiles

bípedos, trepadores. Este dinosaurio quese posaba en las ramas de los árbolesnaturalmente interesó a GerhardHeilmann, cuya especialidad eran losorígenes de las aves, y en un principioestuvo inclinado a coincidir con Abel elos hábitos arbóreos del ypsilophodon. Sin embargo, demostró
que el primer dedo no era losuficientemente largo como paraoponerse al resto, de forma que eldinosaurio no pudo haberse agarrado alas ramas como un pájaro. Pensó quesería mejor buscar la ascendencia de losdinosaurios entre los ágiles reptiles bípedos que vivían en el suelo. Primerocomparó los pies del Hypsilophodon

con los de los monos y al propiodinosaurio con un canguro arbóreo, peromás tarde cambió de opinión y sacó al ypsilophodon completamente de los
árboles, ya que pensaba que estaba másadaptado a la vida en el suelo como susancestros. Sin embargo, la analogía coel canguro arbóreo había echado raíces, en 1936, W. E. Swinton reintegró al ypsilophodon a su antigua existencia
trepadora. Debido a que no era muyveloz, arguyó, el pequeño dinosaurio sehabía subido a los árboles pararefugiarse de los carnívoros. El ypsilophodon, actualmente es bastante
conocido debido a la existencia de 20esqueletos completos o parciales

encontrados en la isla de Wight. Elestudio minucioso de todos losesqueletos hizo que, en 1974, Peter M.Galton de la Universidad de Bridgeport, permitiera volver a salir al dinosauriode los árboles[25]. Heilmann tenía razócuando pensó que el dedo gordo no podía estar opuesto al resto de los dedos para posarse en las ramas, pero Swintose equivocó cuando creía que el ypsilophodon no podía correr. Debido
a que vivía en el suelo, debía de ser capaz de dejar atrás a los carnívoros yel estudio de Galton sobre sus miembrosestableció, tal como se describirá másadelante, que no sólo podía correr sinoque en realidad era muy rápido.

Durante el cretáceo tardío hubo uespectacular apogeo de grandesanimales ramoneantes de dos patas,entre los cuales el Hadrosaurus deJoseph Leidy sólo era un ejemplo demuchos. En realidad, el Hadrosaurus yel Trachodon estaban situados en elextremo más normal del espectro dehadrosaurios, ya que el grupoliteralmente explotó dando una seriediversa de formas poco después dedesarrollarse a partir de losiguanodontes. Los hadrosaurios eran ugrupo extremadamente afortunado que sedesarrolló en gran número, en el nortede América, hacia el final del largoreinado de los dinosaurios. Por

consiguiente, los hadrosaurios son unode los dinosaurios más conocidos. Losesqueletos frecuentemente eradesenterrados completamentearticulados y se ha encontrado más deuna «momia» envuelta por la pielencogida y tirante. Una de estasextraordinarias «momias» fue hallada por Charles Sternberg en Kansas e1908 y es, actualmente, uno de losobjetos expuestos más apreciados en elMuseo Americano de Historia Natural.Este cadáver de 70 millones de años deedad que yacía de espaldas fueencontrado con ambas patas estiradascomo si estuviese agonizando. Para quela bestia pudiera conservarse con la piel

los órganos intactos, el cuerpo tuvoque estar sometido a una particular secuencia de procesos antes defosilizarse. El animal, obviamente, nohabía sucumbido a un depredador ya quesu cuerpo estaba intacto; debió morir por causas naturales. El animal muerto
ació de espaldas expuesto al soldurante un largo período de tiempo en elcual se produjo la deshidratación.Mientras el cadáver se desecaba, la piel los músculos se encogieron alrededor
del cuerpo como la piel marchita de loscadáveres faraónicos, sobresaliendo lascostillas y los huesos de lasextremidades. Posteriormente, la«momia» deshidratada quedó

rápidamente enterrada, quizátransportada por las aguas de lainundación y cubierta de fina arenisca ysedimentos antes de que la piel se pudiera reblandecer. Las arenas yarcillas moldearon la frágil piel yacentuaron sus rasgos prominentes. Loque ahora poseemos no es la piel en sí,sino una impresión en la roca, si bien e perfecto detalle. Por el examen de la«momia», sabemos que el hadrosauriono estaba cubierto de escamas o placasóseas reptiles sino de un mosaico detubérculos córneos incrustados en lacurtida piel. Los tubérculos diferían etamaño a lo largo de la superficie deldinosaurio, con acúmulos de tubérculos

más grandes en la espalda expuesta alsol y las extremidades y otros más pequeños en el vientre[26]. Estasdiferencias de tamaño posiblementereflejen patrones de color opuestos. Noexiste ninguna razón por la cual losdinosaurios no pudieran haber desarrollado patrones de camuflajecomo los mamíferos que viveactualmente.
Los hadrosaurios herbívoros siduda pastaban, para lo cualdesarrollaron un ancho pico de pato, queen vida estaba cubierto por una fundacórnea. La presencia de este pico de pato, durante mucho tiempo hizo pensar que los hadrosaurios eran anfibios y

que, anticipándose a los patos, nadabao caminaban por las regiones pantanosas, tragando la blandavegetación acuática de las orillas delagua. Se pensaba que en vida presentaban una membrana de piel entrelos dedos de las manos que les ayudabaa nadar: estas membranas realmentefueron «vistas» en las «momias», aunqueestas observaciones siempre fuerodiscutidas apasionadamente.
La sorpresa ocasionada por la visióde un dinosaurio con pico fue eclipsada por el descubrimiento de unasextraordinarias protuberancias en lascabezas de muchos de ellos. CuandoCharles Sternberg volvió de la región de

Ciervo Rojo en Alberta en 1913, le presentó al mundo científico su primer hidrosaurio con cresta. Éste le fueentregado a Lambe del GeologicalSurvey de Canadá para su estudio quiele dio el nombre de Stephanosaurus,aunque más tarde fue rebautizado con elnombre de Lambeosaurus. El cráneo dela bestia presentaba una vistosa crestaque se alzaba en la coronilla como usombrero[27]. Más tarde resultó que loshadrosaurios adornados de esta manerano eran en absoluto raros. Prontosurgieron todo tipo de criaturasencapuchadas y rápidamente se hizomanifiesto que, en la época tardía delcretáceo, en Norteamérica, el grupo

había sufrido un período de evolucióacelerada. Aparecieron ornamentos muydiversos. Algunos como el Kritosaurus
tenían la cabeza plana; otros como elSaurolophus tenían sólidas bóvedas;otro grupo tenía «cascos corínticos» bulbosos como el Corythosaurus; yfinalmente existían los que tenían crestascaídas hacia atrás como los penachos delos sombreros de caballero como el
arasaurolophus. Todos estos tiposornamentados aparecieron hacia finesdel cretáceo y muchos de los exóticosdinosaurios crestados de pico de patofueron descubiertos en Alberta por elGeological Survey de Canadá y lasexpediciones del Museo Americano. E

1920, Lambe podía demostrar quemuchas de las capuchas eran huecas yque el conducto nasal tenía un tortuosotrayecto a través de la cresta, quefrecuentemente seguía estrechamente sestrafalario contorno, subiendo hasta la punta del casco o siguiendo el penachohasta el extremo posterior, antes devolver a la garganta[28]. ¿Por quéalgunos hadrosaurios tenían la cabeza plana como el Hadrosaurus y otros presentaban una cresta?
Esta cuestión intrigaba al baróopcsa, una de las figuras más
extravagantes que ha tenido la causa delos dinosaurios. A pesar de sus 99 publicaciones paleontológicas, muchas

de ellas relacionadas con losdinosaurios, Nopcsa es más conocido por su imperiosa obsesión por Albania,sus gentes, costumbres y geografía, yfueron sus estrafalarias hazañas eAlbania las que le valieron un puesto ela leyenda paleontológica. De hecho, talcomo observó una nota necrológica,
opcsa fue «casi rey de Albania».opcsa, al igual que tantas otras
personas brillantes, tenía un carácter extraordinariamente complejo, cuyofrágil estado mental en ocasiones parecía rayar en la locura. Antes de laPrimera Guerra Mundial, vivió como ulord baronial en Hungría aunque mástarde su interés se centró en la provincia

turca de Albania y pronto se convirtió eun experto en la cultura de este país. Alfinal de la guerra de los Balcanes e1913, Albania se convirtió en un peódel juego disputado por las fuerzasimperiales con los países satélites deunos y otros tras la retirada militar. Sedisputaba apasionadamente la cuestióde a quién debía pertenecer Albania.Los austrohúngaros apoyaban lafundación de un Estado independiente para satisfacer las peticionesnacionalistas, pero querían asegurarsede que éste tuviese una posturafavorable respecto al Imperioaustrohúngaro imponiéndole un monarcatítere de su propia elección. Nopcsa,

que poseía los antecedentes feudalesobligatorios y una extraordinaria pasió por el país, se consideraba a sí mismocomo el candidato más adecuado para el puesto de monarca. Escribió alcomandante en jefe del Ejército, pidiéndole quinientos soldados, unoscuantos cañones y dos buques de vapor rápidos. Después de establecer unacabeza de playa invadiría rápidamenteel país y se proclamaría a sí mismocomo el rey Nopcsa. ¡Posteriormentesencillamente se casaría con la hija deun multimillonario norteamericano paraconseguir el capital de inversiónecesario para poner a flote laeconomía! En su lugar, las fuerzas

imperiales instalaron al príncipeWilhelm zu Wied como gobernantetítere, al cual los albaneses tolerarousto durante seis meses antes de
forzarlo a escapar para salvar su vida.Durante la Primera Guerra Mundial,
opcsa sirvió en el Ejército del Imperioaustrohúngaro, no en un cargo ordinario,naturalmente, sino como espía. Se vistiócomo un campesino rumano, se dejócrecer el pelo y realizó una serie deosadas hazañas a lo largo de la fronterarumano-húngara. Simultáneamente fueescribiendo artículo tras artículo sobredinosaurios y reptiles fósiles. Despuésde la derrota, Nopcsa perdió la mayoríade sus propiedades feudales sin recibir

indemnización alguna, de forma que, aldisminuir sus recursos económicos,recurrió a la vida de paleontólogo profesional en el Geological Survey deHungría. Pero la arrogancia delaristócrata hizo inevitable el conflictocon sus colegas más humildes y Nopcsasólo pudo conservar el puesto durantetres años. Emprendió un viaje emotocicleta con su secretario por todaEuropa y recorrió miles de kilómetroshasta que se le acabó el dinero. En 1933le dio a su secretario una taza de té co polvos somníferos y después le disparóun tiro. Seguidamente apuntó con laescopeta a su propia boca y se pegó utiro[29].

En su vida tragicómica, como siviviera en otra esfera casi esotérica, fueentremezclando una efusión de artículos, publicaciones e incluso libros quetrataban sobre los pobladores reptilesde la Tierra mesozoica. Extrañamente,los artículos científicos de Nopcsa eramuy realistas y frecuentemente escritosen un estilo pedante. Nopcsa, elrenegado paleontológico, el inadaptado brillante, incluso arrogante, era básicamente un hombre de ideas. Así, e1929, sólo cuatro años antes de ssuicidio y temiendo ya otro colapsomental, el barón de Transilvania estudióel problema de los extraños dinosaurioscon casco y penacho con pico de pato de

Lambe. Othenio Abel, en Viena, ya habíasugerido que estos ornamentos cranealesdebían ser cuernos: la solución de
opcsa fue una nueva ampliación deesta idea. Lambe había dividido todoslos tipos en nuevos géneros y especies, pero Nopcsa replicó que lo único quehabía hecho Lambe era separar losmachos de las hembras. Loshadrosaurios, pensaba, presentabacomplicadas diferencias sexuales que,en los machos, se manifestaban por losdecorativos ornamentos óseos. Por lotanto, la «especie». Parasaurolophus
con penacho sólo era la versiómasculina del Kritosaurus de cabeza plana, Corythosaurus era el macho del

iclonius, etc. Nopcsa estaba dispuestoa eliminar a la mitad de especiesconocidas de hadrosaurios y lo hubiesehecho si sus ideas hubiesen tenidoalguna popularidad. Pronto extendió susrevolucionarias ideas a los carnívoros eiguanodontes. No le parecía preocupar en absoluto que en Bernissart por lovisto tenía (usando su criterio) un macho guanodon entre 23 hembras. ¡Esto erameramente un reflejo de la tendencia dereunirse en manadas y de la naturalezaaltamente polígama de los bípedosherbívoros[30]! Para hacer que elconjunto de la teoría fuera aún másinverosímil, se podría observar que ealgunas regiones siempre sólo se ha

encontrado hadrosaurios «hembra». Dehecho, se trataba de especies distintas,no puede haber dudas acerca de esto. Lateoría de Nopcsa de que la cresta eratotalmente ornamental, ignoraba lacaracterística más reveladora: que erahueca y albergaba un conducto nasalalargado. Estudiando la función de lacresta y especulando sobre los posibles beneficios de una nariz tan complicadase sacarían más conclusiones.
Debido a que se pensaba que loshadrosaurios eran criaturas nadadoras y buceadoras, se creía que la aperturanasal estaba situada en la punta de lacresta para actuar como tubo derespiración permitiéndole al animal

permanecer sumergido. En 1938, eAlemania, Martin Wilfarth inclusosugirió que la nariz y la cresta sólo eraáreas de adhesión muscular y que todaesta estructura había soportado unatrompa como la de los elefantes. Estedispositivo le permitiría al animalrespirar aire mientras se alimentaba eel fondo del lago, aunque también podríafuncionar como instrumento para coger alimentos como el elefante. El curiosodinosaurio de Wilfarth no sobreviviómucho tiempo. Tal como destacóSternberg, el desarrollo de una trompa prensil anularía completamente los beneficios derivados de un pico de patocórneo. Además, no existían cicatrices

en el cráneo fósil que sugirieran laexistencia de músculos en vida. Decualquier manera, ¿por qué desarrollar un conducto nasal tortuoso dentro delcráneo si en el exterior había una trompa para respirar [31]? De hecho, no habíaningún orificio nasal en la parte superior del cráneo; éste se hallaba en lasituación normal en la punta del hocico.Wilfarth también se había dado cuentade esto, pero eludió la conclusión obviaarguyendo que la apertura del hocico eraun vestigio ancestral cubierto de piel eel animal vivo mientras que el orificioverdadero se hallaba en la partesuperior del cráneo. De hecho, es fácilabandonar las bonitas teorías propuestas

por uno mismo si éstas no concuerdacon un inquietante hecho. El propioSternberg creía que, la curva en formade S que hacía el conducto nasal al pasar a través de la cresta, evitaba laentrada de agua en la garganta al actuar como un cierre de aire. Esta teoríatambién era insostenible por la simplerazón de que la presión de aguaalrededor del hadrosaurio sumergidohabría sido mucho mayor que la presióde aire en el interior de la cresta y elagua hubiese podido entrar. Aparte deesto, los hadrosaurios indudablementeusaban músculos voluntarios para cerrar la nariz, mecanismo de adaptació presente en todas las criaturas acuáticas

para evitar el ahogo.Para un dinosaurio acuático, que
respirase aire, el suministro auxiliar deaire sería una gran ventaja, permitiéndole al animal permanecer eel fondo del lago durante largos períodos. Así, no resulta sorprendenteque, en los años 1940, se desarrollarauna escuela de pensamiento alrededor de la idea de que los cascos y penachosóseos eran escafandras autónomasllevadas en la parte superior de lacabeza. Sin embargo, si medimos elvolumen de este suministro adicional deaire, éste constituye un insignificante 4%de la capacidad pulmonar en elCorythosaurus. Además, habría

existido grandes dificultades paratransferir este suministro de aire vital alos pulmones. La aspiración de este airea los pulmones hubiese creado un vacíoen el casco que sólo podría haber sidoaliviado mediante la entrada de agua siel animal estaba sumergido, o de aire siel animal estaba en la superficie, siendoambos mecanismos contraproducentes.En el primer caso, el dinosaurio sehabría ahogado y en el segundo, elescafandro se habría vuelto superfino.
La total insuficiencia de estasexplicaciones hizo que John Ostrom, profesor de geología en el MuseoPeabody de la Universidad de Yale,volviera a analizar de nuevo el

problema. Tras investigar el aparatonasal y su función en los reptilesmodernos, concluyó que algunasespecies habían desarrollado un sentidomuy agudo del olfato. Sin embargo, éstenunca habría sido tan agudo como en losmamíferos, en los que existecomplicados rollos de hueso revestidosde piel sensorial que multiplican variasveces las áreas de superficie sensitiva.Los hadrosaurios, aparentementeincapaces de plegar primorosamente la piel alrededor de estos rollos de hueso,habían logrado el mismo objetivosimplemente alargando los conductosnasales y ampliando los conductos edistintos puntos a lo largo de su trayecto.

Tanto los hadrosaurios como losmamíferos habían desarrollado un agudosentido del olfato mediante el aumentodel área total del epitelio sensitivo. Laresultante estructura en los mamíferosera compacta y podía ubicarsefácilmente en la nariz. En loshadrosaurios, los conductos tenían queenrollarse alrededor de la cabeza ydebían ser protegidos mediante crestasóseas[32]. ¿Por qué los hadrosaurioshabían desarrollado un sentido agudodel olfato? Ostrom supuso que estosherbívoros eran criaturas pasivas y,debido a que no poseían armadura,carecían de defensas contra los poderosos depredadores. En lugar de

ello habían logrado la capacidad deadvertir anticipadamente laaproximación de algún peligro,incorporándose sobre las patas traseras aspirando los olores transportados por
el viento. De esta forma podían alejarseincluso antes de que el carnívoro fueravisible. Probablemente este buen sentidodel olfato era necesario debido a la pérdida del método característico de losreptiles de detectar depredadores. Loslagartos apoyan sus mandíbulas en elsuelo para captar las vibraciones,transmitiéndolas a través de los huesosdel cráneo a los huesecillos del oído. Udinosaurio erecto, que se alzaba unos 3o 6 metros por encima del suelo, habría

perdido totalmente esta capacidad ycontaba únicamente con las ondas desonido que viajan a través del aire (umedio mucho menos eficiente para oír,especialmente cuando sólo existe uhuesecillo óseo en el oído como es elcaso de los reptiles: los mamíferosdisponen de tres huesecillos queamplifican las vibraciones). Con elsiempre presente peligro deltiranosaurio gigante, el pico de patodebía equiparse con algún sentidoalternativo igualmente eficaz para poder permanecer vigilante. No solamente ssentido del olfato era agudo; su vistatambién era excelente y cuando estabaapoyado sobre sus patas traseras, el

animal podía, al mismo tiempo, oler elviento y explorar el horizonte en buscade peligro. Estos dinosaurios pacíficos yvulnerables vigilaban constantemente la presencia de algún peligro y tenían muydesarrollada la excelente vista de los bípedos, presentando también un buesentido del olfato para poder escapar anticipadamente si aparecían loscarnívoros gigantes del día. Este nuevorazonamiento suscita una interesante pregunta: ¿Qué es lo que producía elolor característico de los dinosaurios si,en realidad, no producían ninguno deimportancia? El olor del sudor y de losaceites corporales en los mamíferos secapta desde lejos y puede ser

rápidamente detectado con un agudosentido del olfato. ¿Qué pasaba con losdinosaurios? ¿Realmente elTyrannosaurus producía aceites paralubricar su curtida piel que, sin querer,servirían para alertar a los dinosauriosherbívoros? Si éste no era el caso, sólonos queda pensar que el agudo sentidodel olfato del hadrosaurio funcionaba para detectar frutas tropicalesaromáticas o, como en el caso del agudosentido del olfato de los elefantes, paradetectar la presencia de agua.
Ostrom se dispuso a continuación areforzar la teoría, sacando totalmente aestos dinosaurios de pico de pato de ssupuesto hogar pantanoso y situándolos

en terreno seco donde, naturalmente, eltiranosaurio reinaba sin tener rivalalguno. En este medio ambiente, un bueolfato habría sido una clara ventaja.Ostrom aportó muchos argumentos paraapoyar su tesis. Las localidades dondefrecuentemente se han encontradoesqueletos de hadrosaurios presentaabundantes restos de plantas terrestres pero muy pocas plantas acuáticas. Loshadrosaurios vivían en las llanuras de lacosta situadas a poca altitud, cruzadas por grandes ríos serpenteantes ycubiertas de extensos bosques deconiferas. La estructura dentaria delhadrosaurio quizás era la másespecializada de todos los dinosaurios y

se adaptaba perfectamente a la materiavegetal leñosa, abrasiva. Los maxilares picudos del hadrosaurio presentaban unacompleja serie de muelas entrelazadas;las placas de molienda se componían dequinientos dientes en cada maxilar, deforma que en la boca había unos dos mil
dientes en total. Los dientes en forma de prisma eran comprimidos formando unamuela con una superficie de trituracióirregular. Estos dientes siempremuestran un alto grado de desgaste, y elhecho de que continuamente eraremplazados desde abajo, una vez rotos por el uso, también sugiere que losalimentos eran muy abrasivos. Desdeluego no se trataba de las suculentas y

blandas plantas acuáticas que habíasido sugeridas en un principio por el pico de pato del animal. Se había perfeccionado una complicada acciómasticadora de los maxilares para moler el material vegetal entre las baterías dedientes. A diferencia del movimientocortante y lateral de la vaca, lasmandíbulas se movían hacia delante yatrás una sobre la otra, de forma parecida a como lo hacen los actualesroedores. La teoría de Ostrom, de quelos hadrosaurios eran habitantesterrestres y de que ramoneaban losárboles coníferos se vio confirmadacuando descubrió un artículo, escrito e1922, acerca del contenido gástrico de

un Anatosaurus momificado, o «reptil pato», llamado así debido a su pico dehadrosaurio. Aparte de gran cantidad deagujas de coníferas, el animal habíaconsumido ramas, frutas y semillas.Debido a que estos hechos fuerodescubiertos durante el apogeo delhidrosaurio acuático, los resultadosfueron ignorados en aquella época. E1964, Ostrom volvió a centrar laatención en estos hallazgos como pruebaconcluyente de que los hadrosaurioshabían sido ramoneantes. Sin duda,multitudes de estas bestias había pasado sus días apoyados sobre sus patas traseras para pacer los árboles deconiferas y los álamos en los extensos

bosques costeros del cretáceo[33].La explosión de grandes
hadrosaurios erguidos en el cretáceotardío atrajo a una multitud dedepredadores, los cuales, asimismo,caminaban sobre sus patas traseras. Siembargo, en los herbívoros bípedos, lasextremidades anteriores nunca fueron ta pequeñas como en los carnívoros. Aligual que los ungulados (mamíferos co pezuñas) vivos, muchos de losdinosaurios herbívoros tenían pezuñas,lo cual sugiere que fácilmente podíahacer uso de sus cuatro patas[34],irguiéndose sobre sus patas traserasquizá para inhalar el viento o paraalcanzar las hojas más altas.

El descubrimiento de la primera bestia preadanita medible causo sensación yenfrento al mundo científico con un enigma.Se ve a Hoffman dirigiendo la operación en1770. Los trabajadores de las minas sacanlos imponentes maxilares, de 1,20 metros delongitud, del mosasaurio.


El estudio del Crystal Palace de WaterhouseHawkins, tal como lo debió ver la reinaVictoria durante su visita en 1853. La figuracentral es el Iguanodon, flanqueado por un Hylaeosaurus (derecha) y el mamífero delterciario Anoplotherium (izquierda). En un primer plano está el dicinodonte (derecha),ligeramente disfrazado de tortuga, y ellaberintodonto anfibio, restaurado tomandola forma de un sapo gigante (izquierda).

El Megalosaurus en una isla del CrystalPalace.

Teniendo como única opción viable laanalogía con el canguro, Cope representó aeste formidable Laelaps como dinosauriosaltador.

Aspecto que hubiera tenido el MuseoPaleozoico en Central Park si hubiese sidoacabado. A la izquierda, un Laelaps ataca aun Hadrosaurus, mientras que otros dos Laelaps se pelean sobre un cuerpo. En un primer plano está escondido el Elasmosaurus de cuello de serpiente. A laderecha se observan armadillos gigantes y perezosos terrestres. El museo debía ser unedificio grande de cristal con una estructura

de hierro, parecido al Sydenham CrystalPalace.
Los vestigios momificados del Trachodon,expuestos en el Museo Americano.


En 1920, Lawrence Lambe y Barnum Browndesenterraron una serie de curiososhadrosaurios con crestas y cascos.
El hadrosaurio de pico de pato canadiense Hypacrosaurus medía 7,6 metros delongitud y andaba, como la mayoría dedinosaurios bípedos, con la cola estirada.

Allosaurus, de la exposición del MuseoAmericano, que se inclina sobre la columnavertebral de un brontosaurio. Las vértebrasdel brontosaurio fueron desenterradasllevando las marcas de los dientes delalosaurio.

El Tyrannosaurus, cuyos maxilares de 1,20metros de largo estaban armados con dientesde 15 centímetros de longitud, era, sin duda,un predador implacable que matabaactivamente a sus presas.
A diferencia de un hadrosaurio

grande, que «sólo» pesaba tanto comoun elefante pequeño (aproximadamente 3toneladas), el Tyrannosaurus
probablemente superaba las 8 toneladas consiguientemente estaba construido
mucho más rígidamente. Las patastraseras eran macizas. El cinturó pélvico era ancho, permitiendo la vastasujeción de los músculos de losmiembros, y estaba sólidamente fundidocon la columna vertebral para soportar la tensión cuando el animal estaba de pie. Era en este punto donde el cuerpomantenía su equilibrio y el peso de 8toneladas era soportado únicamente por las patas traseras. Durante muchos años,las extremidades anteriores y manos

escaparon a los equipos del MuseoAmericano que buscaban en Montana.Cuando finalmente se recobraron losrestos, éstos resultaron ser taridículamente enanos que hacían pensar que las extremidades anteriores habíasido totalmente inútiles. Nadie podíaimaginarse función alguna para talesvestigios.
Las largas y fuertes piernas paracaminar elevaban la región de la caderade la bestia unos 3 metros por encimadel suelo; su rodilla se alzaría por encima de una cabeza humana. Econtraste, los débiles brazos medíaapenas 76 centímetros de longitud. Losmiembros delanteros del Allosaurus

carnívoro del antiguo jurásico eranotablemente más largos y fuertes,aunque en este caso también eratotalmente inútiles para caminar ydebían de haber sido usados únicamente para luchar o para despedazar las presas. Al menos, las largas y fuertesgarras hacen pensar que tenían sfunción ofensiva.
Los primeros restauradores delTyrannosaurus lo retrataron coextremidades anteriores que acababaen los tres dedos de costumbre. Algunasdécadas más tarde, cuando finalmenteaparecieron las patas delanteras, seobservó que sorprendentemente sólo poseía dos dedos, ambos en forma de

garra. Los miembros, de hecho, eradébiles y tan cortos que ni siquierallegaban a la boca de la bestia. Este gradepredador, el carnívoro terrestre másgrande de la historia de la Tierra, habíaaparentemente prescindido de susextremidades anteriores, dejando lafunción de matar a sus contemporáneosgigantes a sus maxilares y garrastraseras[35]. Así, el último carnívoro deeste tipo había desarrollado latendencia, ya observada en susantecesores, los allosaurios ymegalosaurios, a su máxima expresión.El tiranosaurio era totalmente incapaz deadoptar una postura normal cuadrúpeda estaba obligado a mantener el

equilibrio sobre sus enormes patastraseras. Este gigante se balanceabasobre el cinturón pélvico, utilizando scola para compensar el peso de lacabeza y del tronco cuando caminaba.Probablemente, de vez en cuandodescansaría sobre sus piernas utilizandosu cola como apoyo. El ligeramente másantiguo Gorgosaurus muestra uestrecho parecido con elTyrannosaurus, especialmente por poseer sólo dos dedos en la mano.Lambe representó a esta bestiacaminando con una clara inclinación, elugar de adoptar la imponente posturaerguida, atribuida siempre alTyrannosaurus. En las articulaciones de

las piernas también encontró señales deque, en vida, había sido estevado; postura que difícilmente se podríaconvertir en una posición majestuosa[36].Aunque, por el tamaño y peso taenormes del Gorgosaurus, el pasoinclinado con la cola levantada por encima del suelo a modo decontraequilibrio debía de ser másrentable pero también debía ser agotador. La práctica pérdida de lasextremidades anteriores en elGorgosaurus y Tyrannosaurus
demuestra que irrevocablemente estabaobligado a mantener una postura bípeda.¿Pero realmente las extremidadesanteriores rudimentarias eran tan inútiles

como se pensaba en general?Extrañamente, estos diminutos miembros presentan garras relativamente largas, locual resulta inexplicable siverdaderamente carecían de funcióalguna. Pero resultaba incluso aún mássorprendente, que el cinturón pectoralfuera grande lo cual indicaba que los brazos estaban bien dotados demúsculos. Esto es muy sugestivo: el brazo no podía ser tan inútil como sehabía pensado anteriormente.
En una radical reinterpretación delTyrannosaurus, Barney Newman delMuseo Británico ha sugeridorecientemente que los brazos, erealidad, desempeñaban un papel

importante en la vida de la bestia.Aunque el brazo recubierto de carnetenía sólo el grosor de un muslo dehombre, era lo suficientemente fuertecomo para ayudar al tiranosaurio alevantarse del suelo después de haber descansado. El dinosaurio agachado, deocho toneladas de peso, plegaba sus patas traseras debajo de su cuerpo comohacen los pollos. Estiraba sus diminutos brazos hacia delante y bajaba la cabezade forma que sus maxilares reposaran eel suelo tal como en ocasiones hacen losgatos. Cuando la bestia se levantaba,cualquier intento de levantar los cuartostraseros extendiendo las patas traserassólo habría provocado que la cabeza

arrastrara por el suelo. Es aquí dondeayudan las extremidades delanteras. Lasgarras flexionadas se usaban para dirigir los esfuerzos de las patas traseras paralevantar los cuartos traseros en vez deempujar el animal hacia delante. Acontinuación, y al igual que las vacas, eltiranosaurio se esforzaba para alzar suscuartos delanteros aún postrados en elsuelo. El pequeño tamaño de losmiembros anteriores significaba que eltiranosaurio estaba obligado a tirar scabeza hacia atrás mientras enderezabasus piernas. Con las patas traserascompletamente extendidas eltiranosaurio ya podía marchar a cazar asus presas. Al igual que el Gorgosaurus,

en la nueva representación de Newman,el Tyrannosaurus caminaba inclinado.Sugiere que el cuerpo se mantenía euna posición horizontal, en lugar deestar inclinado 45 grados como en lasrestauraciones convencionales. Lasarticulaciones flexibles de las vértebras permitían un cuello curvado como «loscuellos de cisne» y hacían posible lamaniobrabilidad tan esencial como unamáquina de matar de este tipo. Newmaincluso cortó 3,6 metros de la cola deltiranosaurio para obtener una criaturamás simétrica. En realidad, esto no estan arbitrario como pueda parecer yaque la existencia de estos 3,6 metros decola sólo eran conjeturas, no existe

especímenes en los que realmente sehaya podido comprobar este hecho. ElTyrannosaurus presentaba ahora unadistribución del peso más equilibrada etodas partes. En vez de arrastrar lascolas por el suelo detrás de sí, donde nohubiesen podido actuar comocontrapeso, los Tyrannosaurus laslevantaban del suelo para reducir elesfuerzo de sostener el tronco. La ideadel elegante movimiento atribuido eotros tiempos al rey de los asesinos fuedestruida de golpe por el estudio de
ewman sobre las articulaciones de losmiembros. Cuando la pierna era alzadahacia delante, los tendones que llegabahasta los dedos de los pies

automáticamente se estiraban y losdedos se cerraban como los de un pájaroen una percha. Esto le permitía al piesaltar por encima del suelo. Cuando la pierna libre avanzaba, balanceaba senorme cola hacia un lado, haciendo quela marcha del monstruo fuera torpe, parecida a la de los patos. Unas huellasde megalosaurio de Swanage, ahoraexpuestas en el Museo Británico,muestran cómo los dedos están dirigidoshacia dentro. Al igual que elmegalosaurio, el Tyrannosaurus
probablemente tenía los pies torcidoshacia dentro.

c. Primer dibujo del Tyrannosaurus rex, elcarnívoro terrestre más grande de la historiade la Tierra, realizado en 1906.
Inmediatamente se puede apreciar, sise echa un vistazo al anadeo delTyrannosaurus, que el solo esfuerzo de

mantenerse en pie debía de ser enorme.El solo hecho de balancear un cuerpo deocho toneladas sobre las patas traserasdebía requerir mucha energía, energíaconsumida por los músculos de las piernas, cola y vértebras para oponersea la gran fuerza gravitacional. Adiferencia del lagarto, que correhaciendo un breve esfuerzo antes dedejarse caer sobre su vientre pararecuperarse, el Tyrannosaurus no setumbaba con el vientre en el suelo sinoque caminaba con sus piernasescondidas debajo del cuerpo y elvientre levantado al aire. No estabahecho para dejarse caer sobre el vientre.
ormalmente, se veía obligado a

permanecer equilibrado sobre sus pies.Ésta es justo la situación que nosencontramos en los mamíferos y lasaves. Aproximadamente, de la mitad alos tres cuartos de la vida activa de losmamíferos o aves transcurre en estadoerguido. Lo mismo probablemente eraválido para el Tyrannosaurus (y paratodos los dinosaurios). Indudablemente,el tiranosaurio podía ahorrar algo deenergía apoyándose sobre sus piernas.Pero incluso así no se parece en nada ala posición de reposo total de un lagartodespatarrado. El problema naturalmente parece aún mayor si tomamos comoejemplo a un titánico brontosaurio:cuarenta o cincuenta toneladas de carne

destinadas a permanecer de pie durantegran parte del tiempo que el animalestaba despierto para continuar la búsqueda de alimentos. La energíanecesaria debía de haber sido enorme.La energía consumida por los músculos para mantener firmes las articulacionesde los miembros y la columna vertebral,manteniendo al animal erguido sobre sus pies no debe ser subestimada. Losmúsculos deben tener un tono constante para evitar que el esqueleto se colapse;cuanto mayor es el peso del animal,mayor es el esfuerzo. Un lagarto tiene poco de qué preocuparse ya que pasamuy poco tiempo erguido, más bieahorra energía colapsándose en el suelo.

Tiene que hacerlo ya que no tiene la
energía para permanecer erguido. ¡Estáobligado a pasar hasta un 90% de svida «activa» tendido inmóvil en elsuelo!
John Ostrom fue el primero ecomprender las implicaciones de estehecho, implicaciones que sugerían quedebíamos volver a replantear algunas denuestras ideas convencionales. En 1969se pronunció en contra de la aceptadaidea de que los dinosaurios eran reptilesde sangre fría y perezosos. La ocasióera apropiada, ya que fue en la PrimeraConvención Paleontológica
orteamericana, que tuvo lugar en elMuseo Field de historia natural,

Chicago, en 1969. El tema escogido por Ostrom era la interpretació paleoclimatológica a partir de fósilescontemporáneos. Previamente se habíadado por supuesto, casi sin haber analizado críticamente la cuestión, deque la presencia de los dinosauriosgigantes de sangre fría —dependientescomo eran del sol— en latitudes altasimplicaban la existencia de un climatropical uniforme en una gran parte de laTierra en la época mesozoica. Es verdadque los reptiles y anfibios son buenosindicadores térmicos, pero Ostrorehusó utilizar dinosaurios para este fin.Los mamíferos y las aves, destacóOstrom, tienen un gran gasto de energía.

Una rata, por ejemplo, quema diez vecesmás combustible por unidad de volumeque un caimán; el músculo del colibríutiliza quinientas veces más oxígeno queel músculo de una rana del mismotamaño, incluso cuando ambos están eestado de reposo. Estas cifrasdemuestran gráficamente la cantidad deoxígeno consumida para producir lavasta cantidad de energía requerida por los mamíferos y las aves, energía que e parte es utilizada para mantener internamente una temperatura corporalconstantemente alta. Con un alto índicemetabólico y una rápida producción deenergía, los mamíferos y las aves socapaces de mantenerse y caminar

erguidos durante casi todo el tiempo queestán en estado de vigilia. Paraayudarles, los miembros han sidotrasladados debajo del cuerpo de formaque puedan mantenerse erguidos sobre
sus piernas sin estar colgados entre ellascomo en los lagartos. De esta forma, loslagartos pasan la mayor parte de su vida pasivamente mientras que los mamíferos
las aves realmente la pasan de formaactiva, estando de pie, caminando,limpiando, cazando presas,sometiéndose a ritos, etc. «Lacorrelación entre una elevadatemperatura corporal…, un elevadometabolismo y una postura y locomocióerguidas no es accidental —dijo Ostro

—. La evidencia parece indicar que la postura y la locomoción erguidas probablemente no son posibles sin umetabolismo elevado y una alta yuniforme temperatura corporal[37]». Ylos dinosaurios andaban erguidos:manadas de Triceratops con postura derinoceronte eran cazados en las llanurasmesozoicas por carnívoros bípedos detamaño gigante similar. Estosdominantes reptiles mesozoicos probablemente compartieron el secretodel éxito de los mamíferos.Probablemente también eran animales desangre caliente. Como corolario, lamarcha de los dinosaurios a latitudesmás frías del norte, a las que podía

hacer frente con su propio suministro decalor, hace que prácticamente seainservibles como indicadorestérmicos[38].
La prudencia de la postura deOstrom se hizo patente, en 1973, cuandoDale Russell, del Museo Nacional deCiencias Naturales en Ottawa, anuncióel descubrimiento de hadrosaurios de pico de pato dentro del círculo ártico
cretáceo del Canadá (situado a la alturadel Yukón[39]). Las cicádidas y cipresesfósiles indican que en aquella épocadebía de haber un clima más cálido en elártico. No obstante, los meses deoscuridad invernal dejaban a losdinosaurios dos opciones: existir sin sol

o migrar hacia climas soleados. Ambasson difíciles de explicar usando elmodelo de dinosaurio arcaico. Si losdinosaurios eran animales que tomabael sol para calentarse, ¿podríasobrevivir varios meses de noche perpetua? Como alternativa, losdinosaurios de pico de pato, que everano pastaban en el Yukón debían dehaber viajado 800 kilómetros paraalcanzar el borde ártico del cretáceo.(Debido a que la oscuridad inhibe elcrecimiento de las plantas, probablemente marcharían aún más lejos para satisfacer su voraz apetito). ¿Dedónde sacaban la resistencia para lasarduas migraciones de otoño? El

descubrimiento de Russell apoyafuertemente los datos de que losdinosaurios no eran en modo algunoanimales exoteros tradicionales.
Nadie antes había sugerido unainextricable relación entre metabolismoelevado, temperatura estable y posturaerguida, pero una vez expuestaexplícitamente, esta relación parecíaobvia y natural. Resolvía las viejascontradicciones inherentes al absurdomodelo del brontosaurio tomando el sol,descartando el conjunto del modelo ysustituyéndolo por un dinosaurioendotérmico. Naturalmente, estorequiere una apreciación radicalmentenueva de la fisiología de los

dinosaurios; y nos vemos obligados atener como referencia para nuestromodelo a los mamíferos y las aves. Los pulmones de los lagartos son capaces deextraer mucho menos oxígeno del aireque los de un mamífero, donde lostejidos pulmonares son esponjososdebido a una multitud de vesículasdiminutas con paredes ultrafinasdiseñadas para exponer una gran área desuperficie al aire. En los lagartos, estas bolsas de aire son mucho más grandes yel área de superficie estácorrespondientemente reducida. Por lotanto, la sangre recibe menos oxígeno,defecto que además, tal como hemosvisto, es potenciado por un corazó

menos eficaz. El corazón del lagartotiene un solo ventrículo, que sirve para bombear tanto la sangre que va delcuerpo a los pulmones como la quevuelve de los pulmones y va al cuerpo.Debido a que la sangre de estos dossistemas tiene que pasar por una únicacámara, se mezcla: así, la sangre rica eoxígeno que llega de los pulmonesqueda adulterada con la sangre venosaconsumida antes de ser bombeada a losmúsculos donde es requerido el oxígeno.Los mamíferos y las aves presentan uventrículo dividido en dos cámaras, una para recibir específicamente la sangreconsumida recibida de los tejidos y bombearla a los pulmones; la otra está

diseñada para enviar sólo sangreoxigenada no adulterada, recibida de los pulmones, de vuelta al cuerpo. Esto proporciona una gran cantidad deoxígeno esencial necesario para lascriaturas de metabolismo rápido, desangre caliente. Probablemente, éste fueel sistema empleado por losdinosaurios. De hecho, el cocodrilo, que podría ser considerado un vestigio delancestro de los dinosaurios,funcionalmente presenta un corazón decuatro ventrículos, con un septoventricular que divide los sistemasarteriales y venosos.
El alto nivel de metabolismo yrápida producción de energía supuestos

en el dinosaurio, no lo sitúan junto a loslagartos de sangre fría, sino posiblemente junto a los mamíferos y lasaves de sangre más caliente. RichardOwen hubiese estado sorprendido yencantado. Sorprendido, de que un reptilno solamente tuviera algunascaracterísticas comunes con losmamíferos sino que incluso fuera desangre caliente; y encantado por laespectacular confirmación de su teoríade que el dinosaurio realmente era lacumbre de la creación reptiliana.

III. LA CARRERAES PARA LOSVELOCES, LA
LUCHA PARA LOSFUERTES
Esta, aún muy controvertida, ideainmediatamente comenzó a tener partidarios que veían en ella uno de losconceptos más fructíferos y productivos.Sugería hacer nuevas preguntas y plantear nuevas formas de abordar lacuestión que, inesperadamente,mostraban al dinosaurio bajo un punto

de vista completamente nuevo.Por una notable coincidencia, este
nuevo enfoque había surgidosimultáneamente al otro lado delAtlántico. Armand de Ricqlés en laUniversidad de París había llegadoindependientemente a la mismaconclusión de Ostrom en unos pocosmeses aunque su modo de abordar el problema había sido bastante distinta.(Este tipo de feliz coincidencia es unacaracterística recurrente de losdescubrimientos científicos. En ciertasépocas, parece que las ideas nuevasfloten en el aire esperando que alguietenga el valor de cogerlas; recuérdese lalucha entre Newton y Leibniz acerca de

quién inventó el cálculo, o el sobresaltode Darwin cuando recibió un resumede Wallace acerca de su propia teoría dela evolución mediante selecciónatural). Así, en 1969, después de haber estudiado la estructura interna demuchos tipos de huesos fósiles yrecientes, Ricqlés, sugirió quefisiológicamente los dinosaurios eramás parecidos a los mamíferos que a loslagartos. El hueso, aparte de su obvio papel como andamio interno, es unaestructura activa, en crecimiento yfisiológicamente importante: en lamédula se fabrican células sanguíneas yel tejido óseo actúa como un importantedepósito de calcio (las hormonas

desencadenan la liberación del calciodel hueso y lo envían a los lugares de lacontracción muscular donde desempeñaun importante papel). Los animalesactivos con un alto nivel metabólico ysistemas fisiológicos aceleradosexigirán un mayor rendimiento de loshuesos, lo cual se reflejará en su finaestructura. Ricqlés se dio cuenta de quela estructura ósea era un sensibleindicador del nivel de actividad de uanimal y que, en los huesos fósiles,disponíamos de todos los ingredientes para evocar la fisiología de los animalesmuertos hacía mucho tiempo. En losmamíferos y las aves, los vasossanguíneos que penetran en los huesos

largos son numerosos y estádensamente agrupados (permitiendo umayor transporte de materiales hacia ydesde regiones activas); en los reptiles,los vasos sanguíneos son escasos. Losconductos de Havers, los lugares quecontrolan el intercambio de calcio entreel esqueleto y la sangre, son menosnumerosos en los huesos de los reptilesque en los huesos de los mamíferos, y sfalta limita la velocidad de intercambio.Ricqlés afirmó que, los huesos de los
dinosaurios se parecen a los huesos de
los mamíferos, pero que sediferenciaban claramente de los huesosde reptiles y anfibios de sangre fría.Interpretó esta sorprendente

«convergencia» como prueba de laexistencia de un metabolismo activo, yque los dinosaurios posiblemente eraanimales de sangre caliente[1].
Estos estudios independientes deOstrom y Ricqlés convirtieron a losmamíferos en el nuevo paradigma paralos dinosaurios, eliminando así laslimitaciones asociadas al obsoletomodelo de lagarto. Estos nuevosmodelos centraron la atención sobreaspectos hasta entonces inexplorados delos dinosaurios, aspectos que nuncahubiesen salido a la luz bajo el antiguoorden. La tarea de estudiar a fondo todasestas nuevas ideas estuvo en manos deun estudiante de Ostrom en Yale, Robert

T. Bakker. Bakker se llevó consigo estasradicales ideas al Museo de ZoologíaComparada en la Universidad deHarvard y fue aquí, en el museo deLouis Agassiz, donde analizó lasimplicaciones de la endotermia para losdinosaurios en una serie de artículosescritos a principios de la década de1970. Esta nueva forma de abordar el problema también provocó que los problemas existentes fueran analizadosdesde una nueva perspectiva.Frecuentemente estos problemas había pasado inadvertidos y las paradojashabían sido aceptadas sin levantar crítica alguna. Las desventajasfisiológicas de los lagartos no podía

ser ignoradas, y aunque se reconocía quelos reptiles vivos están impedidos por su limitada capacidad para correr oincluso para mantener una posicióerguida, se suponía que los dinosauriosrealizaban prodigiosas proezas de grafuerza y resistencia. Aun a pesar de queestas limitaciones se habríaincrementado desproporcionalmente eun lagarto del tamaño de un elefante, secontinuaba pensando que estosformidables carnívoros se había pasado la vida persiguiendo activamentea sus presas, mientras que, a su vez, losherbívoros habían sido capaces deescapar de sus garras. Si un pequeñolagarto, que pesa pocos gramos,

solamente es capaz de tener breves períodos de actividad, ¿cómo ucarnívoro de ocho toneladas de pesocomo el Tyrannosaurus habría podidoaguantar una lucha feroz y prolongadacon su presa? La muerte de uTriceratops en las garras deltiranosaurio no podía haber sido tainstantánea como lo es la de una moscaatrapada por un lagarto. Un dinosauriogrande tenía un volumeaproximadamente un millón de vecessuperior al de un lagarto pequeño, y aúasí se creía que éste había cazado sus presas y mantenido combates a muerte.¿De dónde sacaban la energía?
Una forma de salir de esta paradoja

era suponer, como hicieron algunoscientíficos en el caso de los gigantescarnívoros, que eran demasiado grandes pesados como para haber cazado
activamente sus presas y luego haber luchado. Lawrence Lambe, eldescubridor del monstruo canadienseGorgosaurus, era el más ferviente partidario de esta teoría. Eldescubrimiento de un nuevo dinosauriocarnívoro siempre es un acontecimiento.Los carnívoros no eran ni con mucho taabundantes como sus inofensivoscontemporáneos herbívoros en la épocamesozoica, pero al ser bestias taimponentes, su escaso número pronto seveía compensado por el interés que

suscitaba en sus descubridores.Frecuentemente, una comunidad de unadeterminada localidad tendrá una seriede herbívoros, pero sólo a un únicocarnívoro gigante para tenerlos a raya.Al explorar la región canadiense del ríodel Ciervo Rojo de Alberta, a principiosdel siglo, se descubrieron muchosherbívoros nuevos, y en 1913 Lambeentró en posesión de un carnívorocompletamente nuevo. El Gorgosaurus
era un gigante del cretáceo tardío:alcanzaba una longitud de 8,8 metros yuna altura de 3,3 metros. Aunque estabaarmado con los obligatorios dientes ymaxilares fuertes, el gorgosaurio teníauna constitución más ligera que el

Tyrannosaurus, con patas traseras másdelgadas y más gráciles. Con todo,obviamente pesaba algunas toneladas yLambe pensaba que la criatura había pasado la mayor parte de su vidatumbada en el suelo. Podía caminar erguida si la situación lo requería, y eeste caso la cola actuaba comocontrapeso al tronco y el peso delcuerpo era soportado por las patastraseras. Sin embargo, esto sucedía muyraras veces, provocado sólo por umotivo muy fuerte. En posició postrada, sentada o agachada, el pesodel gorgosaurio pasaba directamente alsuelo a través del cinturón pélvico.«Esta posición de reposo —aventuró

Lambe—, y particularmente la posturacon el cuerpo acostado en toda slongitud (cuando la criatura estabacompletamente estirada con la cabezaapoyada sobre el suelo) probablementeeran las más frecuentemente adoptadas por un reptil con la forma, y naturalezasupuestamente perezosa delgorgosaurio». Extrañamente, los largosdientes aserrados en los maxilares delgorgosaurio estaban completamentenuevos sin estrenar, a pesar de que elejemplar no era joven. Las puntas de loscolmillos eran casi perfectas, codelicados bordes dentados en los agudosfilos cortantes. Lambe dedujo que losalimentos ingeridos por este carnívoro

eran blandos y no abrasivos, y que la presa no podía haber ofrecidoresistencia alguna.
Por consiguiente, se cree queel gorgosaurio se limitaba aalimentarse de los cadáveres deanimales que no habían sidomatados recientemente, que noera un intrépido cazador sino unanimal que se alimentaba decarroña que desempeñaba su papel en la Naturaleza y sin dudasus servicios eran requeridos amenudo, si tenemos en cuenta lainmensa cantidad de tracodontes,ceraptosianos, estegosaurios y

otros dinosaurios y reptiles quevivían y morían en este período particular del cretáceo[2].
Sumergido, como Lambe creía, emontones de potencial alimento que sedesplazaban lentamente, el Gorgosaurus
no necesitaba ser un depredador activo.A su alrededor debían de haber muertode forma natural todo tipo deherbívoros. La bestia engullía «loscadáveres que encontraba o con los quese tropezaba en sus excursiones provocadas por el hambre». Elmonstruoso gorgosaurio era un animalde carroña casi inofensivo. Arrancaba

los blandos tejidos del cadáver con susgarras antes de agacharse sobre la carneen proceso de descomposición paraconsumir los sabrosos tejidos. Almenos, eso es lo que pensaba Lambe.
Desgraciadamente para la teoría deLambe, los dientes se pierden odesgastan, no durante el acto de matar,sino al arrancar la carne de los huesos.El Museo Americano tiene el esqueletocompleto de un brontosaurio, cuyoshuesos de la cola han sido roídos por ualosaurio. Al comparar un maxilar dealosaurio con las muescas de los dientesse vio que éstas encajaban exactamente.Obviamente, un Allosaurus había tenidoun banquete con el cadáver del

brontosaurio. Es más, durante este proceso había perdido varios dientesque fueron encontrados al lado de lacola del brontosaurio cuando fueextraída de las rocas[3]. Resultaincomprensible por qué los dientes delgorgosaurio canadiense están comonuevos, ya que por el mero hecho decomerse el cadáver se hubiesedesgastado. Naturalmente, esto no puedeser utilizado como prueba parademostrar un comportamiento cobardedel gorgosaurio. La serie de armamentosque llevaba era tan temible como la deun alosaurio o tiranosaurio. Laalimentación a base de carroña parecehaber sido una ocupación poco probable

para esta bestia; tiene el aspecto comosi estuviese hecho para la agresióactiva. Además, la mayoría de losinvestigadores que estudiaban a losgrandes carnívoros como el Allosaurus
Tyrannosaurus de Norteamérica y elTarbosaurus, el homólogo mongol deltiranosaurio, no compartían elescepticismo de Lambe, y en lugar deello preferían creer en la evidencia delas garras y maxilares, éstas indicabaque se trataba de formidables asesinos.En la vida (y la muerte), naturalmentelas cosas nunca están tan claras: un leó puede alimentarse de carroña y unahiena en ocasiones matar. Sería pocorealista imaginarse a un gorgosaurio o

tiranosaurio renunciar a una comidagratuita si se encontraran con un animalmuerto. No obstante, parece ser más probable que, al igual que los leones, setratara principalmente de depredadoresactivos.
El Tyrannosaurus rex, el «rey de loslagartos tiranos», era la expresióextrema de su tipo. Su impresionantetamaño, de poco menos de 12 metros delongitud si aceptamos la estimaciórevisada de Newman, y 6 metros dealtura cuando el animal estaba erguidosobre sus patas traseras, no deja lugar adudas de que se trataba del reymesozoico de las bestias. Fueencontrado por primera vez como

resultado de un intenso período deactividad en el Museo Americano afinales del siglo pasado. Bajo la presidencia de Henry Fairfield Osborn,una gran cantidad de expediciones bieequipadas fueron enviadas a lasMontañas Rocosas, que como resultadohicieron descubrimientos tan prolíficosque aún ahora estamos cosechando los beneficios. El Tyrannosaurus surgió desu tumba de Hell Creek en Montana, e1902, cuando el equipo de BarnuBrown estaba explorando la región e busca de dinosaurios. Brown,acompañado de Lull, localizó el cráneo,maxilares, columna vertebral y patastraseras de la bestia, pero la dura piedra

caliza circundante del cretáceo superior obligó a los equipos del museo a volver durante dos o tres años consecutivos para desenterrar por completo al animalmientras que Lull, ya de vuelta en elmuseo, preparaba el esqueleto paramontarlo. El Tyrannosaurus sorprendióa todo el mundo. El Allosaurus, a pesar de su gran tamaño, parecía pequeño allado del más grande de todos lostiranos. Un colega de Osbom en elmuseo, William Diller Matthew, intentóhacer un dibujo preliminar de la bestia y para destacar sus dimensiones de laforma más dramática, la colocó junto aun diminuto esqueleto humano. Esteespécimen, y gran parte de otros dos

esqueletos hallados en las sucesivasincursiones a Hell Creek, fueroestudiados por Osborn, quien, ereconocimiento a esta criatura que habíasido el carnívoro terrestre más grandeque jamás pisó la Tierra, la bautizó coel nombre de Tyrannosaurus rex[4]. Unacriatura, además, que estaba construidasólidamente e indudablemente paraluchar, con largos dientes letales y unosterroríficos maxilares y unas garras de20 centímetros de longitud que armabalos dedos de los pies. Al igual queCope, que al hallar al Laelaps carnívoroen la misma área que al hadrosaurio, sehabía visto obligado a considerar aambos como enemigos, Osbom sugirió

que el tiranosaurio había sido elenemigo implacable del Triceratops
cornudo. La armadura del herbívorotricornio había alcanzado tal grosor queel Tyrannosaurus había desarrolladodientes de 15 centímetros de longitud para penetrar en ella y unos maxilaresde 1,20 metros de longitud para poder engullir los pedazos de carne.
El estudio del cráneo delTyrannosaurus revela sus hábitosdepredadores. Debido a que las patasdelanteras no servían para desgarrar lacarne de la forcejeante presa, el cráneohabía asumido la función principal deagresión. Consiguientemente, habíacrecido desproporcionalmente e

comparación con el tamaño del animal yestaba armado con una serie de dientescurvados, aserrados, parecidos a lanzas paleolíticas. La longitud de susmaxilares permitía una mordedurarealmente efectiva y el Tyrannosaurus
era así capaz de engullir considerablesmasas de carne de dinosaurio. El
llosaurus, más pequeño incluso,superaba al tiranosaurio en su capacidadde tragar cantidades considerables de presas. En la región de la sien, el huesocuadrado que se articulaba con elmaxilar inferior era ligeramente móvilmientras que los dos maxilares y eltecho del cráneo podían flexionarse esus puntos centrales. Todo ello

constituía una serie de adaptacionesnotables que hacían que la boca se pudiera abrir más y el Allosaurus
pudiera engullir vorazmente enormesmasas de carne.
Dos de los tiranosaurios rescatados por Barnum Brown en Montana, fuerorestaurados en la sala cretácea delMuseo Americano en Nueva York. Las posturas adoptadas por Osborn para susreconstrucciones eran las dibujadas por el director del departamento de reptilesdel parque zoológico de Nueva York,tras un intensivo estudio de losmovimientos de los lagartos. Estasrestauraciones, dijo Osbom,representaban «animales antes del salto

convulsivo y mordedura que distingue lalucha de los reptiles[5]».
Barnum Brown describió la escenamás vivamente.
Son las primeras horas de lamañana en las orillas de un lagodel cretáceo hace tres millonesde años. (En la época de BarnumBrown —este artículo fueescrito en 1915— se le concedíaun período mucho más corto a laduración de la historia de laTierra: actualmente se cree quedesde el final del cretáceo hantranscurrido 64 millones de

años). Un dinosaurio herbívoro,un Trachodon, que se haaventurado fuera del agua paraconseguir un desayuno a base devegetación suculenta, ha sidoapresado y devorado parcialmente por unTyrannosaurus gigante,carnívoro. Mientras estemonstruo se agacha sobre elcadáver, descuartizándoloatareadamente, otroTyrannosaurus es atraído a laescena. Se aproxima irguiéndose para agarrar al más afortunadocazador y disputarle la presa. Lafigura agachada, de mala gana

deja de comer y acepta eldesafío levantándose parcialmente para saltar sobre suadversario.
El momento psicológico deesta tensa inercia, antes delcombate, fue escogido paramostrar mejor las posiciones delos miembros y de los cuerpos y para describir un incidente en lahistoria de la vida de estosreptiles gigantes[6].
Cope había sentado el precedentecolocando a sus Laelaps agarrados unoa la garganta del otro, de modo que

Brown le siguió el ejemplo enfrentandoa un tiranosaurio con otro. Sin embargo,esta agresión intraespecífica pareceimprobable. El hecho de que muchosautores hayan asumido que las limitadascapacidades mentales de los dinosauriosdepredadores hacían que éstos atacaratodo lo que se moviera, es razón de más para creer que existía algún tipo decomportamiento territorial entre loscarnívoros más grandes, similar al queactualmente existe entre los leones. Losdinosaurios depredadores más pequeños probablemente cazaban en manadascomo los lobos[7], existiendo en estecaso muy probablemente algún tipo deerarquía —una jerarquía social de los

dinosaurios— para evitar que se produjeran estas reyertas destructivasque Cope y Brown habían imaginado.
Brown pensaba que elTirannosaurus era «una criatura poderosa, activa y de movimientoságiles si la ocasión lo requería». Dehecho, cuando Brown desenterró uesqueleto de Gorgosaurus en la regiódel río del Ciervo Rojo en Alberta, unadécada más tarde, lo envió al museo para que fuese restaurado en una posturacomo si estuviese corriendo, comocazando una manada de dinosaurios de pico de pato[8].
Osborn y Matthew, conscientes de lalimitada capacidad del lagarto para

mantener una agresión activa, eran másreservados en sus afirmacionesconcernientes a las agresivas luchas yagilidad de estos carnívoros de gratonelaje. Los lagartos matan con rapidez,una embestida y el destino del insectoestá sellado, y Matthew se imaginabaalgo parecido en el antiguo mundo. «Elo referente a las probables costumbres —dijo del Allosaurus (cuyo nombresignifica «reptil saltador», un legado dela época de Cope)—, es sensato deducir que era un depredador, activo y poderoso, adaptado a la vida terrestre.Sus métodos de ataque y lucha deben dehaber sido más parecidos a los de losreptiles modernos que a los de los

mamíferos carnívoros más inteligentes».Se imaginaba al Allosaurus echado en elsuelo a la espera (¿ciertamente uno delos métodos inteligentes de losdepredadores sofisticados actuales?),«luego un repentino y rápido ataque, unaferoz mordedura de los enormesmaxilares y un salvaje ataque codientes y garras hasta que la víctimahabía sido descuartizada o engullidaentera[9]». Era como si un carnívoro deocho toneladas de peso pudiera imitar aun lagarto cuando ataca su presa y seretira rápidamente para volver arecuperar sus fuerzas. Esto,naturalmente, es muy improbable. Elenorme volumen del Allosaurus y de sus

enemigos habrían impedido una victoriade este tipo. Las presas no eradiminutas criaturas inofensivas, como loson las presas del lagarto, ya que en elcaso del Tirannosaurus cabe suponer que se trataba de un ceratopsianoarmado tricornio o de un hadrosaurio de pico de pato más ágil.
Sorprendentemente, Matthew secontradijo a sí mismo cuando volvió arevisar las probables costumbres deltiranosaurio. «Su volumen imposibilitala rapidez y agilidad —declaró, siadvertir que esta afirmación tambiédebía haber sido aplicable al Allosaurus
que a su vez alcanzaba los 10 metros delongitud—. Debe de haber estado

diseñado para atacar y devorar a los pesados y lentos dinosaurios cocuernos y armadura con los que han sidoencontrados sus restos, y cuyas macizascorazas y armas de defensa seemparejaban bien con sus dientes ygarras. El momento implicaba unaacción aparentemente lenta y pesada desu enorme cuerpo, una inercia demovimientos, difícil de iniciar y difícilde cambiar o parar. Estos movimientosse diferencian mucho de la ágil rapidezque asociamos con las bestiasdepredadoras». «Ningún animal de lasdimensiones de un tiranosaurio podíasaltar sobre su presa —concluyó—, y uavance rápido hacia la lucha bien podría

haber acabado en un inevitableempalamiento en los puntiagudoscuernos del Triceratops». Entonces,¿cómo describía Matthew los «combatesde los titanes del antiguo mundo»?Probablemente eran luchas largas contraadversarios bien armados, luchas quedejaban poco respiro a los protagonistas para recuperar sus fuerzas. ¿Podría ureptil de sangre fría, fisiológicamenteineficiente, de ocho toneladas de pesohaber resistido estas luchas con umetabolismo que producía pocaenergía? Un tiranosaurio ectotérmico, dehecho, no habría tenido que retirarseantes que un lagarto, cuyo diminuto peso, después de todo, somete los

músculos a un esfuerzo mucho menor?W. E. Swinton, al discutir las
costumbres de los tiranosaurios en 1934,se imaginaba las luchas de estos primitivos reptiles comoenfrentamientos prolongados y feroces.«Los Tyrannosaurus —dijo de estossuperdepredadores del cretáceo—, no podían haber sido otra cosa que torpes ydesgarbados gigantes que luchabacontra formas de peso similar».
No era una criatura astuta ointeligente sino que en gran partese guiaba por el instinto y lasreacciones automáticas frente alos estímulos visuales y

olfativos, si bien la larga práctica a través de incontablesancestros carnívoros había provocado que esta coordinaciónalcanzara un elevado nivel deeficacia. A pesar de ello, laslimitaciones mecánicas delcuerpo se oponían a losrepentinos saltos, persecucionesveloces o contiendas entre dosinteligencias que caracterizan alas luchas de los mamíferos; y,sin duda, las contiendas delmundo cretáceo, si las pudiéramos ver ahora, parecerían tener la rigidez de lasactividades de aficionados más

que la suavidad de los profesionales aunque lasdisputas eran reales yterribles[10].
La deducción de la función de lasestructuras (tales como dientes y garras)a partir de su forma, sólo es uno de losmedios, si bien el principal, parademostrar que los dinosaurios terópodos(carnívoros) eran agresores activos. Afinales de la década de los años 1930,Roland T. Bird, un explorador que buscaba dinosaurios para el MuseoAmericano y compañero de BarnuBrown, causó bastante revuelo con sus

descubrimientos de dinosaurios. Bien,no exactamente de los mismosdinosaurios sino de sus huellas, dejadas para la posteridad cuando los antiguosleviatanes cruzaron superficies fangosas.En aquellas ocasiones, en las que elfango era lo suficientemente duroquedaron impresas en el suelo huellas bien definidas que se llenaron cosedimentos de aluviones. Cuando elsuelo se elevó y las rocas sobrepuestasse erosionaron millones de años mástarde, el esquisto del interior de lashuellas se desgastó dejando impresionescasi perfectas de las antiguas huellas. Laciencia que se ocupa de las huellasfósiles, o iconografía, recibió u

inmensurable impulso debido a lasnumerosas huellas encontradas por Birden los Estados de las MontañasRocosas.
Hacia finales de noviembre de 1938,Bird llegó al escenario donde seencontraban las enormes huellas decarnívoros con tres dedos — perfectamente impresas en los lechos—,después de haber visto algunas planchasde piedra que contenían huellas fósiles yque estaban a la venta en un tenderete derecuerdos indios. Las huellas dedinosaurios con tres dedos no erararas, ya que se habían descubiertomuchas en el siglo transcurrido desdeque Edward Hitchcock coleccionara sus

huellas de «aves» en el valle delConnecticut para su museo en elAmherst College. Hitchcock habíadescubierto las huellas de dinosaurios primitivos que habitaban la región de
ueva Inglaterra en la época triásica,mientras que Bird estaba tratando co piedras mucho más jóvenes, deaproximadamente 120 millones de añosde antigüedad en Glen Rose a 128kilómetros al sudoeste de Fort Worth eTexas. Glen Rose era territorio virge para las expediciones del MuseoAmericano de Historia Natural. Bird,viendo una piedra de mampostería quecontenía la impresión de una huella deun carnívoro de medio metro de longitud

perteneciente al edificio de laadministración de justicia local, se diocuenta que la región sería provechosa.Parece ser que durante mucho tiempo laciudad había dado por supuesto queestas «huellas eran de hombre». Lashuellas eran muy numerosas en el lechodel río Palaxy, unos kilómetros ríoarriba donde las aguas atraviesan lasmarismas del cretáceo inferior y dejabaal descubierto las huellas. Bird pasó losúltimos días de aquella temporadaexcavando estas huellas de carnívorocon garras de águila, pero tambiédesenterró, casi de forma accidental, la primera sucesión conocida de huellascon cuatro dedos de brontosaurios

gigantes que se movían pesadamente.Estas huellas de saurópodos de 96centímetros de longitud lo hicierovolver a toda prisa a Glen Rose en lasiguiente temporada. Los equipos delMuseo Americano y de la Universidadde Texas extrajeron de este lugar unas40 toneladas de planchas que conteníahuellas, proceso durante el cualencontraron una sucesión de huellas particularmente espectaculares. Uimponente saurópodo había atravesadolas marismas sin darse cuenta que ucarnívoro lo estaba siguiendo de cerca.Cuando el brontosaurio torció hacia laizquierda, el gran carnívoro lo siguió.La tensión iba aumentando incluso entre

los trabajadores más impasibles amedida que se iba exponiendo el drama descubriendo más huellas. Existía
incluso cierto grado de amistosarivalidad al desenterrar las huellas y sehacían especulaciones sobre el resultadode esta cacería prehistórica. ¿Podríaescapar el brontosaurio a la seguridadde las profundidades o se encontraría sesqueleto medio comido en algún lugar más adelante? El decepcionante final dela persecución se produjo cuando laroca quedó enterrada debajo de un murode piedra caliza. El resultado sigue siconocerse[11]. ¿En primer lugar, huborealmente una cacería? Muchasautoridades parecen haber aceptado este

hallazgo como prueba,estremecedoramente dramática, de quelos brontosaurios habían sido perseguidos y caído presas de losrapaces carnívoros que les seguían la pista. Debido a que, tal como dijo unavez Richard Swann Lull, las huellas«son fósiles de seres vivos, mientras quelos demás restos pertenecen a seresmuertos», las pruebas de Bird han sidode gran ayuda[12]. Bird le presentó almundo científico un breve episodio de lavida de un brontosaurio en particular; secentró en un día, día que podría haber sido testigo del final de su vida.
Evidentemente había cierto númerode contradicciones. Por un lado, a los

reptiles actuales se les reconocíagraves limitaciones cuando debíamantener algún tipo de actividad y, a pesar de ello, los glorificados reptilesdel pasado aparentemente eran capacesde las más extraordinarias hazañas. La paradoja sólo era reconocida como tal por unas pocas personas y nunca fuemanifestada explícitamente, aunqueimplícitamente era aceptada por todos.El resultado de la lucha podría haber permanecido incierto, pero existía pocas dudas de que realmente había
habido una lucha. Los carnívoros nohabían sido los cobardes animales decarroña que Lambe había imaginadosino asesinos de sangre fría que

aterrorizaban a sus víctimas.Se conoce el resultado de una lucha.
En agosto de 1971, la expedició paleontológica conjunta polaco-mongolaal desierto del Gobi encontró losesqueletos fósiles de dos dinosauriosque, aparentemente, habían perecidomientras estaban disputando un combatea muerte. El depredador, un pequeño yágil Velociraptor con delgadas piernas ylargas manos cerradas (de aquí elnombre que significaba «ladrórápido»), fue hallado agarrado al cráneode un pequeño Protoceratops
acorazado, abrazado inextricablementesobre el herbívoro en una postura quesugería que ambos habían muerto en la

lucha[13]. (Es un poco difícil de imaginar cómo dos combatientes podían haber muerto simultáneamente y luego haber sido conservados como congelados en lalucha). La paradoja del dinosaurio comoagresor activo fue resueltaingeniosamente por Bakker, en 1972,cuando examinó la ecología de lascomunidades de dinosaurios. Larelación entre depredadores y presas elas comunidades de reptiles difieresignificativamente de la existente en lasde los mamíferos, situación resultante delas diferentes necesidades energéticasde los dos tipos de criaturas. El lagartovivo más grande, el dragón de Komodo(de hecho un monitor estrechamente

emparentado con el mosasaurio marinodel cretáceo) que sobrevive en la remotaisla de Komodo al este de Bali, alcanzalos 3,6 metros de longitud y un pesosuperior a los 450 kilogramos. Estelagarto come el equivalente al cadáver de un cerdo aproximadamente una vez almes. Debido a que esto constituyeaproximadamente la mitad de su peso,cada sesenta días habrá consumido s peso en animales de presa. Un leopardo,en cambio, necesita quemar una mayor cantidad de alimentos para satisfacer susmayores demandas energéticas. Ingerirásu peso en alimentos en sólo diez días.Los leones consumen incluso más, s peso en tan sólo ocho días, y los perros

salvajes en menos de una semana. Por lotanto, las criaturas de sangre calienterequieren aproximadamente diez vecesla cantidad de comida ingerida por ulagarto. ¿Cómo podemos aplicar estoshallazgos a las comunidades dedinosaurios de hace 70 millones deaños? Una comunidad de criaturas sólo podía soportar un determinado númerode depredadores para que la poblacióse pueda mantener estable. Debido a quelos carnívoros de sangre caliente matafrecuentemente, la población sólo podrásoportar a unos pocos carnívoros erelación con los inofensivos herbívoros para no alterar el equilibrio. Lascomunidades de reptiles pueden tolerar

un mayor número de depredadores sique disminuya su población debido aque éstos causan un menor número devíctimas. Bakker demostró que, en lascomunidades reptiles fósiles demediados del período pérmico, existíamuchos más depredadores que en lascomunidades mamíferas fósiles, deloligoceno y plioceno. ¿Qué pasaba colos dinosaurios? Éstos presentaban u patrón análogo al de las relaciones presa-depredador de los mamíferos. Aligual que sucede actualmente en lasllanuras de África, donde los antílopesson mucho más numerosos que losleones, en el cretáceo tardío loshadrosaurios de pico de pato y los

ceratópsidos cornudos eran mucho másabundantes que los tiranosaurioscontemporáneos. Los cálculos de Bakker sobre el flujo energético en lascomunidades de dinosaurios, hacían que pareciese probable que la baja relacióentre carnívoros y herbívoros se debieraa que éstos eran de sangre caliente y noa alguna peculiaridad en la estructura dela comunidad o incluso conservacióselectiva. «El análisis de los flujos deenergía indica claramente —concluyóBakker—, que los balances de energíade los dinosaurios eran similares a losde mamíferos grandes y no a los de
lagartos del tamaño de un elefante[14]».Se han dirigido críticas contra la

precauciones adoptadas por Bakker, por no haber evaluado las pasiones de losrecolectores de fósiles o por lasirregularidades en su conservación; o,tal como destaca Alan Charig del MuseoBritánico, la posibilidad dediscrepancias en las expectativas devida entre los dinosaurios. Todos estosfactores podían afectar drásticamente lasconclusiones de Bakker. A pesar de ello,este enfoque parece ser básicamenteválido, y con algunas modificaciones podría proporcionar unos excelentesresultados.
Los asesinos verdaderamentegrandes de la época mesozoica tardía,los tiranosaurios y alosaurios,

obviamente no eran tan manifiestos (almenos en lo concerniente a su número)como sería de esperar tratándose delagartos monstruosos. No obstante, talcomo se nos había enseñado, se tratabade bestias rapaces muy activas que cofrecuencia necesitaban matar. LosTyrannosaurus al menos, con susgrandes maxilares y pies con garras deáguila, estaban capacitados para abatir alos dinosaurios con pico de pato o a losceratópsidos. Y es muy probable queéstos fueran sus presas. Es improbableque pudieran haber cazado a losdinosaurios más pequeños y ágiles. Laserie de armamentos de los tiranosauriosestaban destinados a combatir

eficazmente a los herbívoros másgrandes y bien protegidos como elTriceratops.
El Tyrannosaurus no estabaincapacitado para correr; probablemente, en ocasiones se veíaobligado a ello para poder cazar a losgalopantes Triceratops. El tiranosaurio probablemente caminaría como los patos, pero incluso los patos pueden ser rápidos cuando es necesario. Tal comosuponía Barnum Brown, tanto elTyrannosaurus como el Gorgosaurus,indudablemente eran bastante rápidoscuando mataban. Los ceratópsidos másgrandes como el Chasmosaurus, quetenía el tamaño de un rinoceronte, y el

Torosaurus y Triceratops, de dieztoneladas de peso, ciertamente podíagalopar a una velocidad notable. Aligual que los rinocerontes, losceratópsidos estaban armados con unaserie de cuernos en el cráneo (algunosde los cuales podrían haber alcanzadouna longitud de 1,20 metros en vida: elnúcleo óseo estaba recubierto de unafunda córnea puntiaguda que se ha perdido en el fósil) y probablementesólo se retiraban si el primer ataque nodebilitaba al agresor. Bakker estimó, a partir de su estudio sobre lasarticulaciones de los miembros de losdinosaurios, que los grandes dinosaurioscornudos podían galopar a velocidades

similares a las de los rinocerontes.«Armados con largos cuernos sobre unacabeza de gran maniobrabilidad, fuertes picos y la capacidad de galopar avelocidades de hasta 48 km/h, estosgrandes ceratópsidos debieron ser losherbívoros terrestres más peligrosos queamás hayan existido[15]». A diferencia
de los tranquilos rinocerontes, losceratópsidos tenían motivos para ser peligrosos ya que compartían su mundocon un terrible enemigo que los igualabaen fuerza.
Aquí encontramos un ejemplo gravede la endiablada hipocresía de la paleontología desde la época victoriana.Leidy y Cope no tuvieron dudas en dejar

que sus dinosaurios «canguro» saltaran, la idea de que los dinosaurios era
activos y que en ocasiones se movíacon rapidez, fue arraigando de formaque, desde entonces, siempre se haaludido a la misma. En realidad, el primer esqueleto genuino de un fósil dedinosaurio montado en los EstadosUnidos fue un hadrosaurio en plenovuelo. El problema siempre había sidolograr posturas que fueran reales. «En lamayoría de reconstrucciones —dijoCharles Beecher de la Universidad deYale en 1902, quizá refiriéndose a loshadrosaurios de Hawkins—,generalmente se observa que elresultado concreto difiere en muchos

aspectos del ideal». Las restauracionesde Hawkins se habían convertido ecuriosidades históricas, así que Beecher comenzó de nuevo la rehabilitación delos dinosaurios en nuestro mundo. Ladificultad era que, al no existir descendientes vivos, no se disponía demodelos para escoger la postura quedebía darse al dinosaurio. Lo que senecesitaba, decía, era un esqueleto quehubiera permanecido intacto desde lamuerte de la bestia, hallándose en una posición que pudiera haber adoptado evida. Afortunadamente, los deseos deBeecher se hicieron realidad con laaparición de un magnífico espécimen delhadrosaurio Claosaunis, que había sido

encontrado por los recolectores deMarsh en 1891. Sus miembros aúestaban articulados y en especial elfémur izquierdo había permanecidoarticulado a la pelvis. Así, el hecho deque la pierna estuviera dirigida haciadelante, como congelada a mediazancada, planteó la pregunta sobre qué postura se le debía dar al animal.Beecher modeló el hadrosaurio como siestuviese corriendo, basándose en las posturas de los animales existentes emovimiento, y prestando especialatención a las fotografías del lagartovivo Chlamydosaurus que,ocasionalmente, en trayectos cortos,corre sobre sus patas traseras.

Se ha intentado (dijoBeecher acerca de larestauración acabada delClaosaurus corriendo) llevar acabo la idea del movimientorápido y hacer que todas las partes del esqueleto contribuyana la perfección y realismo delefecto general. Para lograr esteobjetivo debe existir unadecuado equilibrio y un balanceo verdadero del animalvivo.
Por lo tanto, se pretende queeste enorme espécimen, tal comoestá montado ahora, transmita alobservador la impresión de

precipitación de una bestiamesozoica. La cabeza estálevantada y girada hacia fuera.Los maxilares están ligeramenteseparados. Los antebrazos están balanceando los vaivenes de loshombros. La extremidad posterior izquierda se halla en unextremo de la zancada y llevatodo el peso del animal. El piederecho ha completado el paso yha dejado el suelo preparándose para el balanceo hacia delante.La pesada y poderosa cola estálevantada en el aire ydoblemente curvada paraequilibrar el peso y compensar

el vaivén del cuerpo y de las piernas. El aspecto global es deacción y el espectador fácilmente podrá imaginarse aesta criatura con muchos de susatributos en vida[16].
De forma similar, Barnum Brown,dos décadas más tarde, restauró elGorgosaurus en el Museo Americano deHistoria Natural como si estuviesecorriendo. Esta elección del modo deandar no es sorprendente ya que, inclusoechando una ojeada rápida a losmiembros y articulaciones de losdinosaurios, se puede observar que

muchos dinosaurios estaban construidosde forma similar a los mamíferosungulados modernos. Las articulacionesde los miembros de los ceratópsidos, por ejemplo, eran parecidas a las de losrinocerontes, al igual que lo era el gradode balanceo del antebrazo, lo cualinevitablemente llevaba a la conclusióde que los dos hubiesen podido llevar elmismo paso.
Los dinosaurios bípedosfrecuentemente eran restaurados con las piernas en plena zancada. Una vezreconocido el hecho de que estosdinosaurios «erguidos» podían moversecon bastante rapidez, los paleontólogoscomenzaron a estudiar la mecánica de

los miembros y las posturas máseficaces para poder correr. Actualmente,tal como hemos visto, es corrientereconstruir estos bípedos con la colaextendida y rígida como un tendón queactúa como contrapeso y con la columnavertebral horizontal, enfundada etendones en muchos dinosaurios paraevitar que ésta se hunda. Estosrevestimientos eran de máximaimportancia para contrarrestar losefectos de la gravedad. En el
einonychus, un carnívoro de poco másde 2,4 metros de largo que vivía eMontana a principios del cretáceo —ysin duda el dinosaurio másextraordinario hallado en los últimos

años—, estos revestimientos habíallevado a extremos sin precedentes. Lasfacetas de articulación de las vértebrasde la cola habían crecido formandovarillas, que alcanzaban los 45centímetros de longitud y que encajabaen las vértebras formando una unidadrígida que servía de ayuda paramantener la estabilidad dinámica. Larazón de por qué la bestia necesitaba uestabilizador tan inusual se hizoevidente cuando se analizarofuncionalmente los pies. John Ostrom,que descubrió el dinosaurio en 1964,hizo estas audaces afirmaciones cuandolo presentó al mundo en 1969:

El pie del Deinonychus es, posiblemente, la prueba másreveladora, desde el punto devista anatómico, de los hábitosde los dinosaurios en el sentidode que éstos debieron de ser cualquier cosa menos «reptiles»en su conducta, en sus reaccionesy en su forma de vida. Estedinosaurio tuvo que ser unanimal de carrera rápida,altamente depredador,extremadamente ágil y muyactivo, sensible a muchosestímulos y rápido en susreacciones de respuesta. Esto, asu vez, indica un nivel de

actividad poco corriente para unreptil y sugiere la existencia deun índice metabólico alto. Las pruebas en favor de esa teoríaradican principalmente, aunqueno de modo exclusivo, en el pie[17]».
¿Qué es lo que asombraba tanto elos pies? El Deinonychus eraforzosamente un bípedo ya que laestructura de sus brazos y manosmostraban claramente que el animal no podía, en ningún caso, andar sobrecuatro patas. El pie del animal sedistinguía por presentar únicamente dos

dedos en contacto con el suelo. Existíaun tercer dedo más corto que, siembargo, había sufrido algunasmodificaciones, convirtiéndose en unaestructura ofensiva que contenía unagarra letal de 12 centímetros de largo eforma de hoz curvada. Este dedo sehabía especializado, exclusivamente,como arma extraordinaria cuya funcióinequívocamente era la de cortar ydesgarrar.
Sin embargo, para poder matar ydesgarrar a sus presas con esta garra,debía de tener un sentido del equilibriosin par, debido a que el animal debía
mantenerse de pie sobre una pata
durante el proceso al mismo tiempo que

luchaba con la presa. El hecho demantener precariamente el equilibriosobre un pie probablemente sólo se podía conseguir si la bestia disponía deuna cola rígida, que actuara como unavara de balanceo incorporada, similar ala de los funámbulos. La presa debía ser mantenida alejada del cuerpo a fin de poder ser matada a golpes propinados por las patas traseras. Por esta razón,los brazos eran largos y desgarbados ylas manos estaban mejor adaptadas queen ningún otro dinosaurio para agarrar ysujetar objetos. Esto le permitía al
einonychus agarrar a la desgraciadavíctima y mantenerla a distanciamientras las garras de sus pies la

descuartizaban. Debido a que lascicatrices en las vértebras de este ferozasesino eran exactamente iguales a lasde las grandes aves no voladoras comoel emú y el avestruz, Ostrom concluyóque la columna vertebral del
einonychus debía de habersemantenido en una posición rígidamentehorizontal mediante ligamentos. Laslargas piernas revelaban al
einonychus como veloz depredador,que fácilmente era capaz de dar alcancea sus víctimas. Esta espectacular y pequeña criatura, que se diferenciaba bastante de todas las encontradas hastael momento (de hecho es tan diferenteque Osbom no está seguro acerca de

cuáles son las afinidades), apareció eun momento oportuno. En una décadaanterior probablemente hubiese sidovisto de forma bastante distinta, y sehubiese omitido totalmente laimportancia funcional de suscaracterísticas más notables.Coincidiendo su aparición con la«revolución» en nuestro pensamiento, el
einonychus encarna todo aquello queclaramente no asociamos con losreptiles en los dinosaurios. Nos proporciona, declaró Ostrom al presentar a la bestia, una visiócompletamente nueva de lassorprendentemente sofisticadascapacidades de algunos de los

dinosaurios carnívoros depredadores.
d. El Deinonychus, quizás el dinosaurio másextraordinario descubierto en los últimosaños. Al igual que otros dromeosáuridos erasorprendentemente sofisticado.Reconstrucción realizada por Robert T.Bakker.

e. Deinonychus. Obsérvense las peculiaresvaras óseas a lo largo de toda la cola quehacían que ésta fuera rígida.
En 1970, Galton atacó de formasimilar las reconstrucciones antiguas delos herbívoros bípedos, en las cuales lacriatura corría con el cuerpo inclinadoen un ángulo de 45 grados, con una colafláccida arrastrando por el suelo (locual no solamente habría retardado

considerablemente a la bestia sino quetambién habría causado lesionesinnecesarias en la cola). En lugar deello, también reconstruyó a estos bípedos inclinados hacia delante; estanueva reconstrucción del hadrosaurio
natosaurus, de cabeza plana «quecorre de prisa», guarda poca relaciócon los intentos anteriores pero, desdeluego, transmite la impresión develocidad que anteriormente siemprefaltaba[18].

f. El Anatosaurus de pico de pato de Peter Galton «corriendo».
Sin embargo, hasta finales de ladécada de 1960, estas criaturas fueroconsideradas básicamente comoreptiles. Por muy ágil que fuera saspecto, en vida estaban seriamentelimitadas por una falta de un suministroadecuado de energía. Las aparienciasnaturalmente decepcionaron a muchos

paleontólogos que se vieron obligados aatribuir una función a aquellas piernaslargas y colocarlas de forma natural para poder escapar rápidamente cuandoacechara algún peligro. Incluso atemperaturas óptimas (el momento demayor producción de energía), losreptiles sólo pueden reunir una décima ovigésima parte de la energía de la quedisponen los mamíferos de peso similar.Por lo tanto, un reptil que pese 900gramos puede alcanzar una velocidadde 1609 km/hora con la energíadisponible para ello mientras que umamífero puede alcanzar los 16,09km/hora o más. El Tyrannosaurus
gigante quizá podía alcanzar los 4,8 o

6,4 km/hora si era ectotérmico y elnatosaurus, incluso corriendo, no
alcanzaría una velocidad muchomayor [19].
Las criaturas más grandes y decomplexión más fuerte, no muestran taclaramente lo absurdo de esta situaciócomo lo hacen los imitadores de lasaves no voladoras, con miembros largos cuellos alargados, entre los
dinosaurios, los celurosaurios. Era uhecho universalmente aceptado que loscelurosaurios realmente eran criaturasmuy ágiles. Colbert escribió sobre elOrnithomimus, en 1962, que «losmiembros traseros son largos y estáóptimamente adaptados para correr a

gran velocidad[20]». El Ornithomimus,el «imitador de las aves», poseíamuchas de las características queactualmente asociamos con las grandesaves no voladoras. Presentaba unoshuesos finos y huecos, un cuello largo yun cráneo pequeño con un pico revestido por una vaina córnea en vida y piernasmuy largas. En el imitador de losavestruces, el Struthiomimus, se había perdido todos los dientes, lo cual aúaumentaba más su parecido con elavestruz. Al igual que los avestruces, elStruthiomimus tenía la espaldaacortada, debido al acortamiento de ladistancia entre la espalda y el cinturó pélvico, y los metatarsianos de los pies

se habían alargado para proporcionarleun segmento funcional adicional a la pierna, permitiéndole correr sobre tresdedos (muchas criaturas veloces hasufrido estas modificaciones paraincrementar la zancada). Únicamente lalarga cola, los largos brazos y las manoscon tres dedos hacían que este parecidosuperficial entre el Struthiominus y elavestruz fuera incompleto.
El Struthiomimus medía 3,6 metrosde largo desde el pico hasta la punta dela cola y 2,1 metros de altura, siendo u poco más grande que el avestruz.Debido a que tenía un pico córneo sidientes, tenemos motivos para pensar que, aunque técnicamente perteneciera al

grupo de los terópodos carnívoros, la bestia se había pasado a una dietaherbívora. Los largos y delicados dedos probablemente servían para coger frutosde los árboles o para doblar las ramashacia su pico córneo[21]. Cuando losestrudiomímidos fueron identificados por primera vez por Osborn en 1916,William K. Gregory, del MuseoAmericano, sugirió que las fuertesgarras de los dedos, parecidas a las delos osos, posiblemente habían servido para descascarar los frutos o para coger los frutos de las palmeras, cicádidas ehigueras[22]. De esta forma, los débilesmaxilares desdentados podrían hacer frente a la blanda fruta carnosa. Gerhard

Heilmann estaba considerablementeimpresionado por las ideas de Gregory ehizo una notable reconstrucción de ugrupo de estrutiomímidos, con uaspecto bastante parecido al de losavestruces (para muchas personas el parecido era incluso excesivo[23]),alimentándose de estos frutosdescascarados[24]. El Struthiomimus nonecesariamente debió de ser uvegetariano estricto. El avestruz esomnívoro en estado salvaje y losestrutiomímidos posiblemente tambiéingerían pequeños lagartos o mamíferos,que cazaban con sus manos aferradoras(ventaja que se ha perdido en elavestruz).

g. Una reconstrucción moderna delesqueleto del dinosaurio-avestruzStruthiomimus (derecha). Obsérvese elasombroso parecido con el avestruz(izquierda).
Las poderosas garras han sidoconsideradas por algunos como armasdefensivas, pero es más probable queeste dinosaurio con aspecto de avestruz

huyera al primer indicio de peligro.Gregory no dudaba de que elStruthiomimus fuera un corredor velozcuando estaba amenazado de peligro.Incluso lo invistió con un colgajo de pielque unía los brazos al cuerpo y que podía actuar como un planoaerodinámico para contrarrestar el pesodel tronco, inclinado hacia delantecuando corría.

h. Restauración de Gerhard Heilmann deldinosaurio-avestruz.
El Struthiomimus, en realidad, nonecesitaba esta ayuda aerodinámica. Lacola medía 90 centímetros más que el

tronco y era rígida (como en otrosdinosaurios) en su mitad posterior paracompensar el peso del tronco. Lamaravillosa similitud estructural entre elavestruz y el dinosaurio-avestruz proporciona una valiosa información alanatomista funcional acerca de loshábitos de este último. La convergenciade aspecto entre los animales siempreva unida a una estrecha similitud de lasfunciones. Las criaturas que tienen uaspecto parecido presentan hábitossimilares; sólo hay que pensar en losThylacosmilus marsupiales con dientesde sable y el «tigre». Smilodon codientes de sable, ictiosaurios y delfines,etc. Estas criaturas, frecuentemente

emparentadas, que en ocasiones hatenido orígenes completamentediferentes, finalmente han adquirido uaspecto similar debido a que haadoptado una forma de vida parecida.Por lo tanto, si el avestruz posee loshuesos de las piernas largos paraincrementar la zancada, yconsiguientemente la velocidad, esrazonable pensar que lo mismo es válido para el Struthiomimus, que tambiéalargó su zancada para poder incrementar la velocidad. Debido a quelos avestruces pueden alcanzar velocidades de hasta 80 km/hora, losdinosaurios-avestruces también podíaalcanzar esta velocidad: al menos

poseían la mecánica necesaria paraalcanzarla. ¡Pero si sólo disponían delmetabolismo de los reptiles, con lafisiología y producción de energíacaracterística de los lagartos, losestrutiomímidos sólo podrían haber alcanzado la vertiginosa velocidad de 3km/hora[25]! Esta situación paradójica,en la cual un dinosaurio tamanifiestamente construido para lavelocidad estaba limitado al paso de ucaracol, es equivalente a la construcciódel «Concorde», equipándolo posteriormente con una maquinariarenovada de la firma «Sopwith Camel».
Gregory era un experto elocomoción animal. La velocidad, tal

como demostró, está relacionada con lalongitud de la zancada y la rapidez del paso. Para incrementar la longitud de lazancada, la mayoría de los mamíferosrápidos han incrementado la longitud delos huesos en la parte distal de la pata(región de la espinilla y del pie), y lacomparación de las relaciones entre laslongitudes de los huesos proximales ydistales de la pata es un buen indicador de la velocidad que puede alcanzar uanimal. El fémur de un elefante (muslo)es mucho más largo que su tibia(espinilla); el elefante es un mamíferolento y su relación T/F es de sólo 0,60.En los caballos de carreras, las tibias sehan alargado considerablemente y la

relación se ha incrementado a 0,92. Elos velocistas realmente rápidos comolas gacelas, la tibia es incluso más largaque el fémur y la zancada se acerca a slímite máximo: aquí la relación es de1,25[26]. Los colosales brontosauriostenían una relación T/F casi idéntica a lade los elefantes modernos; los macizosmiembros y la estructura de lasarticulaciones de los miembros en losdos tipos de criaturas, muy separadas eel tiempo, eran asombrosamentesimilares y estaban diseñadas parasoportar un peso aplastante. Losenormes dinosaurios también poseíaunos miembros mucho más rectos quelos de sus contemporáneos más ágiles y

la capacidad de flexión de las rodillas yde los codos era menor, lo cual tambiésucede en los miembros semejantes acolumnas de los elefantes. Esto hacíaque los dinosaurios saurópodoscaminaran muy despacio. Por elcontrario, la mayoría de loshadrosaurios presentaban una relaciósuperior a 0,80, y algunos de estos bípedos tenían las mismas proporcionesque los caballos de carreras y, por lotanto, está justificado concluir que setrataba de corredores veloces. Losdinosaurios-avestruz, tal como era deesperar, tenían una tibia que superaba elongitud al fémur, con una relación de1,12 y, por consiguiente, la criatura

probablemente podría haber competidocon los mamíferos más rápidos. Elsingular Ornithomimus, con unarelación espinilla-muslo de 1,24, erauna verdadera gacela en el mundo de losdinosaurios[27].
El estudio de Dale Russell delesqueleto y de los músculos de las patasde los dinosaurios-avestrucescanadienses del cretáceo tardío, y scomparación funcional con losestrutiomímidos y avestruces, ha proporcionado recientemente una pruebacrucial que apoya la teoría de que losdinosaurios-avestruces eran corredoresveloces. Russell concluyó que, en loconcerniente a la velocidad, estos

dinosaurios rara vez habían sidoigualados en la historia de la vida. Almenos un miembro del grupo, el
romiceiomimus, probablemente habíasido más rápido que el avestruz[28].Otros dinosaurios bípedos sin duda eramuy rápidos. El factor crítico que provocó que Galton sacara de una vez por todas al Hypsilophodon de losárboles fue la larga región de laespinilla. Incluso para los dinosaurios,este factor ya se había convertido en unacaracterística indicadora de losanimales corredores. El Hypsilophodon,con una relación espinilla-muslo de1,18, debía ser capaz de alcanzar velocidades muy elevadas.

Posiblemente era uno de los pocosherbívoros capaces de dejar atrás a loscelorosaurios más pequeños de pataslargas. El Hypsilophodon, al igual queotros dinosaurios, mantenía el cuerpohorizontal mientras corría, con la colareforzada con tendones y alzada al aire.Al igual que otros muchos corredores,corría sobre los dedos en lugar deapoyar las plantas de los pies[29].
Estas cifras no son criteriosabsolutos para evaluar la velocidad. Elhecho de tener unas relaciones idénticasno necesariamente implicaba que sus propietarios pudieran alcanzar velocidades idénticas en vida, ya quehay que tener en cuenta otros muchos

factores (siendo uno de éstos los ajustesque deben hacerse al comparar loscorredores bípedos y cuadrúpedos). Noobstante, estos índices nos proporcionauna directriz. Y lo que es aún másimportante, el uso de estas cifras,obtenidas a partir del estudio demamíferos vivos y fósiles, ilustra laforma cómo los paleontólogos estáempezando a estudiar a los mamíferos yaves, analizando las adaptacionessufridas por estos animales para poder correr, a fin de comprender mejor, lascriaturas que deambularon por el planetadurante la «Era de los Reptiles».

i. El Hypsilophodon, un dinosaurio pequeñoy ágil, era una verdadera gacela en el mundode los dinosaurios.

IV. LA ERA DE LAOSCURIDAD
Debido a que somos mamíferosresulta difícil no adoptar una actitudchauvinista respecto a la Historiageológica, y considerar el origen ysubsiguiente evolución de nuestrosancestros como un suceso de primordialimportancia. La actual posiciódominante del hombre provoca que noconciba la posibilidad de que sus predecesores fueran eclipsados por sus primos reptiles y dinosaurios. Losmamíferos finalmente triunfaron sobre

los reptiles y estamos dispuestos aadmitir que hubo un tiempo en el que losmamíferos fueron dominadoscompletamente, y no por criaturasinferiores sino por criaturasfisiológicamente superiores. Sería uerror pensar que los dinosauriostuvieron poco que ver en la evolución delos mamíferos. Éstos fueron de máximaimportancia en nuestra historia.
Los mamíferos existen desde eltriásico tardío, hace 190 millones deaños. Sin embargo, durante los primerosciento veinte millones de años de sHistoria, desde el final del triásico hastael cretáceo tardío, fueron una razasuprimida, incapaz durante este período

de tiempo de producir algún carnívoroque fuera más grande que un gato o uherbívoro de tamaño superior al de unarata. ¿Por qué los mamíferos fueromantenidos persistentemente diminutos,la mayoría de ellos pequeñosinsectívoros parecidos a las musarañasque estaban obligados a salir de susescondites bajo la protección de laoscuridad y pasar las horas del día en laseguridad de sus madrigueras? ¿Por quéno hubo una radiación hacia la serie deformas que encontramos hoy en día?¿Por qué no vivían «elefantes» en elurásico, «ballenas» en el wealdeniense «gatos con dientes de sable» en el
cretáceo tardío? ¿Por qué el hombre no

aterrizó sobre la luna en el cretáceo?(Estas bestias, naturalmente, no habríasido réplicas de las versiones actuales, pero sí criaturas que cumplirían elmismo papel ecológico. Quizá sería másapropiado denominar a estos mamíferosarcaicos «elefantoides», «humanoides»,etc). Si nos observamos a nosotrosmismos nos damos cuenta del potencialde los mamíferos para triunfar. Éstos poseen todos los ingredientes: umagnífico mecanismo de locomoción,una eficaz fisiología y una inteligenciaque supera con mucho la de cualquier reptil. A pesar de ello, durante los primeros dos tercios de la historia delgrupo, prácticamente no sucedió nada, y

fue a partir del ocaso de los dinosaurios,a finales del cretáceo, cuando finalmentelos mamíferos empezaron a tener éxito.Los mamíferos no desempeñaron papelalguno en la extinción de losdinosaurios. Éstos tuvieron que esperar pasivamente entre bastidores hasta quelos dinosaurios desaparecieron paraocupar el mismo nicho ecológico. Fuecomo si de repente se quitaran losfrenos, liberando a los mamíferos de s posición subordinada.
Nuestros ancestros estuvierorelegados a un papel insignificantedurante el mesozoico. Los dinosaurioseran los amos de ese mundo, criaturascon una fisiología y locomoción ta

eficientes que arrebataron a los«mamíferos» el control del mundo,monopolizándolo durante 120 millonesde años. Es ahora, al comenzar asondear el origen de los dinosaurios,cuando nos damos cuenta de cómofueron capaces de lograrlo, retrasandoasí durante tanto tiempo nuestro propiodesarrollo.
La piedra arenisca de Elgin, eEscocia, es una localidad clásica derecolección de fósiles, en la cual en elúltimo siglo se han descubiertonumerosos reptiles del triásico superior.Muchos de éstos eran etosaurios comoel Aetosaurus y Stagonolepis, «cerdos»reptiles recubiertos de una armadura

ósea inexpugnable, adornada con púasaserradas protuberantes, con hocicossimilares a los de los cerdos para hozar el suelo. Aunque, sin ninguna duda, setrataba de animales terrestres queremovían la tierra en busca de alimentoscaminando a cuatro patas, las patastraseras eran sensiblemente más largasque las delanteras, un posible indicio deque sus ancestros habían sido habitantesde los pantanos, que necesitaban tener fuertes patas traseras para dar coces yabrirse paso por el lodo. De hecho, elos pantanos del triásico tardío aúvivían parientes de los etosaurios, bestias con aspecto de cocodrilos dehocico largo conocidos como

fitosaurios. Estas formas constituyen uimportante grupo del triásico que secaracteriza por presentar dientes coalvéolos, por lo cual Owen les dio elnombre de tecodontes paradiferenciarlos de los lagartos, en loscuales los dientes están fundidos con losmaxilares. En la piedra arenisca«reptilífera», como la llamóexpresivamente E. T. Newton delGeological Survey, se encontró otro deestos tecodontes en la década de 1890,aunque éste era diferente en variosaspectos cruciales. Newton descubrió aeste nuevo reptil triásico en 1894llamándolo Ornithosuchus[1]. A pesar dela naturaleza friable de los huesos

fósiles, Newton pronto se dio cuenta delos rasgos distintos de este tecodonte. ElOrnithosuchus, o «cocodrilo ave»,nombre que revela su carácter intermedio, había conservado algunas delas características primitivas de losetosaurios, pero Newton observómuchas similitudes con los dinosaurioscarnívoros, particularmente los dientesaserrados (el Ornithosuchus eraevidentemente un reptil muydepredador) y el cráneo que seasemejaba mucho al de un megalosaurioen miniatura. Determinadascaracterísticas recordaban a las aves y alos cocodrilos, aunque el esqueleto teníaun notable parecido con el pequeño

dinosaurio Compsognathus de Huxley.Las similitudes entre los tecodontes
los dinosaurios que apareciero posteriormente, siempre habían sidoadvertidas, y el nuevo fósil llenaba elhueco admirablemente. La «dificultad deseparar estos dos grupos —observó
ewton—, se ve incrementada por uestudio de este nuevo reptil de Elgin queocupa… una posición más intermediaentre las dos series». Aunque el cráneo los dientes eran los de un dinosaurio,
el cinturón pélvico y los miembros delOrnithosuchus eran, por su aspecto,definitivamente los de un tecodonte. Setrataba pues de una de las primeras pistas para descubrir la ascendencia de

los dinosaurios. Newton se inclinaba aconsiderar al Ornithosuchus como eldinosaurio más primitivo, más que comoel tecodonto más avanzado, aunqueadmitía que tenía poco en común coéste. El Ornithosuchus de Newton eramuy pequeño, ya que medía menos de 90centímetros de longitud, pero en otracantera de Elgin se encontró un ejemplar de 1,82 metros de longitud en 1901[2]. Eldirector del departamento de reptiles eel zoológico de Londres hizo una nuevaevaluación de sus características primitivas, y lo devolvió a lostecodontes donde se expuso junto a losetosaurios y fitosaurios. El debate, erealidad, sólo se debía a las

preferencias personales, tal como loveía Robert Broom en 1913:
Aunque a primera vista parece ser que estas dosopiniones están reñidas, enrealidad son bastante similares.Prácticamente se trata de que losseudosuquianos (el grupo al cual pertenece el Ornithosuchus),constituyen un grupo de reptiles primitivos que, aunque no seajustan a ninguno de los órdenesespecializados ulteriores, presentan afinidades con unaserie de otros grupos. Pienso queno puede haber la más mínima

duda de que los seudosuquianostenían grandes afinidades con losdinosaurios o, al menos, con losterópodos (carnívoros)… Dehecho, pienso que existen pocasdudas de que el dinosaurioancestral era unseudosuquiano[3].
Broom, el paleontólogo sudafricanomás destacado, que estaba especializadoen reptiles triásicos y antepasados delos mamíferos, se decantaba a favor dela teoría de que el Ornithosuchus nohabía pertenecido al grupo de losdinosaurios. En aquella época, Broo

estaba describiendo un seudosuquianoaún más antiguo al que llamó
uparkeria, que había vivido en eltriásico inferior en África del Sur. ElOrnithosuchus y el Euparkeria,obviamente, estaban estrechamenteemparentados y, debido a que Broo poseía gran parte del esqueleto de estaespecie sudafricana, pudo hacerse unaidea del antepasado de los dinosaurios.Un hecho de máximas consecuencias eraque las patas delanteras del Euparkeria
eran mucho más cortas que las traseras.Broom observó que, mientras loshúmeros y radios medían un total de 68mm, el fémur medía 58 mm y la tibia«aparentemente era un poco más corta

que el fémur y considerablemente másdelgada», es decir, que, en conjunto, elmiembro trasero medía poco más de 100mm. Broom estaba preparando el terrenoal concepto de que los antepasados delos dinosaurios entre losseudosuquianos, habían comenzado a
andar erguidos a principios de la
época triásica, hace 210 millones deaños.
No hubo discusiones acerca de la posición del Euparkeria;definitivamente se trataba de utecodonto. No obstante, dijo Broom, «eel esqueleto poscraneal no existe nadaque no sea justo aquello queesperaríamos encontrar en el antepasado

del dinosaurio». Broom evaluó loshábitos de la bestia: «En mi opinión, el
uparkeria es potencialmente bípedo, y probablemente era parcialmente bípedoen sus hábitos… el animal posiblementecorría sobre sus patas traseras[4]».Broom se imaginaba a este pequeñotecodonto cazando grandes insectos talescomo las langostas que atrapaba en elaire. «El Ornithosuchus —concluyó—, probablemente tenía hábitos muysimilares a los del Euparkeria y estabaincluso un poco mejor adaptado paracorrer velozmente sobre sus patastraseras». De esta forma sentó el precedente para la teoría de que estoságiles bípedos habían sido los

antepasados de los dinosaurios, yconcomitantemente, que la posturaerguida fue adquirida antes de alcanzar el verdadero estado de dinosaurio.Broom creía que los tecodontes parecidos al Euparkeria constituían uestado intermedio entre los dinosauriosverdaderos y los etosaurios acorazados,que conservaban una armadura muy pequeña en el dorso pero que había perdido la mayoría de las característicasarcaicas de los tecodontes.
Los tecodontes seudosuquianos noespecializados dieron lugar a otrosgrupos en el imaginario árbolgenealógico de Broom. Broom creía queel pterodáctilo se originó a partir de

estos ágiles tecodontes por medio de unafase intermedia de transición. Inclusosugirió que este versátil grupo detecodontes habían dado lugar a las aves,que conservaban el modo de andar asaltos de los antepasados de losdinosaurios. También se sospechaba quelos versátiles seudosuquianos(literalmente «falsos cocodrilos») eralos antepasados de los cocodrilos.Aunque algunas de las teorías de Broosiguen siendo controvertidas hoy en día(por ejemplo, que las aves se hubieradesarrollado hace ya tanto tiempo), sus puntos de vista tuvieron una influenciaduradera durante el siguiente mediosiglo. Su teoría de que el Euparkeria y

el Ornithosuchus eran los antepasadosde los dinosaurios fue aceptada pronto,unto con su convicción de que los
ancestros de los dinosaurios ya habíatenido una postura erguida. Colbert haresumido la opinión actual:
En estos tecodontes, losmiembros traseros estánalargados y los miembrosdelanteros son relativamente pequeños. El cuerpo gira sobrela articulación de la cadera, y el peso del cuerpo delante de estaarticulación era contrapesadoeficazmente en el animal vivomediante una larga cola.

Compare mentalmente elmodo de andar de este tecodonte primitivo con el paso veloz deun ave terrestre corredora, comoel correcaminos de los estadosdel Sudoeste, y podrá hacerseuna idea de la locomoción primitiva de los arcosaurios[5].
Existen pocas dudas de que elOrnithosuchus del triásico tardío era uanimal bípedo. La construcción delcinturón pélvico y de la cabeza lateraldel fémur testifican que los miembros semovían hacia delante y atrás, en lugar de proyectarse horizontalmente a ambos

lados del cuerpo. Debido a que losmiembros delanteros eran más cortos, siel animal hubiese intentado caminar acuatro patas, las patas traseras habríadejado atrás a las delanteras, y lacolumna vertebral habría estadoincómodamente arqueada por encima dela pelvis. En la última década, elOrnithosuchus ha sido clasificado unasveces como dinosaurio y otras comotecodonto, pero ello sólo destaca lasimilitud entre los tecodontes avanzados los dinosaurios primitivos. Algunos
paleontólogos quisieran verlo como elantepasado directo del Tyrannosaurus yGorgosaurus[6], mientras que otros piensan que estaba demasiado

especializado como para ser uantepasado directo de los dinosaurios.
El Euparkeria del triásico inferior era una bestia mucho más generalizada yse ha sugerido que dio lugar, no sólo alos carnívoros gigantes a través delOrnithosuchus, sino también a losenormes herbívoros del jurásico comoel Brontosaurus[7]. Debido a que losmiembros del Euparkeria tenían unas proporciones muy similares a las delOrnithosuchus (el húmero constituía el66% de la longitud del fémur en elOrnithosuchus y poco más del 65% eel Euparkeria), existen fundadasrazones para pensar que éste tambiéhabría tenido dificultades para caminar

a cuatro patas. En el caso de que hubiesecaminado como un bípedo, debía dehaber tenido que colocar las piernasdebajo del cuerpo. (Estas doscaracterísticas están interrelacionadastal como ya observó claramente AlfredSherwood Romer: «El cuerpo no podía,naturalmente, ser sostenido en un bípedosi las piernas estaban separadas a ambosflancos del cuerpo[8]»). Se ha sugeridoque cuando corría, doblaba las piernashacia el flanco del cuerpo pero, cuandoandaba despacio, volvía a colocar las piernas en una posición extendida[9].Ciertamente sería lógico pensar quehubo una fase de transición en el origede los bípedos erguidos, pero sigue

siendo discutible si ya la hemosencontrado en el Euparkeria.
Incluso admitiendo que este pequeñotecodonto del triásico inferior no era permanentemente bípedo, sino queadoptaba el ancestral modo de andar desgarbado cuando no corría, aú parece que se trate de una bestia e proceso de volverse erguida. Eldesplazamiento del fémur hacia un planovertical para incrementar la zancada erael primer paso hacia una posturaerguida. Obviamente, estamos presenciando una transición en elmétodo de locomoción y posiblementeen la fisiología del tecodonto. Debido aque una postura erguida probablemente

esté correlacionada con un mayor gastoenergético, tal como destacó Ostrom, parece probable que también se produjouna transformación fisiológica a principios del triásico. Las piernas sedesplazaron debajo del cuerpo paraincrementar la zancada, mientras que elmetabolismo se desvió hacia una producción de energía elevada ysostenida. Esta transformación
suministraría la energía absolutamente
necesaria para que la zancada pudiera
ser utilizada en la fase de máximo
rendimiento, es decir, para la carrera
veloz y sostenida.En el triásico medio, hace 200
millones de años, los tecodontes había

completado el proceso, probablementeeran bestias de sangre caliente con ualto nivel de metabolismo.
¿Por qué estos antepasados de losdinosaurios se volvieron bípedos hace200 millones de años? Aparentementehabían adquirido sus patas traseras máslargas para un fin bastante distinto. Lostecodontes muy primitivos erahabitantes de los pantanos[10]. Al igualque los cocodrilos, habían desarrolladounos miembros traseros más fuertes paramoverse y dar patadas en el agua y unafuerte cola muscular para s propulsión[11]. Estos habitantes de los pantanos compartían su mundo coespecies de animales que nos so

completamente desconocidas hoy en día.En estos pantanos se escondíalaberintodontes anfibios gigantes,criaturas que recordaban mucho acocodrilos aplanados. Muchos eran ta planos que eran incapaces de bajar susmaxilares, éstos ya estaban apoyados eel suelo. ¡En lugar de ello, cuando elanimal quería comer debía levantar elcráneo[12]! En las aguas poco profundasde estos pantanos triásicos pastabahipopótamos reptiles en miniatura, cocolmillos similares a los de las morsas.Estos corpulentos herbívorosevidentemente eran buenos para comer a que los tecodontes acuáticos
primitivos fueron hallados generalmente

unto a ellos en forma de fósiles. Siembargo, cuando el clima del triásico sevolvió más cálido, los pantanos sefueron secando lentamente; los anfibiosa eran escasos a mediados de este
período y la comida habitual de lostecodontes carnívoros, los«hipopótamos», se fue extinguiendo.Eran necesarias medidas drásticas paraque los tecodontes pudieran sobrevivir.Por lo tanto, éstos se aventuraron haciatierra firme por primera vez en busca denuevas presas donde encontrarocomida en abundancia[13]. En tierra,repentinamente se encontraron e posesión de una enorme ventajaheredada de sus días acuáticos, largas

patas traseras y una fuerte cola. Lostecodontes se habían adaptado ya parcialmente para correr sobre sus patastraseras y esto es lo que hacían,inicialmente carreras cortas para cazar sus presas o cuando amenazaba algú peligro[14]. El pequeño Euparkeria,como incipiente bípedo fuetransformando su estilo de vida y,adentrándose en áreas secas altas, pudohacer buen uso de sus veloces miembrossobre suelo más fírme. Esto obviamenteconstituía un gran éxito. En el triásicomedio ya vivían tecodontes «conejo»muy rápidos, en Argentina; se trataba de pequeños reptiles con patas traserasextremadamente largas, que tenían aú

más aspecto de zancas que las de losdinosaurios-avestruz[15]. Sin lugar adudas, estos precursores de losdinosaurios ya habían adquirido umetabolismo que producía un alto nivelde energía tan esencial para las criaturaserguidas corredoras. Estos animaleshabían roto la barrera térmica. Lostecodontes podían dejar atrás a todossus contemporáneos y alcanzar acualquier presa de su elección. Con las patas situadas debajo del cuerpo elugar de a los lados, estas bestias podíacrecer hasta alcanzar proporcionesgigantes. El haber alcanzado por primera vez una postura erguidaconstituía la clave para un crecimiento

sin precedentes. Este acto fue una de lasinnovaciones clave más decisivas en laHistoria de los vertebrados, y lasrepercusiones serían inmensas. Permitióa los tecodontes y a sus antepasadosconquistar un mundo.
En algún punto entre la salida de los primitivos tecodontes del agua y sevolución hacia veloces corredores,hicieron su aparición los dinosaurios,que surgieron en sucesivas oleadas delas filas de los tecodontes[16]. Este áreade estudio está cambiandocontinuamente en la actualidad. Emuchos de los distintos tipos dedinosaurios se puede remontar sascendencia, sin que se observe

grandes cambios hasta el triásico medio.Esto le sugiere a algunos expertos quetuvieron antepasados tecodontesdistintos. Sin embargo, si realmentehubo orígenes múltiples, los respectivosantepasados probablemente estaríaestrechamente emparentados.Recientemente se ha sugerido, en base alas numerosas características — especialmente las relacionadas con laarticulación del tobillo—, comunes atodos los dinosaurios, que el grupo teníaun antepasado único[17].


j. La recientemente descubierta pata traseradel desconocido tecodonte «conejo». Lagerpeton. La cabeza femoral orientadahacia dentro y las largas tibias y pies son pruebas incontestables de que, a mediadosdel triásico, vivían en Argentina tecodontescompletamente erguidos y veloces. Estoindica que ya se había superado la barreratérmica.
Harry Govier Seeley, profesor deGeografía Física en el Bedford College,Universidad de Londres, fue el primeroen pensar en 1887 que los dinosauriosen realidad consistían en dos tiposdistintos; los saurisquios (carnívoros y brontosaurios) y ornitisquios

(iguanodontes y hadrosaurios,ceratópsidos, anquilosaurios yestegosaurios), diferenciándose estosdos grupos principales por el cinturó pélvico[18]. Por lo tanto, parecía que eltérmino «dinosaurio» era antinatural, uagrupamiento artificial de dos tipos degigantes lejanamente emparentados. Elos últimos años la cuestión se ha vueltoenormemente más compleja, yactualmente existen teorías que abarcatoda la gama que va desde un únicoantepasado hasta muchos gruposancestrales diferentes para losdinosaurios. El criterio importante paradecidir si el término «dinosaurio» poseealguna validez, es la relación que tenga

entre sí diversos antepasados (si se da por válida la teoría de los orígenesmúltiples). Los orígenes múltiples noson un obstáculo para agrupar a losrespectivos descendientes; de hecho, losorígenes únicos son la excepción másque la regla en la aparición de los principales grupos de animales. Si todoslos descendientes proceden de un grupoancestral restringido —incluso si sedesarrollan a partir de diferentesmiembros del mismo—, está justificadoel hecho de agruparlos. Debido a quetodos los dinosaurios probablementeeran endotermos completamenteerguidos, resulta más sencillo suponer que estas innovaciones clave existía

como potencial o ya estaban presentesen un limitado grupo ancestral de lostecodontes, y que las sucesivas oleadasde dinosaurios las transmitieron hastalos principales linajes. Si se admite estaidea, el término «dinosaurio» es natural.
El potencial inherente a una posturaerguida, unido a un nuevo logrofisiológico, provocó una explosión delas diversas formas de animales querápidamente llenaron y monopolizarocualquier tipo concebible de nichoterrestre idóneo para animales grandes yactivos. Los pequeños reptiles del tipo
uparkeria hicieron su aparición en umundo en el cual sus descendientes pudieron alcanzar la velocidad de los

estrutiomímidos, el tamaño de los brontosaurios y la ferocidad de lostiranosaurios. Las consecuencias de lamigración de oleadas de tecodontesactivos de tamaño pequeño y medianodesde las regiones húmedas y bajashacia las tierras altas y secas, seríacatastróficas para los reptiles queentonces habitaban estas regiones, entrelos cuales se hallaban los antepasadosde los mamíferos que aún caminabacon las extremidades semiextendidas.Los tecodontes y dinosaurios se estabatrasladando a su terreno y losantepasados de los mamíferos no pudieron competir con estos ágiles yactivos endotermos.

Los reptiles terápsidos, entre loscuales estaban los antepasados de losmamíferos, tuvieron una Edad de Oro afinales del pérmico y principios deltriásico. Durante los treinta millones deaños anteriores a la aparición de lostecodontes y dinosaurios, fueron estosterápsidos o reptiles semejantes amamíferos los que dominaron totalmentela Tierra. En el sistema de Karoo, eÁfrica del Sur, compuesto por una seriede estratos dispuestos en forma desedimentos que llenan una cuencasubyacente, encontramos un registro casicompleto de la vida desde el pérmico

hasta el triásico tardío. La secuencia delos estratos que albergan a susrespectivas faunas es tan completa querealmente podemos ser testigos de cómolos diversos animales se disputaron eldominio durante un período crítico queduró 50 millones de años. Lacaracterística más curiosa de la vida ela cuenca del Karoo, en el pérmicotardío, es el predominio de losterápsidos que prácticamente excluyeroa todas las demás especies de animales.Este orden de animales que prosperócon gran diversidad de formas, y que seextendió hacia todos los nichosecológicos, iba desde los herbívorosenormes, arcaicos hasta los pequeños

insectívoros que removían la tierra e busca de alimentos. Los herbívoros másnotables eran los enormes dicinodontesque tenían unos colmillos parecidos alos de las morsas, grupo que también sediversificó dando una variedad de tiposque abarcaban desde los grandeshabitantes de las llanuras hasta lasrechonchas formas acuáticas que sealimentaban de juncos. El Karoosimplemente rebosaba de estos animalesde dos colmillos en el pérmico tardío ytriásico inferior. En algunos lugares, susestrafalarios cráneos con colmillos y picos estaban esparcidos por todo el paisaje, sugiriendo que estas corpulentas bestias vagaban por el Karoo en grandes

manadas. En algunos estratos, losdicinodontes constituyen alrededor delochenta por ciento de los esqueletosencontrados.
Estas manadas de animalesnaturalmente atraían a los carnívorosque, a su vez, eran terápsidoslejanamente emparentados. Muchos deestos depredadores ligeros teníaaspecto de lobos con miembrosdesparramados. Estos carnívoros decuerpo más grácil y miembros másdelgados, obviamente causaban granúmero de víctimas en las manadas deestos reptiles arcaicos. En muchosdepredadores, los «caninos» superioresestaban alargados a modo de sable y,

obviamente, funcionaban como puñales para penetrar en la gruesa piel de losdicinodontes. Unos pocos carnívoroseran pequeños; la gran mayoría eran deltamaño de un perro o más grandes ycapaces de cazar a los voluminososherbívoros. Posteriormente, en el pérmico y especialmente durante eltriásico, estos carnívoros fuero predominando y adquirieron aspectosmás diversos y muchos de elloscomenzaron a experimentar nuevasformas de incrementar su eficacia.
Existe al menos una pruebacircunstancial de que estos carnívorosestaban intentando incrementar el calor producido internamente. En los cráneos

de estos reptiles de aspecto demamífero, los pómulos empiezan acrecer a lo largo del paladar paraseparar la boca del conducto nasal. Lasventajas de un paladar secundario somúltiples. El animal ya no tenía quedejar de respirar para comer. Con u paladar secundario, la comida podía permanecer más tiempo en la bocamientras el animal continuabarespirando. Esto permitía que la comida pudiera ser masticada antes de ser deglutida. Los dientes en estos cráneosmostraban los primeros indicios de unamayor complejidad, ya que los dientesde las mejillas habían desarrolladocoronas adicionales. Esta serie de

caracteres sugiere que se estabahaciendo necesarias una respiracióininterrumpida y la adecuadamasticación de la comida. El animalestaba aumentando su producción deenergía[19]. En qué fase —si es querealmente hubo una— estos antepasadosde los mamíferos empezaron a tener pelo es una mera conjetura, aunque hayun dato que nos proporciona algunainformación. A principios del triásico,vivían en el Karoo al menos dosterápsidos diminutos que presentabauna característica adicional. En elcráneo, el pómulo estaba perforadoexternamente por numerosas aberturasdirigidas hacia delante. D. M. S. Watso

, que fue el primero en dirigir laatención sobre estos terápsidos en 1931,especuló sobre la función de estas perforaciones. Formuló la teoría de queéstas llevaban una serie de vasossanguíneos muy largos y nervios hacia elhocico. El hocico debía de prolongarseconsiderablemente más allá del cráneoóseo y su morro, similar al de lasmusarañas, probablemente estaría provisto de músculos y pelos de la barba sensibles[20]. Este complicadorazonamiento parece muy audaz, pero eel caso de que en el hocico creciera pelos, indudablemente el cuerpo estaríacubierto de pelos: los pelos de la barbasólo son pelos modificados. Estas

pruebas circunstanciales indican que losterápsidos estaban experimentando couna fisiología endotérmica que, en esafase, probablemente aún era imperfecta.Se cree que los monotremas, como elornitorrinco ovíparo con pico de pato yel oso hormiguero espinoso, hadescendido de los terápsidosindependientemente de otros mamíferos que han heredado su constitució
fisiológica. Debido a que losmonotremas no tienen sistema derefrigeración, y por lo tanto soincapaces de sudar o jadear, no esirrazonable pensar que lo mismo sucedíacon los reptiles semejantes a mamíferosdel triásico[21].

El Karoo pérmico sufrió períodos deintenso frío, de forma que el incipienteintento de elevar la producción de calor del cuerpo y evitar la pérdida de calor através de la piel, aunque de modoineficaz, proporcionó a los terápsidosuna clara ventaja sobre los reptiles detipo lagarto[22]. Los terápsidos habíasido, a lo sumo, animales con una postura semiextendida[23], mientras queel ornitorrinco australiano conserva la postura de miembros extendidoscaracterística de los reptiles. Parece ser que, a finales del pérmico, losterápsidos estaban más interesados eelevar su temperatura para combatir elfrío que en proporcionar más fuerza a

sus miembros.Fue en este mundo del Karoo,
dominado por los terápsidos, dondeirrumpieron los tecodontes. Durante eltriásico, el clima había mejorado y latemperatura se había vuelto menos propensa a sufrir fluctuacionesdrásticas, de forma que los grandestecodontes y dinosaurios no tuvieronecesidad de aislar su piel tal comohabían hecho los mamíferos. Lostecodontes, además, habían adoptadorápidamente una postura erguida yhabían elevado su producción deenergía para potenciar específicamentelos largos miembros, obteniendo comoresultado un marcado incremento de la

velocidad y agilidad. (Debido a que elincremento de la fuerza exigía unafisiología endotérmica para ufuncionamiento bioquímico óptimo, éstafue adquirida simultáneamente). Cuandolas sucesivas oleadas de tecodontes penetraron en el terreno de losterápsidos, hubo un período de intensacompetición[24]. Los antepasados de losdinosaurios estaban mucho mejor equipados y su diversificación estuvoacompañada de un igual descenso delnúmero de terápsidos; en el triásicomedio quedaban ya muy pocos y, afinales de este período, se extinguiero por completo. La Edad de Oro llegóabruptamente a su fin. Estaba

comenzando la Era de la Oscuridad delos mamíferos.
En el triásico medio, los tecodontesse encontraron a sí mismos compitiendocon sus propios descendientes, losdinosaurios, y tuvieron que retirarse.Esta sucesión no fue violenta. Lostecodontes expulsaron a los terápsidosde sus nichos, ocupándolos ellosmismos de forma similar a como laardilla gris más resistente usurpó elnicho de la ardilla roja en Gran Bretaña.El «robo» de un nicho es un sucesohabitual en la sucesión de la fauna, en elcual las criaturas mejor adaptadas almedio ambiente predominante derrocaa sus competidores menos dotados e

una contienda incruenta. Manadasenteras de dicinodontes dejaron paso amanadas de dinosaurios prosaurópodos.Hacia finales del triásico medio, el
elanorosaurus, un precursor del brontosaurio de 12 metros de largo, eraun llamativo ejemplar del paisajesudafricano. De forma similar, losterápsidos «lobos» fueron sustituidos asu vez por tecodontes carnívoros ydinosaurios que comían carne. Era unacarrera hacia una postura erguida, en lacual los vencedores heredarían umundo. Nuestros antepasados perdieron.
Los dinosaurios conquistaron tantolas tierras altas como las bajas.Explotaron todos los nichos ecológicos

que podían ser habitados por criaturasde tamaño medio y grande.Fisiológicamente, presentaban un gradode sofisticación que no tenían losterápsidos y los mamíferos primitivos.diferencia de estos últimos,indudablemente los dinosaurios eracapaces de eliminar el exceso de calor después del ejercicio, posiblemente,como las aves, mediante un sistema desacos aéreos que se extendía por todo elcuerpo. Esto les permitía alcanzar univel de actividad casi inaudito para uanimal terrestre en aquella época.
Mientras tanto, los mamíferosrecientemente evolucionados, se vierosumergidos en su Era de la Oscuridad.

El único nicho que permanecióinaccesible para los dinosaurios era elde las criaturas del tamaño de un ratón;debajo de un tamaño crítico, losdinosaurios habrían perdido demasiadocalor a través de su piel desnuda. Éstaera la única oportunidad de la quedisponían los mamíferos parasobrevivir. Así que se retiraron a estenicho. Por lo tanto, los primerosmamíferos verdaderos (losdescendientes de al menos dos líneas deterápsidos que pasaron al grado demamíferos), eran, muy diferentes de loslobos. Durante los 120 millones de añosque duró la Era de la Oscuridad, losmamíferos estuvieron limitados a u

tamaño muy pequeño y vivieron ehábitats secos de tierras altas. Loshabitantes de este medio ambientesiempre están mal representados en losregistros de fósiles; es en los estuarios bajos y en las regiones costeras dondese acumulan los sedimentos que formalas rocas. Por consiguiente, los restos delos mamíferos primitivos son sumamenteraros, y en su mayor parte se trata sólode dientes. «Hasta hace poco tiempo — estimó A. W. Crompton, que actualmentees el director del Museo de ZoologíaComparada de la Universidad deHarvard—, la colección entera defósiles mamíferos de la “Era de laOscuridad” podía ser guardada en una

caja de zapatos de tamaño medio». Estoes una reflexión que no sólo ilustra laescasez del número de fósiles sinotambién el diminuto tamaño de losdientes de los mamíferos más antiguos.
Una región donde se han encontradodientes de mamíferos muy primitivos esGlamorgan, en el sur de Gales. Cuandoel período triásico dio paso al jurásico,el gran mar de Tethis, que se extendíadesde el Caribe hasta Rusia, comenzó atrasladarse hacia el Norte, inundandogran parte del sur de Gran Bretaña yempujando a los mamíferos hacia el
orte con su avance. Hubo al menos dostipos de diminutos mamíferos quequedaron abandonados en una cadena de

pequeñas islas donde actualmente está elsur de Gales. Estas islas estabacompuestas de piedra caliza plagada dehendiduras y, cuando los mamíferos perecieron, sus restos fueron arrastrados por el agua de lluvia hacia estashendiduras[25]. Los trabajadores que sehallaban en las canteras extrayendo piedra caliza para la construcción decarreteras, pusieron al descubierto estasfisuras y se encontraron toneladas deroca que contenían dientes. Los dientesde bestias llamadas Morganucodon y
uehneotherium miden poco más de 1mm. En el University College deLondres se han procesado muestras delmaterial de las fisuras galesas debido a

que, veinticinco años antes, se habíaadvertido su importancia[26]. En este proceso se utilizan tinas de ácido paradisolver la roca, a fin de obtener losdientes resistentes al ácido (yfragmentos de huesos) que acontinuación son ordenados yclasificados. Los dientes, que presentaunos patrones complejos de las coronas los molares superiores, se adaptan de
forma precisa a los inferiores, ajuste que permite masticar eficazmente la comida.La masticación adecuada de losalimentos es importante para losmamíferos ya que permite una liberaciómás rápida de la energía.
A través del estudio exclusivo de los

dientes podemos saber que se trata demamíferos y no de terápsidos. En losreptiles, incluyendo los terápsidos, losdientes son continuamente sustituidos elos maxilares a lo largo de su vida, yaque los reptiles crecen continuamentemientras viven. Cada sucesión dedientes debe ser más grande que laúltima para poder mantener el mismoritmo de crecimiento que los maxilares.Este sistema, sin embargo, no permite uajuste exacto entre los dientessuperiores e inferiores ya que todos sehallan en diferentes fases de erupción.En los mamíferos, los dientes sólo sosustituidos una vez, en la vida lasegunda dentadura permanece hasta la

muerte del animal como característica permanente y se ajusta perfectamente.Esto es posible debido a que losmamíferos alcanzan rápidamente stamaño máximo. Debido a que losmamíferos jóvenes (que nacen con unagran cabeza) se alimentan con la lechematerna, pueden retrasar la erupción desus dientes definitivos hasta que son casiadultos. En el Morganucodon y el
uehneotherium de las fisuras galesasse han encontrado pruebas de laexistencia de dientes de leche y molaresdefinitivos. Debido a que los molares deleche implican la existencia de cuidados por parte de los padres, por definición,estamos tratando con mamíferos[27]. Si

embargo, como en todas lastransiciones, el proceso debió de ser gradual ya que estos primitivosmamíferos presentan vestigios de sherencia reptil. Los molares«definitivos», y a veces también losmolares, se crían cuando los animaleseran adultos para dejar paso a otradentadura[28]. Sin embargo, ésta nuncaapareció, dejando al diminuto mamíferocon un agujero en la boca. Esto pruebaque estos primeros mamíferos estabaintentando suprimir la última de suscaracterísticas reptiles.
Es notable la ausencia de esqueletosarticulados en las fisuras galesas. Por muy abundantes que sean los hallazgos

de los dientes de los mamíferos primitivos, es difícil encontrar cráneoscompletos. Sin embargo, en otro rincóde la «caja de zapatos» de Crompton, se
udieron encontrar tres cráneos muy preciosos debajo de una capa dedientes. El primero fue hallado por el padre Oehler, de la UniversidadCatólica de Pekín, mientras buscabareptiles semejantes a mamíferos en loslechos del triásico superior en la provincia de Yunán en China en 1948. El padre Harold Rigney, rector de laUniversidad y organizador de laexpedición, consiguió pasar el cráneo decontrabando a Occidente en 1949 antesde ser encarcelado por los comunistas

después de la caída de Pekín. Estecráneo presentaba unos dientes parecidos a los hallados en las fisurasde Gales, por lo que la pequeña bestiarecibió el nombre de Morganucodon
oehleri[29]. Los otros dos cráneos, deuna antigüedad similar, proceden de loslechos rojos del África del Sur. El
egazostrodon y Erythrotherium,descubiertos en 1962 y 1966, son tadiminutos como los mamíferos de Gales,con cráneos que no superan los 20 mde longitud. Es sorprendente quellegaran a ser encontrados. De hecho, el primero de los cráneos africanos fuereconocido por casualidad; se hallabaincrustado en un bloque de roca que

había sido llevado al MuseoSudafricano, debido a que contenía udinosaurio ornitisquiano[30].
Aunque se han encontrado diversosfragmentos, sólo existe un cráneo parcial procedente de rocas que representan lossiguientes 100 millones de años. En loslechos mongoles del cretáceo tardío sevuelven a encontrar cráneos de este tipo.Los tiempos realmente eran difíciles para los mamíferos. Al no poder extenderse hacia nichos aptos paraacoger a animales grandes, ya que éstosa estaban monopolizados por los
dinosaurios, los mamíferos se vieroobligados a esperar tiempos más propicios para ellos. Sin embargo, no

fue una época improductiva. Duranteeste período perfeccionaron sfisiología endotérmica, desarrollando umecanismo de refrigeración eficaz queimplicaba el jadeo y la sudoración ylentamente fueron adquiriendo una postura erguida como los dinosauriosdominantes. Sin embargo, durante todoeste tiempo continuaron siendo una parteinsignificante de la fauna mesozoica.Probablemente debido a su reducidotamaño estaban obligados a salir de susescondites de noche para evitar ser matados y comidos. En aquella épocahabía muchas criaturas grandes, con unafisiología similar a la de los lagartos,que cazaban durante el día, pero que,

con el aire más frío de la noche, sevolvían perezosas. El «lagarto» tuatarade Nueva Zelanda es una reliquia de u primitivo grupo de animales que prosperó durante el mesozoico. Aunque,actualmente, estos reptiles se reducen auna única especie, en otros tiemposfueron extremadamente comunes.Muchos dinosaurios simplemente erademasiado grandes como para preocuparse de estos animales menudos, pero los celurosaurios más pequeños,como el Compsognathus, sí eracapaces de cazar a estos mamíferos. Siembargo, es a partir del cretáceo cuandoencontramos indicios de que losmamíferos eran cazados incluso por la

noche. Los mamíferos eran aterrorizadosademás por criaturas más astutas queellos.
El tenaz Deinonychus era uantepasado de una variedad dedinosaurios carnívoros pequeños perono menos agresivos, en el cretáceotardío, que incluían al Velociraptor
mongol que fue encontrado abrazadosobre su víctima. Las presas delVelociraptor obviamente eran los pequeños dinosaurios herbívoros yOstrom sugiere que el Deinonychus
también tenía preferencia por los pequeños comedores de plantas. Si

embargo, existen motivos fundados paracreer que algunos de los descendientesdel Deinonychus cazaban mamíferos.
Es un tópico que los dinosaurios poseían un cerebro tan pequeño que sólotenían una conciencia muy limitada. Siembargo, el grado de sofisticacióhallado en el Deinonychus —sagilidad, velocidad y capacidad demantener el equilibrio sobre una pierna — parece estar en discordancia con la popular concepción de la mentalidad delos dinosaurios. La coordinaciónerviosa necesaria para estas proezasrequería un cerebro mucho máscomplejo que el que poseían la mayoríade dinosaurios. La complicada

manipulación de los dedos por sí solaconstituye por sí sola un fuerteargumento en favor de un centro decoordinación bien desarrollado, ya queel animal presentaba, además, unosenormes ojos para supervisar laoperación. La caja craneana del
einonychus es desconocida, de formaque nos tenemos que basar en algunos desus parientes de similar talento paraconocer este factor vital. Uno de losúltimos descendientes del Deinonychus
del cretáceo era una criatura pococonocida llamada Dromaeosaurus, el«reptil emú», denominado así por ssupuesto homólogo entre las aves. Esta bestia actualmente ha dado su nombre a

todo un grupo, los dromeosáuridos. Sushuesos fueron hallados originariamente por Barnum Brown, durante su incursióa la región del río del Ciervo Rojo, eAlberta, en 1914, pero fueron descritos por primera vez por Edwin H. Colbert yDale A. Russell en 1969. Lo único queBrown pudo encontrar de este carnívorofueron algunos huesos de los pies y partede un cráneo que, por fortuna, tenía lacaja craneana conservada en su mayor parte. La característica mássorprendente de esta caja craneana erasu forma, ya que era bastante atípica para los dinosaurios carnívoros. Lacavidad ósea que albergaba el cerebroera ancha, especialmente en su regió

posterior o medular. «Si la mitad posterior del cerebro llenaba, aunquesólo fuera aproximadamente, esteespacio —exclamaron Colbert yRussell, evidentemente cogidos por sorpresa por esta forma tan pococaracterística para los dinosaurios—,realmente tuvo que ser muy ancha[31]».Éste fue el primer indicio de queexistían dinosaurios con cerebros queeran más grandes que la proverbial nuez.Al igual que el Deinonychus, el
romaeosaurus era un experto asesino,con dientes incisivos y una garra traserafuertemente curvada diseñada paralacerar, de forma que resulta interesantecorrelacionar su indudable agilidad y

destreza con una mayor capacidadmental. Los dromeosáuridos,obviamente, eran una fuerza a tener ecuenta en los períodos cretáceos posteriores; los dromeosáuridos eracasi tan rápidos como losestrutiomímidos, relativamente másterribles que cualquier otro carnívoro, ycon una capacidad de planificacióinaudita en sus antepasados ocontemporáneos, por lo cual, a smanera, eran incluso más terroríficosque los enormes tiranosaurios. Y posiblemente fueron más numerosos delo que sus raros restos nos hacen pensar.Fueron sus preferenciasmedioambientales las que evitaron que

se conservaran como fósiles.El interés por los dromeosáuridos
comenzó a despertarse a finales de ladécada de 1960, cuando se hizo patenteque existía un grupo totalmente nuevo efósiles de dinosaurio aún relativamente poco explotado. Así, en 1968, DaleRussell llevó un equipo del Museo
acional de Canadá a Albertaespecíficamente para que buscaran aestas raras criaturas. Durante seissemanas, su equipo no recogió otra cosaque huesos aislados, pero en los últimosdías de la exploración en el campotuvieron la buena suerte de localizar elesqueleto desparramado delStenonychosaurus, otro de los

descendientes poco conocidos deleinonychus[32], Este pequeño
depredador sólo medía de 1,5 a 1,8metros de longitud, pero, al igual queotros dromeosáuridos, tenía una cabeza bastante grande, de quizá 22 centímetrosde largo. El cráneo destacaba por presentar unas órbitasdesproporcionadamente grandes que, silos globos oculares las hubieran llenadocompletamente (y existen razones paracreer que así fue), ello hubiesesignificado que el pequeño dinosauriotenía unos ojos de 5 centímetros deancho. Los ojos serían comparables alos de un avestruz que, en la actualidad,son los más grandes de todos los

animales terrestres. Debido a que ladistancia entre los ojos eraanormalmente grande y el hocicocorrespondientemente triangular, losojos estaban dirigidos hacia delante,superponiéndose de esta forma suscampos visuales. Ello resultaba en unavisión binocular . La vistaevidentemente era aguda, y la capacidadde juzgar distancias cercanas de formaexacta era utilizada conjuntamente codedos que podían oponerse a los otros.Una visión aguda y esta nuevamodificación estereoscópica eraesenciales para que el dinosaurio pudiera cazar y manipular a sus presascon las manos. Este estado ta

extraordinariamente avanzado,obviamente requería un control centraladecuadamente desarrollado, de formaque no constituye ninguna sorpresa quela ancha región interorbital del cráneoalbergara un cerebro muy aumentado detamaño, con lóbulos ópticos que seextendían por la región ventrolateral(como en las aves), y hemisferioscerebrales relativamente enormes en laregión dorsolateral. Hasta hace poco,éste era el máximo desarrollo cerebralconocido en los dinosaurios. Nodebemos perder de vista el hecho de quela vasta mayoría de dinosaurios estabadotados de aptitudes insignificantes.Esto es asombrosamente cierto en la

mayoría de los casos. El hadrosauriotenía un peso cerebral que constituíasólo el 1/20 000 de su peso corporal; eel enorme brontosaurio, la proporcióera de 1/100 000[33]. Con cifras comoéstas, es fácil entender por qué se aceptóla idea de que los dinosaurios eracriaturas «no pensantes», casi totalmenteautomatizados, incapaces de presentar lacoordinación sensitiva y capacidad decálculo del frío que se encuentran en losmamíferos y las aves. «El cerebro — afirmó Romer, refiriéndoseespecíficamente a los brontosaurios—,es pequeño en todos los reptiles peroexcesivamente pequeño en estosdinosaurios en proporción a su tamaño.

Con gran probabilidad, el cerebro nohacía otra cosa que mover los maxilares,recibir impresiones de los órganossensitivos y transmitir los mensajes através de la médula espinal a la regióde la cadera, de la cual partían losnervios que hacían funcionar a losmiembros traseros[34]». Losdromeosáuridos era un tipo de animalescompletamente distintos. Éstos habíaalcanzado un nivel de inteligenciasimilar al de las aves y habían dejadotras de sí a los otros dinosaurios. Elmolde del cerebro de uStenonychosaurus, obtenido mediante launión de varios fragmentos de la cajacraneana, sugiere que este

dromeosáurido tenía un volumecerebral de 49 cc y que,consiguientemente, el cerebro debía de pesar aproximadamente 45 gramos evida. Debido a que los emúes deltamaño del Stenonychosaurus
generalmente no pesan más de 45 kg(siendo éste un valor alto), el cerebrodel dromeosáurido posiblemente pesabamás del 1/1000 de su peso corporal. Elemú de grandes ojos tiene un cerebroque pesa, aproximadamente, el 1/1227de su peso corporal, lo cual sugiere queel emú ocuparía un segundo lugar corespecto al Stenonychosaurus si no egrado de inteligencia, al menos etamaño cerebral[35]. Un dinosaurio co

un cerebro de este tamaño ciertamentese opone abiertamente a las ideastradicionales. Incluso reconociendo quelas criaturas pequeñas tenían cerebros proporcionalmente más grandes, eldromeosáurido seguía siendoincomparable en el mundo de losdinosaurios, ya que tenía un cerebro 100veces más pesado, en términos relativos,que el del brontosaurio. Además, habíaáreas particulares en el cerebro de losdromeosáuridos, sobre todo loshemisferios cerebrales, que estabaaumentados de tamaño para controlar elcomportamiento sofisticado del animal ydirigir su complicada coordinaciómotora. (En 1972, Russell procedió a

demostrar que los cerebros de losdinosaurios avestruz eran aún másgrandes, y que la capacidad craneal del
romieiomimus era mayor que la delavestruz[36]). Estos imitadores de lasaves y dromeosaurios del mesozoico,desde luego poseían una inteligenciacompatible con una complejaorganización social.
Probablemente, los herbívoros comoel Struthiomimus eran dinosaurios queformaban bandadas («manadas» pareceun término poco adecuado para estasdelicadas criaturas con aspecto de aves, tanto los carnívoros como los
herbívoros eran capaces de proporcionar cuidados paternales y

formar un grupo familiar estructurado.El Stenonychosaurus probablemente
sólo era uno de muchos de estosdepredadores, pero sabemos muy pocoacerca de las criaturas que compartierosu mundo, a excepción de una especiemongol. El Saurornithoides (el «reptilde aspecto de ave», nombre que hacereferencia a su aspecto) hallado por unaexpedición del Museo Americano aMongolia, en 1922, era un parientecercano del Stenonychosaurus con ucerebro de tamaño similar y loscaracterísticos ojos grandes y manosaferradoras[37]. Al igual que en tantos deestos dinosaurios, su importancia no fuevalorada hasta finales de la década de

1960. Russell cree que ambosdinosaurios —que presentan dedos cocapacidad de manipulación, visió binocular y una clara inteligencia— eracapaces de cazar y comerse a losmamíferos. Anteriormente, se pensabaque los mamíferos habían escapado desus enemigos saliendo de noche de susescondites, cuando la temperatura bajaba y los reptiles se volvía perezosos. Pero debido a que losdinosaurios probablemente eran desangre caliente, éstos presuntamentetambién podían cazar durante la noche.Quizás estos dromeosáuridos más pequeños dormían durante el día, aligual que muchos mamíferos vivos, y

sólo eran activos al amanecer yatardecer. Sus grandes ojos (un bue pero no incontrovertible indicio de laexistencia de hábitos nocturnos)servirían para vislumbrar a losmamíferos en la penumbra. Las manosaferradoras probablemente serviría para coger al mamífero y llevarlo a la boca, o para sujetar a la víctima en elsuelo de forma que los maxilares la pudieran agarrar.
Visto desde una perspectiva másamplia, la depredación activa sólo tuvoun papel secundario en la supresión delos mamíferos durante la Era de laOscuridad. El control de los dinosauriosse basaba más en la naturaleza de la

subyugación pasiva; éstos controlabalos nichos de los animales de tamañogrande e impidieron la irradiación delos mamíferos. Ésta era la situación afinales del cretáceo, cuando umisterioso desastre eliminó el únicoobstáculo que impedía que losmamíferos se desarrollaran con éxito.Algo exterminó la raza de losdinosaurios.

V. LA AGONÍA DELOS TITANES
En julio de 1877, Othniel CharlesMarsh, profesor de Paleontología en elrecientemente inaugurado MuseoPeabody de la Universidad de Yale, publicó una «Nota sobre un nuevogigantesco dinosaurio» en las páginasdel American Journal of Science. Estemodesto artículo, de poco más de una página de longitud y escrito en lahabitual prosa de Marsh inició la décadamás productiva que jamás haya vivido laPaleontología americana. «El museo de

la Universidad de Yale —escribióMarsh— ha recibido recientemente unacolección de restos reptiles de losdepósitos cretáceos de Colorado que esde gran interés. Entre estos especímeneshay partes de un enorme dinosaurio detamaño superior al de cualquier animalterrestre descubierto hasta ahora[1]».Marsh sólo tenía unas pocas vértebras yotros fragmentos, pero su tamaño sugeríaque se trataba de un animal de proporciones gigantescas. Cada vértebramedía 30,48 centímetros de largo y presentaba procesos laterales quemedían hasta 90 centímetros de longitud.Marsh estimó una longitud total dequince o dieciocho metros para la

criatura y la bautizó con el sugestivonombre de Titanosaurus montanus, el«reptil titánico».
El artículo de Marsh marcó un hito,no por sus revelaciones (éstas eraescasas exceptuando el tamaño delreptil), sino porque señaló el comienzode una era. El Nuevo Mundo estaba a punto de descubrir los restos dedinosaurios gigantes cuadrúpedos en talcantidad que los hallazgos anterioresquedarían completamente eclipsados.
Marsh tampoco fue el primero edarse cuenta de la existencia de estosgigantes mesozoicos. En el momento eel que escribió el artículo, los reptilesgigantes ya eran conocidos desde hacía

tiempo en Inglaterra, aunque hacía pocotiempo que se había reconocido el hechode que los más grandes especímeneseran dinosaurios del tipo Iguanodon y
egalosaurus. Antes de pronunciar sdiscurso clásico sobre los caracteres delos dinosaurios en el congreso de laAsociación Británica en 1841, RichardOwen habló acerca de su intento declasificar el revoltijo de grandesvértebras que caracteriza a casi todaslas colecciones. Después de eliminar lasvértebras de los saurios marinos y sustres especies de dinosaurios, Owen seocupó de lo que parecía la vértebra deun cocodrilo enorme. Le impresionó unavértebra de cocodrilo en particular

debido a que ésta era hueca,«reticulada» o «seudoneumática» comoél la llamaba, característicacompletamente desconocida en un sauriogrande. La primera vez que vio unavértebra de este tipo fue en unacolección privada que pertenecía a utal Mr. Saull. Los fósiles habían sidosacados de los estratos delwealdeniense de principios de la eracretácea por el mar y arrastrados por lasolas hasta que finalmente estos huesoslisos habían llegado a las playas de laisla de Wight. Estas vértebras huecas le permitieron a Owen diferenciar otraespecie de saurio gigante de susrecientemente encontrados dinosaurios,

añadiendo así una criatura enorme más asu espectacular zoológico de monstruosresucitados. Owen pensaba que estasvértebras cavernosas eran los restos decocodrilos arcaicos, aunque susantecesores habían llegado aconclusiones bastante distintas. Amediados de la década de 1830, DeaBuckland entró en posesión de un huesodel brazo y de una costilla gigante deuna de estas criaturas. Al igual que loshuesos del Megalosaurus, éstos habíasido encontrados cerca de Woodstock eOxford y se habían convertido en partede la colección de Buckland en el museode la Universidad de Oxford. Al ver estos gigantescos huesos, el baró

Cuvier declaró sin vacilar que se tratabade los restos de una ballena prehistórica, diagnóstico que sorprendeun poco[2]. La gran similitud entre estascostillas desmesuradamente largas y lasde las ballenas deben haber anuladototalmente la objeción de que unacriatura de aspecto tan moderno no podía haber vivido en una época taremota como el mesozoico. Owetambién observó la similitud con lascostillas de las ballenas y se dio cuentade que la textura del hueso, cubierto por una corteza externa compacta, eracaracterística de los cetáceos. Cuvier gozaba de máximo prestigio y susopiniones tenían fuerza de persuasió

incluso después de su muerte (en 1832).o obstante, Owen estaba en posesió
de las vértebras y éstas no se parecíaen nada a las de una ballena, de formaque fue capaz de eliminar esteanacronismo bastante evidente delantiguo mundo. Sin embargo, aunque laidea de la ballena fuera desechada, lassupuestas características cetáceas delfósil mesozoico persistían. Lasvértebras, observó Owen, eran de igualtamaño que las de una ballena adulta y parecían «indicar que el presente sauriomarino gigantesco debió de tener utronco grande y voluminoso, pero propulsado por una cola más larga y más parecida a la de los cocodrilos que a la

de las ballenas modernas[3]». Lasimilitud observada por Owen tuvo sreflejo en el nombre escogido para lacriatura: Cetiosaurus o «lagarto ballena». Owen pensaba que estoscolosos reptiles eran seres estrictamenteacuáticos, que vivían en el mar como las ballenas. Estos animales nadaban, segúOwen, con la ayuda de una larga aletavertical situada al final de un cuerpoaerodinámico y desnudo. Sabemos queOwen pensaba que los ictiosaurios poseían este tipo de aleta debido a quelos ictiosaurios supervivientes deWaterhouse Hawkins en el CrystalPalace tenían largas colas en forma delátigo con una aleta vertical. Owe

imaginaba que los ictiosaurios habíasido habitantes de las aguas poco profundas que, en ocasiones, searrastraban a la playa para tomar el sol.Ésta es la postura en la que fuerorepresentados en el Crystal Palace,donde las aguas del lago chapoteabaalrededor de las aletas de losictiosaurios. Indudablemente, Owe pensaba que los cetiosaurios seimpulsaban mediante una larga cola queacababa en una aleta. Sus pies palmeados eran considerados comoórganos de dirección al igual que lo soen la iguana marina de las islasGalápagos (Owen no halló pruebaalguna de la existencia de pies

palmeados en los cetiosaurios aunquedisponía de algunos huesos de losmiembros para su estudio). En opinióde Owen, los lagartos ballena estabamejor adaptados al mar que losictiosaurios con forma de delfín, yllegados a este punto especuló que probablemente se trataba de carnívoroscolocados en los mares prehistóricosespecíficamente para mantener a raya aestos saurios inferiores[4].
Sin embargo, Owen estabaequivocado como él mismo prontocomprobó. El reconocido método del barón Cuvier para la reconstrucción deanimales enteros a partir de un únicohueso no funcionó en este caso,

sencillamente porque Owen no disponíade precedentes que le pudieran servir deayuda. Owen se basaba en cocodrilos,lagartos y ballenas para sus modelos, deforma que sus cetiosaurios, conocidossólo en base a unas pocas vértebras yhuesos de los miembros,inevitablemente adquirieron un aspectomuy híbrido. Resulta bastante fácilreconstruir un ave fósil a partir de uúnico hueso, si ya se dispone de algúmodelo de ave que sirva de guía. (Owehabía realizado una restauración que fuemuy aclamada. A partir de una astilla de
hueso de unas extremidades rotas de
quince centímetros de largo, Owereconstruyó el ave corredora extinguida

inomis en 1839. Tres años más tardese encontró, en Nueva Zelanda, uesqueleto completo de 3,30 metros dealtura que justificó por completo larestauración publicada por el profesor yla teoría de la existencia de primitivasaves terrestres enormes en la isla. LaPrensa popular elevó a Owen al panteócientífico por este acto único dedestreza científica).Desafortunadamente, cuando intentórealizar la misma proeza con losdragones sin par del mesozoico, nodisponía de ningún prototipo y sólo teníaunas pocas pautas de orientación y, por lo tanto, la empresa estuvo destinada alfracaso.

El modelo de Owen del lagarto ballena fue atacado cuando aparecieromás pruebas que sugeríareconstrucciones más realistas yafinidades diferentes. En 1848 seencontraron en Oxford nuevos huesos delos miembros que, finalmente, loobligaron a abandonar la «hipótesiscetácea», de un cocodrilo marino deltamaño de una ballena. Cuando, e1849, comenzó su monumental obracompuesta de seis partes, el British
ossil Reptiles (serie que completaríaen 1884), no hizo alusión alguna a laanalogía de la ballena. Sin embargo,seguía estando convencido de que elcetiosaurio era un cocodrilo, si bien uno

algo diferente. El tamaño de los nuevoshuesos de los miembros sugería uaspecto parecido al de un dinosaurio y, por lo tanto, primero provisionalmente,comenzó a analizar el cetiosaurio desdeeste punto de vista. En la época delwealdeniense, escribió, cuando el guanodon habitaba la Tierra, existía«un reptil saurio de dimensiones almenos tan grandes como las del guanodon, pero que presentabamodificaciones en la columna vertebraldesde la mitad del dorso hasta la cola,que lo diferenciaban de los dinosaurios lo aproximaban a los cocodrilos[5]».
Desde nuestra privilegiada posició posdarwinista resulta difícil comprender

por qué Owen pensaba que se trataba decriaturas de transición. Cuvier estabatotalmente opuesto a la idea de que lanotable similitud existente en todos loscráneos de vertebrados,independientemente de si se trataba de peces, reptiles u hombres, era elresultado de una «ley superior deuniformidad del tipo», como Owen lallamaba y, en lugar de ello, creía que loshuesos que tenían funciones similareseran también similares en su aspecto.Owen estaba en desacuerdo. El patróde los huesos craneanos es tan parecidoen todos los vertebrados que debe deexistir algún plan general. Todos loscráneos vertebrados son variaciones de

un Arquetipo Ideal utilizado por elArquitecto Divino como prototipo en laCreación[6]. Owen se interesaba más por cómo estos huesos homólogos sediferenciaban entre sí que en losmecanismos que habían producido estoscambios. No obstante, Owen estaba preparado para ir más lejos que Cuvier
admitir que las fuerzas naturaleshabían intervenido transformando elarquetipo básico y dando lugar a lasformas particulares. Por lo tanto, unacriatura como el Cetiosaurus, a mediocamino entre los cocodrilos y losdinosaurios, presentaba modificaciones,algunas características de los cocodrilos otras características de los

dinosaurios, que se desviaban del placomún, si bien es una cuestión discutibleque Owen hubiera considerado alCetiosaurus como paso evolutivointermedio en aquella época (1846). (A pesar de su hostilidad pública a la causadarwinista, Owen frecuentemente tuvoideas evolucionistas en sus posterioresartículos sobre los reptiles fósiles.Parece ser que, en realidad, se oponía almecanismo darwiniano que se basaba eel azar más que al hecho de laevolución).
En los mismos estratos en los que sehabía encontrado la «ballena» originalde Buckland se descubrió, en 1868, otroesqueleto. El profesor John Phillips,

sucesor de Buckland en el Museo deOxford, encontró un fémur del tamaño deun hombre en las rocas jurásicas deEnslow, a 13 kilómetros de Oxford. Estefémur, de 1,60 metros de longitud y30,48 centímetros de diámetro, no tenía par en el mundo de los dinosaurios y sdescubrimiento resultó ser un oportunoincentivo para continuar trabajando en lacantera. Durante más de un año, losexploradores no pudieron encontrar elresto de la criatura, pero, finalmente,Phillips fue recompensado con eldescubrimiento de una gran parte delesqueleto del dinosaurio. Desde los díasde las festividades del Crystal Palace,los grandes lagartos extinguidos había

excitado la imaginación del público y laPrensa popular y las revistas eruditasinformaban regularmente sobre losnuevos descubrimientos. Naturalmente,el nuevo cetiosaurio de John Phillipslevantó un considerable interés. En uartículo aparecido en 1870 en el
thenaeum, una revista literaria ycientífica peculiarmente victoriana,Phillips describía vivamente la canteraen la cual había reposado «el animalmás grande que jamás pisó la Tierra[7]».Sobre el suelo había esparcidos huesosde 1,5 metros de longitud, vértebras decasi 30 centímetros de diámetro y«costillas monstruosas» de 1,5 metrosde longitud que estaban fracturadas e

ambos extremos (es fácil comprender cómo se había producido la confusión ela mente de Cuvier al ver estas costillasde tamaño similar al de las ballenas). Econjunto, dijo Phillips, «parecía el lugar de enterramiento del gran padre de loslagartos». Al igual que en los otroshallazgos de cetiosaurios, no se encontróninguna cabeza, ni siquiera un dientecerca del esqueleto que pudieraestablecer de una vez por todas lasafinidades del animal. Sin embargo,Phillips poseía una buena colección demacizos huesos de los miembros quetestificaban que la criatura había sidocuadrúpeda. Al tratarse de una criaturaterrestre con miembros macizos se

convirtió en el candidato principal paralos dinosaurios de Owen. Sin embargo,Owen no pudo estar absolutamenteseguro de que los restos hallados pertenecían a un nuevo dinosaurio hastaque no supo cuántas vértebras fundidas presentaba el animal en la regió pélvica.
Fue el descubrimiento de un nuevoesqueleto, no del Cetiosaurus sino de usupuesto pariente del cetiosáurido, loque provocó la intervención de Owen.En 1874, la Swindon Brick and TileCompany, que estaba trabajando en lasminas de arcilla de Kimmeridge eWiltshire, escribió una carta a Owen ela cual le describía el descubrimiento de

restos de saurios en uno de sus pozos.Los huesos se habían encontrado a 3metros por debajo de la superficie de latierra y los trabajadores de la mina, aldarse cuenta de la fragilidad de losfósiles, los habían cubierto para evitar que los huesos se desintegraran en elaire solicitando la inmediata presenciade Owen. En los meses siguientes, elMuseo Británico adquirió variastoneladas de estas rocas que conteníalos huesos. La extracción de los huesosde la roca no fue una tarea fácil y,durante un año, un «cantero-escultor»estuvo dedicado por completo a estetrabajo. Finalmente, los huesos fueroarrancados de la roca y Owen pudo

admirar la región de la cadera de scetiosaurio. Las vértebras de la colaestaban en perfecto estado y mostrabala presencia de un poderoso apéndice eforma de látigo en vida. La cola,explicó, «probablemente se utilizaría, aligual que en los saurios vivos másgrandes, para proporcionar golpesmortales en tierra y para surcar corapidez el elemento acuoso[8]». Loslagartos-ballena estaban saliendolentamente, incluso indecisamente, delagua para ir a tierra. El húmero eragrande, lo cual le proporcionó elnombre al dinosaurio: Owen llamó a s«cetiosaurio». Omosaurus, queliteralmente significa «saurio húmero».

Los huesos de la mano eran cortos yrechonchos, mientras que el «conjuntode las patas delanteras —observó Owe —, era más macizo y elefantino en sus proporciones» que en cualquier otrogigante del mesozoico: los enormes piesdebieron de haber sido usados para lalocomoción terrestre. Tal como seaveriguó más tarde, el Omosaurus noera un cetiosaurio, ni siquiera u pariente cercano, sino un dinosaurio más pequeño emparentado con elStegosaurus acorazado. Lo que esimportante es que Owen pensaba que setrataba de un cetiosaurio y que actuó deacuerdo con esta creencia. La presenciade cinco vértebras sacras fundidas,

razonó Owen, era una prueba positivade que los cetiosaurios eran dinosaurios.
Sin embargo, Owen se habíaanticipado. Su precaución extremada yadhesión estricta a su propia definicióanticuada del dinosaurio (que otros yahabían comenzado a considerar de formamás flexible), permitieron avanzar a suscontemporáneos más jóvenes deLondres. En particular, Huxley, que erala eterna espina en el costado de Owen,aprovechó rápidamente estaoportunidad. Los dos paleontólogoshabían roto sus relaciones diplomáticasen 1856, después de que Owen asumierael título de Huxley de profesor dePaleontología mientras daba un ciclo de

conferencias en la Escuela de Ingenieríade Minas de Huxley. Cuatro años mástarde, Huxley se tomó la revanchaderrotando a Owen en el áreaacadémica. La cuestión era,naturalmente, el hombre; el último bastión de los devotos. Owen, en sconferencia en la Linnean Society, e1857, había colocado al hombre en ulugar diferenciado del resto de laCreación basándose en razonesanatómicas. Si Dios debía de continuar siendo una proposición viable y si elhombre debía de conservar su alma,entonces el hombre debía ser considerado como único. De forma queOwen intentó demostrar que Dios le

había otorgado al cráneo y al cerebrodel hombre unas determinadascaracterísticas únicas como signo de sespecial favor. Aunque la Tierra ya nosiguiera manteniendo su posició privilegiada en el Universo, y elGénesis ya no fuera el criterio medianteel cual se medía la cronología, al menosel hombre permanecería inviolable.Huxley tuvo un gran placer en destrozar el razonamiento de Owen, señalandoque estas denominadas característicasúnicas existían en los monos superiores.El último bastión había caído. Acontinuación, Huxley dirigió su atencióhacia los dinosaurios y una vez másdesafió al «Hombre Viejo» de la

Paleontología. En 1867, Huxley visitó elMuseo de Oxford por invitación dePhillips y se maravilló con la coleccióde cetiosaurios de Buckland. No podíahaber duda de que estas criaturasmonstruosas eran dinosaurios, aseguróHuxley, y en 1869 publicó una nuevaclasificación de los dinosauriosincluyendo al Cetiosaurus de Phillips,agrupándolo específicamente con losiguanodontes[9]. Esta unión fuecuestionada con pasión por HarryGovier Seeley, aunque éste tampocodudaba de que los cetiosaurios giganteseran de naturaleza dinosauria. Seeley, dehecho, advirtió que los cetiosaurios eraun grupo importante y diferente de

dinosaurios. Quizá, se aventuró a decir,«grupos originales distintos seconfunden bajo el nombre dedinosaurios». Los dinosaurios eran uimportante grupo de animales en laépoca mesozoica pero se componían desubgrupos principales cuyadiferenciación proponía ahora Seeley.Uno de los grupos que separóinmediatamente del resto fueron loscetiosaurios y, en 1874, sugirió quefueran elevados de rango formando uorden[10]. Los lagartos-ballena habíacomenzado a adquirir algunaimportancia. La fructífera línea derazonamiento de Seeley y su búsquedade algún criterio para distinguir un tipo

de dinosaurio de los otros, dio paso auna clasificación más natural de losdinosaurios en la siguiente década.
Durante aproximadamente cuarentaaños, los cetiosaurios habían sidoconsiderados como seres acuáticos quese disputaban el dominio en los océanosmesozoicos, junto con los ictiosaurios ylos plesiosaurios. Sin embargo, losmiembros paquidérmicos macizos y losdedos cortos de los pies habíademostrado hasta qué punto esta idea sehabía vuelto insostenible. A pesar detodo, el salto completo del agua a tierraera un paso difícil a hacer de un tirón.Aunque los enormes cetiosaurios poseían la «facultad de progresió

terrestre en un grado superior», Owellegó a un compromiso entre el «lagarto- ballena» y el dinosaurio mediante sreafirmación de que «el elementohabitual de los cetiosaurios podría haber sido, y pienso que lo fue, el agua delmar o de los estuarios[11]». Owen eraincapaz de romper con sus ideastradicionales. A pesar de que smonstruo estaba provisto de miembrosmacizos para soportar su peso, Owen erealidad nunca le permitió a la criaturasalir del agua que durante tanto tiempohabía sido su hogar. Ello a pesar de sdeclaración anterior de que losdinosaurios, a diferencia de los plesiosaurios marinos y animales

similares, por definición, eran criaturasterrestres. Aferrado a la tradición, Owedejó a su «lagarto-ballena» monstruoso,sin saber qué hacer, en las aguas poco profundas. Sin embargo, alrededor de1870, había hecho progresosimportantes en el conocimiento delhombre de los dinosaurios incluyendo esu grupo a habitantes gigantes,cuadrúpedos de los mares y lagos.
Los cetiosaurios ingleses eran unacolección poco definida en la época deOwen y, desde entonces, elconocimiento acerca de estos fósiles nose ha incrementado en gran medida.Pero, por muy difícil que fuera snacimiento, los dinosaurios

verdaderamente gigantes habían hechosu aparición en la época victoriana.Estos prolongados dolores de partoapenas fueron experimentados por el
uevo Mundo; aquí, los monstruossalían de la tierra con su formacompleta. Era como si de formaespectacular se hubiera levantado uvelo de la criatura. En 1877,O. C. Marsh, del Yale College, recibiódel Oeste los primeros huesosfosilizados del primo americano delcetiosaurio. Los paleontólogos ya no severían forzados a enfrentarse con restosdesmoronados y fragmentarios. A partir de ahora desenterrarían especímenes perfectamente conservados en u

verdadero cementerio de gigantes.El hecho de que estos gigantes jamás
hayan visto la luz del día, se debe más alos caprichos de la motivación humanaque a los estudios científicosdesapasionados. Los huesos que recibióMarsh habían sido encontradoscasualmente por Arthur Lakes, ugraduado de Oxford que ocupaba u puesto de maestro en una escuela deDenver. Habiendo adquirido u profundo interés por la Historia naturalen Oxford, cuyo museo era un depósitoque rebosaba de restos de saurios, Lakeshabía explorado el territorio virgealrededor de su escuela. Un día demarzo de 1877, mientras estaba

buscando hojas fósiles en la dura piedraarenisca de Dakota, cerca de la ciudadde Morrison, Lake descubrió unavértebra gigante que estaba incrustadaen un bloque de piedra. Debido a queestaba familiarizado con los «lagartos- ballena» del profesor Phillips, leescribió una carta a Marsh en Yaleincluyendo un dibujo de la vértebra y deun hueso del brazo. La reputación deMarsh ya se había consolidado por sushallazgos de pterodáctilos y aves fósilesen Kansas, lo cual le convertía en uobjetivo natural para las noticias sobredescubrimientos de esta índole. Lakesexploró el lugar minuciosamente y pronto desenterró un enorme fémur. De

forma que volvió a escribir excitadamente una carta a Marsh, pensando que el monstruo debía tener una longitud no inferior a los 18 o 21metros. No recibió ninguna respuesta de
ew Haven. Lakes envió una terceracarta seguida de una cuarta. En suscartas hablaba de otros huesos enormesque pertenecían, decía, a seis individuoso quizás a seis especies diferentes.Increíblemente, Marsh no estabainteresado. Lakes envolvió los huesos yesperó varias semanas para que Marsle diera la señal para enviárselos, siembargo, no le llegó ningunacontestación. Perseveró y, en mayo de1877, se encargó él mismo de enviar

diez cajas de embalaje llenas de huesosde dinosaurio a New Haven, cuyo pesototal era inferior a una tonelada. La faltade resolución de Marsh finalmentedefraudó a Lakes. Desesperadamentedio un paso que haría que Marsh seinteresara y prestara atención: escribió ala persona que Marsh más temía queentrara en posesión de estos preciadosespecímenes. Lakes escribió a Cope.Marsh había librado una batalla amarga,incluso algo defensiva, contra Copedesde los días de la disputa sobre elelasmosaurio y, constantemente, estabarechazando los ataques insidiosos de soponente en la Prensa. Ahora, Lakes nosólo había escrito a Cope, sino que

incluso le había enviado algunos de sushuesos gigantes a Filadelfia. Marsh seenteró de esta correspondencia einmediatamente entró en acción,enviando un cable a Lakes para evitar (con retraso) que éste enviara sus huesosde Morrison a Cope, y un telegrama auno de sus propios recolectores sobre elterreno en Kansas, Benjamin Mudge,ordenándole que fuera en seguida aMorrison para que llegara a un acuerdocon Lakes y se asegurase lo que Marsahora consideraba como su propiedad.Cuando llegó a Morrison, Mudgerápidamente se dio cuenta de la ricacolección de fósiles cuando Lakes«salió de la casa de fieras» y se le

aproximó. Se llegó a un acuerdo yMudge le cablegrafió a Marsh: «Hemoslogrado un acuerdo satisfactorio parados meses. Jones no puedeinterferir [12]». «Jones», naturalmente,era Cope. Entretanto, Cope había estado preparando a toda prisa un artículosobre esos restos de saurio y le habíaenviado a la American PhilosophicalSociety, en Filadelfia, una relació preliminar. No es difícil imaginarse scólera cuando éstos fueron arrebatadosde nuevo por Lakes quien informó aCope que Marsh ya los había comprado.Para añadir aún más insulto a la afrenta,Lakes le exigió a Cope que enviara sushuesos fósiles a Marsh.

Incluso una mirada superficial aMarsh y Cope durante su guerra,rápidamente refuta el mito del científicosobrio y objetivo. La celosa rivalidadque existía entre estos dos hombres elos días del Oeste Americano no estabaexenta de compensaciones ya que, siduda, fue la batalla más productiva elos anales de la Paleontología. Amboshombres eran unos descriptores prolíficos y su motivación era a menudoque el otro estaba trabajando en lamisma área. ¿Cuánto tiempo hubiesenecesitado Marsh para darse cuenta delos descubrimientos de Lake si Cope nohubiese actuado repentinamente comocatalizador?

Marsh no perdió tiempo en publicar una relación de los huesos fósiles deLakes, a fin de asegurarse su prioridadsobre Cope. En Morrison, Mudgecomenzó a trabajar en el yacimiento deforma fervorosa. El primer hueso queextrajo, un húmero de más de 60centímetros de ancho, fue un presagio delo que se avecinaba. En el transcurso deunas pocas semanas, Lakes y Mudgehabían enviado con el tren otra toneladade huesos a Marsh, entre los cuales sehallaban los primeros vestigiosconocidos del dinosaurio acorazadoStegosaurus. Al principio, Marsh noestaba muy seguro de las afinidades delestegosaurio y pensaba que podía estar

relacionado con las tortugas, alconsiderar que las enormes placasdérmicas del animal (algunas de lascuales medían más de 90 centímetros delongitud) eran como una especie dearmadura en forma de coraza[13]. Mástarde, colocó estas placas en posicióvertical formando una gran cresta ósea alo largo de la línea media de la espaldadel estegosaurio.
El tamaño de algunos de los huesosde los primeros envíos de Marsh es bastante asombroso incluso paranosotros hoy en día. La mayoría de los primeros descubrimientos fueron de elTitanosaurus. Un fémur que se hallabaen posesión de Marsh medía más de

2,44 metros. ¡Si este hueso hubiese pertenecido a un cocodrilo, especulóMarsh, la criatura hubiese tenido laincreíble longitud de 35 metros! Al igualque sus primos ingleses, Titanosaurus
tenía vértebras ahuecadas, pero sólo treso cuatro vértebras fundidas en la regióde la cadera (menos que en la definicióoriginal del dinosaurio de Owen).Cuando esta región se hizo másconocida en los dinosaurios, el númeroresultó ser variable en los diferentestipos.
Debido a un curioso giro deldestino, del tipo que parece haber seguido tenazmente a Marsh y Copedurante la mayor parte de su vida de

trabajo, otro maestro de escuela estabahaciendo excursiones por las colinas deColorado, en marzo de 1877.O. W. Lucas, un botánico aficionado,estaba explorando la región cerca deCanyon City, al sur de Morrison, e busca de plantas. Él también encontrórestos enormes de saurios pero su
reacción inicial fue escribir a Cope.Cope aprovechó esta oportunidad paracolocarse en igual posición que Marsh einmediatamente contrató a Lucas. Lasexcavaciones comenzaron a toda prisa.Los primeros huesos enviados a Cope eFiladelfia consistían en fragmentos de umaxilar de Laelaps. Sin embargo, elsegundo lote contenía vértebras que,

anunció Cope, «aparentementerepresentan a un animal mucho másgigantesco y creo que se trata del animalmás grande o más voluminoso capaz dedesplazarse por tierra del que tengamosconocimiento[14]». (No sorprende muchoque estas palabras fueran pronunciadascon una regularidad monótona en lossiguientes meses a medida que sedescubrían criaturas cada vez másgrandes en los yacimientos). Cope publicó a toda prisa un artículo, el 23 deagosto de 1877, describiendo el reptilgigante de Lucas al que llamóCamarasaurus o «reptil decompartimentos» por la naturalezacavernosa de sus vértebras. A pesar de

ello fue vencido por un estrecho marge por Marsh cuyo artículo sobreTitanosaurus había aparecido el 20 deunio. Sin embargo, Cope tuvo u
remedio para compensar el haber llegado un poco tarde en su publicación.Insistió en que Titanosaurus, el nombreescogido por Marsh, no podía ser adoptado de acuerdo con las reglas de lanomenclatura debido a que no habíaningún diagnóstico o descripción queacompañaran a los huesos. En base alartículo de Marsh, habría sido imposibledistinguir la criatura de un cetiosaurioinglés. De cualquier manera, dijo Cope por añadidura, Titanosaurus era unombre ya adoptado por una criatura

fósil. Cope, que había llegado elsegundo respecto a Marsh, impugnaría ladecisión en base a los detalles técnicoslegales. Cope dijo que fue él quien habíadescrito por primera vez a un herbívorogigante. Por deferencia hacia las reglasde la nomenclatura, Marsh cambió elnombre de su monstruo por
tlantosaurus aunque esto sea una débilustificación por robarle la prioridad.
Hasta este momento, Cope habíaestado tratando sólo con dinosaurios bípedos con aspecto de canguro, deforma que fue una sorpresa para éldescubrir que las patas delanteras delCamarasaurus eran largas y que lacriatura caminaba sobre cuatro patas. La

reacción inicial de Cope al conocer estehecho fue sorprendente. Las largas patasdelanteras, afirmó, «junto con el notablecuello largo que poseía este génerosugieren un parecido en la forma yhábitos entre los reptiles gigantes y lairafa. Mientras que algunos de los
dinosaurios posteriores se alzaban sobrelas patas traseras para alcanzar lascopas de los árboles donde podíaalimentarse (se refiere al Hadrosaurus
del cretáceo), la forma general delcuerpo en algunos de estos tipos primitivos los capacitaba para alcanzar su alimento sin que las patas anterioresabandonaran la tierra». El siguienteenvío de Lucas incluía los restos de una

bestia aún más grande, Amphicoelias,que Cope consideraba como uadecuado rival para el Camarasaurus
en el mundo mesozoico. Cope describióeste inesperado cambio completo en elestilo de vida de los dinosaurios,escasamente dos años después de queRichard Owen reafirmara, en Londres,su creencia de que estos cuadrúpedosgigantes eran animales acuáticos.Parecía como si el primo americano delcetiosaurio hubiera sido emancipadorepentinamente del mar. Sin embargo, elleviatán estaba luchando prematuramente en tierra.
Marsh, entretanto, se enteró del lugar de excavación de su rival a sólo unos

160 kilómetros al sur del suyo propio, ydio instrucciones a Mudge para queabandonara Morrison e investigara.Cope, sin embargo, ya habíamonopolizado el lugar y Lucas estaba alservicio de él. Mudge observódecepcionado que los huesos queestaban excavando se hallaban en mejor estado y eran hasta un 30% más grandesque sus homólogos en Morrison, y que aCope le estaba costando muy pocoexcavarlos. Marsh ordenó a susrecolectores que se dirigiesen a laregión para comenzar a realizar prospecciones en los alrededores;Samuel Williston fue traído desdeKansas para dirigir las operaciones

unto con Mudge. Cuando tuvieron laimpresión de que Lucas estabadesilusionado con Cope, ya que éste sesentía explotado y mal pagado,inmediatamente le hicieron ofertas. Siembargo, Lucas respetó el acuerdo coCope y envió los restos de los huesos desu yacimiento a Filadelfia. Elacimiento alternativo de Mudge y
Williston no tuvo éxito. Sólo extraíahuesos friables y rotos de la dura piedraarenisca y celosamente miraban cómolos hombres de Cope estaban sacandohuesos completos del esquisto más blando de su yacimiento. Después deque fracasara su intento de sobornar aLucas, Marsh ordenó a sus hombres que

volvieran a Morrison para recoger allítodo lo que pudieran, pero un gravedeslizamiento de montaña casi mató atodo el equipo y el trabajo fuesuspendido durante el resto de latemporada.
Una vez más, se sabe que Marstuvo conocimiento de Canyon City antesque Cope. En febrero de 1877, otrorecolector de fósiles envió cartas en lasque describía el descubrimiento dehuesos de reptiles gigantes, algunos cocabezas articuladas de 50 centímetros deancho. En este caso, Marsh tampocosiguió la pista y, sólo cuando Cope seinteresó, Marsh decidió que el lugar erade valor científico.

A mediados de 1877, Marsh yaestaba completamente sobre aviso de lafantástica riqueza de reptiles fósiles queesperaban ser excavados en las rocasurásicas de los Estados de las
Montañas Rocosas. En julio, recibió unacarta firmada con el seudónimo de«Harlow y Edwards», que le relataba eldescubrimiento de un gran número defósiles, que supuestamente pertenecíaal perezoso terrestre gigante
egatherium, en rocas consideradas por los descubridores pertenecientes al período terciario. «Estamos deseosos devender los fósiles que ya hemosconseguido —escribían Harlow yEdwards— y, también, el secreto de los

otros».«Un omóplato —informaron a Mars
—, medía poco menos de 1,5 metros.Aún no le hemos dicho nada a nadie — dijeron—. Nos gustaría mucho recibir noticias suyas que siempre serán bievenidas, puesto que sabemos que esusted un geólogo entusiasta y un hombrede fortuna, cualidades que siemprehemos estado deseosos de encontrar y,en especial, la última». Como signo de buena voluntad enviaron algunos huesosa New Haven para que Marsh losinspeccionara.
Marsh desempaquetó el perezosogigante y fue recompensado con lasvértebras, dientes, maxilares y huesos de

los miembros del enorme Brontosaurus
urásico. Inmediatamente, telegrafió aHarlow y Edwards y les adelantó ucheque. Éste, naturalmente, no podía ser cobrado porque los descubridoreshabían ocultado su identidad verdadera para mantener secreto el hallazgo y slocalización. Volvieron a escribir unacarta a Marsh diciéndole: «Hay muchagente que busca estas cosas y si noslocalizan podrían hallar descubrimientosque ya hemos hecho». Marsh envió ucable urgente a Williston y lo despachóa este nuevo lugar de las excavacionessituado en Como Bluff, en Wyoming.Williston hizo correr la voz de que seiba a Oregón por si alguien intentaba

seguirlo. (Estos sucesos parecen más udrama de espionaje de ficción que laexploración de fósiles; el secreto eincluso el engaño eran sintomáticos dela paranoia que se había apoderado delos campamentos rivales en su búsquedade saurios gigantes). Los huesos estabaesparcidos en un área de 10 a 11
kilómetros, escribió Williston sobre estanueva localidad, «Canyon City yMorrison son sencillamente nada ecomparación». No perdió tiempocontratando a los dos descubridores, losempleados del ferrocarril de Como,William E. Carlin y Bill Reed, y, en elcurso de tres días, ya se habían resueltotodas las formalidades incluso antes de

que Cope pudiera llegar a saber algo deltrato. El acuerdo incluso estipulaba queCarlin y Reed debían tomar «las precauciones razonables para mantener fuera de la región a todos los buscadores de fósiles no autorizados por el profesor Marsh». Una precaucióustificable, naturalmente. Al cabo de
pocos días, un misterioso personajeapareció repetidas veces cerca del lugar haciendo preguntas sobre huesos ycráneos. Uno de los hombres de Cope,informó Williston. «¡Primero dijo queestaba vendiendo víveres!». Willistoincluso obtuvo una muestra de sescritura para compararla con la deCope, en el caso de que se tratara del

archirrival de Marsh en persona.De esta forma, comenzó la
exploración en Como Bluff, donde ulugar albergaba quizá cientos deacimientos en los que se desenterraba
dinosaurios en un espacio de muy pocosaños. Las intrigas continuaban. Los dos paleontólogos desarrollaron laextracción rápida de dinosauriosconviertiéndola en un arte refinado, cola esperanza de descubrir un nuevo lugar de excavación antes de que el otrosupiera de su existencia. Eran comuneslos incidentes violentos. Hombres quedesertaban al campo contrario y seempleaban tácticas de mano dura paraexpulsar a los invasores, o para

introducirse en un yacimiento rival. Losdinosaurios gigantes como el
iplodocus, Camarasaurus yarosaurus fueron un tema común en los
artículos científicos de Marsh y Cope.Se abrían yacimientos, se extraían todoslos huesos y se cerraba todo en uespacio de tiempo de pocos meses oincluso semanas. Cuando el equipo deexcavación abandonaba un yacimiento,machacaba los huesos restantes paraevitar que cayeran en manos de su rival.Esta explotación sacó a la luz muchosfósiles que podrían haber pasadoinadvertidos con otros métodos deexploración. En un yacimiento hubomucha excitación por el descubrimiento

del primer mamífero jurásicoamericano, una criatura «marsupial» primitiva, de la cual se pensaba quetenía parecido con el opossum y a lacual Marsh bautizó con el nombre de
ryolestes. (De hecho, ésta vivió en unaépoca demasiado temprana como parahaber sido un marsupial. Losmarsupiales y los mamíferos placentarios no aparecieron hastamediados del cretáceo, desarrollándosea partir de mamíferos primitivosemparentados con los drioléstidos). Ualiciente tan espectacular como éste hizoque el mismo Cope acudiera a losacimientos en busca de trofeos
similares.

En 1878, la llegada de huesosgigantes de herbívoros a New Haven eratan masiva, que Marsh los escogió paraun estudio especial, dedicándoles variosartículos. Al igual que Seeley antes queél, reconoció que estos dinosauriosgigantes cuadrúpedos eran algo distintosdel resto y, oficialmente, los clasificócomo saurópodos o «pie de lagarto» por sus pies con cinco dedos (en contrastecon los bípedos que generalmente teníatres dedos[15]). Aunque Seeley ya habíadiferenciado estos dinosaurios comocetiosáuridos, ha sido el nombre deMarsh el que ha pasado al uso común.
Las excavaciones continuarodurante años, esforzándose Marsh y

Cope en mantener la paz entre ellos. Seescribieron y publicaron numerososartículos que describían a las criaturasque surgían de las rocas. Sedescubrieron brontosaurios completosunto con estegosaurios acorazados,
carnívoros gigantes como el Allosaurus
e incluso dinosaurios del tamaño de ugato como el Nannosaurus. Sedesenterró gran número de huesos delsaurópodo Morosaurus. (ElCamarasaurus de Cope: debido a quelos dos paleontólogos estabadescribiendo simultáneamente losmismos dinosaurios, la mayoría de lascriaturas acabaron teniendo dosnombres). Cuando Marsh vio por

primera vez el cráneo ridículamente pequeño de esta criatura monstruosa sesorprendió; su cerebro debía haber sidorelativamente más pequeño que el decualquier otro animal vertebrado. Elgran ornitópodo bípedo Camptosaurus
fue hallado en estos yacimientos, aligual que el delicado Coelurus, cohuesos huecos y característicassemejantes a las de las aves (unacriatura peculiar que, en un principio,Marsh no pudo clasificar, aunque a principios de 1880 reconoció que era utipo de dinosaurio carnívoro). La bonanza duró doce años. Alrededor de1889, la gran fiebre de los huesos yahabía pasado. Los yacimientos fuero

agotados y posteriormente muy pocoshan proporcionado gigantescomparables.
Uno de los resultados deldescubrimiento de saurópodos enterosen Como fue que Marsh fue capaz dereconstruir por primera vez estascriaturas con exactitud. El esqueleto de
rontosaurus hallado por Reed era casi perfecto. Todos los huesos procedían deun individuo y éste debía de haber medido en vida 15 metros de longitud.Marsh restauró su esqueleto en 1883 yacompañó su restauración con unadescripción de la criatura.
La cabeza era notablemente

pequeña. El cuello era largo, y,considerando sus proporciones,flexible, siendo la porción másligera de la columna vertebral.El cuerpo era bastante corto, y lacavidad abdominal era detamaño mediano. Las patas y los pies eran macizos y todos loshuesos sólidos. Los pies eran plantígrados, y cada huella del pie debía de tener aproximadamente 0,8 metroscuadrados de extensión. La colaera larga y casi todos los huesos,sólidos.
La diminuta cabeza es lo primero que atrae la atención, ya

que, en proporción con elcuerpo, es más pequeña que lade cualquier otro animalvertebrado conocido hasta ahora.El cráneo entero tiene undiámetro o un peso verdaderoinferior a la cuarta o quintavértebra cervical.
Una estimación prudente deltamaño del Brontosaurus, talcomo ha sido restaurado aquí,demostraba que en vida elanimal debía haber pesado másde veinte toneladas. La cabeza yel cerebro tan pequeños y ladelgada médula espinal, indicanque se trataba de un reptil

estúpido, de movimientos lentos.La bestia carecía completamentede armas ofensivas o defensivaso de una coraza dérmica.
En sus hábitos, el brontosaurio era más o menosanfibio, y su comida eran, probablemente, las plantasacuáticas u otra vegetaciónsuculenta. Los restosgeneralmente se encuentran enlugares donde por lo visto losanimales se habíanencenagado[16].
No fue la aceptación ciega de los

puntos de vista tradicionales lo que provocó la preferencia de Marsh por lateoría de que se trataba de un saurópodoacuático. Había una característica queestos dinosaurios compartían con lascriaturas verdaderamente acuáticas,como los ictiosaurios y las ballenas: losorificios nasales estaban situados en la parte trasera del cráneo, con frecuenciacerca o incluso entre medio de los ojosen lugar de hallarse en la punta delhocico. Las criaturas acuáticas corren el peligro de aspirar agua al respirar, deforma que es deseable tener los orificiosnasales lo más altos posibles en elcráneo. Los orificios nasales estabaincluso situados en bóvedas en el

rachiosaurus, de forma que podíaelevarse por encima del agua mientrasque el resto del animal permanecíasumergido. A diferencia de Marsh, lareacción inicial de Cope respecto alCamarasaurus fue de que se trataba deuna jirafa reptil, provista de un cuellomaravillosamente alargado quecapacitaba al animal para ramonear elas copas de los árboles. Con estateoría, Cope, de buena gana, habíallevado al animal a tierra por primeravez. Sin embargo, pronto abandonó estaidea y se unió a la opinión generalizadade que los saurópodos eran habitantesde los lagos, representándoloscompletamente sumergidos en el agua y

de pie sobre el fondo fírme del lago o paseando debajo del agua en busca dealimentos. Ocasionalmente, levantaríasus cuellos alargados para sacar lacabeza fuera del agua y tragar aire antesde continuar cogiendo la suculentavegetación del fondo del lago[17]. E profundidades incluso mayores, elanimal, mantenido a flote por el agua, selevantaría sobre sus patas traseras paraalcanzar la superficie a unos 9 metros omás del fondo del lago. El enormesaurópodo paseándose por el fondo dellago a salvo de todo peligro, levantandosu cuello verticalmente a modo de periscopio a través de la superficie hasido una noción popular desde los días

de Cope. El biógrafo de Cope, HenryFaifield Osborn, estaba de acuerdo deforma incondicional con su predecesor.Imaginó al Camarasaurus como a ugran cuadrúpedo vadeando por el fondofírme de los ríos arenosos del Wyomingurásico o quizá nadando rápidamente
cuando amenazaba peligro, propulsándose con la ayuda de su larga
fuerte cola. Una extensa cápsulacartilaginosa en las articulaciones de losmiembros y la supuesta naturalezacartilaginosa de algunos huesos de lostobillos y muñecas hicieron pensar amuchos expertos en dinosaurios que lossaurópodos nunca podrían haber caminado sobre tierra firme sin oprimir

las articulaciones de sus miembros ylesionarse a sí mismos. Osbom noestaba de acuerdo con esta opinióextrema. Sin embargo, él tambiédescribió al Amphicoelias viviendo lamayor parte del tiempo bajo el aguadonde el flexible cuello, parecido al deun emú, era capaz de girar y volverse,elevándose verticalmente cada pocosminutos para coger aire. Esta «hipótesisde la función —dijo Osborn— esaplicable a todos los cetiosaurios, esdecir, también al Morosaurus yDiplodocus[18]».
Los brontosaurios, conocidos por

los paleontólogos de finales delsiglo XIX, tenían cuellos largos como lasserpientes, de aproximadamente 5,5metros de longitud, coronados por unoscráneos ridículamente pequeños coorificios nasales dorsales. Se pensabaque éstos actuaban a modo de excelentestubos de respiración que permitían a losreptiles continuar su existenciasubmarina sin estorbos. Sin embargo,los primeros saurópodos descubiertos,aunque enormes según cualquier estándar, en modo alguno eran losdinosaurios con tubos de respiraciómás grandes que jamás hayan existido.El Brontosaurus de Marsh, de hecho,sólo medía 15 metros de longitud.

Osborn estaba trabajando con u«individuo grande» de Camarasaurus,desenterrado por el equipo del MuseoAmericano de Historia Natural en 1897,una criatura que probablementealcanzaba los 18 metros de longitudaproximadamente.
Después de que Marsh hubieradescrito los primeros escasos huesos de
iplodocus, en cada expedición e busca de fósiles se hacían añicos lasgrandes esperanzas de descubrir uespécimen realmente bueno. El caso deOsbom era típico. Encontró la punta deuna cola de Diplodocus perfectamenteconservada y procedió a seguirla,vértebra por vértebra, acortando el risco

en el cual estaba encastrada. Pero, elugar de que los huesos se hicieran másgrandes y numerosos, se acabarodespués de unos cuantos centímetros.Posteriormente, el Museo Carnegie dePittsburgh se sumó a los esfuerzos paradesenterrar estos prestigiosos gigantes, yfueron los equipos recolectores delmuseo, de 1899 y 1900, los quefinalmente recuperaron el primer esqueleto imperfecto pero reconstruíblede Albany County, Wyoming.
Estas expediciones al Oeste estaba patrocinadas por un hombre, tirano yhéroe de la Era Dorada de América,Andrew Carnegie. Carnegie había idoescalando puestos, pasando de ser u

mensajero telegráfico hasta convertirseen un magnate multimillonario del acero.En una época en la que el acero era elrey y el capitalismo no conocía ningunode los refinamientos de los últimostiempos, la implacable explotación quehacía Carnegie del hombre y de la tierra parecía estar completamente en armoníacon la ética de la época, una ética algusto de Carnegie. Carnegie justificó scarrera contra los contrincantes, sinsaciable exigencia de una mayor producción y el sacrificio de sustrabajadores y colegas en aras de laeficiencia, como ejecución del modelosocial darwinista. La marca de HerbertSpencer del darwinismo social halló u

abogado idóneo en el mundoempresarial, después de la Guerra CivilAmericana, y en Andrew Carnegie e particular: la competencia empresarial,que selecciona los elementos másdébiles de la sociedad industrial,garantizaría la supervivencia de los másfuertes. La nueva ética capitalista,considerada por muchos como una leyde la Naturaleza, era el credo deCarnegie. (Herbert Spencer permaneciódesvinculado del duro mundo descritoen sus escritos, prefiriendo elaislamiento de su hogar inglés. CuandoCarnegie finalmente lo introdujo alvisitar Pittsburgh, ávido de enseñarle sUtopía industrial, fundada sobre los

verdaderos principios de Spencer, eldesinflado filósofo inglés se volvió aCarnegie con ojos tristes y se lamentó:«¡Seis meses de permanencia aquíustificarían el suicidio!»). Como
apologista de su riqueza acumulada,Carnegie hablaba con confianza yenérgicamente: no se consideraba comosu propietario sino meramente como s
uardián. Y disgustando mucho aSpencer, Carnegie obtuvo unasatisfacción aún más grande mediante laredistribución de su fortuna. De hecho,en su Gospel of Wealth, Carnegieadjudicó el primer puesto a sus sabiasdisposiciones (estaba motivado por lacreencia de que sus fundaciones

actuarían como un estímulo para losotros para que se esforzaran tal como lohabía hecho él[19]). Así, en 1895, fundóel Instituto Carnegie en su ciudad natalde Pittsburgh y, poco tiempo después,construyó y equipó un museo. Carnegieinvirtió 25 millones de dólares en elinstituto en los primeros diez años,ayuda que le proporcionó a Pittsburgalgunos de los mejores esqueletos desaurópodo jamás encontrados. Siembargo, el más célebre fue con muchoel Diplodocus.
J. B. Hatcher, el director del museo,intentó realizar en 1901 unareconstrucción en papel del Diplodocus, basada en los últimos hallazgos

encontrados en Wyoming, pero rellenólos huecos con los huesos de muchasotras colecciones. Se pensaba que lacriatura resultante debía tener unalongitud algo inferior a los 20 metros pero, tal como se supo posteriormente,ésta era una estimación demasiado baja.Las extrañas proporciones del
iplodocus se hicieron evidentesinmediatamente tras la restauración deHatcher. Tenía un cuelloextraordinariamente largo que conteníamás de quince vértebras alargadas, unacola con más de 37 vértebras y un troncorelativamente corto en medio. Debido aque el espécimen de Carnegie sediferenciaba de las especies conocidas,

fue bautizado con el nombre deiplodocus carnegiei en honor al patró
del museo. Carnegie estaba encantadocon esto y rehusó que se hicierasolamente una restauración en papel,encargándole a los naturalistas preparadores la tarea de realizar unareconstrucción esquelética de tamañonatural que correría a su costa. Mientrastanto, se habían descubierto en Wyomingnuevos especímenes del Diplodocus deCarnegie que estaban en mejor estado y, por lo tanto, fueron incorporados en lanueva restauración realizada por elsucesor de Hatcher, W. J. Holland. Seconstruyó un esqueleto completo, hechocon yeso mate, que fue expuesto en el

Museo Carnegie. Se supo que muchas delas vértebras habían faltado en larestauración anterior de Hatcher. La colano poseía 37 vértebras. Holland ahoratenía 73, y las carillas articulares de laúltima sugerían que aún no se habíaalcanzado la punta de la cola[20]. Larestauración del Diplodocus carnegiei
en el Museo Carnegie medía más de 26metros de longitud. Con casi 9 metrosmás de longitud que los hallazgosanteriores de Marsh y Cope. Constituía,con mucho, el animal terrestre más largoconocido. En relación con su tamaño,tenía uno de los cerebros más pequeños;una inmensa máquina dirigida por udiminuto centro de control, de modo

similar a como el imperio del acero dePittsburgh estaba dominado por un únicohombre (analogía apoyada por los biógrafos de Carnegie).
Un antiguo fideicomisario del MuseoBritánico, que tuvo no poca influenciasobre el industrial de Pittsburgh, fue elrey Eduardo VII. El monarca inglés sehabía hospedado en casa de Carnegie yse maravilló con su Diplodocus
restaurado, expresando de forma casualla esperanza de que algún día el MuseoBritánico tuviera la suerte de poder adquirir un monstruo de este tipo.Carnegie puso a trabajar inmediatamentea sus naturalistas, haciendo otra serie demoldes de su Diplodocus. En un breve

espacio de tiempo, se había realizado uduplicado completo del dinosaurio dePittsburgh que llegó a su debido tiempoal Museo Británico en enero de 1905.En abril también llegó a Londres elnaturalista preparador jefe de Carnegie asumió el control del montaje de los
aproximadamente 300 huesos. Lacriatura era tan grande que no se pudoencontrar ninguna estancia para ella ela sala de paleontología, tal como era eaquel entonces, de forma que se montóen la galería de los reptiles (que mástarde se convertiría en la galería de losdinosaurios). Los fideicomisariosorganizaron la entrega formal el 12 demayo y la galería rebosaba de

científicos y paleontólogos eminentes(con unos cuantos lores) para queayudaran a celebrar el momento. El profesor Ray Lankester, director delmuseo, presentó los actos mientras queLord Avebury aceptaba el Diplodocus
de Andrew Carnegie de parte de losfideicomisarios[21]. El acontecimientoevidentemente impresionó a Carnegie yaque envió a Holland, en calidad deemisario, a los monarcas de otros paíseseuropeos y su Diplodocus fue presentado brevemente con gran pompaal emperador Guillermo y al presidentede la República Francesa. Los moldestambién fueron enviados a los museos deViena, Bolonia, La Plata, en Argentina, y

a Ciudad de México. En cada ciudad,los naturalistas preparadores deCarnegie acompañaban las cajas dehuesos de dinosaurio de escayola a fide montar la criatura y Holland permanecía el tiempo suficiente parasupervisar la instalación y actuar comorepresentante de Carnegie durante laceremonia de presentación. Este mismo
iplodocus es seguramente el objetoexpuesto más impresionante en muchosde los mejores museos del mundo. Cadauno de los esqueletos tiene la fama dehaberle costado a Carnegie 30 000dólares, sumando los costes de sfabricación, el transporte hasta su lugar de destino y posterior montaje. Con una

promoción tan espléndida, difícilmente puede sorprender que el Diplodocus seauno de los habitantes más conocidos delantiguo mundo.
Durante todo este tiempo, nadie pensó en cuestionar el paso erguido delos saurópodos montados. El
iplodocus y su familia siempre habíasido restaurados en una postura similar ala de los elefantes, con las piernascolocadas debajo del cuerpo. Al igualque en los elefantes, las piernasfuncionaban como puntales paramantener el cuerpo lejos del suelo. La primera reconstrucción de Marsh del
rontosaurus, en 1883, marcó el ritmo.Representaba a la criatura caminando

con el cuerpo elevado por encima delcuerpo, sostenido por piernas coaspecto de columnas. «El ejemplo deMarsh fue seguido desde entoncesservilmente», comentó un disidente,Oliver P. Hay, en Washington, en 1908.De hecho, el Brontosaurus, aun siendouna de las bestias más grandes y pesadasde todos los tiempos, había sidomontado en el Museo Americano deHistoria Natural con una ligera curva esus piernas, haciendo que parecieraestevado. Sin embargo, el Diplodocus
construido por Camegie tenía las piernascompletamente rectas.
¿Por qué en un principio se empleóuna postura similar a la de los elefantes?

Los dinosaurios bípedos fueron los primeros en ser conocidos en América ydominaron por completo los estudiossobre dinosaurios durante muchasdécadas. Cuando Leidy y Copeencontraron criaturas fósiles coenormes patas traseras, inmediatamente pudieron observar una analogía con lafauna de nuestros días. Ésta, siembargo, no se hallaba entre los reptilessino entre los mamíferos. De forma queresulta comprensible que utilizaran elúnico modelo que estaba a sdisposición y restauraran el
adrosaurus y el Laelaps a modo decanguro. Esta imagen de dinosaurioestaba tan arraigada que, cuando Cope

halló el primero de los gigantessaurópodos como el Amphicoelias,automáticamente pensó que éstostambién habían caminado erguidos sobresus patas traseras. Con eldescubrimiento de las patas delanterasigual de macizas se vio forzado a asumir una postura cuadrúpeda para la criatura.Debido a que los dinosaurios bípedoscomo el Hadrosaurus eran capaces decaminar sobre sus patas delanteras —silos miembros estuvieran colocadosdebajo del cuerpo como lo están en loscanguros cuando se desplazan sobrecuatro patas—, Marsh y Cope lo usarocomo su modelo. Simplemente alargarolas patas delanteras y adoptaron una

postura resultante similar a la delelefante. Los saurópodos de Marsh, colas piernas colocadas debajo del cuerpo mirando hacia delante, tenían la
postura que Richard Owen habíaotorgado a sus bestias en Londresmuchos años antes.
Cuando Carnegie distribuyó susespecímenes de Diplodocus (y losmodelos a escala en miniatura) por todoel mundo, por primera vez loscientíficos estuvieron cara a cara con ureptil que tenía aspecto de mamífero paquidérmico. Con la llegada del
iplodocus a Londres, un conocidoilustrador inglés, F. W. Frohawk,claramente vio la paradoja en la

reconstrucción de Carnegie y expresósus dudas en The Field , en 1905:
El visitante de la galería delos reptiles del Museo deHistoria Natural seguramentequedará sorprendido por laextraordinaria postura delesqueleto gigantesco y modeloen miniatura del Diplodocus
carnegiei. Sería interesantesaber la razón por la que se hamontado el espécimen con patastan largas, como algunos paquidermos enormes. Ya que setrata de un lagarto gigante, ¿por qué no se le representa en la

postura generalmente adoptada por estos animales? El esqueletotiene el aspecto de pertenecer aalgún mamífero grande. Elmodelo pequeño realizado por elDr. Holland nos muestra a lacriatura con el cuerpo y losmiembros parecidos a los de unelefante, con un cuello deenorme longitud y una cola que parece la de una pitón inmensa.Parece razonable suponer que unanimal con un cuello de longitudtan prodigiosa difícilmente losoportaría de la manerarepresentada, y que la mayor parte del cuello, cuerpo y

conjunto de la cola probablemente reposarían en elsuelo, y los miembros estaríancolocados más lateralmentecomo en otros reptiles. Sin dudaexiste alguna buena razón paramontarlo y modelarlo en esta postura; en el caso de que asífuera, sería grato recibir algunainformación al respecto[22].
Al año siguiente se hizo un intentode reconstruir uno de estos dinosaurioscon miembros desparramados, en estecaso el Brontosaurus. Dos asistentes,Otto y Charles Falkenbach, del

laboratorio paleontológico del MuseoAmericano de Historia Natural,construyeron un pequeño modelo de
rontosaurus tomando como base a loscocodrilos. Los miembros deldinosaurio estaban orientados haciafuera, de forma que la criatura parecíaestar arrastrándose por el suelo con elvientre ligeramente elevado por encimadel mismo. Este modelo fue exhibido ela reunión de la Sociedad Americana dePaleontología de los vertebrados quetuvo lugar en el museo ese año pero, por común acuerdo, se llegó a la conclusióde que representaba algo imposible.
El Diplodocus de Carnegie es elresponsable de precipitar la verdadera

sublevación, una sublevación que seharía tan internacional como el propio
iplodocus. Mientras Holland estabahaciendo la presentación al presidentefrancés, Hay —irónicamente, él mismoera receptor de una beca de otro de los beneficiarios de Camegie, el InstitutoCamegie de Washington—, desafió laelección de la postura de esta criatura(independientemente parece de losauxiliares del Museo Americano). Esmuy improbable, dijo, que el
iplodocus adoptara una posturaerguida como la de un elefante. Loscontemporáneos de Hay habían usado elenorme peso del animal como argumento
ara las piernas rectas. Con columnas a

modo de pilares para sostener el cuerpo,el peso se transmite directamente através de las piernas al suelo. El pesode un animal se incrementa como elcubo de su longitud, mientras que sfuerza varía solamente como la secciótransversal o el cuadrado. Cuandoaumenta el tamaño, llega un momento eel cual el peso del dinosaurio finalmentesupera a su fuerza para permanecer erguido, especialmente si las piernasestán torcidas. El desarrollo decolumnas de soporte rectas es el mediomás eficaz para sostener este enorme peso. Hay, sin embargo, mantenía que elexceso de peso era un argumento en
contra del modo de andar similar al de

los mamíferos. Los dinosaurios no eramamíferos sino reptiles, y los reptiles nocaminan como los elefantes sino que searrastran como los caimanes. «La ideade un dinosaurio reptador eradifícilmente concebible», se lamentóHay. No obstante, con todo el peso quedebía soportar, el Diplodocus debía dehaber tenido extremidades extendidas deforma que la mayor parte del tiempo podía estar tumbado sobre el vientrecomo un caimán. Los pies, supuso,estaban dirigidos hacia fuera en lugar dehacia delante, mientras que la rodilla ylos codos se hallaban formando uángulo recto como en los lagartos. Si el
iplodocus vivía en regiones

pantanosas, además, estaría mejor dotado si se deslizaba como loscocodrilos. Al ser tan pesado, si hubiesecaminado erguido sobre terrenos pantanosos pronto se habría encenagadoinextricablemente y hubiese perecido deforma miserable. Continuó diciendo:
Es difícil entender por quéun animal cuyos antepasadosinmediatos seguramente sedesplazaban como loscocodrilos, un animal que eraestúpido y probablemente lentode movimientos, un animal quemediante su largo cuello podíaalcanzar la superficie desde el

fondo y llegar desde lasuperficie hasta el fondo, por qué un reptil de este tipo tendríala necesidad de desarrollar lacapacidad de vadear los fondosde los ríos como losmamíferos[23].
El Diplodocus era una criatura perezosa (no por elección sino por necesidad) y podía arrastrarse fuera delagua sólo haciendo un gran esfuerzo. Sverdadero hogar eran los pantanos.
aturalmente, al Diplodocus coaspecto de cocodrilo le faltaba algo dela novedad inherente a un ser elefantino

pero esto se podía remediar:
El escritor tiene la impresiónde que nuestros museos,ocupados haciendo soportes yrestauraciones de los grandessaurópodos, han dejado pasar laoportunidad de construir algunasrepresentaciones curiosas deestos reptiles que secorresponderán más a suverdadera naturaleza. El cuerpocolocado en una postura típicade los cocodrilos sería menos,incluso nada impresionante, encomparación con una posturaerguida; mientras que el cuello

largo, flexible como el de unavestruz, podría ser colocado engran variedad de posicioneselegantes.
Hay dio vida a su Diplodocus en udibujo que realizó en 1910[24]. Laescena que evocó podía parecerlerealmente extraña a una generación quese había erigido sobre saurópodoselefantinos aunque probablemente nomás extraña que el monstruo elefantinode Carnegie que vieron los primerosvisitantes ingleses.

k. El Diplodocus «reptante» de Hay,restaurado según especificacionescaracterísticas de los cocodrilos.
A consecuencia de la difusión deliplodocus de escayola por todo el
mundo se alzó una gran controversia. E

Viena, Othenio Abel se inclinaba a favor de la postura similar a la de losmamíferos mientras que, en Alemania,Gustav Tornier se puso al lado de Hay, prefiriendo un cocodrilo que umamífero. Debido a que el Diplodocus
es un reptil, arguyó, nunca debería haber sido reconstruido en su primera formacon aspecto de mamífero y, como pruebade que la postura de cocodrilo no eraimposible, hizo un esbozo de larestauración. Otros paleontólogosalemanes apoyaron la teoría de Tornier yse burlaron de sus homólogosamericanos, por haber hecho unareconstrucción errónea de la criaturadesde la cabeza hasta los dedos del pie.

Esto fue demasiado y los paleontólogosamericanos, puestos en ridículo,lanzaron una ofensiva dirigida contra los«naturalistas alemanes más limitados»como Holland los llamaba. Con la ayudade un lápiz, dijo Holland, Tornier habíadislocado las articulaciones y torcidolos huesos, configurando una formagrotesca para el dinosaurio. Smonstruosidad habría sidocompletamente incapaz de realizar movimiento alguno. Dos décadas detrabajo de los estudiosos americanosmás eminentes del dinosaurio habíasido borrados con éxito de un plumazo por Tornier. «Como contribución a laliteratura de la caricatura, el éxito

logrado es notable», fue la evaluacióridiculizante de Holland sobre la«monstruosidad esquelética» deTornier [25]. La única característica pararecomendar esta reconstrucción, dijouno de los paleontólogos europeos, eraque justificaba la rápida desaparición delos saurópodos. ¡Teniendo todas lasarticulaciones dislocadas, estosanimales debieron de haber sufrido unaindescriptible agonía en vida!

l. Corte transversal a través del esqueleto deun Diplodocus de 10 toneladas restaurado en postura de lagarto. El intento de hacer del Diplodocus un dinosaurio de extremidadesdesparramadas fue un error fatal. Tal comoobservó sarcásticamente Holland, la profunda caja torácica del dinosaurio habría

requerido un sendero en el suelo para que elanimal pudiera reptar.
m. La «monstruosidad esquelética» deTornier. El deforme Diplodocus, con todaslas articulaciones dislocadas, debió de haber tenido una vida de indescriptible agonía.
El gesto de buena voluntad deCamegie había degenerado en una

discusión violenta internacional con no poco orgullo herido por ambas partes.La «cruelmente convincente polémica»de Holland, tal como la llamabaacertadamente Matthew, en efectosilenció a Tornier y a sus seguidores. Elgran conocimiento de Holland de losdinosaurios y de los reptiles modernosno era superado por nadie. El detalladoestudio de las articulaciones de losmiembros, cinturones y pies le llevó aafirmar, de forma bastante categórica,que el Diplodocus era «rectígrado», quecaminaba erguido sobre extremidadescon aspecto de pilares dirigidas haciadelante. El tamaño enorme de losdinosaurios de este tipo había hecho

necesaria la presencia de extremidadescolosales de soporte.
Las ideas de Holland fueroustificadas completamente a finales de
1930, cuando Roland T. Bird encontrólas primeras huellas de saurópodos. E1938 encontró en Glen Rose, Texas, unasucesión completa de huellas impresasen el barro cretáceo dejadas por u brontosaurio de 18 metros de longitud.La zancada del monstruo medíaaproximadamente 3,6 metros delongitud, aunque la anchura de lashuellas era de sólo 1,8 metros[26]. Si el brontosaurio hubiese reptado como ucocodrilo con las piernas extendidas aambos lados del cuerpo, la anchura

hubiese tenido que ser mucho mayor.Ésta era la prueba inequívoca de que los brontosaurios y dinosaurios similaresandaban firmemente erguidos.
La controversia se había convertidoen historia. Desde nuestra perspectivaalternativa, y con la ayuda de una nuevaserie de presuposiciones que apoyanuestro punto de vista de losdinosaurios, podemos apreciar la paradoja con la que se encontraronuestros predecesores. Los reptiles no pueden mantener un elevado gastoenergético durante mucho tiempo y, por consiguiente, se ven obligados adescansar sobre sus vientres gran partede sus vidas. Esto es válido incluso para

los lagartos pequeños de pesoinsignificante. El problema de Hay eraexplicar cómo una criatura cuyo pesoera enormemente superior al de ulagarto podía permanecer erguida. Losdinosaurios más grandes, como el
rontosaurus y el Brachiosaurus, pesaban entre treinta y ochenta toneladascuando eran adultos (las estimacionesdel peso en vida varíaconsiderablemente). Hay habría tenidorazón al creer que los dinosaurios nunca podrían haber permanecido de pie si setrataba de reptiles como los lagartos.Sin embargo, no lo eran. Los saurópodos probablemente eran animales de sangrecaliente con un metabolismo que

producía una gran cantidad de energíaque les proporcionaba la producción deenergía continua que necesitaban paramantenerse erguidos soportando un pesotan enorme. Se necesita un esfuerzoconstante para mantener el cuerpo por encima del suelo y la energía de la quedisponían los dinosaurios debió de ser mucho mayor que la disponible para ulagarto postrado o cocodrilo. Losdinosaurios adoptaban una posturasimilar a la de los mamíferos probablemente porque tenían unafisiología parecida a la de losmamíferos. Es difícil imaginar otra cosa.
La prueba para cualquier teoría buena consiste en su capacidad para

explicar hechos previamente inconexos.Encontramos que la endotermia nosolamente explica la postura de losdinosaurios sino que también nos ayudaa comprender por qué crecían tanto. Losdinosaurios no presentaban aislamientoalguno (excepto placas o escamas óseasen algunos) y perdían calor a través desu curtida piel. Cuanto mayor era elcrecimiento del dinosaurio, más pequeña se hacía el área de superficieen relación con el volumen. Por lo tanto,una bestia más grande pierderelativamente menos calor que una más pequeña. Un elevado índice metabólico
la piel desnuda les impidieron llenar el micronicho ocupado por los lagartos

pequeños mamíferos insectívoros. Loslagartos tienen bajos índicesmetabólicos y pueden tolerar temperaturas más bajas mientras que losmamíferos presentan un abrigo de pielaislante.
Los dinosaurios nunca podrían haber mantenido una temperaturaconstantemente alta si hubiesedisminuido de tamaño por debajo de univel crítico y hubiesen perecido por laexposición. ¿Es por lo tanto de extrañar que nunca se hayan encontradomicrodinosaurios? Los dinosauriosestaban obligados a ser gigantes.
El reciente descubrimiento de udinosaurio sin precedentes de 100

toneladas de peso crea otro problema.Si esta bestia inimaginable hubiesecrecido uniformemente a lo largo de svida como los reptiles, habríatranscurrido muchos siglos antes de quehubiese alcanzado su tamaño final. Dehecho sabemos que los dinosaurios novivían tanto tiempo (contando losanillos de crecimiento en sus huesoslargos). Más bien el estudio de Ricqléssobre los huesos de dinosaurio sugiereque los dinosaurios jóvenes crecían a uritmo rápido, alcanzando rápidamente lamadurez como los mamíferos y las avesóvenes. Esto explica la escasez de
fósiles; la mayor parte de la vida deldinosaurio transcurría siendo el animal

a adulto. Sin embargo, el metabolismode un reptil simplemente no puedesoportar estos brotes sostenidos decrecimiento juvenil: una prueba más deque los dinosaurios producían vástagosactivos y vigorosos.
La mayoría de los expertos edinosaurios han pensado que, a fin de poder soportar tantas toneladas demúsculos y huesos, las extremidades eforma de columnas de los dinosauriosdebieron de haber recibido algunaayuda. Si el elefante es el mayor tamañoque los mamíferos terrestres actuales pueden manejar, preguntó Matthew,

¿cómo los dinosaurios pudieron alcanzar un tamaño tan superior? Suponía que, aligual que las ballenas, los saurópodoscontaban con el agua para poder flotar.Aligerado de esta forma el peso de los pies, estos animales fueron capaces decrecer alcanzando proporcionesgigantescas[27]. El Diplodocus, con ucuello que medía hasta 6,4 metros delongitud, era un saurópodoextremadamente largo. Esta bestia podría haber continuado respirandohallándose en aguas de más de 9 metrosde profundidad, siendo éste el lugar donde generalmente se encontraba. (Eeste sentido, el trotamundos Diplodocus
fue aventajado con mucho por el

rachiosaurus, desenterrado sólo pocosaños más tarde en Tanzania en ÁfricaOriental. Aunque no era tan largo comoel objeto de valor más preciado deCarnegie, el Brachiosaurus
probablemente pesaba hasta ochentatoneladas en vida y las extremidadesdelanteras, desproporcionadamentelargas, se combinaban con un sinuosocuello que sostenía la cabeza que sealzaba a unos 12 metros de altura). Aligual que el Brachiosaurus, el
iplodocus presentaba unos orificiosnasales en la parte superior de la cabezasituados entre sus ojos. También eraconsiderado una criatura de algunamanera indefensa, razón de peso

suficiente como para dejar aliplodocus en el agua donde se supone
que nunca se adentraban los carnívoros.El Diplodocus parecía tener sólo
unos dientes con aspecto de rastrillo yescasos alvéolos situados sólo en la parte delantera de la boca. Se pensabaque este aparato de masticación taineficaz funcionaba cogiendo las blandas y suculentas plantas acuáticasque los pies habían arrancado del fondodel lago. Esto era al menos comoHatcher pensaba que la criatura obteníasu comida. También describió elambiente en el cual vivía el Diplodocus.En la época jurásica, la región queactualmente es Colorado, Wyoming y

Montana probablemente se parecían a lacuenca tropical del Amazonas, conumerosos lagos y ríos largos rodeados por una densa vegetación tropical covalles extensos y llanos sometidos ainundaciones periódicas[28]. En mediode este exuberante follaje y en un climacaluroso y húmedo, vivía el enorme
iplodocus buscando constantementevegetación suculenta. Desgraciadamente,el extraordinario desgaste observado elos dientes de la mayoría de cráneos de
iplodocus difícilmente se puedeatribuir a una vegetación blanda,acuática. Los dientes grandes coaspecto de estaca que frecuentementemedían hasta 4 centímetros de longitud,

estaban completamente desgastados y planos, formando un ángulo recto en losextremos y, debido a que los dientes delos maxilares superiores e inferiores nose rozaban unos con otros, hemos de buscar otra explicación. Holland sugirióque si estos dinosaurios eraherbívoros, como todo el mundo pensaba, el desgaste podría estar producido porque los animales seraspaban los dientes en la superficie de piedras mientras que arrancaban las plantas. Sin embargo, se inclinaba más a pensar que la materia áspera de lostroncos de las cicádidas, tan abundanteen las junglas del mesozoico, era sdieta corriente. Los dientes

peculiarmente aplanados se aventuró adecir, se producían cuando los animalesroían los troncos de los árboles paraintentar llegar a la médula.
Resulta misterioso cómo estosdinosaurios obtenían suficientesalimentos para satisfacer lasnecesidades de sus enormes cuerpos,aunque el hecho de que tuvieran tantoéxito atestiguaba la falta de problemas para los saurópodos. Gustav Tornier sugirió que comían alimentos animales yque no eran en absoluto herbívoros.Esto, de hecho, les hubiese suministradolos alimentos nutritivos de forma muchomás compacta y probablementenecesitarían consumir menos para

satisfacer sus necesidades diarias deingesta de comida. La nueva sugerenciade Tornier era de que se trataba decomedores de pescado (sólo podíahacer pasar peces a través de susdiminutos cráneos), aunque más tarde se propuso que comían cantidades enormesde moluscos bivalvos de los cuales sesabe que vivían en abundancia a lo largode las orillas de los lagos jurásicos. Siel Diplodocus arrancaba los crustáceosdel fondo del lago, somos tambiécapaces de explicar el extraño desgasteque presentaban los dientes de losanimales, desgaste sugestivo delmaterial duro y abrasivo de las conchasde los mejillones. El Diplodocus, dijo

Holland, podría haber cogido losmejillones con sus dientes frontalescortantes y haberlos arrancado del fondodel lago. Es una hipótesis ingeniosa.Explicaría cómo una bestia gigante podía ingerir suficiente comida a travésde una cabeza diminuta y por qué losdientes estaban desgastados de forma ta peculiar. Desafortunadamente, a pesar de una intensiva búsqueda, Hollandnunca pudo encontrar montículos deconchas de mejillones tragadas en laregión de los fósiles donde se suponíaque se hallaba el estómago. Sabía de laexistencia de varios especímenes de
iplodocus que tenían las vértebras ycostillas en su sitio, pero nunca se halló

algún tipo de contenido visceral. «Por lotanto, hasta el momento no existeninguna confirmación de la teoría deTornier —suspiró Holland añadiendo—:ni tampoco, si vamos a eso, de laafirmación del profesor Marsh de queestas criaturas eran herbívoras. Confiesohallarme en la situación de vermeobligado a comunicar que soy uagnóstico en lo que se refiere a la formade cómo estas criaturas obtenían scomida[29]».
Debido a que los dinosaurios vivíaen el agua, siempre se pensaba que lacomida debía ser acuática. Hubo una odos voces que estaban en desacuerdocon esta teoría. Henry Fairfield Osborn,

del Museo Americano, observó que lasvértebras de los saurópodos eraligerísimas, siendo huecas y por lo tantodistintas a las de las ballenas. El MuseoBrooklyn tenía un esqueleto de ballenade 23 metros de largo y un peso de casi80 toneladas. Osborn estimaba que uesqueleto de saurópodo de la mismalongitud pesaba poco más de la mitad deese valor. «El hecho de que el esqueletosea ligero es una adaptación sufrida paraandar, correr o volar —añadió— y no para poder nadar; los animalesnadadores necesitan la gravedad de sesqueleto debido a que, en el agua, lanecesaria capacidad de flotaciósiempre es proporcionada por los

pulmones y los tejidos blandos delcuerpo». Sin embargo, Osborn no quisoseguir esta línea de razonamiento hastallegar a su conclusión lógica, sino quellegó a un compromiso declarando quela «extraordinaria ligereza de estasvértebras de dinosaurio podrían ser consideradas como una prueba de lasuprema capacidad de propulsión de uenorme esqueleto durante lasincursiones ocasionales haciatierra[30]». A los dinosaurios sólo se les permitía salir fuera del agua durante breves espacios de tiempo, según lo quese creía para poner huevos o paramigrar a una charca cercana. Osborn, aligual que Cope en sus anteriores

especulaciones, se imaginaba a losanimales parando para ramonear en lascopas de los árboles en su camino haciaun nuevo lugar de permanencia en elagua, e incluso los veía estirándose,apoyados sobre sus patas traseras, paraalcanzar las hojas más altas. En una posición como ésta, los herbívoros se podrían haber defendido contracarnívoros más pequeños, alzándosecuando fueran atacados para asestar uterrorífico golpe con sus macizas patasdelanteras[31]. Sin embargo, éste no erael medio ambiente preferido por lossaurópodos: la criatura sólo se sentíacompletamente a gusto en el agua. Elreconocimiento de que tuvieron que salir

a tierra para la reproducción fue uimportante paso, aunque sobre lacuestión de la puesta de huevos noexistía una opinión unánime. Matthewsostenía que los saurópodos podríahaber sido vivíparos como losictiosaurios, trayendo al mundo (y alagua) crías vivas.
Estando fuera del agua,indudablemente estaban expuestos alataque de los carnívoros, tal como prueban los restos de al menos un festí prehistórico. Un equipo del MuseoAmericano encontró huesos de brontosaurio del jurásico que presentaban muescas de dientes, a partir de las cuales montaron sus alosaurios

inclinados sobre los restos de esteesqueleto de saurópodo marcado cocicatrices. Debido a que los alosauriossiempre han sido considerados comoanimales terrestres, estos banquetesdebieron de tener lugar en tierra dondeel brontosaurio había muerto de formanatural o había sido abatido por losalosaurios. Cualquier duda concernientea los hábitos anfibios de los brontosaurios fue disipada de formaespectacular por el descubrimiento deBird de una serie única de huellas. Birdera escéptico respecto a la capacidad delos dinosaurios de caminar sobre tierrafírme. El Brontosaurus, cuyo nombreliteralmente significa «lagarto de

trueno», desde luego habría hecho tronar el suelo bajo sus pies cuando pasaba, yaque pesaba lo mismo que media docenade elefantes. Las huellas encontradas eTexas habían quedado impresas en barroduro, que por sí mismas, no nos proporcionan mucha información sobresi los animales caminaban sobre tierra oen el agua. La primera pista de quehabía al menos algo de agua se obtuvocuando, en el barro, no se pudierohallar surcos de colas; la cola debía dehaber flotado detrás del dinosaurio quecaminaba por el agua. Si no hubiesehabido agua, se arguyó, la cola con todaseguridad se habría arrastrado por el barro dejando un surco. Bird sólo llegó

a encontrar un surco de este tipo aunqueexisten pocas evidencias de queestuviera producido por la cola de u brontosaurio. ¿Es que los dinosaurios erealidad nadaban? Bird halló unashuellas notables en Bandera County, eTexas, que proporcionan la pruebainequívoca de que al menos udinosaurio nadaba en el agua profunda.Las huellas pertenecen sólo a las patasdelanteras ligeramente impresas en elfango; el animal estaba flotando yavanzaba dando patadas en el fondo dellago con los miembros delanteros. Acontinuación aparece, repentinamente,como si surgiera de la nada, unaimpresión única de una pata trasera y el

animal cambia de dirección. El brontosaurio había hecho un giro en elagua dando patadas en una direcciódiferente con las patas traseras. Ésta erala prueba completamente independientede que los brontosaurios podíanadar [32]. Estos descubrimientosdesterraron como herejía las opinionessobre los brontosaurios, braquiosaurioso el Diplodocus considerándoloscualquier cosa menos moradores de los pantanos o ciénagas, que sólo podíasalir a tierra para reproducirse. Emuchos casos, los saurópodos erarelegados a las profundidades de loslagos, sacando la cabeza cada pocosminutos para tomar aire.

Mientras tanto, en otra rama de laciencia se estaban realizando estudioscompletamente independientes. Losfisiólogos alemanes de principios deeste siglo, estaban intentando averiguar la presión máxima que un ser humano podía soportar bajo el agua siendo aúcapaz de respirar. Los voluntarios eraatados con correas a tablas de madera ysumergidos en piscinas en una posicióhorizontal, permitiéndoles respirar através de un tubo que salía del agua.Robert Stigler, que inició este tipo deinvestigaciones en el Instituto deFisiología de la Universidad de Vienaen 1911, descubrió que los sujetos(frecuentemente él mismo) era

físicamente incapaces de respirar si latabla se sumergía a más de 90centímetros de profundidad, haciéndosela presión ejercida sobre los pulmonesdemasiado grande. El propio Stigler intentó la misma proeza a una profundidad 1,8 metros, pero lacatastróficamente elevada tensióarterial creada a una profundidad tagrande lesionó permanentemente scorazón. Los fisiólogos británicoscomprobaron los resultados de Stigler,añadiendo que los sujetos sólo podíarespirar cómodamente a una profundidadde 25 centímetros, a partir de los cualesla respiración se volvía dolorosa. La primera (y al parecer la única) que se

dio cuenta de la importancia de estosestudios fisiológicos realizados en sereshumanos, para los, en apariencia,remotamente relacionados dinosaurios,fue Kenneth Kermack del UniversityCollege de Londres. En 1951, Kermack tomó los resultados de los fisiólogosalemanes y los aplicó a los grandessaurópodos con consecuenciasinesperadas (inesperadas para elsistema basado en puntos de vistatradicionales que no habían cambiadodesde la época de Cope aunque, silugar a dudas, Kermack ya habíaanticipado los resultados). Algunos delos saurópodos con largos cuellosalzados verticalmente podían alcanzar

una altura de 12 metros, y se pensabaque en esta postura los animales estabade pie sobre el fondo del lago actuandosus largos cuellos como tubos derespiración. Los animales tienen queexpandir el tórax y los pulmones pararespirar. Así, los pulmones de los
iplodocus habrían estado a una profundidad de aproximadamente 8metros (a pesar de que el Brachiosaurus
era más grande en conjunto, losmiembros delanteros largos elevaban eltórax aproximándolo más a la superficie,situando los pulmones a la misma profundidad, más o menos, que en el
iplodocus). A esta profundidad, la presión es de aproximadamente 8 libras,

la cual, si no aplastaba la tráquea (eltubo que pasa a los pulmones), sin dudahabría impedido la expansión de los pulmones. ¡Para poder respirar co presiones tan altas, el músculorespiratorio tenía que haber empleadouna fuerza inmensa para tantas toneladasde peso[33]! Los problemas de lossaurópodos no acababan aquí. A fin deevitar el colapso y permanecer funcionales, los vasos sanguíneos debíatener una tensión arterialcatastróficamente alta, estando el producto de la tensión arterial normalcompuesto por la presión externa.Debido a que el aire en el interior de los pulmones (suponiendo que fuera

capaces de resistir el efecto aplastantedel agua) tenía una presión atmosféricanormal, la sangre de los vasos pulmonares habría explotado dentro delos pulmones. Esta forma de aliviar lagran presión no podría haber sidoresistida por los tejidos pulmonares y sehabrían producido hemorragias fatales.(Siempre que, como en el desafortunadocaso de Stigler, el corazón de lossaurópodos no hubiese quedado primeroirrevocablemente lesionado).
Kermack concluyó que era«extremadamente improbable» que lossaurópodos hubieran logrado lo que lossujetos de Stigler no fueron capaces deconseguir. Incluso viviendo en aguas

poco profundas, las criaturas habríatenido que alzarse fuera del agua pararespirar. De esta forma, Kermack socavó completamente la hipótesis de la«nariz acuática»: debido a que losanimales tenían que salir fuera del agua para respirar, la nariz difícilmente podíahaber sido un mecanismo de respiració para aguas profundas o un tubo derespiración. Las teorías de Kermack, publicadas en 1951, eran completamenteopuestas al dogma imperante, dogma queno había tenido en cuenta la fisiologíade los animales. Colbert contestó aldesafío. «Por analogía con el hombre, lalógica de Kermack es razonable. Siembargo, se podría cuestionar la

conveniencia de basar este argumentosobre nuestro conocimiento de laslimitaciones humanas en el agua.Después de todo, el hombre no es uanfibio o vertebrado acuático[34]».Colbert, a continuación, procedió a usar un animal acuático como modelo parademostrar que las criaturas puedesobrevivir bajo el agua. Las ballenas,señaló, pueden respirar en la superficiemientras el cuerpo está inclinadoformando un ángulo agudo, dejando los pulmones a cierta profundidad. Siembargo, como el propio Colbertadmite, las ballenas y marsopas presentan una coraza de proteccióalrededor de los tubos de respiració

para evitar el colapso. Quizá, sugirióColbert, «los dinosaurios saurópodos presentaran algunas modificaciones eel aparato respiratorio que también loscapacitaban para respirar mientras elcuerpo estaba completamente sumergidoen el agua».
Kermack siguió teniendo sus dudas.Las ballenas emergen oblicuamente,admitió, pero el silbido que se oyecuando el orificio nasal sale a lasuperficie no se debe a que los pulmones espiran aire sino a la gra presión, existente a esa profundidad, quelo expulsa a la fuerza. Una ballena esuna criatura marina demasiadoespecializada como para ser utilizada

para la comparación con losdinosaurios. Las modificacionesestructurales tales como la pérdida delos miembros para caminar seacompañan de grandes adaptacionesfisiológicas a la presión del agua. Lossaurópodos eran bestias cuadrúpedas, eincluso los defensores acuáticos másapasionados están de acuerdo en que elanimal podía caminar sobre tierra, aún a pesar de que prefería vivir en losestanques. Las ballenas sólo estáadaptadas a un tipo de vida, la delocéano; fuera del agua se ahogan debidoa que los pulmones no se puedeexpandir sin la ayuda de la capacidad deflotación proporcionada por el agua. Los

dinosaurios son tetrápodosespecializados para la vida sobre tierra difícilmente son comparables a los
mamíferos marinos másespecializados[35].
Las refutaciones como la deKermack no tienen una accióinstantánea; la Ciencia sólo abandonasus teorías mimadas con muchareticencia; la vena conservadora prolonga la vida de las teoríastradicionales, incluso después de queéstas se hayan vuelto insostenibles, aligual que el impulso adquirido por ucoche cuando es conducido demasiadode prisa y es incapaz de parar abruptamente. Kermack posiblemente

sembró, en sus propias palabras, una«grave duda» sobre el modo de vidahabitualmente aceptado para lossaurópodos, pero la revisión de loslibros de texto, publicados dos décadasmás tarde, demuestra la poca atencióque recibieron sus teorías. Seguimosencontrando saurópodos representadosde pie en los fondos de los lagosarrancando plantas acuáticas con sus«débiles» dientes[36]. «La mente humana —escribió Antoine Lavoisier, elquímico francés guillotinado durante laRevolución Francesa— se ve doblegadaa una forma de ver las cosas». Se podríaañadir que la resultante mente colectivasufre no menos ya que continúa,

mediante adoctrinamiento, de generacióen generación. Aquí se impone uejemplo chocante. Desde el comienzo dela época victoriana, cuando Owecimentó su reputación en base a u«lagarto ballena» acuático, lossaurópodos han sido perseguidos por esta imagen acuosa. Si Cuvier nohubiese nunca pontificado sobre lascostillas de ballena, ni Owen sobre los«lagartos ballena», es concebible quehasta hoy no habría existido un solo brontosaurio con tubo de respiracióque viviera en los fondos de los lagos.Sin embargo, esta opinión no esobjetiva. Más bien, esta tendencia surgióa raíz de, lo que retrospectivamente

parece ser, una serie de interpretacioneserróneas y los saurópodos «cetáceos»rápidamente se convirtieron en unaverdad científica indiscutible. Estavisión sobrevive hasta nuestros días. El pensamiento científico de cualquier época debe más de lo que se piensa a s pasado, y sólo comprendiendo el origede las creencias heredadas podemosapreciar la continuidad de su valor.
Durante veinte años, Kermack nohalló apoyo alguno. El cambio de laopinión general se produjo en 1971cuando Robert T. Bakker presentó unavisión radical del brontosaurio en uartículo publicado en Nature[37] (aunqueésta no era la primera vez que había

expresado sus ideas). Bakker se habíadesviado aún más de las ideastradicionales, afirmando que los brontosaurios eran habitantes de lasllanuras y de los bosques como loselefantes y las jirafas. Reunió una gracantidad de pruebas para la crítica delas teorías imperantes, socavandosucesivamente todos los argumentos afavor de una existencia acuática. Erelación con la situación elevada de losorificios nasales, Bakker argumenta queesto también se puede observar en laiguana terrestre y los lagartos monitoresdel desierto. Además, los animalesacuáticos hacen poco uso de su sentidodel olfato y sólo poseen unos orificios

nasales pequeños. En los brontosaurios,el órgano nasal y su abertura en el techodel cráneo son grandes; obviamente presentaban un sentido agudo del olfato.El gran desgaste de los dientesobservado en muchos brontosaurios,sugiere que se alimentaban, no de plantas acuáticas blandas, sino de la basta vegetación terrestre. Sin embargo,el principal soporte de la teoría deBakker lo constituía la similitudexistente entre la construcción de los brontosaurios y de los elefantes. Losanimales acuáticos, como loshipopótamos, presentan miembros cortos un tórax poco profundo, a diferencia
de los brontosaurios y elefantes que

tienen miembros largos y un tóra profundo. Los hipopótamos, que flotaen el agua, tienen miembros débiles,flexionados en las rodillas y los codos.En contraposición, las patas con aspectode columnas de los brontosaurios se parecen a las de los elefantes y debierode ser utilizadas para transportar lacriatura por tierra firme. Si los brontosaurios eran moradores del aguano hubiesen necesitado unos miembrostan macizos. Sus pies tampoco estabaadaptados para caminar sobre el barro;los dedos de los hipopótamos estáextendidos para lograr un mayor soporteen el suelo blando, mientras que loselefantes y los brontosaurios tiene

dedos cortos revestidos de unasalmohadillas carnosas (las huellas desaurópodos también presentan estascaracterísticas). En conjunto, parece quelos brontosaurios vivieron una vidasimilar a la de los elefantes en la épocamesozoica. ¿Podemos aproximamos aúmás con nuestra analogía?
¿Por qué los brontosaurios poseíacuellos largos si éstos no se hacíaservir a modo de tubos de respiración?Los miembros largos y un cuello largose dan en los animales terrestres, comola jirafa, donde se utilizan en gramedida para incrementar la capacidadde forrajear, proporcionando una graventaja en la búsqueda de comida. La

irafa es capaz de ramonear las hojasmás altas de un árbol a las que no llegalas otras criaturas y, sin duda, los brontosaurios también encontraroventajoso disponer de este ampliomargen vertical. Esto hacía posible que pudieran ramonear entre las copas de lascicádidas y pinos comunes de su mundo(las cicádidas y las coniferas eran muyabundantes en la flora de Morrisomientras que apenas se sabe de laexistencia de vegetación pantanosa). Lascriaturas acuáticas como loshipopótamos, por el contrario, socompactas y presentan unos cuellos ymiembros cortos debido a que las plantas acuáticas son más abundantes e

las aguas poco profundas.Si los saurópodos hubiese
escapado al agua en caso de peligro, siduda habrían sellado su propio destino.Los alosaurios que los perseguíaestaban provistos de extremidadestraseras largas y fuertes con dedosextendidos, el tipo de pie que está bieadaptado para proporcionar fuertesgolpes en el agua. Los saurópodos no podrían haber escapado de estoscarnívoros nadando ni tampococorriendo. Probablemente, lossaurópodos formarían grupos pararechazar a los atacantes, quizá, tal comosugirió Osbom, alzándose sobre sus patas traseras y usando sus patas

delanteras como armas formidables.El modelo de jirafa del
brontosaurio, modelo que Cope se vioobligado a abandonar por falta de pruebas, ha sido rehabilitadocompletamente por Bakker. A raíz de larevisión benévola del artículo de Bakker publicada en la misma edición de
ature, se hace patente que esteconcepto del saurópodo está logrando laaprobación general. Nature es una de lasrevistas científicas más importantes, deforma que cualquier opinión favorablede sus augustas columnas es de no pocaimportancia. El ataque anatómico deBakker, afirmó el artículo de fondo,dejando fuera de toda duda la

conversión del crítico, «destruyecompletamente» y de una vez por todasla hipótesis acuática[38]. Desde luego el peso combinado de los argumentos deKermack y Bakker es impresionante.
Sin embargo, aún persiste una prueba firme en contra de esta teoría.¿Cómo podemos explicar la fotografíade las impresiones aisladas de las patasdelanteras de un brontosaurio realizada por Bird? Este hecho por sí solodemuestra que los brontosauriosdebieron de aficionarse al agua al menosen algunas ocasiones. Quizá, como loselefantes, se sumergían regularmente para evitar que la piel se agrietase y para eliminar los parásitos externos. La

mayoría de los mamíferos grandesvisitan regularmente charcas de agua.Desgraciadamente, las huellas de brontosaurios, por su naturaleza, sólo pudieron quedar impresas en el barroalrededor de estas charcas, de forma quesería de esperar que encontráramos pruebas de que los animales eocasiones se revolcaban en el barrocomo los elefantes. Pero esto apenasustifica que los brontosaurios sea
declarados animales acuáticos al igualque los elefantes. El debate sin dudacontinuará.
Bakker concluyó su informe sobrelos brontosaurios con una nota sobre elrara vez mencionado papel ecológico

desempeñado por estas bestiasmajestuosas.


Poco después de localizar las primerashuellas de saurópodos en Glen Rose en1938, Roland T. Bird descubrió estaespectacular huella que fue interpretadacomo un brontosaurio perseguido por uncarnívoro. Las huellas de brontosaurio de 90centímetros de largo, están flanqueadas a laizquierda por las huellas de tres dedos de uncarnívoro del tipo Allosaurus.

Las investigaciones de Baker sobre lamecánica de los miembros de losdinosaurios sirven para destacar el parecidoentre los dinosaurios y los grandesmamíferos. El ceratópsido Chasmosaurus
alcanzaba los 5 metros de longitud y se

parecía mucho a los rinocerontes, y probablemente compartía la disposición aatacar de los rinocerontes.
A principios del triásico, hace 210 millonesde años, los pequeños tecodontes pseudosuquianos se convirtieron en los primeros animales que caminaron erguidos.Aunque el hecho de colocar las patas debajodel tronco incrementaba la zancada, ello solo podía ser aprovechado bien si se podíamantener la producción rápida de energía. El

Euparkeria (arriba) estaba transformandosimultáneamente la postura y elmetabolismo. Los tecodontes emparentadosfueron los responsables de la explosión delos dinosaurios en detrimento de los reptilessemejantes a los mamíferos.
El Cynidionathus, que vivió a comienzos deltriásico, era un terápsido con extremidadessemidesparramadas, semejante a un lobo quemedia aproximadamente 1,20 metros delongitud. Los terápsidos comenzaron a

menguar cuando los ágiles tecodontes sedesplazaron hacia su territorio
El maxilar alargado del Morganucodon, unode los primeros mamíferos. El maxilar, de15 mm de longitud, procede de las fisuras definales del triásico de Gales.

El Morganucodon, que quedó abandonadoen su isla galesa en el mar de Tethis deltriásico, probablemente se alimentaba decarroña o comía escarabajos.

El dinosaurio-avestruz Struthiomimus, aligual que tantos celurosaurios gráciles y decuello largo, ha sido encontradofrecuentemente en una posturacontorsionada. Esto se debe al estiramientode los ligamentos del cuello después de lamuerte.

La prueba más persuasiva de que los brontosaurios se metían en el agua son estashuellas de las patas delanteras de un

brontosaurio que sólo pudieron haber sidohechas estando el animal flotando en el agua.
El impacto de los brontosaurios sobre la ecologíaterrestre mesozoica debió de ser enorme. Las manadas de lasdiferentes especies saurópodassimpátricas debió de despejar los espesos bosques y evitar quela maleza se volviera densa, talcomo es el caso de los elefantesen África. Los dinosauriosherbívoros más pequeños,adaptados a los bosques abiertosy a las llanuras, debieron dedepender de los saurópodos que

impedían que estos hábitatsfueran cubiertos por una espesa jungla[39].
La referencia a las manadas esinteresante y nos lleva a otro aspecto delos saurópodos que nada tiene que ver con los reptiles. Hitchcock, tal comorecordarán los lectores, hizoobservaciones sobre las numerosashuellas paralelas de sus «aves» delvalle Connecticut, y las utilizó como prueba de la presencia de bandadas; sus«aves» eran, naturalmente, dinosaurios.Las vidas de los dinosaurios probablemente tenían una organizació

mucho más compleja de lo quegeneralmente se piensa. A diferencia delos lagartos solitarios, muchosdinosaurios eran gregarios, viajando emanadas como los elefantes y antílopes,desplazándose continuamente en buscade pastos frescos para satisfacer svoraz apetito. Bird fue el primero edarse cuenta de esto como resultado desus estudios sobre las huellas dedinosaurios. En su yacimiento deBandera County, en Texas, descubrió una«guarida» de dinosaurios como en u principio la llamó. Al inspeccionarlamás detenidamente, se hizo evidente quetodos los brontosaurios, 23 individuosen total, habían pasado en la misma

dirección y que, tal como sospechó,habían sido conducidos hacia una metacomún. Uno o dos carnívoros solitarioshabían pasado antes pero la manadaclaramente había atravesado el lugar como una unidad. En otras partesencontró las huellas de un gradinosaurio que se había desplazado perezosamente, a veces arrastrando u pie antes de levantarlo, seguido de otrosquince brontosaurios que paseabadespacio, «como vacas que vuelven a svereda[40]». Si los saurópodos eracriaturas de manada, la incapacidad deBird de hallar surcos de colas cerca delas huellas fósiles tiene una explicaciósencilla. Estas bestias seguramente no se

hubiesen atrevido a arrastrar por elsuelo sus largas colas de 9 metros delargo, por miedo a que las pisasen losmiembros de la manada que las seguíade cerca; en lugar de ello, las colasdebían de ser mantenidas rígidas en elaire para que no pudieran lesionarse.
La teoría de que los dinosaurios eraanimales de manada ha sido apoyadarecientemente por un estudio, realizado por John Ostrom, sobre las huellasencontradas en Holyoke, eMassachusetts. Este lugar tiene una edadgeológica más antigua y todas lashuellas se componen de pies con tresdedos. Éstas pueden ser clasificadas etres grupos por su tamaño, siendo las

más grandes las más abundantes (es posible que algunas de las huellas más pequeñas fueran hechas por animales
óvenes de la misma especie). 134huellas individuales forman 28sucesiones de huellas, y 23 de estassucesiones de huellas pertenecen a las bestias de pie grande. 20 de estasúltimas sucesiones de huellas estáorientadas en una misma dirección,hacia el Oeste, mientras que las huellasmás pequeñas parecen estar esparcidasen todas las direcciones.
Los dinosaurios no podían haber estado ramoneando, ya que se hubiese producido huellas erráticas al ir losanimales de un lado a otro. Parece que

estuvieron viajando como grupo quecomparte un objetivo común. Las huellasque se desvían indican que no existía barrera física para los movimientos eotras direcciones. Por lo tanto, el grupono estaba siendo «canalizado», por ejemplo, por tener que pasar por encimade un banco de arena situado entre doslagos adyacentes. La probabilidad deque estas sucesiones de huellasestuvieran orientadas en una mismadirección, si los individuos no eramiembros de una manada, es fenomenal.¡Ostrom calculó que las probabilidadeseran de 2x1021: 1 en contra! Por lo tanto, parece sensato concluir que estosdinosaurios no eran individuos

solitarios sino miembros de una manadacon una meta en perspectiva[41].
Uno de los problemas másfascinantes es el de la jerarquía social.Si los dinosaurios tenían una capacidadmental tan limitada, ¿cómo podríahaber participado en una manadaestructurada? ¿Había un líder y unaerarquía subordinada de machos y
hembras; un harén de hembras perteneciente a un macho dominante; yuna guardería para los jóvenes? ¿Oquizás una organización de este tipoestaba fuera del alcance de la capacidadde los dinosaurios? Bird se imaginaba,que junto con las manadas, viajaba brontosaurios jóvenes. Bakker incluso

propuso una estructura interna para elgrupo, quizá con los animales jóvenes protegidos por un grupo de adultos. Cotoda seguridad, los animales jóvenesdebían de ser muy vulnerables. Los brontosaurios adultos dominaban a todoslos enemigos contemporáneos y, al igualque los elefantes, probablemente sería bastante inmunes a los ataques aunquelos dinosaurios jóvenes no compartíaesa ventaja.
Los restos de saurópodos adultos seencuentran abundantemente y, siembargo, los fósiles de animalesóvenes son muy escasos. Por analogía
con los reptiles, se ha sugerido queexistía una mortalidad explosiva en los

primeros años de vida, pudiendo sólounos pocos individuos alcanzar la edadadulta[42]. La probabilidad desupervivencia de las tortugas jóvenes, por ejemplo, es reducida, y a fin degarantizar que exista un númerosuficiente de animales que sobrevivan,deben ponerse gran cantidad de huevos.Quizá los jóvenes saurópodos eravíctimas fáciles de los alosaurios queacechaban a las manadas. Sólo cuandolos supervivientes alcanzaban un umbralcrítico de tamaño se volvían inmunes alos ataques y su futuro estaba asegurado.
Si los saurópodos ponían nidadas dehuevos, ello hace surgir algunos problemas logísticos interesantes.

Existen limitaciones físicas respecto altamaño que puede alcanzar un huevo. Amedida que el volumen de líquidodentro del huevo aumenta, la cáscara sedebe volver más gruesa a fin de poder resistir la presión, y llegaría un punto eel que la cáscara sería demasiado gruesae impediría a los jóvenes salir delhuevo. Por lo tanto, los saurópodosgigantes podían poner huevos que noeran más grandes que aquellos de losdinosaurios mucho más pequeños. Estocrea dos problemas. En primer lugar, silos huevos eran colocados en la arena osuelo cálido hasta la eclosión de lascrías, ¿cómo los jóvenes dinosaurios podían encontrar el camino hacia la

manada que se encontraría a muchassemanas de distancia en el momento dela salida del huevo? En segundo lugar,¿cómo podía un diminuto dinosauriorecién salido del huevo, de tamaño nosuperior a un lagarto grande, evitar ser pisoteado por los adultos que podíaalcanzar alturas similares a edificios detres pisos? Por lo tanto, uno se preguntasi existían saurópodos que daban a luzcrías vivas mientras estaban emovimiento. Los jóvenes habrían tenidoun mayor tamaño y, rápidamente, sehabrían unido a la manada que lehubiese proporcionado una proteccióinmediata. También significa que en lamanada hubiesen tenido que nacer

menos animales jóvenes debido almayor potencial de supervivencia. Alhaber menos animales jóvenes, ucrecimiento rápido y una expectativa devida de probablemente un siglo, existiríaun relevo lento de la población y una propensión hacia adultosverdaderamente grandes tal como seobserva en los fósiles encontrados.
La cantidad de comida requerida para abastecer a una manada debió deser enorme. Debido a que los brontosaurios podrían haber tenido uíndice metabólico similar al de loselefantes, probablemente necesitarían lamisma cantidad de comida en relaciócon su peso corporal. Un elefante de tres

o cuatro toneladas necesita una toneladade paja al día[43], de forma que udinosaurio de treinta o cuarentatoneladas de peso podría haber consumido media tonelada devegetación cada día. Suponiendo quecada miembro del grupo consumía estacantidad de hojas y frutas, la manada sehabría visto obligada a desplazarseconstantemente a fin de evitar tragarsesu entorno inmediato.
La objeción más seria a esta teoríaes la dificultad de imaginarse a u
iplodocus o Brontosaurus tragar estascantidades inmensas de comida cada díaa través de una cabeza no mayor que unade sus vértebras. (Es interesante

observar que los orificios nasales posteriores le permitían al saurópodo pasar constantemente comida a través desu boca sin tener que interrumpir larespiración, la nariz «acuática», dehecho, hacía las veces de paladar secundario). Al igual que los mamíferosramoneadores de las llanuras que viveactualmente, los brontosaurios probablemente dedicaban muy pocotiempo a dormir durante el día, buscando constantemente forraje. Laausencia de dientes para mascar noconstituye un problema tan difícil deexplicar. El Diplodocus cortaba los brotes, frutos, ramas y hojas con losincisivos con aspecto de estaca situados

en la parte anterior de la boca. Alfaltarle los molares gigantes que elelefante utiliza para moler su comida, el
iplodocus se veía obligado a tragar lavegetación sin masticarla previamente.Las aves tampoco tienen dientes, perologran el mismo efecto utilizando piedras estomacales. En la molleja (unaforma de preestómago), la comida esmolida por estos guijarros por la accióremovedora de las paredes de lamolleja, de forma que lo que pasa alestómago se diferencia poco de lacomida bien masticada de losmamíferos. En un dinosaurio herbívoro —aunque no se trataba de un saurópodo — descubierto por una expedición del

Museo Americano a Mongolia, seencontraron 112 piedras o gastrolitosmuy bien pulidos en la región estomacal.Existen pocas dudas de que éstos erauna parte integral de su mecanismodigestivo[44]. De forma que quizá losgrandes saurópodos también tragabaguijarros para facilitar la digestión. Seha encontrado un espécimen de
arosaurus con siete de estosgastrolitos característicos relacionadoscon él. Estos guijarros tan bien pulidosse encuentran frecuentemente en lascapas rocosas que albergan huesos dedinosaurios. En los lugares donde sonumerosos los esqueletos de dinosaurioscomo, por ejemplo, los lechos del

cretáceo inferior de Montana, existeliteralmente miles de estas piedrasesparcidas por la región, probablementeregurgitadas por los dinosaurios cuandose gastaban al no resultar ya útiles paramoler los alimentos.
Al igual que los elefantes, quedependen mucho de su sentido delolfato, los saurópodos usaban su gracavidad nasal para captar los olorestransportados por el aire. Los elefantessalvajes mueven sus trompas en el aire para captar los olores del follaje de los bosques o de las frutas maduras y el
iplodocus, probablemente, usaba scuello de la misma forma. Para uanimal que presuntamente iba de árbol

en árbol, arrancando los frutos y lashojas, este mecanismo de rastreo debióde ser indispensable, permitiéndoleelegir los lugares para comer. La visióde estas grandes manadas de saurópodosdesplazándose a través de los bosquesurásicos, con animales jóvenes del
tamaño de un elefante adulto protegidos por un anillo de viejos machos, debió deser una de las más imponentes decualquier período en la Historia de laTierra.

VI. UN GRIFORESCATA LAEVOLUCIÓN
A lo largo de la vasta extensión deltiempo geológico, se pueden reconocer inmediatamente los dos saltos prodigiosos hechos por los animalesvertebrados. En ambas ocasiones,estuvo implicada la conquista de umundo nuevo y extraño frente aobstáculos aparentemente insuperables.Uno de estos pasos gigantes ocurriócuando los vertebrados se elevaron por primera vez en los aires, creando así u

medio completamente nuevo decolonización y, en conjunto, una formanueva de existencia que exigíaestructuras locomotoras y sensorialesnuevas y complejas. La forma de cómo y por qué los reptiles terrestresdesarrollaron por primera vez alas co plumas, potentes músculos para volar yextensos centros de coordinación en elcerebro sigue siendo una cuestión muydiscutida. El proceso de elevación en elaire debió de ser gradual, implicandomuchas fases de evolución.
Sin embargo, cada fase en eldesarrollo de un ala debió de ser funcional, habiendo sido escogido entreuna miríada de variaciones aleatorias

que tuvieran alguna utilidad. Laselección natural, después de todo,funciona adoptando todas aquellasestructuras que proporcionen a s propietario una ventaja positiva sobreaquellos que no las posean. ¿Pero cómo pudo haber un desarrollo por etapas elas alas? ¿Seguro que una cosa así esdisparatada? Sólo el producto acabadoes funcional —al menos esto es lo que pensaron algunos de los contemporáneosde Darwin hace un siglo. St. GeorgeMivart pensaba que las modificacionesconstantes y progresivas que finalmentedaban lugar a una estructuracompletamente integrada como un ala,no se ajustaba al modelo propuesto por

Darwin, la Selección Natural. En lugar de ello, Mivart se vio obligado aconsiderar que las alas y todos susatavíos armoniosos surgieron derepente, como si existiera algúmecanismo interno de control[1].
Sin embargo, naturalmente las alasde las aves no pudieron haber surgidode modo espontáneo, mediante elmilagroso salto imaginado por Mivart: por consiguiente, la evolución de lasalas continúa siendo un enigma. El problema es que, para que un ave puedavolar, antes necesita tener plumas,músculos para volar y las adaptacionesesqueléticas asociadas como elesternón. Pero si es necesario que antes

de volar disponga de estascaracterísticas, ¿por qué las desarrolló?Los reptiles no tenían la capacidad de previsión para darse cuenta de que ibaa necesitar plumas si querían volar. Laexplicación del misterio es que cada unode los componentes que, en conjunto,llegarían a volar originariamentedebieron de haber sido desarrollados por algún motivo que no fuera el de permanecer en el aire. Sólo cuandotodas las estructuras indispensablesestuvieron presentes, pudieron ser conectadas para una nueva función.Resultará más fácil entender esto si primero analizamos otro prodigiososalto hecho por los vertebrados: la

colonización de la tierra por los peces.En el período devónico, hace
aproximadamente 400 millones de años,los vertebrados más avanzados era peces revestidos de una coraza ósea. Etierra fírme se estaban rápidamenteestableciendo los primeros insectos y plantas arcaicas. Allí estaba el nuevomundo a explotar. A pesar de que los peces no abandonaron el agua de formavoluntaria —sólo lo hicieron bajo unagran presión— finalmente salieron de él, es a estos pioneros a los que toda la
subsiguiente vida vertebrada terrestredebe su existencia. ¿Cuáles eran losrequisitos previos para la vida en tierra?En lugar de agallas, los peces que se

aventuraran a tierra fírme necesitaría pulmones para respirar el aireatmosférico. Existían dos tipos de pecesen el período devónico que ya poseía pulmones para respirar aire: el pez pulmonado (con descendientes bastanteespecializados que actualmente viven eAustralia, África del Sur y América delSur) y el pez de aleta carnosa (cuyoúnico descendiente, el famoso celacanto —fue pescado al arrastre en el lechooceánico en la costa de East London,África del Sur en 1938, 70 millones deaños después de su supuesta extinció —, ha convertido su pulmón en unavejiga llena de grasa). Aunque hoy edía raros, éstos eran los peces más

comunes en el devónico. Probablementevivían en los lagos pantanosos que, eocasiones, se volvían sucios. Debido aque el agua estancada no tenía suficienteoxígeno, el tragar aire se convirtió euna necesidad para que el pez pudieraseguir vivo. De esta forma las vejigasque, originalmente probablemente eraórganos hidrostáticos llenos de gas, quecapacitaban al pez para seguir flotando,actuaban ahora como pulmones,ayudando a las agallas en caso deescasez de oxígeno. Sin embargo, para poder tragar aire los peces necesitabaalejarse del fondo del agua y es en este punto donde la aleta carnosa encontró s plena justificación. La aleta pectoral de

estos peces estaba compuesta no sólo deun lóbulo carnoso, sino también de uesqueleto óseo interno (que puede ser equiparado, hueso por hueso en sregión proximal con el de los anfibios yde hecho con los miembros de todos losvertebrados que surgiero posteriormente). Cuando finalmente lascharcas se fueron secando lentamente,haciéndose inhabitables para otros peces, los peces con pulmón pudieroseguir con vida tragando aire. Cuando lacharca se secó por completo, y el pezquedó varado fuera del agua, la aletacarnosa podía seguir respirandolevantando la garganta-bomba del suelo para coger aire. (Mientras tanto, el pez

pulmonado tenía que hacer madriguerasen los bancos de arena para pasar elverano en estado de entorpecimientohasta la época de las lluvias: el pez pulmonado había llegado a un callejósin salida de la evolución). Fuera delagua, los pulmones serían inútiles y el pez se ahogaría al no tener un apoyo para levantar el tórax del suelo. Alemplear el movimiento natural de popa a proa de la aleta, el apoyo podría ser utilizado para que el recién instaladoanfibio se desplazara para buscar másagua.
De esta forma, los peces de aletascarnosas del devónico ya estabaadaptados para sobrevivir fuera del

agua, mientras aún estaban viviendo eél. Ya habían desarrollado el equiponecesario. Si ahora volvemos al problema de los vertebrados voladores,hemos de explicar por qué las plumas,alas, etc., fueron desarrollados antes deque estas protoaves fueran capaces devolar. ¿Para qué habían servido al principio? Las aves no pueden lanzarsede los árboles si no tienen las alas co plumas —por lo tanto, debieron dedesarrollar las alas antes de subirse alos árboles. ¿Pero cuál era la funció primitiva de estas estructuras bonitas«preadaptadas»?
A fin de resolver el problema,debemos volver al ave más antigua que

se conoce —y quizás el fósil másfamoso (o, al principio, infame) detodos los tiempos— para observar dequé estaba provisto este animal para poder volar. Antes de 1860 no se habíaencontrado plumas procedentes de rocasmás antiguas que el período terciario y, por lo tanto, como dice un proverbio, lasaves se conocen por sus plumas, se pensaba que las aves no existieron hastael mesozoico. Se argüía que lasupremacía aérea pertenecía a los pterodáctilos, y sólo cuando éstos seextinguieron, al final del períodocretáceo, las aves tuvieron la ocasión dedesarrollarse (algunos evolucionistasincluso se aventuraron a decir que los

pterodáctilos evolucionaron hacia lasaves). Entonces, en 1860, justo un añodespués de la publicación de El origen
de las especies de Charles Darwin,Hermann von Meyer encontró una plumaen los depósitos de caliza litográfica deSolnhofen, en Baviera, que conteníafósiles de pterodáctilos. Describió la pluma con cautela, casi sin atreverse a pensar que ésta perteneciera a un ave — aún siendo su aspecto de ave. Le dio elnombre de Archaeopteryx lithographica
por el «plumífero arcaico de la calizalitográfíca[2]». ¿Realmente esta plumasolitaria podía proceder de un ave del período jurásico tardío, incluso antes deque florecieran los iguanodontes? En u

principio, Von Meyer era escépticorespecto a su autenticidad; de ningúmodo habría sido el primer engaño paleontológico. Sin embargo, el estudiominucioso de la pluma descartó que setratara de una trampa. Era auténtica cotoda certeza.
Al año siguiente, 1861, se produjo eldescubrimiento paleontológico del siglo, una de las mayores ayudas que podría
haber recibido el recién surgidodarwinismo. Se había encontrado uesqueleto sin cabeza, pero casicompleto, de un híbrido de reptil y ave a18 metros bajo tierra en la cantera deOttmann en Solnhofen, Baviera; era uave porque tenía plumas, aunque seguía

siendo un reptil por sus dedos en formade garra y su larga cola ósea. Lasensación que provocó entre lostrabajadores hizo que llamara laatención del médico del distrito, Dr.Karl Haberlein, que rápidamente se diocuenta de su valor científico. El Dr.Haberlein, que había obtenido el fósilde los trabajadores de la cantera como pago de sus servicios médicos, lo puso ala venta. Cualquier museo lo podríaadquirir para sus colecciones: es decir,cualquier museo que pudiese pagar elinaudito precio de setecientas librasesterlinas. El médico invitó a los futuroscompradores para que vieran el reptil-ave, pero no les permitía tomar notas o

hacer dibujos y dejó que presentaran susofertas. Fueron a verle coleccionistaseminentes de todo el mundo para echar un vistazo a los restos fósiles. Haberleirecibió de América a Louis Agassiz, elconfirmado antidarwinista y fundador del Museo de Zoología Comparada de laUniversidad de Harvard. LordEnniskillen, un ávido coleccionista defósiles, cuyas adquisiciones finalmente pasaron a la propiedad del MuseoBritánico, y el duque de Buckingham,constituyeron el contingente británicoque se dirigió hacia Baviera. Siembargo, Haberlein estuvo un año siquerer comprometerse a nada,intentando elevar el precio. Durante este

tiempo se creó tal misterio alrededor delfósil, que los creacionistas empezaron aacusar a sus oponentes de haber fabricado el «eslabón perdido» paraconfirmar su debilitada teoría. AndreasWagner, un antidarwinista de laUniversidad de Munich, que estuvoanalizando el fósil a partir de losrumores y dibujos de segunda mano(siendo éstos producto de la memoria),hizo lo que pudo para robarle al
rchaepteryx cualquier rasgo que pudiera hacerlo parecer un eslabón y, por lo tanto, munición para losdarwinistas. Un ave con una cola largade reptil y dedos en forma de garras,sería idóneo para la causa darwinista,

así que Wagner lo degradó a un reptilque había desarrollado plumasindependientemente de las aves como«adornos peculiares». Wagner lo llamóGriphosaurus, el «lagarto enigmático»,o «lagarto grifo». «La primeracontemplación del Griphosaurus podríacausamos ciertamente la impresión deque tenemos ante nosotros una criaturaintermedia —advirtió Wagner—.Probablemente, Darwin y sus partidarios emplearán el nuevodescubrimiento como un éxito,excesivamente bien recibido, para laustificación de sus extrañas ideas sobre
la transformación de los animales». Eeste caso, preguntó Wagner, ¿dónde

estaban las fases intermedias de esteeslabón y el ave o reptil final? Al haber perdido el primer combate contra elenemigo, los creacionistas exigían ahorala existencia de criaturas intermedias yasí, probablemente, ad infinitum.Convencido de que había propinado ugolpe demoledor a la extraña doctrinaevolucionista, Wagner dejó a susadversarios con sus «sueñosfantásticos[3]».
La descripción que Wagner hizo delgrifo llegó a oídos de Richard Owen,ansioso como siempre de ampliar lacolección de fósiles del MuseoBritánico (que en aquella época estabaalbergada en Bloomsbury; Owen más

tarde sería el encargado de trasladarla aun edificio independiente en SoutKensington). En 1862, Owen envió aGeorge Waterhouse, conservador delDepartamento Geológico, a Baviera co500 libras esterlinas, la suma máximaque estaban dispuestos a pagar losadministradores del museo. «El viejomédico alemán es obstinado respecto al precio —escribió Owen en su Diario—
Mr. Waterhouse ha vuelto con lasmanos vacías. No deberíamos dejar escapar este fósil[4]». No lo dejaroescapar. Los administradores del museofinalmente pagaron la suma completa,que se llevó una cantidad considerabledel presupuesto previsto de dos años.

o fue una adquisición mala para elfósil más sensacional que jamás hayasido descubierto. Cuando, finalmente,los museos alemanes capitularon, lagente se enteró de que el ave habíavolado a Londres[5].
Y realmente fue sensacional emanos de los darwinistas ingleses.Owen, el protagonista de Huxley en laarena evolucionista, posiblemente lohabía descrito formalmente, pero fueThomas Henry Huxley, el campeón deDarwin en la batalla y el hombre queceremoniosamente destronó a Owen, elque advirtió su valor real. CuandoHuxley se levantó para dar una de sus populares conferencias, se produjo u

silencio de expectación en la audienciade la Institución real. ¿Cómo es, preguntó, que si un tipo de animalevoluciona hacia otro, por ejemplo, ureptil hacia un ave, existen los grandessaltos insuperables que hoy en día losseparan? «Nosotros, los que creemos ela evolución, replicamos diciendo queesos huecos fueron antaño inexistentes,que las formas que conectaban unasespecies con otras existieron en épocas previas de la historia de la Tierra, peroque ahora están extinguidas». Unasonrisa maliciosa rompió la duraexpresión de Huxley. «Resulta natural, pues, que ahora se nos pida que presentemos esas formas de vida

extintas». Y Huxley, tan mago comosiempre, procedió a sacar al
rchaeopteryx de su sombreroVictoriano[6].
Al no estar ya a la defensiva, losevolucionistas pudieron salvar elobstáculo creado por la excusa de las«imperfecciones del registro geológico»que Darwin había dado para la falta deeslabones de conexión. Finalmente, sehabía encontrado una criatura que era uverdadero intermedio entre las aves ylos reptiles —un reptil con plumas o uave con dedos en forma de garra y colaósea; independientemente de cómo fueraanalizado el fósil, no se podía negar quese trataba de una criatura de transición.

De hecho, era exactamente el tipo decriatura intermedia que Huxley habíadescrito antes de que fuera desenterrada.Darwin estaba encantado.
Cuando en Solnhofen fue descubiertoel siguiente Archaeopteryx —en lacantera de Dorr en 1877—, se confirmóla naturaleza de grifo de la criatura por la innegable presencia de dientes en susmaxilares (en realidad, ya se habíasugerido que también se podíaobservar en el espécimen de Londres).Una vez más fue la familia Haberlein,esta vez su hijo, la que entró en posesiódel vestigio científico. El precio puestoa su cabeza (este espécimen estaba provisto de una) era de 1800 libras

esterlinas. Se instó al gobierno alemán aque no dejara que el valioso fósilsaliera del país, al igual que s predecesor, e incluso éste fue ofrecidoal Káiser Guillermo I. La falta de interésde éste provocó un torrente deimproperios de Karl Vogt, profesor dezoología en la Universidad de Ginebra.«Su Majestad no quiere tomar parte eesta discusión. ¡Ay, ojalá se tratara, elugar de un ave, de un cañón o un fusil petrificado[7]!». Vogt, un socialista de larevolución de 1848, asumió la direccióde la lucha darwinista. Cuando, en 1879,habló en el congreso de naturalistassuizos intentó conseguir el entusiasmo (ydinero) para traer el trofeo darwinista a

Ginebra. Sin embargo, el discursodirigido a los naturalistas para recaudar fondos degeneró, como resultado de laferoz retórica socialista de Vogt, en uataque global al Káiser, el militarismo eimperialismo alemán, resultando elKáiser muy mal parado. El fósil le fueofrecido al propio Vogt a un precioreducido, pero éste tampoco fue capazde reunir la suma requerida. Siembargo, sus improperios y reprochesde barbarismo cultural impulsaron a uindustrial alemán a comprar la plancha por 1000 libras esterlinas, y luegovenderla a la Universidad de Berlín por el mismo precio.
A pesar de que el debate sobre la

evolución ya pertenece a la historia y alos Estados del sur, el Archaeopteryx
sigue siendo el centro de lacontroversia. El problema ya no es si sedesarrolló a partir de los reptiles, laatención se centra ahora en qué reptilesfueron sus antepasados.
Las similitudes existentes entre elrchaeopteryx y los pequeños
dinosaurios corredores celurosauriosson sorprendentes. Sin sus plumas, el
rchaeopteryx tiene un esqueleto casiidéntico al de estos dinosaurios bípedos.Tal como observó Heilmann en 1920:«Huesos excesivamente largos cometatarsianos muy delgados y un “dedotrasero”, una mano larga y estrecha, una

cola larga y un cuello largo, órbitasgrandes y costillas ventrales —éstas socaracterísticas de las aves queinmediatamente llaman la atención elos celurosaurios[8]». Era imposible queéstas pasaran inadvertidas. Othniel C.Marsh, el prolífico describidor yescritor de la Universidad de Yale, bautizó a uno de sus nuevos hallazgos decelurosaurios del cretáceo con elnombre de Ornithomimus, «imitador delas aves», en reconocimiento a estassimilitudes y, más tarde, Henry FairfielsOsborn llamó a un espécimen similar Struthiomimus, o «imitador de lasavestruces[9]». Pero el hecho de que lasaves en realidad habían descendido de

estos dinosaurios era demasiado difícilde creer. Siempre que se sugería algunaafinidad entre el ave y el dinosaurio,ésta era contestada por un aluvión de proposiciones contrarias. El debate hacontinuado durante un siglo. En ladécada del 1870, Samuel WendellWilliston (que más tarde se convertiríaen uno de los expertos de reptiles fósilesmás eminentes y que en aquella épocaera un recolector de dinosaurios deMarsh) se inclinaba a favor de la teoríade los antepasados dinosaurios, eoposición a otro de los prospectores deMarsh, el profesor Benjamin Mudge deKansas. (La única razón, de hecho, por la cual Marsh fue capaz de mantener el

récord mundial de «descubrimientos» deespecies nuevas de dinosaurios es queél, o más bien el museo de su tíoPeabody, pudo permitirse tener a todo uequipo de recolectores sobre el terreno).
El implacable enemigo de Marsh,Edward Drinker Cope, ya habíaquedado sorprendido por la grasimilitud existente entre el aspecto deestos reptiles fósiles y las aves. Leimpresionó especialmente elCompsognathus, un pequeñocelurosaurio bípedo contemporáneo del
rchaeopteryx, pero no fue capaz dedecidir si este pequeño celurosaurio se parecía más a un pingüino o a uavestruz[10]. Mientras tanto, en Londres,

el Compsognathus había llamado laatención de Huxley que se estabaocupando del mismo problema y ssolución fue comunicada a la SociedadGeológica de Londres (de la cual eraentonces presidente) en 1869. Huxleyiba a suprimir el término general de«dinosaurio» e introducir un nombrenuevo —los ornitoscélidos o «piernasde ave»—, en reconocimiento a losmiembros traseros de los dinosaurios,similares a los de las aves. Losdinosaurios ordinarios seguirían estandoen uno de los grupos subordinados y, eel otro, Huxley incluiría alCompsognathus a fin de destacar sussorprendentes características, parecidas

a las de las aves[11]. Este audaz intentode Huxley de derribar las viejas ideasno duró mucho tiempo. Una de las personalidades de entre la audiencia queestaba escuchando su conferencia era elhombre que, poco después, escribiría elartículo definitivo sobre la clasificacióde los dinosaurios, Harold Govier Seeley. No obstante, este hechodemuestra el reconocimiento queestaban obteniendo en los primeros añoslas cualidades avícolas de losdinosaurios. Y quizá excusa a EdwardHitchcock por persistir en su idea deque las huellas del valle del Connecticut pertenecían a aves gigantes.

n. Aparte del Archaeopteryx, elCompsognathus (arriba) era el dinosauriomás pequeño. ¿Podía ser que tuviera plumas?
El debate prosiguió durante variasdécadas. A pesar de que no se podíaignorar las similitudes, parece que lamayoría de los expertos pensaban quelas hipótesis de la ascendencia

dinosauria para las aves era herética, para no decir más. Pero, en este caso,tenían la responsabilidad de dar razonesconvincentes para estas similitudes siimplicar algún tipo de relación directa.En palabras de uno de los paleontólogosmás respetados y teóricos de laevolución de este siglo, George GaylordSimpson, «casi todos los parecidos particulares de algunos saurisquianoscon las aves, tanto tiempo observados ytan destacados en la literatura, sodemostrablemente paralelismos yconvergencias. Estas formas dedinosaurios corredores desarrollarounas características en el esqueleto y ealgún género u otro que se asemejan de

forma sorprendente a las de las aves, pero en realidad nunca han alcanzado elnivel de las aves[12]». La convergenciaera el recurso adoptado por losortodoxos para evitar las consecuenciasde la naturaleza, tan extraordinariamente parecida, de los celurosaurios y de lasaves. Éstos protestaron diciendo que lasaves y los dinosaurios tenían uantepasado lejano —probablementetriásico—, común entre los tecodontes primitivos que dieron lugar a todos losarcosaurios, ya fueran pterodáctilos,aves, cocodrilos, dinosaurios, etc. Siembargo, los pequeños celurosaurios, y precursores de las aves, habían tenidoun tipo de vida muy similar. Ambos era

corredores ya que ambos tenían pataslargas; ambos eran depredadores comaxilares llenos de dientes puntiagudos manos aferradoras que podían coger
pequeños mamíferos y lagartos. Al vivir en el mismo nicho ecológico, sufrir lasmismas presiones y comer la mismacomida, la selección natural había provocado que estructuras de aspectosimilar tuvieran las mismas funciones eambos grupos. Se trata de un procesohabitual tal como ya hemos visto. Eldelfín presenta una gran convergenciacon los ictiosauros marinos delmesozoico, ya que ha adoptado el mismoestilo de vida en el mismo tipo de medioambiente: ambos perdieron sus

extremidades y desarrollaron aletas parasurcar el mar y ambos desarrollarocuerpos aerodinámicos con contornoscasi idénticos. De la misma forma que elcráneo del lobo de Tasmania hadesconcertado a muchos estudiantes, que pensaban que se trataba de un lobo placentario, de hecho, a primera vista,son casi indistinguibles. Sin embargo, ellobo placentario está más estrechamenteemparentado con el elefante que con ellobo marsupial de Tasmania. El hechode tener una forma de vida tan similar provocó la convergencia de su aspecto.
Los que se inclinaban a favor de lateoría de los antepasados dinosaurios,fueron silenciados eficazmente con la

publicación del libro altamenteinfluyente de Gerhard Heilmann, The
Origin of Birds, que apareció amediados de la década de 1920.(Después de haber tenido grandesdificultades, como se quejóamargamente Heilmann, ya que ningunode los editores ingleses comprendía scontenido o mostraba el más mínimointerés por la empresa; ¡a menos,naturalmente, que el autor sufragaratodos los costes[13]! El libro ha sido uclásico desde entonces. Heilman parecía estar a punto de aceptar a losdinosaurios como antepasados de lasaves; confeccionó una lista innumerablede similitudes, y observó que «los

sorprendentes puntos de similitud entrelos celurosaurios y las aves estabarelacionados con casi todas las partesdel esqueleto. A partir de estosresultados podría parecer unaconclusión obvia buscar entre loscelurosaurios a los antepasados de lasaves». (Compárese esta afirmación cola opinión expresada por Simpsoarriba: demuestra cómo los expertos pueden examinar el mismo material partiendo de puntos de vista bastantedistintos y, en muchos casos, segúdiferentes inclinaciones teóricas). Peroa continuación, Heilmann se detiene bruscamente antes de dar el siguiente paso herético. «Sin embargo, éste sería

demasiado precipitado ya que, laausencia de clavículas (huesos delcuello) sería en sí misma pruebasuficiente de que estos saurios no pueden ser los antepasados de las aves».
inguno de los dinosaurioscelurosaurios presentaba clavículas; siembargo, el Archaeopteryx sí los presentaba. Debido a que la evolucióno puede dar marcha atrás (dictamen quese conoce como ley de Dollo), y a quecuando se pierde una estructura, ésta no puede volver a desarrollarse, esimposible que un grupo que haya perdido la clavícula evolucione hacia ugrupo que la posea. Y esto acabó con la polémica. Hubo uno o dos intransigentes

que se pusieron a favor de losdinosaurios (especialmente Percy Loween la década de 1920, y después, que pensó que el Archaeopteryx era «udinosaurio volador» sin ningunacaracterística avícola. Debido alreducido número de vértebrascervicales pensó que estaba demasiadoespecializado como para haber dado paso a las aves modernas y, por lo tanto,consideró que se trataba de uno de loscallejones sin salida en la evolución dela Naturaleza). Pero estas declaracioneseran aisladas. La búsqueda de losantepasados se trasladó al períodotriásico, cuando aún existían criaturas primitivas que presentaban clavículas y

se escogió a los seudosuquianos deconstitución más fuerte como candidatosmás probables. A pesar de que las aves primitivas y los dinosaurios corredorestenían un aspecto muy similar, de hechose habían originado de formaindependiente a partir de losseudosuquianos menos refinados. Por lotanto, los delicados celurosaurios, ta parecidos en casi todos los aspectos al
rchaeopteryx contemporáneo suyo,fueron descartados debido a la ausenciade clavículas. Cualquier relación era«falsa», según las palabras de Gavin deBeer [14], y creer en ésta era consideradocomo una herejía por los ortodoxos,estado de opinión que duró hasta 1973.


o. Heilmann se imaginó a los antepasados delas aves como reptiles que se deslizaban por los árboles.
Se pensaba que el verdaderoantepasado seudosuquiano de las aveshabía sido el Ornithosuchus bípedo noespecializado. Éste se parecía mucho alos dinosaurios más primitivos, pero snombre —que literalmente significa«ave cocodrilo»— ilustra bien elsupuesto origen común de las aves, losdinosaurios y los cocodrilos. A primeravista, puede parecer sorprendente que el pariente vivo más cercano de las avessea un cocodrilo acuático, acorazado yde aspecto arcaico. Éstos, actualmente,

parecen tener poco en común porquefueron difundiéndose hacia nichosecológicos muy diferentes, querequerían modificaciones muy diversas.De hecho, los cráneos de las aves y delos cocodrilos comparten muchascaracterísticas comunes, lo cualdemuestra su ascendencia común. Siembargo, no existe consenso sobre elgrado de parentesco que existe entre lasaves y los cocodrilos. Recientemente,Alick Walker, de la Universidad de
ewcastle, ha tomado partido a favor dela existencia de lazos mucho másestrechos de lo que hasta el momento se pensaba, como resultado de sus estudiossobre el Sphenosuchus que ha sido

interpretado de diversos modos, comococodrilo muy primitivo del triásicosuperior, o como seudosuquiano en scamino evolutivo hacia el estado decocodrilo. Nadie está seguro de quélado ponerse. Walker encontró quemuchos de los huesos en el cráneo delSphenosuchus funcionaban como los deun cráneo de ave. Incluso parece que elcocodrilo primitivo tenía unas glándulasexcretoras de sal en las órbitas y que, probablemente, era capaz de beber aguasalada, eliminando el exceso de sal dela misma forma como lo hacen las avesmarinas hoy en día. ¡El oído interno delSphenosuchus, dijo Walker, era más parecido al de una perdiz que al de u

cocodrilo moderno! El valor exacto dealgunos de los datos presentados por Walker, que relacionaban a las avesmodernas con los cocodrilos primitivos,es objeto de discrepancias al igual quelo son sus conclusiones de que loscocodrilos evolucionaron a partir de uantepasado avícola. «Por lo tanto, parece lógico concluir seriamente — dijo Walker— que es posible que loscocodrilos, en su conjunto, hayadescendido, quizá en “oleadas”sucesivas, de un tronco desconocido dereptiles del triásico medio o superior que, por comodidad, podrían ser llamados “protoaves[15]”». Debido aeste supuestamente estrecho parentesco

(la palabra clave es «estrecho» ya quenadie niega su afinidad), Walker inclusoestá dispuesto a aceptar la posibilidadde que en otros tiempos los cocodrilosfueron habitantes de los árboles.
El informe de Walker, publicado eature, era un desafío que no podía ser
ignorado. Éste fue contestadoinmediatamente por John Ostrom, cuyacarta de rechazo apareció sólo pocosmeses más tarde en Nature, en 1973.Ostrom estaba en una buena posición yaque se había pasado dos añosestudiando todos los especímenesconocidos de Archaeopteryx (entoncesexistían cuatro, pero más tarde seencontró otro), intentando desenmarañar

el origen del vuelo de las aves. Elrchaeopteryx jurásico, evidentemente
estaba más relacionado con el origen delas aves que la perdiz y, tal comodemostraría Ostrom de formaconvincente, que los cocodrilosesfenosuquios. No era necesarioremontar la ascendencia de las aveshasta los lejanos cocodrilos, otecodontes del triásico, debido a que losantepasados ya eran evidentes en elurásico. «La anatomía del esqueleto delrchaeopteryx —dijo Ostrom de forma
desafiante— es casi enteramente la deun dinosaurio celurosaurio, nada detecodonto, de cocodrilo o de ave[16]».La teoría había dado una vuelta

completa, pero ahora la prueba, talcomo la presentaba Ostrom, sedecantaba de forma arrolladora a favor del origen dinosáurico de las aves yadefendido por Huxley. Los argumentosde Ostrom son tan poderosos que resultadifícil no llamar dinosaurio al propio
rchaeopteryx. Ostrom hizo unarelación de los sutiles rasgos querelacionaban a los celurosaurios pequeños, bípedos, con manosaferradoras. Sin embargo, el motivo delcambio en la opinión general paleontológica se debe más bien a losdescubrimientos nuevos einterpretaciones nuevas. Los recienteshallazgos de dinosaurios, especialmente

los hechos en el desierto de Gobi,recientemente explorado de nuevo,demostraron que existían muchos máscaracteres compartidos de lo que se pensaba. En concreto, fue eliminadacomo objeción la grave ausencia declavícula que había provocado que, eel último momento, Heilmandesestimara el origen dinosáurico de lasaves; los dinosaurios —incluyendo elVelociraptor mongol encontrado por unareciente expedición polaco-mongol aldesierto de Gobi—, han sidodesenterrados con clavícula.
Sin embargo, el ataque de Ostrofue doble. No solamente demostró lascaracterísticas dinosáuricas del ave más

primitiva, sino que también acercó alrchaeopteryx más a los dinosaurios,
eliminando algunas de suscaracterísticas avícolas. El cinturó pélvico típico de las aves, presenta u proceso púbico orientado hacia atrásmientras que, en los dinosaurios, ésteestá dirigido hacia abajo. Esto socaracteres diagnósticos; el cinturó pélvico siempre ha tenido una graimportancia en la clasificación de losdinosaurios, desde que Seeley advirtió, por primera vez, las diferenciasexistentes entre los principales tipos dedinosaurios, en 1887. Siempre se había pensado que el Archaeopteryx de Berlí presentaba esta orientación hacia atrás

del pubis típica de las aves, hasta queOstrom halló la prueba de que, probablemente, éste se había roto en elfósil y que, por lo tanto, podía haber estado orientado hacia abajo en vida,como en los otros dinosaurioscarnívoros[17]. Después de haber puestoen duda esta cuestión, sólo quedaba lafúrcula o «espoleta», formada por lafusión de las clavículas, que indicabaque el Archaeopteryx era un ave — aparte de una posición bastante obvia.«De hecho —afirmó Ostrom—, si no sehubiesen conservado las impresiones delas plumas, todos los especímenes de
rchaeopteryx hubiesen sidoidentificados como dinosaurios

celurosaurios. La única conclusiórazonable es que el Archaeopteryx
debió de descender de un terópodo de principios o mediados del jurásico».
El origen más probable detantos rasgos característicos delos celurosaurios en el
Archaeopteryx es la herenciadirecta de un antepasadocelurosaurio pequeño. Laimportancia adicional deestafilogenia es que los«dinosaurios» no se extinguieronsin dejar descendientes y sugiereque las plumas, como aislantestérmicos, podrían haber sido la

razón principal del éxito de losdescendientes de losdinosaurios.
Las similitudes eran demasiadocompletas, detalladas y numerosas como para haber sido el resultado de laconvergencia. La convergencia puedeservir como explicación mientras nosobrepase el límite de credulidad.Incluso el cráneo del lobo de Tasmania presenta características diagnósticasmuy importantes, que inmediatamente lorevelan como marsupial y, por lo tanto,no tiene ningún parentesco con nuestrolobo del hemisferio Norte. El
rchaeopteryx era un celurosaurio

pequeño que se diferenciaba de suscontemporáneos por la posesión de plumas. No debería haber ya ningunanecesidad de recurrir a la convergencia;la prueba contundente exige ahora u parentesco sanguíneo directo.
Los simpatizantes se alinearon allado de Ostrom, valiéndose de susresultados para apoyar sus propiostrabajos. De repente, parecía que todaslas piezas encajaban: las aves probablemente eran de sangre caliente porque descendían de dinosaurios desangre caliente. La endotermia no sehabía desarrollado de formaindependiente en estos dos grupos, sinosólo una vez en el tronco común de

antepasados dinosaurios. Comocorolario, naturalmente, la certeza deque las aves son de sangre calienterefuerza la teoría de que los dinosauriostambién lo fueran. Esta solución nosolamente es más agradableestéticamente, sino que se ajusta a laexigencia científica de sencillez; no sedeberían invocar dos orígenes para laendotermia cuando uno solo ya satisfacelas evidencias. Las aves heredaron scondición fisiológica de los dinosaurios.
Las implicaciones de estaclasificación animal son verdaderamenterevolucionarias. Sugieren que debemoscambiar radicalmente nuestra forma dever las aves y los dinosaurios.

Si los dinosaurios tuvieron un origeúnico u orígenes múltiples, a partir deun grupo restringido, y si admitimos queel grupo ancestral sufrió una gratransformación pasando de la ectotermiaa la endotermia —una innovación clavede tal importancia que desencadenó laaparentemente repentina explosión dedinosaurios en el mundo triásico—,nuestra clasificación debería reflejar laconsecución de este nuevo nivel dedesarrollo. Es muy difícil abogar por laseparación de los mamíferos de losantepasados reptiles por lo que serefiere a su clasificación sin, al mismotiempo, abogar por la separación de losdinosaurios de sus ancestros reptiles: e

ambos casos, están en litigio las mismascuestiones. Al igual que los mamíferos,los dinosaurios también habían cortadocon sus antepasados y habíaevolucionado hacia un apogeo nuncaalcanzado hasta entonces. Ciertamente,esto también justifica que sean elevadosa la categoría de clase por derecho propio, condición que los equipararíacon los mamíferos, reptiles, anfibios y peces. Todos éstos representan pasosenormes en la historia de la vida: los peces por salir del agua; los anfibios por poner huevos con cáscara fuera delagua; los reptiles por elevar por vez primera su temperatura interna hasta univel alto estable sin la ayuda del sol.

Cada uno de éstos anuncian un salto a unuevo nivel de organización.
¿Pero, qué pasa con las aves? ¿Por qué no figuran en esta lista? Esto es unasorpresa quizás aún mayor que lacreación de la clase dinosaurios. Estehecho fue apreciado, por primera vez esu totalidad, por Robert Bakker y Peter Galton, cuya modesta carta de cuatro páginas a Nature, el 8 de marzo de1974, llevaba el título de «Monofilia delos dinosaurios y una nueva clase devertebrados». El aspecto quizá pueda parecer modesto, pero lo que contiene, probablemente, es una de las propuestasmás revolucionarias —y controvertidas — en el pensamiento paleontológico de

este siglo. A continuación citaré elúltimo párrafo de este artículo clave quese encontraba entre las cartas publicadasen Nature.
La endotermia, y unmetabolismo aeróbico altamenteefectivo, son una justificaciónsuficiente para separar las avesen una clase distinta de los otrostetrápodos saurópsidos vivos.Sin embargo, la endotermia y elmetabolismo altamente efectivo probablemente ya existían en losantepasados dinosaurios de lasaves, y son las característicasclave que diferencian a los

dinosaurios de los cocodrilos yde otros arcosauriosextinguidos… La radiación delas aves es una utilización aéreade la fisiología básica deldinosaurio y de su estructura,como la radiación delmurciélago es una utilizaciónaérea de la fisiología básica, primitiva, de los mamíferos. Losmurciélagos no están separadosen una clase independiente,simplemente, porque vuelan.Pensamos que ni el vuelo ni ladiversidad de especies de lasaves merecen la separación delos dinosaurios como clase. La

adaptación más profundaexperimentada por los amniotasfue la transición de la ectotermiaa la endotermia, que tuvo lugar durante el origen de losmamíferos y de los dinosaurios.Por esta razón, proponemos lacreación de una clase llamadadinosaurios que incluiría lassubclases saurisquianos, aves yornitisquianos.
Los arcosaurios, como loscocodrilos, tecodontes y pterosaurios,tendrían con los dinosaurios un lazo de parentesco similar al que tienen losreptiles semejantes a mamíferos con los

mamíferos. «Esta nueva clasificación — concluyeron— refleja con mayor fidelidad los más importantes pasosevolutivos. Ectotermos y formastransicionales en el camino hacia laendotermia, se conservan en los reptiles los dos grupos endotérmicos altamente
triunfantes, mamíferos y dinosaurios,deben tener categorías de clasesseparadas[18]».
En efecto, proponen que las aves yano existan como clase rival de losmamíferos. Este privilegio correspondeahora a los dinosaurios, siendo las avessus representantes vivos. Las aves sóloson uno de los tres grupos dedinosaurios.

Las profundas implicaciones de estaidea no podían pasar inadvertidas en laPrensa popular británica. «Lasorprendente afirmación de que las avesson dinosaurios —exclamó el The
Sunday Times en un artículo titulado“Los dinosaurios voladores”— no puede ser ignorada por los compañeros paleontólogos[19]».
Bakker y Galton —continuóel The Sunday Times — proponen la revisión drástica dela clasificación zoológica de losanimales, algo tan controvertidocomo cambiar la Sagrada

Escritura. Incluso los que noestán de acuerdo deben tomar enserio esta teoría.
Sin embargo, a pesar de laresistencia intrínsecamenteconservadora al cambio que caracterizaa toda actividad humana, la ciencia —econtraste con la Sagrada Escritura— es
una disciplina en constante evolución y, por lo tanto, es susceptible a ser actualizada de acuerdo con los últimoshallazgos. La ciencia es una búsquedasin límites fijos. Su objetivo es lograr una mayor comprensión del mundo. Hahabido muchos momentos en los que elhombre ha tenido que cambiar

radicalmente su concepto del mundo, yquizá ninguno tan importante comocuando la Tierra dejó de ser el centrodel mismo. Las teorías científicas posiblemente se acerquen cada vez mása la verdad pero nunca la alcanzarán. Silas pruebas han sido interpretadascorrectamente, y la conclusión se realizade forma lógica, con el tiempo laoposición vencerá a la idea. AlaCharig, director del Departamento deReptiles Fósiles y Aves del MuseoBritánico, admite que las aves podríahaber descendido de los dinosaurios yque los dinosaurios podrían haber sidode sangre caliente, pero no acepta lateoría del origen único para todos los

dinosaurios. Los tres puntos, arguyeCharig, deben ser conciliados. Tal comodice Charig, «los paleontólogos nocomenzarán a llamar oficialmente a lasaves dinosaurios hasta que todos estéunánimemente de acuerdo en estos puntos y estamos lejos de ello[20]». Siembargo, el argumento es igualmenteválido si se acepta que la clasedinosaurios es polifilética, es decir, queun mismo grupo ancestral tenga orígenesmúltiples, de forma que el tercer argumento de Charig no esverdaderamente crucial para estacuestión. De hecho, ninguna de lasclases existentes (con la sola excepcióde las aves discutiblemente difuntas) es

considerada monofilética, de modo quedifícilmente esto puede ser un requisito previo para el reconocimiento de unaclase. El punto importante de lacuestión, en relación con losdinosaurios, es que probablemente una omás líneas estrechamente emparentadasalcanzaron un nivel de organizació basado en la endotermia, y que todos lossubsiguientes linajes se originaron a partir de una clase de dinosaurios desangre caliente. En realidad, lasobjeciones de Charig podrían ser refutadas, y nuestro punto de vista ser considerado más justificable si el grupoancestral —los tecodontesseudosuquianos— también fuese

incluido en la clase de los dinosaurios.De hecho, existen razones fundadas paraello. Tal como siempre se haargumentado, las innovaciones clave(postura erguida, metabolismo rápido ysangre caliente), que asociamos con laradiación de los dinosaurios, probablemente se desarrollaron por primera vez en los seudosuquianos primitivos del triásico. No sería lógicoestablecer la clase dinosaurios y luegoincluir a los seudosuquianos en el grupode los reptiles. Para que nuestra nuevaclase sea válida y refleje el incrementodel nivel de organización, deberíaincluir a los seudosuquianos. La clasede los dinosaurios podría acabar

comprendiendo a los ornitisquianos,saurisquianos y seudosuquianos y —siBakker y Galton impusieran su punto devista— las aves.
Si el Archaeopteryx realmenteestaba emparentado con loscelurosaurios —y el estudio de Ostrosugiere que lo estaba muy estrechamente —, y si presentaba tan pocos de losrasgos que asociamos con las aves, pareciéndose a los pequeñosdinosaurios en casi todos los detalles,¿qué ocurría con su supuesta capacidad para volar? Después de todo, era muyimprobable que los celurosaurios

pudieran volar y parece ser que elrchaeopteryx solamente era u
celurosaurio con plumas sin que presentara las modificacionesesqueléticas que uno asocia con lacapacidad de volar. El Archaeopteryx, por ejemplo, presentaba sólo unasmínimas áreas de adhesión de losmúsculos de vuelo. Le faltabacompletamente la enorme quilla, oesternón, que proporciona a las avesvoladoras el área de adhesión principal para los músculos de vuelo[21]. Además,las crestas en el antebrazo donde seinsertan estos músculos son muy pequeñas en el Archaeopteryx de formaque, sin duda, éste no podía haber tenido

músculos de vuelo potentes. Laarticulación del hombro es típica de losdinosaurios y la fosa articular estáorientada hacia abajo, en lugar de haciafuera como en las aves, lo cual habríadificultado el movimiento de las«alas[22]». Después de descender el ala,el animal no hubiese sido capaz devolver a levantarla ya que los músculoselevadores aún no habían sidodesarrollados[23]. En vez de tener menosmúsculos que las aves modernas, el
rchaeopteryx habría necesitado más afin de poder elevarse en el aire debido asu mayor peso (sus huesos eran pesados,el pesado cráneo estaba lleno de dientes tenía una larga cola ósea).

Los huesos del Archaeopteryx eracompletamente inadecuados para unaexistencia voladora. Al igual que loscelurosaurios, las articulaciones de loscodos, muñecas y dedos eran móviles yno fundidas, mientras que, en las aves,los codos y las muñecas sólo tienen unalimitada libertad de movimientos y losdedos están fundidos, de forma que, econjunto, forman un sólido puntal en elala a fin de soportar el esfuerzo delaleteo. Incluso suponiendo que el
rchaeopteryx se hubiera convertido eun animal volador prematuramente, nohabría sido capaz de aterrizar coseguridad, ya que no disponía de uadecuado tren de aterrizaje[24]. El

cinturón pélvico de las aves estáreforzado y fundido con la columnavertebral en la región de la cadera, paraabsorber el impacto mientras que el
rchaeopteryx sólo tenía elequipamiento estándar de losdinosaurios que, naturalmente, no servía para realizar proezas tales como saltar de los árboles. Empieza a parecer comosi L. B. Halstead tuviera razón alafirmar que las aves más primitivas noeran voladoras[25]. Volar es la forma másdelicada —además de peligrosa— delocomoción jamás inventada por losanimales vertebrados y, a pesar de todo,¡el Archaeopteryx lo había logrado, o almenos esto es lo que frecuentemente nos

han hecho creer, sin estar incapacitado por su falta de músculos de vuelo!
Resulta extremadamente tentador olvidar la herencia dinosáurica de esteextraordinario fósil pequeño, co plumas, expuesto (de forma bastanteapropiada) en el exterior de la Galeríade Dinosaurios en el Museo de Historia
atural de Londres e imaginárselo,como han hecho muchos, volar de árbolen árbol buscando insectos.Recientemente, el problema ha sidoenfocado desde otro ángulo,demostrando las consecuencias quehubiese tenido el intento del
rchaeopteryx de volar. Parece extrañoque tuviera que pasar un siglo antes de

aplicar los principios aerodinámicos aeste supuesto volador. En 1970,W. G. Heptonstall, de la Universidad deEdimburgo, extrapoló algunos hallazgosrecientes basados en el vuelo de la paloma para el antepasado más antiguode la paloma. Naturalmente, el
rchaeopteryx —inmóvil como está esu plancha rocosa— no es tan apto comouna paloma para la técnica experimental, pero aun así se pueden aplicar algunasde las consideraciones teóricas. Debidoa que el antebrazo tiene que soportar la principal fuerza de sustentación, resultaser la estructura anatómica sometida almayor grado de esfuerzo durante elvuelo. Conociendo el grosor del hueso,

su resistencia a la tensión, Heptonstall pudo calcular el peso máximo que podían elevar y sostener en el aire lasalas. Estimó que las alas-extremidadesdel Archaepteryx sólo podrían haber resistido el deslizamiento, ya que elvuelo aleteante hubiese requerido uesfuerzo demasiado grande y el huesodel brazo se habría roto mucho antes deque el ave llegara a flotar en el aire[26].El Archaeopteryx tendría que haber aleteado con una velocidad dos vecessuperior a la de las palomas para permanecer en el aire, pero, debido aque ni siquiera tenía la fuerza de una paloma, forzosamente tenía que permanecer en tierra.

Todos aquellos a los que les gustaríaver al Archaeopteryx como un animalvolador, critican las estimaciones quehizo Heptonstall del peso del
rchaeopteryx de 500 gramos,señalando que las aves modernas detamaño similar sólo pesan la mitad[27].Además, se podría haber logrado ciertogrado de elevación por la larga colacomo, por ejemplo, en el «Concorde».Sin embargo, difícilmente puede parecer razonable esperar que un dinosauriohubiera descubierto la forma de reducir el peso desarrollada por susdescendientes a lo largo de 140 millonesde años de evolución. Las avesdispusieron de mucho tiempo, y de u

largo período de entrenamiento para perder peso y así aumentar la eficacia:la presencia de huesos completamenteneumáticos, la pérdida de la cola y elaligeramiento del cráneo por la pérdidade los dientes sólo son algunas de lasventajas no compartidas por el
rchaeopteryx. Las estimaciones deHeptonstall probablemente se acercamás a las características de udinosaurio que apenas ha empezado a pensar en volar.
De forma que nos encontramos en la paradójica situación de que el
rchaeopteryx, probablemente, aún no podía volar a pesar de que tenía alas.¿Por qué había desarrollado plumas si

no era para volar? La solución de este problema ya había sido hallada hacemucho tiempo, incluso antes decomprender que los primitivosdinosaurios eran de sangre caliente. La pista la proporcionaron las avesmodernas que necesitan las plumas paraconservar el calor, al igual que losmamíferos necesitan el pelo. Durantemucho tiempo, también se haargumentado que la presencia de plumasen el dinosaurio-ave Archaeopteryx eraun indicio de que ya se había instauradola endotermia[28]. Hace un siglo, Oweadvirtió la importancia de las plumas.«Podemos deducir con toda seguridadque el Archaeopteryx era de sangre

caliente —afirmó—, debido a que tenía plumas y no debido a que podía volar».En realidad, analizándolo lógicamente,no podía haber sido de otra forma. ¿Por qué una criatura de sangre fría, quedependía de la temperatura externa,tendría que desarrollar plumas? Las plumas evitan la pérdida de calor ecriaturas que están produciendoactivamente calor a través de smetabolismo, no facilitan la absorciódel calor. Tal como observa Ostrom, las plumas serían un obstáculo positivo paralos reptiles de sangre fría, ya que «las propiedades aislantes de las plumastambién reducen o minimizan laabsorción de calor por temperaturas

ambientales elevadas o radiación solar.Este hecho establece claramente que laselevadas temperaturas corporales,endógenas y la homeotermia efectivafueron logradas antes de que existiera laenvoltura de plumas. ¡La primeracriatura con plumas no podía haber sidoun ectotermo! El Archaeopteryx y sus predecesores inmediatos dependían delcalor metabólico interno, no de lastemperaturas externas. Y lo que es aúmás importante: esto significa que laendotermia debió de preceder alvuelo[29]».
El bello reductio ad absurdum deOstrom, expresado de forma sencilla, esel siguiente: las criaturas de sangre fría

con plumas estarían aisladas de la fuentede energía que resulta tan vital para sexistencia. Por lo tanto, las plumas no podrían haber aparecido en animales desangre fría. El ave-dinosaurio
rchaeopteryx debió de ser de sangrecaliente. Las plumas son el argumento
más fuerte para la aceleración
metabólica de los ágiles celurosaurios.Las plumas proporciona
aislamiento. Al igual que el pelo de losmamíferos, están diseñadas para atrapar una capa de aire junto a la piel que puede actuar como un amortiguador dela temperatura. Este aire es calentado por el cuerpo y enfriado por laatmósfera, de forma que la piel nunca

queda expuesta directamente al aire frío.Debido a esto, el cuerpo está rodeado por una capa de aire caliente a dondequiera que vaya. También significa queun animal de sangre caliente, quecontinuamente está produciendo calor para mantenerse caliente, no pierdeenergía debido a una pérdida excesivade calor. Cuanto más frío sea el tiempo,más plumas son «encrespadas» por losmúsculos superficiales, paraincrementar el volumen de aireatrapado. Los que piensan que las plumas fueron desarrolladas para volar,argumentan que, si su función originalera el aislamiento, el Archaeopteryx
debería haber tenido plumones. Si

embargo, esto presupone que los plumones sean un aislante más eficazque las plumas de contorno queenvuelven el cuerpo, lo cual no escierto[30]. Los plumones nunca estásituados por debajo de las plumas decontorno en las aves modernas, y son las plumas de contorno las que estácontroladas por músculos paraincrementar o reducir los espacios deaire aislantes. Aunque el Archaeopteryx
de Londres no presenta plumas decontorno visibles (éstas, sin duda, se perdieron después de la muerte tal comoa veces sucede con las gaviotas muertasen la costa, quedando sólo las plumas delas alas y de la cola), el espécimen de

Berlín sí que las tiene en el dorso,cuello, pecho y piernas, quedando asídemostrado que el Archaeopteryx estabacompletamente cubierto de plumas evida[31].
Las plumas se desarrollaron paraque los pequeños dinosaurios de sangrecaliente pudieran hacer frente a la pérdida de calor. La palabra clave aquíes «pequeño», ya que es el tamaño de
los celurosaurios lo que explica por
qué aparecieron las plumas en primer
lugar . Los dinosaurios, tal como ellector recordará, estaban limitados a ugran tamaño por su fisiologíaendotérmica. Cualquier intento devolverse más pequeños habría sido

frustrado por la excesiva pérdida decalor a través de su desnuda piel, ya queun descenso del tamaño también habríaido acompañado de un incremento delárea relativa de superficie. Sin embargo,el Archaeopteryx era pequeño, no másgrande que un cuervo, y con mucho, eldinosaurio más pequeño que conocemos(medía 30 centímetros menos que elCompsognathus, su rival más próximo).Sólo podría haber sobrevivido alaumento de la exposición aislando s piel y evitando así la pérdida de calor.
as aves fueron el resultado del intento
de los dinosaurios de reducir su
tamaño, y las plumas el legado querepresentaba su éxito.

Es posible que otros celurosaurios pequeños del jurásico también tuviera plumas, y que las plumas no se hubieraconservado debido a que los sedimentoscircundantes no eran tan finos como losde Solnhofen[32]. No tenemos ningúindicio de la envoltura de piel de lamayoría de dinosaurios, sólo deaquellos encontrados como momias queson pocos y pertenecientes sólo a lasespecies más grandes. El Archaeopteryx
proporciona la prueba crucial de que almenos algunos de los dinosaurios más pequeños tenían plumas. Una vez todaslas aves fueran completamente plumadas, constituirían una duracompetencia para todos los pequeños

no-voladores con plumas.Sólo después de que los dinosaurios
pequeños hubieran adquirido plumas para hacer frente a una mayor área desuperficie, éstas pudieron cambiar susfunciones permitiéndole a las aves evías de desarrollo romper sus lazos cotierra firme. La importancia de loscomentarios de introducción relativos ala «adaptación previa» es ahoraevidente. Las plumas de contorno fuerodesarrolladas como aislantes térmicos, pero proporcionaron a los pequeñosdinosaurios una ventaja aerodinámicaque los capacitó para conquistar el aire.
Las plumas de contorno del cuerposólo podían ser levantadas o bajadas

para aumentar los espacios aislantes deaire, si poseían un fuerte eje central eel cual se pudieran insertar losmúsculos. Sólo este tipo de plumaapoyada podría haber estado colgadadetrás del brazo, formando el aparatovolador. Debido a que la presencia de plumas largas, colgantes, hubiese proporcionado escaso, si acaso algúaislamiento, ¿podemos suponer que la posesión de estas plumas implica unaexistencia aérea? El Archaeopteryx
estaba adornado con estas largas plumasen las alas y si éstas no eran para dar calor debemos dar una explicación parasu función.
El Archaeopteryx pasa por ser u

planeador, y hemos visto que sus alas,mecánicamente eran lo suficientementefuertes como para sostenerlo pasivamente en el aire. Sin embargo, uelevado peso corporal, junto con laexistencia de alas aerodinámicamenteineficaces (las garras debían de generar muchas turbulencias), resultaban en unavelocidad rápidamente declinante que, probablemente, produciría un choque bastante fuerte con su desprotegida pelvis. En vez de como grácil planeador,sería más adecuado imaginarse a este pequeño dinosaurio cayendo al suelocomo un plomo de manera poco digna.La presencia de nodos en el esqueletodel antebrazo de los voladores

modernos demuestra que las plumasestán fuertemente adheridas al hueso,mientras que, en el Archaeopteryx,Ostrom observa la ausencia casi total deestos nodos, indicando con ello que sus plumas sólo estaban implantadassuperficialmente en la piel. Ello,indudablemente, hacía del salto e paracaídas, desde cualquier altura, unaocupación peligrosa.
Estas especulaciones presuponeque el Archaeopteryx era, en primer lugar, un trepador de árboles. Aunque parezca paradójico, los árboles no soel mejor lugar para desarrollar plumas.Para las criaturas que trepen sobre uárbol, como lo hacen los murciélagos, la

presencia de plumas en los brazos seríaun obstáculo positivo; y tanto más parael Archaeopteryx ya que éste no eracapaz de retraer completamente sus alas.Si lo esencial era la velocidad, tanto para escapar de depredadores terrestres,como para intentar coger insectosvoladores, su probabilidad desobrevivir no hubiese sido muy elevada.
o obstante, había ventajas positivas para, en un momento determinado, trepar a los árboles como, por ejemplo, lacolonización de algún nicho ecológicovacío. Los primitivos anfibios salierodel agua (algunos dirían que forzados por la feroz competencia), y prosperaroen las deshabitadas orillas de los lagos

mientras que los reptiles cambiarocompletamente su forma de vida,alejando sus huevos (y por lo tantocrías) de los peligros de una existenciaacuática, proporcionándoles una cáscara desovando en tierra firme. La
competencia es la que proporciona losmotivos para la innovación. Las avesreaccionaron de la misma forma frente ala presión, poniendo sus huevos en losárboles, donde los podrían incubar relativamente seguros. Sin embargo, esmuy improbable que el Archaeopteryx
diera este paso.La compleja coordinación entre los
ojos y las manos que se necesita paratrepar a un árbol, sin mencionar la

agudización de los sentidos que debeacompañar al salto de paracaídas,hubiese dejado algún signo revelador eel cerebro de un Archaeopteryx arbóreo.Sólo hay que pensar en el efecto quetuvo el trepar a los árboles, unido a ladestreza manual sobre la evolución delcerebro de nuestros antepasados, paradarse cuenta de la inextricable relación.En realidad, al principio parecía que elcerebro del Archaeopteryx, investigadode nuevo en 1968 después de haber sidodeclarado completamente característicode los reptiles una generación antes,mostraba incipientes característicasavícolas[33]. Harry Jerison encontró queel cerebro del Archaeopteryx estaba a

medio camino entre el de un avemoderna y el de un reptil moderno, e base a su tamaño mientras que su forma parecía ser más avícola que reptil,sugiriéndole a Jerison que ya se habíaestablecido reacciones como las de lasaves. Sin embargo, ¿son los reptilesmodernos realmente una buenacomparación? No solamente tienecerebros increíblemente pequeños, sinoque están (con la excepción parcial delos cocodrilos) sólo muy remotamenteemparentados con las aves o el
rchaeopteryx. Es cierto que la mayoríade los dinosaurios también teníacerebros notoriamente pequeños, perocada vez se hace más patente que los

celurosaurios y dromeosaurios, amboscorredores ágiles y rápidos con manosaferradoras, estaban provistos decerebros relativamente gigantes y ojosenormes para coordinar sus sofisticadosreflejos. Debido a que el Archaeopteryx
se originó a partir del tronco de loscelurosaurios, sería de esperar quetuviera un cerebro más grande de lonormal como rasgo estándar. El tamañodel cerebro ya no puede ser utilizadocomo prueba de los hábitos trepadoresdel Archaeopteryx.
Los pequeños dinosaurios bípedostenían un aspecto tan parecido al
rchaeopteryx que, tal como hemosvisto, los paleontólogos ortodoxos se

tomaron mucho trabajo en enfatizar lomaravillosa que es la convergencia (y,de hecho, lo es), pudiendo desarrollarseestructuras casi idénticas en animalesemparentados lejanamente: estasestructuras debieron desempeñar funciones similares. Sin embargo, estamisma ortodoxia había llegado aconfirmar la hipótesis arbórea de losorígenes de las aves, según lo cual losantepasados de las aves eran trepadoresde los árboles, que en un principiovolaban como con la ayuda de u paracaídas. Como prueba de ello,observaron que las patas del
rchaeopteryx, similares a las de lasaves, presentaban dedos opuestos para

poder trepar y que las manos, en formade garras, los capacitaban para subir alos árboles, etc. Tal es la lógica paleontológica que en estas dos teoríastan inconsistentes mutuamente pudieromantenerse durante mucho tiempo. ¡El
rchaeopteryx había, por un lado,convergido funcionalmente con losdinosaurios corredores y, por otro lado,era una criatura que trepaba a losárboles! El dedo opuesto del
rchaeopteryx también estaba presenteen los dinosaurios carnívoros. La únicaforma de explicar esta paradoja, en elcaso de que se tratara de equivalentesconvergentes o funcionales, era suponer que el Tyrannosaurus había trepado a

los árboles o que el Archaeopteryx
había sido un depredador terrestre.Debido a que la primera opción es bastante improbable, John Ostrom —quefue el primero en resolver convincentemente la paradoja— escogióla segunda[34]. Las adaptaciones comoson los dedos opuestos y las manos eforma de garras, que estaban presentesen los celurosaurios, también existían elas protoaves que, supuestamente, podían trepar a los árboles debido a queéstas también eran corredoras bípedas.
El barón Nopcsa, pintoresco espíatransilvano, experto en dinosaurios yautoproclamado heredero de la coronade Albania, había llegado a

conclusiones similares, en 1907 (aunquela teoría cursorial fue propuesta por Samuel Williston en 1879). Nopcsaargumentó que, debido a que las patasdel Archaeopteryx eran idénticas a lasde los dinosaurios, las aves primitivastambién debieron de ser criaturas quehabitaban en el suelo. Creía que las plumas se originaron a partir de prominencias escamosas en los bordesde las extremidades anteriores, paraincrementar la velocidad de carrera delanimal mientras movía los brazos haciadelante y hacia atrás como remos. Estas plumas permitían a las protoaves dar largos saltos como los gansos,elevándose parcialmente en el aire.

Pensaba que esta ascendencia corredorade las aves quedaba reflejada en losavestruces que habían conservado esteestado primitivo[35]. A pesar de que lahipótesis «cursorial» es aceptable, lacausa de la adquisición de las plumas parece improbable. Debido a que elobjetivo del desarrollo de las alas eraincrementar la velocidad, el ave habríaescogido el camino equivocado paralograrlo. ¡Si se estudia la formaaerodinámica de un coche de carrerasmoderno, se observará que los planosaerodinámicos están diseñados paramantener el coche «en» el suelo y no para elevarlo en el aire! La pérdida detracción es igual a la pérdida de

velocidad.Ostrom ha evitado estos problemas
partiendo de la idea de que las largas plumas de los brazos no tenían nada quever con la capacidad de volar olocomoción y que, en lugar de ello,estaban relacionadas con ladepredación. El Archaeopteryx era udepredador ligero de pies, con una buena visión y fuertes manos.Probablemente cazaba insectos grandes la mejor forma para hacerlo era
usando una red de plumas: las plumas delos antebrazos actuarían a modo detrampa para atrapar los insectosvoladores. De esta forma, el ave prácticamente podría rodear los insectos

antes de cogerlos con sus garras. Mástarde comenzó a dar saltos como lasgallinas para cazar los insectosvoladores. De esta forma, el aleteo sehizo importante para la caza de las presas y debió de acompañarse de unacoordinación nerviosa más precisa. Elave-dinosaurio, provisto de endotermia, plumas, alas y un cerebro capaz decoordinar maniobras complicadas, yaestaba completamente «preadaptado» para volar.
De esta forma nacieron las aves: noa partir de los pterodáctilos como habíaimaginado St. George Mivart en el siglo pasado. Mivart estaba tratando deeconomizar sus hipótesis —o, en ese

caso, inventos—, permitiendo que elvuelo surgiera sólo una vez durante elmesozoico entre los antepasados pterodáctilos de las aves voladoras. Aligual que el barón Nopcsa, después deél, se imaginó a aves no voladoras comolos avestruces, originándosedirectamente a partir de los dinosauriosindependientemente de las avesvoladoras. El origen pterodáctilo fue puesto en duda rápidamente, aunque laascendencia de las aves no voladorasfue una idea que Percy Lowe encontróatractiva en la década de 1920. Lasteorías de Lowe, alimentadas durante u período de un cuarto de siglo, fueroformuladas por primera vez como

reacción al lamarckismo. Las creenciasa las que se oponía Lowe habían surgidode África del Sur, donde un tal doctorJ. E. Duerden, un colega experto eavestruces con el detectivesco título de«Director de las Investigaciones sobreAvestruces», en la Universidad deRhodes, había cometido la herejía biológica de negar una explicación deDarwin referente a las callosidades eel pecho y rabadilla de losavestruces[36]. Los avestruces caen haciadelante cuando se agachan y estasalmohadillas engrosadas le proporcionan algo de protección al ave.Duerden pensaba que estas callosidadesno están codificadas genéticamente sino

que aparecían como respuesta a lafricción, como ocurre con los calloshumanos. Sin embargo, debido a queaparecieron por primera vez eembriones de avestruz (obviamenteantes de que se pudiera producir fricción alguna), el doctor concluyó quese trataba de caracteres adquiridosdurante la vida de los padres y quehabían sido transmitidos a ladescendencia. El comportamiento de los padres literalmente modificó laestructura de sus descendientes[37]. Elcaso clásico y más repetido del supuestolamarckismo es el de la jirafa. Esteanimal, en algún momento de su historia(cuando poseía un cuello truncado),

comenzó a estirarse para alcanzar lashojas más altas y allí encontró que lacomida era tan buena que de algunamanera informó a sus descendientes efase de desarrollo sobre las obviasventajas de tener un cuello largo, y laoven jirafa apareció debidamente co
uno. Se trata de un proceso un tantomágico, ya que no se conoce ningúmecanismo a través del cual las célulascorporales puedan influir a las célulasreproductoras. Tanto Darwin comoLamarck creían en este tipo de proceso, pero es comprensible ya que, por aquelentonces, no existía ninguna alternativa.Sin embargo, a comienzos de este sigloel neodarwinismo ofreció una

explicación alternativa: las mutacionesque actúan sobre el material hereditario podrían ser las responsables de loscambios observados en los animales y, posteriormente, estos cambios aleatorios(como un cuello ligeramente más largoen una jirafa) podrían ser seleccionadossi resultaban ser ventajosos. Habíasido desterrados eficazmente los últimosvestigios del lamarckismo.
Así, las teorías anacrónicas deDuerden podían ser criticadas en elsentido de que no existía ningúmecanismo que pudiera explicar latransmisión hereditaria de los caracteresadquiridos. Lowe, sin embargo, atacódesde otro punto de vista. Las

callosidades, dijo, estaban presentes elos primitivos dinosaurios corredoresque también habían pasado muchotiempo agachados[38]. Los avestruceseran un grupo arcaico distinto del restode aves, ya que nunca se habían elevadoen el aire; después de todo no se habíaencontrado fósiles de avestrucesvoladores. Lowe creía que se originabadel ave buceadora cretácea Hesperomis
(que el propio Marsh había descrito unavez no demasiado en serio como«avestruz carnívoro nadador») que, aligual que el avestruz, no podía volar.Los ñandúes, que baten las alas cuandocorren a través de las Pampas, tampocoapoyaban la teoría de que, en otros

tiempos, habían sido aves voladores;Lowe pensaba que los dinosaurioscorredores también se habíadesplazado moviendo las extremidadeshacia delante y atrás, a modo de remos, nadie había sugerido nunca que los
dinosaurios habían sido alguna vezcriaturas voladoras. Los avestruces,simplemente, no se elevaron en el aire.En opinión suya, los avestruces habíacambiado poco desde que salieron delas filas de los reptiles bípedos. Paraapoyar su teoría, citó la presencia de plumones, similares a los de los polluelos, en los avestruces adultos quesuponía eran una característica primitiva(tal como hemos visto, probablemente se

originaron secundariamente a las plumasde contorno con eje). Las plumassimilares a las de los polluelos presentes en los avestruces adultosindicaban que no había cambiado desdeel período mesozoico, conservando uestado primitivo y sin evolucionar haciaun estado volador. «En lo que respecta asus plumas, los estrutiones (avestruces)son polluelos grandes que han crecidodemasiado. Son los “Peter Pan” delmundo avícola. No se han hecho adultosnunca[39]».
Es un concepto bastante agradable:dinosaurios que degeneraron aavestruces. Naturalmente, hay una partede verdad en esto, pero sólo en el

sentido de que todas las aves seoriginaron a partir de los dinosaurios. Elavestruz no se parece en nada a losantepasados de las aves, que era bastante pequeños y parecidos al
rchaeopteryx, si es que no lo era elmismo fósil famoso. Hoy la opiniógeneral es que las aves primitivas podían volar y que sólo más tardealgunas volvieron al suelo paraconvertirse en aves corredoras. Las plumas similares a las de los polluelos presentes en los avestruces, se deben aque los animales adultos sigueconservando rasgos juveniles: procesoque se conoce con el término deneotenia. (Los humanos también so

neotenios porque conservan cabezasgrandes embriónicas y no tienen pelo).
Las aves, al igual que los mamíferos,fueron una raza dominada durante elmesozoico y continuaron siendo uelemento poco llamativo de la fauna. Seconvirtieron en hábiles voladores, perofueron incapaces de explotar el suelocomo corredores, por miedo a ser cazados por los depredadores gigantes.Muchas aves se refugiaron en el mar yalgunas, como el Hesperomis cretáceode Marsh[40], redujeron el tamaño de susalas a muñones y desarrollaron fuertes patas para impulsarse en el agua. Sólocon la desaparición de los dinosauriosal final del cretáceo, las aves pudiero

prosperar en las llanuras vacías. Debidoa que la forma más rápida paradesplazarse por terreno llano es correr,durante el período terciario yaencontramos aves terrestres grandes deextremidades largas. Éstas estabatratando de llenar el nicho dejadovacante por los dinosaurios, y asísucederlos como la clase dominante deanimales terrestre. A comienzos deleoceno ya existía un gigante, el
iatryma de 2,10 metros de altura, cohuesos macizos y un enorme cráneo co pico que vivía en las llanuras deWyoming. En la misma época existieroen Inglaterra y Francia gansos deltamaño de avestruces que vagaban por

el campo[41]. Durante el eoceno, las aves proliferaron hasta tal punto que parecíaque se estaban apoderando del mundo.Las aves llegaron antes que losmamíferos a los nichos destinados a losanimales de cuerpo grande, en otrostiempos monopolizados por losdinosaurios, y florecieron en gravariedad de formas grandes. Las aves novoladoras continuaron aumentando detamaño durante el período terciario,desarrollaron robustas patas corredoras,cuellos largos y una buena visión y sevolvieron más parecidas a losdinosaurios bípedos. Sin embargo, estono duraría mucho tiempo. Losdepredadores mamíferos causaron gra

número de muertes entre las grandesaves no voladoras y, gradualmente,fueron limitando su zona de distribución.En los últimos tiempos, el hombre hademostrado ser el más destructivo detodos los mamíferos. La primitiva faunade Madagascar incluía, en otros tiempos —y en épocas más recientes—, el
epyomis titan, el pájaro elefante.


p. Poco después de la desaparición de losdinosaurios, fuertes aves terrestres como el Diatryma (arriba), encontrados en el eocenoinferior de Wyoming, penetraron en el nichoecológico vacío en un intento de suceder alos dinosaurios.
Los huevos de esta ave de 3 metrosde altura, que tenían una circunferenciade aproximadamente 90 centímetros, aúse encuentran ocasionalmente en la costade Madagascar. Los antiguoscomerciantes árabes que llegaron a estascostas, y encontraron estos enormeshuevos, no pudieron encontrar a los padres e inventaron cuentos sobre elfabuloso Roc, un pájaro gigante lo

suficientemente fuerte como parallevarse a elefantes incautos. El otrosantuario de las criaturas separadas por el tiempo es el continenteaustralasiático, donde los extinguidosmoas gigantes incluso superaban a lostitanes de Madagascar, Dinomis
maximus, cuyo nombre superlativoliteralmente significa «el más grande delas aves gigantes», medía hasta 3,5metros de altura. Los moas aún eran losseñores de su reinado cuando losmaoríes llegaron a Nueva Zelanda, peroencontraron la muerte como comida delos maoríes. Es un final bastanteignominioso para una odisea quecomenzó cuando los dinosaurios trataro

de volverse más pequeños.

VII. FANTASMASDEL INFIERNO
Cuando, en 1784, se produjo eldescubrimiento del primer pterodáctiloen la piedra caliza de la localidad bávara de Eichstätt, hecho por elnaturalista italiano Cosmo AlessandroCollini, el hallazgo constituyó el últimoenigma para todos los curiososcientíficos de Europa. El pterodáctilosería, junto con el mosasaurio, el fósilque probaría de forma dramática larealidad de la extinción. Además, dejó bastante manifiestas las grandes

desigualdades que existen entre lasformas actuales de vida y las del lejano pasado. Cuvier, que sería el que reuniríalas pruebas fósiles que harían creíble laextinción, tenía graves dudas acerca delas interpretaciones contemporáneas deeste extraño animal prehistórico. El fósilfue llevado al Grand Ducal Museum, eMannheim, del cual Collini era eldirector. Collini daba por sentado que lacriatura era un habitante de los antiguosmares, pero los doctores alemanes quevisitaron Mannheim tenían otras ideas.S. T. Sömmering sostenía que se tratabade una criatura intermedia entre las aves los murciélagos, mientras que Lorenz
Oken no se podía decidir entre u

mamífero y un reptil. Cuvier se diocuenta del parecido superficial con losmurciélagos y los vampiros en su formageneral, su pico era alargado como el delas chochas pero provisto de dientescomo el hocico de los cocodrilos, susvértebras y las patas eran similares a lasde un lagarto, sus dedos en forma degarras eran como los de los murciélagos su cuerpo estaba cubierto de escamas
como en las iguanas. Cuvier eraconsiderado como uno de los paleontólogos más perspicaces deEuropa, reputación que se había ganadogracias a su brillante técnica deanatomía comparada. Por consiguiente,era de él de quien se esperaba co

impaciencia la solución del enigma.Después de haber examinado el fósil deCollini, a principios del siglo XIX, prestando una especial atención a scráneo y articulación de los maxilares,Cuvier afirmó que, con toda seguridad,se trataba de un saurio, un reptil delantiguo mundo. Sus cuatro dedosalargados sostenían las alas que locapacitaba para volar y, por esta razón,lo llamó Pterodactylus o «dedo de ala».Aunque pertenecía a la clase de lossaurios y, por lo tanto, estabaemparentado con los reptiles modernos,Cuvier hizo la siguiente advertencia: detodos los seres del mundo antiguo, ésteera el más extraordinario y, si se le

devolviera la vida, no se parecería aningún otro tipo de forma de vida[1].
Ya no se podía dudar durante mástiempo que razas enteras de animaleshabían desaparecido de la faz de latierra, o que la vida primitiva habíatenido un aspecto arcaico. A pesar detodo, la idea de que el pterodáctilohabía sido una criatura marina estaba taarraigada que muchos se opusieron aCuvier. Los pterodáctilos traeríamuchos problemas en el futuro. «EInglaterra —dijo Harry Govier Seeleymás tarde—, son clasificados comoreptiles, principalmente debido a lainfluencia que tuvo sobre ellos ladisertación del barón Cuvier [2]». Si

embargo, los doctores alemanes noaceptaron las ideas de Cuvier,simplemente, debido a la grareputación de la que gozaba el científicofrancés. En 1830, Johannes Wagler vioen el pterodáctilo un parentesco con losictiosaurios y los plesiosaurios, quehabitaban los océanos del mesozoico. Aligual que éstos, nunca abandonó el agua«pero nadaba en la superficie como ucisne y buscaba los alimentos en elfondo del mar». Se imaginaba que loslargos brazos funcionaban como lasaletas de los pingüinos o de las tortugas, que los tres pequeños dedos en forma
de garras servían para agarrar a «lashembras en el proceso reproductor [3]».

Al año siguiente, 1831, Georg AugustGoldfuss, profesor de zoología en laUniversidad de Bonn y una eminenteautoridad alemana en animales fósiles,confirmó el diagnóstico de Cuvier: los pterodáctilos eran reptiles. Sin embargo,añadió Goldfuss, se trataba de reptilesde aspecto muy parecido a las aves y, probablemente a consecuencia de estacreencia, dijo haber observado crestas parecidas a las plumas grabadas en elfósil, adorno que fue rechazado covehemencia por sus contemporáneos.
Durante la década de 1820, muchosde los recolectores ingleses de fósilescreyeron que los delgados huesos de suscolecciones no pertenecían a aves, sino

que eran restos de esta extraña criaturareptil descrita por Cuvier. Pero no fuehasta diciembre de 1828 cuando sedescubrió un esqueleto casi completo.Prácticamente, todos los hallazgos delos primitivos reptiles dominantes eInglaterra fueron hechos por aficionados, y muchos de éstos por unoen particular, una joven y talentosa damallamada Mary Anning. Loscontemporáneos menos dotados tienen latendencia de otorgar a las personas cotalento alguna desgracia ocurrida en suventud que justifique este don anormal, Mary Anning no fue ninguna
excepción. A la tierna edad de un año,ella y su niñera fueron alcanzadas por u

rayo. La niñera falleció pero la niña«pudo ser reanimada metiéndola en u baño de agua caliente; no había sufridolesión alguna. Antes había sido una niñainactiva, pero después de este accidente,se convirtió en una muchacha másvivaracha e inteligente[4]». Habiéndosedado cuenta de su talento, abrió unatienda con su hermano en Lyme Regis,Dorset, y allí vendía los fósiles querecogía durante sus paseos diarios a lolargo de los vecinos acantilados. E1811, cuando sólo contaba 11 años deedad, halló un esqueleto de ictiosaurio y,en 1824, el primer plesiosauriocompleto. Todos los geólogos de aquellaépoca le ofrecieron su amistad y muchos

fueron a Lyme Regis, entonces un lugar de moda, a pasar sus vacaciones para poder pasear con Mary por losacantilados. Cuando, en 1828,desenterró el primer pterodáctilo inglésse lo mostró a un frecuente visitantesuyo, el reverendo William Buckland dela Universidad de Oxford. Buckland ledescribió el fósil a la SociedadGeológica de Londres al año siguiente(aunque la descripción no fue publicadahasta 1835, una ilustración gráfica de lalentitud de la ciencia a principios delsiglo XIX; el ritmo sería aceleradoconsiderablemente cuando Marsh yCope comenzaron a disputarse la prioridad). Aunque ya había sido

preparado por Cuvier para ver algoasombroso, la total sorpresa que sellevó Buckland cuando le fue entregadala ilustración hizo que repitiera las palabras de Cuvier. Era, dijo, «umonstruo que no se parecía a nada jamásvisto o conocido en la tierra,exceptuando los dragones de las novelaso libros de caballerías[5]». La técnicaanatómica de Cuvier, tan inspiradora para Buckland, había revelado al animalcomo una criatura voladora. Siembargo, Buckland estaba claramenteinfluido por algunas de las ideas rivales volvieron a aparecer viejas
confusiones. Buckland estabadesconcertado por los cortos dedos e

forma de garras: debieron de ser fuertesgarras, pensó, que permitirían al animalreptar, o trepar o colgarse de losárboles. ¡Al año siguiente, en scontribución a los Bridgewater
Treatises on the Power Wisdom an
Goodness of God as manifested in the
Creation, agotó todas las posibilidadesde hacer de él también una criaturaacuática! «Así, como el demonio deMilton, cualificado para todos losservicios y todos los elementos, lacriatura era un compañero idóneo paralos emparentados reptiles que pululabaen los mares o se arrastraban en lasorillas de un turbulento planeta:

El demonio,a través de los pantanos,
cuestas empinadas, estrechos,terrenos quebrados y vegetacióndensa o escasa, sigue su caminocon la cabeza, manos, alas o pies, y nada o se sumerge, ovadea, o se arrastra o vuela.
EL PARAÍSO PERDIDO, Libro II
»Con bandadas de criaturas de estetipo, volando en el aire, y multitudes deictiosaurios y plesiosaurios no menosmonstruosos en los mares, y cocodrilosgigantes y tortugas arrastrándose por lasorillas de los primitivos lagos y ríos, elaire, el mar y la tierra debieron de estar

habitados de forma extraña en los primeros períodos de nuestro jovemundo[6]».
La imagen evocada por Buckland fuereflejada de forma ilustrativa por otroaficionado bastante excéntrico deDorset, sólo cuatro años más tarde.Thomas Hawkins, un coleccionista defósiles saurios marinos, reprodujo deforma gráfica la confusiócontemporánea e hizo una interpretacióteológica que resultaba bastanteanacrónica, incluso en 1840. La portadadel libro de Hawkins, Book of the Great
Sea-Dragons, inchthyosauri an
lesosiosauri, Gedolim Taninim o
oses, Extinct Monsters of the Ancient

arth, había sido confeccionada por John Martin y representaba la carniceríaque se producía cuando estosmonstruosos seres se juntaban en elmundo preadanita. Los grabados de JohMartin también adornaban la obra deGideon Mantell, Wonders of Geology,donde ilustraba a un Megalosaurus y u guanodon luchando encarnizadamente.Estos grabados sólo recibieron elogiosde Hawkins y Mantell a pesar de que,como reconstrucciones científicas, ausiendo las primeras de este tipo, más bien parecían caricaturas de aparicionesgrotescas. Othenio Abel, profesor de paleobiología en la Universidad deViena, a principios de este siglo, y

experto en reconstrucciones, las censuró por ser tan atroces como la fantasía deJulio Verne. Nos recuerdan, dijosarcásticamente, una «figura fantasmalde uno de los infiernos de Breughel[7]».
Hawkins era un recolector infatigable y, en su afán de «sacar a losantiguos saurios de sus tumbas de roca»,tal como los describió Richard Owen,acabó llevándose yacimientos enteros.Mediante este método de demoliciógeneral, recogió algunos de los mejoressaurios marinos jamás hallados que, coel tiempo, pasaron a ser propiedad delMuseo Británico donde aún estáexpuestos hoy en día. Afortunadamente,los monstruos que habitaba

inofensivamente la tierra sólo era«efigies de razas extinguidas», razas que«eran ajenas a las actuales como lo son, probablemente, las de Saturno[8]». Erauna creación antigua de Jehová, quehacía mucho tiempo ya habíaabandonado el proyecto. «Perpetúan udiseño que ya no se usa», dijo Hawkins,de forma que sería imprudente intentar emparentar las formas pasadas y presentes. Se trataba de mundos, o almenos reinos, diferentes. ¡Estos gigantesmarinos ni siquiera pertenecían al reinoanimal y Hawkins, de acuerdo con esto,creó un nuevo reino bíblico para ellos,el Gedolim Taninim!
Cuando, finalmente, se impusiero

las ideas de Cuvier y se determinódefinitivamente la forma de locomocióde los pterodáctilos, se disipó laconfusión. Antes de que Richard Owemandara reconstruirlos en el CrystalPalace, en 1854, ya habían sidoreconocidos por todos como criaturasvoladoras. Los representó posadosmajestuosamente en la cima de un riscoa punto de elevarse en el aire, uno colas alas plegadas y el otro con las alasextendidas. Después de haber llegado ala opinión unánime de que se trataba devoladores, surgió un nuevo problema.Debido a que tanto los pterodáctiloscomo las aves se habían originado a partir de arcosaurios ancestrales, y

ambos tenían alas, algunos pensaron que podrían estar directamenteemparentados. Owen rehusócautelosamente equiparar los pterodáctilos a las aves, a diferencia deSt. George Mivart que, comoapasionado evolucionista, deseaba hacer de los pterodáctilos los antepasadosinmediatos de las aves voladoras. Los pterodáctilos habían aparecido en rocasde épocas mucho más antiguas, observócomo prueba de su teoría, y los largos picos y forma general recordaban a lasaves. La realización se hizo aún másverosímil cuando los paleontólogosempezaron a obtener pruebasfragmentarias del cerebro del

pterodáctilo, del cual se habíasospechado desde hacía tiempo que eramás grande que el de los reptilesmodernos.
q. Ilustración contemporánea de los

pterodáctilos realizada por Owen y Hawkinsen el Crystal Palace. Hawkins se imaginaba alos reptiles voladores como prototipos delmítico dragón y los construyó de acuerdocon esta idea.
Los esqueletos fósiles bieconservados de pterodáctilos querevelen detalles del cerebro son bastanteraros aunque, a lo largo del siglo XIX, sefueron reuniendo pruebas a partir dediferentes fragmentos. Posteriormente,en la década de 1880, el reverendoD. W. Purdon encontró un esqueleto deaspecto prometedor en el esquisto dealumbre del jurásico inferior en Whitby,en Yorkshire, que envió al Geological

Survey, en Londres, para su examen. Eel Survey, en 1888, E. T. Newton llamóa la criatura Scaphognathus purdoni, ereferencia a su descubridor, y,aprovechándose de su estado nofragmentado, sometió el esqueleto a udetallado estudio[9]. Una fractura en elcráneo permitió estudiar la cavidadcraneana y sugirió que se habíaconservado en las rocas un molde delcerebro durante 170 millones de años.
ewton, sacrificando los huesoscraneanos superficiales, fuerecompensado por una impresiócraneana parcialmente impecable y, a partir de ésta, confeccionó una réplicadel cerebro del «pterosaurio». (El

término colectivo de «pterodáctilo» yaestaba siendo abandonado pues sólo serefería a uno de los grupos principalesde reptiles voladores; el conjunto delgrupo se denomina ahora pterosaurios).
El diminuto cerebro de los reptilesmodernos nunca llena por completo lacavidad ósea del cráneo. Por esta razón,los moldes tomados del interior de lacaja craneana no son realmenterepresentativos del cerebro que siemprees más pequeño. En las aves y losmamíferos, en cambio, el cerebro muyexpandido ejerce presión contra las paredes del cráneo de forma que losmoldes internos proporcionan un modeloexacto del tamaño, y tanto del tamaño

como de la forma. Newton, que por primera vez examinó detenidamente elcerebro «reptil» de un pterosaurio,estaba sorprendido. «Es indudable quela forma del molde —escribió— representa la forma del cerebro del pterodáctilo, al igual que lo haría umolde tomado del cráneo de un ave o umamífero; y estos moldes muestran laforma externa natural del cerebroincluso mejor que el mismo cerebro[10]».El cerebro del pterodáctilo tambiéhabía llenado completamente su cajacraneana.
Esto no era el único parecido. Elcerebro se divide en tres regiones principales. El cerebelo, situado en la

parte posterior del primitivo cerebrovertebrado, está relacionado con elmovimiento y el equilibrio y es un área poco desarrollado en los reptiles. Loslóbulos ópticos del mesencéfalocoordinan la visión y, una vez más, somás pequeños en los reptiles ydinosaurios que en los mamíferos y lasaves. Lo mismo es válido para loshemisferios cerebrales en el prosencéfalo, que albergan los centrosde coordinación superior (una excepcióde esto, tal como hemos visto, son loságiles y listos dromeosaurios ydinosaurios avestruces). El cerebro posterior, o cerebelo, obviamente tieneuna gran importancia en las aves ya que

controla y coordina las complicadasmaniobras necesarias para el vuelo (ytambién para posarse en algún lugar yaque requiere un sentido del equilibriomucho más desarrollado que elnecesario para mantenerse sobre cuatro patas). El cerebelo se ha desarrolladotanto en las aves, que los lóbulosópticos han sido desplazados hacia ulado mientras que el dilatado prosencéfalo ha crecido hacia atrás. Loslóbulos ópticos agrandados han tenidoque expandirse lateral y ventralmentehacia la base propia del cerebro.
Esto es totalmente desconocido elos reptiles normales y, a pesar de todo,
ewton encontró exactamente la misma

condición en el Scaphognathus. Elcerebro posterior estaba aumentando detamaño y había adquirido un aspecto bastante avícola, al igual que el cerebroanterior. Estos dos se habían encontrado excluido a los lóbulos ópticos de la
superficie dorsal del cerebro. Loslóbulos ópticos eran grandes perohabían sido desplazados hacia los ladosdel cerebro. La completa reducción del bulbo olfatorio demuestra que los pterosaurios, al igual que las aves, prácticamente habían perdido su sentidodel olfato y que ambos dependían, elugar de ello, de una vista excelente. Econjunto, las semejanzas entre elcerebro del reptil volador y el de un ave

son notables. Los pterosaurios tenían,con mucho, el cerebro más grande(hablando en términos relativos) quecualquier otro reptil y, por lo que serefiere a la inteligencia, habrían hechoque sus primos reptiles pareciera bastante estúpidos, aunque entre losdinosaurios podrían ser equiparados alos dromeosaurios y a los dinosauriosavestruz. Los pterosaurios,supuestamente reptiles inferiores, habíadesarrollado de forma independiente ucerebro casi tan eficaz como el de lasaves. «Por sí solo, —dijo Seeley e1901—, el cerebro con forma avícola eun animal sería una prueba, tan válidacomo cualquier otra, de que su grado de

organización era el de una ave[11]».La razón de la semejanza de
cerebros es fácil de comprender. Los pterosaurios tenían tamaños muyvariables, que iban desde el diminuto«dragón alado», Ptenodracon, definales del Jurásico, tan pequeño comoun gorrión, hasta los gigantescos
teranodon y criaturas aún más grandesde finales del cretáceo. La mayoría delos pterosaurios pequeños tenían lasalas cortas y anchas que caracterizan alas aves que baten las alas (econtraposición con las aves planeadoras), de forma que es sensatosuponer que ellos también batían lasalas. (Las alas largas y delgadas, como

las de los albatros, de los pterosauriosque aparecieron posteriormente indicahábitos planeadores). Debido a que los pterosaurios adoptaron el mismo tipo deexistencia aérea que las aves, debieronecesitar las mismas áreas decoordinación en el cerebro paracontrolar el equilibrio y poder realizar maniobras complejas. Al igual que elas aves, su vista era obviamenteexcelente (esto también lo sabemos e base al gran tamaño de las órbitas) y es posible que incluso tuvieran una visióen color.
La idea de que los pterosauriostenían cerebros como las aves fueconfirmada completamente cuando, e

1941, fue examinado otro excelenteespécimen. El pterosaurio de Newtohabía perdido parte del cerebro, pero elespécimen descrito por el eminenteneuropaleontólogo Tilly Edinger pudoser reconstruido completamente[12]. Elnuevo espécimen, un diminuto
terodactylus de sólo unos cuantoscentímetros de longitud, y perfectamenteconservado en su bloque de Solnhofen,había sido donado al Museo de ZoologíaComparada de la Universidad deHarvard por su descubridor, LouisAgassiz, en el siglo XIX. Un gra«lóbulo temporal» en el cerebro anterior le proporcionaba a la criatura uaspecto con cabeza oviforme. En vida,

probablemente incluso había tenido uaspecto inteligente. Edinger tambiéquedó sorprendido por la forma decómo el cerebro posterior y el cerebroanterior habían crecido hasta juntarsecomo en las aves. Obviamente, observó,«éste es uno de los caracteres quediferencia a todos los pterosaurios deotros reptiles». Cualquier duda que aú pudiera quedar en relación con lainteligencia de los pterosaurios (Newtono quiso destacar demasiado estacaracterística para no apoyar la teoríade que los pterosaurios evolucionarohacia las aves), fue rápidamentedisipada por Edinger. El Scaphognathus
de Newton tenía un pico

extraordinariamente largo que hacía queel cerebro pareciese más pequeño erelación con la longitud del cráneo. Eotros especímenes, en cambio, las proporciones eran como las de las avescantoras. Cuando los pterosauriosevolucionaron, sus cerebros se hicieroaún más parecidos a los de las aves, deforma que, en el Pteranodon, la anchuradel cerebro anterior era incluso mayor que su longitud al igual que sucede elas aves. Los pterosaurios incluso poseían surcos cerebrales eexactamente las mismas posiciones queen los cerebros de las aves, mientrasque los lóbulos ópticos de los pterosaurios que apareciero

posteriormente como el Pteranodon
eran similares a los de las aves másavanzadas. En los reptiles, la parte posterior del cráneo está modelada deacuerdo con su necesidad de proporcionar áreas de adhesión para losmúsculos, mientras que, en los pterosaurios, aves y mamíferos, elcerebro posterior se ha agrandado tantoque ha formado una cúpula cerebelosaque determina la forma de la partetrasera del cráneo. «Por lo que serefiere a la longitud —concluyó TillyEdinger—, los cerebelos de los pterosaurios se hallan en el límitesuperior de variación reptil y en ellímite inferior de variación avícola»,

aunque su aspecto fuera puramenteavícola. ¡El pterosaurio debió de ser ureptil tan extraordinario como lo es uave o un mamífero! Sin embargo, aúveremos cuán extraordinario era.
r. Cerebro agrandado del Pterodactylus que

hacía presión contra su caja craneana.
Debido a que el funcionamiento deun avión se equipara a sus controles, podemos, suponer, que los pterosaurioseran voladores muy maniobrables. Losmecanismos de control estaban tasumamente bien desarrollados para elvuelo complejo que, durante algútiempo, los pterosaurios mantuvieron lossuyos en competencia con las aves evías de desarrollo. La extremadamenteactiva existencia aleteadora de las avesrequiere una gran cantidad de energía.El delicado equilibrio, la complejacoordinación y la pura fuerzarequeridos, significan que los músculos

deben recibir constantemente oxígeno para que puedan oxidar rápidamente losazúcares, a fin de liberar un flujocontinuo de energía. Estas reaccionesquímicas que producen energíarequieren una temperatura alta, constante para una liberación óptima y rápida dela energía en todo momento. Por lotanto, no es una sorpresa que las aves desangre caliente, con un elevado índicemetabólico, tengan temperaturascorporales medias uno o dos grados máselevadas que en el hombre. Al disponer de un metabolismo rápido, las aves socapaces de permanecer activas durantelargos períodos de tiempo. Han hechotodo lo posible para desarrollar

estructuras nuevas que permitan que los pulmones puedan absorber la máximacantidad de oxígeno: oxígeno requeridotanto para mantener una elevadatemperatura como para producir laenergía necesaria para volar.
La mecánica respiratoria de losmamíferos (una bomba constituida por las costillas-diafragma-pulmones), proporciona una solución al problemade la energía. Sin embargo, las avesemplean un sistema distinto, igualmenteeficaz. Aquí, los pulmones son másrígidos pero conducen a extensos sacosaéreos que se ramifican por toda lacavidad corporal. Frecuentemente,existen diez de estos sacos de aire

aunque éstos también van formandosacos más pequeños, en forma de vejiga,que se distribuyen por los brazos y las piernas. Los huesos de muchas aves sohuecos y tienen pequeñas aberturasespecíficamente para permitir la entradade los sacos aéreos (que revisten elinterior de los huesos). Los huesos delas extremidades, columna vertebral,etc., son literalmente estructuras llenasde pulmón. Se cree que estos sacos, eforma de globo, actúan como depósitosde aire para el tórax, como un fuelle, bombeados mediante el esternón y el batido de las alas (durante el vuelo, las palomas sincronizan la respiración y el batido de las alas). El bombeo impulsa

el flujo de aire a través de los diminutostubos pulmonares, donde se produce laverdadera absorción de oxígeno yeliminación del dióxido de carbonoresidual. Y, lo que es de sumaimportancia, este flujo de aire a travésdel cuerpo facilita el enfriamiento.Frecuentemente, se puede ver a las aves«jadear» después de haber hechoejercicio.
Es razonable pensar que los pequeños pterosaurios batían las alas,despegando activamente. En este caso, probablemente necesitaban un rápidointercambio de oxígeno-dióxido decarbono para obtener una producción deenergía constante. Al igual que las aves,

necesitarían eliminar rápidamente elexceso de calor. Debido a que los pterosaurios poseían pequeños orificiosen sus huesos huecos, probablementetambién tenían sacos pulmonares, y una bomba de los sacos aéreos. En 1837,Hermann von Meyer utilizó estasaberturas como prueba de que los pterosaurios eran voladores. Los huesosneumáticos de los pterosaurios parecehuesos avícolas llenos de aire, excepto por el hecho de que presentan unaenvoltura ósea aún más delgada. Losfinos huesos de las aves parecemacizos en comparación. Los huesos delos pterosaurios contenían sacos másgrandes con más aire. Quizá por esta

razón las aberturas, colocadas de formasimilar en las aves y en los pterosaurios,siempre eran más grandes en estosúltimos.
Seeley dedicó toda su vida alestudio de los pterosaurios mesozoicos, en 1901, resumió los hallazgos en el
libro Dragons of the Air . Aparte de unaobra especializada anterior suya, ésteera el primer libro dedicadoexclusivamente a estas extraordinarias bestias[13]. En éste, estaba pocodispuesto a admitir que los pterosaurioseran reptiles. El cerebro era como el delas aves, prueba del elevado nivel deinteligencia necesario para el vueloaleteante de un vertebrado. El sistema

respiratorio probablemente era similar al de las aves, demostrando una avidezde aire que revelaba una aceleraciómetabólica. Naturalmente, la conclusióde Seeley fue que los pterosaurios erade sangre caliente. De otra manera, esdifícil imaginárselos volando durante períodos más largos sin producir udemoledor déficit de oxígeno. Ulagarto aleteante, aún estando equipadocon la musculatura y coordinaciómental necesarias, habría caído prematuramente a tierra por agotamiento. Es especialmente poco probable que los pterosauriosmalgastaran la ventaja de tener unaventilación avícola, no modificando el

sistema circulatorio. El corazón de loslagartos, con tres cámaras, mezcla lasangre arterial rica en oxígeno con lasangre venosa pobre en oxígeno antes de bombearla al cuerpo. Los cocodrilos,que se originaron a partir de losancestrales arcosauriosaproximadamente al mismo tiempo quelos pterosaurios, tienen un corazón quefuncionalmente posee cuatro cámaras.Probablemente los dinosaurios, que seoriginaron a partir de un mismo tronco,también desarrollaron este tipo decorazón. Éste está presente en losdescendientes avícolas de losdinosaurios. Como observó Seeley: «Eltipo de corazón que siempre se asocia a

estructuras vitales, como las que se pensaba tenía el pterodáctilo, habiendosido éstas deducidas a partir del moldecerebral y de los orificios neumáticos elos huesos, es el corazón de cuatrocámaras de las aves y de losmamíferos[14]». El sistema circulatoriode los pterosaurios no presentaba ningúrasgo inconfundiblemente reptil,concluyó Seeley.
Sin duda, Seeley tenía razón. Unoseficaces pulmones avícolas proporcionaban el oxígeno y eliminabael calor y el dióxido de carbono residualnecesarios para los vuelos largos.Probablemente, un corazón de cuatrocámaras garantizaba el suministro

constante de sangre saturada de oxígenoa los tejidos. Un vuelo sostenido implicauna aceleración metabólica. Los pterosaurios eran de sangre calientecomo los mamíferos y las aves.
Debido a que los pterosaurios eraendotermos, y muchos de ellos era pequeños (el Ptenodracon sólo tenía eltamaño de un gorrión), debieron de estar protegidos por un abrigo de piel o plumas, o quizás incluso disponían dealgún tipo de aislamiento que no ha podido sobrevivir hasta nuestros días.Una criatura de sangre caliente deltamaño de un ave no habría resistidonunca la pérdida de calor, si no hubiesedispuesto de plumas. No existen aves

desnudas y tan sólo uno o dos ejemplosde pequeños mamíferos sin pelo(algunos animales que viven emadrigueras como los topos carececasi completamente de pelo pero estáaislados del frío extremo por susmadrigueras). La afirmación deGoldfuss de que se podían divisar mechones de pelo en los pterodáctilosfosilizados, nunca fue tomada muy eserio en Alemania. Esto no essorprendente, ya que Cuvier había dichoque la bestia era un reptil y los reptilesson escamosos por definición. Hasta ellúcido estudio realizado por Seeley erainconcebible, al pensar que un reptil
pudiera ser de sangre caliente.

Anteriormente, ya se habían hecho una odos sugerencias alternativas, pero eltema era abordado con extrema cautela ysin confianza alguna. Las ideas deGoldfuss impresionaron mucho alzoólogo inglés Edward Newman. Siembargo, suponiendo que fuera ciertoque los pterosaurios tuvieran pelo,
ewman también tendría que admitir que no podían haber sido reptiles.Cuando, en 1843, Newman escribió sartículo tenía conocimiento delcontrovertido maxilar de «opossum» deBuckland hallado en la pizarra jurásicade Stonesfield treinta años antes (dehecho fue el primer mamífero mesozoicoen ser descubierto). De forma que se

aventuró a sugerir que los pterodáctiloscontemporáneos también habían sidomarsupiales, sólo que murciélagosmarsupiales. Estando expuesto a la mofa pública, insistió en que los anatomistasno adoptaran una actitud reverencialhacia las autoridades: Cuvier yBuckland podrían estar equivocados.«Ahora creo que es posible —dijo
ewman cautelosamente— que Cuvier yBuckland no tuvieran razón. Confiesoque esto es muy improbable, perosostengo que es posible. Sea cual sea elconcepto que tengamos de ellos, existeel factor humano que nos induce a pensar que ellos también son capaces decometer errores[15]». La herejía de

ewman fue contestada con un silenciotorrencial, destino que acontecedemasiadas veces a las especulacionesiconoclásticas.
Sólo cuando Seeley comenzó a echar las bases para el cambio en la opiniórespecto a la condición del pterosaurio,la cuestión del aislamiento se hizocrucial. Si, como sugería Seeley, setrataba de una criatura de sangrecaliente, ¿no debería haber tenido algútipo de aislamiento? En 1908, KarlWanderer, de la Universidad de Munich,examinó los pterosaurios en el Museo deDresde para ver si él podía ver pelo.Goldfuss afirmaba que su fósil presentaba impresiones de piel con una

superficie verrugosa, lo cual interpretócomo prueba de la presencia de pelo o plumas en vida. Wanderer lo dudaba.Estos «hoyuelos» distribuidosirregularmente no eran debido a otracosa que a la naturaleza desmenuzablede la roca[16]. Más tarde, en 1927, unode los paleontólogos alemanes másrespetados, Ferdinand Broili, mostróenérgicamente su disconformidad coWanderer insistiendo en queefectivamente se trataba de impresionesde mechones de pelo y, parademostrarlo, comparó la piel verrugosade un murciélago con las impresiones dela piel de los pterosaurios. No cabeduda, exclamó, de que los pterosaurios

poseían un abrigo de piel[17]. Siembargo, el artículo cayó oportunamenteen el olvido y, si no fuera por algunosrecientes hallazgos espectaculares, probablemente se habría perdido por completo.
En 1970, A. G. Sharov, de laAcademia de Ciencias Soviética, estabarecolectando fósiles en los depósitos delagos del jurásico superior en Karatay,Kazajstán, una provincia del sur de la eURSS y descubrió un pequeño pterosaurio extraordinariamente bieconservado. Las rocas de esta regióque contienen los fósiles, al igual quelas de Solnhofen y Eichstätt, son degrano muy fino al haberse formado a

partir de los sedimentos de un antiguolago y, al igual que la piedra caliza deBaviera, registran las impresiones de la piel y otras partes blandas de losanimales que quedaron incrustados eellas. El pterosaurio ruso de Sharovestaba completo, mostrando lasimpresiones de las membranas de lasalas y un abrigo de piel. Todo el cuerpoestaba revestido de pelo largo y espeso;en vida, probablemente era blando ylanudo ya que los pelos y mechonesindividuales parecen estar curvados einclinados en el fósil. También había pelo en los dedos y las membranas entrelos dedos del pie, aunque aquí máscorto, e incluso se puede observar en la

misma membrana de un ala. Sharovllamó enigmáticamente su bestia Sordes
ilosus, que literalmente significa«abrigo de pieles sucio». Sin embargo,también empleó el equivalente ruso deSordes que, en esta lengua, tiene elsignificado alternativo de «demonio» o«espíritu maligno», por lo cual elnombre también podría ser traducido,más sugestivamente, como «demonio peludo[18]».
Este descubrimiento demostróinequívocamente que los pterosaurioseran de sangre caliente.Retrospectivamente, es fácil decir queno podría haber sido de otra forma. Siembargo, el proceso para llegar a esta

conclusión fue largo y penoso,abarcando casi dos siglos. Durante estetiempo, nuestra opinión sobre estaextraña bestia ha sufrido una radicaltransformación. Con la autoridad deCuvier, el pterodáctilo fue clasificadocomo reptil, lo cual hacía imposible laexistencia de pelo. (Oficialmente aún
sigue siendo clasificado como reptil etodos los textos corrientes). Después dela publicación de los estudios de Seeleya principios del siglo XIX, se fueaceptando cada vez más la idea que uvertebrado volador no podría haberseelevado nunca en los aires, a menos quedispusiera de una fisiología con umetabolismo rápido y con gra

producción de energía, al menosequivalente a la de un murciélago. Estetipo de fisiología se acompaña de unaelevada temperatura interna, que esmantenida estable para garantizar ufuncionamiento bioquímico óptimo. E base a estos antecedentes teóricos, elhallazgo de Sharov no debería haber causado gran sorpresa; ya estábamos prevenidos. Con el fin de evitar la pérdida de calor y el desperdicio deenergía, los pterosaurios habíadesarrollado pelo de forma paralela alos mamíferos. Al igual que losmamíferos, los pterosaurios probablemente pasaban muchas horascada día limpiándose el pelo. En los

pterosaurios primitivos, los dientes y lasgarras posiblemente habían servido de peines. Cuando en los tipos quesurgieron posteriormente se perdierolos dientes, con el fin de perder peso, ylas garras ya no llegaron al tronco, posiblemente el pico asumió estafunción.
Debido a que los pterosaurios era bastante inteligentes y de sangrecaliente, ¿cuál era su forma dereproducción? ¿Ponían huevos o daban aluz a crías vivas? Y lo que es aún másimportante, ¿existían cuidados posnatales? Incluso los cocodrilos de pequeño cerebro son padres dedicadosque desentierran a sus crías que salen de

los huevos para transportarlas hasta elagua (en sus maxilares). Incluso irán esu ayuda si las oyen chillar cuando estáen manos de algún científico incauto.Apenas cabe menos esperar de la formade atención paterna de los lejanos primos voladores de los cocodrilos.
Debido a que los pterosaurioshabían desarrollado el cuerpo volador más ligero de todos los vertebrados, yse habían tomado mucho trabajo parareducir el peso a gran escala, esimprobable que las hembras pudierahaber volado llevando a sus críascuando éstas hubieran alcanzado unafase muy avanzada de su desarrollo. Silos pterosaurios eran vivíparos, las

crías probablemente nacían en estadoinmaduro, conclusión que también essugerida por la pelvis muy pequeña dela madre. O, quizá, ponían huevos aligual que las aves. De cualquier forma,los pterosaurios que salían de sushuevos, debido a que eran criaturasinmaduras y de sangre caliente, debierode depender de sus padres para que lesdieran calor, alimentos y más tarde probablemente un entrenamiento devuelo. Las hembras, al tener un abrigode piel y ser una fuente de calor, posiblemente empollaban los huevoshasta que salían las crías (el único caso paralelo en el mundo reptil es el de lahembra pitón de la India, que se enrolla

alrededor de sus huevos y se calienta así misma enviando espasmos decontracción muscular a lo largo de scuerpo). Los pterosaurios probablemente permanecían junto a suscrías hasta que éstas podían volar [19].Esto podría significar que los pterosaurios se apareaban, al igual quelas aves, durante una época del año —oincluso de por vida—, de forma que elmacho pudiera suministrarle a lahembra, que estaría empollando loshuevos, los alimentos almacenados en s bolsa. Los pterosaurios habíaascendido a un nuevo nivel social,caracterizado por la unidad familiar temporal. Puede incluso que hubiera

existido una organización colonial másavanzada, en la cual los vulnerables polluelos —especialmente delgigantesco Pteranodon —, formaríamanadas y estarían protegidos por uno odos adultos. (En la arenisca verde deCambridge, en Inglaterra, se haencontrado miles de fragmentos de pterosaurios más pequeños, sugiriendoque posiblemente habían sido gregarios, pasando la noche juntos en coloniascomo los murciélagos. Sin embargo, eeste caso es más probable que estoshuesos fueran arrastrados por el aguaquedando amontonados mucho tiempodespués de haberse producido la muertede los individuos vastamente

distribuidos[20]).Seeley concluyó su clásico informe
sobre los dragones voladores,«ornitosaurios» como él los llamaba,con una reafirmación de su teoría de queeran algo más que humildes reptiles:«estos fósiles han demostrado que losornitosaurios tienen órganos vitales blandos en común con los dinosaurios ylas aves», escribió y que habíadesarrollado «formas especiales deórganos respiratorios y cerebros que losdistinguían de los reptilesexistentes[21]». Estas ideas fueroformuladas a principios de este siglo ySeeley, que murió sólo ocho años mástarde, no llegaría a saber nunca que sus

ornitosaurios eran peludos, demostrandoasí fuera de toda duda que eran, talcomo él siempre había pensado, desangre caliente. Su nivel de organizacióera sumamente alto y, aunque se habíaoriginado a partir del mismo troncoancestral que los dinosaurios, habíaevolucionado más que sus primos. Losdinosaurios también eran de sangrecaliente, es cierto, pero los pterosaurioshabían desarrollado el pelo como losmamíferos y un cerebro como las aves,de forma que, aun no siendo endotermosmás eficientes, con toda seguridad eramás inteligentes. Habían superado la barrera térmica y evolucionado a partir de la clase reptil en la misma medida

que las aves o los mamíferos. Bakker yGalton afirmaron, cuando establecierola clase dinosaurios, que entre losanimales vertebrados superiores «laadaptación más importante fue el pasode la endotermia[22]». Esto habíaocurrido en los reptiles semejantes a losmamíferos y en los antepasados de losdinosaurios, dando lugar a nuevosniveles de organización y provocando profundos cambios en los tejidos blandos, que permitieron la produccióconstante de energía y una mayor actividad. Bakker y Galton continuaron:«Probablemente los antepasadosdinosaurios de las aves ya eraendotérmicos y tenían un metabolismo

de alto rendimiento, característicasclave que diferencian a los dinosauriosde los cocodrilos y de otros arcosauriosextinguidos». Esto no es del todoverdad, tal como ya se ha comentadoanteriormente, ya que los mismoscriterios que le permiten a Bakker declarar a los dinosaurios endotermos, probablemente también son atribuibles alos antepasados seudosuquianos de losdinosaurios, en el caso de que fueran desangre caliente. Finalmente, estadiferencia podría ser ahora ampliada para incluir a los pterosaurios. Laendotermia posiblemente diferencie alos dinosaurios de los cocodrilos, y seauna justificación para mantener a estos

últimos como reptiles y elevar a los primeros a una nueva clase. Siembargo, los pterosaurios tambiéhabían evolucionado. Se habían vueltoendotermos y, además, tan eficientescomo las aves. Parece bastante absurdodejarlos en la clase de los reptiles y, almismo tiempo, elevar de clase a losdinosaurios. Seeley no lo hubieseaprobado.
Si los pterosaurios aún viviesen,inmediatamente reconoceríamos que noeran reptiles. Aristóteles —el grarecolector y clasificador de laantigüedad— nunca habría incluido a uanimal de sangre caliente, peludo einteligente, en el grupo de las serpientes.

Esta tradición, de no proceder de estaforma, se habría transmitido hastanuestro siglo XX, tal como ocurre colas aves y los mamíferos.Desafortunadamente, los pterosaurios seextinguieron a finales del cretáceo ysólo a partir de los huesos fósiles pudieron ser revividos en la mente delos hombres; huesos, además, queinicialmente proporcionaron pocas pistas en relación con la fisiología ointeligencia. Estas características hasido determinadas laboriosamente a lolargo de varias décadas, de forma queahora nos podemos hacer una image bastante clara del pterosaurio en vidaque se parece muy poco a los reptiles.

Reconocemos que, aunque losmamíferos se originaron a partir de losreptiles, se trata de criaturas distintas.Asimismo, a los pterosaurios, cuyafisiología tan distinta a la de los reptileses la prueba de que hacía ya muchotiempo que se habían distanciado de losreptiles, se les debería otorgar lacondición de clase en reconocimiento asu elevado nivel de organización.
Ahora disponemos de todas las pruebas y podemos especular acerca delorigen y de las subsiguientesradicaciones de los endotermos nomamíferos del planeta mesozoico y, almismo tiempo, sugerir por quédeterminados grupos fueron capaces de

«explotar» cuando lo hicieron. En
resumen, podemos exponer una teoría
unificada que relaciona estas
radiaciones de los vertebrados
mesozoicos con los logros fisiológicos
subyacentes. Probablemente, losseudosuquianos lograron por primeravez una fisiología de sangre caliente a principios del triásico —hace 210millones de años— y evolucionarohacia los paradinosaurios, algunos decuyos miembros más tardíos (por ejemplo, el Ornithosuchus) llegaron atener un gran parecido con losdinosaurios carnívoros bípedos. Losdinosaurios ornitisquianos ysaurisquianos apareciero

aproximadamente a mediados deltriásico, explotando al máximo slegado fisiológico y posturalseudosuquiano y asumiendo rápidamenteuna posición dominante en el orden delmundo mesozoico. Sin embargo, otrosseudosuquianos bípedos del triásicomedio, con brazos libres de la carga desu función de soporte y locomoción,empezaron a planear y desarrollaron unamembrana de piel que se extendía entrelos brazos y el tronco. Existen pruebasde la existencia de esta fase intermedia,el seudosuquiano planeador Podopteryx
(literalmente «pie alado»), encontrado por Sharov en sedimentos del triásicosuperior en Rusia[23]. (El Podopteryx

vivió en una época demasiado tardíacomo para haber sido el verdaderoantepasado de los pterosaurios). Losseudosuquianos estaban «preadaptados» para volar si, en efecto, ya poseían unafisiología de metabolización rápida. Deeste modo, pudieron elevarse en losaires como criaturas que batían las alas, a principios del triásico tardío,
aparecieron los primeros pterosauriosen los cielos[24]. Es bastante probableque hubiera un único origen de laendotermia entre los «saurios» delmesozoico que se remontaba a lostecodontes de principios del triásico, y
que esto desencadenó dos explosiones
rincipales, la de los dinosaurios y la

de los pterosaurios. Estos descendientesendotermos seudosuquianos se dierocuenta del potencial inherente a slegado fisiológico y se convirtieron eunos de los vertebrados terrestres quemás éxito han tenido en la historia.(Véase el árbol genealógico de losvertebrados superiores en el apéndice I).La sustitución de un origen único por dos o incluso tres orígenesindependientes de la endotermia,simplifica en gran manera nuestra visióde la historia de los vertebrados ademásde satisfacer la exigencia científica desencillez. El resultado es una síntesisestéticamente grata que satisfaceampliamente, por su amplia envergadura

gran poder aclaratorio (que, al fin y alcabo, es el objetivo final de laexplicación científica).
A pesar de que los pterosaurios más pequeños eran aleteadores, a finales delmesozoico se desarrollaron extraños planeadores gigantescos, entre ellos losanimales voladores más grandes que haexistido en el mundo. Anteriormente a1840, ninguna especie conocidasuperaba, en tamaño, a un gramurciélago de la fruta. Esto no eraextraño, dijo Richard Owen en aquellaépoca, ya que «a priori erafisiológicamente improbable que la

organización de sangre fría de un reptil pudiera ser capaz de elevar en los airesuna masa más grande de la que podíalevantar los mamíferos de sangrecaliente[25]». En 1845, el secretario dela Sociedad Paleontográfica, JamesScott Bowerbank, describió un nuevoespécimen de la creta de Kent que presentaba una envergadura de, almenos, 2,44 o 2,74 metros. En lugar demodificar su teoría de que los pterodáctilos eran de sangre fría, Oweexclamó que Dios una vez más estabademostrando su omnipotencia al haber logrado lo que parecía imposible. Esteespécimen, dijo Owen, «demuestra quelas manifestaciones de la fuerza del

Creador en tiempos pasados sobrepasacualquier cálculo basado en lanaturaleza real[26]». Posteriormente, e1847, Bowerbank excavó u pterodáctilo que no superaba al otro elongitud, pero que tenía casi el doble detamaño, con una envergadura quealcanzaba los 4,5 o 4,8 metros de puntaa punta y al que llamó Pterodactylus
iganteus. Fue una criatura de estetamaño la que Owen mandó reconstruir a escala natural en el Crystal Palace eSyndenham.
Después de la exploración delOeste, la atención se desvió hacia el
uevo Mundo. En el otoño de 1870,O. C. Marsh (acompañado de Col.

W. F. Cody, más conocido como«Buffalo Bill») dirigió una expediciódel Yale College en la región de lasMontañas Rocosas en busca de nuevosfósiles y, como siempre, al eminente paleontólogo le aguardaba una gra bonanza. El equipo de Marsh estableciósu campamento en la bifurcación nortedel río Smokey, al oeste de Kansas, ycomenzó a explorar los depósitos decreta del cretáceo superior.
Kansas era el territorio de Cope y lacreta había proporcionado una ricavariedad de espectaculares criaturas alrecién descubierto rival de Marsh, queincluían muchos mosasaurios y el plesiosaurio marino de cuello largo,

lasmosaurus. En realidad, Cope nohabía visitado Kansas. Los fósiles, dehecho, le habían sido enviados aFiladelfia donde los describió. El
lasmosaurus fue enviado en 1868 y elmomento de su llegada a Filadelfiacoincidió con el del experto inglés ereconstrucciones, Benjamin WaterhouseHawkins, que estaba buscando reptiles prehistóricos para su Museo Paleozoico.De esta forma, ocurrió que, apenas Copehabía descrito el animal, Hawkins yahabía construido un modelo de tamañonatural en el centro de Nueva York. Unavez desencadenada la guerra entreMarsh y Cope (provocada por Cope queno había sido capaz de distinguir entre

la cabeza y la cola del Elasmosaurus),comenzó la apasionada carrera en buscade nuevos descubrimientos. Marsh cruzólas Montañas Rocosas por primera vezen 1868 y, pensando que la recoleccióde huesos en Kansas era buena, volviócon el equipo de Yale en 1870. Marsinmediatamente desenterró tres nuevasespecies de mosasaurios, uno de loscuales alcanzaba una longitud de 9metros. Durante las excavaciones, setropezó con algunos huesos problemáticos; éstos eran largos,delgados y, a primera vista, se parecíaa los huesos de las patas de las aveszancudas. Al igual que los huesosavícolas eran ligeros, tenían paredes

delgadas y eran neumáticos. Los restosde los huesos obviamente procedían dedos especies distintas. Marsh estaba preocupado porque las articulacioneseran completamente diferentes a las delas aves y, a pesar de su gran tamaño,sólo podían ser comparadas con laarticulación de los dedos de las alas delos pterosaurios ingleses. Sin embargo,el dedo era desproporcionado; a pesar de que Marsh sólo tenía un diminutofragmento de un metacarpiano, éstemedía 17 centímetros de longitud. «¡Estoindicaría —dijo Marsh proféticamente — una envergadura de las alas de nomenos de 6 metros de largo[27]!». Latemporada de recolección estaba

llegando a su fin, de forma que elentusiasmo tuvo que ser contenido.Marsh se vio obligado a esperar hasta elverano siguiente para poder confirmar sus especulaciones. En el veranosiguiente, 1871, el talento de Marsh paradescubrir fósiles resultó en un hallazgoque incluso eclipsó parcialmente a susreptiles alados. Cuando volvió a la cretade la región del río Smokey, en Kansas,descubrió el primer esqueleto de un aveno voladora del cretáceo, el
esperomis, o «ave del Oeste», en loslechos que contenían los fósiles de pterosaurios[28]. (Los simpatizantes deCope insisten en que no fue Marsh sinoBenjamin Mudge quien, en realidad,

encontró los restos del Hesperomis yadvirtió su importancia. Mudge estaba a punto de enviárselos a Cope cuandoapareció en escena Marsh y convenció aMudge para que se los entregara). Losenormes pterosaurios obviamente habíavivido junto con aves de aspecto bastante moderno (moderno ecomparación con el arcaico
rchaeopteryx a pesar de que elesperomis, al igual que los
dinosaurios-ave, tenía dientes). La profecía de Marsh acerca del gigantescotamaño de los reptiles voladores seconfirmó dramáticamente en este viaje.Encontró nuevos especímenes con losque pudo sumar la longitud de los

diversos huesos aislados de alas. Elmetacarpiano de la mano, del cuál yahabía encontrado un pequeño fragmento,medía 40 centímetros mientras que el primero de los cuatro huesos del dedoalargado del ala medía 43 centímetros.El brazo era corto y medía 18centímetros. Esto hacía un total de másde 1,20 metros sin contar los tres huesosde los dedos de las alas que faltaban.«Las mediciones reseñadasanteriormente de los huesos de las alasindicarían una longitud total de las alasde, al menos, 2,6 metros y unaenvergadura de ambas alas de 5,5 a 6,1metros. Por lo tanto, la presente especieincluye a algunos de los “dragones

voladores” más grandes jamásdescubiertos[29]». Aun hallándose en los primeros días de su carrera en busca defósiles, Marsh difícilmente podría haber encontrado algo más espectacular paraaventajar a su rival. En una posicióalgo más elevada del mismo yacimiento,desenterró un pterosaurio aún másgrande cuyos huesos de las alas eran lamitad más anchas que las del espécimeanterior, que ya había sido bautizado coel nombre de Pterodactilus
occidentalis. Marsh, al que obviamentese le acabaron los superlativos, veía alnuevo espécimen como «uno de los pterosaurios más gigantescos. ¡Tenía almenos el doble de volumen que el

terodactylus occidentalis y probablemente la distancia, medida de punta a punta de las alas completamenteextendidas, sería de aproximadamente6,7 metros!».
Sin duda fue un gran triunfo personal para Marsh. El hallazgo de los primerosPterosaurios americanos le habíacorrespondido a él, como era justo.Cope, entretanto, no sería dejado fuera.Cuando, al final de la segundatemporada de excavaciones, Marsabandonó Kansas, Cope inmediatamentese dirigió a los mismos lugares ycomenzó a excavar frenéticamente a fide conseguir su propia colección devaliosos fósiles de la creta. (Cuando,

una década más tarde, el Popular
Science Monthly publicó un artículosobre la vida de Cope, éste adelantó lafecha de la época de sus exploracionesal Oeste a 1870 para que coincidieracon la de Marsh). Tan pronto como,hubo marchado Marsh, Cope trabajó corapidez en un intento de extraer losfósiles y asignarles nombres antes deque lo pudiera hacer Marsh. La prioridad en la asignación del nombrede las bestias era de suma importancia para Cope. El 1 de marzo de 1872 leyóun informe ante la Sociedad FilosóficaAmericana, describiendo sus hallazgosde pterosaurios. Llamó a los dos sauriosgigantes alados Ornithochirus o «pies

de ave», en oposición al Pterodactylus
de Marsh. Pero llegó demasiado tarde.Tal como señaló con gran placer el portavoz de Marsh —el American
ournal of Science publicado en laciudad natal de Yale, New Haven—, elartículo de Cope fue distribuido eforma impresa el 12 de marzo mientrasque la contribución de Marsh habíaaparecido cinco días antes, el 7 demarzo. Marsh había conservado la prioridad por un pelo.
La creta de Kansas resultó ser uverdadero cementerio de estos enormessaurios voladores y Marsh, seguido mástarde por Willinston, excavaron docenasde especímenes fragmentarios durante

las siguientes décadas. Marsh rebautizóa su gigantesca especie con el nombrede Pteranodon, que literalmentesignifica «alado y sin dientes». Setrataba de un pterosaurio de aspectoextraordinario, con mandíbulas queformaban un largo pico sin dientes y quemantenía el equilibrio con la ayuda deuna cresta ósea alargada que selevantaba en la parte trasera del cráneo.La criatura era literalmente toda alasque, probablemente, superaban los 7metros de envergadura mientras que eltamaño de su diminuto cuerpo nosuperaba el de un pavo. Ucontemporáneo más pequeño de Kansas,el Nyctosaurus, o «lagarto de la noche»,

ilustra la misma gran desproporción coun cuerpo de 15 centímetros de longitud alas de 2,4 metros de envergadura.
s. El esqueleto del pterosaurio Pteranodon
de alta mar. Estaba construido como un planeador artificial. El fuselaje en forma decaja, reforzado, soportaba la tensión de alasde 3,6 metros de longitud. El hueso

pteroideo de la muñeca probablementesostenía una membrana elevadora que llegabaal cuello.
Las largas extremidades traserascolgantes del Pteranodon eradefinitivamente inadecuadas para elmovimiento terrestre normal y es difícilimaginarse a estas torpes criaturascaminar para el suelo. A diferencia delos murciélagos, que pueden doblar susalas a lo largo de su cuerpo (e inclusoasí tienen dificultades para caminar sobre cuatro patas), el cuerpo del
teranodon no podría de ningunamanera haber sido envuelto por suslargas alas. Debido a que los dedos de

las alas sólo podían ser flexionadas elos nudillos, en el punto medio de lasalas, los dedos de 1,8 metros de longitud probablemente estarían dirigidos haciaarriba cuando el animal avanzaba arastras. Cualquier forma de desgarbadalocomoción habría resultadoextremadamente difícil. Los primeros pterosaurios tenían una articulacióentre el hombro y el ala consistente ecóndilos y carillas articulares. Siembargo, las especies que aparecieromás tarde presentaban una charnela quesólo permitía movimientos ascendentes descendentes[30]. «Quizás el método de
desplazamiento en tierra más factible para ellos —dijeron Hankin y Watson e

1914— era que, una vez se había posado sobre tierra, cayeran sobre susestómagos y se fueran arrastrando por elsuelo, como hacen los pingüinos,dándose impulso con sus patas traseras ysalvando los obstáculos con uocasional alzamiento de las alas[31]».
aturalmente, en algún momento debíade posarse sobre tierra y, por lo tanto,debían de ser capaces de desplazarse por tierra aunque resulta difícilimaginárselos caminar a modo de pingüinos aunque sólo sea porque poseían unos enormes picos. Quizásempleaban los dedos de las alas paralevantar el cuerpo del suelo. Con unalocomoción en tierra tan torpe, el

teranodon habría sido una fácilvíctima para cualquier dinosauriocarnívoro que estuviera cerca. Siembargo, parece más probable que los pterosaurios más grandes volaran casiconstantemente, al igual que lo hacen losalbatros, y quizá se posaban sobre tierrasólo una vez al año para reproducirse.Las patas traseras, extraordinariamente parecidas a las de los murciélagos,sugieren que el Pteranodon se podíacolgar con la cabeza hacia abajo comohacen los murciélagos cuando se hallabaen tierra. Debido a que era demasiadogrande como para haber sido utrepador de árboles, se pensaba quedescansaba en salientes de las rocas. Si

embargo, probablemente cerca de loslagos del interior de Kansas no habíaacantilados, por lo cual resulta difícilaceptar la idea popular de que los
teranodon habían sido habitantes de lacosta donde podrían estar seguros[32].
Al igual que los albatros, elteranodon probablemente pasaba gra
parte de su vida adulta en el aire. Para poder hacer esto, y sin embargo crecer tanto, era necesaria una reducció bastante drástica del peso.Generalmente, no se apreciasuficientemente lo ligero que era erealidad este planeador con alas de 7metros de envergadura. Debido a esto,se han imaginado todo tipo de ideas

fantásticas para explicar cómo elteranodon permanecía en vuelo. Se ha
supuesto, por ejemplo, que la atmósferamoderna era demasiado ligera para estascriaturas y que, por consiguiente, en elcretáceo la atmósfera debió de haber sido el doble de densa, haciendo posibleque el Pteranodon literalmente flotaraen el aire[33]. El Pteranodon en realidadno necesitaba esta ayuda atmosférica. Evida el animal pesaba unos 18 kg(algunas estimaciones son de 9 kg), u25% aproximadamente del peso de uhombre. El Pteranodon había perdidosus dientes, cola y parte de lamusculatura de vuelo y sus patastraseras se habían vuelto largas y

delgadas. Fue, sin embargo, en loshuesos donde se logró la mayor reducción de peso. Los huesos de lasalas, columna vertebral y extremidadestraseras eran tubulares, como los puntales de soporte de los aviones, que proporcionan resistencia perodisminuyen el peso. En el Pteranodon
estos huesos, de hasta 2,5 centímetros dediámetro, no eran otra cosa que espaciosaéreos cilindricos limitados por unaenvoltura ósea externa no más gruesaque una carta. Barnum Brown, delMuseo Americano, informó sobre ufragmento del hueso de un brazo de unaespecie desconocida de pterosaurio delCretáceo Superior de Texas, en la cual

se había alcanzado «la culminación del pterosaurio… la cima de la construccióligera[34]». ¡Aquí, la tendencia habíallegado hasta tal punto que la pared óseadel cilindro tenía el increíble grosor de0,50 centímetros! En el interior del tubohabía puntales óseos al través, no másgruesos que alfileres, que ayudaban areforzar la estructura; otra «innovación»en el diseño de aviones anticipado por los pterosaurios del mesozoico.
La combinación de un gran tamaño y peso insignificante necesariamente tuvoque provocar cierta fragilidad. Es fácilimaginarse que estos huesos tubulares,delgados como una hoja de papel, quesostenían alas gigantes, posiblemente

hicieron difícil el aterrizaje. ¿Cómo podían estas criaturas aterrizar siromperse todos los huesos? En primer lugar, ¿cómo podían elevarse en losaires? Evidentemente no eran capacesde batir estas alas de 3,6 metros delongitud ensartadas entre tubos delgadoscomo pajitas. Las aves más grandestienen que alcanzar una cierta velocidadcorriendo y batiendo las alas para poder elevarse en el aire, y otras tienen que producir una velocidad de aleteo quecasi les permita permanecer flotandoantes de poder elevarse en el aire. Para poder permanecer flotando en el airecon una envergadura de alas de 7metros, el Pteranodon habría necesitado

99 kg de músculos de vuelo taeficientes como los de los colibríes. Siembargo, el Pteranodon había reducidosu musculatura a unos 3,6 kg, de formaque es inconcebible que pudieraelevarse activamente en el aire[35].
Por lo tanto, el Pteranodon no erauna criatura que batía las alas ya que nodisponía ni de los músculos ni de laresistencia a la fuerza resultante. Susalas largas, delgadas como las de losalbatros lo revelan como planeador, el planeador más avanzado jamás producido por el reino animal. Con u peso de sólo 18 kg, la carga de las alasera de sólo 0,45 kg por 929 cm2. Esto le proporcionaba una velocidad de

descenso incluso inferior a la de los planeadores fabricados por el hombre,cuyas alas deben soportar un peso de, almenos, 1,8 kg por 929 cm2[36]. Larelación entre el área de las alas y el peso total en el Pteranodon sólo essuperada en algunos insectos. El
teranodon estaba construido como u planeador con el esternón, articulaciódel hombro y columna vertebralsoldados formando un fuselaje rígidocomo una caja, capaz de absorber latensión de las gigantescas alas. El bajo peso, combinado con una enormeenvergadura de alas, significaban que el
teranodon podía planear avelocidades extremadamente bajas si

que hubiera peligro de que se parase.Cherrie Bramwell, de la Universidad deReading, ha calculado que podía permanecer en el aire con una velocidadde sólo 24 km por hora. De este modo,le habría resultado relativamente fácilelevarse en el aire. Lo único quenecesitaba el Pteranodon era una brisade 24 km por hora para que se pusieracara al viento, extendiera sus alas y seelevara como un trozo de papel. No eranecesario ningún esfuerzo. Si la criaturase hubiese visto obligada a aterrizar sobre el mar, sólo habría tenido queextender sus alas para atrapar el aire yasí emerger suavemente del agua.
Parece extraño que un animal, que se

había extremado tanto en reducir su pesoa un mínimo, desarrollara una crestaósea, alargada en su cráneo. El hecho deque la cresta estuviera ausente en otros pterosaurios sugiere que no eraindispensable para el animal. Se hasugerido que la cresta era principalmente ornamental[37] perosencillamente es absurdo imaginarse uornamento tan engorroso en una criaturavoladora tan especializada. Ucomentario editorial en el The
eronautical Journal de 1914, escrito por un científico que no era biólogo, pero que evidentemente estabafamiliarizado con la acción de laevolución y economía del diseño de los

aeroplanos, puso en ridículo esta idea.«Es obvio que un animal volador que hareducido su peso al máximo,difícilmente habría desarrollado unalarga cresta a costa de, o sin efectosobre, su capacidad para volar. Es decir,la cresta debió de tener alguna ventajaaerodinámica para su propietario paraque pudiera evolucionar en la medidaque lo hizo. No es difícil ver que estetipo de cresta pudo ser muy útil desde el punto de vista de la estabilidad.
A todos aquellos que estéfamiliarizados con la extremaimportancia que posee la disposición delas aletas para la estabilidad de losaeroplanos, no será necesario

explicarles que, cuando el Pteranodon
dirigía su pico hacia abajo, movía unagran área de aleta (el mismo pico) haciaatrás y abajo[38]». Sin embargo, cuandoel pico bajaba, concomitantemente seelevaba la cresta para compensar elefecto: el Pteranodon estaba equipadocon un mecanismo automático decontracción. Más tarde, se demostró quela cresta, en realidad, ahorraba peso.Los músculos adicionales del cuellonecesarios para mantener el pico rectocara al viento, habrían excedido emucho el peso de la cresta[39]. Esto hasido demostrado mediante experimentoscon tanques de descarga y túneles deviento con modelos a escala reducida de

cabezas de Pteranodon con cresta y sicresta. Estos experimentos tambiérevelan otra función de la cresta.Cuando se eliminaba la cresta, la cabezaera desviada constantemente hacia ulado por la fuerza de la corriente y, evida, los músculos del cuello habríatenido que actuar continuamente paramantener la cabeza dirigida haciadelante. Cuando se volvía a colocar lacresta en su sitio, se producía unacorrección automática si la cabeza sedesviaba oblicuamente a la direcciódel flujo de aire. En otras palabras, el pico y la cresta actuaban conjuntamentecomo una veleta estando dirigidossiempre en dirección al viento. La cresta

probablemente también funcionaba comoun timón en un planeador para evitar movimientos de derrape lateralesdurante los giros[40].
El vuelo de los pterosaurios siempreha llamado la atención de los ingenierosaeronáuticos. En 1914, el año de lamuerte de Wilbur Wright, se publicó uinforme de un equipo de trabajo muycurioso compuesto por D. M. S. Watson,el especialista en fósiles de reptiles delUniversity College de Londres yE. H. Hankin, un experto en aeronáutica.En éste analizaban la complicada seriede articulaciones del hombro, codo,muñecas y nudillos en el Ornithodesmus
del cretáceo inferior, con una

envergadura de 4,5 metros. En vista dela delicada naturaleza de los huesos, yde la ausencia de extensos músculos devuelo en estos pterosaurios gigantes,Watson y Hankin llegaron a laconclusión de que «no volaban peor queotras aves sino que volaban de formamás científica[41]». La complicada seriede articulaciones les proporcionaba uincremento de la maniobrabilidad y, posiblemente, les permitía frenar suavemente mientras se elevaban haciael cielo.
Recientemente, se ha atribuido unafunción similar al enigmático hueso pteroideo, el pequeño sobrehueso en eldedo del ala para el cual no se había

encontrado ninguna función. CherrieBramwell sugiere que sostenía unamembrana que se extendía hasta elcuello. Cuando la criatura se eleva en elaire, el hueso podía ser levantado y lamembrana actuaba como un elevador,como en los aviones modernos. Cuandoel animal quería posarse sobre tierra, elhueso era descendido para reducir lavelocidad a un mínimo sin crear turbulencias y paradas del vuelo. El
teranodon podría así aterrizar si peligro de lesionarse sus frágileshuesos.
Cuando Cherrie Bramwell introdujolos datos del Pteranodon en elordenador de la Escuela de Aeronáutica,

se descubrió hasta qué punto estos planeadores naturales estabaavanzados. El ordenador estaba programado para examinar elcomportamiento de vuelo de los planeadores construidos por el hombre yno necesitó ningún ajuste para el
teranodon[42]. Parece ser que covelocidades superiores a 18 nudos(aproximadamente 32 km por hora), el
teranodon podía girar sus alas haciaatrás en forma de V (a diferencia de losmurciélagos podía hacerlo debido a quesus alas estaban sostenidas por una largaserie de huesos que podían ser flexionados en los nudillos). Esto provocaba una disminución de la

velocidad de descenso y es el principioempleado por los aviones a reacciócon alas de geometría variable: u principio descubierto de nuevo, despuésde haber estado dormido durante 70millones de años.
Los sedimentos marinos quecontienen esqueletos de Pteranodon
también albergan una gran diversidad dereptiles marinos, especialmente plesiosaurios y mosasaurios. Algunos delos depósitos que contenían pterosauriosse formaron a cientos de kilómetros dedistancia de la costa del mar máscercana, que invadió Tejas y Kansas ela era cretácea. Esto sugiere que el
teranodon podía recorrer grandes

distancias lejos de tierra como losalbatros[43]. Evidentemente, tuvo unaexistencia idílica en el aire dependiendosólo de los peces que recogía. Al igualque los pelícanos modernos, poseía una bolsa en el cuello (que en ocasiones puede ser observada en los fósiles coluz ultravioleta), en la cual guardaba los peces. Un espécimen en el MuseoAmericano tiene los esqueletos de dosespecies de peces que aún estaban en s bolsa, dando fe de sus gustos dietéticos.
El modo cómo recogía los peces nos plantea algunos problemas. Existealgunos murciélagos que también sealimentan de peces. Al igual que losPterosaurios, poseen enormes patas

traseras con garras que cuelgan cuandoestán volando. El murciélago alano(Noctilio leporinus), de cuyos hábitosconocemos algo, vuela por encima delagua al atardecer y desciende para coger los peces con sus garras. Éste probablemente también es el métodoempleado por los otros murciélagos pescadores aunque sus costumbres so poco conocidas. Existe un murciélago pescador en California ( Pizonyx vivesi),que durante el día vive junto a petrelesen grietas rocosas, del cual, ya antes dehaber sido observado durante la noche,se sospechaba que era un pescador debido a sus largas garras traseras. Estofue confirmado posteriormente por u

análisis del contenido de su estómagoque reveló que sólo contenía peces. Siembargo, ¿era posible que el
teranodon dejara colgar sus garras eel mar para coger peces? Si éste era elmétodo, y vivía a cientos de kilómetrosde tierra, ¿cómo llegaban los peceshasta su boca? Es bastante más probableque el Pteranodon planeara por encimade las crestas de las olas atrapando los peces que vivían en la superficie delagua con su pico extremadamente largo,montado a un cuello móvil. Quizá se posaba sobre la superficie para coger los peces. Últimamente, se ha sugeridoque el dedo del ala podía girar produciendo con el ala un plano e

forma de M que iniciaría la inmersióinmediata[44]. Si esto fuera posible,debió de haber sido desde una altura bastante baja a fin de reducir el impactode los extremadamente ligeros huesoscon el agua. Una generación anterior,quizá más influenciada por losaeroplanos como instrumentos dedestrucción, vio el Pteranodon desdeotro punto de vista. Barnum Brown serefería a los pterosaurios como «losaeroplanos de la época prehistórica».En un artículo publicado en octubre de1943, Brown veía un paralelismo entrelas aterradoras técnicas aéreascontemporáneas y el estilo de vida delos primitivos pterosaurios. El

«bombardero de inmersión».teranodon, como lo llamaba Brown,
era la máquina voladora másdesarrollada de su época. La cresta era«una especie de timón que le era útil alanimal cuando, con las alas dobladas,cruzaba el aire como un rayo en buscade presas… El cuello eramoderadamente largo, fuerte y flexible.Presentaba una notable serie dearticulaciones adicionales nunca vistasen los huesos del cuello de otrosanimales, que proporcionaban una grafuerza al pico para asestar golpes[45]».Sin embargo, Barnum Brown había sidoinfluido excesivamente por los sucesossiniestros de su época. El incremento de

la sofisticación de la guerra aérea lehabía llevado a investir al Pteranodon
de una agresividad que en vida noexistía. Es poco probable que el frágil
teranodon pudiera haber resistido elchoque de precipitarse al agua desdecualquier altura. A diferencia de lasgaviotas, que pueden recoger sus alas, el
teranodon no tenía cóndilos y carillasarticulares en la articulación delhombro, de forma que no podía recoger sus enormes alas y, al estar éstasextendidas, no podrían haber resistido elimpacto. ¡Tal como sugirió el mismoWatson, en 1914, en una época en la cualla guerra era un poco menos sofisticada, cuando los aviadores sabían lo que

significaba la fragilidad en el aire, probablemente el enorme pterosaurio sehabría desintegrado al intentar enderezarse después de unainmersión[46]! En lugar de ser eldiabólico bombardero buceador deBarnum Brown, el Pteranodon era unacriatura grácil, muy inteligente, que sededicaba a volar plácidamente sobre losgrandes mares del cretáceo.
El Pteranodon se pasaba los días planeando sobre el océano en busca de peces que vivían en la superficie.Tradicionalmente, se pensaba que estosPterosaurios gigantes eran negros coalas curtidas como las de losmurciélagos. Desde que sabemos que el

«demonio peludo» de Sharov, Sordes
ilosus, tenía un grueso abrigo de piel, yque las membranas de sus alas tambiéestaban cubiertas de pelo parece más probable —aunque a primera vistaresulte sorprendente— que el
teranodon estaba cubierto de peloblanco. Esta piel le habría proporcionado a la criatura un parecidoaún mayor con el albatros. Las gaviotasson blancas, lo cual constituye una formade camuflaje; al presentar un perfilmenos evidente para los peces, evitaahuyentar al potencial alimento. Estohabría constituido un problema aúmayor para el pescador Pteranodon.Sus enormes alas habrían tapado el sol a

medida que sobrevolara el agua y unasalas negras habrían agravado seriamentela situación.
Decir que, como criatura voladora,el Pteranodon era grande sería unadescripción insuficiente. De hecho,existen razones fundadas para creer quese trataba del animal más grande queamás pudo elevarse en los aires. Co
cada incremento del tamaño y, por lotanto, también del peso, un animalvolador necesita un incrementoconcomitante de fuerza (para batir lasalas en los animales aleteadores y parasostenerlas y manipularlas en los planeadores). Sin embargo, la fuerza essuministrada por músculos que a su vez

añaden aún más peso a la estructura.Cuanto más grande se hace un volador,más desproporcionadamente pesado sevuelve por la adición de su propiosuministro de fuerza. Llegará un punto eel cual el peso será demasiado grande para permitir que la máquina se eleve eel aire. Los cálculos realizados, erelación con el tamaño y la fuerza,sugieren que el peso máximo que puedealcanzar un vertebrado volador es deaproximadamente 22 kg: por esta razón,se piensa que el Pteranodon, y shomólogo jordano Titanopteryx,ligeramente más grande pero menosconocido, eran los animales voladoresmás grandes[47].

Sin embargo, en 1972 se produjo el primero de una espectacular serie dehallazgos que sugieren que debemoscambiar drásticamente nuestras ideasacerca del tamaño máximo permisibleen vertebrados voladores. A pesar deque las excavaciones aún prosiguen, losdescubrimientos hechos durante trestemporadas —desde 1972 hasta 1974—, por Douglas A. Lawson de laUniversidad de California, han reveladoesqueletos parciales de tres pterosauriosextremadamente grandes en el Parque
acional de Big Bend, en el condado deBrewster, Texas. Estos esqueletos pertenecen a criaturas que hacen queincluso el Pteranodon parezca pequeño.

Lawson halló los restos de cuatro alas,un largo cuello, extremidades posteriores y maxilares carentes dedientes en yacimientos no marinos; secree que estos antiguos sedimentossepultados se originaron debido aaluviones en llanuras sujetas ainundaciones. El inmenso tamaño de los pterosaurios de Big Bend, que en elmundo paleontológico se conocen como«los 747» o «Jumbos», podrían ser calibrados colocando los húmeros deTexas al lado de los de un Pteranodon:el húmero del «Jumbo» tiene el doble delongitud que el del Pteranodon. ¡Elordenador de Lawson estimó unaenvergadura de más de 75 metros para

este planeador! «No es ninguna sorpresa —dijo Lawson al anunciar el animal eScience en 1975— que los restos deesta criatura fueran hallados eTexas[48]».
A diferencia del Pteranodon, estascriaturas fueron encontradas en rocasque se formaron a 400 kilómetros en el
interior de la costa cretácea. Laausencia de yacimientos de lagos en losalrededores, habla en contra de queestos pterosaurios hubieran sido pescadores. Lawson sugiere que habíasido carroñeros, hartándose de montonesde carne podrida abandonados despuésdel desmembramiento del cadáver de udinosaurio. Quizás, al igual que los

buitres y los cóndores, estosPterosaurios volaban por encima delcadáver esperando su turno. Después dehaberse posado sobre el cadáver, sus picos sin dientes sólo les habría permitido alimentarse de los blandosórganos internos. Cómo pudieroelevarse en el aire después de habersehartado de comida es un enigma. Elanimal no podría haber batido alas de utamaño tan extraordinario estando en elsuelo. Debido a que los pterosaurioseran incapaces de correr a fin deemprender el vuelo, debieron deelevarse verticalmente en el aire. Las palomas sólo pueden despegar verticalmente, reclinando sus cuerpos y

batiendo las alas delante de ellos. En elcaso de que fueran aleteadores, ¡los pterosaurios de Texas habríanecesitado tener patas muy largas eforma de zancos a fin de elevar elcuerpo lo suficiente como para permitir que sus alas de 7 metros de longituddespejaran el suelo! La principalobjeción, sin embargo, sigue siendo laausencia de una musculatura adecuada para una operación de este tipo. ¿Es quela única solución es suponer que estascriaturas, con las alas completamenteextendidas y los elevadores alzados,fueran levantadas pasivamente del suelo por el viento? Si Lawson tuviera razón,
los pterosaurios de Texas fuera

carroñeros (lo cual comienzo a dudar),se podría prever otro problema. Laexistencia de cadáveres de dinosauriosimplica la presencia de dinosaurios. Losgigantescos y torpes pterosaurios eravulnerables a ser atacados estando en elsuelo. Así que, ¿cómo podían escapar delos formidables dinosaurios? Estando amerced de las corrientes de viento, eldespegue habría sido una empresaarriesgada[49]
Los exóticos pterosaurios deLawson plantean algunas cuestionesintrigantes. Sólo la prosecución de lainvestigación nos dará las respuestas.

VIII. LA LLEGADADE ARMAGEDÓN:¿UN CATACLISMO
CÓSMICO?
A finales del cretáceo, hace 64millones de años, la Tierra fuedevastada. La vida fue destruida por unade las peores catástrofes que hayaasolado el planeta. La causa de estadestrucción siempre ha sido en ciertomodo un enigma. Después de este sucesono siguió viviendo ninguno de losgrandes animales terrestres,

desaparecieron los plesiosaurios ymosasaurios de los mares y los pterosaurios de los cielos y nosobrevivió ninguna amonita en losfondos del mar ni el plancton formador de la creta en la superficie. Todos fueroaniquilados simultáneamente. Lasformas de vida que aún persisten — mamíferos, aves, unos cuantos reptiles, plantas terrestres, etc.—, sufrieron unadrástica reducción de su número.Heredaron un mundo que debió de parecer vacío.
Esta destrucción resulta aún mássorprendente por el hecho de que lossupervivientes eran las formas modestasdel mesozoico. Fueron precisamente los

animales y plantas que habían florecido
durante el mesozoico los que sufrierolas mayores pérdidas. Los dinosauriosestaban entre las criaturas de más éxitoque jamás hayan habitado la Tierra. «Sitener en cuenta el peculiar fenómeno dela evolución humana —declaró el paleontólogo argentino Oswaldo Reig —, debemos reconocer que el triunfo delos dinosaurios y de sus parientes hasido el mayor logro de la evolución delos vertebrados terrestres[1]…». Sreino abarcó 150 millones de años. Si elDivino Artífice de Owen hubiera tenidoen mente un proyecto para la Creación,seguramente habría sido el brontosaurioen su Edén mesozoico.

¿Por qué entonces se produjo esterepentino cambio de la política Divinahace aproximadamente 64 millones deaños? ¿Qué es lo que pudo causar talcrisis? Las pruebas son escasas y lamagnitud de la destrucción ha generadoun interés morboso sin precedentes, quetiene como resultado que las teoríassean numerosas. Una generación anterior de paleontólogos arguyó que, debido ala longevidad racial, los dinosaurios,como grupo, habían alcanzado edadesmuy avanzadas hacia finales delmesozoico. Afirmaron que, al igual queun individuo nace y muere, una raza decriaturas se podía volver senil despuésde un período largo de existencia. La

prueba de esta senilidad fue«observada» en la aparente osificacióexcesiva del cráneo de los dinosaurios:en el desarrollo de golas óseasanormales en la parte trasera del cráneodel Triceratops, el peculiar penachonasal en el Parasaurolophus de pico de pato, en el grotesco cráneo en forma de bóveda y de hueso sólido del
achycephalosaurus. Sobre loscontemporáneos viejos de losdinosaurios también actuaban fuerzasextrañas. Algunos de los miembros delas vistosas amonitas del cretáceo tardíoadoptaron formas distorsionadas,desenrollando y torciendo sus espirales.Estos eran supuestos ejemplos de rasgos

seniles producidos en el caso de losdinosaurios por una hipotética alteraciófuncional de la glándula hipofisaria.Hacia finales del cretáceo, incluso losanimales jóvenes nacían ya viejos,saliendo de los huevos con caras eminiatura arrugadas. El grupo, al igualque el individuo cuando ha pasado eltiempo previsto, estaba agotado y muriódebido a su avanzada edad[2]. Siembargo, la analogía es errónea. El proceso de envejecimiento de uindividuo no puede tener paralelo en lahistoria de un grupo. Además, ¿quésignifica que un grupo de animalesexistentes en un determinado momento,tanto si se trata de serpientes, aves,

mamíferos o dinosaurios, tienen sorigen en un océano primitivo, todostienen exactamente la misma edad? Sólonuestra creación artificial de un grupo e base a rasgos característicos —aves co plumas, mamíferos que amamantan, etc. —, nos permite adjudicarle una edad.Sin embargo, este procedimiento disociade manera poco natural el grupo ecuestión de sus antepasados. De hecho,se podría remontar la ascendencia detodos los animales contemporáneoshasta el origen de la vida misma.Además, los dinosaurios «seniles» quevivieron a finales del reino, incluso losde aspectos extraños o monstruosos,eran notablemente exitosos. El

Triceratops ramoneaba formandograndes manadas mientras que loshadrosaurios de pico de pato era posiblemente los dinosaurios másnumerosos a finales del cretáceo.Muchas de estas presuntas adaptaciones«monstruosas» representaban una graventaja para sus propietarios. La gola, o«escudo del cuello» del Triceratops,servía como área de inserción para sus potentes músculos de masticación[3], y lacresta de los hadrosaurios era unaadaptación para incrementar el sentidodel olfato.
Lejos de ser «seniles», losdinosaurios eran unas de las criaturasterrestres más avanzadas. Durante s

permanencia como gobernantes de laTierra, produjeron una serie de formas para llenar los nichos ecológicos queactualmente están ocupados por mamíferos y aves tan diversos como solos elefantes, tigres y avestruces. Losdinosaurios eran los modernos queconstantemente estaban evolucionandomientras que las arcaicas tortugas ycocodrilos apenas avanzaban. Por lo quese refiere al estancamiento, las tortugasllevaban ventaja. Su primitivo cráneoera parecido al del antepasado de todoslos reptiles. Y mientras los dinosaurioscolocaban sus patas debajo del cuerpo para lograr una locomoción eficiente yveloz, las tortugas conservaban la

marcha torpe y desgarbada de los yamuy lejanos antepasados de losdinosaurios. Para ser justo, deberíahaber sido la tortuga la que habríatenido que desaparecer. Sin embargo, lasupervivencia bajo condicionescambiantes le hace el juego a los menosespecializados; y las tortugas y loscocodrilos aún están entre nosotrosmientras que los dinosaurios murieron,cuando nuestros antepasados aún eratrepadores de árboles y musarañas quecomían insectos.
Los habitantes más intrigantes delcretáceo tardío eran los imitadoresinteligentes desenterrados en los últimosaños: «dinosaurios-avestruces» de ojos

grandes y dromeosáuridos como eleinonychus y el Saurornithoides, co
una visión binocular funcionalmenteadaptada para los pulgares opuestos.Estos dinosaurios, capaces de realizar más hazañas diestras que ningún otroanimal terrestre hasta ahora, estabaseparados de los dinosaurios por uabismo comparable al que divide alhombre y las vacas: la disparidad deltamaño cerebral es asombrosa. El potencial inherente a los dromeosaurios
celurosaurios para lograr unaevolución explosiva al principio de laera terciaria, no puede ser puesto eduda, quién sabe qué nuevas cumbreshubiesen alcanzado los sofisticados

«imitadores de las aves» si hubiesesobrevivido hasta la «Era de losMamíferos». Sin embargo,aparentemente, ni una sola población decrías de estos dinosaurios hermosos ylistos sobrevivió a loscomparativamente voluminosos y lerdosgigantes. Esto constituye un misterio tagrande como lo es la naturaleza globalde las extinciones. El agente que buscamos no respetaba a la inteligencia.
Los dinosaurios no fueron, de ningúmodo, un grupo estático. Después dehaber evolucionado a partir de losseudosuquianos del triásico, siguióhabiendo cambios constantes en elgrupo. Para citar un ejemplo, la

formación de Lance del cretáceo tardíose caracteriza por albergar Triceratops
tricornios, que no se encuentran en rocasmás antiguas; el pequeño y primitivo
eptoceratops, que sólo tenía una golaincipiente, era un contemporáneomientras que el Anchiceratops seríaencontrado en la formación ligeramentemás antigua de Lower Edmonton, en lastierras desgastadas por la erosión delrío del Ciervo Rojo; el Chasmosaurus yel Monoclonius de un solo cuerno eraceratópsidos de la formación del ríoBelly, aún más antigua[4]. Cada una deestas formaciones tenía susrepresentantes característicos quegeneralmente no podían ser encontrados

en otro lugar. En las amonitascontemporáneas se puede observar u«cambio aún más revolucionario». Aquíse produjo un cambio constante y casicompleto de comunidades enteras quesólo necesitó un millón de años para larevolución[5]. Se ha dicho muchas vecesque estos reemplazos son una regla en la
aturaleza y que, al final del mesozoico,sencillamente no hubo sucesoresdisponibles. Pero ¿por qué no habíasucesores? ¿Por qué formas de vida tadiversas como dinosaurios y amonitas, plesiosaurios y muchos tipos de plancton, pterosaurios y numerosas plantas terrestres fueron eliminadassimultáneamente en una de las

extinciones en masa más universales dela Historia geológica?
La extinción de los dinosaurios debeser analizada desde este contexto másamplio. Demasiadas «soluciones» parael enigma de la extinción se hacentrado exclusivamente en losdinosaurios, descuidando elfallecimiento de otros animales y plantascontemporáneos. Y demasiadas veces la«solución» propuesta para explicar la pérdida de las bestias más grandes no se puede aplicar de ningún modo a las más pequeñas. Sólo analizando losacontecimientos acaecidos en elcretáceo desde un punto de vista másextenso, podemos esperar reunir todas

las piezas para explicar la causa únicade todas las extinciones sincrónicas eaquella época.

Tanto Cope como Osborn jugaron con la idea

de que el enorme saurópodo había sido undinosaurio terrestre semejante a una jirafa.Sólo últimamente, sin embargo, Bakker hasuministrado una plétora de pruebasdetalladas que apoyan esta teoría. El Barosaurus (arriba), al igual que otrossaurópodos, usaba su extraordinario cuello para ramonear en las copas de los árboles adonde no llegaban los otros dinosaurios.


El Archaeopteryx es el dinosaurio conocidomás pequeño cubierto de plumas paraenfrentarse a la pérdida de calor. Éste es elespécimen de Berlín.

El Archaeopteryx no tenía músculos devuelo adecuados. Ostrom lo retrata como un predador terrestre que atrapaba insectos con

las plumas extendidas de las alas.


Un Pterodactylus perfectamente conservadode la caliza del jurásico superior de Eichstätten Baviera.
El aspecto de huevo de la cabeza del Pterodactylus era debido a su cerebro deltamaño de un ave.

Al aceptar que los pterodáctilos eran peludos, Newman se vio obligado areconocer que no podían haber sido reptiles.

Así que en 1843 los restauró comomarsupiales voladores, carnívoros.
El Pteranodon, un enorme pterosauriovolador, restaurado aquí con unrevestimiento con abrigo de piel, blanco.

Diplodocus carnegiei de Holland, de más de26 metros de longitud, en postura erguidacomo un mamífero.
Si estudiamos los dinosaurios in
vacuo, el resultado es unadesconcertante serie de teorías paraexplicar la extinción, algunas de lascuales reciben más atención que otras.Tiene especialmente muchos partidariosla teoría de que el cambio de lavegetación afectó negativamente a losenormes herbívoros. Esta idea ha sido

muy popular aunque sólo sea porque, a principios del cretáceo, haceaproximadamente 120 millones de años,hicieron su aparición las plantas coflores y la extinción de los dinosaurios«le siguió» a finales del cretáceo. Y, siembargo, los hadrosaurios yceratopsianos herbívoros tuvieron sapogeo durante el cretáceo tardío, locual es un fuerte argumento en contra deque hubieran sucumbido a algún tipo deguerra química librada por las plantascon flores.
Tony Swain, un sistemático bioquímico de los Jardines BotánicosReales de Kew, es el principal defensor de la teoría de la «agresión química de

las plantas». Hasta el cretáceo, loselementos florales dominantes era plantas sin flores, con semillas como loshelechos, coniferas y cicadáceas. Swaianalizó los complejos productosquímicos sintetizados por las plantasdurante su historia, y en especial la deaquellas que habían actuado como frenoalimenticio para los grandes herbívorosdestructivos. Los helechos y lasconiferas de principios del mesozoicoempleaban para este fin taninoscondensados, pero éstos sencillamenteno eran de ningún modo nocivos a graescala para los dinosaurios del precretáceo. Hace aproximadamente 120millones de años, las primeras plantas

con flores las sustituyeron con un nuevotipo de veneno, los potentes alcaloides.Los alcaloides tienen un sabor amargo, yalgunos como la estricnina son altamentetóxicos, otros como la morfina produceefectos psíquicos, así que ¿pudieroéstos haber afectado adversamente a losdinosaurios? La función de estos productos químicos repelentes eradisuadir a los probables herbívoros, nomatarlos. Swain estudió las reaccionesde las tortugas y mamíferos a los que ellímite de tolerancia para los taninos erasimilar en ambos: una determinadaconcentración anulará cualquier deseode comer más. Sin embargo, en el casode los alcaloides hubo una gra

diferencia. Las tortugas necesitan unaconcentración 40 veces superior a la delos mamíferos antes de quedar sensibilizadas y reducir su ingesta.Swain extrapola estos hallazgos para losdinosaurios y sugiere que — debido a
que tanto las tortugas como los
dinosaurios son reptiles — losdinosaurios eran «incapaces de detectar compuestos (venenosos) econcentraciones lo suficientemente bajascomo para resultar inofensivas, (y) posiblemente comieron la suficientecantidad como para sufrir gravesalteraciones fisiológicas e incluso lamuerte[6]». Debido a su voraz apetito,las cantidades de alcaloides consumidas

por los dinosaurios habrían sidoconsiderables. Al no darse cuenta de lasituación ellos mismos se fueroenvenenando hasta morir y extinguirse.
Había graves errores en losargumentos de Swain; algunas de sussuposiciones son injustificables; y unagrave discrepancia en su datación de loseventos del drama hace que su teoría parezca sospechosa. Si los alcaloidestóxicos aparecieron hace 120 millonesde años con las primeras plantas coflores, ¿por qué los dinosaurios«persistieron» durante 50 millones deaños más antes de morir? ¿Y por qué,cuando llegó el momento, la extinción se produjo tan repentinamente? Incluso

Swain admite que, en su fase finalacelerada, la aniquilación de toda laraza de los dinosaurios sólo tardó cincomillones de años, que, en el tiempogeológico, es rápido. ¿Cómo podemosexplicar la desaparición simultánea delos pterosaurios comedores de peces delos cielos, de los plesiosaurios ymosasaurios de los mares y de losdinosaurios más pequeños que comíamamíferos al igual que una multitud deotras extinciones? Sin embargo, elinconveniente más grave que puede ser formulado en contra de esta teoría esque fue después de la introducción delas plantas con flores, probablementeunos 30 millones de años más tarde,

cuando se produjo el apogeo de sdiversidad, una época en la cual existíamanadas inmensas de hadrosauriosherbívoros de pico de pato,ceratópsidos cornudos y anquilosauriosacorazados. Muchos de éstos, probablemente aparecieron debido a ladisponibilidad de este nuevo materialvegetal. No puede haber sido ningunacoincidencia que después de laaparición de las plantas con floresaparecieran tantos herbívoros. Colbortafirma en The Age of Reptiles:
Los dinosaurios ornítopodos,que habían gozado de un gradomoderado de variedad evolutiva

a finales del jurásico y a principios del cretáceo, comolos camptosaurios eiguanodontes, aumentaron cincoveces su número durante el período cretáceo tardío en gran parte como resultado de laradiación diversa de losdinosaurios de pico de pato.Además, surgieron dos nuevosgrandes grupos, o subórdenes, dedinosaurios herbívoros duranteel período cretáceo, losdinosaurios acorazados oanquilosaurios y los dinosaurioscornudos o ceratópsidos. Ambosgrupos de dinosaurios se

hicieron variados y numerososdurante la fase final de lahistoria cretácea, sumándose asía la serie de dinosauriosherbívoros en todo el mundo. Ytal como era de esperar, contantos herbívoros habitando laTierra también se produjo unincremento entre los carnívorosque cazaban los inofensivosconsumidores de plantas[7].
Sin embargo, en el caso delhadrosaurio, tal como había demostradoOstrom, la masiva batería de dientes y elcomplejo mecanismo de masticació

estaban diseñados para poder hacer frente a material abrasivo como son lasagujas de coniferas pero no para las blandas plantas con flores. Las plantascon flores no pudieron haber sido elveneno de los hadrosaurios. Si estehubiera sido el caso, los argumentos deSwain no habrían tenido ningunarelación con este grupo y deberían haber sobrevivido al cretáceo.
La tortuga no es un buen modelo — aun a pesar de que sea uno de los pocosreptiles herbívoros modernos—, por lasencilla razón de que está lejanamenteemparentada con los dinosaurios al igualque lo está con los mamíferos. Además,existen razones para creer que la

fisiología de los dinosaurios era mássemejante a la de los mamíferos o lasaves que a la de los reptiles arcaicos[8].
Uno de los argumentos sacado arelucir para apoyar el envenenamiento por alcaloides se centra en la retorcida postura adoptada por muchos de losdelicados dinosaurios de cuello largo(especialmente los celurosaurios)después de morir. El envenenamiento por estricnina, en efecto induce uestado opistotónico, en el cual losespasmos de la contracción muscular creada por los trastornos del sistemanervioso provocan un tensamiento de losmúsculos del dorso y cuello. Esto provoca que la cabeza quede tirada

hacia atrás. Muchos de los dinosauriosmás pequeños y más ágiles fueroencontrados fosilizados en este estado,lo cual ha llevado a pensar en repetidasocasiones que la muerte se había producido debido a un envenenamientodespués de angustiosas convulsiones.Este estado «patológico», sin embargo,se encuentra en todos los tipos dedinosaurios a lo largo de su historia; así,no solamente ocurría en losStruthiomimus de cuello largo delcretáceo sino también en el carnívorourásico Compsognathus que vivió antes
de la aparición de las plantas con flores.Muchos mamíferos y aves han sidohallados en posturas retorcidas

similares[9]. Existe una explicaciómucho más sencilla. La flexióesquelética después de la muertetambién está ocasionada por eldesecamiento de los largos ligamentosdel cuello, provocando que éstos seencojan y tiren hacia atrás la cabeza,simulando así una muerte opistotónica.La ubicuidad de los hallazgos mostrandoesta postura hace que la explicació patológica sea inverosímil,especialmente si el proceso normal dedeshidratación postmortem producirá uestado idéntico.
Es innegable que la característicamás destacada de la frontera cretáceo-era terciaria es la pérdida de los

dinosaurios; de hecho, esteacontecimiento frecuentemente define lafrontera. La repentina y evidentedesaparición del elemento másdramático de la fauna ha provocado unacierta miopía para buscar la causa.Tendemos a pasar por alto el hecho deque, en las profundidades del océano,también fueron destruidos organismosmenos espectaculares. Si queremosanalizar la extinción de los dinosauriosen un contexto —como parte de unacatástrofe mundial que afectó a la fauna al medio ambiente—, sería instructivo
desviar la atención de los miembros másllamativos del paisaje cretáceo y, en vezde esto, centrarla en los miembros

menos dramáticos de su marina.El problema de la frontera que
separa el final del mesozoico del principio del período terciario era eltema de la 21a Sesión del CongresoGeológico Internacional, que tuvo lugar en 1960. Los informes presentados por los delegados ponían gran énfasis en larevolución del mundo microscópico delos cocolitos y foraminíferos formadoresde la creta en aquella época[10]. Setrataba de organismos unicelulares,similares al plancton, que debido a queestaban envueltos por concha dura pueden ser localizados en las rocas a pesar de su diminuto tamaño. Nos proporcionan una visión de las

condiciones marinas cuando en loscontinentes vivían los últimosdinosaurios. Los delegados del congresollegaron a una conclusión unánime: etodas las partes de la transición desdelos estratos más tardíos del cretáceo(denominados de Maestricht), hasta losestratos más bajos del terciario(Dánico) existe un marcado cambio dela microfauna en cada región. El finaldel cretáceo fue testigo de la expulsióde los foraminíferos existentes y del plancton formador de la creta, mientrasque, en la base de la era terciaria,hacían eclosión foraminíferos totalmentediferentes. Los dos tipos planctónicoseran completamente distintos, au

cuando a ojos de los no expertos noexistan muchas diferencias. La magnitud brusquedad del cambio era una
evidencia de que se había producido unarevolución en el mar. Obviamente estosdiminutos organismos, a merced de lascorrientes del mar, habían sufrido nomenos que los dinosaurios. Más de udelegado pensó que la solución delenigma había sido un descenso repentinode la temperatura. William Hay, cuyaárea especial de estudio era México,resumió la opinión imperante:
La fauna planctónica (de principios del período terciario)está compuesta por relativamente

pocas especies, todas de tamañosingularmente pequeño. Se parece a la fauna planctónicamoderna septentrional(templada) y da la impresióncomo si el agua de los océanosera mucho más fría durante elDaniano que inmediatamenteantes o después.
El estudio realizado enMéxico apoya la idea de que, afinales del período deMaestricht, se produjo una de lasmayores catástrofes de laHistoria de la Tierra. A finalesdel período de Maestricht hubouna regresión mayor del mar,

registrada no sólo en Méxicosino en gran parte de la región deTethis y otros lugares. Al mismotiempo, se produjo unenfriamiento drástico de lasaguas de los océanos. Losgrandes cambios climáticos quedebieron de producirse probablemente fueron la causade la extinción de los grupos biológicos termofílicos («quegustan del calor») tancaracterísticos del mesozoicotardío. La fauna dánica de aguafría representa el comienzo deuna nueva secuenciaevolutiva[11].

En otras palabras, a pesar de queestaban acogedoramente aisladas en losocéanos, estas diminutas criaturas planctónicas notaron los efectos de udeterioro del clima. Eran pocas las que podían resistir el enfriamiento del agua
se parecían al plancton de las zonastempladas modernas. Fueron las que nofallecieron junto con los dinosaurios.
Las plantas no lo estaban pasandomejor en tierra; fueron profundamenteafectadas cuando finalizó el cretáceodando paso al terciario. Las plantastropicales fueron expulsadas por sus parientes más resistentes que procedíade las regiones templadas situadas másal Norte.

Debido a que el polen es la partemás resistente de las plantas, éstefrecuentemente se conserva en las rocasdejando un registro de la vida vegetal decada período. En el este de Montana, losanálisis del polen fosilizado revelan quela mitad de todas las plantas delmesozoico tardío eran plantas coflores, termofílicas; a principios del período terciario ya sólo representabamenos de la tercera parte. Las coniferasde hoja perenne, cuyas agujas les permitían tolerar condiciones más frías,habían incrementado su dominio sobrela tierra como resultado del cambiorepentino del clima mundial[12]. Lasconiferas no sólo remplazaron a las

plantas con flor. Las plantas típicas delmesozoico como las cicadáceas — durante tanto tiempo la principal comidade muchos dinosaurios herbívoros— preferían regiones con temperaturasaltas y estables y, por esta razón, hoy edía están limitadas a áreas uniformescomo son Nueva Zelanda, América delSur y África del Sur. Cuando a finalesdel mesozoico la temperatura descendió,éstas también perdieron su dominio elas regiones más septentrionales siendoun ejemplo las coniferas como el
etasequoia[13]. Estos estudios sobre el plancton y las plantas terrestres fósilessugieren provisionalmente que la Tierrase vio inmersa en un período de frío y

que los efectos se notaron tanto en elmar como en tierra. Las pruebas socircunstanciales, obtenidas en base a lacomparación de especies del mesozoicocon otras formas vivas estrechamenteemparentadas; conociendo las preferencias medioambientales de lasespecies existentes, podemos deducir las condiciones climáticas de hace 64millones de años. Con todo, laconclusión de que el clima se estabadeteriorando parece ineludible.
La destrucción de plancton, amonitas reptiles en los océanos y dinosaurios,
pterosaurios y plantas en tierra pareció producirse en un espacio de tiempocatastróficamente corto. En casi todos

los casos, su desaparición de las rocases tan repentina que muchas autoridadesincluso hablan del diezmamientosincrónico de los grupos ubicuos emenos de un millón de años.M. N. Bramlette, del Instituto Scripps deOceanografía, en un estudio sobre ladestrucción masiva del plancton estimaque sólo fueron necesarios unos milesde años para completarla[14]. Otros piensan que unos cuantos días ya seríasuficientes.
La aniquilación simultánea que tuvolugar en toda la superficie de la Tierraha llevado a algunos a formular explicaciones, a primera vista,extravagantes. Han vuelto a surgir de

forma dramática teorías cataclísmicas,descartadas de la Geología desde laépoca del barón Cuvier, cuya teoría delas extinciones masivas que implicabarevoluciones o catástrofes inexplicablescoincidían de forma bastante adecuadacon los años más vigorosos de laRevolución Francesa. Un acontecimientoque podía poner un brusco fin a grupostan diversos como dinosaurios y plancton de forma simultánea en todo el planeta, necesariamente tenía que tener proporciones verdaderamente cósmicas.
La explosión de una supernova,como la observada por los astrónomoschinos en 1054, emitiría radiaciócósmica de inimaginable magnitud a la

galaxia. La explosión de una estrella dediez masas solares liberaría una ondaexpansiva equivalente a los efectoscombinados de1 000 000 000 000 000 000 000 000000 bombas de hidrógeno de diez megatones.La «estrella huésped» medieval china,cuyos restos ahora conocemos como laconstelación de Cáncer, estabademasiado alejada como para haber representado un peligro de radiació para la Tierra. Pero si explotara unaestrella cercana, a unos cien años luzaproximadamente de nuestro sistemasolar, los niveles de radiación en la zonaalta de la atmósfera —que normalmente

es de unos 0,03 roentgens por año— ascenderían hasta alcanzar losasombrosos 3000 roentgens. Debido aque la radiación sería recibida en formaconcentrada y se extendería durantealgunos días a lo sumo, el efecto seríacatastrófico. Los niveles de radiación ela atmósfera se incrementarían diezmillones de veces y tendrían un efectosimilar al de un holocausto nuclear universal. A pesar de que las supernovastienen una frecuencia de una explosiócada cincuenta años en nuestragalaxia[15], la mayoría, al igual queCáncer, están tan lejos que soinofensivas. Sin embargo, la Tierrarecibe una dosis aguda de radiación de

500 roentgens en intervalos deaproximadamente 50 millones de añoscuando explota una estrella más cercana; probablemente cada 300 millones de
años una dosis de 1500 roentgens.Desde el Período Precámbrico, hace600 millones de años, las probabilidades indican que debió dehaber una ocasión en la cual la Tierrafue inundada de radiación de pocomenos que dos veces esta intensidad.
La dosis letal para los animales delaboratorio oscila entre 100 y 700roentgens, de forma que una explosióde radiación de la magnitud de unasupernova, en el caso que llegara deforma no diluida a la superficie del

planeta, haría estragos. Los dinosaurioshabrían sido particularmentevulnerables; al ser grandes y carecer de protección, incapaces de esconderse emadrigueras, habrían sido irradiadoscon dosis masivas. Aquellos que nohubiesen perecido debido a alteracionescelulares y cáncer seguramente habríagenerado descendientes mutantes. Estoen el caso de que hubiera habido unaexplosión de una supernova en lavecindad, hace aproximadamente 64millones de años.
Resulta muy atractivo jugar con estascifras astronómicas, especialmente siestán relacionadas con sucesoscataclísmicos en el Universo. El

morboso atractivo de los cataclismosatrae la mente como espectadores a uincendio. El encaprichamientointelectual es un pobre criterio para saceptación. Las anomalías que seobservan en esta teoría de la radiaciócósmica directa sencillamente sodemasiado importantes, inclusosuponiendo que los científicosespaciales dispongan de cifrascorrectas. Para matar a un sencilloorganismo unicelular, por ejemplo, senecesita aproximadamente diez veces ladosis letal de un ratón. Aun así, losmamíferos sobrevivieron el ataqueviolento del cretáceo mientras que el plancton fue afectado profundamente.

Debido a que incluso una capa delgadade agua superficial protegeríaeficazmente contra la radiación, la teoríade la radiación directa no es válida yaque murieron las criaturas equivocadas.El modelo ha sido criticado incluso por algunos físicos que insisten en que eltortuoso camino tomado por la radiaciócósmica para un recorrido de diez añosluz, habría provocado que llegase en uespacio de tiempo alargado de 40años[16].
El bombardeo cósmico directo puede ser descartado. Sin embargo, nose habían tenido en cuenta lasfluctuaciones de la temperatura que probablemente fueron la causa principal

del desastre. Al menos dos trabajadoresinteresados por el tema de la extincióde los dinosaurios han sido convencidos por los argumentos de los partidarios dela teoría de la supernova. Dale Russell,del Museo Nacional de Ciencias
aturales de Ottawa, y Wallace Tucker,del American Science and Engineeringen Boston, han modificado la teoría paradestacar la importancia de los efectosclimáticos de la radiación incidente. Laatmósfera absorbería el equivalente a lafuerza de 100 000 bombas de hidrógenode diez megatones, si se produjera laexplosión de una estrella cercana anuestro planeta. La radiación adoptaríaen gran parte la forma de rayos X que

serían absorbidos por la capa de ozono la ionosfera. Los efectos climáticos
resultantes serían desastrosos. Laturbulencia generada alteraríadrásticamente la capacidad de retenciódel calor de la atmósfera. El aire más bajo, saturado de agua, sería desplazadohacia regiones más altas donde seformarían cristales de hielo y actuaríadesviando los rayos solares, provocando así un descenso de lastemperaturas en todo el planeta de formaalarmantemente imprevista. Esinteresante señalar, en este contexto, queel incremento agudo de la emisión derayos X del Sol provoca un cambio en lacirculación atmosférica hacia una de

características claramente glaciales[17].¿Era posible que el bombardeo de lasupernova hubiera provocado elrepentino período de frío temporalresponsable de la aniquilación de todaforma de vida oceánica y terrestre? Losefectos duales de la radiación a niveldel suelo, resultantes de la interaccióde rayos cósmicos y rayos gamma con laatmósfera, y del descenso de lastemperaturas, podrían ser la razón detodos los cambios observados segúRussell:
Los sucesos que marcan elfinal de la era de los dinosaurios

parecen haber sido semejantes alos que podrían seguir a lallegada del estallido inicial deenergía de una supernovacercana a nosotros. Losorganismos terrestres másgrandes y más evolucionados(dinosaurios, plantas con flores),habrían sido diezmados por losefectos combinados de laradiación y el frío. Losorganismos marinos probablemente habrían sidoafectados más severamente por el enfriamiento de los océanos,aunque algunas formas quevivían en aguas poco profundas

posiblemente habrían sidoexterminadas por la radiación odebido a las turbulencias de aguacreadas por tormentas. La Tierraestuvo fría el suficiente tiempocomo para enfriar los océanos, pero no lo suficientemente como para permitir el desarrollo de placas de hielo continentales enlas latitudes altas, ya que no hayevidencias de glaciación enningún sitio del planeta enaquella época[18].
Esto ilustra bien la gravedad y brusquedad de la devastació

concebidos por los teóricos de lasupernova. Niveles de radiaciósemejantes a los que siguen a una guerranuclear, tormentas en todo el planeta deextrema intensidad, temperaturasdesastrosamente bajas y la destruccióde los elementos primarios de la cadenaalimenticia en tierra (plantas con flores cicadáceas), y en el mar (plancton y
amonitas), debieron de hacer estragosincalculables. Y todo esto en el espaciode una semana. ¿Por qué, se preguntauno, sobrevivieron los mamíferos? ¿Ylos cocodrilos y aves sumamentevulnerables, estando todos tan expuestoscomo los dinosaurios?
Existe un dato obstinado, si

resultara ser verdad , quemanifiestamente no concuerda con lateoría de la exterminación sincrónica yrepentina de la vida. Si se examinan comás detalle las rocas que albergan a losúltimos dinosaurios, parece como si ladesaparición hubiera sido uacontecimiento progresivo en vez derápido. (Esto es cuestionado por los partidarios de la teoría de la supernovaque ven en el aparente titubeo elresultado del sondeo selectivo). ElTriceratops rinocerino vagaba en otrostiempos formando manadas, como las delos bisontes americanos antes de laaparición del rifle Winchester. Sushuesos se encuentran en los últimos

estratos del Cretáceo, la formación deLance (por ejemplo, en la regióoriental de Wyoming[19]). Las conchas yareniscas de esta formación tienen ugrosor de más de 60 metros de arribaabajo, y dejan un registro de lascriaturas que vivieron hasta finales delmesozoico. El Triceratops, que se hallaen abundancia en el fondo de laformación, se hace progresivamente másraro a medida que ascendemos hacia lasrocas terciarias sobrepuestas.Finalmente, ya no se encuentra ningúfósil, indicando así la desaparición delos dinosaurios. El Triceratops debió deser el dinosaurio más resistente. Siembargo, incluso su curtida piel no pudo

protegerlo contra la llegada delinvierno. El Triceratops sobrevivió amuchos de sus contemporáneos más primitivos y vivió hasta el final delcretáceo, convirtiéndose en una figurasolitaria del paisaje. Colbert econcreto ha argumentado que poco antesdel final del cretáceo, hubo un períodode declinación entre los dinosaurios: notanto una disminución de su número sinode su variedad. El número de géneros dedinosaurios disminuyó hasta tal puntoque, tal como observa, los grandescarnívoros debieron de haber tenido unadieta cada vez más monótona[20]. Eltópico de la confrontación entre losTriceratops y Tyrannosaurus

probablemente no era tanto una cuestióde libre elección para el carnívorogigante, sino el resultado de la falta dealternativas adecuadas. Al igual que casitodos los grupos, afirma Colbert, loshadrosaurios comenzaron a escasear hacia fines de este período. De hecho, laformación Lance ha proporcionadomenos de la cuarta parte de géneros dehadrosaurios que la formación BellyRiver más antigua. Lo mismo se puededecir de los anquilosaurios acorazados,cuyos representantes hallados en Lance,que forman nuestras colecciones,constituyen menos de la tercera parte delnúmero encontrado en estratos másantiguos. En la época de Lance,

aparentemente el número de géneros deceratópsidos había sido reducido a lamitad[21]. Si esta disminución de ladiversidad es real, esto significará queel comienzo de las condiciones adversasfue temprano, eliminando las formas comenor tolerancia. Sin embargo, lamayoría de los dinosaurios dejaroalgunos supervivientes resistentes que,al igual que el Triceratops, explotarola falta de competición y se extendieroformando grandes manadas.
De forma similar, parece que losictiosaurios marinos perecieron antesdel final de este período y es posibleque los plesiosaurios fueradesapareciendo en los mares cada vez

más fríos, antes de la desaparición delos últimos dinosaurios en tierra. Éstas pérdidas en la parte superior de lacadena alimenticia marina reflejan ladesaparición de la base de plancton. Lasamonitas, cuyas conchas ornamentalesen espiral habían dominado las rocas primitivas, también estaban sufriendouna reducción de su diversidad antes delfinal aunque el ritmo de extinción seincrementó drásticamente hacia el finalde éste período[22]. Existe u paralelismo sorprendente entre la pérdida progresiva de las amonitasmarinas y la disminución de la variedadde dinosaurios en el mundo cretáceotardío. Si estos hallazgos so

confirmados, desde luego invalidaríamuchas de las tonterías de losargumentos aportados por los partidarios de la supernova. Noobstante, incluso suponiendo que estosresultados reflejen fielmente la gradualdecadencia de la vida durante elcretáceo, queda el problema de explicar la extraordinaria aceleración del proceso de extinción en su fase final.¿Por qué se aceleraría tan rápidamenteesta progresiva extinción? ¿Y por qué leocurrió a tantas criaturas noemparentadas de forma casi simultánea,llevando a una aparente crisis global?¿Hubo un período de frío efímero perograve hacia el final del Cretáceo que

tuvo como resultado la creación de ufuerte gradiente entre el ecuador y los polos y el desplazamiento de lasregiones tropicales situadas más al
orte hacia la zona templada, tal comoatestiguan el plancton y las coniferas?¿Se estableció un fuerte gradiente diariocon una aguda caída de las temperaturasal anochecer? Las plantas deciduas coflores son una prueba de que, hacia elfinal del cretáceo, las estaciones estaba bien constituidas. Por lo tanto, ¿es posible que las estaciones se hicieramás marcadas hacia el final, coinviernos cada vez más fríos?
Los reptiles terrestres quecompartían el final del cretáceo con los

dinosaurios nos serían familiares hoy edía. En aquella época, los cocodrilostenían un aspecto moderno. Las iguanas los monitores prosperaban y había
serpientes, aunque raras. Incluso algunasde las tortugas que vivían en los mares,del cretáceo, tardío tienen actualmentedescendientes vivos. Todossobrevivieron mientras que una gravariedad de tipos de dinosaurios — ceratópsidos, anquilosaurios,hadrosaurios de pico de pato,tiranosaurios, dromeosaurios,celurosaurios y muchos más— desaparecieron simultáneamente. ¿Quédiferenciaba a los dinosaurios de losreptiles?

Una respuesta es evidente: losreptiles son de sangre fría mientras quelos dinosaurios probablemente eran desangre caliente. Cuando la temperaturadesciende por debajo de un gradoóptimo, los reptiles se vuelve perezosos. Ellos mismos se vuelvefríos y su índice metabólico disminuyeagudamente, sumiéndolos en un estadode letargo y finalmente hibernación.Pero permanecen vivos, aun atemperaturas ambiente por debajo del punto de congelación y, de esta manera,son capaces de sobrevivir en zonastempladas hoy en día. Así, no es ningunasorpresa que los reptiles quesobrevivieron al Cretáceo tambié

fueran los que podían esconderse debajode leños para hibernar o, como en elcaso de las tortugas y los cocodrilos,excavar túneles en las orillas de los ríos para buscar protección, como hace elcaimán chino hoy en día. Sin embargo,los dinosaurios eran incapaces dehibernar. No solamente eran demasiadograndes[23], sino que probablemente noestaban adaptados fisiológicamente paraello. La razón del por qué es fácil decomprender. Los lagartos, con srelación más grande de área desuperficie/volumen perdían rápidamentecalor cuando descendía la temperatura yesto los inactivaba rápidamente. Loslagartos debían estar preparados para

hibernar. Sin embargo, los pequeñoscambios de la temperatura ambientaleran compensados fácilmente por losvoluminosos dinosaurios con su área desuperficie proporcionalmente más pequeño. Todos los dinosaurios eragrandes, pocos pesaban menos de 50,8kg en vida[24], y algunos alcanzaban las80 toneladas. Incluso los animalesóvenes parecen haber nacido co
tamaños bastante considerables. Cocuerpos tan grandes, fácilmente podía permanecer calientes en condicionesuniformes y no necesitaban uaislamiento superficial como pelo o plumas. Al igual que los grandesmamíferos que viven en los trópicos

como los elefantes, hipopótamos yrinocerontes, los dinosaurios nonecesitaban protección alguna. El hechode que los dinosaurios no presentabaaislamiento indica que se habíadesarrollado bajo condicionesclimáticamente uniformes. Por otro lado,los reptiles semejantes a los mamíferos,nacidos en el duro clima del Pérmico,habían aislado sus cuerpos con pelo yesta característica había sido heredada por los mamíferos[25]. Esto les permitíareducir su tamaño y, a pesar de ello,sobrevivir a la exposición. Esto tambiéhizo posible que sobrevivieran alcretáceo. Si a finales del cretáceo lastemperaturas disminuyeron en todo el

mundo, los dinosaurios habrían sidoatrapados por su propio ingenio. Elfactor que había significado el éxitodurante 150 millones de años militaríacontra ellos. Eran de sangre caliente:debían mantener una temperatura internaconstante a fin de permanecer con vida, pero no tenían aislamiento alguno contrael frío[26]. En un principio, losdinosaurios posiblemente fuerocapaces de generar suficiente calor para permanecer calientes, pero cuando lasestaciones se hicieron más marcadas ylas heladas se convirtieron en un sucesohabitual cada año, incluso en latitudes bajas, y los lagartos hibernaron mientrasque los pequeños mamíferos cubiertos

de pelo encontraron madriguerascalientes donde refugiarse, losdinosaurios se quedaron fuera,expuestos al frío. Incapaces de hibernar debido a su gran tamaño, estosleviatanes posiblemente muriero«congelados».
Los pterosaurios alados plantean u problema completamente diferente. Era peludos y de sangre caliente como losmamíferos, y aun así perecieron juntocon los dinosaurios. La perdición finalde los pterosaurios fue el castigo quetuvieron que pagar por su extremaespecialización. Las grandes alas posiblemente hicieron que la pérdida decalor fuera un factor crítico. Si

embargo, el deterioro del clima posiblemente afectó a los pterosauriosde otra forma. El Pteranodon era umaestro volando lentamente, siendocapaz de permanecer suspendido en elaire con velocidades de hasta 24 km por hora. El clima uniforme del cretáceo,con un tiempo templado hasta latitudesmuy altas, habría provocado vientossuaves ideales para volar. El
teranodon, como recordará el lector,sólo necesitaba una brisa de 24 km por hora para elevarse en el aire sin ayudaalguna. Se ha sugerido que, debido a queaumentó el gradiente de temperaturasentre los polos y el ecuador, se alteró lafuerza de los vientos del Oeste y vientos

alisios. Un incremento de sólo 16 k por hora de los vientos predominantes probablemente habría sido demasiadotempestuoso para el apacible planeador,haciendo que perdiera el control[27]. Elgigantesco pterosaurio de Lawsoencontrado en Big Beng habría padecido, además, el problema delsuministro alimenticio cada vez másescaso. Si la teoría de Lawson era cierta el nuevo pterosaurio era un animal
carroñero —que dependía de loscadáveres de los grandes dinosaurios para vivir—, como consecuencia de laextinción de los dinosaurios se habría producido también su propiadesaparición. Tiene una gra

importancia que ningún animal grandesobreviviera al Cretáceo. Los animales pequeños heredaron el mundo. El pterosaurio de Texas, habituado a comer cantidades enormes de carne putrefacta,habría sucumbido en un mundo donde lacarroña tenía el tamaño de una rata.
Los partidarios de la teoría de lasupernova siguen siendo inflexibles. Loscriaturas (específicamente losictiosaurios) que desaparecieron antesdel final de este período, estádesligadas de las extincionescatastróficas finales. Además, ladisminución progresiva de la diversidadentre los dinosaurios observada por Colbert es considerada ilusoria. El

último estudio de Russell sobre el predominio de dinosaurios durante elCretáceo tardío, publicado en 1975,revela que, en efecto, existe un factor dedistorsión en la recogida de muestras.Durante generaciones, los estratossuperiores del cretáceo superior habíasido explorados más frecuentemente yde modo más exhaustivo que los estratosmás jóvenes, provocando así una mayor producción de especies de dinosaurios.La correlación de Russell entre eltiempo de trabajo empleado en cualquier formación y el número de especiesextraídas de ésta, es muy sugestiva.Ignorando esta anomalía en larecolección de muestras, los

paleontólogos supusieron falsamente quelos dinosaurios estaban escaseandohacia el final del cretáceo; lo quesucedía es que los mismos paleontólogos habían recogido menosmuestras de las rocas más recientes. Unavez corregido este error, Russelldeterminó que la diversidad de los
dinosaurios posiblemente no disminuyó
notablemente incluso hacia el final del
cretáceo[28]. Posiblemente, no unoscuantos rezagados sino toda la estirpe delos dinosaurios, floreció hacia el finalde este período. ¿Era posible que lasamonitas también hubieran sido objetode una tendencia similar en larecolección de las muestras?

De acuerdo con los catastrofistasmodernos, los dinosauriosdesaparecieron con la explosión másespectacular desde la Creación y, a pesar de la persistente aversión de losgeólogos a los cataclismos, resultadifícil no estar de acuerdo. La caza hacomenzado buscando los vestigios deuna supernova cercana, y es alentador observar que efectivamente se hadetectado indicios de una explosióestelar a finales del cretáceo[29]. Sea loque sea, una supernova o cualquier otrofactor, lo que trastornó el planeta hace64 millones de años, provocó elrepentino final del orden mundialmesozoico y cerró uno de los capítulos

más importantes de la Historia de laTierra. A pesar de que la datación deeste acontecimiento aún sigue siendodiscutida apasionadamente, hay pocosdesacuerdos respecto a su brusquedad.El debate se centra ahora en qué espaciode tiempo. La magnitud de ladevastación no puede ser infravalorada,a que la destrucción de todos los
animales de tamaño grande y mediano,unido a la interrupción de la vidamarina, alteró la dirección de la vidasobre la Tierra. Sea cual sea lanaturaleza del acontecimiento del cualnos ocupamos, debió de tener proporciones cataclísmicas.
¿Podemos imaginarnos el

sufrimiento de los dinosaurios hacia elfinal? ¿Qué evidencia nos podría revelar los tormentos de criaturascompletamente extrañas separadas denosotros por 64 millones de años? Elos últimos años, Heinrich Erben, delInstituto de Paleontología de laUniversidad de Bonn, ha estadoexaminando las cáscaras de huevos delos dinosaurios de finales del cretáceo,midiendo su grosor en diferentesestratos y registrando las tendencias. Elanálisis estadístico de Erben, obtenidoen base a miles de fragmentos de capassucesivas de rocas en Aix-en-Provence Corbières, en los Pirineos franceses,
ha proporcionado resultados

asombrosos e imprevistos. Los estratosde rocas que contienen las cáscarasfosilizadas representan las partessuperiores del estrato superior deMaastricht y, por lo tanto, abarcan u breve período de tiempo inmediatamenteanterior a la desaparición de losdinosaurios. Los huevos de los estratosmás antiguos tienen cáscaras gruesas, dehasta 2,5 mm, perfectamente aceptables para huevos de gran tamaño. Pero alllegar a estratos más altos (y por lo tantomás jóvenes), Erben encontró que lascáscaras se hacían cada vez másdelgadas mientras que las de las rocasmás jóvenes ya eran tan lastimosamentedelgadas —frecuentemente sólo 1 mm—

que debieron de haber padecido unaextrema fragilidad[30].
Debido a que Erben encontrómuchos huevos enteros, podemossuponer que sobrevivieron a laexperiencia de haber sido puestos; peroestas mismas cáscaras intactas indicaque las crías no salieron de los huevos.Las implicaciones son obvias. Losdinosaurios reaccionaron frente a u período de estrés breve perocontinuado[31] de la misma forma comolo hacen las aves modernas, poniendohuevos con cáscaras cada vez másdelgadas. El estrés en las aves, ya sea provocado por el frío, envenenamiento por DDT o sobrepoblación, resulta e

un desequilibrio del delicado sistemahormonal que altera el tamaño de lasnidadas y el grosor de las cáscaras. Lascáscaras pueden llegar a ser tadelgadas que se rompen o no tienesuficiente calcio que es absorbido por los embriones para construir susesqueletos. En los últimos años de sdominio, el dinosaurio estuvo sometidoa una presión insoportable. Esto escierto. Las solitarias bestias quelograron seguir con vida nos dan unaidea de la agonía que debieron de haber sufrido. Los dinosaurios, con sussistemas hormonales completamentedesafinados, reaccionaron poniendohuevos de cáscara débil y, con ello,

sellaron el destino de sus propiosdescendientes.
En 1973, el equipo de Bonconfirmó sus anteriores hallazgoscuando localizó ocho huevos más —dosde ellos intactos— en un peñasco cercade Corbières. Los huevos intactos eragrandes, de aproximadamente 23centímetros de largo y 18 centímetros deancho, y procedían del horizonte másoven. Los fragmentos, observados a
través de un microscopio electrónico,tenían una cáscara tan delgada que losembriones de los dinosaurios posiblemente no pudieron absorber bastante calcio para completar laformación de sus esqueletos. Los

majestuosos dinosaurios, señores delmesozoico que disfrutaron de un nivelde organización semejante al de losmamíferos o de las aves, no se habíaido con una explosión pero sí con ugemido, el gemido de las crías que perecieron encerradas en sus diminutascárceles.
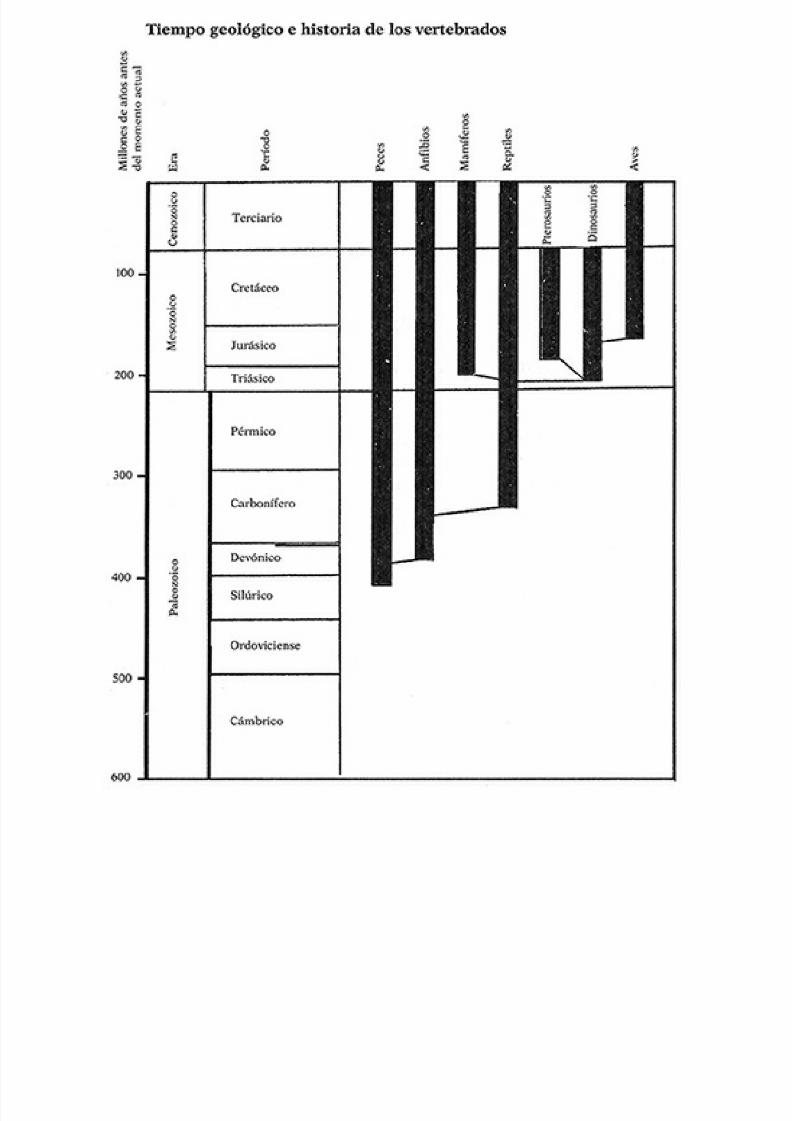

En el Apéndice I se incluye un glosario delos términos técnicos que aparecen en elGlosario y árbol genealógico de losvertebrados superiores.

Apéndice 1
NOTA SOBRE LACLASIFICACIÓN
Los animales del mesozoico, quetienen un papel crucial en la presentetesis —dinosaurios, tecodontes y pterosaurios— siguen siendodenominados oficialmente reptiles. Laclase de los reptiles fue creadaoriginalmente para incluir a las

serpientes, los lagartos y las tortugas,tienen mucho antes de que reaparecieralos saurios del mesozoico a comienzosdel siglo XIX. Los reptiles eran, por definición, animales de sangre fría couna envoltura escamosa y extremidadesdesparramadas en sentido horizontal,incapaces de mantener una actividadenérgica y generalmente limitados aregiones climáticamente uniformes.Cuvier justificó la inclusión de los pterodáctilos, y Owen la de losdinosaurios, en base a algunos huesos elos esqueletos fosilizados queaparentemente eran semejantes a los delos lagartos. Dado el nivel deconocimiento actual, este intento de

relacionar las formas de vida pasadas y presentes es loable por su valor heurístico.
Pero la Paleontología no ha estado parada durante el siglo intermedio o másque ha pasado. Lentamente, a veces casiimperceptiblemente y otras muchas penosamente, las viejas ideas han sidoinvalidadas poco a poco. Consideren elcreciente conocimiento de la fisiologíaendotérmica, de tipo avícola de los pterosaurios, su gran cerebro y pelaje;las controversias sobre la postura de losdinosaurios, semejante a la de losmamíferos, y la provisional correlacióde este rasgo con la calidad de sangrecaliente; y la postura de los tecodontes y

posiblemente un metabolismo activo. Laclasificación, sin embargo, no hallevado el mismo paso que el cambio denuestras ideas, teniendo como resultadouna situación anómala, según la cual laclase de los reptiles comprendecriaturas tan diversas como lo son losvoladores peludos, inteligentes yendotérmicos; los ectotermos arcaicos,de extremidades desparramadas ycerebro pequeño (lagartos y tortugas);los endotermos terrestres gigantes yerguidos; y los dinosaurios corredoresveloces y extremadamente sofisticados, al menos uno ( Archaeopteryx) co
plumas. La naturaleza arbitaria de esteagrupamiento es prueba del desconcierto

de muchos paleontólogos.La clasificación no solamente
debería reflejar el progreso evolutivo dela vida, sino también la adquisición denuevos niveles importantes deorganización y su ulterior explotación.La clasificación corriente de losvertebrados «superiores» fracasarotundamente, al no reflejar los logrosde los dinosaurios y pterosaurios.
La clase de los reptiles es un sacovariado de formas singularmentediversas y, aunque en un principio no eraesto lo que se pretendía, se haconvertido en una colección arbitrariade animales. Hablando claramente, hayque hacer algo.

Uno de los cambios más importantesen la historia de la vida se produjo alcomienzo del triásico —haceaproximadamente 210 millones de años —, cuando algunos de los tecodontesseudosuquianos adoptaron por primeravez una postura erguida, alargaron susextremidades posteriores para aumentar la zancada y probablemente aumentarosu capacidad para producir energía. Estaserie de caracteres fueron unainnovación clave de consecuenciasmayores, ya que permitió a susdescendientes conquistar la tierra y elaire. Desencadenó las explosiones delos dinosaurios y pterosaurios.
Esto debe ser reconocido en la

clasificación. No se puede justificar durante más tiempo la inclusión dedinosaurios y lagartos en un mismogrupo. Los «reptiles» de sangre calientedel mesozoico habían roto las relacionescon sus antepasados de la misma formaque lo han hecho los mamíferos. Debidoa que los mamíferos viven, somosconscientes de que son distintos y lesotorgamos una clase propia. Lo mismoes válido en el caso de las aves. Losdinosaurios y pterosaurios —a pesar dela extensión de su reino— nosobrevivieron, y tuvieron que ser clasificados en un principio en base avestigios fragmentarios y uconocimiento insuficiente. Es ahora

cuando nos damos cuenta de su unicidad.


Árbol genealógico esquemático de losvertebrados superiores. Los grupos en negroson considerados aquí como de sangrecaliente, los rayados como de sangre fría. Laendotermia probablemente se desarrollósólo dos veces durante la historia de losvertebrados, entre los terápsidos y lostecodontes. Los dinosaurios y pterosauriosheredaron la condición de los tecodontes yésta se transmitió a las aves a través de losdinosaurios.
A los «saurios» de sangre calientedel mundo mesozoico debería otorgarseun rango a fin de hacer justicia a simportancia fisiológica, anatómica yecológica. Por esta razón, proponemosla creación de las clases dinosaurios

(incluyendo a los seudosuquianos) y pterosaurios. Debido a que ambosgrupos probablemente heredaron suscaracterísticas fisiológicas de lostecodontes primitivos del triásico, losdinosaurios y pterosaurios podrían ser agrupados naturalmente en la superclasede los endosaurópsidos.

Apéndice 2
ALGUNOSPROBLEMAS
«La crítica de nuestras conjeturas tieneuna importancia decisiva: el hecho desacar a luz nuestros errores nos hace
comprender las dificultades del problema que estamos intentando
resolver».
K ARL POPPER , Conjectures an

Refutations[1]
Lo relatado en este libro no pretendede ninguna manera ser una declaraciódefinitiva. Eso es la desafortunadasuerte de los libros de texto. Essolamente un informe provisional delestado actual de la Ciencia, laPaleontología está continuamentecambiando. Quizás algunas de las ideas especulaciones sean confirmadas pero,
inevitablemente, muchas resultarán ser incorrectas. Algunos problemas ya hasido resueltos. Éstos, sin embargo, norestan valor a la fuerza aclaratoria denuestra teoría endotérmica de losdinosaurios. No obstante, estas

dificultades requieren ser expuestas yesperamos que ello sea un estímulo parala investigación ulterior. Por lo tanto, yen resumen, de acuerdo con los principios de la filosofía de Popper,debemos exponer nuestra teoría a lascríticas más severas a fin de poder estimar su valor.
Una de las críticas más fuerteshechas contra nuestra teoría, formulada por primera vez por Alan Feduccia e1972, se refiere al tamaño cerebral delos dinosaurios. Las aves y losmamíferos poseen cerebrosrelativamente grandes en marcadocontraste con los notoriamente pequeñoscerebros de la mayoría de dinosaurios.

A pesar de ello, Ostrom y Bakker postulan para los dinosaurios niveles deactividad como los que se observan elos mamíferos y las aves. Seguramente,arguye Feduccia, estos niveles deactividad habrían dado como resultadouna mayor complejidad del cerebro delos dinosaurios[2]. Feduccia no sabía eese momento que Dale Russell habíademostrado de forma bastanteconcluyente que algunos dinosaurios, dehecho los más sofisticados, losdromeosaurios y estrutiomímidos,desarrollaron cerebros relativamentegigantescos (algunos incluso másgrandes que los de las aves) a fin decoordinar el equilibrio, agilidad,

agudeza visual y destreza manual, deforma que éstos deben ser excluidos dela objeción[3]. Más tarde, Feducciaconcedió que «quizás éstos (refiriéndosea los dinosaurios avestruces ydromeosaurios de Russell) eraendotermos o, al menos, animalescretáceos inteligentes[4]». Sin embargo,queda por aclarar el problema de ladisparidad entre el cerebro y el tamañocorporal en la mayoría de los grandesdinosaurios. ¿Podría ser que, aunque losdinosaurios pequeños y grácilesnecesitaban cerebros grandes, losimponentes gigantes no tenían elsofisticado comportamiento que exige laexistencia de un cerebro agrandado

aunque al mismo tiempo sean animalesde sangre caliente? La endotermia, per
se, posiblemente no requiera laexistencia de un cerebro grande. HarryJerison ha sugerido recientemente en sexhaustivo estudio de la Evolution o
the Brain que, «a priori, sería deesperar que los animales con un controlextrínseco de la temperatura, como loslagartos que deben tener respuestas alsol o a la sombra… utilizarían más
tejido nervioso que los (endotermos) para controlar su temperaturacorporal[5]». En otras palabras, lasrespuestas de la conducta de los lagartosorientadas, en función del sol, posiblemente requieran un mayor

volumen cerebral que el mecanismo deregulación de la temperatura interna puramente automático de losendotermos. En base a hipótesis, u brontosaurio de sangre calientenecesitaría un volumen cerebral más pequeño que un reptil heliotrópico paracontrolar su temperatura corporal.Jerison advierte que no se debe llegar precipitadamente a la conclusión que laendotermia, por sí sola, requiere ucerebro agrandado. El control de unafisiología de sangre caliente sólorequiere un área muy pequeña delhipotálamo, los dinosaurios podían ser endotermos aun teniendo cerebrosdiminutos. De forma similar, los

terápsidos probablemente desarrollarouna fisiología de sangre caliente duranteel pérmico tardío, a pesar de continuar teniendo cerebros pequeños que nollenaban sus cajas craneanas. Losmamíferos primitivos (triásicos)heredaron estos cerebros pequeños ysólo mucho más tarde desarrollaron elcerebro. Una vez más parece que laendotermia se desarrolló antes de quehubiera un cerebro agrandado.
La evolución de un cerebroaumentado de tamaño se correlacionamejor con el desarrollo de lasofisticación de la conducta. No es unacasualidad que los pterosaurios ydromeosaurios de cerebros grandes

fueran notablemente sofisticados en susrespectivas maneras. En lugar de que laendotermia exija la presencia de ucerebro grande, más bien es al revés:sólo un endotermo de metabolismorápido puede llevar a cabo la complejaconducta que observamos, por ejemplo,en las complicadas maniobras de vuelode los pterosaurios o la capacidad demanipulación y conservación delequilibrio de los dromeosaurios. Estasconductas complejas se desarrollaconcomitantemente con los controles decoordinación del cerebro. La endotermia posiblemente sea uno de los requisitos previos para la expansión del cerebro yno viceversa.

Un problema relacionado, aunque nocrucial para la tesis general, es lareconciliación de la tendencia a formar manadas de los dinosaurios con suscerebros pequeños. Los estrutiomímidos dromeosáuridos, sin duda, era
capaces de formar una organizaciósocial (quizá incluyendo una jerarquíasocial como la de las aves), proporcionar cuidados a sus crías, etc., pero ¿es el diminuto cerebro de losTriceratops un argumento en contra dela teoría de que era un animal demanada?
Un leit-motiv del libro ha sido lainextricable relación existente entre laendotermia y el tamaño de los

dinosaurios: los dinosaurios carentes deaislamiento estaban obligados a ser titanes y por esta razón, quedaroexcluidos de los micronichos ecológicosque fueron dominados por los mamíferos los lagartos. Pero, si los dinosaurios
no eran capaces de reducir su tamaño por debajo de un nivel crítico, ¿cómo podemos explicar que sus descendientescomenzaran su vida por debajo de estenivel umbral sin perecer debido a laexposición? Un dinosaurioindudablemente relacionado con sushuevos es el pequeño Protoceratops
mongol. Es significativo que los huevoseran muy grandes (mucho más grandesrelativamente que los de los reptiles), de

forma que los descendientescomenzaban su vida con un tamañosuperior al de muchos mamíferos ylagartos contemporáneos. El momentode la eclosión del huevo también habríacoincidido con la época más templadadel año a fin de reducir la severidad dela explosión[6]. Queda por ver si estosargumentos son lo suficientementesólidos como para responder a lasobjeciones.
Resulta costoso mantener lacondición de sangre caliente; requiere lacombustión de una gran cantidad decombustible. Un hombre consumeaproximadamente cuarenta veces másalimento que un lagarto del tamaño de u

hombre y alrededor de un 80-90% de laenergía liberada se utiliza para mantener una temperatura constante. Por lo tanto,un brontosaurio necesitaría, debido a stamaño, más comida que un lagarto deltamaño de un brontosaurio: quizá mediatonelada de vegetación al día. Esto plantea la intrigante pregunta de cómo podía pasar tanta comida a través de unacabeza que, como en el caso del
rontosaurus, era del tamaño de unavértebra. A continuación, para liberar laenergía, la comida debía ser descompuesta mecánicamente lo quehace que aún sea más misteriosa laausencia de dientes para moler. El
iplodocus sólo poseía unos cuantos

«incisivos» cortantes. Probablementetenían piedras estomacales quesustituían a los dientes para moler losalimentos.
Finalmente, aunque tampoco sea unacuestión crucial para la tesis central,está el persistente problema de laausencia de todo dinosaurio en el período poscretáceo. Si el frío era elresponsable de su aniquilación, inclusoacompañando a la explosión inicial deradiación procedente de una supernovacercana, ¿por qué no sobrevivierodinosaurios en las regiones ecuatorialesmás cálidas? ¿Es posible que todo el planeta estuviera sometido a un intensofrío? Parece improbable. Aun cuando

las dos masas de tierra en el hemisferioorte, América y Eurasia, estaba
situadas en latitudes medias y altas en el período cretáceo tardío, y lindaban en elsur con el mar de Tethis, América delSur y África, climáticamente estabasituadas más favorablemente. La brusquedad, ubicuidad y gravedad de lasextinciones cretáceas hace cada vez másdifícil descartar una teoría cataclísmica.
«El destino habitual de las nuevasverdades —observó el irascible Huxley,que conocía bien las estrangulacionescientíficas— es comenzar como herejías acabar como supersticiones».aturalmente, Huxley estaba pensando
en la herejía darwinista de la cual él era

el máximo exponente. Y a pesar de quesu opinión del mundo a la larga cayó ela ortodoxia, nunca perdió las ganas deluchar. La cause célebre de la década de1970 posiblemente no tenga lasconnotaciones morales tan queridas delos científicos Victorianos, pero unoapenas se habría enterado de ellodebido a las apasionadas protestas. Lossosegados museos una vez más se haceeco de las voces de descontento. Elcoloso Victoriano es una bestiademasiado querida como para renunciar a ella sin luchar. Tal como dijo C. P.Snow, «los arranques de cólera se hadado en ambos bandos». Pero ¿dóndeestá la belleza del dinosaurio ortodoxo?

Retrasado, minusválido y con la estéticade sus reliquias congeladas de losmuseos apenas evoca la era doradacuando reinaban los gigantes. Loscríticos insisten en que cada argumentoen favor de la endotermia se presta a unaexplicación alternativa. (Así, por ejemplo, las plumas podrían habersedesarrollado para proteger a losectotermos del sol abrasador en losdesiertos). Estoy de acuerdo en que cadaargumento tiene su interpretacióalternativa. Pero esto pasa por alto el punto crucial. La hipótesis de la sangrecaliente es una amplia red que abarcauna impresionante serie de hechos que,de otra forma, serían inconexos. Puede

explicar (aunque por el momentotoscamente) la decisiva dominación delos dinosaurios, su tamaño, su postura,la incesante supresión de los mamíferos,el origen de las plumas y de las aves, lasupremacía aérea de los pterosaurios,etc. Esto es, en efecto, una síntesisextraordinaria, tantas cuestionesaparentemente explicadas mediante unaúnica teoría. Y es éste el logrocientífico más importante de la teoría.Como dijo Darwin al final de El origen
de las especies, es difícil imaginarseque una teoría errónea explique unaserie de hechos tan numerosos ydiversos con tanta facilidad. Este puntode vista de los dinosaurios tambié

entraña grandiosidad. No obstante persiste la tergiversación coronada por la reciente afirmación de que losvestigios mortales de los dinosaurios,sencillamente, son inadecuados pararevelar su fisiología. Sin embargo,incluso huesos de 70 millones de añosde edad tienen una historia que contar. Yla Historia enseña el puro disparate de prescribir límites a nuestroconocimiento. En efecto, la ironía máscruel de la Historia, tras la imprudentedemanda de Comte de que debíamoscontinuar ignorando para siempre laquímica estelar, proporcionaríainmediatamente a los físicos el análisisespectral que sondearía la composició

de los lejanos cielos. Hoy también nosencontramos en el umbral: nuestroherético dinosaurio que es la clave de lacompleja red de vida ancestral de u pasado igualmente remoto.

GLOSARIO
alcaloide.— Sustancia químicaorgánica sintetizada por las plantas coflores (incluye a la morfina y estricnina).
amonita.— Molusco con tentáculosemparentado con el calamar, pero couna concha enrollada y multicameral.Muy abundante en los mares delmesozoico.
anquilosaurio.— Dinosaurioacorazado como un tanque.Ornitisquiano cuadrúpedo.
arcosaurio.— Agrupamiento devertebrados superiores basado edeterminadas características

esqueléticas compartidas. Comprende adinosaurios, pterosaurios, aves,cocodrilos y tecodontes.
arenisca roja nueva.— Rocas deltriásico.
batracio.— Anfibio.belemnita.— Molusco co
tentáculos emparentado con el calamar,con una protección o concha en forma de bala.
bípedo.— Animal que está erguido ycamina sobre sus miembros traseros.
brontosaurio.— Dinosauriosaurisquiano, cuadrúpedo, gigante (por ejemplo Brontosaurus).
bulbo olfatorio.— Región delcerebro anterior de los vertebrados

relacionado con el olfato.camptosaurio.— Dinosaurio
ornitópodo primitivo del jurásicosuperior de América del Norte.
ceratópsido.— Dinosaurioornitisquiano cornudo. El grupo florecióa finales del cretáceo (por ejemploTriceratops, Protoceratops).
celurosaurio.— Uno de losdinosaurios terópodos. Loscelurosaurios eran dinosaurios bípedos,veloces, con una constitución ligera. Sesabe que los ornitomímidos teníacerebros más grandes de lo corriente.Estos dinosaurios tenían un aspectosemejante al de las aves y eran erealidad los antepasados de las aves

(por ejemplo Ornithomimus,
Compsognathus).cerebelo.— Región del cerebro
posterior de los vertebrados, estárelacionado con la coordinación delmovimiento.
cetáceo.— Ballena.cetiosaurio.— Saurópodo.cicádidas o cicadáceas.— Planta
semejante a las palmeras con un troncoúnico y una corona de hojas similares alas de los helechos que floreció en eltriásico y jurásico. Está representada por muy pocos géneros supervivientes.
clavícula.— Hueso del cuello.cocolito.— Planta planctónica
microscópica, unicelular, con una

concha dura. Cuando estos diminutosorganismos mueren, la concha sedeposita en el fondo del mar, dondefinalmente se hace compacta y formacreta.
convergencia.— Las criaturas quehan adquirido un aspecto similar aunquesus antepasados hayan sidocompletamente diferentes (y sólo estélejanamente emparentadas) sedenominan convergentes. Un bueejemplo son el delfín mamífero y elictiosaurio reptil. Los animales quedesarrollan estructuras similares lohacen como respuesta a unas demandassimilares del estilo de vida.
cretáceo.— Último período del

mesozoico, hace 140-64 millones deaños.
cuadrúpedo.— Animal que está de pie y anda sobre cuatro patas.
cursorial .— Con extremidadesesbeltas adaptadas para correr, adiferencia de graviportal.
Dánico.— Estratos más primitivosdel terciario.
de sangre fría.— Animales cuyatemperatura interna puede variar con latemperatura ambiente. Cuando latemperatura externa es fría, latemperatura interna disminuye y elanimal se vuelve perezoso. Cuando estascriaturas de sangre fría toman el sol, stemperatura se eleva por encima de la

del aire (por ejemplo lagartos,cocodrilos, anfibios). Véase ectotermia.
Devónico.— Período del paleozoico, hace 395-345 millones deaños. Aparición en tierra del primer pez.
dicinodonto.— Terápsido o reptilsemejante a los mamíferos. Losdicinodontos eran herbívoroscorpulentos, abundantes en el pérmico ytriásico, que posiblemente vivían emanadas. Muchos presentaban colmillossemejantes a las morsas.
Diplodocus.— es el género,iplodocus camegiei la especie. Puede
haber más de una especie deiplodocus.
Diplodocus camegiei.— es una

especie. Pueden haber muchas especiesdentro de un género ( Diplodocus es elgénero).
dromeosáurido.— Uno de losdinosaurios terópodos. Poco conocido.Muy ágil, con un gran cerebro y enormesojos. Principios hasta finales delcretáceo (por ejemplo Deinonychus ySaurornithoides).
ectotermia.— Dependencia de unafuente de calor externa (el Sol) paraelevar la temperatura interna. Véase desangre fría.
endotermia.— Dependencia de lasfuentes de calor internas; calor generado por los músculos y los tejidos. El animalse conserva caliente a pesar de las

fluctuaciones de la temperaturaambiente. Véase de sangre caliente.
eoceno.— Época en el períodoterciario, hace 55-37 millones de años.
especie.— Unidad más pequeña dela clasificación de uso corriente.
estegosaurio.— Dinosaurioornitisquiano, cuadrúpedo con una seriede escamas óseas a lo largo de la líneacentral del dorso.
Estratos de Maastricht .— Estratosde finales del cretáceo.
etosaurio.— Reptil tecodonte deltriásico con un morro semejante al delos cerdos. Cuadrúpedo terrestre queremovía la tierra buscando raíces.
falange.— Hueso de los dedos de la

mano o de los pies. fémur .— Hueso de la pierna. fisiología.— Ciencia que se ocupa
de los procesos de la materia viva y delas funciones de los órganos y tejidos.
fitosaurio.— Tecodonte acuático delTriásico con un largo morro semejanteal de los cocodrilos.
foraminífero.— Organismo planctónico unicelular con una conchadura
fósil .— Vestigios petrificados de uanimal o planta. Cuando se acumulan lossedimentos que forman las rocas,atrapan las partes duras de los animales de las plantas (y, a veces, si los
sedimentos son lo bastante finos,

vestigios de tejidos más blandos),conservándolos.
gastrolito.— Piedra usada paramoler los alimentos en la molleja oestómago.
género.— Unidad de clasificación.Grupo de especies emparentadas.
graviportal .— Animales pesados deconstrucción robusta, frecuentementecon patas en forma de columnas.Generalmente andan muy despacio (por ejemplo los elefantes y brontosaurios).
hemisferios cerebrales.— Par deexcrecencias en el cerebro anterior delos vertebrados, centro de coordinaciósuperior.
hueso neumático.— Hueso hueco,

en vida lleno de sacos aéreos. Seencuentra en los pterosaurios, las aves yalgunos dinosaurios.
hueso pteroideo.— Pequeño huesoque sobresale en la muñeca de los pterosaurios. Probablemente sosteníauna membrana que iba al cuello yactuaba como elevador.
húmero.— Hueso del brazo.ictiosaurio.— Reptil marino, co
forma de delfín que vivió durante el período Mesozoico.
iguana.— Lagarto tropical.iguanodonto.— Dinosaurio
ornitisquiano, bípedo, herbívoro. jurásico.— Período medio del
mesozoico, hace 190-140 millones de

años.laberintodonto.— Anfibio
acorazado, cuadrúpedo, rechoncho,corriente en el pérmico y triásico.
Lamarck .— Lamarck promulgó umecanismo de evolución (1744-1829).Creía que el uso incrementado de unaestructura provocaba que ésta creciera.Actualmente remplazado por elneodarwinismo, que implica lasmutaciones y la selección natural.
Lías.— Comienzos del jurásico.lóbulo óptico.— Región del cerebro
medio de los vertebrados relacionadocon la visión.
Mar de Tethis.— Oceáno que seextendía en dirección Este-Oeste durante

el mesozoico. Tenía un área muy extensa separaba los supercontinentes Norte y
Sur.marga.— Caliza fangosa.marsupial .— Mamífero con bolsa
(por ejemplo opossum, canguro, lobomarsupial).
médula.— Estructura situada en la parte posterior del cerebro de losvertebrados.
mesozoico.— Una de las principaleseras geológicas, hace 215-64 millonesde años, constituida por tres períodos, eltriásico, jurásico y cretáceo.Coloquialmente nos referimos a ellacomo la «Era de los reptiles».
metabolismo.— Este término se

refiere a los procesos químicos quetienen lugar en el cuerpo, especialmentela descomposición de los alimentos paraliberar energía. Un metabolismo rápidosupone la capacidad de liberar másenergía y a una mayor velocidad. Estoimplica una mayor cantidad de oxígeno para «quemar» los alimentos y unaaceleración de las reacciones químicasen los tejidos. La velocidad metabólicadepende de la temperatura; lasreacciones son más lentas cuando latemperatura desciende por debajo de u punto óptimo para las enzimascatalizadoras. La mayoría de los reptilesmodernos son capaces de tener umetabolismo rápido durante breves

espacios de tiempo, pero son incapacesde mantenerlo durante espacios más prolongados. Los mamíferos y las aves,en cambio, pueden mantener un nivelalto de producción de energía y, por lotanto, son capaces de tener una mayor actividad.
metacarpiano.— Hueso largo en la palma de la mano, situado entre lamuñeca y los dedos.
metatarsiano.— Hueso largo en lasuela del pie, situado entre el tobillo ylos dedos.
miembros desparramados.— Extremidades extendidas a ambos ladosdel cuerpo, los huesos de la partesuperior de las extremidades (fémur y

húmero) están paralelos al suelo.moa.— Ave grande, no voladora,
actualmente extinguida, de Australasia(por ejemplo Dinomis).
molleja.— Estómago muscular elas aves y posiblemente también en losdinosaurios. Tiene una fuerte paredmuscular; a veces contiene gastrolitos.La comida puede ser tragada sin haber sido masticada (como en las aves) y lamolleja se encargará de moler la comidahasta formar una pulpa antes de pasar alestómago.
monitor .— Lagarto tropical osubtropical que vive en el Viejo Mundo, puede alcanzar un gran tamaño (por ejemplo el dragón de Komodo en las

Indias Orientales). Emparentado con elmosasaurio cretáceo.
monofilia.— Origen único de ugrupo de animales (las aves so probablemente monofiléticas, losmamíferos no lo son: los monotremas sedesarrollaron a partir de los terápsidos,independientemente de los mamíferos placentarios y marsupiales).
monotrema.— Mamífero muy primitivo, ovíparo, con escasacapacidad para perder calor. Sedesarrolló independientemente de otrosmamíferos a partir de los terápsidos, por ejemplo el platypus de pico de pato.
nicho.— Tipo de medio ambiente preferido por un animal. Por ejemplo las

llanuras para los corredores de largosmiembros, malezas para las musarañas,etc.
oligoceno.— Época del períodoterciario, hace 37-27 millones de años.
oolito.— Jurásico medio. Oolitosignifica «piedra de huevo» y muchas delas calizas de color claro estácompuestas por pequeños granitos que parecen huevas de pez. Las rocasoolíticas del jurásico están distribuidasa lo largo de Inglaterra, desdeYorkshire, en el Noreste, hasta Dorset eel Sudoeste. La pizarra de Stonesfield,que albergaba el Megalosaurus, es parte de la serie de los oolitos.
opistótonos.— Trastorno nervioso

por envenenamiento que provocaespasmos musculares y frecuentementela muerte. Los músculos se vuelverígidos y el animal muere en una posturacontorsionada. Puede estar producido por alcaloides como la estricnina.
ornitisquianos.— Uno de los dosgrupos principales de dinosaurios;comprende a los ornitópodos,estegosaurios, anquilosaurios yceratópsidos.
ornitópodo.— Dinosaurioornitisquiano, herbívoro, bípedo (por ejemplo Iguanodon, Hadrosaurus e ypsilophodon).
paladar secundario.— Huesos queforman el techo de la boca en los

mamíferos, cocodrilos y algunosdinosaurios, separando la boca de lacavidad nasal.
Paleontología.— El estudio de lavida pasada tal como se revela por losregistros fósiles. Implica lareconstrucción del aspecto de losorganismos primitivos, su forma defuncionar, ecología y parentescos cootras criaturas.
paquidermo.— Término queactualmente se aplica a los grandesmamíferos de piel gruesa, no rumiantes,especialmente los elefantes.
pérmico.— Último período del paleozoico, hace 280-215 millones deaños. Dio paso al triásico.

pes.— Pie. placentario.— Mamífero co
placenta (todos los mamíferossupervivientes excepto los marsupiales los monotremas).
plancton.— Organismos marinos pequeños de diverso tipo que sotransportados por las corrientes.
plantígrado.— Andar sobre lassuelas de los pies; a diferencia de andar sobre los dedos (digitígrado).
plesiosaurio.— Reptil marino o deagua dulce del Mesozoico con aletas.
Plioceno.— Última época del período terciario, hace 12-2 millones deaños.
polifilia.— Orígenes múltiples de u

grupo de animales: véase monofilia. preadanita.— Término común en el
siglo XVIII y comienzos del siglo XIX
para los animales y las plantas que,según se creía, habían vivido en épocasanteriores a Adán.
preadaptación.— Desarrollo de unaestructura en un medio ambiente (por ejemplo los pulmones en un pez), queresulta ser aún más útil en otro (por ejemplo en tierra).
prediluviano.— Término utilizadocomúnmente en el siglo XVIII y a principios del siglo XIX para describir los artefactos (especialmente fósiles)que se pensaba procedían de una fechaanterior a la Inundación Universal de

oé. prosaurópodo.— Dinosaurio
saurisquiano primitivo del triásico.Incluye tanto a las formas bípedas comoa las cuadrúpedas.
pseudosuquiano.— Tecodonte deltriásico. A veces bípedo, aunquetambién existían formas cuadrúpedas.Aquí encontramos el desarrollo de una postura completamente erguida por primera vez en la Historia de losvertebrados. Antepasado de losdinosaurios.
radio.— Hueso del antebrazo. Roentgen.— Nombre que recibe por
el descubridor de los rayos X WilhelRöntgen (1845-1923), es la medida

estándar de la radiación. La vida sobrela tierra está expuesta anualmente a pocomás de 0,1 roentgens que proceden delsuelo, la atmósfera e incluso ladesintegración del potasio y carbonoradiactivo en los tejidos vivos. Laradiación cósmica adopta la forma derayos X de alta energía y rayos gamma;cuando están en exceso provocan ladesintegración celular en los tejidos yoriginan cánceres. Las célulasreproductivas son especialmentevulnerables y la irradiación puede provocar esterilidad o el nacimiento demutantes.
sacos aéreos (sacos pulmonares).. — Extensión de los pulmones a través

de la cavidad corporal de las aves, pterosaurios y quizás algunosdinosaurios. Estos sacos llenos de aire pueden penetrar en los huesos.
sacro.— Vértebras en la regió pélvica, fundidas entre sí y unidas con elcinturón pélvico.
saurio.— Reptil (término empleadoa principios del siglo XIX).
saurisquianos.— Uno de los dosgrupos principales de dinosaurios quecomprende a los terópodos (todoscarnívoros) y saurópodos(brontosaurios, etc).
saurópodo.— Dinosauriossaurisquianos, cuadrúpedos, gigantesque tuvieron su apogeo durante el

urásico pero que sobrevivieron hastafinales del cretáceo (por ejemplo
rontosaurus y Diplodocus). supernova.— Explosión de una
estrella.tanino.— Sustancia química
orgánica sintetizada por las plantas. Escorriente en la corteza de los árboles.
tecodonte.— Reptil triásico codientes con alvéolos; comprenden a los pseudosuquianos, etosaurios yfitosaurios.
terápsido.— Reptil semejante a losmamíferos. Entre los terápsidos seencontraban los antepasados de losmamíferos. Los terápsidos fueron losanimales terrestres dominantes a finales

del pérmico y comienzos del triásico, pero fueron expulsados por lostecodontes seudosuquianos.
terciario.— Período que sigue alcretáceo. Coloquialmente nos referimosa éste como la «Era de los mamíferos»,hace 64-2 millones de años.
terópodo.— Dinosauriosaurisquiano, carnívoro. El grupocomprende a los carnosaurios (por ejemplo Tyrannosaurus), celurosaurios(por ejemplo Ornithomimus) ydromeosaurios (por ejemplo
einonychus).triásico.— Primer período del
mesozoico, hace 215-190 millones deaños.

ventrículo.— Cámara en el corazóresponsable de bombear la sangre alcuerpo y a los pulmones.
vértebras cervicales.— Vértebrasdel cuello.
Wealden.— Depósitos del cretáceoinferior. El nombre Wealden se derivadel «Weald» (bosque) de Kent donde fueestudiado por primera vez. Estácompuesto de areniscas y arcillas queoriginalmente se depositaron a principios del cretáceo, hace más de100 millones de años, en un enormeestuario de agua dulce o salobre que seextendía por todo el sur de Inglaterra. El guanodon y el Hylaeosaurus vivieroen la época del Wealden o

wealdeniense.

BIBLIOGRAFÍAGENERAL
Alan Charig y Brenda Horsfield,1975. Before the Ark . BBC, Londres.
Edwin H. Colbert, 1962. Dinosaurs:
Their Discovery and their World .Hutchinson, Londres.
Edwin H. Colbert, 1965. The Age o
eptiles. Weidenfeld & Nicolson,Londres.
Edwin H. Colbert, 1969. Evolution
of the Vertebrades. 2.a ed., Wiley,ueva York.
Edwin H. Colbert, 1971. Men an

inosaurs: The Search in Field an
aboratory. Penguin.Edwin H. Colbert, 1974. Wanderin
ands and Animals. Hutchinson,Londres.
L. B. Halstead, 1969. The Pattern o
Vertebrate Evolution. Oliver y Boyd,Edimburgo.
L. B. Halstead, 1975. The Evolution
and Ecology of Dinosaurs. Eurobook.Url Lanham, 1973. The Bone
unters. Columbia University Press,ueva York y Londres.
Alfred Sherwood Romer, 1966.Vertebrate Palaeontology. 3.a ed.,University of Chicago Press, Chicago yLondres.

Alfred Sherwood Romes, 1971.an and the Vertebrates. 2 vols.,
Penguin.M. J. S. Rudwick, 1972. The
eaning of Fossils: Episodes in the
istory of Palaeontology. Macdonald,Londres; American Elsevier, NuevaYork.
Harry Govier Seeley, 1901.ragons of the Air . Methuen, Londres.
(Reimpresión 1967, Dover, bolsillo).Barbara J. Stahl, 1974. Vertebrate
istory: Problems in Evolution. MeGraw-Hill, Nueva York.
W. E. Swinton, 1934. The
inosaurs: A Short History of a Great
Group of Extinct Reptiles. Thomas

Murby & Co., Londres.W. E. Swinton, 1965. Fossil Birds.
2.a ed., British Museum (Historiaatural).
W. E. Swinton, 1969. Dinosaurs. 4.a
ed., British Museum, (Historia Natural).W. E. Swinton, 1970. The
inosaurs. George Allen & Unwin Ltd.,Londres; John Wiley & Sons, Inc.,
ueva York.Herbert Wendt, 1968. Before the
eluge. Doubleday & Co., Nueva York.

CRÉDITOS DE LASILUSTRACIONES
1. Faujas-Saint-Fond B., 1799.istoire Naturelle de la Montagne de
Saint-Pierre de Maestricht . Paris. Coel permiso de los administradores delMuseo Británico.
2. & 16. Illustrated London News:1853, 23 págs. 599-600.
1854, 24, pág. 22. Restauración por eave Parker, 1955, 227, pág. 1065.
3. Fotografía tomada especialmente para el libro por Keith Penny.
4. Restauración de Charles R.

Knight, en Osborn, H. F., 1931. Cope:
aster Naturalist . Princeton.5. Board of Commissioners of the
Central Park, 1868. Twelfth Annual
eport . Nueva York.6. Osborn, H. F., 1911. Amer. Mus.
our ., 11, págs. 7-11.7. Lambe, L., 1920. Canada Dept.
Mines, Geol. Surv., 120 págs. 1-79.8. Restauración de Robert T. Bakker,
en Russell, D. A., 1973. Cañad. Geog.
., 87, págs. 4-11.9. Matthew, W. D., 1915. Dinosaurs.
ueva York: Museo Americano deHistoria Natural.
10. Matthew, W. D., 1915.inosaurs. Nueva York: Museo

Americano de Historia Natural.11. Bird, R. T., 1954, National
Geographic, 105, págs. 707-722.12. 13. 14 & 19. Restauración de
Robert T. Bakker en Crompton, A. W.,1968. Optima, 18, págs. 137-151.
15. Kermack, K. A., 1965. Science
ournal , 1 (9), págs. 66-72.17. Holland, W. J., 1905, Mem.
Carnegie Mus., 2, Págs. 225-279.18. Bird, R. T., 1944. Natural
istory, 53 (2), págs. 60-67. 20.Heilmann, G., 1926. The Origin o
irds. Londres.12 & 23. Augusta, J., y Burian, Z.,
1962. Prehistoric Reptiles and Birds.Londres.

22. Broili, F., 1938. Sitzgsb. d.
math.-naturw. Abt ., págs. 139-154.24. Newman, E., 1843. The
Zoologist I. págs. 129-131.25. Preparado especialmente para el
libro por Jessica Gwynne.26. Osbom, H. F., 1917. Bull. Am.
us. Nat. Hist ., 35, págs. 733-771.

CRÉDITOS DE LOSDIBUJOS
a. Owen, R., 1854. Geology an
nhabitants of the Ancient World .Londres.
b. Heilmann, G., 1926. The Origin
of Birds. Londres.c. Matthew, W. D., 1915. Dinosaurs.
ueva York: Museo Americano deHistoria Natural.
d. & e. Restauraciones de Robert T.Bakker, en Ostrom, J. H., 1969. Bull.
eabody Museum of Natural History,
Yale University, 30, págs. 1-165.

f. Gabon, P. M., 1970. /. Paleo. 44, págs. 464-473.
g. Izquierda: Gabon, P. M., 1970. J.
aleo. 44, págs. 464-473. Derecha:Russell, D. A., 1972. Can. J. Earth Sci.,9. págs. 375-402.
h. Heilmann, G., 1926. The Origin
of Birds. Londres.i. Restauración de Robert T. Bakker,
en Gabon, P. M., 1974. Bull. Br. Mus.
nat. Hist. (Geol.), 25 (1), págs. 1-152. j. Romer, A. S., 1971. Breviora,
378, págs. 1-10.k. Hay, O. P., 1910. Proc. Wash.
cad. Sci., 12, págs. 1-25.1. & m. Holland, W. J., 1910. Am.
at ., 44, págs. 259-283.

n. Heilmann, G., 1926. The Origin
of Birds. Londres.o. Heilmann, G., 1926. The Origin
of Birds. Londres. p. Matthew, W. D., y Granger, W.,
1917. Bull. Amer. Mus. Nat. Hist ., 37, págs. 307-326.
q. Phillips, S., 1856. Guide to the
Crystal Palace and Park . Londres, pág.190.
r. Edinger, T., 1941. Am. Jour. Sci.239, págs. 665-682.
s. Eaton, G. F., 1910. Mem.
Connecticut Acad. Sci., 2, págs. 1-38.

ADRIAN DESMOND (Reino Unido,1947). Adrian John Desmond es uescritor especializado en Historia de laCiencia. Estudió Fisiología en laUniversidad de Londres y continuó susestudios en Historia de la Ciencia yPaleontología de los Vertebrados en laUniversidad Colegio de Londres, y

posteriormente en la Universidad deHarvard. Sus estudios de doctoradotuvieron como tema el contextovictoriano de la Teoría de la Evolució propuesta por Darwin. Desmond esInvestigador Honorario delDepartamento de Biología de laUniversidad Colegio de Londres.
El primer libro de Desmond, The Hot-
looded Dinosaurs ( Los dinosaurios de
sangre caliente) (1975), fue traducido asiete idiomas. Siendo el primer libro afavor de que los dinosaurios eran desangre caliente y dieron lugar a las aves,fue ampliamente divulgado y recibiónumerosos elogios. Basándose en el

libro, la BBC realizó un documentaltitulado también The Hot-Bloode
inosaurs. Su segundo libro dedivulgación científica, The Ape’s
eflexion (1979), sobre losexperimentos con chimpancés usando ellenguaje de signos, fue elegido comouno de los mejores libros del año por sychology Today. Seguidamente,
Desmond publicó dos libros decontenido académico: Archetypes an
ncestors (1982), que trata del papelque jugaron los fósiles en los debatessobre la evolución en los tiempos deDarwin, y The Politics of Evolution
(1989), que analiza el contexto en que sedesarrollaron las teorías de la evolució

antes de Darwin, y que ganó el PremioPfizer en 1991 de la History of ScienceSociety (EEUU). Darwin (1991), escrito por Desmond y Moore, recibió el premio James Tait Black, el premioComisso en Italia, el premio WatsoDavis de la History of Science Society yel Premio Dingle de la British Societyfor the History of Science. Su siguiente biografía, Huxley: From Devil’s
isciple to Evolution’s High Priest
(dos volumenes 1994-7), fue uno de losmejores libros de 1997 según el New
York Times, y fue escogido como uno delos mejores libros de todos los tiempos por la Science Books and Films (1999).

Desmond ha escrito numerosos artículoscientíficos sobre la historia de lasteorías evolucionistas, así como lasentradas del Oxford Dictionary o
ational Biography de Darwin (con JiMoore y Janet Browne) y Huxley, y laentrada de la Enciclopedia Británica deDarwin.
Desmond es investigador de laZoological Society of London, miembrode la British Society for the History oScience, la History of Science Society(EEUU), y la Society of VertebratePalentology (EEUU). En 1993 le fueconcedida la Medalla del Fundador dela Society for the History of Natural

History. En la actualidad vive eBerkshire, Inglaterra.

Notas
I. LA CUMBRE DE LA CREACIÓN

[1] Faujas-Saint-Fond, B., 1799. Histoire
aturelle de la Montagne de Saint-
ierre de Maastricht . París, págs.59-67. Este relato sobre el mosasauriode Maastricht fue escrito durante la erarevolucionaria por un profesor deGeología del museo de París y,naturalmente, apoya al científicoHoffmann y describe al clérigo Godicomo el villano.<<

[2] Para un informe sobre Cuvier,Lamarck y la extinción véase: Rudwick,M. J. S., 1972. The Meaning of Fossils.Londres y Nueva York. El trabajozoológico de Cuvier es tratado en laobra de Coleman, W., 1964. Georges
Cuvier: Zoologist . Cambridge, Mass. Elconcepto del plenum y su influenciasobre el pensamiento científico,especialmente en lo referente a la ideade la extinción se puede encontrar en:Lovejoy, A. O., 1964. The Great Chain
of Being . Cambridge, Mass., capítulos 8 9.
Cuvier demostró no sólo la desaparició

de los mastodontes sino que continuóinvestigando al Megatherium
paraguayo, o perezoso terrestre gigante, al mamut siberiano. Cuvier sugirió que
se había producido una revolucióviolenta en el pasado reciente de laTierra que había exterminado gran partede la vida animal. Cuando Cuvier supode la existencia de reptiles mesozoicosmás remotos modificó su teoría,diciendo que había habido repetidascatástrofes y que, tras cada una de éstas,había aparecido una fauna nueva deaspecto más moderno. El progresionismo de Cuvier tuvo que ser rectificado tantas veces con eldescubrimiento de un número creciente

de animales, extinguidos que, en 1849,a se necesitaban 29 catástrofes.aturalmente, cada creación nueva se
iba asemejando cada vez más a lasiguiente, de forma que, finalmente, elconjunto del edificio se derrumbó bajosu propio peso. Fue remplazado por lateoría de la evolución de Darwin e1859. Véase: Bourdier, F., 1969.Geoffroy Saint-Hilare versus Cuvier: lacampaña en favor de la evolució paleontológica (1825-1838). In Schneer,C. J. (ed.), 1969. Toward a History o
Geology. Cambridge, Mass., págs.36-61. Esp. pág. 60.<<

[3] Cuvier, G., 1836. Recherches sur les
Ossimens Fossiles. 4.a ed., París, pag.175.<<

[4] Buckland, W., 1837. Geology an
ineralogy. 2.a ed., vol. 1, pág. 217.<<

[5] Ibid ., pág. 220<<

[6] Buckland, W., 1824. «Notice on theMegalosaurus or great fossil lizard oStonesfield». Trans. Geol. Soc. Londres,serie 2, vol. I, págs. 390-96.<<

[7] Mantell, G., 1825. «Notice on theIguanodon, a newly discovered fossilreptile, from the sandstone of Tilgateforest, in Sussex». Phil. Trans. Roy.
Soc., 115, págs. 179-86. Las opinionesde Mantell sobre estos grandes«Lagartos Fosilizados», como losllamaba, se pueden encontrar en:Mantell, G., 1833. The Geology of the
South East of England . Londres. Estelibro también contiene el primer informe publicado del Hylaeosaurus
desenterrado el año anterior en el bosque de Tilgate.<<

[8] Buckland, W., 1835. «On thediscovery of fossil bones of theIguanodon, in the iron sand of theWealden formation in the Isle of Wight,and in the Isle of Purbeck». Trans. Geol.
Soc. Londres, serie 2, vol. 3, págs.425-32.<<

[9] Once años antes, en 1830, Hermanvon Meyer ya se había anticipado aOwen. Meyer había dividido los reptilesfosilizados y había agrupado al
egalosaurus e Iguanodon debido asus pies paquidérmicos. Definió a estos«paquipodos» como «saurios coextremidades semejantes a las de losmamíferos terrestres voluminosos».Owen, R., 1874-1889. Monograph o
the Fossil Reptilia of the Mesozoic
ormations. Sociedad Paleontográfica, págs. 201, 595 et seq. El gra protagonista de Owen, Thomas HenryHuxley, insistió en otorgar a Meyer el

crédito de haber apreciado laimportancia del grupo con gran disgustode Owen. De hecho, como arguyó Owen,la clasificación de Meyer erasolamente un orden tabular y Meyer nohabía publicado sus razones paraagrupar a los dinosaurios. Owen, por otro lado, se había esforzado mucho para descubrir las diferenciasanatómicas existentes entre losdinosaurios y otros reptiles.<<

[10] Owen, R., 1841. «Report on BritisFossil Reptiles». Report of the Eleventh
eeting of the British Association for
the Advancement of Science, págs.60-204. Véase especialmente la pág.103.<<

[11] Ibid ., págs. 142-3.<<

[12] Ibid ., pág. 110.<<

[13] Ibid ., pág. 200.<<

[14] Lamarck. J. B., 1809. Philosophie
Zoologique. París. Traducido al ingléscomo Zoological Philosophy por Elliot,H. (1914, Londres; 1963, Nueva York).Véase el prólogo de la pág. 2 de latraducción de Elliot.<<

[15] Owen, op. cit . (10), pág. 196.<<

[16] Ibid ., pág. 202.<<

[17] Ibid ., pág. 204 (nota al pie de página).<<

[18] Desmond, A. J., 1974. «CentralPark’s fragüe dinosaurs». Natural
istory, 83 (8), págs. 64-71. Paraconsultar las fuentes originales, véaseHawkins, B. W., 1854. «On visualeducation as applied to geology».ournal of the Society of Arts, 2, págs.
444-9. Owen, R., 1854. Geology an
nhabitants of the Ancient World .Londres.<<

[19] The Illustrated London News, 31Die. 1853, No. 661. Vol. 23, págs.599-600. (El Crystal Palace deSydenham).<<

[20] Owen, obv. cit . (18).<<

[21] The Illustrated London News, 7 deenero de 1854, No. 662, Vol. 24, pág.22. (El Crystal Palace de Sydenham).<<

[22] The London Quarterly Review,1854, 3, (5), págs. 232-79. «La cena coel fósil» y la valoración de losmonstruos se encuentran en las páginas237-9. Para la reacción popular véaseThe Westminster Review, 1854, 62 (6), págs. 540-41.<<

[23] Dickens, C. J. H., 1853. Bleak
ouse, Londres.<<

[24] Bakker, R. T., 1968. «The superiorityof dinosaurs». Discovery (New Haven),3 (2), págs. 11-22. Véase la pág. 11.<<

[25] Bogert, C. M., 1959. «How reptilesregulate their body temperature».Scientific American, 200 (4), págs.104-20.<<

[26] Ibid .<<

[27] Colbert, E. H., Cowles, R. B., yBogert, C. M., 1946. «Temperaturetolerances in the American alligator andtheir bearing on the habits, evolution andextinction of the dinosaurs». Bull. Am.Mus. Nat. Histo., 86, págs. 331-73.<<

[28] Colbert, E. H., Cowles, R. B., yBogert, C. M. 1947. «Rates otemperature increase in the dinosaurs».Copeia, 2, págs. 141-2.<<

[29] Colbert, E. H., 1962. Dinosaurs:
Their Discovery and their World .Londres, pág. 207.<<

[30] Bogert, op. cit . (25).<<

[31] Ibid .<<

II. EL TIRANO ENCUENTRA SUS PIES

[1] Hayden, F. V., 1860. «Geologicalsketch of the estuary and fresh water deposit of the bad lands of the Judith,with some remarks upon the surroundingformations». Trans. Am. Philos. Soc.,11, págs. 122-38.<<

[2] Leidy, J., 1860. «Extinct vertebratafrom the Judith River and great ligniteformations of Nebraska». Ibid., págs.139-54. Este artículo, leído ante laSociedad Filosófica Americana, e1859, apareció tras la descripción del
adrosaurus de Leidy, a pesar de queLeidy había adquirido el Deinodon y elTrachodon algunos meses antes que elesqueleto del Hadrosaurus; simportancia se vio obviamenteincrementada por el descubrimiento delesqueleto.<<

[3] Leidy, J., 1858. « Hadrosaurus and itsdiscovery». Proc. Acad. Nat. Sci.,Filadelfia, 14 de diciembre, págs.213-18. Véase pág. 217.<<

[4] Ibid ., pág. 217.<<

[5] Carta de Cope a su padre, con fechadel 15 de agosto de 1866. Reproducidaen Osbom, H. F… 1931. Cope: Master
aturalist, The Life and Letters o
dward Drinker Cope, Princeton, pág.157.<<

[6] Cope, E. D., 1866. «Remains of agigantic extinct dinosaur». Proc. Acad.
at. Sci., Filadelfia, 21 de agosto, págs.275-9.<<

[7] Junta de administración del CentralPark, 1868. Twelfth Annual Report ,Apéndice F, págs. 125-41.<<

[8] Parece ser que el padre deWaterhouse Hawkins había sido amigode Charles Willson Peale, el fundador del museo de Historia natural deFiladelfia, en 1786. A principios delsiglo XIX, Peale descubrió y reconstruyóel primer esqueleto completo de umastodonte americano y, en 1802,muchos de los amigos de Peale loacompañaron en una cena en el interior
del esqueleto del «mamut ». Entre los presentes estaba el padre de Hawkins,un inventor cuyo «gran piano portátil patentado» había sido traído para lasfestividades. El mismo año, el padre de

Hawkins volvió a su Inglaterra nativa.La extraordinaria semejanza entre esteepisodio en Filadelfia y la cena que dioWaterhouse Hawkins en el interior del guanodon, en el Crystal Palace 52 añosmás tarde, nos lleva a la conclusión deque Waterhouse Hawkins había oído delacontecimiento de su padre. Agradezcola ayuda del biógrafo de Peale, CharlesColeman Sellers por haberme facilitadoesta información. Comunicació personal, 3 de noviembre de 1974.<<

[9] Desmond, A. J., 1974. «CentralPark’s fragile dinosaurs». Natural
istory, 83 (8), págs. 64-71.<<

[10] Marsh, O. C., 1890. Contestación al profesor Cope. New York Herald , 19 deenero.<<

[11] Marsh, O. C., 1877. «Notice of anew and gigantic dinosaur». Am. Joum.
Sci., 14, págs. 87-8.<<

[12] Cope, E. D., 1877. «On theVertebrata of the Dakota Epoch oColorado». Proc. Amer. Philos. Soc.,16, págs. 233-47.<<

[13] Olmstead se convirtió posteriormente en el patrón de laarquitectura de jardines. Fue elresponsable de la mayoría de grandes proyectos de la época, que incluyeron elHarvard Yard y el Central Park. Al igualque muchas otras personas cultas se puso a malas con la administración deTweed.<<

[14] Callow, A. B., 1966. The Twee
ing . Nueva York.<<

[15] The New York Times, 7 de marzo de1871, pág. 5.<<

[16] Wingate, C. F. 1875. «An episode imunicipal government». 3. «The RingCharter». North American Review, 111, págs. 113-55. La muerte del CentralPark se describe en las páginas 120-23.<<

[17] Junta de administración deldepartamento de parques públicos,1871. First Annual Report , págs. 18-20.Éste fue el primero (y único) informedel círculo de Tweed.<<

[18] Hawkins aparentemente habíaconstruido otro Hadrosaurus de tamañoreal antes de su estancia en Princeton.Probablemente, cuando su estudio delCentral Park estaba intacto. El director del Museo de Historia Natural, el Dr.Donald Baird, escribe: «Anteriormentea había montado en nuestro museo u
esqueleto de yeso del Hadrosaurus
oulkii (el primer esqueleto dedinosaurio que fue montado en un museouniversitario), y ahora, entre 1875 y1877, produjo una serie de 17 grandes pinturas de escenas prehistóricas. Estas pinturas eran el resultado directo de s

investigación para el Crystal Palace y para el Museo Paleozoico e incluían losmismos animales y otros. Durante treintaaños, estuvieron colgados desde las barandillas del balcón en Nassau Hall,en la estancia que sirvió sucesivamentecomo capilla, lugar de reunión para elCongreso Continental, biblioteca, museo (actualmente) como claustro».
«Quince de las originalmente diecisiete pinturas aún están colgadas en GuyotHall en diversos estados deconservación. Algunas están acabadas yfirmadas, algunas están sin firmar,algunas están inacabadas». Comen. pers.del 2 de noviembre de 1974.<<

[19] Schuchert, C., y Le Vene, C. M.,1940, O. C. Marsh: Pioneer in
alaeontology, New Haven, pág. 385.<<

[20] Huxley, T. H., 1868. «On the animalswhich are most nearly intermediate between birds and reptiles». Geol. Mag.5, págs. 357-65.<<

[21] Hitchcock, E… 1848. «An attempt todiscriminate and describe the animalsthat made the fossil footmarks of theUnited States, and especially of NewEngland». Memoirs of the American
cademy of Arts and Sciences, seriesnuevas 3, págs. 129-256. Véanse las páginas 250-51.<<

[22] Hitchcock, E., 1836.«Ornithichology. Description of the footmarks of birds (Ornithichnites) on newRed Sandstone in Massachusetts». Am.
oung. Sci., 29, págs. 307-40. Véase la página 313.<<

[23] Dollo, L., 1882. «Première note sur les dinosauriens de Bemissart». Bulletin
de Musée Royal d’Histoire Naturelle de
elgique, 1, págs. 161-80. Éste fue unode una larga serie de artículos. Dupont,El, 1870. «Sur la découverted’ossements d’iguanodon, de poissons etde végétaux dans la Fosse Saint-Barbedu Charbonnage de Bernissart». Bulletin
de l’Académie Royale, des Lettres et
des Beaux Arts de Belgique, segundaserie, 46, págs. 387-408.<<

[24] Brown, B., 1938. «The mysterydinosaur». Natural History, 41, págs.190-202, 235.<<

[25] Galton, P. M., 1974. «Theornithischian dinosaur Hypsilophodofrom the Wealden of the Isle of Wight».Bull. Br. Mus. nat. Hist. (Geol.), 25 (1), págs. 1-152.<<

[26] Osborn, H. F., 1911. «A dinosaur mummy». Amer. Mus. Jour ., 11, págs.7-11.<<

[27] Lambe, L. M., 1914. «On a newgenus and species of carnivorousdinosaur from the Belly River formatioof Alberta, with a description oStephanosaurus marginatus from thesame horizon». Ottawa Nat., 28, págs.17-20.<<

[28] Lambe, L. ML, 1920. The hadrosaur dmontosaurus from the Upper
Cretaceous of Alberta. Canada Dept.
ines. Geol. Surv., Memoria 120, págs.1-79.<<

[29] Para algunos recuerdos personales, yun relato de los episodios más pintorescos de la vida de Nopcsa,véase: Edinger, T., 1955. «Personalitiesin palaeontology: Nopcsa. Society oVertebrate Palaeontology»; News
ulletin, No. 43, págs. 35-9. Véasetambién Colbert, E. H., 1971. Men an
inosaurs. Pelican, pág. 128 ysiguientes.<<

[30] Nopcsa, F., 1929. «Sexualdifferences in ornithopodous dinosaurs».
alaeobiologica, 2, págs. 187-201. Lasdiferencias anatómicas en los animalesfosilizados del mismo tipo han sidoconsideradas frecuentemente comodimorfismos sexuales. Los pelicosaurioscomo el Dimetrodon, con una cresta ela espalda (sostenida por las espinasalargadas de las vértebras dorsales) hasido considerados como la expresiómasculina del Sphenacodon sin cresta.Colbert señala que el Dimetrodon
«macho» se encuentra en Texas y elSphenacodon «hembra» en Nuevo

México. Estas dos localidades estabaseparadas por un mar a principios delPérmico cuando hubo un apogeo deestos reptiles. Un «obstáculo paradesanimar a los reptiles más persistentesen busca de amoríos», añade Colbert.Colbert, E. H., 1969. Evolution of the
Vertebrates. Nueva York, página 131.<<

[31] Wilfarth, M., 1938. «Gab esriisseltragende Dinsaurier?». Zeits-chr.
eutsch. Geol. Ges., tomo 40, cuaderno2, págs. 88-100. Para la contestacióvéase: Sternberg, C. M., 1939. «Werethere proboscis-bearing dinosaurs?».
nn. Mag. Nat. Hist ., serie 2 (3), págs.556-60.<<

[32] Ostrom, J. H., 1962. «The cranialcrests of hadrosaurian dinosaurs».
ostilla, n.o 62, págs. 1-29.<<

[33] Ostrom, J. H., 1964. «Areconsideration of the paleocology ohadrosaurian dinosaurs». Am. Jour. Sci.,262, págs. 975-97.<<

[34] Halstead, L. B., 1969. The Pattern
of Vertebrate Evolution. Edimburgo, pág. 124.<<

[35] «Las patas delanteras soridículamente pequeñas —dice Romer en su manual estándar de laPaleontología de los vertebrados— y parecen haber sido prácticamenteinútiles; eran tan cortas que no llegabahasta la boca y, aparentemente, erademasiado débiles como para haber sido de ayuda para agarrar o desgarrar alas presas». Romer, A. S., 1966.Vertebrate Palaeontology. 3. ed.,Chicago y Londres, pág. 153.<<

[36] Lambe, L. M., 1917. «TheCretaceous theropodous dinosaur Gorgosaurus». Canada Dept. Mines.
Geol. Surv., memoria 100, págs. 1-84.Véase la página 64.<<

[37] Newman, B. H., 1970. «Stance andgait in the flesh-eating dinosaur Tyrannosaurus». Biol. J. Linn. Soc., págs. 119-23. La radical nuevaorientación de Newman, criticando a lasrestauraciones ortodoxas, sin duda seráatacada. Sin embargo, sus puntos devista sobre la forma de mantenerseerguido y andar del Tyrannosaurus
coinciden mucho con el análisis deLambe sobre el Gorgosaurus.<<

[38] Ostrom, J. H., 1969. «Terrestrialvertebrates as indicators of Mesozoicclimates». Proc. North Amer.
alaeontol. Conv., págs. 347-76. Laimportancia de la correlación entre una postura completamente erguida, laendotermia y la capacidad paramantener un metabolismo rápido fuereconocida por primera vez por Ostroaunque el tema fue desarrollado por Robert T. Bakker. Bakker, R. T., 1971.«Dinosaur physiology and the origin omammals». Evolution, 27, págs. 636-58.La correlación fue propuesta en 1973 e base a la aparente ausencia de cualquier

conexión casual. Véase Feduccia, A.,1973. «Dinosaurs as reptiles».
volulion, 27, págs. 166-70. Bennett,A. F., y Dalzell, B., 1973. «Dinosaur physiology: a critique». Ibid., págs.170-74. Ostrom contestó que, debido aque todos los vertebrados terrestresmodernos con posturas erguidas soendotérmicos, y a que los dinosauriostambién tenían una postura erguida,existía una gran probabilidad de que losdinosaurios también fueraendotérmicos. Independientemente de sise ha identificado o no la relaciócausal, contestó Ostrom, la correlacióentre la postura y el metabolismo semantiene en vigor. Ostrom, J. H., 1974.

Contestación a «Dinosaurs as reptiles».volution, 28, págs. 491-3. Véanse
también las contestaciones de Bakker yPeter Dodson a los ataques y susréplicas a Bennett y Feduccia en elmismo volumen.<<

[39] Russell, D. A., 1973. «Theenvironments of Canadian dinosaurs».Cañad, geog . J., 87, (1), págs. 4-11.<<

III. LA CARRERA ES PARA LOS
VELOCES, LA LUCHA PARA LOS FUERTES

[1] Ricqlès, A. de, 1969. «L’histologieosseuse envisagée comme indicateur dela physiologie thermique chez lestétrapodes fossiles». Comptes Rendus
cad. Scie., París, 268D, págs. 782-5.Este artículo fue sólo una nota de lasconclusiones de Ricqlès; el informecompleto de sus hallazgos en relaciócon los huesos de dinosaurio fue publicado el año anterior, pero, eaquella época, sacó pocas conclusiones.Ricqlès, A. de, 1968. «Recherches paléohistologique sur les os longs destétrapodes. I. Origine du tissu osseu plexiforme des dinosauriens

sauropodes». Ann. Paléontol.
(Vertébrés), 54, págs. 133-45.<<

[2] Lambe, L. M., 1917. «The Cretaceoustheropodus dinosaur Gorgosaurus».
Canada, Départ. Mines; Geol. Surv.,Memoria 100, Ottawa, págs. 1-84.<<

[3] Matthew, W. D., 1915. Dinosaurs.ueva York, Museo Americano de
Historia Natural: Manual no. 5, págs.40-41.<<

[4] Osborn, H. F., 1905. «Tyrannosaurus
and other Cretaceous carnivorousdinosaurs». Bull. Amer. Mus. Nat. Hist .,21, págs. 259-65.<<

[5] Osborn, H. F., 1913. «Tyrannosaurus,restoration and model of the skeleton».
ull. Amer. Mus. Nat. Hist ., 32, págs.91-2.<<

[6] Brown, B., 1915. «Tyrannosaurus, aCretaceous carnivorous dinosaur. Thelargest flesh-eater that ever lived».Scientific American, 113, págs. 322-3.<<

[7] Ostrom, en su descripción delextraordinario carnívoro Deinonychus,afirma que «los múltiples vestigios del( Deinonychus) sugieren que podríahaber sido gregario y que cazaba emanadas». Ostrom, J. H., 1969.«Osteology of Deinonychus
antirrhopus, an unusual theropod frothe Lower Cretaceous of Montana».
ull. Peabody Museum of Natural
istory, Yale University, 30, págs.1-165. Véase la página 144.<<

[8] Matthew, W. D., y Brown, B., 1923.«Preliminary note of skeletons andskulls of Deinodontidae from theCretaceous of Alberta». Amer. Mus.
ovit ., 89, págs. 1-10.<<

[9] Matthew, op. cit . (3), pág. 45.<<

[10] Swinton, W. E., 1934. The
inosaurs: A Short History of a Great
Group of Extinct Reptiles. Londres, pág.70.<<

[11] Bird, R. T., 1954. «We captured a“live” brontosaur». Nat. geogr. Mag .,105 (5), págs. 707-22.<<

[12] Lull, R. S., 1917. Organic
volution. Nueva York, págs. 414-15.<<

[13] Kielan-Jaworowska, Z., y Barsbold,R., 1972. «Results of the Polish-Mongolian palaeontological expedition part IV». Palaeontol. Polonica, 27, págs. 5-13.<<

[14] Bakker, R. T., 1972. «Anatomicaland ecological evidence of endothermyin dinosaurs». Nature, 238, págs. 81-5.Véase la página 85 (en la cual pongo elénfasis). Hay que señalar que en algunoscasos, especialmente en Mongolia, loshallazgos consisten casi por completo ecarnívoros. En estos casos es posibleque las arenas movedizas o losacimientos de alquitrán atraparan a
estas criaturas. De los yacimientos dealquitrán del pleistoceno de California,en los cuales se han encontradonumerosos cazadores como los «tigres»dientes de sable, se sabe que los

hallazgos no son representativos de lascomunidades como conjunto. Lascriaturas atrapadas atraen a un númerodesproporcionado de carnívoros allugar.<<

[15] Bakker, R. T., 1968. «The superiorityof dinosaurs». Discovery (New Haven),3 (2), págs. 11-22. Véase la pág. 21.<<

[16] Beecher, C. E., 1902. «Thereconstruction of a Cretaceous dinosaur,Claosaurus annectans». Marsh. Trans.
Conn. Acad. Arts and Sci., 11, págs.311-24.<<

[17] Ostrom, obr. cit .. (7), pág. 139.<<

[18] Galton, P. M., 1970. «The posture ohadrosaurian dinosaurs». J. Paleo., 44 págs. 464-73.<<

[19] Bakker, op. cit . (14), pág. 81.<<

[20] Colbert. E. H… 1962. Dinosaurs:
Their Discovery and their World .Londres, pág. 87.<<

[21] Osbom, H. F., 1916. «Skeletaladaptations of Ornitholestes,
Struthiomimus, Tyrannosaurus». Bull.
mer. Mus. Nat. Hist ., 35, págs. 733-71.<<

[22] «Notes on the habits oStruthiomimus by Dr. William K.Gregory». Ibid., págs. 758-60.<<

[23] Esta restauración de Heilman provocó el siguiente comentario deSwinton: «El nombre genérico significaimitador de las avestruces y, sin duda, laestructura del esqueleto se parece muchoa la de las aves semejantes a losavestruces modernos que conocemos ta bien hoy en día. Estas características, y posiblemente una traducción demasiadoliteral del nombre, han provocado quealgunos artistas, notablemente G.Heilmann (1926), hicieran unareconstrucción de aspecto tan parecido alos avestruces que, a pesar de la presencia de las patas delanteras y una

larga cola, la primera impresiódifícilmente recuerda a un reptil…».Pero, no resulta sorprendente que elestrutiomímido no tuviera aspecto dereptil ya que, como dice Swinton, «hayque recordar que no había muchascaracterísticas realmente típicas de losreptiles en el aspecto de su estructuraesquelética». Swinton, obr. cit .. (10), pág. 77.<<

[24] Heilmann, G., 1925. The Origin o
irds. Londres, pág. 183, fig. 132,reproducida aquí como fig. h. Este libroes un tesoro que contiene magníficasilustraciones de dinosaurios corredores.<<

[25] Bakker, op. cit . (14), pág. 81.<<

[26] Gregory, W. K., 1912. «Notes on the principies of quadrupedal locomotioand on the mechanism of the limbs ihoffed animals». Annals N. Y. Acad. Sci.,22, págs. 267-94. Véase la lámina 34.<<

[27] Dibujos extraídos de: Galton, P. M.,1974. «The ornithischian dinosaur ypsilophodon from the Wealden of the
Isle of Wight». Bull Br. Mus. Nat. Hist .(Geol.), 25 (1), págs. 1-152. Véase latabla V, pág. 139.<<

[28] Russell, D. A., 1972. «Ostricdinosaurs from the Late Cretaceous oWestern Cañada». Can. J. Earth. Sci., 9, págs. 375-402. Russell pudo demostrar que estos dinosaurios tenían cerebrosgrandes (en el Dromiceiomimus, elcerebro era más grande que el de uavestruz). Los ojos eran enormes, elárea de la retina era más grande que elde cualquier vertebrado terrestre y lavista era extremadamente buena. Loscuidados de sus crías no estaban fueradel alcance de la capacidad mental deestos dinosaurios y Russell especulasobre éste y otros misteriosos

problemas.<<

[29] En 1912, Gregory también observóla «conexión existente entre la forma delocomoción y la longitud delmetatarsiano medio en comparación coel fémur». Gregory, obr. cit .. (26), págs.284 et seq. Esta relación es incluso másextrema que la de T/F y expresa el gradohasta el cual la criatura se ha levantadosobre los dedos de los pies como ayuda para correr con mayor rapidez. Por ejemplo, en los pesados mamíferosgraviportales, el MT/F oscila entre 0,10 0,20 (0,13 en los elefantes), indicando
que los metatarsianos de los dedos delos pies casi no se han alargado; en los

rinocerontes es de 0,37; en los caballos primitivos subcursoriales es de0,50-0,60; mientras que en los caballosmodernos los metatarsianos se haalargado hasta tal punto que losanimales andan sobre las puntas de losdedos y tienen una relación MT/F de0,78. Para los estrutiomímidos larelación es de 0,77, en el ypsilophodon de 0,62 y ambos
dinosaurios andan sobre los dedos delos pies. En los dinosaurios saurópodosgigantes la cifra desciende hasta 0,13( Apatosaurus). Estas cifras describen lomismo que las relaciones tibia; fémur.Las cifras de los dinosaurios han sidotomadas de Galton, ob. cit . (18), págs.

467-8; y Galton, op. cit . (27).<<

IV. LA ERA DE LA OSCURIDAD

[1] Newton, E. T., 1894. «Reptiles frothe Elgin Sandstone. Description of twonew genera». Phil. Trans. Roy. Soc.,185 B, parte 1, págs. 573-604.<<

[2] Boulenger, C. A., 1904. «On reptiliaremains from the Trias of Elgin». Phil.
Trans. Roy. Soc., 196 B, págs. 175-89.<<

[3] Broom, R., 1913. «On the SoutAfrican pseudosuchian Euparkeria andallied genera». Proc. Zool. Soc. Lond .,(2), págs. 619-33.<<

[4] Ibid ., pág. 630.<<

[5] Colbert, E. H., 1962. Dinosaurs:
Their Discovery and their World .Londres, págs. 66-7.<<

[6] Walker lo ha incluido en el grupo delos dinosaurios carnívoros, aunque este punto sea muy discutido. Walker, A. D.,1964. «Triassic reptiles from the Elgiarea: Ornithosuchus and the origin othe carnosaurs». Phil. Trans. Roy. Soc,248 B, págs. 53-134. Aunque Romer lesiguió pronto e incluyó alOrnithosuchus en el grupo de loscarnosaurios en su Vertebrate
aleontology (1966), más tarde cambióde opinión y lo clasificó junto a losseudosuquianos. Romer, A. S., 1972.«The Chañares (Argentina). Triassicreptile fauna. XIII. An early

ornithosuchid pseudosuchian,Grascilisuchus stipanicicorum», gen. etsp. nov. Breviora, 389, págs. 1-24.Véase la pág. 23.<<

[7] Ewer, R. F, 1965. «The anatomy othe thecodont reptile Euparkeria
capensis». Broom. Phil. Trans. Roy.
Soc, 428 B, págs. 379-435.<<

[8] Romer, A. S., 1966. Vertebrate
alaeontology, tercera edición, Chicago Londres, pág. 137.<<

[9] Ewer, op. cit . (7).<<

[10] Reig, O., 1970. «The Proterosuchiaand the early evolution of thearchosaurs; an essay about the origin oa major taxon». Bull. Mus. Comp. Zool .,Harvard, 139 (5), págs. 229-92.<<

[11] Olson, E. H., 1971. Vertebrate
aleozoology. Nueva York, págs. 664 y666.<<

[12] Watson, D. M. S., 1951.aleontology and Modem Biology.ew Haven, págs. 38-78.<<

[13] Reig, op. cit . (10), págs. 259-62.<<

[14] Romber alude a este punto. «Se hasugerido que el comienzo de estoscambios (se refiere al desarrollo de lasextremidades traseras largas y la colaque actúa como contraequilibrio en los bípedos) estaba relacionado con el perfeccionamiento de la capacidad denadar de los antepasados de losarcosaurios que tenían hábitos anfibios.Si éste fuera el caso, se trataría de una“preadaptación ventajosa”». Romer, obr.
cit .. (8), pág. 136. Charig expone lacuestión de forma mucho másespecífica: «¿No podría ser que los primitivos arcosaurios eran anfibios —

como los mesosaurios y las nutrias— que utilizaban su cola para propulsarseen el agua y sus extremidades traseras para remar, cambiar de dirección eimpulsarse en el fondo del agua? La graimportancia de las extremidades traserasen la locomoción acuática se haceevidente por la mayor disparidad de lasextremidades en los cocodrilos marinos por el hecho de que en los cocodrilos
modernos sólo los pies traseros so palmeados». Charig, A. J., 1966.«Stance and gait in the archosaur reptiles». Advmt. ScL, Lond ., 22, pág.537 (resumen). En ocasiones, cuandoestán asustados, los cocodrilos seencabritan y hacen breves carreras sobre

las patas traseras.<<

[15] Romer, A. S., 1971. «The Chañares(Argentina). Triassic reptile fauna. X.Two new but incompletely known long-limbed pseudosuchian». Breviora, 378, págs. 1-10.<<

[16] La innovación clave quedesencadenó la aparición de losdinosaurios probablemente fue el bipedalismo (y, por lo tanto, la posturacon extremidades verticales), asociado auna fisiología con un metabolismorápido continuo. Charig cree que fuesolamente la postura erguida la queotorgó una especial ventaja a losdinosaurios. Por esta razón piensa quelos cuadrúpedos erguidos dieron lugar alos bípedos y a los dinosaurios persistentemente cuadrúpedos. Debido aque los seudosuquianos bípedos, ágiles de larga zancada, hicieron su aparició

durante el Triásico Inferior-Medio, yobviamente tuvieron un gran éxito,incluso las formas en evolución (comoel Ornithosuchus) que corrían parejascon los dinosaurios carnívoros, bípedos,resulta difícil creer que no poseían ya lainnovación clave, heredada más tarde y perfeccionada por los dinosaurios.Charig, A. J., 1972. «The evolution othe archosaur pelvis and hindlimb: aexplanation in functional terms». EJoysey, K. A., y Kemp, T. S. (ed).Studies in Vertebrate Evolution.Edimburgo, págs. 1221-55.<<

[17] Bakker, R. T… y Galton, P. M.,1974. «Dinosaur monophyly and a newClass of vertebrates». Nature, 248, págs. 168-72.<<

[18] Seeley, H. G., 1887. «Theclassification of the Dinosauria». Report
of the British Association for the
dvancement of Science, Manchester,57, págs. 698-9.<<

[19] Brink, A. S., 1956. «Speculations osome advanced mammaliacharacteristics in the higher mammal-like reptiles». Palaeont. Africana, 4,
págs. 77-95. Armand de Ricqlés apoyacompletamente la teoría de que losterápsidos eran endotermos. Ricqlés hademostrado, a partir de sus estudioshistológicos, que los huesos de losterápsidos, con numerosos vasossanguíneos y canales de Haver, puedeser clasificados como huesos demamíferos en contraposición con loshuesos de los reptiles y anfibiosfosilizados y vivos. Ricqlés, A. de,

1974. «Recherches paléohistologiquessur les os longs des tétrapodes. IV.Eothériodontes et pélycosaurs». Ann.
aleontol. (Vertébrés), 60, págs. 3-39.Véase también Ricqlés, A. de, 1969. «L’histologie osseuse envisagée commeindicateur de la physiologie thermiquechez les tétrapodes fossiles». Comptes
endus Acad. Sci., París, 268D, págs782-5. Ricqlés, A. de, 1969.«Recherches paléohistologiques sur lesos longs des tétrapodes. II. Quelquesobservations sur la structure des oslongs des thériodontes». Ann. Paleontol.
(Vertébrés), 55, págs. 1-52.<<

[20] Watson, D. M. S., 1931. «On theskeleton of a bauriamorph reptile».
roc. Zool. Soc. Lond ., págs. 1163-1205. Véase la pág. 1169.<<

[21] Bakker, R. T., 1971. «Dinosaur physiology and the origin of mammals».
volution, 25, págs. 636-58.<<

[22] Crompton, A. W., 1968. «The enigmaof the origin of mammals. Optima»(Anglo American Corp.), 18, págs.137-51. Véase la página 144.<<

[23] Jenkins, F. A., Jr, 1971. «The postcranial skeleton of Africacynodonts: problems in the evolution omammalian postcranial anatomy». Yale
eabody Mus. Bulletin, 36, págs. 1-216.Bakker, obr. cit .. (21), habría dejado quetuvieran los miembros aún másdesparramados que lo permitido por Jenkins.<<

[24] Crompton, op. cit . (22), pág. 143.<<

[25] Robinson, P. L., 1969. «A probleof faunal replacement on Permo-Triassiccontinents». Palaeontology, 14, págs.131-53. Véanse las páginas 135-7.<<

[26] Kermack, K. A., y Mussett, F., 1959.«The first mammals». Discovery
(Londres), 20, págs. 144-51. Kermack,D. M., Kermack, K. A., y Mussett, F.,1968. «The Welsh pantothers
uehneotherium praecursoris». J.
inn. Soc. (Zool.), 47, págs. 407-23.<<

[27] Hopson, J. A., 1971. «Postcaninereplacement in the gomphodont cynodont
iademodon». En Kermack, D. M., yKermack, K. A. (ed). Early Mammals.
Zool. J. Linn. Soc. Lond ., 50, supl. 1, páginas 1-21. Véanse las páginas 18-20.También Brink, obr., cit. (19).<<

[28] Gill, P. G., 1974. «Resorption o premolars in the early mammal
uehneotherium praecursoris». Archs.
oral Biol ., 19, págs. 327-8.<<

[29] Kermack, K. A., Mussett, F., yRigney, H. W., 1973. «The lower jaw o
organucodon». Zool. J. Linn. Soc.
ond ., 53, págs. 87-175. En este artículotambién hay un relato de lasexcavaciones del material en la fisuragalesa.<<

[30] Crompton, A. W., 1968. «In searcof the “insignificant”». Discovery (NewHaven), 3 (2), págs. 23-32.<<

[31] Colbert, E. H., y Russell, D. A.,1969. «The small Cretaceous dinosaur
romaeosaurus». Amer. Mus. Novit .,2380, págs. 145-62.<<

[32] Russell, D. A., 1969. «A newspecimen of Stenonychosaurus from theOldman Formation (Cretaceous) oAlberta». Can. J. Earth. Sci., 6, págs.595-612.<<

[33] Colbert, op. cit . (5), pág. 199.<<

[34] Romer, A. S., 1962. Man and the
Vertebrates. Penguin, vol. 1, págs. 95-6.<<

[35] Cobb, S., y Edinger, T., 1962. «The brain of the emu ( Dromaeus
novaehollandiae, Lath.)». Breviora, 170, págs. 1-18. Véase la página 5.<<

[36] Russell, D. A., 1972. «Ostricdinosaurs from the Late Cretaceous oWestern Canada». Can. J. Earth Sci., 9, págs. 375-402.<<

[37] Russell, op. cit . (32)<<

V. LA AGONÍA DE LOS TITANES

[1] Marsh, O. C., 1877. «Notice of a newand gigantic dinosaur». Am. Jour. Sci,14, págs. 87-8.<<

[2] Buckland, W., 1837. Geology an
ineralogy. Londres, pág. 115.<<

[3] Owen, R., 1841. «Report on Britisfossil reptiles, Part 2». Report of the
leventh Meeting of the British
ssociation for the Advancement o
Science, Londres, véanse las páginas94-103.<<

[4] Owen, R., 1841. «A description of a portion of the skeleton of theCetiosaurus, a gigantic extinct SauriaReptile occurring in the Ooliteformations of different portions oEngland». Proc. Geol. Soc. Londres,vol. 3, parte 2, no. 80, págs. 457-62.<<

[5] Owen, R., 1874-1889. Monograph o
the Fossil Reptilia of the Mesozoic
ormations. Sociedad Paleontográfica, pág. 412.<<

[6] Owen, R., 1847. «Report on thearchetype and homologues of thevertebrate skeleton». (Publicado por separado; originalmente apareció en el
eport of the British Association for
the Advancement of Science de 1846),Londres, págs. 169-340. Owen mástarde adoptó una posición evolucionistamás firme, por ejemplo, en s
onograph, obr. cit . (5), págs. 87-93.<<

[7] Phillips, J., 1870. Cetiosaurus.
thenaeum, no. 2214, 2 de abril, pág.454. Véase también Phillips, J., 1871.Geology of Oxford and the Valley of the
Thames. Oxford, págs. 254-94.<<

[8] Owen, op. cit . (5), pág. 564.<<

[9] Huxley, T. H., 1870. «Further evidence of the affinity between thedinosaurian reptiles and birds». Proc.
Geol. Soc. Londres, 26, pág. 12.También Huxley, T. H., 1870. «On theclassification of the Dinosauria, witobservations on the Dinosauria of theTrias». Ibid ., pág. 35.<<

[10] Seeley, H. G., 1874. «On the base oa large lacertilian cranium from thePotton Sands, presumably dinosaurian».Quart. Jour. Geol. Soc., 30, págs.690-92.<<

[11] Owen, op. cit . (5), pág. 593.<<

[12] Este relato de la controversiaMarsh-Cope está basado en Schuchert,C., y LeVene, C. M., 1940. O. C. Marsh,
ioneer in Palaeontology. New Haven, págs. 189 y sigs.; Ostrom, J. H., yMcIntosh, J. S., 1966. Marsh’s
inosaurs. New Haven y Londres, págs.2-47; y Williston, S. W., 1915. «The firstdiscovery of dinosaurs in the West».Matthews, W. D., 1915. En Dinosaurs.
ueva York, A. M. N. H., págs. 124-31.<<

[13] Marsh, O. C., 1877. «A new order oextinct Reptilia (Stegosauria) from theJurassic of the Rocky Mountains». Am.
our. Sci., 14; págs. 513-14.<<

[14] Cope, E. D., 1877. «On theVertebrata of the Dakota Epoch oColorado». Proc. Amer. Philos. Soc.,17, págs. 233-47.<<

[15] Marsh, O. C., 1878. «Principalcharacters of American Jurassicdinosaurs». Am. Jour. Sci., 16, págs.411-16.<<

[16] Marsh, O. C., 1883. «Principalcharacters of American Jurassicdinosaurs, Part VI, Restoration oBrontosaurus». Am. Jour. Sci., 26, págs.81-5.<<

[17] Ballou, W. H., 1897. «Strangecreatures of the past». The Century
llustrated Monthly Magazine. NuevaYork, 55, págs. 15-23. Cope hizo otrodibujo del Amphicoelias de pie sobre elfondo de un lago. Éste ha sidoreproducido en Osbom, H. F., 1931.Cope: Master Naturalist . PrincetoUniv. Press, en la página de enfrente dela página 302. La fecha de Osbom paraeste dibujo (1878) parece un pocotardía, ya que los saurópodos erarepresentados con extremidadesdelanteras diminutas y, obviamente, se pensaba que eran bípedos. En 1877,

Cope ya sabía que el Camarasaurus
tenía extremidades delanteras diminutas, como es natural, se pensaba que era
bípedos. En 1877, Cope ya sabía que elCamarasaurus tenía extremidadesdelanteras macizas.<<

[18] Osbom, H. F., 1898. «Additionalcharacters of the great herbivorousdinosaur Camarasaurus». Bull. Am.
us. Nat. Hist ., 10, págs. 219-33.<<

[19] Véase Wall, J. F., 1970. Andrew
Carnegie. Nueva York, esp. cap. 11 y12. Carnegie, A., 1965. The Gospel o
Wealth, and Other Timely Essays.Harvard Univ. Press, Cambridge, Mass.(Reimpresión de la primera edición,1900).<<

[20] La restauración preliminar deHatcher apareció en: Hatcher, J. B.,1901. « Diplodocus Marsh, its osteology,taxonomy, and probable habits, with arestoration of the skeleton». Mem.
Carnegie Mus., 1, pág. 1-63. Larestauración modificada de Holland fue publicada en: Holland, W. J., 1905.«The osteology of Diplodocus Marswith a special reference to therestoration of the skeleton of Diplodocus
camegiei Hatcher presented by Mr Andrew Carnegie to the BritisMuseum», 12 de mayo de 1905. Mem.
Carnegie Mus., 2, págs. 225-64.<<

[21] Anónimo (Holland, W. J.), 1905.«The presentation of a reproduction o
iplodocus camegiei to the Trustees othe British Museum». Ann. Carnegie
us., págs. 443-52.<<

[22] Frohawk, F. W., 1905. «The attitudeof Diplodocus carnegiei. The Field »,106, pág. 388. Para la contestación delos americanos, justificando la eleccióde la postura, véase The Field , 106, pág.466.<<

[23] Hay, O. P., 1908. «On the habits andthe pose of the sauropodous dinosaurs,especially of Diplodocus». Am. Nat .,42, págs. 672-81.<<

[24] Hay, O. P., 1910. «On the manner olocomotion of the dinosaurs especially
iplodocus, with remarks on the origiof birds», Proc. Wash. Acad. Sci., 12, págs. 1-25. Lámina 1.<<

[25] Holland, W. J., 1910. «A review osome recent criticisms of therestorations of sauropod dinosaursexisting in the museums of the UnitedStates, with special reference to that o
iplodocus camegiei in the CarnegieMuseum». Am. Nat ., 44, págs. 259-83.<<

[26] Bird. R. T., 1954. «We captured a“live” brontosaur». National
Geographic, 105, págs. 707-22.<<

[27] Matthew, W. D., 1910. «The pose osauropodous dinosaurs». Am. Nat ., 44, págs. 547-60.<<

[28] Hatcher, op. cit . (19), pág. 60.<<

[29] Holland, W. J… 1924. «The skull oDiplodocus». Mem. Carnegie Mus., 9, págs. 379-403.<<

[30] Osborn, H. F., 1915. «The dinosaursof the Bon Cabin quarry». Matthew,W. D., 1915. En Dinosaurs. NuevaYork, A. M. N. H., págs. 131-52.<<

[31] Ibid ., pág. 141.<<

[32] Bird, R. T., 1944. «Did Brontosaurusever walk on land?». Natural History,53, no. 2, págs. 60-67.<<

[33] Kermack, K. A., 1951. «A note othe habits of the sauropods». Ann. Mag.
at. Hist ., 12 (4), págs. 830-32. Para elartículo de Stigler, en el quecorrelaciona la profundidad de lainmersión, la presión del agua y lacapacidad (o incapacidad) del hombre para respirar, véase: Stigler, R., 1911.«Die Kraft unserer Inspirationsmuskulatur». Pflügler’s
rchiv für Physiologie, 139, págs.234-54. De hecho, Oliver P. Hay e1910, obr. cit .. (24), pág. 25, posiblemente ya se adelantó a la líneade razonamientos de Kermack. Hay

escribió:
«En el artículo publicado por Mr.Ballou… hay una ilustración querepresenta a un grupo de cuatroindividuos de Amphicoelias latus…
Estos animales han sido representados paseándose por el fondo del río,comiendo vegetación y alzándose sobresus patas traseras para coger aire… Esdifícil imaginarse que el Diplodocus
pudiera pasearse sumergido en el aguaal igual que lo es en el caso delhombre». <<

[34] Colbert, W. H., 1952. «Breathinghabits of the sauropod dinosaurs». Ann.
ag. Nat. Hist ., 12 (25), págs. 708-10.<<

[35] Kermack conversando con el autor el18 de octubre de 1974.<<

[36] Swinton escribe, en 1967, que losgrandes saurópodos «probablementevivieron en las aguas de los estuarios olagos donde la capacidad de flotacióreduciría el enorme peso corporal» yque «en el Diplodocus… la nariz era unaabertura en la parte superior del cráneo.Los saurópodos tenían un medioambiente satisfactorio en las aguas profundas, rodeados por abundantecomida y sin estar expuestos a losataques de los carnívoros». Swinton,W. E., 1967. Dinosaurs. B. M. (N. H.),Londres, págs. 17, 19. La obra estándar más frecuentemente utilizada e

Paleontología de los vertebrados es laVertebrate Palaeontology de Romer. Eella se expresan las mismas ideas: «La posición de estos órganos (nasales)sugiere una forma de vida anfibia paralos saurópodos; el animal podía respirar mirar a su alrededor con sólo la parte
superior de la cabeza sobresaliendo por encima del agua». Romer, A. S., 1966.Vertebrate Paleontology. Chicago yLondres, pág. 155.<<

[37] Bakker, R. T., 1971. «Ecology of the brontosaurs». Nature, 229, págs. 172-4.Las insólitas ideas de Bakker concernientes a los saurópodos fuero publicadas por primera vez unos tresaños antes: Bakker, R. T., 1968. «Thesuperiority of dinosaurs». Discovery
(New Haven), 3, (2), págs. 11-22.<<

[38] «News and Views, 1971. Changingdinosaurs –but not in midstream».
ature, 229, pág. 153.<<

[39] Bakker, 1971, op. cit . (37), pág. 174.<<

[40] Bird, obr. cit .. (32)<<

[41] Ostrom, J. H., 1972. «Were somedinosaurs gregarious?». Palaeogreogr.,
alaeoclimatol, Palaeoecol ., 11, págs.287-301.<<

[42] Richmond, N. D., 1965. «Perhapsuvenile dinosaurs were always scarce».. Palaeo, 39, págs. 503-5.<<

[43] Sikes, S. K., 1971. The Natural
istory of the African Elephant .Londres, págs. 102-7.<<

[44] Brown, B., 1941. «The lastdinosaurs». Natural History, 48, págs.290-95.<<

VI. UN GRIFO RESCATA A LA EVOLUCIÓN

[1] Mivart, St. G., 1871. The Genesis o
Species. Londres. Véanse el capítulo I
especialmente las páginas 106-7.<<

[2] Meyer, H. von, 1862. «On therchaeopteryx lithographica from the
lithographic slate of Solnhofen». Ann.
ag. nat. Hist., 9, pág. 366-70.<<

[3] Wagner, A., 1862. «On a new fossilreptile supposed to be furnished witfeathers». Ann. Mag. nat. Hist., 9, págs.261-7.<<

[4] Citado por de Beer, G., 1954.« Archaeopteryx lithographica: A study based upon the British Museuspecimen». Museo Británico (Historia
atural), pág. 2.<<

[5] Un deliciosamente malicioso juego de palabras de Herbert Wendt, 1970.
efore the Deluge. Paldin bolsillo, pág.240.<<

[6] Huxley, T. H., 1868. «On the animalswhich are most nearly intermediate between birds and reptiles». Ann. Mag.
nat. Hist . (4), 2, páginas 66-75.<<

[7] Vogt, C., 1880. « Archaeopteryx
macrura, an intermediate form betwee birds and reptiles». Ibis, 4, págs. 434-56.<<

[8] Heilmann, G., 1925. The Origin o
irds, Londres, pág. 165.<<

[9] Osborn, H. F., 1916. «Skeletaladaptations of Ornitholestes,
Struthiomimus, Tyrannosaurus». Bull.
m. Mus. Nat. Hist ., 35, págs. 733-71.<<

[10] Cope, E. D., 1867. «An account othe extinct reptiles which approached the birds». Proc. Acad. Nat. Sci
hiladelphia, págs. 234-235.<<

[11] Huxley, op. cit . (6) y 1870. «On theclassification of the Dinosauria, witobservations on the dinosaurs of theTrias». Q. Jour. Geol. Soc., 26, págs.32-51.<<

[12] Simpson utilizó este recurso paradesacreditar a Lowe (que creía que lasaves no voladoras se habían originadodirectamente a partir de losdinosaurios). Haciendo que loscelurosaurios y las aves fueraconvergentes, Simpson eliminó alantepasado dinosaurio e hizo que fueramás verosímil que todas las aves sehubieran originado a partir del
rchaeopteryx (al cual Lowe habíadescartado como vástago estéril).Simpson, G. G., 1946. «Fossil penguins». Bull. Am. Mus. Nat. Hist .,
ueva York, 87, págs. 1-99. Para la

polémica contra Lowe, véase «A note orchaeopteryx and Archaeomis», págs.
92-5.<<

[13] Fleilmann se vio obligado a pedir una carta de recomendación de Arthur Smith Woodward del Museo Británico.Carta de Woodward del 10 de octubrede 1925, acompañada de una copia dellibro: colección de Smith Woodwardque se encuentra en la Biblioteca deCiencias Naturales D. M. S. Watson, eel University College de Londres.<<

[14] De Beer, op. cit . (4), véanse las págs. 42-50.<<

[15] Walker, A. D., 1972. «New light othe origin of birds and crocodiles».
ature, 237, págs. 257-63.<<

[16] Ostrom, J. H., 1973. «The ancestryof birds». Nature, 242, página 136.<<

[17] De hecho, esta dislocación se hace patente en la ilustración del cinturó pélvico del espécimen de Berlín en lamonografía de Beer, op. cit . (4),ilustración 7, pág. 29.<<

[18] Bakker, R. T., y Galton, P. M., 1974.«Dinosaur monophyly and a new Classof vertebrates». Nature, 248, págs.168-72.<<

[19] The Sunday Times, 17 de marzo de1974, pág. 13.<<

[20] Ibid .<<

[21] Ostrom, J. H., 1974. « Archaeopteryx
and the origin of flight». Q. Rev. Biol .,49, págs. 27-47.<<

[22] Bakker y Galton, op. cit . (18).<<

[23] Walker, obr. (15) señala que elmúsculo elevador supracoracoideo delala no era funcional en el
rchaeopteryx, y Ostrom (comentario personal del 4 de marzo de 1975)observa la reducción o pérdidacompleta de los elevadores corrientes,los deltoides, tal como se evidencia por la estrecha escápula.<<

[24] De Beer, obr. cit .. (4), pág. 24.<<

[25] Halstead, L. B., 1969. The Pattern
of Vertebrate Evolution, Edimburgo, pág. 143.<<

[26] Heptonstall, W. B., 1970.«Quantitative assessment of the flight o
rchaeopteryx». Nature, 228, págs.185-6. La estimación del peso deHeptonstall de 500 gramos está basadaen la de una paloma de tamaño similar (400 gramos), más una cantidadadicional que tenga en cuenta laausencia de huesos neumáticos, etc.<<

[27] Yalden, D. W., 1971. «Flying abilityof Archaeopteryx». Nature, 231, pág.127. Bramwell, C., 1971. Nature, 231, pág. 128. Véase la contestación deHeptonstall a ambas críticas, Ibid. Laestimación de Yalden del peso de
rchaeopteryx es de 200 gramos, esdecir, el de un ave moderna ligera perode tamaño similar. Esto parece que es u peso demasiado ligero para udinosaurio celurosaurio apenasmodificado.<<

[28] Mayr, E., 1960. «The emergence oevolutionary novelties». S. Tax (ed). eThe Evolution of Life. Chicago. Bock,W. L, 1969. «The Origin and radiation o birds». Ann. N. Y. Acad. Sci., 167, págs.147-55.<<

[29] Ostrom, op. cit . (21), págs. 43-4.<<

[30] Bock, op. cit . (28)<<

[31] De Beer, obr. cit .. (4), pág. 38.<<

[32] Ostrom, J. H., 1974. Contestación a«Dinosaurs as reptiles». Evolution, 28, págs. 491-3. El Compsognathus, ausiendo más grande que el
rchaeopteryx, era pequeño y seencontró en los sedimentos deSolnhogen. Es concebible que estecelurosaurio tuviera plumas. De acuerdocon Ostrom (comen. pers. del 4 demarzo de 1975), el bloque que conteníael Compsognathus fue pulido cocuidado alrededor del fósil para que nose destruyese ninguna impresión de plumas. El Compsognathus fuedescubierto antes del primer espécime

de Archaeopteryx (Londres), de formaque, en el momento de su aparición,nadie había podido ser alertado de la posibilidad de existencia de plumas. Lasimpresiones de plumas en los últimostres especímenes de Archaeopteryx sotan débiles que, al principio, lascriaturas no fueron reconocidas como
rchaeopteryx. Por último, quierodestacar el parecido existente entre los pequeños celurosaurios y el
rchaeopteryx. Hay que tener en cuentaque durante mucho tiempo se pensó queel quinto espécimen de Archaeopteryx
era un Compsognathus, ya que no seencontraron las impresiones de plumasdistintivas.<<

[33] Jerison, H. J. 1968. «Brain evolutioand Archaeopteryx». Nature, 219, págs.1381-2.<<

[34] Ostrom, op. cit . (21).<<

[35] Nopcsa, barón F., 1907. «Ideas othe origin of flight». Proc. Zool. Soc.,Londres, págs. 223-36. Véanseespecialmente las páginas 234-6.<<

[36] Duerden, J. E., 1920. «Inheritance ocallosities in the ostrich». Am. Nat., 54, págs. 289-312.<<

[37] Para una explicación darwinistamoderna de cómo puede imitarse ellamarckismo, véanse las ideas deWaddington sobre la canalización.Maynard Smith, J., 1972. The Theory o
volution. Penguin, págs. 294-9.<<

[38] Lowe, P. R., 1926. «On thecallosities of the ostrich (and other Palaeognathae) in connection with theinheritance of acquired characters».
roc. Zool. Soc. Londres, págs. 667-79.Lowe, P. R., 1935. «On the relationshipof the Struthiones to the dinosaurs and tothe rest of the avian class, with specialreferente to the position o
rchaeopteryx». Ibis, 5, págs. 398-432.Lowe, P. R., 1944. «Some additionalremarks on the phylogeny of theStruthiones». Ibis, 86, págs. 37-42.<<

[39] Lowe, 1935, op. cit . (38), pág. 420.<<

[40] Marsh, originariamente, encontró elesperornis en Kansas, pero, más
recientemente, Dale Russell halocalizado vestigios tan numerosos en laregión ártica del Canadá (donde el
esperornis representa entre un tercio ytres cuartos de todos los hallazgos), que parece como si hubiera encontrado ellugar de reproducción del Hesperornis.Esto parece posible en base al númerode especímenes inmaduros que seencuentran en el Norte. Russell, D. A.,1973. «The environments of Canadiadinosaurs». Cañad. Geog. J., 87, (1) págs. 4-11.<<

[41] Swinton, W. E., 1965. Fossil Birds.Museo Británico (Historia Natural), pág.41.<<

VII FANTASMAS DEL INFIERNO

[1] Cuvier, G., 1824. Recherches sur les
Ossimens Fossiles. París, vol. 5, parte2, págs. 379-80.<<

[2] Seeley, H. G., 1870. The
Ornithosauria. Cambridge, pág. 7.<<

[3] Descrito en Ibid ., pág. 11.<<

[4] Citado de Lang, W. D., 1939. «MaryAnning (1799-1847) and the pioneer geologists of Lyme». Proc. Dorset nat.
ist, archaeol. Sole., 6, págs. 142-64.<<

[5] Buckland, Rev. W., 1835. «On thediscovery of a new species o pterodactyl in the Lias at Lyme Regis».Geol. Trans., Londres, serie 2, vol. 3, págs. 217-22. Véase la pág. 218.<<

[6] Buckland, Rev. W. 1836. Geology
and Mineralogy, Considered with
reference to Natural Theology, 1, págs.224-5. Éste es el volumen 5 de The
ridgewater Treatises on the Power
Wisdom and Goodness of God as
manifested in the Creation, Londres.<<

[7] Abel, O., 1918. «Uber dieVerwertung von Fossilrekonstruktioneim naturwissenschaftlichen Unterricht».Aus der Natur, cuaderno 1, págs. 17-23.Véase la pág. 20. También Aberl, O.,1911. «Die Bedeutung der fossileWirbeltiere für die Abstammungslehre».Die Abstammungslehre, págs. 198-250.Véanse las págs. 207-13.<<

[8] Hawkins, T., 1840. The book, of the
Great Sea-Dragons, Ichthyosauri an
lesiosauri, Gedolim Taninim o
oses, Extinct Monsters of the Ancient
arth. Londres, pág. 26. Sexcentricidad se estaba haciendo másevidente en aquella época. Su trabajoanterior sobre los saurios marinos habíaconsistido en una serie de descripcionesrelativamente pedantes. Abandonó esteestilo después de la obra publicada e1840 y en 1844 produjo The Wars o
ehovah in Heaven, Earth and Hell !<<

[9] Newton, E. T., 1888. «On the skull, brain and auditory organ of a newspecies of pterosaurian (Scaphognathus
urdoni), from the Upper Lias near Whitby, Yorkshire». Phil. Tras. Soc.,Londres, 179 B, págs. 503-37.<<

[10] Ibid ., pág. 510<<

[11] Seeley, H. G., 1901. Dragons of the
ir . Nueva York (reimprimido en 1967,Dover, bolsillo), pág. 54.<<

[12] Edinger, T., 1941. «The brain oterodactylus». Am. Jour. Sci., 239,
págs. 665-82.<<

[13] Seeley, op. cit . (11). Su libroanterior, The Ornithosauria, sólo sehabía ocupado de los pterodáctilos delas areniscas verdes de Cambridge.<<

[14] Seeley, op. cit . (11), pág. 57.<<

[15] Newman, E., 1843. «Note on the pterodactyle tribe considered asmarsupial bats». The Zoologist , 1, págs.129-31.<<

[16] Wanderer, K., 1908.« Rhamphorhynchus gemmingi H. v. Meyer. Ein Exemplar mit teilweiseerhaltener Flughaut a. d. Kgl.Mineralog.-Geol. Museum zu Dresden».
alaeontographica, volumen 55, págs.195-216. La piel «con hoyuelos» del pterosaurio es descrita en las páginas198 y 205.<<

[17] Broili, F., 1927. «Eihamphorhynchus mit Spuren vo
Haarbedeckung». Sitzgsb. d. math.-
naturw. Abt ., 5 de febrero, págs. 49-67.<<

[18] Sharov, A. G., 1971. «New flyingreptiles from the Mesozoic deposits oKazakhstan and Kirgizia». Trudy. Pal.
nst . AN S. S. S. R., 130, págs. 104-13.(En ruso). Mr. Jack Saxon, que tuvo laamabilidad de traducir este artículo paramí, también es el responsable de latraducción alternativa del Sordes
ilosus.<<

[19] Halstead propone un argumentoligeramente diferente para los cuidadosde las crías de los pterosaurios. «Se hasugerido que los pterosaurios como el
teranodon cuidaban a sus crías, ya quelas hembras no podrían haber llevadohuevos equivalentes al tamaño esperado.Tendrían que haber puesto huevos muy pequeños y, una vez salidas de loshuevos, las crías no habrían sidocapaces de cuidar de sí mismas; por lotanto los padres tenían que cuidar deellas». Halstead, L. B., 1969. The
attern of Vertebrate Evolution.Edimburgo, pág. 142.<<

[20] Seeley, op. cit . (2)<<

[21] Seeley, op. cit . (11), pág. 230.<<

[22] Bakker, R. T., y Galton, P. M., 1974.«Dinosaur monophyly and a new classof vertebrates». Nature, 248, pág. 171.<<

[23] Sharov, op. cit . (18)<<

[24] Zambelli, R., 1973.« Eudimorphodon ranzii gen. nov., sp.nov., uno pterosauro Triassico». IstitutoLombardo (Rend. Sc.), B, 107, págs.27-32. Este primer pterosaurioconocido, encontrado en los sedimentosdel triásico superior en Bérgamo, Italia,es extraordinario ya que posee dientescon múltiples protuberancias en losmolares como los de los mamíferos primitivos. Los pterosaurios queaparecieron más tarde tenían dientes couna única corona.<<

[25] Owen, R., 1841-1884. A History o
ritish Fossil Reptiles. Londres, vol. 1, págs. 245-6.<<

[26] Ibid .<<

[27] Marsh, O. C., 1871. «Note on a newand gigantic species of Pterodactyle».
m. Jour. Sci. (3), 1, pág. 472.<<

[28] Marsh, O. C., 1872. «Discovery of aremarkable fossil bird». Am. Jour. Sci.(3), 3, pág. 56-7.<<

[29] Marsh, O. C., 1872. «Discovery oadditional remains of Pterosauria, witdescriptions of two new species» Am.
our. Sci., (3), 3, págs. 241-8.<<

[30] Hankin, E. H., y Watson, D. M. S.,1914. «On the flight of pterodactyls».Aeronaut. J, 18, págs. 324-35.<<

[31] Ibid.<<

[32] Bramwell y Whitfield, teniendo ecuenta el parecido existente entre los pies del Pteranodon y los de umurciélago, concluyen que el
teranodon se colgaba con la cabezadirigida hacia abajo de los acantilados.Bramwell, C. D., y Whitfield, G. R.,1974. «Biomechanics of Pteranodon».
hil. Trans. Roy. Soc., Londres, 267 B, págs. 503-92. Ostrom (comen. pers., 4de marzo de 1975) señala la ausencia dedetritos en la creta de Niobrara, eKansas, como prueba de que losacantilados habían estado muy lejos.<<

[33] Esta idea, tal como ha sido descritaen el capítulo 1, al parecer se debe aRichard Owen en 1841 aunque siguióapareciendo en la Prensa popular e1914; véase Hankin y Watson, obr. cit ..(30).<<

[34] Brown, B., 1943. «Flying reptiles».atural History, 52, páginas 104-11.<<

[35] Heptonstall, W. B., 1971. «Aanalyisis of the flight of the Cretaceous pterodactyl Pteranodon ingens
(Marsh)». Scott. J. Geol ., págs. 61-78.<<

[36] Bramwell, C., 1970. «The first hot- blooded flappers». Spectrum, 69, págs.12-14. Véase también Bramwell, B., yWhitfield, G. R., 1970. «Flying speed othe largest aerial vertebrate». Nature,225, págs. 660-61. Heptsonstall, obr.
cit . (35), pensaba que el Pteranodon
pesaba más del doble del peso estimado por Bramwell en sus cálculos.Utilizando sus cifras, el pterosauriohabría resultado ser muy inferior a los planeadores modernos. Bramwell yWhitfield atacaron posteriormente lasestimaciones de Heptonstall.<<

[37] Short, G. H., 1914. «Wingadjustments of pterodactyls». Aeronaut.
., 18, pág. 336-43.<<

[38] Comentario editorial a ibid .<<

[39] Bramwell y Whitfield, op. cit . (32).<<

[40] Heptonstall, obr. cit .. (35)<<

[41] Hankin y Watson, op. cit . (30).<<

[42] Bramwell, C., 1970. «Thoseflappers again». Spectrum, 72, página 7.<<

[43] Bramwell y Whitfield, op. cit . (32),sin embargo, sugieren que, al ser ultraligeros, los cuerpos de los pterosaurios posiblemente fueroarrastrados por el agua mar adentro.<<

[44] Heptonstall, op. cit . (35).<<

[45] Brown, op. cit . (34)<<

[46] Véase el comentario de Short, obr.
cit .. (37), pág. 340, sobre el artículo precedente de Hankin y Watson.<<

[47] Halstead, op. cit . (19), pág. 142.<<

[48] Lawson, D. A., 1975. «Pterosaur from the latest Cretaceous of WestTexas: discovery of the largest flyingcreature». Science, 187, págs. 947-8.Quisiera dar las gracias al Sr. Lawso por mostrarme un borrador de esteartículo antes de su publicación.
El nuevo pterosaurio de Texas no tieneun nombre oficial y aún no se haexcavado ningún fósil que se parezca alespécimen de Lawson. Aún existemuchos conceptos erróneos en tomo aestos planeadores que se hacen patentes por las afirmaciones que aparecen en laPrensa en relación con el hallazgo de

Lawson. «El anuncio del descubrimiento —afirmó el The Times — seguramentevolverá a reavivar el viejo debate entrelos paleontólogos de si estos reptilesvoladores batían sus alas sin plumascurtidas o si solamente se subían aramas altas y saltaban hacia lascorrientes de aire para encumbrarsecomo los planeadores». The Times, 13de marzo de 1975, pág. 1. Es evidenteque la idea de un pterosaurio peludo aútiene que ser asimilada popularmente.Véanse mis comentarios en el The
Times, 14 de marzo de 1975, pág. 16.<<

[49] Desmond, A. J., 1975. «The puzzleover the pterosaur: how did the world’s biggest bird (!) ever manage to fly?».The Times, 12 de abril, pág. 12.<<

VIII. LA LLEGADA DE ARMAGEDÓN: ¿UN
CATACLISMO CÓSMICO?

[1] Reig, O. A., 1970. «TheProterosuchia and the early evolution othe archosaurs; an essay about the origiof a major taxon». Bull. Mus. Comp.
Zool . Harvard, 139 (5), pág. 229-92.Véase la pág. 230.<<

[2] Simpson, G. G., 1953. Major
eatures of Evolution. Nueva York, pág.291-3.<<

[3] Frecuentemente se observacicatrices en la gola lo cual indica lainserción de músculos en vida. La golatambién protegía la región del cuelloque siempre era el lugar más vulnerable.Ostrom, J. H., 1964. «A functionalanalysis of jaw mechanics in thedinosaur Triceratops». Postilla, 88, págs. 1-35. Véanse las págs. 11-13.<<

[4] Russell, D. A., 1967. «A census odinosaur specimens collected in Wester Canada». Nati. Mus. Can. Hist. Pap.,36, págs. 1-13.<<

[5] Newell, N. D., 1966. «Problems ogeochronology». Proc. Acad. Nat. Sci.
hiladelphia, 118, págs. 63-89. Véasela pág. 74. Este «cambiorevolucionario» en sólo un millón deaños demuestra la rapidez con la cual se puede extinguir toda una comunidad sino existen sucesores.<<

[1] Swain, T., 1974. «Cold-bloodedmurder in the Cretaceous». Spectrum,120, pág. 10-12.<<

[7] Colbert, E. H., 1965. The Age o
eptiles. Londres, págs. 16 y sigs.<<

[8] La tortuga es un reptil muy primitivo.Se cree que su origen, a pesar de que es bastante desconocido, se sitúa cerca delos reptiles más primitivos, pocodespués de su evolución a partir de losanfibios. Tanto los mamíferos como losdinosaurios tenían orígenes mucho mástardíos y están alejados por igual de latortuga. Debido a que los mamíferos, y probablemente los dinosaurios, eraendotermos (un factor no tenido ecuenta por los dinosaurios) deberían ser incluidos en el grupo de los mamíferos yno en el de las tortugas de sangre fría eel estudio de Swain.<<

[9] El Museo Británico (Historiaatural) tiene una exposición de
anomalías patológicas halladas edinosaurios que incluye un esqueleto dedinosaurio bípedo flexionado. Siembargo, en la exposición cercana del
rchaeopteryx, los moldes deldinosaurio-ave jurásico presentaexactamente los mismos síntomas, sique se hayan encontrado evidencias quedemuestren que se tratara deespecímenes patológicos. Existedemasiados esqueletos flexionados demodo que se puede descartar lahipótesis de que todos ellos fuera

patológicos y, actualmente, la mayoríade las autoridades aceptan que esteestado era el resultado de la desecaciódel cadáver.<<

[10] Rosenkrantz, A., y Brotzen, F., 1960.Congreso Geológico Internacional.
eport of the Twenty First Session,
orden. Parte 5. «The CretaceousTertiary boundary».<<

[11] Hall, J. W., 1960. «The Cretaceous-Tertiary bounday in the TampicoEmbayment, Mexico». Ibid . págs. 70-77.<<

[12] Hall, J. W., y Norton, N. J., 1967.«Palynological evidence of floristicchange across the Cretaceous-Tertiary boundary in eastern Montana (U. S. A.)». Palaeogeography, Palaeoclimatol,
alaeoecol , 3, págs. 121-31. A finalesdel mesozoico, las coniferasrepresentaban sólo el 12% de la flora, pero hacia comienzos del terciariohabían aumentado esta cifra hasta el30%. En cifras absolutas, los autoresencontraron 11 especies degimnospermas en la formación HellCreek (finales del cretáceo) y 16especies en la formación Tullock

(comienzos del terciario). Lasdicotiledóneas (plantas con flores)estaban representadas con 46 especiesen Hell Creek pero sólo con 17 eTullock. El número total de especiesvegetales se redujo a la mitad en lafrontera.<<

[13] Axelrod, D. I., y Bailey, H. P., 1968.«Cretaceous dinosaur extinction».
volution, 22, págs. 595-611. Losetasequoia y otros «fósiles vivos»,
como los ginkos o el cabello de Venuscultivados en los templos chinos yaponeses, han encontrado un refugio e
nuestro mundo en algunas áreasmontañosas de China donde estáasociados con árboles de hoja perenneresistentes y árboles de hojas caducas.En altitudes más bajas dan paso aexuberantes bosques de robles, palmeras laurel.<<

[14] Bramlette, M. N., 1965. «Massiveextictions in biota at the end oMesozoic time». Science, 148, págs.1696-9.<<

[15] Terry, K. D., y Tucker, W. H., 1968.«Biologic effects of supemovae».Science, 159, págs. 421-3. Estas y lassiguientes cifras del texto han sidoextraídas de esta fuente. Existe unamedida de prueba independiente de quela radiación solar directa (pero demenor cantidad) puede provocar cambios en la fauna, al menos entre losorganismos unicelulares. Cada millón deaños, aproximadamente, el campomagnético de la Tierra cambia dedirección de forma que el polo Norte seconvierte en el polo Sur y viceversa.Cuando está a punto de producirse la

inversión, la intensidad del campodesciende a un 15%, el cambio se produce durante mil años y el campovuelve a recuperar su potencia. Duranteel cambio se colapsa el cinturón de VaAllen que protege la Tierra de laradiación cósmica y la Tierra es bañadaen radiación solar. Hay un notable gradode correlación entre estos cambiosmagnéticos y las vicisitudes de las poblaciones de radiolarios. Cuando los polos cambian, los radiolarios — organismos unicelulares deaproximadamente 0,1 mm de tamaño ycon una concha dura— cambiaconcomitantemente su tipocaracterístico. Uffen, R. J., 1963.

«Influence of the earth’s core on theorigin and evolution of life». Nature,198, págs. 143-4. Véase también: Black,D. I., 1971. «Polarity reversal andfaunal extinction». En Gass, I. G., Smith,P. J. y Wilson, R. C. L. (ed.),Understanding the Earth, Artemis, págs. 257-61.<<

[16] Laster, H., 1968. «Cosmic rags fronearby supernova: biologic effects».Science, 160, pág. 1138.<<

[17] Russell, D., y Tucker, W., 1971.«Supemovae and the extinction of thedinosaurs». Nature, 229, págs. 553-4.<<

[18] Russell, D., 1971. «Thedisappearence of the dinosaurs». Cañad.
Geog. J., 83, págs. 204-15.<<

[19] Clemens, W. A., 1960. «Stratigraphyof the Type Lance formation».Rosenkrantz and Brotzen, obr. cit .. (10), págs. 7-13.<<

[20] Colbert, op. cit . (7), pág. 200.<<

[21] Ibid ., págs. 197-203. Véansetambién las págs. 169-179.<<

[22] Entre el período cenománico(cretáceo superior) y período deMaastricht inferior, el número defamilias de amonitas registradasdescendió de 22 (conteniendo 78géneros) a 11 (con 34 géneros). Duranteel período Maastricht, sólo 11 de estosgéneros pudieron sobrevivir hasta elfinal cuando fueron destruidos todos.También existe una notable contraccióde la extensión de las amonitas; alcomienzo del período de Maastricht, lasamonitas se hallaban por todo el planeta, pero hacia el final ya habíadesaparecido del Hemisferio Sur.

Esencialmente, lo mismo es válido paralas emparentadas belemnitas. Hancock,J. M., 1967. «Some Cretaceous-Tertiarymarine faunal changes». Harland, W. B., otros (ed), The Fossil Record .
Sociedad Geológica, Londres, págs.91-104.<<

[23] Cys, J. M., 1967. «On the inability othe dinosaurs to hibernate as a possiblekey factor in their extinction». J.
aleontol ., 41, pág. 226.<<

[24] Bakker, R. T., 1972. «Anatomicaland ecological evidence of endothermyin dinosaurs». Nature, 238, págs. 81-5.Cys, obr. cit . (23), estimó que losdinosaurios más pequeños del cretáceosuperior, es decir los Leptoceratops,
Stegoceras, Dromaeosaurus,
Saurornithoides y Thescelosaurus nomedían menos de 1,5 a 3,6 metros delongitud.<<

[25] Bakker, R. T., 1971. «Dinosaur physiology and the origin of mammals».
volution 25 (4), págs. 636-58. Eltamaño de las crías es analizado en la pág. 651.<<

[26] Russell, L. S., 1965. «Bodytemperature of dinosaurs and itsrelationships to their extinction». J.
aleontol ., 39, pág. 497-501. En esteartículo, Russell explora lasconsecuencias que habría habido si losdinosaurios hubiesen sido de sangrecaliente. Concluyó que la endotermia era«posible» y, de hecho, «verosímil». La piel desnuda de los dinosaurios,especialmente si las bestias eraendotérmicas, habría provocado unacierta susceptibilidad al frío. El artículode Russell, corto pero elemental, publicado en 1965, está lleno de ideas

que fueron ampliadas y desarrolladas por otros autores. La piel desnudatambién es tratada por Bakker, obr. cit ..(24), pág. 650.<<

[27] Bramwell, C. D., y Whitfield, G. R.,1974. «Biomechanics of Pteranodon».
hil. Trans. Roy. Soc., Londres, 268 B, págs. 503-92.<<

[28] Russell, D. A., 1975. «Reptiliadiversity and the Cretaceous-Tertiarytransition in North America». Caldwell,W. G. E. (ed). «Colloquium on theCretaceous System in the Wester Interior of North America». Geological
ssociation of Canada, Special Paper
(en prensa).<<

[29] Descrito por Russell, D. A., 1973.«The environments of Canadiadinosaurs». Cañad. Geolog. J., 87 (1), págs. 4-11.<<

[30] Erben, H. K., 1972. «Ultrastruktureund Dicke der Wand pathologischer Eischalen». Abh. Akad . Wiss. Lit. math.-
nat. Kl., 6, págs. 191-216. Véasetambién la tabla I, figuras 5 y 6.<<

[31] El profesor Erben me ha informadoque el descenso del grosor de lascáscaras no fue repentino sino gradual.
o obstante, el profesor opina que todoel proceso se desarrolló con gran
rapidez. La prueba de Erben sugiere quela tensión sufrida por los dinosaurios nose produjo debido a la brusquedad deuna «supernova», a pesar de que eltiempo implicado era rápido segúestándares geológicos. Comen. pers. del27 de febrero de 1975.<<

APÉNDICE 2

[1] Popper, K. R., 1972. Conjetures an
efutations. Londres.<<

[2] Freduccia, A., 1973. «Dinosaurs asreptiles». Evolution, 27, págs. 166-9. LaDra. Pamela Robinson sostiene unaopinión (aunque extrema) similar corelación a la adquisición de la calidadde sangre caliente en los mamíferos.Arguye que la endotermia de losmamíferos, probablemente se desarrollóa finales del mesozoico o comienzos delterciario, concomitantemente con elagrandamiento del cerebro de losmamíferos. Robinson, P. L., 1971. «A problem of faunal replacement oPermo-Triassic continents».
alaeontology, 14, págs. 131-53.<<

[3] Russell, D. A., 1969. «A newspecimen of Stenonychosaurus from theOldman Formation (Cretaceous) oAlberta». Can. J. Earth. Sci., 6, págs.595-612. Russell, D. A., 1972. «Ostricdinosaurs from the late Cretaceous oWestern Canada». Ibid ., 9, págs.375-402.<<

[4] Feduccia, A., 1974. «Endotermy,dinosaurs and Archaeopteryx».
volution, 28, págs. 503-4.<<

[5] Jerison, H. J., 1973. Evolution of the
rain and Intelligence. Nueva York yLondres, pág. 408.<<

[6] Bakker, R. T., 1971. «Dinosaur physiology and the origin of mammals».
volution, 25, págs. 636-58.<<


















