LA INCORPORACIÓN DE LO COTIDIANO A LA ... -...
51
1 LA INCORPORACIÓN DE LO COTIDIANO A LA ENSEÑANZA DE LA FISIOLOGÍA VEGETAL MARÍA ESTRELLA LEGAZ GONZÁLEZ BLANCA FONTANIELLA LÓPEZ CARLOS VICENTE CÓRDOBA Grupo de Investigación Validado UCM-CM: “Interacciones celulares en Simbiosis Vegetales” (Ref. 910081). Facultad de Biología, Universidad Complutense Con la colaboración de: Dra. María Blanch Rojo, Dra. Mara Sacristán San Cristóbal, Dra. Rocío Santiago Tejero, MSc. Borja Alarcón Aguareles, Lcda. Eva María Díaz Peña y MSc. Elena Sánchez Elordi. LA INCORPORACIÓN DE LO COTIDIANO A LA ENSEÑANZA DE LA FISIOLOGÍA VEGETAL MARÍA ESTRELLA LEGAZ GONZÁLEZ BLANCA FONTANIELLA LÓPEZ CARLOS VICENTE CÓRDOBA Grupo de Investigación Validado UCM-CM: “Interacciones celulares en Simbiosis Vegetales” (Ref. 910081). Facultad de Biología, Universidad Complutense Con la colaboración de: Dra. María Blanch Rojo, Dra. Mara Sacristán San Cristóbal, Dra. Rocío Santiago Tejero, MSc. Borja Alarcón Aguareles, Lcda. Eva María Díaz Peña y MSc. Elena Sánchez Elordi.
Transcript of LA INCORPORACIÓN DE LO COTIDIANO A LA ... -...

1
LA INCORPORACIÓN DE LO COTIDIANO A LA ENSEÑANZA DE LA FISIOLOGÍA
VEGETAL
MARÍA ESTRELLA LEGAZ GONZÁLEZBLANCA FONTANIELLA LÓPEZCARLOS VICENTE CÓRDOBA
Grupo de Investigación Validado UCM-CM: “Interacciones celulares en Simbiosis Vegetales”(Ref. 910081).Facultad de Biología, Universidad Complutense
Con la colaboración de: Dra. María Blanch Rojo, Dra. Mara Sacristán San Cristóbal, Dra. Rocío Santiago Tejero, MSc. Borja Alarcón Aguareles, Lcda. Eva María Díaz Peña y MSc. Elena Sánchez Elordi.
LA INCORPORACIÓN DE LO COTIDIANO A LA ENSEÑANZA DE LA FISIOLOGÍA
VEGETAL
MARÍA ESTRELLA LEGAZ GONZÁLEZBLANCA FONTANIELLA LÓPEZCARLOS VICENTE CÓRDOBA
Grupo de Investigación Validado UCM-CM: “Interacciones celulares en Simbiosis Vegetales”(Ref. 910081).Facultad de Biología, Universidad Complutense
Con la colaboración de: Dra. María Blanch Rojo, Dra. Mara Sacristán San Cristóbal, Dra. Rocío Santiago Tejero, MSc. Borja Alarcón Aguareles, Lcda. Eva María Díaz Peña y MSc. Elena Sánchez Elordi.

2
MAESTRO
ALUMNO
FORMACIÓN
Actividadescomplementarias
Flujo de Informaciónprimaria
Lecturassugeridas
Interiorización Tutorías
Fig. 1. El sistema actual de transmisión del conocimiento.
1. La materia, Fisiología vegetal La Fisiología vegetal es la ciencia que estudia los mecanismos mediante los cuales las plantas desarrollan sus procesos vitales, cómo se nutren, cómo crecen, cómo se reproducen, cómo se comunican. La Fisiología Vegetal trata de establecer relaciones causa-efecto, en una dinámica principalmente experimental, que aclaren dichos procesos vitales, estableciendo modelos explicativos para cada familia de fenómenos, modelos susceptibles de verificación y, eventualmente, modificación. La ciencia establece modelos que son válidos en tanto en cuanto vencen al criterio de falsabilidad: aquellas condiciones que invalidarían el modelo jamás se cumplen (Popper, 1994). En este sentido, no sería válida la premisa de Stenger (2007) cuando afirma:
Nótese que el principal propósito de los modelos científicos consiste en describir y no tanto en explicar los fenómenos.
La ciencia que el hombre ha desarrollado, que permite explicar (a pesar de lo que opine Stenger) casi todo lo que existe o sucede, que aclara las relaciones causa-efecto de los fenómenos naturales y sus manifestaciones, ha sustituido en gran parte al miedo que surge del desconocimiento y la ignorancia. Precisamente por esto no se debería definir la ciencia con premisas falsas, incompletas o susceptibles de interpretaciones tendenciosas.
2. El método docente Si en la formación tradicional el esquema de transmisión del saber otorga al profesor un papel predominante, un proceso de este tipo podría definirse como un fenómeno de “desconocimiento de lo lento”, según lo define Mühlmann (2006). “Lo lento” es, en este caso, lo adquirido para ser transmitido o mejor, la adquisición de saberes para ser posteriormente transmitidos. Mühlmann
aclara que
Lento es un movimiento que dura más de una generación. Para observarlo, hemos de recurrir a la colaboración de seres humanos que han vivido antes que nosotros y de seres humanos que vivirán después.

3
Léase por vivido lo realizado en los años precedentes, susceptible de transformarse en conocimiento transcendente. Profesor y alumno se transforman así en eslabones de una cadena que elabora conocimiento al mismo tiempo que lo transmite, en la vía por la que “lo lento” (en cuanto a ciencia se refiere) transcurre. Lo que hasta hace unos 50 años resultaba estructuralmente casi imposible, la colaboración con generaciones anteriores y posteriores, hoy es un hecho. La velocidad con la que el conocimiento científico se genera, especialmente el conocimiento biológico, es tan rápida que la colaboración se establece entre tres generaciones solapadas en el tiempo y laboralmente activas. Lo único que puede retrasar el proceso de transmisión del conocimiento es el rechazo de uno de los eslabones, generalmente segundo o tercero (es decir, profesor meramente transmisor y discípulo) a la aceptación de los saberes.
No podemos criticar el sistema docente actual acusándolo de maldad intrínseca o de falta absoluta de vigencia. Hace muy pocos años, ante la pregunta de un periodista sobre qué desearía para la Universidad de Oxford, su Canciller, de visita en Madrid, respondió sin vacilar: “Más Erasmus españoles”. Al margen de la cortesía obligada del visitante al país que visitaba, la frase tiene un significado real y profundo. Los actuales universitarios españoles, estudiantes de Licenciatura y quizá también de Grado, son tan buenos como los mejores. El sistema, qué duda cabe, ha formado muchos y excelentes profesionales a la altura de los mejores de cualquier país del mundo. Hoy, por mor de facilitar la transferencia de especialistas entre los países de la Unión Europea, el sistema debe ser reformado, adjudicando al discípulo un papel de mayor protagonismo en su formación cultural, convirtiéndolo en administrador de su propia ansiedad de progreso, incardinada ésta en un sistema de formación pautada (Mühlmann, 1996). Y esto no es nada fácil.
Si el antiguo sistema tiene bondades objetivas, destruirlo sería un despilfarro, pero su adaptación a los nuevos intereses precisaría una “deconstrucción” para que las piezas de su esqueleto descoyuntado fuesen reorganizadas de forma distinta e incluso dejaran espacio para nuevas opciones susceptibles de ser injertadas sin rechazo. Una deconstrucción, en el sentido que le dio Derrida (2003), de un cuidadoso desmontaje de la metafísica de un sistema para tratar de privarlo de sus defectos.
2.1. Las nuevas tendencias
La biología moderna está plagada de descubrimientos maravillosos: la funcionalidad del sistema nervioso, los virus, los antibióticos, la doble hélice, los sistemas de comunicación celular. Sin duda, lo maravilloso es un acicate, un motivo de esperanza para el alumno que comienza sus estudios en Biología. ¿A qué estudiante de Biología no se le aceleró el pulso leyendo ¿Qué es la vida? (Schrödinger, 1944), La doble hélice (Watson, 2000) o Los cazadores de microbios (de Kruif, 2006)? Cada uno de esos deslumbrantes descubrimientos entraña una promesa de protagonismo, ¿por qué no? Pero este estímulo de lo maravilloso cuesta mucho trabajo de mantener. La rutina cotidiana, el

4
ALUMNO MAESTRO
Insertar experiencias vitales propias en el aprendizaje
Ansiedad por adquirir bienes culturales
Búsqueda de su propio rol en el futuro: importancia de lo aprendido
Clarificación de su posición exacta en la vida académica
Respuestas formativas
Garantía(Auctoritas)
Integración socio-científica del conocimiento
Fig. 2. Las nuevas tendencias o lo que Bolonia demanda.
esfuerzo del estudio, la multiplicidad de asignaturas, otras alternativas de más inmediata satisfacción personal para un joven hacen que el efecto de lo maravilloso se diluya o se posponga para mejor ocasión. Pasa lo mismo con el arte. Lo sublime en el arte proporciona momentos de satisfacción intensa, pero se diluyen con la ausencia del objeto (no todo el mundo puede tener un da Vinci en su casa) y con el tiempo. Quizá entonces habría que buscar en lo cotidiano una fuente de estímulos para el alumno que se enfrenta a los estudios de Biología, probablemente en el mismo sentido que Hegel (1989) le atribuye al arte:
Por mucho amor a lo maravilloso que se tenga, la tarea del arte es en último término hacer justicia a la cotidianeidad.
¿No podría decirse lo mismo de la ciencia? Claro que, llegados a este punto, la pregunta debe ser distinta: ¿la curiosidad sobre el hecho biológico entra dentro de los intereses cotidianos de un estudiante universitario? La respuesta podría ser indistintamente sí o no, aunque en ambos casos con matizaciones. Partiendo de la base de que el porcentaje de alumnos que muestran una curiosidad primaria por algún hecho biológico que concierne a
nuestra materia suele ser bajo, la mayor parte los integrantes de este bajo porcentaje muestran interés por problemas y dentro de estos problemas, la mayor parte de ellos son de una naturaleza netamente patológica. ¿Ejemplos? Muchos. “En mi casa hay un castaño que tiene las hojas llenas de manchas rojas, ¿qué podría hacerse?” La pregunta no es inventada, sino real, y nótese que ni siquiera el interés primario es por la relación causa-efecto, sino una simple demanda de solución. Un bajo (bajísimo) porcentaje de estos alumnos con curiosidad tienen inquietudes más básicas y, por tanto, plantean preguntas con mucha mayor opción propedéutica. “Mi madre fregó con

5
amoniaco una mesa de piedra sobre la que tenía una planta de narciso y sus flores blancas se volvieron azules, ¿a qué puede deberse esto?”
La observación de hechos cotidianos, sorprendentes o demasiado familiares, pertenece al acervo de la memoria viva de cualquier individuo. Otra cosa es que dichas observaciones no entren dentro del cuadro primario e inmediato de sus intereses. Si fuera así, perfecto; si no, siempre es posible obligarles a la concatenación del hecho observado con su explicación científica si el maestro incorpora lo cotidiano a su programa docente, bien ilustrando la explicación de procesos fisiológicos con ejemplos de este tipo, bien suscitando en el alumno la capacidad de relación mediante el recuerdo asociativo. Esto, que parece a simple vista una empresa difícil de abordar, no lo es tanto si estudiamos algunos de los ejemplos que a continuación se exponen.
POSIBLES PREGUNTAS QUE LOS ALUMNOS PUEDEN HACERSE
1º ¿Por qué en Andalucía dejan el agua “solearse” (templarse al sol) antes de regar las plantas con ella?
2º ¿Por qué las macetas se recubren durante el invierno con un plástico transparente para evitar que se hielen?
3º ¿Por qué se dice que en verano es mejor regar las macetas al atardecer?
4º ¿Por qué se recomienda no exponer a los ciclámenes a las corrientes de aire?
5º ¿Por qué los suelos excesivamente pisoteados (compactados) suelen ser pobres en vegetación?
6º ¿Por qué muchas especies de plantas costeras tienen las hojas gruesas?
7º ¿Por qué hay plantas que cuando comienzan a secarse enrollan sus hojas sobre su eje longitudinal hasta solapar sus bordes, formando una especie de cilindros?
8º ¿Por qué hay setas que forman hileras o arcos más o menos abiertos sobre el suelo?
9º ¿Por qué un plátano de sombra mantiene en diciembre un penacho de hojas verdes justo debajo de una farola del alumbrado público mientras que el resto de la copa se ha desfoliado completamente?
10º ¿Por qué los geranios de varias macetas en la baranda de una terraza orientan ramas y flores hacia el exterior?
11º ¿Por qué el tronco de un pino rodeno en la Sierra de Albarracín dibuja el contorno de una peña inmediata a donde ha crecido?
12º ¿Por qué el pie de un árbol se derrama sobre el cemento del borde del alcorque?
13º ¿Por qué la luz del atardecer es púrpura en Madrid y rosa en Granada?

6
Fig. 3. Higos primeros del mes de mayo
Fig. 4. La “cama” de los melones maduros.
Fig. 5. ¿Cuáles son las semillas de los cocos?
14º Si las flores del diente de león y del hibisco se abren durante el día y se cierran durante la noche, ¿por qué las primeras se cierran a mediodía si se las riega y las segundas no?
15º ¿Por qué almendros y cerezos florecen en el frío febrero?
16º ¿Se da el fenómeno del fotoperiodismo en el ecuador?
17º ¿Por qué una misma higuera da brevas e higos?
18º ¿Por qué hay higueras que sólo dan higos?
19º ¿Por qué una higuera puede dar higos en abril, brevas en julio e higos en Septiembre?
20º ¿Por qué las flores de
los jazmines huelen de noche?
21º ¿Por qué se podan los árboles en invierno?
22º ¿Por qué las frutas se guardan en las cajoneras del frigorífico mientras que el resto de los alimentos, incluso vegetales, se colocan en las baldas?
23º ¿Por qué las chirimoyas o los plátanos maduran más rápidamente envueltos en papel de estraza, o de periódico, al aire libre?
24º ¿Por qué suele decirse que un melón está maduro cuando su “cama” es amarilla?
25º ¿Dónde están las semillas de los cocos y cómo germinan?
25º ¿Por qué hay pinos y cedros que forman un círculo en el suelo desnudo de vegetación herbácea alrededor de su tronco?
26º ¿Por qué algunas patatas, al ser

7
Fig. 6. Hojas hendidas de Monstera deliciosa.
cortadas, tienen puntos negros bien definidos mientras que el resto del tubérculo aparece blanco?
27º ¿Por qué las hojas adultas de Monstera deliciosa están profundamente hendidas mientras que la hoja juvenil muestra un limbo continuo?
28º ¿Por qué se oscurecen las manos al pelar cardos antes de cocinarlos?
29º ¿Por qué no son verdes los espárragos blancos, mientras que sí lo son los llamados trigueros?
30º ¿Es cierto que mordisquear una ramita de adelfa puede resultar mortal?
Sin duda, La Fisiología Vegetal puede dar respuesta cumplida a estas y otras muchas curiosidades de un alumno. ¿Qué no tienen importancia? Sí que la tienen, y mucha. Por eso es importante que las sientan. Y si no tienen estas curiosidades, pueden despertárseles. Recibirlas directamente, o suscitarlas si no afloran de forma espontánea, son maneras de insertar lo maravilloso del conocimiento biológico en lo cotidiano de la vida del estudiante, una forma de llevar un aspecto de la ciencia a la vida real del individuo. Los resultados de esta inserción solo pueden ser positivos para su formación integral.
2.2. Las armas del profesor
Bolonia requiere que los alumnos desarrollen habilidades específicas en relación con la ciencia que estudian. El término “habilidades” tiene muchas interpretaciones. Abordemos las más corrientes.
1ª Clases o seminarios.
Una práctica usual en los grupos piloto preparatorios a la implantación del Plan de Convergencia Europea ha sido dejar a la iniciativa de los alumnos el desarrollo de temas específicos que después exponían en una puesta en común con sus compañeros. Estos temas específicos podían ser, bien una lección del programa, elegida por el profesor en función de parámetros tales como facilidad, accesibilidad a la información etc., bien seminarios complementarios a una lección del programa. Los resultados de estas experiencias han sido muy diversos y, a veces, contradictorios. Como norma general, nos inclinaríamos por la opción de desechar la preparación de un tema del programa y por aceptar los seminarios complementarios en los cuales los alumnos puedan desarrollar habilidades tales como su capacidad de relación con la línea directriz del problema tratado, el manejo de la bibliografía, la elección de experimentos claramente demostrativos, la

8
capacidad de síntesis (el seminario debe tener un tiempo de exposición limitado), el orden mental, la capacidad de comunicación etc.
Por otro lado, la división de la materia en tres partes bien diferenciadas que se propone en el programa necesita una aclaración adicional. La primera parte, Nutrición, está enfocada de forma que requiere un formalismo físico-químico riguroso. Esto implica que, quizá en esta parte, las habilidades del alumno estén muy limitadas y su misión fundamental sea adaptarse a la dinámica de unas clases muy conceptuales. Esto no es excluyente de los criterios fundamentales del espíritu de Bolonia: simplemente, las partes pueden ser inhomogénas dentro de un todo homogéneo. Sinceramente, la explicación de la expresión de Onsager, ecuación fenomenológica que aplica principios de termodinámica irreversible para explicar situaciones muy alejadas del equilibrio en sistemas de permeación en medio acuoso, no debería ser encargada sin más a un alumno. Las habilidades que comportan la incardinación del estudiante en los procesos de elaboración de la materia, en mayor o menor proporción, pueden ejercerse con mucha mayor libertad y con mucha mayor eficacia en las otras dos partes del programa, Fotosíntesis y Metabolismo (2ª Parte) y Crecimiento y Desarrollo (3ª parte).
Suele ser útil también proporcionar al alumnado en general, a través de los sistemas virtuales al uso en cada Universidad, resúmenes de cada uno de los temas, incluyendo una bibliografía mínima pero muy significativa, con tiempo suficiente como para que lleguen a clase con el resumen leído o, dicho de otra forma, sabiendo de qué va la lección del día. Desgraciadamente, la frase “para que lleguen a clase con el resumen leído” no implica ni de lejos una seguridad absoluta de que suceda, sino más bien, posibilitar que el alumno tenga la opción de saber de qué va la lección del día, si es que quiere saberlo. Si quisiera, y con el resumen leído, la clase le sería mucho más provechosa.
Los seminarios no deben ser demasiados (un máximo de cuatro-seis para el cuatrimestre), adjudicados con tiempo suficiente como para que su preparación no engendre estrés, preparados por no más de tres alumnos, supervisados por el profesor antes de la puesta en común y generadores de discusión. Aquí el papel del tutor es decisivo porque el estudiante que recibe el seminario, compañero de los ponentes, tiende a no intervenir por un mal entendido sentido de solidaridad con sus condiscípulos (cree que evita ponerlos en evidencia). El profesor debe promover el debate y encaminar la discusión de tal manera que los alumnos intervinientes asuman un papel antes de colaboradores que de críticos de lo realizado, dar a entender que el trabajo no está terminado hasta que sea pulido y aceptado por la totalidad de la clase.
2º Elaboración de mapas conceptuales.
Un mapa conceptual es un diseño gráfico en el que las relaciones causa-efecto de un fenómeno particular se concatenan en compartimientos superponibles de tal manera que el desarrollo lógico de los acontecimientos desde la causa

9
desencadenante hasta el efecto final quedan clara y secuencialmente expuestos. Es una especie de mapa conceptual global, en el sentido descrito por Novak (1998), el cual puede descomponerse en subconjuntos coherentes, de extensión limitada y de más fácil acceso, conectados a su vez entre sí. Un mapa conceptual es, pues, un resumen gráfico de un concepto de tal forma que sus múltiples pasos, perfectamente ordenados e interrelacionados, pueden ser visualizados fácilmente (Vicente, 2009):
El alumno parte de una bibliografía mínima que contenga los elementos experimentales en que basarse y se le anima a encontrar fuentes bibliográficas complementarias. Su misión es asimilar la experimentación, encontrarle la concatenación lógica a los diferentes sucesos, establecer las interrelaciones y sacar las conclusiones precisas. El tratamiento experimental puede resultarle suficiente pero también puede pensar que hubiera sido mejor de otra forma, que a tal experimento le falta algo que lo haga enlazar mejor con el siguiente, que tal conclusión podría haber sido de otra manera si el enfoque hubiese sido distinto.
Por resumirlo en una imagen, se trata antes de crear un collage, una obra propia utilizando elementos ajenos, que de ordenar un puzzle, colocar las piezas en una posición única y decidida de antemano. Con todos estos elementos, puede construir un esquema topográfico (un mapa conceptual) en el que:
“los diferentes conceptos aparecen jerárquicamente distribuidos en el espacio de modo que permite la representación de los diversos tipos de relaciones entre ellos” (Sánchez y Gairín, 2008)
Un ejemplo servirá para explicar mejor esta nueva “habilidad”. Supongamos que en clase se ha desarrollado el tema de la formación de los aerénquimas. Un resumen convencional podría ser el siguiente:
La diferenciación de los aerénquimas en las raíces y tallos de plantas sensibles a la anoxia. En raíces de maíz, por ejemplo, se puede desencadenar un proceso de formación de aerénquimas radiculares sometiendo el órgano a hipoxia. Una concentración de oxígeno del 4%, por ejemplo, hace que grupos de células parenquimatosas incrementen la producción de glucanasas que digieren las paredes celulares, con lo que consecuentemente se desorganiza el protoplasto y la célula muere (Subbaiah y Sachs, 2003). Se forman así cavidades concretas dentro del tejido parenquimatoso que se rellenan de aire para asegurar, al menos temporalmente, el aporte de oxígeno al resto de la raíz. La señal para la síntesis de glucanasas consiste en un incremento de la concentración de calcio citoplásmico que viene mediada por una cascada de reacciones que transmiten la señal (baja disponibilidad de oxígeno) hasta lograr la apertura de un canal iónico. La transducción de la señal involucra proteínas G de membrana, activables por unión de GTP, activación de

10
Fig. 7. Mapa conceptual de la formación del aerénquima
adenilato ciclasa y proteína quinasas y finalmente fosforilación de proteínas que abren el correspondiente canal iónico. Esto ha sido demostrado al provocar diferenciación de aerénquima en raíces de maíz en normoxia (alta concentración de oxígeno que no dispararía la apoptosis conducente a la muerte celular) tratadas con GTPγS (un análogo estructural del GTP no metabolizable por la actividad GTPasa de las proteínas G) o por ácido okadaico, un inhibidor de las proteína fosfatasas, enzimas que desfosforilan las proteínas-P de canal iónico e inducen su cierre. Ambas manipulaciones, al mantener los canales iónicos abiertos, incrementan constantemente la concentración de calcio citosólico e imponen la apoptosis que desencadena la formación del aerénquima aunque no haya sido percibida por la planta una falta de oxígeno (Dangl et al., 2000).
Se propone a los alumnos realizar el mapa conceptual de este fenómeno desglosando cada uno de los pasos que lo componen, ordenándolas en el tiempo y, cuando sea posible, en el espacio, buscando relaciones causa-efecto entre dos pasos consecutivos y encadenándolos entre sí hasta lograr tener ante los ojos un cuadro lo más completo posible de la evolución del mecanismo. Evidentemente, este mapa conceptual, quizá mínimo, puede complicarse hasta el punto que se quiera, por ejemplo, introduciendo cuñas entre las diferentes elipses explicando la esencia de cada uno de los mecanismos. La construcción de mapas conceptuales sirve entonces básicamente para fijar conceptos y facilitar el repaso de los temas en los casos que se requiera.
El resultado podría ser, entre otras muchas posibilidades, el expuesto en el cuadro siguiente.
3º Problemas propuestos.
Tradicionalmente, aquellas materias que poseen una extensa conceptualización matemática (las Matemáticas en sí mismas) o las ciencias experimentales recurren a la proposición de problemas como acción docente

11
complementaria de la exposición teórica. En el caso de la Fisiología Vegetal, las actividades complementarias consistentes en seminarios para la resolución de problemas propuestos han sido desarrolladas con una cierta extensión desde comienzo de los años 70 del pasado siglo (Legaz y Vicente, 1989; Sabater, 1998).
Conviene hacer aquí algunas aclaraciones previas. Un seminario teórico sobre un asunto específico de un tema concreto puede ser en sí mismo un problema, sensu lato. Para desarrollar tal seminario, hay que preparar un guión (una ruta intelectual que vaya desde el planteamiento hasta la solución), buscar la bibliografía de apoyo más adecuada, establecer una secuencia lógica de hipótesis y demostraciones hasta llegar a la solución final. Esto, sin duda, requiere varios días de trabajo. Por problema, aquí, queremos usar la concepción clásica del mismo, la desarrollada durante todo el siglo XX para materias científicas, como la matemática o la física. Un problema, en este contexto, es el enunciado de una incógnita de forma que contenga en sí mismo todos los datos precisos para que un alumno pueda desvelar dicha incógnita con sus conocimientos actuales y en un tiempo relativamente corto. Un ejemplo clásico sería el de los dos trenes que van a encontrarse en un punto determinado, algo que todos hemos podido ver en nuestro primer curso de Física. Un tren sale de A con una velocidad de 120 km h-1. Otro tren sale de B, distante 600 km de A, a una velocidad de 150 km h-1. Si ambos trenes parten simultáneamente a las 9:00 am, ¿Cuánto tiempo tardarán en encontrarse? ¿A qué distancia de A se producirá el encuentro? Todos recordamos algo semejante, ¿verdad? Encontrar ejemplos análogos en la Fisiología Vegetal es relativamente fácil, teniendo siempre en cuenta que hay cosas que nunca deben hacerse. Por ejemplo, un problema nunca puede ser una adivinanza. ¿Qué pensaríamos de los siguientes enunciados?
Caso 1. Dos trenes salen de dos lugares distintos a velocidades de 120 y 150 km h-1. respectivamente …….
Caso 2. Un tren sale de A a una velocidad dada. Otro tren sale de B, a 600 km de A, a una velocidad distinta …..
Caso 3. ……… ¿Cómo se llama la hermana del maquinista?
Ya que estos problemas, por su propia naturaleza, son elementos inmediatos de evaluación, tienen que ser un acto docente, no una trampa. Deben ser algo inherente a la enseñanza, no algo de lo que el profesor pueda desprenderse hasta recibir la solución por parte de terceros. Tienen que ser posibles, resolvibles en un tiempo determinado. Tienen que servir a quien los resuelve como prueba de sus propios conocimientos y deberán aportar al alumno confianza en sí mismo y satisfacción al ser capaz de resolverlos con un esfuerzo razonable, usando unos saberes que ya le son propios. La búsqueda de carencias en su propia formación nunca debe ser un objetivo para sentarse frente a un problema e intentar solucionarlo: un problema nunca debe constituir el paso inmediato a una posible evaluación. Aquí el papel del profesor es decisivo: jamás debe ser un mero guía en la búsqueda de información (un profesor no es Google, ni siquiera su manual de uso), jamás debe hacer dejación de su autoridad como maestro, tiene la estricta

12
obligación de dejar solo al alumno frente al problema únicamente cuando le haya provisto de todos los recursos necesarios. Y aún a partir de este momento, nunca debería perderlo de vista. Solo entonces el problema podrá ser el camino en solitario para el que el maestro ha preparado al alumno y de su éxito el maestro debe sentirse orgulloso.
Para evaluar correctamente la capacidad resolutiva del alumno, deberían valorarse:
1. Comprensión del enunciado: dimensión precisa del problema propuesto.
2. Pasos iniciales: es muy positivo encontrar el hilo que desmadeja el ovillo.
3. Economía de recursos: es mejor un camino corto y recto que dar mil vueltas y acabar perdido.
4. Correcto entendimiento de los signos: en fisiología, + puede ser algo más que más, y – puede ser algo más que menos.
5. Uso correcto de unidades: las dimensiones son singulares.
6. Capacidad de relación: un fenómeno fisiológico siempre tiene conexiones con otros relacionados.
7. Abstenerse de argumentos innecesarios: contestar a un problema completo no debe transformarse en una exposición global de saberes.
8. Llegar a la solución correcta.
Estos problemas son siempre elaborados por el profesor y pueden ser agrupados en dos clases diferentes:
a) Problemas de cálculo, en la que se agrupan todas aquellas cuestiones que se refieren a régimen hídrico, permeabilidad de membrana, difusión pasiva o transporte activo, velocidad de flujo, velocidad de transpiración etc., en incluso en la misma clase podrían considerarse problemas prácticos, como cálculo de concentraciones, potenciometría, cálculo de la acidez total, energía de la luz, efecto fotoquímico, demanda cuántica, rendimiento cuántico etc.
b) Problemas sobre hechos experimentales, consistentes en la exposición de datos, bien sean en tablas o en gráficas, describiendo con claridad y concisión los antecedentes, las bases experimentales y los protocolos utilizados y solicitar del alumno el enunciado de las conclusiones mediante el uso de preguntas concatenadas. Esta clase abarca prácticamente todo el contenido de la materia.
Un ejemplo de cada una de estas posibilidades se ofrece a continuación:
A) Cierta disolución adherida a un microcapilar se inserta dentro del citoplasma de células epidérmicas de raíz de Quercus pyrenaica. Dos horas

13
después, la concentración de soluto es 0,3 molal en el plano de inserción y 0,111 molal a una distancia de 8 mm.
Calcular:
a) El coeficiente de difusión del soluto.
b) ¿Cuándo alcanza el soluto la concentración del 37% del valor en el plano de inserción a 64 cm de distancia?
c) ¿Cuántos moles de soluto por unidad de área existen a 8 mm de distancia del plano de inserción?
d) Suponiendo que una pequeña cantidad de cierta substancia con un coeficiente de difusión 100 veces menor que del soluto principal se introduzca inicialmente, ¿dónde caerá su concentración al 37% de la del plano de inserción para el tiempo calculado en b)?
B) Se ha analizado mediante Northern blot la producción de mRNA para invertasa por nodos de tallos de avena gravitrópicamente estimulados. Un RNA-poliA parcialmente purificado fue sometido a electroforesis en agarosa al 1%, transferido a una membrana de nylon e hibridado con un α−32PdCTP-cDNA
de invertasa producido por PCR, usado como prueba. En A) se muestra el mRNA aislado de nodos caulares intactos graviestimulados de avena, en B), el mRNA aislado de la cara superior de los nodos graviestimulados y en C), el mRNA aislado de la cara inferior del nodo. Las diferentes calles indican tiempos diferentes de graviestimulación, correspondiendo el tiempo 0 al control vertical.
Se pregunta:
a) ¿Conduce la estimulación gravitrópica a inducción de invertasa y en qué zona principalmente?
b) ¿Cuál puede ser la relación entre actividad invertasa y reorientación gravitrópica?
Los seminarios de problemas no deben ser bajo ningún concepto sesiones en las cuales el profesor los explica a sus alumnos, sino que deben ser enfocados en el sentido de la participación activa del estudiante, ofreciendo al resto de sus compañeros los cálculos realizados, las pautas lógicas que han aplicado para la resolución de la cuestión, y las conclusiones obtenidas.
Vicente (2010) describe un ensayo con estudiantes de 4º y 5º Cursos de Licenciatura en el cual el sistema se invierte y son los alumnos los que proponen a sus compañeros problemas a resolver. Para ello, el estudiante cuenta con una separata propuesta por el profesor, de la cual elige la gráfica o tabla, o ambas, que será sujeto de análisis. El estudiante debe entonces analizar detenidamente la necesidad y el planteamiento del experimento,

14
Fig. 8. Uno de los bocetos pintados por Salvador Dalí, dedicado a Severo Ochoa, para el Congreso de la Sociedad Española de Bioquímica, celebrado en Madrid en 1970 y utilizado para la cartelería.
elegir cuáles son las condiciones experimentales más significativas que deben ser destacadas y, sobre sus propias conclusiones extraídas de los resultados elegidos, construir una serie de preguntas, no más de tres, que conduzcan pautadamente a un análisis lógico de lo expuesto y a unas conclusiones acertadas.
También podría ser innovador, e incluso estimulante para el alumno, plantear problemas más amplios en los cuales se relacionen aspectos tanto científicos como culturales. Un ejemplo de lo dicho podría ser:
Al olmo viejo, hendido
Por el rayo
Y en su mitad podrido,
Con las lluvias de abril y el sol de mayo
Algunas hojas verdes le han salido.
Se pregunta:
¿Cuál es el fundamento fisiológico de estos versos de Antonio Machado?
4º Conexión con otros elementos culturales.
De seguirse por completo el mandato de la Unión, el aprendizaje de la ciencia debe incardinarse en el acervo cultural común como forma más eficaz para el enriquecimiento intelectual del individuo y la comunidad y la consecución de su bienestar. En un mundo en el que el libre albedrío puede comenzarse a explicar en términos de plasticidad neuronal, donde la variablidad hace desaparecer el predeterminismo causal mientras que la epigenética anula la determinación genética absoluta y entre ambas dan paso a la libertad del individuo (Evers, 2011), en un mundo en que lo eterno e
inmutable puede intentar explicarse por la teoría de las supercuerdas (Hawking y Mlodinow, 2010), en un mundo,
finalmente, en el que llenamos nuestros cuerpos de sustancias naturales o de síntesis que nos curan nuestras enfermedades y nos alivian nuestros sufrimientos, es difícil considerar la ciencia como un mundo aparte de lo cotidiano, al margen de la cultura. Las relaciones entre Dalí y la Biología

15
Molecular, la transcendencia artística de la doble hélice a finales de los años 60 podría ser un hermoso ejemplo de este maridaje irrenunciable. El conocimiento científico de la fisiología de las plantas está presente en nuestra agricultura, en nuestra cultura gastronómica (hasta la forma de cortar las verduras puede tener su importancia en la conservación de su composición química y de sus propiedades organolépticas, según explica MacGee en 2004), en el cuidado del cuerpo a través de cosméticos dermatológicos, en el mantenimiento de nuestra salud combatiendo la enfermedad, en la decoración verde de nuestros hogares, en la correcta ordenación del territorio urbano. Podría pensarse que las plantas y su buena función están presentes en todos los órdenes de la vida. Cuando las técnicas de mercado se adelantan al consumidor y constantemente le bombardean con conceptos como cultivo ecológico o cultivo orgánico, cuando en las tiendas de moda intentan crear la conciencia del consumo responsable de productos naturales anunciando vestidos de “algodón orgánico”, el individuo debe estar preparado para responder a tanto reto. Debe ser capaz de discernir si el “mercado” es veraz o si por el contrario engaña, exagera o solo le interesa vender sin ningún tipo de escrúpulos.
Claro que se trata de querer hacerlo o no querer, como todo en esta vida, como la formación de los propios estudiantes. Lo mismo que se puede “estar” en la Universidad pero no estudiar, se puede estar simplemente en la vida actual y dejarse llevar por ella sin profundizar en su sentido y significado. Se puede comprar una camiseta para hacer deporte e inclinarse por comprarla de marca, Nike, por ejemplo. Buena marca, pensamos. Tenemos entonces dos opciones:
a) Hemos comprado una camiseta Naic (Nike), un nombre estrictamente comercial, una palabra inventada que no significa nada, pero que transmite la idea de marca, de producto confiable y caro. Aunque no haga deporte, siempre podremos presumir con ella de nuestro buen gusto y, sobre todo, de nuestro poder adquisitivo.
b) Hemos comprado una camiseta Nike, una camiseta fabricada pensando en la élite del deporte, en los triunfadores, y en hacer llegar ese espíritu de triunfo a todo el mundo porque Niké, en griego, significa Victoria y esa ha sido la intención del fabricante al bautizar sus productos, inducir al consumidor a creer en la victoria como algo posible en cualquier competición y anunciarlo desde su marca.
También dentro de la cultura está el considerar la ciencia en sus exactas dimensiones, ocupando su lugar preciso en la filosofía de la vida. La ciencia, en tanto que traducción de la naturaleza a un idioma comprensible para el ser humano, no es ni buena ni mala, sino neutra. No puede culparse a la ciencia de su utilización errónea por parte de los hombres. Después de la Gran Guerra se culpó a la química de millones de muertes por la utilización de gases tóxicos en la contienda. Después de la II Guerra Mundial la física fue la bestia negra de la conciencia colectiva tras el doble holocausto de Hiroshima y Nagasaki. Hoy día la Biotecnología es la bestia negra de ecologistas, fundamentalistas y otros movimientos de la conciencia de las masas (Vicente, 2004). El deber del docente en este caso es poner las cosas en su sitio. La

16
ciencia hace avanzar al hombre. Siempre, sin paliativos. Que el hombre en su locura utilice la ciencia para destruirse a sí mismo no es un problema de la ciencia, sino de la psicología y la política, que tienden a la tergiversación de una manera automática. En las políticas innovadoras y, en algún sentido, postmodernas actuales existe una inevitable tendencia a explicar cualquier cambio de las conductas sociales tradicionales porque el objeto del cambio es “algo que se da en la naturaleza”. Este es otro ejemplo palmario de cómo el concepto básico puede ser tergiversado. La naturaleza no está sometida a la ética, sino a la evolución y en ella podemos encontrar conductas seleccionadas que, bajo el prisma de la psicología humana, no pueden ser consideradas de otra forma que horrendas. A Gould (2000) le era particularmente atractivo el ejemplo de los icneumónidos, que ponen sus huevos en el interior de orugas vivas, a las que previamente han paralizado, de tal forma que el joven icneumónido recién nacido tiene alimento fresco a su disposición. Ni la conducta del icneumónido posee la genialidad de la inteligencia superior (característica que debemos seguir suponiendo como humana), ni el sufrimiento de la oruga ni la crueldad de la cría sirven para enseñarnos algo, porque carecen de cualquier sentido ético. Cada día es más aceptado el hecho de que el hombre vive separado de la naturaleza, de que el mundo natural no está hecho a nuestra medida y que los rasgos netamente humanos como serían (Aguirre, 1980)
“la conciencia de la responsabilidad y la libertad, de los valores éticos, se explican peor en la dinámica general de la vida y constituyen desde sus primeros desarrollos un nivel de integración cualitativamente distinto de todos los logrados por aquella fuera del hombre”
El profesor debe también saber, y el estudiante entender, que el conocimiento puede ser expresado y transmitido de diversas formas, todas ellas válidas. Puede ser expresado, además de con rigor científico, con humor, con alegría y con sentimiento. Los cambios cualitativos de la luz solar de principio y final del día, las variaciones en la razón rojo lejano/rojo, definido por Harry Smith como ζ, fueron descritos en el último cuarto del siglo XX mediante la utilización de complejos y sofisticados aparatos de medida denominados espectro-radiómetros, cuyo costo era de muchos, pero muchos miles de euros. Casi cinco siglos antes, D. Luis de Góngora escribía en la octava real inicial de su Polifemo y Galatea:
Oh excelso conde, en las purpúreas horas
Que es rosas la alba y rosicler el día.
sin aparatos millonarios, sin funciones de onda, sin saber que E = hc/λ, solamente con la percepción de sus sentidos, con el lenguaje de su poesía. Estaba ya definiendo qué cambios cualitativos sufre la luz del mediodía (rosicler) frente a la del amanecer (rosas). Todos estos acercamientos a entender la ciencia, no solamente como una profesión que nos permite un cierto nivel de bienestar, sino como una forma cultural que nos posibilita una relación más completa con los demás y con el mundo que nos rodea, debería ser una de las tareas de esa adquisición de habilidades que plantea el nuevo

17
sistema de titulaciones universitarias. El empeño en conseguir un mundo tecnificado y cómodo no puede desdeñar la formación humanística del individuo y es muy deseable que ese humanismo necesario en una sociedad técnica esté presente en todos los órdenes de la vida.
Se ha dicho que un país que no produce está condenado a importar productos del exterior y, por tanto, a vivir perpetuamente endeudado. Esto obliga a que el Grado no sea una reducción en el nivel de información ni en la calidad de la formación de los estudiantes egresados con respecto a las antiguas Licenciaturas. Pero, además, si un país no acrecienta y extiende su nivel cultural está abocado a un tercermundismo indeseable. Huntington (2000) explicaba que cuando dos países parten de idéntico PIB y uno de ellos destaca significativamente del otro en un periodo de unos diez años, lo único que justifica y avala esta diferencia en la desigual evolución de ambos es la cultura del primero respecto a la del segundo. La base cultural de un país es, en último término, lo que posibilita su emergencia como potencia política, como una voz a tener en cuenta en el conjunto de las naciones.
6º Tutorías
Las tutorías son una herramienta docente que, por desgracia, difícilmente es utilizada de una forma correcta. Tiene tres inconvenientes de entrada para nuestra idiosincrasia. El primero, la actitud de rechazo del alumno. La tutoría lo significa delante del profesor, el alumno deja de ser un bulto anónimo perdido en la clase y esto puede tener sus ventajas y sus inconvenientes. Mejor ser cauto entonces. El segundo que, el alumno, por regla general, duda de sus dudas. ¿Cuántas veces no habremos oído a nuestros alumnos la frase inicial: “Bueno, no sé si lo que voy a preguntar es una tontería”? El tercero, el temor a ser mal recibido o incluso rechazado, en el cual podrían incluirse como una de sus causas los horarios demasiado rígidos impuestos por el profesorado. Las tutorías tienen unos horarios fijos y en algunos casos, el mantenimiento de estos horarios es una cuestión de fuero, aunque todos los alumnos de ese grupo tengan otras clases a esas horas. Y aunque debe haber más, citemos por último el peor de todos, la desgana, la falta de interés, la apatía. Cierto es que hay alumnos que recurren a las tutorías con regularidad, pero son los menos.
Algunos de estos escollos al uso de la tutoría por el alumno tienen su origen en la actitud del profesor. Para que una tutoría sea efectiva, el profesor tiene que ser asequible, cercano, paciente y comprensivo, debe tener autoridad para sacar al alumno de sus dudas y rechazar el autoritarismo como elemento de distanciamiento. El autoritarismo destroza la autoridad. Incluso nos atrevemos a decir que algo, alguna vez, podrá el profesor aprender de las dudas de sus alumnos. Un diálogo entre la curiosidad y la experiencia, entre dos ansias por saber, aunque de distinto grado, no deberían ser afrontadas con tantos prejuicios.
El uso de la tutoría como elemento en la docencia nos plantea una enorme cantidad de dudas y preguntas. La tutoría, como el seminario, nunca

18
debería ser un sustitutivo de la docencia. Es responsabilidad del profesor enseñar su disciplina e intentar que el alumno aprenda lo máximo posible de ella. Es la responsabilidad del profesor frente a la sociedad y, por tanto, nunca debería hacer dejación de la misma, ni siquiera en aras de una pretendida “innovación pedagógica”. Por otra parte, el uso convencional de la tutoría como elemento utilizado por el alumno para resolver dudas de cara a un examen inminente no deja de ser una degradación conceptual de la función tutorial. Porque entendemos la tutoría como un compromiso continuo, duradero y mutuo entre profesor y alumno, un compromiso que trata de completar el desarrollo intelectual y académico del segundo, un compromiso leal entre ambas partes. Es transformar al alumno en discípulo y al profesor en maestro. Es algo más profundo y más sutil que lo que se logra bajando sesenta veces por año a un aula ocupada por cien alumnos para dar una clase de cincuenta minutos. Es algo más que resolver cuatro dudas en una entrevista de diez minutos porque mañana o pasado hay un examen de la materia. Es el seguimiento continuo, durante un periodo de tiempo acordado, del desarrollo del aprendizaje de un tema concreto, de un interés mutuo, la ayuda y la orientación en la búsqueda, el respeto por la metodología del discípulo e, incluso, ante sus equivocaciones para extraer entonce conclusiones más acertadas. Es, en último término, tiempo. Tiempo que hay que tener, que hay que sacar de una manera compensada para que el discípulo pueda seguir estudiando y el maestro investigando. Fácil no es. Por lo tanto, solo quedan dos opciones: o es imposible o es difícil. Si es imposible más valdría olvidarlo. ¿Para que vamos a engañar a nadie, incluidos nosotros mismos? Si es difícil, hagámoslo bien y no una chapuza impuesta por la falta de tiempo, el agobio y las prisas. Existen varias opciones:
1º Tutoría voluntaria: El alumno no es obligado a elegir un tutor y someterse a tutorías. Esta libertad mejora el compromiso y facilita al alumno un mejor rendimiento académico sin entrar en conflicto con la libre capacidad de elección de aquellos otros cuyos intereses primordiales pueden no concernir a nuestra propia materia, de la cual les sobra, por elección propia, con obtener un conocimiento básico.
2º Tutoría compartida: Un pequeño colectivo va a ser tutorizado por un profesor, dedicándose a la ampliación de conocimientos, desarrollo de temas transversales, implicaciones de nuestra materia con otras afines. ¿Ejemplos? Miles. ¿Por qué los jasmonatos parecen controlar procesos fisiológicos tan aparentemente alejados como tuberización en patata, tolerancia a la salinidad en tomate o actuar como un mensajero volátil que avisa de la presencia de un patógeno en un viñedo? ¿Qué significa que fitocromo y rodopsina presenten tantas semejanzas, no solo estructurales sino también funcionales? ¿Cuál es la analogía entre el potencial de acción generado por las hojas de Dionaea mecánicamente estimuladas y el generado en mi antebrazo por un estímulo semejante?
Sinceramente creemos que, antes de seguir ahondando en un sistema de tutorías falseado por las circunstancias, convendría más conceptualizar este acto docente y elegir un sistema que nos permita ejercerlo de una manera positiva, clara y honesta.

19
El resultado de este trabajo debería tomar cuerpo en forma de un texto o de una exposición a la cual tuviesen acceso el resto de los alumnos. Un diez por ciento de la nota final solo es un toma y daca, el pago de un esfuerzo y, a mi modo de ver las cosas, este esfuerzo necesitaría de una mayor recompensa. El cambio de concepción de las tutorías quizá requiriera la producción de una especie de boletín, semejante al de la SEFV, que recogiera aquellos esfuerzos que realmente merezcan la pena. Un boletín trimestral, semestral o anual, alojado en la página web del Departamento, de libre acceso a cualquier persona. Podríamos pensar que añadimos una dificultad al sistema si hay que discriminar en función de la calidad del trabajo. Pero esto es parte del sistema. ¿O es que acaso no discriminamos entre un alumno calificado con un notable y otro con un simple aprobado? En último término, unos trabajos, los mejores, podrían ser publicados in toto mientras que otros podrían ser incluidos en la web como resumen. A fin de cuentas, lo que proponemos es un estímulo más para alcanzar la excelencia en el trabajo.
4º Prácticas de Laboratorio.
Al ser la Fisiología Vegetal una ciencia eminentemente experimental, el paso de los alumnos por el laboratorio es imprescindible. En él, el alumno debe desarrollar diversos tipos de habilidades, desde las meramente manuales (manipulación del material vegetal, utilización de aparatos y equipos como balanzas, centrífugas, pHmetros, cámaras de cultivo, espectrofotómetros o equipos de electroforesis, material de vidrio, manejo de disoluciones) hasta las intelectuales (perfecta compresión de los objetivos del experimento, protocolos y desarrollo del experimento, elaboración de los resultados incluyendo el cálculo cuanto éste sea necesario y análisis de los mismos del tal manera que se facilite la obtención de conclusiones válidas, significativas y pertinentes). Por supuesto que la asistencia de un tutor es absolutamente necesaria, aunque sin coartar la creatividad del alumno siempre que éste se comprometa a consultar cualquier iniciativa antes de ponerla en práctica.
Dado que la cantidad de créditos ECTS dedicados a las prácticas es limitada, estas deben ser seleccionadas con cuidado para tratar de dotar de una “expresión experimental” puntos clave de la teoría. Sería deseable incluir en un programa práctico un acercamiento experimental, siquiera sea mínimo, a cuestiones como permeabilidad, transporte xilemático, transpiración, propiedades ópticas de los pigmentos fotosintéticos, naturaleza fotoquímica de la fotosíntesis, diferencias funcionales entre plantas C3 y C4, germinación, crecimiento y senescencia con especial referencia a su fotocontrol y a su control hormonal.
Entre las habilidades personales que el alumno puede aportar en esta parte práctica de su formación en Fisiología Vegetal podría encuadrarse el proyecto de innovación educativa que subvencionó la UCM el curso 2011-2012 y que podría resumirse de la manera siguiente. Las clases prácticas convencionales de una materia experimental, como es la Fisiología Vegetal, requieren de un laboratorio con instalaciones adecuadas, de un equipamiento costoso, de un material fungible caro y de condiciones de seguridad en el desarrollo de los diferentes experimentos. También se requiere de un profesor que dirija este desarrollo, atento a la manipulación de plantas, reactivos y

20
equipos para evitar equivocaciones, gastos innecesarios y posibles accidentes, además de hacerse cargo del aspecto meramente formativo de explicación de la naturaleza del experimento, de sus objetivos y protocolos, del cálculo y el análisis de los resultados y de extraer las condiciones adecuadas. Todo ello requiere necesariamente de la Institución y el papel del alumno no es excesivamente creativo.
Sin embargo, hay muchísimos experimentos que podrían calificarse como caseros, que no entrañan riesgo, ni un gasto considerable, ni siquiera necesidad de equipamiento alguno y que podrían ser diseñados y realizados personalmente por los estudiantes a poco que reciban los estímulos necesarios. Con un ejemplo podría entenderse mejor el desarrollo de estas habilidades personales. En clase se les ha explicado el flujo xilemático ascendente, acrópeto y las diferentes variables que lo determinan: composición del fluido xilemático, gradientes, tensión-cohesión, naturaleza y sección de los elementos xilemáticos, evapotranspiración etc. Todo ello puede ser “visualizado” formalmente y formalmente comprendido. Pero hay otra forma de visualizarlo y de, intuitivamente, entenderlo. Existen plantas que en alguna fase de su desarrollo o incluso durante periodos muy largos de su vida poseen tallos translucidos, plantas que pueden incluso formar parte de la vida familiar del alumno, como especies ornamentales que pueden decorar sus casas. Por ejemplo, las llamadas vulgarmente alegrías. Bastaría con tomar una planta joven, de tallo suficientemente translúcido, sumergir sus raíces en una disolución de tinta china de color intenso y acelerar su transpiración con un sencillo ventilador manual de aspas de plástico blando y movido a pilas. En poco tiempo, el alumno puede observar cómo la tinta negra va ascendiendo por el interior del tallo gracias a su transparencia. Incluso podría obtener fotografías secuenciadas con una sencilla cámara digital y producir un corto video con el programa adecuado. Todo con el mínimo coste y totalmente exento de riesgo. Y como este ejemplo podrían encontrarse cientos, sencillos, seguros, formativos e interesantes. De esta forma, el alumno no solo podría desarrollar habilidades experimentales sino que muy probablemente afianzaría la confianza en sus propias capacidades, aumentaría su interés por la materia y contribuiría decididamente a su formación científica.
En este Proyecto, hemos desarrollado algunas de estas aproximaciones experimentales caseras con el único objeto de señalar que el camino a seguir es del todo factible y formativo.
3. Referencias Aguirre, E. (1980): El misterio de la vida. En Biología, Alvarado, R. (ed.). Carroggio, Barcelona.
Dangl, J.L., Dietrich, R.A., Thomas, H. (2000): Senescence as programmed cell death. En Biochemistry and Molecular Biology of

21
Plants (Buchanan, B.B., Gruisem, W., Jones, R.L., eds.), pp. 1044-1100. American Society of Plant Physiologists, Rockville, Maryland.
Derrida, J. (2003): Espectros de Marx. Trotta, Madrid.
Evers, K. (2011): Neuroética. Katz Editores, Buenos Aires
Gould, S.J. (2000): Ciencia versus Religión. Un falso conflicto. Crítica, Barcelona.
Hawking, S., Mlodinow, L. (2010): El gran diseño. Crítica, Barcelona.
Heisenberg, W. (1969): Der Teil und das Ganze. Gespräche im Umfreis der Atomphysik. R. Piper & Co. Verlag, München (Existe una traducción al castellano: Diálogos sobre la física atómica. BAC, Madrid, 1972).
Hegel, G.W.F. (1989): Lecciones de Estética. Ediciones 62 S.A., Barcelona.
Huntington, S.P. (2000): Introduction. In: Harrison, L.E., Huntington, S.P. (eds.): Culture Matters. Basic Books, New York.
Husserl, E. (1996): Die Krisis der Europäischen Wissenschaften und die Transzentale Phänomenologie: Eine Einleitung in die Phänomenologische Philosophie. Felix Meiner Verlag, Hamburg.
Kruif, P. (2006): Los cazadores de microbios. Editorial Porrúa, México.
Legaz, M.E., Vicente, C. (1989): 123 Problemas de Fisiología Vegetal. Editorial Síntesis, Madrid.
McGee, H. (2004): On Food and Cooking, Scribner, N.Y.
Mühlmann, H. (1996): Die Natur der Kulturen. Entwurfeiner kulturgenetischen Theorie. Springer, Viena, NY.
Mühlmann, H. (2006): Die Ökonomiemaschine. En de Bruyn, G. (Ed.): 5 Codes. Architektur, Paranoia und Risiko in Zeiten des Terrors, pp. 227. Birkhäuser-Verlag für Arkitechtur, Basel.
Novak, J.D. (1998): Conocimiento y aprendizaje. Los mapas conceptuales como herramientas facilitadoras para las escuelas y las empresas. Alianza Editorial, Madrid.
Popper, K.R. (1994): Conjeturas y refutaciones: el desarrollo del conocimiento científico. Paidós Ibérica, Barcelona.
Sabater, B. (1998): Problemas resueltos de Fisiología Vegetal. Servicio de Publicaciones de la Universidad de Alcalá de Henares.
Sánchez, P., Gairín, J. (2008) Planificar la formación en el Espacio Europeo de Educación Superior. ICE de la Universidad Complutense, Madrid.

22
Schrödinger, E. (1944): What is Life? The Physical Aspect of the Living Cell. Cambridge University Press, Cambridge (¿Qué es la vida?, última edición en castellano publicada por Tusquests, Barcelona, 2008)
Stenger,V.J. (2007): God: the Failed Hypothesis. Prometheus Books Publishers, Amherst (Edición castellana con el título ¿Existe Dios? El gran enigma. Ediciones Robinbook, Barcelona, 2008).
Subbaiah, C.C., Sachs, M.M. (2003): Calcium-mediated responses of maize to oxygen deprivation. Research Journal of Plant Physiology, 50: 752-761.
Vicente, C. (2004): El papel de lo humanístico en la enseñanza profesionalizada. En: Ante los problemas de la Universidad española, 65 propuestas para conectarla con el futuro (F. Parra, coord.), pp. 171-182. Entrelíneas Editores, Madrid.
Vicente, C. (2009): El uso de mapas conceptuales en la programación de la asignatura de Fisiopatología Vegetal y en su desarrollo interactivo. Reduca , Ser. Fisiología Vegetal, 2: 48-60.
Vicente, C. (2010): Enseñanza participativa de la asignatura Bioproducción de metabolitos vegetales de interés industrial: Una experiencia con quince años de vida. Reduca (Biología). Ser. Fisiología Vegetal. 3: 1-17.
Watson, J.D. (2000): La doble hélice. Alianza Editorial, Madrid.

23
OBSERVACIÓN Y EXPERIMENTACIÓN

24
RAÍCES ADVENTICIAS.
Las raíces de las plantas tienen su origen en el extremo radical del eje embrionario. Después de la germinación de la semilla, crece del extremo radical una raíz primaria
que, según las especies, puede ramificarse o diversificarse. Sin embargo, las raíces adventicias no nacen del eje embrionario, sino de las partes aéreas de la planta o de los tallos subterráneos o postrados, como es el caso de la fresa. Estas raíces adventicias pueden ramificarse o no, pero suelen presentar un tamaño homogéneo cuando alcanzan su máximo desarrollo vegetativo. Son raíces fasciculadas o sistemas radicales fibrosos.
Aunque su principal función es ampliar el área edáfica de la cual la planta obtiene agua y nutrientes, existen algunos casos, como las palmeras de los géneros Socratea y Pandanus, o las especies arbóreas de gran porte del género Ficus, como F. benghalensis per ejemplo, que producen raíces adventicias para lograr una mayor estabilidad, dado su altura y la extensión de su dosel. Estas raíces reciben también el nombre de raíces fúlcreas o raíces zancos. Algunas gramíneas, como el sorgo y el maíz, pueden también desarrollar raíces fúlcreas. Suelen ser gruesas, se forman en los nudos basales y penetran al suelo donde cumplen la doble función de sostén y de absorción de nutrientes.
En la serie de fotografías adjuntas puede verse la el tronco de un ejemplar de F. macrophylla, situado en una céntrica plaza de la ciudad de Benidorm, de cuyas ramas horizontalmente extendidas nacen múltiples raíces adventicias. Estas raíces van creciendo con orientación gravitrópica positiva (hacia la superficie del suelo) hasta que finalmente penetran en el sustrato. Se desarrollan formando columnas que crecen en diámetro y además pueden unirse por concrescencia, adquiriendo el aspecto de nuevos troncos. Como las ramas quedan sostenidas por estas raíces columnares, pueden alcanzar una longitud mucho mayor que las ramas de otras especies, que tienen soporte sólo en el punto de unión con el tronco.

25
TRANSPORTE DE AGUA Y NUTRIENTES POR EL XILEMA
Las plantas absorben agua y nutrientes, tanto inorgánicos como orgánicos, desde el suelo en el que están ancladas. La absorción se realiza a través de la epidermis radicular y los pelos radicales y el transporte de masa lleva el conjunto de solutos y disolvente hasta el xilema. El xilema es un conjunto de vasos, derivados de células cuyas paredes se han lignificado y el protoplasto muere. Por tanto, se trata de un espacio vacío que forma un continuum desde la raíz hasta las nervaduras de las hojas
A lo largo de esta vía de transporte en los vasos no vivos del xilema ocurren
interacciones importantes entre los solutos, disolvente, las paredes celulares de los vasos y las células parenquimatosas que rodean el xilema. Las principales interacciones son el intercambio de cationes polivalentes en las paredes celulares, y la resorción (toma) y liberación de elementos minerales y de solutos orgánicos por las células vivas circundantes (parénquima del xilema y floema).
Hay muchas formas de visualizarlo este transporte acrópeto y de entenderlo intuitivamente. Existen plantas que en alguna fase de su desarrollo o incluso durante periodos muy largos de su vida poseen tallos translúcidos, plantas que pueden incluso formar parte de nuestra vida familiar, como las especies ornamentales que decoran nuestras casas. Por ejemplo, las llamadas vulgarmente alegrías. Bastaría con tomar una planta joven, de tallo suficientemente translúcido, sumergir sus raíces en una disolución de tinta china de color suficientemente intenso y acelerar su transpiración con un sencillo ventilador manual de aspas de plástico blando y movido a pilas. En poco tiempo, se puede observar cómo la tinta negra va ascendiendo por el interior del tallo gracias a su transparencia. Incluso podría obtener fotografías secuenciadas con una sencilla cámara digital y producir un corto video con el programa adecuado.
En la secuencia de fotografías se muestra una joven plántula de garbanzo (tres semanas de vida) cuyas raíces han sido sumergidas en agua (dos primeras fotografías de la izquierda). En un momento determinado se añade tinta china a esta agua y al cabo de unas tres horas se observa como la base del tallo se va ennegreciendo tras la penetración de la tinta en sus elementos xilemáticos para iniciar la ascensión hasta las hojas.

26
TRANSPORTE DE FOTOASIMILADOS (TRANSPORTE BASÍPETO VIA FLOEMA)
James Bonner y Arthur W. Galston, del Instituto Tecnológico de California, describen en su clásico texto Principios de Fisiología Vegetal, publicado en castellano en 1961, que los primeros experimentos realizados para el estudio del transporte de
azúcares por el floema consistieron en el descortezamiento anular de troncos leñosos. Si se separa de un tronco leñoso un anillo cortical que contenga los tubos cribosos, tiene lugar, inmediatamente después del descortezamiento, un crecimiento e hinchazón acompañados de una acumulación de azúcares y
esto solo sucede cuando en la parte superior del tallo existen hojas verdes fotosintéticamente activas. Es fácil intuir entonces que los azúcares formados en las hojas, al ser transportados basípetamente por el floema, se acumulan en los espacios anteriores a la herida dado que la continuidad de los vasos conductores ha sido traumáticamente rota. En la figura superior a la izquierda de la página, tomada del libro de Bonner y Galston, se ilustra este fenómeno.
Sin embargo, no es necesaria la cirugía para obtener esta información. En un vivero de La Cabrera (Madrid) hemos observado troncos de plátano de sombra (Platanus x hispanicus, un híbrido de P. orientalis y P. occidentalis) que ha sido anillado mediante una abrazadera metálica, forrada de caucho para no hendir el tejido, fuertemente apretada sobre el tronco. Esta presión mecánica constriñe el floema externo, impidiendo que el flujo basípeto circule a la velocidad normal, o anulándolo incluso, con lo cual los azúcares fotosintetizados por las hojas sobre el anillamiento se van acumulando progresivamente en la parte superior de la zona comprimida (fotografía de la derecha), formando un rodete engrosado semejante al descrito previamente.

27
GUTACION
La gutación, fenómeno observable como pequeñas gotas de agua en la epidermis foliar, se produce cuando la planta está en condiciones que favorecen la absorción rápida de agua y minerales y una transpiración mínima, como sucede c durante la noche on las plantas que crecen en suelos húmedos. Esta exudación se realiza a través de hidatodos, un tipo especial de estoma.
Los iones minerales absorbidos de noche son bombeados al apoplasto que rodea a los elementos del xilema. Esta pérdida de solutos hace que disminuya el potencial agua en los elementos del xilema, generando un ingreso de agua desde las células circundantes. Al aumentar la presión dentro del xilema el agua es forzada eventualmente a salir a través de los hidatodos foliares. Las gotas pueden resbalar por la superficie de las hojas y caer al suelo, lo que supone una pérdida neta de agua y sales minerales, pueden evaporarse, dejando un depósito de sales sobre la epidermis foliar, o pueden ser reabsorbidas.
Los hidatodos se hallan distribuidos en muchas especies de Angiospermas (350 géneros pertenecientes a 115 familias botánicas). En plantas tropicales como Colocasia (Araceae), la cantidad eliminada por noche supera los 300 ml de agua.
En las fotografías adjuntas se muestran hojas de Dieffenbachia bowmanii, planta de interior, perteneciente a la familia de las Araceas, una hora después de haber sido abundantemente regada. Las gotitas exudadas de los hidatodos han ido resbalando sobre el haz inclinado hasta reunirse y comenzar a gotear al suelo desde el extremo apical del limbo foliar.

28
Atmósfera libre Atmósferas de saturación de vapor de agua Capa límite
hidrodinámica
Célula de sostén
Células oclusivas
Ostiolo
Atmósfera libre Atmósferas de saturación de vapor de agua Capa límite
hidrodinámica
Célula de sostén
Células oclusivas
Ostiolo
ESTOMAS
La transpiración tiene lugar en su casi totalidad a través de los estomas. Al ser un fenómeno de pérdida de vapor de agua proporcional al perímetro del poro, su grado de apertura es el primer parámetro determinante del volumen de vapor difundido. El grado de apertura puede ser relacionado con el volumen del par de células de guarda.
Los estomas son grupos de dos células epidérmicas especializadas cuya función es regular el intercambio gaseoso y la transpiración Se hallan en los órganos verdes aéreas de la planta, particularmente en las hojas, en las que se distribuyen en el haz y más frecuentemente en el envés. Su número oscila entre 22 y 2.230 por mm2. Las raíces, por tanto, no tienen estomas, ni las plantas parásitas etioladas, como Monotropa y Neottia. Los órganos etiolados de las plantas, como las hojas variegadas, pueden tener estomas pero no son funcionales, al igual que los de los pétalos.
Cada estoma está formado por 2 células especializadas llamadas oclusivas que dejan entre sí una abertura llamada ostiolo o poro. En muchas plantas hay 2 o más células adyacentes a las oclusivas y asociadas funcionalmente a ellas, denominadas células de sostén. .Las células oclusivas muestran una estructura diferencial de sus paredes celulares de tal forma que éstas están fuertemente engrosadas en la cara que conforma la cavidad central del estoma mientras que la zona de contacto con las células de sostén es más delgada. Esto hace que la presión de turgor se ejerza fundamentalmente sobre la cara más delgada de la pared, provocando la retracción parcial de la cada más engrosada, con lo que el ostiolo se abre. La apertura del estoma se lleva a cabo mediante un aumento del volumen celular. De acuerdo con esto, el mecanismo comienza con una fase de tensión

29
(Spannungsphase) a la que sigue una fase de apertura del estoma (Motorphase). Los cambios en el volumen de las células de guarda van a incrementar la presión de turgor que, ejerciéndose sobre las paredes celulares, causarán una extensión de sus polímeros, de naturaleza muy distinta a la lograda durante el crecimiento por aumento del volumen celular ya que aquí no hay ruptura micelar, sino cambios de configuración inducidos por tensiones físicas. Estos cambios pueden resumirse en un acercamiento de los polímeros vecinos y un alejamiento de sus sitios de cruzamiento. El ostiolo conduce a un amplio espacio intercelular llamado cámara subestomática, que ponen en comunicación el sistema de espacios intercelulares del parenquima con el aire exterior.
Dado que el vapor de agua sale de la cámara subestomática a la atmósfera exterior por los bordes del ostiolo (eflujo de vapor de agua proporcional al perímetro del poro, no a su área) es necesario que los estomas estén lo suficientemente alejados entre sí para evitar la superposición de sus atmósferas individuales de saturación en ausencia de viento, dado que esta superposición sobresaturaría el aire en los bordes del ostiolo y opondría una mayor resistencia a la salida de nuevas moléculas de agua en estado de vapor.
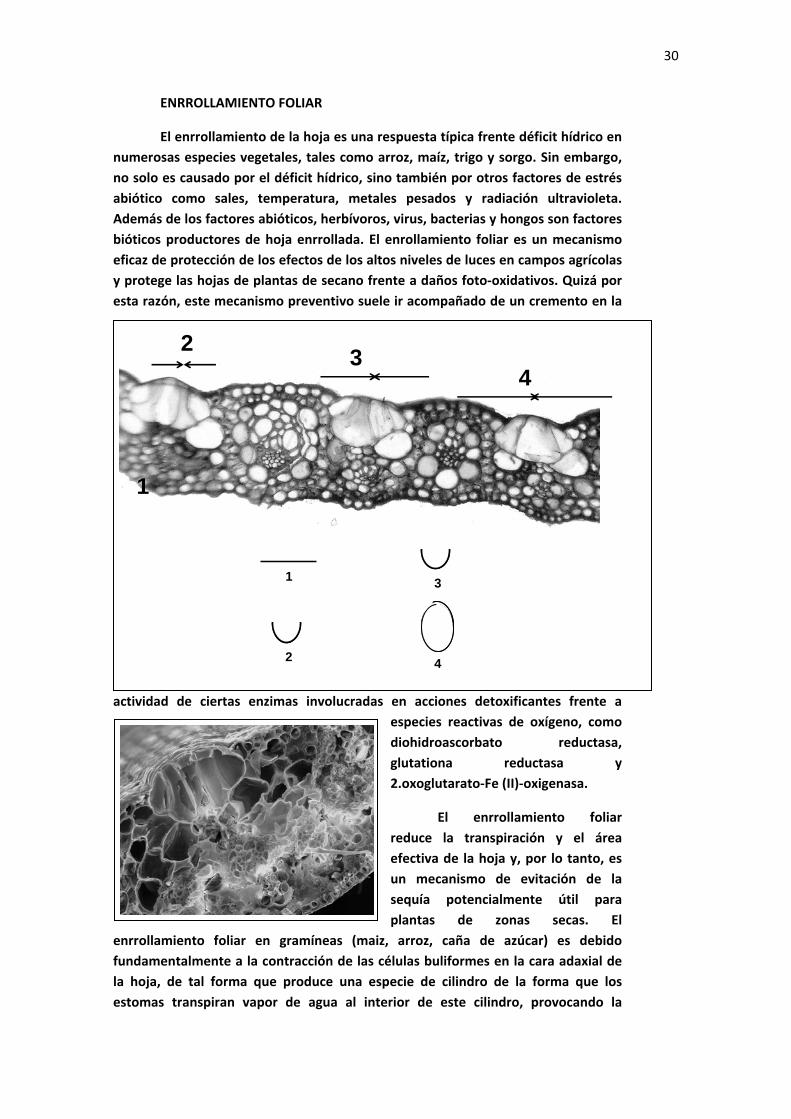
30
ENRROLLAMIENTO FOLIAR
El enrrollamiento de la hoja es una respuesta típica frente déficit hídrico en numerosas especies vegetales, tales como arroz, maíz, trigo y sorgo. Sin embargo, no solo es causado por el déficit hídrico, sino también por otros factores de estrés abiótico como sales, temperatura, metales pesados y radiación ultravioleta. Además de los factores abióticos, herbívoros, virus, bacterias y hongos son factores bióticos productores de hoja enrrollada. El enrollamiento foliar es un mecanismo eficaz de protección de los efectos de los altos niveles de luces en campos agrícolas y protege las hojas de plantas de secano frente a daños foto‐oxidativos. Quizá por esta razón, este mecanismo preventivo suele ir acompañado de un cremento en la
actividad de ciertas enzimas involucradas en acciones detoxificantes frente a especies reactivas de oxígeno, como diohidroascorbato reductasa, glutationa reductasa y 2.oxoglutarato‐Fe (II)‐oxigenasa.
El enrrollamiento foliar reduce la transpiración y el área efectiva de la hoja y, por lo tanto, es un mecanismo de evitación de la sequía potencialmente útil para plantas de zonas secas. El
enrrollamiento foliar en gramíneas (maiz, arroz, caña de azúcar) es debido fundamentalmente a la contracción de las células buliformes en la cara adaxial de la hoja, de tal forma que produce una especie de cilindro de la forma que los estomas transpiran vapor de agua al interior de este cilindro, provocando la
1
2
3
4
1
2 34

31
saturación de una atmósfera tan cerrada. Esto trae como consecuencia la drástica disminución de la transpiración.
En la siguiente página se muestra un claro ejemplo de este proceso. Una especie del género Dracaena que, en condiciones de buen suministro de agua muestra sus hojas abiertas y expandidas, las va enrollando progresivamente conforme va avanzando un proceso de sequía experimental.

32
LENTICELAS
Las lenticelas son partes delimitadas de la felodermis en las que el felógeno es más activo que en otras y produce un tejido que posee numerosos espacios intercelulares. El propio felógeno de la lenticela está provisto de dichos espacios. El felógeno es un meristemo lateral que forma el súber (= corcho o felema), tejido protector con células muertas en su madurez, constituido por células hijas localizadas hacia fuera tras la división de la célula meristemática, y la felodermis, constituida por células hijas hacia dentro tras la división de la célula meristemática, un tejido parenquimatoso formado por células vivas. Debido a esta estructura, la lenticela
permite el intercambio de gases con la atmósfera desde raíces, tallos y ramas. Hacia fuera, la lenticela se presenta generalmente como una masa alargada horizontal o verticalmente formada por células laxas que sobresalen de la superficie a través de una fisura de la peridermis. La peridermis es el tejido de protección secundario que reemplaza a la epidermis en tallos y raíces que tienen crecimiento secundario. Se encuentra también en dicotiledóneas herbáceas, en las partes más viejas de tallos y raíces. Es la corteza externa.
El felógeno de una lenticela se continúa con el de la peridermis suberosa, pero normalmente se curva hacia dentro, de manera que parece alcanzar mayor profundidad. El tejido laxo formado por el felógeno de la lenticela hacia el exterior es el llamado tejido de relleno mientras que hacia el interior constituye la felodermis. Durante la estación de crecimiento, las lenticelas están abiertas pero, aún así, son muy pocas las bacterias y hongos que penetran en los tejidos a través de ellas y avanzan posteriormente entre las capas celulares más profundas. La mayor parte de los patógenos que penetran a través de lenticelas también lo hacen a través de heridas de una forma más eficiente.
En las peridermis iniciadas en capas subepidérmicas, las primeras lenticelas se forman con frecuencia debajo de los estomas. Las células parenquimáticas que
CutículaEpidermisCélulas del corchoFelógenoFelodermo
Corteza
Cutícula
Epidermis
Corcho
Felógeno
Felodermo
Corteza

33
rodean la cámara subestomática se dividen activamente y forman el felógeno de la lenticela, que produce felodermis hacia adentro y células de relleno hacia afuera. Estas últimas se acumulan en cantidad, rompen la epidermis y sobresalen hacia afuera.
Muchas veces las lenticelas se forman antes de que se inicie el desarrollo de la peridermis, como ocurre en los tallos de Aristolochia y Menispermum. Las manzanas presentan lenticelas, pero el fruto está protegido por epidermis, nunca forma una peridermis. En otros casos las lenticelas son independientes de los estomas, igual que en las peridermis iniciadas en capas profundas del tallo: una porción del felógeno deja de producir súber, y forma tejido de relleno
hacia afuera, como en Hedera helix.
Las lenticelas de las dicotiledóneas pueden ser de tres tipos: 1) Sin capa de cierre, en las que el tejido de relleno está formado por células suberizadas bastante compactas, con espacios intercelulares. Pueden presentar capas anuales de crecimiento. Ejemplos: Persea, Liriodendron, Magnolia, Salix.
2) Con una capa de cierre, capa suberizada que se forma al final de la estación. El tejido de relleno está formado por una masa laxa de células. Ejemplos: Fraxinus, Quercus, Tilia, Sambucus
3) Con varias capas de cierre, siendo dichas capas y producidas anualmente. Son suberizadas y alternan con tejido laxo no suberizado. Las capas de cierre pueden tener una a varias líneas celulares de espesor y sujetan el tejido de relleno, de varias células de espesor. Ejemplos: Betula, Fagus, Prunus, Robinia. Son las lenticelas más especializadas.
En el corcho comercial, las lenticelas se observan en corte transversal o corte radial como largos canales rellenos de polvo parduzco. Un corcho con demasiadas lenticelas pierde porosidad (B y D en la fotografía) y no es conveniente para conservar la calidad de la botella de vino. Lo mejor es el corcho bien prensado (A) hasta soldar la lenticela, aunque también se utilizan prensados de corcho granulado.
A B C D

34
MADERA DE REACCIÓN
La madera de reacción es un recurso de diferenciación celular que emplean algunas leñosas para reorientar sus troncos o ramas en el espacio o para evadir un obstáculo que se presenta en su desarrollo. Cuando un tronco o rama es obligado a adoptar una posición horizontal, se produce una acumulación de madera de compresión en la zona inferior del órgano de tal manera que le obliga a retomar su
crecimiento vertical. Por ejemplo, un pino rodeno (Pinus pinaster) de la Sierra de Albarracín, al encontrar en su crecimiento vertical un saliente rocoso, trata de evitarlo adaptándose a su contorno, según se observa en la fotografía adjunta. Un movimiento de tierras
puede hacer que un tronco se incline y tras la formación de madera de compresión, retomar su crecimiento vertical, según se observa en la fotografía de la derecha.
La madera de compresión se produce por un aumento en la actividad mitótica del cambium vascular, seguido de un incremento del patrón de lignificación, en la parte inferior del órgano inclinado. Esto hace que se formen anillos de crecimiento excéntricos, los inferiores más anchos que los superiores, lo cual conduce al enderezamiento del órgano (ver dibujo). Esta madera de compresión es
diferente tanto anatómica como químicamente, de la madera normal. Sus traqueadas son más cortas y el depósito de lignina le confiere un color más oscuro que el que muestra la madera normal. El obstáculo que se opone al crecimiento

35
geotrópico negativo de los troncos puede ser meramente artificial, como sucede para las Bougainvillea que se muestran en la siguiente fotografía.
En estos casos, la formación de madera de compresión es un aditivo a la reorientación geotrópica negativa en leñosas, que se suma a la distribución lateral de auxina revelada en herbáceas.
En Angiospermas, la madera de reacción se forma en la cara superior de la rama o en la zona que toma contacto con un obstáculo insalvable para el crecimiento en grosor, hablándose en este caso de madera de tensión. Este fenómeno puede observarse, por ejemplo, en el aumento del diámetro de la base de los troncos de Platanus x hispanicus que adornan paseos y avenidas de nuestras ciudades. Cuando el crecimiento en grosor determina que la circunferencia de la base del tronco choca con el alcorque de cemento u hormigón, la formación de madera de tensión “desborda” el alcorque “derramándose” sobre la acera, como se muestra en la figura. O también puede observarse en los chopos de la avenida de Ramiro de Maeztu, cómo uno de ellos, presionado por el alambre de una valla, ha ido formando madera de tensión hasta envolver el obstáculo en el interior de su tronco.
La madera de tensión también colabora en reorientaciones geotrópicas, como en el caso de plantas trepadoras leñosas, en las que cada giro de circumnutación va acompañado de depósitos diferenciales de lignina en la cara superior de la espiral, como es el caso de Wisteria sinensis, una leguminosa arbórea de la
familia Fabaceae, observable en el arboretum de la E.T.S.I. de Montes, en la Ciudad Universitaria madrileña. Este depósito de lignina, que forma la madera de tensión, facilita tanto el giro como la estabilidad del la espiral trepadora, que se va cerrando sobre un soporte adecuado.

36

37
LA ADQUISICIÓN DE LA FOTOERGONIA
Una planta se genera a partir de una semilla que germina, haciendo que su eje embrionario crezca en dos direcciones del espacio, hacia arriba para formar el tallo y hacia abajo para formar la raíz o raíces. Hay muchas semillas que germinan bajo tierra, lo cual implica que en sus primeras fases del desarrollo, la plántula recién formada no recibe luz solar. Este hecho genera plántulas blancas o amarillentas, etioladas, que no pueden realizar fotosíntesis y que, por tanto, deben crecer utilizando las sustancias nutritivas almacenadas en los cotiledones y el agua
y las sales minerales que toma del suelo. El ejemplo cotidiano más claro que se puede aducir es la diferencia entre los espárragos blancos y los verdes, los llamados trigueros. No es que se trate de dos especies distintas. El agricultor sepulta el tallo del espárrago durante su crecimiento, acumulando tierra sobre el tallo para evitar la percepción de la luz. De esta manera, el tallo (el espárrago) que se forma es blanco.
La planta comienza a enverdecer cuando rompe la superficie del suelo y comienza a acrecer al aire libre, recibiendo la luz del sol. Ya que el color verde es debido a la producción de clorofilas, debemos buscar en este proceso la causa de la desetiolación a la luz. En la síntesis de clorofilas, hay un paso ya muy al final de la ruta, cuando ya se ha incluido el magnesio en el núcleo tetrapirrólico, que requiere específicamente luz para llevarse a cabo. Se trata, por tanto de una reacción fotoquímica que transforma la protoclorofilida‐holocromo en clorofilida. La planta empleará una enzima, una esterasa para unir el alcohol denominado fitol a un resto ácido de la clorofilida y formar finalmente clorofila. Pero en ausencia de luz no hay formación de clorofilida y, por tanto, la planta no puede sintetizar las clorofilas necesarias para sustentar su fotoergonia.
Esta síntesis procede paulatinamente. Así, puede verse como en caducifolias (pinos, abetos y especies relacionadas) las acículas jóvenes recién

38
formadas tienen un color verdoso pálido, casi amarillento, frente al color verde oscuro de las acículas del año anterior. Las hojas de Ficus benjamina van enverdeciendo lentamente a lo largo de dos meses o más desde su brotación, aunque algunas zonas cercanas a su nervadura media puedan sintetizar clorofilas más rápidamente que el resto del limbo foliar en función de su menor reflectancia.
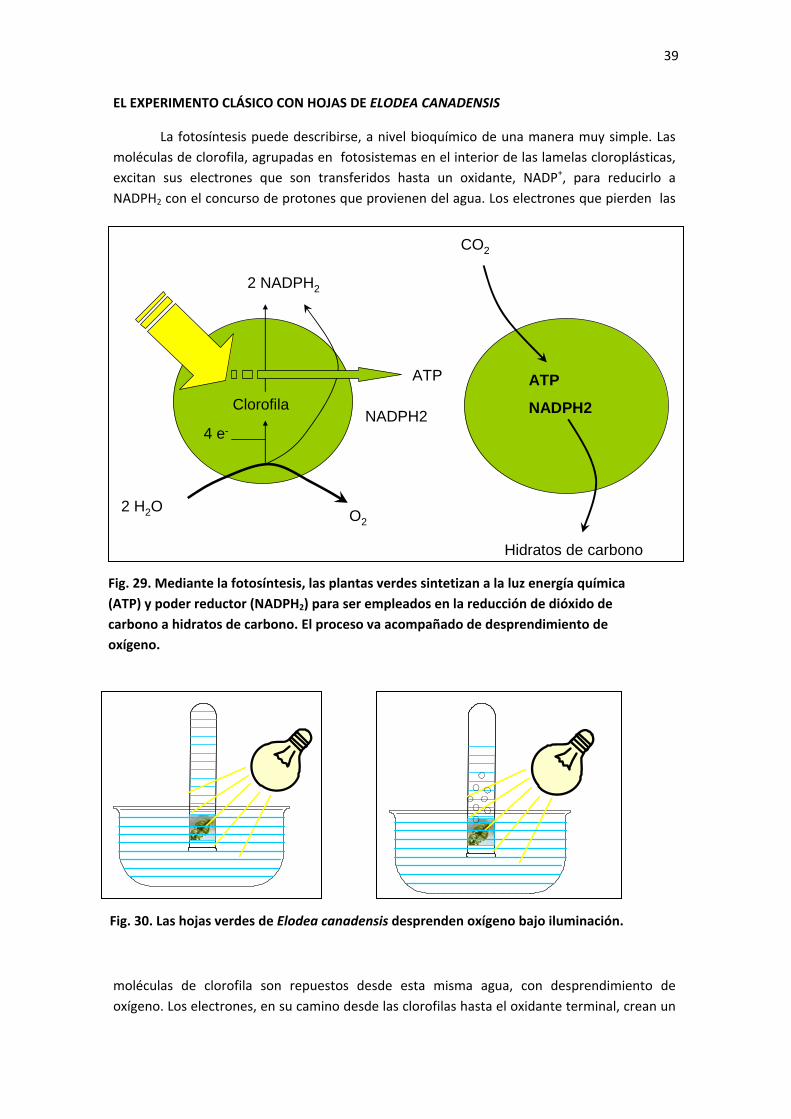
39
2 H2O O2
Clorofila
4 e-
2 NADPH2
ATP
CO2
Hidratos de carbono
ATP
NADPH2NADPH2
Fig. 29. Mediante la fotosíntesis, las plantas verdes sintetizan a la luz energía química (ATP) y poder reductor (NADPH2) para ser empleados en la reducción de dióxido de carbono a hidratos de carbono. El proceso va acompañado de desprendimiento de oxígeno.
Fig. 30. Las hojas verdes de Elodea canadensis desprenden oxígeno bajo iluminación.
EL EXPERIMENTO CLÁSICO CON HOJAS DE ELODEA CANADENSIS
La fotosíntesis puede describirse, a nivel bioquímico de una manera muy simple. Las moléculas de clorofila, agrupadas en fotosistemas en el interior de las lamelas cloroplásticas, excitan sus electrones que son transferidos hasta un oxidante, NADP+, para reducirlo a NADPH2 con el concurso de protones que provienen del agua. Los electrones que pierden las
moléculas de clorofila son repuestos desde esta misma agua, con desprendimiento de oxígeno. Los electrones, en su camino desde las clorofilas hasta el oxidante terminal, crean un

40
gradiente electroquímico a través de la membrana tilacoidal que se emplea en la síntesis de ATP. El cloroplasto, mediante su maquinaria enzimática, es capaz de reducir el CO2 atmosférico a hidratos de carbono, utilizando el ATP y el poder reductor previamente generados (figura 29).
Macroscópicamente hablando, podemos entonces “visualizar” la fotosíntesis como un intercambio de gases en el que se consume dióxido de carbono y se produce oxígeno. Un experimento clásico y demostrativo usando hojas de Elodea canadensis consiste en lo siguiente. Se introducen unas hojas de la planta en un tubo de vidrio transparente y se llena completamente de agua. Tapando su boca con la yema del dedo pulgar, se introduce invertido en un recipiente que contiene agua, de tal forma que el fondo del tubo, ahora en la zona superior, no contenga nada de aire. Se iluminan las hojas con una luz potente. El aparato fotosintético comenzará a consumir el dióxido de carbono disuelto en el agua y comenzará a desprender burbujas de oxígeno, de tal forma que, al cabo del tiempo, aparecerá un volumen “vacío” realmente llego de oxígeno, en el fondo del tubo. Este fenómeno no se dará si las hojas permanecen en oscuridad absoluta durante el mismo tiempo que en el experimento anterior (figura 30).

41
FOTOSÍNTESIS Y PRODUCCIÓN DE ALMIDÓN
Las conocidas como plantas C3 son aquellas que fijan el dióxido de carbono atmosférico sobre un azúcar de cinco átomos de carbono, doblemente fosforilado, para producir azúcares tres átomos de carbono (de ahí el nombre de plantas C3) que son empleados para la producción de hexosas (fructosa, glucosa, galactosa etc.). La planta puede emplear estos azúcares con diversos fines. El primero de ellos es producir sacarosa como disacárido exportable al resto de la planta y ser empleado como sustrato de la respiración aerobia previa su hidrólisis en los dos monosacáridos constituyentes. La síntesis de sacarosa es citoplásmica. También puede utilizarlos para formar almidón en los cloroplastos y acumular este almidón como polisacárido de reserva. Por último, muchas pratenses pueden formar, a partir de la sacarosa, polímeros de fructosa, de significación osmótica y de reserva, conocidos con el nombre de fructanos.
El almidón es un compuesto que reacciona con el lugol, una disolución de I2‐KI desarrollando un color azul‐morado. La reacción es tan específica que puede usarse para la detección citoquímica de granos de almidón en cortes transversales de hojas (micrografía superior).
Un experimento sencillo para demostrar la relación entre fotosíntesis y acumulación de almidón en hojas se ilustra en la
fotografía de la izquierda. Ocultando la zona izquierda de una hoja de trébol (Trifolium repens) con papel de aluminio, la luz no penetra y por tanto esa valva no puede realizar fotosíntesis. Al cabo de unas horas de iluminación, las hojas se sumergen en una disolución de lugol. Al rato, se observa que la valva que no ha realizado fotosíntesis sigue mostrando su coloración verde característica mientras que la otra mitad, que ha sido iluminada al igual que las hojas circundantes, se tiñe de morado tras la reacción del almidón acumulado con el de I2‐KI.

42
AUXINAAUXINA
TROPISMOS
Los tropismos son reorientaciones de órganos vegetales en el espacio siguiendo la dirección de un estímulo vectorial. Si al tallo de una plántula de garbanzo se la sitúa ceca de una ventana que le proporciona luz lateral, sus tallos se orientan hacia esa luz lateral, como muestra la fotografía. Unas macetas de geranio, colocadas
en el barandal de una terraza que solo recibe luz solar directa por la tarde, orientan el haz de sus hojas hacia el oeste.
El receptor de la luz direccional es una proteína de membrana celular, denominada fototropina, la cual, al ser
activada por luz azul, fosforila las proteínas PIN, colocadas en membrana celular de la cara inferior de las células, provocado su fosforilación y su desplazamiento hacia la cara lateral de la membrana mas alejada de la fuente de luz. Igual sucede con las proteínas AUX transportadoras de auxina,
originariamente ubicadas en la membrana de la cara superior de las células. Las proteínas PIN facilitan el transporte de auxina desde una célula donadora, mientras que las proteínas AUX internalizan la auxina en la célula siguiente, la célula receptora. De esta manera, la auxina se va acumulando en las células laterales del órgano vegetal más alejadas de la fuente direccional de luz, las células en sombra. Este aporte suplementario de auxina por transporte lateral hace que las células en sombra crezcan más deprisa que las células iluminadas y, por tanto, el tallo se incline hacia la fuente lateral de luz, dado que la auxina es un efector positivo de la elongación celular.

43
Tiempo 0 30 min
60 min 2 h
La orientación en vertical de los tallos y las raíces se denomina geotropismo, porque se orientan en la dirección del campo gravitatorio que perciben los órganos sensoriales de las plantas, situados en sus células más apicales. Ya que las raíces crecen hacia abajo, no solo siguen la dirección del campo gravitatorio, sino también su sentido, mientras que el crecimiento de los tallos hacia arriba sigue la dirección
del campo pero en sentido contrario. Por ello, se dice que las raíces muestran un geotropismo positivo mientras que los tallos siguen desarrollan un geotropismo negativo. El mecanismo de reorientación cuando, por ejemplo, se coloca una plántula de garbanzo en horizontal, es casi idéntico al descrito para el fototropismo, salvo en lo que se refiere al órgano receptor del estímulo
externo. Para la percepción de la dirección del campo gravitatorio, las plantas han diferenciado amiloplastos en las células de la cofia radicular y en las células apicales de sus tallos que cambian de orientación con cada cambio que sufre el órgano, tendiendo siempre a ocupar una posición basal en la célula. Este movimiento de desplazamiento interacciona con el citoesqueleto de tal manera que se genera un potencial de acción que cambia el sentido del transporte de la auxina.
El cambio de orientación del órgano comienza a ser percibido visualmente
AUXINAAUXINA

44
pocos minutos después de recibido el estímulo gravitrópico, completándose la reorientación en un periodo máximo de 2‐3‐h para órganos no lignificados

45
FOTONASTIAS
Las fotonastias son reorientaciones de órganos vegetales en el espación como respuesta a la perceopción de variaciones en la intensidad de luz percibida. Fotonastias típicas serían a apertura de flores durante el día y su cierre al anochecer. Las fotonastias. requieren fotorreceptores específicos denominados fototropinas. Son proteínas con cromóforos de naturaleza flavínica que inducen, tras la percepción del cruce de los umbrales de fotosensibilidad, una cascada de transducción de energía que permite el traslado de otra clase de proteínas, las llamadas PIN, de la
zona basal de la membrana celular a la cara lateral de la misma. En esta cascada de señalización, el desplazamiento viene provocado por la actividad proteína quinasa (serina‐treonina quinasa) de las fototropinas. Las proteínas PIN funcionan como

46
“facilitadoras” del transporte de auxinas y actúan asociadas a las proteínas AUX, permeasas transportadoras de auxina, localizadas en la zona apical de la membrana celular. La acción conjunta de ambas clases de proteínas, PIN y AUX, obligan a un transporte basípeto de la auxina. Sin embargo, este sentido basípeto del transporte puede ser cambiado transitoriamente si las proteínas PIN son desplazadas a las caras laterales de la célula. Si las células de la cara externa de los sépalos de una flor de Hybiscus reciben más auxina al percibir las fototropinas la disminución de la intensidad luminosa del atardecer, estas células crecerán más deprisa que las células de la cara interna del sépalo y éste se curvará hacia dentro. En el caso del hibisco, flor que ilustra este ejemplo, el crecimiento diferencial de los sépalos hace que estos se imbriquen entre sí en un proceso semejante al cierre de las láminas de un diafragma de cámara fotográfica. El resultado es que la corola cierra durante la noche. El incremento en la intensidad luminosa tras el amanecer desencadenaría el proceso inverso y la corola abre durante el día.

47
LA FLORACIÓN DEL ALMENDRO
La floración de las plantas depende de la percepción de la longitud relativa del día y la noche. Una vez que se ha completado la fase de crecimiento vegetativo,. La planta tiene suficientes recursos energéticos para hacer frente a la transición a la fase reproductiva. Las plantas de día largo son aquellas que florecen cuando la longitud del día supera un valor umbral, denominado longitud crítica del día, que es genéticamente impuesto. Las plantas de día corto, por el contrario, son aquellas que florecen cuando la duración del día es inferior a este valor crítico. Por ello, en nuestras latitudes, es sorprendente que almendros y cerezos comiencen a florece en pleno invierno, finales de enero y comienzos de febrero, cuando el pie
de planta ha perdido todas sus hojas y las temperaturas son realmente bajas. Sin embargo, el diseño floral ha comenzado a finales del verano. El desarrollo de estambre continua durante el otoño, y las yemas florales entran en letargo con un tejido esporogénico totalmente desarrollado. Este pre‐desarrollo se lleva a cabo cuando aún sobre la planta persisten sus hojas fotosintéticamente activas, por lo que su ausencia en las primeras semanas del invierno no afecta al desarrollo de la yema. Aunque no se
observa ningún cambio anatómico durante el letargo, la rotura del mismo ocurre tras una secuencia clara de acontecimientos. El primero de ellos es el alargamiento sensible del fotoperiodo. Los días comienzan a alargarse, lo que significa que almendros y cerezos son plantas de día largo, con un bajo valor de longitud crítica del día. La percepción de un fotoperiodo de 10 horas es suficiente para romper el letargo de la yema. El almidón se ha acumulado durante el otoño en sitios particulares, previniendo cualquier desarrollo en aquellas áreas hasta su movilización. Los haces vasculares se desarrollan entonces y las células de madre de polen entran en meiosis seguida del desarrollo de microspora. La percepción del alargamiento de los días, aún a pesar de las bajas temperaturas, permite la activación de una caja de genes consistente al menos en AGAMOUS (AG), SEEDSTICK (STK) y SEPALLATA (SEP), genes de identidad del órgano floral que juegan un papel fundamental en el desarrollo de los estambres, carpelo y óvulo.

48
LOS TALLOS FLORALES
La transición del crecimiento vegetativo a la etapa reproductora en plantas con flores se realiza tras la percepción de señales, tanto externas como internas, que determinan la suspensión de la actividad de algunos genes e inician la actividad de
otros. Se sabe, por ejemplo, que el gen denominado TERMINAL FLOWER (TFL1) determina la actividad del crecimiento vegetativo continuo de los meristemos. El desencadenamiento de la producción de transcritos por los genes CONSTANTS (CO), FLOWERING LUCUS T (FT), FLOWERING LOCUS C (FLC) y FLOWERING LOCUS D (FLD) inhibe la actividad de TFL1 y permiten la transición del meristemo
vegetativo al meristemo floral. La mayor parte de estos genes se activan mediante percepción de señales tanto externas como internas, es decir, son genes para la percepción de señales. Así, CO y FT se activan mediante la percepción de señales luminosas, como son la duración relativa del día o el color dominante de la luz incidente. El gen FLC se activa por bajas temperaturas, mientras que el gen FLD es activado por un factor de crecimiento endógeno, de naturaleza hormonal, que se conoce con el nombre genérico de giberelina. Sin embargo, no está claro en este último caso que las giberelinas sean realmente señales de iniciación de la floración o factores requeridos para una respuesta de crecimiento vegetativo particular, necesaria para la formación ulterior de flores.
Algunas plantas tienen el tallo tan corto que parece no existir, y sus hojas crecen muy juntas a ras del suelo. Son las llamadas plantas en roseta. En algunos casos, como la violeta africana (Saintpaulia ionantha), la planta presenta una roseta basal de donde surgen las hojas y los tallos florales, ambos de la misma longitud. Los tallos florales no presentan hojas y en su ápice aparecen las flores. En otros, como las especies crasas, del centro de la roseta sale un tallo floral que puede alcanzar una longitud de varios metros, sobre el que se diferencian los órganos florales. Un ejemplo clásico es la pita (género Agave). El crecimiento de este tallo es un requerimiento absoluto para que los meristemos florales comiencen su actividad. El crecimiento de este tallo requiere, a su vez, un aporte especial de giberelinas, necesarias para la división vegetativa de sus células meristemáticas.

49
Cianidina‐3‐O‐diglucósido‐5‐O‐glicósido
El principal componente de la fracción fenólica de la lombarda
OHO
O
OH
OH
OO
OH
HO
HOO
OHOHHO
OH
O
O
OH
HO
OH
HO
PIGMENTOS DE LAS PLANTAS
Muchos órganos vegetales incluso las hojas adquieren coloraciones diferentes gracias a la síntesis y acumulación de una gran variedad de pigmentos, la mayor parte de ellos de naturaleza fenólica: Taninos, fenilpropanos, flavonoides etc. Incluso esa coloración puede cambiar según las circunstancias ambientales o la manipulación que el hombre haga de dichas plantas, ya que muchas de estas sustancias pueden comportarse como indicadores de pH o indicadores redox. Por
ejemplo, todo el mundo sabe que la lombarda tiene un color morado característicos. En muchas ocasiones, el comensal que va a
comer un planto de lombarda, simplemente hervida y rehogada, suele añadirle un poco de vinagre para acentuar el sabor y observa entonces como el color morado intenso vira a rosado. Pero podemos complicar un poco más este sencillo experimento. Lo mismo que el color de la hoja de lombarda cocida vira a rosa, igual el agua en el que hemos cocido la planta vira a rosado tras añadirle unas gotas de vinagre. Ahora, podemos añadir a ese caldo algún compuesto alcalino que tengamos a mano en casa. Puede ser sosa, o el bicarbonato que usamos para
corregir la hiperclorhidria, o simplemente perborato del que se usa para blanquear la ropa. Observaremos que el color del líquido vira instantáneamente a un violeta‐azul muy denso.
El mecanismo es fácil. El color morado de la hoja de lombarda viene dado por la acumulación de las vacuolas de sus células parenquimatosas
de un flavonoides denominado cianidina, principalmente en su forma glicosilada. Este flavonoides es un indicador de pH, de tal manera que cuando añadimos vinagre, la acidificación provoca la protonación del pigmento, que vira a un color rojo‐rosado. Sin embargo, cuando se añade suficiente sosa como para, no solo neutralizar el pH sino virarlo a básico, la forma desprotonada del compuesto proporciona el color morado‐azul que observamos en nuestro pequeño experimento casero.

50
Y como éste, hay muchas más transformaciones observables en la cocina‐ Los cardos hay que “pelarlos” con cuchillo y guantes, ya que sí no, las manos se ennegrecen y es dificilísimo desprenderse de ese color. Hay muchos alimentos vegetales que ennegrecen cuando se les corta y se les deja al aire. Suele decirse que se trata de una oxidación y es muy cierto. Hagamos otro experimento sencillo.
Tomemos una alcachofa y cortémosla por la mitad según el eje transversal. Una de las dos mitades se colocará en un plato con la cara de corte hacia arriba. Con la otra
mitad se hará lo mismo, pero será rociada con abundante zumo de limón. Al cabo de muy pocos minutos, uno o dos, veremos cómo la mitad seca de la alcachofa se va
OHO
OH
OH
OH
OH
OHO
OH
OH
O
OH
pH ácido (rojo) pH básico (violeta‐azul)
Cianidina

51
COOH
OH
OH
COOH
O
OH
H
COOH
O
OH
e
H
COOH
O
O
COOH
O
O
e
.
.
oscureciendo progresivamente mientras que la mitad rociada con limón mantiene su color claro.
La explicación de este oscurecimiento es sencilla. La alcachofa, como tantas otras verduras y frutas, tiene una importante
concentración de fenoles. También, como en el caso de
la lombarda, produce flavonoides, principalmente luteolina, que es el pigmento que proporciona ese color rosado de la base de las brácteas más externas y de las que conforman el corazón de la alcahofa. El resto acumula compuestos de tipo fenilpropanoide, principalmente ácido cafeico y compuestos relacionados. Estos fenoles son sustancias indicadoras del estado redox, de tal manera que pueden
oxidarse con facilidad en contacto con el aire, mientras que esa oxidación es eficientemente evitada si se añade sobre la zona de corte una disolución de algún antioxidante fuerte, como es el zumo de limón, que contiene ácido ascórbico.
En la secuencia adjunta se ilustra la oxidación (el oscurecimiento) en cuatro pasos del ácido 3,4‐dihidroxicinámico, el precursor inmediato del ácido cafeico, hasta llegar a la forma doblemente oxidada, de color pardo.
OHO
OH O
OH
OH
Luteolina