LEOQ Clase V Catabolismo - Anabolismo Aquellas reacciones ...

INTRODUCCIÓN AL
METABOLISMO
CELULAR
APUNTES DE ANATOMÍA APLICADA
IES LAS VIÑAS MANILVA. CURSO 15-16
Susana Serradilla

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
2
ÍNDICE
1.- INTRODUCCIÓN ……………………………………………………………………………………………… 3
2.- CONCETOS CLAVE
A. Metabolismo ……………………………………………………………………….…. 4
B. La energía como enlace entre las rutas metabólicas ………………...……………….. 5
C. Enzimas: biocatalizadores de las reacciones metabólicas ……………………………..8
D. Moléculas que intervienen en el metabolismo ………………......………………….. 11
E. Catabolismo ………………………………………………………………………….11
F. Anabolismo …………………………………………………………………………..14
3.- CATABOLISMO DE GLÚCIDOS A. Glucólisis …………………………………………………………………………… 15
B. Respiración aerobia ………………………………………………………………… 17
C. Fermentación ………………………………..……………………………………… 21
4.- CATABOLISMO DE LÍPIDOS.……………………………………………………………………………22
5.- CATABOLISMO DE PROTEÍNAS……………………………………………………………………….24
6.- CATABOLISMO DE ÁCIDOS NUCLEICOS ……………………………………………………….…27
7.- ANABOLISMO DE LAS PRINCIALES BIOMOLÉCULAS……………………………………….28
8.- CARACTERÍSTICAS METABÓLICAS DE LOS TEJIDOS Y ÓRGANOS…………………….30
9.- REGULACIÓN DEL METABOLISMO……………………………………………………………..…...36
10.- METABOLISMO EN REPOSO Y EN EJERCICO………………………………………………….38

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
3
TEMA 2: INTRODUCCIÓN AL METABOLISMO CELULAR
1.- INTRODUCCIÓN:
Como resultado de la función digestiva a las células les llegan moléculas simples:
monosacáridos, aminoácidos y triacilglicéridos… La obtención de energía por las células se realiza
mediante la oxidación de estas moléculas, siendo necesario que el oxígeno aportado por el sistema de
aporte de oxígeno sea el adecuado a las necesidades del organismo. Los mecanismos de regulación
cardiovascular y respiratoria tienen que ajustar las variables correspondientes, a fin de suministrar el
oxígeno en función de las necesidades celulares y eliminar el dióxido de carbono producido.
Los seres vivos son considerados SISTEMAS ABIERTOS, en EQUILIBRIO DINÁMICO
que realizan un TRABAJO, es decir:
Existe un intercambio de sustancias y energía entre el ser vivo y el entorno que lo
rodea.
Los seres vivos mantienen su equilibrio interno (homeostasis). Mantienen sus
variables (temperatura, salinidad, pH…) dentro de unos intervalos de confianza o sin
variación a lo largo del tiempo.
Las células, pese a ser sistemas en equilibrio (las leyes de la termodinámica postulan
que los sistemas en equilibrio no realizan trabajo), deben realizar diversos trabajos,
mecánico, osmótico, eléctrico y químico para crecer, reproducirse… su intercambio
continuo de materia y energía con el entorno posibilita que realicen ese trabajo, sin
dejar de estar en equilibrio.
Según la forma de obtener energía, oxígeno y la fuente de carbono (materia) podemos hablar
de los siguientes tipos de seres vivos:
Según su fuente de carbono:
AUTÓTROFOS: Su fuente de carbono es inorgánica (CO2 de la atmósfera)
HETERÓTROFOS: su fuente de carbono es orgánica, es decir, la obtienen de
otros organismos.
Según su fuente de energía:
FOTOTROFOS: luz como fuente de energía
QUIMIOTROFOS: reacciones de oxidación – reducción
Según su fuente de oxígeno:
AEROBIOS: El oxígeno molecular (O2) es el último receptor de electrones
ANAEROBIOS: utilizan otra molécula diferente al oxígeno como aceptor
de electrones.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
4
2.- CONCEPTOS CLAVE:
A.- METABOLISMO
Conjunto de reacciones bioquímicas que se producen en las células, en los tejidos, y por
extensión, en el organismo, con un doble objetivo:
Obtener y utilizar la energía para desarrollar trabajo
Formar compuestos macromoleculares.
De forma concreta definimos METABOLISMO CELULAR como el conjunto de
reacciones altamente organizadas que ocurren en el interior de la célula, mediante las cuales la célula:
Obtiene energía a partir de los materiales captados del medio.
Fabrica moléculas de recambio para sustituir las gastadas.
Sintetiza otras moléculas necesarias para el crecimiento o para realizar cualquier otra
función celular.
Los organismos pluricelulares han desarrollado tal grado de especialización, que el
metabolismo individual se adapta a la función que deben llevar a cabo, así por ejemplo, un adipocito
tiene su metabolismo desplazado hacia el almacenamiento de lípidos.
El primer objetivo, la obtención de energía se realiza a través de reacciones bioquímicas, que
de forma conjunta reciben el nombre de CATABOLISMO.
La formación
de compuestos
orgánicos complejos
(segundo objetivo)
recibe el nombre de
ANABOLISMO, se
trata de un proceso que
consume energía.
La ENERGÍA,
es el nexo de unión
entre las dos partes del
metabolismo. Las
células heterótrofas
han rentabilizado la
forma de obtención de
energía, mediante la formación de ATP. Su escisión permite la liberación de una gran cantidad de
energía.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
5
Los organismos animales pueden considerarse un sistema termodinámico abierto, es decir,
que intercambian materia y energía con el medio. La materia la obtienen a través del alimento y la
transforma en energía y compuestos macromoleculares.
Si el alimento no está disponible, tenemos la capacidad de almacenar energía, esta capacidad
se logra a través de la especialización metabólica de los tejidos y órganos.
Cualquier célula presenta mecanismos reguladores para controlar las diferentes rutas
metabólicas, es imprescindible la interrelación metabólica entre los diferentes tejidos y órganos. En
los mamíferos, los tejidos y órganos establecen interrelaciones mediante la síntesis de mediadores
químicos denominados HORMONAS, que actúan sobre las enzimas reguladoras o las reacciones que
intervienen en el control de los niveles de sustrato de una determinada vía metabólica. Un ejemplo de
interrelación metabólica mediada por hormonas lo constituye el ejercicio físico dinámico y de
duración prolongada (carrera de larga duración).
B.- LA ENERGÍA COMO ENLACE ENTRE LAS RUTAS METABÓLICAS
Las oxidaciones biológicas:
Las reacciones químicas son en esencia transformaciones energéticas en las cuales la energía
almacenada en los enlaces químicos se transfiere a otros enlaces recién formados. En estas
transferencias los electrones pasan de un nivel energético a otro.
En muchas reacciones los electrones se transfieren de un átomo o molécula a otro. Estas
reacciones, muy importantes en los sistemas vivientes se conocen como reacciones de oxidación –
reducción (rédox).
La pérdida de uno o más electrones se conoce como OXIDACIÓN y se dice que el átomo o la
molécula que los ha perdido se ha oxidado. La REDUCCIÓN, es la ganancia de uno o más
electrones.
Al compuesto que se oxida se lo denomina AGENTE REDUCTOR, y al que se reduce,
AGENTE OXIDANTE.
La oxidación y la reducción siempre ocurren simultáneamente, porque el electrón que pierde
el átomo oxidado es aceptado por otro átomo, que se reduce en el proceso.
Frecuentemente los electrones que se transfieren en las reacciones rédox de los seres vivos
van acompañados de un protón (es decir, forman un átomo de hidrógeno), por lo que podemos
considerar como una oxidación la pérdida de átomos de hidrógeno y como una reducción la ganancia
de átomos de hidrógeno.
Los compuestos orgánicos tienen un mayor contenido energético cuanto más reducidos estén,
de lo que se puede deducir que las reacciones biológicas de oxidación liberan energía.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
6
Características que presentan las reacciones metabólicas:
Actúan secuencialmente, originándose rutas metabólicas constituidas por varias de
ellas. En estas rutas el producto final de una ruta constituye la molécula de partida de
la siguiente ruta.
Existen rutas convergentes, divergentes, cíclicas, ramificadas…
Algunas rutas son reversibles, si existe reversibilidad se habla de rutas
ANFIBÓLICAS.
Todas las rutas metabólicas son CATALIZADAS, necesitan la presencia de
ENZIMAS.
Existen dos rutas metabólicas generales:
CATABOLISMO: fase degradadora.
ANABOLISMO: biosíntesis.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
7
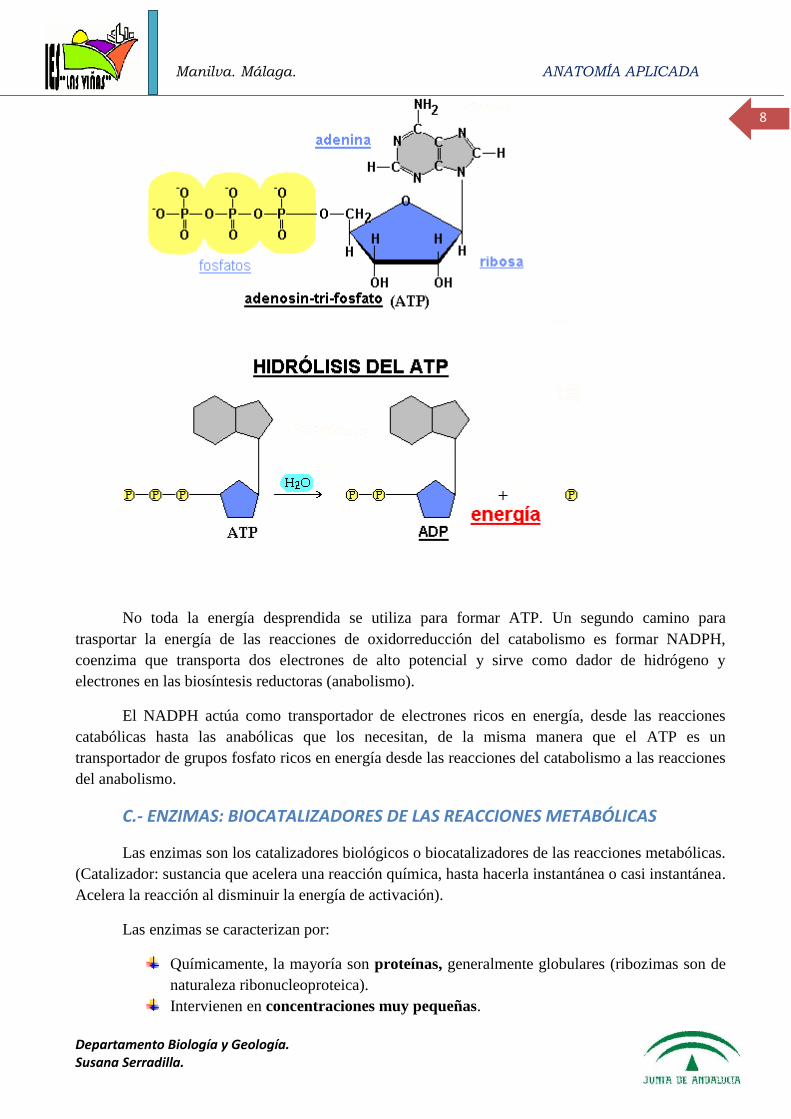
Papel del ATP como transportador de energía
La estrategia básica del metabolismo es formar ATP, NADPH y precursores
macromoleculares. La energía química de las sustancias alimenticias está en las diversas uniones
covalentes entre los átomos de una molécula. Dentro de la célula viva, esta gran cantidad de energía
no puede ser desprendida de golpe, pues la célula no podría utilizarla de una manera eficaz. Por eso,
la célula degrada poco a poco, de manera gradual y controlada mediante la intervención de enzimas.
Las reacciones catabólicas provocan la oxidación de los sustratos, por deshidrogenación, y las
enzimas que catalizan estas reacciones son deshidrogenasas ligadas a las coenzimas NAD, NADP y
FAD, principalmente. Los electrones desprendidos en estas reacciones de oxidación son captados por
otras moléculas transportadoras de electrones que se encuentran organizadas de tal manera que la
oxidación de un transportador libera más energía de la necesaria para reducir al siguiente. Si el
excedente de energía es suficiente se utiliza para fosforilar el ADP y formar ATP.
La formación de ATP proviene de la energía química aportada por la rotura de los enlaces de
las biomoléculas (carbohidratos, triacilglicéridos y aminoácidos). Las biomoléculas se encuentran
formadas por carbono, hidrógeno y oxígeno (las proteínas además contienen nitrógeno) y uno de los
productos finales del metabolismo es el AGUA. Ello implica que, de alguna manera, durante el
metabolismo se “eliminan” los hidrógenos de los combustibles y se “recogen” en la molécula de
agua, mediante reacciones rédox, en las que el agente oxidante es el oxígeno molecular.
Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
8
No toda la energía desprendida se utiliza para formar ATP. Un segundo camino para
trasportar la energía de las reacciones de oxidorreducción del catabolismo es formar NADPH,
coenzima que transporta dos electrones de alto potencial y sirve como dador de hidrógeno y
electrones en las biosíntesis reductoras (anabolismo).
El NADPH actúa como transportador de electrones ricos en energía, desde las reacciones
catabólicas hasta las anabólicas que los necesitan, de la misma manera que el ATP es un
transportador de grupos fosfato ricos en energía desde las reacciones del catabolismo a las reacciones
del anabolismo.
C.- ENZIMAS: BIOCATALIZADORES DE LAS REACCIONES METABÓLICAS
Las enzimas son los catalizadores biológicos o biocatalizadores de las reacciones metabólicas.
(Catalizador: sustancia que acelera una reacción química, hasta hacerla instantánea o casi instantánea.
Acelera la reacción al disminuir la energía de activación).
Las enzimas se caracterizan por:
Químicamente, la mayoría son proteínas, generalmente globulares (ribozimas son de
naturaleza ribonucleoproteica).
Intervienen en concentraciones muy pequeñas.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
9
No sufren modificaciones en el proceso de reacción.
Se recuperan indefinidamente. Al finalizar la reacción quedan libres sin alterarse,
como cualquier otro catalizador y pueden funcionar otras veces.
No llevan a cabo reacciones que sean energéticamente desfavorables, no modifican el
sentido de los equilibrios químicos, sólo aceleran su consecución.
Elevada especificidad de sustrato y acción. Cada reacción está catalizada por una
enzima específica. Las enzimas ejercen su acción biológica uniéndose selectivamente
a sustratos (moléculas sobre la cual la enzima ejerce su acción catalítica). Cada enzima
posee en su superficie una zona activa, como una especie de hendidura u oquedad
denominada CENTRO CATALÍTICO, a la cual se adapta perfectamente la molécula
de sustrato con la geometría complementaria a la conformación espacial de centro
activo.
Una reacción bioquímica enzimáticamente catalizada transcurre siempre mediante la
unión del sustrato a la enzima, formándose el complejo enzima – sustrato
imprescindible para que la reacción química pueda llevarse a cabo. Cualquier reacción
enzimática puede representarse mediante la ecuación:
Dada la naturaleza proteica de las enzimas, han de ser sintetizadas por el propio
organismo, como el resto de las proteínas lo que implica su codificación genética.
Cada enzima presenta su temperatura y su pH óptimo.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
10
Algunas enzimas no son proteínas exclusivamente, sino que están asociadas con otro tipo de
moléculas que tienen naturaleza no proteica y de las cuales depende su actividad. Estas asociaciones
o enzimas conjugadas se denominan holoenzimas, las moléculas con las que se asocian, cofactores y
la parte proteica de la enzima, apoenzima. Los cofactores tienen diversa naturaleza, y pueden ser:
- Cationes metálicos, como Zn2+
, Ca2+
, Fe 2+
o Mg2+
, que se unen a la apoenzima o regulan
su activación.
- Moléculas orgánicas complejas. Se denominan coenzimas cuando se unen débilmente a la
apoenzima (NAD+, FAD, NADP
+).

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
11
D.- MOLÉCULAS QUE INTERVIENEN EN EL METABOLISMO
Una ruta o vía metabólica es un proceso formado por una cadena de reacciones enzimáticas
sucesivas. Cada una de las sustancias que intervienen en una ruta metabólica, y sufre
transformaciones durante el proceso recibe el nombre de metabolito. Son las moléculas que ingresan
en las diferentes rutas del metabolismo, ya sea para su degradación (catabolismo) o para participar en
la síntesis de otras moléculas más complejas (anabolismo). La glucosa y los ácidos grasos son
ejemplos de metabolitos de degradación que actúan como verdaderos nutrientes de la célula viva. Los
aminoácidos son metabolitos necesarios para la síntesis de moléculas complejas.
En las rutas metabólicas además de los metabolitos y las enzimas específicas de cada ruta
distinguimos otras tres moléculas indispensables:
Nucleótidos: son las moléculas como NAD+, FAD, NADP
+ y el FMN
- que posibilitan
la oxidación o la reducción de los metabolitos según en qué ruta se encuentren. Desde
el punto de vista funcional suelen ser coenzimas asociadas a la parte proteica de las
enzimas que intervienen en el metabolismo, y actúan como transportadores de
electrones.
Moléculas con enlaces ricos en energía. Generalmente, los enlaces energéticos están
vinculados al grupo fosfato. Al formarse, se almacena en energía química; al romperse
se libera la misma cantidad de energía. De este modo, acoplándose a determinadas
reacciones del metabolismo, moléculas como el GTP, el ATP y la coenzima A, sirven
de almacén y actúan como proveedores de energía según sean las necesidades.
Moléculas extremas ambientales: se encuentran al comienzo o al final de un proceso
metabólico. Proceden del ambiente celular o son cedidas a él. Son moléculas sencillas
como el oxígeno, el agua y el dióxido de carbono, o más complejas, como el alcohol
etílico o el ácido láctico. Con su ingreso o salida de la célula, garantizan que el sistema
sea abierto y que, sin perder su equilibrio de flujos, siga realizando trabajo.
E.- CATABOLISMO
Fase degradativa del metabolismo, en la que a partir de sustratos reducidos se obtiene energía
y productos oxidados. Su principal objetivo es obtener energía para que las células, los tejidos y el
organismo desarrollen trabajo: mecánico, eléctrico, osmótico y químico.
Los nutrientes son degradados a elementos más sencillos a través de la acción del aparato
digestivo.
El primer factor limitante del metabolismo lo constituye la función digestiva.
El metabolismo comienza en el aparato digestivo donde se produce la transformación de las
biomoléculas complejas que se ingieren con el alimento a moléculas sencillas. Éstas pasan al hígado
(central metabólica del organismo).

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
12
Con cada uno de los elementos esenciales (monosacáridos, triacilglicéridos y aminoácidos), el
hígado puede realizar las siguientes acciones:
Liberarlos a la sangre.
Almacenarlos. El hígado tiene la capacidad de almacenar la glucosa en forma de
glucógeno.
Metabolizarlo para su funcionamiento o para funciones generales para el
organismo. Una de las funciones más importantes del hígado es realizar el
metabolismo de los aminoácidos. El hígado interviene en la síntesis de proteínas
del plasma, en la degradación, liberando nitrógeno en forma de urea.
Los ácidos grasos, la glucosa y los aminoácidos liberados por el hígado, son utilizados por las
células de forma muy rentable, ya que todas conducen a un proceso común que ocurre en las
mitocondrias de las células. La naturaleza ha logrado “agrupar” el catabolismo de moléculas distintas
en una sola molécula: ACETILCOENZIMA A (Acetil-CoA), compuesto se incorpora a varias rutas
metabólicas:
Ciclo del ácido cítrico o ciclo de Krebs. Libera dióxido de carbono y pares de
hidrógeno
Cadena respiratoria
Fosforilación oxidativa.
Como resultado de estas reacciones acopladas se genera ATP y agua.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
13

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
14
F.- ANABOLISMO
El anabolismo constituye la fase constructiva o biosintética del metabolismo. Tiene por objeto
formar compuestos macromoleculares. Se produce la biosíntesis enzimática de los componentes
moleculares de las células tales como los ácidos nucleicos, las proteínas, los polisacáridos y los
lípidos a partir de sus precursores sencillos.
La biosíntesis de las moléculas orgánicas a partir de éstos, precisa el consumo de energía
química aportada por el ATP y generado durante el catabolismo.
También se desarrolla en tres fases, pero en sentido divergente, es decir, desde sustratos
parcialmente oxidados comunes hasta productos diferentes altamente reducidos (aminoácidos,
glucosa y ácidos grasos). La adicción de hidrógenos durante el anabolismo consume energía.
No todas las rutas anabólicas se realizan por la inversión simple de las reacciones catabólicas,
sino por mecanismos complejos que salvan determinadas reacciones energéticamente inviables.
El catabolismo y el anabolismo se desarrollan simultáneamente y de modo concurrente en las
células, pero son reguladas independientemente para que el control del metabolismo sea más
eficiente.
3.- CATABOLISMO DE GLÚCIDOS
Los procesos clave del catabolismo de los glúcidos son las siguientes:
Glucólisis: ruta metabólica en la que, a partir de la glucosa y sin que se requiera
oxígeno, se forma piruvato y ATP, aunque esta última molécula con bajo rendimiento.
Ocurre en el citoplasma celular.
Respiración celular: Proceso que ocurre tras la glucolisis. Se produce la oxidación
completa de los átomos de carbono mediante el ciclo de Krebs. Los electrones
procedentes de estas oxidaciones son transferidos a las cadenas de transporte
electrónico. La síntesis de ATP ocurre en la membrana mitocondrial interna gracias a
la acción de las ATPasas.
Fermentación: Proceso que ocurre después de la glucólisis, presenta una fase de
reducción para generar NAD+. Esta fase origina varios productos finales según el tipo
de fermentación. Es una oxidación incompleta que tiene lugar en el citoplasma celular.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
15
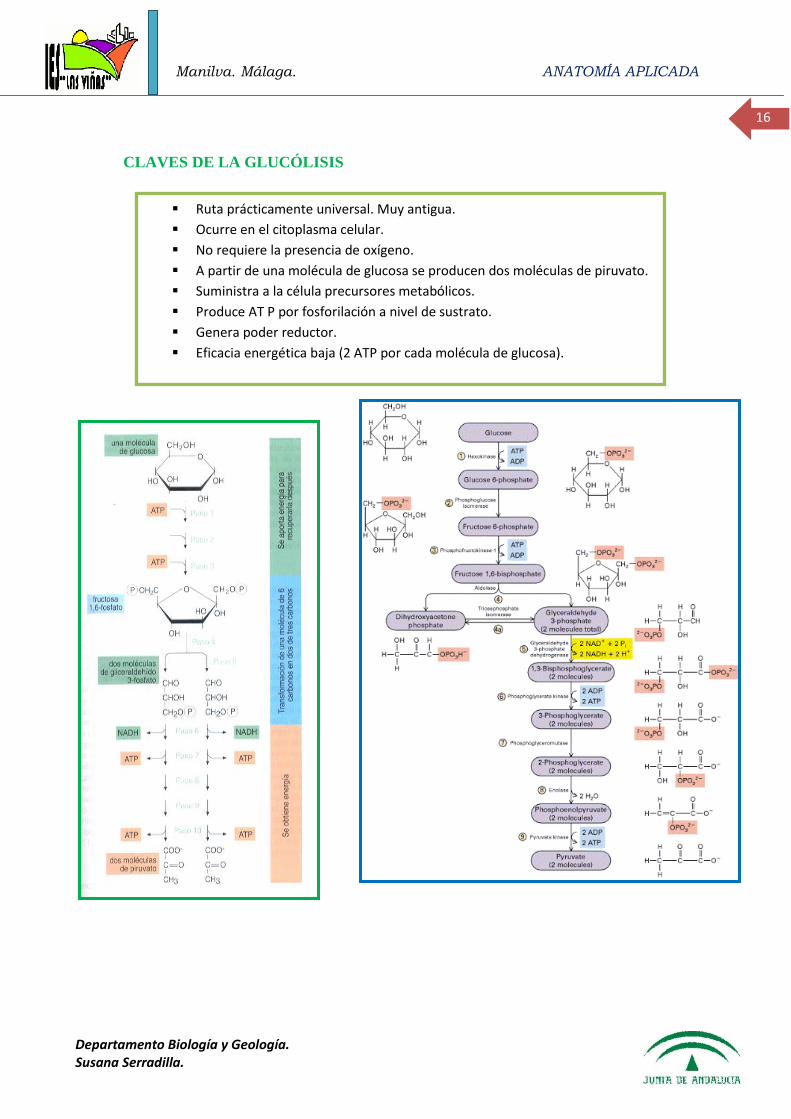
A.- GLUCÓLISIS
Se cree que el conjunto de reacciones químicas que constituyen la glucólisis constituyen una
de las rutas metabólicas más antiguas. Se cree que pudo darse en una ambiente anaerobio como la
atmósfera primitiva y servir como sistema biológico de obtención de energía celular a partir de la
glucosa. Apoyan la idea de su antigüedad el hecho de que prácticamente todos los seres vivos la
realizan.
La síntesis de ATP tiene lugar exclusivamente mediante fosforilaciones a nivel de sustrato.
La glucólisis produce dos moléculas de piruvato (es más correcto hablar de piruvato que de
ácido pirúvico) por cada molécula de glucosa de partida. El proceso se realiza en tres etapas:
1. Etapa de fosforilación que requiere aporte energético. Para la escisión del esqueleto
carbonado es preciso activar la molécula de glucosa mediante fosforilaciones, para ello es
necesario la hidrólisis de moléculas de ATP de la reserva celular. Se producen dos
fosforilaciones que proporcionan la energía necesaria para el desarrollo de las siguientes
etapas.
2. Etapa de oxidación que rinde energía y poder reductor: Tiene lugar la oxidación del grupo
aldehído a grupo carboxilo. Esta etapa requiere la incorporación de fosfato inorgánico en una
reacción catalizada por una enzima que requiere NAD+ como coenzima. La energía liberada
en esta reacción se “almacena” en el enlace fosfato rico en energía de la molécula 1,3
difosfoglicerato. Se obtiene poder reductor en forma de NADH. En la última reacción de
esta etapa se produce la primera síntesis de ATP.
3. Etapa en la que se restituye a la célula el ATP consumido en la primera fase. El 3
fosfoglicerato se transforma en piruvato y se libera un grupo fosfato de cada una de las
moléculas. Los dos fosfatos se emplean para producir dos moléculas de AT mediante
fosforilaciones a nivel de sustrato. La energía “almacenada” en estos enlaces fosfato y
utilizada inicialmente para activar las hexosas, se devuelva la reserva energética de la célula.
La eficacia de la glucólisis como ruta energética es muy baja ya que se obtiene un rendimiento
neto de dos moléculas de ATP por cada molécula de glucosa. También se obtienen dos moléculas de
NADH, que originarían más ATP, en el caso de que se siga posteriormente un proceso de respiración.
Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
16
CLAVES DE LA GLUCÓLISIS
Ruta prácticamente universal. Muy antigua.
Ocurre en el citoplasma celular.
No requiere la presencia de oxígeno.
A partir de una molécula de glucosa se producen dos moléculas de piruvato.
Suministra a la célula precursores metabólicos.
Produce AT P por fosforilación a nivel de sustrato.
Genera poder reductor.
Eficacia energética baja (2 ATP por cada molécula de glucosa).

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
17
Las etapas de la respiración aerobia son:
1. Formación de acetil CoA
2. Ciclo de Krebs o de los ácidos tricarboxílicos o
ciclo del ácido cítrico.
3. Fosforilación oxidativa:
a. Transporte electrónico.
b. Formación del gradiente quimiosmótico
c. Síntesis de ATP
B.- RESPIRACIÓN AEROBIA
Los organismos eucariotas y gran parte de los procariotas realizan la respiración aerobia en la
cual los electrones que se obtienen de la glucosa son cedidos al oxígeno. En este proceso participan
varias reacciones encadenadas, de forma que los electrones de la glucosa se transfieren a ciertas
coenzimas (NAD+.
FAD). Estas, posteriormente, cederán los electrones a unos transportadores
electrónicos. Su destino final es el oxígeno.
La respiración es el conjunto de procesos catabólicos que tienen lugar tras la glucólisis. La
respiración aerobia, en el catabolismo de los glúcidos, es la oxidación completa del producto de la
glucólisis (piruvato) hasta CO2. Para ello el piruvato debe ingresar en la mitocondria y transformarse
en acetil CoA mediante una reacción de descarboxilación oxidativa.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
18
1.- Formación de Acetil CoA
Para que la molécula de piruvato generada durante la glucólisis continúe su oxidación
incorporándose al ciclo de los ácidos tricarboxílicos (CAT) o ciclo de Krebs, debe sufrir una reacción
de descarboxilación oxidativa y convertirse en un resto acetilo en forma de acetil CoA.
Todas las biomoléculas que sirven de combustible a la célula (glúcidos, ácidos, algunos
aminoácidos) tienen que convertir sus esqueletos carbonados en grupos acetilo en forma de acetil
CoA, para poder incorporarse al ciclo de Krebs y ser oxidados hasta CO2 y H2O.
El piruvato debe ser conducido desde el citoplasma hasta la mitocondria y atravesar las dos
membranas mitocondriales, para ello debe unirse a transportadores específicos. En el interior de la
mitocondria se produce la descarboxilación oxidativa, reacción catalizada por el complejo
multienzimático denominado piruvato – deshidrogenasa. En este proceso se origina una molécula de
NADH.
2.- Ciclo de Krebs
Conjunto cíclico de reacciones
que producen la oxidación completa del
acetil CoA hasta CO2.
Consta de ocho pasos.
Los electrones cedidos en esta
oxidación son captados por las
coenzimas NAD+ y FAD, liberándose
las correspondientes moléculas
reducidas NADH y FADH2.
Se lleva a cabo en la matriz
mitocondrial y desempeña las
siguientes funciones:

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
19
Obtención de poder reductor: 3 moléculas NADH y una de FADH2, que permiten la
formación de moléculas de ATP durante el proceso de fosforilación oxidativa.
Obtención de precursores metabólicos.
Obtención de energía en forma de GTP (1 molécula) por fosforilación a nivel de
sustrato (convertible en ATP)
Dos moléculas de CO2, que corresponden a los carbonos de una molécula de acetil
CoA completamente oxidados.
El ciclo de Krebs es el núcleo del metabolismo intermediario. Algunos compuestos
procedentes de otras rutas catabólicas se incorporan a este ciclo para su degradación. Por otra parte,
algunas moléculas del ciclo de Krebs sirven como punto de partida para las rutas biosintéticas. Es,
por tanto, una vía anfibólica, resultando clave tanto para procesos catabólicos como anabólicos.
3.- Fosforilación oxidativa
Es el mecanismo de síntesis de ATP en la respiración. Tiene lugar en la mitocondria, en la
membrana interna. La enzima que cataliza la síntesis de ATP es la ATPasa o ATP sintetasa.
La síntesis de ATP se realiza por la unión de un grupo fosfato al ADP mediante un enlace de
alta energía. Esta reacción es endergónica, requiere un aporte energético importante ara producirse,
el cual es suministrado por el transporte de los electrones liberados en las oxidaciones que han
ocurrido previamente y la formación de un gradiente de protones en la membrana mitocondrial
interna.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
20
La cadena transportadora de electrones que interviene en la respiración mitocondrial está
formada por un conjunto de moléculas, asociadas a la membrana interna de la mitocondria, capaces
de reducirse y oxidarse, es decir, aceptar electrones de una molécula anterior y cedérselos a la
siguiente molécula. Para que el proceso tenga lugar de forma espontánea las moléculas
transportadoras tienen que estar situadas según un gradiente de potenciales de óxido-reducción, de
manera que al pasar de una molécula de la cadena a la siguiente, los electrones “descienden” a
niveles energéticos inferiores y liberan energía que se emplea para establecer un gradiente
quimiosmótico ente los dos lados de la membrana mitocondrial interna.
Los transportadores de electrones de la cadena respiratoria están organizados en cuatro
grandes complejos enzimáticos.
El oxígeno actúa, por consiguiente, como aceptor final de electrones, ya que, en último
término, recoge todos los electrones que se han liberado en las diferentes etapas de la oxidación de la
glucosa, o de otras moléculas donadoras de electrones.
La energía que los electrones van perdiendo al pasar por estas moléculas transportadoras se
emplea en bombear protones (H+) a través de la membrana mitocondrial interna, que se acumulan en
el espacio intermembranal que originan un potencial eléctrico de membrana.
De esta forma se forma entre las dos caras de la membrana interna se produce una diferencia
de concentración de protones y una separación de cargas eléctricas; esta situación de gradiente
electroquímico de protones es un almacenamiento temporal de energía denominada fuerza protón-
motriz que constituye el motor energético de la fosforilación del ADP en la síntesis de ATP.
La respiración aerobia es un proceso
extremadamente eficaz desde el punto de vista de
rendimiento energético
1 NADH (oxidación) ------------ 3 ATP
1 FADH2 (oxidación) -----------2 ATP

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
21
C.- FERMENTACIÓN
El metabolismo fermentativo es un proceso de oxidación incompleta de los compuestos
orgánicos, ya que no se libera toda la energía química que contienen. Las reacciones de oxidación se
producen en ausencia de oxígeno.
La síntesis de ATP tiene lugar exclusivamente por fosforilación a nivel de sustrato, es decir,
no intervienen mecanismos quimiosmóticos.
Los sustratos de fermentación son generalmente glúcidos, en particular, glucosa.
Las fermentaciones de los glúcidos comienzan con la glucólisis. Sin embargo, la vía
glucolítica, se interrumpiría en poco tiempo, ya que el NAD+ utilizando no se recupera al no existir
cadena respiratoria. La solución es la regeneración del NAD+ acoplando la oxidación del NADH a la
reducción del piruvato formado en la glucólisis. Por tanto, tienen lugar dos etapas:
Etapa de oxidación de la glucosa hasta piruvato. Se consume NAD+ y se produce
NADH.
Etapa de reducción del piruvato para dar los productos finales. Se regenera el NAD+
La reducción del piruvato da lugar a diversos productos finales. Si se origina lactato tiene
lugar la fermentación láctica y si se produce etanol y CO2, se trata de la fermentación alcohólica.
El rendimiento energético de las fermentaciones es muy bajo si se compara con el
rendimiento de un metabolismo respiratorio que actúe sobre los mismos sustratos.
Al tratarse de una oxidación incompleta del sustrato, los productos finales son moléculas
orgánicas que todavía conservan un contenido energético considerable.
En el metabolismo fermentativo de la glucosa hay una producción neta de dos moléculas de
ATP por molécula de glucosa.
La mayoría de las fermentaciones son realizadas por bacterias, sin embargo, algunas células
eucariotas también son capaces de llevar a cabo fermentaciones en condiciones de anaerobiosis.
Cuando a las mitocondrias de las células no llega el oxígeno requerido, el ácido pirúvico se
reduce a ácido láctico. Esta alternativa energética es muy utilizada por el tejido muscular.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
22
Sin embargo, tiene un serio inconveniente: provoca un aumento de la concentración de
protones pues el ácido láctico es un ácido fuerte. Por tanto, aunque el músculo es un tejido eficaz en
la amortiguación de los ácidos, un recurso es eliminarlo a la sangre, aunque ello signifique
“trasladar” el problema de la acidosis a todo el organismo.
4.- CATABOLISMO DE LÌPIDOS
Los lípidos, también llamados triacilglicéridos son moléculas muy adecuadas como
combustibles de reserva, pues su catabolismo libera mucha energía, son insolubles en agua (por lo
que se agregan en gotas lipídicas en el citoplasma) y, como tienen poca reactividad química, no
provocan reacciones indeseadas en la célula, aunque se presenten en ella en grandes cantidades.
La insolubilidad en agua lleva consigo la necesidad de ser emulsionados por enzimas del
intestino y para ser transportados por la sangre deben unirse a determinadas proteínas. No obstante,
estas mismas características hacen que su degradación no sea fácil.
Cuando la célula necesita un aporte energético mayor al habitual o no dispone de glúcidos,
degrada las grasas.
Como un triacilglicérido está formado por una molécula de glicerina y 3 moléculas de un
mismo ácido graso, el proceso de degradación tiene tres pasos:
1. Liberación de la glicerina: las lipasas de las células adiposas efectúan este paso. La glicerina,
compuesta por tres carbonos, se transforma en un intermediario de la glucólisis.
2. Entrada de los ácidos grasos al interior de la mitocondria.
3. Oxidación de los ácidos grasos, β oxidación.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
23
El paso de los ácidos grasos desde el citoplasma a la matriz mitocondrial se realiza por medio de
enzimas presentes en las dos membranas mitocondriales que actúan como transportadores. Para
atravesar las dos membranas mitocondriales, las cadenas de ácidos grasos deben unirse a una
molécula transportadora, la CARNITINA. Una vez en el interior de la mitocondria, se unen a la CoA,
antes de iniciar los ciclos de oxidación. La molécula de carnitina sale al citoplasma para ser
reutilizada.
La β oxidación consiste en la oxidación del carbono β, consiguiéndose la ruptura del enlace que
une este carbono con el α, así se van desprendiendo unidades de dos átomos a partir del extremo
carboxilo.
La degradación de los ácidos grasos comienza con su activación, lograda por la unión a la CoA
mediante un enlace éster para formar acil CoA, que requiere la hidrólisis del ATP.
El acil-CoA entrará en el ciclo de Krebs, el acil CoA con dos átomos menos de carbono
continuará el ciclo.
La degradación de los ácidos grasos suministra a la célula:
Energía todas las moléculas de ATP formadas como consecuencia de:
Incorporación al ciclo de Krebs del acetil CoA que se producen en los
sucesivos ciclos de β oxidación
La entrada a la cadena respiratoria de los electrones de un FADH2 y un
NADH por cada ciclo de rotura.
Un precursor metabólico: acetil CoA
Poder reductor: las coenzimas reducidas FADH2 y NADH

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
24 Las células pueden obtener ácidos grasos de tres
formas distintas:
A partir de grasas procedentes de los
alimentos.
De las grasas almacenadas en células
denominadas adipocitos.
En el hígado, donde el exceso de glúcidos se
convierte en grasas que posteriormente se
exportan a otros tejidos.
5.- CATABOLISMO DE PROTEÍNAS Las proteínas constituyen una parte importante de la dieta de los animales como fuente
nitrogenada y de aminoácidos esenciales. Las proteínas desempeñan una gran variedad de funciones
en los seres vivos pero no suelen ser buenos “carburantes” metabólicos. En las dietas que no aportan
glúcidos o grasas (“dietas de ayuno”) se emplean los aminoácidos como fuentes de carbono y
energía.
Ante una situación de ayuno total y teniendo en cuenta que el cerebro sólo puede consumir
glucosa, se produce la degradación de las proteínas del propio individuo para obtener glucosa a partir
de los aminoácidos.
El catabolismo de las proteínas comienza por la hidrólisis de los enlaces peptídicos que
liberan los aminoácidos. Esta hidrólisis se realiza mediante la acción de enzimas proteolíticas.
Plantas: no existe ingestión de proteínas, aunque si existen reservas de proteínas en
semillas y algunos frutos. En ellos si aparecen enzimas proteolíticas como la papaína en la
papaya, bromelaína en la piña o la ficina en la higuera. En las plantas fotosintéticas
raramente oxidan aminoácidos. Las concentraciones de los aminoácidos de los tejidos
vegetales están reguladas con la finalidad exclusiva de cubrir las necesidades para la
biosíntesis de proteínas, ácidos nucleicos y unas cuantas moléculas necesarias para el
crecimiento. En las plantas existe un catabolismo de aminoácidos relacionado
generalmente con la producción de metabolitos para otras rutas metabólicas.
Animales. La parte de energía metabólica generada a partir de los aminoácidos varía según
el tipo de organismo y la situación metabólica en la que se encuentran. Los herbívoros solo
pueden obtener una pequeña parte de sus necesidades energéticas a partir de esta fuente.
Los carnívoros, inmediatamente después de una comida, pueden obtener hasta un 90% de
las necesidades metabólicas mediante la oxidación de los aminoácidos. En los animales el
proceso de degradación de las proteínas tiene lugar en varios puntos del aparato digestivo
en los que aparecen proteasas y peptidasas que hidrolizan las cadenas polipetídicas
presentes en los alimentos.
La mayor producción de energía
presenta un inconveniente: la
necesidad de consumir más oxígeno.
La relación entre la energía producida
y el oxígeno consumido es
ligeramente peor que la
correspondiente a la oxidación de la
glucosa.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
25
En los animales, los aminoácidos se pueden degradar oxidativamente en tres situaciones
metabólicas diferentes:
1. Durante la síntesis y degradación normales de proteínas celulares (recambio proteico)
algunos de los aminoácidos liberados durante la degradación de las proteínas se
degradan oxidativamente si no se necesitan para la síntesis de nuevas proteínas.
2. Cuando una dieta es rica en proteínas ingiriéndose aminoácidos en exceso de las
necesidades corporales se puede catabolizar el excedente. Los aminoácidos no se
pueden almacenar
3. Durante la inanición o la diabetes mellitus, en las que no hay glúcidos o éstos no son
utilizables adecuadamente, se recurre a las proteínas corporales como combustible.
En estas condiciones diferentes los aminoácidos pierden sus grupos amino y los α cetoácidos
así formados pueden oxidarse a CO2 y H2O. Los esqueletos carbonados de los aminoácidos
proporcionan las unidades de tres y cuatro carbonos que se pueden convertir en glucosa la cual, a su
vez, puede proporcionar la energía para el funcionamiento del cerebro, músculo y otros tejidos.
Las rutas degradativas de los aminoácidos son muy semejantes en la mayoría de los
organismos. Al igual que ocurre con las rutas catalíticas de los azúcares y los ácidos grasos, los
procesos de degradación de aminoácidos convergen en las rutas catabólicas centrales del metabolismo
carbonado. Los esqueletos carbonados de los aminoácidos generalmente van a parar al ciclo del ácido
cítrico y de allí se oxidan para producir energía química o se canalizan hacia la gluconeogénesis.
No obstante, hay un factor importante que distingue la degradación de los aminoácidos de los
procesos catabólicos descritos con anterioridad, cada aminoácido contienen un grupo amino. Por tanto,
todas las rutas degradativas pasan a través de una etapa clave en la que se separa el grupo α amino del
esqueleto carbonado desviándolo hacia rutas especializadas para el metabolismo del grupo amino.
Las proteínas no son buenos “carburantes” metabólicos pero en las “dietas de ayuno”
pueden usarse como fuente de carbono y energía. El catabolismo de las proteínas comienza con
la hidrólisis de los enlaces peptídicos que requieren enzimas proteóliticas que liberen los
aminoácidos. Los aminoácidos no pueden almacenarse, o se utilizan o se eliminan. El
catabolismo de los aminoácidos ocurre en dos etapas: la eliminación del grupo amino y la
oxidación de la cadena carbonada.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
26
1.- ELIMINACIÓN DEL GRUO AMINO
Los aminoácidos presentes en las proteínas de la dieta son la fuente de la mayor parte de los
grupos amino. La mayoría de los aminoácidos se metabolizan en el hígado. Parte del amoníaco
generado se recicla y se utiliza en diversos procesos biosintéticos; el exceso se excreta directamente o
se convierte en ácido úrico o urea para su excreción según el organismo. El exceso de amoníaco
generado en otros tejidos se transporta al hígado para su conversión en la forma de excreción
adecuada según el organismo.
RECUERDA: los peces de agua dulce y los invertebrados son capaces de excretar directamente al
medio acuático el amoníaco producido en el catabolismo de las proteínas y se denominan
amoniotélicos. Las aves, los insectos y los reptiles excretan el nitrógeno amínico en forma de ácido
úrico y se les denomina uricotélicos. La mayoría de los animales terrestres (mamíferos, anfibios
adultos y peces de agua salada) eliminamos el nitrógeno amínico en forma de urea y somos
denominados ureotélicos.
Existen dos vías posibles para llevar a cabo el proceso de eliminación del grupo amino:
la transaminación y la desaminación oxidativa.
La transaminación consiste en la transferencia del grupo amino desde el aminoácido hasta una
molécula aceptora de los grupos amino que suele ser α cetoglutarato, el cual se transforma en
glutamato. Al ceder el grupo amino, la cadena carbonada del aminoácido se oxida quedando
convertida en un α cetoácido.
Los aminoácidos constituyen una fuente esencial de nitrógeno para las células animales. Por
ello, en el catabolismo de estas moléculas del grupo amino se reutiliza con frecuencia en la síntesis
de nuevos aminoácidos.
El glutamato y la glutamina constituyen la reserva de los grupos amino que la célula emplea
para la síntesis de nuevos aminoácidos. Las reacciones de transaminación están catalizadas por
enzimas denominadas transaminasas que llevan vitamina B6 como coenzima, y tiene lugar
principalmente en el hígado.
La desaminación oxidativa da lugar la eliminación del grupo amino del glutamato en forma
de amoniaco o ión amonio. Esta reacción se lleva a cabo en el hígado y en los riñones y está
catalizada por la enzima glutamato – deshidrogenasa.
La producción catabólica de amoníaco plantea un problema bioquímico serio, el amoníaco es
tóxico para las células, ya que aumenta el pH intracelular.
En los organismos ureotélicos, el amoníaco de las mitocondrias de los hepatocitos se
convierte en urea mediante el ciclo de la urea. En él se incorporan dos grupos NH4+ y una molécula
de CO2, formándose una de urea por cada vuelta del ciclo. El ciclo de la urea requiere energía.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
27
La producción de urea tiene lugar casi exclusivamente en el hígado y representa el destino de
la mayor parte del amoníaco allí canalizado.
La urea generada pasa a la sangre para su eliminación a través de los riñones.
El ser humano, a pesar de ser ureotélico, también produce ácido úrico, pero no procede del
metabolismo de las proteínas, del catabolismo de los aminoácidos, sino del metabolismo de las bases
nitrogenadas púricas.
2.- OXIDACIÓN DE LA CADENA CARBONADA.
La molécula α cetoácido formada tras la eliminación del grupo amino sigue unos procesos de
transformación que la incorporan a otras rutas metabólicas, tanto catabólicas como anabólicas. Según
la ruta seguida, los aminoácidos se clasifican en dos grupos: aminoácidos glucogénicos (la cadena
carbonada sufre oxidaciones que originan piruvato a algunos intermediarios del ciclo de Krebs, que
ueden permitir sintetizar glucosa a través de la gluconeogénesis) y aminoácidos cetogénicos (dan
lugar a la formación de acetil CoA que puede incorporarse al ciclo de Krebs para producir ATP o
desviarse hacia otras rutas para sintetizar por ejemplo ácidos grasos). Algunos aminoácidos
aromáticos pueden ser tanto cetogénicos como glucogénicos simultáneamente.
6.- CATABOLISMO DE ÁCIDOS NUCLEICOS
Los ácidos nucleicos, ADN y ARN son hidrolizados por enzimas nucleasas, liberándose los
nucleótidos que los constituyen. Estos nucleótidos se escinden, a su vez en sus componentes: pentosa,
fosfato y la base nitrogenada, que son reutilizados para la síntesis de nuevos nucleótidos. Si esto no
es así, estas moléculas se catabolizan y los productos finales se eliminan.
La degradación de las bases nitrogenadas pirimidínicas (citosina, uracilo y timina) origina
CO2 y NH3 (en algunos animales puede formar urea o ácido úrico).
Las bases púricas (adenina y guanina) se catabolizan y forman ácido úrico, alantoína o ácido
alantoico, según el animal de que se trate.
La enfermedad conocida como “gota” es una deficiencia del metabolismo de las purinas, por
la cual se acumula ácido úrico en la sangre y en los tejidos. Como consecuencia, se forman cristales
de urato sódico que se depositan en el riñón y las articulaciones.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
28
7.- ANABOLISMO DE LAS PRINCIALES BIOMOLÉCULAS
Cualquier organismo vivo sintetiza macromoléculas y estructura supramoleculares y también
tiene sistemas de coordinación y regulación de sus actividades. Para conseguir este equilibrio
requiere un aporte de energía del exterior y materiales para construir nuevas estructuras o reparar las
que se van desgastando.
El anabolismo es la parte del metabolismo encargada de la síntesis de moléculas complejas a
partir de precursores más sencillos. Las reacciones biosintéticas originan polisacáridos, ácidos
nucleicos, proteínas y otras moléculas complejas imprescindibles para el funcionamiento del
organismo. Las rutas anabólicas son endergónicas y reductoras, es decir, requieren ATP y poder
reductor en forma de NADPH.
Los seres humanos, al igual que el resto de los animales somos quimioheterótrofos, es decir,
nuestra fuente de carbono es a partir de moléculas orgánicas, como fuente de energía empleamos la
oxidación de moléculas orgánicas y los compuestos orgánicos también son los donadores de
electrones. Sintetizamos moléculas orgánicas a partir de moléculas también orgánicas.
BIOSÍNTESIS DE POLISACÁRIDOS: todos los seres vivos sintetizan
polisacáridos a partir de monosacáridos. Los animales producimos glucógeno.
Cuando una célula necesita glucosa y no dispone de ella puede realizar su síntesis a
partir de otros intermediarios metabólicos. Este proceso se denomina gluconeogénesis
y constituye una ruta metabólica que es inversa a la glucólisis. Los metabolitos
intermediarios son iguales a los que se producen durante la glucólisis aunque
intervienen enzimas diferentes, lo que permite regular ambos procesos de forma
independiente. Se trata de un proceso energéticamente desfavorable ya que consume
energía, 6ATP por cada molécula de glucosa que sintetiza. El proceso tiene lugar a
lo largo de varias reacciones químicas que ocurren tanto en la mitocondria como en el
citoplasma.
BIOSÍNTESIS DE LÍPIDOS: la capacidad para biosintentizar lípidos es esencial
para los organismos por la gran cantidad de funciones variadas que presentan:
energéticas, estructurales, hormonales…
La biosíntesis de ácidos grasos tiene lugar en el citoplasma porque es donde se
encuentra el complejo enzimático ácido-graso sintetasa que lleva a cabo el
proceso. Esta ruta biosintética queda físicamente separada de la catalítica que ocurre
en la mitocondria.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
29
RECUERDA: Sólo las plantas y algunas bacterias sintetizan todos los
aminoácidos proteicos. Los aminoácidos que no podemos sintetizar se
denominan aminoácidos esenciales y deben ser ingeridos con la dieta.
Para iniciar la biosíntesis de una cadena de ácido graso, una molécula de acetil CoA
actúa de cebador. Otra molécula de acetil CoA sufre una carboxilación y origina una
molécula de tres carbonos, malonil CoA. Esta molécula se condensa con el acetil CoA
cebador, provocándose una descarboxilación que origina una cadena de un ácido
carboxílico de cuatro carbonos, unido a la CoA. Las demás unidades de dos carbonos
se van añadiendo a la cadena en forma de malonil CoA.
Los triglicéridos o grasas se forman por esterifricación (las reacciones de
esterificación se realizan entre un ácido orgánico y un alcohol, y originan un éster y
agua) de tres moléculas de ácidos grasos con una de glicerina. Para que se produzca
la esterificación, ambos componentes tienen que ser activados. Los ácidos grasos se
activan uniéndose a la coenzima A y la glicerina a un grupo fosfato.
Los ácidos grasos se sintetizan en el citoplasma de las células hepáticas y en los
adipocitos, en los que se puede almacenar una gran cantidad de grasas como material
de reserva. Las células del músculo cardíaco pueden almacenar asimismo, cierta
reserva de grasas, ya que parte de la energía que obtienen procede de la
oxidación de los ácidos grasos en la mitocondria.
BIOSÍNTESIS DE COMPUESTOS NITROGENADOS: el nitrógeno es un
elemento esencial para los seres vivos y se encuentra fundamentalmente como
componente de aminoácidos y nucleótidos. Salvo las bacterias fijadoras de nitrógeno
el resto de los seres vivos necesita utilizar compuestos nitrogenados para
incorporarlos a sus biomoléculas.
La biosíntesis de los aminoácidos tiene lugar en el citoplasma celular y en
ella intervienen dos procesos fundamentales: la síntesis del esqueleto carbonado
a partir de precursores metabólicos y la incorporación del grupo amino
mediante reacciones de transaminación.
Los aminoácidos suelen agruparse en familias según el precursor del que parte
la biosíntesis del esqueleto carbonado.
Ninguno de los precursores metabólicos contiene nitrógeno y dado que los
aminoácidos son compuestos nitrogenados, su biosíntesis requiere una fuente de este
elemento, los organismos heterótrofos sólo pueden sintetizar aminoácidos a partir de
los aminoácidos adquiridos con la dieta, por medio de transaminaciones catalizadas
por transaminasas.
La unión de aminoácidos mediante enlaces peptídicos sigue una secuencia
concreta determinada por el material genético y constituye la síntesis de proteínas.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
30
RECUERDA: las purinas y las pirimidinas son las bases nitrogenadas de los
nucleótidos. Están presentes en los ácidos nucleicos, las coenzimas como NAD+ y
FAD, los nucleótidos trifosfato como ATP y UTP y algunas vitaminas.
Las purinas y las pirimidinas que constituyen los nucleótidos pueden obtenerse
mediante dos vías:
- a partir de precursores metabólicos, síntesis de novo.
- vías de recuperación que reciclan las bases liberadas en el catabolismo de los
ácidos nucleicos.
AMPLIACIÓN: Las células cancerosas debido a su rápido crecimiento tienen
unos requerimientos de nucleótidos mayores que las células normales. Muchos de los
medicamentos empleados en la quimioterapia anticancerosa actúan inhibiendo la
síntesis de nucleótidos.
8.- CARACTERÍSTICAS METABÓLICAS DE LOS TEJIDOS Y ÓRGANOS
HÍGADO
Presenta un papel central en el procesamiento y distribución en el metabolismo y proporciona
a todos los demás órganos y tejidos una mezcla de nutrientes adecuados a través del torrente
sanguíneo.
Los hepatocitos transforman los nutrientes obtenidos de la dieta en los combustibles y en los
precursores necesarios para cada tejido y los exportan a la sangre. Los tipos y cantidades de
nutrientes que alcanzan el hígado dependen de diversos factores entre los que destacan el tipo de
dieta y el intervalo entre comidas. El hígado presenta una flexibilidad metabólica increíble para
adaptarse a las circunstancias variables que pueden ocurrir en un organismo. Por ejemplo, en una
dieta rica en proteínas, los hepatocitos contienen niveles elevados de enzimas para el catabolismo de
los aminoácidos y la gluconeogénesis. Al cambiar a una dieta rica en glúcidos, los niveles de estas
enzimas caen en pocas horas y empiezan a sintetizarse las enzimas esenciales ara el metabolismo de
los glúcidos.
TEJIDO ADIPOSO
El tejido adiposo, compuesto por adipocitos, se encuentra ampliamente distribuido por el
organismo (bajo la piel, alrededor vasos sanguíneos, cavidad abdominal…). Suele representar el 15%
de la masa de un individuo adulto.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
31
Los adipocitos son metabólicamente muy activos y responden rápidamente a estímulos
hormonales en conexión metabólica con el hígado, el músculo esquelético y el corazón.
Los adipocitos presentan un metabolismo glucolítico activo, utilizan el ciclo del ácido cítrico
para oxidar piruvato y ácidos grasos y realizan la fosforilación oxidativa mitocondrial.
Durante los periodos de elevada ingesta de glúcidos, el tejido adiposo puede convertir glucosa
en ácidos grasos (formación de piruvato y acetil CoA) y a partir de ellos formar triacilgliceroles que
se almacenan como grandes glóbulos de grasa.
En el ser humano, la mayor parte de la síntesis de ácidos grasos tiene lugar en los hepatocitos
y no en los adipocitos. Los adipocitos almacenan triacilgliceroles procedentes del hígado y del tubo
intestinal, especialmente después de comidas ricas en grasas.
Cuando se precisan, los triacilgliceroles almacenados en el tejido adiposo son hidrolizados por
las lipasas en el interior de los adipocitos y liberan ácidos grasos, que pueden ser transferidos a los
músculos esqueléticos y al corazón a través del torrente sanguíneo.
La adrenalina acelera la liberación de los triacilgliceroles de los adipocitos al estimular la
conversión de las enzimas lipasas a su forma activa. La insulina contrarresta el efecto de la adrenalina
al disminuir la actividad de la lipasa.
MÚSCULO ESQUELÉTICO
El músculo esquelético consume más del 50% del total le oxígeno en un ser humano en
condiciones de reposo y hasta un 90% durante el ejercicio muscular muy intenso. El metabolismo del
músculo esquelético está en buena parte especializado en producir ATP como fuente inmediata de
energía. El músculo esquelético está especializado además en satisfacer las necesidades energéticas
de forma inminente según la demanda de energía. El músculo esquelético debe realizar a veces un
gran trabajo en un periodo de tiempo corto (carrera de 100 metros lisos), en ocasiones se requiere un
esfuerzo prolongado (maratón, durante el parto…).
Los músculos esqueléticos pueden utilizar como combustible:
- Cuerpos cetónicos.
- Glucosa
- Ácidos grasos
El uso de uno u otro dependerá del
grado de actividad muscular que se requiera.
En el músculo en reposo los princiales
combustibles son los ácidos grasos y los
cuerpos cetónicos que vienen del hígado.
Estos compuestos se oxidan y se degradan ara

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
32
dar acetil CoA que ent5ra en el ciclo de Krebs para ser oxidado hasta CO2. La transferencia de
electrones aloxñigeno que reswulta de este proceso roorciona energía para la síntesis de ATP por
fosforilación oxidativa.
Los músculos moderadamente activos utilizan glucosa además de ácidos grasos y cuerpos
cetónicos. La glucosa se fosforila, a través de la glucólisis se degrada hasta piruvato y éste se
convierte en acetil CoA que se oxida a tráves del cilco de Krebs.
En los músculos más activos la demanda de ATP es tan grande que el flujo sanguíneo no
puede
suministrar el oxígeno y los combustibles lo suficientemente deprisa ara producir el ATP necesario
tan sólo a travñes de la resiración aeróbica. En estas circunstancias, el glucógeno alamacenado en el
músculo se degrada hastra lactato or fermentación, proporcionando dos moléculas de ATP por cada
unidad de glucosa degradada. La frmentación del ácido láctico proporciona así, de forma rápida, un
suplemento de energía bajo fomra de ATP, que refuerza la producción basal de ATP que resulta de la
oxidación aeróbica de otros combustibles or la viá del ácido cítrico o ciclo de Krebs.
El músculo esquelético no contiene glucosa 6 fosfatasa y no puede convertir la glucosa 6
fosfato en glucosa libre transortable a otros tejidos. El glucógeno muscular está dedicado enteramente
al suministro de energía en el músculo a través de la degradación glucolítica.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
33
Tras una intensa actividad muscular, la respiración se mantiene profunda por un cierto tiempo.
La mayor parte del oxígeno que se obtiene de esta forma se utiliza para la producción de ATP por
fosforilación oxidativa en el hígado. El AT se utiliza en el hígado
para la gluconeogénesis a partir de lactato, que, a partir de los
músculos, es transportado en la sangre hasta el hígado. La glucosa
así formada vuelve a los músculos para restablecer su contenido de
glucógeno, completando el ciclo de Cori.
Los músculos esqueléticos presentan una cantidad
considerable de FOSFOCREATINA, que puede regenerar
rápidamente ATP a partir de ADP mediante una reacción
catalizada por la creatina quinasa. Durante la contracción activa y
la glucólisis esta reacción está orientada hacia la síntesis de ATP,
pero durante la recuperación del ejercicio físico el mismo enzima
se utiliza para sintetizar de nuevo fosfocreatina a partir de creatina
y ATP.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
34
SISTEMAS PARA LA PRODUCCIÓN
DE ENERGÍA ANAERÓBICO
ALÁCTICO
ANAERÓBICO
LÁCTICO
AERÓBICO
METABOLISMO Reservas
Fosfágenas
Glucólisis Oxidación de
Principios
Inmediatos
NECESIDAD DE
OXÍGENO
No precisa
oxígeno
No precisa
oxígeno
Necesita
oxígeno
DÓNDE SE
PRODUCE
Sarcoplasma Sarcoplasma Mitocondria
NIVEL DE
POTENCIA QUE
DESARROLLA
Muy Alto Alto Medio
NIVEL DE
CAPACIDAD QUE
DESARROLLA
Hasta 20 seg Entre 20” y 180 “ + 3-5 minutos
INTENSIDAD DE
ESFUERZO QUE
PERMITE
Muy Alta Alta Medio-Alta
DURACIÓN DEL
ESFUERZO QUE
PERMITE
Breve y Corta Media Prolongada
RENDIMIENTO
ENERGÉTICO
Rendimiento
Energético Muy
Alto
Rendimiento
Energético Alto
Rendimiento
Energético
Medio – Alto
FATIGA O
AGOTAMIENTO
DEL SISTEMA DE
PRODUCCIÓN DE
ENERGÍA
El sistema pierde
eficacia en poco
tiempo debido al
rápido
agotamiento de
las reservas de
fosfágeno
existentes.
El sistema pierde
eficacia en un
tiempo
relativamente
corto debido a
una acumulación
elevada de
lactato.
El sistema es
muy eficaz
porque permite
la realización de
esfuerzos muy
prolongados
con niveles de
fatiga
pequeños.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
35
MÚSCULO CARDÍACO
A diferencia del músculo esquelético presenta una actividad continua, debiendo mantener un
ritmo regular de contracción y relajación. El corazón tiene un metabolismo aeróbico de forma
permanente. Las mitocondrias son mucho más abundantes en el músculo cardíaco que en el
esquelético, llegando a representar casi la mitad del volumen de la célula.
El corazón utiliza como combustible una mezcla de glucosa, ácidos grasos libres y cuerpos
cetónicos que llegan de la sangre. Estos combustibles se oxidan a través del ciclo del ácido cítrico
para proporcionar la energía necesaria para generar el ATP por fosforilación oxidativa.
El músculo cardíaco no almacena lípidos ni glucógeno en grandes cantidades. Se almacenan
pequeñas cantidades de reservas energéticas bajo forma de fosfocreatina. Puesto que el corazón es
normalmente aeróbico y obtiene su energía mediante fosforilación oxidativa, la dificultad del oxígeno
para alcanzar una zona del músculo cardíaco cuando los vasos sanguíneos se bloquean
(aterosclerosis, trombosis coronaria…) puede provocar la muerte del tejido cardíaco (infarto de
miocardio).
CEREBRO
El metabolismo del cerebro presenta varios aspectos característicos:
El cerebro de los mamíferos adultos utiliza normalmente tan sólo glucosa como
combustible.
El cerebro presenta un metabolismo respiratorio muy activo. Utiliza por lo menos un
20% del total de oxígeno consumido por un organismo adulto en reposo. La tasa de
utilización de oxígeno por el cerebro es bastante constante y no se modifica de forma
significativa ni durante el sueño ni durante la actividad mental.
El cerebro contiene poco glucógeno, por lo que depende de forma permanente de la glucosa
que recibe vía sanguínea. Si la glucosa de la sangre disminuye drásticamente, aunque sea en un
periodo muy corto de tiempo, por debajo de un umbral crítico puede causar efectos irreversibles en
la función cerebral.
Aunque el cerebro no puede utilizar directamente los ácidos grasos libres o los lípidos de la
sangre como combustible celular, puede, si es necesario, utilizar un cuerpo cetónico (β
hidroxibutirato) producido en los hepatocitos a partir de ácidos grasos. La capacidad del cerebro de
oxidar el β hidroxibutirato por la vía del acetil CoA es de esencial importancia en periodos de ayuno
prolongado o inanición ya que permite utilizar la grasa del organismo como fuente de energía y
preservar las proteínas musculares. Representa el último recurso para obtener glucosa, a través de la
gluconeogénesis durante la inanición grave.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
36
9.- REGULACIÓN DEL METABOLISMO
Para entender el significado de las rutas metabólicas y su regulación, debemos considerar
estas rutas en el contexto del organismo en su conjunto. Una característica esencial de los organismos
pluricelulares es la diferenciación celular y la distribución de funciones. Además de las rutas
centrales del metabolismo, que proporcionan energía y que tienen lugar en todas las células, los
tejidos y órganos en los organismos complejos como los seres humanos presentan funciones
especializadas y tienen, por tanto, requerimientos energéticos y unos patrones metabólicos
característicos.
Las señales hormonales integran y coordinan las actividades metabólicas de los distintos
tejidos de manera que regulan la distribución de precursores y sustratos energéticos en cada órgano.
El hígado es el órgano que centraliza la distribución y el procesamiento de los nutrientes. Los
azúcares y aminoácidos formados en la digestión atraviesan el epitelio intestinal y entran en la
sangre, a través de la cual llegan al hígado. Algunos triglicéridos derivados de lípidos de la dieta
también son procesados en el hígado, donde sus ácidos grasos constituyentes se utilizan para toda una
serie de procesos.
La glucosa 6 fosfato es el intermediario clave en el metabolismo de los glúcidos. Puede
polimerizar para dar glucógeno, ser desfosforilada para proporcionar glucosa a la sangre o convertirla
en ácidos grasos a través del acetil CoA. Puede ser degradada por la glucólisis y el ciclo del ácido
cítrico para proporcionar energía en forma de ATP o por la vía de las pentosa fosfato para
proporcionar pentosas y NADPH. Los aminoácidos se utilizan para sintetizar proteínas hepáticas y
plasmáticas, o bien sus esqueletos carbonados pueden ser convertidos en glucosa y glucógeno, a
través de la gluconeogénesis, el amoníaco formado por su desaminación se convierte en urea. Los
ácidos grasos pueden convertirse en el hígado en otros triacilgliceroles, colesterol o lipoproteínas
plasmáticas que se transportan y almacenan en el tejido adiposo. Pueden igualmente ser oxidados
para dar ATP y para formar cuerpos cetónicos que se utilizarán en otros tejidos.
El músculo esquelético está especializado en producir ATP para el trabajo mecánico. Durante
un esfuerzo muscular el glucógeno es el combustible fundamental proporcionado ATP por
fermentación hasta lactato. Durante la recuperación el lactato se reconvierte a glucógeno y a glucosa
en el hígado, a través de la gluconeogénesis. La fosfocreatina es una fuente inmediata de ATP
durante la contracción activa. El músculo cardíaco obtiene todo su ATP a partir de la fosforilación
oxidativa. El cerebro utiliza sólo glucosa y β hidroxibutirato como fuentes de energía, el segundo
resulta importante durante el ayuno y la inanición.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
37
El cerebro utiliza la mayor parte de su energía en forma de ATP para el transporte activo de
NA+ y K+ y el mantenimiento del potencial eléctrico de las membranas neuronales. La sangre pone
en comunicación todos los órganos, transportando nutrientes, productos de desecho y señales
hormonales entre ellos.
Las hormonas son mensajeros químicos segregados a la sangre por determinados tejidos para
regular la actividad de otros tejidos. Actúan según una jerarquía de funciones. Impulsos nerviosos
estimulan el hipotálamo ara enviar hormonas a la hipófisis, estimulando o inhibiendo la liberación de
hormonas trópicas. La hipófisis anterior, a su vez, estimula otras glándulas endocrinas (tiroides,
suprarrenales, páncreas) para segregar sus hormonas características que, a su vez estimulan tejidos
diana específicos.
La concentración de glucosa en sangre está controlada hormonalmente. Las fluctuaciones en
la glucosa de la sangre debidas a la ingestión de alimentos o a ejercicio intenso, se compensan por
una serie de cambios en el metabolismo de varios órganos desencadenados por hormonas.
La adrenalina prepara el organismo para un aumento de actividad, movilizando la glucosa de
la sangre a partir de glucógeno y otros precursores. Unos niveles de glucosa en sangre reducidos
provocan la liberación de glucagón, que estimula la liberación de glucosa a partir de glucógeno
hepático y desvía el metabolismo energético en el hígado y el músculo hacia la utilización de ácidos
grasos, ahorrando la glucosa ara su utilización por el cerebro.
Durante el ayuno prolongado los triacilgliceroles se convierten en la principal fuente
energética; el hígado transforma los ácidos grasos en cuerpos cetónicos que son utilizados en otros
tejidos, incluyendo el cerebro.
Unos niveles elevados de glucosa en sangre estimulan la liberación de insulina, que acelera la
captación de glucosa por los tejidos y favorece el almacenamiento de compuestos de reserva
energética como el glucógeno y los triacilgliceroles. En la diabetes no tratada puede ser que no se
produzca insulina o que no se reconozca por los tejidos, de forma que se hace difícil la utilización de
la glucosa. Cuando los niveles de glucosa en sangre son altos, la glucosa se excreta tal cual en la
orina. Los tejidos pasan a depender entonces de los ácidos grasos como combustible, dando lugar a
cuerpos cetónicos y degradan proteínas celulares para fabricar glucosa a partir de sus aminoácidos
gluconeogénicos. La diabetes no tratada se caracteriza por los niveles altos de glucosa en la sangre y
en la orina y por la producción y excreción de cuerpos cetónicos.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
38
10.- METABOLISMO EN REPOSO Y EJERCICIO
El ejercicio físico es una situación en la que la actividad metabólica se puede multiplicar más
de diez veces, con independencia de las características del ejercicio es fundamental la regulación
hormonal de las diferentes rutas y su interrelación. A consecuencia de la mayor actividad metabólica,
se produce una mayor cantidad de calor, que se pierde a través del sudor, y por tanto también es
importante el mantenimiento del equilibrio hidroelectrolítico del organismo y el control renal del
agua y los electrolitos.
El conocimiento del metabolismo durante el ejercicio ha sido y es uno de los objetivos
fundamentales de la fisiología. Una idea intuitiva consiste en relacionar la duración y la intensidad
del ejercicio con la utilización de las diferentes alternativas de las rutas metabólicas y su regulación.
Desde un punto de vista pedagógico pueden dividirse los esfuerzos en:
- Esfuerzos submáximos de duración prolongada.
- Esfuerzos máximos de duración media.
- Esfuerzos supramáximos de breve duración.
Habitualmente se considera como el máximo esfuerzo aquel que requiere la máxima cantidad
de oxígeno que el organismo es capaz de consumir.
El tipo de ejercicio que se suele rescribir como un objetivo de salud o terapéutico suele ser el
ejercicio realizado de forma continua con variaciones de la intensidad. En el campo del rendimiento,
se realiza un tipo de esfuerzo con periodos de descanso absoluto o relativo. Es el esfuerzo
intermitente.
Todos y cada uno de los tejidos y órganos contribuyen al metabolismo global, sin embargo,
no todos poseen la misma actividad metabólica: el miocardio, el cerebro o el hígado presentan un
metabolismo en reposo muy activo, mientras que el músculo esquelético en reposo presenta una
actividad metabólica muy baja pero en actividad multiplica su metabolismo unas 20 veces. Durante el
ejercicio el miocardio y el cerebro intentan mantener o incluso aumentar su actividad metabólica. Por
tanto, el metabolismo global depende de las necesidades articulares en un momento determinado.
Para medir la actividad metabólica del organismo durante un determinado tiempo existen dos
posibilidades:
- Medir el calor desprendido. Calorimetría directa.
- Medir la cantidad de oxígeno consumido. Calorimetría indirecta.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
39
Los procedimientos de calorimetría directa son técnicamente complejos por lo que lo más
sencillo es medir el oxígeno consumido, que puede ser una buena estimación del metabolismo global.
El consumo de oxígeno en reposo es de 300 mL/min
Debemos tener en cuenta que el organismo puede “quemar” diversos combustibles y por
tanto, el metabolismo global pueda diferir en función de qué sustrato o sustratos esté utilizando. El
valor del consumo de oxígeno de forma aislada aporta una buena información pero es incompleta.
Metabolismo basal
Se define el metabolismo basal como el gasto mínimo necesario para mantener las funciones
vitales cuando se establecen unas determinadas condiciones denominadas baales. Estas condiciones
son:
- Reposo psicofísico absoluto
- Temperatura ambiente de 22 a 25ºC
- Estado de ayuno de corta duración (<12horas)
La dificultad de asegurar estas condiciones hace difícil la determinación del metabolismo
basal. El metabolismo basal se expresa en referencia a la superficie corporal. Para el varón el
metabolismo basal es de 40kcal/m2/h y para la mujer 36kcal/m
2/h.
Desde el punto de vista conceptual, el metabolismo basal es muy importante, por ejemplo,
para calcular la ingesta mínima en los programas de pérdida de peso. Parece lógico que una dieta no
debe ser inferior al valor del metabolismo basal. El metabolismo basal depende de los siguientes
factores:
- Edad: va incrementándose desde el nacimiento (30 kcal/m2/h) hasta los 6 años cuando
alcanza su máximo valor (55 kcal/m2/h). A partir de esa edad desciende de forma
progresiva, salvo durante la adolescencia, periodo en que alcanza un valor de meseta.
- Sexo: las mujeres poseen un valor que es un 10% menor al de los varones.
- Temperatura: el incremento de la temperatura ambiente por encima de 30º aumenta el
metabolismo basal y el descenso por debajo de los 15º lo disminuye.
- Altitud sobre el nivel del mar: la altura influye cuando la presión parcial de oxígeno
desciende a un valor de la mitad respecto al valor correspondiente al nivel del mar. Cuando
la presión parcial de oxígeno es de 75mm de Hg, el metabolismo basal disminuye.
- Factores fisiológicos y patológicos: a consecuencia de realizar ejercico físico de forma
regular y continua se produce un reajuste de la actividad metabólica del organismo,
traducido en un aumento del metabolismo basal. Los individuos entrenados pueden tener
un metabolismo basal más elevado, en comparación con los sedentarios.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
40
Los descensos patológicos del metabolismo basal se dan en enfermedades como la
hipofunción tiroidea (descensos del 25 al 40%), hiponutrición y anorexia nerviosa. Los
ascensos se producen en la hiperfunción tiroidea, enfermedades infecciosas y procesos
febriles.
Cualquier actividad cotidiana entraña un coste adicional o gasto adicional del metabolismo
basal. Este gasto adicional se denomina metabolismo energético.
Estados nutricionales
- Estado alimentario o posprandrial: según el tipo de alimento ingerido, aproximadamente
después de 3 o 4 horas, el hígado recibe todas las moléculas simples que han sido absorbidas a través
del intestino. Se produce un incremento de la glucosa sanguínea, que en el hígado determina un
incremento de la fosforilación de la glucosa que, a su vez condiciona a) un incremento de la relación
insulina/glucagón (estimulación de la síntesis de glucógeno) b) su utilización para la síntesis de
ácidos grasos y su liberación al tejido adiposo y c) su utilización como combustible.
Como el organismo no siempre dispone de alimentos para llevar a cabo sus funciones vitales
es necesario que almacene energía.
Cuantitativamente, el mayor almacenamiento de energía es en forma de grasa:
1- La liberación de energía de un gramo de grasa hipotética de seis átomos de carbono es de
45 ATP, mientras que la misma cantidad de glucosa libera 38 ATP.
- La acumulación de energía en forma de carbohidratos (glucógeno) determinaría dos
efectos negativos: elevada desproporción del peso corporal (el peso se elevaría al doble) ya
que la acumulación de glucógeno requiere hidratación y
- dificultad para mantener la presión osmótica intracelular.
2.- Aunque el almacenamiento de grasa permite el ayuno prolongado durante un mes, se
requieren otras formas de almacenamiento de energía ya que determinados tejidos son dependientes
de la glucosa de forma relativa como el tejido nervioso o funcionan prioritariamente en condiciones
anaerobias como el caso de los eritrocitos.
- Estado de ayuno de corta duración: el que habitualmente se realiza entre la última comida
de un día y la primera del día siguiente. Menos de 24 horas. Cuando no se suministran sustratos al
organismo, el hígado libera a sangre glucosa procedente de la glucogenólisis, la cual es utiliza
preferentemente por el cerebro. En esta situación, el tejido adiposo se encarga de aportar ácidos
grasos utilizados por el músculo y el riñón. La regulación se efectúa a través del ciclo glucosa/ácidos
grasos y por la acción hormonal. El descenso de la relación insulina/glucagón por disminución de la
insulina y aumento del glucagón interviene en el control de la glucemia.

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
41
- Estado de ayuno de larga duración: Aquel que se prolonga en el tiempo y oscila entre 24
horas hasta 24 días. La adaptación metabólica es extraordinaria y debe atender a dos principios: a)
mantener la glucemia lo más estable posible y b) controlar la degradación proteica.
Durante los primeros días de ayuno (del primer al tercer día) se producen los siguientes
efectos:
- Incremento de la gluconeogénesis a partir de alanina, glicerol y lactato.
- Incremento de la proteólisis, principalmente muscular.
A partir del tercer día, el tejido adiposo incrementa la liberación de ácidos grasos, los cuales,
al llegar al hígado, estimulan la producción de cuerpos cetónicos. La regulación se produce or la
acción del ciclo glucosa/ácidos grasos/cuerpos cetónicos.
A lo largo del periodo de ayuno se produce un descenso de la utilización de glucosa por el
cerebro. Al mismo tiempo aumenta el consumo de cuerpos cetónicos. Se produce además una
reducción drástica de la proteólisis muscular, lo que constituye un mecanismo de autodefensa del
organismo. Al descender la proteólisis desciende por un lado el aporte de sustratos para la
gluconeogénesis, de manera que se produce un descenso de la glucosa, y también disminuye la
producción de urea, lo que desencadena un estado de coma, debido al descenso de la glucosa y al
aumento del amonio.
- Esfuerzos submáximos de duración prolongada: es el tipo de esfuerzo que se aconseja
cuando se considera el ejercicio físico como fuente de salud o de prevención de enfermedades. Desde
el punto de vista fisiológico es la máxima expresión de rendimiento.
Este tipo de esfuerzos requiere una gran producción de energía a una velocidad relativamente
lenta.
La energía para este tipo de esfuerzos se puede obtener del propio músculo (depósitos de
energía en forma de glucógeno y ácidos grasos) y de tejidos y órganos extramusculares (tejido
adiposos e hígado)
El tejido muscular dispone de glucógeno lo que le confiere cierta capacidad metabólica
independiente, relativa porque sus depósitos son finitos. Presenta una baja actividad de la enzima
lipoproteína lipasa, lo que obliga al tejido adiposo y al hígado a mantener una elevada actividad
metabólica para suministrar combustible a la fibra muscular.
A medida que avanza la duración del esfuerzo:
Se incrementa la utilización de los ácidos grasos y disminuye la del glucógeno
muscular.
El aporte de glucosa por parte del hígado se mantiene constante a lo largo del tiempo.
La interrelación entre los tejidos impide un “vacío metabólico”

Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
42
El agotamiento del glucógeno muscular es gradual durante los esfuerzos continuos, si bien las
reservas de glucógeno en músculo no llegan a agotarse completamente. El rendimiento mecánico
baja ostensiblemente cuando la energía se obtiene sólo de las grasas. Por consiguiente, la necesidad
de coordinación entre músculo, hígado y tejido adiposo es imprescindible. La concentración de
ácidos grasos en sangre aumenta durante el ejercicio físico prolongado, se desconoce su importancia
de su combustión en el músculo y otros tejidos.
En este tipo de esfuerzos, el músculo utiliza una doble fuente de energía: la propia
(glucógeno muscular) y la ajena (glucógeno hepático y ácidos grasos del tejido adiposo)
- Esfuerzos máximos o supermáximos de corta duración: el otro extremo del movimiento
lo constituyen los esfuerzos muy breves pero de una intensidad superior a la que se podía desarrollar
de forma constante. Este tipo de esfuerzos no son exclusivos del deporte, por ejemplo al realizar un
sprint para coger un autobús. Las unidades motoras utilizadas en este tipo de esfuerzos son las fibras
de tipo II, o de contracción rápida, ya que poseen el complejo enzimático necesario para acelerar la
glucogenólisis.
Se considera al fosfágeno como la suma de las reservas de ATP y fosfocreatina. La escisión
de la fosfocreatina produce AT P a gran velocidad, mientras exista sustrato. Gracias a la existencia en
el músculo de este compuesto se pueden realizar las primeras contracciones musculares. La escisión
de la fosfocreatina permite además enlazar de forma inmediata la glucólisis, de manera que ésta
acentúa su velocidad disminuyendo la utilización de la fosfocreatina.
La concentración de lactato aumenta con la distancia recorrida, lo que demuestra la
participación creciente de la glucogenólisis.
Los esfuerzos de intensidad supramáxima se pueden realizar gracias a la disponibilidad
de depósitos de energía en el músculo (fosfágeno y glucógeno). La obtención de energía de estos
dos sustratos es simultánea, de manera que desde los primeros segundos del movimiento se
activan las dos rutas metabólicas.
- Esfuerzos intermitentes: en muchas actividades deportivas y en el entrenamiento se
alternan periodos de esfuerzo con periodos de inactividad absoluta o relativa. A este tipo de esfuerzos
se les denomina intermitentes y su comprensión es fundamental para entender los métodos de
entrenamiento.
Desde el punto de vista metabólico, el esfuerzo intermitente persigue incrementar la obtención
de energía por el músculo en un mayor grado que si el esfuerzo se realizara de forma continua. Los
combustibles utilizados dependerán de la intensidad y duración del ejercicio. Y del número de
esfuerzos realizados.
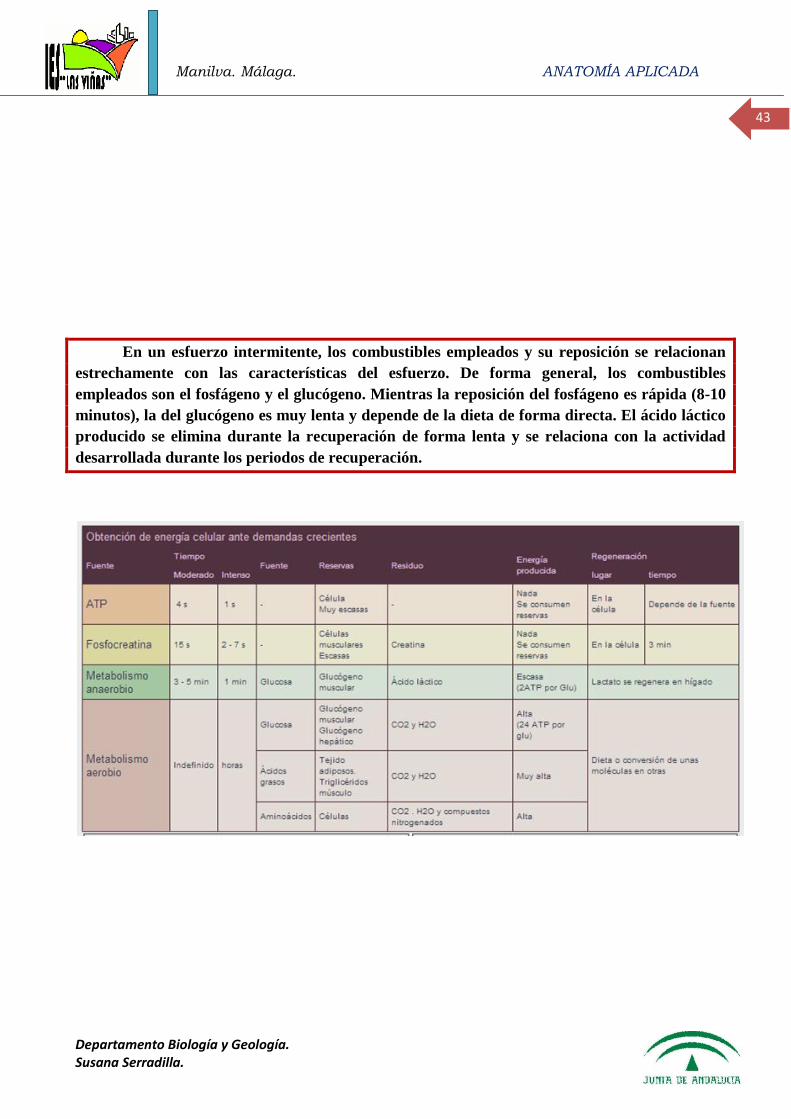
Manilva. Málaga. ANATOMÍA APLICADA
Departamento Biología y Geología. Susana Serradilla.
43
En un esfuerzo intermitente, los combustibles empleados y su reposición se relacionan
estrechamente con las características del esfuerzo. De forma general, los combustibles
empleados son el fosfágeno y el glucógeno. Mientras la reposición del fosfágeno es rápida (8-10
minutos), la del glucógeno es muy lenta y depende de la dieta de forma directa. El ácido láctico
producido se elimina durante la recuperación de forma lenta y se relaciona con la actividad
desarrollada durante los periodos de recuperación.