
Fisiología vegetal
37
Manual de Sesiones Prácticas Laboratorio de Relación Suelo-Agua-Planta Facultad de Ciencias Agronómicas Universidad de Chile
Transcript of Fisiología vegetal

Manual de Sesiones Prácticas
Laboratorio de Relación Suelo-Agua-Planta Facultad de Ciencias Agronómicas
Universidad de Chile

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 1 CONTENIDO Y ESTADO ENERGÉTICO DEL AGUA EN EL SUELO
En esta sesión analizaremos tres aspectos del agua en el suelo: el estado del agua en el suelo, métodos de medición del contenido de agua del suelo y las relaciones entre el contenido de agua del suelo y el potencial hídrico. Para profundizar el tema revisar Rawlins (1976), Hillel (1980), Klute (1986). 1. Estado del agua en el suelo Puede ser descrito en dos formas: en términos de la cantidad presente y en términos del estado energético del agua. La cantidad presente puede ser expresada en términos gravimétricos y/o volumétricos. El contenido gravimétrico es la masa de agua en una unidad de masa de suelo seco. Se registra el peso húmedo de la muestra de suelo, luego la muestra es secada a 100-110°C hasta peso constante y nuevamente pesado (Gardner, 1986). El contenido volumétrico de agua se expresa en términos de volumen de agua por volumen de suelo. Puede ser calculado a partir del contenido gravimétrico de agua que se multiplica por la densidad aparente del suelo (kg de suelo seco/volumen que ocupa). El estado energético del agua en el suelo puede ser expresado como sigue (Hillel, 1980; Hanks y Aschroft, 1980):
gspm ψ+ψ+ψψ=Ψ )( (1)
Donde Ψ es el potencial del agua en el suelo, ψm es el potencial mátrico del agua en el suelo, ψp es el potencial debido a presión (o carga de agua), ψs es el potencial de solutos y ψg es el potencial gravitacional. Las unidades de potencial hídrico en el suelo pueden ser expresadas en base masa, volumen o peso. Utilizaremos unidades de presión, en términos de pascal (newton m-2) y megapascal (106 pascal), MPa. Para una discusión de la interconversión de estas unidades, ver Hillel (1980) o Nobel (1983). 2. Medición del contenido de agua y del potencial de agua del suelo o sus componentes Se describen a continuación seis técnicas para la medición del contenido y estado energético del agua en el suelo: método gravimétrico, aspersor de neutrones y reflectometría, que miden el contenido de agua del suelo; tensiómetros y bloques de resistencia que miden el potencial mátrico del agua del suelo y finalmente los psicrómetros de suelo miden el potencial hídrico total.
2.1. Contenido gravimétrico de agua Es el método más simple de medición del agua en el suelo, representado por el símbolo (w). Este valor se determina secando la muestra de suelo a 100-110°C hasta peso constante. La determinación gravimétrica del contenido de agua está compuesta por dos mediciones independientes, el peso húmedo (Mt) y el peso seco (Ms), combinadas de la siguiente forma:
MsMww = (2)
Donde Mw = Mt-Ms 2.2. Aspersor de neutrones Los neutrones rápidos son termalizados cuando ellos chocan con un cuerpo de masa similar, tales como los núcleos de hidrógeno. La energía de los neutrones es transmitida a los protones y el “rebote o choque” neutrónico es mucho más bajo. Este principio ha sido adoptado para estimar la densidad del núcleo de hidrógeno en el suelo. En la mayoría de los suelos el hidrógeno es asociado con el agua del suelo, aunque en suelos orgánicos o densamente enraizados este no sería el caso. Los neutrones rápidos son emitidos desde una fuente (Am-Be) en un tubo instalado en el suelo; los neutrones lentos son contabilizados por un detector. El número de neutrones lentos detectados es proporcional al número de colisiones entre neutrones y núcleos de hidrógeno, los cuales reflejan el contenido de agua del suelo. (Van Bavel et al., 1954).
1 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
La técnica neutrónica ha sido utilizada en una amplia variedad de sistemas especialmente agrícolas, ecosistemas naturales y pastizales. Esta técnica es más adecuada para suelos profundos, en suelos con estructura fina donde los tubos de acceso para la sonda sean fácilmente instalados. Los tubos de acceso pueden ser de acero, aluminio o PVC y los diámetros usados generalmente son de 4 cm (1,625”) o de 5 cm (2”) que es de mayor disponibilidad. Dependiendo de las propiedades del suelo, el contenido de agua del suelo y el tipo de material del tubo utilizado (acero, cobre o PVC) utilizado, los neutrones de la sonda muestrean una esfera de suelo con un radio de 10-30 cm. Este radio hace difícil una medida adecuada del contenido de agua del suelo en la superficie, obteniéndose conteos extremadamente bajos (Hanna y Siam, 1980). Para evitar este problema, deben realizarse curvas de calibración para lecturas cercanas a la superficie o medirlas en forma diferente, por ejemplo a través de muestreos gravimétricos. En todos los casos, deben adoptarse precauciones y evitar la exposición a los neutrones rápidos ya que ellos representan riesgo probable a la salud. Los operadores de un aspersor deben manipular el instrumento cuidadosamente y medir la exposición mediante un test de neutrones, incluso se recomienda situar una fuente de agua entre la sonda de medición y el operador del sistema. Calibración: Cada tubo debe ser calibrado para cada tipo de suelo. La distribución y tamaño de las partículas de suelo, la estructura y la composición mineralógica, pueden afectar la relación entre la tasa de conteo y el contenido de agua del suelo. Por estas razones la calibración de fábrica generalmente no se utiliza. Puede ser calibrado directamente en el campo y en el laboratorio. Podrían utilizarse grandes depósitos o contenedores (por ejemplo 200 litros) que pueden ser llenados con el suelo del campo, se instala un tubo de acceso y el sistema llevado a un contenido de agua y/o potencial hídrico del suelo conocido, o bien, directamente en un sector contiguo al de mediciones estableciendo una parcela sembrada que se riega y luego se evalúa paralelamente la humedad por gravimetría y el registro del aspersor y con estos valores se establece una curva de calibración. Obtención de datos: Generalmente se toman diez conteos con la sonda a la profundidad establecida, considerando cinco que varían menos del 5%. Este conteo es usado para establecer una curva estándar. Los resultados del conteo de campo y la curva de calibración pueden incorporarse a una planilla electrónica y luego con lo conteos siguientes calcular el contenido de agua a distintas profundidades, las cantidades estacionales y la distribución del agua en el suelo. 2.3. Reflectometría Desde 1980, se ha estado desarrollando una técnica para medir el contenido de agua en el suelo, denominada reflectometría basada en la constante dieléctrica. La reflectometría es aconsejable en estudios de balance hídrico del suelo a nivel de campo y para la medición en grandes volúmenes de suelo, ya que evitaría los efectos de la heterogeneidad espacial sobre los valores medidos (Topp, 1987; Dalton, 1987). La constante dieléctrica del material de suelo varía directamente con el contenido de agua del suelo y es proporcional al tiempo requerido para el paso de un voltaje dado a través de una longitud conocida de suelo (Topp et al., 1980). La constante dieléctrica del suelo ha sido medida con cables coaxiales orientados verticalmente y horizontalmente. Topp et al., (1980) desarrollaron una relación empírica entre la constante dieléctrica y el contenido de agua del suelo. 2.4. Tensiómetros Los tensiómetros son ampliamente utilizados para medir el contenido de agua disponible del suelo cuando el Ψ es alto (Richards, 1965). Estos aparatos son simples, no caros, siendo muy prácticos en los sistemas agrícolas. Los tensiómetros miden la tensión del agua del suelo o potencial mátrico desarrollada a medida que el agua “es forzada fuera” del terminal cerámico hacia el suelo por las fuerzas mátricas. La cápsula de cerámica se conecta a un tubo lleno de agua. El vacío desarrollado se mide usando un vacuómetro, el vacío obtenido o la tensión es numéricamente igual al potencial mátrico del agua en el suelo. Los tensiómetros funcionan mejor en el rango de 0 a -0,07 MPa (Devitt et al., 1983) debido a que el aire comienza a entrar por los poros de la cápsula de cerámica en suelos más secos que -0,07 a -0,08 MPa. Los tensiómetros son insensibles al potencial osmótico del agua en el suelo y por lo tanto no proporcionan una adecuada medición del potencial hídrico en suelos con salinidad significativa. Los tensiómetros frecuentemente se utilizan en combinación con el aspersor de neutrones, bloques de resistencia o psicrómetros para cubrir el rango total de humedad del suelo. 2.5. Bloques de resistencia Los bloques de resistencia representan un tipo de aparato importante en la medición del potencial mátrico del agua en el suelo como una analogía del potencial hídrico del suelo. Su operación se basa en el hecho
2 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
de que la conductividad eléctrica de muchos materiales varía en función del contenido de agua. Los más comunes, son los bloques de yeso que contienen dos electrodos de acero inoxidable, cada uno soldado a un cable, ambos salen del bloque y se dejan en la superficie del suelo. La conductividad de los bloques aumenta a medida que la cantidad de agua del suelo absorbido por los bloques aumenta, mientras que la solución salina permanece constante (producto de solubilidad del yeso). Las mediciones se realizan conectando un microamperímetro a los cables que salen del bloque de resistencia. La resistencia medida es proporcional a la cantidad de agua en el bloque, la que depende del agua del suelo. Cada bloque debe ser calibrado para obtener una curva que relacione el potencial mátrico del suelo y la medida de resistencia. Una técnica útil es calibrar los bloques en el suelo sobre un sistema de olla a presión. En esta forma la resistencia, el contenido de agua y el potencial hídrico pueden ser determinados simultáneamente en cada muestra. Los bloques también se construyen en material sintético (fibra de vidrio) y otros materiales ya que la conductancia eléctrica de la solución del suelo es función de su composición, no trabajan bien en suelos salinos o cuando el riego se hace con aguas salinas. Los bloques de resistencia trabajan mejor en suelos más secos que -0,07 MPa, siendo complementarios en el rango de operación con los tensiómetros de suelo. Son recomendables para potenciales de suelo de -2,0 a -3,0 MPa. Debido a que la respuesta de los bloques de resistencia es lenta, no son útiles para seguir un evento rápido de desecamiento. Se pueden observar efectos significativos de histéresis entre calibraciones de humedecimiento y desecamiento. Por estas razones, los bloques de resistencia son útiles para seguir ciclos de desecamiento lentos. a) Psicrómetros de suelo El potencial hídrico total del suelo (mátrico más osmótico) puede ser determinado usando psicrómetros de suelo. Estos son psicrómetros de termocuplas encerrados en un bulbo cerámico. El bulbo permite el equilibrio entre la solución del suelo y la atmósfera al interior del bulbo. En el equilibrio, la HR en el interior del bulbo es igual a aquel de la atmósfera de la muestra. Dado que ambas fuerzas, mátrica y de soluto contribuyen al equilibrio de la humedad relativa, el aparato mide el potencial hídrico total del suelo, no considerando el gravitacional. El psicrómetro de termocuplas es efectivo para suelos húmedos a suelos secos. Los suelos húmedos están cercanos a saturación, suelos sobre -1,2 MPa no son efectivamente monitoreados. En realidad, en el rango de disponibilidad de agua para las plantas, la humedad relativa del suelo varía solo entre 0,99 y 1,00 (Rawlins y Campbell, 1986). Cuando los suelos son salinos, las sales se pueden depositar en las paredes de la cápsula a medida que el suelo pasa desde una condición húmeda a una seca, inactivando el sistema. Finalmente, debido a que la humedad relativa es afectada por temperatura, los gradientes de temperatura en el suelo o mediciones inadecuadas pueden alterar significativamente la lectura especialmente en las capas superficiales de suelo. En tales situaciones las medidas psicrométricas debieran realizarse dos veces durante el día cuando los gradientes térmicos son cercanos a 0 a una profundidad dada. La psicrometría de termocuplas se basa en que la presión de vapor de agua en una atmósfera en equilibrio con una muestra, tiene un potencial conocido definido por la ecuación 3 y que es igual al potencial hídrico de la muestra. La forma en la cual los solutos y otros componentes reducen el potencial hídrico puede ser expresado en términos de sus efectos en la actividad química del agua. La actividad, es la tendencia del agua a reaccionar o moverse en el sistema y su valor afecta directamente a la presión de vapor del agua en el sistema (Brown y Oosterhuis 1992). Por lo tanto, el potencial hídrico es descrito en términos de presión de vapor del agua en el sistema en comparación a aquel del agua libre a la misma temperatura y presión que se expresa a través de la ecuación:
⎟⎟⎠
⎞⎜⎜⎝
⎛=Ψ
s
a
ee
VRT ln (3)
donde Ψ es el potencial hídrico en MPa, R es la constante universal de gases (8,31*10-6 m3 MPa mol-1 K-1 ), T es la temperatura en grados Kelvin, V es el volumen molal del agua a la temperatura T (18,069 x 10-6 m3 mol-1), e es la presión actual de vapor (ea) y es es la presión de vapor a saturación. Un término gravitacional se incluye como componente del Ψ, el cual es despreciable dado que el aumento de este valor es aproximadamente 0,0098 MPa*m-1 (Nobel, 1983). Algunos autores recomiendan su utilización como un parámetro separado del Ψ.
3 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
REFERENCIAS BIBLIOGRÁFICAS BROWN R.W. & OOSTERHUIS D.M. (1992) Measuring Plant and Soil Water Potentials with Thermocouple Psychrometers: Some Concerns. Agronomy Journal 84:78-86. DALTON, F.N. 1987. Measurement of soil water content and electrical conductivity using time domaine reflectometry. R.W. Pearcy, J. Ehleringer, H.A. Mooney and P.W. Rundell (eds) Plant Physiological Ecology: 161-183. DEVITT, D., JURY, WA., STENBERG, P & LH STOLZY (1983). Comparison of methods used to estimate evapotranspiration for leaching control. Irrig. Sci. 4:59-69. GARDNER, WH (1986) Water content. In: Methods of soil analysis Part 1 Physical and mineralogical Methods, Agronomy Number 9 (Part 1), American Society of Agronomy Madison, pp. 493-544. HANKS, RJ & GL ASCHROFT (1980) Applied Soil Physics, Springer Verlag, Berlin, 159 pp. HANNA G.W. and SIAM N. (1980). The estimation of moisture content in the top 10 cm of soil using a neutron probe. J. Agr. Sci. 94:251-254. HILLEL D (1980) Fundamental of Soil Physics, Academic Press, New York, 413 pp. HULSMAN RB (1985) The neutron probe and the microcomputer. Soil Sc. 140:153-157. KLUTE A (1986) Methods of soil analysis. In A. Klute (ed) Part 1 Physical and Mineralogical Methods, 2nd edn, Agronomy Number 9 (Part 1). American Society of Agronomy, Madison, pp 635-652. NOBEL P (1983) Biophysical Plant Physiology and Ecology, WH Freeman, San Francisco 603 pp. RAWLINS SL (1976) Measurement of water content and the state of water in soils. In: T Kozlowski (ed) Water deficits and plant growth : 1-55. Academic Press. RAWLINS SL & GS CAMPBELL (1986) Water potential: thermocouple psychrometry. In: A. Klute (ed) Methods of Soils Analysis. Part 1 Physical and Mineralogical Methods, 2nd edn. Agronomy Number 9. American Society of Agronomy, Madison, pp 597-518. RICHARDS SJ. (1965) Soil suction with tensiometers. In: CA Black (ed) Methods of Soil Analysis. Part 1 Physical and Mineralogical Properties, Including Statistics of measurement and Sampling. American Society of Agronomy, Madison, pp 153-163. TOPP GC (1987) The application of time-domain reflectrometry (TDR) to soil water content measurement. In: Proceedings of the International Conference on Measurement of Soil and Plant Water Status, vol 1, Utha State University, pp 85-93. VAN BAVEL CHM., EC HOOD & N UNDERWOOD (1954) Vertical resolution in the neutron method for measuring soil moisture. Trans. Am. Geophys. Union 35: 595-600.
4 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 2 CONTENIDO DE AGUA EN EL SUELO
PROBLEMAS 1. Se tomó una muestra de suelo a los 15 cm de profundidad que pesó 350 g, luego fue secada en una estufa a 105°C por 24 horas y su peso fue de 280 g. Si la densidad aparente de dicho suelo es de 1,3 g cm-3 calcule:
a) El volumen de agua que tenía la muestra de suelo en el campo. b) El contenido gravimétrico de agua que tenía la muestra de suelo en el campo. c) El contenido volumétrico de agua que tenía la muestra de suelo en el campo. d) La altura de agua que tienen los primeros 30 cm de profundidad de dicho suelo.
2. Un suelo húmedo tiene un contenido de agua de 0,15 g g-1. Si se necesitan 200 g de suelo seco para un experimento ¿Cuántos gramos de suelo húmedo se necesitan? 3. Una muestra de suelo tiene un contenido de humedad de 35%. ¿Qué volumen de agua se debe añadir a una muestra de 1 kg de ese suelo para que su contenido de humedad aumente a un 50%? 4. Se debe llenar una maceta de 2,5 L con un suelo que tiene un contenido de agua de 24% y una densidad aparente de 1,2 g cm-3 ¿Cuántos gramos de dicho suelo se requerirán para llenar la maceta? Si la CC del suelo es de 30% ¿Cuántos litros de agua hay que añadir a la maceta para que quede a CC? 5. Una muestra de suelo con 40% de porosidad tiene un volumen total de 120 cm3 y un contenido de humedad de 0,3 cm3 cm-3. Si la densidad de la fase sólida es de 2,5 g cm-3. ¿Cual es la densidad aparente? 6. ¿Cuál es la carga que debe soportar un camión tolva de 5 m3 de capacidad si la arena que transporta está saturada con agua? Da= 1,5 g cm-3. 7. Un terrón natural de suelo húmedo tiene un volumen de 150 cm3, un peso húmedo de 240 g y un espacio ocupado por aire de 0,15 cm3 cm-3. Si la densidad de partículas es de 2,65 g cm-3. Calcular:
a) Densidad aparente b) Contenido de humedad en base a peso c) Contenido de humedad en base a volumen. d) Porosidad total.
8. Calcular el volumen de agua en media hectárea de un suelo que tiene un contenido de agua de un 23%, Da= 1,35 g cm-3 y una profundidad de 75 cm. 9. Un suelo tiene una porosidad de 0,4 cm3 cm-3. Si el contenido de humedad del suelo es de 0,30 g g-1 y la densidad real de 2,6 Mg m-3, calcule la altura de agua en una profundidad de 40 cm de suelo. 10. Se tiene un suelo en el que un volumen de 1000 cm3 tendría 340 cm3 de agua a CC. La porosidad es de 54%. ¿Qué volumen de agua requiere la capa arable (15 cm de profundidad) por hectárea para quedar a CC si el suelo se encuentra a PMP?
5 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 3
CONTENIDO DE AGUA EN EL SUELO PROBLEMAS
11. Con el objetivo de conocer la Da de un suelo homogéneo, franco, de 80 cm de profundidad que se encontraba saturado, se sacó una muestra con forma de cubo de 10 cm por lado, que pesó 1900 g. La muestra se llevó inmediatamente a una estufa a 105°C hasta un peso constante de 1400 g. Para conocer el contenido de agua a CC, dicho suelo se dejó drenar por 24 horas y se sacó una muestra que dio los siguientes resultados: 1570 g y 1250 g en peso húmedo y seco, respectivamente. Para determinar el PMP se colocó una muestra en un plato de presión a 15 bares que pesó 1320 g en húmedo, al secarla en estufa a 105°C pesó 1175 g. Conociendo el contenido de agua de dicho suelo a CC y PMP conteste:
a) Si el U.R. es de 30%, ¿cuántos gramos de agua habría que agregar a un macetero cilíndrico de 40 cm de alto y 30 cm de diámetro que se llena con este suelo?.
b) ¿Cuantos m3 de agua por hectárea existen en el suelo en el momento en que ha cesado el drenaje del exceso de agua?
12. un cultivo de papas tiene una profundidad de enraizamiento de 0,7 m, en un suelo homogéneo. La CC y el PMP de ese suelo es de 30% y 15% respectivamente, y la Da es 1,3 g cm-3. Si se riega con un UR de 25%:
a) ¿Cuál será la humedad aprovechable de dicho suelo? b) ¿Cuál es el contenido de agua que tiene este suelo al momento de regar? c) ¿Qué altura de agua habría que aplicar a dicho suelo para dejarlo a CC? d) ¿Qué volumen de agua habría que aplicar para dejar a CC 1 ha de dicho suelo? e) Si antes del riego cae una precipitación de 28 mm ¿qué altura de agua se debería aplicar en el
riego?
13. Se desea llevar un suelo a CC. Se sabe que el suelo alcanza CC con un valor de 0,4 cm3 cm-3 (contenido de agua en base a volumen). Si el suelo esta a PMP, ¿Qué altura de agua (cm) hay que aplicar para que el suelo quede a CC hasta una profundidad de 80 cm? Porosidad= 50%; Densidad real= 2,6 g cm-3 14. Se sabe que regando un suelo que esta a PMP con una altura de agua de 16 cm queda a CC hasta una profundidad de 80 cm. A CC el contenido de agua en base a volumen es de 0,4 cm3 cm-3. Indique:
a) Capacidad de campo del suelo. b) Punto de marchitez permanente. c) Humedad aprovechable (%). d) ¿Qué profundidad del suelo queda a CC después de regar durante 20 h, si la velocidad promedio
de infiltración es de 0,8 cm h-1 y en el momento en que se inicia el riego, el suelo tiene un contenido de agua de 10% (base a peso)?
Porosidad del suelo = 50%; Dr = 2,6 g cm-3 15. Determinar Hd y Hsat en un suelo de 60 cm de profundidad que presenta una CC de 27,7%, PMP 13%, Pw 23% y una porosidad de 50,1% 16. Un suelo presenta las siguientes características:
Profundidad CC (%) PMP (%) Da (g/cm3) E (%) Pw (%) 0-25 30 15 1,35 48 22 25-50 28 12 1,4 47 21 50-88 26 11 1,41 47 20
6 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
a) Determinar la humedad aprovechable. b) Determinar el déficit de humedad. c) ¿Hasta que profundidad moja a CC una lluvia de 30 mm? d) Determinar humedad de saturación a partir de Pw.
17. Se tienen 3 ha de tomates con una profundidad radicular de 40 cm que se esta regando con un UR de 40%. El suelo es homogéneo y esta con un contenido de agua de 35%, su Da, CC y PMP es de 1,2 g cm-3, 0,4 g g-1 y 0,2 g g-1, respectivamente.
a) Si el contenido de agua actual es de 35% ¿qué altura de agua dispone antes de realizar el riego? b) Después de regar ¿qué altura de agua dispone antes de realizar un nuevo riego? c) ¿Qué altura de agua se debe aplicar en cada riego? d) ¿Qué volumen de agua aplicará en el riego para las 3 ha de tomate?
18. Un suelo con las siguientes características:
Profundidad CC (%) PMP (%) Da (g cm-3) 0-30 30 15 1,35 30-80 28 13 1,4 80 Estrata impermeable
a) Calcular la lamina de agua que debería aplicarse para dejar un nivel freático a 40 cm desde la
superficie. b) Si llueve 45 mm antes del riego ¿Se forma o no nivel freático?. Si se forma ¿A qué profundidad
desde la superficie?. Considere un U.R. 25%. 19. Se tienen 5 ha de maravilla con una profundidad radicular de 80 cm se está regando con un UR de 30% en un suelo con las siguientes características:
Profundidad CC (%) PMP (%) Da (g cm-3) 0-25 30 12 1,41 25-45 30 12 1,42 45-80 28 11 1,35
a) Si antes del riego cae una lluvia de 25 mm ¿Hasta qué profundidad se humedece el suelo? b) Si a los 80 cm existe un hardpan impermeable ¿Qué monto de lluvia dejará un nivel freático a 25
cm de la superficie? 20. Se tomaron muestras de dos estratas de suelo con un cilindro de 10 cm de largo y 5 cm de radio, luego se les determinó lo siguiente:
Profundidad CC (%) PMP (%) Da (g cm-3) 0-30 30 15 1,35 30-80 28 13 1,40
a) Si el UR es 30%, ¿cuántos metros cúbicos de agua se deben aplicar por ha? b) Si antes del riego cae una lluvia de 45 mm, ¿Se forma o no nivel freático? Si se forma, ¿hasta
qué profundidad? c) Si se saca una muestra de 100 cc desde la primera estrata antes del riego, ¿cuánto pesaría dicha
muestra?
7 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
FORMULARIO
profDaPEHsatPCCURHaHd
URprofDPMPCCHdPMPCCHa
profH
DDVV
E
VVVVMMM
VMD
VMD
DWVV
MMW
w
w
a
rat
p
sawt
swT
s
sr
t
sa
at
w
s
w
))((*
**)(
*
)/1(
65,2
*
−=−==
−=−=
=
−==
++=+=
==
=
==
=
θ
θ
8 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 4 MOVIMIENTO DE AGUA EN EL SUELO
PROBLEMAS
1. Hay dos tensiómetros de bourdon instalados en un suelo a diferentes profundidades: A= 40cm, B= 70cm. Ambos tensiómetros marcan 80 cb ¿Cuál es el potencial del agua en los puntos A y B? ¿Hacia donde se mueve el agua? ¿Qué valor debieran indicar estos tensiómetros para asumir que el perfil esta a CC? 2. Hay 2 tensiómetros con manómetros de mercurio ubicados a diferente profundidad en el perfil del suelo A= 30cm, B= 60cm. Ambos tensiómetros marcan 18,8 cm de mercurio. ¿Cuál es el potencial total del agua en A y B?¿Hacia donde se mueve el agua? 3. Si en un suelo hay una napa freática ubicada a 2,5 m de profundidad y existe un tensiómetro de bourdon ubicado a 90 cm de profundidad que marca 15 cb. Calcule el potencial hidráulico a 3m de profundidad (A), el potencial hidráulico en el límite de la napa freática (B) y el potencial hidráulico en el punto donde esta ubicado el tensiómetro (C). ¿Hacia donde se mueve el agua ? 4. En el siguiente sistema de cañería cerrada con soluciones y agua pura separadas por membranas semipermeables calcule los potenciales hidráulicos ubicados en los puntos A, B, C, D, E y F. ¿Hacia donde se mueve? ¿Cuando cesará la entrada de agua en C y en F?
40cm
B C D 10cm Agua Pura 20cm
A
N.R
Solución ψs=-15cb
30cm
F E
60cm
5. Una cubeta está dividida por una membrana semipermeable. A un lado hay agua pura y al otro una solución con ψs = -0,24 MPa. ¿Qué potencial total del agua tiene el punto A ubicado en el agua pura a 30 cm de altura de un punto B ubicado en la solución? ¿Hacia donde se mueve el agua ?¿Qué potencial de presión debiera aplicarse a la solución para que no ocurra movimiento de agua ?
agua pura solución
A 25cm
B
30cm N.R.
6. Una planta de trigo tiene raíces hasta una profundidad de 60 cm, en un suelo donde un tensiómetro marca 30 cb (A), tiene en su quinta hoja, ubicada a 30 cm de altura, una célula del mesófilo cuya vacuola ejerce una presión de 0,2 MPa sobre la pared celular. ¿Qué potencial osmótico tiene dicha célula para que su potencial hídrico sea de -0,8 MPa (B)? ¿Hacia donde se mueve el agua? Según el resultado
9 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
obtenido ¿por qué cree usted que no se considera el ψg como un componente importante en el potencial hídrico del tejido vegetal? 7. El potencial hídrico de una célula en un matraz con agua pura es de -0,6 MPa sus componentes son: ψp = 0,5 MPa y ψs = -1,1 MPa. Si la misma célula se coloca en una matraz con solución salina -1 MPa ¿Cual será el nuevo potencial hídrico de la célula y sus componentes? 8. Hay un suelo de 60 cm de profundidad en una ladera de un cerro con una pendiente de 20%. Se instalan 2 tensiómetros a 50 cm de profundidad y a una distancia horizontal de 1 metro uno de otro. El tensiómetro A (el más alto) marca 50cb y el tensiómetro B (el más bajo) marca 150cb. ¿Cuál es el potencial total del agua en cada punto? ¿Hacia donde se mueve el agua? (Ocupe como N.R. el punto B). 9. Calcular los potenciales hidráulicos y sus componentes en los puntos A, B, C, D y E. Considere que el suelo esta saturado y no esta en equilibrio. Si la conductividad del suelo es de 0,00023 m s-1 ¿Cual será el flujo?. Si el diámetro de la cañería es de 10 cm ¿Cual será el caudal?.
NR A 6cm B
36cm
E
18cm C D
27cm
FORMULAS
sgpm ψψψψΨ +++=
gm ψψΨ +=
gp ψψΨ +=
xk
tAQJ
ΔΔΨ
−== **
)(*5,13ψm Hhh +−= (tensiometro de mercurio)
EQUIVALENCIAS: 0,1Mpa ≈ 1 bar = 100cb = 1000 cm 1 mca = 1 metro columna agua = 10 cb = 100 cm
10 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 5 CONTENIDO Y ESTADO ENERGÉTICO DEL AGUA EN LA PLANTA
CONTENIDO RELATIVO DE AGUA (CRA) El CRA es una medida apropiada del contenido de agua de la planta en términos del déficit hídrico celular. Mientras que el potencial hídrico (Ψ) es útil en el transporte de agua en el contínuo S-P-A. Todos los componentes de las relaciones hídricas foliares cambian durante el día a medida que la radiación y temperatura cambian. Generalmente se usan entre 4 a 6 muestras de un tratamiento o genotipo de hojas totalmente expandidas: 1.- En función de la hoja señalada se sacan entre 5 y 10 cm2 por muestra. Evitando las grandes nervaduras y podrían ser de 1,5 cm de diámetro con el uso de sacabocados. Es importante que el muestreo sea rápido para evitar excesiva deshidratación. 2.- En hojas compuestas más pequeñas. Varias hojas podrían representar un muestreo homogéneo y rápido. 3.- En cereales. la muestra podría ser la sección media de la hoja de 5 a 10 cm2 cortada con tijeras. En campo cada muestra debe ser puesta en depósitos sellados y si es necesario, usar depósitos plásticos con ice-pack en su interior para mantener la temperatura entre 10 y 15° sin congelar la muestra y trasladar al laboratorio. 4.- En el laboratorio se pesa para obtener el peso fresco (PF) luego la muestra es totalmente hidratada a turgor máximo durante 4 horas en agua destilada bajo luz y temperatura ambiente. Si se trabaja con ramillas terminales u hojas de gran tamaño es recomendable hacer una curva de calibración de peso versus tiempo en agua destilada Después de este tiempo se sacan las muestras se secan con papel absorbente e inmediatamente pesada para obtener el peso a turgor máximo (PT). 5.- Las muestras se colocan en estufa a 80° por 24 horas para obtener el peso seco de la muestra (PS). Con los datos de PF, PT y PS se calcula el CRA de acuerdo a Barr y Weatherley (1962).
100*secsec
oPesoPesototaloPesoPesofrescoCRA
−−
= (1)
Estado energético del agua en la planta El agua líquida, en la planta, existe como un continuo desde el suelo a través de las raíces, tallo y hoja. El movimiento del agua en este continuo está determinado por la diferencia de potencial hídrico (ΔΨ), es decir, el agua se mueve desde zonas de mayor potencial a zonas de menor potencial hídrico. El equilibrio se produce cuando ΔΨ= 0, es decir, el Ψ es igual en todas las partes del sistema vegetal y por lo tanto no existe movimiento de agua. Los componentes del potencial hídrico total (Ψ) son el potencial de gravedad (ψg), el potencial de presión (ψp), el potencial mátrico (ψm) y el potencial osmótico (ψ s). En el caso de los vegetales el potencial mátrico como el de gravedad son despreciables.
Ψ = ψ g + ψ p + ψ s + ψ m (2) Ψ = ψ p + ψ s (3)
Es posible medir el potencial hídrico total (ψ) a través de varias técnicas, como por ejemplo: psicometría de termocuplas, cámara a presión, método de la variación de volumen, etc. En este práctico se medirá ψ con la cámara a presión, su uso es ampliamente generalizado por la facilidad de manejo y por la obtención de las curvas presión-volumen (Schollander et al. , 1961, 1965; Steudle 2001; Salisbury y Ross, 1992).
CÁMARA A PRESIÓN TIPO SCHOLLANDER Medición del potencial hídrico total La medición del potencial hídrico total en la cámara a presión está basado en la teoría coheso-tenso-transpiratoria. La tensión en el xilema está dada por las fuerzas de cohesión entre moléculas de agua unidas por puentes de hidrógeno y fuerzas de adhesión entre moléculas de agua y las paredes de los vasos del xilema.
11 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
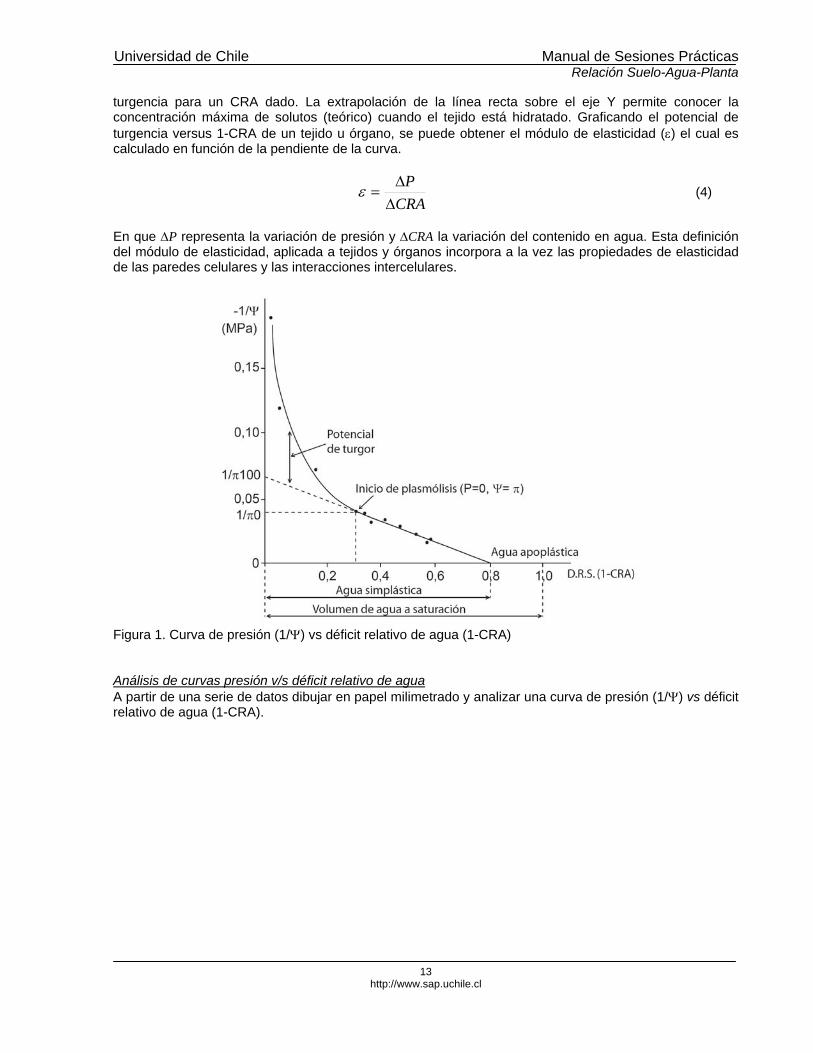
El sistema de la cámara a presión está constituido por una cámara donde se introduce parte del vegetal (tallo. Hoja o raíz) al que se medirá su Ψ, un balón de gas comprimido (usualmente N2), un manómetro, un tapón de goma que permite que la cámara quede cerrada en forma hermética. En el interior de la cámara se introduce un tallo o rama en el que se quiere medir la tensión presente en el xilema (ψ), con la zona de corte del vegetal hacia el exterior. El agua en el xilema está bajo tensión. Cuando se corta una rama o tallo desde un vegetal que está transpirando, la columna de agua presente en los vasos del xilema retrocede por debajo de la superficie cortada, debido a que se iguala la presión interna y la presión atmosférica. Al introducir la muestra vegetal en la cámara y aplicar presión, se reestablece la columna líquida y es posible visualizar exudación en la zona de corte. Por lo tanto, la presión que se necesita para forzar la columna de agua hasta la zona de corte es igual a la tensión que existía en la rama o tallo antes de ser seccionado, equivalente a su valor de Ψ, el cual se lee en el manómetro en el momento de la exudación, expresado en unidades de presión (bares o MPa). En hojas es posible obtener dos valores de potencial hídrico: el foliar y el xilemático. El potencial foliar se obtiene midiendo directamente sobre una hoja transpirante y el potencial xilemático se obtiene encerrando la hoja en papel aluminio y bolsa plástica a lo menos 1 hora antes de su medición. Curvas presión versus déficit relativo de agua Una de las ventajas de la técnica de la cámara a presión es que no sólo mide el potencial hídrico total (Ψ), sino que también se puede utilizar para medir la relación presión-volumen del tejido vegetal o curvas presión-déficit relativo de agua (-1/P versus 1-CRA). La interpretación de este tipo de curvas permite inferir diversos parámetros fisiológicos (Figura 1)
Procedimiento experimental 1. Se realiza un muestreo, generalmente al finalizar el día, de ramas u hojas situadas a una distancia de
10 y 20 cm del ápice, dependiendo de la especie. 2. Se mide el potencial hídrico total de las ramas en la cámara a presión y se pesan (peso fresco PF). 3. Las ramas son mantenidas en agua destilada durante un día hasta saturación, con la zona de corte
hacia el exterior. 4. Se vuelven a pesar las ramas y se obtiene el peso a máxima saturación (PT). Es recomendable
establecer una curva de calibración que relacione el cambio de peso con el tiempo y cuando los valores se estabilicen se puede determinar gráficamente el tiempo de saturación el cual depende de la especie
5. Siguiendo el procedimiento clásico desarrollado por Tyree y Hammel (1972), una rama es colocada en la cámara a presión donde es sometida a presiones crecientes desde 0,5 MPa a 6,0 MPa. El volumen exudado por efecto de la presión, que corresponde a líquido xilemático, es removido con uso de papel absorbente y es posible conocer su volumen.
6. Cada intervalo de presión tiene un valor de 1,0 MPa para obtener los primeros cinco puntos y luego de 0,5 MPa los siete puntos siguientes, la presión es aplicada durante 10 min. El peso fresco de muestreo es medido después de cada presión aplicada en una balanza de precisión de 0,001g. Este último aspecto constituye una variante respecto al procedimiento clásico.
7. Finalmente se obtiene el peso seco (PS) de las ramas manteniéndolas en estufa a 80ºC por 48 horas. Con estos datos se obtiene el valor de contenido relativo de agua (CRA) para cada presión aplicada.
12 http://www.sap.uchile.cl
La curva presión volumen, -1/P en función de 1-CRA, posee dos tramos, una zona no lineal donde existe una fluctuación de Ψ, que es la resultante de variaciones de presión osmótica (ψS), y de presión de turgencia (ψP), y una zona lineal que puede ser atribuida solamente al componente osmótico (ψS) (Silva y Acevedo, 1993; Lo Gullo et al., 2003). En la curva presión volumen, cuando la presión de turgencia alcanza un valor cero, es decir ψP= 0, la representación de -1/P en función de 1-CRA se hace lineal y el potencial hídrico total se iguala al potencial de soluto, esto es, Ψ= ψS. De acuerdo a la Figura 1, ψP= 0 corresponde al punto de plamólisis incipiente o inicio de plasmólisis que corresponde a una etapa en la cual la vacuola presente en la célula deja de ejercer presión sobre la pared celular. La extrapolación de la línea recta sobre el eje x permite estimar el volumen de agua apoplástico y por diferencia al volumen total de agua, el volumen de agua simplástico. La presión de equilibrio (P), en la curva -1/P= f(1-CRA) representa el potencial hídrico promedio de la rama. Despreciando el potencial osmótico del agua apoplástica, las variaciones de CRA corresponden a la cantidad neta de agua osmóticamente activa. En función de estos parámetros, es posible deducir los valores de potencial osmótico y del potencial de
Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
turgencia para un CRA dado. La extrapolación de la línea recta sobre el eje Y permite conocer la concentración máxima de solutos (teórico) cuando el tejido está hidratado. Graficando el potencial de turgencia versus 1-CRA de un tejido u órgano, se puede obtener el módulo de elasticidad (ε) el cual es calculado en función de la pendiente de la curva.
CRAP
ΔΔ
=ε (4)
En que ΔP representa la variación de presión y ΔCRA la variación del contenido en agua. Esta definición del módulo de elasticidad, aplicada a tejidos y órganos incorpora a la vez las propiedades de elasticidad de las paredes celulares y las interacciones intercelulares.
Figura 1. Curva de presión (1/Ψ) vs déficit relativo de agua (1-CRA) Análisis de curvas presión v/s déficit relativo de agua A partir de una serie de datos dibujar en papel milimetrado y analizar una curva de presión (1/Ψ) vs déficit relativo de agua (1-CRA).
13 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
TRABAJO EN TERRENO
REFERENCIAS BIBLIOGRÁFICAS BARR HD. and P.E. WEATHERLEY (1962) A re-examination of the relative turgidity technique for estimating water deficit in leaves. Australian Jopurnal Biological Science 15:413-428- LO GULLO M.A., SALLEO S.. ROSSO R. and P. TRIFILO (2003) Drought resistance of 2 year-old saplings of Mediterranean forest trees: relations between water relations. Hydraulics and productivity. Plant and Soil 250:259-272. SALISBURY, F.B. and C.W. ROSS (1992). Plant Physiology. Chapter 2, 3, 4. Wadsworth Publishing. 682 p. SILVA H y E. ACEVEDO (1993) Relaciones hídricas de Atriplex repanda Phil., a dos niveles de disponibilidad de agua. Revista Chilena de Historia Natura l66:467-477. SCHOLANDER, P.F., HEMMINGSEN, E.A. and W. GAREY (1961). Cohesive lift of sap in the rattan vine. Science 134: 1835-1838. SCHOLLANDER, P.F., HAMMEL, H.T., BRADSTREET, E.T. and E.A HEMMINGSEN, E.A. 1965. Sap pressure in vascular plants. Science 148: 339-346. STEUDLE E. (2001) The cohesion-tension mechanism and the acquisition of water by plants roots. Annual Review Plant Physiology Plant Mol Biology 52:847-875. TYREE, M.T. and H.T. HAMMEL (1972). The measurement of the turgor pressure and the water relations of plants by the pressure bomb technique. Journal Experimental Botany 23 : 267-282. WEATHERLEY, P.E. (1970). Some aspects of water relations. Advances in Botanical Research, 3 : 171 - 206.
14 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 6 ESTADO ENERGÉTICO DEL AGUA EN LA PLANTA
PSICROMETRIA DE TERMOCUPLAS I. COMPONENTES DE POTENCIAL HÍDRICO (Ψ) La psicrometría de termocuplas se basa en que la presión de vapor relativa (pa/p0) de una solución, o muestra de material vegetal, se halla relacionada con su potencial hídrico (Ψ), según la siguiente ecuación:
⎟⎟⎠
⎞⎜⎜⎝
⎛=Ψ
o
a
pp
VRT ln (1)
Donde, Ψ es el potencial hídrico en MPa, R es la constante universal de gases (8,31*10-6 m3 MPa mol-1 K°-1), T es la temperatura en grados Kelvin, V es el volumen molal del agua a la temperatura T (18,069 x 10-6 m-3 mol-1), pa es la presión actual de vapor y p0 es la presión de vapor a saturación (Brown, 1970; Brown & Oosterhuis, 1992). La psicrometría de termocuplas se refiere a la estimación del potencial hídrico, (Ψ) a través de mediciones de cambio de temperatura (ΔT) durante el proceso de evaporación del agua desde una superficie que se enfría. La diferencia de temperatura (ΔT) corresponde a una presión de vapor determinada y un microvoltímetro registra las variaciones de voltaje asociadas a las diferencias de presión de vapor del agua. El enfriamiento se produce debido al escape de moléculas de agua desde la fase líquida hacia la fase gaseosa. En la fase líquida, se encuentran moléculas de agua que tienen una alta energía cinética y que les permite romper las fuerzas intermoleculares que las retienen en la fase líquida. Al pasar a la fase de vapor quedan en la fase líquida aquellas moléculas de agua de menor energía cinética y por lo tanto baja la temperatura de la fase líquida produciéndose el enfriamiento antes mencionado.
Psicrómetros
Los psicrómetros están constituidos por una o dos termocuplas (sensores de temperatura) unidas a alambres conductores y conectadas a un microvoltímetro. Los sensores de temperatura se encuentran en el interior de una cámara cerrada en forma hermética que permite la transferencia de vapor de agua y a la vez evita el contacto de la muestra con la termocupla. Bajo estas condiciones isotérmicas, la presión de vapor en el interior de la cámara sobre la muestra y alrededor de la termocupla alcanzará el equilibrio con el potencial hídrico de la muestra, generalmente después de 2 a 4 horas. Una vez que se alcanza el equilibrio de la presión de vapor y de temperatura en el sistema, se registra el potencial hídrico de la muestra. Psicrómetro Richards & Ogata y Técnica Isopiéstica El psicrómetro de Richards y Ogata (1958) tiene una estructura similar al ya descrito, pero además consiste en que en el terminal de las uniones de chromel-constantan de la termocupla va unido un pequeño anillo de plata. (Figura 1). En el interior de la cámara se coloca una muestra de tejido vegetal cuyo potencial se desconoce, y en el anillo se coloca una gota de solución de potencial osmótico conocido (ψs), luego se introduce en la cámara, se cierra herméticamente y se sumerge en baño de agua para mantener constante la temperatura. De acuerdo a lo anterior es posible obtener dos situaciones: 1. El potencial hídrico de la gota de solución conocida, es mayor que el potencial hídrico de la muestra de
tejido vegetal. Entonces habrá moléculas de agua presentes en la solución de concentración conocida que pasarán a la fase de vapor produciendo el humedecimiento del aire que se encuentra en el interior de la cámara sellada, difundirán y serán absorbidas por la muestra de tejido. Producto de esta evaporación se produce una disminución de la temperatura en la solución que es captada por la termocupla y que genera una diferencia de potencial eléctrico que es registrada por el microvoltímetro
15 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
y cuya magnitud es directamente proporcional a la diferencia de temperatura y por lo tanto, al potencial hídrico de la muestra.
2. El potencial hídrico de la gota de solución conocida, es menor que el potencial hídrico de la muestra de
tejido vegetal. Entonces se producirá condensación de agua en la gota de concentración conocida y producto de la condensación ocurrirá un calentamiento de la termocupla. Nuevamente esta diferencia de temperatura será captada por la termocupla y generará una diferencia de potencial eléctrico que es registrada por el microvoltímetro.
Para obtener el valor del potencial de la muestra se colocan diferentes soluciones en el anillo en intervalos de 2 horas, hasta obtener igualdad de potencial hídrico entre la gota de solución conocida y la muestra. En este caso la diferencia de temperatura en la termocupla es 0 y el flujo neto de vapor de agua también es 0, es decir, que la cantidad de agua que pierde la solución de concentración conocida es igual a la cantidad de agua que pierde la muestra problema. Luego, el punto en que la presión de vapor entre la solución de concentración conocida y la muestra problema es igual, se denomina PUNTO ISOPIÉSTICO. Debido a que la relación entre la diferencia de potencial gota-muestra y la diferencia de voltage registrada es lineal, en la práctica bastan 2 ó 3 soluciones y luego se realiza una interpolación lineal (Figura 2). El Ψ y sus componentes (ψp y ψs), pueden ser medidos en cámaras bajo condiciones isotérmicas, a nivel de hojas y otros tejidos vegetales. Para este efecto, deben adoptarse importantes precauciones en laboratorio y en terreno. La superficie foliar debe ser limpia y seca (libre de polvo, humedad o agua) anterior al muestreo. Se ha observado gradiente de potencial hídrico a través de hojas, especialmente cuando se usan discos foliares y/o trozos de tejido (Oosterhuis y Wullsschleger, 1987; 1989). Por lo tanto, es esencial ser consistente en la selección de la muestra dentro y entre hojas. La velocidad y consistencia entre el tiempo de muestreo foliar y el sellado de la muestra en la cámara para evitar excesiva pérdida de agua. Un lapso de 1 minuto puede resultar en errores de 0,5 a 0,7 MPa (Boyer y Knipling, 1965). La magnitud de estos errores está en relación al tamaño de la muestra foliar y de la demanda evaporativa de la atmósfera. Las hojas requieren un lapso de equilibrio bastante largo en comparación a aquél para la calibración de soluciones debido a la resistencia de la superficie foliar. Por lo que se requiere precisar el tiempo de calibración foliar, el cual es variable en función de la especie. Procedimiento de Calibración y Medición de Potenciales a) Preparar soluciones de potencial conocido con KCl, NaCl o sacarosa de 0 - 0,5 - 1,0 - 2,0 y 3,0 MPa. b) Limpiar cuidadosamente la cámara y el vástago. La argolla de plata se lava con agua destilada y luego se seca con cloroformo. (Figura 1). c) Colocar una capa delgada de vaselina sólida en la base del vástago y en el interior de la cámara. d) Cubrir con papel filtro la pared y la base de la cámara, y luego humedecerlo con agua destilada y, posteriormente con cada una de las soluciones. e) Colocar agua destilada en la argolla después de 2 horas en que se supone se alcanzó el equilibrio inicial. Una vez introducida la termocupla se espera 1,5 h y se registra la primera lectura. f) Se continúa con el resto de las soluciones, lavando muy bien la argolla entre mediciones. Una vez realizada la última lectura, se gráfica la curva en función de los datos obtenidos (μVol versus potencial hídrico solución conocida en MPa). Esta fase de calibración permite detectar errores de lectura o de funcionamiento y seleccionar psicrómetros. Una vez obtenida una curva de calibración confiable, se realiza un ensayo preliminar con tejido vegetal cubriendo la totalidad de la superficie de la cámara y así obtener el tiempo promedio en que se alcanza el equilibrio de presión de vapor entre la muestra vegetal y la solución en la argolla, repetir fases a, b, c, e y f, en d reemplazar papel filtro por el tejido. El Ψ foliar es medido en tejido fresco como ya se ha explicado, mientras que el componente osmótico es normalmente determinado por congelamiento y descongelamiento del tejido en la cámara mediante N líquido o hielo seco, procedimiento que permite destruir las membranas celulares. En estas condiciones el ψp= 0 y puesto que Ψ= ψs + ψp, entonces, Ψ= ψs. Enseguida la muestra es retornada a las condiciones isotérmicas hasta restablecer el equilibrio del sistema y proceder a la medición del potencial osmótico, procediendo de acuerdo a las diferentes fases señaladas. Una vez graficados el potencial hídrico (curva inicial) y el potencial osmótico (curva final), el turgor se obtiene por método gráfico. El potencial de presión o turgor, se obtiene por sustracción:
16 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
ψp = Ψ - ψs (2)
0
50
100
150
200
250
300
350
0 0.5 1 1.5 2 2.5
Concentración M
uVol
3
ΨT ΨS
Figura 2: Gráfico de Relación entre lecturas en μVol versus concentración de soluciones.
17 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
II. DETERMINACIÓN DEL POTENCIAL DE SOLUTOS POR OSMOMETRÍA DE PRESIÓN DE VAPOR En la mayoría de las soluciones, los cambios en concentración son acompañados por cambios proporcionales y lineales en las propiedades coligativas del solvente: la presión de vapor, el punto de congelación y el punto de ebullición. La medición de cada una de estas propiedades proporciona un índice indirecto de la osmolalidad, entre ellas solamente la presión de vapor puede ser determinada sin un cambio forzado en el estado físico de la muestra. La medición de la presión de vapor es el método más fácil y rápido para determinar la osmolalidad, y las limitaciones termodinámicas de la osmometría de presión de vapor son menos restrictivas. Por esta razón la osmometría basada en presión de vapor es el método más adecuado para la mayoría de los fluidos en que el agua es el solvente (Sweeney y Beuchat, citado por Wescor, 1998). En el vapro-osmómetro, la presión de vapor se determina mediante una termocupla suspendida en un pequeño espacio sobre la muestra en el interior de la cámara sellada. Durante un ciclo de mediciones, la termocupla experimenta una serie de cambios (microprocesados) controlados por la temperatura. Etapa 1. Después de la inserción de la muestra, la presión de vapor y la temperatura llegan a un equilibrio en la cámara sellada. La termocupla sensa la temperatura exacta de aire sobre la muestra y el microprocesador fija este valor como el punto nulo o punto de referencia para la medición (TA). Etapa 2. La termocupla es enfriada por efecto Peltier por debajo de la temperatura del punto de rocío (TD). Gotas de agua microscópicas comienzan a condensarse sobre la superficie de la termocupla. Etapa 3. El microprocesador permite que la temperatura de la termocupla sea controlada exclusivamente por la condensación de agua sobre ella. El calor de condensación provoca un aumento de la temperatura en la termocupla, convergiendo en al temperatura exacta a la cual cesa la condensación. Esta condición estable es alcanzada aproximadamente 1 minuto después de la inserción de la muestra. Etapa 4. El punto de estabilidad de la temperatura es la temperatura del punto de rocío. La temperatura final sobre el display del instrumento es proporcional a la depresión de la temperatura del punto de rocío con una resolución de 0,0003ºC. Debido a que la depresión de la temperatura del punto rocío es función explícita de la presión de vapor, el instrumento puede ser calibrado para proporcionar directamente la osmolalidad en unidades del SI (mmol/kg). Descripción del sistema. La ventaja principal del método de presión de vapor es que no requiere alteración del estado físico de la muestra. Requiere de un tamaño de muestra de 10 ul, la operación de rutina es a micro-escala, incluye muestras complejas tales como muestras de tejido. Mayor confiabilidad debido a que las mediciones involucran mínima complejidad mecánica. Una muestra de 10 ul es aspirada mediante una micropipeta. Luego ésta es inoculada sobre un disco de papel filtro sobre el cabezal de muestreo, el cual es empujado al interior del instrumento iniciando la secuencia automática de medición. El elemento sensible es un higrómetro de termocupla (de alambres muy finos). La termocupla entonces, es suspendida en una base metálica, la cual se une con el portador de la muestra formando una pequeña cámara que la encierra. A medida que se equilibra la presión de vapor en la cámara, la termocupla sensa la temperatura ambiente del aire, estableciendo el punto de referencia para la medición. Bajo control electrónico la termocupla determina la temperatura del punto de rocío en el interior de la cámara, dando una salida proporcional al diferencial de temperatura. La diferencia entre la temperatura ambiente y la temperatura del punto de rocío es la depresión de la temperatura del punto de rocío. Una función explícita de la presión de vapor de una solución. Las mediciones de control de microprocesador para un ciclo de medición requiere de 75 segundos. En 1887, J.H. Van Hoff descubrió una relación empírica que permite el cálculo del potencial osmótico a partir de la concentración molal de la solución. Graficó el potencial osmótico de lecturas directas desde un osmómetro como función de la concentración molal, obteniendo la siguiente relación, forma que es idéntica a la ley de los gases perfectos:
ψs = CI··R·T (3) C = Concentración de la solución en molalidad (mol de soluto / Kg de agua)
18 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
I = Constante que considera la ionización y otras desviaciones de las soluciones perfectas. R = Constante universal de los gases ( 0.0083 kg MPa mol-1 K-1) T = Temperatura absoluta (K) ψs = Potencial de osmótico (MPa) REFERENCIAS BIBLIOGRÁFICAS BOYER, J. & KNIPLING E. 1965. Isopiestic technique for measuring leaf water potentials with a thermocouple psychrometer. Proceedings of National Academy of Sciences 54(4): 1044-1051. BROWN, R.W. 1970. Measurement of water potential with thermocouple psychrometers: Construction and applications. USDA Forest Service Research Paper INT 80. 27 p. BROWN, R.W. & OOSTERHUIS, D.M. 1992. Measuring Plant and Soil Water Potentials with Thermocouple Psychrometers: Some Concerns. Agronomy Journal 84: 78-86. OOSTERHUIS, D.M. & WULLSCHLEGER, S.D. 1987. The use of leaf discs in thermocouple psychrometers for measurement of water potential. p 77. In R.J. Hanks and R.W. Brown (ed) Proc. Int. Conf. on Measurement of Soil and Plant Water Status, Vol. 2 Utah State Univ., Logan UT. 6-10 July 1987. Utah State Univ., Agricultural Experiment Station. OOSTERHUIS, D.M. & WULLSCHLEGER, S.D. 1989. Psychrometric water potential analysis in leaf discs. p. 113-133. In H.F. Linkens, (ed). New Series, Vol. 9. Springer Verlag, Berlin. RICHARSD, L.A. & OGATA, G. 1958. Thermocouple for water pressure measurent in biological and soil systems and high humidity. Science 128:1089-1090. SALISBURY, F.B. & ROSS.C.W. 1992. Plant Physiology. Chapter 2, 3, 4. Wadsworth Publishing. 682 p. WESCOR. 1998. VAPRO. Vapor Pressure Osmometer. User´s manual. Model 5520.100p WIEBE, H.H. 1984. Water condensation on Peltier-cooled thermocouple psychrometers: a photographic study. Agronomy Journal 76: 166-168.
19 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 7 INTERCAMBIO GASEOSO
TRANSPIRACIÓN Y FOTOSINTESIS Las mediciones de transpiración foliar y los cálculos de conductancia foliar al vapor de agua son parámetros importantes en el estudio del continuo suelo-planta-atmósfera. La transpiración es determinante en el balance energético a nivel foliar y en el estado hídrico de la planta. Junto al intercambio de CO2 determina la eficiencia en el uso del agua (EUA) o eficiencia transpiracional, parámetro utilizado como criterio de selección de especies para zonas de pluviometría limitada. La estrecha relación entre la captación de CO2 y la pérdida de H2O a través de los estomas ha permitido una separación de los factores que limitan el proceso fotosintético en estomáticos y/o bioquímicos. TRANSPIRACIÓN En sistemas de intercambio gaseoso, la transpiración se calcula midiendo el vapor de agua en una atmósfera cerrada que rodea a la hoja. Una hoja puede ser introducida en una cámara (extremo de la pinza) o cuveta (pinza) transparente, donde es posible medir la humedad, la temperatura y el volumen de gas que entra y aquel que sale de la cubeta. Con estos datos es posible calcular la tasa de transpiración, la conductancia estomática (o su resistencia) y la temperatura. La humedad absoluta (densidad de vapor o presión) en moles o g por m3 o en Pascal, es medida directamente con un higrómetro de punto de rocío o bien calculada usando ecuaciones apropiadas. Conociendo el volumen de aire en la cubeta y los cambios en la densidad del vapor de agua, la cantidad de agua transpirada desde la hoja es igual a la cantidad de agua aportada al aire. La transpiración es expresada como g o moles de agua por m2 de hoja por segundo (moles m-2 s-1). Si los datos están siendo comparados a la fotosíntesis es lógico usar moles en lugar de g de agua. Una de las expresiones más simples para calcular la tasa transpiratoria (T) a nivel de tejidos foliares, se expresa como:
sva gCT *Δ= o rCT va /Δ= (1) Los instrumentos más utilizados para la medición de la conductancia estomática son los porómetros de difusión. En principio todos los porómetros de difusión miden transpiración, la cual es utilizada para inferir un valor de conductancia. Los porómetros más utilizados son aquellos que mantienen la humedad constante a través de un flujo de aire seco en la cámara para balancear el vapor de agua que pierde la hoja, denominados porómetros de balance nulo. Beardsell et al. (1972), fue el primero en desarrollar un porómetro de balance nulo, utilizando un flujo de aire seco a traves de una cámara para balancear la tasa de transpiración de una hoja y por lo tanto, mantener una humedad constante. Debido a que la humedad del aire que entra a la cámara es conocida (generalmente 0% o cercano al 0%), puede demostrarse que bajo condiciones isotérmicas, la determinación de la conductancia al vapor de agua, requiere solamente la medición de humedad del aire dentro de la cámara, la tasa de flujo de aire seco que entra a la cámara y el área foliar. En la práctica, las condiciones no son totalmente isotérmicas por lo tanto es necesario medir la temperatura foliar y la del aire. Uno de los porómetros de difusión más simples es el DeltaT, modelo Mk3, provisto de un sensor de humedad, este sensor tiene una capacitancia eléctrica que cambia linealmente con la humedad relativa. En la cubeta de intercambio se controla la temperatura de la hoja con un termistor. El sensor fija la HR ambiental y un contador de 200 Hz, permite medir el “tiempo relativo” en que se alcanza la HR previamente fijada. Esta calibración se realiza en base a placas perforadas de resistencias conocidas para temperaturas que varían entre 15 y 35ºC, de acuerdo a procedimiento establecido por Morrow y Slatyer (1971). El gráfico de calibración relaciona las resistencias conocidas (s cm-1) con el “conteo de tiempo” correspondiente. A partir de este gráfico se convierten los conteos obtenidos de las mediciones foliares en resistencia difusiva o conductancia. A 20°C los valores de resistencia de la placa de calibración son los siguientes: 22,5 - 10,9 - 6,5 - 2,9 - 1,3 y 0,4 (s cm-1) para 1, 2, 3, 4, 5 y 6 “perforaciónes” que simulan a los estomas. Estos valores fueron obtenidos aplicando la siguiente fórmula para resistencia difusiva: Para posiciones 1 a 5
20 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
)4/()8/(
2dDnLArππ+
= (2)
y para 6
DLr = (3)
Donde: r es la resistencia en (s cm-1), A es el área de la cubeta cm2 (0,559 cm2); n es el número de perforaciones, L es el grosor de la placa (cm); d es el diámetro de las perforaciones y D es el coeficiente de difusión (0,242 a 20°C). El LI-COR 1600, utiliza un sensor tipo VAISALA, montado en una cámara ventilada. Las aperturas de cámara son intercambiables para una amplia variedad de formas y dimensiones foliares. Un soporte externo con un segundo ventilador ayuda a mantener la temperatura de la cámara cercana a la temperatura del aire ambiente. Un flujo de aire seco es suplementado a la cámara mediante una bomba y tubos desecantes con silica-gel, esta tasa de flujo es medida con un medidor de flujo de masa eléctrico. Un sistema de válvulas conectadas regula el flujo en la cámara para mantener una humedad en equilibrio al nivel pre-seleccionado por el operador. En la práctica, la humedad ambiental es generalmente medida con la cámara abierta. El ajuste manual se requiere para llevar el flujo en el rango donde la válvula automática pueda controlarla convenientemente. Un micropocesador basado en un sistema de adquisición de datos, mide las señales mediante termocuplas desde la hoja y ambiente de la cámara, desde el medidor de flujo y sensor de humedad y proporciona la conductancia estomática sobre un LCD display. Las mediciones de conductancia estomática generalmente requiere entre 15” y 1’. Una batería recargable permite uso continuo hasta 6 horas. El Li-1600 usa mediciones primarias para calcular la resistencia o conductancia por lo tanto, no requiere ni curvas de calibración ni coeficientes empíricos. El agua perdida por transpiración desde una hoja colocada en la cámara de intercambio es determinada por la medición de la tasa de flujo de aire seco necesario para mantener una HR constante en el interior de la cámara. En este caso la HR ambiente es usada como el punto nulo, y el aire seco es incorporado a la cubeta a una tasa suficiente para balancear el flujo transpiratorio foliar, lo cual mantiene la HR en el punto nulo (set-point). La resistencia o conductancia es medida directamente de los valores medidos de HR, temperatura del aire y temperatura foliar y de la tasa de flujo. La tasa transpiratoria (T) se relaciona al flujo volumétrico de acuerdo a:
AFT ac )( ρρ −= (4)
Donde: ρc (μ cm-3) es la densidad del vapor de agua en la cuveta, ρa es la densidad del vapor de agua del flujo de aire seco que entra a la cuveta (se asume un HR constante de 2%) y A es el área foliar en cm2. La transpiración foliar también puede ser expresada en términos de gradiente de vapor de agua entre la hoja y el aire dividido por la suma de las resistencias estomática y de la capa límite:
cle
cl
rrT
+−
=ρρ
(5)
combinando las ecuaciones 4 y 5:
clac
cle r
FAr −⎟⎟
⎠
⎞⎜⎜⎝
⎛−−
==ρρρρ
(6)
El Li-1600 usa la ecuación 6 con rcl= 0,15 s cm-1 para calcular la resistencia estomática. La memoria del equipo proporciona la información de la densidad de vapor de agua en función de la temperatura (tabla psicrométrica). Se asume que la atmósfera foliar interna está a saturación, así la densidad del vapor de agua corresponde a la densidad de vapor de agua a saturación a la temperatura foliar medida.
21 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
FOTOSINTESIS SISTEMA DE INTERCAMBIO GASEOSO (IRGA) PARA TEJIDO FOTOSINTÉTICO LAMINAR EN PLANTAS C3, C4. Para el estudio de la fotosíntesis a nivel foliar, de la planta o de la cubierta vegetal, el (los) instrumento(s) utilizado (s) son identificados como sistemas de intercambio gaseoso o simplemente sistemas. Es posible medir la fotosíntesis sin transpiración y viceversa, pero la efectividad de medición del proceso aumenta si simultáneamente se mide la transpiración por dos razones. Primero las dos mediciones son sinérgicas debido a que los dos procesos son interdependientes. El CO2 difunde en las hojas y el agua difunde fuera de la hoja. La respuesta estomática disminuye la pérdida de agua y disminuye la fotosíntesis, y las condiciones que aumentan la fotosíntesis también aumentan la pérdida de agua. Segundo, la conductancia foliar al vapor de agua, calculada a partir de las mediciones de intercambio de H2O, es un parámetro determinante para el calculo de la concentración interna de CO2 (Ci). Ci puede ser medido directamente pero la medición es técnicamente difícil (Sharkey et al., 1982). La solución más simple es calcular Ci usando la conductancia foliar al vapor de H2O. Debido a que el modelo de difusión para CO2 y H2O son esencialmente el mismo, podemos calcular la conductancia foliar para CO2 a partir de la conductancia al vapor de agua (Nobel, 1983). Estas mediciones pueden efectuarse a diferentes escalas: desde el nivel de órganos (toda o parte de la hoja) como a nivel de la planta entera o del cultivo. Para poder efectuar esto, el principio general de medición es colocar el órgano o la planta dentro de una cámara, hacer pasar aire a través de esta a un flujo conocido y luego medir la diferencia de concentración del CO2 entre el aire que entra y el que sale de la cámara. Así, la tasa de intercambio de CO2 puede ser expresada en función del área foliar, de la planta entera o de la materia seca del cultivo. De igual forma, la escala de tiempo de la medición también es importante. Esta puede ir desde el segundo hasta el día. Para relacionar la medición con las condiciones del medio, las condiciones en la cámara deben ser medidas y controladas cuidadosamente por ejemplo, composición del aire, temperatura, flujo fotónico y humedad. MEDICIÓN DE CO2 La producción total de un cultivo depende de la diferencia entre fotosíntesis bruta (Fgross) y las pérdidas por respiración o fotorespiración (Rphot) más la respiracion oscura (Rdark). En el caso de una hoja en la luz, la fotosíntesis neta es igual a: Fn = Fb - (Rphot + Rdark). Para medir los diferentes componentes de esta ecuación existen varios métodos: a) Fotosíntesis bruta. La fotosíntesis bruta puede ser medida usando trazadores marcados con 14CO2 de actividad específica conocida cuya absorción puede ser medida por períodos cortos (normalmente menos de 1 segundo). Esto se efectúa midiendo la disminución de la cantidad de radioactividad del 14C en el aire de la cámara, o bien determinando el aumento del isótopo en la materia seca (Calvin & Benson, 1948). Los isótopos del carbón no son usados de la misma forma por la fotosíntesis. Hay una discriminación contra el 13C (también la hay en el 14C). La relación entre 13C y 12C puede ser usada para analizar diferentes procesos fotosintéticos (eficiencia transpiracional). b) Fotosíntesis neta. La fotosíntesis neta es normalmente medida mediante el uso de un analizador de gases infrarrojo (IRGA). Este determina la cantidad de moléculas de CO2 en un determinado volumen de aire. El CO2 absorbe la radiación infrarroja, especialmente aquella de longitud de 4,26 μm, previa eliminación del agua y otros gases con filtros adecuados. La cantidad de radiación que llega al detector es función de la cantidad de moléculas de CO2 que se encuentran entre la fuente de radiación y el detector. De manera similar, para la medición de la transpiración, el vapor de agua absorbe radiación IR en la longitud de 2,56 μm. La forma absoluta permite la obtención de la cantidad real de CO2 en el aire. Sin embargo, es poco sensible para análisis en fisiología vegetal. Por el contrario la forma diferencial es mucho más sensible, pero es menos estable. Actualmente, en el comercio se pueden encontrar diferentes tipos de IRGA, con diferentes sensores, fuentes infrarrojas, mostrando diferente sensibilidad al CO2 y al vapor de agua. Todos los IRGAS deben ser calibrados con gases estándares y la sensibilidad al vapor de agua debe también ser controlada. El efecto de la temperatura y presión debe ser considerada en los cálculos. Para la calibración, algunos de los equipos actuales presentan tubos divididos en diferentes proporciones, que permiten calibrar rápidamente, como el modelo 125 de la marca ADC. Otros poseen una referencia fija en el instrumento que se usa para calibrar tanto el gas como la referencia usada. El sistema PMS usa la técnica no dispersiva de medición de IR asociada con un microprocesador de señales para lograr estabilidad y especificidad para CO2 y H2O. Utiliza 4 analizadores IR, 2 para CO2 y 2 para vapor de agua. Un par
22 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
CO2/H2O tiene una entrada y salida comunes y es definido como referencia y el otro par es definido como de análisis. Las mediciones son expresadas como concentraciones absolutas para la referencia y como la diferencia entre la referencia y la concentración del análisis. Cálculo: 1.- Cálculo del flujo en masa del aire por unidad de área foliar que entra a la cubeta (W). El sistema está calibrado para leer el volumen de flujo a 20°C y a 1 bar (V20). Dado que el volumen molar es 22,4 a 0°C a 1 atm entonces:
)/10000(*)013,1/1(*)293/273(*)4,22/1(*)1000/( 20 aVW = (7) Donde a es el área foliar . 2.- La tasa de T se calcula a partir de la presión de vapor del aire que entra (een) y deja la cubeta (esal). El flujo molar del agua en la cubeta es:
12**)/( −− smmolWPeen (8) el flujo molar de aire fuera de la cubeta, debido a la adición del agua transpirada, es (E + W). Por lo tanto, el flujo molar de agua fuera de la cubeta es:
12 **)(*)/( −−+ smmolWTPesal (9) Pero la diferencia entre flujo molar que entra y sale de la cubeta debe ser igual a la transpiración:
)/*())/(*)(( PeWPeTWT ensal −+= (10) Por lo tanto,
)/())(*( salensal ePeeWT −−= (11) La tasa de fotosíntesis (F) se calcula a partir de la diferencia en la concentración de CO2 que entra y aquél que deja a esa tasa de flujo a través de la cubeta. Las lecturas de CO2 son corregidas por la temperatura del vapor de agua y de la presión atmosférica. También la adición de vapor de agua por transpiración en la cubeta diluye el aire de salida, y ésta debe ser compensada en el cálculo:
)(** TWCWCF salen +−= (12) Finalmente:
TCCCWF salensal *)(*( +−−= (13) Pero (Csal – Cen) es la diferencia de CO2 que es calculada por el sistema. Definiciones:
Símbolo Parámetro medido Unidades V Volumen de aire seco a la cubeta cm3 s-1 A Area foliar cm2 rcl Resistencia de la capa límite al vapor de agua m2 s mol-1 P Presión atmosférica bar V20 Flujo de masa de aire a la cubeta a 20°C y 1
bar cm3 s-1
23 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
FOTOSINTESIS EN TEJIDOS NO LAMINARES ACIDEZ TITULABLE TEJIDO FOTOSINTÉTICO DE PLANTAS CAM En plantas con metabolismo ácido crasuláceo, la captación de CO2 durante la noche significa la fijación neta de CO2 en forma de ácido málico. Por lo tanto, el cambio de acidez titulable durante el período de oscuridad corresponde en gran parte a un cambio en el contenido de este ácido (Kluge & Ting, 1978; von Willert & Kluge, 1973), entre otros, sugieren que la acumulación de ácido málico puede estar integrada el mecanismo que controla la abertura estomática de las especies CAM. Después de un período de fijación de CO2 con la acumulación de ácidos y la disminución de carbohidratos hay una fuerte disminución del contenido de este ácido cuando la planta recibe luz. Las variaciones del contenido de ácido málico del tejido fotosintético pueden ser medidas a partir de una titulación: variación de acidez titulable. Esta variación se evalúa en función de los cambios del contenido de ácido málico por medio de la titulación de acidez del tejido fotosintético entre al atardecer y el amanecer del día siguiente. Este método se fundamenta en el hecho que el CO2 es incorporado en ácido orgánico (4 átomos de carbono) luego acumulado en las vacuolas del clorénquima, lo cual conduce a los cambios de acidez (Nobel & Hartsock, 1983). PROCEDIMIENTO Para medir los niveles de acidez se muestrea parte del tejido fotosintético a nivel del cladodio con un sacabocados de 1,13 cm2 de superficie (n = 3). El clorénquima de cada lado de la muestra es separado y molido en arena esterilizada agregando 30 ml de H2O destilada y enseguida titulada a pH 6,4 con NaOH 0,01 N con la ayuda de un pH metro portátil (WTW pH90). Los resultados experimentales indican que el 95% de los ácidos orgánicos que contribuyen a la variación de acidez son neutralizados a pH 6,4 (Nobel, 1983). Los datos son expresados en moles de CO2 por m2 de tejido fotosintético por día (Kluge y Ting, 1978; Nobel y Hartsock, 1983), suponiendo una estequiometría aceptada por diversos autores de 2H+ por un CO2 fijado (Luttge & Ball, 1980; Nobel y Hartsock, 1983). Para el cálculo de la eficiencia transpiracional o EUA instantánea, los valores pueden expresarse en g MS m-2 dia-1, utilizando el factor de conversión de 0,027 kg de MS por mol de CO2 fijado (Garcia y Nobel, 1986).
24 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 8 INTERCAMBIO GASEOSO
RESISTENCIAS ASOCIADAS AL INTERCAMBIO GASEOSO
RESISTENCIAS ASOCIADAS A LA PERDIDA DE VAPOR DE AGUA. COMPORTAMIENTO ESTOMÁTICO. Es fundamental conocer el rol que cumplen los estomas, ya que regulan más del 90% del intercambio gaseoso entre la planta y su medio. Analizaremos el comportamiento estomático en términos de resistencia a la pérdida de vapor de agua haciendo una analogía con la ley de Ohm de transporte eléctrico. Esta serie de resistencias dadas por las características anatómicas y estructurales del órgano foliar es posible cuantificarlas mediante el uso de porómetros de flujo difusivo. 1.- Movimientos estomáticos. Los estomas se encuentran en todas las especies vegetales superiores y su función es el intercambio gaseoso de vapor de agua y CO2. Las plantas se enfrentan a un dilema de prioridades opuestas cómo obtener el máximo posible de CO2 desde una atmósfera en la cual se encuentra extremadamente diluido (0,035% en volumen) y al mismo tiempo retener el máximo de agua posible. La comprensión de los factores ambientales y cómo ellos influyen en la transpiración y absorción de CO2 a nivel foliar en el campo y en función del tiempo es difícil debido a que los factores interactúan en muchas formas. Los factores ambientales no sólo influyen en el proceso físico de evaporación y difusión, sino también en la apertura y cierre de los estomas. El aumento de la temperatura foliar, por ejemplo, promueve considerablemente la evaporación y ligeramente la difusión. Sin embargo, puede determinar el cierre o la apertura máxima de los estomas dependiendo de la especie y de otros factores. Al amanecer los estomas se abren en respuesta al aumento de la luz, y la luz aumenta la temperatura foliar, lo que determina que el agua se evapore más rápidamente. Si la hoja es calentada por sobre la temperatura del aire por la luz del sol, el viento disminuye la temperatura, disminuyendo la transpiración. Cuando la humedad del suelo es limitante, la transpiración y captación de CO2 se inhiben debido al cierre estomático. Las plantas tienen mecanismos de regulación temporal para la abertura estomática, de tal forma de evitar la pérdida de agua innecesa. La salida de vapor de agua a través de los estomas posibilita que el agua entre desde el suelo transportando nutrientes minerales hacia la parte aérea y además, permite que la vaporización de este elemento, desde su estado líquido, mantenga la temperatura foliar por debajo de la temperatura ambiente. No obstante lo anterior, en condiciones de restricción de agua las plantas deben evitar la pérdida de este elemento, cerrando sus estomas. Las células oclusivas poseen propiedades que la hacen particularmente eficiente en la formación del poro: a) Pueden alterar rápida y reversiblemente su turgencia. b) La forma de la célula oclusiva, a baja y elevada turgencia depende de las propiedades de la pared celular (disposición, orientación, de las microfibrillas de celulosa en la pared celular. c) Las células oclusivas no se encuentran comunicadas con las células adyacentes a través de plasmodesmos. Los movimientos estomáticos dependen de cambios en la presión de turgencia los que resultan de una modificación del potencial hídrico de las células oclusivas: mecanismo pasivo, o de cambios en su potencial osmótico: mecanismo activo. En consecuencia las células oclusivas sensan ya sea una señal hidráulica o una señal química representada por el ABA. Se desconoce la forma de transducción y cómo se produce la interacción de ambas señales. En ambos casos ocurre un movimiento de agua hacia o desde las células oclusivas, aumentando o disminuyendo el tamaño del poro estomático, respectivamente. En el caso del cierre estomático observado al mediodía, éste se debe a la pérdida de agua desde las células oclusivas, disminuyendo el turgor, lo que cierra el poro estomático. En muchos casos, sin embargo, el agua se pierde lentamente desde el suelo, de tal forma que la disminución de este potencial hídrico del suelo, es “advertido primariamente por las raíces”. Desde aquí se envían señales a la parte aérea posiblemente por medio de reguladores de crecimiento, particularmente el ácido abscísico, los que inducen el cierre estomático. Todo lo anterior ha conducido a postular una serie de hipótesis que pretenden explicar el comportamiento estomático basado en señales hidráulicas (Comstock, 2002) y señales químicas (Zhang y Davies, 2003). Cowan y Farqhuar (1982) postularon la teoría de la optimización del comportamiento estomático: “habría un momento del día en el cual los estomas maximizan su abertura y esta abertura condiciona la mayor o menor eficiencia de transpiración a escala instantánea”. Resultados registrados en Vitis vinífera cv Crimson Seedless señalan máxima abertura de
25 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
estomas en las primeras horas del día. En estas condiciones el déficit de presión de vapor es mínimo por lo tanto la tasa transpiratoria es baja y la carboxilación es máxima lo que condiciona una mayor eficiencia de transpiración, lo cual explicaría la resistencia a la sequía que poseen las especies de V. vinifera (datos no publicados). RESISTENCIAS AL FLUJO DE VAPOR DE AGUA. Los estomas controlan aproximadamente el 90% de la salida del agua desde las hojas y la entrada del CO2 a ella. Aunque las células epidérmicas ocupan un mayor porcentaje de la superficie foliar que los poros estomáticos, la cutícula que cubre la superficie externa de las células epidérmicas, lo que reduce la pérdida de agua desde las paredes celulares al aire turbulento que rodea a la hoja. La vía normal para el vapor de agua que sale de la hoja durante la transpiración es a través de los estomas. Consideremos la transpiración de agua y la fijación fotosintética de CO2. Las fuerzas generadoras de tales flujos son las diferencias de concentración o fracción molar de H2O y CO2. Trataremos de discutir cuantitativamente el movimiento de gases fuera y dentro de las hojas en términos de resistencias y/o conductancias involucradas. Las resistencias o conductancias son aquéllas encontradas por el vapor de agua a medida que difunde desde los poros de las paredes celulares de las células del mesófilo a otros sitios de evaporación de agua hacia el aire turbulento que rodea a la hoja. Esto se define cuantitativamente a nivel de espacios intercelulares, el estoma, la cutícula y la capa límite adyacente a la hoja. Dado que el mayor intercambio ocurre a nivel de los estomas analizaremos sólo la resistencia a este nivel. Como se verá más adelante el CO2, difunde a través de las mismas resistencias en fase gaseosa que para el vapor de agua y también atraviesa otros componentes en fase líquida. Como punto de partida para nuestra discusión de flujo a nivel de estomas, vamos a considerar la forma unidimensional de la primera Ley de Fick de Difusión, cuya fórmula general es: Flujo = “fuerza”/ resistencia:
xCDJδδ*−= (14)
reemplazando el gradiente de concentración por la diferencia de concentración, así la densidad de flujo depende linealmente de una fuerza. La fuerza puede ser representada por la diferencia de concentración y la distancia que nos interesa es la profundidad del poro estomático δes. La cantidad de partículas por unidad de tiempo hacia o desde la hoja debe ser igual a la cantidad de partículas moviéndose por unidad de tiempo a través del estoma D ΔC/δes veces el área estomática Aes que ocurre en un área foliar dada A. Reconociendo que es importante el efecto restrictivo que el estoma tiene sobre el área disponible para la difusión de moléculas nos da la siguiente relación:
es
eseses
es
es naCDA
ACDJδδ
*** Δ=
Δ= (15)
donde n es el número de estomas por unidad de área foliar y aes es el área promedio por poro estomático; por ejemplo, naes es igual a la fracción de área ocupada por los poros estomáticos, Aes/A. El área disponible para la difusión de vapor de agua cambia bruscamente de Aes al A de la superficie foliar. Este aspecto geométrico podría introducir complicaciones, pero afortunadamente podemos aún usar la forma unidimensional de la primera ley de Fick para describir el movimiento de gases. Finalmente se necesita hacer una corrección: la distancia entre estomas es generalmente 100 μm, la cual es considerablemente menor que la capa límite. Así, la concentración de vapor de agua que rodea a estomas adyacentes tiende a superponerse en la capa límite. Debido a esto, la concentración de vapor de agua varía ligeramente en los planos paralelos a la superficie foliar, pero cambia sustancialmente en la dirección perpendicular a la superficie de la hoja. Sin embargo, la forma en ambos terminales del poro estomático determinan que el poro tenga una profundidad efectiva más grande por lo que se considera el radio efectivo: rades. Debido a que los poros estomáticos son aproximadamente elípticos, no circulares, definimos rades como: π (rades)2 donde rades es el área efectiva del poro. Por lo tanto, definimos la resistencia de estomas para la difusión de gases como sigue:
26 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
es
eses
naDrad
r**δ
= (16)
REFERENCIAS BIBLIOGRÁFICAS. BEARDSELL, M.F., JARVIS P.G. AND DAVIDSON, B. 1972. A null balance diffusion porometer suitable for use with leaves of many shapes. Journal Applied Ecology 9:677-190. Calvin, M. & Benson A. 1948. The path of carbon in photosynthesis. Science 107:476-480. FIELD, C.B., BALL, T. & BERRY, J. 1991. Photosynthesis: principles and field techniques.In R:Pearcy, J. Ehleringer, H. Mooney & P. Rundell (eds) Plant Physiological Ecology, Field Methods and Instrumentation. Chapman and Hall, Chapter 11. GARCÍA DE CORTAZAR V & PS NOBEL (1986) Modelling of PAR interception and productivity of a prickly pear cactus Opuntia ficus indica L., at various spacings. Agronomy Journal 78:80-85. KLUGE M & IP TING (1978) Crassulacean Acid Metabolism Analysis of an Ecological Adaptation. Ecological Studies 30. Springer Verlag. Berlin-Heildelberg-New York, 209 p. MORROW P.A. & SLATYER R.A. 1971. Leaf resistance measurements with diffusion porometers precautions calibration and use. Agriculture Meteorology 8:223-233. NOBEL PS & TL HARTSOCK (1983) Relationships between photosynthetically active radiation nocturnal acid accumulation and CO2 uptake a crassulacean acid metabolism plan, Opuntia ficus indica. Plant Physiology 71:71-75. Nobel, P. 1983. Biophysical Plant Physiological and Ecology, W.H. Freemsn, San Francisco. PEARCY, R.W. SCHULZE, E.D. & ZIMMERMAN R. 1991. Measurement of transpiration and leaf conductance. In: R.W. Pearcy, J. Elheringer, H.A. Mooney and Rundel, P. (eds). Fields Methods and Instrumentation. Plant Physiological Ecology III: 137-160. Salisbury, F.B. & Ross C.W. 1992. Plant Physiology. Chapter 2,3,4 Wadsworth Publishing 682 p. SILVA, H. & ACEVEDO, E. 1995. Eficiencia en el uso del agua en diez taxa de Opuntia, introducidas en la región mediterránea de Chile. Revista Chilena de Historia Natural 68:271-282. SHARKEY, T., IMAI, K., FARQUHAR, G.D. & COWAN, I.R. 1982. A direct confirmation of the standard method of estimating intercellular partial pressure of CO2. Plant Physiology 69:657-659. VON WILLER D & M KLUGE. (1973). Studies on malate fluxes in leaf slices of Bryophyllum daigremontianum. Plant Sciences Letters1 :341-34.
27 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N° 9 RESISTENCIAS Y FLUJOS EN LAS PLANTAS
PROBLEMAS 1. Calcule la resistencia total desde el mesófilo al aire turbulento del vapor de agua. Para ello considere las siguientes resistencias:
r (s m-1) capa límite 25 estoma 300 cutícula 4000 espacio intercelular 15
2. Tenemos 2 hojas (A y B). La hoja A presenta mayor área ocupada por estomas su rst es de 53 s m-1. La hoja B presenta menor área ocupada por estomas su rst es de 600 s m-1. Ambas hojas presentan los mismos valores de rcl, rcut y rei de 20 s m-1, 5000 s m-1 y 30 s m-1, respectivamente. a) Calcule la resistencia de la hoja y la resistencia total en ambas hojas, compare. b) Si la concentración de vapor de agua en la atmósfera es de 18 g m-3 y el flujo de transpiración en ambas hojas es de 100 mg m-2s-1 ¿Cuál debiera ser la concentración del vapor de agua en las paredes del mesófilo en la hoja A y en la hoja B? 3. Si el ΔCH2O entre la superficie de una hoja de 5 cm de largo y el aire turbulento es de 22,9 g m-3 y la velocidad del viento es de 0,1 m s-1. Asuma una temperatura de la hoja y del aire igual a 20°C (D20°C H2O= 2,4x10-5 m2 s-1) a) ¿Cual será el grosor de la capa límite? b) ¿Cual será el flujo de vapor de agua? c) Si la velocidad del viento aumentara a 10 m s-1 ¿cual sería el nuevo grosor de la capa límite? ¿Cual será el flujo de transpiración? ¿Aumento o disminuyo? ¿Por qué? ¿El flujo de fotosíntesis se altera? Si se altera, ¿aumenta o disminuye? ¿Por qué? Explique en detalle. 4. Dadas las siguientes resistencias al vapor de agua, de una hoja que esta transpirando en forma continua 3,4x10-5 kg m-2 s-1, calcule: - La conductancia de la capa límite, estoma y del espacio intercelular. - La concentración del vapor de agua sobre el estoma (A), en la cavidad subestomática (B) y sobre la pared celular de una célula del mesófilo (C). Considere una concentración de vapor de agua en el aire turbulento de 8,65x10-3 kg m-3.
r (s m-1) gs (mm s ) Capa Limite 100 Estoma 500 Espacio Intercelular 40
5. Se tiene una hoja en la oscuridad, los estomas están cerrados. En estas condiciones la hoja transpira a una tasa de 4 x10-4 mg cm-2 s-1 y la diferencia de vapor de agua entre las paredes de las células del mesófilo y el aire turbulento es de 1,01 mg cm-3. La rc es de 2500 s cm-1. a) Calcule la rcl b) La misma hoja se pone a la luz y los estomas se abren. La tasa de transpiración sube a 1x10-3 mg cm-2 s. La diferencia de vapor de agua, rc y la rCL son los mismos que en a), asuma rei = 15s cm-1. Calcule la rs c) ¿Cual sería la tasa de transpiración si la rs calculada en b) se reduce a la mitad?
28 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN Nº 10 EFICIENCIA EN EL USO DE LA RADIACIÓN, TEMPERATURA DE CANOPIA, FLUJO DE
SAVIA Y DENDROMETRIA. En este práctico conoceremos el uso de medidores discretos y medidores continuos de respuestas fisiológicas con distintas aplicaciones, desde estimar el comportamiento de estomas, hasta la aplicación del riego. Estas son la eficiencia en el uso de la radiación, la temperatura de la canopia, sensores de contracción y dilatación de tallos y sensores de flujo de savia. Los niveles de luz tienen una marcada influencia en el crecimiento de la planta, como se ha observado al comparar plantas de sombra con aquéllas creciendo a plena luminosidad. Sólo en los últimos años se ha estado estudiando las relaciones cuantitativas entre el crecimiento del cultivo y los niveles de luz. Inicialmente el crecimiento se evaluó en función del tiempo y actualmente el análisis de crecimiento se basa en su relación con el nivel de luminosidad. Sinclair y Muchow (1999) la definieron como eficiencia en el uso de la radiación (EUR), constituyéndose en uno de los criterios importantes de selección de especies e integrador como la eficiencia de uso de agua (EUA). La EUR, corresponde a la biomasa producida por radiación interceptada (g MJ-1 o µMol de fotones de PAR). La producción de biomasa puede ser modelada como una función lineal de la radiación fotosintéticamente activa interceptada. De manera similar a la EUA, la EUR es relativamente estable dentro de una especie, por ejemplo, el maíz (C4) tiene uno de los valores altos de EUR 3,5 g MJ-1 de PAR interceptado (o 1,75 g MJ-1 de radiación solar interceptada), el sorgo 2,8, para arroz, maravilla y trigo (C3) se han determinado 2,3; 2,2 y 2,8 g MJ-
1.respectivamente. Es posible predecir la producción de materia seca a partir de la radiación interceptada y puede ser definida mediante la siguiente identidad:
MS = RAD x EUR x DA (1) Donde: MS : Rendimiento (o el producto de interés, grano, fruto o materia seca) RAD : Radiación total incidente recibida por el cultivo durante el periodo de tiempo de crecimiento. EUR : Eficiencia en el uso de la radiación o la eficiencia fotosintética total del cultivo (producto comercializable por % radiación interceptada). IC : Indice de cosecha o fracción de materia seca cosechada como rendimiento. (IC es expresado en términos de producción aérea, excluyendo el sistema de raíces). Para estimar el uso de la radiación por los cultivos se usan captores lineales de radiación. La biomasa total de un cultivo, puede ser entendida, en sentido fisiológico, como el resultado de la fotosíntesis neta del cultivo, por lo que puede ser incrementada aumentando cada uno de los factores que la componen. Específicamente EUR puede ser incrementada: • Aumentando la distribución de PAR entre varias hojas. • Aumentando la fotosíntesis bajo condiciones de estrés hídrico por temperatura alta (el estrés hídrico
produce gran disminución en el crecimiento foliar y promueve la senescencia temprana de las hojas). • Reduciendo la fotorrespiración, aumentando la afinidad de la enzima RUBISCO por el CO2 y
disminuyendo la actividad oxigenasa, por selección de genotipos. • Maximizar la intercepción de la luz productiva evitando el exceso de radiación que puede causar
fotoinhibición y marchitez. El establecimiento rápido de la canopia puede ser beneficioso para aumentar la radiación y reducir la evaporación del agua del suelo si las condiciones son favorables al inicio del ciclo. Bajo estrés hídrico puede ser beneficioso tener un bajo IAF (índice de área foliar) y hojas “enrolladas” para evitar el daño producido por exceso de radiación, o bien un bajo contenido de clorofila en las hojas superiores (aquellas que están expuestas directamente al exceso de radiación) en estados tardíos de llenado de granos y si no hay más agua disponible. Otra forma de reducir la radiación captada, bajo estrés, corresponde a hojas pubescentes y cerosas.
En resumen la optimización de la fotosíntesis requiere un balance entre la maximización de la asimilación en los estados críticos de crecimiento y cuando las condiciones son favorables en el campo, evitando los efectos destructivos del exceso de la radiación.
29 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
DETERMINACIÓN DE TEMPERATURA FOLIAR EN LA PLANTA, MEDIANTE TERMOMETRÍA INFRAROJA La temperatura de la cubierta vegetal, particularmente la de las hojas, ha sido propuesta como un indicador de la respuesta de la planta a factores ambientales adversos. La escasez de agua en el suelo, causa que las plantas transpiren a una tasa menor que la demanda evaporativa de la atmósfera, lo que genera un calentamiento de la hoja, debido a que disminuye el efecto refrigerante de la transpiración, afectando el balance energético de la planta, al perder energía sólo por radiación y convección. La radiación emitida por la hoja se caracteriza por estar compuesta principalmente por la parte infrarroja del espectro, y su magnitud depende de la temperatura de la hoja. Esto se expresa en la ley de Stefan-Boltzmann, la cual postula que la cantidad de energía emitida por un cuerpo es función de la cuarta potencia de su temperatura absoluta:
4T·d·eQ = (2) donde Q es la cantidad de radiación emitida, e corresponde a la emisividad, d es una constante y T es la temperatura absoluta. Se puede calcular la temperatura de la planta determinando la radiación infrarroja que emite. La termometría infrarroja se basa en esta teoría, desarrollando sensores capaces de medir la radiación emitida por los cuerpos. Sin embargo, se presentan ciertas reticencias en su uso, debido principalmente al campo visual que abarca el sensor. Con el fin de minimizar los errores, se debe estandarizar y regular el ángulo y la dirección de la medición, de manera de no tener como objetivo parte del horizonte o la fracción de suelo en el campo visual. Aplicaciones. La temperatura de la canopia es un indicador indirecto del estado hídrico de la planta, ya que al encontrarse la planta sometida a condiciones de estrés hídrico disminuye su transpiración debido al cierre parcial de sus estomas, lo cual se traduce en un aumento de la temperatura foliar (Gallardo y Thompson, 2003). Las primeras aproximaciones para encontrar un indicador del estado hídrico en la planta a partir de la temperatura de la canopia, resultaron variar con una gran cantidad de factores ambientales adversos de manera que su interpretación y aplicación en terreno se hacía muy difícil. Con el fin de solucionar este problema, muchos investigadores plantearon diferentes modelos para lograr utilizar la temperatura de canopia con fines prácticos, como la programación de riego. Apreciando que si la planta tiene un buen suministro de hídrico transpirará a su potencial, por lo que estará más fría que el ambiente, y si presenta un estrés hídrico total se suprime la transpiración siendo la temperatura de la hoja mayor a la del ambiente. A partir de esto Jackson et al (1981) desarrollaron un Indice de Estrés Hídrico del cultivo (CWSI). FLUJO DE SAVIA El proceso transpiratorio provoca un flujo de agua líquida desde el suelo hasta los sitios de evaporación en las hojas. El flujo de agua líquida o flujo de savia varia durante el día de manera proporcional a la radiación solar, debido al movimiento de agua que ocurre desde las raíces hasta los estomas donde se pierde como vapor. El flujo de savia disminuye al disminuir la disponibilidad de agua en el suelo, ya que la planta en respuesta a un posible estrés hídrico cierra parcial o totalmente sus estomas, reduciendo así la evaporación de agua desde las hojas y por consecuencia también habrá una disminución en el flujo de savia. En la actualidad existen métodos que permiten mediciones continuas del flujo de savia directamente de la planta bajo condiciones de campo, así las reacciones de la planta frente a diferentes condiciones ambientales pueden ser monitoreadas. Uno de estos métodos es el propuesto por Granier (1987), el cual fue desarrollado para grandes árboles en estudios de ecosistemas forestales y que fue adaptado satisfactoriamente para troncos más pequeños, como los de vides.
30 http://www.sap.uchile.cl
Los sensores de flujo de savia aún están en etapa experimental, por lo tanto es difícil encontrar valores de referencia para especies específicas que permitan detectar cuando su valor es reducido por causa de un déficit hídrico y así poder corregir el manejo del riego. Ginestar et al. (1998) utilizó sensores de flujo de savia para establecer criterios de riego en vides viníferas. Estos autores encontraron que la velocidad de flujo de savia en el tronco sigue estrechamente los cambios que se producen a nivel de la evapotranspiración potencial. En la Figura 1 se observa una clara respuesta del flujo de savia (Fs) a los cambios del dpv durante el día en uva de mesa sometida a diferentes tratamientos (T1, T2, T4) de déficit hídrico, 143 días después del brote. Los valores máximos de Fs se observarion entre las 12:30 y 13:30 h, los que fluctuaron entre los 0,0076 y 0,0108 cm s-1 para T4 y T1 respectivamente. El dpv alcanzó su
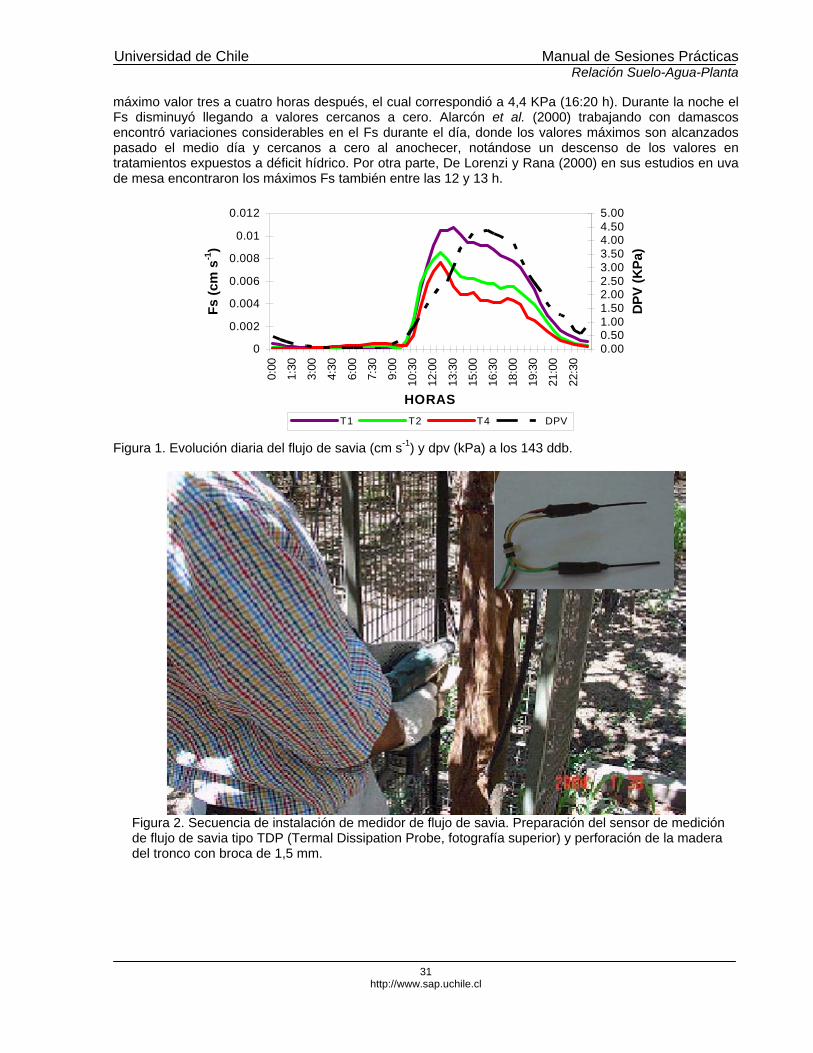
Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
máximo valor tres a cuatro horas después, el cual correspondió a 4,4 KPa (16:20 h). Durante la noche el Fs disminuyó llegando a valores cercanos a cero. Alarcón et al. (2000) trabajando con damascos encontró variaciones considerables en el Fs durante el día, donde los valores máximos son alcanzados pasado el medio día y cercanos a cero al anochecer, notándose un descenso de los valores en tratamientos expuestos a déficit hídrico. Por otra parte, De Lorenzi y Rana (2000) en sus estudios en uva de mesa encontraron los máximos Fs también entre las 12 y 13 h.
0
0.002
0.004
0.006
0.008
0.01
0.012
0:00
1:30
3:00
4:30
6:00
7:30
9:00
10:3
0
12:0
0
13:3
0
15:0
0
16:3
0
18:0
0
19:3
0
21:0
0
22:3
0
HORAS
Fs (c
m s
-1)
0.000.501.001.502.002.503.003.504.004.505.00
DPV
(KPa
)
T1 T2 T4 DPV
Figura 1. Evolución diaria del flujo de savia (cm s-1) y dpv (kPa) a los 143 ddb.
Figura 2. Secuencia de instalación de medidor de flujo de savia. Preparación del sensor de medición de flujo de savia tipo TDP (Termal Dissipation Probe, fotografía superior) y perforación de la madera del tronco con broca de 1,5 mm.
31 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
a
b
Figura 3. Secuencia de instalación de medidor de flujo de savia. a) inserción del sensor en la madera. b) Protección del sensor de las variaciones externas de temperatura.
32 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
DENDROMETRÍA: DETERMINACIÓN DE LA VARIACIÓN EN EL DIÁMETRO DEL TRONCO. El diámetro del tronco es un indicador sensible del estado hídrico de la planta, presentando variaciones a lo largo del día. Estas variaciones se presentan por la entrada y salida de agua desde los tejidos vivos como el floema y el cambium vascular (Genard et al., 2001; Fereres y Goldhamer, 2003). Los máximos valores se alcanzan precisamente antes del amanecer y los valores mínimos de diámetro de tronco se alcanzan durante la tarde cuando la transpiración es más elevada. A medida que disminuye la disponibilidad de agua en el suelo como consecuencia de la extracción que realiza la planta, la amplitud de la contracción diurna aumenta y el crecimiento o expansión nocturna decrece, pudiendo llegar a ser nulo o incluso negativo. La diferencia entre el máximo y el mínimo condiciona la contracción máxima diaria (CMD), parámetro que muestra el incremento del déficit hídrico en diversas especies frutales como manzano, durazneros y uva. Para desarrollar este tipo de mediciones es necesario un instrumento que registre continuamente el nivel de agua en la planta, o algún aspecto del funcionamiento que esté directamente relacionado al nivel de agua en la planta. Existen en el mercado diversos sistemas que miden las variaciones del diámetro de troncos como los de Ivens, Dynamac y Phytec. Asociado a esto, encontramos una de las ventajas en las mediciones de diámetro de tronco que es proporcionar registros continuos que permitan calcular o dimensionar la tasa de crecimiento del tronco para una temporada. Descripción del sistema Este instrumento es un captor de desplazamiento lineal que es colocados en el tronco y se conecta mediante una antena a una central de adquisición de datos que permite registrar la variación en forma continua, a lo largo de toda la temporada de crecimiento de la planta. El equipo va acompañado de un programa de PC para su configuración, mediante el cual se puede recoger los datos y visualizar, de forma gráfica, los valores de las lecturas. El sistema puede funcionar a batería, con o sin panel solar. Este sistema llamado bioprogramador permite una automatización del riego en caso de sistemas presurizados.
Figura 4. Dendrómetro electrónico y sistema de adquisición y envío de datos.
33 http://www.sap.uchile.cl
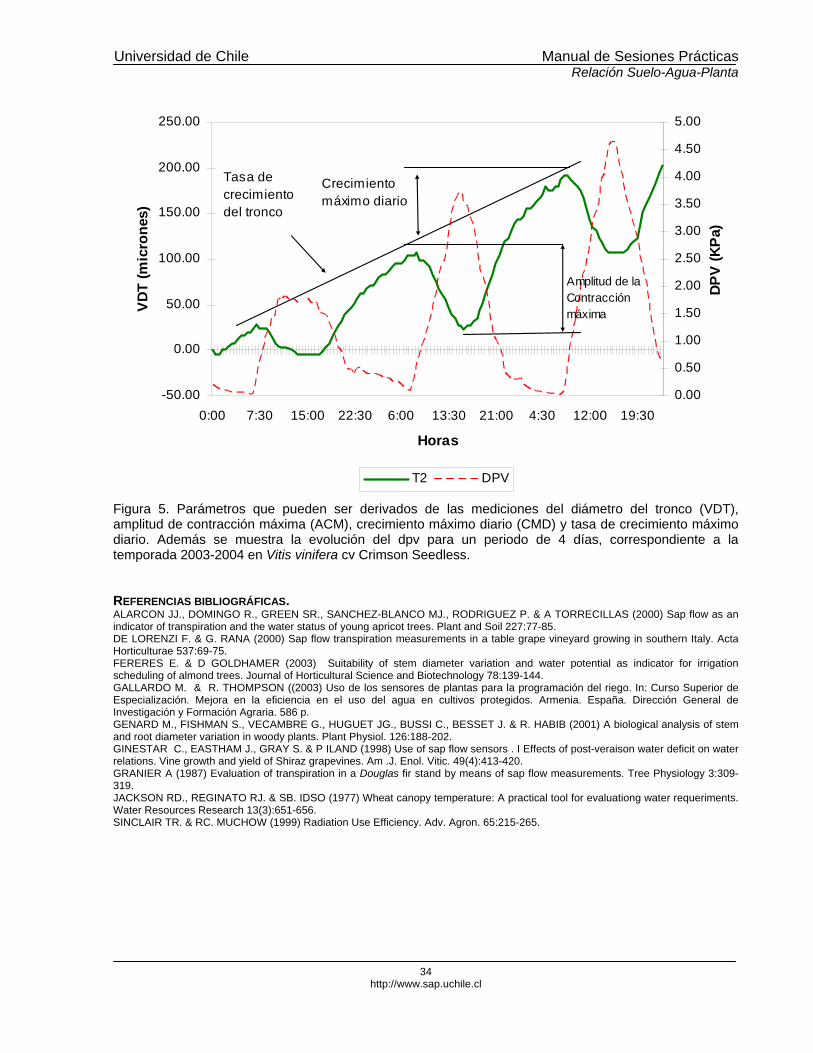
Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
-50.00
0.00
50.00
100.00
150.00
200.00
250.00
0:00 7:30 15:00 22:30 6:00 13:30 21:00 4:30 12:00 19:30
Horas
VDT
(mic
rone
s)
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4.50
5.00
DPV
(KP
a)
T2 DPV
Amplitud de la Contracción máxima
Crecimiento máximo diario
Tasa de crecimiento del tronco
Figura 5. Parámetros que pueden ser derivados de las mediciones del diámetro del tronco (VDT), amplitud de contracción máxima (ACM), crecimiento máximo diario (CMD) y tasa de crecimiento máximo diario. Además se muestra la evolución del dpv para un periodo de 4 días, correspondiente a la temporada 2003-2004 en Vitis vinifera cv Crimson Seedless. REFERENCIAS BIBLIOGRÁFICAS. ALARCON JJ., DOMINGO R., GREEN SR., SANCHEZ-BLANCO MJ., RODRIGUEZ P. & A TORRECILLAS (2000) Sap flow as an indicator of transpiration and the water status of young apricot trees. Plant and Soil 227:77-85. DE LORENZI F. & G. RANA (2000) Sap flow transpiration measurements in a table grape vineyard growing in southern Italy. Acta Horticulturae 537:69-75. FERERES E. & D GOLDHAMER (2003) Suitability of stem diameter variation and water potential as indicator for irrigation scheduling of almond trees. Journal of Horticultural Science and Biotechnology 78:139-144. GALLARDO M. & R. THOMPSON ((2003) Uso de los sensores de plantas para la programación del riego. In: Curso Superior de Especialización. Mejora en la eficiencia en el uso del agua en cultivos protegidos. Armenia. España. Dirección General de Investigación y Formación Agraria. 586 p. GENARD M., FISHMAN S., VECAMBRE G., HUGUET JG., BUSSI C., BESSET J. & R. HABIB (2001) A biological analysis of stem and root diameter variation in woody plants. Plant Physiol. 126:188-202. GINESTAR C., EASTHAM J., GRAY S. & P ILAND (1998) Use of sap flow sensors . I Effects of post-veraison water deficit on water relations. Vine growth and yield of Shiraz grapevines. Am .J. Enol. Vitic. 49(4):413-420. GRANIER A (1987) Evaluation of transpiration in a Douglas fir stand by means of sap flow measurements. Tree Physiology 3:309-319. JACKSON RD., REGINATO RJ. & SB. IDSO (1977) Wheat canopy temperature: A practical tool for evaluationg water requeriments. Water Resources Research 13(3):651-656. SINCLAIR TR. & RC. MUCHOW (1999) Radiation Use Efficiency. Adv. Agron. 65:215-265.
34 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
SESIÓN N°11 NECESIDADES DE AGUA DE LOS CULTIVOS
PROBLEMAS 1. Determine las necesidades de agua para cada una de las fases del período vegetativo de una
variedad de maíz, sembrada el 10 de octubre en Santiago y cuya información fenológica es:
Fase fenológica Duración (días) Kc (promedio de la fase)
Fase inicial 20 0,6 Fase de crecimiento activo 40 0,8 Fase de pleno desarrollo 50 1,5 Fase de maduración 30 0,7 TOTAL 140
Los datos evaporimétricos y los coeficientes kp determinados de acuerdo a las condiciones climáticas son:
S O N D E F M Ev (mm) 2,3 4,0 4,5 6,7 7,0 6,0 4,0 Kp 0,8 0,70 0,70 0,70 0,70 0,70 0,70
Exprese los resultados en m3ha-1 (considere que 1 mm = 10 m3 ha-1) 2. Un cultivo de trigo tiene una profundidad de enraizamiento de 1 m en un suelo homogéneo que a 1,5
m tiene una estrata impermeable. La CC y PMP de este suelo es 0,28 g·g-1 y 0,13 g·g-1 respectivamente, y la Da de 1,25 g cm-3. Si se riega con un UR de 25% y su ETc es de 5 mm día-1. a) ¿En cuántos días se regará? b) ¿Qué altura de agua se aplicará? c) ¿Qué volumen de agua se aplicará en una ha?
3. De acuerdo a los datos presentados, calcule el número de riegos y las fechas de riego (grafique).
Considere que la temporada de riego comienza en septiembre y el cultivo se cosecha el 15 de enero. CC = 30% ETc Septiembre: 4 mm día-1 PMP = 18% ETc Octubre: 5 mm día-1 Da = 1,4 g cm-3 ETc Noviembre: 6 mm día-1 UR = 50% ETc Diciembre: 3 mm día-1 Profundidad de raíces 50 cm ETc Enero: 0
4. Se tienen plantas de poroto cuya ETc es de 1 mm día-1 en una maceta de 30 cm de diámetro y 50 cm
de profundidad, llena de suelo homogéneo con un contenido de humedad de 0,4 g·g-1. a) ¿En cuantos días el contenido de humedad del suelo llegará a 0,25g·g-1? b) A partir de este ultimo contenido de humedad ¿Qué volumen de agua se deberá agregar para que el suelo quede a CC (0,5 g·g-1)? Da =0,9 g cm-3.
5. Un cultivo de trigo en floración crece en un suelo franco con una profundidad de arraigamiento de 95
cm. El contenido de humedad del suelo a CC y PMP es de 0,3 g·g y 0,15 g·g-1, respectivamente. La Da del suelo es 1,2 g cm-3. La evapotranspiración promedio diaria para un cultivo de trigo en floración es de 5 mm día-1. a) Si el contenido actual de humedad es de 0,3 cm3 cm-3 desde la superficie hasta una profundidad de 120 cm ¿En cuantos días se regará el cultivo? b) Si el UR es de 40% ¿En cuantos días se volverá a regar el cultivo? c) Haga un gráfico altura de agua v/s tiempo en que aparezcan los riegos de a y b.
6. De acuerdo a la siguiente información, calcule el número y las fechas de riego para un cultivo de maíz (profundidad de arraigamiento de 1,2 m). Considere que la temporada de riego comienza el 10 de
35 http://www.sap.uchile.cl

Universidad de Chile Manual de Sesiones Prácticas Relación Suelo-Agua-Planta
Septiembre, el cultivo se siembra el 1 de Octubre y se cosecha el 15 de Abril. CC= 35%; PMP= 18%; Da= 1,35 g cm-3; UR= 50%,
ETc Septiembre 4 mm día-1 ETc Octubre 5 mm día-1 ETc Noviembre 6 mm día-1 ETc Diciembre 6,5 mm día-1 ETc Enero 6 mm día-1 ETc Febrero 5 mm día-1 ETc Marzo 4 mm día-1 ETc Abril 0 mm día-1
7. Un cultivo de maíz en estado de llenado de grano (enero y febrero) tiene una ETc promedio de 8 y 10
mm día-1, respectivamente. En la mañana del día 20 de enero el cultivo de maíz, que tiene 1 m de profundidad de arraigamiento, inicia su llenado de grano el que se prolongará por 30 días. En dicha fecha el suelo (franco y homogéneo) esta a CC con 0,38 g·g-1 (Da = 1,5 g cm-3). a) ¿Qué contenido de humedad tiene dicho suelo cuando el U.R. es de 30%? ¿Qué día se regará? ¿Qué altura de agua se debe aplicar en el riego? b) Si no se regara, ¿qué día la humedad del suelo llegará a PMP (0,15 g·g-1)?
8. Durante la temporada 97/98 se realizó un ensayo de quínoa en la Estación Experimental Antumapu.
La profundidad de enraizamiento de este cultivo fue de 60 cm. Las características del suelo fueron:
Profundidad CC (%) PMP (%) Da (g cm-3) 0 - 15 23,43 15,00 1,31 15 - 45 19,23 13,31 1,23 45 - 75 13,30 10,89 1,36
La ETc mensual de la quinoa fue: Septiembre: 13,4 mm mes-1 Octubre: 23,5 mm mes-1 Noviembre: 42,4 mm mes-1 Diciembre: 58,3 mm mes-1 Enero: 65,5 mm mes-1 El suelo estaba a CC el 1 de septiembre, no hubo lluvia entre el 1 de Septiembre y el 31 de Enero y el U.R. es de 50%, conteste: a) ¿Qué contenido de humedad tiene el suelo cuando el umbral de riego alcanza al 50%? b) ¿Cuántos riegos se dieron a la quínoa entre el 1 de Septiembre de 1997 y el día 31 de Enero de 1998? c) ¿Cada cuantos días se realizaron los riegos para dicho período? d) ¿Qué altura de agua se debió aplicar en cada riego? e) ¿Qué volumen de agua se aplicó durante todo el período en 1 hectárea de quínoa? f) Grafique la variación del contenido de agua versus tiempo e indique con una flecha los riegos hechos durante el período
36 http://www.sap.uchile.cl


















