
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE...
89
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO TROPICAL (BH-T) DE COLOMBIA 1 ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO TROPICAL (BH-T) DE COLOMBIA JENY ALEJANDRA CASTRILLÓN ARBOLEDA LORENA LIZETH PINZÓN CASTILLO UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS FACULTAD DEL MEDIO AMBIENTE Y RECURSOS NATURALES PROYECTO CURRICULAR DE INGENIERÍA FORESTAL 2015
Transcript of ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE...

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
1
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE
BOSQUE HÚMEDO TROPICAL (BH-T) DE COLOMBIA
JENY ALEJANDRA CASTRILLÓN ARBOLEDA
LORENA LIZETH PINZÓN CASTILLO
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
FACULTAD DEL MEDIO AMBIENTE Y RECURSOS NATURALES
PROYECTO CURRICULAR DE INGENIERÍA FORESTAL
2015

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
2
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE
BOSQUE HÚMEDO TROPICAL (BH-T) DE COLOMBIA
AUTORES:
Jeny Alejandra Castrillón Arboleda
Lorena Lizeth Pinzón Castillo
Trabajo de grado para optar a el título profesional de ingeniera forestal bajo la
modalidad de investigación
DIRECTORA: Esperanza Nancy Pulido Rodríguez
JURADOS: Ingrid Amórtegui
Mario León
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
FACULTAD DEL MEDIO AMBIENTE Y RECURSOS NATURALES
PROYECTO CURRICULAR DE INGENIERÍA FORESTAL
2015

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
3
TABLA DE CONTENIDO
RESUMEN ........................................................................................................................ 8
ABSTRACT ...................................................................................................................... 9
INTRODUCCIÓN .......................................................................................................... 10
PLANTEAMIENTO DEL PROBLEMA ....................................................................... 13
JUSTIFICACIÓN ........................................................................................................... 15
OBJETIVOS ................................................................................................................... 16
OBJETIVO GENERAL .............................................................................................. 16
OBJETIVOS ESPECÍFICOS ...................................................................................... 16
MARCO DE REFERENCIA .......................................................................................... 17
MARCO TEÓRICO .................................................................................................... 17
ESTADO DEL ARTE ................................................................................................. 23
METODOLOGÍA ........................................................................................................... 27
ÁREA DE ESTUDIO .................................................................................................. 27
ESPECIES EVALUADAS .......................................................................................... 28
VARIABLES EVALUADAS ..................................................................................... 29
ELABORACIÓN DE LAMINILLAS HISTOLÓGICAS .......................................... 31
MACERACIÓN/DISOCIACIÓN ............................................................................... 34
DESCRIPCIÓN ANATÓMICA ................................................................................. 38
MEDICIÓN DE ELEMENTOS .................................................................................. 38
PROCESAMIENTO Y ANÁLISIS DE DATOS ........................................................ 43
RESULTADOS ............................................................................................................... 44
RASGOS ANATÓMICOS CUALITATIVOS DE LA MADERA ............................ 44
RASGOS ANATÓMICOS CUANTITATIVOS DE LA MADERA ......................... 57
ANÁLISIS DE RESULTADOS ..................................................................................... 68
CONCLUSIONES .......................................................................................................... 80
RECOMENDACIONES ................................................................................................. 81
AGRADECIMIENTOS .................................................................................................. 82
BIBLIOGRAFÍA ............................................................................................................ 84

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
4
LISTA DE TABLAS
Tabla 1. Especies a evaluar en el estudio. Estas especies han sido colectadas para la
realización de los proyectos CAR USAID, y se encuentran registradas en las
colecciones biológicas de la Universidad Distrital Xiloteca JaLR UD y Herbario
Forestal UDBC ................................................................................................................ 29
Tabla 2. Variables cuantitativas del estudio. ................................................................... 30
Tabla 3. Variables cualitativas del estudio (Vasos, fibras, parénquima, radios y otros.) 30
Tabla 4. Características cuantitativas de los vasos, fibras y radios de las 25 especies
estudiadas (estadísticos de media, desviación y coeficiente de variación) ..................... 57
Tabla 5. Índices de vulnerabilidad y mesomorfía para las especies objeto de estudio. .. 63
Tabla 6. Matriz de correlación de rasgos anatómicos cuantitativos................................ 65
Tabla 7. Estudios revisados y nomenclatura empleada en el presente estudio. .............. 68

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
5
LISTA DE FIGURAS
Figura 1. Mapa Conceptual; Metodología seguida en el proceso de Investigación. ....... 27
Figura 2. Área de estudio (Cuenca del Río Atrato). Municipios de Bojaya, Vigía del
fuerte y San Miguel. Fuente. OCHA, 2003. .................................................................... 28
Figura 3. Realización de cortes histológicos transversal, tangencial y radial en
micrótomo de rotación Leica RM 2255. ......................................................................... 32
Figura 4. Proceso de tinción, deshidratación y aclaramiento de los cortes transversales,
tangenciales y radiales de las muestras. a. Reactivos empleados. b. Tinción con
safranina. c y d. Lavado con alcohol. e. Deshidratación de las especies. f. Ubicación de
los cortes. ........................................................................................................................ 33
Figura 5. Proceso de montaje de laminillas histológicas (Acomodación de cortes,
adición de Eukitt y colocación de cubreobjetos. a. Ubicación de los cortes. b.
Aplicación de Eukitt. c. Ubicación de la laminilla. y d. Montaje final. .......................... 34
Figura 6. Procedimiento para maceración de la madera (a y b. corte de astillas, c y d.
inmersión en reactivos, e. Reactivos empleados y f. calentamiento de los tejidos). ....... 35
Figura 7. Proceso de tinción del material disociado (a. Tinción, b. limpieza con alcohol
al 90% c, d y e. almacenamiento). .................................................................................. 36
Figura 8. Proceso para montaje del material disociado (a. y b. disposición de material
macerado, c. Disposición en láminas y d. colocación de cubrobjetos). ......................... 37
Figura 9. Rotulado preliminar de laminillas (Trazabilidad)............................................ 37
Figura 10. Laminillas etiquetadas en colección biológica de la Xiloteca JaLR UD. ...... 38
Figura 11. Medición de diámetro de vaso. Ecclinusa ramiflora Mart. ........................... 39
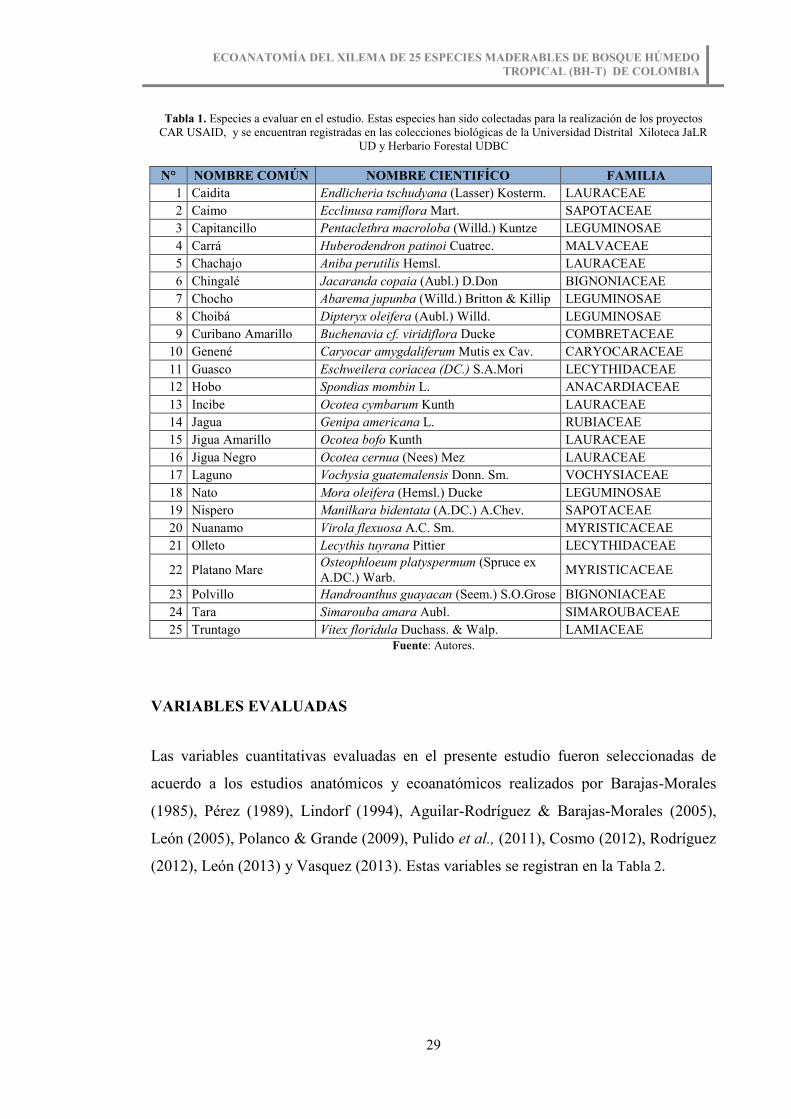
Figura 12. Medición de número de vasos/mm2. Lecythis tuyrana Pittier. ..................... 40
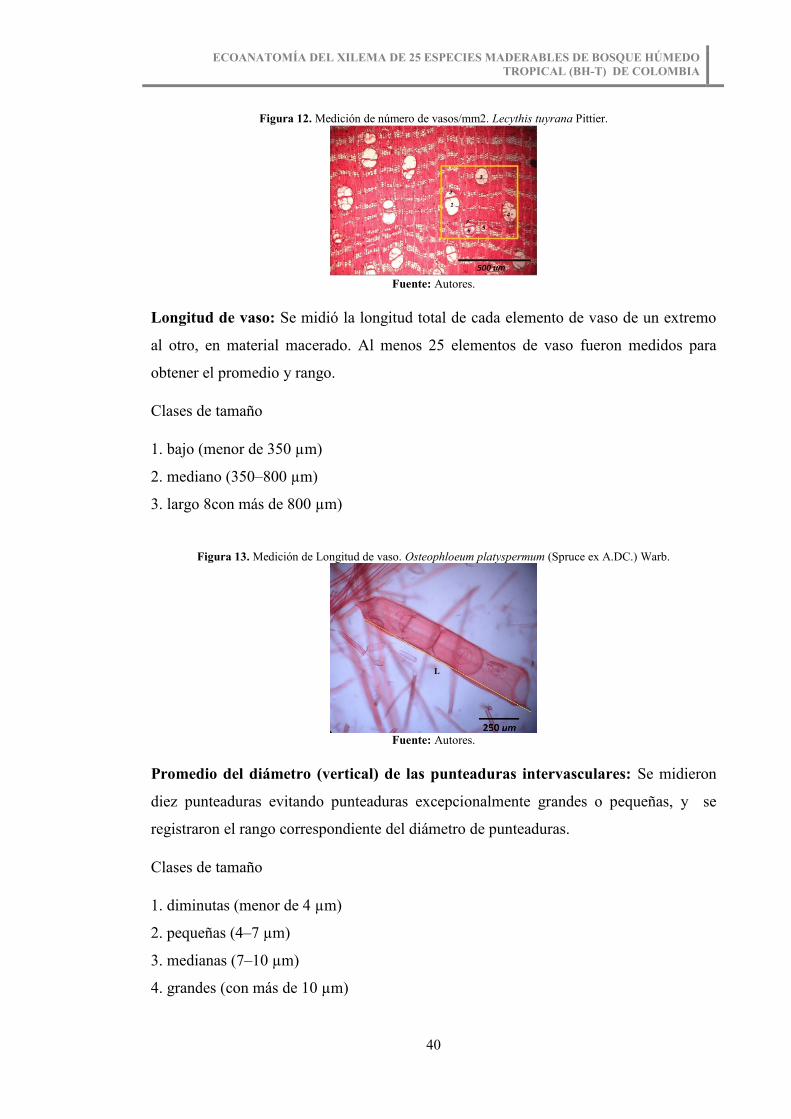
Figura 13. Medición de Longitud de vaso. Osteophloeum platyspermum (Spruce ex
A.DC.) Warb. .................................................................................................................. 40
Figura 14. Medición del diámetro de punteadura intervascular. Endlicheria tschudyana
(Lasser) Kosterm. ............................................................................................................ 41
Figura 15. Medición de Longitud de fibra. Aniba perutilis Hemsl. ................................ 41
Figura 16. Medición de grosor de la pared celular de la fibra.Mora oleifera (Hemsl.)
Ducke .............................................................................................................................. 42
Figura 17. Medición altura de radios. Huberodendron patinoi Cuatrec. ........................ 42
Figura 18. Medición de número de radios por mm lineal. Ecclinusa ramiflora Mart. ... 43
Figura 19. Mapa conceptual. Proceso para el análisis de los datos obtenidos para
variables cualitativas y cuantitativas. .............................................................................. 43
Figura 20. Tipo de porosidad. a. Porosidad semicircular (Genipa americana L.) b.
Porosidad difusa (Virola flexuosa A.C. Sm.) .................................................................. 44
Figura 21. Disposición de poros. a. Sin patrón de disposición (Vochysia
guatemalensis).b. Disposición dendrítica (Buchenavia cf. viridiflora), c. Disposición
diagonal (Ocotea bofo). ................................................................................................... 45
Figura 22. Agrupación de poros. a. Poros solitarios (Genipa americana), b. Múltiplos
radiales cortos (Virola flexuosa) ..................................................................................... 45

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
6
Figura 23. Forma de los poros. a. Poro ovalado (Ecclinusa ramiflora), b. Poro redondo
(Spondias mombin). ........................................................................................................ 46
Figura 24. Tipos de platinas de perforación. a. Escaleriforme (Endlicheria tschudyana),
b. Reticulada (Virola flexuosa), c. Simple (Ocotea cernua), d. Simple (Genipa
americana). ..................................................................................................................... 46
Figura 25. Arreglo de las punteaduras intervasculares. a. Punteaduras areoladas opuestas
(Pentaclethra macrolaba), b. Punteaduras areoladas alternas (Ocotea cymbarum) ....... 47
Figura 26. Tipo de punteaduras intervasculares. a. Punteadura intervascular poligonal
(Eschweilera coriacea), b Punteadura intervascular ovalada (Vochysia guatemalensis),
c. Punteadura intervascular redonda (Handroanthus guayacan). a, b y c. Abertura
incluida. ........................................................................................................................... 47
Figura 27. Tamaño de las punteaduras intervasculares. Punteaduras intervasculares
grandes (Ocotea bofo), b. Punteaduras intervasculares medianas (Ocotea cernua), c.
Punteaduras intervasculares pequeñas (Simarouba amara), d. Punteaduras
intervasculares diminutas (Jacaranda copaia). .............................................................. 48
Figura 28. Punteaduras ornadas. a. Punteaduras intervasculares ornadas (Caryocar
amygdaliferum), b. Punteaduras intervasculares NO ornadas (Spondias mombin). ....... 49
Figura 29. Inclusiones a. Gomas (Vochysia guatemalensis), b. Tílides (Eschweilera
coriacea) c. Esclerotilides (Ecclinusa ramiflora) ........................................................... 49
Figura 30. Grosor de la pared celular. a. Pared celular gruesa (Buchenavia cf.
viridiflora), b. Pared celular mediana (Endlicheria tschudyana), Pared celular delgada
(Spondias mombin). ........................................................................................................ 50
Figura 31. Engrosamientos espiralados en fibras a. presentes (Buchenavia cf.
viridiflora), b. ausentes (Huberodendron patinoi) .......................................................... 51
Figura 32. Presencia de fibras septadas. a. Fibras septadas (Ocotea bofo), b. Fibras no
septadas (Simarouba amara)........................................................................................... 51
Figura 33. Tipos de parénquima. a. Parénquima apotraqueal difuso y en agregados
(Genipa americana), b. Parénquima aliforme confluente y aliforme de ala ancha y
corta,(Mora oleifera), c. Parénquima en bandas finas (Manilkara bidentata), d.
Parénquima reticulado (Lecythis tuyrana), e. Parénquima de ala fina y extendida
(Jacaranda copaia), f. Parénquima apotraqueal difuso y paratraqueal unilateral
(Buchenavia cf. Viridiflora). ........................................................................................... 52
Figura 34. Ancho de los radios. a. Radios biseriados (Mora oleifera), b. Radios
multiseriados (Vochysia guatemalensis), c. Radios uniseriados (Abarema jupunba), d.
Radios de 1 a 3 células (Eschweilera coriacea). ............................................................ 53
Figura 35. Tipo de radios. a. Radio tipo I (Vitex floridula), b. Radio tipo III
(Osteophloeum platyspermum), c. Radio tipo VI (Manilkara bidentata). ...................... 54
Figura 36. Modificaciones en células radiales a. Células radiales perforadas
(Huberodendron patinoi), b. Células tipo baldosa (Ecclinusa ramiflora). ..................... 54
Figura 37. Tipo de punteaduras radiovasculares. a. Similares a las intervasculares
(Simarouba amara), b. de dos tamaños (Virola flexuosa), c. Con borde reducido
aparentemente simple rendondeadas a angulares (Spondias mombin), d. Con borde muy
reducido con abertura horizontal y vertical (Aniba perutilis) ......................................... 55
Figura 38. Estructuras estratificadas. a. Radios estratificados (Handroanthus guayacan),
b. Fibras, radios y parénquima estratificado (Simarouba amara). .................................. 55

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
7
Figura 39. Tubo laticifero. (Osteophloeum platyspermum). ........................................... 56
Figura 40. Inclusiones minerales. a. Cristales prismáticos (Caryocar amygdaliferum), b.
Sílice (Eschweilera coriacea). ........................................................................................ 56
Figura 41. Biometría diámetro de vasos. a. Diagrama de barras de diámetro de vasos por
clases de tamaño, b. Boxplot de diámetro de vasos. ....................................................... 58
Figura 42. Biometría longitud de vasos. a. Diagrama de barras de longitud de vasos por
clases de tamaño, b. Boxplot de longitud de vasos. ........................................................ 59
Figura 43. Biometría frecuencia de vasos. a. Diagrama de barras de frecuencia de vasos,
b. Boxplot de frecuencia de vasos. .................................................................................. 59
Figura 44. Biometría diámetro de las punteaduras intervasculares. a. Diagrama de barras
de diámetro de punteaduras intervasculares por clases de tamaño, b. Boxplot de
diámetro de punteaduras intervasculares. ....................................................................... 60
Figura 45. Biometría de grosor de fibras. a. Diagrama de barras de grosor de fibras por
clases de tamaño, b. Boxplot de grosor de fibras. ........................................................... 60
Figura 46. Biometría de longitud de fibras. a. Diagrama de barras de longitud de fibras
por clases de tamaño, b. Boxplot de longitud de fibras. ................................................. 61
Figura 47. Biometría de altura de radios. a. Diagrama de barras de altura de radios por
clases de tamaño, b. Boxplot de altura de radios. ........................................................... 62
Figura 48. Biometría de frecuencia de radios. a. Diagrama de barras de frecuencia de
radios, b. Boxplot de frecuencia de radios. ..................................................................... 62
Figura 49. Índice de vulnerabilidad. a. Histograma de índice de vulnerabilidad, b.
Boxplot de índice de vulnerabilidad. .............................................................................. 64
Figura 50. Índice de mesomorfía. a. Histograma de índice de mesormorfía, b. Boxplot
de índice de mesormorfía. ............................................................................................... 65
Figura 51. Análisis de componentes principales para variables cuantitativas. ............... 66
Figura 52. Diagrama de barras para diámetro de vasos, en diferentes ecosistemas. ....... 69
Figura 53. Diagrama de barras para longitud de vaso, en diferentes ecosistemas. ......... 70
Figura 54. Diagrama de barras para poros/mm2 para diferentes ecosistemas. ............... 71
Figura 55. Diagrama de barras para el diámetro de las punteaduras intervasculares para
diferentes ecosistemas. .................................................................................................... 72
Figura 56. Diagrama de barras para la longitud de la fibra en diferentes ecosistemas. .. 73
Figura 57. Diagrama de barras para la altura del radio en diferentes ecosistemas. ........ 74
Figura 58. Diagrama de barras para radios/mm en diferentes ecosistemas. ................... 75
Figura 59. Variación de los elementos anatómicos. a. Variación de acuerdo a la
precipitación. b. Variación de acuerdo a la temperatura. ................................................ 78
Figura 60. Variación de la longitud de la fibra de acuerdo a la altitud ........................... 79

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
8
RESUMEN
La ecoanatomía se fundamenta en la correlación que existe entre los factores climáticos
y algunas características anatómicas, las cuales pueden ser consideradas como
estrategias adaptativas. Para conocer la tendencia en la variación de los elementos
anatómicos de la madera de acuerdo a factores climáticos en un bosque húmedo
tropical de Colombia, se estudiaron 25 especies. Los resultados sugieren que los rasgos
anatómicos cuantitativos que más se ven afectadas por las condiciones ambientales
(precipitación, temperatura, altitud) de una zona son: diámetro de vaso, longitud del
elemento vascular, frecuencia de poros y longitud de fibra. Por el contrario, los rasgos
anatómicos que no presentaron una relación evidente con las condiciones ambientales
fueron el diámetro de la punteadura intervascular, el grosor de la pared celular de la
fibra y la altura y frecuencia de los radios. Por otra parte se determinó mediante los
índices de vulnerabilidad y mesomorfía que las especies poseen un comportamiento
mesomórfico y por lo tanto un sistema de conducción orientado hacia la eficiencia en el
transporte del agua, con vulnerabilidad al estrés hídrico.
Palabras clave:
Ecoanatomía, madera, factores ambientales, índices de vulnerabilidad y mesomorfía,
bosque húmedo tropical.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
9
ABSTRACT
Ecoanatomy is based on the relationship between climatic factors and some anatomical
features, which can be considered as adaptive strategies. To know variation trends of
wood anatomical elements according to weather in a tropical rain forest of Colombia, 25
species were studied. The results suggest that the quantitative anatomical features most
affected by environmental conditions (precipitation, temperature, altitude) of an area
are: vessel diameter, vessel element length, pores frequency and fiber length. By
contrast, the anatomical features that did not present an obvious relationship with the
environmental conditions were intervascular bordered pits diameter, fiber wall thickness
and rays height and frequency. On the other hand, vulnerability and mesomorphy index
indicate that species present a mesomorphic behavior and therefore a conductive system
oriented to efficiency on water transport with vulnerability to water stress.
Key words:
Ecoanatomy, wood, environmental factors, mesomorphy and vulnerability index,
tropical rain forest.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
10
INTRODUCCIÓN
La madera es el producto de la división celular del cambium vascular, tejido
meristemático responsable del crecimiento secundario, se encarga de la producción de
xilema hacia el interior y floema hacia el exterior del tronco. Este meristemo lateral
genera células iniciales fusiformes y células iniciales radiales que mediante su división
dan origen a los sistemas celulares axial y radial respectivamente (Carrillo et al., 2013).
El tejido xilemático originado a partir del cambium, está constituido por un conjunto de
células o elementos leñosos que han pasado a través de diferentes etapas de desarrollo;
desde la división celular hasta la diferenciación y maduración de células (Webster &
McKetchine, 1980). Al terminar el proceso, la madera se constituye de los siguientes
tipos de células: vasos, traqueidas, parénquima y fibras, las cuales poseen una pared
celular compuesta de celulosa, hemicelulosa y lignina (Montaño et al., 2011), y un
componente químico secundario formado por diferentes tipos de extractos, azucares y
otras sustancias orgánicas e inorgánicas (Zobel & Van Buijtenen, 1989).
Durante su formación las células son afectadas por muchos factores, tales como el sitio,
condiciones ecológicas, manejo, genética y edad de los árboles. Los factores de sitio
interactúan e influyen directa o indirectamente en los procesos de desarrollo de las
plantas, ejerciendo una presión de selección a un mayor o menor grado. En
consecuencia, los factores ambientales también tienen influencia en el patrón de
distribución de la flora en diferentes ecosistemas, debido a que influyen en la capacidad
competitiva de las especies (Cosmo, 2012). Según Walter (1986), en un área geográfica
con características relativamente uniformes, con ligeras diferencias en las condiciones
ambientales y el suministro de agua se observan efectos en el suelo y la vegetación que
alteran todo el ecosistema. Por lo tanto, se puede suponer que la vegetación en
equilibrio con el ambiente, representa una expresión integrada de los efectos de tales
factores ambientales y su componente genético por lo que se pueden determinar
cambios cualitativos o cuantitativos en la vegetación, en la morfología de sus especies
vegetales y en la anatomía de sus tejidos vegetales.
Por su parte, Carrillo et al. (2013) mencionan que los procesos mediante los cuales se
producen el crecimiento y desarrollo celular de los elementos anatómicos que

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
11
constituyen la madera, están influenciados por la una amplia variedad de genes que
modulan la producción de enzimas y proteínas, para finalmente otorgar las dimensiones
y características a cada uno de dichos elementos anatómicos; por lo tanto, los genes que
participan en el crecimiento y desarrollo de las células, inciden directamente sobre las
características físicas, químicas y anatómicas de la madera.
La composición y arreglo de estas células, le otorgan a la madera características físicas
y mecánicas que hacen posible su uso como materia prima para construcción,
generación de energía, fibras para la producción de pulpa y papel, paneles, tableros,
biocombustibles y biomateriales (Carrillo et al., 2013). Por otra parte, el estudio de la
variación anatómica en especies de importancia económica proporciona información
útil que puede emplearse como registro de las condiciones ambientales de las zonas en
las que se desarrollan los individuos y son fundamentales ya que proporcionan las bases
para un manejo forestal adecuado que favorezca la restauración regional de los
ecosistemas y una mejor calidad de la madera. (Chávez-Romero et al., 2010).
Debido a la importancia de la incidencia de los factores ambientales en el desarrollo de
la madera, se han realizado varios estudios en los que se evalúa la relación entre la
anatomía de la madera (diámetro de poros, frecuencia de vasos, agrupamiento de los
vasos, longitud de los elementos vasculares y tamaño de punteaduras intervasculares) y
factores climáticos (humedad, temperatura, estacionalidad, altitud y distribución
geográfica); en los que se encontró que las características anatómicas mencionadas si se
encuentra influenciadas por los factores climáticos (León, 2005; Prida et al., 2006;
Moya & Tomazello, 2008; Polanco & Grande, 2009; Meena & Gupta, 2014). Por
ejemplo, Castro et al., (1998) y Montaño et al., (2011) encontraron que los vasos con
diámetros y longitudes mayores se encuentran en localidades húmedas y cálidas,
mientras que los diámetros y longitudes menores se hallan en localidades frías y secas, y
que los elementos que más influyen en la anatomía de la madera son la altitud, la
latitud y el clima.
El objetivo de este trabajo es determinar la influencia de los factores climáticos del
bosque húmedo tropical, en el desarrollo de las características anatómicas del xilema de
25 especies a partir de las descripciones anatómicas, la medición de características

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
12
cuantitativas como diámetro de poros, longitud de fibras, diámetro de las punteaduras,
entre otros y evaluación de características cualitativas.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
13
PLANTEAMIENTO DEL PROBLEMA
La estructura anatómica de la madera, es influenciada por la genética de la especie y por
el comportamiento adaptativo que estas tengan en condiciones naturales cambiantes
como: cambio de uso del suelo, cambio climático, afectación de ecosistemas naturales,
entre otros. Estas condiciones actúan sobre las especies generando variaciones en su
estructura anatómica, por lo que se considera importante identificar las características
que se afectan más fácilmente por factores ambientales, y que pueden dar información
de la especie, su crecimiento y por lo tanto su adaptación a los cambios producidos por
estos fenómenos.
La revisión de estudios sobre los efectos de la variación de los factores ambientales
como el clima sobre los elementos anatómicos de la madera, en la región tropical son
pocos. Sin embargo las investigaciones realizadas por autores como Carlquist (1977),
Barajas-Morales (1985), Pérez (1989), Lindorf (1994), Aguilar-Rodríguez & Barajas-
Morales (2005), León (2005), Araque & León (2006), Luostarinen (2006), Moya &
Tomazello (2008), Terrazas et al., (2008), Polanco & Grande (2009), Chávez-Romero et
al., (2010), Cosmo (2012), Gámez (2013), Meena & Gupta (2014) y Barrios & Rondón
(2014) entre otros, demuestran como los elementos anatómicos de la madera se ven
afectados por las condiciones ambientales en las que se desarrollan las especies.
Es importante, resaltar que se encuentra muy poca información reportada para los
ecosistemas húmedos tropicales. Sin embargo, León (2013) realizó un estudio en un
bosque húmedo tropical en la reserva forestal Ticoporo (Barinas, Venezuela) que tuvo
como propósito el estudio de las características ecoanatómicas de los elementos de
conducción del xilema secundario para 87 especies de bosque húmedo tropical, en
donde se demostró que las especies estudiadas poseen características anatómicas que
desarrollan un sistema de conducción de comportamiento mesomórfico y con
orientación hacia la eficiencia. En Colombia, en cambio, se puede afirmar que aún no
se han reportado resultados de investigaciones en esta área, por lo que se considera
necesario desarrollar líneas de investigación que aporten al conocimiento de las
relaciones entre los factores climáticos y los rasgos anatómicos de las especies leñosas
representativas de nuestros ecosistemas húmedos tropicales, tan afectados por

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
14
múltiples factores como la alta tasa de deforestación, la sobreexplotación de especies
valiosas y el cambio climático, entre otros.
Debido a este vacío de información, surge la necesidad de identificar los elementos
anatómicos de la madera que se encuentran afectados por la influencia de los factores
ambientales en especies maderables de bosque húmedo tropical (bh-T) en Colombia y
su variabilidad, ya que estos elementos son los que otorgan propiedades físicas y
mecánicas que dan valor a dichas especies; además de aportar información sobre su
adaptación a las condiciones ambientales producidas por efecto de los fenómenos de
fragmentación, degradación, deforestación, sobreexplotación de especies y cambio
climático, entre otros.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
15
JUSTIFICACIÓN
El bosque húmedo tropical es un depósito grande de la diversidad biológica del mundo
(Padilla, 2003) por lo que las especies maderables que allí se encuentran son de gran
importancia ecológica, cultural y económica. Su alto valor económico ha llevado a una
explotación desmedida sobre algunas de ellas, lo cual amenaza sus poblaciones. Por ello
es urgente estudiar y analizar las estrategias que desarrollan las especies maderables
para sobrevivir a los cambios ambientales (Norman et al., 2013) y con base en ello
poder establecer medidas de conservación y restauración de los ecosistemas.
El comportamiento de la madera va a ser un reflejo de sus características anatómicas,
las mismas van a determinar en gran medida sus propiedades físicas, químicas y
mecánicas y por ende su posible uso y valor económico (León & Espinoza, 2001).
El estudio de las características anatómicas de la madera es muy importante a nivel
ambiental y socioeconómico; ya que al determinar cómo estas características afectan el
crecimiento en el medio húmedo tropical, se puede emplear esta información en la toma
de decisiones en el manejo adecuado del bosque, teniendo en cuenta la ecología de la
especie.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
16
OBJETIVOS
OBJETIVO GENERAL
Evaluar la ecoanatomía xilemática de 25 especies del bosque húmedo tropical.
OBJETIVOS ESPECÍFICOS
Evaluar las características anatómicas de la madera de cada una de las especies
seleccionadas.
Determinar qué características anatómicas de las especies dan respuesta al clima.
Identificar la variación en los elementos anatómicos de la madera que dan
respuesta al clima.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
17
MARCO DE REFERENCIA
MARCO TEÓRICO
Ecoanatomía
Como resultado de la interacción entre su material genético (genotipo) y los diversos
factores ambientales, las plantas presentan diferentes cambios que pueden manifestarse
mediante la modificación de sus características anatómicas o funcionales (fenotipo), lo
que les permite adaptarse a las condiciones del ambiente (Montaño et al., 2011).
El análisis del efecto de algunos factores ambientales como humedad, temperatura,
estacionalidad, altitud y distribución geográfica, sobre características anatómicas
cuantitativas y cualitativas ha permitido entender cómo se desarrollan cambios
adaptativos funcionales por parte de las plantas a los cambios ambientales (León, 2005).
Por ejemplo, Castro et al., (1998) citados por Montaño et al., (2011) encontraron que la
altitud y el clima son dos factores que tienen influencia directa sobre la anatomía de la
madera, en donde los elementos vasculares más cortos se encuentran en localidades
frías y secas y los más largos en localidades húmedas y cálidas. Por su parte, Carlquist
(1977) estudió la flora maderable en Australia en los ecosistemas sotobosque, laderas
graníticas costeras, pantano, brezal de arena y desierto para determinar si la anatomía de
la madera estaba relacionada con la ecología y así los elementos vasculares podían
adquirir un valor predictivo con respecto los factores ambientales; encontrando que los
elementos vasculares de la madera de las especies estaban influenciados por la
xeromorfia y mesomorfia de sus hábitats.
La influencia de los factores ambientales es principalmente estudiada en elementos
vasculares, ya que una de las principales funciones del tejido xilemático es el transporte
de agua (León, 2005). De acuerdo a Panshin & Zeeuw (1980), la variación de los
elementos anatómicos es uno de los aspectos más estudiados en especies forestales de
importancia económica, ya que, como lo menciona León & Espinoza (2001), estas
características anatómicas influencian directamente las propiedades físicas y mecánicas
de la madera.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
18
Es debido a esto que los estudios ecoanatómicos adquieren importancia, ya que
permiten determinar la variación de las estructuras anatómicas de la madera en
diferentes ambientes, además de que permiten conocer la seguridad y eficiencia del
transporte del agua (León, 2005).
Ambiente
Para su crecimiento y desarrollo, las plantas requieren suelos y temperaturas favorables,
disponibilidad adecuada de agua y aireación del suelo, luz y dióxido de carbono para la
fotosíntesis, y elementos esenciales para el equilibrio nutricional satisfactorio. Estos
factores influenciaran directamente el tipo de comunidad vegetal que se pueda
establecer en el sitio (Cosmo, 2012). El éxito que tengan las especies en establecerse en
un lugar dependerá de las condiciones anatómicas que esta tenga para adaptarse a las
condiciones ambientales del sitio, especialmente a la disponibilidad de humedad (León,
2013). Medina et al. (2013) señalan que la arquitectura hidraúlica de la madera de una
especie puede ser entendida como una de las estrategias de adaptación al ambiente
donde se desarrolla y por ello puede ser interpretada desde una perspectiva ecológica
que permite estimar el grado de seguridad o eficiencia de conductividad que presentan
las especies en determinado sitio o etapa de su desarrollo.
Bosque húmedo tropical
Según la clasificación de zonas de vida de L. R. Holdridge se establecen por medio de
factores como temperatura, precipitación o altitud, parámetros que permiten el
desarrollo de cierto tipo de vegetación, característica en cada una de las zonas. El
bosque húmedo tropical según esta clasificación, es el que se encuentra en lugares con
altitud entre 0 y 1000 metros sobre el nivel del mar, precipitación entre 2000 y 4000
mm al año y temperaturas entre 24 y 35 grados centígrados (Palacios, s.f.)
En cuanto a la vegetación, se encuentran cinco estratos (superior de árboles emergentes,
árboles que alcanzan el dosel, subdosel, sotobosque y basal); se presentan formas de
crecimiento tales como: epifitismo, lianas, estranguladoras, parásitas, saprófitas. Las
características comunes en los árboles incluyen bambas, o raíces tablares, hojas grandes,
ápices acuminados en las hojas, corteza delgada, y frutas grandes y carnosas. Los suelos
que presentan son predominantemente los oxisoles, suelos estériles, muy lavados y

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
19
lixiviados. La fauna es altamente diversa en los diferentes grupos de vida animal:
mamíferos, aves, reptiles, anfibios y se presentan adaptaciones a la vida arbórea. Las
principales formaciones son neotropical, africano e indo –malaysiana (Palacios, s.f.).
Madera
La madera es el producto de las divisiones del cambium vascular y está constituido por
una serie de células o elementos leñosos que han pasado a través de diferentes etapas de
desarrollo, desde la división celular hasta la diferenciación y maduración de células,
para formar el tejido leñoso (Webster & McKetchine, 1980). Este tejido se compone de
células como: Vasos, traqueidas, parénquima y fibras con pared celular además de
celulosa, hemicelulosa y lignina (Montaño et al., 2011), y un componente químico
secundario formado por diferentes tipos de extractos, azucares y otras sustancias
orgánicas e inorgánicas (Zobel & Van Buijtenen, 1989). Estas propiedades hacen de la
madera un material que puede ser usado con diferentes propósitos incluyendo papel,
materiales de construcción, químicos, energía e incluso de comida (Zobel & Van
Buijtenen, 1989). El estudio anatómico de la madera se realiza desde sus características
estructurales, anatómicas y físicas desde lo macroscópico, mientras que el estudio de
sus características celulares se realiza desde lo microscópico (León & Espinoza, 2010)
Características anatómicas
Vasos
Las maderas latifoliadas presentan vasos, estos son elementos prosenquimatosos y
cumplen como función el transporte de agua y sales minerales. Dichos elementos se
pueden observar como orificios, cuando realizamos un corte transversal en la madera
los mismos reciben el nombre de poro. Al momento de realizar la descripción anatómica
de los poros se deben tener en cuenta características tales como porosidad, disposición,
tamaño de los poros, agrupación de los poros y los contenidos depositados en el interior
de los mismos (León & Espinoza, 2001).
Tamaño de los poros: se refiere al tamaño promedio de los poros cuando se
observan en la sección transversal.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
20
Disposición de los poros: se refiere al arreglo u orientación que presentan los
poros, con respecto a la dirección de los radios, a lo ancho de la sección transversal. Se
clasifican en los siguientes tipos: Radial, radial oblicua, dendrítico, bandas tangenciales
y ningún patrón definido.
Agrupación de los poros: se refiere al grado de contacto que existe entre los
poros. Incluye las siguientes categorías: Exclusivamente solitarios, poros múltiples
radiales y arracimados.
Platina de perforación: al extremo de cada elemento vascular se encuentran unas
aberturas que reciben el nombre de platinas de perforación; la forma de estas es
variable, lo cual les otorga un carácter útil para diferenciar ciertas maderas. Según
IAWA committe (1989), citado por León & Espinoza (2001), se presentan los
siguientes tipos: simple, reticulada, escaleriforme y foraminada.
Contenido de los poros: se refiere a la presencia o ausencia de cualquier tipo de
depósito en el interior de los poros. Pueden ser los siguientes: Tílides, gomas, otras
sustancias.
Traqueidas
Las traqueidas son células largas que presentan lúmen celular y son cerradas en los
extremos por lo que se comunican por pares de punteaduras areoladas con torus. En las
coníferas su función primaria es la conducción y función secundaria el sostén. (Licoln
& Zeiger, 2006). Sin embargo en las latifoliadas no tienen función de transporte y sólo
proporcionan resistencia; tienen paredes celulares más delgadas que las fibras, por lo
que zonas con alta presencia de traqueidas parecen más claras (Sacarello, 2010).
Las traqueidas no se encuentran presentes en todas las latifoliadas, pero cuando están
presentes se categorizan en dos tipos principales: Traqueidas vasculares y traqueidas
vasicéntricas (León & Espinoza, 2001).
Fibras
Las fibras son células alargadas, con extremos puntiagudos y cerrados, comúnmente de
paredes gruesas (Léon & Espinoza, 2001). Las fibras son el principal componente de
las maderas latifoliadas, algunas pueden alcanzar hasta el 50% del volumen; son células

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
21
largas (1-2 mm) con diámetro entre 0,01mm y 0,05 mm, su función principal es dar
soporte al tronco (Sacarello, 2010).
Las paredes de las fibras pueden ser muy delgadas o tan gruesas que la cavidad celular
prácticamente no se pueda observar. El grosor de las paredes de las fibras depende de
las condiciones climáticas y de las características propias de la especie. Es una
característica muy importante ya que tiene una gran influencia sobre las propiedades de
resistencia, contracción y trabajabilidad de la madera (León & Espinoza, 2001).
Parénquima axial
El parénquima axial consiste en células axiales alargadas o hileras de células vivas en la
madurez, derivadas de las células iniciales fusiformes del cambium. Las células del
parénquima axial son usualmente de pared más delgada que las fibras con las que se
encuentran asociadas. Cada célula en una hilera de parénquima axial está usualmente
rodeada por una pared secundaria.
La distribución de las células del parénquima axial siempre se puede observar en la
sección transversal, sin embargo al momento de diferenciarlas de otro tipo de células es
mejor recurrir a la sección radial. El parénquima axial se clasifica principalmente en
ausente y presente; cuando está presente puede ser apotraqueal (distribuido sin relación
con los vasos) o paratraqueal (relacionado con los vasos). Los tipos de parénquima son:
ausente, difuso, difuso en agregados, apotraqueal en bandas, marginal, adyacente al
radio, vasicéntrico escaso, vasicéntrico abundante, abaxial, aliforme, confluente e
intervascular (Carlquist, 1988).
Parénquima radial
Los radios son el conjunto de células parenquimatosas que se arreglan de forma tal que
adquieren la apariencia de líneas o bandas y que tienen como función el
almacenamiento y conducción de sustancias alimenticias (García et al., 2003). Estas
células parenquimatosas son alargadas y se distinguen dos tipos: procumbentes (en
disposición radial) y decumbentes (en disposición vertical), las cuales de acuerdo a su
disposición definen el tipo de radios (Alonso, 2011).
Los radios de las latifoliadas están constituidos solo por células parenquimáticas. Sin
embargo, dichas células pueden presentar una gran variación, en cuanto a forma y

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
22
tamaño por lo que se establecen diferentes tipos de radios en base a su composición.
Con respecto a la composición de los radios (IAWA comitte. 1989) ha establecido 6
categorías.
A nivel microscópico, las características dimensionales de los radios se definen por su
longitud, ancho y altura. La longitud se mide desde el cambium hasta el extremo interno
del radio. El ancho corresponde a su extensión tangencial y se expresa con el número de
células en esa dirección. La altura se mide en dirección paralela al eje longitudinal del
tallo (Leon & Espinoza, 2001).
Contenidos en las células y elementos secretores
Contenidos en los poros: cuando los poros presentan contenidos, podemos
encontrar los siguientes: Tílides; que se presentan cuando ocurre la transformación de
albura en duramen y se puede observar como almacenes brillantes o vidriosos; gomas:
son depósitos de coloración oscura que obstruye total o parcialmente los poros cuando
ocurre la transición entre albura y duramen; otros tipos de sustancias (León & Espinoza,
2001).
Cristales: Cuando éstos aparecen en la madera, usualmente se describe la
abundancia, forma, tamaño y distribución (radios, parénquima axial o fibras).
(Carlquist, 1988).
Sílice: Este contenido se puede observar principalmente en forma esférica o
pueden ser alargados o emparejados y presentan tamaños diminutos, medianos o
grandes (Carlquist, 1988).
Células oleaginosas y mucilaginosas: Estas células dan origen en su interior a
otro tipo de contenidos que pueden ser aceites o mucílagos (Carlquist, 1988).
Conductos: Son espacios intercelulares que se pueden desarrollar en los sentidos
longitudinal (se observa en el plano transversal) y transversal (se observa en el plano
tangencial) (García et al., 2003).
Índice de vulnerabilidad
Este índice permite evaluar la eficiencia y la seguridad en la conducción del agua a
través del xilema de una planta, al igual que permite evaluar la resistencia o

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
23
vulnerabilidad a condiciones de estrés hídrico. Valores bajos indican una alta
redundancia de vasos, una capacidad para resistir al estrés hídrico y al congelamiento y
por consiguiente una menor vulnerabilidad. De igual forma a través de este índice es
posible determinar el comportamiento xeromórfico (IV < 1) o mesomórfico (IV > 1) de
la planta (Carlquist, 1977 y León, 2005). Este índice fue propuesto por Carlquist (1977)
y se calcula mediante la siguiente ecuación:
IV =𝐷𝑖á𝑚𝑒𝑡𝑟𝑜 𝑑𝑒 𝑣𝑎𝑠𝑜𝑠 (µ𝑚)
𝑁 𝑣𝑎𝑠𝑜𝑠/𝑚𝑚2
Índice de mesomorfía
Al igual que con el índice de vulnerabilidad, mediante este índice es posible determinar
el comportamiento xermórfico o mesomórfico de las especies. Cuando el índice arroja
valores inferiores de 75 se considera un comportamiento xeromórfico, mientras que
cuando arroja valores superiores a 200 se considera un comportamiento mesomórfico
(Calrquist, 1977). Se calcula de acuerdo a la siguiente ecuación:
IM = IV ∗ Longitud de Elemento Vascular (µm)
ESTADO DEL ARTE
La ecoanatomía estudia la relación entre la estructura de la madera y factores
ambientales. León (2005) afirma que se han realizado algunos estudios con énfasis en el
efecto de ciertos factores ecológicos como humedad, temperatura, estacionalidad, altitud
y distribución geográfica sobre caracteres anatómicos cuantitativos como diámetro de
poros, frecuencia de vasos, agrupamiento de los vasos, longitud de elementos vasculares
y tamaño de punteaduras intervasculares.
Los estudios realizados en este tema han tenido varios enfoques; algunos autores han
realizado estudios de ecoanatomía en zonas de vida específicas, empleando diversas
especies y obteniendo así, el resultado de las variaciones anatómicas provocadas por la
zona de vida (Barajas-Morales, 1985; Pérez, 1989; Lindorf, 1994; Aguilar-Rodríguez &
Barajas-Morales, 2005; León, 2005; Polanco & Grande, 2009; Cosmo, 2012 y León,
2013). Por otro lado, otros autores se enfocan en la ecoanatomía de las familias, géneros

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
24
o especies en ciertas zonas de vida (Araque & León, 2006; Luostarinen, 2006; Moya &
Tomazello, 2008; Terrazas et al., 2008; Chávez-Romero et al., 2010; Gámez, 2013;
Barrios & Rondón, 2014; Meena & Gupta, 2014).
En cuanto a ecosistemas, Barajas-Morales (1985) realizó una comparación de las
estructuras de la madera entre especies de un bosque húmedo tropical y un bosque
deciduo, teniendo en cuenta variables tanto cualitativas como cuantitativas para 27
especies en cada bosque, obteniendo diferencias significativas en algunos elementos
como diámetro de vasos, abundancia de poros, longitud de vasos y fibras, altura de
radios y espesor de la pared de los vasos entre los dos ecosistemas. Por su parte, Pérez
(1989) realizó una investigación en un bosque húmedo premontano, en el que encontró
que las 40 especies estudiadas poseían estructuras anatómicas e índices de
vulnerabilidad y mesomorfía orientados hacia la eficiencia en la conducción del agua.
En un bosque muy seco tropical, Lindorf (1994) estudió la econatomía de la madera,
encontrando especies con comportamiento xeromórfico y características anatómicas
orientadas hacia la seguridad en la conducción del agua. Por su parte, Aguilar-
Rodríguez & Barajas.Morales (2005) estudiaron 29 especies de un bosque de niebla,
teniendo en cuenta características anatómicas cualitativas y cuantitativas en vasos,
radios y fibras, en las que encontraron elementos anatómicos con características
orientadas hacia la seguridad de la conducción de agua. Por su parte León (2005),
estudió la anatomía ecológica del xilema secundario de un bs-T de Venezuela, para ello
se basó en 51 individuos de especies arbóreas y se correlacionó la estructura anatómica
con las condiciones del ambiente; finalmente se determinaron los índices de
vulnerabilidad, mesomorfía y agrupamiento de vasos.
Por otra parte, Polanco & Grande (2009) realizaron la caracterización anatómica de 40
especies de bosque altoandino encontrando que además de la influencia que hay por
parte de las características ecológicas, también existe una amplia relación entre las
características anatómicas y el grado de evolución de las especies. Para un bosque
húmedo premontano, Cosmo (2012) estudió la anatomía de la madera de 19 especies
mediante la medición de diámetro de vasos, frecuencia de vasos, agrupación de vasos,
área del xilema ocupado por vasos, longitud de elementos vasculares, longitud de fibras,
grosor de fibras, ancho de fibras, altura y ancho de radios y el cálculo de los índices de

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
25
vulnerabilidad y mesomorfía, encontrando que las especies se poseían características
anatómicas orientadas hacia la eficiencia del transporte de agua.
Por último, León (2013) realizó un estudio econanatómico de la madera de un bosque
húmedo tropical, tomando como muestra 87 especies para las que se determinaron
medidas de diámetro, frecuencia y longitud de vasos y a partir de allí se calcularon los
índices de vulnerabilidad y mesomorfía; lo cual permitió concluir que dichas especies
poseen un sistema de conducción de comportamiento mesomórfico orientado hacia la
eficiencia hídrica. Por otra parte, en bosque húmedo tropical en Colombia, Rodríguez,
(2012) realizó un estudio de rasgos funcionales donde correlacionó la estructura
anatómica de la madera con las condiciones del ambiente, y encontró un predominio de
caracteres orientados hacia la eficiencia en conducción de agua, mostrando poros con
diámetros grandes y con baja frecuencia.
Para ilustrar las características ecoanatómicas por familia, Gámez (2013) estudió la
ecoanatomía una familia (MALVACEAE), teniendo en cuenta cuatro especies en el
ecosistema de bosque húmedo tropical en transición al bosque seco tropical, que arrojó
como resultados a partir de los índices de vulnerabilidad y mesomorfía una condición
mesomórfica y un sistema de conducción orientado a la eficiencia. Igualmente, Barrios
& Rondón (2014), estudiaron la anatomía al interior de una familia (ERICACEAE.
subfamilia Vaccinioideae), tomando como base tres especies en un páramo en
Venezuela e incluyendo mediciones en vasos, punteaduras y poros se hallaron índices
que dieron como resultado el comportamiento xeromórfico de las especies, además de
encontrar que el sistema de conducción de las mismas está orientado hacia la seguridad.
Por otra parte, otros autores han realizado investigaciones a nivel de género, es el caso
de Terrazas et al., (2008), quienes evaluaron la anatomía de la madera en relación con el
tamaño de la planta y la latitud para el género Buddleja perteneciente a la familia
BUDDLEJACEAE, encontrando que las especies que crecían en lugares con
condiciones extremas poseen hábitos principalmente arbustivos y presentan poca
variación en el tamaño celular, en ambientes heterogéneos poseen diferentes hábitos y
así mayor variación en el tamaño celular de la madera. Igualmente Montaño-Arias et al.
(2013) estudiaron la ecoanatomía de elementos vasculares en madera de cinco especies
del género Mimosa, tomando tres individuos por especie en los cuales se midieron

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
26
diámetro, frecuencia, grosor de la pared y longitud de los vasos y se calcularon los
índices de vulnerabilidad y mesomorfía; dando como resultado un sistema de
conducción mesofítico y una baja resistencia de las especies al estrés hídrico.
En cuanto a nivel de especie, Araque & León (2006), realizaron un estudio sobre la
ecoanatomía de la madera de Spondias mombin en una reserva en Venezuela para dos
posiciones fisiográficas, en el que se identificó que la especie poseía un comportamiento
xilemático mesomórfico y un sistema de conducción orientado a la eficiencia tanto en
banco como bajío en la zona de estudio; lo cual permitió concluir que el microclima no
afectaba significativamente el leño de la especie y por lo tanto tiene una alta
adaptabilidad.
Moya & Tomazello (2008), realizaron una investigación con Gmelina arbórea,
buscando las variaciones que se producían en la anatomía del xilema debido a las
diversificaciones de las condiciones ecológicas. Chávez-Romero et al., (2010),
realizaron una investigación similar, estudiando la variación anatómica en la madera de
Quercus obstusata; para ello se describieron las características anatómicas de la madera
y se evaluó estadísticamente la variación anatómica de nueve caracteres cuantitativos de
su madera en función de 11 parámetros climáticos en cuatro localidades. Meena &
Gupta (2014) realizaron un estudio para Albizia procera, en el que se evalúa la anatomía
de la madera de dicha especie en diferentes localidades de la región de la India y en el
que se encontraron diferencias significativas entre la estructura anatómica de la madera
en las zonas estudiadas y una alta adaptabilidad ecológica en clima subtropical y
tropical templado debido a la relación entre los índices de vulnerabilidad y mesomorfía.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
27
METODOLOGÍA
El proyecto se realizó en tres fases. La primera fase consiste en un diagnóstico, la
segunda fase en actividades en laboratorio y la tercera fase en procesamiento y análisis
de los datos. El procedimiento se muestra a continuación:
Figura 1. Mapa Conceptual; Metodología seguida en el proceso de Investigación.
Fuente: Autores.
ÁREA DE ESTUDIO
La cuenca del Río Atrato, comprendida entre los 5° y 8° de latitud Norte y los 76° y 78°
de longitud Oeste, en el extremo noroccidental de Colombia y de Sudamérica. La
cuenca se encuentra delimitada al este por la cordillera occidental y al oeste por las
serranías del Baudó y del Darién, al sur se encuentra la divisoria de aguas con el Río
San Juan definida por el istmo de Istmina (MMA et al., 2002).
Evaluación ecoanatomía xilemática especies bh-T
Fase I: Diagnóstico
Revisión bibliográfica
Identificación problema
Identificación área de estudio, especies
y variables
Fase II: Laboratorio
Elaboración laminillas
histológicas
Ablandamiento madera
Realización cortes y selección
Tinción, deshidratación y
aclaramiento
Montaje
Rotulado final
Maceración/ Disociación
Obtención astillas
Inmersión químicos y
calentamiento
Tinción, limpieza
Montaje
Descripción anatómica
Descripción microscópica
Medición de elementos
Toma de fotografías
Medición en el software
Fase III: Procesamiento y análisis de datos
Cálculo índices
Análisis cualitativo
Análisis cuantitativo
Análisis de resultados
Publicación de la información

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
28
Los materiales que conforman la planicie aluvial del Río Atrato, son primordialmente
sedimentos aluviales, arenas, limos y arcillas (Alcaldía de Vigia del fuerte, 2006),
generadas en el proceso denudativo de la cordillera occidental y de las serranías del
Baudó y del Darién que se ha producido desde finales del terciario y a través del
cuaternario, acompañado de procesos de tectonismo y levantamiento litoral. Los suelos
son superficiales, pobremente drenados y de baja fertilidad (Instituto de Investigaciones
en Geociencia, Minería y Química, 1989)
Figura 2. Área de estudio (Cuenca del Río Atrato). Municipios de Bojaya, Vigía del fuerte y San Miguel. Fuente.
OCHA, 2003.
Fuente: Autores.
ESPECIES EVALUADAS
Las especies evaluadas fueron colectadas para proyectos ejecutados por el laboratorio
de maderas en convenio con la CAR y USAID, en los cuales se tuvo en cuenta que estas
especies maderables fueran comercializadas en el territorio nacional.
Las especies seleccionadas para la realización del estudio pertenecen al ecosistema de
bosque húmedo tropical y se encuentran registradas en las colecciones biológicas de la
Universidad Distrital Francisco José de Caldas, estas se relacionan en la Tabla 1.
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
29
Tabla 1. Especies a evaluar en el estudio. Estas especies han sido colectadas para la realización de los proyectos
CAR USAID, y se encuentran registradas en las colecciones biológicas de la Universidad Distrital Xiloteca JaLR
UD y Herbario Forestal UDBC
N° NOMBRE COMÚN NOMBRE CIENTIFÍCO FAMILIA
1 Caidita Endlicheria tschudyana (Lasser) Kosterm. LAURACEAE
2 Caimo Ecclinusa ramiflora Mart. SAPOTACEAE
3 Capitancillo Pentaclethra macroloba (Willd.) Kuntze LEGUMINOSAE
4 Carrá Huberodendron patinoi Cuatrec. MALVACEAE
5 Chachajo Aniba perutilis Hemsl. LAURACEAE
6 Chingalé Jacaranda copaia (Aubl.) D.Don BIGNONIACEAE
7 Chocho Abarema jupunba (Willd.) Britton & Killip LEGUMINOSAE
8 Choibá Dipteryx oleifera (Aubl.) Willd. LEGUMINOSAE
9 Curibano Amarillo Buchenavia cf. viridiflora Ducke COMBRETACEAE
10 Genené Caryocar amygdaliferum Mutis ex Cav. CARYOCARACEAE
11 Guasco Eschweilera coriacea (DC.) S.A.Mori LECYTHIDACEAE
12 Hobo Spondias mombin L. ANACARDIACEAE
13 Incibe Ocotea cymbarum Kunth LAURACEAE
14 Jagua Genipa americana L. RUBIACEAE
15 Jigua Amarillo Ocotea bofo Kunth LAURACEAE
16 Jigua Negro Ocotea cernua (Nees) Mez LAURACEAE
17 Laguno Vochysia guatemalensis Donn. Sm. VOCHYSIACEAE
18 Nato Mora oleifera (Hemsl.) Ducke LEGUMINOSAE
19 Nispero Manilkara bidentata (A.DC.) A.Chev. SAPOTACEAE
20 Nuanamo Virola flexuosa A.C. Sm. MYRISTICACEAE
21 Olleto Lecythis tuyrana Pittier LECYTHIDACEAE
22 Platano Mare Osteophloeum platyspermum (Spruce ex
A.DC.) Warb. MYRISTICACEAE
23 Polvillo Handroanthus guayacan (Seem.) S.O.Grose BIGNONIACEAE
24 Tara Simarouba amara Aubl. SIMAROUBACEAE
25 Truntago Vitex floridula Duchass. & Walp. LAMIACEAE
Fuente: Autores.
VARIABLES EVALUADAS
Las variables cuantitativas evaluadas en el presente estudio fueron seleccionadas de
acuerdo a los estudios anatómicos y ecoanatómicos realizados por Barajas-Morales
(1985), Pérez (1989), Lindorf (1994), Aguilar-Rodríguez & Barajas-Morales (2005),
León (2005), Polanco & Grande (2009), Pulido et al., (2011), Cosmo (2012), Rodríguez
(2012), León (2013) y Vasquez (2013). Estas variables se registran en la Tabla 2.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
30
Tabla 2. Variables cuantitativas del estudio.
Fuente: García et al., (2005) citado por Pulido et al., (2011).
A nivel cuantitativo, adicionalmente se calcularon los índices de vulnerabilidad (IV) y
mesormorfía (IM) propuestos por Carlquist (1977).
En cuanto a las variables cualitativas, se evaluaron de acuerdo a IAWA (1989) Estas
son presentadas a continuación:
Tabla 3. Variables cualitativas del estudio (Vasos, fibras, parénquima, radios y otros.)
Elemento Variable
Vasos
Porosidad
Disposición de poros Elemento Variable
Agrupación de poros
Parénquima
Tipo de parénquima axial
Forma de poro Tipo de células en parénquima
axial
Platina de perforación
Radios
Ancho de radios
Arreglo punteaduras
intervasculares Composición de radios
Forma punteaduras intervasculares Tipo punteaduras
radiovasculares
Abertura punteaduras
intervasculares Variantes en células radiales
Tamaño punteaduras
intervasculares
NA
Presencia de estratificación de
elementos
Punteaduras ornadas Presencia de elementos
secretores
Presencia inclusiones Presencia cristales
Presencia de traqueídas Presencia de sílice
Fibras Tamaño punteaduras de fibras Anillos de crecimiento
Ubicación punteaduras de fibras Olor

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
31
Presencia de engrosamientos
espiralados
Presencia de fibras septadas
Grosor de pared de la fibra
Fuente Adaptado de IAWA (1989).
ELABORACIÓN DE LAMINILLAS HISTOLÓGICAS
Ablandamiento de la madera
Para la realización de los trabajos anatómicos de la madera, se necesita contar con
preparaciones fijas de los cortes transversal, tangencial y radial. En el caso de las
maderas blandas, la obtención de los cortes fue relativamente fácil, ya que con sólo
hervirlas en agua destilada durante unas cuantas horas, pudieron obtenerse cortes hasta
de 20 µm (Huertas, 1963). Sin embargo para las maderas duras, fue necesario aumentar
las horas de cocción o someter los cubos de madera a la acción del vapor en una
autoclave con presión variable de acuerdo a la dureza de los cubos, como lo recomienda
Guridi, (1977).
Corte y selección
Luego de haber pasado la madera por el proceso de ablandamiento, se realizaron cortes
finos de los tres planos de estudio (transversal, tangencial y radial), utilizando un
micrótomo de rotación Leica RM 2255, este equipo permite graduar el espesor de las
secciones entre 5 y 100 µm lo cual es útil, teniendo en cuenta que el grosor
recomendado para los montajes histológicos de madera es entre 12 y 45 µm (Guridi,
1977).
Posterior a la obtención de los cortes, se seleccionaron los más adecuados para la
observación de las características anatómicas que se van a evaluar, se realizó un corte
de alrededor 7mm x 7mm, en el cual se pueden observar las características de relevancia
para este estudio (fibras, platinas de perforación, vasos y punteaduras, entre otras).

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
32
Figura 3. Realización de cortes histológicos transversal, tangencial y radial en micrótomo de rotación Leica RM
2255.
Fuente: Autores.
Tinción, deshidratación y aclaramiento
Para el proceso de tinción, se empleó lo sugerido por Sandoval (2005). Se llevaron los
cortes realizados en la etapa anterior (transversal, radial y tangencial) a una cámara
extractora y se aplicó una solución de safranina al 1% para adherir color al corte y así
poder realizar una mejor observación de las estructuras a analizar.
Para eliminar los residuos de safranina y deshidratar los tejidos, se realizó un lavado con
etanol en concentración de 50% y se esperó a que el etanol se evaporara, luego se
repitió el procedimiento dos veces más para posteriormente aumentar la concentración
de etanol a 96%, dejarlo evaporar y lograr su deshidratación. Para el aclaramiento, se
empleó xileno puro, que es aplicado sobre los tejidos luego de la deshidratación y les
otorga a las células un aspecto más claro para la observación en el microscopio.
Igualmente se empleó lo sugerido por Sandoval (2005).

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
33
Figura 4. Proceso de tinción, deshidratación y aclaramiento de los cortes transversales, tangenciales y radiales de las
muestras. a. Reactivos empleados. b. Tinción con safranina. c y d. Lavado con alcohol. e. Deshidratación de las
especies. f. Ubicación de los cortes.
Fuente: Autores.
Montaje
Para el montaje de los tejidos, se dispuso de ellos en láminas portaobjetos de colocando
en la parte superior el corte transversal, en la parte inferior derecha el corte radial y en la
parte inferior izquierda el corte tangencial. Luego se aplicó una gota del conservante
adhesivo Eukitt en el centro de los tres tejidos y se colocó la laminilla cubreobjetos
(25mm x 25mm) desde el lado izquierdo con un ángulo de 45º para evitar que se
formaran posibles burbujas sobre los tejidos.
Para cada especie se realizaron un total de cinco (5) montajes, con el fin de obtener
información precisa de las estructuras anatómicas de cada especie.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
34
Figura 5. Proceso de montaje de laminillas histológicas (Acomodación de cortes, adición de Eukitt y colocación de
cubreobjetos. a. Ubicación de los cortes. b. Aplicación de Eukitt. c. Ubicación de la laminilla. y d. Montaje final.
Fuente: Autores.
MACERACIÓN/DISOCIACIÓN
Con el fin de realizar las mediciones de los elementos anatómicos objeto de estudio, se
realizó a la madera un proceso conocido como maceración; esto debido a que los cortes
histológicos muestran las estructuras en dos dimensiones y para obtener una vista
tridimensional, los elementos celulares deben ser aislados, mediante la técnica conocida
como disociación o maceración. Esta técnica consiste en separar las células mediante
técnicas mecánicas y químicas por medio de reactivos que las desintegran.
Existen varios métodos que pueden ser empleados, tales como: el método de Jefrey
(mezcla de ácido nítrico al 10% y ácido crómico al 10% en proporción 1:1), el método
de Foster (mezcla de etanol al 70% y ácido clorhídrico, en proporción 3:1) y el método
de Franklin (peróxido de hidrógeno al 30% y ácido acético glacial, en proporción 1:1).
(Safdari & Devall, 2012).
Obtención de astillas
Antes de iniciar la disociación por medio de técnicas químicas, es necesario emplear
técnicas mecánicas, por lo que con un escalpelo se obtuvieron astillas de la madera
cortando en sentido longitudinal.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
35
Inmersión en químicos y calentamiento
Se empleó el método de Franklin, el cual consiste en colocar el tejido con una mezcla
de peróxido de hidrógeno al 30% y ácido acético glacial, en proporción 1:1 en un tubo
de ensayo con tapa, y llevarlo al horno por 48 horas a una temperatura de 60°C (Safdari
& Devall, 2012).
Tinción y limpieza
Posterior a la disociación del tejido, se procedió a hacer limpieza y tinción, para lo que
se empleó un beaker de 100 mL con un papel filtro en la apertura. En este se colocó el
contenido del tubo de ensayo y se lavó con agua destilada hasta que se eliminó la
mezcla de ácido acético glacial y peróxido de hidrogeno. Luego se realizó el proceso de
tinción igual que el empleado con los cortes. Finalmente se recogió el material
disociado en viales de 25 mL con alcohol al 96%.
Figura 6. Procedimiento para maceración de la madera (a y b. corte de astillas, c y d. inmersión en reactivos, e.
Reactivos empleados y f. calentamiento de los tejidos).
Fuente: Autores.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
36
Figura 7. Proceso de tinción del material disociado (a. Tinción, b. limpieza con alcohol al 90% c, d y e.
almacenamiento).
Fuente: Autores.
Montaje
Para el montaje de las células, se tomó del vial una gota del material disociado y se
dispuso en láminas portaobjetos, se esperó a que el alcohol se evaporara, posteriormente
se aplicó una gota del conservante adhesivo Eukitt en el centro y se colocó la laminilla
cubreobjetos (25mm x 25mm) desde el lado izquierdo -con un ángulo de 45º para evitar
que se formen posibles burbujas sobre las células.
Para cada especie se realizó un total de tres (3) montajes, lo cual permitiría obtener
información representativa para realizar su análisis estadístico.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
37
Figura 8. Proceso para montaje del material disociado (a. y b. disposición de material macerado, c. Disposición en
láminas y d. colocación de cubrobjetos).
Fuente: Autores.
Rotulado preliminar
Con el fin de llevar seguimiento a las muestras, se realizó una marcación de cada una de
las laminillas histológicas obtenidas. Para esto, se coloca al lado derecho de la laminilla
el nombre común de la especie o el nombre científico, dependiendo de cada caso con un
marcador.
Figura 9. Rotulado preliminar de laminillas (Trazabilidad).
Fuente: Autores.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
38
Rotulado final
Al finalizar el proceso de montaje de todo el material, se generaron las etiquetas de
acuerdo al protocolo establecido por el Laboratorio de Maderas de la Universidad
Distrital Francisco José de Caldas, el cual establece que estas deben contener la
siguiente información: Nombre común, nombre científico, familia y determinador.
Figura 10. Laminillas etiquetadas en colección biológica de la Xiloteca JaLR UD.
Fuente: Autores.
DESCRIPCIÓN ANATÓMICA
Para la identificación de los elementos anatómicos presentes en cada una de las especies
y su clasificación, se realizó una descripción a partir de las laminillas histológicas,
teniendo en cuenta la matriz presentada en el Anexo 1, la cual contiene descripción de
vasos, fibras, radios, parénquima y otras variantes en las células. Para este
procedimiento se emplearon microscopios Carl Zeiss.
MEDICIÓN DE ELEMENTOS
Toma de fotografías
Para efectuar las mediciones de los elementos de nuestro interés, se tomaron fotografías
en diferentes aumentos (5x, 10x, 40x y 100x) de acuerdo al nivel de detalle requerido
por cada elemento, esto mediante el software Remote Capture DC, cámara Canon
PowerShot G9 y microscopio Axiostar Plus trilocular.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
39
Medición en software
Para la medición de las variables cuantitativas, se empleó el software ImageJ (Rasband,
2014), en el que a partir de una escala conocida en las fotografías anteriormente
mencionadas, se efectuaron mediciones de dichas variables. La medición de los
elementos anatómicos se realizó de acuerdo con lo sugerido por Richter & Dallwitz
(2000) de la siguiente manera:
Diámetro de vasos: El diámetro de vasos es medido en secciones transversales. Los
vasos son seleccionados teniendo en cuenta los más grandes o más pequeños. El
diámetro tangencial de los lúmenes de vaso, excluyendo la pared, es medido en la parte
más ancha del orificio. Se deben medir por lo menos 25 vasos.
Clases de tamaño
1. bajo (menor de 50 µm)
2. mediano (50–100 µm)
3. grande (100–200 µm)
4. muy grande (más de 200 µm)
Figura 11. Medición de diámetro de vaso. Ecclinusa ramiflora Mart.
Fuente: Autores.
Promedio del número de vasos/mm2: Todos los vasos son contados individualmente.
Por ejemplo: un vaso múltiple radial de 4 deberá ser contado como de cuatro. Si sólo
hay medio poro debe contarse uno sí, y uno no.
Clases de tamaño
1. muy pocos (menos de 5)
2. pocos (5–20)
3. moderadamente frecuentes (20–40)
4. frecuentes (40–100)
5. muy frecuentes (con más de 100)
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
40
Figura 12. Medición de número de vasos/mm2. Lecythis tuyrana Pittier.
Fuente: Autores.
Longitud de vaso: Se midió la longitud total de cada elemento de vaso de un extremo
al otro, en material macerado. Al menos 25 elementos de vaso fueron medidos para
obtener el promedio y rango.
Clases de tamaño
1. bajo (menor de 350 µm)
2. mediano (350–800 µm)
3. largo 8con más de 800 µm)
Figura 13. Medición de Longitud de vaso. Osteophloeum platyspermum (Spruce ex A.DC.) Warb.
Fuente: Autores.
Promedio del diámetro (vertical) de las punteaduras intervasculares: Se midieron
diez punteaduras evitando punteaduras excepcionalmente grandes o pequeñas, y se
registraron el rango correspondiente del diámetro de punteaduras.
Clases de tamaño
1. diminutas (menor de 4 µm)
2. pequeñas (4–7 µm)
3. medianas (7–10 µm)
4. grandes (con más de 10 µm)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
41
Figura 14. Medición del diámetro de punteadura intervascular. Endlicheria tschudyana (Lasser) Kosterm.
Fuente: Autores.
Longitud de las fibras: Se utilizaron maceraciones de madera de tronco adulto, y se
midió la longitud de por lo menos 25 fibras para determinar el promedio, rango, y la
desviación estándar.
Clases de tamaño
1. bajo (menor de 900 µm)
2. mediano (900–1600 µm)
3. largo (con más de 1600 µm)
Figura 15. Medición de Longitud de fibra. Aniba perutilis Hemsl.
Fuente: Autores.
Espesor de las fibras: Es el cociente que se da entre la diferencia del diámetro externo
de la fibra (d1) y el diámetro interno-lumen (d2) dividido por dos.
Clases de tamaño
1. Delgada
2. Mediana
3. Gruesa

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
42
Figura 16. Medición de grosor de la pared celular de la fibra.Mora oleifera (Hemsl.) Ducke
Fuente: Autores.
Altura de radios: Se miden los radios en el plano tangencial alrededor de su eje.
Clases de tamaño
1. hasta 500 µm
2. comúnmente de 500 hasta 1000 µm
3. comúnmente con más de 1000 µm
Figura 17. Medición altura de radios. Huberodendron patinoi Cuatrec.
Fuente: Autores.
Número de radios por mm: El número de radios por unidad linear es mejor
determinarlo de una sección tangencial a lo largo de una línea perpendicular al eje de
los radios.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
43
Figura 18. Medición de número de radios por mm lineal. Ecclinusa ramiflora Mart.
Fuente: Autores.
PROCESAMIENTO Y ANÁLISIS DE DATOS
En lo que respecta al análisis de datos se empleó el software R-Project con el cual se
realizaron diferentes tipos de análisis estadísticos descriptivos, análisis de similitud y
análisis de componentes principales. Ver mapa conceptual para el análisis de datos en la
Figura 19
Figura 19. Mapa conceptual. Proceso para el análisis de los datos obtenidos para variables cualitativas y
cuantitativas.
Fuente: Autores.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
44
RESULTADOS
RASGOS ANATÓMICOS CUALITATIVOS DE LA MADERA
Las características microscópicas de las especies objeto de estudio (vasos, fibras, radios,
parénquima y otras variantes en las células) se pueden observar en el Anexo 1.
Vasos
De las especies evaluadas Genipa americana L presentó porosidad semicircular (Figura
20a) y en las restantes se presentó porosidad difusa (Figura 20b).
Figura 20. Tipo de porosidad. a. Porosidad semicircular (Genipa americana L.) b. Porosidad difusa (Virola flexuosa
A.C. Sm.)
Fuente: Autores.
En cuanto a la disposición de los poros diecisiete especies no presentan un patrón
definido, cinco especies presentan disposición dendrítica (Buchenavia cf. viridiflora,
Vochysia guatemalensis, Ecclinusa ramiflora, Aniba perutilis, Manilkara bidentata,) y
tres especies presentan disposición diagonal (Genipa americana, Ocotea bofo, Ocotea
cernua)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
45
Figura 21. Disposición de poros. a. Sin patrón de disposición (Vochysia guatemalensis).b. Disposición dendrítica
(Buchenavia cf. viridiflora), c. Disposición diagonal (Ocotea bofo).
Fuente: Autores.
Para la agrupación de poros se observó que una especie presenta poros exclusivamente
solitarios (Genipa americana), veintitrés especies presentan poros solitarios
acompañados de otro tipo de agrupación ya sea múltiplos radiales o arracimados, y una
especie presenta poros agrupados exclusivamente en múltiplos radiales cortos (Virola
flexuosa).
Figura 22. Agrupación de poros. a. Poros solitarios (Genipa americana), b. Múltiplos radiales cortos (Virola
flexuosa)
Fuente: Autores.
La forma de los poros que se presenta en la mayoría de los casos es ovalada con
dieciocho especies, y siete especies presentan poros redondos (Ocotea bofo,
Handroanthus guayacan, Huberodendron patinoi, Spondias mombin, Ocotea cymbarum
y Vitex floridula).
a.
a.
b.
b.
c.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
46
Figura 23. Forma de los poros. a. Poro ovalado (Ecclinusa ramiflora), b. Poro redondo (Spondias mombin).
Fuente: Autores.
Las platinas de perforación en su mayoría son simples, ya que se presentan en veintidós
especies, mientras que simples y escaleriformes se presentan en dos especies (Ocotea
cernua y Endlicheria tschudyana) y simples y reticuladas en una especie (Virola
flexuosa).
Figura 24. Tipos de platinas de perforación. a. Escaleriforme (Endlicheria tschudyana), b. Reticulada (Virola
flexuosa), c. Simple (Ocotea cernua), d. Simple (Genipa americana).
Fuente: Autores.
En el arreglo de las punteaduras intervasculares se encontró que son principalmente
areoladas alternas ya que se presentan en veinticuatro especies y tan sólo una de las
especies posee punteaduras areoladas opuestas (Pentaclethra macroloba).
a. b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
47
Figura 25. Arreglo de las punteaduras intervasculares. a. Punteaduras areoladas opuestas (Pentaclethra macrolaba),
b. Punteaduras areoladas alternas (Ocotea cymbarum)
Fuente: Autores.
Por otra parte, la forma de las punteaduras es predominantemente poligonal,
presentándose en quince especies, mientras que la forma ovalada se presenta en cinco
especies (Lecythis tuyrana, Vochysia guatemalensis, Ocotea cernua, Endlicheria
tschudyana y Virola flexuosa) y la forma redonda igualmente se presenta en cinco
especies (Ecclinusa ramiflora, Dipteryx oleífera, Handroanthus guayacan, Ocotea
cymbarum y Vitex floridula). En cuanto a la abertura, todas las especies presentaron
abertura incluida.
Figura 26. Tipo de punteaduras intervasculares. a. Punteadura intervascular poligonal (Eschweilera coriacea), b
Punteadura intervascular ovalada (Vochysia guatemalensis), c. Punteadura intervascular redonda (Handroanthus
guayacan). a, b y c. Abertura incluida.
Fuente: Autores.
Las punteaduras son principalmente medianas, encontrándose en nueve especies
(Eschweilera coriacea, Huberodendron patinoi, Ocotea cernua, Aniba perutilis,
Dipteryx oleífera, Lecythis tuyrana, Ocotea cymbarum, Caryocar amygdaliferum y
Spondias mombin, siete especies presentaron punteaduras pequeñas (Virola flexuosa,
a.
a.
b.
b. c.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
48
Vochysia guatemalensis, Endlicheria tschudyana, Simarouba amara, Vitex floridula
Manilkara bidentata y Ecclinusa ramiflora), seis especies prsentaron punteaduras
diminutas (Jacaranda copaia, Genipa americana, Abarema jupumba, Handroanthus
guayacan, Mora oleifera y Pentaclethra macroloba) y por último sólo tres especies
presentaron punteaduras grandes (Ocotea bofo, Buchenavia cf. Viridiflora y
Osteophloeum platyspermum).
Figura 27. Tamaño de las punteaduras intervasculares. Punteaduras intervasculares grandes (Ocotea bofo), b.
Punteaduras intervasculares medianas (Ocotea cernua), c. Punteaduras intervasculares pequeñas (Simarouba amara),
d. Punteaduras intervasculares diminutas (Jacaranda copaia).
Fuente: Autores.
Por último, dieciséis especies poseen punteaduras ornadas y las nueve restantes no
(Genipa americana, Simarouba amara, Spondias mombin, Endlicheria tschudyana,
Jacaranda copaia, Aniba perutilis, Manilkara bidentata, Huberodendron patinoi y
Ecclinusa ramiflora).
a. b.
c. d.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
49
Figura 28. Punteaduras ornadas. a. Punteaduras intervasculares ornadas (Caryocar amygdaliferum), b. Punteaduras
intervasculares NO ornadas (Spondias mombin).
Fuente: Autores.
Inclusiones
En diez especies analizadas no se presentaron inclusiones en los vasos, mientras que en
ocho especies se presentaron gomas (Endlicheria tschudyana, Vochysia guatemalensis,
Mora oleífera, Abarema jupunba, Pentaclethra macroloba, Osteophloeum
platyspermum, Lecythis tuyrana y Dipteryx oleifera), en cinco tílides (Ocotea cernua,
Buchenavia cf. viridiflora, Caryocar amygdaliferum, Ocotea cymbarum y Eschweilera
coriacea) mientras que la presencia de tílides y esclerotílides solo se encuentra en dos
especies (Manilkara bidentata y Ecclinusa ramiflora).
Figura 29. Inclusiones a. Gomas (Vochysia guatemalensis), b. Tílides (Eschweilera coriacea) c. Esclerotilides
(Ecclinusa ramiflora)
Fuente: Autores.
Fibras
Las especies evaluadas presentaron principalmente grosor delgado (11 especies), ocho
especies presentaron pared celular gruesa (Mora oleifera, Ecclinusa ramiflora, Lecythis
tuyrana, Manilkara bidentata, Dipteryx oleifera, Buchenavia cf. viridiflora, Caryocar
a.
a.
b.
b. c.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
50
amygdaliferum, Eschweilera coriacea y Handroanthus guayacan) y por último seis
especies pared celular mediana (Endlicheria tschudyana, Genipa americana, Vochysia
guatemalensis, y Vitex floridula).
Figura 30. Grosor de la pared celular. a. Pared celular gruesa (Buchenavia cf. viridiflora), b. Pared celular mediana
(Endlicheria tschudyana), Pared celular delgada (Spondias mombin).
Fuente: Autores.
La mayoría de las especies (15) poseen punteaduras diminutas en las fibras, mientras
que en diez especies se presentan punteaduras areoladas distinguibles (Endlicheria
tschudyana, Ecclinusa ramiflora, Pentaclethra macroloba, Jacaranda copaia, Caryocar
amygdaliferum, Genipa americana, Ocotea cernua, Lecythis tuyrana, Simarouba amara
y Vitex floridula). Se registró que las punteaduras en las fibras están principalmente
restringidas a la pared radial (en 15 de las especies) y en diez de las especies las
punteaduras son visibles en la pared radial y tangencial (Huberodendron patinoi,
Ocotea bofo, Vochysia guatemalensis, Osteophloeum platyspermum, Handroanthus
guayacan, Endlicheria tschudyana, Ecclinusa ramiflora, Pentaclethra macroloba,
Caryocar amygdaliferum y Lecythis tuyrana).
Por otro lado, en dieciséis de las especies no se presentaron engrosamientos espiralados
en las fibras, mientras que en las nueves especies restantes sí se presentaron (Dipteryx
oleífera, Buchenavia cf. viridiflora, Eschweilera coriacea, Mora oleífera,
Handroanthus guayacan, Endlicheria tschudyana, Ecclinusa ramiflora, Pentaclethra
macroloba y Caryocar amygdaliferum).
a.
b.
c.
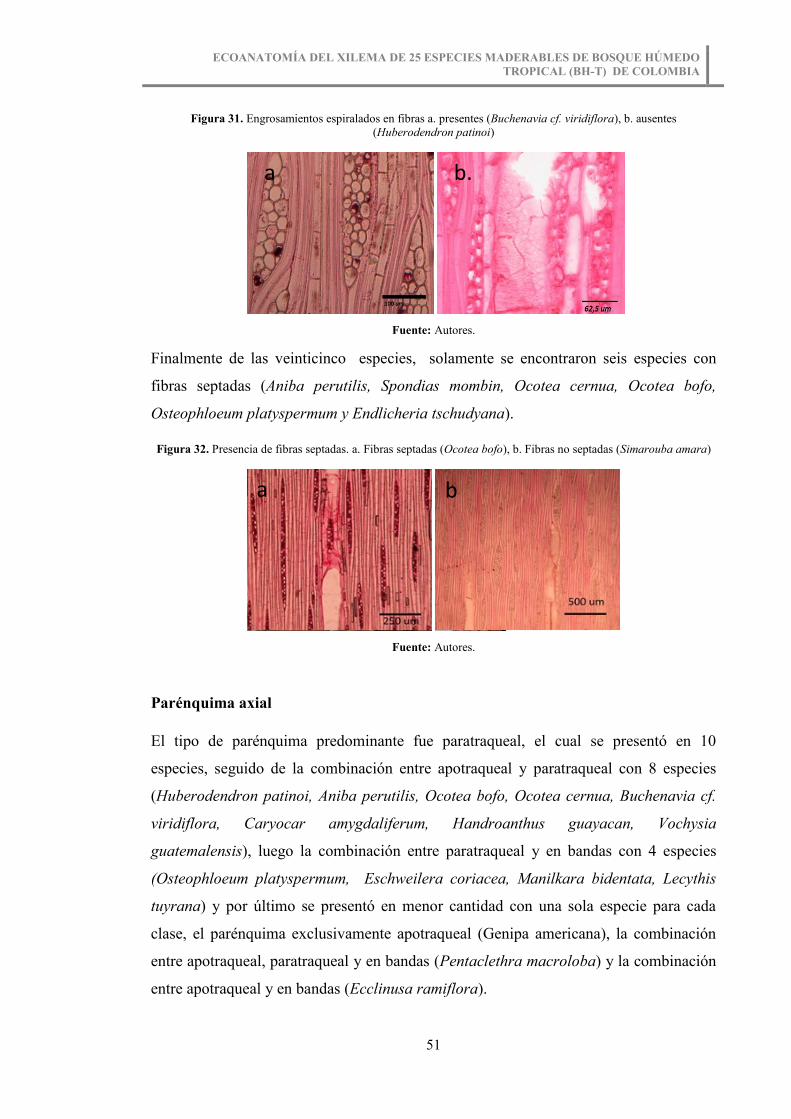
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
51
Figura 31. Engrosamientos espiralados en fibras a. presentes (Buchenavia cf. viridiflora), b. ausentes
(Huberodendron patinoi)
Fuente: Autores.
Finalmente de las veinticinco especies, solamente se encontraron seis especies con
fibras septadas (Aniba perutilis, Spondias mombin, Ocotea cernua, Ocotea bofo,
Osteophloeum platyspermum y Endlicheria tschudyana).
Figura 32. Presencia de fibras septadas. a. Fibras septadas (Ocotea bofo), b. Fibras no septadas (Simarouba amara)
Fuente: Autores.
Parénquima axial
El tipo de parénquima predominante fue paratraqueal, el cual se presentó en 10
especies, seguido de la combinación entre apotraqueal y paratraqueal con 8 especies
(Huberodendron patinoi, Aniba perutilis, Ocotea bofo, Ocotea cernua, Buchenavia cf.
viridiflora, Caryocar amygdaliferum, Handroanthus guayacan, Vochysia
guatemalensis), luego la combinación entre paratraqueal y en bandas con 4 especies
(Osteophloeum platyspermum, Eschweilera coriacea, Manilkara bidentata, Lecythis
tuyrana) y por último se presentó en menor cantidad con una sola especie para cada
clase, el parénquima exclusivamente apotraqueal (Genipa americana), la combinación
entre apotraqueal, paratraqueal y en bandas (Pentaclethra macroloba) y la combinación
entre apotraqueal y en bandas (Ecclinusa ramiflora).
a
a
b.
b

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
52
Figura 33. Tipos de parénquima. a. Parénquima apotraqueal difuso y en agregados (Genipa americana), b.
Parénquima aliforme confluente y aliforme de ala ancha y corta,(Mora oleifera), c. Parénquima en bandas finas
(Manilkara bidentata), d. Parénquima reticulado (Lecythis tuyrana), e. Parénquima de ala fina y extendida
(Jacaranda copaia), f. Parénquima apotraqueal difuso y paratraqueal unilateral (Buchenavia cf. Viridiflora).
Fuente: Autores.
En cuanto al tipo de células presentes en el parénquima axial, es predominante el
parénquima en serie de cinco a ocho células (once especies), en menor proporción el
parénquima en serie de tres a cuatro células con siete especies (Pentaclethra macroloba,
Huberodendro patinoi, Aniba perutilis, Ocotea cernua, Abarema jupunba, Vitex
floridula y Eschweilera coriacea), el parénquima en serie de más de ocho células con 5
especies (Ecclinusa ramiflora, Caryocar amygdaliferum, Jacaranda copaia, Ocotea
cymbarum, Manilkara bidentata) y por último el parénquima en serie de dos células con
2 especies (Handroanthus guayacan y Dipteryx oleifera).
Radios
Los radios presentan un ancho entre una y tres células principalmente con trece
especies, siente especies presentan radios biseriados (Ocotea bofo, Endlicheria
tschudyana, Mora oleifera, Osteophloeum platyspermum, Caryocar amygdaliferum,
Aniba perutilis y Ocotea cernua), tres especies presentan radios multiseriados (Vochysia

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
53
guatemalensis, Spondias mombin y Lecythis tuyrana) y dos especies presentan radios
uniseriados (Pentaclethra macroloba y Abarema jupunba).
Figura 34. Ancho de los radios. a. Radios biseriados (Mora oleifera), b. Radios multiseriados (Vochysia
guatemalensis), c. Radios uniseriados (Abarema jupunba), d. Radios de 1 a 3 células (Eschweilera coriacea).
Fuente: Autores.
En cuanto a la composición de los radios, diez especies presentan radios
heterocelulares tipo IV (células del cuerpo del radio procumbentes con 2-4 rutas de
células decumbentes), ocho especies (Abarema jupunba, Pentaclethra macroloba,
Buchenavia cf. viridiflora, Handroanthus guayacan, Dipteryx oleifera, Eschweilera
coriácea, Endlicheria tschudyana, Vitex floridula) presentan radios homocelulares tipo I
(todas las células procumbentes), 6 especies (Osteophloeum platyspermum, Aniba
perutilias, Ocotea cernua, Simarouba amara, Lecythis tuyrana, Mora oleifera) poseen
radios heterocelulares tipo III (células del cuerpo del radio procumbentes con 1 ruta de
células decumbentes) y una especie (Manilkara bidentata) presenta radios
heterocelulares tipo VI (células procumbentes y decumbentes mezcladas en el cuerpo
del radio).
b.
c.
d.
a.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
54
Figura 35. Tipo de radios. a. Radio tipo I (Vitex floridula), b. Radio tipo III (Osteophloeum platyspermum), c. Radio
tipo VI (Manilkara bidentata).
Fuente: Autores.
No se observaron variantes en las células radiales de veinte especies, mientras que dos
especies (Huberodendron patinoi y Endlicheria tschudyana) presentan células radiales
perforadas, dos especies (Pentaclethra macroloba y Genipa americana) presentan
parénquima radial disyuntivo y una especie (Ecclinusa ramiflora) presenta células de
tipo baldosa.
Figura 36. Modificaciones en células radiales a. Células radiales perforadas (Huberodendron patinoi), b. Células tipo
baldosa (Ecclinusa ramiflora).
Fuente: Autores.
Por último, las punteaduras radiovasculares predominantes son las similares a las
intevasculares, presentándose en diez especies, seguidas de las punteaduras en dos
tamaños con nueves especies (Buchenavia cf. viridiflora, Endlicheria tschudyana, Vitex
floridula, Osteophloeum platyspermum, Ocotea cernua, Ocotea bofo, Virola flexuosa,
Caryocar amygdaiferum y Ecclinusa ramiflora), las punteaduras con borde reducido
aparentemente simple rendondeadas a angulares con cuatro especies (Eschweilera
coriacea, Spondias mombin, Ocotea cymbarum y Huberodendron patinoi) y por último
a.
a.
b.
b.
c.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
55
las punteaduras con borde muy reducido con abertura horizontal y vertical en dos
especies (Aniba perutilis y Manilkara bidentata).
Figura 37. Tipo de punteaduras radiovasculares. a. Similares a las intervasculares (Simarouba amara), b. de dos
tamaños (Virola flexuosa), c. Con borde reducido aparentemente simple rendondeadas a angulares (Spondias
mombin), d. Con borde muy reducido con abertura horizontal y vertical (Aniba perutilis)
Fuente: Autores.
Estructuras estratificadas
Del total de las especies, veintidós de ellas no presentan ningún tipo de estructura
estratificada; mientras que Handroanthus guayacan y Dipteryx oleifera presentan radios
estratificados y Simarouba amara presenta fibras, radios y parénquima estratificado.
Figura 38. Estructuras estratificadas. a. Radios estratificados (Handroanthus guayacan), b. Fibras, radios y
parénquima estratificado (Simarouba amara).
Fuente: Autores.
a.
a.
b.
b.
d.
c.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
56
Elementos secretores y Variantes del cambium
No se presentan elementos secretores ni variantes del cambium dieciocho especies,
Ocotea cernua, y Osteophloeum platyspermum presentaron tubos laticíferos,
Huberodendron patinoi presentó canales intercelulares y Aniba perutilis, Buchenavia
viridiflora, Ocotea bofo, Ocotea cernua y Ocotea cymbarum presentaron ceulas
oliferas.
Figura 39. Tubo laticífero. (Osteophloeum platyspermum).
Fuente: Autores.
Inclusiones minerales
En las especies Abarema jupunba, Mora oleifera, Jacaranda copaia, Manilkara
bidentata, Caryocar amygdaliferum, Huberodendron patinoi, Pentaclethra macroloba,
Dipteryx oleifera, Spondias mombin y Ocotea cymbarum se presentan cristales
prismáticos. Por otra parte, de las 25 especies estudiadas, el sílice se encuentra presente
en Lecythis tuyrana, Vochysia guatemalensis, Eschweleira coriacea, Vitex floridula,
Osteophloeum platyspermum, Virola flexuosa, Ecclinusa ramiflora, Ocotea cernua,
Huberodendron patinoi, Dipteryx oleifera y Ocotea cymbarum.
Figura 40. Inclusiones minerales. a. Cristales prismáticos (Caryocar amygdaliferum), b. Sílice (Eschweilera
coriacea).
Fuente: Autores.
a. b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
57
RASGOS ANATÓMICOS CUANTITATIVOS DE LA MADERA
En la Tabla 4 se presentan los resultados de media, desviación estándar y coeficiente de
variación para las características cuantitativas de los rasgos anatómicos de la madera:
Vasos (diámetro, longitud, frecuencia y diámetro de punteaduras intervasculares), fibras
(grosor y longitud) y radios (altura y número de radios por mm lineal).De acuerdo a esta
información se puede observar que se presenta una alta dispersión en las variables
frecuencia de poros, grosor de fibras y frecuencia de radios con un coeficiente de
variación de 81%, 52% y 52% respectivamente. Estos resultados se pueden observar
con mayor detalle en el Anexo 2 donde se muestra cada variable por especie y su
respectiva media, desviación estándar, mínimo, máximo y coeficiente de variación.
Tabla 4. Características cuantitativas de los vasos, fibras y radios de las 25 especies estudiadas (estadísticos de
media, desviación y coeficiente de variación)
Nombre Científico
Vasos Fibras Radios
Diá
met
ro (
µm
)
Lo
ng
itu
d (
µm
)
Fre
cuen
cia
(po
ros/
mm
2)
Diá
met
ro p
un
ted
ura
s
inte
rva
scu
lare
s (µ
m)
Gro
sor
(µm
)
Lo
ng
itu
d (
µm
)
Alt
ura
(µ
m)
Nú
mer
o (
Ra
d/m
m)
Endlicheria tschudyana 145,09 435,72 12,50 6,54 2,10 1271,57 527,67 4,40
Ecclinusa ramiflora 173,82 349,36 6,60 5,84 4,02 769,14 416,99 16,10
Pentaclethra macroloba 202,73 301,87 3,60 3,74 3,65 1063,38 158,93 10,50
Huberodendron patinoi 308,90 400,21 2,00 7,28 5,91 796,19 603,14 5,50
Aniba perutilis 137,10 410,74 9,60 7,78 1,43 1043,21 354,42 5,60
Jacaranda copaia 231,65 467,43 2,00 3,16 2,31 1029,88 388,84 5,30
Abarema jupunba 216,33 325,57 3,60 3,35 2,71 969,22 191,09 8,00
Dipteryx oleífera 167,60 314,91 7,00 8,30 3,29 887,76 234,09 6,80
Buchenavia cf. Viridiflora 167,08 465,73 11,20 10,94 5,67 909,14 487,51 7,40
Caryocar amygdaliferum 252,67 618,00 3,40 9,18 2,64 1159,81 638,60 19,80
Eschweilera coriácea 300,18 385,82 7,30 7,10 3,32 710,36 479,41 9,30
Spondias mombin 156,31 357,98 13,70 9,76 1,19 888,13 583,50 3,80
Ocotea cymbarum 186,42 524,47 5,90 8,95 2,18 1154,18 365,19 4,90
Genipa americana 121,45 498,33 23,60 3,35 5,94 1301,21 498,30 7,50
Ocotea bofo 161,08 732,42 9,50 10,41 2,36 1336,37 465,76 4,10
Ocotea cernua 179,09 584,06 7,60 7,57 2,05 1138,32 360,13 4,30
Vochysia guatemalensis 347,31 327,93 3,40 6,44 5,55 891,00 729,10 4,30
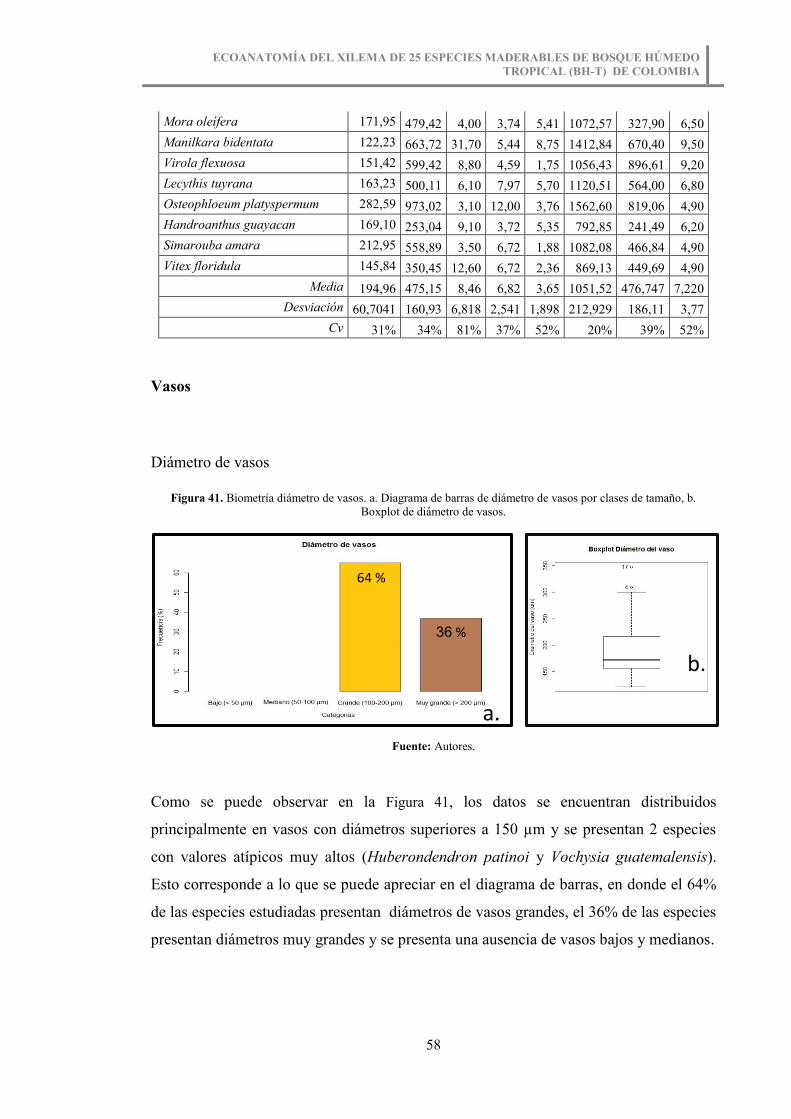
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
58
Mora oleífera 171,95 479,42 4,00 3,74 5,41 1072,57 327,90 6,50
Manilkara bidentata 122,23 663,72 31,70 5,44 8,75 1412,84 670,40 9,50
Virola flexuosa 151,42 599,42 8,80 4,59 1,75 1056,43 896,61 9,20
Lecythis tuyrana 163,23 500,11 6,10 7,97 5,70 1120,51 564,00 6,80
Osteophloeum platyspermum 282,59 973,02 3,10 12,00 3,76 1562,60 819,06 4,90
Handroanthus guayacan 169,10 253,04 9,10 3,72 5,35 792,85 241,49 6,20
Simarouba amara 212,95 558,89 3,50 6,72 1,88 1082,08 466,84 4,90
Vitex floridula 145,84 350,45 12,60 6,72 2,36 869,13 449,69 4,90
Media 194,96 475,15 8,46 6,82 3,65 1051,52 476,747 7,220
Desviación 60,7041 160,93 6,818 2,541 1,898 212,929 186,11 3,77
Cv 31% 34% 81% 37% 52% 20% 39% 52%
Vasos
Diámetro de vasos
Figura 41. Biometría diámetro de vasos. a. Diagrama de barras de diámetro de vasos por clases de tamaño, b.
Boxplot de diámetro de vasos.
Fuente: Autores.
Como se puede observar en la Figura 41, los datos se encuentran distribuidos
principalmente en vasos con diámetros superiores a 150 µm y se presentan 2 especies
con valores atípicos muy altos (Huberondendron patinoi y Vochysia guatemalensis).
Esto corresponde a lo que se puede apreciar en el diagrama de barras, en donde el 64%
de las especies estudiadas presentan diámetros de vasos grandes, el 36% de las especies
presentan diámetros muy grandes y se presenta una ausencia de vasos bajos y medianos.
64 %
36 %
a.
b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
59
Longitud de vasos
Figura 42. Biometría longitud de vasos. a. Diagrama de barras de longitud de vasos por clases de tamaño, b. Boxplot
de longitud de vasos.
Fuente: Autores.
La Figura 42b permite apreciar que la distribución de las longitudes de los vasos es
asimétrica a la izquierda, se observa un valor atípico que se refiere a la longitud máxima
registrada, la cual corresponde a la especie Osteophloeum platyspermum. Por su parte,
en la Figura 42a se observa que la mayoría de los vasos presentan longitudes
consideradas como medianas con un 72% y un 24% con longitudes menores a 350 µm,
es decir vasos cortos.
Frecuencia de vasos
Figura 43. Biometría frecuencia de vasos. a. Diagrama de barras de frecuencia de vasos, b. Boxplot de frecuencia de
vasos.
Fuente: Autores.
En relación a la frecuencia de los vasos, en la Figura 43 se observa que los datos son
poco dispersos, ya que el 92% de los datos se encuentran entre 0 y 15 vasos/mm2
aproximadamente, también se puede observar la presencia de 2 datos atípicos máximos,
que corresponden a las especies (Genipa americana y Manilkara bidentata). Por
24 %
36 %
72 %
56 %
4 %
8 %
a.
a.
b.
b.
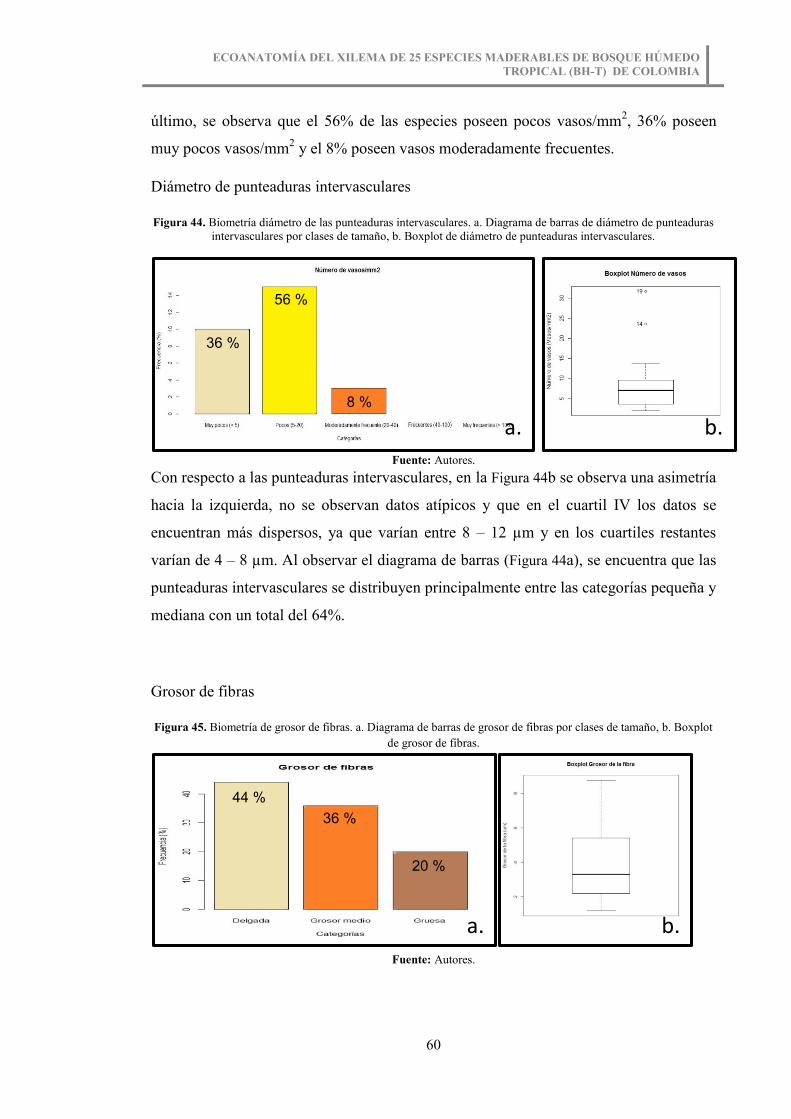
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
60
último, se observa que el 56% de las especies poseen pocos vasos/mm2, 36% poseen
muy pocos vasos/mm2 y el 8% poseen vasos moderadamente frecuentes.
Diámetro de punteaduras intervasculares
Figura 44. Biometría diámetro de las punteaduras intervasculares. a. Diagrama de barras de diámetro de punteaduras
intervasculares por clases de tamaño, b. Boxplot de diámetro de punteaduras intervasculares.
Fuente: Autores.
Con respecto a las punteaduras intervasculares, en la Figura 44b se observa una asimetría
hacia la izquierda, no se observan datos atípicos y que en el cuartil IV los datos se
encuentran más dispersos, ya que varían entre 8 – 12 µm y en los cuartiles restantes
varían de 4 – 8 µm. Al observar el diagrama de barras (Figura 44a), se encuentra que las
punteaduras intervasculares se distribuyen principalmente entre las categorías pequeña y
mediana con un total del 64%.
Grosor de fibras
Figura 45. Biometría de grosor de fibras. a. Diagrama de barras de grosor de fibras por clases de tamaño, b. Boxplot
de grosor de fibras.
Fuente: Autores.
36 %
44 %
56 %
36 %
8 %
20 %
a.
a.
b.
b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
61
En la Figura 45b, se observa que los datos agrupados en el tercer y cuarto cuartil
presentan una alta dispersión y por consiguiente una asimetría hacia la derecha, no se
encuentran valores atípicos para el grosor de la pared celular de las fibras y el rango de
los grosores varía entre 1 – 9 µm. Por otro lado, en la Figura 45a, se observa que la
mayoría de las especies poseen pared celular delgada con un 44%, que un 36% poseen
pared mediana y el 20% restante poseen pared gruesa.
Longitud de fibras
Figura 46. Biometría de longitud de fibras. a. Diagrama de barras de longitud de fibras por clases de tamaño, b.
Boxplot de longitud de fibras.
Fuente: Autores.
En cuanto a la longitud de la fibra, la Figura 46b permite apreciar que los datos se
encuentran distribuidos entre 600 y 1600 µm aproximadamente con una asimetría hacia
la izquierda, se observa un dato atípico que se refiere a la longitud máxima de las 25
especies, la cual corresponde a la especie Osteophloeum platyspermum. Al observar la
Figura 46a, se puede apreciar que la mayoría de las especies poseen fibras categorizadas
con largo mediano con un 68% y que el 32% restante poseen fibras cortas.
32 %
68 %
a. b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
62
Altura de radios
Figura 47. Biometría de altura de radios. a. Diagrama de barras de altura de radios por clases de tamaño, b. Boxplot
de altura de radios.
Fuente: Autores.
De acuerdo a la Figura 47b, la altura de los radios posee un comportamiento ligeramente
asimétrico a la derecha, el bigote de la derecha es más largo que el de la izquierda, lo
cual sugiere que los datos se encuentran más dispersos a la derecha que a la izquierda.
En la Figura 47a, la altura de los radios se concentra en la clase pequeña con un 64% y el
36% restante pertenece a la clase mediana.
Frecuencia de radios
Figura 48. Biometría de frecuencia de radios. a. Diagrama de barras de frecuencia de radios, b. Boxplot de frecuencia
de radios.
Fuente: Autores.
Desde el punto de vista de la frecuencia de los radios, en la Figura 48b se observa una
baja dispersión de los datos y una asimetría hacia la derecha, en donde adicionalmente
se observan dos datos atípicos, que corresponden a las especies Ecclinusa ramiflora y
Caryocar amygdaliferum, también se puede observar que la mayor parte de los datos se
encuentran en un rango entre 5 y 8 radios/mm2, lo cual corresponde a las frecuencias
64 %
64 %
36 %
24 %
a.
12 %
b.
a. b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
63
presentadas en la Figura 48a, en donde el 64% de las especies tiene radios
moderadamente numerosos y el 24% radios numerosos.
ÍNDICES DE VULNERABILIDAD (IV) Y MESOMORFÍA (IM)
En la Tabla 5 se muestran los resultados obtenidos para los índices de vulnerabilidad y
mesomorfía de las especies estudiadas.
Tabla 5. Índices de vulnerabilidad y mesomorfía para las especies objeto de estudio.
Nombre Científico Índice de
vulnerabilidad
Índice de
mesomorfía
Endlicheria tschudyana 11,607 5057,589
Ecclinusa ramiflora 26,337 9201,181
Pentaclethra macroloba 56,313 16999,553
Huberodendron patinoi 154,452 61813,538
Aniba perutilis 14,281 5865,890
Jacaranda copaia 115,825 54139,936
Abarema jupunba 60,091 19564,272
Dipteryx oleífera 23,943 7539,897
Buchenavia cf. Viridiflora 14,918 6947,948
Caryocar amygdaliferum 74,313 45925,751
Eschweilera coriácea 41,120 15865,118
Spondias mombin 11,409 4084,367
Ocotea cymbarum 31,596 16571,469
Genipa americana 5,146 2564,414
Ocotea bofo 16,956 12418,889
Ocotea cernua 23,564 13763,008
Vochysia guatemalensis 102,150 33497,605
Mora oleífera 42,988 20609,244
Manilkara bidentata 3,856 2559,101
Virola flexuosa 17,207 10314,094
Lecythis tuyrana 26,759 13382,755
Osteophloeum platyspermum 91,157 88697,434

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
64
Handroanthus guayacan 18,583 4702,120
Simarouba amara 60,842 34003,869
Vitex floridula 11,575 4056,391
Media 42,280 20405,817
Desviación 38,839 21570,571
Cv 91,9% 105,7% Fuente: Autores.
Figura 49. Índice de vulnerabilidad. a. Histograma de índice de vulnerabilidad, b. Boxplot de índice de
vulnerabilidad.
Fuente: Autores.
En la Tabla 5 se puede observar una gran dispersión entre los valores de Vulnerabilidad
obtenidos para las especies objeto de estudio ya que se encuentran en un rango de 3,8 a
154. A pesar de ello, todas presentaron un comportamiento homogéneo de tipo
mesormórfico, lo que significa que son vulnerables al estrés hídrico y que no tienen
características anatómicas que les permitan resistir a condiciones de sequía. Como se
puede observar en la Figura 49b dentro de los datos obtenidos solo se encuentra un valor
atípico que corresponde a la especie Huberodendron patinoi la cual tiene un índice de
Vulnerabilidad de 154,45 por lo que se puede considerar la especie con mayor eficiencia
en la conducción de agua, dentro de las especies estudiadas y con menor resistencia al
estrés hídrico.
a.
b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
65
Figura 50. Índice de mesomorfía. a. Histograma de índice de mesormorfía, b. Boxplot de índice de mesormorfía.
Fuente: Autores.
En cuanto al Índice de mesormorfía en la Tabla 5 se puede observar que existe una gran
dispersión entre los datos obtenidos con un coeficiente de variación del 105,7%, ya que
se encuentran en un rango de 2259 a 88697. Al igual que el Índice vulnerabilidad, este
presenta un comportamiento homogéneo, ya que, de acuerdo con Carlquist (1977) las
especies que tengan IM superior a 200, tienen un comportamiento mesofítico. Teniendo
en cuenta la Figura 50b, se observan 4 datos atípicos que se refieren a los valores
máximos de IM obtenidos, y que corresponden a las especies Osteophloeum
platyspermum, Huberodendron patinoi, Jacaranda copaia y Caryocar amygdaliferum,
las cuales tienen características anatómicas que facilitan la conducción, como vasos
grandes, baja frecuencia de vasos y vasos largos.
Correlación de los rasgos anatómicos cuantitativos
Tabla 6. Matriz de correlación de rasgos anatómicos cuantitativos.
Fuente: Autores.
En la Tabla 6 se muestra la correlación existente entre las variables cuantitativas de las
especies evaluadas, se resalta la relación existente entre la longitud de fibra y la longitud
a.
b.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
66
de vaso, en donde existe una relación positiva con un valor de 0,82 y a medida que los
vasos son más largos la longitud de las fibras será mayor. Adicionalmente, se resalta la
relación negativa entre diámetro de vaso y frecuencia de vasos con un valor de -0,66 en
donde a medida que los vasos son más grandes la frecuencia es menor.
Análisis de componentes principales
Mediante el análisis de componentes principales se espera encontrar cuales son las
variables que explican en mayor medida el comportamiento de las especies y así
concluir cuáles son las variables más representativas para este estudio. Se presenta un
análisis entre las variables diámetro de vasos, longitud de vasos, frecuencia de vasos,
diámetro de punteaduras intervasculares, grosor de la fibra, longitud de la fibra, y altura
y frecuencia de radios. Los números al interior del cuadro indican las especies referidas
en la Tabla 1.
Figura 51. Análisis de componentes principales para variables cuantitativas.
Fuente: Autores.
De acuerdo a la Figura 51, el diámetro de vaso, la frecuencia de vaso, longitud del vaso y
longitud de fibra son las variables que más explican el comportamiento de las especies,
por el contrario las variables frecuencia de radio, altura de radio, grosor de fibra y

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
67
diámetro de las punteaduras intervasculares son poco representativas para explicar el
comportamiento de las especies.
En cuanto a las variables representativas se observa que la variable diámetro de vaso
esta inversamente relacionada con la variable frecuencia de vaso y la variable longitud
de vaso está directamente relacionada con la variable longitud de fibra, tal como se
observó en la matriz de correlación (Tabla 6) en dónde se observó que estas variables
tuvieron la mayor correlación con -0,66 y 0,82 respectivamente.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
68
ANÁLISIS DE RESULTADOS
Para el análisis de resultados cuantitativos, éstos se compararon con los valores
obtenidos para las características anatómicas de la madera en diferentes tipos de
bosques, reportados por otros autores (Barajas-Morales, 1985; Pérez, 1989; Lindorf,
1994; Aguilar-Rodríguez & Barajas-Morales, 2005; León, 2005; Polanco & Grande,
2009: Cosmo, 2012; Vásquez, 2013 y León, 2013). Los valores más descritos en otras
investigaciones son el diámetro de vasos, longitud de vasos, frecuencia de vasos,
diámetro de punteaduras intervasculares, longitud de fibras, altura de radios y
frecuencia de radios, en cuanto al grosor de la fibra no se realizó comparación debido a
que se encontraron clasificaciones distintas y no se pudieron establecer parámetros para
su comparación, adicionalmente el análisis de componentes principales demostró que
esta no es una variable representativa para establecer el comportamiento de las especies
del bosque húmedo tropical y como lo mencionan León & Espinoza (2001), a pesar de
que esta característica es importante para realizar estudios comparativos, no es
importante para el estudio de la relación de la anatomía de la madera con la ecología.
Es importante aclarar, que en los estudios empleados para la comparación, no siempre
se encontraban todas las variables que se evaluaron en el presente estudio y por lo tanto
no todas las variables fueron comparadas con el mismo número de ecosistemas.
A continuación se presentan los estudios y las respectivas convenciones empleadas en
los gráficos realizados para las comparaciones de las variables:
Tabla 7. Estudios revisados y nomenclatura empleada en el presente estudio.
Tipo de Bosque Autor Convención
Bosque muy húmedo premontano. Barajas-Morales, 1985 Bmhp-B
Bosque muy seco tropical Barajas-Morales, 1985 BmsT-B
Bosque húmedo premontano Pérez, 1989 Bhp-P
Bosque muy seco tropical Lindorf, 1994 BmsT-L
Bosque de niebla Aguilar-Rodríguez & Barajas-Morales, 2005 Bn-AB
Bosque seco tropical León, 2005 BsT-L
Bosque altoandino Polanco & Grande, 2009 Baa-PG
Bosque húmedo premontano Cosmo, 2012 Bhp-C
Bosque húmedo tropical León, 2013 BhT-L
Transición seco a húmedo León, 2013 (Datos sin publicar) Btsh-L
Bosque altoandino Vásquez, 2013 Baa-V
Bosque húmedo tropical Castrillón & Pinzón, 2015. BhT-CP
Fuente: Autores.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
69
Vasos
Diámetro de vaso
Figura 52. Diagrama de barras para diámetro de vasos, en diferentes ecosistemas.
Fuente: Autores.
El diámetro de vasos pudo ser comparado con 10 ecosistemas como se observa en la
Figura 52. Se determinó que existen semejanzas entre los datos reportados en este
estudio y los reportados por Cosmo (2012) para bosque húmedo premontano, León
(2005) para bosque seco tropical y León (2013) para bosque húmedo tropical, en donde
se encontró que los diámetros de los vasos son principalmente grandes y muy grandes,
es decir que son superiores a 100 µm, lo cual según Carlquist (1977) es una
característica de la madera orientada a la eficiencia en el transporte del agua. Se observa
que esta es una característica que predomina en las especies de ecosistemas con
condiciones de humedad favorables. La semejanza que se presenta con el bosque seco,
puede asociarse a que en esa área de estudio la precipitación se encuentra entre 1400 y
1500 mm, la cual no representa un elevado déficit hídrico para las plantas que implique
arriesgar la eficiencia y adaptar sus estructuras anatómicas a condiciones de sequía.
En comparación a zonas con menor disponibilidad hídrica como el bosque muy seco
tropical estudiado por (Barajas-Morales,1985; Lindorf, 1994) los diámetros de los
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
70.00%
80.00%
90.00%
Diámetro de vasos
Bajo(menor 50 um) Mediano(50-100) Grande (100-200) Muy grande (más de 200)
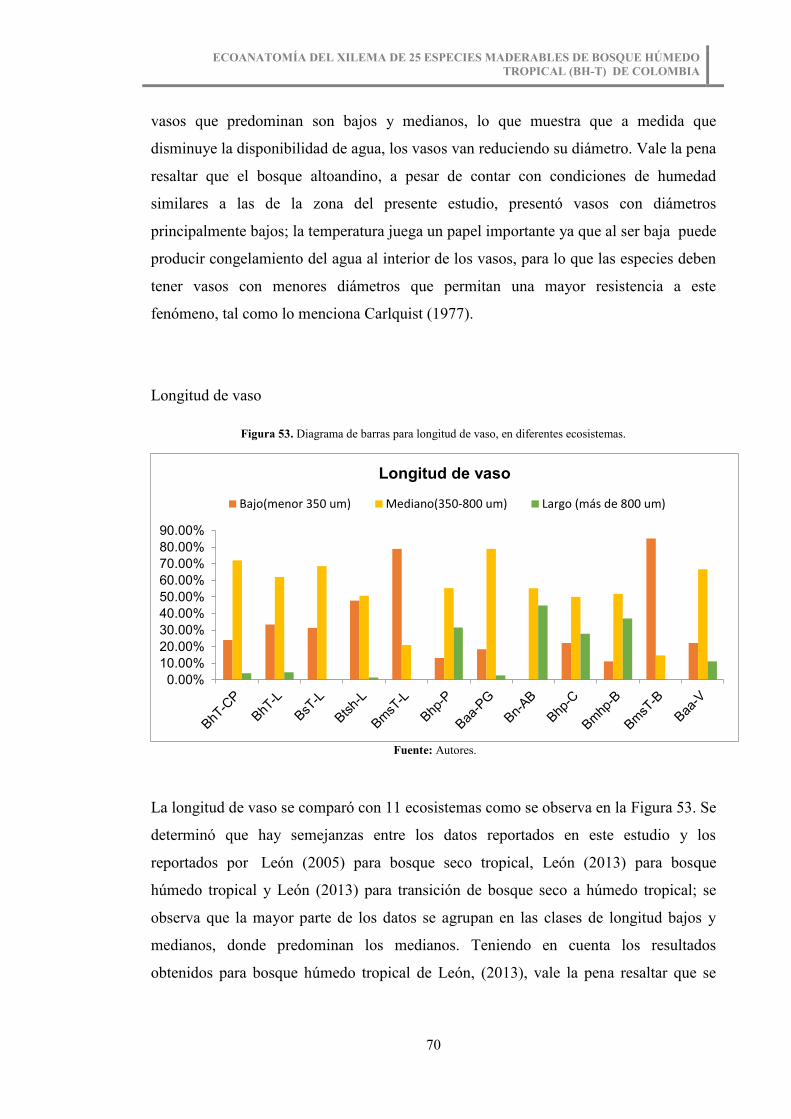
ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
70
vasos que predominan son bajos y medianos, lo que muestra que a medida que
disminuye la disponibilidad de agua, los vasos van reduciendo su diámetro. Vale la pena
resaltar que el bosque altoandino, a pesar de contar con condiciones de humedad
similares a las de la zona del presente estudio, presentó vasos con diámetros
principalmente bajos; la temperatura juega un papel importante ya que al ser baja puede
producir congelamiento del agua al interior de los vasos, para lo que las especies deben
tener vasos con menores diámetros que permitan una mayor resistencia a este
fenómeno, tal como lo menciona Carlquist (1977).
Longitud de vaso
Figura 53. Diagrama de barras para longitud de vaso, en diferentes ecosistemas.
Fuente: Autores.
La longitud de vaso se comparó con 11 ecosistemas como se observa en la Figura 53. Se
determinó que hay semejanzas entre los datos reportados en este estudio y los
reportados por León (2005) para bosque seco tropical, León (2013) para bosque
húmedo tropical y León (2013) para transición de bosque seco a húmedo tropical; se
observa que la mayor parte de los datos se agrupan en las clases de longitud bajos y
medianos, donde predominan los medianos. Teniendo en cuenta los resultados
obtenidos para bosque húmedo tropical de León, (2013), vale la pena resaltar que se
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
70.00%
80.00%
90.00%
Longitud de vaso
Bajo(menor 350 um) Mediano(350-800 um) Largo (más de 800 um)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
71
evidencia una mayor semejanza, ya que en ambos se presentan vasos largos, es decir
con longitudes mayor a 800 µm, manteniéndose la mayor representatividad en vasos
medianos.
Por el contrario, en ecosistemas con condiciones de déficit hídrico como el bosque muy
seco tropical estudiado por Barajas-Morales (1985) y Lindorf (1994), los vasos se hacen
más cortos ya que esta característica contribuye a la seguridad en la conducción del
agua y otorga mayor resistencia a la especies ante el estrés hídrico.
En comparación, zonas con temperaturas inferiores a las del área del presente estudio
como bosque muy húmedo premontano (Barajas-Morales, 1985), bosque húmedo
premontano (Pérez, 1989), bosque de niebla (Aguilar-Rodríguez & Barajas-Morales,
2005), bosque húmedo premontano (Cosmo, 2012), y bosque altoandino (Vásquez,
2013) mostraron vasos más largos y medianos, lo cual puede indicar que las bajas
temperaturas contribuyen a la adaptación de estructuras anatómicas con fines de
garantizar la eficiencia de la conducción del agua ya que a medida que el vaso es más
largo, la condición mesofítica de la especie es mayor (Carlquist, 1977).
Frecuencia de vaso
Figura 54. Diagrama de barras para poros/mm2 para diferentes ecosistemas.
Fuente: Autores.
La frecuencia de vasos se comparó con 9 ecosistemas como se observa en la Figura 54.
Se observa una semejanza entre los datos reportados en el presente estudio y los datos
0.00%
20.00%
40.00%
60.00%
80.00%
100.00%
BhT-CP BhT-L BsT-L Btsh-L BmsT-L Bhp-P Baa-PG Bn-AB Bhp-C Bmhp-B
Frecuencia de poros
muy pocos (menos de 5) pocos(5-20)
moderadamente frecuente (20-40) frecuentes(40-100)
muy frecuentes (más de 100)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
72
reportados por Barajas-Morales (1985) para bosque muy húmedo premontano, León
(2005) para bosque seco tropical y León (2013) para bosque húmedo tropical, la cual
consiste en que la frecuencia de los vasos es principalmente de pocos a muy pocos, es
decir, menor a 20 vasos/mm2. Se observa que estos ecosistemas, son los mismos que
presentaron vasos de diámetros grandes; esto se explica con lo encontrado en la Figura
51 en donde se evidencia una correlación inversa entre el diámetro de los vasos y la
frecuencia de los mismos. Debido a esto, es de esperarse que los valores de índice de
vulnerabilidad de las especies de estos ecosistemas sean altos y por consiguiente
presentan comportamiento mesofítico en donde su sistema de conducción esté orientado
hacia la eficiencia con poca resistencia al estrés hídrico.
En comparación a zonas con menor disponibilidad hídrica como el bosque muy seco
tropical estudiado por Barajas-Morales (1985) y Lindorf (1994), se observó que las
especies estudiadas por Lindorf (1994) presentaron poros más frecuentes por mm2,
mientras que las especies estudiadas por Barajas-Morales (1985) presentaron poros
principalmente poco frecuentes. Debido a esto se puede decir que no sólo la
precipitación y la temperatura influencian los rasgos anatómicos de las especies en un
ecosistema, sino que existen otras variables que propician la adaptación de las especies,
por ejemplo, la latitud como lo mencionan Montaño et al., (2011).
Diámetro de punteaduras intervasculares
Figura 55. Diagrama de barras para el diámetro de las punteaduras intervasculares para diferentes ecosistemas.
Fuente: Autores.
0.00%
20.00%
40.00%
60.00%
80.00%
BhT-CP BsT-L Baa-V
Diámetro de las punteaduras Intervascualres
Diminutas (menos de 4 um) Pequeñas (4-7 um)
Medianas (7-10 um) Grandes (con más de 10 um)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
73
El diámetro de punteaduras se comparó con 2 ecosistemas como se observa en la Figura
55. Los datos del presente estudio presentaron una semejanza con los reportados para
bosque seco tropical por León (2005) ya que el patrón de distribución en las clases de
tamaño se comporta en forma de campana en ambos casos, presentándose en todas las
clases de tamaño. Por el contrario, para el bosque altoandino estudiado por Vásquez
(2013), las especies presentaron diámetros de punteaduras exclusivamente pequeños y
medianos. Sin embargo hay que tener en cuenta que en el análisis de componentes
principales se determinó que esta variable no es representativa para explicar el
comportamiento de las especies en el ecosistema, confirmando lo que expresa León
(2005), que el tamaño de las punteaduras intervasculares se encuentra principalmente
relacionado con el grupo taxonómico al que pertenecen las especies más que con el sitio
en el que se desarrollan.
Longitud de fibra
Figura 56. Diagrama de barras para la longitud de la fibra en diferentes ecosistemas.
Fuente: Autores.
La longitud de fibra se comparó con 5 estudios como se puede observar en la Figura 56.
Se observa que el ecosistema que presentó longitudes de fibras similares (fibras
medianas y cortas) a las del presente estudio fue el bosque muy seco tropical estudiado
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
70.00%
80.00%
BhT-CP Baa-PG Bn-AB Bmhp-B BmsT-B Baa-V
Longitud de fibra
Bajo (menor 900) Mediano (900 - 1600) Largo (más de 1600 um)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
74
por Barajas-Morales (1985). En este caso se observó que las similitudes no
corresponden a ecosistemas con condiciones ambientales similares a las del área de
estudio. Por el contrario en las variables evaluadas para los elementos vasculares, se
encontró que era precisamente con el ecosistema evaluado por Barajas-Morales (1985),
que se presentaban mayores diferencias. Esto demuestra que no todas las características
anatómicas son afectadas de igual forma por las condiciones ambientales (precipitación
y temperatura) como se observó en los elementos vasculares, ya que estos se encuentran
afectados por la disponibilidad de agua debido a la función que desempeñan dentro de la
planta, mientras que las fibras pueden estar más influenciadas por otras condiciones de
acuerdo a la función de soporte que cumplen dentro de la misma.
Para la longitud de fibras, Montaño et al., (2011) señalan que esta es afectada por la
altitud (msnm) de la zona donde crecen las especies y a medida que la altitud es mayor
la longitud de las fibras disminuye. Sin embargo como se puede ver en la Figura 56. no
se observa claramente esta tendencia entre fibras y altitud, ya que no en todos los
bosques con altitudes bajas predominan o se presentan fibras largas y no en todos los
bosques con altitudes altas predominan o se presentan fibras cortas.
Altura de radios
Figura 57. Diagrama de barras para la altura del radio en diferentes ecosistemas.
Fuente: Autores.
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
70.00%
BhT-CP Baa-PG Bmhp-B BmsT-B Baa-V
Altura de radios
Pequeño (<500 µm) Mediano (500 - 1000 µm) Grande (>1000 µm)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
75
La altura de radios se comparó con 4 estudios como se puede observar en la Figura 57.
Se observa que al igual que los radios de las especies evaluadas en este estudio, los
ecosistemas de bosque altoandino estudiado por Polanco & Grande, (2009) y bosque
muy seco tropical estudiado por Barajas-Morales (1985) presentaron radios
principalmente pequeños es decir, menores a 500 µm. En cuanto esta variable no se
observa una relación por condiciones ambientales (precipitación, temperatura, altitud),
ya que estos bosques presentan características ambientales diferentes, por lo que estas
no pueden el explicar el comportamiento de los radios en las especies presentes en un
determinado ecosistema, es posible que este rasgo este más afectado por características
propias de la especie, o por la combinación de otros factores que no fueron evaluados en
el presente estudio.
Es difícil caracterizar la altura de radios incluso en los mismos tipos de bosque ya que el
estudio realizado por Vásquez (2013), se realizó en un bosque altoandino al igual que el
estudio de Polanco & Grande (2009) y estos no presentaron similitudes entre sí; el
primero presentó altura de radios principalmente pequeños, mientras que el segundo
presentó radios en las tres clases de altura (pequeños, medianos y grandes).
Frecuencia de radios
Figura 58. Diagrama de barras para radios/mm en diferentes ecosistemas.
Fuente: Autores.
0.00%
20.00%
40.00%
60.00%
80.00%
BhT-CP Bmhp-B BmsT-B Baa-V
Frecuencia de radios
Poco numerosos (< 4 radios/mm)
Moderadamente numerosos (4 - 6 radios/mm)
Numerosos (7-9 radios/mm
Muy numerosos (>10 radios/mm)

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
76
En cuanto a la frecuencia de los radios, se realizó una comparación con tres estudios
como se presenta en la Figura 58. Esta variable no presentó similitudes notables en su
comportamiento con respecto a los ecosistemas de bosque muy húmedo premontano y
bosque muy seco tropical estudiados por Barajas-Morales (1985) y bosque altoandino
estudiado por Vásquez (2013); ya que para el presente estudio predominan frecuencias
de radios moderadamente numerosos, mientras que en los otros estudios se presentan
radios desde poco numerosos hasta muy numerosos en proporciones similares.
Como se pudo observar, esta variable no proporciona información valiosa con respecto
al comportamiento de los rasgos anatómicos de las especies en función de las
características ambientales; esto se respalda de acuerdo al análisis de componentes
principales (Figura 51), el cual permitió determinar que esta es la variable menos
representativa para explicar el comportamiento de las especies.
Índice de vulnerabilidad (IV) e Índice de mesomorfía (IM)
Los índices de vulnerabilidad y mesomorfía para las especies estudiadas, representan
una anatomía orientada a la eficiencia del transporte del agua, lo que muestra su
adaptación al hábitat de bosque húmedo tropical y las condiciones de humedad que este
presenta. De acuerdo con León (2005), mientras más se aleja el Índice de vulnerabilidad
del valor correspondiente a la unidad, mayor será la orientación de la especie a
condiciones que garanticen la eficiencia de la conductividad en sacrificio de la
seguridad. El 8% de las especies tienen un IV menor a 10 pero mayor que 1, lo que
significa una eficiencia a la conducción ligada a la seguridad y el 92% de las especies
restantes mostraron IV superiores a 10 lo que significa más eficiencia en la conducción
del agua.
Estas especies presentaron en su mayoría poros grandes, frecuencias de poros bajas y
vasos largos por lo que es de esperarse que los valores de índice de vulnerabilidad de las
especies de estos ecosistemas sean altos y por consiguiente presenten comportamiento
mesofítico en donde su sistema de conducción esté orientado hacia la eficiencia con
poca resistencia al estrés hídrico.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
77
Lo anterior concuerda con los resultados obtenidos por Parra (2010) en los que
concluyó que las maderas que crecen en ecosistemas húmedos tienen varias
características que van orientadas hacia la eficiencia del transporte de agua como son:
diámetro de los vasos medianos, frecuencia de poros baja, longitud de los elementos
vasculares medianamente largos, diámetros de punteaduras intervasculares grandes y
platinas de perforación simples; además con los obtenidos por León (2013) en un
bosque húmedo tropical de Venezuela donde, los índices de vulnerabilidad y
mesomorfía indicaron comportamientos mesomórficos enfocados a la eficiencia y con
los obtenidos por Rodríguez (2012), en un bosque húmedo tropical en el medio Atrato,
donde también se presentaron índices de vulnerabilidad y mesomorfía que indican
especies con un comportamiento mesomórfico y por lo tanto un sistema de conducción
orientado hacia la eficiencia.
Variabilidad de caracteres anatómicos de acuerdo a factores climáticos
De acuerdo a los resultados obtenidos se observó que el diámetro de vaso, la longitud de
vaso, la frecuencia de vaso y la longitud de fibras son las características anatómicas
más influenciadas por condiciones ambientales como se puede observar en las Figuras
52, 53 y 54. Igualmente, Barajas-Morales (1985), Carlquist (1977), León (2005), Moya
& Tomazello (2008), Chávez-Romero et al., (2010), Montaño et al., (2011), Montaño-
Arias et al., (2013) y Meena & Gupta (2014) entre otros determinaron que son
precisamente estos elementos los que más varían de acuerdo a las condiciones
climáticas como precipitación, temperatura y altitud.
Esto puede explicarse debido a que los vasos son los encargados de la conducción del
agua por lo que la disponibilidad de está implica la adaptación de las estructuras
anatómicas de las especies para sobrevivir a diferentes condiciones como lo mencionan
Norman et al., (2013).
Teniendo en cuenta estos resultados es posible realizar una aproximación de la
tendencia de la variación de los elementos anatómicos en función de las condiciones
climáticas.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
78
Figura 59. Variación de los elementos anatómicos. a. Variación de acuerdo a la precipitación. b. Variación de
acuerdo a la temperatura.
Fuente: Autores.
El diámetro de los vasos aumenta a medida que aumenta la precipitación y la
temperatura en la zona donde se desarrolla la planta. Esto se evidenció en este estudio y
en los ecosistemas húmedos y cálidos estudiados por Cosmo (2012) y León (2013) ya
que las condiciones humedad son favorables y por lo tanto la estructuras se adaptan para
que el transporte del agua sea más eficiente. Esto concuerda con lo mencionado por
Castro et al., (1998) y Carlquist (1977) quienes dicen que los diámetros grandes se
encuentran en localidades húmedas. Mientras que en los ecosistemas secos estudiados
por Barajas-Morales (1985) y Lindorf (1994) y los ecosistemas fríos estudiados por
Aguilar-Rodríguez & Barajas-Morales (2005), Polanco & Grande (2009) y Vásquez
(2013) el diámetro de los vasos es menor.
Teniendo en cuenta la relación inversa que existe entre el diámetro de vasos y la
frecuencia de vasos es de esperarse que a medida que la precipitación y la temperatura
de la zona en la que se desarrolle la planta sean altas la frecuencia de los vasos
disminuya, y por el contrario cuando la precipitación y la temperatura sean bajas la
frecuencia de los vasos aumente.
La temperatura influencia la longitud del vaso en la medida en que, esta al ser menor, la
longitud del elemento vascular aumenta y viceversa como se puede observar al
comparar los ecosistemas cálidos estudiados por Barajas-Morales (1985), Lindorf
(1994), León (2005) y León (2013) donde se presentan vasos más cortos, con los
ecosistemas fríos estudiados por Aguilar-Rodríguez & Barajas-Morales (2005), Polanco

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
79
& Grande (2009) y Vásquez (2013) donde se presentan vasos más largos. Por el
contrario cuando la precipitación de un ecosistema es baja la longitud del elemento
vascular disminuye como en el caso de los ecosistemas muy secos estudiados por
Barajas-Morales (1985) y Lindorf (1994) y cuando la precipitación es alta, la longitud
del elemento vascular aumenta como se puede observar en el presente estudio y en los
ecosistemas húmedos estudiados por Barajas-Morales (1985), Cosmo (2012) y León
(2013).
Figura 60. Variación de la longitud de la fibra de acuerdo a la altitud
Fuente: Adaptado de Montaño et al. (2011)
Montaño et al., (2011) mencionan que existe una relación inversa entre la altitud y la
longitud de fibra por lo que a medida que la altitud es mayor se presentan fibras más
cortas y viceversa. Sin embargo, en los resultados obtenidos no se pudo evidenciar
claramente esta relación.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
80
CONCLUSIONES
Los rasgos anatómicos cuantitativos que más se ven afectados por las condiciones
ambientales (precipitación, temperatura, altitud) de una zona son aquellos que están
relacionados a los elementos vasculares, tales como: diámetro, longitud y frecuencia y
la longitud de la fibra.
Los rasgos anatómicos que no presentaron una relación evidente con las condiciones
ambientales fueron: el diámetro de la punteadura intervascular, el grosor de la pared
celular de la fibra y la altura y frecuencia de los radios.
Se determinó que existe una relación positiva entre la longitud de fibra y la longitud de
vaso, por lo que a medida que los vasos son más largos la longitud de las fibras será
mayor.
Existe una correlación negativa entre el diámetro y la frecuencia de los poros, por lo que
a mayor diámetro de poros se presentan frecuencias bajas, lo cual indica que las plantas
poseen una mayor eficiencia en la conducción del agua.
Las especies de bosque húmedo tropical tienen un comportamiento mesomórfico y de
vulnerabilidad al estrés hídrico, con características anatómicas orientadas a la eficiencia
del transporte del agua, lo que muestra su adaptación a las condiciones de humedad que
el ecosistema presenta.
Las rasgos anatómicos cuantitativos (diámetro de vasos, longitud de vasos, frecuencia
de vasos, diámetros de las punteaduras intervasculares) de la madera de bosque húmedo
tropical presentan un comportamiento similar al de bosque seco tropical debido a que el
bosque seco tropical no posee un alto déficit hídrico ya que tienen precipitaciones
entre 1000 - 2000 mm.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
81
RECOMENDACIONES
El grosor de las fibras no presenta parámetros de clasificación que permitan realizar
comparaciones con la mayoría de los estudios, ya que en algunos se presentan de
manera cualitativa y en otros de manera cuantitativa; por lo tanto se recomienda
establecer un parámetro de clases de tamaño que permita realizar comparaciones en
futuros estudios.
Se recomienda que para futuros estudios ecoanatómicos se verifique la importancia de
evaluar las variables grosor de fibra, altura de radios y frecuencia de radios, ya que en el
presente estudio dichas variables no se encontraron afectadas en un alto grado por las
condiciones ambientales.
Se recomienda realizar estudios con una mayor cantidad de especies en diferentes zonas
de vida con el fin de determinar de forma más precisa el comportamiento anatómico de
las especies dependiendo de la ecología. Teniendo en cuenta la diversidad de cada
ecosistema.
Se recomienda incluir variables como el suelo, gradientes de temperatura, latitud,
factores endógenos, distribución geográfica u otras que se consideren pertinentes.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
82
AGRADECIMIENTOS
Alejandra Castrillón Arboleda
Agradezco a Dios por permitirme llegar a este momento tan especial en mi vida. Por los triunfos
y los momentos difíciles que me han enseñado a valorar todo cada día más.
A mis padres Alba Arboleda y Carlos Castrillón por ser los promotores de mis sueños, gracias
por su esfuerzo y apoyo en mi formación como profesional; por sufrir conmigo en la
elaboración de trabajos interminables, en los días de parciales y en las entregas de notas donde
no se sabía si pasaba o no, por festejar las buenas notas, los reconocimientos, el fin de cada
semestre y cada buena noticia que les contaba. ¡¡¡¡Gracias infinitas!!!!.
A mis hermanos Omar, Luci, Adrián, Diana y Jimmy por su apoyo incondicional y motivación
para seguir adelante y lograr el objetivo trazado para un futuro mejor y ser orgullo para ellos y
toda la familia.
A la Universidad Distrital Francisco José de Caldas y al proyecto Curricular de Ingeniería
forestal porque nos está formando como excelentes profesionales.
A la docente Nancy pulido, por su guía durante toda la investigación; por compartir sus
conocimientos, su sentido de responsabilidad y rigor académico, por su paciencia y por todos
sus aportes a mi formación en el maravilloso mundo de las maderas.
A los jurados Mario León e Ingrid Amórtegui por sus valiosos aportes durante este proceso, sus
recomendaciones fueron invaluables en la elaboración del documento.
Lorraine gracias por tu apoyo y amistad. Dios permitió que hiciéramos este trabajo juntas para
aprender y conocer muchas cosas nuevas y poder terminar con éxito nuestras carreras. Sin ti,
esto no sería lo mismo.
A mis compañeros y amigos Jully Forero, Lorena Pérez, Loren Baquero, Karen Amaya, Esteban
Moreno, por compartir sus conocimientos, alegrías y tristezas y a todas aquellas personas que
durante estos 6 años estuvieron a mi lado apoyándome y lograron que este sueño se haga
realidad, a pesar de que por momentos parecía una tarea titánica interminable.
Por último y no por eso menos importante, a una persona muy especial que ha estado
apoyándome incondicionalmente en todo momento, que me ha dado fuerzas para seguir
adelante cuando pensaba que no podía, por eso y más, Rayid Rodríguez, Gracias.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
83
Lorena Pinzón Castillo
A Dios, quien me permitió culminar esta etapa y conseguir alcanzar tan anhelada meta.
A mis padres José Pinzón y Tilcia Castillo, por su constante apoyo durante toda mi vida
y en esta importante etapa, por ser mi fuerza y darme ánimo y consuelo en los
momentos más difíciles con sus palabras y su amor y a quienes debo todo lo que soy. Al
igual que a mis hermanos José, Leidy y Angel por acompañarme y apoyarme en los
momentos difíciles y compartir conmigo los momentos felices. En especial a mi
hermana gemela Bibiana por ser mi compañía en los mejores y peores momentos y por
ser mi más grande apoyo y estar siempre conmigo. También quiero agradecer a mis
sobrinos Esteban, Jonathan y Sarah por su amor y ser mi motivación muchas veces.
A todos mis amigos dentro y fuera de las aulas de clase, quienes hicieron de mi paso por
la universidad una etapa inolvidable, por los gratos momentos que vivimos y el apoyo
recibido durante estos años.
En especial a mi compañera de tesis y amiga Alejandra Castrillón, por contar con su
apoyo constante, por entenderme, soportarme durante este largo trabajo, por escucharme
y ser más que mi compañera en los días de trabajo. Muchas gracias por todo.
También a mis amigas Lorena Pérez por su amistad y cariño, por estar ahí siempre y ser
la compañera de noches en vela haciendo trabajos excelentes y Jully Forero por su
especial amistad durante toda la carrera y por su comprensión y cariño en esta
importante etapa.
A mis amigos, Vanesa Velásquez, Óscar Rozo, Julián Piraján y Óscar Gómez, con
quienes no compartí muchos momentos académicos pero que hicieron de la universidad
un sitio más agradable en esos momentos de ocio y que me dieron su apoyo cuando lo
necesitaba.
A la Universidad Distrital Francisco José de Caldas y en especial al proyecto curricular
de Ingeniería Forestal por contribuir a mi formación profesional.
A mis profesores, por los conocimientos compartidos y sus valiosas enseñanzas en el
ámbito profesional. En especial a los docentes Nancy Pulido y Mario León y a la
ingeniera Ingrid Amórtegui por su importante contribución en el desarrollo de este
trabajo.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
84
BIBLIOGRAFÍA
Aguilar-Rodríguez, S. & Barajas-Morales, J. (2005). Anatomía de la madera de
especies arbóreas de un bosque mesófilo de montaña: un enfoque ecológico-evolutivo.
Boletín de la Sociedad Botánica de México 77: 51-58.
Alcaldía de Vigia del Fuerte. (2006). Plan de Gestión Integral de residuos sólidos del
municipio de Vigia del Fuerte. pp 52-54
Alonso, J. (2011). Manual de Histología Vegetal. Ediciones Mundi-Prensa. España.
Araque, A., & León, W. (2006). Anatomía Comparada del Leño de Spondias mombin
L. (ANACARDIACEAE) que crece en Zonas de Banco y Bajío de la Reserva Forestal
Caparo (Barinas, Venezuela). Revista Forestal Venezolana. 50(1): 9-17.
Argollo, J., Soliz, C., & Villalba, R. (2004). Potencialidad dendrocronológica de
Polylepistarapacana en los Andes Centrales de Bolivia. Ecología en Bolivia. 39(1): 5–
24.
Barajas-Morales, J. (1985). Wood structural differences between trees of two tropical
forests in Mexico. IAWA Bulletin. 6 (4): 355-364.
Barrios, N., & Rondón, D. (2014). Anatomía de la Madera de tres especies de
ERICACEAE en el Parámo de la Culata, Mérida, Venezuela. Pittieria. 38 (1): 135–146.
Carlquist, S. (1977). Ecological factors in wood evolution: A floristic approach.
American Journal of Botany. 64(7): 887-896.
Carlquist, S. (1988). Comparative Wood anatomy, systematic, ecological and
evolutionary aspects of dicotyledonous wood.Springer-Verlag, Berlín, Alemania.
Carrillo, I., Elissetche, J., Valenzuela, S., & Teixeira, R. (2013). Formación de
elementos anatómicos en maderas duras: Una revisión desde una perspectiva genómica.
Maderas, ciencia y tecnología. 15(1):93-104.
Castro, D., Villar, S., Guerrero-Campo, J., Pérez, R. y Marti, G. (1998).
Variaciones morfoanatómicas en las comunidades de encinar lo largo de un gradiente
climático en el NE de la Península Ibérica. Acta Botánica de Barcelona 45: 577-586.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
85
Chávez-Rómero, R., Aguilar-Rodríguez, R., & Terrazas, T. (2010). Variación
anatómica en la madera de Quercus obtusata (Fagaceae). Madera y Bosques. 16(2): 69-
87
Cosmo, N. (2012). Ecología do lenho de 19 espécies nativas do estado do Paraná. (Tese
apresentada como requisito à obtenção do título de Doutor em Ciências Florestais.)
Universidade Federal do Paraná. 54 p
Gámez, A. (2013). Estudio Ecoanatómico de cuatro especies arbóreas de Malvaceae en
la Estación Experimental Caparo, Estado Barinas (Venezuela). Pittieria. 37: 41-51.
García, L. Guindeo, C. Peraza & Palacios, P. (2003). La madera y su anatomía:
Anomalías y defectos, estructura microscópica de coníferas y frondosas. Asociación de
Investigación Técnica de las Industrias de la Madera y Corcho. Editorial Mundi –
Prensa. Madrid.
Guridi, L. (1977). Metódo de Ablandamiento de Madera Dura y Muy Dura para la
obteción de cortes en xilotomo. Ciencia Forestal. 2(10):59 - 64
Huertas, C. (1963). Anatomía de la madera de 12 especies de coníferas de Mexicanas
(Secretaría de Agricultura y Recursos Hidráulicos). Insitituto Nacional de
Investigaciones Forestales de México. Vol 8.
IAWA Committee. (1989). IAWA list of microscopic features for hardwood
identification. IAWA Bull. n.s. 10 (3): 221–332.
Instituto de Investigaciones en Geociencia, Minería y Química. (1989). Mapa
Geológico de Antioquia. Escala 1.000.000. Bogotá.
León, W. (2005). Anatomía ecológica del xilema secundario de un bosque seco tropical
de Venezuela. Acta Botánica Venezuelica. 28 (2) : 257 – 273.
León, W. (2013). Ecoanatomía de la madera en un bosque húmedo tropical de la
Reserva Forestal Ticoporo (Barinas, Venezuela). Revista Forestal Venezolana. 57:(2).
115-129.
León, W., & Espinoza, N. (2001). Anatomía de la madera. Universidad de los Andes,
Consejo de publicaciones. Mérida, Venezuela.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
86
Liconln, T., & Zeiger, E. (2006). Fisiología Vegetal. Castello de la Plana. Publicación
de la Universitat Jaume.
Lindorf, H. (1994). Eco-Anatomical Wood Features of Species from a Very Dry
Tropical Forest. IAWA Journal. 15(4): 361 – 376.
Luostarinen, K. (2006). Effects of enviromental and internal factors of trees and timber
treatment on colour of dried birch (Betula pendula) Wood. University of Joensuu.
Faculty of Forestry.
Medina, A., Razquin, M., & Andia, I. (2013). Estrategia conductiva del leño de
Nothofagus alpina (Nothofagaceae), cuenca Lacar, Neuquén, Argentina. Bosque
(Valdivia). 34(1):81-88
Meena, V., & Gupta, S. (2014). Wood Anatomy of Albizia procera Correlation
Between Tropical and Subtropical From different Geographical Zones of Indian
Subcontinent. International Journal of scientific & Technology Research. 3(5): 2-18.
Ministerio del Medio Ambiente., Codechoco., IIAP & Municipio de rio Sucio.
(2002). Plan de Ordenamiento Territorial de las tierras Colectivas de las comunidades
negras del bajo Atrato: municipio de Rio Sucio-Bajira-Carmén del Darién. Medellín.
Moglia, J., Giménez, A., González, D., & Gerez, R. (2010). Caracterización de los
anillos de crecimiento y su relación conla densidad básica de la madera en Eucalyptus
camaldulensis. Quebracho. 18 (1):47-57.
Montaño, S., Camargo, S., Grether, R., & Pérez, C. (2011). La Madera en función
del Ambiente: Un análisis ecoanatómico. Elementos: Ciencia y cultura. 18 (83):19-23.
Montaño-Arias, S., Camargo-Ricalde, S., & Pérez-Olvera, C. (2013). Ecoanatomía
de los elementos de vaso de la madera de cinco especies del género Mimosa
(Leguminosae-Mimosoideae). Botanical Sciences. 91(1): 1-10
Moya, R., & Tomazello, M. (2008). Variation in the Wood anatomical structure of
Gmelinaarborea (Verbenaceae) trees at different ecological conditions in Costa Rica.
Revista Biologica Tropical. 56 (2): 689-704

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
87
Nuñez, C. (2005). Programa de Investigación de Celulosa y Papel (PROCYP). Facultad
de Ciencias Exactas, Químicas y Naturales. Universidad Nacional de Misiones.
República Argentina. Disponible en http://www.aulavirtual-
exactas.dyndns.org/claroline/backends/download.php?url=L01hZGVyYV9wdWxwYV
95X3BhcGVsL0FuaWxsb3NfYW51YWxlc195X2RlX2NyZWNpbS4ucGRm&cidRese
t=true&cidReq=INDEPa643. Visto el 03 de febrero de 2015.
OCHA, United nations office fot the cordinattion of humanitarian affairs. (2003).
Mapa físico del Chocó. Escala 1:1315000.
Palacios, P. s.f. Biogeografía. Instituto de investigaciones científicas SINCHI –
Universidad Nacional de Colombia. Disponible en :
http://www.virtual.unal.edu.co/cursos/sedes/leticia/80123/lecciones/cap3/leccion7.html
#veintiun. Visto el 4 de febrero de 2015
Panshin, A. & Zeeuw, C. (1980). Textbook of wood technology. Structure,
identification, properties, and uses of the commercial woods of the United States and
Canada, Editorial McGraw-Hill.New York
Parra, J. (2010). Determinación de los índices de vulnerabilidad y mesormorfía en
especies de Laurales de la selva San eusebio (Merida, Venezuela). Pittieria 34: 13-22.
Pérez, M. (1989). Caracterización ecoanatómica del leño de 40 especies del bosque La
Mucuy , Estado de Mérida, Venezuela. Revista Forestal Venezolana. 33: 43-51.
Polanco, C., & Grande, D. (2009). Análisis ecoanatómico, evolutivo y comparativo de
la madera de 40 especies de dos asociaciones del bosque alto Andino Colombiano.
Revista Colombia Forestal 12: 183-203.
Prida, A., Boulet, J., Ducousso, A., Nepveu, G., & Puech, J. (2006). Effect of species
and ecological conditions on ellagitannin content in oak wood from an evenaged and
mixed stand of QuercusroburL. and QuercuspetraeaLiebl.
Pulido, E., Mateus, D., & Lozano, I. (2011). Anatomía xilemática de
CAESALPINIACEAE registradas en la xiloteca de la Universidad Distrital Francisco
José de Caldas. Colombia Forestal. 14(2): 145 -171.

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
88
R Core Team. (2014). R: A language and environment for statistical computing.
Vienna, Austria: R Foundation for Statistical Computing. ISBN: 3-900051-07-0,
recuperado de http://www.R-project.org.
Rasband, W. (2014). ImageJ, U. S. National Institutes of Health, Bethesda, Maryland,
USA, http://imagej.nih.gov/ij/, 1997-2014.
Richter, H.G., & Dallwitz, M. (2000). Onwards. Commercial timbers: descriptions,
illustrations, identification, and information retrieval. In English, French, German,
Portuguese, and Spanish. Version: 25th June 2009. http://delta-intkey.com’.
Rodríguez, I. (2012). Determinación de rasgos Funcionales del xilema en especies
forestales tropicales de la cuenca media del Rio Atrato, Colombia. Tesis de grado para
optar al título de Ingeniero Forestal. Universidad Distrital Francisco José de Caldas.
Bogotá D.C.
Sacarello, M. (2010). LA MADERA, Desde su conocimiento a la conservación.
Editorial Gente común. Bolivia.
Safdari, V. & Devall, M. (2012). Identification of important Iranian Hardwoods by
Vessel-Ray Pits and Vessel Element Shapes.(MacerationProcess). Lignocellulose.
1(1):55-70.
Sandoval, E. (2005). Técnicas aplicadas al estudio de la anatomía vegetal. Universidad
Nacional Autónoma de México. Primera edición. México D.F., México. 281pp
Terrazas, T., Aguilar-Rodríguez, S. & López-Mata, L. (2008). Wood Anatomy and
Its relation to Plant size and Latitude in Budleja L. (BUDLEJACEAE). Interciencia.
33(1): 46 – 50.
Vásquez, M. (2013). Dinámica y biomasa fustal de un bosque alto andino: Una
aproximación a su función ecológica (Trabajo de pregrado, Ingeniería Forestal). Bogotá:
Universidad Distrital Francisco José de Caldas. 80 p.
Walter, H. (1986). Vegetação e zonas climáticas: tratado de ecologia global. São Paulo.
Ed. Pedagógica e UniversitáriaEPU. 326p

ECOANATOMÍA DEL XILEMA DE 25 ESPECIES MADERABLES DE BOSQUE HÚMEDO
TROPICAL (BH-T) DE COLOMBIA
89
Webster, N., & McKetchine, J. (1980). Webster new twentieth century dictionary of
the English language unabridged. William Collins, 2129 p.
Wheeler, E., Baas, P., & Gasson, P. (1989). IAWA Listof microscopic Features for
hardwood identification. IAWA bulletin n.s. 10(3):219-332.
Zobel, B., & Van Buijtenen, J. (1989). Wood Variation: Its Causes and Control
Springer Verlag. New York.


















