Clase N 9 Citoesqueleto
10
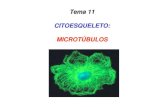
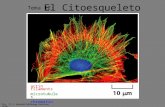
7/25/2019 Clase N 9 Citoesqueleto http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 1/10 CLASE Nº9: CITOESQUELETO Prof. Andrea Paula Lima El Citoesqueleto es la estructura de la célula, el cual le permite desplazarse, y mediar el movimiento de sus organelos intracelularmente. En la imagen, se puede apreciar marcado el núcleo (rosado), los microtúbulos o la tubulina (amarillo) y los microfilamentos de actina (azul). Entre las funciones que cumplen están: Separa los cromosomas durante la mitosis. Divide la célula en dos. Soporta la membrana plasmática y le da el sostén mecánico a la célula. Desplaza a los espermatozoides y fibroblastos. Desplaza a los espermatozoides y fibroblastos. A los fibroblastos y glóbulos blancos les permite pasar entre células Contrae las células musculares. Transporta organelos de un extremo a otro en las células. Extiende axones y dendritas en las células nerviosas. Dirige el crecimiento de la pared celular vegetal. La dinámica o movimiento de las células (como los queratinocitos) se debe a las proteínas estructurales del Citoesqueleto. Al llegar una señal a la célula, se da una despolimerización de proteínas y luego una polimerización de estas hacia el lado donde están los nutrientes; es decir la actividad del Citoesqueleto depende de una despolimerización y una polimerización, estas características son lo que lo permiten ser dinámico. Los tres filamentos que conforman el Citoesqueleto se forman a partir de subunidades, y se estructuran en ensamblajes helicoidales, a través de uniones no covalentes. La asociación de monómeros, por ejemplo de actina, forma un polímero y esa asociación se da gracias a estas uniones no covalentes débiles, las que permiten ensamblarse y desensamblarse fácilmente, es decir debe ser un proceso reversible. Existen tres tipos de filamentos en el Citoesqueleto, los cuales poseen funciones muy relacionadas. Sin embargo se puede decir que, los filamentos de actina determinan la forma de la superficie celular, y son fundamentales en la locomoción. Luego existen los filamentos de tubulina, o microtúbulos los cuales cumplen funciones como posicionar los organelos de membrana, como por ejemplo el retículo y las vesículas, y dirigir el transporte intracelular. Y por ultimo los filamentos intermedios que proporcionan la fuerza y resistencia al estrés mecánico.
-
Upload
valentina-aichele-petridis -
Category
Documents
-
view
223 -
download
0
Transcript of Clase N 9 Citoesqueleto

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 1/10
CLASE Nº9: CITOESQUELETOProf. Andrea Paula Lima
El Citoesqueleto es la estructura de la célula, el cual le permite desplazarse, y
mediar el movimiento de sus organelos intracelularmente. En la imagen, sepuede apreciar marcado el núcleo (rosado), los microtúbulos o la tubulina(amarillo) y los microfilamentos de actina (azul). Entre las funciones quecumplen están:
Separa los cromosomas durante la mitosis. Divide la célula en dos. Soporta la membrana plasmática y le da el sostén mecánico a la
célula. Desplaza a los espermatozoides y fibroblastos.
Desplaza a los espermatozoides y fibroblastos. A los fibroblastos y glóbulos blancos les permite pasar entre células Contrae las células musculares. Transporta organelos de un extremo a otro en las células. Extiende axones y dendritas en las células nerviosas. Dirige el crecimiento de la pared celular vegetal.
La dinámica o movimiento de las células (como los queratinocitos) se debea las proteínas estructurales del Citoesqueleto. Al llegar una señal a lacélula, se da una despolimerización de proteínas y luego una polimerización de estas hacia el lado donde están los nutrientes; es decir la actividad del
Citoesqueleto depende de una despolimerización y una polimerización,estas características son lo que lo permiten ser dinámico. Los tres filamentos
que conforman el Citoesqueleto se forman a partir de subunidades, y se
estructuran en ensamblajes helicoidales, a través de uniones no covalentes. Laasociación de monómeros, por ejemplo de actina, forma un polímero y esaasociación se da gracias a estas uniones no covalentes débiles, las quepermiten ensamblarse y desensamblarse fácilmente, es decir debe ser unproceso reversible.
Existen tres tipos de filamentos en el Citoesqueleto, los cuales poseen
funciones muy relacionadas. Sin embargo se puede decir que, los filamentos
de actina determinan la forma de la superficie celular, y son fundamentales
en la locomoción. Luego existen los filamentos de tubulina, o microtúbulos loscuales cumplen funciones como posicionar los organelos de membrana, como
por ejemplo el retículo y las vesículas, y dirigir el transporte intracelular. Y
por ultimo los filamentos intermedios que proporcionan la fuerza y resistencia
al estrés mecánico.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 2/10
Carmen María Pirela
Page 2
Los microfilamentos están ensamblados a partir de monómeros de actina,que son muy finos teniendo un diámetro de alrededor de 7 nm, en las célulasdel intestino se les puede observar en las microvellosidades. Los filamentosintermedios por otro lado, están formados por un tipo de proteína, a pesarde que exista una gran variedad proteínas que pueden componer losfilamentos intermedios (laminina, vimentina, etc), asociándose y formándoseun filamento de 10nm de diámetro. Y los microtúbulos tienen una formahueca y esta formado por dos proteínas la alpha tubulina y la betatubulina, ambas son tubulinas con ligeras diferencias, determinando laasociación de dímeros de tubulina una estructura de este filamento de25nm de diámetro, y su organización es a través de un centro organizadorde microtúbulos. Dependiendo de la célula los filamentos que componen elcitoesqueleto suelen variar, por ejemplo los microfilamentos de actina enlas células intestinales se encuentran a nivel de las microvellosidadesformando cilios, sin embargo en células como espermatozoides, esosmismos cilios son de microtúbulos.
MICROFILAMENTOS
Son estructuras polares ¿Qué quiere decir? Que en uno de los extremoshay una polimerización rápida mientras que en el otro hay unadespolimerización. De una manera general el proceso de polimerizaciónde actina empieza con una fase de nucleacion, que es a través de tresmonómeros, en los cuales se hace un gasto de energía muy grande para
lograr estabilizarlos. Luego de la formación del trímero, la primeraasociación de actina es mucho más energéticamente favorable, por lo quela incorporación de nuevos monómeros de actina sucede de una formamucho más rápida.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 3/10
Carmen María Pirela
Page 3
En términos gráficos se tiene el número de monómeros y filamentos deactina (eje Y o ordenada) y el tiempo de adhesión de una sal que catalizala formación de los núcleos. Hay una etapa en la cual no hay ningúnmonómero, pero luego que se da la nucleacion de tres monómero deactina, la curva se hace exponencial y por un lado se va a tender a
adicionar monómeros y por el otro lado una perdida de ellos. Laformación de microfilamentos consiste en la nucleacion y la adición demonómeros para la formación de filamentos, hasta que se llega a un platoen el cual esta en un estado critico, en el cual hay una concentración alta y
el polímero favorece la adhesión por un lado y la separación por otra, de
manera que es un equilibrio, de un lado la constante se da para polimerizar(ATP) y otro por despolimerización (ADP).
Esta característica de polaridad le permite desplazarse a la célula,incorporando monómeros a sus filamentos, a aquellas direcciones a lascuales debe movilizarse. El equilibrio de polimerización- despolimerización
da como resultado un lado del filamento que es positivo, es decir para ATPdonde la constante de asociación de los monómeros es mucho mayor , y unlado negativo para ADP. A medida que el polímero crece y se le
adicionan moléculas de actina con ATP, se van hidrolizando aquellas quevan quedando posteriores quedando ADP. Proteínas que controlan la
dinámica del citoesqueleto de actina pueden mediar ese intercambio de ATP a
ADP, las proteínas unidas a alguno de los extremos, modulando lapolimerización de actina, al modificar la estructura del monómero o nopermiten el recambio de ATP por ADP.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 4/10
Carmen María Pirela
Page 4
Durante la hidrólisis de ATP se libera energía la cual puede ser utilizadapara generar el movimiento de determinadas bacterias y virus, porejemplo la listeria que es un parasito intracelular que en su maquinariamolecular permite modificar la polimerización de moléculas de actina,permitiendo la liberación de ATP y utilizar esa energía para desplazarsedentro de la célula.
DESPLAZAMIENTO CELULAR
El cual también tiene que ver con los microfilamentos, y hay que hablar delamelipodios. Los lamelipodios presentan un sustrato que es lapolimerización del citoesqueleto de actina, lo que hace que la célula“camine” o se desplace, por otro lado a través de la proteína Miosina 2 lacual es una proteína motora de los microfilamentos, se permite laretracción de la célula del lado contrario a la polimerización o movimientode la célula. Molecularmente se ve como prolongación de fibras hacia
adelante, y la retracción de la parte trasera de la célula, en esa retracción se
permite la separación de la célula y el sustrato.
El mismo citoesqueleto de actina puede formar distintos polímeros, y seorganización desde lo más externo hasta las más internas, de tal forma
que las fibras que van en sentido paralelo y muy juntos, están másexternamente y son llamados filopodios, más internamente también seorganizan de forma entrecruzada o lamelipodio (con ángulos de 90º). Ymás cerca del núcleo están las fibras de estrés, las cuales son hacescontráctiles y con organización antiparalela, pudiendo moverse uno sobrelos otros. Cuando la célula se mueve, los organelos también se desplazanhacia donde la célula se dirige, y esto se debe a que en cierta forma losmicrofilamentos están asociados a la tubulina
PROTEÍNAS REGULADORAS
Existen varias proteínas, como la cofilina que promueve la disociación en elextremo negativo, la serverina forma un “capuchón” del lado positivo
impidiendo la polimerización. El cap Z que se encuentra en el sarcomero,limitando los extremos de hasta donde queda la actina, conformando lalínea Z del sarcomero.
[Nota: no es importante saber cuales son las proteínas, pero si saber comose modifica el citoesqueleto en la perdida o agregación de ellas]

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 5/10
Carmen María Pirela
Page 5
También existen proteínas que ayudan al ensamblaje por ejemplo la filamina,
que promueve el entrecruzamiento de actina en los lamelipodios, dando una
solubilidad de gel en esa parte del citoplasma. ¿Si se inactiva la filaminaque sucede? No se forman entrecruzamiento, no se forman loslamelipodios y se pierde esa estructura de gel, por lo tanto la célula no semueve. Ahora, la formación de haces paralelos y muy junto se asocian afimbrina, que los mantiene bastante unidos, mientras que la Alpha actinina,relacionada con la fibra contráctil, se dan haces antiparalelos más
separados.
Las proteínas también asocian a la célula desde su interior, con respecto alexterior celular, por ejemplo la integrina, la cual posee una cara citosólicapequeña, un dominio transmembrana y una cara extracelular con undominio unido a las proteínas de la MEC. Por lo que parte deldesplazamiento celular se debe a esta “conversación” con el medio
extracelular, mediada por estos puentes que forma la actina a través dela integrina y sus dominios. Por lo que cuando la actina se asocia conproteínas que aumentan la adhesión con integrina se tienen adhesionesfocales, pero cuando hay una despolimerización mediada por proteínas deactina se pierden esas adhesiones focales. También la célula intestinal, lasmicrovellosidades están representadas por actina unidas a proteínas que sellaman vilina y fibrina
MIOSINAS
Son proteínas que interactúan con los filamentos de actina, y se acoplan ala hidrólisis de ATP para cambios conformacionales, resultantes de losmovimientos de actina-miosina en el sarcomero. A través del cambio de sus
dos cabezas, se asocia a los microfilamentos mediante la hidrólisis de ATP, lo
que luego origina un cambio conformacional de la miosina, recambiando
ADP por ATP soltándose la cabeza desde el monómero, corriendo el
filamento de actina uno sobre el otro. Hay varios tipos de miosinas, quedeterminan que movimientos ocurren con las actinas; por ejemplo lamiosina II promueve el desplazamiento de filamentos pero la V eltransporte de vesículas, y la VI la de la exocitosis, pero siempre utilizandola hidrólisis de ATP y sus dos cabezas. La miosina I tiene una sola cabezaque se asocia a una sola vesícula y desplaza una sola fibra de actina,mientras que la dos tiene dos cabezas y desplaza dos filamentos deactina, por lo que distintas miosinas promueven distintos movimientos.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 6/10
Carmen María Pirela
Page 6
SARCOMERO
Son microfilamentos de actina, que por un lado tienen un CAP-Z que limitaun margen de polimerización. Existen proteínas que conectan el CAP conlas miosinas y cada una de esas moléculas de miosina, asociadas con
microfilamentos de un lado, mientras que del otro extremo se asocian alotro lado del microfilamento. De manera que con la hidrólisis de ATP cada
una de esas cabezas se desplaza hacia el centro del sarcomero,
contrayéndose la célula muscular.
Este proceso es mediado por calcio y ATP, uniéndose a proteínasasociadas a los microfilamentos y la miosinas, promoviendo eldeslizamiento del sarcomero. La importancia del CAPZ o disco Z es que
mantiene todos los elementos o filamentos del sarcomero unidos, de manera
que todos los filamentos de un lado desplazan en el mismo sentido. Del mismomodo no solo se presenta microfilamentos a nivel del sarcomero en lacontracción muscular, si no que también en la división celular; durante la
división se polimerizan de tal forma que estrangulan, formando un anillocontráctil que separa la célula y las divide en dos.
La polimerización se puede modular también mediante drogas, como lafaloidina que estabiliza los filamentos de actina es decir limita elmovimiento celular debido a la rigidez que esta entrega. La citocalasina hace que en los extremos + se detenga la despolimerización, mientras queen los extremos – continua, es decir acorta el movimiento. En generalcuando las drogas estabilizan, no permiten la dinámica de la actina,debido a que no haga ni despolimerización y polimerización.
[Nota: No es necesario aprenderlos, pero si como se relacionan con losprocesos, y como la ausencia de estos puede afectar la polimerización]

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 7/10
Carmen María Pirela
Page 7
MICROTUBULOS
Estos filamentos son muy importantes en la interfase, en las células ciliadas,en la división celular e incluso estructura el axón de las células nerviosas. El
microtubulo también es una estructura polarizada, es decir que tiene un
extremo + y un extremo -, lo que le permite polimerizarse. Se componende dos proteínas, una tubulina alpha y una tubulina beta, formando unheterodímero y la secuencia de estos varios heterodímero conforman un
protofilamento. La agrupación de 13 protofilamentos alineados en paralelo
forma el microtúbulo, el cual tiene un lumen.
También hay una polimerización hacia el extremo positivo, hay unapreferencia en ambos, microtúbulos y microfilamentos, por polimerizarsehacia este extremo, esta asociación es mediada a diferencia de losmicrofilamentos, a través de GTP y GDP. Hay un fenómeno que seinestabilidad dinámica de los microtúbulos en la que en algunos momentos ellado positivo hay una constante de asociación muy baja, por lo que se
tiende a acortar, esto se debe a que la hidrólisis de GTP cambia la subunidad
beta del heterodímero, debilitando sus uniones permitiendo ladespolimerización.
¿De que manera? La subunidad beta posee un sitio de unión a GTP, si setiene uno o varios GTP en el protofilamento, este se mantiene recto. Sinembargo al realizar una hidrólisis, y pasar de GTP a GDP, el monómeroconsigue un cambio en ese protofilamento de manera que la unión entre
los heterodímero se debilita, promoviendo la inestabilidad del microtúbulo,permitiendo que en el extremo + haya perdida de polimerización.¿Cuándo ocurre? En la división celular, cuando los cromosomas se unen alos microtúbulos, pero estos para separarlos deben acortarse, es decir sedespolimerizan desde su lado positivo. De manera que si la adición deldímero es rápida se polimeriza mientras que si es lenta se vadespolimeriza.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 8/10
Carmen María Pirela
Page 8
PROTEÍNAS REGULADORAS
Entre ellas se encuentran la TAU y la MAP2, esa asociación tiende aestabilizar el microtubulo, lo cual en la fibra nerviosa le da más
estabilidad.
CENTROSOMA
Es el conjunto de un par de centriolos, los cuales son cuerpos basales para los
cilios y flagelos de las células móviles, y una matriz que se asocia a los
centros negativos del microtubulo. De manera que los lados positivosquedan externos al centrosoma, y los negativos están anclados haciadentro, lo que permite que exista una polimerización y no unadespolimerización. Cuando se duplican los centrosomas permite que hayan
dos centros organizadores, a partir de los que parte el crecimiento de los
microtúbulos, pero gracias a su inestabilidad corta promueve que la se de
la despolimerización de lado positivo y vayan arrastrando de cada ladolos cromosomas. Hay que saber también que si en una célula que poseeestos microtúbulos en forma de astro, se corta en una parte el centrosomay en el otro quedan los microtúbulos de forma libre, rápidamente se formaun centro organizador de microtúbulos en el fragmento.
De manera que los centrosomas promueven muchas actividades en las
células, por ejemplo en los peces el cambio de color o “camuflaje” que
presentan algunos de estos para adaptarse al ambiente, se debe almovimiento de vesículas que contienen los pigmentos o la melanina, através de los microtúbulos hacia al centrosoma o hacia al extremo positivodel microtubulo, lo que permite metizarse.
PROTEÍNAS MOTORAS
Son las kinesinas y dineinas, donde las kinesinas transportan vesículascargo al lado positivo y las dineinas vesículas cargo al lado negativo delmicrotúbulo.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 9/10
Carmen María Pirela
Page 9
Esto permite que mediante el microtubulo se transporten vesículas adistintos extremos del interior de la célula. A las proteínas motoras seasocian otras proteínas (ejm anquirina, espectrina, etc) forman una especiede puentes con las vesículas cargo.
AXONEMAS
Son estructuras básicas de los cilios y flagelos, y están formados por unpar de centriolos independientes centrales, con un par de microtúbulos de
los cuales uno es completo y el otro incompleto, y todos asociados al tubulocentral del axonema a través de espinas radiales. Y entre los paresperiféricos se encuentran unidos por nexina, y además hay dos brazos dedineinas. Las nexinas y espinas radiales son importan porque matienen todos
los microtúbulos unidos entre sí, entonces al mover las dineinas se mueve todo
el complejo junto o todo el flagelo, de manera que sin estas proteínas
habrían movimientos independientes de los túbulos. En los flagelos siempreexiste ese movimiento de látigo, y se debe al movimiento de dineinas,caminando sobre los microtúbulos.

7/25/2019 Clase N 9 Citoesqueleto
http://slidepdf.com/reader/full/clase-n-9-citoesqueleto 10/10
Carmen María Pirela
Page 10
SEMEJANZAS ENTRE MICROFILAMENTOS Y MICROTÚBULOS
1. Tanto los microfilamentos como los microtúbulos están constituidos por proteínas globulares con actividad NTPasa (ATP o GTP)
2. En ambos casos, ~ 50% de la proteína constituyente se encuentra en
soluble y el otro 50% en forma de filamentos3. Forman estructuras muy dinámicas, con un intercambio rápido desubunidades entre el "pool" soluble y el insoluble (filamentoso).
4. Ambos tipos de filamentos son estructuras polarizadas.
FILAMENTOS INTERMEDIOS
Estos filamentos pueden variar en las distintas proteínas fibrilares que lascompongan, de hecho hay 50 polímeros diferentes en base a proteínascomo keratina, vimentina, laminina, etc. Se forman a partir de unmonómero alpha hélice que se enrolla con otro monómero, cada dímeroentonces se enrolla con otros dímeros, luego se forma un tetrámero, y la
agrupación de los tetrámeros de forma antiparalela se forma unprotofilamento, donde ocho tetrámeros u ocho protofilamentos es lo queconforma el filamento intermedio final, el cual no es hueco como losmicrotúbulos, de hecho es muy denso debido a que es el enrollamiento dedímeros uno sobre otro. De manera que las proteínas fibrosas varían, perose caracterizan porque todas poseen el dominio ROD el cual permite elenrollamiento de una sobre la otra. Entre estas proteínas que conformanlos filamentos, quizás la más importan son lamininas que forman parte dela estructura interna de la membrana nuclear, también las queratinas,vimentinas, neurofilamentos, etc.
Estas proteínas también se asocian a uniones que median la interación célula –
célula o uniones célula matriz, es decir desmosomas y hemidesmosomas
respectivamente. Se organizan también en forma de una red desde la zonanuclear con la membrana plasmática jugando roles adherentes entre lascélulas epiteliales, y algunas especializadas en células musculares ynerviosas.