#Nanocristalizacion catalizada aplicada en Baldosa hidráulica
Upload
nguyendienCategory
view
219download
0
Cinética Enzimática
Enrique Rivera González

Importancia
• Las enzimas son proteínas capaces de catalizar específicamente reacciones bioquímicas.
• La actividad catalítica de las enzimas depende de su estructura.
Pueden requerir:
1) Sólo su secuencia de aminoácidos y su conformación
2) Un cofactor (iones inorgánicos como Fe2+, Mg2+, Cu2+)
3) Un grupo prostético (porfirinas)
2

• Las catálisis enzimática es esencial para sistemas vivos.
• Permite que procesos químicos no favorables energéticamente se lleven a cabo en condiciones biológicas:
Medio acuoso, pH neutro, temperatura y presión bajas.
• Cuando la enzima se desnaturaliza, pierde su estructura y por lo tanto su actividad catalítica.
3
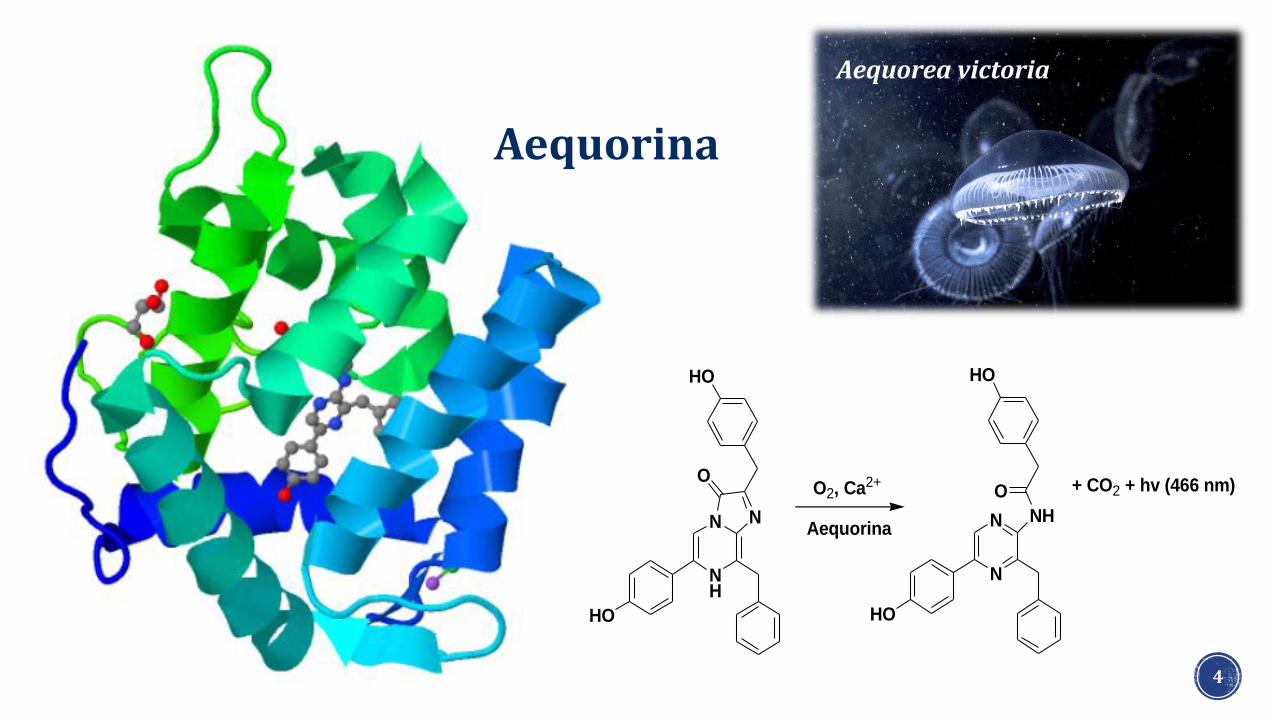
4
NH
N N
O
HO
HO
N
N NH
O
HO
HO
O2, Ca2+
Aequorina
+ CO2 + hv (466 nm)
Aequorea victoria
Aequorina
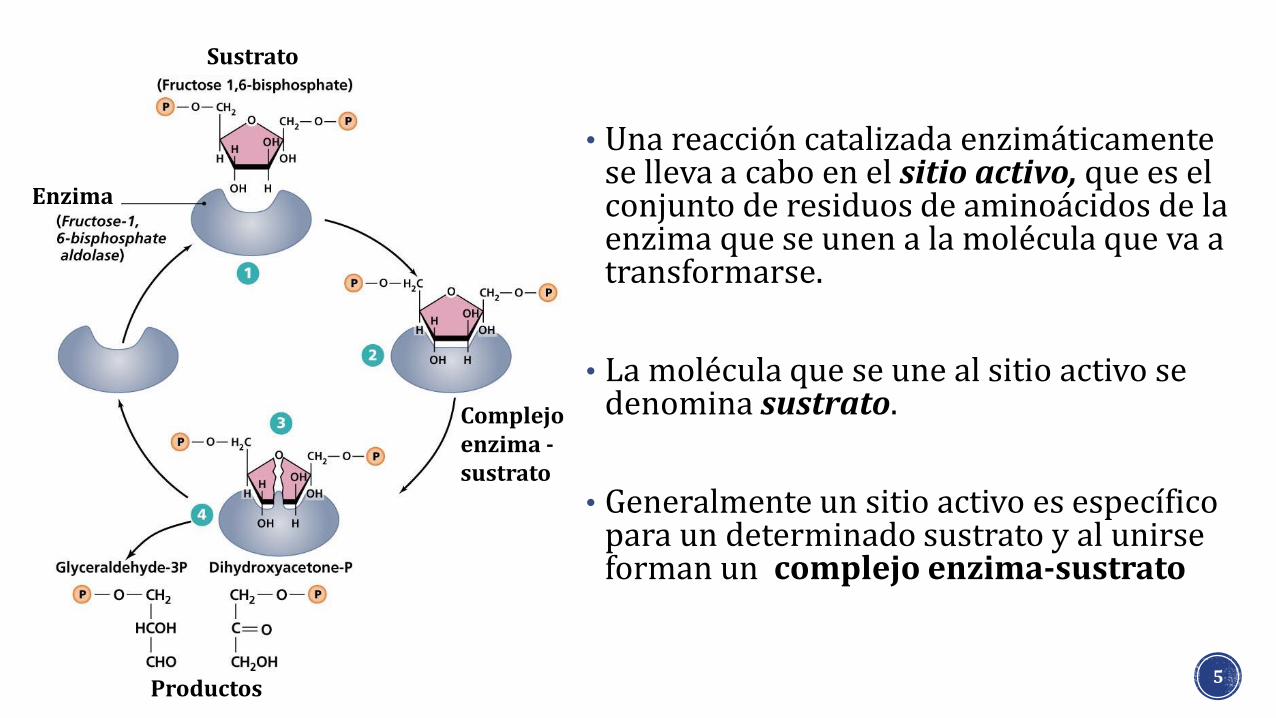
• Una reacción catalizada enzimáticamente se lleva a cabo en el sitio activo, que es el conjunto de residuos de aminoácidos de la enzima que se unen a la molécula que va a transformarse.
• La molécula que se une al sitio activo se denomina sustrato.
• Generalmente un sitio activo es específico para un determinado sustrato y al unirse forman un complejo enzima-sustrato
5
Enzima
Sustrato
Complejo enzima -sustrato
Productos

Factores energéticos
6
𝑆 → 𝑃
𝐸 + 𝑆 ⇌ 𝐸𝑆→𝑘2𝑃 + 𝐸
Coordenada de reacción
En
ergí
a li
bre
G Estado de transición
no catalizada
Reacción sin catalizar
Reacción catalizada por enzimas
𝑘1
𝑘−1
catalizada
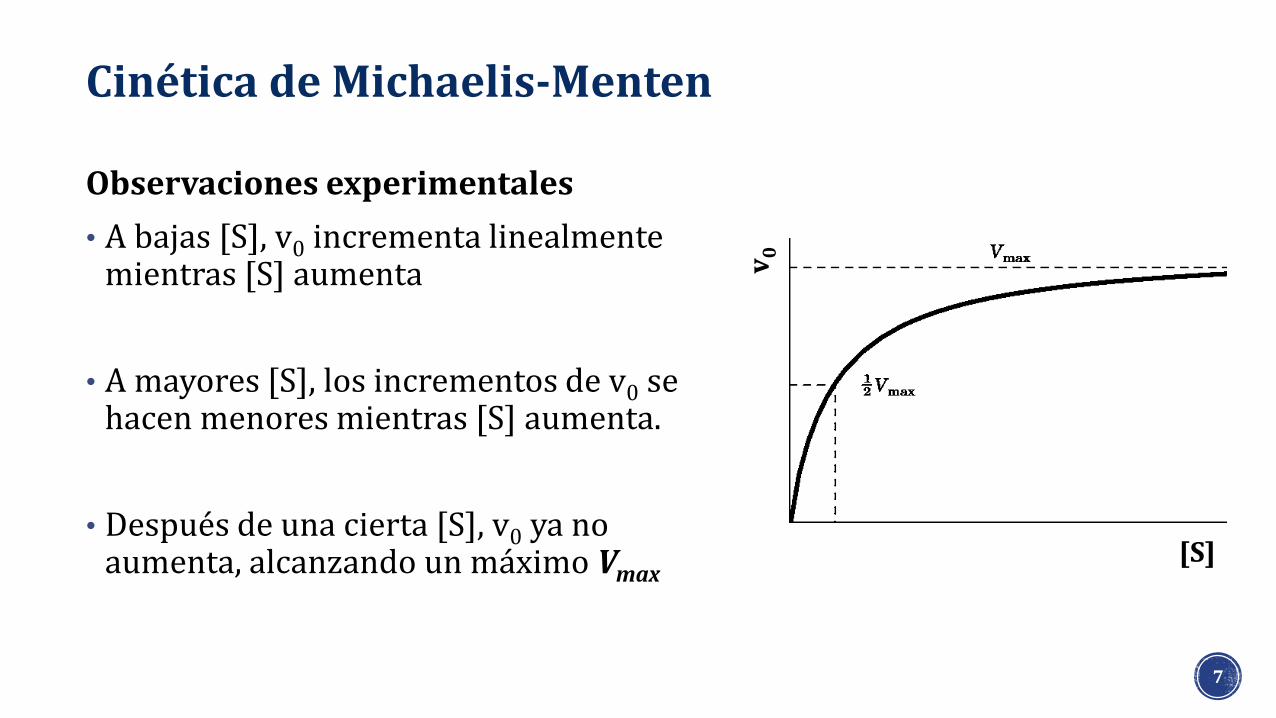
Cinética de Michaelis-Menten
Observaciones experimentales
• A bajas [S], v0 incrementa linealmente mientras [S] aumenta
• A mayores [S], los incrementos de v0 se hacen menores mientras [S] aumenta.
• Después de una cierta [S], v0 ya no aumenta, alcanzando un máximo Vmax
7
[S]
v0
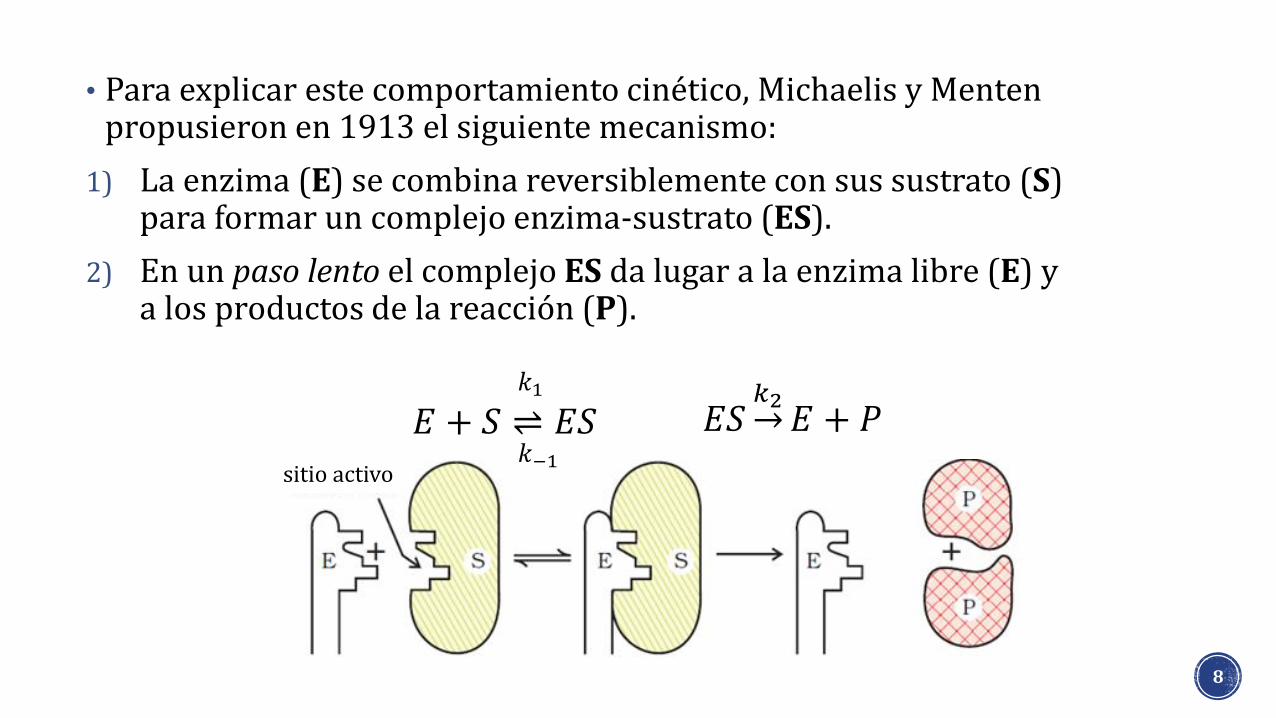
• Para explicar este comportamiento cinético, Michaelis y Mentenpropusieron en 1913 el siguiente mecanismo:
1) La enzima (E) se combina reversiblemente con sus sustrato (S) para formar un complejo enzima-sustrato (ES).
2) En un paso lento el complejo ES da lugar a la enzima libre (E) y a los productos de la reacción (P).
8
𝐸 + 𝑆 ⇌ 𝐸𝑆 𝐸𝑆→𝑘2𝐸 + 𝑃
𝑘−1
𝑘1
sitio activo

Se define la ecuación de velocidad total.
En la primera reacción se plantea una situación de equilibrio y las velocidades hacia ambas direcciones se igualan.
9
𝑣0 =𝑑[𝑃]
𝑑𝑡0
= 𝑘2[𝐸𝑆]
𝑘1 𝐸 [𝑆] = 𝑘−1[𝐸𝑆] 𝐾𝑠 =𝑘−1𝑘1
=𝐸 [𝑆]
[𝐸𝑆]
𝐸 + 𝑆 ⇌ 𝐸𝑆→𝑘2𝑃 + 𝐸
𝑘−1
𝑘1
Deducción de la ecuación de velocidad

A cualquier tiempo en una reacción catalizada enzimáticamente, la enzima existe en dos formas: E y ES y su concentración total se expresa:
Despejando [ES].
Sustituyendo [ES] en la expresión de velocidad.
10
𝑣0 =𝑑[𝑃]
𝑑𝑡0
= 𝑘2 𝐸𝑆
𝐾𝑠 =𝐸 [𝑆]
[𝐸𝑆]=
( 𝐸𝑆 − 𝐸 0)[𝑆]
[𝐸𝑆]
[𝐸]0= 𝐸 + [𝐸𝑆] 𝐸 = 𝐸𝑆 − [𝐸]0
𝐸𝑆 =𝐸 0[𝑆]
𝐾𝑠 + [𝑆]
=𝑘2 𝐸 0[𝑆]
𝐾𝑠 + [𝑆]
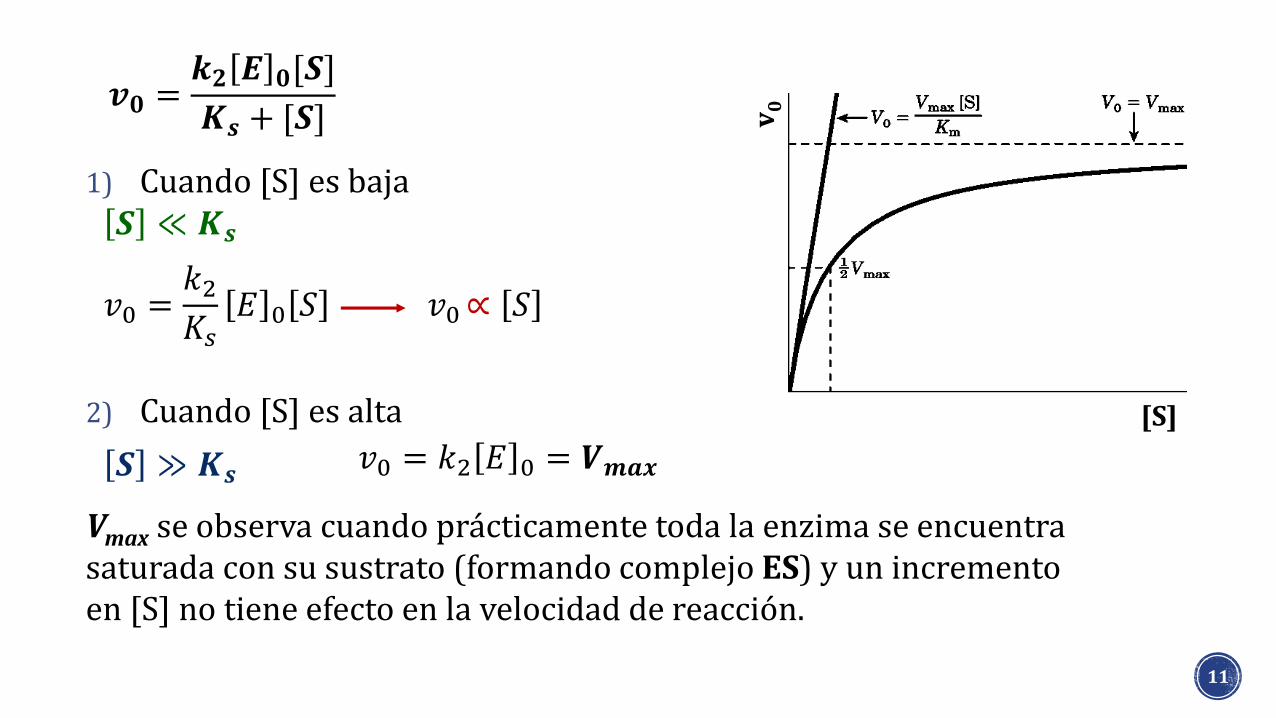
1) Cuando [S] es baja
2) Cuando [S] es alta
Vmax se observa cuando prácticamente toda la enzima se encuentra saturada con su sustrato (formando complejo ES) y un incremento en [S] no tiene efecto en la velocidad de reacción.
11
𝒗𝟎 =𝒌𝟐 𝑬 𝟎[𝑺]
𝑲𝒔 + [𝑺]
𝑣0 =𝑘2𝐾𝑠
𝐸 0 𝑆 𝑣0∝ 𝑆
𝑺 ≪ 𝑲𝒔
𝑺 ≫ 𝑲𝒔𝑣0 = 𝑘2 𝐸 0 = 𝑽𝒎𝒂𝒙
[S]
v0
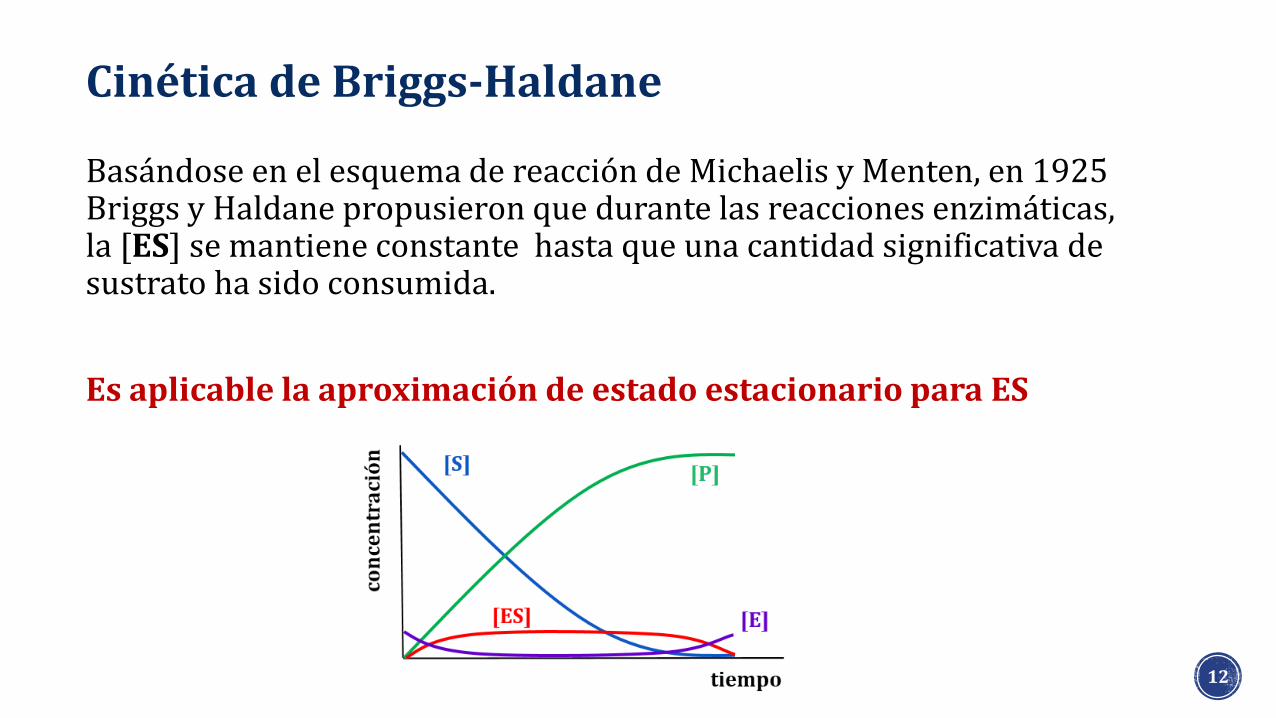
Basándose en el esquema de reacción de Michaelis y Menten, en 1925 Briggs y Haldane propusieron que durante las reacciones enzimáticas, la [ES] se mantiene constante hasta que una cantidad significativa de sustrato ha sido consumida.
Es aplicable la aproximación de estado estacionario para ES
12
Cinética de Briggs-Haldane

Se define la ecuación de velocidad total.
Se plantea la diferencial de ES.
13
𝑣0 =𝑑[𝑃]
𝑑𝑡0
= 𝑘2[𝐸𝑆]
𝑑[𝐸𝑆]
𝑑𝑡= 𝑘1 𝐸 𝑆 − 𝑘−1 𝐸𝑆 − 𝑘2 𝐸𝑆 = 0
𝐸 + 𝑆 ⇌ 𝐸𝑆→𝑘2𝑃 + 𝐸
𝑘−1
𝑘1
Deducción de la ecuación de velocidad

La concentración total de la enzima se expresa:
Sustituyendo [E] en la ecuación de estado estacionario.
Despejando [ES].
Sustituyendo [ES] en la expresión de velocidad.
14
[𝐸]0= 𝐸 + [𝐸𝑆] 𝐸 = 𝐸𝑆 − [𝐸]0
𝐸𝑆 =𝑘1 𝐸 0 𝑆
𝑘−1 + 𝑘2 + 𝑘1 𝑆
𝑘1( 𝐸𝑆 − 𝐸 0) 𝑆 − 𝑘−1 𝐸𝑆 − 𝑘2 𝐸𝑆 = 0
𝑘1 𝐸 𝑆 − 𝑘−1 𝐸𝑆 − 𝑘2 𝐸𝑆 = 0
𝑣0 =𝑑[𝑃]
𝑑𝑡0
= 𝑘2 𝐸𝑆=𝑘2𝑘1 𝐸 0 𝑆
𝑘1 𝑆 + 𝑘−1 + 𝑘2=
𝑘2 𝐸 0 𝑆
𝑘−1 + 𝑘2𝑘1
+ 𝑆

Aproximación al equilibrio: Aproximación de estado estacionario:
• Michaelis y Menten asumieron que la unión al sustrato y la disociación del complejo ES ocurrían más rápido que la formación de producto (𝑘−1 ≫ 𝑘2).
• Briggs y Haldane no asumieron nada acerca de los valores de 𝑘−1 𝑦 𝑘2.
El modelo cinético de M-M es un caso especial del modelo de B-H.15
𝐸 + 𝑆 ⇌ 𝐸𝑆→𝑘2𝑃 + 𝐸
𝑘−1
𝑘1
𝑣0 =𝑘2 𝐸 0 𝑆
𝑘−1 + 𝑘2𝑘1
+ 𝑆=𝑘2 𝐸 0[𝑆]
𝐾𝑚 + [𝑆]
𝐾𝑚 =𝑘−1 + 𝑘2
𝑘1𝐾𝑠 =
𝑘−1𝑘1
𝑣0 =𝑘2 𝐸 0[𝑆]
𝐾𝑠 + [𝑆]
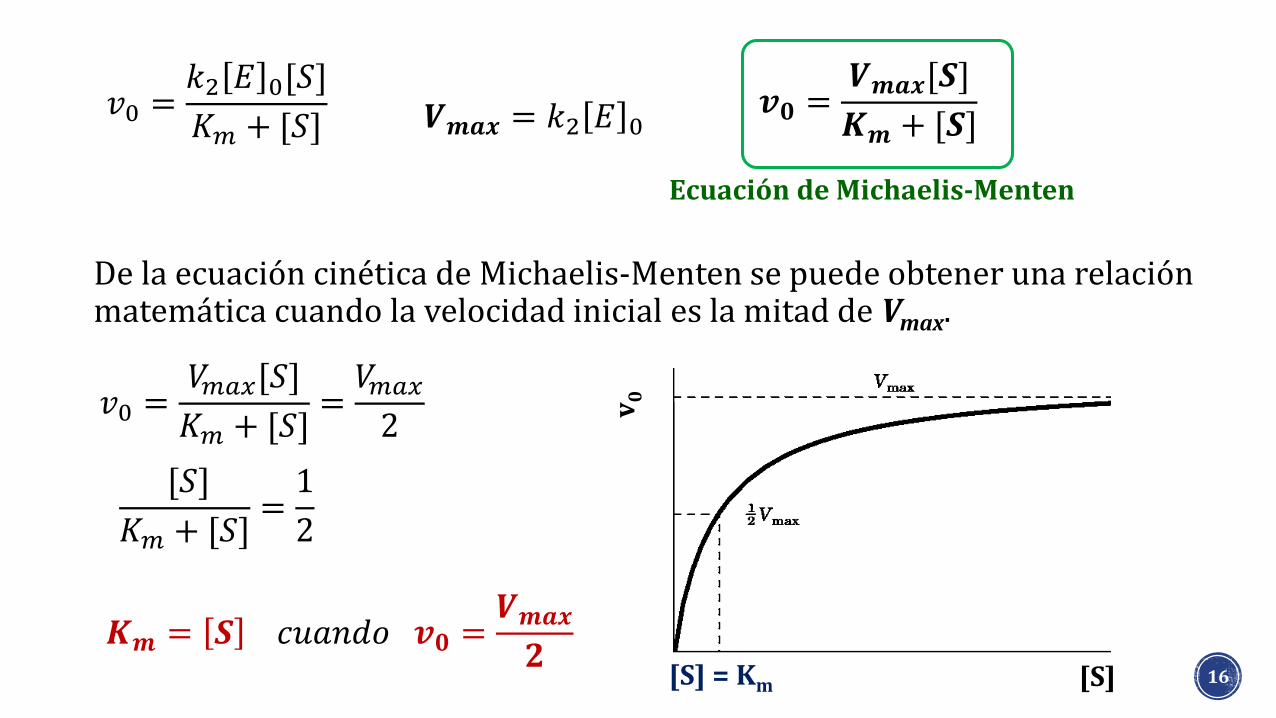
De la ecuación cinética de Michaelis-Menten se puede obtener una relación matemática cuando la velocidad inicial es la mitad de Vmax.
16
𝑣0 =𝑘2 𝐸 0[𝑆]
𝐾𝑚 + [𝑆]
𝑣0 =𝑉𝑚𝑎𝑥[𝑆]
𝐾𝑚 + [𝑆]=𝑉𝑚𝑎𝑥
2
𝑽𝒎𝒂𝒙 = 𝑘2 𝐸 0𝒗𝟎 =
𝑽𝒎𝒂𝒙[𝑺]
𝑲𝒎 + [𝑺]
Ecuación de Michaelis-Menten
[S]
v0
[𝑆]
𝐾𝑚 + [𝑆]=1
2
𝑲𝒎 = 𝑺 𝑐𝑢𝑎𝑛𝑑𝑜 𝒗𝟎 =𝑽𝒎𝒂𝒙
𝟐[S] = Km
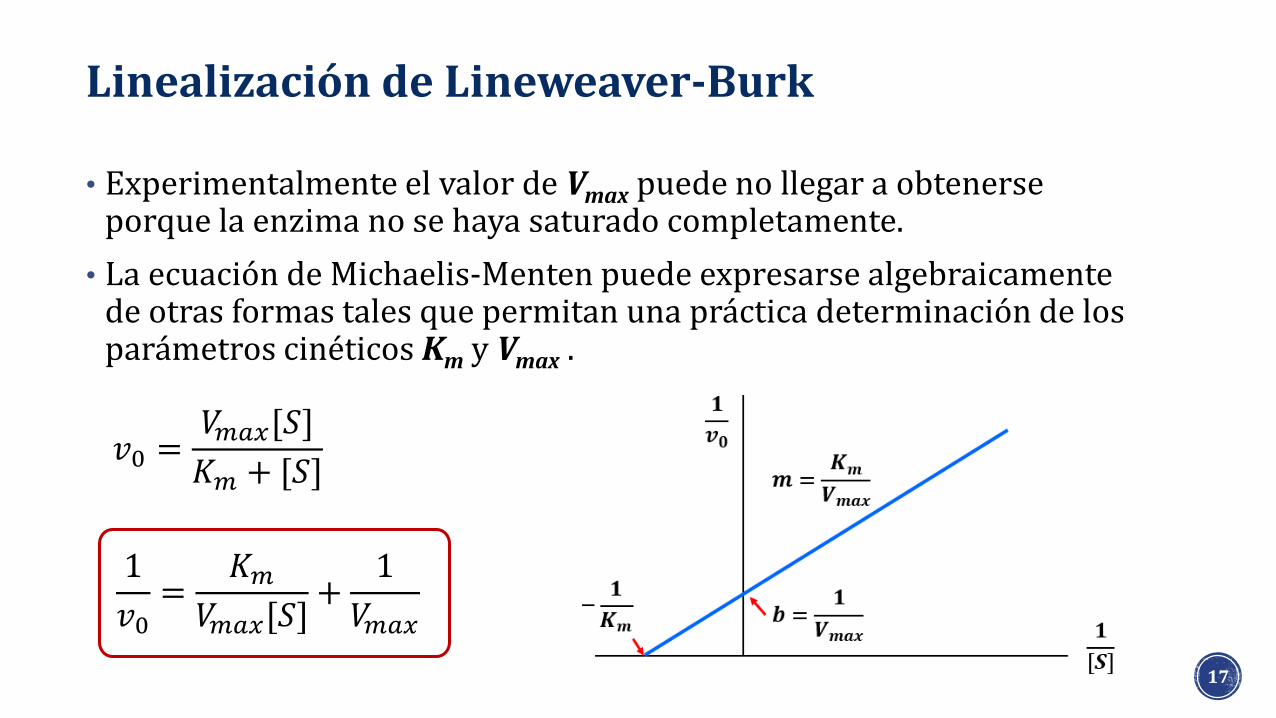
Linealización de Lineweaver-Burk
• Experimentalmente el valor de Vmax puede no llegar a obtenerse porque la enzima no se haya saturado completamente.
• La ecuación de Michaelis-Menten puede expresarse algebraicamente de otras formas tales que permitan una práctica determinación de los parámetros cinéticos Km y Vmax .
17
𝑣0 =𝑉𝑚𝑎𝑥[𝑆]
𝐾𝑚 + [𝑆]
1
𝑣0=
𝐾𝑚𝑉𝑚𝑎𝑥[𝑆]
+1
𝑉𝑚𝑎𝑥
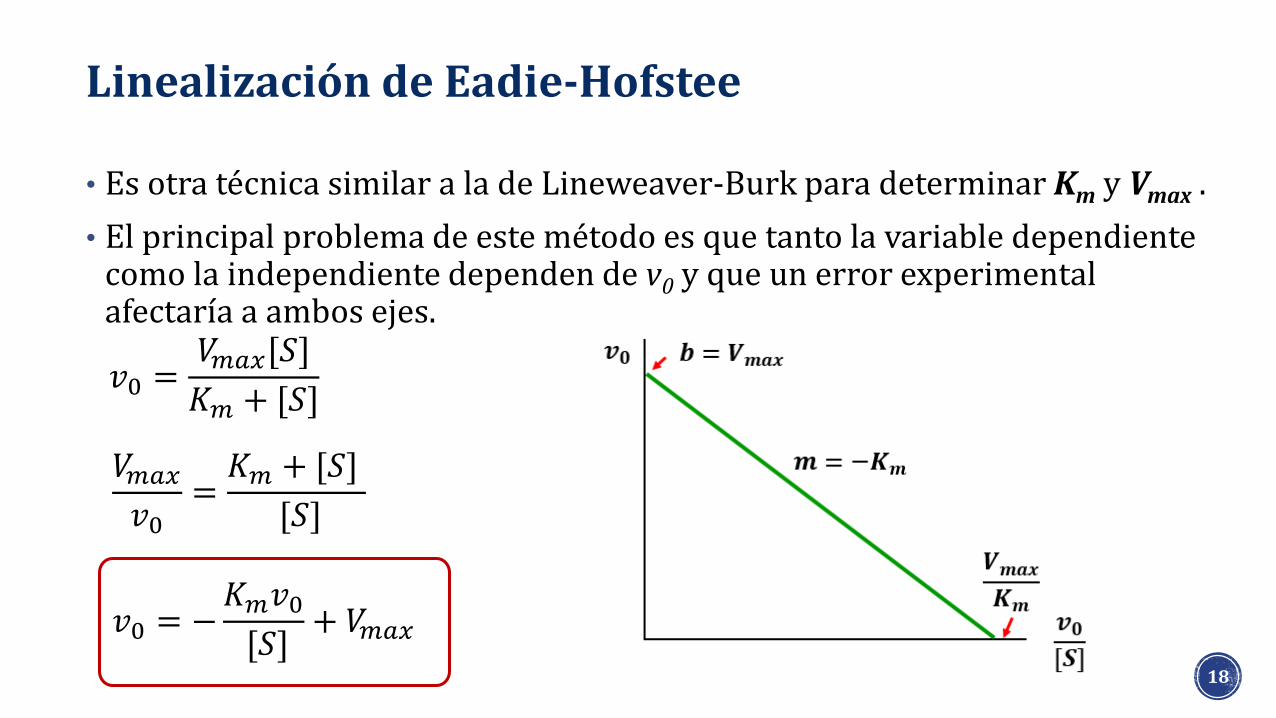
Linealización de Eadie-Hofstee
• Es otra técnica similar a la de Lineweaver-Burk para determinar Km y Vmax .
• El principal problema de este método es que tanto la variable dependiente como la independiente dependen de v0 y que un error experimental afectaría a ambos ejes.
18
𝑣0 =𝑉𝑚𝑎𝑥[𝑆]
𝐾𝑚 + [𝑆]
𝑉𝑚𝑎𝑥
𝑣0=𝐾𝑚 + [𝑆]
[𝑆]
𝑣0 = −𝐾𝑚𝑣0[𝑆]
+ 𝑉𝑚𝑎𝑥

Interpretación de Km
• Representa la [S] a la cual la mitad de las moléculas de enzima se encuentran saturadas, formando complejo ES.
• El valor de Km sugiere la afinidad de la enzima con sus sustrato.
Un valor bajo de Km indica mayor afinidad y por lo tanto una mayor velocidad a cualquier [S].
• Km es utilizado para identificar isoenzimas, las cuales presentan diferente afinidad por el mismo sustrato.
• Km es sensible a condiciones como pH y temperatura, lo cual permite identificar cambios estructurales de la enzima.
19

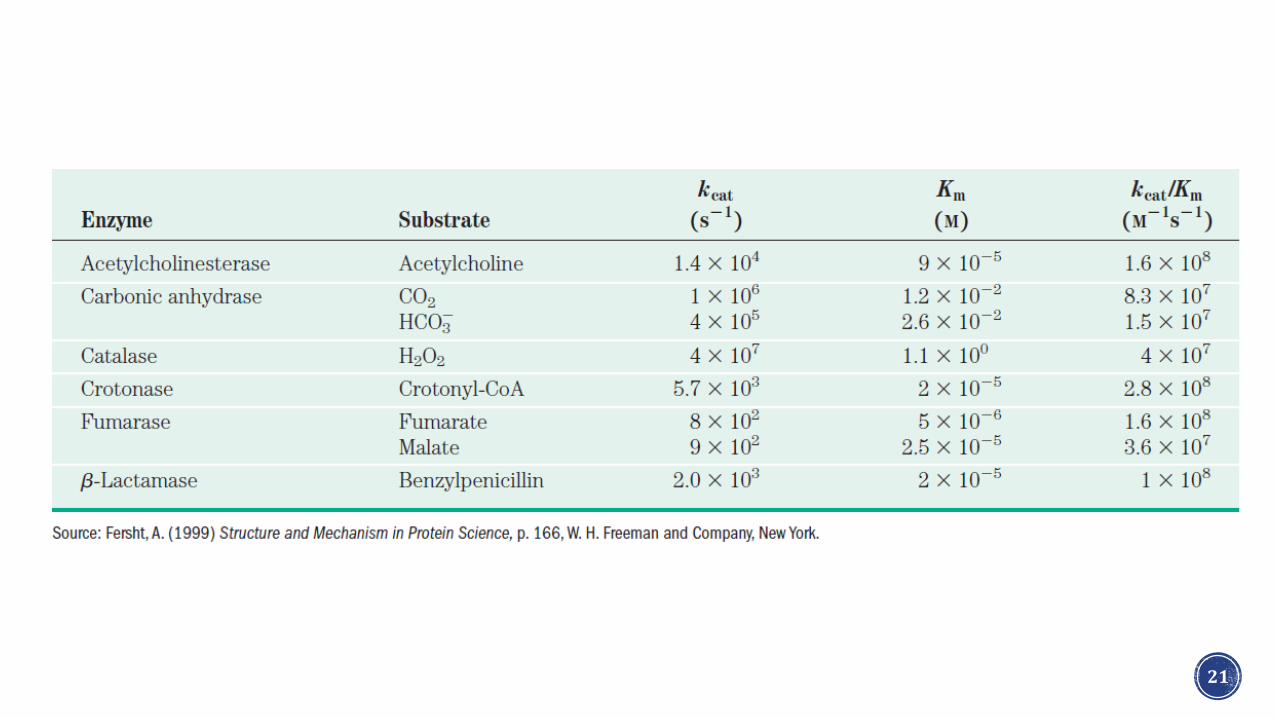
Interpretación de k2
• k2 también conocida como kcat o número de recambio es la constante de velocidad para el paso determinante de velocidad.
• kcat se define como el número de moléculas de producto formadas a partir del sustrato por una enzima en una unidad de tiempo.
• El cociente kcat
K𝑚
, denominado como cociente de especificidad permite
comparar la eficiencia catalítica de diferentes enzimas o de la transformación de dos sustratos diferentes por la misma enzima.
20
𝑣0 =𝑘2 𝐸 0[𝑆]
𝐾𝑚 + [𝑆]
21
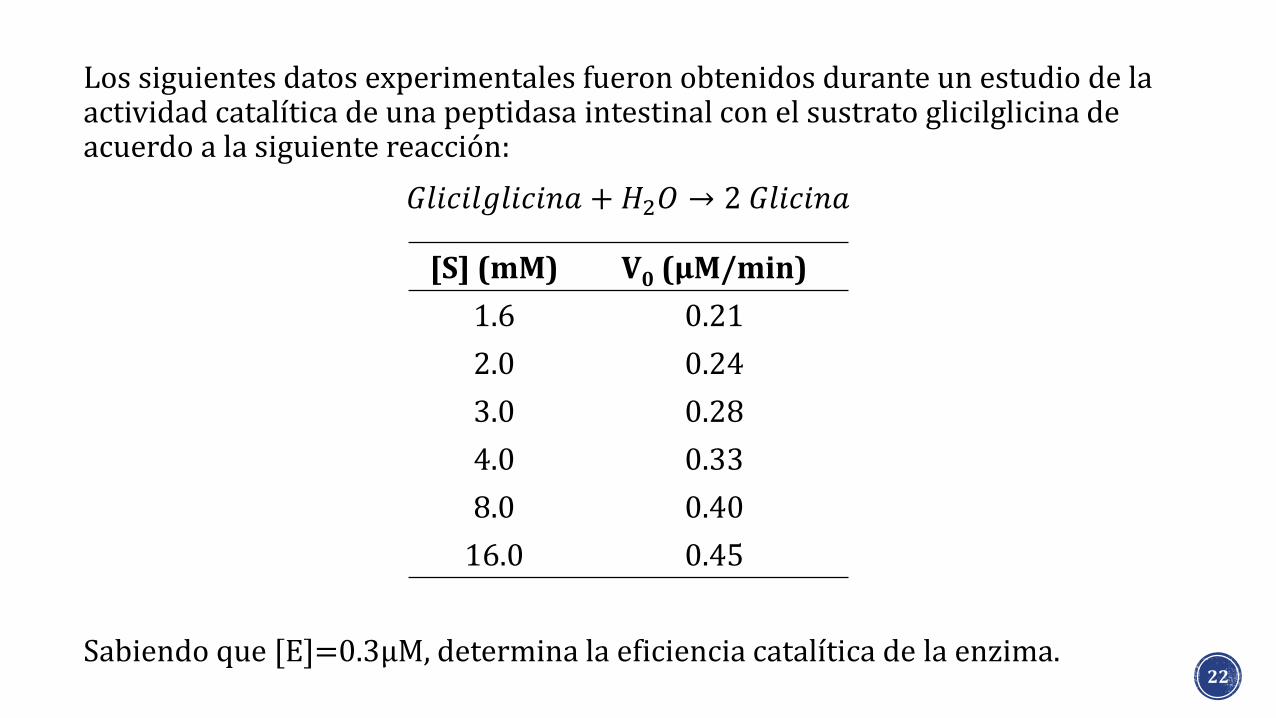
Los siguientes datos experimentales fueron obtenidos durante un estudio de la actividad catalítica de una peptidasa intestinal con el sustrato glicilglicina de acuerdo a la siguiente reacción:
Sabiendo que [E]=0.3μM, determina la eficiencia catalítica de la enzima.22
[S] (mM) V0 (μM/min)
1.6 0.21
2.0 0.24
3.0 0.28
4.0 0.33
8.0 0.40
16.0 0.45
𝐺𝑙𝑖𝑐𝑖𝑙𝑔𝑙𝑖𝑐𝑖𝑛𝑎 + 𝐻2𝑂 → 2 𝐺𝑙𝑖𝑐𝑖𝑛𝑎

Se determinó la velocidad inicial a varias concentraciones de sustrato para una reacción catalizada enzimáticamente de acuerdo con los siguientes datos:
23
[S] /105(M) V0 /106(mol/min)
2.5 0.21
4.0 0.24
6.0 0.28
8.0 0.33
16.0 0.40
20.0 0.45
a) Menciona si la reacción sigue una cinética de acuerdo al modelo de Michaelis-Menten.
b) Si kcat es 8.0 𝑥 102, ¿Cuál fue la concentración de enzima utilizada?
c) Calcula v0 cuando [𝑆] = 5.0 𝑥 10−5 y [𝑆] = 5.0 𝑥 10−1
d) Si se duplicara la concentración de la enzima, ¿Cómo se verían afectados los valores de Km, kcat y Vmax?