CARACTERIZACIÓN MOLECULAR, DETECCIÓN Y RESISTENCIA … · caracterizaciÓn molecular,detecciÓn y...
234
5 9 Iker Sevilla Agirregomoskorta CARACTERIZACIÓN MOLECULAR, DETECCIÓN Y RESISTENCIA DE MYCOBACTERIUM AVIUM SUBESPECIE PARATUBERCULOSIS
Transcript of CARACTERIZACIÓN MOLECULAR, DETECCIÓN Y RESISTENCIA … · caracterizaciÓn molecular,detecciÓn y...

59
Iker Sevilla Agirregomoskorta
CARACTERIZACIÓN MOLECULAR, DETECCIÓNY RESISTENCIA DE MYCOBACTERIUM AVIUM
SUBESPECIE PARATUBERCULOSIS



CARACTERIZACIÓN MOLECULAR, DETECCIÓN
Y RESISTENCIA DE MYCOBACTERIUM AVIUM
SUBESPECIE PARATUBERCULOSIS
TESIS DOCTORALESN.º 59
Vitoria-Gasteiz, 2007
NEKAZARITZA, ARRANTZAETA ELIKADURA SAILA
DEPARTAMENTO DE AGRICULTURAPESCA Y ALIMENTACIÓN
Iker Sevilla Agirregomoskorta
UNIVERSIDAD DEL PAÍS VASCO

Edición: 1.ª junio 2007
© Administración de la Comunidad Autónoma del País VascoDepartamento de Agricultura, Pesca y Alimentación
Internet: www.euskadi.net
Edita: Eusko Jaurlaritzaren Argitalpen Zerbitzu NagusiaServicio Central de Publicaciones del Gobierno VascoDonostia-San Sebastián, 1 - 01010 Vitoria-Gasteiz
Impresión: XXXXXXXXXXXXXXXXXXXXXXXX
ISBN: 978-84-457-2602-0
D. L.: XXXXXXXXX
SEVILLA AGIRREGOMOSKORTA, IkerCaracterización molecular, detección y resistencia de “Mycobacterium avium” subespecie
“Paratuberculosis”/ Iker Sevilla Agirregomoskorta. - 1ª ed. - Vitoria-Gasteiz : Eusko JaurlaritzarenArgitalpen Zerbitzu Nagusia = Servicio Central de Publicaciones del Gobierno Vasco, 2007
p. ; cm. - (Tesis doctorales ; 59)Tesis-Universidad del País VascoISBN 978-84-457-2602-0
1. Paratuberculosis-Tesis doctorales. I. Euskadi. Departamento de Agricultura, Pesca y Alimentación.II. Título. III. Serie636.2/.3:616.98:579.873.2(043)

Caracterización Molecular, Detección y Resistencia de Mycobacterium avium
subespecie paratuberculosis
Tesis Doctoral
Memoria del trabajo realizado en Neiker-Tecnalia
presentado por Iker Sevilla Agirregomoskorta
para optar al grado de Doctor en Biología
por la Universidad del País Vasco
2007

El presente trabajo de Tesis Doctoral ha sido realizado en el Departamento de Sanidad Animal del Instituto Vasco de Investigación y Desarrollo Agrario–Neiker-Tecnalia.
Su desarrollo ha sido posible gracias a las becas de Formación de Tecnólogos del Departamento de Agricultura, Pesca y Alimentación, y de Formación de Doctores en centros de la Red Vasca de Ciencia, Tecnología e Innovación del Departamento de Industria, Comercio y Turismo y del Departamento de Educación, Universidades e Investigación del Gobierno Vasco, y a la beca de la Fundación Cándido Iturriaga.

Ainarari
Amatxu ta Amamari
Patxi, Elixabete ta Aitari
Familia ta Lagunei


AUTORIZACION DEL/LA DIRECTOR/A DE TESIS
PARA SU PRESENTACION
Los Dres. Ramón A. Juste Jordán y Joseba M. Garrido Urkullu como Directores de la
Tesis Doctoral: Caracterización Molecular, Detección y Resistencia de Mycobacterium
avium subespecie paratuberculosis realizada en el Departamento/Instituto Instituto
Vasco de Investigación y Desarrollo Agrario (NEIKER-Tecnalia) por el Doctorando
Don/ña. Iker Sevilla Agirregomoskorta, autorizan la presentación de la citada Tesis
Doctoral, dado que reúne las condiciones necesarias para su defensa.
En Derio a 11 de Diciembre de 2006
LOS DIRECTORES DE LA TESIS
Fdo.: Ramón A. Juste Jordán Fdo.: Joseba M. Garrido Urkullu


RATIFICACION DEL/LA PONENTE PARA LA
PRESENTACION DE LA TESIS
El Dr. Mikel Iriondo Orensanz como Tutor y Ponente de la Tesis Doctoral del
Doctorando Don Iker Sevilla Agirregomoskorta, ratifica la autorización de los
Directores para la presentación de la Tesis Doctoral que lleva por título Caracterización
Molecular, Detección y Resistencia de Mycobacterium avium subespecie
paratuberculosis, realizada en el Departamento/Instituto Departamento de Sanidad
Animal del Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER-Tecnalia).
En Leioa a 26 de Diciembre de 2006
EL TUTOR DE LA TESIS
Fdo.: Mikel Iriondo Orensanz


CONFORMIDAD DEL
DEPARTAMENTO/INSTITUTO
El Consejo del Departamento/Instituto de
en reunión celebrada el día ____ de de ha acordado dar la
conformidad a la admisión a trámite de presentación de la Tesis Doctoral titulada:
Caracterización Molecular, Detección y Resistencia de Mycobacterium avium
subespecie paratuberculosis dirigida por los Dres. Ramón A. Juste Jordán y Joseba M.
Garrido Urkullu y presentada por Don/ña. Iker Sevilla Agirregomoskorta ante este
Departamento/Instituto.
En a de de
Vº Bº DIRECTOR/A DEL SECRETARIO/A DEL DEPARTAMENTO/INSTITUTO DEPARTAMENTO/INSTITUTO
Fdo.: __________________________ Fdo.: _______________________


ACTA DE GRADO DE DOCTOR
ACTA DE DEFENSA DE TESIS DOCTORAL
DOCTORANDO DON/ÑA. Iker Sevilla Agirregomoskorta
TITULO DE LA TESIS: Caracterización Molecular, Detección y Resistencia de
Mycobacterium avium subespecie paratuberculosis
El Tribunal designado por la Subcomisión de Doctorado de la UPV/EHU para calificar
la Tesis Doctoral arriba indicada y reunido en el día de la fecha, una vez efectuada la
defensa por el doctorando y contestadas las objeciones y/o sugerencias que se le han
formulado, ha otorgado por___________________la calificación de: unanimidad ó mayoría
En a de de
EL/LA PRESIDENTE/A, EL/LA SECRETARIO/A,
Fdo.: Fdo.:
Dr/a: ____________________ Dr/a: ______________________
VOCAL 1º, VOCAL 2º, VOCAL 3º,
Fdo.: Fdo.: Fdo.:
Dr/a: Dr/a: Dr/a:
EL/LA DOCTORANDO/A,
Fdo.: Iker Sevilla Agirregomoskorta


AGRADECIMIENTOS


Dicen que para ser feliz en la vida como mínimo es necesario plantar un árbol, escribir un libro y tener un hijo. No creo que todo ello sea necesario, pero yo por si acaso, ya he cumplido la primera premisa, y con esta tesis intento dejar cumplida también la segunda. De la tercera... hablaremos. El camino ha sido duro y difícil, lleno de subidas y bajadas, pero los logros obtenidos y el apoyo recibido han permitido mantener el rumbo trazado. Por eso, quiero agradecer a todos la ayuda que me habéis brindado para lograr el cachito de mi felicidad que tenéis entre las manos.
Antes de nada, quiero dar las gracias a las personas a quien está dedicado este trabajo. A Ainara, a ama, a amama, a Patxi, a Elixabete, a aita, a mis familiares más cercanos y a mis amigos de toda la vida. Espero que después de soportarme durante este tiempo, todavía os queden ganas de seguir haciéndolo por mucho más tiempo.
Les doy las gracias a Ramón y a Joseba, por darme esta oportunidad, por dirigir mi trabajo, por discutirlo, por disfrutar de él, por buscar soluciones y por confiar en mí incluso cuando ni siquiera yo era capaz de hacerlo. A Mikel, por hacer de puente entre Neiker y la Universidad y por conseguir las respuestas a todas mis preguntas. A los tres, gracias por transmitirme vuestra tranquilidad.
Agradezco mucho la labor de las personas que durante este tiempo han contribuido de una u otra forma al trabajo del grupo de micobacterias de Neiker. A Mertxe, Mariví G., Mariví L., Sara, Natalia, Olaia, Ainara, Zuriñe, Galder, Amaia, y nuestro último fichaje pero no menos importante, Elena. Sin su trabajo esta tesis nunca habría salido adelante. Además, me alegra haber hecho muy buenas migas con algunos de ellos. En este sentido, también me gustaría dar las gracias por su colaboración al grupo de micobacterias de León, formado por Francisco, Valentín, Andrea, Jorge, Carlos, María, Laetitia y otros que no conozco pero que seguramente habrán aportado su trabajo. También aprovecho para dar las gracias al grupo de micobacterias del IREC, a JM Plazaola, a Shoor Vir, a Ana Claudia, a RJ Whittington, a RH Whitlock, a Tomás, a Buenaventura, a Artur, a María Jesús, a Martha, y a todos los ganaderos y veterinarios que nos han suministrado muestras e información, por su importantísima colaboración.
A Gorka, que no ha dudado en prestarme toda la ayuda que haya podido necesitar, y cuyo trabajo ha permitido parte de esta tesis. A Ana G., Marta, Inés y Jesse por dejarme dar el primer paso en el mundo de la investigación y echarme un cabo cuando lo he necesitado. A Ana H. por contestar a mis preguntas e incentivar nuestra curiosidad.
Muchas gracias a Itziar, Xeider, Jon, Iratxe, David, Bea, Vega, Josune, Mara, Idoia, y Leyre, mis compañeros de laboratorio de siempre, los que más se han implicado, los que más veces han soportado mi mala uva cuando las cosas se torcían, pero también con los que más he reído y disfrutado. Entre ellos cuento ya con auténticos amigos. También a las recientes incorporaciones Laura E., Félix, y Sonia, y a otros que dejaron Neiker, como Aitzol, Laura L., Laura M., Itziar K., y Dani. Muchas gracias a todos por vuestro apoyo, interés, y ayuda, por hacer el día a día más ameno y agradable.
Gracias a José Carlos por “prestarme” material con una sonrisa y ayudarme con las contaminaciones. A Lucía por ayudarme con los medios y los pedidos, y por surtirnos de ositos-gominola para no perder las energías. A Rakel, Gloria y Argiñe, por ayudarme en mis inicios en el laboratorio, a Nieves, Arrate, Esme y Nico por conseguir las mejores muestras y echar un cable. A Iñigo, Jon Paul, Leire, Iranzu, Yolanda, Nekane, Jon A. y Rosa por su apoyo administrativo, y a Gontzal y Aitor por su apoyo informático. A José Ángel, Unai, Sergio, Fernando y Fidel por mantener, muchas veces desde la “sombra”, el funcionamiento general de Neiker, y hacer lo posible por satisfacer nuestras necesidades. A Mari y a Claudina, que además de ocuparse de que todo esté limpio, una me echa piropos y otra me trae pan recién horneado los días de bocadillo. A Julián y a Fer por ayudarme con la encuadernación y diseño del libro de tesis.
Creo que es imposible no dejarme a alguien en el tintero, pero espero que sepa perdonarme. Me alegro mucho de haber contado con vosotros en este período tan importante.
ESKERRIK ASKO DANORI !


ÍNDICE


LISTA DE ABREVIATURAS .......................................................................................................................25 INTRODUCCIÓN GENERAL ......................................................................................................................27 ESCENARIO Y PLANTEAMIENTO DEL TRABAJO DE LA TESIS.................................................33
OBJETIVOS ................................................................................................................................................36 ESTUDIOS LLEVADOS A CABO .........................................................................................................36
REVISIÓN DE LA BIBLIOGRAFÍA ...........................................................................................................39 ETIOLOGÍA .................................................................................................................................................41
Género Mycobacterium......................................................................................................................41 Morfología externa ..........................................................................................................................41 Metabolismo ......................................................................................................................................42 Composición genética ...................................................................................................................42 Clasificación taxonómica..............................................................................................................43 Identificación.....................................................................................................................................45 Tipado molecular .............................................................................................................................48
Complejo M. avium ..............................................................................................................................50 Historia ................................................................................................................................................51
Mycobacterium avium subsp. paratuberculosis .......................................................................52 Morfología y metabolismo ............................................................................................................52 La micobactina .................................................................................................................................53 Los medios de cultivo ....................................................................................................................54 Resistencia y persistencia............................................................................................................55 Composición genética ...................................................................................................................56
EPIDEMIOLOGÍA ......................................................................................................................................57 Hospedadores .......................................................................................................................................57 Transmisión ...........................................................................................................................................59 Distribución y prevalencia ................................................................................................................61 Trascendencia Económica ...............................................................................................................65 Implicaciones en salud pública ......................................................................................................66
Enfermedad de Crohn ....................................................................................................................66 Map en la cadena alimentaria del ser humano. La leche, el agua y otros productos. Inactivación de Map en leche ......................................................................................................71
Epidemiología molecular...................................................................................................................75 PATOGÉNESIS Y PATOLOGÍA ............................................................................................................83
Entrada e invasión de Map en el hospedador............................................................................83 Factores de virulencia....................................................................................................................85
Respuesta inmune del hospedador...............................................................................................86 Cuadro clínico .......................................................................................................................................90 Lesiones macroscópicas ..................................................................................................................91 Lesiones microscópicas ...................................................................................................................92
DIAGNÓSTICO...........................................................................................................................................93 Diagnóstico basado en las técnicas histopatológicas convencionales ...........................93 Diagnóstico basado en la detección bacteriológica................................................................94
Tinción y detección al microscopio ..........................................................................................94 Análisis inmunohistoquímico de preparaciones histológicas ........................................95 Aislamiento ........................................................................................................................................95 Detección de DNA............................................................................................................................98
Diagnóstico basado en la respuesta inmunológica del hospedador ..............................102 Detección de la inmunidad celular in vivo: Intradermorreacción .................................103 Detección de la inmunidad celular in vitro ...........................................................................103 • Prueba del IFN-γ ................................................................................................................104
Detección de la inmunidad humoral .......................................................................................105 • Prueba de AGID .................................................................................................................105 • Prueba de ELISA................................................................................................................106
TRATAMIENTO Y MEDIDAS DE CONTROL...................................................................................108 Programas de detección y eliminación de positivos ............................................................110 Programas que incluyen la vacunación.....................................................................................111
ESTUDIOS .....................................................................................................................................................115 ESTUDIO I. Comparison of blood polymerase chain reaction and enzyme-linked immunosorbent assay for detection of Mycobacterium avium subsp. paratuberculosis infection in cattle and sheep ..............................................................................................................117

Abstract .................................................................................................................................................119 Introduction..........................................................................................................................................119 Materials and methods .....................................................................................................................120 Results ...................................................................................................................................................122 Discussion and conclusions..........................................................................................................123 Acknowledgements ...........................................................................................................................125
ESTUDIO II. Molecular typing of Mycobacterium avium subspecies paratuberculosis strains from different hosts and regions .......................................................................................127
Summary ...............................................................................................................................................129 Introduction..........................................................................................................................................129 Materials and methods .....................................................................................................................129 Results ...................................................................................................................................................132 Discussion............................................................................................................................................133 Acknowledgements ...........................................................................................................................135
ESTUDIO III. Pulsed-field gel electrophoresis profile homogeneity of Mycobacterium avium subsp. paratuberculosis isolates from cattle and heterogeneity of those from sheep and goats .....................................................................................................................................137
Abstract .................................................................................................................................................139 Background..........................................................................................................................................139 Methods .................................................................................................................................................141
Sources and growth of isolates ................................................................................................141 Cultures for PFGE..........................................................................................................................143 Isolate identification: IS900 PCR and Locus 251 PCR ......................................................143 IS1311 PCR-REA ............................................................................................................................143 SnaBI-SpeI PFGE analysis..........................................................................................................143 Phylogenetic analysis of PFGE profiles and estimation of genetic diversity...........144
Results ...................................................................................................................................................145 IS900 PCR and Locus 251 PCR .................................................................................................145 IS1311 PCR-REA ............................................................................................................................145 SnaBI-SpeI PFGE analysis..........................................................................................................146 Phylogenetic analysis of PFGE profiles and estimation of genetic diversity...........150
Discussion............................................................................................................................................151 Conclusions .........................................................................................................................................155 Authors' contributions .....................................................................................................................156 Acknowledgements ...........................................................................................................................156
ESTUDIO IV. Detection and pathological implications of the presence of IS900 in human Mycobacterium avium isolates ...........................................................................................157
Abstract .................................................................................................................................................159 Introduction..........................................................................................................................................159 Methods and results .........................................................................................................................160 Discussion and conclusions..........................................................................................................161
ESTUDIO V. Inactivation of Mycobacterium avium subsp. paratuberculosis in cow’s milk by means of high hydrostatic pressure at mild temperatures......................................163
Abstract .................................................................................................................................................165 Introduction..........................................................................................................................................165 Materials and methods .....................................................................................................................166
Strains and preparation of inocula ..........................................................................................166 Inoculation .......................................................................................................................................166 HHP treatments ..............................................................................................................................166 Microbiological analysis .............................................................................................................167 Statistical analysis ........................................................................................................................167
Results, discussion and conclusions.........................................................................................167 Acknowledgements ...........................................................................................................................170
DISCUSIÓN GENERAL .............................................................................................................................171 CONCLUSIONES GENERALES..............................................................................................................179 RESUMEN; LABURPENA; SUMMARY.................................................................................................183
RESUMEN .................................................................................................................................................185 LABURPENA ............................................................................................................................................188 SUMMARY ................................................................................................................................................191
BIBLIOGRAFÍA CITADA ...........................................................................................................................195

25
LISTA DE ABREVIATURAS A: adenina ABC: adenosine triphosphate-binding cassette transporter ABTS: 2,2'-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid) AFLP: amplified fragment length polymorphism Ag: antigen/ antígeno AGID: agar gel immunodiffusion AP: alkaline phosphatase ASA: 5-amino salicylic acid ATCC: American type culture collection BHI: brain heart infusion BSA: bovine serum albumin (k)bp: (kilo) base pair/ pares de bases C: citosina CARD: caspase-activating and recruitment domain CD: cluster of differentiation CF: complement fixation CFU: colony forming unit CIP: Collection de l'Institut Pasteur CMI: cell mediated immunity CS: complementary sensitivity CTAB: cetyl-trimethylammonium bromide dATP: (2’-deoxyadenosine 5’-triphosphate) dCTP: (2’-deoxycytidine 5’-triphosphate) dGTP: (2’-deoxyguanosine 5’-triphosphate) DMSO: dimethyl sulfoxide DNA: deoxyribonucleic acid DR: direct repeats dTTP: (2’-deoxythymidine 5’-triphosphate) EDTA: ethylene-di-amine-tetra-acetic acid EEUU: Estados Unidos de América ELISA: enzyme-linked immunosorbent assay EST: expressed sequence tag FAP: fibronectin-attachment protein FET: Fisher exact test FPA: fluorescence polarization assay FR: flanking region G: guanina GLC: gas-liquid chromatography GM-CSF: granulocyte macrophage colony stimulating factor HEY(M): Herrold’s egg yolk (medium) HHP: high hydrostatic pressure HIV: human immunodeficiency virus HPC: hexadecyl pyridinium chloride HPLC: high performance liquid chromatography HRP: horse radish peroxidase HTST: high temperature short time hsp: heat shock protein IFN: interferón IL: interleukin IS: insertion sequence ITS: internal transcribed spacer IVIAT: in vivo induced antigen technology JNK/SAPK: c-Jun N-terminal kinase/stress activated protein kinase LAM: lipoarabinomanano LFA: leukocyte function antigen LB: linfocito B LJ: Lowënstein-Jensen LT: linfocito T LTh: LT helper MAC: Mycobacterium avium complex
MAIC: Mycobacterium avium-intracellulare complex Maa: Mycobacterium avium subespecie avium Mah: Mycobacterium avium subespecie hominissuis MALDI-tof: Matrix-assisted laser desorption/ionization-time of flight Map: Mycobacterium avium subespecie paratuberculosis MAPK: mitogen-activated protein kinase Mas: Mycobacterium avium subespecie silvaticum MGIT: mycobacterial growth indicator tube mig: macrophage-induced gene MIRU: mycobacterial interspersed repetitive unit MLSSR: multi-locus short sequence repeat MLVA: multi-locus VNTR analysis MMP: matrix metalloproteinase MOTT: mycobacteria other than tuberculosis MPIL: multiplex PCR of IS900 insertion loci MSPD: matrix-solid phase dispersion MTC: Mycobacterium tuberculosis complex NK: natural killer NOD: nucleotide oligomerization domain NRAMP: natural resistance-associated macrophage protein NTM: nontuberculous mycobacteria OADC: oleic acid-albumin-dextrose-catalase OD: optical density OPD: ortho-phenylenediamine ORF: open reading frame PBS: phosphate-buffered saline PCR: polymerase chain reaction PFGE: pulsed field gel electrophoresis PNPP: para-nitrophenyl phosphate PPA: protoplasmatic paratuberculosis antigen PPD: protein purified derivative PPE: Pro-Pro-Glu PRA: PCR-REA Q-RT-PCR: quantitative-real time-PCR RAPD: randomly amplified polymorphic DNA rDNA/rRNA: DNA/RNA ribosómico RFLP: restricted fragment length polymorphism REA: restriction endonuclease analysis RNA: ribonucleic acid ROC: receiver operation characteristic RT: room temperature SDS: sodium dodecyl sulfate SIDA (AIDS): síndrome de inmunodeficiencia adquirida S/P: sample to positive SPARC: secreted protein, acidic and rich in cysteine T: timina TE: Tris-EDTA TGF: transforming growth factor TIMP: tissue inhibitor of metalloproteinase TLC: thin layer chromatography TMB: tetra-methylbenzidine TNF: tumor necrosis factor TRAF: TNF receptor-associated factor tRNA: RNA transferente UK: United Kingdom USA: United States of America VNTR: variable-number tandem repeat


INTRODUCCIÓN GENERAL


INTRODUCCIÓN GENERAL
29
La paratuberculosis o enfermedad de Johne es una enfermedad crónica del intestino
causada por Mycobacterium avium subsp. paratubeculosis (Map). Esta bacteria
patógena pertenece al género Mycobacterium, grupo en el que también se incluyen los
agentes etiológicos de la tuberculosis y la lepra. Es una micobacteria del complejo M.
avium, de gran importancia en veterinaria y medicina por su implicación en infecciones
de animales de granja y pacientes inmunodeprimidos. Sus rasgos distintivos son la
dependencia de micobactina, la baja velocidad de crecimiento, la resistencia a
condiciones adversas, y el elemento genético IS900.
La paratuberculosis afecta principalmente a rumiantes domésticos y salvajes,
adquiriendo mayor importancia entre las especies típicas explotadas por el hombre
(bovino, ovino y caprino) y entre otras menos habituales como el ciervo. No obstante
Map ha sido aislado de infinidad de animales, de otras especies monogástricas como el
hombre, el conejo o el jabalí, y de insectos, lombrices, nemátodos y amebas.
El mecanismo de transmisión principal de la enfermedad es la vía fecal-oral, por
ingestión de alimentos contaminados con heces de animales infectados. También se
conocen otras formas de transmisión como la intrauterina, por ingestión de leche o
calostro de madres infectadas o por semen de toros infectados.
La paratuberculosis se encuentra extendida por todo el mundo, y a pesar de que la
dificultad de su diagnóstico enmascara la realidad, las prevalencias que se calculan son
muy elevadas. En el estado español, donde se conoce su existencia desde hace tiempo,
se considera ampliamente distribuida, y según las aproximaciones de los estudios más
recientes la prevalencia se sitúa entre el 10 y el 86% de explotaciones infectadas según
la especie y la región. Las principales pérdidas económicas ocasionadas por la
paratuberculosis se deben a la reducción de la producción láctea, al sacrificio prematuro
y a la devaluación de los animales sacrificados para su venta. Las pérdidas calculadas
son muy elevadas en países como EEUU, Canadá, Australia, Holanda o Dinamarca.
La posible vinculación de Map en el proceso inflamatorio del intestino humano
conocido como enfermedad de Crohn, fue propuesta hace mucho tiempo, sin embargo,
su frecuente detección mediante el cultivo o técnicas moleculares en pacientes de esta
enfermedad, incluso a partir de muestras de leche y sangre de estos pacientes, han
incrementado notablemente el interés sobre este microorganismo. A pesar de la
existencia de múltiples estudios, ninguno de ellos ha conseguido demostrar o desmentir
totalmente la hipotética implicación de la bacteria causante de paratuberculosis en la
enfermedad de Crohn. La preocupación por el posible riesgo en salud pública, ha
promovido la realización de estudios sobre la presencia de Map en la cadena alimentaria
humana.
La presencia de esta micobacteria ha sido investigada principalmente en leche,
queso y agua como posibles vehículos de transmisión a la especie humana. Gracias a

INTRODUCCIÓN GENERAL
30
este tipo de estudios se ha comprobado la relativamente alta incidencia de Map en estos
productos, así como la ineficacia de tratamientos como la pasterización o la cloración a
la hora de eliminar sus células viables. En consecuencia, la preocupación en relación
con la enfermedad de Crohn sigue en aumento, y se desarrollan nuevos estudios para
garantizar la ausencia de Map en los productos destinados al consumo humano.
El genotipado de cepas es esencial para comprender la epidemiología de
microorganismos patógenos. En el caso de Map la técnica más utilizada en este campo
es la IS900-RFLP, gracias a la que fueron diferenciados por primera vez dos grupos
generales de cepas, las bovinas y las ovinas, así denominados por la aparente mayor
prevalencia de cada tipo en la especie hospedadora. Posteriormente se han desarrollado
otros procedimientos mucho más sencillos para diferenciar esos dos grandes grupos,
como la IS1311 PCR-REA. Además, según algunos trabajos, otra técnica similar a la
del RFLP como es la PFGE también da buenos resultados para llevar a cabo un tipado
más detallado. Antes que la caracterización genotípica, ya se habían observado ciertas
dificultades en el cultivo de esta bacteria de muestras de algunos casos ovinos de
paratuberculosis, y algunos investigadores recomendaban el uso de medios de cultivo
diferentes para sembrar ese tipo de muestras. La distribución regional y de
hospedadores de las diferentes cepas varía según los autores, pero es evidente que las
cepas de tipo bovino están más extendidas y son capaces de infectar a una gran variedad
de especies hospedadoras.
La patogénesis de la paratuberculosis comienza con la entrada de las bacterias a
través del tejido linfoide del intestino delgado (placas de Peyer principalmente) por
mediación de las células M epiteliales. Los bacilos son fagocitados por macrófagos, en
cuyo interior sobreviven y se multiplican, evitando los mecanismos bactericidas de estas
células. Aún no están claros todos los eventos que suceden en la interrelación
hospedador-parásito, ni los posibles factores de virulencia de Map. Según los
conocimientos actuales, tras la infección se abre un período de dos años o más sin
signos de enfermedad, estadios iniciales en los que predomina una respuesta inmune
celular (Th1). Según avanza la enfermedad a estadios clínicos, la respuesta inmune va
haciéndose más específica e incluye ya un componente humoral con producción de
anticuerpos específicos.
El cuadro clínico clásico de la paratuberculosis es una diarrea más o menos
intermitente según la especie, acompañada de pérdida de peso y condición corporal
progresiva, y muerte, pero en estadios subclínicos, los signos son más sutiles y
normalmente pasan desapercibidos. Esta enfermedad provoca una enteritis
granulomatosa cuyas lesiones más evidentes se localizan en los últimos tramos del
intestino delgado y en los nódulos linfáticos asociados.
Entre las técnicas de diagnóstico de la paratuberculosis, se pueden diferenciar las
histopatológicas que identifican las lesiones tisulares, las etiológicas que detectan la

INTRODUCCIÓN GENERAL
31
presencia de la micobacteria (cultivo, PCR,...), y las inmunológicas que tratan de
detectar la respuesta inmune específica del hospedador (interferón-γ,
intradermorreación, ELISA,...). Los métodos más ampliamente utilizados in vivo son el
cultivo fecal y el ELISA, aunque otros como la detección de interferón-γ (IFN-� ) van
tomando mayor protagonismo ya que trata de identificar animales infectados antes de
que presenten sintomatología clínica. No obstante, los métodos actuales más específicos
como la histopatología o el ELISA adolecen de falta de sensibilidad precisamente por su
ineficacia en la identificación de animales con paratuberculosis subclínica, mientras que
los más sensibles como el cultivo, (que es considerado como técnica de referencia pero
que es muy lento y costoso), la estimulación de la producción de IFN-� o la PCR
parecen ser demasiado inespecíficos al menos en términos prácticos de predicción de la
enfermedad clínica.
En general los tratamientos con fármacos en animales no son ni eficaces ni
rentables, por lo que no se recomienda su uso. En el control de esta enfermedad lo más
importante es llevar un manejo encaminado a evitar la introducción y la transmisión de
la enfermedad en el rebaño, haciendo especial hincapié en la higiene en los partos, la
separación de la reposición y de los animales infectados. Las estrategias para erradicar
la enfermedad basadas en programas de detección y eliminación de positivos, han
fracasado principalmente por la falta de sensibilidad de los métodos de diagnóstico y la
mala aplicación de las medidas higiénicas, lo que ha provocado grandes pérdidas. La
estrategia de vacunación ha proporcionado resultados satisfactorios en relación con el
mantenimiento de la producción y la economía del rebaño, sin embargo no es capaz de
eliminar la infección. Actualmente se trabaja en programas que subrayan la
caracterización de los factores de riesgo y la concienciación de ganaderos y veterinarios,
así como en el estudio de nuevos programas de vacunación.


ESCENARIO Y PLANTEAMIENTO DEL TRABAJO DE LA TESIS


ESCENARIO Y PLANTEAMIENTO DEL TRABAJO DE LA TESIS
35
Tal y como refleja la revisión bibliográfica, el trabajo de investigación llevado a
cabo en torno a la paratuberculosis es muy extenso, y el interés sobre ésta y otras
micobacteriosis continúa creciendo. A pesar de este esfuerzo, también quedan reflejadas
las dificultades que entraña la búsqueda de soluciones al gran problema que supone la
paratuberculosis, camino en el que todavía queda mucho por avanzar. El impacto
mundial de esta enfermedad infecciosa en el sector primario es muy importante, como
demuestran su amplia distribución y las pérdidas económicas que derivan de la misma.
Map, la micobacteria responsable de esta afección crónica típica de los rumiantes, ha
sido detectada en infinidad de medios y hospedadores (ya sean transitorios o
definitivos), lo que confirma su éxito de diseminación en ecosistemas diferentes, y hace
más compleja su epidemiología. Los modelos sobre su patogénesis son poco
explicativos, y gran parte del conocimiento sobre este aspecto, está basado en hipótesis
aún sin confirmar. Los mecanismos por los que esta bacteria esquiva las defensas del
organismo hospedador no están claros, pero sus características fisiológicas parecen estar
estrechamente relacionadas con el fallo de la respuesta inmunitaria. Hasta la actualidad
no se han realizado suficientes estudios que contemplen un potencial componente
genético del hospedador que pueda explicar la resistencia/susceptibilidad al desarrollo
de la enfermedad, ni la asociación de ciertos genotipos de Map a determinadas especies
o razas hospedadoras. Por el momento, ninguna de las estrategias de control que se han
llevado a la práctica ha proporcionado resultados totalmente satisfactorios. Una parte
muy importante del fracaso en el entendimiento y el control de esta micobacteriosis,
radica precisamente en la dificultad que implica su correcto diagnóstico, en la
incapacidad de diferenciar a tiempo animales infectados o portadores de animales libres
de la bacteria, en la falta de especificidad y sensibilidad de los métodos empleados. A
raíz de la preocupación generada por el hipotético nexo entre Map y la enfermedad de
Crohn, la paratuberculosis ha adquirido aún más importancia si cabe. La presencia y el
aislamiento de este patógeno en muestras (incluso de sangre) procedentes de pacientes
de Crohn con mayor frecuencia que de personas no aquejadas de ese mal, han
fortalecido esa idea, y la detección de células viables de Map en vehículos potenciales
de transmisión para la especie humana, ha puesto en evidencia la ineficacia de los
tratamientos de desinfección que se aplican y la exposición del ser humano a la bacteria.
Estas cuestiones obligan a la cautela y a establecer una actitud preventiva para evitar ése
contacto.
Neiker lleva muchos años trabajando en el diagnóstico de la paratuberculosis y de
otras micobacteriosis, durante los que ha adquirido una gran experiencia y conocimiento
sobre la enfermedad. Ha trabajado conjuntamente con otros centros nacionales e
internacionales de investigación en este campo y ha organizado reuniones y congresos
también de nivel nacional e internacional. Conoce la situación de la enfermedad en las
explotaciones ganaderas del País Vasco y de otras zonas incluidas en su radio de acción,
además de estudiar la fauna silvestre. Aparte de diagnosticar la enfermedad, en Neiker

ESCENARIO Y PLANTEAMIENTO DEL TRABAJO DE LA TESIS
36
se ha trabajado en la obtención de indicadores de la prevalencia de la enfermedad, en la
mejora de las técnicas de diagnóstico, en la caracterización de la respuesta inmune
frente a la infección, y en la creación y evaluación de medidas de control,
principalmente la vacunación.
OBJETIVOS Como fruto de los conocimientos previamente adquiridos en Neiker y de las
inquietudes que estos han generado en la investigación de la paratuberculosis y de su
agente etiológico Mycobacterium avium subsp. paratuberculosis, se marcaron una serie
de nuevos objetivos:
1. Confirmar la existencia de bacteriemia (o “DNA-hemia”) en el caso de la
paratuberculosis y estudiar su potencial para el diagnóstico epidemiológico
en comparación con el método ELISA.
2. Desarrollar, validar y aplicar métodos de caracterización genética de
aislamientos de Mycobacterium avium subsp. paratuberculosis.
3. Evaluar el uso de métodos alternativos al tratamiento térmico para la
inactivación de Mycobacterium avium subsp. paratuberculosis en leche
destinada al consumo humano.
ESTUDIOS LLEVADOS A CABO
El presente trabajo aborda varias cuestiones de interés diagnóstico y epidemiológico
en las que el nexo es su perspectiva microbiológica convencional o molecular. Éste se
ha llevado a cabo mediante cinco estudios diferentes no contemplados en la revisión
bibliográfica, por ser el objeto de esta tesis:
I. Ramón A. Juste, Joseba M. Garrido, Mariví Geijo, Natalia Elguezabal, Gorka
Aduriz, Raquel Atxaerandio, Iker Sevilla. 2005. Comparison of blood polymerase
chain reaction and enzyme-linked immunosorbent assay for detection of
Mycobacterium avium subsp. paratuberculosis infection in cattle and sheep.
Journal of Veterinary Diagnostic Investigation, 17:354–359: Comparación entre la
PCR y el ELISA de sangre para la detección de la infección de Map en bovino y
ovino. Estudio relacionado con el diagnóstico de la enfermedad en vacas y ovejas,
en el que se evalúa la detección de Map en la sangre de los animales y se compara
con los resultados de ELISA.

ESCENARIO Y PLANTEAMIENTO DEL TRABAJO DE LA TESIS
37
II. Iker Sevilla, Shoor V. Singh, Joseba M. Garrido, Gorka Aduriz, Sara Rodríguez,
María V. Geijo, Richard J. Whittington, Vanessa Saunders, Robert H. Whitlock,
Ramón A. Juste. 2005. Molecular typing of Mycobacterium avium subspecies
paratuberculosis strains from different hosts and regions. Revue Scientifique et
Technique (International Office of Epizootics), 24(3):1061-1066: Tipado
molecular de cepas de Map procedentes de diferentes hospedadores y regiones.
Estudio preliminar de tipado de cepas de Map por medio de IS1311 PCR-REA,
para evaluar su utilidad en el estudio de la epidemiología molecular de la
paratuberculosis.
III. Iker Sevilla, Joseba M. Garrido, Mariví Geijo, Ramón A. Juste. 2006. Pulsed-field
gel electrophoresis profile homogeneity of Mycobacterium avium subsp.
paratuberculosis isolates from cattle and heterogeneity of those from sheep and
goats. BMC Microbiology, 2007, 7:18: Homogeneidad de perfiles de
electroforesis en gel de campo pulsado (PFGE) de los aislados bovinos de Map, y
heterogeneidad de los de ovejas y cabras. Caracterización exhaustiva de un
importante número de aislados de Map por medio de PFGE e IS1311 PCR-REA,
para conocer aspectos epidemiológicos de las cepas de paratuberculosis aisladas
en varias regiones (principalmente en la Península Ibérica).
IV. Iker Sevilla, Martha I. Murcia, Joseba M. Garrido, Douwe Bakker, Maria J.
García, Ramón A. Juste. 2006. Detection and pathological implications of the
presence of IS900 in human Mycobacterium avium isolates. Preparando su envío a
Journal of Clinical Microbiology: Detección e implicaciones patológicas de la
presencia de IS900 en aislados humanos de Mycobacterium avium. Estudio en el
que se informa de la presencia de la secuencia de inserción IS900 específica de
Map en aislados de Mycobacterium avium procedentes de pacientes de SIDA de
Colombia, y las consecuentes implicaciones en el diagnóstico y la patología de las
técnicas de detección molecular basadas en IS900.
V. Tomás López-Pedemonte, Iker Sevilla, Joseba M. Garrido, Gorka Aduriz,
Buenaventura Guamis, Ramón A. Juste, Artur X. Roig-Sagués. 2006. Inactivation
of Mycobacterium avium subsp. paratuberculosis in cow’s milk by means of high
hydrostatic pressure at mild temperatures. Applied and Environmental
Microbiology, 72(6):4446–4449: Inactivación de Map en leche de vaca por medio
de altas presiones hidrostáticas a temperaturas moderadas. Estudio sobre la
alternativa del uso de la presión para inactivar Map en la leche, como medida de
control en relación con la enfermedad de Crohn, que podría ser aplicado también a
otros productos.


REVISIÓN DE LA BIBLIOGRAFÍA


REVISIÓN DE LA BIBLIOGRAFÍA
41
ETIOLOGÍA
Género Mycobacterium
El agente causal de la paratuberculosis o enfermedad de Johne, Mycobacterium
avium subsp. paratuberculosis (Map), es una bacteria perteneciente al género
Mycobacterium (Lehmann y Neumann 1896). En ocasiones, las micobacterias presentan
un crecimiento filamentoso o pseudomiceliar, y aunque no forman estructuras de tipo
fúngico (esporas, conidias, cápsulas, hifas), el género debe su denominación al aspecto
fungoide de las colonias de M. tuberculosis. Además de micobacterias patógenas
estrictas de los humanos y los animales como M. tuberculosis, M. leprae, M. bovis, M.
caprae o Map, también existen micobacterias patógenas oportunistas y micobacterias
saprofitas ambientales. M. avium y M. intracellulare son importantes patógenos
causantes de diversas enfermedades que también afectan a un amplio rango de
hospedadores, entre ellos, el hombre. Estas y otras especies como M. haemophilum, M.
kansasii, M. malmoense, M. simiae, M. szulgai, M. xenopi, M. genavense son causantes
de infecciones en individuos inmunodeprimidos o inmunocompetentes con
predisposición a la infección. M. scrofulaceum provoca linfoadenitis en niños, M.
ulcerans es responsable de lesiones graves de la piel, y M. marinum causa el granuloma
de estanque. Otras como M. fortuitum y M. abscessus son responsables de infecciones
más esporádicas relacionadas con heridas traumáticas o quirúrgicas, e infecciones de
piel o de los pulmones (Falkinham III 1996, Rastogi y cols. 2001, Tortoli 2003).
Morfología externa
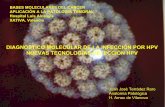
Se trata generalmente de bacilos rectos o
ligeramente curvados de 1-10 µm de largo y
0,2-0,6 µm de ancho, inmóviles y
considerados Gram positivos a pesar de la
escasa penetración de colorante por medio de
esta técnica. Esto es debido a la presencia de
ácidos micólicos en la compleja envoltura
celular de que están provistos, gracias a la cual
poseen su característica resistencia (Figura 1).
Esta envoltura está constituida por tres
componentes (Rastogi y cols. 2001): La
cápsula, cuya síntesis parece estar controlada
por mecanismos reguladores dependientes del
huésped en micobacterias patógenas. La pared
celular, formada por unión covalente entre
peptidoglicano, arabinogalactano y los ácidos
Figura 1. Detalle de los componentes de la pared celular de M. tuberculosis: ácidos micólicos, lipoarabinomanano (LAM), peptidoglicano, fosfatidil-inositol-manósido (PIM),… [http://oregonstate.edu/~mahmudt/pictures/Mycobacterium%20cell%20envelope.jpg]

ETIOLOGÍA
42
micólicos. Y por último, la membrana citoplasmática, cuya composición es similar a la
de otras bacterias. El lipoarabinomanano es un fosfolípido unido a la membrana que
atraviesa toda la envoltura celular y cuya composición difiere entre las distintas especies
micobacterianas (Daffe y cols. 1998).
Metabolismo
Las micobacterias son microorganismos aerobios o microaerófilos,
quimioorganotrofos, y su temperatura óptima de crecimiento se sitúa entre 30 y 45ºC
(Wayne y cols. 1986, Levy-Frebault y cols. 1992). Utilizan glicerol o piruvato como
fuente de carbono y energía, y como fuente de nitrógeno pueden utilizar asparagina,
glutamato y amonio (Goodfellow y cols. 1998). Son prototrofas para todos los
aminoácidos, purinas, pirimidinas y vitaminas del complejo B. Producen exoquelina y
micobactina, dos sideróforos necesarios para metabolizar el hierro.
Los medios sólidos de cultivo en los que son aisladas son con base de huevo
(Herrold, Löwenstein-Jensen, Stonebrink, Petragnani, Coletsos, Ogawa kudoh,...) o con
base sintética (Middlebrook 7H10 o 7H11). También crecen en medios líquidos como
Middlebrook 7H9, Watson-Reid, Dubos, Sula, Sauton, o Proskauer-Beck. Dependiendo
de la especie que se pretenda cultivar se puede suplementar los medios con micobactina,
hemina, ácido oleico, albúmina, dextrosa, catalasa u otras sustancias (Goodfellow y
cols. 1998). A pesar de ello, la aplicación de métodos convencionales de microbiología
resulta dificultosa, debido a su lentitud de crecimiento y a su tendencia a formar grumos
o a crecer mal en suspensión (Juste y cols. 2000).
Composición genética
El genoma micobacteriano es rico en guanina (G) y citosina (C) (61-71%) (Levy-
Frebault y cols. 1992). Se conoce la secuencia completa de dos cepas de M. tuberculosis
(Cole y cols. 1998, Fleischmann y cols. 2002), y las de M. leprae (Eiglmeier y cols.
2001, Cole y cols. 2001), M. bovis (Garnier y cols. 2003) y Map (Li y cols. 2005)
(Tabla 1). Contiene secuencias de inserción (IS), elementos que han resultado muy
útiles en la identificación y diferenciación de especies. Estos pequeños tramos de DNA
transponible que pueden encontrarse varias veces repetidas a lo largo del genoma,
tienen en sus extremos secuencias flanqueantes repetidas que están involucradas en su
transposición. En medio de esas secuencias terminales repetidas hay genes involucrados
en la transposición y secuencias que pueden controlar la expresión de los genes, pero no
presentan otros genes que no sean esenciales (Mayer 2005). Las de mayor interés en
medicina son IS6110 (Thierry y cols. 1990), IS1081 (Collins y cols. 1991), IS1245
(Guerrero y cols. 1995), IS1547 (Fang y cols. 1999), IS1311 (Guerrero y cols. 1994),
IS900 (Green y cols. 1989), IS901 (Kunze y cols. 1991), IS902 (Moss y cols. 1992a) y
ISMav2 (Strommenger y cols. 2001).

REVISIÓN DE LA BIBLIOGRAFÍA
43
Tabla 1. Características de los genomas de Map, M. avium, M. tuberculosis, M. bovis, M. leprae, y M. smegmatis (Li y cols. 2005).
Organismo Map M. avium M. tuberculosis M. bovis M. leprae M. smegmatis
Tamaño genoma (bp) 4.829.781 5.475.738 4.411.532 4.345.492 3.268.203 6.988.209 Contenido G+C (%) 69,30 68,99 65,61 65,63 57,79 67,40 Codifican proteínas (%) 91,30 NA 90,80 90,59 49,50 92,42 ORFs 4.350 NA 3.959 3.953 1.604 6.897 Densidad génica (bp/gen) 1.112 NA 1.114 1.099 2.037 1.013 Longitud génica media (bp) 1.015 NA 1.012 995 1.011 936 tRNAs 45 45 45 45 45 47 Operones rRNA 1 1 1 1 1 2
Aunque no se conoce bien su importancia biológica, en muchas especies
micobacterianas (M. avium, M. intracellulare, M. scrofulaceum, M. fortuitum, M.
peregrinum, M. abscessus, M. chelonae, M. xenopi, M. celatum, M. branderi) pueden
encontrarse comúnmente plásmidos, de forma notable en M. avium, pero en otras como
M. tuberculosis o Map no es así (Crawford y cols. 1981, Erardi y cols. 1987, Jensen y
cols. 1989, Jucker y cols. 1990, Dale 1995, Picardeau y cols. 1997). A pesar de ello, se
han podido demostrar ejemplos de transferencia genética lateral en Map (Marri y cols.
2006), tal y como ocurre en la mayoría de bacterias.
Clasificación taxonómica
En cuanto a la clasificación taxonómica de las micobacterias, el género es el único
representante de la familia Mycobacteriaceae, que se integra en el suborden
Corynebacterinae, orden Actinomycetales, subclase Actinobacteridae, clase
Actinobacteria, división Firmacutes, y superreino Bacteria (Stackebrandt y cols. 1997).
Durante años, los estándares mínimos para la inclusión en este género han sido
resistencia al alcohol ácido, un rango del 61-71% de contenido G+C en el DNA
cromosómico, y la presencia de ácidos micólicos que son degradados por pirolisis a
metil-ésteres de ácidos grasos de C22 a C26 (Levy-Frebault y cols. 1992). Las
recomendaciones actuales para la descripción de nuevos taxones micobacterianos
incluyen varias pruebas (Tortoli 2003):
- Pruebas de cultivo: temperatura de crecimiento, velocidad de crecimiento, tipo de
pigmentación, morfología de colonias, y para los organismos de crecimiento
rápido, tolerancia a NaCl y crecimiento en MacConkey.
- Pruebas bioquímicas: arilsulfatasa de 3 días, reducción de nitrato, hidrólisis de
Tween 80, hidrólisis de urea, catalasa a 68ºC, y catalasa semicuantitativa como
más utilizados, pero sin restar importancia al análisis lipídico por medio de
cromatografía de capa fina (TLC), cromatografía gas-líquido (GLC) y
cromatografía líquida de alta resolución (HPLC).
- Pruebas genéticas: hibridación DNA-DNA (en desuso), y secuenciación del gen
16S rRNA completo, o de al menos las primeras 500 bp.
En los últimos 50 años han sido propuestos muchos agrupamientos y modelos para
la clasificación de agentes micobacterianos atendiendo a propiedades de crecimiento,

ETIOLOGÍA
44
bioquímicas y moleculares (Shinnick y cols. 1994, Wayne y cols. 1996, Falkinham III
1996, Rastogi y cols. 2001, Tortoli 2003). No obstante el esquema basado en
características de crecimiento y pigmentación propuesto por el grupo de Runyon (Timpe
y cols. 1954, Runyon 1959), sigue vigente en la actualidad, principalmente debido a la
concordancia generalizada de éste con los modelos genotípicos.
De este modo, han sido descritos dos grandes grupos de micobacterias: las de
crecimiento rápido, que forman colonias claramente visibles en menos de 7 días, y las
de crecimiento lento, que precisan más de 7 días para conseguirlo (Wayne y cols. 1986,
Stahl y cols. 1990, Shinnick y cols. 1994). Las especies patógenas más importantes,
tanto para el humano como para los animales, se encuentran entre las de crecimiento
lento. Es el caso de las especies responsables de tuberculosis, lepra, linfadenitis cervical
juvenil, infecciones oportunistas en pacientes inmunodeprimidos, y otras enfermedades
relacionadas (Stahl y cols. 1990, Shinnick y cols. 1994). Sin embargo, la infección con
micobacterias de crecimiento rápido, también ha sido descrita en la especie humana
(Griffith y cols. 1993, Tortoli 2003, Freudenberger y cols. 2006) así como en especies
animales (Green y cols. 2000, Malik y cols. 2000, Tortoli 2003, Malik y cols. 2004). En
Figura 2. Árbol filogenético de las micobacterias de crecimiento lento. No se incluyen las especies M. visibilis ni M. lepraemurium, de las cuales sólo se conoce parte de la secuencia del 16S rDNA. sqv., secuevar; MSR, relacionados con M. simiae; MTR, relacionados con M. terrae; SG, de crecimiento lento (Tortoli 2003).

REVISIÓN DE LA BIBLIOGRAFÍA
45
la mayoría de casos M. fortuitum, M. chelonae y M. abscessus son los agentes
oportunistas implicados. Este tipo de infecciones están frecuentemente relacionadas con
heridas postraumáticas y/o postoperatorias, además de las asociadas a enfermedades
pulmonares crónicas e infecciones cutáneas en personas inmunodeprimidas (Falkinham
III 1996).
Otra clasificación basada en la importancia en medicina humana es la que distingue
entre el grupo de micobacterias del complejo M. tuberculosis (MTC: M. tuberculosis,
M. africanum, M. bovis, M. microti y M. caprae) y las micobacterias no tuberculosas,
atípicas o distintas de la tuberculosis (NTM ó MOTT) (Inderlied y cols. 1993,
Falkinham III 1996, Rastogi y cols. 2001).
Actualmente, el abordaje taxonómico y filogenético más ampliamente aceptado es el
de la secuenciación del gen que codifica el 16S rRNA (Bercovier y cols. 1986,
Boddinghaus y cols. 1990, Rogall y cols. 1990, Kirschner y cols. 1993, Turenne y cols.
2001), centrado en dos secuencias hipervariables conocidas como regiones A y B.
Como resultado del análisis de todas las secuencias de 16S rDNA descritas en el género
Mycobacterium, Tortoli obtuvo un árbol filogenético que clasifica las cerca de 100
especies conocidas en 5 grandes grupos (Tortoli 2003) (Figuras 2 y 3).
Además, también ha sido propuesta la clasificación del género en 4 grupos,
dependiendo del número de operones ribosómicos por genoma y de la longitud de la
hélice 18 del 16S rDNA (Menendez y cols. 2002). Sin embargo, la concordancia entre
estos grupos y los caracterizados en base a su velocidad de crecimiento no es total.
Identificación
Debido a la similitud entre especies, la identificación de miembros del género es a
veces dificultosa, a lo que se suma la incapacidad de los métodos microbiológicos
clásicos. Los avances en métodos moleculares están facilitando este proceso, algo que
resulta imprescindible a la hora de intentar controlar las infecciones causadas por estos
microorganismos.
Los eventos evolutivos de los organismos quedan impresos en las secuencias del
rRNA. El análisis de estas secuencias, forma la base para la identificación de los
microorganismos (Kirschner y cols. 1998). La secuenciación de ciertas regiones del 16S
rDNA permite identificar con fiabilidad la especie a la que pertenece una bacteria en
particular. Sin embargo, existen entre las micobacterias excepciones como M. kansasii y
M. gastri cuya secuencia es idéntica, a pesar de ser fenotípicamente diferentes
(Kirschner y cols. 1998, Soini y cols. 2001). Wilton y cols. describieron un método de
PCR por el que se puede determinar si un aislado pertenece al género Mycobacterium e
identificar la especie concreta simultáneamente (Wilton y cols. 1992). Se basa en la
amplificación de regiones conservadas y variables del 16S rDNA, método que en
combinación con la amplificación del gen que codifica para la proteína MPB70

ETIOLOGÍA
46
(específica del MTC) permite la identificación de micobacterias clínicamente
importantes en una sola PCR. Este gen ribosómico también ha resultado de utilidad por
medio de análisis de restricción con endonucleasas de los productos de PCR (PCR-REA
o PRA) (Thierry y cols. 1990).
Otra posibilidad con el material genético ribosómico, es la secuenciación de la
región intergénica 16S-23S (ITS-1), zona muy variable aun entre grupos taxonómicos
relacionados (Gurtler y cols. 1996). Mediante esta técnica es posible diferenciar varios
taxones intra-especie (secuevares) en función de las distintas secuencias que se obtienen
Figura 3. Árbol filogenético de las micobacterias de crecimiento rápido. sqv., secuevar; TTRG, termotolerantes de crecimiento rápido; RG, de crecimiento rápido (Tortoli 2003).

REVISIÓN DE LA BIBLIOGRAFÍA
47
de esa región (De Smet y cols. 1995). Esta zona es de gran variabilidad entre las
micobacterias de crecimiento lento (Roth y cols. 1998). Los resultados obtenidos dentro
del complejo M. avium-intracellulare han sido satisfactorios (Frothingham y cols.
1994), y se han descrito 7 secuevares de M. avium (de Mav-A a Mav-G), y 35
secuevares en total dentro del complejo (De Smet y cols. 1995, Novi y cols. 2000, Mijs
y cols. 2002, Tortoli y cols. 2004, Lebrun y cols. 2005, Murcia y cols. 2006). Se ha
observado una fuerte asociación entre M. avium secuevar Mav-B, con SIDA y con
linfoadenitis en niños (De Smet y cols. 1995), aunque en pacientes HIV positivos de
países como Colombia no parece que esa distribución sea tan clara (Murcia 2004). La
restricción por endonucleasas de amplificados de esta región ha sido también objeto de
estudio (Dvorska y cols. 2001). Existe un kit comercial renovado llamado INNO-LiPA
MYCOBACTERIA v2 (Innogenetics) que se basa en la detección de esos secuevares
por hibridación inversa (Tortoli y cols. 2003).
La secuenciación y/o PCR-REA de los genes recA (asociado a reparación del DNA)
y rpoB (codificante para la RNA polimerasa) ha resultado de utilidad en la
identificación de M. leprae, M. aurum, M. mucogenicum, M. kansasii, M. gastri, M.
xenopi, M. asiaticum, M. shimoidei, M. gordonae, M. celatum, M. fortuitum y miembros
del MTC (Dvorska y cols. 2001).
La caracterización del gen que codifica la proteína de choque térmico de 65 kDa
(hsp65) también ha sido utilizada a la hora de identificar y diferenciar especies y
subespecies de micobacterias, ya sea por medio de análisis de restricción de
amplificados o de secuenciación (Telenti y cols. 1993, Swanson y cols. 1997, Devallois
y cols. 1997, Ringuet y cols. 1999, Leao y cols. 1999, Oliveira y cols. 2000, Leclerc y
cols. 2000, Smole y cols. 2002, Oliveira y cols. 2003, McNabb y cols. 2004, Murcia y
cols. 2004, Cavusoglu y cols. 2005, Turenne y cols. 2006). Los mismos métodos han
dado resultados parecidos al ser aplicados al gen dnaJ, que codifica para una proteína de
choque de frío (Dvorska y cols. 2001).
Algunos genes relacionados con la resistencia-sensibilidad de las micobacterias a
ciertas drogas y medicamentos (katG, pncA, oxyR, ahpC) también han sido utilizados
para diferenciar cepas, principalmente del MTC (Dvorska y cols. 2001).
Otra posibilidad en la identificación de aislados de interés clínico (MTC, M. avium-
intracellulare, M. kansasii, M. gordonae) es la utilización de sondas comerciales
AccuProbe (GenProbe) (Reisner y cols. 1994, Devallois y cols. 1997, Hall y cols.
2004), en las que sondas quimioluminiscentes de DNA de cadena simple hibridan con
RNA de la especie problema. Su inconveniente principal es su elevado coste (Soini y
cols. 2001).
Como ya se ha indicado anteriormente, las secuencias de inserción también han
demostrado su validez a la hora de identificar especies, por medio de la amplificación
de un fragmento por PCR (Dvorska y cols. 2001). Otros genes o secuencias específicas

ETIOLOGÍA
48
de especie o subespecie también son utilizados en la identificación de algunas
micobacterias, como en el caso del gen mtp40 exclusivo de M. tuberculosis y la región
JB21-JB22 de M. bovis, o PCR-REA del gen gyrB del MTC (Dvorska y cols. 2001).
Una buena manera de determinar la especie o subespecie es hacer varias PCRs que
amplifiquen fragmentos del 16S rDNA, secuencias de inserción (IS901, IS1245,
IS900,...) y genes específicos (hspX), como el panel de PCRs descrito por Ellingson y
cols. (Ellingson y cols. 2000).
Tipado molecular
El tipado molecular de los microorganismos tiene por objeto determinar la relación
entre aislamientos de un mismo grupo, saber si proceden del mismo foco, cómo circula
en el ecosistema, y cómo es transmitido, en definitiva, estudiar parte de su
epidemiología. En micobacteriología, el tipado de cepas por métodos basados en la
detección de secuencias de inserción ha sido de gran ayuda.
El análisis de polimorfismos en la longitud de fragmentos de restricción (RFLP)
cortados con diferentes endonucleasas, y detectados con diferentes sondas, es el método
más extendido y estandarizado. Existen protocolos estándares para el tipado de
miembros del MTC (IS6110-RFLP) (Thierry y cols. 1990, Van Embden y cols. 1993),
algunos del complejo M. avium (IS1245-RFLP, IS1311-RFLP, IS901-RFLP) (Roiz y
cols. 1995, van Soolingen y cols. 1998, Dvorska y cols. 2003), y Map (IS900-RFLP)
(Pavlik y cols. 1999a). En el caso del IS6110-RFLP, la técnica no se puede aplicar a
todas las cepas de ese complejo, pues hay algunas que no poseen ninguna copia de la
misma (Cave y cols. 1991, Agasino y cols. 1998). Según Kremer y cols. (Kremer y cols.
2002) esta técnica junto con la PCR “mixed-linker” relacionada con esta misma
secuencia, poseen el mayor poder discriminatorio cuando se trata de cepas con muchas
copias de IS6110. Cuando son menos copias por genoma, puede obtenerse un resultado
mejor por medio de otras técnicas. En el caso de Map, fue diseñada posteriormente la
PCR múltiple de los loci de IS900 (MPIL), que daba resultados parecidos a los del
RFLP (Bull y cols. 2000).
Una de las técnicas más populares que se ha desarrollado es el espoligotipado
(Groenen y cols. 1993), que se basa en polimorfismos en un locus cromosómico
caracterizado por su elevado número de repeticiones directas (DR) conservadas.
Mediante este método han sido tipadas cepas de M. tuberculosis, M. bovis, M. caprae,
M. microti y M. canetti (Dvorska y cols. 2001).
Recientemente, ha sido demostrada la utilidad de otro método denominado
“unidades repetitivas intercaladas micobacterianas-repeticiones en tándem de número
variable” (MIRU-VNTR), en estudios de tipado de cepas de M. tuberculosis y M. bovis
(Supply y cols. 2006, Allix y cols. 2006). Está basado en la variabilidad del número de
repeticiones en tándem que al mismo tiempo conforman lo que se conoce como

REVISIÓN DE LA BIBLIOGRAFÍA
49
unidades intergénicas micobacterianas (Supply y cols. 1997, Frothingham y cols. 1998).
Estos son similares a los minisatélites de las células eucariotas (Supply y cols. 1997).
En el caso de MTC, se trata de amplificar y determinar por el tamaño el número de
copias en tándem en 12 loci polimórficos conocidos que se encuentran presentes en
todas las cepas (Supply y cols. 2000, Mazars y cols. 2001). Este método también ha sido
empleado con Map y M. avium (Bull y cols. 2003b), especies en las que se han
detectado 18 loci conservados, de los cuales 6 son diferentes para las dos especies. Por
este método las cepas de Map y las de M. avium se diferenciaban en 2 y 5 grandes
grupos respectivamente. Overduin y cols. consiguieron subdividir el grupo de IS900-
RFLP mayoritario de Map en 6 subgrupos, por medio de análisis multilocus-VNTR
(MLVA) (Overduin y cols. 2004). Sin embargo, 17 tipos diferentes de RFLP
compartían un mismo tipo de MLVA. Con este método, detectaron diferencias en el
número de repeticiones en tándem, así como mutaciones en las secuencias de esas
unidades y en las regiones que las flanquean. También se ha usado recientemente en el
tipado de aislados de M. ulcerans (Ablordey y cols. 2005). Por un método muy similar
de amplificación y secuenciación de zonas repetitivas en busca de polimorfismos, ha
sido descrito el análisis de repeticiones multilocus de secuencia corta (MLSSR) para el
tipado de Map (Amonsin y cols. 2004), y posteriormente se ha reducido al análisis de
los loci con mayor poder de discriminación (Motiwala y cols. 2003, Motiwala y cols.
2004, Ghadiali y cols. 2004, Motiwala y cols. 2005, Corn y cols. 2005). En la puesta a
punto, se detectaron 78 repeticiones perfectas de las que 11 polimórficas fueron
seleccionadas, con 3,2 posibles alelos por locus. Según el índice de Simpson calculado
(Amonsin y cols. 2004), este es el método con mayor poder de discriminación para el
tipado de cepas de Map, por encima del IS900-RFLP, el MPIL y el análisis de
polimorfismos en la longitud de fragmentos amplificados (AFLP). A pesar de ello, su
fiabilidad deberá ser probada comparando sus resultados con los del resto de técnicas en
un número representativo de cepas y de orígenes.
Otra técnica utilizada frecuentemente en el tipado de micobacterias, es el análisis de
DNA polimórfico aleatoriamente amplificado (RAPD) (Mazurier y cols. 1992). Su
ventaja es que se basa en la amplificación arbitraria de fragmentos de DNA no
determinados previamente, por lo que no es necesario conocer bien el genoma de las
bacterias. Ha sido de utilidad en el tipado de M. tuberculosis (Linton y cols. 1994) y
otras especies como M. avium (Matsiota-Bernard y cols. 1997, Matsiota-Bernard y cols.
2000, Pillai y cols. 2001), M. abscessus, M. chelonae (Zhang y cols. 1997), M. szulgai
(Zhang y cols. 2002) y M. malmoense (Kauppinen y cols. 1994). No obstante, puede
resultar poco reproducible, y en el caso de Map no aporta información adicional (Pillai
y cols. 2001).
Otra posibilidad muy extendida en el tipado de micobacterias es la electroforesis en
gel de campo pulsado (PFGE). Se extrae todo el DNA genómico del aislamiento dentro
de bloques de agarosa, se digiere con un enzima de restricción, y los fragmentos

ETIOLOGÍA
50
resultantes se separan en un gel de agarosa por medio de pulsos eléctricos alternos en
tiempo y dirección. Esto permite separar fragmentos de gran tamaño, y se puede realizar
con diferentes enzimas sobre la misma cepa, con lo que se obtienen perfiles con
patrones múltiples. Ha sido aplicado con éxito en el tipado de M. fortuitum (Burns y
cols. 1991), M. tuberculosis (Zhang y cols. 1992, Singh y cols. 1999), M. avium, M.
intracellulare (Levy-Frebault y cols. 1989, Coffin y cols. 1992, Mazurek y cols. 1993,
Feizabadi y cols. 1997a), M. chelonae, M. abscessus (Wallace, Jr. y cols. 1993), M.
haemophilum (Yakrus y cols. 1994), M. bovis (Feizabadi y cols. 1996), M. kansasii
(Iinuma y cols. 1997) y Map (Stevenson y cols. 2002, de Juan y cols. 2005). La
reproducibilidad es muy buena y sus resultados son muy consistentes ya que explora el
genoma completo. Gracias al PFGE fueron demostradas infecciones policlonales de M.
avium en pacientes de SIDA (Arbeit y cols. 1993, Slutsky y cols. 1994). Para emplear
este método es necesario contar con cultivos puros de calidad y cantidad, lo que se
complica en el caso de algunas micobacterias. El inconveniente principal es su elevado
coste, tanto en material y reactivos, como en tiempo y mano de obra.
Complejo M. avium
Un importante grupo entre las micobacterias de crecimiento lento es el complejo M.
avium o MAC (complejo M. avium-intracellulare o MAIC, para algunos), donde se
incluye Map. Todos los componentes del complejo son de crecimiento lento, y
pertenecen al grupo NTM (Inderlied y cols. 1993). En la actualidad, se acepta que MAC
está compuesto de las especies M. avium y M. intracellulare, aunque una importante
proporción de las cepas incluidas en el complejo no pueda ser clasificada en ninguna de
las dos (Wayne y cols. 1993, Frothingham y cols. 1994), y se han denominado como
“otras MAC” (Smole y cols. 2002). Entre estas “otras MAC” han sido recientemente
elevadas a nivel de especie M. chimaera (Tortoli y cols. 2004) y M. colombiense
(Murcia y cols. 2006).
M. avium es responsable de cerca del 90% de las infecciones por MAC en pacientes
infectados por el HIV, y provoca infecciones en animales (muy importantes en aves)
(Thorel y cols. 1990, Falkinham III 1996, Pavlik y cols. 2005). M. intracellulare se
asocia con infecciones pulmonares menos patógenas que las producidas por M. avium, y
se aísla más frecuentemente de enfermos no afectados por el SIDA (Yamori y cols.
1992, Falkinham III 1996). También infecta animales, a veces de manera persistente,
pero normalmente la infección no desencadena enfermedad clínica, y sólo se detecta en
la necropsia (Thorel y cols. 2001). M. chimaera ha sido únicamente aislado de humanos
con infecciones pulmonares no infectados por el HIV, y su denominación se debe a que
comparte características de las dos especies principales del complejo (Tortoli y cols.
2004). Se diferencia de éstas en base a su HPLC, pues los secuevares de 16s rDNA y de
ITS-1 se corresponden con secuevares incluidos en alguna de esas dos especies. Por
último, M. colombiense ha sido aislada en pocas ocasiones de sangre (o esputos) de

REVISIÓN DE LA BIBLIOGRAFÍA
51
pacientes colombianos infectados por el HIV (Murcia y cols. 2006). Produce ureasa, a
diferencia del resto de miembros de MAC. También se diferencia del resto por su
HPLC, su secuevar de ITS-1, su secuevar de 16s rDNA, y por medio de hibridación
DNA-DNA.
Historia
En la primera descripción de la enfermedad de la paratuberculosis (Johne y cols.
1895), el agente fue caracterizado como una micobacteria de tipo aviar. Posteriormente,
debido a las particularidades de la afección, pasó a considerarse como una enteritis
pseudotuberculosa bovina, y la bacteria, una entidad diferenciada de la de tipo aviar y
designada Mycobacterium enteriditis chronicae pseudotuberculosae bovis (Twort y
cols. 1912). Más tarde, el microorganismo recibió la denominación de M.
paratuberculosis (Bergey y cols. 1923) y M. johnei (Francis 1943), nombres que
actualmente aún son utilizados. En general, la identificación de las micobacterias es
dificultosa, y el desacuerdo surgido a la hora de determinar si Map es una especie
independiente de M. avium o no, provocó una gran polémica en torno a su nomenclatura
y situación taxonómica (Juste y cols. 2000).
Las peculiaridades fenotípicas de la bacteria hicieron que ésta fuese contemplada
como un agente diferente de la micobacteria aviar durante largo tiempo, hasta que la
alta homología genética detectada entre estas dos entidades y las llamadas micobacterias
de la paloma torcaz (Saxegaard y cols. 1988), reabriera el debate al sugerir que se
trataba de la misma especie. Los resultados de ese estudio concluían en la
recomendación de clasificar los tres agentes como subespecies diferentes dentro del
mismo complejo. Finalmente, en 1990, Thorel y cols. propusieron la taxonomía
oficialmente aceptada en la actualidad, apoyándose en los estudios previos de
caracterización genética y fenotípica, y en su exhaustivo trabajo de taxonomía numérica
(Thorel y cols. 1990). De este modo, M. avium quedaba dividido en las subespecies M.
avium subsp. avium (Maa), Map y M. avium subsp. silvaticum (Mas), garantizando así
la representación de la homología observada entre ellas, pero sin olvidar las propiedades
distintivas de cada una.
Dejando a un lado las características comunes a las 3 subespecies, las diferencias
más marcadas son las relativas a sus requerimientos de cultivo, su poder patógeno y su
rango de hospedadores (Thorel y cols. 1990). Maa se aísla frecuentemente de muestras
ambientales y crece sin necesidad de micobactina. Es la responsable de tuberculosis en
aves, puede causar infección a un amplio espectro de animales, y en humanos las
principales patologías son infección pulmonar en adultos, adenopatías submandibulares
en niños y septicemia en pacientes de sida. La cepa tipo es la ATCC 25291 ó CIP
104244. El distintivo más práctico de Map es su dependencia de micobactina para
crecer in vitro y su extremadamente baja velocidad de crecimiento. Es patógena
obligada, afecta principalmente a rumiantes causándoles enteritis crónica

ETIOLOGÍA
52
(paratuberculosis) y parece estar implicada en la enfermedad de Crohn. La cepa tipo es
la ATCC 19698 ó CIP 103963. En Mas destaca su incapacidad de propagación en
medios de cultivo con base de huevo, la estimulación de crecimiento inducida por un
pH 5,5 y su independencia de micobactina (excepto en el aislamiento primario, según
distintos autores). Es también patógena obligada, provocando tuberculosis en aves y
paratuberculosis en mamíferos (Thorel y cols. 1990).
Recientemente, se ha planteado distinguir taxonómicamente las cepas de Maa
halladas en humanos y otros mamíferos como el cerdo, de las cepas de Maa típicamente
responsables de la tuberculosis aviar (Mijs y cols. 2002), sustentándose en diferencias
genéticas y fenotípicas. Se propone mantener la designación de Maa para las cepas
típicas aisladas de aves, y llamar a las de origen humano-porcino M. avium subsp.
hominissuis (Mah). Según Mijs y cols. además de la preferencia por el hospedador, las
características distintivas de Maa serían un IS1245-RFLP típico de 3 bandas (8.7, 3.2,
2.9 kbp), ser tipo Mav-A en ITS-1 (C en la posición 228), poseer 7-11 copias de IS901,
y la incapacidad de crecer a 24 y 45 ºC. Mientras que las características de Mah serían
patrones polimórficos en IS1245-RFLP con múltiples copias de la IS, predominancia
del secuevar Mav-B (G en la posición 228) en la ITS-1, ausencia casi siempre de la
IS901, y capacidad de crecer tanto a 24 como a 45 ºC. Parece estar ya ampliamente
aceptado el nombre de Mah para designar las cepas humano-porcinas (Mobius y cols.
2006, Bartos y cols. 2006), sin embargo, la descripción de cepas de M. avium que no
encajan completamente en esas definiciones (Pavlik y cols. 2000e, Murcia 2004),
mantiene abierta la posibilidad de más cambios, a medida que se vayan descubriendo
nuevos agrupamientos. El grupo de Pavlik y cols. además de usar el serotipado,
distinguen los miembros de MAC por métodos moleculares: Maa da positivo en IS901-
PCR y IS1245-PCR, y negativo en FR300-PCR (secuencia flanqueante de IS901; aquí
negativo es cuando se obtiene una banda de 1742 bp), Mah en cambio no posee IS901, y
es positivo en IS1245-PCR y FR300-PCR (Pavlik y cols. 2005, Bartos y cols. 2006).
Para añadir mayor controversia a este asunto, se ha descubierto recientemente que 2 de
las 3 bandas del patrón de IS1245-RFLP típico de las cepas aviares se debe a la
hibridación inespecífica de la sonda utilizada con copias de la IS1311 (Johansen y cols.
2005). Además, los resultados de un detallado estudio en el que son caracterizadas (ITS-
1, IS901, hsp65, RFLP) varias cepas de M. avium, sugieren que Maa y Mas son
diferentes cepas de una misma entidad y que su diferenciación en subespecies puede no
estar suficientemente justificada (Turenne y cols. 2006).
Mycobacterium avium subsp. paratuberculosis
Morfología y metabolismo
El agente causal de la paratuberculosis (Figura 4) es un bacilo de 1-2 µm de
longitud por 0,5 µm de anchura, que normalmente forma colonias rugosas.

REVISIÓN DE LA BIBLIOGRAFÍA
53
Generalmente presenta una tinción homogénea con los métodos de tinción y
decoloración con alcohol ácido (como el de Ziehl-Neelsen), aunque en el caso de
formas de pared deficiente o inexistente (McGee y cols. 1971, Imaeda 1984, Chiodini y
cols. 1986b) y en formas más largas (Wayne y cols. 1986) no sea así. Estas células de
pared deficiente o esferoplastos parecen tener gran importancia en la patogénesis de la
paratuberculosis (García Marín y cols. 1992a, Hulten y cols. 2000c, Hines y cols. 2003),
y probablemente en la enfermedad de Crohn (Hulten y cols. 2000b, Hulten y cols.
2001). Tal y como puede observarse en extensiones de heces y tejidos, los bacilos
tienden a agruparse en grumos, supuestamente debido a la disposición que adoptan al
replicarse dentro de los macrófagos que infectan (Thoen y cols. 1979).
La mayoría de las cepas son no fotocromógenas y el
crecimiento de algunas se estimula con piruvato. Carece de
fosfatasa alcalina y es resistente a la D-cicloserina y la
ansamicina. Los requerimientos nutricionales para el
cultivo de Map son similares a los del resto de
micobacterias. Aunque generalmente no necesiten factores
de crecimiento, algunos componentes como detergentes,
triglicéridos, ácidos palmítico y oleico, albúmina sérica
bovina, yema de huevo o dióxido de carbono, pueden
potenciarlo. La fuente de carbono más utilizada
tradicionalmente es el glicerol, aunque el piruvato puede
ser el elegido en algunas circunstancias. Pueden también utilizar hidrocarburos,
especialmente alcanos (excepto metano), y los ácidos grasos derivados, así como
alcoholes, cetonas, y ácidos mono-, di- o tricarboxílicos. La asparagina es la fuente de
nitrógeno preferida, sin embargo, ésta puede ser sustituida por glutamina, glutamato,
ácido aspártico, o sales de amonio. Entre los minerales, los macroelementos que
requieren en grandes cantidades son potasio, magnesio, azufre y fósforo. Mientras, los
microelementos y oligoelementos que cabe destacar son hierro, zinc, manganeso, y
molibdeno, muchas veces añadidos al medio como impurezas (Ratledge 1982).
La micobactina
Tal y como se ha comentado anteriormente, existe un ingrediente que ha sido
considerado factor de crecimiento en el aislamiento de Map (Twort y cols. 1913,
Francis y cols. 1949, Snow 1970): la micobactina. Las micobactinas son un grupo de
compuestos estrechamente relacionados entre ellos y asociados a la célula, que
producen las micobacterias en condiciones limitantes de hierro, para poder transportar el
metal al interior de la célula (Snow 1965, Snow y cols. 1969). Está demostrado que Map
puede crecer perfectamente sin aporte exógeno de micobactina si el medio contiene
suficiente hierro y otros nutrientes disponibles (Merkal y cols. 1974, Juste y cols. 1993),
y que además tiene la facultad de sintetizar el sideróforo (Merkal y cols. 1982a, Barclay
Figura 4. Micrografía de Map obtenida con microscopio electrónico de barrido en la Universidad de Wisconsin (MT Collins).

ETIOLOGÍA
54
y cols. 1985). Por lo tanto, la incapacidad de crecer sin micobactina añadida que
muestran algunas micobacterias (Map, ciertas cepas de M. avium, y algunas especies no
patógenas), no residiría en la imposibilidad real de ser sintetizada, sino en la represión
de su producción, o en la presencia de sustancias quelantes en el medio (Merkal y cols.
1974).
Existen cepas de campo de paratuberculosis que son aisladas incluso con mayor
facilidad en medios sintéticos sin micobactina añadida (Aduriz y cols. 1995a, Motiwala
y cols. 2004), y otras de las que se ha comprobado su independencia tras sucesivos
subcultivos (Morrison 1965, Merkal y cols. 1974, Pavlas 1990, Lambrecht y cols.
1992). Mientras algunos autores achacaban la pérdida de tal dependencia en los casos de
pases sucesivos a la estimulación inducida por pH ácido (5,5) o a la formación de
compuestos promotores del crecimiento tras el autoclavado del medio (Morrison 1965),
otros han afirmado que se debe a que el compuesto es transportado en la propia pared
bacteriana desde el medio del cultivo primario a los siguientes (Lambrecht y cols.
1992). Sin embargo, la cepa de referencia de Map ATCC 19698 crece bien en el medio
líquido Middlebrook 7H9 sin micobactina suplementado con oleico-albúmina-dextrosa-
catalasa (OADC) (Aduriz 1993) y en subcultivos posteriores en el mismo medio, lo que
descartaría esta última justificación, al menos en ciertos casos. Al comparar el grupo de
genes que controlan la síntesis de la micobactina en M. tuberculosis (Quadri y cols.
1998), M. avium y la cepa K10 de Map, sólo se ha detectado una diferencia en un gen
(mbtA) supuestamente iniciador del proceso, que podría encontrarse atenuado en Map
(Li y cols. 2005), pero no ha sido probado aún. Al margen de todas estas observaciones,
la adición de micobactina es muy eficaz a la hora de identificar el agente en medios a
base de huevo como son el de Herrold (HEY) o el de Lowënstein-Jensen (LJ),
resultando de gran utilidad a la hora de descartar otro tipo de micobacterias en estos
medios, que por otro lado han demostrado ser muy sensibles para el aislamiento
primario de ciertas cepas (Juste y cols. 1991b).
Aparte de las micobactinas, las exoquelinas son imprescindibles en la captación de
hierro de las micobacterias, y su estudio está posibilitando la comprensión del
metabolismo férrico micobacteriano (Macham y cols. 1975, Barclay y cols. 1988,
Ratledge y cols. 1996, Gobin y cols. 1996, Matzanke y cols. 1997, Ratledge 2004).
Además, Homuth y cols. descubrieron una reductasa extracelular de Map capaz de
movilizar hierro a partir de citrato férrico de amonio, ferritina y transferrina, que podría
suponer una alternativa para la captación de hierro (Homuth y cols. 1998). Incluso se
propuso que esta reductasa podría tener un papel importante en la evasión de los
mecanismos de defensa intracelulares de los macrófagos que infecta Map.
Los medios de cultivo
Los medios de cultivo aptos para Map son los mencionados anteriormente para el
resto de micobacterias, con la salvedad de la adición de micobactina exógena. De todos

REVISIÓN DE LA BIBLIOGRAFÍA
55
modos, los más ampliamente extendidos son el HEY y el LJ enriquecidos con
micobactina, y distintas variantes de los de Middlebrook (7H11, 7H9,...) con OADC y
micobactina. En medios sólidos, las colonias aparecen normalmente pequeñas, firmes,
convexas, húmedas y de color blanco-crema (menos firmes y más blancas cuando
comienzan a aparecer) (Merkal y cols. 1974). Pero algunas cepas pueden presentar
colonias más lisas, suaves, y brillantes, coloreadas de un pigmento amarillo anaranjado
(Benazzi y cols. 1996, Stevenson y cols. 2002). La aparición de colonias puede tardar
entre 3 semanas y un año (Taylor 1951, Merkal y cols. 1974, Wayne y cols. 1986,
Whittington y cols. 2001d), aunque el crecimiento puede ser perceptible en un par de
semanas en medios líquidos (Collins y cols. 1990b).
Resistencia y persistencia
En comparación con otras bacterias, incluidas el resto de micobacterias, Map se
caracteriza por su extremadamente baja actividad metabólica, lo que conlleva una
velocidad de crecimiento en condiciones óptimas 24 veces inferior que la de bacterias
como Escherichia coli (Juste y cols. 1993). Esa cualidad de metabolismo ralentizado en
combinación con la defensa que le confiere la compleja pared celular (Minnikin 1982),
hace de Map una bacteria muy resistente a condiciones adversas, al mismo tiempo que
le permite ser la primera en estar allí cuando estas sean más favorables (Ratledge 1982).
Es capaz de sobrevivir en agua de estanque esterilizada 9 meses y 163 días en agua de
río (Lovell y cols. 1944), en agua corriente con pH no neutro hasta 14 meses (Larsen y
cols. 1956) y en agua destilada hasta 15 meses, reduciéndose la concentración inicial en
1 Log10 cada 68.5 días (Sung y cols. 1998). Un reciente estudio realizado en el Reino
Unido, ha demostrado la presencia de células viables de la bacteria en aguas de un
embalse del río Taff (Pickup y cols. 2005). La supervivencia en heces puede llegar hasta
los 246 días (Lovell y cols. 1944), en purines hasta 287 días (Jorgensen 1977) y entre 21
y 28 días durante la digestión anaeróbica de purines de una planta de biogás (Olsen y
cols. 1985), aunque supuestamente la mezcla de heces y orina produce un efecto
bactericida (Chiodini y cols. 1984a). También permanece 55 semanas en ambientes
secos y sombríos, y 48 semanas en agua de presa no expuesta a luz solar (Whittington y
cols. 2004, Whittington y cols. 2005). Pueden recuperarse células viables después de
largos períodos de congelación (Richards y cols. 1977, Juste y cols. 1990a), y es capaz
de sobrevivir e incluso replicarse en el interior de protistas ambientales (Mura y cols.
2006). Además, su resistencia le permite soportar tratamientos de esterilización
aplicados a la leche y al agua destinada al consumo humano (Whan y cols. 2001, Grant
y cols. 2002a) (tema revisado en el apartado de Epidemiología. Implicaciones en
salud pública).
Por el contrario, parece sensible a la luz solar, las altas temperaturas, la desecación,
la elevada concentración de calcio y el hervido (Merkal y cols. 1982c, Blood y cols.
1983, Schroen y cols. 2003, Whittington y cols. 2004, Whittington y cols. 2005).

ETIOLOGÍA
56
También puede ser inactivado por medio de desinfectantes en suspensión acuosa como
el amonio (3%), el formol (5%), compuestos cresólicos (1:32), el fenol (1:40), el
bicloruro de mercurio (1:1000), el hipoclorito cálcico (1:50), el cloro, cianamida cálcica
o el etanol (Merkal y cols. 1982c, Chiodini y cols. 1984a, Böhn y cols. 1994, Katayama
y cols. 2003). El efecto de estas sustancias es mucho menor en heces secas, aunque
puede incrementarse si se combinan con detergentes (Merkal y cols. 1982c).
Composición genética
La secuenciación del genoma completo de la cepa de origen bovino K10 ha revelado
un cromosoma circular de 4.829.781 pares de bases que codifica para 4.350 ORFs, 45
tRNAs, y un operón rRNA. El análisis in silico ha identificado más de 3.000 genes con
homólogos en M. tuberculosis, y 161 regiones genómicas únicas que codifican 39 genes
de Map desconocidos hasta el momento (Li y cols. 2005) (Figura 5).
Figura 5. Representación linear de las regiones repetidas y de las regiones únicas en el genoma de la cepa K10 de Map. De arriba a abajo: repeticiones VNTR/DR histograma azul; repeticiones SSR histograma rojo; regiones únicas histograma verde; secuencias de inserción histograma multicolor (IS900, azul; MAP01, aguamarina; MAP02, gris; MAP03, rosa; MAP04, verde; MAP05, púrpura; MAP06, negro, MAP07, naranja; MAP08, rojo; MAP09, rosa claro; MAP10, púrpura claro; MAP11, beige; MAP12, marrón; MAP13, fucsia; MAP14, aguamarina claro; MAP15, gris claro; MAP 16, verde oscuro; IS1311, granate; Mav2, azul claro; REP, verde claro); contenido GC histograma púrpura. Figura generada con el programa informático GENESCENE (DNAstar, Madison, WI). (Li y cols. 2005).

REVISIÓN DE LA BIBLIOGRAFÍA
57
Esta cepa de referencia no presentaba plásmidos. Map tiene insertadas en su genoma
entre 14 y 18 copias de IS900 (Green y cols. 1989, Bull y cols. 2000), de 7 a 10 copias
de IS1311 (Whittington y cols. 1999a), al menos tres copias de ISMav2 (Strommenger y
cols. 2001), 3 copias de ISMpa1 (Olsen y cols. 2004), y 6 copias de ISMap02 (Stabel y
cols. 2005). Map no contiene la IS1245 a pesar de que en algunos trabajos se obtuvieran
bandas en el IS1245-RFLP, bandas producidas en realidad por hibridación inespecífica
con la IS1311 (Johansen y cols. 2005). Otros genes o dianas de interés específicos de
Map son el locus 251 (Bannantine y cols. 2002a, Motiwala y cols. 2003), el hspX
(Ellingson y cols. 1998, Ellingson y cols. 2000) y el F57 (Poupart y cols. 1993, Cocito y
cols. 1994, Coetsier y cols. 2000). La IS900 pertenece a una familia de elementos IS
relacionados entre sí que se encuentran en microorganismos del orden Actinomycetales,
y entre las que se incluyen IS901/902, IS110, IS116, IS1110, IS1626 y IS1613 (Bull y
cols. 2003a).
Además, el conocimiento progresivo del genoma de Map incluyendo la secuencia
completa de la cepa K10 está proporcionando el conocimiento de regiones genéticas
únicas de la bacteria, que además de servir para la identificación y el diagnóstico,
permitirán revelar los procesos genéticos de la interacción patógeno-hospedador
(Bannantine y cols. 2002a, Bannantine y cols. 2003b, Paustian y cols. 2004, Bannantine
y cols. 2004a, Bannantine y cols. 2004b, Li y cols. 2005, Paustian y cols. 2005, Marsh y
cols. 2006, Bannantine y cols. 2006b).
EPIDEMIOLOGÍA
Hospedadores
La paratuberculosis afecta principalmente a rumiantes tanto domésticos como
salvajes (Chiodini y cols. 1984a). Adquiere mayor importancia en las especies bovina
(Bos taurus), ovina (Ovis aries) y caprina (Capra hircus) debido a los intereses
adquiridos por el ser humano sobre ellas.
Últimamente, la enfermedad ha pasado a ser junto con la tuberculosis una de las más
preocupantes en cérvidos (familia Cervidae) (Mackintosh y cols. 2004), infectando
distintas especies (Cervus elaphus, Capreolus capreolus, Odocoileus virginianus, Axis
axis, Dama dama, Cervus nippon, Rangifer tarandus, Cervus unicolor, Alces alces) de
orígenes variados (Hillerma 1966, Libke y cols. 1975, Temple y cols. 1979, Riemann y
cols. 1979, Jessup y cols. 1981, Chiodini y cols. 1983, Richards y cols. 1983, Shulaw y
cols. 1986, Weber y cols. 1992b, de Lisle y cols. 1993, Power y cols. 1993, Fawcett y
cols. 1995, Rohonczy y cols. 1996, Stehman 1996, Cook y cols. 1997, Manning y cols.
1998, Moreira y cols. 1999, Nebbia y cols. 2000, Godfroid y cols. 2000, Pavlik y cols.
2000a, Quist y cols. 2002, Marco y cols. 2002, Motiwala y cols. 2004, Machackova y
cols. 2004). Su presencia también ha sido confirmada en otros rumiantes como el

EPIDEMIOLOGÍA
58
bisonte (Bison bison) (Whitlock y cols. 1999, Ellingson y cols. 1999, Buergelt y cols.
2000, Mackintosh y cols. 2002, Leid y cols. 2002, Ellingson y cols. 2005b), el búfalo
(Bubalus bubalis) (Lillini y cols. 2003, Sivakumar y cols. 2005), el borrego cimarrón
(Ovis canadensis), la cabra de las montañas rocosas (Oreamnos americanus) (Williams
y cols. 1979, Richards y cols. 1983), el muflón (Ovis musimon) (Boever y cols. 1974,
Richards y cols. 1983, Weber y cols. 1992a, Pavlik y cols. 2000a, Machackova y cols.
2004, Deutz y cols. 2005), el íbice (Capra ibex) (Richards y cols. 1983, Weber y cols.
1992a, Giacometti y cols. 1995, Ferroglio y cols. 2000), la cabra salvaje (Capra
aegagrus) (Machackova y cols. 2004), el borrego audad (Ammotragus lervia) (Richards
y cols. 1983), el camello (Camelus bactrianus) (Richards y cols. 1983, Paling y cols.
1988, Stehman 1996), la llama (Lama glama) (Blood y cols. 1983, Richards y cols.
1983, Belknap y cols. 1994, Miller y cols. 2000), la alpaca (Lama pacos) (Ridge y cols.
1995a, Miller y cols. 2000, Cousins y cols. 2000, Lucas y cols. 2003), distintas especies
de antílopes (Saiga tatarica, Addax nasomaculatus, Damaliscus albifrons, Damaliscus
lunatus, Kobus ellipsiprymnus, Oryx gazella, Antidorcas marsupialis, Aepyceros
melampus, Tragelaphus angasi, Tragelaphus spekei, Gazella thomsoni, Connochaetes
gnou) (Richards y cols. 1983, Steinberg 1988, Dukes y cols. 1992, Burton y cols. 2001,
Motiwala y cols. 2004), el urial (Ovis vignei), el duíquero bayo (Cephalophus dorsalis),
el muntjac de Reeve (Muntiacus reevesi), y el markhor (Capra falconeri) (Motiwala y
cols. 2004).
Map también ha sido aislado de otros animales no rumiantes, siendo el conejo uno
de los más importantes (Oryctolagus cuniculus) (Fuentes y cols. 1988, Angus 1990,
Fernández de Luco y cols. 1995, Greig y cols. 1997, Greig y cols. 1999, Beard y cols.
2001b, Daniels y cols. 2003d, Raizman y cols. 2005, Judge y cols. 2006). Debido a las
altas prevalencias detectadas en este animal (Greig y cols. 1997), se ha sugerido que
esta especie puede jugar un importante papel en la epidemiología de la enfermedad en el
Reino Unido (Greig y cols. 1999, Daniels y cols. 2001, Daniels y cols. 2003a, Daniels y
cols. 2003b, Daniels y cols. 2003c, Daniels y cols. 2003d, Judge y cols. 2005, Judge y
cols. 2006). También han sido hallados casos naturales de infección en cerdo y jabalí
(Sus domesticus y Sus scrofa) (Ringdal 1963, Richards y cols. 1983, Thoen y cols.
1996b, Machackova y cols. 2003b, Machackova y cols. 2004, Alvarez y cols. 2005,
Aurtenetxe y cols. 2005), en burro (Equus asinus) (Dierckins y cols. 1990), en perro
(Canis familiares) (Vogel 1977), en gatos asilvestrados (Palmer y cols. 2005), en
primates (Pitcock y cols. 1961, McClure y cols. 1987), en liebre (Lepus europaeus), en
zorro (Vulpes vulpes), en mustélidos (Mustela erminea, Mustela minuta, Meles meles),
en ratón de campo y rata (Apodemus sylvaticus, Rattus norvegicus), y en córvidos
(Corvus corone, Corvus frugilegus, Corvus monedula) (Beard y cols. 1999, Beard y
cols. 2001a).
A parte de los experimentos llevados a cabo con las tres especies principales de
rumiantes domésticos (Stuart 1965, Angus y cols. 1971, Allen y cols. 1974a, Thoen y

REVISIÓN DE LA BIBLIOGRAFÍA
59
cols. 1983, Williams y cols. 1983, García Marín y cols. 1988a, García Marín y cols.
1988b, Juste 1990, Saxegaard 1990, Juste y cols. 1994, Begara-McGorum y cols. 1998,
Sigurdardottir y cols. 1999, Waters y cols. 2003, Reddacliff y cols. 2003b, Chacon y
cols. 2004, Emery y cols. 2004), la infección ha sido inducida artificialmente en
experimentos con rumiantes salvajes (Williams y cols. 1983) y no rumiantes como
animales de laboratorio, cerdos, caballos, gallinas, y otras especies (Francis 1943,
Jorgensen 1969, Larsen y cols. 1971, Larsen y cols. 1972a, Larsen y cols. 1972b, Larsen
y cols. 1978a, Mokresh y cols. 1990, Van Kruiningen y cols. 1991, Hamilton y cols.
1991, Chiodini y cols. 1993, Mutwiri y cols. 2001, Mutwiri y cols. 2002, Stabel y cols.
2002a, Chacon y cols. 2004, Vaughan y cols. 2005).
Por otro lado, Map ha sido señalado como agente etiológico de la enfermedad de
Crohn humana, materia que se revisa en el apartado de Epidemiología. Implicaciones
en salud pública.
Transmisión
A pesar de que el mecanismo más importante de transmisión de la paratuberculosis
es el fecal-oral, es decir, ingiriendo heces procedentes de animales infectados
contaminadas con la bacteria (Sweeney 1996), también existen otras vías. Los animales
portadores pueden contaminar con sus heces los pastos, la comida y el agua, lo que
provoca un alto riesgo de exposición para el resto de animales, ya sea en rumiantes
domésticos o salvajes (Boever y cols. 1974, Riemann y cols. 1979, Jessup y cols. 1981,
Chiodini y cols. 1984a, Steinberg 1988, Stehman 1996, Manning y cols. 1998, Daniels y
cols. 2003c). Los animales recién nacidos, que son los más susceptibles, pueden estar
expuestos a la ingesta de grandes cantidades de la micobacteria vía calostro, leche, o al
mamar de las ubres sucias de las madres (Riemann y cols. 1983, Chiodini y cols. 1984a,
Thoen y cols. 1988, Sweeney 1996). El riesgo de infección es muy grande en
explotaciones intensivas donde existe un contacto frecuente entre recién nacidos y
animales eliminadores. Según Whittington y cols., en la especie ovina, la cantidad de
bacterias eliminadas en heces parece estar directamente relacionada con el tipo de
lesiones. De este modo, ovejas con lesiones multibacilares excretarían los bacilos de
manera constante, mientras que con otro tipo de lesiones la excreción sería intermitente
(Whittington y cols. 2000c). Así, si cada oveja elimina al día más de 108 unidades
formadoras de colonia (CFU) por gramo de heces de media (según el cultivo fecal), el
medio de estos rebaños está permanentemente contaminado con la bacteria.
La transmisión vertical prenatal ha sido suficientemente documentada y demostrada.
Map ha sido aislada de tejidos fetales (membranas, cotiledones,…) y de tejidos
maternos (endometrio, mucosa uterina,…), aunque esta posibilidad parece ceñirse en su
gran mayoría a casos de madres clínicamente enfermas (Alexejeff-Goloff 1929, Hole
1953, Pearson y cols. 1955, Lawrence 1956, Doyle 1958, Seitz y cols. 1989, Rohde y

EPIDEMIOLOGÍA
60
cols. 1990, Lambeth y cols. 2004). Sin embargo, algunos autores la han observado en
casos de madres subclínicas (Lawrence 1956, Kopecky y cols. 1967), mientras que otros
correlacionan la presencia de la bacteria en fetos con madres aparentemente sanas, pero
que excretan grandes cantidades de Map en las heces (Sweeney y cols. 1992b). En esta
misma línea, un reciente estudio retrospectivo de Aly y cols. (Aly y cols. 2005)
demuestra que la probabilidad de que una vaca sea seropositiva es 6,6 veces mayor para
hijas de vacas seropositivas que para hijas de vacas seronegativas, ya sea por
transmisión congénita o vía calostro/leche posparto. Así, se podría pensar que los
ovocitos y embriones pueden ser portadores del patógeno a la hora de ser implantados
en madres receptoras, no obstante, los resultados de un trabajo sobre esta materia,
sugieren que los ovocitos y fetos derivados de madres eliminadoras moderadas no
suponen una vía de transmisión, a pesar de la pequeña muestra utilizada en ese estudio
(Kruip y cols. 2003). Por el contrario, los recientes hallazgos de van Kooten y cols. (van
Kooten y cols. 2006), indican un alto riesgo de transmisión intrauterina en ciervos, y
sugieren que ésta puede jugar un importante papel en la amplia distribución de la
paratuberculosis. La placenta puede también suponer un vehículo de contagio en época
de partos (Pearson y cols. 1955, Lawrence 1956, Doyle 1958) especialmente en
explotaciones intensivas, por lo que se debe evitar el contacto con los recién nacidos.
Map puede aislarse de leche de vacas clínicamente afectadas con relativa frecuencia
(Alexejeff-Goloff 1929, Smith 1960, Taylor y cols. 1981, Giese y cols. 2000), pero
también de vacas asintomáticas o subclínicas con cultivo fecal positivo (Sweeney y
cols. 1992a, Streeter y cols. 1995). En definitiva, la leche puede servir como vía de
transmisión de la paratuberculosis, especialmente si se trata de animales que eliminen
grandes cantidades del bacilo en las heces (Sweeney 1996). Una vaca seropositiva
puede eliminar una media de 42,33 ± 19,67 CFUs por 100 ml de leche (Herman y cols.
2006). El semen de toros afectados por la enfermedad puede contener la bacteria
(Larsen y cols. 1970, Larsen y cols. 1981, Eppleston y cols. 2001, Glawischnig y cols.
2004), provocando infección intrauterina en hembras receptoras, que al mismo tiempo
puede ser trasladada al cigoto, y según Merkal y cols. (Merkal y cols. 1982b), genera
hipersensibilidad en vacas receptoras. Como en el caso de Mycobacterium bovis
(Serraino y cols. 1999), otra hipotética ruta de contagio sería por las vías respiratorias
(Corner y cols. 2004) o por ingestión de presas o cadáveres contaminados en animales
carnívoros o carroñeros como el zorro y el jabalí (Beard y cols. 2001a, Machackova y
cols. 2004).
En cuanto al conejo se refiere, la transmisión vertical, pseudovertical (por la
ingestión de leche y/o heces maternas) y horizontal (tanto intra-especies como inter-
especies) son posibles, como demuestran los resultados de un estudio que afirma que la
existencia de tales vías de transmisión contribuyen al mantenimiento de la enfermedad
en las poblaciones de estos lagomorfos, y como resultado, en el medio (Judge y cols.
2006). Además de todas las fuentes mencionadas, Map también puede ser aislada de

REVISIÓN DE LA BIBLIOGRAFÍA
61
insectos (Fischer y cols. 2001, Fischer y cols. 2003b, Fischer y cols. 2004a, Fischer y
cols. 2004b), lombrices (Fischer y cols. 2003a), nemátodos (Lloyd y cols. 2001,
Whittington y cols. 2001a) y amebas de vida libre (Mura y cols. 2006), por lo que estos
invertebrados también podrían tener un papel en la transmisión de la paratuberculosis, y
consecuentemente, en su epidemiología.
Distribución y prevalencia
La paratuberculosis es una enfermedad de creciente prevalencia en especies
ganaderas que está ampliamente distribuida por todo el mundo (Chiodini y cols. 1984a,
Kennedy y cols. 2001). Sin embargo, la prevalencia real de esta enfermedad puede estar
enmascarada por las dificultades que plantea su diagnóstico. Las técnicas utilizadas en
la detección de animales infectados en general no son especialmente sensibles, a lo que
hay que añadir el largo período de latencia desde la infección hasta la aparición de
síntomas, y la variabilidad del curso de la enfermedad entre animales. Entre las técnicas
serológicas utilizadas para estimar la prevalencia destaca la del inmunoanálisis ligado a
enzimas o ELISA, aunque la inmunodifusión en gel de agarosa (AGID) también haya
sido popular con pequeños rumiantes (Juste y cols. 1991a, Aduriz 1993, Mainar-Jaime y
cols. 1998, Reviriego y cols. 2000, Michel y cols. 2000, Sergeant y cols. 2002b). Las
técnicas microbiológicas más importantes para ese mismo fin son el cultivo y la PCR de
muestras fecales o de tejidos (en caso de estudios de matadero). Las técnicas
histopatológicas también proporcionan resultados satisfactorios (Juste y cols. 1991a,
Aduriz 1993, García Marín y cols. 1994, Pérez y cols. 1997, González 1998, Pérez y
cols. 1999, Corpa y cols. 2000a, Garrido 2001, Balseiro y cols. 2003, González y cols.
2005). En la Tabla 2 se indica el porcentaje de animales y/o rebaños positivos
detectados por medio de diferentes técnicas en varias especies hospedadoras
procedentes de numerosos países de todo el mundo.
A nivel estatal, aunque fue por primera vez mencionada en 1915 (Farreras 1915) y
posteriormente en 1935 (Roca Soler 1935), la paratuberculosis no fue adecuadamente
descrita hasta 1973 en ovino de León (Aller y cols. 1973), 1983 en bovino del País
Vasco (Juste y cols. 1983) y 1984 en caprino de Andalucía (León y cols. 1984) (aunque
en esta especie fueron comunicados resultados positivos de serología en 1979 (Garrido
y cols. 1979)). Se considera ampliamente distribuida por todo el estado (Aduriz y cols.
1991, Corpa y cols. 1997), tal y como indica la descripción de la enfermedad en varias
comunidades y especies hospedadoras (Badiola y cols. 1980, Badiola y cols. 1986,
Villanueva 1989, Díaz 1989, Carrera y cols. 1989, Santamaría 1989, Garrido y cols.
1989, Guijarro 1990, Juste y cols. 1990c, Molina y cols. 1991, Aduriz y cols. 1991,
Pérez 1992, Aduriz 1993, Mainar-Jaime y cols. 1998, Reviriego y cols. 2000, Rodríguez
2000, Garrido 2001, Marco y cols. 2002, Balseiro y cols. 2003, González 2003, Balseiro
2004, Alvarez y cols. 2005, de Juan y cols. 2006a). Varios de estos estudios aportan
además información sobre la prevalencia de la enfermedad.

Tab
la 2
. Por
cent
aje
de a
nim
ales
y/o
reb
años
pos
itivo
s se
gún
las
técn
icas
, esp
ecie
s ho
sped
ador
as y
reg
ione
s m
enci
onad
as.
R
egió
n
Añ
o(s
) T
écn
ica(
s)
Esp
ecie
A
nim
ales
(%
) R
eba
ño
s (%
) B
ibli
og
rafí
a A
lem
ani
a <
200
4 E
LI
Bov
12
,2
84.7
(H
acke
r y
cols
. 2
004)
A
rge
ntin
a 19
93-2
001
E
LI+
TT
C
Bov
0,
07-
1,7
5
(Hol
zma
nn y
col
s. 2
004
) A
ustr
alia
N
uev
a G
ales
del
Sur
Z
ona
Sur
T
ota
lida
d
<1
996;
has
ta 2
000
19
90-9
3 20
01;
200
4
HP
; HP
mat
ad
ero
CF
/ ELI
/ T
FC
to
das
Ovi
B
ov
Ovi
; Bov
(le
che/
car
ne)
9-2
3; -
0,
43/
-/ -
-; 0
,04
-39
1,3
9/ 0
,64
/ 7,4
7 0-
43;
11
,9/
0,08
(Ma
rsha
ll y
cols
. 19
97);
(S
erg
eant
y c
ols.
20
02a)
(V
ande
graa
ff y
col
s. 1
995)
(S
erg
ean
t 2
003)
; (K
enn
edy
y co
ls.
2006
) A
ustr
ia
1995
-97
; 200
2-03
E
LI
Bov
2;
19
7; -
(G
aste
ine
r y
cols
. 1
999)
; (K
hol
y c
ols.
20
06)
Bél
gica
19
97-9
8 E
LI
Bov
1,
18
0,8
7 (B
oela
ert y
col
s. 2
000
) B
rasi
l R
ío d
e Ja
neiro
R
ío G
rand
e do
Su
l
<2
002
<2
005
ELI
E
LI
Bov
B
ov
18
44,6
82
97,2
(Fer
reir
a y
cols
. 200
3)
(Gom
es y
col
s. 2
006)
C
anad
á A
lber
ta
Not
ario
Q
uéb
ec
Sas
katc
hew
an
Zo
nas
atlá
ntic
as
<2
003
1986
-89
; 200
3 19
99-2
000
20
01;
199
9 19
98-9
9; 2
001
ELI
/ C
FL
CF
/ CF
/ ELI
; E
LI/E
LIL
H
P
ELI
E
LI;
HP
+C
T
Bov
B
ov
Ovi
B
ov (
lech
e);
Bov
(ca
rne)
B
ov
7/ 1
,1-3
,4
5,5
/ 0,1
/ 1
5,2;
2,6
/ 1,7
3 2,
7; 0
,8
2,6
; 15,
1
40/
20,
4 -;
58/
34
43,3
; 15
,2
43,3
; -
(Sor
ens
en
y co
ls. 2
003)
(M
cNa
b y
cols
. 1
991)
; (H
end
rick
y c
ols.
20
05a)
(A
rsen
ault
y co
ls. 2
003)
(V
anLe
euw
en
y co
ls.
200
5);
(Wal
dn
er y
col
s. 2
002)
(V
anLe
euw
en
y co
ls.
200
1);
(McK
enna
y c
ols.
20
04)
Cor
ea
1999
E
LI
Bov
3,
2-5,
3 84
,7-9
0,6
(Pak
y c
ols.
200
3)
Cos
ta R
ica
1990
E
LI
Bov
11
,9
18,7
(D
olz
y co
ls. 1
999)
C
roac
ia
1987
-96
ELI
+T
FC
B
ov
1,3
6
(Cve
tnic
200
0)
Din
amar
ca
1998
E
LIL;
ELI
B
ov
-; 8
,8
70;
86
(Nie
lsen
y c
ols.
20
00b)
; (J
ako
bsen
y c
ols.
20
00)
Est
ados
Un
idos
A
lab
am
a C
alif
orn
ia
Co
lora
do
Flo
rida
G
eor
gia
L
ouis
iana
M
aine
M
ichi
gan
Mis
sour
i N
uev
a Y
ork
Tex
as
Wis
cons
in
Sur
este
S
in e
spec
ifica
r
1998
-99
1988
-89
; 200
0 19
99
1986
-87
2000
19
88
2001
19
96
1993
-94
1989
-93
2000
-01
1989
-90
1998
-20
00
1996
;97
ELI
E
LI
ELI
E
LI
ELI
E
LI
HP
+C
T
ELI
E
LI
CF
E
LI
ELI
C
T/ E
LI/
AG
ID
ELI
Bov
(ca
rne)
B
ov (
lech
e/c
arne
); B
ov
Bov
B
ov (
lech
e/ c
arne
) B
ov (
lech
e/ c
arne
) B
ov
Bov
B
ov
Bov
(le
che
/ car
ne)
Bov
B
ov (
carn
e)
Bov
C
ierv
o d
e V
irgin
ia
Bov
(le
che
); B
ov (
carn
e)
8 7/ 6
; 4,
6 0-
7,8
2 17
,1/
8,6
9,5
8/ 3
,95
4,4
21,7
6,
87
8/ 5
0,
7-2
8,2
3 7,2
9 0,
3/ 2
/ 0
3,4
; 0,4
63,2
9 60
/ 6
0; 7
7,3
3 73
,33
30
55
74/
40
57,5
7 43
,8
50
21,6
; 7,
9
(Hill
y c
ols.
20
03)
(B
ehym
er y
col
s. 1
991)
; (A
das
ka y
col
s. 2
003)
(H
irst y
col
s. 2
004)
(B
rau
n y
cols
. 19
90)
(Pen
ce y
col
s. 2
003)
(O
lcot
t y c
ols.
19
92)
(McK
enn
a y
cols
. 20
04)
(Joh
nson
-Ife
aru
lund
u y
cols
. 19
99a)
(T
hor
ne
y co
ls. 2
000)
(O
basa
njo
y co
ls. 1
997)
(R
ouss
el y
col
s. 2
005)
(C
ollin
s y
cols
. 19
94)
(Dav
idso
n y
cols
. 20
04)
(We
lls y
col
s. 2
000)
; (D
arga
tz y
col
s. 2
001
b)
Esc
ocia
19
96
CF
óT/ H
P/
CF
óT+
HP
C
onej
o 7,
14/
16
,19/
19
,52
18,1
8/ 5
0/ 5
0 (G
reig
y c
ols.
199
9)
Esl
ova
quia
19
82-8
7 C
FóT
B
ov (
con
sosp
echa
) 41
,2
(H
anzl
ikov
a y
cols
. 19
89)
Esl
ove
nia
1997
/ 9
8;20
00-0
1 E
LI
Bov
2,
84/
6,4
8; 0
,44
0,4
1/ 1
,16
; 11,
59
(Oce
pek
y co
ls. 1
998
); (
Oce
pek
y c
ols.
200
3)
Fin
land
ia
2001
C
F ó
EL
I ó H
P
Bov
(le
che
/ car
ne)
0/ ≈
0
(Seu
na y
col
s. 2
003)

F
ranc
ia
Yo
nne
1998
-99
ELI
B
ov
3,3
68
(Pet
it 20
01)
Hol
anda
19
95;9
8;
<2
000
CF
; ELI
L;
- B
ov
-; 2
,5;
5-1
3 51
;55
; -
(Kal
is y
col
s. 1
999
a);
(Mus
kens
y c
ols
. 20
00);
(N
iels
en y
col
s. 2
000
a)
Ing
late
rra
Sur
oest
e T
ota
lida
d
1994
19
97
CT
/ PC
RT
en
cues
ta
Bov
B
ov
2,6
/ 3,5
17,4
(Çet
inka
ya y
col
s. 1
996
) (Ç
etin
kaya
y c
ols.
19
98)
Ind
ia
Utt
ar P
rad
esh
<2
005
ELI
B
úfal
o/ B
ov/ C
ap
25,5
-40,
3/ 2
5-4
2,6
/ 15-
50
(S
ingh
y c
ols.
20
06)
Irla
nda
1997
C
F/ E
LI
Bov
(im
port
ada)
4,
07/
3,5
5 37
,5/
25
(O'D
ohe
rty
y co
ls.
2002
a)
Isra
el
2003
C
F y
EL
I B
ov
61
(K
ore
n y
cols
. 20
06)
Italia
T
rent
ino
Ve
neto
C
ent
ro
Nor
te
1998
-20
02
2000
-01
<2
005
1995
-20
02
CT
E
LI
ELI
C
F/ E
LIS
A
Cie
rvo
Bov
B
ov
Bov
48,2
3,
5 1,
8
65
42
1,7-
10,
4/ 7
,5-6
0,3
(Fra
quel
i y c
ols.
200
6)
(Rob
bi y
col
s. 2
003
) (L
illin
i y c
ols.
20
06)
revi
sad
o en
(C
avir
ani
y c
ols
. 20
06)
Mar
ruec
os
<1
996
CF
/ TF
C
Ovi
29
,8/
29,2
(Ben
azzi
y c
ols.
199
6)
Nor
ueg
a 19
92-9
9 E
LI
Alc
e/ C
ierv
o/ C
orzo
/ R
eno
1,9
/ 3,8
/ 1
2,2/
3,4
(Try
land
y c
ols.
20
04)
Nue
va Z
ela
nda
19
86-2
000
; 20
01-0
4; 1
995
CT
ó P
CR
; C
T ó
PC
R;-
C
ierv
o;
Cie
rvo
; Ovi
6;
12;
6,4
(de
Lisl
e y
cols
. 20
03);
(d
e Li
sle
y co
ls. 2
006
); (
Wes
t 19
97)
Por
tug
al
Nor
oest
e L
isbo
a S
erra
da
Est
rela
2003
-04
<2
005
2004
ELI
E
LI+
AG
ID
ELI
Ovi
O
vi
Ovi
0-1
0,3
5,6
46,7
27
(Coe
lho
y co
ls. 2
006)
(M
end
es y
col
s. 2
006)
(S
eixa
s y
cols
. 20
06)
Rep
úblic
a C
hec
a 19
92-9
8; 1
999-
2001
T
FC
; CF
óT
Bov
(im
port
ada)
; ci
ervo
/ co
rzo/
ga
mo
/ m
ufló
n/
bezo
ar
-; 1
5/ 0
,2/
1/ 3
,8/ 0
12
,2; -
(P
avlik
y c
ols
. 20
00b)
; (M
acha
ckov
a y
cols
. 2
004)
S
udáf
rica
19
96
AG
ID
Ovi
2,7
2 (M
ich
el y
col
s. 2
00
0)
Sue
cia
1993
-98
CF
/ ELI
B
ov
1,3
/ 2
(B
olsk
e y
cols
. 19
99)
Sui
za
1994
; <
95
2001
; 0
2
ELI
; E
LI
PC
RL;
PC
RL
Bov
; Bov
B
ov; C
ap/
Ovi
0,7
; 6
8; -
1,
7-4
9,2
; 23/
23
,8
(Sta
rk y
co
ls. 1
997)
; (M
eyla
n y
cols
. 19
95)
(Cor
ti y
cols
. 2
002)
; (M
uehl
herr
y c
ols.
20
03)
Uru
gua
y <
200
2 E
LI
Bov
16
72
(P
iagg
io y
col
s. 2
003
)
Abr
evia
cion
es: B
ov=b
ovin
o, C
ap=c
apri
no, C
F=cu
ltivo
fec
al, C
FóT
=cul
tivo
feca
l o d
e te
jidos
, CF
L=
culti
vo f
ecal
por
lote
s, C
T=
culti
vo d
e te
jidos
, EL
I=E
LIS
A d
e sa
ngre
, E
LIL
=E
LIS
A d
e le
che,
HP=
anál
isis
his
topa
toló
gico
, A
GID
=in
mun
odif
usió
n en
gel
de
agar
osa,
Ovi
=ovi
no,
PC
RL
=PC
R d
e le
che,
PC
RT
=PC
R d
e te
jidos
, T
FC=t
est d
e fi
jaci
ón d
el c
ompl
emen
to, T
TC
=tes
t de
la tu
berc
ulin
a co
mpa
rada
, <=a
ntes
de.

EPIDEMIOLOGÍA
64
En un estudio realizado sobre 201 vacas de desvieje procedentes del matadero de
Bilbao, se detectaron porcentajes de positivos individuales del 0,5, 0,5, 4,7, 15,4, y
19,4% por medio del cultivo de tejidos y heces, AGID, histopatología, PCR y ELISA,
respectivamente, lo que situó la prevalencia en un 31,3% al combinar los resultados de
las distintas técnicas (Garrido 2001). Otro estudio similar, realizado sobre 134 vacas
sacrificadas en mataderos asturianos, arroja una prevalencia individual del 28,35%
mediante histopatología, inmunohistoquímica y PCR (Balseiro y cols. 2003).
Posteriormente, se ha ampliado este estudio sobre bovino procedente de Asturias,
detectando una prevalencia (ELISA) media del 1,23% entre los años 1995 y 2001, que
asciende hasta un 44,39% por medio de técnicas histopatológicas (Balseiro 2004). En el
mismo, también se señala una prevalencia intra-rebaño de 0-22% detectada por medio
de ELISA. Según un estudio preliminar sobre la prevalencia de Map en explotaciones
bovinas lecheras en España por medio de PCR de muestras de leche de tanque, hasta un
18% (10 ± 8%) de las mismas podía estar infectado (Sevilla y cols. 2003).
En Aragón se observó que el 46,7% de los más de 150 rebaños ovinos estudiados
estaban infectados, tal y como indicó la combinación de técnicas bacteriológicas,
anatomopatológicas y serológicas empleadas (Juste y cols. 1991a). Los resultados de
ELISA de rebaños caprinos procedentes de Huelva, Córdoba e Islas Canarias, indicaron
porcentajes de animales positivos del 41, 13 y 0,5% respectivamente (Molina y cols.
1991). El estudio anatomopatológico del desecho de rebaños ovinos afectados en León,
señaló un 46% de animales con lesiones (Pérez 1992). En el País Vasco, tras el análisis
de muestras de 4.504 ovejas procedentes de 226 rebaños, según los resultados de AGID
y ELISA, el 2% de animales fueron positivos con la primera técnica y el 5,8% con la
segunda (Aduriz 1993), mientras que la seroprevalencia de rebaños fue del 23,5 y
31,4% respectivamente. En pequeños rumiantes de Zaragoza se observó que el 10,8%
de ovejas y el 22,5% de cabras era positivo a alguno de los métodos diagnósticos
empleados (cultivo, AGID y anatomía patológica) (García Marín y cols. 1994). Otro
estudio lesional realizado en León sobre ovejas de desecho detectó un 24,3% de estos
animales con lesiones compatibles con esta enfermedad (González 1998). La utilización
de AGID reveló un 44% de pequeños rumiantes infectados en Madrid (Mainar-Jaime y
cols. 1998), y un 56% de rebaños afectados en Ávila (Reviriego y cols. 2000). Más
tarde, a pesar de la posible interferencia por vacunaciones frente a paratuberculosis no
declaradas, se han observado prevalencias del 97% de rebaños con una incidencia intra-
rebaño del 49,7% en la especie caprina en Córdoba (Rodríguez 2000). Las altas
seroprevalencias observadas en caprino de Salamanca (44,12% de animales y 86,66%
de explotaciones) reflejan la importante incidencia de la paratuberculosis en esta especie
en España (Fernández y cols. 2001).

REVISIÓN DE LA BIBLIOGRAFÍA
65
Trascendencia Económica
A la hora de calcular el impacto económico de la paratuberculosis, las pérdidas
pueden ser clasificadas como directas, indirectas o inaparentes (Merkal 1983,
Hutchinson 1996, Kennedy y cols. 2001). Dentro de los costos directos, entrarían las
pérdidas derivadas de la reducción productiva de animales clínicos y subclínicos, bien
por descenso en las cantidades de leche producidas, por descenso del valor del animal
en el matadero, o por el sacrificio prematuro de los mismos. Los costos derivados del
control de la enfermedad, como la contratación de servicios veterinarios, la realización
de pruebas diagnósticas o los generados por cambios de manejo del rebaño, también
pueden clasificarse como directos. Se pueden considerar costos indirectos, aquellos
derivados de restricciones de acceso al mercado (muy importantes en algunas regiones
de Australia), los gastos de testado de animales para venta o exportación, así como los
originados por la adopción de medidas preventivas de manejo en los grupos de cría.
Entre los costos inaparentes, se encuentran la pérdida de potencial genético debido al
sacrificio prematuro (Buergelt y cols. 1978a) y las restricciones en la compraventa de
animales de alto valor genético. Debido tanto al desconocimiento del alcance real de la
enfermedad, como a las diferencias de producción y de valoración de precios existentes
entre países, no existen datos absolutamente fiables de las pérdidas que provoca la
paratuberculosis. La reducción en la producción de leche en vacas clínicamente
afectadas, varía de un 14,6% a un 15,9%, alcanzando hasta un 19,5% en la última
lactación (Buergelt y cols. 1978a, Benedictus y cols. 1987, Spangler y cols. 1992a). En
la penúltima lactación de animales con paratuberculosis subclínica, se ha observado una
reducción en la producción de leche del 4-6% (Nordlund y cols. 1996), ascendiendo la
pérdida hasta un 14% en la última lactación antes del sacrificio (Benedictus y cols.
1985b, Wilson y cols. 1997). Sin embargo, según Johnson y cols., en rebaños con una
media de dos partos o menos, las pérdidas de producción lechera apenas tienen
importancia (Johnson y cols. 2001). En cambio, un reciente estudio llevado a cabo sobre
9 explotaciones lecheras, concluye que la paratuberculosis es responsable de una
importante reducción en la producción de leche y en la longevidad de las vacas
(Hendrick y cols. 2005c). Las pérdidas asociadas a animales subclínicos pueden estar
relacionadas con el incremento en la incidencia de mamitis e infertilidad, y con las tasas
de eliminación de animales del rebaño (Merkal y cols. 1975, Wilson y cols. 1997). Se
ha estimado que las pérdidas económicas derivadas de la paratuberculosis en las granjas
lecheras de Estados Unidos puede exceder los 1,5 billones de dólares por año (Jones
1989). Un reciente estudio con datos de 1996 sobre el impacto económico producido
por la reducción en la producción de leche debida a la paratuberculosis, determina que
ésta fue de 580 ± 460 millones de kg, lo que supuso una pérdida total de 200 ± 160
millones de dólares para la economía de Estados Unidos (Losinger 2005). Los rebaños
positivos a paratuberculosis (≥1 animal positivo a ELISA) experimentan una pérdida de
casi 100 dólares por vaca en comparación con los libres de paratuberculosis, y en los

EPIDEMIOLOGÍA
66
rebaños con al menos un 10% declarado de bajas por esta enfermedad, las pérdidas
ascienden hasta 200 dólares y 700 Kg de leche por vaca y año (Ott y cols. 1999). La
presencia de paratuberculosis en las explotaciones lecheras del estado de Michigan,
supondrían unas pérdidas directas anuales de 1.600-4.400 dólares (Johnson-Ifearulundu
y cols. 1999a, Johnson-Ifearulundu y cols. 1999b). En Canadá, el promedio de los
costos anuales directos para un rebaño lechero tipo de 50 vacas asciende a 2.472 dólares
según lo calculado por Chi y colaboradores (Chi y cols. 2002). La depreciación en la
venta de los animales en el matadero puede variar de un 5 a un 100% en función del
estado de la infección (de temprana a clínica) (Weber y cols. 2006).
En pequeños rumiantes no se cuenta con tantos estudios sobre el impacto económico
de esta enfermedad. En los años 80, en Inglaterra se realizó una estimación de las
pérdidas basándose en el seguimiento de un rebaño ovino con una tasa de bajas anuales
del 9%, lo que reveló unas pérdidas por sacrificio prematuro y reducción del potencial
genético de 6.000 libras en dos años (Cranwell 1988). Denholm y colaboradores
pronosticaron unas pérdidas de 92 millones de dólares australianos anuales para el año
2010 en Nueva Gales del Sur (Australia) (Denholm y cols. 1997). Los problemas y las
pérdidas derivadas de la enfermedad de Johne ovina en este país, pasaron a tener gran
relevancia en la industria, y promovieron un programa nacional de control de la
enfermedad de 6 años de duración que comenzó en 1999, y con una inversión de 40
millones de dólares australianos (Sergeant 2001). En Nueva Zelanda, el promedio anual
de pérdidas se estimó en 20-22 dólares por oveja (Brett 1998). Estudios llevados a cabo
en el País Vasco demostraron un descenso del 10% en la producción de leche de oveja
Latxa seropositiva, cuya venta habría producido un beneficio de 25 millones de pesetas
en 1991 (Aduriz y cols. 1995b). Asimismo, se calculó una pérdida de 100.000 pesetas
anuales en rebaños destinados a producción cárnica. Se ha estimado que las pérdidas
ocasionadas por casos clínicos en ovino de carne y de leche ascienden a 75 y 125
dólares respectivamente (Juste 1997b).
En explotaciones intensivas de ciervos, la paratuberculosis puede llevar a
importantes pérdidas económicas por la muerte de animales (Temple y cols. 1979,
Fawcett y cols. 1995).
Implicaciones en salud pública
Enfermedad de Crohn
La enfermedad de Crohn es un proceso inflamatorio del intestino que afecta a la
especie humana (Dalziel 1913, Crohn y cols. 1932), que ha sido relacionada con la
paratuberculosis debido a las semejanzas entre ambas (Patterson y cols. 1972, Chiodini
1983, Chiodini y cols. 1984c, Chiodini 1997). Se agrupa dentro de las denominadas
enfermedades inflamatorias digestivas (IBD), junto con la colitis ulcerosa y otras.

REVISIÓN DE LA BIBLIOGRAFÍA
67
Los síntomas más típicos de la enfermedad son la diarrea, el dolor abdominal y la
pérdida de peso. La lesión característica es una inflamación granulomatosa en el tramo
final del íleon y zona ileocecal, caracterizada histológicamente por agregados de células
epiteliales, linfocitos, y células plasmáticas, que afectan a la mucosa y submucosa
intestinal (Anderson 1980). Los jóvenes de entre 16 y 25 años son los típicamente
afectados, pero puede aparecer antes o después en la vida (Podolsky 2002). Su
impredecible curso clínico se caracteriza por una alta incidencia de recurrencia
(Yamamoto 2005). Cerca del 80% de los pacientes que la padecen requieren cirugía
durante su vida (Olaison y cols. 1990). Pero los pacientes de Crohn son un grupo
heterogéneo, y la etiología puede no ser la misma para todos los individuos (Grant
2005).
Con datos obtenidos en Olmsted (Minnesota, EEUU) entre 1940 y 1993, se calculó
que cada año había 5,8 casos más por cada 100.000 habitantes, mientras que la
prevalencia en 1993 ascendía a 133/100.000, un 46% más elevada que en 1980 (Loftus,
Jr. y cols. 1998). El número de nuevos casos anuales en Croacia pasó de 0,34/100.000
en 1973 a 3,47/100.000 en 1994 (Jovanovic 1999). La incidencia anual de casos
pediátricos de Crohn en Suecia aumentó en más del doble en una década (Askling y
cols. 1999). En Manitoba (Canadá) registraron un incremento anual de 14,6/100.000 en
el período 1984-1995, y una prevalencia de 198,5/100.000 en 1994 (Bernstein y cols.
1999). La prevalencia en una población de Israel era de 25,5/100.000 en 1987 y de
65,1/100.000 en 1997, lo que supone una incidencia anual de nuevos casos para ése
período de 5/100.000 (Niv y cols. 1999).
El estudio del tratamiento de la enfermedad de Crohn ha dado como fruto un ingente
número de publicaciones, siendo menos los trabajos dedicados a determinar su
etiología. Esta última materia parece indispensable a la hora de tratar de conocer y
controlar cualquier enfermedad, y en el caso de la de Crohn, la comunidad científica no
ha llegado a un consenso (Van Kruiningen 1999, Hermon-Taylor y cols. 2000,
Greenstein 2003). Existen tres teorías principales sobre su etiología (Chamberlin y cols.
2006), o combinación de varias (Shanahan 2002): la teoría de autoinmunidad, la de
inmunodeficiencia y la micobacteriana.
Según la teoría de autoinmunidad o de desregulación de la inmunidad, las bacterias
comensales de la flora normal provocan una activación inapropiada y dañina del sistema
inmune de la mucosa, en un huésped genéticamente susceptible. La teoría de la
inmunodeficiencia sugiere que el fenotipo de Crohn se debe a que defectos de la
inmunidad innata comprometen la funcionalidad de los neutrófilos, lo que hace posible
que los microbios sobrevivan por más tiempo, activando asimismo una respuesta Th1
compensatoria excesiva. Por último, la teoría micobacteriana, ha venido sugiriendo
desde los inicios del descubrimiento de esta patología humana la implicación de un
agente micobacteriano, hasta el punto de señalar a Map como agente causal. Map estaría

EPIDEMIOLOGÍA
68
localizado en el interior de los macrófagos, en forma de esferoplasto, y con una
extremadamente baja velocidad de crecimiento (Hermon-Taylor 2002).
Para Chamberlin y cols. (Chamberlin y cols. 2006), las tres teorías tienen mucho en
común, pero difieren a la hora de interpretar los datos o de dar mayor énfasis a los
diferentes aspectos del proceso de la enfermedad. A pesar de haberse llevado a cabo
numerosos trabajos para apoyar o desmentir esta hipotética relación entre Map y el
síndrome de Crohn, ninguno de ellos ha sido concluyente hasta hoy.
Algunos han tratado de demostrar la presencia de Map buscando su DNA por medio
de PCR, hibridación in situ u otros métodos, en muestras de tejido, de biopsias y de
fluidos biológicos de pacientes. Aunque los resultados de algunos de estos trabajos,
apoyarían la relación Map-Crohn (Moss y cols. 1991, Sanderson y cols. 1992a, Moss y
cols. 1992b, Wall y cols. 1993, Fidler y cols. 1994, Lisby y cols. 1994, Dell'Isola y cols.
1994, Mishina y cols. 1996, Collins y cols. 2000, Ikonomopoulos y cols. 2000, Sechi y
cols. 2001, Hulten y cols. 2001, Naser y cols. 2002, Ryan y cols. 2002, Bull y cols.
2003a, Naser y cols. 2004, Sechi y cols. 2004, Romero y cols. 2005, Autschbach y cols.
2005, Sechi y cols. 2005), los resultados de otros tantos o no lo apoyarían o irían en
contra (Yoshimura y cols. 1988, Suenaga y cols. 1995, Rowbotham y cols. 1995,
Murray y cols. 1995, Kreuzpaintner y cols. 1995, Dumonceau y cols. 1996, Frank y
cols. 1996, Cummins y cols. 1996, Al Shamali y cols. 1997, Clarkston y cols. 1998,
Chiba y cols. 1998, Cellier y cols. 1998, Kallinowski y cols. 1998, Kanazawa y cols.
1999, Fujita y cols. 2002, Ellingson y cols. 2003, Baksh y cols. 2004, Ryan y cols.
2004, Tzen y cols. 2006). En los trabajos en que se ha detectado el DNA de esta
bacteria, el porcentaje de positivos en el grupo de pacientes de Crohn varía en el rango
4,7-100%, en el grupo de colitis ulcerosa el rango es 0-100%, y en los controles es 0-
87,5%. Las diferencias más significativas probablemente sean las publicadas por Bull y
cols., al obtener amplificación en el 92% de los pacientes de Crohn frente al 20% de los
controles (Bull y cols. 2003a). Es necesario señalar que la gran mayoría de estos
estudios se basa en la detección del elemento genético IS900.
Algo similar sucede con los estudios serológicos. Mientras unos detectan
anticuerpos específicos (p35, p36, antígeno protoplasmático, 65KD HSP, MPP14,…)
frente a Map en enfermos de Crohn (Elsaghier y cols. 1992, Vanuffel y cols. 1994, El-
Zaatari y cols. 1995, Juste y cols. 1995, Suenaga y cols. 1999, El-Zaatari y cols. 1999,
Collins y cols. 2000, Naser y cols. 2000a, Olsen y cols. 2001c, Shafran y cols. 2002b,
Olsen y cols. 2003, Barta y cols. 2004, Nakase y cols. 2006), lo que demostraría una
respuesta inmunológica frente a esa infección, otros o no los detectan o no observan
diferencias significativas con los grupos control (McFadden y cols. 1988, Kobayashi y
cols. 1989, Brunello y cols. 1991, Ibbotson y cols. 1992, Stainsby y cols. 1993,
Walmsley y cols. 1996, Bernstein y cols. 2004).

REVISIÓN DE LA BIBLIOGRAFÍA
69
Esta divergencia de resultados también se refleja en ensayos clínicos con
tratamientos antimicobacterianos (Hulten y cols. 2000a). Así, algunos apoyan la
implicación de Map tras comprobar que estos tratamientos mejoran la situación de los
pacientes (Warren y cols. 1986, Hermon-Taylor 1998, Hermon-Taylor 2002, Borody y
cols. 2002, Shafran y cols. 2002a), y otros en cambio, no detectan diferencias
significativas (Thomas y cols. 1998).
Ni siquiera el aislamiento de Map de muestras humanas ha podido esclarecer
definitivamente este polémico asunto. Los primeros aislamientos de Map de muestras
procedentes de pacientes de Crohn, se obtuvieron en 1984 a partir de secciones de íleon
terminal extraídas quirúrgicamente a dos niños de 15 y 12 años, y a un anciano de 78
años (cepas Linda, Dominic y Ben, respectivamente) (Chiodini y cols. 1984b, Chiodini
y cols. 1984c, McFadden y cols. 1987). En un trabajo posterior sobre muestras de
tejidos de 18 pacientes de Crohn, 5 con colitis ulcerosa y 6 controles, Map fue aislado y
confirmado por PCR en el 33,3% de los enfermos de Crohn y en el 16,7% de los
controles (Moss y cols. 1992b). Además también se obtuvieron cultivos positivos de
Mas en 2 pacientes de Crohn, 2 de colitis ulcerosa y 2 controles. La bacteria no era
detectada en su forma habitual de bacilo ácido-alcohol resistente, sino en forma de
esferoplasto, de la que tras su cultivo in vitro podía revertir a la bacilar (Chiodini y cols.
1984b, Chiodini y cols. 1986b, Graham y cols. 1987, Gitnick y cols. 1989, Naser y cols.
2000b, Markesich y cols. 2004, Naser y cols. 2004). Esta forma pleomórfica que
presenta Map tanto en animales como en humanos es de crecimiento
extraordinariamente lento y podría ser importante en la interacción con el sistema
inmunitario del hospedador, posiblemente en la presentación de antígenos, haciendo
posible su infección persistente (Imaeda 1984, García Marín y cols. 1992a, Hulten y
cols. 2000b, Hulten y cols. 2000c, Hulten y cols. 2001). Map fue después aislado
incluso de leche materna de mujeres afectadas por este síndrome (Naser y cols. 2000b).
El mismo equipo de investigación, aisló el microorganismo en tejidos del 37% de los
pacientes y del 5,6% de los controles cuando estudiaron muestras de 27 pacientes y 36
controles (Schwartz y cols. 2000). El grupo de Bull y Hermon-Taylor, presentó junto
con los resultados de PCR de tejidos ileocolónicos, unos porcentajes de aislamiento de
un 42% en los enfermos y un 9% entre los controles (Bull y cols. 2003a). Sechi y cols.
(Sechi y cols. 2005) obtuvieron recientemente un 63,3 y un 10,3% de cultivos positivos,
respectivamente.
Pero sin duda el hallazgo más interesante de los últimos tiempos sobre el
aislamiento de Map en relación con esta enfermedad, ha sido el cultivo de la bacteria a
partir de células blancas procedentes de sangre periférica de pacientes (Naser y cols.
2004). El 50% de los enfermos de Crohn y el 22% de los afectados por colitis ulcerosa
albergaban células viables de Map en su sangre, mientras que los cultivos realizados de
controles sanos eran todos negativos. Esto demostraría una bacteriemia en enfermos de
Crohn (y en menor número en pacientes con colitis ulcerosa), y consecuentemente,

EPIDEMIOLOGÍA
70
contribuye a la idea de que Map es una causa de la enfermedad de Crohn. Sin embargo,
muchos investigadores no han conseguido aislar la bacteria de muestras de pacientes
con enfermedad digestiva inflamatoria (Graham y cols. 1987, Kreuzpaintner y cols.
1995, Clarkston y cols. 1998, Kallinowski y cols. 1998, Collins y cols. 2000), por lo que
algunos de ellos no apoyan esa hipótesis. Algunos trabajos (Kallinowski y cols. 1998,
Tiveljung y cols. 1999, Fujita y cols. 2002) sobre la detección de otros microorganismos
a parte de Map (Escherichia coli, Yersinia spp., Helicobacter spp., Listeria
monocytogenes, Bacteroides vulgatus, …), sugieren la posible implicación de otros
agentes, o el rechazo de la hipótesis de la relación Map-Crohn, pudiéndose clasificar la
presencia de Map como secundaria.
Si existiera esa relación, se podría pensar que la exposición a paratuberculosis
clínica bovina, supone un factor de riesgo, y que la prevalencia de Crohn en grupos de
personas expuestas sería mayor. No obstante, esta hipótesis no encuentra confirmación
en un reciente trabajo (Jones y cols. 2006). Por otro lado está el factor de
susceptibilidad-resistencia que explicaría por qué esta micobacteria, a pesar de ser
detectada tanto en pacientes como en controles, sólo provoca la enfermedad en ciertos
individuos. En el año 2001, tres grupos de investigación independientes publicaron el
hallazgo de mutaciones en el gen NOD2/CARD16 del cromosoma 16, asociadas con la
enfermedad de Crohn, pero no con la colitis ulcerosa (Grant 2005). Este gen parece
estar relacionado con el reconocimiento de muropéptidos bacterianos que derivan del
peptidoglicano de la pared celular. Las tres mutaciones principales detectadas, han
demostrado alterar profundamente las vías metabólicas pro-inflamatorias en respuesta a
muropéptidos sintéticos. NOD2 ha sido propuesto como factor antibacteriano en las
células epiteliales del intestino humano, provocando la muerte directa de Salmonella
typhimurium intracelular. De todos modos, la estimación de la proporción de casos de
Crohn atribuibles a mutaciones en este gen es del 15-30%, lo que implicaría otros
factores genéticos y/o ambientales en la patogénesis de esta enfermedad (Grant 2005).
Tal y como concluyen los grupos de expertos de la Comisión Europea y otros
organismos, atribuir la responsabilidad etiológica de Crohn a Map, simplemente por ser
detectada con mayor frecuencia en el aparato digestivo de estos enfermos, no está
suficientemente justificado (European Commission 2000). Se puede pensar en varios
motivos que posiblemente explicarían ese hecho. Sin embargo, existen evidencias que
sugieren algún tipo de asociación entre Map y al menos algunos casos de Crohn, por lo
que ésa vinculación continúa siendo convincente (Grant 2005). Más información acerca
de la epidemiología y la posible relación Map-Crohn puede ser consultada en el informe
de la “Food Standards Agency” (Rubery 2001).
Map también ha sido relacionado con la sarcoidosis (El Zaatari y cols. 1996).
Además, en el último congreso internacional sobre paratuberculosis (8th International
Colloquium on Paratuberculosis, agosto 2005, Copenhague, Dinamarca), un estudio que

REVISIÓN DE LA BIBLIOGRAFÍA
71
detectó el DNA de Map (IS900 y hspX) (Ellingson y cols. 1998) en 6 pacientes con
síndrome de Blau ha atribuido a esta bacteria parte de la etiología de esta afección, una
enfermedad autosómica dominante, sistémica, e inflamatoria, similar a una sarcoidosis
juvenil (Dow 2006b). El propio autor postuló que Map es un “súper-antígeno”, y que
también podría estar implicado en la diabetes mellitus tipo 1 (Dow 2006a).
Map en la cadena alimentaria del ser humano. La leche, el agua y otros
productos. Inactivación de Map en leche
Debido al creciente interés y preocupación suscitados en torno a la posible
asociación entre Map y la enfermedad de Crohn, han sido investigados varios
hipotéticos vehículos de transmisión para el ser humano. El producto más veces
estudiado hasta el momento es la leche. Tal y como se ha mencionado en el apartado de
Transmisión, Map ha sido cultivada de leche de vacas con paratuberculosis clínica o
subclínica (Giese y cols. 2000, Pillai y cols. 2002). La contaminación de ésta en las
explotaciones puede producirse por eliminación directa en la leche de vacas infectadas,
o por contacto con heces infectadas en el procesado. Ha sido calculado que la leche de
tanque de un rebaño con casos de paratuberculosis puede encontrarse contaminada con
11,67 ± 12,33 CFUs por cada 100 ml (Herman y cols. 2006).
Varios estudios demuestran por medio de PCR y/o cultivo la presencia de Map en
leche destinada al consumo humano (Tabla 3). El primer trabajo publicado, reveló la
presencia de IS900 en el 7% de 312 muestras de leche pasterizada procedentes del
Reino Unido, además de encontrar la micobacteria en el 4,8% de los cultivos realizados
(Millar y cols. 1996). Posteriormente, Map ha sido detectada en leche ovina y/o caprina
de tanque o individual en el Reino Unido (Grant y cols. 2001a), Noruega (Djonne y
cols. 2003) y Suiza (Muehlherr y cols. 2003), así como en leche bovina de tanque o
individual en Suiza (Stephan y cols. 2002, Corti y cols. 2002, Bosshard y cols. 2006),
España (Sevilla y cols. 2003), Estados Unidos (Pillai y cols. 2002, Stabel y cols. 2002b,
Jayarao y cols. 2004), Reino Unido (Grant y cols. 2002a), Irlanda (O'Reilly y cols.
2004) y República Checa (Ayele y cols. 2005).
En el único estudio que se ha publicado sobre carne picada para consumo humano,
no han sido halladas células viables de Map (Maher y cols. 2004). Tampoco se
obtuvieron resultados positivos en el estudio realizado sobre muestras de músculo de
100 bovinos procedentes del matadero de Bilbao (Neiker, datos sin publicar).
Dada la alta persistencia de esta bacteria en el medio, también está siendo
investigado el posible papel del agua como vehículo de infección para el ser humano.
Experimentos del efecto del cloro utilizado para potabilizar el agua sobre la viabilidad
de Map, indican una reducción de células viables en el rango 1,32-2,82 log10 (2 mg/ml
cloro) (Whan y cols. 2001), que se ven reducidas a 0,94 log10 al encontrarse en el
interior de protozoos de vida libre, donde incluso pueden hasta multiplicarse (Mura y

EPIDEMIOLOGÍA
72
cols. 2006, Whan y cols. 2006). Map ha sido detectado por PCR en el 32,3% de las 96
muestras de agua del río Taff (Reino Unido) analizadas y aislado del 12,5% de las
mismas (Pickup y cols. 2005). En Irlanda del Norte se aisló en el 4,16% de 192
muestras de agua del afluente de entrada a plantas potabilizadoras, además de encontrar
su DNA en el 8% de ellas (Whan y cols. 2005b). Pickup y cols. han demostrado la
presencia de este patógeno en zonas de captación de aguas, en agua de río para uso
doméstico y en el efluente de plantas de aguas residuales (Pickup y cols. 2006).
Tabla 3. Estudios sobre la detección de Map en productos lácteos.
Estudio País Tipo de muestra PCR+/total (%) Cultivo+/total (%) descontaminación
(Millar y cols. 1996) Reino Unido
Leche comercial pasterizada bovina 22/312 (7)
9/18 (50) PCR+ 6/36 (16) PCR-
0,1% Cloruro de Benzalkonio ó 0,1/0,7% HPC, 18 h.
(Grant y cols. 2001a) Reino Unido
Leche tanque ovina-caprina 0/14 (0) - 1/90 (1,1) 0/14 (0) - 0/90 (0) 0,75% HPC, 5 h.
(Muehlherr y cols. 2003) Suiza Leche tanque ovina-caprina
15/63 (23,8) - 79/344 (23) - -
(Stephan y cols. 2002) Suiza Leche tanque bovina 112/501 (22,4) - -
(Corti y cols. 2002) Suiza Leche tanque bovina 273/1384 (19,7) - -
(Bosshard y cols. 2006) Suiza Leche tanque bovina 3/100 (3) - -
(Sevilla y cols. 2003)
España (Gral.-Norte)
Leche tanque bovina
7/70 (10) - 16/200 (8) - -
(Pillai y cols. 2002) EEUU Leche individual bovina 10/20 (50) 1/20 (5) 0,75% HPC, 18 h.
(Stabel y cols. 2002b) EEUU Leche tanque bovina 35/52 (67,3) 0/52 (0) 0,9% HPC, 18 h. (37ºC)
(Grant y cols. 2002a) Reino Unido
Leche tanque bovina 19/244 (7,8) 4/244 (1,6) 0,75% HPC, 5 h.
(Grant y cols. 2002a) Reino Unido
Leche comercial pasterizada bovina 67/567 (11,8) 10/567 (1,8) 0,75% HPC, 5 h.
(O'Doherty y cols. 2002b) Irlanda
Leche tanque bovina - 0/310 (0) 0,75% HPC, 5 h.
(O'Doherty y cols. 2002b) Irlanda
Leche comercial pasterizada bovina-caprina - 0/77 (0) – 0/9 (0) 0,75% HPC, 5 h.
(Djonne y cols. 2003) Noruega Leche individual caprina 24/340 (7,1) 0/340 (0) 0,75% HPC, 5 h.
(Jayarao y cols. 2004) EEUU Leche tanque bovina 8/29 (27,5) 6/29 (20,6) 0,75% HPC, 18 h.
(O'Reilly y cols. 2004) Irlanda Leche tanque bovina 50/389 (12,9) 1/389 (0,25) 0,75% HPC, 5 h.
(O'Reilly y cols. 2004) Irlanda
Leche comercial pasterizada bovina 35/357 (9,8) 0/357 (0) 0,75% HPC, 5 h.
(Ayele y cols. 2005) República Checa
Leche comercial pasterizada bovina - 4/244 (1,6) 0,75% HPC, 5 h. (37ºC)
(Ellingson y cols. 2005a) EEUU
Leche comercial pasterizada bovina 452/702 (64) 20/702 (2,84) no
(Ikonomopoulos y cols. 2005) Grecia Queso Feta 21/42 (50) 2/42 (4,7) 0,75% HPC, 5 h. (Ikonomopoulos y cols. 2005)
República Checa
Queso de leche bovina 5/42 (11,9) 1/42 (2,4) 0,75% HPC, 5 h.
(Hruska y cols. 2005) Países europeos
Leche en polvo infantil 25/51 (49) 18/51 (35,3) 0,75% HPC, 4 h.
(Paolicchi y cols. 2006) Argentina
Leche comercial pasterizada bovina 2/70 (2,9) 2/70 (2,9) 0,75% HPC, 18 h.
(Clark, Jr. y cols. 2006) EEUU Requesón de leche bovina 5/98 (5) 0/98 (0) no

REVISIÓN DE LA BIBLIOGRAFÍA
73
Como consecuencia de todos estos hallazgos la preocupación sobre la salud pública
ha crecido considerablemente, y los estudios sobre inactivación de Map se han
multiplicado. En 1996 fue demostrada su persistencia en leche pasterizada por dos
métodos diferentes (estándar y estático: 63,5ºC durante 30 min.; HTST o alta
temperatura corto tiempo: 71,7ºC durante 15 s.) al ser inoculada con grandes cantidades
de la bacteria (Grant y cols. 1996). Posteriormente, Stabel y cols. publicaron un trabajo
en el que se afirmaba que el método HTST era suficientemente efectivo para la
inactivación de Map (Stabel y cols. 1997). Mientras algunos autores afirmaban aislar el
bacilo de leche tratada por HTST hasta con concentraciones iniciales de 10 CFU/ml
(Sung y cols. 1998, Grant y cols. 1998a), otros defendían la total inactivación de Map al
menos cuando se tata de concentraciones esperables en leche cruda natural (Keswani y
cols. 1998, Stabel 2000b). La ampliación de los tiempos de pasterización (72ºC, 25 s.) sí
parecía ser suficiente para acabar con todas las CFUs (Grant y cols. 1999), pero el
incremento en la temperatura y duración de este tipo de tratamientos incide
negativamente en las propiedades y características organolépticas de la leche.
En estudios recientes (a nivel de laboratorio, en plantas piloto o en plantas
industriales) sobre una variedad de condiciones térmicas y temporales, la HTST ha
conseguido una reducción considerable de células viables de Map en general (4-7 log10).
Sin embargo, exceptuando los trabajos de Pearce y cols. y de Stabel y cols. (Pearce y
cols. 2001, Stabel 2001) donde descontaminan la muestra tratada antes de ser cultivada,
se ha podido observar la supervivencia de pequeñas cantidades de Map, tanto en
estudios donde se ha inoculado artificialmente la bacteria (Gao y cols. 2002, Hammer y
cols. 2002, Stabel y cols. 2004b, McDonald y cols. 2005, Grant y cols. 2005a, Hammer
y cols. 2006), como en los que se ha utilizado leche infectada de modo natural (Grant y
cols. 2002b, Ayele y cols. 2005). Estos hallazgos invitan a proseguir la búsqueda de
métodos más eficaces de esterilización.
En vista de las diferencias que se observan entre los resultados y conclusiones de
todos estos trabajos, la propuesta de varios grupos de investigación ha sido la adopción
de métodos consensuados que contemplen las posibles causas de esas diferencias (Klijn
y cols. 2001, Hasting y cols. 2001, Lund y cols. 2002). El trabajo realizado para tratar
de mejorar y/o unificar los métodos de detección de Map en leche, en otros productos
lácteos, y en agua, es muy abundante (Grant y cols. 2000, Odumeru y cols. 2001,
Dundee y cols. 2001, Stratmann y cols. 2002, Smith y cols. 2003, Grant y cols. 2003,
Donaghy y cols. 2003a, Donaghy y cols. 2003b, Grant y cols. 2004, Sasahara y cols.
2004, Rodriguez-Lazaro y cols. 2004, O'Mahony y cols. 2004, Khare y cols. 2004,
Wiszniewska y cols. 2004, Tasara y cols. 2005, Rodriguez-Lazaro y cols. 2005, Whan y
cols. 2005a, Stratmann y cols. 2006, Bosshard y cols. 2006, Metzger-Boddien y cols.
2006).

EPIDEMIOLOGÍA
74
Map es también resistente al curado de los quesos, aunque durante el proceso los
recuentos de células viables van disminuyendo lenta pero progresivamente (Sung y cols.
2000, Spahr y cols. 2001, Donaghy y cols. 2004). En la viabilidad de Map durante el
curado influyen factores como las temperaturas de procesado, el pH, la cantidad de
agua, y evidentemente, el grado de contaminación de la leche utilizada para su
elaboración. Es necesario tener en consideración el efecto concentrador del curado, lo
que puede suponer pasar de 10 CFU Map/ml de leche a 100 CFUs Map/g de queso
(Donaghy y cols. 2004). El tiempo necesario para obtener una reducción de 1 log10 en
los recuentos de Map (valor de supervivencia D10) es de 36,5-59,9 días para el queso
fresco (Sung y cols. 2000), 45,5 días para el Tilsiter suizo (Spahr y cols. 2001), 27,8
días para el Emmental suizo (Spahr y cols. 2001), 90-107 días para el Cheddar
(Donaghy y cols. 2004).
Según Grant y cols., el fracaso observado en el aislamiento de Map tanto en estudios
de detección en productos lácteos como de supervivencia a tratamientos de
esterilización, puede deberse a bajas concentraciones de células viables, métodos de
cultivo ineficaces, o al mal procesamiento de muestras (Grant y cols. 2004). Factores
como el volumen de muestra analizado, la descontaminación química de la muestra, o el
tiempo transcurrido desde el tratamiento térmico hasta el análisis (inmediatamente o
refrigeración hasta su uso) parecen tener efecto en la eficacia de aislamiento de Map a
partir de leche pasterizada (Grant y cols. 2004, Gao y cols. 2005). La tendencia de
formar grumos que presenta esta micobacteria, ha sido señalada como posible
mecanismo de resistencia a los tratamientos térmicos (Klijn y cols. 2001), proponiendo
que las bacterias del interior del grumo estarían protegidas del calor (>104
bacterias/grumo). Aunque esta hipótesis ha sido ampliamente descartada, sí parece que
este fenómeno pueda influir en la supervivencia de Map, como demostraría la mayor
inactivación de células observada al romper estos grumos por medio de homogenización
antes de ser pasterizadas (Grant y cols. 2005a). Herman y cols. descartan totalmente la
hipótesis de Klijn y cols. (2001), pues dicen no detectar grumos tan grandes en leche ni
en heces de vaca en casos de paratuberculosis natural (Herman y cols. 2006). Además,
demuestran una activación de Map en leche por medio de calentamiento, lo que
explicaría su persistencia a este tipo de tratamientos, hipótesis previamente propuesta
por Hammer y cols. (2002).
La homogenización que se aplica a la leche para prevenir la formación de nata
rompiendo los glóbulos de grasa, favorece la inactivación de Map en el proceso de
pasterización, sin embargo no garantiza la muerte del 100% de estas bacterias (Grant y
cols. 2002b, McDonald y cols. 2005, Grant y cols. 2005a). Otro proceso normalmente
aplicado a la leche es la centrifugación (bactofugación), con el fin de precipitar esporas
de clostridios (Torres-Anjel y cols. 1971) y otras partículas. En condiciones de
laboratorio, una centrifugación de 10 segundos a 7.000 g (precalentada a 60ºC) también
parece ser capaz de eliminar una media de 85 a 93% o de 74 a 79% de las células de

REVISIÓN DE LA BIBLIOGRAFÍA
75
Map de leche artificialmente inoculada, dependiendo de si el inóculo estaba formado
por células libres o grumos (Grant y cols. 2005b).
Es necesario estudiar otros procesos de esterilización que no alteren las cualidades
organolépticas y nutritivas de la leche, y que consigan eliminar completamente las
células viables de Map. En algunos países ya se vende leche microfiltrada (PurFiltreTM),
y aunque las primeras pruebas de laboratorio con Map parezcan prometedoras (Grant y
cols. 2005b), el principal inconveniente es que la microfiltración sólo es aplicable a
leche desnatada. El uso de irradiación gamma (Stabel y cols. 2001a), de campos
eléctricos pulsados (Rowan y cols. 2001) y de luz de láser de ultravioletas (Smith y cols.
2002), son algunos de los métodos potenciales para la esterilización de la leche, que ya
han sido objeto de estudios preliminares con Map. La aplicación de altas presiones
también ha sido empleada en la inactivación de microorganismos patógenos
(Staphylococcus aureus, Listeria sp., Escherichia coli,...) que se encuentran en
alimentos (Timson y cols. 1965, Butz y cols. 1986, Hoover y cols. 1989, Mertens y cols.
1992, Cuoghi 1993, Trujillo 2002), sin embargo no existían trabajos sobre sus efectos
sobre Map. La leche cruda presurizada con 400-600 MPa posee una calidad
microbiológica similar a la de la pasterizada (72 ºC, 15 s) (Mussa y cols. 1997, Buffa y
cols. 2001, Trujillo 2002), y aunque no destruye las esporas bacterianas (por ejemplo
Bacillus sp.), se puede estimular su germinación con presiones entre 50-300 MPa,
convirtiéndose en susceptibles a los tratamientos térmicos o de presión posteriores
(Smelt 1998). La efectividad de los tratamientos de leche con altas presiones puede
incrementarse si se combinan con el uso de bacteriocinas (nisina, lacticina, lisocima,...)
y/o con temperaturas de 30-50 ºC (García-Graells y cols. 1999, Morgan y cols. 2000,
Lopez-Pedemonte y cols. 2003, Rodriguez y cols. 2005, Arques y cols. 2005).
Epidemiología molecular
Las estrategias de prevención y control de enfermedades deben asentarse en el
entendimiento epidemiológico de la enfermedad, para lo que es necesario conocer las
posibles adaptaciones evolutivas entre el hospedador y el parásito, que permiten a éste
multiplicarse en las poblaciones susceptibles. Estas adaptaciones se reflejan en el
genoma del parásito y son susceptibles de ser detectadas por medio de técnicas llamadas
genéricamente de genotipado. Las micobacterias han sido objeto de numerosos trabajos
de investigación en este campo debido principalmente a su implicación en importantes
enfermedades animales y humanas. La dificultad en la diferenciación fenotípica por
medio de los métodos estándares clásicos ha generado una gran controversia en la
clasificación taxonómica de las especies y subespecies de este género. La distinción a
nivel de cepa, ha sido posible gracias a la caracterización genética de estos
microorganismos por medio de técnicas moleculares. Tal y como se ha comentado
anteriormente, la identificación de los aislamientos de Map se basa en la alcohol-ácido
resistencia, su extremadamente baja tasa de crecimiento, la dependencia de micobactina

EPIDEMIOLOGÍA
76
exógena, y a la presencia de varias copias de la secuencia IS900 insertadas en su
genoma. No obstante, esto podría no ser suficiente debido a la descripción de algunas
cepas no dependientes de micobactina (Aduriz y cols. 1995a, Motiwala y cols. 2004), y
a la presencia de elementos similares a la IS900 (IS900-like) en otras bacterias (Cousins
y cols. 1999, Naser y cols. 1999, Englund y cols. 2002, Motiwala y cols. 2004).
En el caso de Map, la técnica de tipado genético más utilizada es el análisis RFLP
por medio de sondas específicas de IS900, tras la digestión del DNA cromosómico con
diferentes enzimas (IS900-RFLP). En el primer trabajo publicado de IS900-RFLP,
fueron analizadas 4 cepas de referencia y 46 aislamientos de vacas, ovejas y cabras
procedentes de Nueva Zelanda, Australia, Canadá, Islas Feroe y Noruega (Collins y
cols. 1990a). Tras digerir el DNA micobacteriano con la endonucleasa BstEII,
obtuvieron 10 patrones de hibridación diferentes que fueron clasificados en dos grandes
grupos. Estos pasaron a denominarse tipo bovino (“cattle type”, C) y tipo ovino (“sheep
type”, S), debido a su aparente predominancia en esas especies hospedadoras, a pesar de
no ser estrictamente específicas. Dos ovejas procedentes de Canadá, estaban infectadas
por un tipo de cepa que designaron intermedio (I). La dificultad en el cultivo de las
cepas de tipo S quedó claramente patente. Desde entonces se ha mantenido esta
hipótesis de que existen dos principales grupos de cepas de Map.
Un estudio estadounidense posterior que incluía 29 cultivos de origen bovino, y sólo
4 procedentes de pequeños rumiantes, no pudo identificar los dos grupos de cepas
detectados en el trabajo de Collins (Whipple y cols. 1990), probablemente por ser una
muestra más sesgada. No obstante, se subrayó el poder discriminatorio de BstEII sobre
el de otros enzimas de restricción (BclI, PstI y PvuII). Más tarde, el genotipado de 5
aislamientos de ovejas surafricanas reveló que estos pertenecían al tipo intermedio (de
Lisle y cols. 1992). En Nueva Zelanda, tras analizar 20 aislamientos obtenidos de
ciervos de explotaciones establecidas en pastos previamente utilizados por vacas y
ovejas, se observó que el 15% de las cepas se incluía en el tipo S, mientras el 85%
restante lo hacía en el tipo C (de Lisle y cols. 1993). El análisis PvuII-IS900-RFLP de
aislamientos procedentes de 16 cabras de Noruega, 5 vacas y 2 ciervos de Dinamarca,
una oveja de Grecia, y una oveja de las Islas Feroe, demostró que todos ellos eran
similares entre sí, exceptuando el de la oveja de las Islas Feroe, que era marcadamente
diferente (Thoresen y cols. 1994b). Pavlik y cols. detectaron 3 grupos de patrones PstI-
IS900-RFLP consistentes con los descritos hasta el momento con otros enzimas, al
estudiar 84 cultivos aislados de 70 animales (56 vacas, 13 ovejas y una cabra)
procedentes de Europa (principalmente Eslovaquia y República Checa), uno de un
paciente de Crohn estadounidense, y 5 de vacas australianas (Pavlik y cols. 1995). En
otro trabajo, fueron tipados y clasificados 28 aislamientos ovinos procedentes de África
(26 de Marruecos y 2 de Sudáfrica), 7 caprinos y 9 bovinos de Alemania, y 12 bovinos
de Marruecos y EEUU, en base a la velocidad de crecimiento y el análisis PvuII-IS900-
RFLP (Bauerfeind y cols. 1996). Los cinco patrones diferentes hallados fueron

REVISIÓN DE LA BIBLIOGRAFÍA
77
clasificados en dos grupos. Los del primer grupo, los denominados 1, 2 y 3, sólo fueron
detectados en ovejas, y fenotípicamente eran de crecimiento lento. Contrariamente,
todos los cultivos bovinos y caprinos fueron de crecimiento rápido, y todos compartían
el mismo patrón de hibridación (4) a excepción de uno (5). Las micobacterias obtenidas
de raspados de mucosa intestinal de 5 ovejas australianas pertenecían al tipo S1
previamente descrito por Collins y cols. (Choy y cols. 1998). El tipado de 15
aislamientos de conejo en Escocia reveló que 14 de ellos tenían el mismo patrón PstI-
BstEII-IS900-RFLP (B-C17) de tipo bovino (Greig y cols. 1999).
Posteriormente, este método (PstI-BstEII-IS900-RFLP) fue propuesto por Pavlik y
cols. como técnica estándar para el tipado de cepas de Map, siendo aplicado a 1008
aislamientos de diversos hospedadores y regiones (Pavlik y cols. 1999a). En ese trabajo
fueron detectados los mismos dos grupos principales y el intermedio (Figuras 6 y 7),
pero según los autores no existía relación de especificidad o correlación entre cepas,
velocidad de crecimiento y hospedadores. Entre 61 aislados de vacas y ciervos
procedentes de Argentina, fueron diferenciados 4 patrones de BstEII y 2 de PstI, todos
de tipo bovino (Moreira y cols. 1999). Además el 75% de los mismos pertenecían a un
único PstI-BstEII-IS900-RFLP, el B-C17, prevalente en el Reino Unido.
En el año 2000 se llevaron a cabo dos importantes trabajos sobre epidemiología
molecular en Australia, y ambos subrayaron la restringida diversidad de cepas
observada (Cousins y cols. 2000, Whittington y cols. 2000b). Todos los cultivos ovinos
analizados fueron cepas de tipo S, y casi todos tenían exactamente el mismo patrón (S1),
al contrario que los obtenidos de bovino, caprino, alpaca, rinoceronte y humano, que
eran de tipo C. En el trabajo de Cousins y cols. el 66% de todos los aislados y el 59% de
los de origen bovino eran del tipo C3. Mientras, las cepas de tipo C3 y C1
conjuntamente, representaron el 85% de los aislados en el de Whittington y cols., en el
Figura 6. A la derecha, representación gráfica de los patrones de bandas IS900-RFLP obtenidos con la endonucleasa PstI por Pavlik y cols., al analizar más de 1000 aislamientos de Map (Pavlik y cols. 1999a). A cada patrón se le asigna una letra correlativa. Las flechas indican bandas que aparecen o desaparecen en comparación con el tipo C1. A la izquierda, el dendrograma que representa el grado de similitud entre los distintos patrones de bandas.

EPIDEMIOLOGÍA
78
que además detectaron la ausencia de cepas C3 en bovino de carne y otras diferencias
interesantes en la distribución de las cepas de vacas de leche y de vacas de carne que
relacionaron con la diferenciación entre el mercado lechero y el cárnico. En la
República Checa, han sido analizados por el método RFLP propuesto previamente como
estándar aislamientos obtenidos de animales salvajes libres o de explotaciones (Pavlik y
cols. 2000a, Machackova y cols. 2004). Estos estudios sugieren que la prevalencia del
tipo B-C1 es la más alta entre las especies ganaderas tradicionales, y que va en aumento
entre especies salvajes. Más recientemente, la idea de una diversidad genética
restringida entre aislamientos de Map se ha visto reforzada en Noruega al haber
detectado que el 84% de los cultivos tipados (51 de caprinos y 4 bovinos) eran del
mismo tipo (B-C1) (Djonne y cols. 2005).
Figura 7. A la derecha, representación gráfica de los patrones de bandas IS900-RFLP obtenidos con la endonucleasa BstEII por Pavlik y cols., al analizar más de 1000 aislamientos de Map (Pavlik y cols. 1999a). A cada patrón se le asigna un nombre compuesto de una letra (indica si es tipo C, S o I) y un número correlativo. Las flechas indican bandas que aparecen o desaparecen en comparación con el tipo C1. A la izquierda, el dendrograma que representa el grado de similitud entre los distintos patrones de bandas.

REVISIÓN DE LA BIBLIOGRAFÍA
79
Un grupo de investigación australiano desarrolló en 1999 una técnica rápida y
sencilla (IS1311 PCR-REA) que permite diferenciar entre M. avium y las cepas C y S de
Map basada en la detección de mutaciones puntuales en la secuencia IS1311 presente en
estas bacterias (Marsh y cols. 1999). En un trabajo posterior se pudo observar que
existía un cuarto polimorfismo en la IS1311 en aislados de Map obtenidos de bisontes
de EEUU (Whittington y cols. 2001b), que denominaron tipo B (“bison type”, B). Las
cepas tipo B dan patrones de RFLP agrupables con las de tipo C.
Otra técnica prometedora para realizar el tipado genético de esta bacteria parásita, es
la electroforesis en gel de campo pulsado del genoma digerido. Tal y como se ha
revisado anteriormente, el PFGE ha sido de gran utilidad para el tipado de otras
importantes micobacterias como M. avium y M. tuberculosis. En el primer trabajo
publicado de tipado de Map por medio de PFGE, fueron comparados los patrones de
restricción de 35 aislamientos (27 bovinos, 5 de alpacas, 2 ovinos y uno caprino) con
varias endonucleasas (Feizabadi y cols. 1997b). No se detectó ninguna variabilidad con
DraI, sin embargo, XbaI y SpeI diferenciaron los cultivos en dos grupos muy similares
entre sí. Un estudio escocés sobre conejos mencionado anteriormente, obtenía
resultados parecidos al del IS900-RFLP (Greig y cols. 1999).
En un intento de estandarizar el PFGE para el tipado de cepas de Map, Stevenson y
cols. llevaron a cabo la caracterización molecular de 5 aislamientos pigmentados (todos
ovinos) y 88 no pigmentados (18 ovinos y el resto procedentes de vacas, conejos,
cabras, ciervos y otras especies silvestres) por varios métodos (Stevenson y cols. 2002).
Los enzimas elegidos para PFGE fueron SnaBI y SpeI, y los patrones distintos con cada
Figura 8. Fotografía de los patrones de bandas PFGE obtenidos con la endonucleasa SnaBI por Stevenson y cols. al analizar 93 aislamientos de Map (Stevenson y cols. 2002). Los rótulos de encima corresponden al nombre del aislamiento. Los números de abajo corresponden al nombre asignado a cada pulsotipo (patrón de bandas) diferente. Los patrones que figuran en el panel de la izquierda se agrupan en el Tipo II, y los de la derecha en el Tipo I.

EPIDEMIOLOGÍA
80
enzima se designaron con un número, lo que da como resultado un perfil múltiple de
campo pulsado con dos números para cada aislado (nº patrón SnaBI-nº patrón SpeI). De
ese modo, en esa muestra fueron descritos 16 perfiles múltiples que se agrupaban en dos
grandes grupos o tipos (Tipo I y II; Ver Figuras 8 y 9).
Figura 10. Fotografías de los nuevos patrones de bandas PFGE obtenidos con las endonucleasas SnaBI (izquierda) y SpeI (derecha) por de Juan y cols. al analizar 44 aislamientos de Map (de Juan y cols. 2005). Los números de arriba corresponden al nombre asignado a cada pulsotipo. Patrones SnaBI nuevos, del 12 al 19, y patrones SpeI nuevos, del 10 al 13.
Figura 9. Fotografía de los patrones de bandas PFGE obtenidos con la endonucleasa SpeI por Stevenson y cols. al analizar 93 aislamientos de Map (Stevenson y cols. 2002). Los rótulos de encima corresponden al nombre del aislamiento. Los números de abajo corresponden al nombre asignado a cada pulsotipo diferente. Los patrones que figuran en el panel de la izquierda se agrupan en el Tipo II, y los de la derecha en el Tipo I.

REVISIÓN DE LA BIBLIOGRAFÍA
81
Las cepas comprendidas en el Tipo I, son de crecimiento más lento, con una fuerte
preferencia de hospedador por la especie ovina, y aparentemente más virulenta para este
tipo de hospedador. Las cepas comprendidas en el Tipo II son de crecimiento más
rápido, y son comúnmente aisladas de vacas, cabras, ciervos y otras especies,
incluyendo la humana.
El tipado por medio de SnaBI-SpeI-PFGE de 44 nuevos aislamientos procedentes de
cabras de España, Escocia, y Noruega (30, 11 y 3, respectivamente), reveló 10 nuevos
perfiles múltiples (de Juan y cols. 2005) (Figura 10). Uno de ellos fue hallado sólo en
dos cabras españolas, y al diferir de los comprendidos en los Tipos I y II, fue descrito el
Figura 11. Dendrograma representando la similitud entre todos los perfiles múltiples de PFGE de Map (nº patrón SnaBI-nº patrón SpeI) descritos hasta el momento (de Juan y cols. 2005). Según éste, las cepas se agrupan dentro de tres tipos diferentes: el Tipo I, donde se incluyen todos los aislados pigmentados ovinos descritos por Stevenson y cols. (Stevenson y cols. 2002), el Tipo II, que incluye cepas de varias especies hospedadoras, y el Tipo III, sólo detectado en dos aislamientos caprinos de España (de Juan y cols. 2005).

EPIDEMIOLOGÍA
82
Tipo III, un tipo intermedio considerado más cercano al Tipo I (Figura 11). Así, el Tipo
I se podía considerar comparable al tipo S, el Tipo II al C, y el Tipo III al I. Más
recientemente, de Juan y cols. han observado por medio de PCR cepas de Tipo I/III o
IS1311-S infectando tanto a la especie ovina como a la caprina y a la bovina en España
(de Juan y cols. 2006a). En la Tabla 4 se puede consultar el tipo de cepa que se ha
descrito en varias especies hospedadoras de Map procedentes de varios países,
independientemente de la técnica utilizada.
Tabla 4. Distribución de los tipos de cepas de Map en hospedadores y regiones, según la bibliografía.
CEPAS DE TIPO BOVINO CEPAS DE TIPO OVINO O INTERMEDIO Hospedadores Países Bibliografía Países Bibliografía Bovino Nueva Zelanda,
Australia, EEUU, Dinamarca, República Checa, Eslovaquia, Hungría, Alemania, Francia, Marruecos, Reino Unido, Austria, Italia, Países Bajos, Eslovenia, España, Suecia, Argentina y Noruega
(Whipple y cols. 1990, Collins y cols. 1990a, Thoresen y cols. 1994b, Pavlik y cols. 1995, Bauerfeind y cols. 1996, Moreira y cols. 1999, Greig y cols. 1999, Pavlik y cols. 1999a, Cousins y cols. 2000, Whittington y cols. 2000b, Djonne y cols. 2005)
Australia, Islandia y España
(Whittington y cols. 2001d, de Juan y cols. 2006a)
Ovino Canadá, EEUU, República Checa, Francia, Grecia, Australia, España y Reino Unido
(Whipple y cols. 1990, Collins y cols. 1990a, Thoresen y cols. 1994b, Pavlik y cols. 1995, Pavlik y cols. 1999a, Stevenson y cols. 2002)
Canadá, Nueva Zelanda, Islas Feroe, Sudáfrica, Marruecos, Australia, Islandia, Noruega, Reino Unido, EEUU y España
(Collins y cols. 1990a, de Lisle y cols. 1992, Thoresen y cols. 1994b, Bauerfeind y cols. 1996, Choy y cols. 1998, Cousins y cols. 2000, Whittington y cols. 2000b, Whittington y cols. 2001d, Stevenson y cols. 2002, Motiwala y cols. 2004, de Juan y cols. 2006a)
Caprino Nueva Zelanda, Noruega, Argentina, Alemania, Dinamarca, Italia, Reino Unido, Australia, EEUU, y España
(Whipple y cols. 1990, Collins y cols. 1990a, Thoresen y cols. 1994b, Bauerfeind y cols. 1996, Pavlik y cols. 1999a, Cousins y cols. 2000, Djonne y cols. 2005, de Juan y cols. 2005, de Juan y cols. 2006a)
Nueva Zelanda, República Checa, y España
(Collins y cols. 1990a, Pavlik y cols. 1995, de Juan y cols. 2005, de Juan y cols. 2006a)
Cérvidos Nueva Zelanda, República Checa, Reino Unido y Argentina
(de Lisle y cols. 1993, Moreira y cols. 1999, Pavlik y cols. 1999a, Pavlik y cols. 2000a, Machackova y cols. 2004)
Nueva Zelanda y República Checa
(de Lisle y cols. 1993, Machackova y cols. 2004)
Lagomorfos Reino Unido y República Checa
(Greig y cols. 1999, Beard y cols. 2001a, Machackova y cols. 2004)
Alpacas Australia (Cousins y cols. 2000) Rinocerontes Australia (Cousins y cols. 2000) Jabalí República Checa (Machackova y cols.
2004)
Muflón República Checa (Machackova y cols. 2004)
Humanos EEUU, República Checa, Países Bajos y Australia
(Collins y cols. 1990a, Pavlik y cols. 1999b, Naser y cols. 2000b, Whittington y cols. 2000b, Ghadiali y cols. 2004)

REVISIÓN DE LA BIBLIOGRAFÍA
83
Se han desarrollado otras técnicas de caracterización genética para intentar aumentar
el poder de discriminación de los análisis de RFLP y de PFGE (Bull y cols. 2000, Pillai
y cols. 2001, Motiwala y cols. 2003, Ghadiali y cols. 2004, Overduin y cols. 2004,
Amonsin y cols. 2004). Sin embargo, algunas de ellas no proporcionan suficiente
información adicional, y la proporcionada en otros casos no parece del todo fiable.
Otras pueden ser interesantes, pero deben ser examinadas con una muestra
representativa de cepas de distintos orígenes que tengan RFLP y/o PFGE conocidos,
con el fin de verificar su fiabilidad y reproducibilidad para ser utilizadas en posteriores
estudios de epidemiología molecular de paratuberculosis.
Tal y como se comentaba en el apartado de Etiología. Tipado molecular, el método
con mayor poder discriminatorio es el análisis MLSSR, según indicó el índice de
diversidad de Simpson (hasta 0,96), por encima de IS900-RFLP, MPIL y AFLP
(Motiwala y cols. 2003, Motiwala y cols. 2004, Amonsin y cols. 2004). En la actualidad
se está trabajando en la comparación de resultados de PFGE y MLSSR (Neiker-
Universidad de Minesota).
PATOGÉNESIS Y PATOLOGÍA
La patogénesis estudia la manera de originarse y desarrollarse una enfermedad. El
agente patógeno entra en el hospedador y trata de replicarse para después ser liberado y
poder diseminarse, mientras que el hospedador lucha por minimizar los daños causados
y por eliminar el patógeno. Es necesario comprender con claridad la patogénesis de la
enfermedad para poder llegar a prevenir la infección o tratar adecuadamente los
animales infectados. En el proceso patogénico de la paratuberculosis influyen varios
factores entre los que destacan los factores genéticos y el estatus inmune del hospedador
9(Clarke y cols. 1996b, Clarke 1997a, Koets y cols. 2000). Map es una bacteria
intracelular facultativa que al infectar a especies rumiantes causa la enteritis
granulomatosa fatal que conocemos como paratuberculosis o enfermedad de Johne. La
dificultad en el aislamiento de la bacteria (tiempo, cepas de cultivo complicado,…), el
largo período de incubación de la enfermedad (normalmente más de 2 años) y la
dificultad en reproducir las características típicas de ésta de manera experimental (a
pesar del éxito de Map en la naturaleza o precisamente por ello), han obstaculizado el
estudio de sus mecanismos patogénicos, con lo que el conocimiento sobre esta materia
es reducido (Coussens 2004, Stevenson 2006).
Entrada e invasión de Map en el hospedador
Está ampliamente aceptado que el mecanismo natural y principal de introducción de
Map en el organismo es oral, por transmisión fecal-oral, en animales de menos de un
año (Chiodini y cols. 1984a, Blood y cols. 1989, Sweeney 1996), aunque también
existen otras vías de infección (materia revisada en el apartado Transmisión). El

PATOGÉNESIS Y PATOLOGÍA
84
motivo de que animales jóvenes sean más susceptibles a la infección que aquellos de
más edad, parece residir en la extensa superficie que abarcan las placas de Peyer
intestinales en animales jóvenes, que va reduciéndose gradualmente con la edad, ya que
ése parece ser el punto de entrada de la bacteria al organismo (Momotani y cols. 1988,
Liebler y cols. 1988a, Liebler y cols. 1988b).
Map se localiza y reside preferentemente en fagosomas o endosomas inmaduros de
los macrófagos del hospedador, con predominio de aquellos asociados a las placas de
Peyer ileales y yeyunales (Bendixen y cols. 1981, Momotani y cols. 1988, Pérez 1992,
Peris 1992, Corpa y cols. 2000a), después de traspasar las barreras de la mucosa por
endocitosis (Lugton 1999). A pesar de que algunos autores proponen como localización
inicial las tonsilas y ganglios retrofaríngeos (Payne y cols. 1961, Lugton 1999), estas
parecen ser transitorias y desaparecen sin ocasionar trastornos (García Marín y cols.
2000). No obstante, según avanza la enfermedad, los fagocitos infectados diseminan las
micobacterias por otras partes del cuerpo al ser transportados en los vasos sanguíneos y
linfáticos (Merkal 1984, van der Giessen y cols. 1995, Lugton 1999). Se ha detectado la
presencia de Map en sangre, hígado, nódulos retrofaríngeos y mandibulares, bazo,
pulmones, riñones, ubres, glándulas supramamarias, útero, testículos, epidídimo, y
semen (Alexejeff-Goloff 1929, Doyle 1954, Tunkl y cols. 1965, Larsen y cols. 1970,
Merkal y cols. 1982b, Merkal 1984, Koenig y cols. 1993, van der Giessen y cols. 1995,
Collins 1997, Whitlock y cols. 1997, Pavlik y cols. 2000c, Eppleston y cols. 2001,
Ayele y cols. 2004).
La entrada de Map en el organismo está mediada por las células M epiteliales que
recubren las cúpulas de las placas de Peyer del íleon y yeyuno (Momotani y cols. 1988,
García Marín y cols. 1992a), y depende de la expresión de la proteína de membrana de
35 kDa de la bacteria (Bannantine y cols. 2003a), del puente de unión de fibronectina
entre la proteína de acoplamiento de fibronectina (FAP) de Map y las integrinas de las
células M (Secott y cols. 2002, Secott y cols. 2004), o de ambos eventos. Schleig y cols.
mediante un modelo in vitro han demostrado la capacidad de Map para acoplarse a
diferentes zonas del tracto intestinal sin encontrar diferencias significativas, aunque sí
las han observado al emplear diferentes cepas del patógeno (Schleig y cols. 2005).
Parece que también los enterocitos yeyunales no asociados a placas de Peyer son vía de
entrada para los bacilos, al menos en cabras (Sigurdardottir y cols. 2005). Una vez que
las bacterias han logrado entrar, son transportadas transcelularmente de manera libre o
en leucocitos dentro de vacuolas, y posteriormente se introducen en fagosomas de
macrófagos sub- e intra-epiteliales adyacentes a la placa de Peyer (Momotani y cols.
1988, Kaufmann 1995, Fujimura y cols. 1996, Lugton 1999, Valentin-Weigand y cols.
1999, Sigur-Dardottir y cols. 2001, Tessema y cols. 2001). Hipotéticamente, los
antígenos se presentan a los linfocitos T (LT) “de memoria” en ese momento
(Momotani y cols. 1988, García Marín y cols. 1992a), lo que se asocia a una resistencia

REVISIÓN DE LA BIBLIOGRAFÍA
85
de animales de más edad, coincidiendo con la involución del tejido linfoide del íleon
(Larsen y cols. 1975).
La supervivencia y replicación de Map dentro de los macrófagos depende del grado
de activación y diferenciación de los mismos. A pesar de que los macrófagos son
capaces de controlar la propagación de Map, grandes cantidades de la micobacteria son
citotóxicas y provocan su apoptosis (Bannantine y cols. 2002b). Durante la respuesta
inmune celular (mediada por los LT auxiliares 1; LTh 1) se da la multiplicación y
eliminación intracelular de bacilos de forma simultánea (Zhao y cols. 1997). Estos
bacilos son capaces de sobrevivir inhibiendo la acidificación del fagosoma, lo que
impide su maduración a fagolisosoma (Cheville y cols. 2001, Kuehnel y cols. 2001,
Pieters 2001, Hostetter y cols. 2002, Hostetter y cols. 2003, Weiss y cols. 2004a). Map
soporta los enzimas lisosómicos, el óxido nítrico y los demás mecanismos bactericidas
que producen los macrófagos, ayudada entre otras cosas por su envoltura celular lipídica
(Zhao y cols. 1997, Harris y cols. 2001, Tessema y cols. 2001). La bacteria libera súper-
óxido dismutasa (Liu y cols. 2001), suprime la presentación de antígenos a los LT
(Weiss y cols. 2001), y altera la expresión de moléculas del complejo mayor de
histocompatibilidad tipo II y de integrinas tipo LFA-1 (Alzuherri y cols. 1997). Los
mecanismos de defensa que muestran los macrófagos hasta este punto, pueden ser
considerados como inmunidad innata. La iniciación de la respuesta inmune (de
inmunidad adquirida) frente a Map puede verse obstaculizada por una producción
defectuosa de citoquinas proinflamatorias (como el factor de necrosis tumoral, TNF), y
por un incremento en la expresión de interleuquina-10 (IL-10) (Weiss y cols. 2002,
Coussens y cols. 2004). Las proteínas de choque térmico (hsp) probablemente actúan
como activadoras de las células presentadoras de antígeno, pues parece ser que la hsp de
70 kDa micobacteriana es reconocida por estas células y provoca un incremento en la
producción de IL-10 (Langelaar y cols. 2005a, Langelaar y cols. 2005b). Las células
“natural killer” (NK) bovinas han sido recientemente caracterizadas, y podrían jugar un
importante papel en la transición de inmunidad innata a inmunidad adquirida, por medio
de la producción de interferón (IFN) en respuesta a los antígenos micobacterianos
(Storset y cols. 2003, Olsen y cols. 2005).
Factores de virulencia
Se ha tratado de identificar factores de virulencia de Map principalmente a través de
estudios sobre proteínas antigénicas de la bacteria, pues se cree que éstas son la llave de
los mecanismos patogénicos, además de ser específicos (no todos) y potencialmente
útiles en el desarrollo de vacunas y métodos diagnósticos (Stevenson 2006). Al
principio este proceso se llevó a cabo creando bibliotecas de expresión (“expression
libraries”) con antisueros de animales experimentalmente o naturalmente infectados
(método hoy día conocido como IVIAT; tecnología de antígenos inducidos in vivo).
Algunas de las proteínas identificadas por IVIAT han sido utilizadas posteriormente en

PATOGÉNESIS Y PATOLOGÍA
86
pruebas serológicas, pero no han elucidado los mecanismos patogénicos (Stevenson
2006). Tal y como ha sido comentado anteriormente, la inmunogenicidad de las
proteínas de choque térmico ha sido ampliamente estudiada (Koets y cols. 1999,
Langelaar y cols. 2005a, Langelaar y cols. 2005b). La hsp de 70kDa parece jugar un
importante papel con respecto a la protección celular, además de haber sido probada su
utilización para producir una vacuna (Koets y cols. 2006a). La hsp de 34kDa es una
posible serina proteasa, que aumenta la supervivencia de Map en el interior de
macrófagos (Stevenson 2006). La proteína de membrana de 35 kDa interviene en la
invasión de las células epiteliales bovinas (Stevenson 2006). Miembros de entre 30 y 32
kDa de la familia micolil-transferasa (Ag85) de Map son antígenos inmunodominantes
para las células T, y son rápidamente reconocidos tanto en infecciones experimentales
como naturales, lo que los hace buenos candidatos para su uso como vacuna (Rosseels y
cols. 2006). El operón de proteínas transportadoras ABC específico de Map que se
encuentra en una isla genética de patogenicidad de 38 kbp junto a otros genes
relacionados con el metabolismo del hierro y con un posible papel patogénico, es
expresado in vivo e in vitro en la superficie micobacteriana, y dichas proteínas son
reconocidas por el sistema inmune de animales infectados (Stevenson 2006). Gracias a
la secuenciación del genoma completo de Map, es posible llevar a cabo arrays de
proteínas (para identificar aquellas con poder inmunogénico), como han hecho
Bannantine y cols. al haber detectado recientemente 10 proteínas (de las cuáles 7 no
eran conocidas) reconocidas por el sistema inmune de vacas infectadas (Bannantine y
cols. 2006a). Por medio de la prueba de IFN-gamma (IFN-γ) con células mononucleares
de sangre periférica bovina, Nagata y cols. han identificado tres proteínas
recombinantes de las cuales 2 son de la familia PPE (Pro-Pro-Glu) (Stevenson 2006,
Nagata y cols. 2006). La versión recombinante de otra lipoproteína recientemente
identificada de 22 kDa (P22) provocó secreción de IFN-γ en la sangre de 8 de 9 ovejas
vacunadas con la cepa atenuada de Map 316F, que no provocó el mismo efecto en el
grupo de 5 no vacunadas (Dupont y cols. 2005). También se están dando avances en la
identificación de posibles factores de virulencia mediante análisis de electroforesis en
dos dimensiones y espectrometría de masas por medio de ionización-desorción por laser
asistida por matriz acoplada a un analizador de tiempo de vuelo (MALDI-tof) (Hughes
y cols. 2006). Otro abordaje posible es la creación de bibliotecas genéticas a partir de
cepas mutantes, método hasta ahora no usado por la incapacidad de manipular
genéticamente a Map, pero que ya está dando los primeros resultados utilizando
transposones (Stevenson 2006, Shin y cols. 2006).
Respuesta inmune del hospedador
Como es típico en casos de paratuberculosis natural, tras la infección se abre una
fase asintomática de dos años o más, durante la que no es posible observar una respuesta
inmunológica celular ni humoral diferenciada, ni detectar la excreción de la bacteria en

REVISIÓN DE LA BIBLIOGRAFÍA
87
heces (Clarke 1997b). En casos de paratuberculosis inducida experimentalmente, se ha
observado la producción de anticuerpos anti-Map que incluso preceden a la respuesta
celular inicial típica (Waters y cols. 2003). Los estadios iniciales y subclínicos de la
enfermedad se caracterizan por una alta respuesta inmune celular (CMI), mientras en
estadios clínicos avanzados predomina la respuesta inmune humoral (Bendixen 1978,
Chiodini y cols. 1984a, Pérez y cols. 1996, Burrells y cols. 1998, Stabel 2000a, Stabel
2000c). Los primeros signos de la infección son la producción de IFN-� y la
proliferación de linfocitos en respuesta a la inoculación de antígenos micobacterianos
como el derivado proteico purificado (PPD). Esto indica la activación de la inmunidad
celular, y la excreción intermitente de la bacteria en heces. Esta respuesta irá en
incremento junto con la carga micobacteriana excretada en las heces, mientras, los
anticuerpos de la respuesta humoral pueden ir apareciendo. Esta fase de aparente
equilibrio entre patógeno y hospedador puede durar años. Al final, la respuesta celular
cae y la excreción de bacterias aumenta, lo que lleva al estado clínico de la enfermedad
(Chiodini 1996). En la especie bovina se distinguen bajos excretores, excretores
moderados y grandes excretores (o súper-excretores) (Whitlock y cols. 2006b). En
comparación con los menos excretores, el grupo de grandes excretores se asocia con
animales de baja respuesta celular (sobre todo en las fases iniciales de infección),
asentamiento más temprano de la enfermedad y títulos de anticuerpos (respuesta
humoral) más altos (Koets y cols. 2006b).
Se considera que la inmunidad de protección contra Map viene dada por la respuesta
inmune celular, con lo que esa protección depende de las interacciones entre las células
T y las células presentadoras de antígeno infectadas (Kaufmann 2001). En la respuesta
inmune celular, las células T excretan citoquinas que activan la eliminación de Map en
los macrófagos (Stabel 2000c). Los macrófagos presentan los antígenos de Map a las
células T, que liberan IL-2. Esta interleuquina impulsa la propagación de los LT CD8+
citolíticos y los CD4+LTh1 (Chiodini y cols. 1992), y estos últimos, liberan más IL-2,
� -IFN, TNF y factor estimulante de colonias de granulocitos de macrófago (GM-CSF),
con el objetivo de activar el poder bactericida de los macrófagos (Kaufmann 1995,
Stabel 1995, Begara-McGorum y cols. 1998, Valentin-Weigand y cols. 1999, Flesch y
cols. 1999, Lee y cols. 2001, Zur y cols. 2003). Después, la población de CD4+LTh1 cae
impulsada al final por la proliferación de las células T gamma-delta (γδ), entrando así en
una fase anérgica (Chiodini y cols. 1992, Koets y cols. 2002, Langelaar y cols. 2002).
Los cometidos de estos LT-γδ no están del todo claros, y parece que hay distintas
subpoblaciones de estas células con comportamientos diferentes (Park y cols. 2000,
Koets 2006). Hipotéticamente, el descenso de la respuesta inmune celular refleja un
cambio de respuesta de tipo 1 (con LTh1) a tipo 2 (con LTh2) (Chiodini 1996, Clarke
1997b), de acuerdo con la dicotomía de las células T auxiliares (Mosmann y cols.
1986). Los estudios realizados con animales con paratuberculosis no han sido capaces
de demostrar ese hipotético cambio de Th1 protector a Th2 permisivo, sin embargo el

PATOGÉNESIS Y PATOLOGÍA
88
descenso de IFN-γ, y el incremento de IL-10 y TGF-β parecen más evidentes (Lee y
cols. 2001, Stabel y cols. 2002a, Coussens y cols. 2005, Pérez y cols. 2006). La
neutralización de la IL-10 con anticuerpos monoclonales provoca un importante
incremento en la producción de IFN-γ, lo que demuestra el efecto supresor de esta
citoquina sobre la respuesta inmune (Buza y cols. 2004). En macrófagos bovinos
cultivados en presencia de Map se ha observado además el incremento de GM-CSF e
IL-8, así como el descenso de IL-12 y TNF-α (Weiss y cols. 2002). En la última fase, la
fase clínica, los CD4+LTh2 producen citoquinas (IL-4 e IL-10 entre otras) que suprimen
la respuesta celular y activan la respuesta humoral (González 1998), con el aumento de
los linfocitos B (LB) y la producción de anticuerpos (Burrells y cols. 1999, Ostrowski y
cols. 2003, Langelaar y cols. 2005b), quedando establecida la respuesta humoral.
Coussens y cols. han llevado a cabo varios estudios para comprender las
interacciones hospedador-patógeno valiéndose de la tecnología de la PCR a tiempo real
cuantitativa (Q-RT-PCR) y los “expressed sequence tag microarrays” (EST) y cDNA
(Yao y cols. 2001, Coussens y cols. 2002, Coussens y cols. 2003, Coussens 2004,
Coussens y cols. 2004, Coussens y cols. 2005). Demostraron la represión de 83 genes
relacionados con la respuesta inmune celular (que codifican para la proteína de
microesférula 1, el factor de crecimiento de fibroblastos, la proteína quinasa Lyn B,...)
en células mononucleares de sangre periférica de vacas clínicamente afectadas, mientras
que en vacas con paratuberculosis subclínica 71 de estos genes mostraron
sobreexpresión (codificantes para CD40L, varias metaloproteinasas de matriz, y la
proteína secretada ácida y rica en cisteína o SPARC) (Coussens y cols. 2002).
Posteriormente también se observaron diferencias en la expresión de estos genes al
comparar células mononucleares de sangre periférica de vacas infectadas y de no
infectadas (Coussens y cols. 2003). La ampliación del estudio por parte de Aho y cols.
demostró la activación de la citoquina proinflamatoria IL-1 (que se asocia a la
formación y mantenimiento de granulomas) y del señalizador proapoptótico intracelular
denominado proteína asociada al receptor TNF 1 o TRAF1, así como la expresión de
proteínas en íleon de vacas infectadas (Aho y cols. 2003). TRAF1 se encuentra en altos
niveles ante todo en los macrófagos del infiltrado, mientras IL-1α parece ser secretada
por células de dentro de la lesión en animales infectados (Coussens y cols. 2006).
En el último trabajo publicado de este grupo sobre las células de dentro de células
mononucleares de sangre periférica en casos de paratuberculosis bovina, se resaltan tres
conceptos (Coussens y cols. 2005): La adopción del fenotipo predominante Th2 por las
células T. Aparente incremento de los inhibidores de tejido de la metaloproteinasa de
matriz (TIMP) 1 y 2 y descenso de la metaloproteinasa de matriz (MMP) 14, que
ocasiona deficiencias de remodelación de tejidos. Y por último, que estas células están
probablemente preparadas para una rápida apoptosis. Weiss y cols. también han
utilizado esta tecnología para caracterizar la respuesta inmune de macrófagos derivados

REVISIÓN DE LA BIBLIOGRAFÍA
89
de monocitos bovinos (Weiss y cols. 2004a, Weiss y cols. 2004b). Sus resultados tienen
puntos de discrepancia con los de Coussens y cols., muy probablemente por el uso de
diferentes equipos y protocolos, y por la mezcla de linfocitos y monocitos utilizada por
Coussens y cols. (a diferencia de los macrófagos derivados de monocitos usados por
Weiss y cols.). En los últimos trabajos de este grupo se habla de un posible estado de
tolerancia en el intestino de vacas subclínicamente afectadas (Weiss y cols. 2006), y del
posible papel de la quinasa de terminal-Jun N/proteína quinasa activada por estrés
(JNK/SAPK) y de la proteína quinasa p38 activada por mitógeno (MAPK p38) de Map
en la regulación de los mecanismos inmunitarios de los monocitos para evitar sus
efectos antimicobacterianos (Souza y cols. 2006a, Souza y cols. 2006b).
A nivel histopatológico se compara la paratuberculosis con la lepra, en la que fueron
establecidos 5 tipos inmunopatológicos principales que correlacionan las lesiones
histológicas y la respuesta inmune (Ridley 1974): tuberculoide, tuberculoide intermedio,
intermedio, lepromatoso intermedio y lepromatoso. En las formas tuberculoides existe
una fuerte resistencia a la infección detectable por pruebas de inmunidad celular y una
baja (o muy baja) cantidad de bacilos, mientras en las formas lepromatosas se da todo lo
contrario. Estudios en paratuberculosis confirman un modelo similar para esta patología
(Buergelt y cols. 1978b, Chiodini y cols. 1984a, Clarke 1997b), y sugieren que se da
una evolución de una respuesta tuberculoide (lesiones iniciales en el punto de infección)
hacia otra de tipo lepromatoso dependiendo del estado inmunitario del hospedador
(Stamp y cols. 1954, Buergelt y cols. 1978b, Paliwal y cols. 1985, Pérez 1992, Pérez y
cols. 1997, Pérez y cols. 1999, Corpa 1999, Corpa y cols. 2000a, González 2003,
González y cols. 2005). Aunque nunca se ha descartado que esta evolución siga una
secuencia temporal. En el caso de la paratuberculosis se han diferenciado las formas
focales, las multifocales, y las formas difusas/graves, entre las que se distinguen las
multibacilares, las linfocíticas y las intermedias, en relación al espectro
inmunopatológico (Tabla 5) y a la especie.
Tabla 5. Relación entre formas lesionales de paratuberculosis y espectros inmunopatológicos. PARATUBERCULOSIS ESPECTRO INMUNOPATOLÓGICO
Formas focales en la placa de Peyer • formas tuberculoides extremas • respuesta inmune celular y no humoral (o ausencia de ambas periféricamente) • presencia muy escasa de micobacterias
Formas multifocales • formas tuberculoides • respuesta inmune celular y ausencia o débil de la humoral • presencia escasa de micobacterias
Formas difusas y graves: a. Multibacilar • formas intermedias-lepromatosas
• respuesta inmune humoral intensa y ausencia o débil de celular • presencia de numerosas micobacterias
b. Linfocítica • formas intermedias-tuberculoides • respuesta inmune celular intensa y escasa o nula humoral • presencia escasa de micobacterias
c. Intermedias • formas intermedias • respuesta inmune humoral y celular • presencia moderada de micobacterias
*Las formas focales lepromatosas puras no han sido descritas en paratuberculosis. Extraído de García Marín y cols. (García Marín y cols. 2000).

PATOGÉNESIS Y PATOLOGÍA
90
Como en el caso de la enfermedad de Crohn, otro aspecto muy interesante es la
hipotética influencia de la genética del hospedador en la resistencia a la
paratuberculosis, campo en el que ya se están dando los primeros pasos que servirán
como base para futuros estudios (Koets y cols. 2000, Mortensen y cols. 2004, Estonba y
cols. 2006, Gonda y cols. 2006). Tras observar diferencias significativas en la
distribución de un alelo del gen NRAMP1 entre animales seropositivos y seronegativos,
Estonba y cols. sugieren que éste puede estar envuelto en la evolución de las lesiones
(Estonba y cols. 2006).
Cuadro clínico
El cuadro clínico clásico de la paratuberculosis es una diarrea más o menos
intermitente según la especie, acompañada de consunción progresiva y muerte. Sin
embargo, estas formas representan un porcentaje relativamente bajo de la casuística de
la paratuberculosis, ya que la fase más prolongada es subclínica. Durante el estadio
subclínico, los síntomas que pueden llevar a su sospecha son principalmente pérdidas de
producción, que en bovino lechero como ya se ha mencionado puede variar de un 4
hasta un 14% de leche dependiendo de la lactación en que se encuentre (Benedictus y
cols. 1985b, Nordlund y cols. 1996, Wilson y cols. 1997). Otros síntomas relacionados
con paratuberculosis subclínica son problemas reproductivos y mastitis (Merkal y cols.
1975, Wilson y cols. 1997, Valentin-Weigand y cols. 1999). Otros autores señalan
además baja forma muscular de ciertas zonas o la aparición intermitente de un edema
mandibular como signos de enfermedad subclínica (Dunkin 1934). La mayoría de
animales infectados en una explotación son animales subclínicos que no muestran
signos evidentes de la enfermedad (aparte de los mencionados de producción,
infertilidad y mastitis, que pueden ser más o menos evidentes), aunque sí lesiones
microscópicas (Pérez y cols. 2000). En la especie bovina los síntomas clínicos no suelen
aparecer hasta los dos años de edad, coincidiendo con los partos y el inicio de las
lactaciones, y aunque en pequeños rumiantes pueden comenzar antes, su aparición
también está asociada a la introducción de estos estreses (Chiodini y cols. 1984a, Blood
y cols. 1989, García Marín y cols. 1994).
El signo clínico de paratuberculosis común a todas las especies es la pérdida
progresiva de peso y condición corporal (Taylor 1945, McEwen y cols. 1958, Jakob y
cols. 1969, Aller y cols. 1973, Newsholme y cols. 1981, Seaman y cols. 1984, Chiodini
y cols. 1984a, Gilmour y cols. 1991, García Marín y cols. 1994), aparentemente como
consecuencia de la insuficiente absorción de proteínas que causan los infiltrados
celulares y edemas del intestino (Allen y cols. 1974b). Este extremo no está claro pues
se ha podido observar que no hay pérdidas sustanciales de digestibilidad en ovinos con
formas clínicas (Juste y cols. 1994). La formación constante o intermitente de heces
diarreicas homogéneas, no hemorrágicas y no mucosas, es otro síntoma típico en vacas
con paratuberculosis clínica (Merkal y cols. 1970, Buergelt y cols. 1978b). No obstante,

REVISIÓN DE LA BIBLIOGRAFÍA
91
en las especies ovina y caprina sólo aparece en algunos casos en la fase final de la
enfermedad, cuando las lesiones afectan ampliamente el intestino, y muchas veces como
un sutil reblandecimiento de las heces (Stamp y cols. 1954, Brugère-Picoux 1987,
Carrigan y cols. 1990, García Marín y cols. 1994, Clarke y cols. 1996a). La caída de la
producción (principalmente en bovino lechero) es otro rasgo indicativo (Blood y cols.
1989, Aduriz y cols. 1995b, Clarke 1997b), así como el deterioro del pelo o la lana.
Otros posibles síntomas más avanzados son la formación de edemas submandibulares o
en otras zonas, empalidecimiento de las mucosas, fiebre intermitente, alteración de
algunos valores hemáticos y séricos, sangre en heces, anemia, caquexia extrema y
atrofia muscular (Behrens 1955, Kopecky y cols. 1971, Badiola y cols. 1980, Chiodini y
cols. 1984a, Sherman 1985, Scott y cols. 1995, Clarke 1997b). Finalmente el animal
queda postrado y muere (Pérez y cols. 2000).
Lesiones macroscópicas
La paratuberculosis provoca lesiones macroscópicas comunes a las que se producen
en otros procesos caquectizantes. En la necropsia de animales con avanzados signos
clínicos puede observarse una falta muy importante de depósitos grasos que pueden
mostrar atrofia de la serosa, atrofia de la masa muscular, formación de edemas, ascitis y
acumulación de líquidos en otras cavidades (Buergelt y cols. 1978b, Chiodini y cols.
1984a, Cranwell 1988, Gilmour y cols. 1991, Clarke y cols. 1996a). Más específicos de
la paratuberculosis son los fenómenos relacionados con el sistema digestivo,
principalmente las lesiones que afectan al íleon, al yeyuno, a la válvula ileocecal, a los
nódulos linfáticos mesentéricos, e incluso al ciego y a la primera parte del colon
(McEwen 1939, Stamp y cols. 1954, Prudvi Reddy y cols. 1984, Chiodini y cols. 1984a,
Van Kruiningen y cols. 1986, Carrigan y cols. 1990).
El engrosamiento de la pared intestinal es el rasgo característico de la enfermedad
en la especie bovina, principalmente en el íleon. Presenta unas mucosas edematosas,
tumefactas, con aspecto carnoso, formando gruesos pliegues, que pueden presentar
grietas, áreas congestivas, petequias o ulceraciones (Buergelt y cols. 1978b, Juste y cols.
1983, Chiodini y cols. 1984a, Jubb y cols. 1985, Blood y cols. 1989). Los vasos
linfáticos que conectan la serosa con los ganglios, pueden aparecer visiblemente
inflamados o dilatados, a modo de cordón retorcido con aspecto blanquecino (McEwen
1939, Buergelt y cols. 1978b, Badiola y cols. 1980, Chiodini y cols. 1984a, Blood y
cols. 1989, Carrigan y cols. 1990), aunque a menudo no sean tan evidentes (Juste y cols.
1983, Pérez y cols. 2000, González 2003). Los ganglios mesentéricos (los yeyunales e
ileocecales sobre todo) presentan normalmente un incremento en su tamaño,
tumefacción y aspecto edematoso, y en la sección de los mismos la zona medular
adquiere un color marrón verdoso. La aparición de lesiones en otras zonas es menos
frecuente. Pueden aparecer granulomas en el hígado y ganglios asociados, depleción

PATOGÉNESIS Y PATOLOGÍA
92
linfoide del timo, o lesiones arterioscleróticas en corazón o aorta (Clarke 1997b,
González 2003).
En pequeños rumiantes las lesiones intestinales pueden suponer desde pequeños
nódulos hasta áreas centrales con caseificación, el engrosamiento de la pared y los
pliegues suelen ser menos evidentes. En cambio, la tumefacción de la mucosa, la
linfangitis y linfangiectasia son normalmente muy marcadas, especialmente en cabras,
especie en la que puede observarse calcificación. Los nódulos linfáticos presentan un
aspecto similar al de los bovinos, pero aparecen caseificaciones y calcificaciones con
mayor frecuencia, sobre todo en cabras (Stamp y cols. 1954, Rajya y cols. 1961,
Badiola y cols. 1980, Seaman y cols. 1981, Lepper 1989, Carrigan y cols. 1990, García
Marín y cols. 1994, Pérez y cols. 1996, Clarke y cols. 1996a, Tafti y cols. 2000, Corpa y
cols. 2000a). En algunos casos ovinos el intestino puede aparecer coloreado de un tono
amarillento, debido a la presencia masiva de bacilos pertenecientes a cepas pigmentadas
de Map (Taylor 1945, Taylor 1951, Taylor 1953, Stamp y cols. 1954, Benazzi y cols.
1996, Clarke y cols. 1996a, Stevenson y cols. 2002).
Lesiones microscópicas
Se trata de una enteritis granulomatosa cuyas lesiones más evidentes se localizan en
los últimos tramos de intestino delgado, primeros del grueso, y nódulos y vasos
linfáticos asociados. La lesión característica es un infiltrado celular compuesto por
macrófagos, linfocitos, y células epiteloides y plasmáticas. Los macrófagos aparecen
normalmente llenos de bacilos, acompañados frecuentemente de células gigantes
también con numerosas bacterias en su interior (Buergelt y cols. 1978b, Badiola y cols.
1980, Chiodini y cols. 1984a, Jubb y cols. 1985).
Las formas lesionales muestran una gran variabilidad, de focales a difusas, y
dependen de factores como la dosis infectante, el estado inmunológico y la
susceptibilidad/resistencia del hospedador, y el tipo de cepa implicado (Chiodini y cols.
1984a, Aduriz 1993, Clarke 1997b). Debido a esa variabilidad, las lesiones han sido
clasificadas de acuerdo con la especie afectada, la gravedad de las mismas, y los tipos
celulares implicados. De este modo, las clasificaciones más completas serían:
- en bovino, las de Buergelt y cols. (Buergelt y cols. 1978b) y González y cols.
(González y cols. 2005),
- en ovino, las de Stamp y Watt (Stamp y cols. 1954), Rajya y Singh (Rajya y cols.
1961), García Marín y cols. (García Marín y cols. 1992c), Clarke y Little (Clarke
y cols. 1996a) y Pérez y cols. (Pérez y cols. 1996),
- en caprino, las de Paliwal y Rajya (Paliwal y cols. 1984), García Marín y cols.
(García Marín y cols. 1992b), Menchén (Menchén 1995) y Corpa y cols. (Corpa y
cols. 2000a).

REVISIÓN DE LA BIBLIOGRAFÍA
93
En este sentido, a partir de un primer estudio con corderos vacunados (Juste y cols.
1994), los trabajos que ha venido desarrollando el grupo de anatomía patológica
veterinaria de la Universidad de León (Pérez 1992, García Marín y cols. 1995a, Pérez y
cols. 1996, Pérez y cols. 1997, Corpa 1999, Corpa y cols. 2000a, González 2003,
González y cols. 2005, García-Pariente y cols. 2006, Muñoz y cols. 2006), proporcionan
un modelo inmunohistopatológico general y específico de hospedador de gran utilidad
en la evaluación de las lesiones naturales o experimentales de la paratuberculosis. Tal
como se ha mencionado anteriormente, éstas se clasifican en focales o multifocales
relacionadas con formas iniciales, latentes o subclínicas con baja carga micobacteriana,
y en difusas o severas con alta carga micobacteriana en los tejidos. También han
introducido el recuento de granulomas, la estimación de su área y su localización para
caracterizar la gravedad de las lesiones.
DIAGNÓSTICO
Actualmente, los métodos de diagnóstico más utilizados son aquellos basados en la
detección e identificación de Map y los basados en la detección de la respuesta
inmunológica originada en el hospedador. Los métodos clínico-epidemiológicos
resultan poco específicos, y principalmente son de utilidad para asentar la sospecha,
mientras que los anatomopatológico-histopatológicos se realizan mayoritariamente post
mortem (Garrido y cols. 2000a). La visualización de lesiones macroscópicas
relacionadas con la paratuberculosis y de los signos clínicos típicos de ésta, es un
método bastante específico que sirve para establecer un diagnóstico de rebaño, pero
puede ser insuficiente o simplemente impracticable para la detección de infecciones
subclínicas (Vialard y cols. 1994, Pérez y cols. 1996, Clarke y cols. 1996a, Corpa
1999).
Diagnóstico basado en las técnicas histopatológicas
convencionales
Las zonas donde más frecuentemente se asientan las lesiones son los últimos tramos
del intestino delgado, y primeros del grueso, nódulos linfáticos mesentéricos y vasos
linfáticos aferentes, en correspondencia con las alteraciones macroscópicas. Para
detectar las alteraciones microscópicas típicas de esta enfermedad granulomatosa, se
utilizan normalmente métodos rutinarios como la tinción hematoxilina-eosina, que
complementada con la tinción de Ziehl-Neelsen puede revelar la presencia de los
bacilos asociados a las lesiones (Garrido y cols. 2000a). El método de Ziehl-Neelsen no
es capaz de teñir las micobacterias con pared celular defectuosa o de esferoplasto. En
tales casos, es posible recurrir a técnicas de inmunohistoquímica u otras técnicas
(Massone y cols. 1990, Thoresen y cols. 1994a, Condron y cols. 1995, Balseiro y cols.
2003). Dada la especificidad de las características histológicas de las lesiones

DIAGNÓSTICO
94
paratuberculosas, este método también goza de una buena especificidad, y realizando
una buena selección de la muestra, también puede presentar una buena sensibilidad
(Jubb y cols. 1985, Whitlock y cols. 1992a, Clarke 1997a), incluso en animales con
paratuberculosis subclínica, y a veces superior a la del cultivo (Huchzermeyer y cols.
1992, Pérez y cols. 1996, Corpa y cols. 2000a, Dimareli-Malli y cols. 2001, González
2003). Alternativamente al análisis post mortem, ha sido propuesto como método de
diagnóstico el análisis histopatológico y bacteriológico de biopsias de ganglios o íleon
(Benedictus y cols. 1985a, Benedictus y cols. 1986, Coetsier y cols. 1998, Brumbaugh y
cols. 2000). No obstante, es de momento un método poco interesante fuera de su uso
experimental o con animales de gran valor (Garrido y cols. 2000a).
Diagnóstico basado en la detección bacteriológica
Tinción y detección al microscopio
Se realiza un frotis o impronta de heces o de cualquier tejido susceptible de contener
las micobacterias, o bien se utilizan preparaciones de cortes histológicos.
Se tiñe por medio de la técnica de Ziehl-Neelsen que se basa en la retención de la
fucsina (colorante) por las bacterias ácido-alcohol resistentes, en las que a diferencia de
otras bacterias, el decolorante (ácido-alcohol) no penetra, y se visualiza en el
microscopio, pudiéndose observar la morfología típica de las células micobacterianas
Figura 12. Fotografía de la vista al microscopio (x 1000) de la tinción Ziehl-Neelsen de una impronta intestinal ovina en la que se pueden apreciar claramente los bacilos de Map (fucsia) formando grumos.

REVISIÓN DE LA BIBLIOGRAFÍA
95
(Garrido y cols. 2000a) (Figura 12). Es útil, rápido y barato, pero tiene una baja
sensibilidad y no es específico de Map.
Análisis inmunohistoquímico de preparaciones histológicas
Es una técnica basada en la unión específica entre anticuerpos marcados y los
antígenos que estos reconocen, y se aplica sobre cortes histológicos. En el caso de la
paratuberculosis, se utilizan anticuerpos monoclonales o policlonales frente a Map, que
generalmente se obtienen de forma artesanal y que pueden dar reacciones cruzadas en
caso de infección con otras micobacterias (Stabel y cols. 1996, Coetsier y cols. 1998,
Brees y cols. 2000, González 2003). Aun así es más específica que la tinción Ziehl-
Neelsen, y es especialmente útil en la detección de esferoplastos o cuando la presencia
de bacilos es mínima (Massone y cols. 1990, Navarro y cols. 1991, García Marín y cols.
1992a, Thoresen y cols. 1994a, Brees y cols. 2000, Balseiro y cols. 2003, González
2003).
Aislamiento
Se considera como la técnica de referencia para el diagnóstico de la
paratuberculosis. El primer aislamiento de Map se consiguió en 1911 (Twort y cols.
1911). Desde entonces se ha tratado de mejorar la técnica del cultivo, con el objetivo de
hacerlo más rápido, eficaz y sensible. Igual que en la tinción, las muestras patológicas
normalmente utilizadas para diagnóstico son heces, ganglios mesentéricos o raspados de
mucosa intestinal (Garrido y cols. 2000a), y a veces, leche. Cuando se trata de muestras
intestinales obtenidas en la necropsia de un animal, algunos autores obtienen mejores
resultados con raspados de válvula ileocecal y ganglios ileocecales o yeyunales, que con
otras localizaciones (Chiodini y cols. 1986a, Merkal y cols. 1987), pero también puede
influir la especie del animal. El cultivo tradicional puede llegar a un límite de detección
de 50-100 células de Map por gramo de heces y se considera 100% específico (Merkal
1973, Jorgensen 1982, Whitlock 1998). Aunque la sensibilidad del método varía mucho,
esta suele ser relativamente baja, sobre todo por falsos negativos con animales
subclínicamente infectados, que se ve aún más reducida al emplear menos de cuatro
tubos de cultivo o por la aparición de cepas de difícil aislamiento (Merkal 1973,
Chiodini y cols. 1984a, Chiodini y cols. 1986a, Juste y cols. 1991b, Huchzermeyer y
cols. 1992, Benazzi y cols. 1996, Whitlock 1998, Whittington y cols. 1999b, Eamens y
cols. 2000, Whitlock y cols. 2000, Whittington y cols. 2000a, Corpa y cols. 2000a,
Whittington y cols. 2000c, Whittington y cols. 2001d, Crossley y cols. 2005). Además
es un método caro, lento y laborioso (Collins y cols. 1990b, Kalis y cols. 2002).
Generalmente el cultivo requiere un tratamiento previo de la muestra para evitar su
contaminación por microorganismos que crecen más rápido que Map. Han sido varios
los tratamientos probados con Map aprovechando su elevada resistencia a ciertos
agentes descontaminantes. Se han utilizado distintos compuestos corrosivos (Merkal y

DIAGNÓSTICO
96
cols. 1964), cloruro de benzalconio (Merkal y cols. 1964, Merkal y cols. 1972, Lagadic
y cols. 1983), ácido oxálico (Brotherston y cols. 1961a, Karpinski 1972, Gunnarsson y
cols. 1979, Saxegaard 1985), ácido oxálico con hidróxido sódico (Jorgensen 1982), y
finalmente el más extendido de todos, el cloruro de hexadecil piridinio (HPC) (Whipple
y cols. 1983, Merkal 1984, Juste y cols. 1991b, Whitlock y cols. 1992a, Stabel 1997). A
pesar de que el HPC es el agente utilizado prioritariamente, la concentración de uso, y el
tiempo y condiciones térmicas de incubación empleadas pueden variar sustancialmente,
hecho que puede influir en el éxito del cultivo de determinado tipo de muestra o cepa
(Whitlock y cols. 1992a, Stabel 1997, Pavlik y cols. 1999a, Whitlock y cols. 2000,
Dundee y cols. 2001, Stabel y cols. 2002b, Whitlock y cols. 2003, Reddacliff y cols.
2003a, Donaghy y cols. 2003a, Grant y cols. 2004, Ayele y cols. 2005). En algunos
métodos, la muestra se incuba adicionalmente con infusión de cerebro y corazón (BHI)
para favorecer la germinación de estructuras resistentes (esporas,...) haciéndolas
susceptibles al HPC (Whitlock 1992). En otros protocolos tras realizar una
concentración de las micobacterias por centrifugación, paso que al mismo tiempo parece
incrementar el número de contaminaciones (Whipple y cols. 1991, Aduriz y cols. 1995a,
Eamens y cols. 2000), tratan la muestra con antibióticos antes de ser cultivada (Stabel
1997, Whittington y cols. 1999b, Whitlock y cols. 2000). Los métodos principales de
concentrar las micobacterias que se hallan en la muestra son la sedimentación, la
centrifugación (Kim y cols. 1988, Whitlock y cols. 1992a) y la captura
inmunomagnética (Grant y cols. 1998b). Para los medios sin base de huevo es necesario
centrifugar y lavar con agua la muestra una vez descontaminada para eliminar el HPC,
ya que éste no es neutralizado por los fosfolípidos del huevo, y sigue actuando
inhibiendo la propagación de Map en el medio (Garrido y cols. 2000a).
Los medios más ampliamente utilizados para el aislamiento de Map son el HEY
(Merkal y cols. 1964) y el LJ (Jorgensen 1982) adicionados con micobactina J, y que
están hechos a base de huevo. El medio de Dubos a base de suero y con micobactina es
otra alternativa (Gunnarsson y cols. 1979, Saxegaard 1985). Los medios sintéticos
sólidos de Middlebrook (7H10 y 7H11) también dan buenos resultados, ya sea con
(Merkal y cols. 1974) o sin micobactina (Aduriz y cols. 1995a). Para el cultivo en medio
líquido han sido utilizadas varias versiones de Middlebrook (7H9, 7H10, 7H12)
(Damato y cols. 1982, Merkal y cols. 1982a, Damato y cols. 1984, Damato y cols.
1987), y a veces el medio Watson-Reid. En los casos positivos, Map puede tardar entre
3 semanas y un año en crecer de manera visible en los medios sólidos (Taylor 1951,
Merkal y cols. 1974, Wayne y cols. 1986, Whittington y cols. 2001d). Los sistemas
automáticos de cultivo líquido que detectan variaciones en la concentración de ciertas
sustancias (dióxido de carbono, oxígeno,...) como resultado de la respiración de las
micobacterias, que en un principio fueron diseñados para aislar las especies que infectan
al ser humano (M. tuberculosis principalmente), están basados en el caldo Middlebrook
7H9. La composición de los mismos ha ido modificándose para cubrir los

REVISIÓN DE LA BIBLIOGRAFÍA
97
requerimientos especiales de Map (Whittington y cols. 1998, Whittington y cols.
1999b). Normalmente todos estos medios líquidos son suplementados con micobactina,
y enriquecidos con albúmina sérica bovina, ácido oleico, dextrosa, catalasa y otros
ingredientes como yema de huevo y antibióticos. Los de mayor renombre son los
radiométricos BACTEC (Damato y cols. 1982, Damato y cols. 1990, Collins y cols.
1990b), y los no radiométricos MGIT (fluorométrico) (Schwartz y cols. 2000, Bull y
cols. 2003a), TREK ESP (barométrico) (Kim y cols. 1988, Ellingson y cols. 2004), y
MB/BacT (colorimétrico) (Stich y cols. 2004). La ventaja de estos sistemas es que
detectan más rápido y de manera automatizada el crecimiento micobacteriano, además
de ser teóricamente más sensibles (Collins y cols. 1988, Damato y cols. 1990, Collins y
cols. 1990b, Cousins y cols. 1995, Whittington y cols. 1999b, Eamens y cols. 2001,
Grant y cols. 2003, Gumber y cols. 2006), sin embargo, tienen varios inconvenientes.
Por una parte, suelen contaminarse con mayor frecuencia (Collins y cols. 1990b,
Whittington y cols. 1999b), y para evitarlo se utilizan antibióticos que al final pueden
retrasar el crecimiento de las propias micobacterias (Gumber y cols. 2006). Son aún más
caros que los tradicionales, y requieren un mayor equipamiento (Whitlock 1998). No
resultan tan específicos y normalmente se necesita confirmar la identidad del
crecimiento por métodos moleculares (Cousins y cols. 1995, Whittington y cols. 1998).
Para demostrar la independencia de micobactina es necesario realizar un subcultivo en
un medio sólido sin este ingrediente, y algunos autores repiten esta operación desde el
primer medio sólido a un segundo tubo para confirmar que no se arrastre el sideróforo
del medio líquido al sólido (Eamens y cols. 2000, Motiwala y cols. 2004). Y por último,
señalar que no se obtienen colonias puras por lo que es necesario su subcultivo en
medios sólidos si se precisa de organismos clónicos para estudios posteriores.
Con el fin de evadir los costos económicos y de procesado del cultivo individual,
algunos investigadores han optado por realizar cultivos por lotes (incluyendo muestras
de distintos animales en un mismo cultivo), que aunque menos sensible, es una
herramienta útil para detectar rebaños afectados. De todos modos, la necesidad de
realizar el cultivo individual de todos los componentes de los lotes con resultado
positivo, y la influencia del número de animales del rebaño y de la prevalencia del
mismo en la sensibilidad, son aspectos que limitan su uso (Vialard y cols. 1993, Kalis y
cols. 1999a, Kalis y cols. 2000, Whittington y cols. 2000a, Wells y cols. 2002, Sergeant
y cols. 2002b, Van Schaik y cols. 2003b).
Desde los inicios de los métodos de cultivo de la paratuberculosis, se han detectado
diferencias importantes en el éxito de aislamiento de Map. Normalmente, las muestras
bovinas producen colonias más rápido, que crecen mejor en HEY con micobactina y
piruvato que en cualquiera de los restantes medios sólidos (Merkal y cols. 1964,
Chiodini y cols. 1984a, Juste y cols. 1991b, Nielsen y cols. 2004). Sin embargo, tal y
como refleja la bibliografía, las muestras procedentes de pequeños rumiantes resultan
más problemáticas, especialmente las ovinas. Las colonias suelen tardar más en

DIAGNÓSTICO
98
aparecer, e incluso se habla de cepas incultivables (Dunkin y cols. 1935, Taylor 1951,
Brotherston y cols. 1961a, McCausland 1980, Seaman y cols. 1981, Juste y cols. 1991b,
Huchzermeyer y cols. 1992, Shulaw y cols. 1993, Benazzi y cols. 1996, Dimareli-Malli
y cols. 2001, Machackova y cols. 2003a). Juste y cols. obtenían mejores resultados
utilizando LJ con micobactina pero sin piruvato, q con cualquiera otra combinación de
los medios HEY y LJ (Juste y cols. 1991b), lo que alimentaba la idea de una posible
diferencia entre hospedadores, cepas y preferencias. Algunos autores no se muestran de
acuerdo con estas observaciones (Pavlik y cols. 1999a) (materia revisada en el apartado
de Epidemiología molecular), pero estudios más recientes parecen confirmarlo (de
Juan y cols. 2006b). En un estudio posterior del grupo de Neiker, quedó demostrada la
eficacia del medio Middlebrook 7H11 con OADC y sin micobactina para el aislamiento
de Map a partir de muestras ovinas (Aduriz y cols. 1995a). En la Figura 13 se muestra
el aspecto de colonias de tipo bovino y de colonias pigmentadas ovinas sobre la
superficie de diferentes medios sólidos.
Detección de DNA
La detección de DNA específico de un microorganismo implica su presencia en la
muestra que se está analizando. Existen otras técnicas como los diferentes sistemas de
hibridación (algunos, como la hibridación in situ, muy utilizados en los casos de Crohn)
(Hulten y cols. 2001, Chamberlin y cols. 2001, Harris y cols. 2001), pero la técnica
reina en este campo es la reacción en cadena de la polimerasa o PCR (Mullis y cols.
1986), que permite amplificar in vitro y de manera específica cantidades mínimas de
DNA gracias a la acción de la DNA polimerasa, haciendo que puedan ser detectadas
con facilidad. Su especificidad diagnóstica depende de que el fragmento de DNA
escogido sea específico del microorganismo en cuestión, y de la especificidad de los
cebadores (fragmentos de unas 18-24 pares de bases de DNA que hibridan con el DNA
molde o diana y funcionan como iniciadores de la amplificación) por ése fragmento
concreto. La selección de la muestra, el proceso para concentrar los organismos que se
Figura 13. A la izquierda, fotografía del crecimiento de una cepa ovina pigmentada en medio LJ, que fue aislada de los intestinos amarillentos (debido al color del aislamiento) de una oveja procedente del País Vasco. A la derecha, dos tubos HEY (el de arriba más viejo que el de abajo) con colonias típicas bovinas.

REVISIÓN DE LA BIBLIOGRAFÍA
99
quieren detectar, y la extracción y purificación del DNA, condicionarán la cantidad de
DNA diana obtenida, y por lo tanto la sensibilidad de la técnica.
El descubrimiento de la secuencia de inserción específica de Map IS900 (Green y
cols. 1989, Collins y cols. 1989) abrió todo un horizonte de posibilidades para la
detección de esta bacteria por métodos genéticos. Gracias a este elemento genético la
PCR puede ser una técnica de gran sensibilidad, ya que cada bacilo de Map cuenta con
14-18 copias del mismo (Bull y cols. 2000), lo que multiplica las posibilidades de
detección en comparación con genes o secuencias con una única copia por genoma
bacteriano. A pesar de que se cuenta con otras técnicas, la PCR es el método molecular
más utilizado en el diagnóstico de la paratuberculosis (Vary y cols. 1990, Moss y cols.
1992b, Sanderson y cols. 1992b). La principal ventaja que confiere la PCR sobre el
cultivo es la velocidad en la obtención de resultados (Garrido y cols. 2000a), además es
capaz de detectar la presencia de la bacteria aun cuando no es viable en cultivo. Sin
embargo difícilmente puede suplantarlo, ya que la sensibilidad del cultivo parece
todavía mayor (Whitlock 1998), y que la presencia de inhibidores de la PCR sobre todo
en muestras de heces producen falsos negativos (Collins y cols. 1993a, Harris y cols.
2001). Estos problemas van siendo solventados con la introducción de controles
internos de amplificación que garantizan el buen funcionamiento de la reacción sobre la
muestra descartando falsos negativos (Englund y cols. 1999, Chevallier y cols. 2003,
Ozbek y cols. 2003, Rodriguez-Lazaro y cols. 2004, Bartos y cols. 2006, Brey y cols.
2006), además de mejorar los protocolos de extracción y purificación del DNA para
evitar la copurificación de esas sustancias inhibidoras. La sensibilidad de la PCR
aumenta con la realización de una segunda PCR que amplifica una región interna del
producto de la primera reacción, lo que se conoce como PCR anidada (Bascuñana y
cols. 1996, Frank y cols. 1996, Englund y cols. 1999, Gao y cols. 2002, Bull y cols.
2003a, Buergelt y cols. 2004, Stabel y cols. 2005, Romero y cols. 2005).
No hay que olvidar que existen otras bacterias distintas de Map con secuencias
parecidas (IS900-like) que pueden dar amplificados que se toman por positivos en la
PCR IS900, aunque son distinguibles por secuenciación (Cousins y cols. 1999, Englund
y cols. 2002, Motiwala y cols. 2004). En casos de aislados humanos de M. avium no ha
quedado claro si se trataba de secuencias parecidas, o de si realmente se trataba de la
IS900 (Roiz y cols. 1995, Naser y cols. 1999). Se ha buscado y actualmente se sigue
buscando regiones o secuencias genéticas que sean específicas de la micobacteria y que
puedan suplir a la IS900, o al menos complementarla. Han sido descritas unas cuántas
secuencias alternativas (revisado en Map. Composición genética), pero de momento
ninguna goza de la sensibilidad que proporciona la IS900. En caso de identificación en
cultivos, esto no supone un problema pues el DNA micobacteriano es abundante, o al
menos suficiente. Pero en muestras patológicas el número de micobacterias puede ser
mínimo, y el número de copias por genoma de la secuencia diana puede ser
determinante a la hora de dar un diagnóstico (van der Giessen y cols. 1992b). En

DIAGNÓSTICO
100
cultivos, se consigue una identificación correcta confirmando la presencia de la IS900 y
del locus 251 (Rajeev y cols. 2005). La alternativa con mayores posibilidades para
detectar DNA de Map en otro tipo de muestras es complementar la PCR IS900 con la
PCR ISMap02 (Stabel y cols. 2005), o incluso con la PCR-REA IS1311 (Marsh y cols.
1999).
Además de los aislamientos, las muestras que se suelen analizar por medio de la
PCR, son las heces, tejidos y leche. La PCR de heces no ha conseguido reemplazar al
cultivo fecal, a pesar de ser muy eficiente para algunos autores, principalmente en la
especie ovina por la dificultad que entraña el cultivo a partir de muestras ovinas (Collins
y cols. 1993a, Pérez y cols. 1995a, Stevenson y cols. 1997, Garrido y cols. 2000a,
Garrido y cols. 2000b, Sivakumar y cols. 2005). La sensibilidad en heces varía mucho
debido a la heterogeneidad de técnicas que se usan en diferentes laboratorios (Garrido y
cols. 2000a). Lo que sí parece conferir rapidez y solidez al diagnóstico es el uso del
cultivo y posterior análisis por PCR, antes de que haya crecimiento visible o
confirmando los tempranos cultivos positivos de los sistemas automáticos de detección
(Cousins y cols. 1995, Whittington y cols. 1998, Secott y cols. 1999, Paolicchi y cols.
2003). La leche parece también una buena muestra para la detección de DNA de Map,
aunque parece más interesante para ser utilizada en muestras agrupadas o de tipo lote,
como la leche de tanque, con la aplicación potencial de estimar prevalencias de
explotaciones o realizar estudios de vigilancia (Stephan y cols. 2002, Corti y cols. 2002,
Grant y cols. 2002a, Sevilla y cols. 2003, Muehlherr y cols. 2003, O'Reilly y cols. 2004,
Jayarao y cols. 2004). En el análisis de muestras individuales el cultivo parece seguir
siendo más eficaz (Giese y cols. 2000). En algunos trabajos de PCR IS900 se habla de
límites de detección de hasta 1-100 CFU/g heces, o de 10-20 CFU/ml leche (Grant y
cols. 2000, Garrido y cols. 2000b, Odumeru y cols. 2001, Halldorsdottir y cols. 2002,
Stabel y cols. 2004a). En caso de utilizar tejidos, las muestras más interesantes son las
que mayor número de micobacterias puedan contener, como la válvula ileocecal,
porciones de íleon y yeyuno, y los ganglios asociados, como en el caso del cultivo. El
uso de estas muestras en comparación con las heces mejora la sensibilidad de la prueba,
además de haber menos riesgo de inhibiciones (Challans y cols. 1994, Gwozdz y cols.
1997, Stevenson y cols. 1997), aunque cuando se usan biopsias de hígado no se obtiene
el mismo resultado.
Otra muestra posible para PCR sobre la que existen pocos trabajos publicados es la
sangre, más concretamente las células blancas obtenidas de sangre periférica, fracción
de la sangre donde cabe esperar encontrar las micobacterias con mayor facilidad (van
der Giessen y cols. 1995, Hanna y cols. 1995) (y según lo revisado en el apartado de
Patogénesis). Esta es una muestra fácil de tomar, y que se recoge habitualmente para
llevar a cabo pruebas serológicas, lo que la hace muy interesante. La sensibilidad de la
PCR en sangre de ovejas clínicamente afectadas puede ser del 66% con una
especificidad del 100% (Gwozdz y cols. 1997), y del 40% en vacas subclínicamente

REVISIÓN DE LA BIBLIOGRAFÍA
101
afectadas (Barrington y cols. 2003). Es un método con grandes posibilidades, que
incluso puede servir para un primer tipado de las cepas infectantes que se detectan
(Bhide y cols. 2006), a pesar de los resultados contradictorios de Gwozdz y cols.
(Gwozdz y cols. 2000c). Además la PCR de sangre puede servir de método precoz de
detección de animales subclínicos o expuestos antes de que comiencen a excretar
bacterias en sus heces y supongan un foco de infección para el resto de animales del
rebaño (Buergelt y cols. 2004). En la especie humana también ha sido escasamente
empleado este método, aunque con resultados de gran importancia y calado en relación
con la etiología de la enfermedad de Crohn (Naser y cols. 2004).
Otra manera de analizar tejidos, es realizar la PCR a partir de tejidos fijados e
incluidos en parafina preparados para estudios histológicos, cuya sensibilidad está
limitada entre otras cosas por el efecto degradante de la fijación con formol sobre el
DNA (Plante y cols. 1996). Esto permite el análisis de tejidos ya testados por otras
técnicas (tinción hematoxilina-eosina + Ziehl-Neelsen, inmunohistoquímica,...) a modo
de confirmación, o realizar estudios de carácter retrospectivo, incluso utilizando
cebadores para diferentes dianas, posterior hibridación, o enzimas de restricción que
diferencian especies y subespecies (Plante y cols. 1996, Miller y cols. 1999, Marsh y
cols. 1999, Whittington y cols. 1999d, Coetsier y cols. 2000, Ikonomopoulos y cols.
2000, Sanna y cols. 2000, Miller y cols. 2002, Ellingson y cols. 2005b). En los trabajos
con muestras de pacientes de Crohn, la PCR IS900 de tejidos fijados e incluidos en
parafina, junto con la hibridación in situ con porciones de IS900, son las técnicas
moleculares más extendidas (Fidler y cols. 1994, Lisby y cols. 1994, Hulten y cols.
2000b, Sechi y cols. 2001, Hulten y cols. 2001, Naser y cols. 2002, Ellingson y cols.
2003, Sechi y cols. 2004, Baksh y cols. 2004) para detectar DNA de Map. Cabe destacar
que en la técnica de hibridación in situ, la sonda IS900 marcada es capaz de penetrar en
los esferoplastos o formas de pared defectuosa de esta bacteria e hibridar con su DNA
haciéndolas visibles, pero no detecta los bacilos ácido-alcohol resistentes (Hulten y cols.
2000b, Hulten y cols. 2000c, Hulten y cols. 2001).
Los métodos de extracción y purificación de DNA cada vez son más sofisticados,
aunque parte del éxito depende de los métodos de concentración de las bacterias de la
muestra que suelen ser similares a los empleados en el cultivo (Garrido y cols. 2000a).
Entre los métodos de concentración micobacteriana destacan los que utilizan la
centrifugación (Vary y cols. 1990, Garrido y cols. 2000b, Odumeru y cols. 2001,
Stephan y cols. 2002, Corti y cols. 2002, Halldorsdottir y cols. 2002, Ozbek y cols.
2003), la separación inmunomagnética (Grant y cols. 2000, Mason y cols. 2001, Djonne
y cols. 2003, O'Reilly y cols. 2004, Khare y cols. 2004, Whan y cols. 2005a, Whan y
cols. 2005b, Metzger-Boddien y cols. 2006), el procesado C18-carboxipropilbetaina
(Thornton y cols. 2002, Ozbek y cols. 2003), y la separación magnética mediada por un
péptido (Stratmann y cols. 2002, Stratmann y cols. 2006).

DIAGNÓSTICO
102
Los problemas iniciales para extraer el DNA de las resistentes estructuras celulares
micobacterianas se superaron con el uso de métodos más agresivos, por ejemplo, las
perlas de zirconio (van der Giessen y cols. 1992a, Collins y cols. 1993b, Challans y
cols. 1994, Odumeru y cols. 2001), tratamiento con tampones de lisis (Marsh y cols.
1999, Odumeru y cols. 2001, Chevallier y cols. 2003), el hervido (Whittington y cols.
1998, Marsh y cols. 1999, Whittington y cols. 1999d, Odumeru y cols. 2001, Chevallier
y cols. 2003, O'Mahony y cols. 2004) y la congelación-hervido (Miller y cols. 1997,
Garrido y cols. 2000b, Odumeru y cols. 2001).
En la purificación de DNA se han probado varios métodos que incluyen el uso de
otros reactivos u otros sistemas de captura de DNA. Por ejemplo, sustancias como el
dodecil-sulfato de sodio (SDS), el TE-Triton X100 (Garrido y cols. 2000b, Garrido
2001), el bromuro de cetil-trimetil-amonio (CTAB) (Pavlik y cols. 1999a), ácido etilen-
diamin-tetra-acético (Christopher-Hennings y cols. 2003) o el isotiocianato de guanidina
(Whittington y cols. 1998, Garrido y cols. 2000b). Para captar el DNA extraído se han
empleado métodos como la dispersión en matriz con fase sólida (MSPD; que también
consta de un paso de lisado mecánico de células) (Hines y cols. 1995), la captura por
hibridación en fase sólida y/o líquida (Millar y cols. 1995, Marsh y cols. 2000, Marsh y
cols. 2001), columnas de sílice (Whittington y cols. 1998, Chevallier y cols. 2003),
otros sistemas magnéticos patentados (Chui y cols. 2004), filtros que retienen el DNA
(Blanchard y cols. 2005), o las tarjetas de tecnología FTA® de Whatman (Jaravata y
cols. 2006). Incluso se ha probado la adición de sustancias como la albúmina sérica
bovina (BSA) (Odumeru y cols. 2001), el dimetil sulfóxido (DMSO) (Bull y cols.
2003a) o el mercaptoetanol (Marsh y cols. 1999) a la reacción de PCR.
La PCR a tiempo real está siendo ampliamente introducida en la detección de Map,
con la ventaja que supone la cuantificación del DNA, o la amplificación del cDNA
complementario al RNA que se halla en células vivas por medio de la RT-PCR. Son
abundantes las publicaciones al respecto, y aunque todavía no parecen mostrar
sensibilidades mucho mayores que las PCRs tradicionales (Kim y cols. 1989, Kim y
cols. 2002, Fang y cols. 2002, Christopher-Hennings y cols. 2003, Khare y cols. 2004,
Rajeev y cols. 2005, Stabel y cols. 2005, Bogli-Stuber y cols. 2005, Brey y cols. 2006,
Jaravata y cols. 2006), deberá transcurrir cierto tiempo mientras se obtienen los
primeros resultados de diagnóstico a gran escala, con lo que se podrá determinar qué
métodos son los más apropiados. Aún así, algunos ya le atribuyen una mayor
sensibilidad que al cultivo (Bogli-Stuber y cols. 2005).
Diagnóstico basado en la respuesta inmunológica del
hospedador
Las pruebas diagnósticas de inmunidad tratan de detectar la respuesta inmunológica
que se produce en el hospedador a consecuencia de la infección. La infección con Map

REVISIÓN DE LA BIBLIOGRAFÍA
103
provoca respuestas inmunes de tipo espectral, empezando con una de tipo celular que
evoluciona hacia una de tipo humoral, donde el tiempo de transición de una a otra es
variable, aunque suele ser bastante largo. Por ello, existen pruebas enfocadas a la
detección de la infección en sus fases iniciales donde predomina la respuesta inmune
celular, y pruebas enfocadas a la detección de infecciones más avanzadas que muestran
un respuesta fuerte de tipo humoral.
Detección de la inmunidad celular in vivo: Intradermorreacción
Se basa en la reacción inflamatoria local que se produce al inyectar
intradérmicamente un antígeno en el animal infectado, como respuesta retardada de
hipersensibilidad (Chiodini y cols. 1984a). Es similar a la prueba que se realiza para
detectar la tuberculosis bovina (Real Decreto 1047/2003), con la diferencia de que los
extractos proteicos purificados (PPD) que se utilizan a modo de antígeno, son la johnina
o la PPD (o tuberculina) aviar, derivados de Map o de M. avium respectivamente
(Johnson y cols. 1949, Stuart 1962, Collins 1996). El uso de la tuberculina aviar está
más extendido ya que los resultados son muy similares a los de la johnina pero su
producción es más sencilla (Juste y cols. 1990a). También se ha probado el uso del
complejo A36 de Map con buenos resultados (Gilot y cols. 1992, De Kesel y cols.
1992). El punto de inoculación recomendado en las especies bovina y caprina es la zona
de las tablas del cuello, y en la ovina el pliegue de la cola, y el resultado se suele
chequear a las 48 horas (Chiodini y cols. 1984a, Blood y cols. 1989, Juste y cols.
1990a). La sensibilidad de la prueba es bastante baja, y debido a las reacciones cruzadas
que surgen con otras micobacterias y otros microorganismos, también se la considera de
baja especificidad (Larsen y cols. 1965, Larsen 1972, de Lisle y cols. 1980, Chiodini y
cols. 1984a, Blood y cols. 1989). Esta prueba puede dar falsos positivos en animales
vacunados de paratuberculosis (Blood y cols. 1989). Sin embargo, hasta el momento los
problemas más significativos son los derivados de los falsos positivos a la
intradermorreacción simple con tuberculina bovina (tuberculosis bovina, M. bovis) por
animales infectados con Map o vacunados de paratuberculosis, interferencias que tratan
de evitarse por medio de la intradermorreacción comparada (inoculando al animal con
ambas tuberculinas, la aviar y la bovina) (Chiodini y cols. 1984a, Blood y cols. 1989,
Kohler y cols. 2001, Balseiro 2004). Aunque también han sido propuestos otros medios,
como la prueba del interferón-γ y la prueba de aglutinación con bolas de látex (Balseiro
2004, Koo y cols. 2004).
Detección de la inmunidad celular in vitro
Algunas de las pruebas como la de la transformación linfocitaria o la de la inhibición
de la migración leucocitaria están en desuso (Garrido y cols. 2000a), y otras como la del
receptor de IL-2 (Whist y cols. 2000) prácticamente no se utilizan, y no se revisan en
este trabajo.

DIAGNÓSTICO
104
• Prueba del IFN-γ
En este caso la técnica consiste en la detección de la citoquina IFN-� que liberan
los linfocitos T sensibilizados al ser estimulados con un antígeno, por medio de un
ELISA con anticuerpo primario monoclonal específico. Como siempre, en el caso de la
paratuberculosis los antígenos de elección son PPDs, que parecen dar mejores
resultados que los sonicados celulares crudos (Robbe-Austerman y cols. 2006c), al
menos en ovino. La prueba fue desarrollada primero para la detección de infecciones
subclínicas de tuberculosis bovina (Rothel y cols. 1990, Wood y cols. 1990a, Wood y
cols. 1990b), y fue modificada para su uso en el diagnóstico de la paratuberculosis
(Billman-Jacobe y cols. 1992). La cantidad de IFN-� que producen los linfocitos T
varía de acuerdo con el tipo de respuesta y con el estadio de la infección, siendo más
alta en respuestas iniciales de tipo celular en animales subclínicamente afectados (Wood
y cols. 1990b, Pérez y cols. 1995b, Stabel 1996, Sweeney y cols. 1998, McDonald y
cols. 1999, Corpa 1999, García Marín y cols. 1999a, Stabel 2000a, Stabel 2000c, Stabel
y cols. 2001b, Jungersen y cols. 2002, Huda y cols. 2003b), o en infecciones más
avanzadas pero de tipo tuberculoide o focal (Pérez y cols. 1999, Burrells y cols. 1999,
Corpa 1999, González 2003, Huda y cols. 2003b). En general, al contrario que en
animales con infección clínica avanzada, a la hora de revelar la manifestación de la
respuesta celular, la sensibilidad de la técnica en relación con el cultivo o las
observaciones patológicas, es bastante alta (75-100%) (Billman-Jacobe y cols. 1992,
Pérez y cols. 1999, Corpa 1999, González 2003). En otros trabajos donde la técnica es
comparada con el cultivo, la sensibilidad es más baja (Stabel y cols. 2001b). La
especificidad puede ser elevada, sin embargo, puede verse reducida por las
interferencias que provocan las reacciones cruzadas con otros microorganismos más o
menos relacionados (Billman-Jacobe y cols. 1992, Pérez y cols. 1999, Gwozdz y cols.
2000b, Gwozdz y cols. 2002, Jungersen y cols. 2002, Kalis y cols. 2003a). Existen otros
factores como la temperatura o el tiempo que pasa antes de realizar la prueba, que
afectan a la viabilidad de los linfocitos, problema que se pretende controlar con el uso
de la concanavalina (Stabel y cols. 2001b) o la IL-12 (Jungersen y cols. 2006). Otros
opinan que de todos modos la sangre debe estar a temperatura ambiente y ser estimulada
dentro de las 12 horas después de su recolección para que los resultados sean fiables
(Robbe-Austerman y cols. 2006b). Como conclusiones de los estudios de IFN-γ con
muestreos repetidos de los mismos animales, en los que esta prueba se compara con el
cultivo y las pruebas de inmunidad humoral, Huda y cols. recomiendan el uso de este
test en animales de 1-2 años, y el ELISA para los de 3 años (Huda y cols. 2003a, Huda y
cols. 2004). No obstante, la variabilidad en los resultados que proporcionan estas
pruebas inmunológicas también ha quedado patente (Huda y cols. 2003a), lo que da pie
a otros investigadores para pensar que actualmente la prueba del IFN-γ continúa siendo
una promesa de análisis válido más que una realidad (Collins 2006). En estudios
recientes se han probado otros antígenos de Map (Olsen y cols. 2000, Olsen y cols.

REVISIÓN DE LA BIBLIOGRAFÍA
105
2001a, Vesosky y cols. 2004, Olsen y cols. 2005, Rosseels y cols. 2006), lo que ha
demostrado entre otras cosas la posible interferencia de la inmunidad innata de células
NK que inducen algunas proteínas excretadas de Map en las pruebas diagnósticas de
IFN-γ (Olsen y cols. 2001a, Olsen y cols. 2005).
Detección de la inmunidad humoral
Se trata de revelar la presencia de los anticuerpos que genera el sistema inmune del
hospedador infectado. En la paratuberculosis actualmente se utilizan dos tipos de prueba
de inmunidad humoral, la AGID y el ELISA, aunque esta última es la más extendida
(Harris y cols. 2001). La fijación del complemento (CF) fue una de las técnicas más
utilizadas hasta los años 90, pero actualmente no se emplea (Garrido y cols. 2000a) y no
será revisada. Son técnicas rápidas, baratas, fáciles de ejecutar sin mucho equipamiento,
automatizables para su uso a gran escala (ELISA), y la toma de muestras no entraña
dificultades (Juste y cols. 1990a, Garrido y cols. 2000a). La respuesta humoral suele ser
fuerte y fácil de detectar en los estadios avanzados de paratuberculosis, por la que la
sensibilidad de estas pruebas alcanza su máximo en animales con lesiones lepromatosas,
con síntomas clínicos o que excretan grandes cantidades de la bacteria en sus heces. En
cambio, son incapaces de identificar con precisión animales en fases iniciales de la
infección, aunque por lo general son bastante específicas (Harris y cols. 2001).
• Prueba de AGID
En un gel de agarosa se enfrenta el suero problema con un antígeno específico.
Éstos difunden por el gel, y en caso de haber anticuerpos para ese antígeno, el complejo
antígeno-anticuerpo precipita haciéndose visible. El antígeno utilizado normalmente
para la AGID de paratuberculosis es el de la proteína protoplasmática o PPA3 de la cepa
Strain 18 (St18) (Juste y cols. 1990a), cepa de M. avium que fue identificada en un
principio como Map (Chiodini 1993). En comparación con la histopatología y el cultivo
la técnica presenta una sensibilidad baja tanto en bovino (Sherman y cols. 1984,
Colgrove y cols. 1989, Kormendy 1990, Sockett y cols. 1992, Ferreira y cols. 2003),
como en pequeños rumiantes (Hope y cols. 2000, Sergeant y cols. 2003), aunque parece
mejor en estos últimos (Juste y cols. 1988). Su baja sensibilidad se debe a la
incapacidad de detectar animales con paratuberculosis subclínica, ya que detecta
correctamente animales con síntomas clínicos que excretan grandes cantidades de la
bacteria en las heces (Juste y cols. 1988, Huchzermeyer y cols. 1992, García Marín y
cols. 1992b, García Marín y cols. 1992c, Clarke y cols. 1996b, Pérez y cols. 1997,
García Marín y cols. 1999a, Pavlik y cols. 1999c). Esta prueba presenta mejor
correlación con el cultivo fecal que la CF o la intradermorreacción (Thomas 1983), pero
es menos sensible que el ELISA (Cocito y cols. 1994, Collins 1996, Whittington y cols.
2003). Para algunos autores, el ELISA no puede reemplazar completamente a la AGID
porque cada uno de ellos detecta una población diferente de animales infectados

DIAGNÓSTICO
106
(Robbe-Austerman y cols. 2006a). La especificidad obtenida en estos estudios es muy
alta, aunque parece ser incapaz de diferenciar claramente las infecciones producidas por
Map, Maa, y Mas (Kormendy y cols. 1984, Chávez 1993, Pérez y cols. 1995b).
• Prueba de ELISA
Existen dos tipos de ELISA, el directo, en el que se detecta un determinado antígeno
por medio de anticuerpos, y el indirecto, en el que lo detectado son los anticuerpos
frente a un determinado antígeno. El ELISA indirecto consiste en capturar por unión
Ag-Ac los anticuerpos presentes en un fluido biológico (suero, plasma, orina, leche) por
medio de un antígeno fijado a un sustrato que sea reconocido por estos. Su revelado se
realiza mediante un conjugado entre anticuerpo o proteína con afinidad específica por
las inmunoglobulinas y un enzima. Al unirse este conjugado al complejo Ag-Ac, éste
queda igualmente fijado, y tras añadir un sustrato para el enzima, se produce un cambio
de color, cuya densidad óptica es proporcional a la cantidad de complejo fijado. Fue
introducida en el diagnóstico de la paratuberculosis por Jorgensen y cols. (Jorgensen y
cols. 1978), y es actualmente la técnica serológica más utilizada (Harris y cols. 2001).
Su éxito radica en su bajo coste, rapidez, versatilidad, y adaptabilidad (Juste y cols.
1990a, Collins 1996, Pérez y cols. 1997, Garrido y cols. 2000a, Collins 2002).
Probablemente, los antígenos más extendidos para el ELISA de paratuberculosis
sean el PPA3 (Lyle y cols. 1983, Yokomizo y cols. 1983) y el lipoarabinomanano
(LAM) (Sugden y cols. 1989). Como se ha dicho antes, el PPA3 es un derivado crudo
de una cepa de M. avium que al principio fue considerada Map, aún así no da resultados
menos específicos que otro antígeno derivado de la cepa VRI 316 de Map (Nielsen y
cols. 2001). Se ha recurrido a la elaboración de antígenos no purificados similares por
medio de tratamientos físico-químicos de cultivos de Map (Milner y cols. 1987,
Colgrove y cols. 1989, Cox y cols. 1991, Hilbink y cols. 1994, Sweeney y cols. 1995,
Pavlik y cols. 1999c, Hope y cols. 2000, Speer y cols. 2006). Hoy en día se pueden
encontrar unos cuántos de manera comercial (Allied Monitor, CSL,...). En la busca de
antígenos más específicos, han sido probados otros muchos candidatos: A y D (Sugden
y cols. 1988, Sugden y cols. 1991), polipéptido a362 (Vannuffel y cols. 1994), las
proteínas constitutivas AhpC y AhpD, y la secretada de 14 kDa (Olsen y cols. 2001b),
proteínas de choque térmico (Stevenson y cols. 1991, Stevenson y cols. 1992, Colston y
cols. 1994, Koets y cols. 2001), proteína de 35 kDa (Shin y cols. 2004), IS900-
glutatione-S-transferasa (Nakase y cols. 2006), y otros muchos que se están
investigando actualmente (Griffin y cols. 2005, Bannantine y cols. 2006a, Cho y cols.
2006a, Bannantine y cols. 2006b, Cho y cols. 2006b). La parte del conjugado encargada
de unirse a los anticuerpos captados por el antígeno de la placa de ELISA pueden ser
anti-IgG o anti-IgM específicos de especie, o proteínas con alta afinidad por las
inmunoglobulinas de diferentes especies como la proteína A o la G (Garrido y cols.
1999a, Garrido y cols. 2000a). Como enzima suele utilizarse o peroxidasa de rábano

REVISIÓN DE LA BIBLIOGRAFÍA
107
(HRP) o fosfatasa alcalina (AP), y como sustrato orto-fenil-diamina (OPD), tetrametil-
bencidina (TMB), ácido sulfónico 2,2’-anzino-di(3-etil)benzotiazolina (ABTS), ácido 5-
aminosalicílico (ASA), o para-nitrofenil fosfatasa alcalina (PNPP) (Garrido y cols.
2000a).
En comparación con otras técnicas serológicas, es más sensible y específica
(Yokomizo y cols. 1991, Sockett y cols. 1992, Meylan y cols. 1994, Sweeney y cols.
1995, Kalis y cols. 2002), aunque su sensibilidad puede variar considerablemente,
presentando valores en el rango de 15-98% en bovino (Colgrove y cols. 1989, Milner y
cols. 1990, Collins y cols. 1993c, Meylan y cols. 1994, Sweeney y cols. 1995, Garrido
2001, Dargatz y cols. 2001a, Ferreira y cols. 2002, Pak y cols. 2003, Garrido y cols.
2003, McKenna y cols. 2005b, Eda y cols. 2006), y de 40-100% en pequeños rumiantes
(Juste y cols. 1988, Sugden y cols. 1989, Molina y cols. 1991, Molina Caballero y cols.
1993, Pérez y cols. 1997, Hope y cols. 2000, Sergeant y cols. 2003). Como en las otras
pruebas de detección de inmunidad humoral, generalmente su sensibilidad aumenta con
el grado de excreción de bacterias en heces y el grado de difusión de las lesiones
(Milner y cols. 1990, Sockett y cols. 1992, Billman-Jacobe y cols. 1992, Ridge y cols.
1992, Spangler y cols. 1992b, Collins y cols. 1993c, Sweeney y cols. 1995, Clarke y
cols. 1996b, Pérez y cols. 1997, Reichel y cols. 1999, García Marín y cols. 1999a,
Pavlik y cols. 2000d, Dargatz y cols. 2001a, Kalis y cols. 2002, González 2003,
Sergeant y cols. 2003). En comparación con el cultivo la sensibilidad general del ELISA
puede ser de un 14-30% (Collins y cols. 2005, McKenna y cols. 2005a), muy baja para
muchos. No obstante, la frecuencia de detección de animales eliminadores en heces es
alta, y además, la magnitud de los resultados (S/P o el valor de densidad óptica) del
ELISA están directamente relacionadas con la probabilidad de que el animal esté
infectado y excretando micobacterias en sus heces, lo que permite detectar los animales
infecciosos de un rebaño (Collins y cols. 2005). También es un buen instrumento para
predecir qué vacas se encuentran probablemente sufriendo una supresión importante en
su producción de leche (Lombard y cols. 2005).
Como la mayoría de antígenos utilizados eran mezclas no específicas, la
especificidad del ELISA fue mejorada de forma revolucionaria gracias al trabajo de
Yokomizo y cols. (Yokomizo y cols. 1983, Yokomizo y cols. 1985). En vista de las
reacciones inespecíficas que detectaban con muestras de campo, pensaron en eliminar
los anticuerpos correspondientes a otras micobacterias y microorganismos mediante la
preadsorción de los sueros con M. phlei, lo que dio muy buenos resultados.
Normalmente su especificidad no baja del 90% (Harris y cols. 2001). Teóricamente si el
antígeno es específico no sería necesario realizar el paso de adsorción de Yokomizo y
cols. Parece que se pueden conseguir mejores resultados con antígenos realmente
específicos de Map, sin preadsorber los sueros (Griffin y cols. 2005). El ELISA con los
antígenos AhpC y AhpD específicos de Map, y la proteína de 14 kDa específica de
MAC, permiten detectar animales infectados de paratuberculosis sin reacciones

DIAGNÓSTICO
108
cruzadas en casos de animales con tuberculosis (Olsen y cols. 2001b). Al miso tiempo,
el ensayo de polarización de fluorescencia (FPA) con el antígeno MPB70 de M. bovis es
capaz de detectar animales infectados por esa micobacteria sin interferencias en
animales con paratuberculosis (Surujballi y cols. 2002).
El ELISA también se puede aplicar a muestras de leche (Sweeney y cols. 1994),
incluso como indicador de la estimación de la prevalencia de rebaños utilizando
muestras de leche de tanque (Nielsen y cols. 2000b, Beyerbach y cols. 2004). En
comparación con los resultados de ELISA de suero sanguíneo, la sensibilidad es menor
aunque parece mantener una concordancia aceptable (Sweeney y cols. 1994, Winterhoff
y cols. 2002, Klausen y cols. 2003, Munjal y cols. 2004, Salgado y cols. 2005, Hendrick
y cols. 2005a, Hendrick y cols. 2005b, Lombard y cols. 2006), pero no para todos los
autores (Hardin y cols. 1996). En cuanto a los resultados del cultivo se refiere, para
algunos, los resultados del ELISA de leche muestran una concordancia superior que los
del ELISA de suero (Hendrick y cols. 2005b). En otros trabajos, ha sido analizada la
probabilidad de ser positivo en suero y leche en relación con la fase de lactación y de
parto. Se ha observado que la positividad aumenta en suero al final de la lactación, en
leche al principio de la misma, y tanto en suero como en leche a partir del segundo parto
(Nielsen y cols. 2002a, Nielsen y cols. 2002b, Kudahl y cols. 2004).
Sigue la mejora del ELISA con antígenos que otorgan mayor especificidad y
sensibilidad (Eda y cols. 2006, Speer y cols. 2006). Una nueva técnica serológica
basada en citometría de flujo, ha mostrado una sensibilidad y especificidad
prometedoras (95,2 y 96,7%) al ser comparada con un ELISA comercial (Eda y cols.
2005), y es capaz de detectar animales con infección subclínica. Gracias al uso de
análisis estadísticos (método Bayesiano, “likelihood ratio”, “receiver operation
characteristic curve” [curva ROC], ...) y de la incorporación de otra información (edad,
datos de producción,...) al análisis, se pueden hacer predicciones más precisas del estado
de los animales (Collins 2002, Nielsen y cols. 2002c, Nielsen y cols. 2003, Van Schaik
y cols. 2003a, Wang y cols. 2006, Nielsen y cols. 2006, Berghaus y cols. 2006a).
En conclusión, el carácter multifásico o espectral de la enfermedad, hace imposible
la adopción de una técnica de referencia única, y si se pretende obtener un diagnóstico
de gran sensibilidad y especificidad, debe aplicarse una combinación de técnicas, y
hacerlo de manera repetida si es posible (Pérez y cols. 1997, García Marín y cols.
1999a, Pavlik y cols. 2000d, Garrido 2001, Barrington y cols. 2003, Toft y cols. 2005).
TRATAMIENTO Y MEDIDAS DE CONTROL
En vista del elevado coste y de la ineficacia de los tratamientos disponibles,
actualmente no se recomienda el uso de ningún tratamiento contra la paratuberculosis en
ganado, a pesar de que ofrezcan buenos resultados in vitro (Aduriz y cols. 2000, Harris
y cols. 2001). Únicamente sería aceptable para la recuperación temporal de animales de

REVISIÓN DE LA BIBLIOGRAFÍA
109
alto valor genético (Gezon y cols. 1988, St Jean 1996), teniendo en cuenta que los
animales tratados pueden suponer un importante foco de contagio (Juste y cols. 1990b),
pues aunque su situación clínica puede mejorar durante el suministro de los fármacos, la
excreción de micobacterias en heces continúa (Larsen y cols. 1953, Gilmour y cols.
1971, Merkal y cols. 1973, Hendrick y cols. 2006). Otros autores en cambio, aseguran
que los animales tratados con estreptomicina, rifampicina, y pirazinamida o levamisol,
no dan cultivos fecales positivos (Das y cols. 1992, Belloli y cols. 1995). Se han
probado muchos fármacos o combinaciones de los mismos in vivo, entre otros,
isoniazida, rifampizina, rifabutina, gentamicina, etambutol, claritromicina, amikacina y
monensina (St Jean y cols. 1991, Chiodini y cols. 1993, Cocito y cols. 1994, St Jean
1996, Williams y cols. 1999, Brumbaugh y cols. 2000, Whitlock y cols. 2006a), pero
ninguno de ellos es capaz de eliminar permanentemente la infección. En el caso de la
enfermedad de Crohn también se han probado varios tratamientos (revisado en
Implicaciones en salud pública. Enfermedad de Crohn), presuponiendo una etiología
en la que Map estaría involucrado. Un tratamiento de 17-54 meses de duración, en el
que se combina rifabutina y claritromicina, con y sin clofaximina, es capaz de inducir la
remisión de los síntomas, aunque ni cura la enfermedad ni garantiza la desaparición de
nuevas recaídas (Hulten y cols. 2000a, Borody y cols. 2002, Shafran y cols. 2002a).
Otra hipotética alternativa sería llevar a cabo tratamientos con micobacteriófagos
(virus bacteriófagos que infectan a las micobacterias), pero su utilización in vivo tiene
muchas limitaciones: necesidad de rebajar la acidez del abomaso para garantizar la
supervivencia de los fagos, monitorización constante de la resistencia-sensibilidad de
Map a los fagos, evidencia experimental de que las bacterias intracelulares se
encuentran protegidas de los fagos (tendrían que atacarlas en fase extracelular), falta de
evidencia experimental in vivo sobre la efectividad de este tratamiento sobre bacterias
intracelulares obligadas causantes de enfermedades crónicas, y el potencial peligro de
que la transducción o transfección con fagos incremente la virulencia de Map (Emery y
cols. 2004).
Entre las medidas de control aplicables, la primera a tener en cuenta y de
cumplimiento constante durante cualquier plan de control de la paratuberculosis, es
llevar correctamente un manejo encaminado a evitar la introducción y la transmisión de
la enfermedad en el rebaño (Juste y cols. 1990b). Son especialmente importantes las
costumbres de manejo que afectan a la reposición, los partos, los terneros, los animales
con infección detectada, la higiene y los purines (Aduriz y cols. 2000). La introducción
de nuevos animales debería hacerse con las máximas garantías (Chiodini y cols. 1984a,
Aduriz y cols. 2000, Carpenter y cols. 2004). Las vías de transmisión fecal-oral y láctea
deberían ser atajadas evitando o restringiendo el contacto entre madres y crías, y entre
adultos y jóvenes en general, evitando el contacto de los purines con los animales, los
alimentos y la bebida, realizando tratamientos efectivos del calostro y de la leche (o usar
preparados artificiales), manteniendo unas buenas medidas higiénicas generales, además

TRATAMIENTO Y MEDIDAS DE CONTROL
110
de eliminando o aislando los animales con sintomatología clínica (Blood y cols. 1983,
Chiodini y cols. 1984a, Goodger y cols. 1996, Rossiter 1996, Obasanjo y cols. 1997,
Johnson-Ifearulundu y cols. 1999a, Muskens y cols. 2000, Pavlik y cols. 2000d, Kalis y
cols. 2001, Beyerbach y cols. 2001, Gerlach 2002, Muskens y cols. 2003, Groenendaal
y cols. 2003b). En casos de madres que excreten micobacterias en heces, puede ser
aconsejable la eliminación de su descendencia pues puede encontrarse infectada por vía
intrauterina (Chiodini y cols. 1984a). La mayoría de estas medidas son económicamente
sostenibles y rentables, y su adopción reduce considerablemente la prevalencia del
rebaño (Groenendaal y cols. 2003a), aunque no siempre funcionan, principalmente por
incumplimiento de alguna. Durante el último congreso internacional sobre
paratuberculosis (8th International Colloquium on Paratuberculosis, 14-17 de agosto de
2005, Copenhague, Dinamarca), se subrayó la importancia de una correcta aplicación de
este tipo de medidas, de la caracterización de los factores de riesgo, del uso de sistemas
de certificación, así como de la concienciación de las partes implicadas. Además de
contemplar estos factores, los planes de control de la paratuberculosis se basan en la
detección y eliminación de animales infectados y/o en la vacunación.
Programas de detección y eliminación de positivos
El objetivo de este método es el de erradicar la enfermedad, por medio del sacrificio
de los animales que den resultado positivo en la prueba (o pruebas) de seguimiento
seleccionada (“test and cull”). Al margen de otras apreciaciones, la primera limitación
de este sistema yace en la propia ineficacia de las técnicas de diagnóstico (Whitlock y
cols. 2000), ya que hay animales infectados que pasan inadvertidos, y otros que no lo
están y pueden ser equivocadamente identificados como positivos. El cultivo fecal ha
sido la base de varios programas de control en los que no se ha logrado alcanzar ese
propósito (Moyle 1975, Rossiter 1992, Whitlock y cols. 1992b, Whitlock y cols. 1995,
Ridge y cols. 1995b, Rossiter 1996, Kalis y cols. 1999a, Kalis y cols. 2000, Benedictus
y cols. 2003, Pavlik y cols. 2003). Las pruebas serológicas utilizadas al principio (CF e
intradermorreacción) tampoco dieron buenos resultados por sí solas (Benedictus 1984).
Posteriormente se introdujo el ELISA combinado con el cultivo y a veces con otras
pruebas serológicas como la del interferón-γ (Rossiter y cols. 1992, Whitlock y cols.
1995, Collins 1995, Ridge y cols. 1995b, Bulaga y cols. 1999, Kalis y cols. 1999a, Kalis
y cols. 2000, Luyven y cols. 2002, Kalis y cols. 2003b), que junto con la aplicación de
medidas higiénico-sanitarias ha dado mejores resultados (Kalis y cols. 2003b). No
obstante, las estimaciones sobre la sensibilidad del ELISA y del cultivo en estadios
preclínicos, no superan el 15-25% y el 30% respectivamente, además de que el ELISA
puede identificar erróneamente como positivos hasta un 4% de animales (Whitlock y
cols. 2000, Collins y cols. 2005). A pesar de la introducción de métodos moleculares
(Thoen y cols. 1996a), en vista de la sensibilidad de estos métodos tampoco es esperable
una gran mejora del sistema, aunque sí más rapidez.

REVISIÓN DE LA BIBLIOGRAFÍA
111
Los costes de la monitorización de rebaños pueden resultar muy elevados (Aduriz y
cols. 2000), por lo que se han buscado varias estrategias para reducirlos, como por
ejemplo, el cultivo fecal de muestras agrupadas (Kalis y cols. 1999a, Whittington y cols.
2000a), el análisis de muestras ambientales del rebaño (Raizman y cols. 2004, Berghaus
y cols. 2006b), o el análisis de subpoblaciones concretas del rebaño (Gardner y cols.
2006). El éxito económico de este tipo de programas depende de la situación específica
de cada rebaño, y no resultan rentables para la mayoría de rebaños que se basan en
ELISAs actualmente disponibles (Groenendaal y cols. 2003a). Aparentemente la
erradicación de la paratuberculosis bovina no es posible con los conocimientos actuales,
pero sí se logra reducir su prevalencia, y haciendo hincapié en la concienciación e
implicación de ganaderos y veterinarios (Kalis y cols. 1999a, Kennedy y cols. 2001,
Whittington y cols. 2001c, Benedictus y cols. 2003, Kennedy y cols. 2006, Caldow
2006, Wells 2006) se puede llegar a un escenario mucho más optimista. A pesar de que
existen casos puntuales en los que se ha alcanzado la erradicación de la enfermedad por
el método de detección y eliminación, todos los programas generales de este tipo han
fracasado (Fridriksdottir y cols. 2000, Kennedy y cols. 2001, Benedictus y cols. 2003,
Bakker y cols. 2006).
Programas que incluyen la vacunación
El objetivo inicial de una vacuna es el de inmunizar a un hospedador potencial
mediante la inoculación de antígenos del agente infeccioso, para que cuando éste último
entre en el organismo del hospedador su respuesta inmunológica no permita que se
produzca la infección. La primera vacuna contra la paratuberculosis, que estaba
compuesta por una mezcla de bacilos atenuados, parafina, aceite de oliva y piedra
pómez, se utilizó en Francia para inmunizar vacas (Vallée y cols. 1926). Tras su
seguimiento, observaron la reducción de animales excretores así como la disminución
de la prevalencia de casos clínicos (Vallée y cols. 1934), resultados que animaron a
continuar con estas pruebas. Su utilización se extendió a otros países como Inglaterra
(Wilesmith 1982), Holanda (Huitema 1967, Benedictus y cols. 1988, Wentink y cols.
1992, Kalis y cols. 1992), Islandia (Sigurdsson 1960) o Dinamarca (Jorgensen 1983),
además de mantenerse en Francia (Marly y cols. 1988, Saint-Marc y cols. 1992a, Saint-
Marc y cols. 1992b). Tras la introducción de la enfermedad en Islandia, la inusitada
difusión que alcanzó en su cabaña ovina sólo pudo ser adecuadamente controlada con el
uso de la vacuna (Sigurdsson 1960, Fridriksdottir y cols. 2003a).
Las reducciones en el número de animales con síntomas clínicos o excretores es
variable según los autores y difícil de comparar en función de los diferentes métodos y
períodos de aplicación, pero en programas de 5 o más años, ésta puede situarse entre un
89 y un 99% (Sigurdsson 1960, Wilesmith 1982, Hurley y cols. 1983, Jorgensen 1983,
Benedictus y cols. 1988, Kormendy 1994). Sin embargo parece que nunca desaperecen
completamente los animales excretores (Kalis y cols. 1992), por lo que parece evidente

TRATAMIENTO Y MEDIDAS DE CONTROL
112
la necesidad de combinar la vacunación con medidas higiénico-sanitarias y de detección
y eliminación de animales excretores (Kalis y cols. 1999b).
Tal y como demuestran diferentes trabajos experimentales y de campo, la
vacunación ha proporcionado mejores resultados en pequeños rumiantes que en bovino,
principalmente en ovejas (Sigurdsson 1960, Brotherston y cols. 1961a, Brotherston y
cols. 1961b, Nisbet y cols. 1962, Gilmour y cols. 1973, Crowther y cols. 1976, Pérez
1992, Aduriz 1993, Juste y cols. 1994, Tellechea y cols. 1994, García Marín y cols.
1995b, Pérez y cols. 1995c, García Marín y cols. 1997, Corpa y cols. 1999, Tharaldsen
y cols. 2003, Reddacliff y cols. 2006, Hines y cols. 2006). Esto se debe en parte a que
las ovejas son resistentes a la tuberculosis, con lo que la vacunación de paratuberculosis
en esta especie no se ha visto restringida por las interferencias en las pruebas
diagnósticas de las campañas de control de la tuberculosis (intradermorreacción),
permitiendo así un amplio seguimiento y contrastado de resultados de múltiples trabajos
(Aduriz y cols. 2000). Sin embargo, los casos positivos pueden volver a aparecer en
zonas supuestamente libres si se deja de vacunar (Djonne 2003, Fridriksdottir y cols.
2003b). Algo semejante ha ocurrido en Holanda, donde gracias al programa de
vacunación en explotaciones bovinas con elevadas prevalencias (vacuna que también
fue utilizada en rebaños de pequeños rumiantes), la prevalencia y las pérdidas fueron
reducidas, pero el programa fue parado por informes conflictivos sobre su eficacia y por
interferencias con la intradermorreacción de la tuberculosis, lo que generó problemas
severos en la industria caprina principalmente (Bakker y cols. 2006). En el estado
español se han otorgado varios permisos especiales para realizar estudios de vacunación
en la especie bovina con una vacuna comercial (SILIRUM®), y parecen obtenerse
resultados equiparables a los obtenidos en trabajos con otras especies hospedadoras
(Garrido y cols. 2006, García-Pariente y cols. 2006, Muñoz y cols. 2006).
Según los hallazgos inmunopatológicos del grupo de Veterinaria de la Universidad
de León, la vacunación reduce la tasa de sacrificios por paratuberculosis, reduce la
incidencia de la enfermedad clínica, y en la mayoría de los casos, limita la extensión y
severidad de las lesiones, todo ello, en las tres especies rumiantes domésticas más
importantes (Pérez 2006). En definitiva, se puede decir que la vacuna no evita la
infección con Map, pero la incidencia de casos clínicos se ve reducida, además de ser
una medida muy económica. Favorece la aparición de respuesta celular y humoral
tempranas, controlando la evolución de las lesiones a tuberculoides, provocando su
focalización, reduciendo la cantidad de bacilos presentes en las mismas, así como la
cantidad que se excreta en las heces (Nisbet y cols. 1962, Jorgensen 1983, García Marín
y cols. 1988b, Juste y cols. 1994, Kormendy 1994, Pérez y cols. 1995c, Van Schaik y
cols. 1996, Gwozdz y cols. 2000a, García-Pariente y cols. 2003). Estas respuestas
interfieren al mismo tiempo con las pruebas serológicas de diagnóstico (Aduriz y cols.
2000).

REVISIÓN DE LA BIBLIOGRAFÍA
113
Los resultados que se obtienen con vacunas inactivadas o vivas atenuadas son
similares (Larsen y cols. 1964, Wilesmith 1982, Marly y cols. 1988, García Marín y
cols. 1999b), aunque actualmente se encuentra más extendido el uso de las inactivadas
por miedo a la reactivación de las vivas (Aduriz y cols. 2000). También se han probado
vacunas compuestas de fracciones de microorganismos (Larsen y cols. 1978b, Hurley y
cols. 1983), antígenos purificados de Map (Mullerad y cols. 2002, Mullerad y cols.
2003, Koets y cols. 2006a), vacunas de DNA (Bannantine y cols. 2001, Huntley y cols.
2005, Sechi y cols. 2006), y a pesar de que algunas de ellas permiten diferenciar entre
animales infectados y vacunados, o no son tan efectivas como las celulares completas, o
su efectividad no ha sido demostrada. Por otro lado, hasta el momento los ensayos de
revacunación no han mostrado una mayor protección, y la vacuna se administra sólo una
vez en la vida del animal, lo que parece ser suficiente (Aduriz y cols. 2000).
Normalmente se recomienda administrar la vacuna durante el primer mes de vida con el
fin de proteger al animal antes de ser infectado y de minimizar la interferencia en el
diagnóstico posterior de tuberculosis (Doyle 1956, Stuart 1962, Leslie y cols. 1988,
Aduriz y cols. 2000). A pesar de ello, la vacunación de animales adultos también es
beneficiosa, incluso más efectiva en algunos casos, aunque los animales ya se
encuentren infectados (Sigurdsson 1960, Crowther y cols. 1976, Benedictus y cols.
1988, Pérez 1992, García Marín y cols. 1995b, García Marín y cols. 1997, Corpa y cols.
2000b, García-Pariente y cols. 2003, García-Pariente y cols. 2006). La vía de
administración más eficiente es la subcutánea (Vallée y cols. 1926, Larsen y cols.
1978b, Marly y cols. 1988) y preferiblemente en el pliegue del pecho para evitar
molestias al animal (Juste y cols. 1990b, Aduriz 1993), pero también se ha inoculado en
la axila (Peris 1992, Corpa y cols. 1999). Aunque las vías pueden ser otras como la oral
(Brotherston y cols. 1961a), la conjuntival (Marly y cols. 1988), o la intradérmica
(Saint-Marc y cols. 1992b), estas no dan mejores resultados (Aduriz y cols. 2000).
Transcurridos varios días desde la aplicación de la vacuna, se forma un nódulo
fibrocaseoso en el punto de inoculación, debido en parte a la composición oleica de los
adyuvantes, que muestran ciertas diferencias entre unos y otros (Hagan 1935, Reyes y
cols. 2003). Si la vacuna se administra adecuadamente, el inconveniente que puede
suponer la formación del nódulo vacunal se ve reducido a un efecto estrictamente
estético, que con el tiempo puede incluso desaparecer (Hagan 1935, Sigurdsson y cols.
1949, Spears 1959, Gilmour y cols. 1966, Reddacliff y cols. 2006), y no produce
pérdidas económicas de importancia (Eppleston 2006). Si se inyecta incorrectamente
pueden aparecer lesiones problemáticas desde el punto de vista tanto patológico como
económico (Windsor y cols. 2006). La autoinoculación en personal encargado de su
administración es otro problema a tener en cuenta, pues los casos más graves requieren
cirugía y drenaje para evitar la necrotización de los tejidos afectados (Sherman 1985,
Patterson y cols. 1988, Windsor y cols. 2005).


ESTUDIOS


ESTUDIO I
Comparison of blood polymerase chain reaction
and enzyme-linked immunosorbent assay for
detection of Mycobacterium avium subsp.
paratuberculosis infection in cattle and sheep
Ramón A. Juste, Joseba M. Garrido, Mariví Geijo, Natalia Elguezabal, Gorka
Aduriz, Raquel Atxaerandio and Iker Sevilla
Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER), Berreaga, 1, 48160 Derio, Bizkaia, Spain.
Journal of Veterinary Diagnostic Investigation, Vol. 17, 2005, pp. 354–359


ESTUDIOS
119
Abstract
A study was carried out to compare the performance of enzyme-linked
immunosorbent assay (ELISA) and blood polymerase chain reaction (PCR) for
diagnosis of paratuberculosis in cattle and sheep. For cattle, a set of 278 samples from
one paratuberculosis affected Friesian farm was used; it included 80 ELISA-positive
samples and 198 ELISA-negative samples from an age-matched group. Ninety-four
samples were from heifers and 184 were from 2-5-year-old cows. The overall analysis
showed a clear association (Fisher exact test [FET] p = 0.0049) but a weak negative
agreement (45.3%, kappa = -0.1665 ± 0.0994) between the two tests. It reflected a
moderate agreement among heifers (87.7%, kappa = 0.4471 ± 0.2435) and a moderate
disagreement among cows (62.7%, kappa = -0.3670 ± 0.1057). For sheep, 496 single
blood samples from 53 Latxa dairy flocks; 180 of the blood samples were from
dam/offspring pairs. The overall association between the 2 tests on ovine samples was
strong (FET, p < 0.0005), whereas the agreement was low (kappa = 0.1622 ± 0.1188).
There was slightly better agreement for ewes (kappa = 0.2135 ± 0.1992) than for lambs
(kappa = 0.1193 ± 0.1301). There was also a highly unlikely proportion of
dam/offspring positive results (FET p < 0.0001, kappa = 0.6269 ± 0.1854). Four of 6
lambs that were necropsied one year after testing had paratuberculosis microscopic
lesions in the ileocecal valve (3 lambs) or a PCR positive result (4 lambs). These results
suggest that blood PCR testing might be a potentially useful new approach in
paratuberculosis diagnosis, especially in young animals.
Introduction
It is widely accepted that the most reliable method for diagnosis of paratuberculosis
in live animals is fecal culture (Collins y cols. 1993b). However, this method has the
drawbacks that it takes several weeks to yield a result and there is no good
interlaboratory standardization (RH Whitlock, personal communication). Although the
use of new growth detection methods can significantly reduce this time (Whittington y
cols. 1998), fecal culture is still an expensive and labor intensive technique. The
alternative use of serological methods such as the enzyme-linked immunosorbent assay
(ELISA) has become more popular because blood is a very convenient sample.
Moreover, ELISA tests are fast and inexpensive and can be easily automated for
processing large numbers of samples. However, the usefulness of serological tests is
compromised by the variability of the immune response depending on the
immunopathological form of paratuberculosis. For this reason, although several
commercial ELISA tests are available, it is generally accepted that their sensitivity in
detecting infected animals is only about 50% (Meylan y cols. 1994). In fact, because of
its low sensitivity, the ELISA test is rarely positive in animals under 2 years of age and
frequently fails to detect individuals in the early phases of infection. Detection of

ESTUDIO I
120
Mycobacterium avium subsp. paratuberculosis (Map) by polymerase chain reaction
(PCR) in fecal samples has been reported as an alternative to fecal culture because it
allows quick detection based on amplification of IS900, an insertion sequence
considered to be highly specific to Map. However, this method has not gained
widespread use probably because of high costs and low throughput as well as problems
caused by PCR inhibitors present in feces (Collins y cols. 1993a, Harris y cols. 2001).
The advantage of PCR methods is that they can be easily adapted to a variety of
samples. Because there have been several reports on the use of PCR in blood samples in
cattle (van der Giessen y cols. 1992b, Gwozdz y cols. 1997), this method was applied to
a set of field samples to determine whether PCR testing of blood could be of any value
in some stages of the infection as a complement to the ELISA test.
Materials and methods
A set of 278 ethylenediaminetetraacetic acid (EDTA) blood samples of 1,016 tested
in a Friesian dairy farm severely affected with paratuberculosis was used. All ELISA-
positive animals were included (80 samples) as well as an age-matched ELISA-negative
set (198 samples). Ninety-four samples originated from 6-month to 2-year-old heifers,
and 184 were from 2- to 5-year-old cows.
A total of 496 single EDTA blood samples from Latxa dairy sheep in 53 flocks in
the Basque Country of Spain were used. A total of 316 samples originated from 3- to 8-
month-old lambs, whereas 180 were from ewes between 2 and 6 years of age. Of these,
180 lamb samples belonged to animals whose dam was also included in the study, and
therefore, allowed a dam/offspring pair comparison.
Ten milliliters of blood was drawn from the caudal vein in cattle, or from the jugular
vein in sheep, into EDTA vacuum tubes. After low-speed centrifugation, the plasma
was transferred to an Eppendorf tube, and the cellular fraction was incubated with
0.83% ammonium chloride (1:2, vol:vol) for 20 minutes to lyse the red blood cells.
Then, the tube was centrifuged for 10 minutes at 200 X g, and the cell pellet was
washed twice with phosphate-buffered saline (PBS) by centrifugation under the same
conditions. Thereafter, the cells were transferred to 2-ml tubes and submitted to 3 freeze
and boiling cycles before DNA extraction by chloroform–octanol separation (Garrido y
cols. 2000b). Both the plasma and the purified DNA were stored at -20ºC until use.
The absorbed ELISA test was performed to detect antibodies against Map in the
bovine and ovine samples following protocols reported previously for each species
(Garrido 2001). In brief, high absorption capacity polystyrene microtiter plates were
coated with 100 � l/well of 0.04 mg/ml solution of paratuberculosis protoplasmatic
antigen 3 (PPA-3) and frozen at -20ºC until use. The plasma samples were adsorbed
(1:1) with a saline suspension of Mycobacterium phlei (5 g/liter) (laboratory grown
strain from Dr. O. Fuentes, INIA, Madrid, Spain) to remove nonspecific anti-

ESTUDIOS
121
Mycobacterium spp. antibodies. After an overnight incubation at 4ºC, the supernatants
were diluted in PBS-Tween (0.05% Tween 80) pH 7.6 to 1/600 and 1/200 for cattle and
sheep, respectively, and 100 � l was added into duplicate wells of the antigen-coated
plate. Cattle and sheep positive and negative control sera were included in every plate in
duplicate according to the species tested. The positive control sera were obtained from 1
single culturally and histopathologically infected cattle or sheep. The negative sera were
obtained from single animals from a herd or flock without any clinical history of
paratuberculosis that were negative both by culture and by histopathological
examination. After a 2-hour incubation period at room temperature (RT) in a humidified
chamber, the plates were washed thrice with PBS-Tween. Protein G horseradish
peroxidase conjugate (0.025 � g/ml) was added and incubation was repeated at RT for 2
hours. After 3 washes with PBS-Tween, 100 � l of 2,2’-azino-di-(ethyl-benzythiazoline
sulfonate) (ABTS) substrate was added and plates were further incubated 20 minutes in
the dark. The reaction was stopped with 50 � l/well (2%) hydro-fluoric acid, and optical
density (OD) was measured in a spectrophotometer at double wave length (405/450
nm). The mean serum OD value was divided by the mean positive control OD, and this
ratio was used as the final serum reading for scoring the sample as negative or positive.
The ELISA cut-off was set by a sensitivity versus specificity plotting method (Dohoo y
cols. 2003) at the highest semi-sum of sensitivity and specificity for a specificity of at
least 90%. It was 0.345 (95.1% specificity) in cattle and 0.200 (96.9% specificity) in
sheep as determined by combined results of fecal culture and microscopic lesions from
a set of 42 and 51 positive and 244 and 26 negative cattle and sheep, respectively
(Garrido 2001).
A 2.5 � l aliquot of extracted DNA was added to a PCR mix containing primers P90
and PT91 and Taq polymerase in a final volume of 25 � l in 200-� l tubes (Garrido y
cols. 2000b). The tubes were placed in a thermal cycler and subjected to a cycling
profile consisting of 1 cycle at 96ºC for 2 minutes, 45 cycles of 96ºC for 15 seconds,
55ºC for 15 seconds, and 72ºC for 15 seconds, followed by an additional extension
period of 10 minutes at 72ºC (Garrido y cols. 2000b). Cross-contaminations were
checked by intercalating 1 distilled water blank after each group of 23 samples during
the extraction procedure. One additional negative control was included for every 46
samples during the PCR setup. The products were resolved by electrophoresis in a 2%
agarose gel for 50 minutes at 150 V. A band of the expected 389 bp size matching the
positive control was considered as a positive result. The target band from 6 positive
samples was cut out, purified with a GFX PCR DNA and Gel Band purification kit,
submitted to a nested PCR (Naser y cols. 2004), repurified, and sent to a commercial
subcontractor for sequencing to verify the specificity of the reaction.
The overall and age-group ELISA and PCR results were compared in a 2X2 table by
using Fisher exact test (FET). The agreement between the 2 methods was tested by the
kappa index test at a 95% confidence interval (Dohoo y cols. 2003). Both analyses were

ESTUDIO I
122
carried out with the FREQ procedure of SAS statistical package. Furthermore, a
variable called complementary sensitivity (CS) was calculated as the ratio of positive
results in only one method to the total number of positive results in the other method.
This variable was meant to represent the additional detection efficacy of one method
over the other, assuming both were highly specific.
Results
A total of 136 cattle samples had a positive result by ELISA or PCR (48.9%). Of
these, only 10 were positive by both methods, whereas 70 were positive only by ELISA
and 56 only by PCR (Table 1). The FET indicated that this distribution had a chance
probability of 0.0049. The kappa coefficient had a value of -0.1665 ± 0.0994 (95%
confidence interval), indicating a low degree of negative agreement (45.3%). The CS
for PCR and ELISA were, respectively, 106.1% and 70.0%. When results were
analyzed according to age, heifers had a total of 18 positive results of which 6 were
positive in both tests, 12 were positive by PCR alone, and none by ELISA alone (Table
1). The FET indicated a probability of less than 0.0001, with a kappa value of 0.4471 ±
0.2435, indicating a moderate agreement (87.7%). For this age group, the PCR CS was
200.0%, whereas the ELISA CS was 0.0%. When only adult cows were considered,
there were 118 samples with a positive result in any method. Of these, only 4 were
positive by both methods, 44 by PCR alone, and 70 by ELISA alone. The FET showed a
probability of less than 0.0001, with a kappa value of -0.3670 ± 0.1057, indicating a
clear disagreement (62.0%). The PCR CS (59.5%) was much lower than the ELISA CS
(145.8%).
Table 1. Cattle results.
ELISA +
PCR +
ELISA +
PCR -
ELISA –
PCR +
ELISA –
PCR -
Total
* FET
‡ Kappa
Agreement
§ CS
ELISA
§ CS
PCR
All 10 †
(3.6%)
70 †
(25.2%)
56 †
(20.1%)
142 †
(51.1%)
278 0.0049 -0.1665
(0.099)
54.7% 106.1% 70.0%
Heifers 6 †
(6.4%)
0 †
(0%)
12 †
(12.6%)
76 †
(80.9%)
94 <0.0001 0.4471
(0.244)
87.7% 0.0% 200.0%
Cows 4 †
(2.2%)
70 †
(38.0%)
44 †
(23.9%)
99 †
(53.8%)
184 <0.0001 -0.3670
(0.106)
38.0% 145.8% 59.5%
* FET: Fisher exact test for comparison of frequencies given as the probability of the distribution resulting from chance. § CS: Complementary sensitivity. † In parentheses, percent of each cell related to total. ‡ Kappa: kappa index of agreement. In parentheses, kappa 95% confidence error.
A total of 74 sheep samples had a positive result in any test. Of these, only 9 were
positive in both tests, whereas 13 were positive by ELISA alone and 52 by PCR alone
(Table 2). The FET indicated a probability of less than 0.0005. The kappa index was
0.1622 ± 0.1188 indicating a low degree of agreement (86.9%). The overall PCR CS
was 236.4%, whereas the ELISA CS was only 21.3%. Among the lambs, 3 were
positive by both tests, 1 by ELISA alone, and 36 by PCR alone (Table 1). The FET

ESTUDIOS
123
indicated that this distribution was unlikely to be due to chance (P = 0.0064), and the
kappa index (0.1193 ± 0.1301) indicated a lack of significant agreement (88.3%). For
this age group, the CS was 900.0% for PCR and only 2.6% for ELISA. The ewes had 6
positive results by both tests, 12 by ELISA alone and 16 by PCR alone. The FET for
these results indicated a probability of 0.0117. The kappa index indicated a fair
agreement (0.2135 ± 0.1992). The PCR and ELISA CS were 88.9% and 54.5%,
respectively.
Table 2. Sheep results. a) PCR and ELISA comparison. b) Dam/offspring PCR results.
a) ELISA +
PCR +
ELISA +
PCR -
ELISA –
PCR +
ELISA –
PCR - Total * FET ‡ Kappa Agreement
§ CS
ELISA
§ CS
PCR
All 9 †
(1.8%)
13 †
(2.6%)
52 †
(10.5%)
422 †
(85.1%)
496 <0.0005 0.1622
( 0.119)
86.9% 21.3% 236.4%
Lambs 3 †
(1.0%)
1 †
(0.3%)
36 †
(11.4%)
276 †
(87.3%)
316 0.0064 0.1193
(0.130)
88.3% 2.6% 900.0%
Ewes 6 †
(3.3%)
12 †
(6.7%)
16 †
(8.9%)
146 †
(81.1%)
180 0.0117 0.2135
(0.199)
84.4% 54.5% 88.9%
b) Dam +
Offs +
Dam +
Offs -
Dam -
Offs +
Dam -
Offs - Total * FET ‡ Kappa Agreement Dam pr Offs pr
13 †
(7.2%)
9 †
(5.0%)
4 †
(2.2%)
154 †
(85.6%)
180 <0.0001 0.6269
(0.185)
92.8% 76.5% 61.9%
* FET: Fisher exact test for comparison of frequencies given in probabilities of the distribution resulting from chance. § CS: Complementary sensitivity. † In parentheses, percent of each cell related to total. ‡ Kappa: kappa index of agreement. In parentheses, kappa 95% confidence error. Pr: probability; Offs: Offspring.
The dam/offspring analysis carried out on 180 ewe/lamb pairs indicated that there
was substantial agreement (92.8%) between PCR results (kappa = 0.6269 ± 0.1854)
indicating a clear association between dam and offspring with regard to Map apparent
presence in peripheral blood (Table 2). Of the 6 PCR-positive lambs necropsied 1 year
after the tests were carried out, 3 showed small focal lesions in the ileocecal valve. All 3
lambs plus a fourth lamb that had no lesions were PCR positive in this tissue. The 6
PCR products that were sequenced showed over 98–99% homology with published
sequences of the IS900 gene (GenBank accession numbers X16293, AF416985).
Discussion and conclusions
Because the absorbed ELISA with PPA-3 (Harris y cols. 2001) has generally been
found to be much more specific than sensitive, it can be assumed that positive results
are likely to correspond to truly infected animals (Meylan y cols. 1994). The specificity
of the PCR test, including the IS900-based test on fecal samples, is generally considered
to be very high, although there are a few reports of false-positive results (Cousins y
cols. 1999, Englund y cols. 2002). Use of PCR on bacteria-free sample such as blood
should be even more specific because unlike feces, blood should normally not contain
bacteria that might share some homology with the Map PCR target sequence. In fact,

ESTUDIO I
124
the cattle samples originated from a herd with a high incidence of clinical
paratuberculosis, which provides a good degree of certainty that the animals were
exposed to Map. The fact that sequencing confirmed all 6 cases tested suggests that
false-positive results were probably not a problem. Therefore, although no gold standard
confirmation method was used, it was assumed that both methods correctly identified
infected animals. Thus, the concept of CS introduced for paratuberculosis in this study
appears to be useful tool when no gold standard is available, and the methods being
compared are known to be highly specific. In this situation, the CS provides a measure
of the efficiency of combining methods to increase sensitivity.
The agreement between the 2 tests was low in general, and this was interpreted to
mean that each method would detect different forms or stages of Map infection because
their respective targets (bacteria and antibodies) might not have parallel dynamics. This
interpretation is consistent with the finding that young animals were more easily
detected by PCR than by ELISA, possibly because Map-loaded phagocytic cells would
physiologically recirculate from the intestinal lymphoid tissue to other lymphoid
locations very early after the infection, reinfection, or reactivation (van der Giessen y
cols. 1995). This should be expected to be more frequent among young animals newly
exposed to Map than in adults who are known to be more resistant to infection. On the
contrary, because the antibody response is slow to develop and highly dependent on the
total number of mycobacteria, only the most advanced cases should have detectable
antibody responses (Juste y cols. 1994). The ELISA-positive and PCR-negative results
are more difficult to explain under this hypothesis because they should correspond to
animals with large numbers of mycobacteria. One likely explanation is that
paratuberculosis pathogenesis might not be a progressive process, but it may rather have
alternating peaks and troughs in the antibody levels (Barrington y cols. 2003) and
bacteremia corresponding to successive relapses or reinfections. It would be important
to further investigate this hypothesis because it implies change in the current Map
pathogenesis paradigm.
The results of this study indicate that blood PCR may have a high potential as a
complementary diagnostic test to ELISA because it might pick up the fraction of
infected animals that ELISA tests usually fail to detect. In cattle, ELISA seemed to
perform at similar levels of detection as PCR. However, in sheep, the blood PCR test
clearly outperformed the ELISA in terms of positive results. This can be most likely
related to the characteristics of the samples which, in the case of cattle, originated from
a single paratuberculosis-affected herd and, in the case of sheep, originated from 53
different flocks of generally unknown Map infection status. On the other hand, the age
seemed to have a major effect on detection by both methods. Indeed, the only test
detecting all positive cases in heifers and almost all the positive cases in lambs was
PCR, whereas in adults, the ELISA test yielded more positives.

ESTUDIOS
125
From an epidemiological standpoint, the high prevalence of PCR-positive results in
sheep blood might indicate that there are many Map-carrier sheep that may sometimes
develop very limited focal lesions but remain clinically normal. This situation would
call into question the effectiveness of full eradication in paratuberculosis-control
programs. The close association between PCR results in ewes and lambs is noteworthy
because it indicates that the offspring of positive animals may be a greater risk of
infection.
In conclusion, the data in this report present a new approach for paratuberculosis
diagnosis on the basis of the complementary use of ELISA on plasma or serum samples
and PCR on blood samples to increase overall sensitivity. There is a need for further
research on the pathogenesis of Map to determine whether bacteremia occurs and to
validate the use of blood as a diagnostic sample.
Acknowledgements
We thank Drs. Belén Extramiana, Mercedes G. Goti, and Lorenzo Gonzalez for their
participation in the sheep study from which some of the samples used in this study were
obtained. We also thank Dr. Eduardo Berriatua for his useful critical comments on this
article.


ESTUDIO II
Molecular typing of Mycobacterium avium
subspecies paratuberculosis strains from
different hosts and regions
Iker Sevilla,1 Shoor V. Singh,2 Joseba M. Garrido,1 Gorka Aduriz,1 Sara
Rodríguez,3 María V. Geijo,1 Richard J. Whittington,4 Vanessa Saunders,5 Robert
H. Whitlock6 and Ramón A. Juste1
1 Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER), Berreaga 1, 48160 Derio, Bizkaia, Spain.
2 Goat Health Division, Central Institute for Research on Goats, Makhdoom, Post Farah Dist. Mathura, (UP) 28 11 22, India.
3 Universidad de Córdoba, Facultad de Veterinaria, Campus Universitario de Rabanales, Ctra. Madrid-Cádiz, km. 396, 14071 Córdoba, Spain.
4 Faculty of Veterinary Science, University of Sydney, NSW 2006, Australia.
5 NSW Agriculture, Camden NSW 2570, Australia.
6 USDA-ARS-National Animal Disease Center, New Bolton Center, Kennett Square, PA 19348-1692, United States of America.
Revue Scientifique et Technique (International Office of Epizootics),
Vol. 24, No. 3, 2005, pp. 1061-1066


ESTUDIOS
129
Summary
The IS1311 polymerase chain reaction-restriction endonuclease analysis was used to
detect genetic differences among 38 Mycobacterium avium subsp. paratuberculosis
(Map) isolates from cattle, sheep, goats and bison from distinct regions of Spain, India
and the United States of America (USA). In Spain, all eight bovine isolates, three out of
six caprine isolates and one of ten ovine isolates were of the C type, while the other nine
ovine isolates and three caprine isolates were of the S type. In India, all five ovine
isolates and six caprine isolates were of the B type, and so were all three isolates from
bison (Bison bison) from the USA. These results show that there are genetic differences
between Map isolates related to geographic and host factors that have a potential use in
the epidemiological tracing of new paratuberculosis isolates.
Introduction
Genotypic differences have been used to characterise isolates of Mycobacterium
avium subsp. paratuberculosis (Map), the agent of paratuberculosis in ruminants.
According to Collins et al. (Collins y cols. 1990a), these differences make possible a
classification of isolates into two groups: cattle-type strains and sheep-type strains.
Although the most widely used typing methods are IS900 restricted fragment length
polymorphisms (RFLP) (Pavlik y cols. 1999a) analysis and restricted genome on pulsed
field gel electrophoresis (Stevenson y cols. 2002), Marsh et al. (Marsh y cols. 1999)
developed a polymerase chain reaction-restriction endonuclease analysis (PCR-REA)
based on polymorphisms in IS1311, an insertion sequence present in Map and in
Mycobacterium avium subsp. avium (Maa) in seven to ten copies. This technique
provides an easy and rapid method of distinguishing between Maa and both cattle and
sheep Map strains. In a later study, Whittington et al. (Whittington y cols. 2001b)
discovered a new IS1311 restriction band pattern in bison isolates from Montana in the
United States of America (USA). In that study, this IS1311 PCR-REA typing method
was applied to a series of mycobacterial isolates in order to detect any genetic
differences related to host or geographical origin.
Materials and methods
The identity and origin of each isolate is shown in Table 1. Sample processing for
culture was performed as described by Aduriz et al. (Aduriz y cols. 1995a). While the
Indian ovine and caprine samples were cultured on Herrold’s egg yolk medium
(HEYM) without sodium pyruvate, the Spanish bovine paratuberculosis isolates were
grown on HEYM supplemented with sodium pyruvate. The Spanish ovine and caprine
paratuberculosis strains were grown on Middlebrook 7H11 medium supplemented with
Oleic acid-albumin-dextrose-catalase (OADC) (7H11) (Difco, Detroit, Michigan, USA)

ESTUDIO II
130
and on Löwenstein-Jensen (L-J) (Difco). Bison Map isolates grew on L-J or on 7H9,
and the remaining mycobacteria were isolated on Coletsos (Bio-Mérieux, Marcy-
l’Etoile, France). All media except Middlebrook and Coletsos were supplemented with
mycobactin J (Allied Monitor, Inc., Fayette, Missouri, USA). Mycobacterium avium
subsp. paratuberculosis was identified on the basis of time of incubation to visible
colonies, colony and bacilli morphology including acid fastness, mycobactin
dependance on egg-based media, and IS900 PCR.
Table 1. Characteristics of the isolates studied.
Geographic origin Host Sample Species Primary culture
No. of isolates Isolate code
IS900 PCR
IS1311 PCR-REA
Spain Basque Country
Cattle Faeces Map HEYM+ 5 341/02, 3515/02, 2059/03, 2226/03, 2702/03
Pos C
Castilla y León
Cattle Tissues Map HEYM+ 2 2484/02-7, 2484/02-18
Pos C
Madrid Cattle Faeces Map HEYM+ 1 181/02 Pos C
Andalucía Goat Tissues Map L-J 3 s/n, 43, 52504
Pos S
Andalucía Goat Tissues Map L-J 3 242, 392, 485
Pos C
Basque Country
Sheep Tissues Map 7H11 1 P38I Pos C
Basque Country
Sheep Tissues Map 7H11 8 9G, 12I, 14I, 17G, 18I, 19I, 21I, 269ov
Pos S
Andalucía Sheep Tissues Map L-J 1 27 Pos S
Basque Country
Cattle Tissues Maa Coletsos 1 3321/00 Neg Maa pattern
Andalucía Goat Tissues Maa Coletsos 1 Cap 120 Neg Maa pattern
Basque Country
Badger Tissues M. intracellulare Coletsos 1 2974/03 Neg 275 + 245 bp
Basque Country
Cattle Tissues M. bovis Coletsos 1 320/03 Neg Neg PCR
Cantabria Cattle Tissues M. chelonae Coletsos 1 858/03 Neg Neg PCR
Castilla y León
Goat Tissues M. bovis Coletsos 1 887/00 Neg Neg PCR
India Farah Goat Faeces Map HEYM– 6 S3, S5, T7, S2, S28, S4
Pos B
Mathura Sheep Faeces Map HEYM– 5 3812, S9, V95, V16, 43
Pos B
Mathura Sheep Faeces M. asiaticum HEYM– 1 V 30 Neg Neg PCR
USA Montana Bison Tissues Map L-J 3 6.1, 8.3, 10.3 Pos B
Montana Bison Tissues Maa 7H9 1 13.1 Neg Maa pattern
HEYM+: HEYM with Na pyruvate; HEYM–: HEYM without Na pyruvate. See rest of abbreviations in introduction and material and methods sections.

ESTUDIOS
131
For DNA extraction, colonies were suspended in 500 µl of TE-Triton X100
(Calbiochem, Bad Soden, Germany), subjected to three cycles of freezing and boiling,
and treated according to the protocol described by Garrido et al. (Garrido y cols.
2000b). Final DNA concentration was measured in a NanoDrop‚ ND-1000
spectrophotometer (NanoDrop Technologies, Inc., Rockland, Delaware, USA).
Five microlitres of the resuspended DNA were used in a IS1311 PCR mix
containing 0.8 µM of primers M56 and M119 (Marsh y cols. 1999), 200 µM of each of
the nucleotides dATP, dTTP, dGTP and dCTP, 20 mM of Tris-HCl, 50 mM of KCl, 2.5
mM of MgCl2 and 2 U of Taq polymerase (Invitrogene Ltd, Paisley, UK). The assay
was performed in a GeneAmp 9,600 PCR system (Applied Biosystems, Foster City,
California, USA) under the following conditions: one cycle of 3 min at 94ºC and 37
cycles of 30 s at 94ºC, 15 s at 62ºC and 1 min at 72ºC. In order to confirm Map
identification by IS1311 PCR, an IS900 PCR was also performed, using primers and
conditions described by Garrido et al. (Garrido y cols. 2000b). DNA bands of 608 bp
and 389 bp were considered PCR positive results for IS1311 and IS900, respectively,
after separation in a 2% agarose gel stained with ethidium bromide.
An IS1311 REA was performed to detect the C/T polymorphism at base pair 223
between Map strains. Eight microlitres of the positive IS1311 PCR solution was
digested for 2 h at 37ºC in a 16 µl reaction containing 2 U of each endonucleases HinfI
and MseI supplemented with buffers provided by the manufacturer (New England
Biolabs, Inc., Beverly, Massachusetts, USA). Band patterns visualised after
electrophoresis on 4% agarose gels stained with ethidium bromide were interpreted in
the manner indicated by Whittington et al. (Whittington y cols. 2001b): Map S strains
were defined by two bands of 285 bp and 323 bp (all IS1311 copies have a C at 223 bp);
Map C strains were characterised by four bands of 67 bp, 218 bp, 285 bp and 323 bp
(some copies with a C and some with a T); Map B strains were identified by three bands
of 67 bp, 218 bp and 323 bp (T in all copies); and finally, isolates showing three bands
of 134 bp, 189 bp and 285 bp were considered as Maa (C at base pair 223 and T at base
pair 422).
After purification with a GFX™ PCR DNA purification kit (Amersham
Biosciences, Buckinghamshire, UK), IS1311 PCR products of two ovine (3812, S9) and
two caprine (S3, S5) isolates from India, and one ovine (269ov.) and one bovine
(3515/02) isolates from Spain were sent to the DNA Automatic Sequencing Service of
the Centro de Investigaciones Biológicas-Consejo Superior de Investigaciones
Científicas in Madrid to confirm the DNA sequence of the amplicons and the point
mutations between strains. Both forward and reverse strands were sequenced with M56
and M119 PCR primers, respectively. Vector NTI v 8.0 software (InforMax Inc.,
Oxford, UK) was used to analyse and compare these sequences with those described

ESTUDIO II
132
earlier for Maa and Map C, S and B strains (GeneBank U16276, and EMBL AJ223974,
AJ223975, AJ308375).
The DNA from cultures of eight mycobacteria other than Map (Maa, M.
intracellulare, M. bovis, M. chelonae, M. asiaticum) was used as a negative control for
IS900 and IS1311 PCR tests. Identification of non-Map mycobacteria was performed by
the National Reference Laboratory at the Centro Nacional de Microbiología (Instituto
de Salud Carlos III, Madrid, Spain) by bacteriological and molecular methods.
Results
Both IS900 and IS1311 PCR and REA results are given in Table 1. As expected, all
Map isolates yielded positive results in both PCR assays, and all Maa isolates were also
positive in IS1311 PCR. None of the other isolates was positive in any of the PCR
assays performed, except M. intracellulare, which yielded a band of approximately 520
bp in the IS1311 PCR. Although the 67 bp band of C and B type digests is difficult to
see, all samples submitted to restriction analysis gave unmistakable band patterns
(Figure 1) which corresponded to the four possibilities described above, except for the
M. intracellulare isolate which produced two bands of approximately 275 bp and 245
bp.
Figure 1. Photograph of IS1311 polymerase chain reaction-restriction endonuclease analysis electrophoresis on 2% agarose gel stained with ethidium bromide. Lanes 1 and 10, 1Kb DNA size marker. Lane 2 shows an S pattern from a Spanish ovine isolate (269ov). Lane 3 shows a C type pattern from a Spanish bovine isolate (3515/02). Lane 4 shows a type B pattern from an American bison isolate (8-3). Lanes 5 and 6 show type B pattern from Indian caprine isolates (S5 and T7). Lanes 7 and 8 show type B pattern from Indian ovine isolates (3812 and S9). Lane 9 corresponds to an avian type isolate from a Spanish goat (Cap120).

ESTUDIOS
133
All Map isolates from India were identified as B type strains regardless of whether
they were of ovine or caprine origin. With regard to the Spanish cultures, while all the
bovine isolates gave a type C REA pattern, only one (P38I) of the ten ovine isolates
analysed was of C type, while all the others were of S type. Three of the six caprine
paratuberculosis isolates were S type and three were C type. Finally, the three bison
Map isolates from the USA were classified as B strains. MseI endonuclease recognised
the T at base position 422 of the two Maa isolates (substituted by a C in
paratuberculosis strains), which produced Maa type REA patterns.
The DNA sequences obtained from the amplified products matched exactly with
those published previously for each strain type. The ovine Map isolate from Spain,
which was S type in restriction analysis, had a C at base position 223. A mixed C/T
signal was detected at the same base position in the sequence of the Spanish bovine
paratuberculosis isolate (Figure 2), classified as C type by REA. And finally, all four
Indian ovine and caprine PCR products sequenced possessed a T in all copies at this
position, which had only been found before in bison isolates from the USA.
Discussion
Results of this study confirm previous findings (Whittington y cols. 1999a) that both
IS900 and IS1311 PCR techniques are suitable for use in Map confirmation and typing.
The unique amplification observed in M. intracellulare IS1311 PCR, is easily
distinguishable and can be confirmed by REA.
Figure 2. Sequences obtained from IS1311 amplicons of S, C and B type isolates matched exactly with those described before. Point mutations are indicated.

ESTUDIO II
134
The cultural characteristics and restriction patterns of Spanish paratuberculosis
isolates analysed in the authors’ laboratory to date are in accordance with part of the
literature (Marsh y cols. 1999, Whittington y cols. 2001b). These results indicate that
cattle are mainly infected with type C Map, which is readily isolated on HEYM,
although the presence of S type strains cannot be ruled out in this species since primary
cultures of bovine samples are usually performed only on this medium. Most of the
ovine Map isolates (nine out of ten) were of the S type and difficult to grow in primary
cultures, but a C type strain (in one case out of ten) was also detected in sheep. Goats
appeared to be affected by both C and S types, much as have been found in other studies
(Pavlik y cols. 1999a, Whittington y cols. 2001b, Stevenson y cols. 2002).
There is a certain discrepancy in the distribution patterns according to host species
that were found in this study and the results reported by other authors (Pavlik y cols.
2000a, Stevenson y cols. 2002). The latter conclude that C type strains are commonly
isolated from sheep in Europe, while in the present study only 10% of the sheep isolates
were found to be of C type. This could be for the same reason that was discussed above
with regard to the fact that only type C strains were isolated from cattle in the present
study. It is likely that our isolates might represent a less biased sample of strains
infecting Spanish sheep since we used L-J and 7H11 for primary isolation. According to
Juste et al. (Juste y cols. 1991b), L-J might improve primary isolation of ovine strains
by about 90%, but decrease primary isolation of bovine strains by approximately 37% in
relation to HEYM. The other authors either did not report the media used for primary
isolation (Stevenson y cols. 2002) or used HEYM (Pavlik y cols. 2000a). A larger, more
specifically designed study that avoids primary isolation bias is needed in order to draw
any definitive conclusions on this subject.
It has been reported that culture of Map from bison samples is more difficult than
from cattle but easier than from sheep, and that it can be improved by the use of HEYM
without sodium pyruvate (Whitlock y cols. 1999). In the case of the Indian isolates,
such differences were not noticed because only HEYM without added pyruvate was
used for primary isolation. The observation of Map colonies produced from sheep
samples cultured in HEYM (Das y cols. 1991, Singh y cols. 1996, Dimareli-Malli y
cols. 2001) contradicts the findings of some other authors, who reported that the culture
of Map colonies from sheep samples was unrewarding (Juste y cols. 1991b, Whitlock y
cols. 1999). These findings led to the suspicion that a strain different from S could be
involved in the Indian cases in the present study, probably a C strain. However the
molecular methods used in this study proved that they were B type strains, which had
previously only been found in bison from Montana (USA). Nevertheless, the
preliminary results from RFLP analysis of genomic DNA indicate that these isolates are
not the same as those from bison and probably represent a new biotype, as yet not seen
outside India (Whittington, unpublished observations). In summary, this study confirms
the existence of Map genetic differences related to geographic and host factors that

ESTUDIOS
135
would help to explain the variable success rates in the primary isolation of Map from
some host species on some culture media in previous reports. These differences could
also prove useful for the epidemiological tracing of new paratuberculosis cases.
Acknowledgements
We thank V. Kumar and S. Hajra, from the College of Veterinary Science and
Animal Husbandry of C. S. Azad University of Agriculture and Technology (Kanpur,
Mathura), for performing primary cultures, and A.K. Bhatia, Director of the Central
Institute for Research on Goats, for providing necessary facilities. Samples from
Castilla-León were provided by the Pathology Unit of the Faculty of Veterinary
Medicine of León. The Centro Nacional de Microbiología of the Instituto de Salud
Carlos III kindly made the non-Map mycobacteria identification. We also thank Mertxe
Bascones for technical help at NEIKER. While conducting this study, I. Sevilla held a
fellowship from the Department of Agriculture and Fisheries, and currently holds a
fellowship from the Department of Industry of the Basque Government.


ESTUDIO III
Pulsed-field gel electrophoresis profile
homogeneity of Mycobacterium avium subsp.
paratuberculosis isolates from cattle and
heterogeneity of those from sheep and goats
Iker Sevilla, Joseba M. Garrido, Marivi Geijo and Ramon A. Juste
Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER), Berreaga, 1, 48160 Derio, Bizkaia, Spain.
BMC Microbiology, 2007, 7:18


ESTUDIOS
139
Abstract
Mycobacterium avium subsp. paratuberculosis (Map) causes paratuberculosis in
animals and is suspected of causing Crohn’s Disease in humans. Characterization of
strains led to classify paratuberculosis isolates in two main types, cattle type strains,
found affecting all host species, and sheep type strains, reported affecting mainly sheep.
In order to better understand the epidemiology of paratuberculosis a large set of Map
isolates obtained from different species over the last 25 years have been characterized.
Five-hundred and twenty isolates from different hosts (cattle, sheep, goats, bison, deer
and wild boar) and origins had been cultured and typed by IS1311 restriction-
endonuclease-analysis. Two-hundred and sixty-nine isolates were further characterized
by pulsed-field gel electrophoresis (PFGE) using SnaBI and SpeI endonucleases.
Differences in strain isolation upon various media conditions were also studied.
All bovines, 4 and 26% of Spanish sheep and goats, respectively, and the deer and
wild boar studied, carried IS1311-Cattle type strains. IS1311-Sheep type encompassed
96% and 74% of Spanish sheep and goats, and all three Portuguese sheep. Thirty-seven
distinct multiplex PFGE profiles were found, giving 32 novel profiles. Profiles 2-1 and
1-1 accounted for the 85% of cattle isolates. Ten distinct profiles were detected in
Spanish sheep, none of them with an incidence higher than 25%. Profile 16-11 (43%)
and another three profiles were identified in Spanish caprine cultures. The hierarchical
analysis clustered all profiles found in cattle, “wild” hosts and some small ruminants
within the same group. The other group included 11 profiles only found in Spanish
sheep and goats, including Spanish pigmented profiles. Differences in growth
requirements associated with isolate genotype were observed.
Cattle in Spain are infected with cattle type strains, while sheep and goats are
mainly infected with sheep type strains. Although 7H9 broth based culture media seem
to broadly cover the growth requirements of most Map strains, the use of various solid
media is recommended to reduce any recovery biases. High genetic homogeneity of
isolates from cattle, and heterogeneity of those from sheep and goats have been
detected.
Background
Mycobacterium avium subsp. paratuberculosis (Map) is an obligate pathogen
causing paratuberculosis in ruminants and other animal species, and suspected of being
responsible for Crohn’s Disease in humans. Paratuberculosis causes significant
economic losses to farmers and is becoming prevalent among livestock populations
world-wide (Kennedy y cols. 2001). Strategies for paratuberculosis prevention and
control require an understanding of the epidemiology of the disease. In particular, strain

ESTUDIO II
140
genotyping is a valuable tool for epidemiological tracing of pathogenic micro-
organisms.
Map isolates are generally identified by acid fast staining, the size of the bacilli,
extremely slow growth, dependence on mycobactin, and the presence of several copies
of the IS900 element in its genome. However, the description of some Map strains that
are not dependent on mycobactin (Aduriz y cols. 1995a, Motiwala y cols. 2004) and the
presence of IS900-like insertion sequences in bacteria other than Map (Cousins y cols.
1999, Naser y cols. 1999, Englund y cols. 2002, Motiwala y cols. 2004) mean that more
reliable methods of characterization would be valuable. The most commonly used
typing method is the analysis of restriction fragment length polymorphism using IS900
specific probes (IS900-RFLP). IS900-RFLP classified isolates from cattle, sheep and
goats into two major groups using BstEII endonuclease (Collins y cols. 1990a). These
were called sheep (S) and cattle (C) types because of their apparent predominance in
those host species. An intermediate (I) group was also detected in two sheep from
Canada. S strains are difficult to culture (Stamp y cols. 1954, Whittington y cols.
1999c), making further strain classification difficult. BstEII-C or cattle types have been
described in cattle from New Zealand, Australia, the USA, Denmark, the Czech
Republic, Slovakia, Hungary, Germany, France, Morocco, the United Kingdom,
Austria, Italy, the Netherlands, Slovenia, Spain, Sweden, Argentina and Norway
(Whipple y cols. 1990, Collins y cols. 1990a, Thoresen y cols. 1994b, Pavlik y cols.
1995, Bauerfeind y cols. 1996, Moreira y cols. 1999, Greig y cols. 1999, Pavlik y cols.
1999a, Cousins y cols. 2000, Whittington y cols. 2000b, Djonne y cols. 2005). These
RFLP types have also been found infecting sheep from Canada, the USA, the Czech
Republic, France, Greece, Australia, Spain and the United Kingdom (Whipple y cols.
1990, Collins y cols. 1990a, Thoresen y cols. 1994b, Pavlik y cols. 1995, Pavlik y cols.
1999a), and goats from New Zealand, Norway, Argentina, Germany, Denmark, Italy,
the United Kingdom, Australia and the USA (Whipple y cols. 1990, Collins y cols.
1990a, Thoresen y cols. 1994b, Bauerfeind y cols. 1996, Pavlik y cols. 1999a, Cousins y
cols. 2000, Djonne y cols. 2005). These strains have also been found in cervids from
New Zealand, the Czech Republic, the United Kingdom and Argentina (de Lisle y cols.
1993, Moreira y cols. 1999, Pavlik y cols. 1999a, Pavlik y cols. 2000a, Machackova y
cols. 2004), in lagomorphs from Scotland and the Czech Republic (Greig y cols. 1999,
Machackova y cols. 2004), in alpaca and rhinoceros from Australia (Cousins y cols.
2000) and in Czech moufflon and wild boar (Machackova y cols. 2004). In contrast, the
literature contains little evidence of BstEII-S strains and BstEII-I strains being as
distributed in animal species: they have been reported in sheep from Canada, New
Zealand, the Faroe Islands, South Africa, Morocco and Australia (Collins y cols. 1990a,
de Lisle y cols. 1992, Thoresen y cols. 1994b, Bauerfeind y cols. 1996, Choy y cols.
1998, Cousins y cols. 2000, Whittington y cols. 2000b), in goats from New Zealand and
the Czech Republic (Collins y cols. 1990a, Pavlik y cols. 1995), cattle from the Czech

ESTUDIOS
141
Republic (Pavlik y cols. 1995), and deer from New Zealand and the Czech Republic (de
Lisle y cols. 1993, Machackova y cols. 2004). IS1311 PCR-restriction endonuclease
analysis (REA) (Marsh y cols. 1999) led to the identification of S strains in Australian
and Icelandic sheep and cattle (Whittington y cols. 2001d) and Spanish sheep and goats
(Sevilla y cols. 2005). A recent study based on IS1311 PCR-REA and other two PCR
methods reported infection of Spanish goats and bullfighting cattle with S (Type I/III)
strains (de Juan y cols. 2006a).
Another promising technique for typing this parasitic bacterium is pulsed-field gel
electrophoresis of digested genomic DNA (PFGE). It has been useful for typing other
important mycobacteria including M. avium and M. tuberculosis (Levy-Frebault y cols.
1989, Coffin y cols. 1992, Feizabadi y cols. 1996, Kim y cols. 1997, Singh y cols.
1999). Pigmented and non-pigmented isolates from a wide variety of hosts and origins
were analyzed by SnaBI-SpeI PFGE in an effort to standardize the technique for
paratuberculosis strain typing (Stevenson y cols. 2002). Subsequently, caprine isolates
from Spain, Scotland and Norway were analyzed (de Juan y cols. 2005). Thus, 26
multiplex profiles have been described and clustered into 3 major types. Other
genotyping techniques have been investigated in an attempt to improve upon the
discriminatory power of RFLP and PFGE (Bull y cols. 2000, Pillai y cols. 2001,
Motiwala y cols. 2003, Overduin y cols. 2004, Amonsin y cols. 2004). However, some
of them do not provide enough additional information and others need to be validated
with larger numbers of strains already characterized by RFLP and/or PFGE and from
diverse host and geographic origins to assess their value for further epidemiological
studies on paratuberculosis.
There have been few studies reporting molecular characterization of Spanish
paratuberculosis isolates (Sevilla y cols. 2005, de Juan y cols. 2005, de Juan y cols.
2006a). Our aim in this work was to characterize by IS1311 PCR-REA and PFGE
typing a large set of Map isolates obtained at Neiker from different species over the last
25 years to serve as a basis for a better understanding of the epidemiology of
paratuberculosis.
Methods
Sources and growth of isolates
Feces and tissue samples from cattle, sheep and goats were collected for routine
paratuberculosis confirmation and for research purposes between 2000 and 2005.
Twelve samples from sheep and one from a goat date from 1984-1999, and isolates
obtained from these samples were maintained as glycerol stocks at -80ºC. Most strains
were from animals in northern regions of Spain, but strains from other Spanish regions
were also included (see Figure 1). Four-hundred and forty-four paratuberculosis
isolates were recovered from cattle (n=409; 178 herds), 27 from sheep (n=27; 15 flocks)

ESTUDIO II
142
and 21 from goats (n=19; 11 flocks). Four of the ovine isolates were yellow pigmented
(Figure 2). Map was also cultured from one deer (2 isolates from 2 different tissue
samples) and one wild boar. Eight bovine cultures from Argentina, one from France and
3 ovine cultures from Portugal were also included, as well as isolates from American
bison (n=3) and Indian sheep (n=2) and goats (n=8). Samples were processed for culture
of Map as described elsewhere (Aduriz y cols. 1995a). Bovine samples were cultured on
Herrold´s egg yolk medium supplemented with mycobactin J (2 mg/l in all media
containing the mycobactin) (Allied Monitor, Inc., Fayette, MO, USA) and sodium
pyruvate (HEY). To avoid any bias in the recovery of strains from cattle, additional
tubes of mycobactin J-supplemented Lowënstein-Jensen medium (LJ) were inoculated
with bovine samples collected after 2002. Some bovine samples were also used to
inoculate Middlebrook 7H11 supplemented with 1% Middlebrook OADC Enrichment
(Becton, Dickinson and Company, MD, USA) (7H11) and mycobactin J.
Ovine and caprine samples were cultured on HEY, on LJ and/or on 7H11
(alternatively with mycobactin J), depending on when they were collected. Some of
these cultures failed in the first attempt, and material stored at -20 ºC was subsequently
cultured again on Middlebrook 7H9 broth supplemented with OADC, 0.05% Tween 80
(Panreac Quimica SA, Barcelona, Spain) (7H9) with or without added mycobactin J,
and then sub-cultured on 7H11. Indian samples were initially grown on HEY without
added pyruvate and bison samples on LJ. Deer samples were cultured on HEY and LJ,
and wild boar samples on these two media and on 7H11 with mycobactin J.
Figure 1. Regions mentioned in the study are indicated as follows: An= Andalucia; Ar= Aragón; As= Asturias; BC= Basque Country; Can= Cantabria; Cat= Cataluña; CL= Castilla y León; CM= Castilla-La Mancha; Ex= Extremadura; Ga= Galicia; IB= Balearic Islands; Ma= Madrid; Na= Navarra.
Figure 2. Pigmented colonies of Mycobacterium avium subsp. paratuberculosis on a Lowënstein-Jensen slant, isolated from the yellow-colored intestines of one sheep with clinical paratuberculosis from the Basque Country (Spain).

ESTUDIOS
143
Cultures for PFGE
High quality DNA could not be prepared from all samples due to loss of viability of
some of the older isolates. Sufficient growth was obtained for PFGE typing with 232
isolates from cattle (n=228; 131 herds), 17 isolates from sheep (n=17; 10 flocks), seven
isolates from goats (n=6; 5 flocks), two from deer, one from wild boar, three from
American bison and seven from Indian small ruminants (two from sheep and five from
goats).
Nearly 85% of the bovine isolates were from Holstein (dairy) cattle, 10% from
Limousin (beef) cows, 3% from Pyrenean (beef) cows, and 2% from Lidia (bullfighting)
cattle. Spanish sheep were Latxa (dairy; n=14), Rasa-Aragonesa (meat; n=2) and
Carranzana (dairy; n=1). Goat breeds were Payoya (dairy/meat; n=3), Murciano-
Granadina (dairy/meat; n=3) and Saanen (dairy; n=1). The Indian sheep
(Muzzafarnagari) and goats (Barbari) were all meat breeds.
Isolate identification: IS900 PCR and Locus 251 PCR
Map was identified on the basis of time of incubation to visible colonies, colony and
bacillus morphology including acid fastness, and mycobactin dependence on egg-based
media. Isolates were screened for the presence of IS900 and Locus 251 (Bannantine y
cols. 2002a) as follows. The method described by Garrido et al. (Garrido y cols. 2000b)
was used to extract DNA from single colonies or if there was no visible growth on solid
media from broth culture pellets obtained by centrifugation of 5 ml at 2000 x g for 15
min. A GeneAmp 9600 PCR system (Applied Biosystems, Foster City, CA, USA) with
specific primers were used for amplification under standardized conditions as described
previously (Garrido y cols. 2000b, Bannantine y cols. 2002a). PCR products were
subjected to electrophoresis in 2 % (w/v) agarose gels and stained with ethidium
bromide to reveal DNA bands.
IS1311 PCR-REA
A segment of IS1311 element was amplified and digested with HinfI and MseI
(MseI only to rule out presence of Mycobacterium avium subsp. avium) endonucleases
as described by Marsh et al. (Marsh y cols. 1999). DNA fragments were separated by
electrophoresis in 4% (w/v) agarose gels stained with ethidium bromide. Isolates were
classified as cattle (C), sheep (S) or bison (B) Map types according to previously
published criteria (Marsh y cols. 1999, Whittington y cols. 2001b).
SnaBI-SpeI PFGE analysis
A protocol described previously (Stevenson y cols. 2002) was used to prepare
mycobacterial DNA. Ten ml of 7H9 broth was inoculated and incubated at 37 ºC in a
static 25 cm3 cell culture flask (Corning Inc., Corning, NY, USA). When sufficient
growth was observed at the bottom of the flask, the culture was homogenized by

ESTUDIO II
144
vigorous shaking and the optical density measured using a Densimat (Bio-Mérieux,
Marcy L’Etoile, France). An appropriate volume of culture was centrifuged at 2000 x g
for 15 min, and the pellet washed and resuspended in modified spheroplasting buffer to
obtain a bacterial suspension of approximately 6 x 109 bacteria/ml. This suspension was
heated to 55 ºC and mixed with an equal volume of molten 1.5 % (w/v) low-melting-
point agarose (Biorad, Hemel Hempstead, Hertfordshire, United Kingdom) in 50 mM
EDTA, then poured into plug molds (Biorad), and allowed to solidify at 4 ºC. The plugs
were incubated in a Tris-EDTA (10 mM Tris-HCl and 1 mM EDTA; pH 8) lysis
solution containing 1.5 mg/ml lysozyme at 37 ºC for 16-20 h. They were then placed
with ESP solution (0.5 M EDTA, 1 % (w/v) lauryl sarcosine and 1.5 mg/ml proteinase
K) and incubated at 55 ºC for at least 3 days. The ESP solution was discarded and the
agarose plugs were washed in pH 8 Tris-EDTA 6 times for 20 min each. The plugs were
equilibrated in the restriction buffer supplied by the enzyme manufacturer (1 x NE
buffer and 0.1 mg/ml bovine serum albumin; New England Biolabs, Inc., Beverly, MA,
USA) and then subjected to digestion with 20 U of the appropriate endonuclease in
fresh buffer at 37 ºC (SnaBI, 3 h; SpeI, overnight). The plugs were placed in pH 8 Tris-
EDTA and loaded into a 1 % (w/v) pulsed field certified agarose gel (Biorad). Lambda
midrange II PFG markers (New England Biolabs) and the type strain ATCC 19698 were
included in every gel. Electrophoresis was carried out in a CHEF-DRII apparatus
(BioRad). SnaBI restricted DNA was separated with initial and final switches of 6.8 and
26.3, respectively, and for separation of SpeI digests the values were 2.2 and 35.4,
respectively. The electrophoresis time for SnaBI was 28 h to allow rapid screening of
isolates, and samples with different and/or doubtful patterns were then electrophoresed
for 40 h under the same conditions. Electrophoresis for SpeI gels was for 23 h. The
gradient applied was always 6 V/cm. DNA bands were visualized and photographed in a
Fluor-S MultiImager (BioRad) after ethidium bromide staining. Nomenclature of novel
patterns was as recommended elsewhere (Stevenson y cols. 2002, de Juan y cols. 2005).
The first number refers to the SnaBI profile and the second to the SpeI profile, giving
consecutive numbers to new profiles. Both molecular size markers and ATCC 19698
SnaBI and SpeI profiles were used as between-gel references for band identification.
Phylogenetic analysis of PFGE profiles and estimation of genetic diversity
PFGE images were analyzed with the Quantity One Software package version 4.5
(BioRad). Similarities between different multiplex profiles were calculated as Jaccard
coefficients on the basis of band presence/absence scores with the SPSS software for
Windows version 11.0 (SPSS Inc., Chicago, IL, USA). An agglomerative hierarchical
cluster analysis of the data using the unweighted pair group method with arithmetic
averages was used to construct a dendrogram. Simpson’s Index of Diversity was
calculated as follows to compare the genetic diversity of the isolates between host
species and to asses the discriminatory power of the SnaBI-SpeI-PFGE typing:

ESTUDIOS
145
1-[Σ(no. of isolates with a particular multiplex profile/total no. of isolates)2]
Results
All bovine isolates grew in 2-4 months on HEY, and some cultured after 2002 also
grew on LJ or in 7H11 with mycobactin J. Ovine isolates grew in 3-7 months on LJ,
7H11 (with or without mycobactin J) and/or 7H9 (with or without mycobactin J), but
only from tissue samples. None of the ovine samples used to inoculate HEY produced
Map colonies. Goat isolates grew in 2-7 months on the same media as sheep isolates,
and some of them were also successfully isolated on HEY.
Mycobacterial cells were not successfully propagated from several cases in small
ruminants, although the presence of Map was identified on the basis of acid fastness,
IS900 and locus 251 PCR results, and IS1311 PCR-REA result. Indian isolates grew in
2-4 months on HEY with no added pyruvate. Deer isolates grew in 3 months on HEY
and LJ, and wild boar isolates grew in 2-3 months on HEY and 7H11 with mycobactin
J. The isolation media for cultures typed by PFGE are given in Table 2. Subcultures in
liquid medium of C and B strains grew faster than those of S strains (data not shown).
IS900 PCR and Locus 251 PCR
PCR amplification detected both IS900 and Locus 251 in all 520 isolates used in the
study. These findings and the phenotypic characteristics (see “Methods” section)
allowed all bacterial isolates used to be identified as Map.
IS1311 PCR-REA
The IS1311 insertion sequence was successfully amplified from all isolates.
Restriction analysis of amplicons assigned each isolate to one of the 3 strain types
previously described by this method.
All bovine isolates, as well as the deer and wild boar isolates, exhibited band
patterns corresponding to C type strains (Table
1, Figure 3). In contrast, 96.3% and 73.7% of
Spanish sheep and goats, respectively, gave
isolates with banding patterns corresponding to
the S type strains; all sheep isolates from
Portugal were also classified in this group. The
other Spanish sheep (3.7%) and goats (26.32%)
were infected by C type strains. Interestingly,
two goat flocks were infected by both types,
some goats carrying a C strain and others an S
strain. All isolates from Indian sheep and goats
and from American bison were typed as B
0
20
40
60
80
100
Sheep Goat Cattle Wildlife
typ
e p
rop
ort
ion
S TYPE C TYPE
Figure 3. Graphic representation showing the clear polarization of C and S strain types found in Spanish bovine, ovine, caprine and “wild” species.

ESTUDIO II
146
strains.
Table 1. IS1311 classification of Map strains. The 520 Map isolates from different host species classified as C (cattle), S (sheep) or B (bison) type according to IS1311 PCR-REA. Numbers of isolates from particular regions in Spain are specified in brackets following the region abbreviation. Abbreviations: An= Andalucía; Ar= Aragón; As= Asturias; BC= Basque Country; Can= Cantabria; Cat= Cataluña; CL= Castilla y León; CM= Castilla-La Mancha; Ex= Extremadura; Ga= Galicia; IB= Balearic Islands; Ma= Madrid; Na= Navarra.
Geographic origin Host sp. no. of isolates
no. of animals(%)
no. of herds/flocks(%)
IS1311 type
Spain ·Ar (19), As (9), BC (228), Can (22), CL (92), CM (2), Cat (9), Ex (1), Ga (13), Ma (4), Na (46)
Cattle 444 409 (100.0) 178 (100.0) C
·Ar Sheep 1 1 (3.7) 1 (6.67) C ·An (1), Ar (2), BC (17), Na (5) Sheep 26 26 (96.3) 14 (93.33) S ·An (4), BC (3; 1 goat) Goat 7 5 (26.32) 4 (36.36) C ·An (11), CL (1), Ga (1), IB (1) Goat 14 14 (73.68) 9 (81.82) S ·CM Deer 2 2 (100.0) 1 (100.0) C ·CM Wild boar 1 1 (100.0) 1 (100.0) C Argentina Cattle 8 8 (100.0) - C France ·Lys Cattle 1 1 (100.0) 1 (100.0) C Portugal ·Trás-os-Montes Sheep 3 3 (100.0) 2 (100.0) S India ·Mathura Sheep 2 2 (100.0) 1 (100.0) B ·Farah Goat 8 8 (100.0) 1 (100.0) B USA ·Montana Bison 3 3 (100.0) 1 (100.0) B Total 520 483
SnaBI-SpeI PFGE analysis
Restriction endonucleases SnaBI and SpeI differentiated 26 and 24 pulsotypes,
respectively (Figures 4 and 5). Thus, a total of 37 different multiplex PFGE profiles
were identified among the 269 Map isolates analyzed.

ESTUDIOS
147
Twenty-two of the patterns identified with SnaBI (20-41) and 19 of those obtained
with SpeI (14-32) have not been described previously; the two endonucleases together
1 5 10 12 14 15 16 19 20 25 26 27 30 1 31 11 17 18 21 22 23 24 28 29 32 M W
M W
M W
M W
cattle type profiles sheep type profiles
48.5
97
145.5
194 242.5
339.5
Figure 5. SpeI restriction profiles. Photographs of two gels comprising the 24 different SpeI pulsotypes detected. Numbers above the lanes correspond to the PFGE nomenclature. Arrows and asterisks indicate differences (band loss and new band, respectively) between type 1 and cattle type profiles, and between type 11 and sheep type profiles. MW= molecular weight standards, in Kilobases (Kb).
cattle type profiles sheep type profiles
37 40 16 25 26 30 34 35 36 38 39 33 M W 41
M W 1 2 15 20 21 22 23 24 27 28 29 31 32
M W
M W
48.5
97
145.5
194
242.5
291
339.5
388
Figure 4. SnaBI restriction profiles. Photographs of two gels comprising the 26 different SnaBI pulsotypes detected. Numbers above the lanes correspond to the PFGE nomenclature. Arrows and asterisks indicate differences (band loss and new band, respectively) between type 1 and cattle type profiles, and between type 16 and sheep type profiles. MW= molecular weight standards, in Kilobases (Kb).

ESTUDIO II
148
giving 32 novel PFGE multiplex profiles in our collection of isolates (see Table 2 and
Figure 6).
Twenty-three different multiplex profiles were detected in Spanish cattle, 10 in
Spanish sheep and 4 in Spanish goats. Profiles 2-1 and 1-1 included 52.16 and 32.33%
of cattle isolates, found in 58.02 and 30.53% of herds analyzed, respectively. The
calculated incidence for each of the other profiles identified in cattle was lower than
1%, except for profile 23-16 which included 3% of bovine isolates. In Spanish sheep,
strain type 39-17 was the most frequently found with four isolates detected in 40% of
ovine flocks. Four pigmented sheep isolates exhibited novel restriction patterns (26-24
and 25-23), and accounted for the 23.5% of sheep isolates. The other types observed in
Spanish sheep appeared only once or twice. Profile 16-11 was found in three goats, and
profiles 2-1, 15-1 and 34-28 were identified in a single animal each. The two isolates
from deer had profile 37-1, and the wild boar isolate was typed as 2-1. All the Indian
isolates gave the same multiplex profile regardless of their host species. All bison
isolates from USA were 2-1.
Several herds/flocks carried strains of different profiles: up to 3 in 3 bovine herds
and 3 sheep flocks (34 bovine herds, 4 sheep flocks and 1 goat flocks gave more than
one isolate). Eleven bovine and one goat herds gave isolates with two different profiles.
The other flocks/herds each gave isolates with a single multiplex pattern within the
same herd/flock. PFGE typing results according to host species breeds are reported in
Table 3.
Table 2. SnaBI-SpeI PFGE classification of Map strains. Details of SnaBI-SpeI multiplex profiles of the 269 Map isolates studied by PFGE: geographic origin, number of isolates, number of herds/flocks of the indicated host species, primary isolation media and IS1311 PCR-REA type. Profiles in boldface correspond to pigmented isolates. *novel profiles found in this study. Abbreviations: An= Andalucía; Ar= Aragón; As= Asturias; BC= Basque Country; Can= Cantabria; Cat= Cataluña; CL= Castilla y León; CM= Castilla-La Mancha; Ex= Extremadura; Ga= Galicia; IB= Balearic Islands; Ma= Madrid; Na= Navarra. HEY= Herrold's Egg Yolk Medium with mycobactin J; LJ= Löwenstein-Jensen Medium with mycobactin J; 7H11= OADC supplemented Middlebrook 7H11 Medium without mycobactin J; 7H11*= OADC supplemented Middlebrook 7H11 Medium with mycobactin J; 7H9+7H11= OADC supplemented Middlebrook 7H9 Broth without mycobactin J, subcultured on 7H11; 7H9*+7H11= OADC supplemented Middlebrook 7H9 Broth with mycobactin J, subcultured on 7H11.
Country PFGE profile Locations Host sp
No. of isolates (%)
No. of herds or flocks (%)
Other types in same herd/flock (No. of herds/flocks) Primary Isolation
IS1311 type
Spain 1-1 BC, As, Ar, Na, Cat
Cattle 75 (32.33) 40 (30.53) 1-20 (1), 2-1 (6), 22-1 (1), 23-16 (1), 28-30 (1), 31-1 (1)
HEY, LJ, 7H11* C type
*1-10 BC Cattle 1 (0.43) 1 (0.76) HEY C type
*1-20 Na Cattle 1 (0.43) 1 (0.76) 1-1 (1), 31-1 (1) HEY C type
*1-27 BC Cattle 3 (1.29) 1 (0.76) HEY C type 2-1 BC, As, Can,
Ga, CL, Ar, Na, Cat, CM
Cattle 121 (52.16) 76 (58.02) 1-1 (6), 2-10 (2), 2-15 (1), 2-31 (1), 22-1 (1), 23-16 (2), 28-30 (1)
HEY, LJ, 7H11* C type
*2-5 As Cattle 1 (0.43) 1 (0.76) HEY C type 2-10 Can, CL, Ex Cattle 3 (1.29) 3 (2.29) 2-1 (2) HEY C type
*2-12 BC Cattle 1 (0.43) 1 (0.76) HEY C type
*2-14 Ga Cattle 1 (0.43) 1 (0.76) HEY C type *2-15 BC, Ma Cattle 3 (1.29) 3 (2.29) 2-1 (1), 23-16 (1) HEY C type
*2-25 CL Cattle 2 (0.86) 1 (0.76) HEY C type
*2-31 CL Cattle 1 (0.43) 1 (0.76) 2-1 (1) HEY C type *20-27 BC Cattle 2 (0.86) 2 (2.29) HEY C type

ESTUDIOS
149
*21-1 Can Cattle 1 (0.43) 1 (0.76) HEY C type
*22-1 Ar Cattle 1 (0.43) 1 (0.76) 1-1 (1), 2-1 (1) HEY C type
*23-16 BC Cattle 7 (3.02) 7 (5.34) 1-1 (1), 2-1 (2), 2-15 (1) HEY C type *24-19 Ma Cattle 1 (0.43) 1 (0.76) HEY C type
*27-26 BC Cattle 1 (0.43) 1 (0.76) HEY C type
*28-30 BC Cattle 1 (0.43) 1 (0.76) 1-1 (1), 2-1 (1) HEY C type *31-1 Na Cattle 1 (0.43) 1 (0.76) 1-1 (1), 1-20 (1) HEY C type
*32-1 BC, Na Cattle 2 (0.86) 2 (1.53) HEY C type
*33-1 BC Cattle 1 (0.43) 1 (0.76) HEY C type *40-1 CL Cattle 1 (0.43) 1 (0.76) HEY C type
2-1 Ar Sheep 1 (5.88) 1 (10.0) LJ C type
*25-23 Ar Sheep 1 (5.88) 1 (10.0) LJ S type *26-24 BC, Na Sheep 3 (17.65) 2 (20.0) LJ, 7H9*+7H11 S type
*30-11 BC Sheep 2 (11.76) 2 (20.0) 39-17 (2), 39-21 (1), 41-32 (1) 7H11 S type
*35-29 BC Sheep 1 (5.88) 1 (10.0) 7H11*, 7H9+7H11 S type *36-18 BC Sheep 1 (5.88) 1 (10.0) 38-22 (1), 39-17 (1) 7H11 S type
*38-22 BC Sheep 1 (5.88) 1 (10.0) 36-18 (1), 39-17 (1) 7H11 S type
*39-17 BC, Na Sheep 4 (23.53) 4 (40.0) 30-11 (2), 36-18 (1), 38-22 (1), 39-21 (1), 41-32 (1)
7H11 S type
*39-21 BC Sheep 2 (11.76) 2 (20.0) 30-11 (1), 39-17 (1) 7H11 S type *41-32 BC Sheep 1 (5.88) 1 (10.0) 30-11 (1), 39-17 (1) 7H11 S type
2-1 BC Goat 2 (28.57) 1 (20.0) HEY, 7H11* C type
15-1 An Goat 1 (14.29) 1 (20.0) 34-28 (1) 7H11*, 7H9+7H11 C type 16-11 CL, An, IB Goat 3 (42.86) 3 (60.0) LJ, 7H9+7H11 S type
*34-28 An Goat 1 (14.29) 1 (20.0) 15-1 (1) 7H9+7H11 S type
*37-1 CM Deer 2 (100.0) 1 (100.0) HEY C type 2-1 CM Wild Boar 1 (100.0) 1 (100.0) HEY, 7H11* C type
India *29-1 Mathura Sheep 2 (100.0) 1 (100.0) HEY B type
*29-1 Farah Goat 5 (100.0) 1 (100.0) HEY B type
USA 2-1 Montana Bison 3 (100.0) 1 (100.0) LJ B type
In five cases, more than one isolate cultured from the same animal was available.
Two cows, one goat and one deer yielded two isolates each one but the restriction
patterns of isolates from the same animal were identical. In contrast, an isolate cultured
in 2002 from the feces of one Holstein bull was of 1-1 type, whereas the two isolates
cultured in 2003 from the small intestine and mesenteric lymph nodes of the same
animal, were of 2-1 and 28-30 profiles, respectively.
Table 3. Distribution of Map genotypes among the different breeds of host species. Number of isolates, animals and farms in Spain giving isolates of each SnaBI-SpeI multiplex profile/IS1311 type according to the host species breed. Percentages are in brackets.
CATTLE
(232 isolates & 228 animals) SHEEP
(17 isolates & 17 animals) GOAT
(7 isolates & 6 animals)
Breed Holstein Limousin Pyrenean Bullfight Latxa Rasa-
Aragonesa Carranzana Payoya Murciano-Granadina Saanen
Isolates 197 (84.91) 23 (9.91) 7 (3.02) 5 (2.16) 14 (82.35) 2 (11.76) 1 (5.89) 3 (42.86) 3 (42.86) 1 (14.29) Animals 193 (84.65) 23 (10.09) 7 (3.07) 5 (2.19) 14 (82.35) 2 (11.76) 1 (5.89) 3 (50.0) 2 (33.33) 1 (16.67)
Herds/Flocks 100 (76.34) 22 (16.79) 6 (4.58) 3 (2.29) 7 (70.0) 2 (20.0) 1 (10.0) 2 (40.0) 2 (40.0) 1 (20.0)
1-1 / C 68 (34.52) 3 (13.04) 4 (57.14) 1-10 / C 1 (14.29)
1-20 / C 1 (0.51)
1-27 / C 3 (1.52) 2-1 / C 103 (52.28) 15 (65.22) 2 (28.57) 1 (20.0) 1 (50.0) 2 (66.67)
2-5 / C 1 (0.51)
2-10 / C 1 (0.51) 2 (40.0) 2-12 / C 1 (0.51)
2-14 / C 1 (0.51)
2-15 / C 3 (1.52) 2-25 / C 2 (40.0)

ESTUDIO II
150
2-31 / C 1 (0.51)
15-1 / C 1 (33.33)
20-27 / C 2 (8.70) 21-1 / C 1 (0.51)
22-1 / C 1 (0.51)
23-16 / C 6 (3.05) 1 (4.35) 24-19 / C 1 (4.35)
27-26 / C 1 (0.51)
28-30 / C 1 (0.51) 31-1 / C 1 (0.51)
32-1 / C 2 (1.02)
33-1 / C 1 (4.35) 40-1 / C 1 (0.51)
16-11 / S 1 (33.33) 1 (33.33) 1 (100.0)
25-23 / S 1 (50.0) 26-24 / S 3 (21.43)
30-11 / S 2 (14.29)
34-28 / S 1 (33.33) 35-29 / S 1 (100.0)
36-18 / S 1 (7.14)
38-22 / S 1 (7.14) 39-17 / S 4 (28.57)
39-21 / S 2 (14.29)
41-32 / S 1 (7.14)
Phylogenetic analysis of PFGE profiles and estimation of genetic diversity
Phylogenetic analysis of the multiplex profiles obtained produced a dendrogram
with two main branches (Figure 6). One includes all profiles found in cattle, deer, wild
boar, Indian small ruminants and American bison, for a total of 26 profiles. Previously
described profiles 1-1, 2-1, 2-5, 2-10 and 15-1, were part of this group. The other main
branch includes 11 profiles only found in sheep and goats from Spain, including the 16-
11 profile identified previously and both profiles detected in pigmented isolates (25-23
and 26-24). The discriminatory power for this PFGE strain typing method calculated as
Simpson’s Index of Diversity, taking into account all isolates and profiles described,
was 0.693. The index value was 0.621 for Spanish cattle, and 0.666 for Spanish goats,
but much higher, 0.865, for Spanish sheep.

ESTUDIOS
151
Discussion
Differences in the success of Map isolation and, more specifically, difficulties in
culturing this micro-organism from ovine specimens, have been repeatedly reported in
many countries (reviewed in reference (Whittington y cols. 1999c)). Some authors
suggest that different host species preferentially carry different strains, and the differing
cultivation requirements of these strains causes the divergent efficacy of isolating the
organism (Collins y cols. 1990a, Juste y cols. 1991b, Aduriz y cols. 1995a, Bauerfeind y
cols. 1996). In contrast, other researchers did not report any links between host species,
strain type and/or culture characteristics (Thoresen y cols. 1994b, Pavlik y cols. 1999a,
Stevenson y cols. 2002). Our study does not resolve this issue because we did not use
the same methods for all cultures, and culture attempts were carried out with the
Figure 6. Dendrogram showing the similarities between multiplex PFGE profiles of isolates in the collection studied. The agglomerative hierarchical cluster analysis of the data using the unweighted pair group method with arithmetic averages divided strain profiles in two main branches, cattle type profiles and sheep type profiles. Profiles in boldface correspond to pigmented isolates. *novel profiles found in this study.
Rescaled Distance Cluster Combine
0 5 10 15 20 25
SnaBI-SpeI+---------+---------+--------- +---------+--------- +
1-1 __
*40-1 __
*31-1 __
*1-27 __
*1-20 ____
*1-10 __ _
2-10 __ _
2-1 __ _
*32-1 __ _
*2-25 __ _
*20-27 ____
*22-1 ____
*2-14 ____
*29-1 ____
*33-1 __ ___
*21-1 ____ _
*2-12 ____ _
*2-5 ______
*27-26 __ _
*2-31 ________
*28-30 ______ ___
*2-15 ______ _ _
15-1 ________ _
*37-1 ____ _________________________________________
*23-16 __________ _
*24-19 __________ _
16-11 ______ _
*30-11 __ _ _
*39-17 __ ___ _
*39-21 ____ _ _ _
*41-32 __ ___ ___ _
*38-22 ____ _ ___ _
*34-28 ________ _ _____ _
*36-18 __________ _ ___ _
*26-24 ____________ _ _________________________________
*25-23 ________________ _
*35-29 __________________
CATTLE TYPE PROFILES
SHEEP TYPE PROFILES
Rescaled Distance Cluster Combine
0 5 10 15 20 25
SnaBI-SpeI+---------+---------+--------- +---------+--------- +
1-1 __
*40-1 __
*31-1 __
*1-27 __
*1-20 ____
*1-10 __ _
2-10 __ _
2-1 __ _
*32-1 __ _
*2-25 __ _
*20-27 ____
*22-1 ____
*2-14 ____
*29-1 ____
*33-1 __ ___
*21-1 ____ _
*2-12 ____ _
*2-5 ______
*27-26 __ _
*2-31 ________
*28-30 ______ ___
*2-15 ______ _ _
15-1 ________ _
*37-1 ____ _________________________________________
*23-16 __________ _
*24-19 __________ _
16-11 ______ _
*30-11 __ _ _
*39-17 __ ___ _
*39-21 ____ _ _ _
*41-32 __ ___ ___ _
*38-22 ____ _ ___ _
*34-28 ________ _ _____ _
*36-18 __________ _ ___ _
*26-24 ____________ _ _________________________________
*25-23 ________________ _
*35-29 __________________
CATTLE TYPE PROFILES
SHEEP TYPE PROFILES

ESTUDIO II
152
concrete objective of isolating Map from paratuberculosis-affected animal specimens
received. Nevertheless, our results are consistent with other reports on the comparison
of different media for Map culture (Whittington y cols. 1999c, de Juan y cols. 2006b),
and confirm the existence of different culture requirements according to Map type.
IS1311 PCR-REA typing with our collection of bovine isolates — presumably the least
biased due to its size and to the use of various culture media — confirms the finding of
a previous study (Sevilla y cols. 2005): Holstein, Limousin and Pyrenean cattle in Spain
are infected with C type strains. These strains are more easily isolated on HEY than on
other solid media. This strain type was found only in one of 27 isolates from Spanish
sheep, the others, like those from Portugal (n=3) being S type. This indicates that sheep
in Spain and probably also in Portugal are mostly infected with S strains. Some of these
were isolated with ease on 7H11 without mycobactin J, others on the same medium with
added mycobactin or on LJ with longer incubation periods, and some grew in 7H9
culture flasks (with or without mycobactin J). We did not obtain any growth of such
strains on HEY, but not all samples were used to inoculate HEY slants. The single C
type strain isolated from sheep dates from 1993 and was cultured only on LJ.
Consequently, our study does not indicate whether C strains can be readily isolated from
ovine samples on HEY, but other authors report that this is feasible (Collins y cols.
1990a, Pavlik y cols. 1995). Choy et al. (Choy y cols. 1998) report that it would appear
that the IS900-RFLP type is not correlated with the ability to culture isolates, as
uncultivable sheep isolates from intestinal-mucosal homogenates in Australia had the
same pattern (S1) as those from New Zealand grown on HEY (Collins y cols. 1990a).
Other factors, including mycobacterial concentration, contamination of inocula,
susceptibility to hexadecyl-pyridinium-chloride (or other decontaminants) and/or
different phenotypic stages could also contribute to the difficulty of culturing these
strains. Only 26% of the Spanish goats analyzed were infected with strains belonging to
the C type: the other 74 % had sheep (S) type paratuberculosis. The cultural
characteristics of caprine isolates depended on the strain type: C strains had bovine-like
characteristics and S strains had ovine-like characteristics. This strain type distribution
in goats disagrees with the findings of other authors in Spain and other countries
(Thoresen y cols. 1994b, Djonne y cols. 2005, de Juan y cols. 2005) who report a very
low incidence of S strains. The proportion of S type isolates previously calculated (de
Juan y cols. 2006a) for Spanish goats (24.7% of goat isolates and 39.1% of flocks
studied) is lower than that detected in our study (73.7% of animals and 81.8% of
flocks). These differences may be due to a culture media bias, but epidemiological
reasons can not be ruled out. We found several goat flocks infected with both C and S
types, although this was not the case for all farms or animal species studied. Our
findings suggest that goats are highly susceptible to C, S and B Map strains.
Although we describe numerous multiplex PFGE profiles, nearly the 85% of bovine
isolates belonged to only two different profiles, 1-1 and 2-1. The homogeneity of

ESTUDIOS
153
paratuberculosis isolates, especially bovine isolates, has been described before
(Bauerfeind y cols. 1996, Feizabadi y cols. 1997b, Cousins y cols. 2000, Whittington y
cols. 2000b, Motiwala y cols. 2003, Djonne y cols. 2005). Apparently, particular Map
strains have developed the ability to infect a wide range of types of cattle, and
international trading has favored the worldwide spread of these strains. This may be
accentuated by the genetic homogeneity of cattle as a result of genetic improvement
programs and international trading. Indeed, bovine genotypes highly susceptible to
certain strains may have spread across the world, and consequently the contact between
other host species and these strains would have increased proportionally.
We noted slight but interesting differences between strain types found in different
bovine breeds coming from the same geographic area. Holstein cattle had the largest
number of different profiles (n=18), but this is unsurprising because it was the most
numerous breed. More remarkable is the absence from Holsteins of 5 profiles detected
in other breeds. Limousine cattle carried more 2-1 type and less 1-1 type than Holstein
cattle. In contrast, the 1-1 strain was much more prevalent in Pyrenean cows. Cattle
destined to bullfighting came from a less represented geographic area. With the
exception of one 2-1 type, strains isolated from these animals (n=5) showed rare
profiles, suggesting a separate epidemiological context. This is in agreement with the
results of the study by de Juan et al. (de Juan y cols. 2006a) reporting a high proportion
of Type I/III isolates in two bullfighting cattle farms. Type 2-10 detected in 2
bullfighting and 1 Holstein animal, has only previously been reported in one goat from
Norway (de Juan y cols. 2005). The novel profile 2-25 was only isolated from two
bullfighting animals. The differences of geographic origin of isolates may contribute to
the differences found between cattle breeds, especially in bullfighting cattle.
The seven farms affected with 23-16 strains, are all in the same geographic area,
some being only 30 km apart, and 5 of them also share the same veterinarian. These
strains were isolated over several years (2000-2003) indicating persistence in this
ecosystem. According to the phylogenetic analysis, this is one of the strains more
different from the others in the bovine type sub-cluster. We are investigating probable
links in food and water resources or historic introduction.
Ours is not the first description of mixed infection evidenced by isolation of
different strains from the same animal (Pavlik y cols. 1995, Cousins y cols. 2000,
Whittington y cols. 2000b), and could be similar to Mycobacterium avium polyclonal
infections in human patients (Arbeit y cols. 1993). The bull with 3 different strains
belonged to a Holstein herd with clinical paratuberculosis cases. The other two isolates
obtained from the herd were typed as 1-1 and 2-1. This is consistent with types found in
the bull, except for the extra profile (28-30) which was not isolated in any other animal.
Re-infection of the animal by different strain types is the most probable explanation,

ESTUDIO II
154
because they were isolated from different biological samples and on different sampling
dates.
Strain diversity in the sheep population was high. All the PFGE patterns of Spanish
sheep isolates were different from those found in cattle, except for one 2-1 strain from a
Rasa-Aragonesa sheep from Aragon. Pigmented isolates exhibited novel restriction
patterns designated 26-24 and 25-23. The first was found in Latxa flocks in Basque
Country and Navarra. These were not related to the Rasa-Aragonesa sheep from Aragon
infected with the other pigmented profile. This is the first report of pigmented strains in
Spain. Pigmented strains were less prevalent than the 39-17 strain, found in Basque
Country and Navarra. Three flocks in neighboring locations in the main area of
prevalence in sheep in Basque Country, gave 3 isolates per flock. For each of the 3
flocks all 3 isolates differed from each other, but 39-17 type was found in all of them.
The diversity index calculated for goats was low. However, including goat isolates
reported by de Juan et al. (de Juan y cols. 2005), the index is 0.817. All isolates we
cultured from goats, except one (34-28), have profiles that have previously been
reported in this host species. The 16-11 type was observed throughout Spain and in
numerous goat breeds. We detected the 15-1 type in goats in Cordoba (Andalucia), and
it is also detected in other breeds in Madrid. In conclusion, goats in Spain are affected
with a wide range of different strains, including C and S type strains.
Deer isolates had a novel profile (37-1) closely related to the 15-1 type (see Figure
1 and 6), previously found in the same area. Wild boar has carrion-eating behavior, and
is a possible reservoir or vector of Map, which has been isolated several times from this
species (Machackova y cols. 2003b, Machackova y cols. 2004, Alvarez y cols. 2005).
However, there is no evidence of diseased animals. The strain we isolated from wild
boar was 2-1 type, a strain type that is widely disseminated. Cultures from India and
bison isolates from USA showed different multiplex profiles, indicating they are
different strains.
The phylogenetic dendrogram we constructed for our collection of paratuberculosis
strains shows two distinct branches. One comprises isolates from cattle sheep, goats,
deer, wild boar, and bison, whereas the other includes isolates only found in sheep and
goats. This is in general agreement with previous work (Stevenson y cols. 2002, de Juan
y cols. 2005). Our first branch (isolates from cattle and other hosts) is the same as the
previously described Type II cluster, and IS900-RFLP Cattle types. The pigmented
isolates from Scotland and Faroe Islands were included in the Type I cluster. However,
type 16-11 found later in Spanish goats, was not included in Type I, and they were
considered to be Type III. Our pigmented strain profiles clustered together with 16-11
strains and other isolates only found in sheep and goats. It is not clear where these novel
profiles should be incorporated, but apparently they are more closely related to the 16-
11 profile than to those previously described for pigmented isolates. Other studies

ESTUDIOS
155
included both Sheep and Intermediate IS900-RFLP types within the same cluster,
composed of two different sub-clusters (Pavlik y cols. 1999a, Cousins y cols. 2000).
This seems to be a more accurate approach because Type III strains are very much
closer to Type I than to Type II strains. Our findings confirm the soundness of the
classification proposed by de Juan et al. (de Juan y cols. 2006a) into 2 major types
(Type I/III and Type II). These results suggest, however, that the original classification
into cattle (C) and sheep (S) strains is more descriptive, has historic priority, and is
substantially confirmed by the associations between bacterial type and host species
(Figure 3). It is also consistent with the widely accepted practice in bacteriology of
using as species name the Latin name of the species in which the parasite is found.
Consequently, we strongly encourage the use of this system with only two groups
corresponding to cattle and sheep.
The overall discriminatory power for this PFGE typing was not high but acceptable
(0.693). The problem with this method appears to be the low degree of diversity of the
bovine isolates rather than a true lack of power of the typing technique. Multiplex PFGE
seems to be one of the most robust and reliable typing methods, because it explores the
entire genome and only depends on the specificity of restriction endonucleases used.
Other approaches depend on the distribution of repetitive DNA fragments, random
amplification of certain fragments or on sequencing of variable repetitive regions; all
these approaches can suffer from problems of reproducibility or reliability. However,
PFGE is expensive and time consuming, in addition to the problem of obtaining
typeable cultures. This method was unable to distinguish the IS1311-B type found in
bison (PFGE 2-1) from other IS1311-C strains. The discrimination between IS1311-B
and C is important, because these B strains (from both India and USA) share the
phenotypic characteristic of preferring egg-based media without added pyruvate (Sevilla
y cols. 2005).
Conclusions
Improved liquid culture media (based on Middebrook 7H9) seem to be appropriate
for primary isolation, because they appear to provide the entire isolation requirement
spectrum of all strains. Nevertheless, we recommend the use of various solid media to
avoid biases in primary isolation, and subculture of liquid cultures (when used for
primary isolation) onto solid media to obtain pure colonies. Alternatively, PCR-based
differentiation of strain type before culture inoculation may help to decide the media to
be used.
High homogeneity of isolates from cattle, and heterogeneity of those from sheep and
goats have been detected. Overall typing may need to be improved and this could be
done by including an additional endonuclease or complementary techniques. PCR-REA
typing is a powerful technique because it can both confirm Map identity and provide

ESTUDIO II
156
clear microbiological and epidemiological information. Systematic use of the more
sophisticated PFGE method used does not seem to be beneficial: it provides genetic
information which does not appear to be clearly related to phenotypic properties but
rather to limited epidemiologic details.
Authors' contributions
IS carried out the laboratory and field work, compiled and analysed information and
data, and drafted the manuscript as part of his PhD dissertation. JMG participated in the
laboratory and field work, compiled information, coordinated the study and helped to
draft the manuscript. MG participated in the laboratory and field work. RAJ conceived
of and coordinated the study, and helped to draft the manuscript. All authors read and
approved the final manuscript.
Acknowledgements
We wish to thank the farmers and veterinarians who provided the samples used in
this study. J.M. Plazaola from the Diputación Foral of Gipuzkoa is acknowledged for
his help with data collection. Numerous samples from Castilla-León were provided by
the Pathology Unit, Faculty of Veterinary Medicine of the University of León (Spain)
headed by V. Pérez and J.F. García-Marín. Samples from wild boar and deer were sent
by the Instituto de Investigación en Recursos Cinegéticos (IREC) in Ciudad Real
(Spain) headed by C. Gortazar. Samples and cultures from Andalucía (Spain),
Argentina, India, Portugal and USA were kindly provided by S. Rodríguez, A. Verna,
S.V. Singh, A.C. Coelho and R. Whitlock, respectively. We are grateful to G. Aduriz
for isolation and information on some strains. We also thank A. Badiola, Z. Pérez, I. del
Pozo, M. Bascones, J.C. Ibabe and O. Aurtenetxe for their technical assistance. The
work of I. Sevilla was supported by two research fellowships from the Department of
Agriculture and Fisheries, the Department of Industry and the Department of Education,
Universities and Research of the Basque Government. This study was partially
supported with EU-FEDER funds.

ESTUDIO IV
Detection and pathological implications of the
presence of IS900 in human Mycobacterium
avium isolates
Iker Sevilla,1 Martha I. Murcia,2,3 Joseba M. Garrido,1 Douwe Bakker,4 Maria J.
García,2 Ramón A. Juste1
1 Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER), Berreaga, 1, 48160 Derio, Bizkaia, Spain.
2 Dept. Medicina Preventiva, Facultad de Medicina, Universidad Autónoma, c/ Arzobispo Morcillo s/n, 28029-Madrid, Spain.
3 Dept. Microbiología, Facultad de Medicina, Universidad Nacional, Carrera 30 No.45-03, Edificio 471, Bogotá, Colombia.
4 Department of Bacteriology and TSE's, CIDC-Lelystad, Edelhertweg 15, 8203 AA Lelystad, The Netherlands.
Processing submission to Journal of Clinical Microbiology


ESTUDIOS
159
Abstract
IS900 insertion sequence is thought to be specific of Mycobacterium avium subsp.
paratuberculosis. In the present work, the presence of IS900 in M. avium isolates from
HIV-infected human patients has been demonstrated. Future studies regarding Crohn’s
disease and M. avium subsp. paratuberculosis detection based on IS900 screening
should be reconsidered.
Introduction
Mycobacterium avium subsp. paratuberculosis (Map) is a mycobacteria that has a
wide host spectrum including humans. Map is mainly known for its role as the causative
agent of paratuberculosis in ruminants, a chronic inflammation of the intestines (Harris
y cols. 2001). Initially considered as a M. avium closely related species, it was described
as M. avium subspecies after the numerical taxonomic differentiation of M. avium
subsp. avium (Maa), M. avium subsp. silvaticum (Mas) and Map by Thorel et al.
(Thorel y cols. 1990). In a further effort on the clarification of the M. avium complex
(MAC) taxonomy, Maa designation was proposed to be reserved for bird type isolates,
while pig and human isolates were classified as the fourth subspecies hominissuis (Mah)
(Mijs y cols. 2002). In spite of the previous statement the clear differentiation of
subspecies within M. avium is still controversial. Thus, some authors consider that Mas
is not different enough from Maa to be considered as a separate subspecies (Turenne y
cols. 2006); besides, not fully agreement was made by other authors on Map
nomenclature (Stevenson 1997, Juste 1997a) and M. paratuberculosis designation is
still in use (Bernstein y cols. 2004).
Map is classically identified as an acid-fast rod with dependence on mycobactin and
slow growth on primary isolation. The description of the insertion sequence IS900 in a
Map human isolate (Green y cols. 1989) allowed adding a positive result in IS900 PCR
testing to that definition (Harris y cols. 2001). Given its rapid turn out results and
specificity, IS900 amplification became the main identification criteria of Map, even
before completion of the culture. In spite of the previous statements, reports on positive
IS900 PCR results were published in mycobacteria other than Map (Cousins y cols.
1999, Englund y cols. 2002, Motiwala y cols. 2004). However, because IS900 is rarely
found outside Map in animal samples, its amplification is still applied with good results
in the veterinary microbiology diagnostic field. A more controversial issue is the use of
IS900 molecular detection as a confirmatory diagnostic tool in studies concerning
Crohn’s disease in humans. Roiz et al. reported hybridization of IS900 probe in more
than 19% of M. avium isolates from AIDS patients (Roiz y cols. 1995). Naser et al.
detected up to 57% of M. avium isolates from HIV-infected patients also carrying that
insertion sequence (Naser y cols. 1999). This finding has not been suitably considered

ESTUDIO II
160
in further IS900 based studies on Crohn’s Disease (Bull y cols. 2003a, Naser y cols.
2004, Sechi y cols. 2005, Elguezabal y cols. 2006).
Methods and results
During the characterization of 70 MAC strains from Colombia and Spain, isolated
from HIV positive and negative human patients (Murcia y cols. 2004), a pool of
different genomic targets were tested. All the mycobacteria were characterized using
molecular methods, selected from those frequently applied in members of the MAC.
Methods applied included commercial AccuProbe (Gen-Probe Inc.), PCR restriction
analysis of the hsp65 gene (hsp65-PRA) (Telenti y cols. 1993), and PCR amplification
and sequencing of several selected targets, such as 16S-23S rDNA internal transcribed
spacer (ITS-1) (Frothingham y cols. 1993, Novi y cols. 2000) and macrophage-induced
gene (mig) (Beggs y cols. 2000). RFLP molecular typing was also performed using
IS1245 (van Soolingen y cols. 1998) and IS1311 (Roiz y cols. 1995) as probes (data not
shown). Finally, a panel of insertion sequences previously described within MAC was
also tested by PCR. The panel and procedures were next: IS900 (Bull y cols. 2003a);
IS901 (Ellingson y cols. 2000, Turenne y cols. 2006); IS1245 (Guerrero y cols. 1995);
and IS1311 (Roiz y cols. 1995). Up to five of the Colombian isolates were positive to
IS900 by PCR. All Spanish isolates were negatives for that target (not shown). The
IS900 positive isolates, as well as two other isolates obtained from a same patient were
further studied to confirm those results. The complete set of results in the molecular
characterization of the seven Colombian isolates is shown in Table 1. Results obtained
using the panel of molecular tests confirmed that the isolates were members of the M.
avium most closely related to the subspecies hominisuis (Table 1).
The IS900 PCR products from the human isolates were sequenced in both directions
using primers AV1 and AV2 (Bull y cols. 2003a), and standard dye terminator
chemistry, on a 3700 Genetic Analyzer (Applied Biosystems). These amplified products
showed a nucleotide sequence identical to that found in the genome of Map K10 bovine
strain. That sequence has been recently proposed as the reference sequence of IS900
(Semret y cols. 2006). To rule out the presence of Map DNA, the isolates were tested
for the presence of the Map specific locus 251, using PCR methods described earlier
(Bannantine y cols. 2002a). Besides, the isolates were analysed using IS1311 PCR-REA
(Marsh y cols. 1999), which allowed the differentiation of the Map cattle, sheep and
bison type strains from other members of M. avium (Sevilla y cols. 2005).
All the seven Colombian isolates were negatives for the locus 251 (Table 1).
Moreover, all they showed IS1311 PCR-REA patterns characteristic of M. avium
(Marsh y cols. 1999). These two characteristics allowed their differentiation of the
subspecies Map. The five IS900-positive M. avium isolates, corresponding to 3
Colombian HIV-positive patients, represented a total of 10% of all M. avium isolates

ESTUDIOS
161
included in the previous study (Murcia y cols. 2004), besides they represented the
17.4% of the HIV infected patients.
Table 1. Detailed description of source (patient and specimen) and genetic results of the selected human mycobacterial isolates from Colombian patients under study.
AccuProbe PCR
patient isolate sample MAC Mav Mint PRA
Hsp65 ITS-1
sequevar IS1245 IS900 IS901 Locus 251
IS1311 PCR-REA
3 20 B stools + + – Mav II Mav-A + (0) + + – avium type
4 22 B blood + + – Mav II Mav-B + (12) + – – avium type
4 23 B blood + + – Mav II Mav-B + (12) + + – avium type
4 24 B stools + + – Mav II Mav-B + (12) – – – avium type
4 25 B stools + + – Mav II Mav-B + (12) – + – avium type
11 45 B urine + + – Mav III Mav-A + (1) + – – avium type
11 46 B urine + + – Mav III Mav-A + (1) + + – avium type
All the patients had HIV positive status. Samples from the same patient were obtained separated from minimum one week to maximum six months. Results from Accuprobe, hsp65 PRA, ITS-1 sequevar, and PCR amplification of several targets are indicated. The number of bands detected by RFLP using IS1245 as probe is indicated between brackets in the corresponding column. Isolates from the same patient (patients 4 and 11) showed identical RFLP pattern. None of the RFLP patterns belonged to the typical three-banded bird pattern proposed to be reserved for Maa (Mijs y cols. 2002). All isolates were PCR positive for the gene mig (not shown). MAC, Mycobacterium avium complex; Mav, Mycobacterium avium; Mint, Mycobacterium intracellulare.
Discussion and conclusions
Our results are in agreement with those of Naser et al. (Naser y cols. 1999) although
the frequency of detection of IS900 was much lower in our set of human isolates. The
distinct selection criteria and different geographical origins of both sets could explain
that difference. We have detected the presence of the IS900 element in human M. avium
isolates other than Map. IS900 is present in all the Map bacteria analyzed thus far,
however, its detection in other M. avium subspecies is rather unusual. This difference
indicates the known usefulness of IS900 in the identification of Map in veterinary
medicine (Sevilla y cols. 2005, Semret y cols. 2006). The usually high copy-number of
IS900 per genome in Map improves the diagnostic sensitivity using that IS as a target
on these clinical samples.
The amplification of IS900 in mycobacteria other than Map, and isolated from
human sources, clearly points out a weakness in the usual identification strategy of the
Map that needs to be taken into account. In this sense, the IS1311 PCR-REA can
advantageously complement the standard IS900 PCR in the diagnostic procedures. That
method directly differentiates the Map strains from other M. avium subspecies in a
single analytical procedure (Sevilla y cols. 2005).

ESTUDIO II
162
A rather puzzling observation is that IS900 was not always detected when
sequential isolates from the same patient were studied (Table 1, patient 4). Polyclonal
infection of that patient could be discarded according to the same pattern obtained by
RFLP using IS1245 as probe (data not shown). It can be hypothesized that IS900 could
be an unstable insertion sequence when inserted outside of its usual environmental
genetics, the Map genome; thus the IS could had been lost as sequential isolates from
the same patient proceeds.
Considerations raised by studies on Crohn’s Disease using molecular detection of
IS900 element are still controversial. The high prevalence of IS900 DNA observed in
specimens from patients suffering or not Crohn’s Disease (Bull y cols. 2003a, Naser y
cols. 2004, Sechi y cols. 2005, Elguezabal y cols. 2006) might be due to some degree of
interference with DNA of M. avium, an ubiquitous bacteria. Our data support that
statement when only HIV-positive patients are considered. On the other hand, presence
of M. avium DNA other than Map could also be detected, at some stage, during IS900
PCR-based surveillance studies, in potential vehicles for human infection such as milk,
dairy products, meat, water, etc. Testing on environmental or food products for Map
cannot rely only on the IS900 PCR detection with full confidence.
The results presented here have two implications in human health. Firstly, the
amplification of this insertion element could not fully indicate detection of Map
genome; however, positive IS900 amplification still appears to be associated to clinical
disease. At this respect, and in addition to animal infections, IS900-carrying
mycobacteria could also be considered as etiological agents in infections of susceptible
human population, such as AIDS patients. As a consequence, analysis of additional
specific targets could help to clarify the identity of the IS900-carrying organisms
detected. Secondly, results regarding the involvement of Map in Crohn’s disease, on the
basis of IS900 identification strategy, need to be re-assessed. Our data and those from
previous publications are suggesting that Crohn’s disease could be associated to any
IS900 or IS900-like carrying mycobacteria, which could probably be identified or be
closely related to the Map bacterium in the majority of the cases.
Further research broadening the scope of mycobacterial isolates studied is needed to
clarify the role of IS900 in the human pathology.

ESTUDIO V
Inactivation of Mycobacterium avium subsp.
paratuberculosis in cow’s milk by means of high
hydrostatic pressure at mild temperatures
Tomás López-Pedemonte,1 Iker Sevilla,2 Joseba M. Garrido,2 Gorka Aduriz,2
Buenaventura Guamis,1 Ramón A. Juste2 and Artur X. Roig-Sagués1
1 Centre Especial de Recerca Planta de Tecnologia dels Aliments, CeRTA, XiT, Departament de Ciència Animal i dels Aliments, Facultat de Veterinària, Universitat
Autònoma de Barcelona, 08193 Bellaterra, Spain.
2 Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER), Berreaga, 1, 48160 Derio, Bizkaia, Spain.
Applied and Environmental Microbiology, Vol. 72, No. 6, 2005, pp. 4446–4449


ESTUDIOS
165
Abstract
Two strains of Mycobacterium avium subsp. paratuberculosis (3644/02 and ATCC
19698) were inoculated (approximately 6 log CFU/ml) into sterilized milk to evaluate
inactivation by high hydrostatic pressure. Reductions of M. avium subsp.
paratuberculosis increased with pressure level. Significant differences were also found
between M. avium subsp. paratuberculosis strains and between the media used. Average
reductions of 4 log CFU/ml after treatment with 500 MPa are comparable to those
caused by thermal treatments.
Introduction
Paratuberculosis (Johne’s disease) is a chronic granulomatous enteritis caused by
Mycobacterium avium subspecies paratuberculosis that affects cattle and other
ruminants. Its prevalence in food-producing animals is increasing globally, causing
significant financial losses (Greenstein 2003). Crohn’s disease is a chronic regional
enteritis affecting around 5 per 100,000 humans in developed countries (European
Commission 2000, Chamberlin y cols. 2001, Chacon y cols. 2004). Due to the clinical
and pathological similarities between the diseases, involvement of M. avium subsp.
paratuberculosis in the etiology of Crohn’s disease has been debated for many years.
The introduction of specific PCR together with optimized processing procedures
significantly increased the implication of M. avium subsp. paratuberculosis in disease
causation (Hermon-Taylor y cols. 2000, Greenstein 2003, Bull y cols. 2003a, Chacon y
cols. 2004). Milk could be a potential vehicle of transmission of M. avium subsp.
paratuberculosis from animals with Johne’s disease to humans. The presence of M.
avium subsp. paratuberculosis DNA in human patients without symptoms of
inflammatory bowel disease suggests that this widespread bacterium is present in
contaminated water, milk, and other sources (Argueta y cols. 2000, Naser y cols. 2004).
Recent reports add evidence to this hypothesis and include the isolation of M. avium
subsp. paratuberculosis from breast milk and intestinal and lymph nodes from patients
with Crohn’s disease. Viable M. avium subsp. paratuberculosis has also been detected
in peripheral blood in a substantial proportion of individuals with Crohn’s disease
(Hermon-Taylor y cols. 2000, Schwartz y cols. 2000, Naser y cols. 2000b, Naser y cols.
2004, Selby 2004). The dairy industry is also concerned about M. avium subsp.
paratuberculosis due to reports that suggest it may not be effectively inactivated by
high temperature, short-time pasteurization (72°C, 15 s). There is controversy regarding
this particular issue, although differences in inactivation may be explained by different
pasteurization times and equipment, different detection methods and selective media,
separation of clumps, different initial loads, variation in heat sensitivity between M.
avium subsp. paratuberculosis strains, and the use of freshly isolated or laboratory-

ESTUDIO V
166
adapted strains of M. avium subsp. paratuberculosis (Grant y cols. 2001b, Lund y cols.
2002, Grant y cols. 2002b, Ayele y cols. 2005).
In the last 15 years, treatment of milk and dairy products with high hydrostatic
pressure (HHP) has been the subject of intensive research (O'Reilly y cols. 2001,
Trujillo 2002), mostly involving gram-positive or -negative cells. However, acid-
resistant microorganisms such as Mycobacterium spp. have never been included in these
studies. Our objective was to determine the possibility of reducing counts of M. avium
subsp. paratuberculosis in milk by means of moderate HHP treatments carried out at
mild temperatures.
Materials and methods
Strains and preparation of inocula
The strains used were Mycobacterium avium subsp. paratuberculosis ATCC 19698
and Mycobacterium avium subsp. paratuberculosis 3644/02, a Neiker isolate from a
natural case of clinical paratuberculosis in cattle, determined to be of cattle type (C) by
IS1311 PCR-restriction endonuclease analysis (Marsh y cols. 1999) and of type 1-1
according to pulsed-field gel electrophoresis analysis (Stevenson y cols. 2002). Both
strains were grown in Middlebrook 7H9 broth (Difco, Detroit, MI) supplemented with
Middlebrook oleic acid-albumin-dextrose-catalase enrichment (Becton, Dickinson and
Company, MD) and 0.05% Tween 80 (Panreac Química SA, Barcelona, Spain) (7H9
medium) for 6 and 12 weeks, and then the MacFarland scale (Densimat; Bio-Mérieux,
Marcy l’Etoile, France) was used to estimate cell concentration. The final inoculums
were 8.5 log CFU/ml and 8.6 log CFU/ml for strains 3644/02 and 19698, respectively.
Inoculation
Sixteen 30-ml aliquots of ultra-high-temperature-treated milk were inoculated with
1 ml of each strain suspension. Afterwards, the milk was dispensed in sterile, hermetic,
30-ml low-density polyethylene bottles (Azlon; Bibby Sterilin, Ltd., United Kingdom)
and vacuum packed in a triple layer of plastic bags (bb4.l; Cryovac Packaging, Sant Boi
de Llobregat, Spain). Inoculated samples were stored at 6 ± 2°C.
HHP treatments
Milk samples were pressurized 24 h after inoculation by using discontinuous HHP
equipment (model S-FL-850-9-W; Stansted Fluid Power, Stansted, United Kingdom).
On each run, two milk samples were placed inside the product canister. They were
stabilized at the required temperature (5 or 20°C) and then immediately subjected to 10
min of HHP treatment at 300, 400, or 500 MPa (as measured on the digital and analogue
pressure indicators). Control samples not subjected to HHP (0.1 MPa) were used as the

ESTUDIOS
167
reference for initial M. avium subsp. paratuberculosis counts. The time of pressure
buildup was 24 s per 100 MPa; the decompression time was 30 to 35 s.
Microbiological analysis
At 24 h after pressurization, microbiological analysis was carried out. After
vigorous shaking, a 1-ml aliquot of each treated spiked milk sample was diluted in 9 ml
of sterile distilled water. Further 10-fold dilutions up to 1:100,000 were made for each
aliquot. Then, 0.2 ml of four dilutions (1:1, 1:10, 1:1,000, and 1:100,000) were
inoculated onto two tubes of Herrold’s egg yolk medium (HEY) supplemented with 2
� g/ml of mycobactin J (Allied Monitor, Inc., Fayette, MO) and onto another two tubes
of agar-solidified 7H9 (Bacto Agar; Becton, Dickinson and Company, MD) and
incubated at 37°C. Colonies of M. avium subsp. paratuberculosis were counted after an
incubation period of 16 weeks. Some tubes were discarded due to contamination or
desiccation during the incubation period. An aliquot of similarly treated, uninoculated
ultra-high-temperature-treated milk and pressurization liquid were used as negative
controls.
Statistical analysis
Colony counts per tube were transformed into number of CFU in the original milk
sample. This calculated inoculum value was assigned to all tubes with 100 colonies or
more because confluent growth made individual colony count beyond this number
impossible. Only dilutions with different counts in at least two levels of pressure were
used for statistical analysis after logarithmic transformation. This resulted in using
dilutions of 1/1, 1/10, and 1/1,000 on both media for strain 3644/02 and dilutions of
1/1,000 and 1/100,000 on 7H9 and of 1/1 and 1/10 on HEY for strain 19698. The
general linear models procedure of the SAS System version 8.0 (SAS Institute Inc.,
Cary, NC) was used to test main effects of strain, pressure, temperature, and culture
medium, as well as their interactions, on the final colony count logarithm. A Dunnett
test was used to assess the significance of differences between the control level and each
other level of each main factor, and a Tukey-Kramer multirange test was used for
testing the differences between paired-interaction marginal means.
Results, discussion and conclusions
The overall analysis of variance including all effects and interactions showed that
strain (P < 0.0006), culture medium (P < 0.0001), and pressure (P < 0.0001) had strong
effects on the colony counts. However, the assayed temperature levels had no
significant effects (P = 0.2783), and therefore this effect was eliminated from further
statistical analysis (data not shown). As can be deduced from Table 1 and Figure 1,
observing data for both strains and culture media, 300-, 400-, and 500-MPa treatments
caused significant (P < 0.05) reductions of M. avium subsp. paratuberculosis counts of

ESTUDIO V
168
0.92, 1.59, and 4.58 log CFU/ml, respectively, compared to control samples as
determined by the Dunnett test. Strain 3644/02 had an average recovery of 4.54 log
CFU/ml, which was 1.68 log CFU/ml higher than the recovery of strain 19698 (P <
0.05), independently of other factors. For both strains and all pressures, the tubes of
7H9 medium had a recovery rate 1.72 log CFU/ml higher than those of HEY medium.
However, there was a strong interaction between strain and culture medium (P <
0.0001); nonsignificant differences in recovery occurred for strain 3644/02 in both
media (5.01 log CFU/ml on 7H9 versus 4.24 log CFU/ml on HEY, for all pressures),
while very few colonies of strain 19698 grew on HEY compared to 7H9 medium (4.49
versus 1.27 log CFU/ml, respectively). Therefore, another analysis was carried out,
considering only results on 7H9 for strain and pressure factors. This analysis confirmed
the significant differences between control and 500-MPa-treated samples for both
strains (4.85 log CFU/ml; P < 0.05), but this was not the case for the other two pressure
levels. The analysis of interactions further supported this result, since the differences
between control and 500-MPa-treated samples were 4.07 log CFU/ml (P = 0.01) and
5.20 log CFU/ml (P < 0.001) for strains 3644/02 and 19698, respectively (Table 1).
Table 1. HHP treatment effects on counts of Mycobacterium avium subsp. paratuberculosis strains 3644/02 and 19698 in different culture media a.
Strain 3644/02 Strain 19698 Both strains
Counts (Log CFU/mL)
Counts (Log CFU/mL)
Counts (Log CFU/mL) Medium
Pressure (MPa)
No. of replicates
Mean S.D.
No. of replicates
Mean S.D.
No. of replicates
Mean S.D.
7H9 0.1 30 5.90 1.66 20 6.10 2.63 50 5.98 2.08
300 7 5.22 2.74 14 5.49 3.02 21 5.40 2.86
400 19 4.37 2.53 13 5.37 3.07 32 4.78 2.76 500 5 1.83 1.03 16 0.90 1.98 21 1.13 1.82
All 61 5.01 2.33 63 4.49 3.37 124 4.75 2.91
HEY 0.1 30 6.28 1.24 20 1.98 1.43 50 4.56 2.50
300 19 5.66 2.00 15 1.23 1.06 34 3.70 2.76 400 24 3.94 2.67 16 1.11 0.93 40 2.81 2.55
500 21 0.39 0.65 14 0.50 0.83 35 0.44 0.72
All 94 4.24 2.87 65 1.27 1.22 159 3.03 2.75
Both 0.1 60 6.09 1.46 40 4.04 2.95 100 5.27 2.39 300 26 5.54 2.17 29 3.29 3.08 55 4.35 2.90
400 43 4.13 2.59 29 3.02 3.02 72 3.68 2.80
500 26 0.67 0.92 30 0.72 1.54 56 0.69 1.28
All 155 4.54 2.69 128 2.86 2.98 283 3.78 2.94 a There were no significant differences between treatments at 5 and 20°C.
The counts of M. avium subsp. paratuberculosis obtained at 5°C were similar to
those obtained at 20°C. This fact turns out to be very useful, since it means that mild
HHP treatments could be applied to milk at refrigeration temperatures, thus minimizing
organoleptic and nutritional alteration. As can be seen in Table 1, HHP inactivation of
both strains of M. avium subsp. paratuberculosis was proportional to the increase in the

ESTUDIOS
169
pressure applied, an effect which was
also observed in both culture media.
This is generally the tendency
observed among vegetative bacteria
(Smelt 1998, Gervilla y cols. 1999).
However, there were strong
interactions between strain and
medium, which indicate that assay
conditions are very important for the
assessment of inactivation of M. avium
subsp. paratuberculosis. This was
quite evident for strain 19698 in HEY,
where even the counts of control
samples were reduced by 4.12 log
CFU/ml relative to their growth on 7H9 (Table 1). In contrast, strain 3644/02 grew
similarly in both media. Preference of different strains of M. avium subsp.
paratuberculosis for primary isolation media has been observed before (Aduriz y cols.
1995a).
Different piezotolerances between strains of the same species have been observed
by many researchers (Alpas y cols. 1999, Ritz y cols. 2000, Lamo-Castellvi y cols.
2005). From the way that both M. avium subsp. paratuberculosis strains responded to
HHP treatments, it seems that strain 3644/02 was slightly more pressure resistant than
strain 19698.
The mean reduction obtained for strain 3644/02 at 500 MPa was 4.07 log CFU/ml.
This value suggests the possibility of inactivating this microorganism by means of HHP
when its load in raw milk is less than 4 log CFU/ml. Many authors estimate that raw
bulk tank milk could reach levels of up to 4 and 5 log CFU/ml of M. avium subsp.
paratuberculosis cells (Meylan y cols. 1996, Grant y cols. 1996, Sung y cols. 1998,
Grant y cols. 1998a, Grant y cols. 2001b, Gao y cols. 2002, O'Reilly y cols. 2004). This
is mostly due to fecal contamination and also, to a lesser extent, to cells secreted within
the udders (Sweeney y cols. 1992a, Streeter y cols. 1995, Ayele y cols. 2005); Sweeney
et al. (Sweeney y cols. 1992a) found 2 to 8 CFU/ml (0.3 to 0.9 log CFU/ml) of M.
avium subsp. paratuberculosis cells in milk samples aseptically obtained from the
udders of nine subclinical animals.
Nevertheless, HHP inactivation values are comparable to those obtained in
pasteurization studies (Gao y cols. 2002, Greenstein 2003) and to those obtained with
pulsed electric fields (Rowan y cols. 2001), but they have the advantage of using
relatively lower temperatures. It is still possible to apply pressures higher than 500 MPa
to raw milk. HHP technology could also be applied to soft-curd cheeses traditionally
made from raw milk in order to improve its safety. The work of Donaghy et al.
Figure 1. Graphic representation of reductions obtained according to the pressure level (MPa), the culture medium and the strain used.

ESTUDIO V
170
(Donaghy y cols. 2004) states that M. avium subsp. paratuberculosis cells survive short-
term curing when present in high (up to 5 log CFU/ml) or low (2 log CFU/ml) numbers
as well.
Due to the isostatic condition of HHP treatments (which means that pressure is
homogeneously applied to every point of the sample), large differences are not expected
when larger samples and HHP machines are used. However, aggregated M. avium
subsp. paratuberculosis cells could still interfere and modify the reductions obtained.
Moreover, temperature gradients due to the adiabatic heating of the pressure chamber
may vary in laboratory and industrial equipment. This is why pilot-scale experiments
such as those made with pasteurization treatments (Pearce y cols. 2001, Grant y cols.
2002b) may be useful. Further research should be done in order to determine the nature
of HHP injuries in this sort of vegetative cell and to evaluate the behavior of pressurized
M. avium subsp. paratuberculosis cells during storage life at refrigeration temperatures.
Acknowledgements
Iker Sevilla held a fellowship from the Department of Industry and the Department
of Education, Universities and Research of the Basque Government.

DISCUSIÓN GENERAL


DISCUSIÓN GENERAL
173
Actualmente los métodos rutinarios de diagnóstico in vivo de la paratuberculosis son
el ELISA y el cultivo fecal, técnicas consideradas mucho más específicas que sensibles
(Meylan y cols. 1994, Whitlock y cols. 2000, Collins y cols. 2005). A pesar de que el
ELISA puede resultar muy rentable, el cultivo fecal es económicamente poco atractivo
y lento, con lo que el diagnóstico completo se ve retrasado y resulta caro, situación
problemática que se ve incrementada cuando se trata de un número importante de
animales en el rebaño. La PCR es una técnica cuyo precio y ejecución es cada día más
asequible, y la que se basa en la detección de un fragmento de la secuencia IS900 a
partir de heces, se utiliza para sustituir o complementar al cultivo fecal (Collins y cols.
1993b, Garrido y cols. 1999b, Chevallier y cols. 2003). A pesar de que se ha informado
de algunos resultados inespecíficos (Cousins y cols. 1999, Englund y cols. 2002), esta
técnica se considera muy específica. Además, se puede adaptar con facilidad a otro tipo
de muestras como la sangre, que puede utilizarse en otros ensayos como es el ELISA, y
en la que una interferencia con DNA de otras bacterias es menos probable. En nuestro
trabajo se ha detectado una falta de concordancia entre los resultados de ELISA y PCR
de sangre, siendo muy evidente en animales jóvenes. Dado el carácter espectral de la
enfermedad, esto puede interpretarse como indicio de que cada técnica detecta
diferentes estadios de la infección. Por eso y por la especificidad que se les atribuye, se
puede hablar de que estas técnicas complementan su sensibilidad mutuamente. En
cualquier caso, nuestro trabajo demuestra la frecuencia de la bacteriemia celular en las
poblaciones susceptibles a la paratuberculosis y el potencial de la PCR específica en
esta matriz como herramienta de diagnóstico etiológico.
La paratuberculosis está considerada como una enfermedad progresiva y la
producción de anticuerpos se asocia a grandes cantidades de la bacteria infectando al
animal, sin embargo, en este estudio hemos observado individuos con anticuerpos
detectados por ELISA con resultado negativo de PCR. Probablemente la patogénesis de
esta micobacteriosis sea más compleja, y los niveles de anticuerpos y bacteriemia se
vean alterados por reinfecciones, recaídas o reactivaciones, que regulen la respuesta
humoral o la circulación por el organismo de células fagocíticas cargadas de
micobacterias. En este trabajo también hemos observado infección mixta por distintas
cepas en distintos momentos de la vida de un animal, que se han aislado de distintas
muestras, lo que da cierto apoyo a la idea anterior. Además el tipo de cepa también
puede ser un factor importante en el desarrollo de la enfermedad, de hecho ciertas cepas
de tipo bovino parecen ser muy patógenas para todas las especies, mientras que otras no
lo son tanto (Taylor 1951, Taylor 1953, Stamp y cols. 1954, Stevenson y cols. 2002).
Esto también puede explicar la alta prevalencia de ovejas positivas en PCR de sangre,
especie que según hemos podido observar se encuentra parasitada mayoritariamente por
cepas de tipo ovino, que quizá sean mejor contenidas por el sistema inmunitario de parte
de los animales, y cuya infección se vea reducida a formas focales sin avanzar a
clínicas. Estos portadores pueden resultar determinantes en los planes de control que

DISCUSIÓN GENERAL
174
buscan la erradicación de la enfermedad, y la PCR de sangre puede mejorar
sensiblemente su detección, pudiendo convertirse así en una buena herramienta de
diagnóstico complementaria. La variabilidad en la detección de Map en las células
fagocíticas de la sangre, deja abiertas otras cuestiones sobre la patogénesis de la
paratuberculosis, como la posible existencia de factores genéticos de resistencia o
susceptibilidad del hospedador, punto ya señalado en la enfermedad de Crohn (materia
ya revisada). En la línea de esta discusión, parece que en el caso de la paratuberculosis
la infección no conduce necesariamente a la enfermedad, hipótesis bajo la que no se
cumplirían los clásicos postulados de Koch sobre la etiología de las enfermedades
infecciosas. Dicho de otra manera, Map sería un factor necesario pero no único o
suficiente en la etiología de la paratuberculosis. Esta hipótesis tiene importantes
implicaciones en lo que se refiere al control de la paratuberculosis, que al menos por el
momento, debería centrarse más en la prevención de la enfermedad que de la infección.
Del mismo modo, la estrategia para demostrar el posible papel de Map en la enfermedad
de Crohn debería estar orientada a la determinación de los mecanismos inmunes que en
unos casos contienen la infección eficazmente y en otros conducen al desarrollo de
lesiones tisulares graves, en lugar de limitarse a comparar la presencia (infección) de
Map entre enfermos clínicos y población clínicamente sana.
Sin embargo, en aislamientos humanos de M. avium parece bastante frecuente la
presencia de secuencias similares a la IS900 (IS900-like) (Roiz y cols. 1995, Naser y
cols. 1999), que pueden interferir en las técnicas de detección de Map basadas en ese
elemento. La gran ventaja que proporciona la IS900 es principalmente su alto número
de copias por bacteria, aumentando de manera importante la sensibilidad de estas
pruebas. Dado que los procedimientos actuales pueden producir amplificaciones
indistinguibles de ciertas regiones de la IS900 en aislamientos de otras micobacterias
procedentes de pacientes de sida (como se ha demostrado en nuestro trabajo), parece
necesario tomar con cautela los resultados de los estudios que las utilizan para detectar
Map en humanos, y complementarlos si es posible incluyendo otra diana específica de
esta micobacteria. Asimismo, los trabajos de vigilancia sobre la presencia de Map en
distintos medios o productos, pueden verse afectados por las mismas conclusiones. Con
toda la cautela necesaria debido al escaso interés que dicha secuencia ha recibido en
estudios microbiológicos del género Mycobacterium desde la perspectiva humana,
puede decirse que no se ha descrito la presencia de este elemento en micobacterias
ambientales demostradamente apatógenas. Este estudio precisamente muestra el interés
de los estudios microbiológicos comparados y confirma hallazgos previos en el sentido
de que IS900 es un elemento presente en una proporción moderada de aislados distintos
de Map. No obstante, todos los aislamientos de Map descritos en éste y en todos los
demás trabajos poseen esta controvertida secuencia de inserción, y su detección es
indicativa de la presencia de Map, o alternativamente, de micobacterias potencialmente
patógenas.

DISCUSIÓN GENERAL
175
Continuando con el trabajo que hemos llevado a cabo en torno a la diferenciación de
cepas de Map, hay que señalar que se ha informado repetidamente de dificultades en el
aislamiento a partir de muestras de ovino principalmente, y en zonas geográficas y
sistemas productivos muy diversos. Esta observación, sin embargo, no es universal y
algunos autores no han encontrado diferencias entre la especie hospedadora, los medios
de cultivo, la velocidad de crecimiento y el tipo de cepa (Pavlik y cols. 1999a). En los
trabajos anteriores de Neiker ya se apuntaban ciertas diferencias en el éxito del cultivo
de muestras procedentes de vacas, cabras y ovejas en diferentes medios de cultivo (Juste
y cols. 1991b, Aduriz y cols. 1995a), sin embargo no se había comprobado la posible
relación entre los fenotipos y los genotipos de tales aislados. Tras comprobar la utilidad
de la IS1311 PCR-REA, como resultado de los trabajos realizados con esta técnica,
hemos podido verificar la asociación entre las características de crecimiento y el
genotipo de estos aislados, viniendo a confirmar las conclusiones de trabajos
precedentes (Bauerfeind y cols. 1996). Así, según nuestros hallazgos las cepas de tipo
IS1311–C parecen tener una preferencia muy fuerte por el medio HEY con piruvato y
micobactina añadidos, medio en el que nunca obtuvimos colonias de tipo IS1311–S.
Estas últimas crecen mejor en LJ con micobactina o en medios Middlebrook con OADC
con o sin micobactina. Las cepas IS1311–B, crecen mejor en HEY con micobactina
pero sin piruvato añadido. Tal y como venimos observando en los últimos tiempos, los
medios de cultivo líquido basados en 7H9 parecen cubrir el amplio abanico de
requerimientos de cultivo para todas las cepas, aunque los problemas de
contaminaciones siguen siendo importantes.
Como ya se ha comentado, existe cierta discordancia en la bibliografía sobre la
distribución de las diferentes cepas en cuanto a hospedadores y regiones. A excepción
de algunos casos en los que se ha confirmado la presencia de cepas ovinas en vacuno
(uno de ellos muy recientemente) (Whittington y cols. 2001d, de Juan y cols. 2006a),
parece que esta especie se ve afectada principalmente por el tipo bovino de Map. En las
especies ovina y caprina de numerosas regiones también se consideran más extendidas
las cepas de tipo bovino que las de tipo ovino (Thoresen y cols. 1994b, Pavlik y cols.
1999b, Pavlik y cols. 2000a, Stevenson y cols. 2002, Djonne y cols. 2005, de Juan y
cols. 2005), sin embargo, a la vista de los resultados de nuestro trabajo, esa no sería la
distribución en la Península Ibérica. Estas diferencias, quizá puedan ser explicadas por
un sesgo causado por la utilización de medios y procedimientos inadecuados para aislar
cepas ovinas, aunque no se pueden descartar cuestiones epidemiológicas. En nuestro
caso, hemos contado con una representación de aislados presumiblemente menos
sesgada, y la polarización observada en la distribución ha sido muy clara (ver Figura 3
en estudio III). El éxito del cultivo también puede verse influenciado por otros factores
(método de descontaminación, presencia de otros microorganismos que interaccionan
con Map, estados metabólicos o de estrés de Map,...), algunos como los señalados para

DISCUSIÓN GENERAL
176
explicar las diferencias en el aislamiento a partir de leche pasterizada (Grant y cols.
2004), o como los que hemos podido observar en el estudio de HHP (ver más adelante).
Este es el trabajo con el mayor número de aislados de Map tipados por medio de
SnaBI-SpeI-PFGE, aislados para los que se han podido describir numerosos patrones de
restricción nuevos en varias especies hospedadoras. Entre los aislados bovinos la
diversidad de cepas es muy baja (en nuestro estudio casi el 85% pertenecen a dos únicos
tipos), al contrario que lo que hemos podido observar entre ovejas y cabras. Sin
embargo en otros lugares los aislados son muy homogéneos en todas las especies
hospedadoras (Pavlik y cols. 1999a, Cousins y cols. 2000, Whittington y cols. 2000b,
Djonne y cols. 2005). La baja diversidad parece ser el reflejo de la homogeneidad en el
manejo y posiblemente en la genética de los animales, provocada por el mercado
internacional y por los programas de mejora genética, que quizá hayan diseminado
genotipos de animales más susceptibles a ciertos genotipos de Map. Esta idea se ve
reforzada por las interesantes diferencias observadas entre razas, que en el caso de la
raza de lidia señala la existencia de un contexto epidemiológico distinto, a pesar de la
pequeña representación de estas cepas. Del mismo modo, la alta diversidad de cepas que
hemos observado en pequeños rumiantes parece reflejar la heterogeneidad genética y de
manejo de los rebaños ovinos y caprinos en nuestra área de estudio. Según nuestro
dendrograma de agrupamiento de genotipos de Map, la pigmentación no sería una
característica fenotípica que produzca el agrupamiento genético de cepas pigmentadas
por un lado y no pigmentadas por otro como se había propuesto anteriormente
(Stevenson y cols. 2002), puesto que algunas pigmentadas tienen patrones más similares
a otras no pigmentadas que al resto de pigmentadas. Nuestro estudio viene a confirmar
el agrupamiento tradicional en dos grupos con subgrupos propuesto inicialmente
(Collins y cols. 1990a). El poder de discriminación general de la PFGE empleada ha
sido bajo, pero debido a la homogeneidad de las cepas bovinas. De todos modos, se trata
de una técnica cuyo uso sistemático en paratuberculosis no es recomendable, ya que en
ocasiones como la descrita en bovino lechero, no proporciona suficiente información
adicional que justifique la dificultad y los elevados costos que origina. Esta observación
apunta una línea de investigación en el desarrollo de otros sistemas de tipado más
simples e informativos para los que la colección de cepas caracterizadas en este estudio
puede ser una importante base de partida.
La observación de diferentes patrones PFGE en un mismo animal plantea una
cuestión inesperada que resulta difícil de explicar a partir de las bases teóricas de la
técnica de genotipado utilizada. Es decir, no parece probable que exista una variabilidad
tan alta en el genoma de Map que justifique la aparición de tantos patrones en un tiempo
relativamente corto de tiempo. Por lo tanto, la explicación más simple sería asumir que
pueden existir infecciones policlonales en un mismo individuo que podrían ser
seleccionadas al hacer el subcultivo a partir de colonias aisladas en medio sólido. Este
hallazgo sería consistente con los resultados obtenidos en colonias de M. avium aisladas

DISCUSIÓN GENERAL
177
de pacientes de SIDA que se discuten más arriba y que en unas ocasiones muestran la
presencia de IS900 y en otras no.
La posible implicación de Map en la enfermedad de Crohn, ha favorecido el
desarrollo de estudios sobre la presencia y resistencia de la bacteria en nuestra cadena
alimentaria. Los niveles de la bacteria en leche de tanque de explotaciones bovinas
afectadas pueden ser muy elevados (Meylan y cols. 1996, Grant y cols. 1998a, Gao y
cols. 2002) debido a la contaminación fecal principalmente, y en menor medida, a la
excreción en leche de vacas infectadas (Sweeney y cols. 1992a, Streeter y cols. 1995,
Sung y cols. 1998). Los tratamientos con altas presiones hidrostáticas (HHP) de leche y
otros productos lácteos han sido objeto de investigación intensiva durante los últimos 15
años con resultados satisfactorios (Trujillo 2002), sin embargo ninguno incluía bacterias
ácido-alcohol resistentes como Map. En nuestro estudio hemos realizado una prueba
preliminar a pequeña escala sobre el uso de tratamientos HHP moderados a
temperaturas de 5 y 20º C, como procedimiento para eliminar de la leche las células
viables de esta bacteria. El hecho de que no se hayan observado diferencias
significativas entre los tratamientos a estas temperaturas es muy interesante, pues indica
que se pueden obtener reducciones del mismo nivel que las de la pasterización a
temperaturas de refrigeración, a las que la leche y otros productos pueden mantener sus
propiedades y características organolépticas intactas. De este modo se pueden evitar
tratamientos térmicos más severos y con mayores efectos no deseables sobre las
cualidades de este importante producto. Como es de esperar, la inactivación es
proporcional al incremento en la presión ejercida, y teniendo en cuenta que todavía
pueden aplicarse presiones superiores a 500 MPa a la leche cruda, probablemente se
pueda llegar a producir leche totalmente libre de células viables de Map. Asimismo, el
tratamiento HHP podría aplicarse al calostro y leche con que alimentar a los animales de
reposición, a la leche cruda que se utiliza para elaborar quesos (con la ventaja de que
esto acelera el proceso de curado y mejora la producción (Trujillo 2002)), a los quesos
directamente, y a otros muchos productos.
El efecto e interacción entre cepas y medios de cultivos también ha quedado patente
en el ensayo de inactivación. A pesar de que las dos cepas empleadas poseen el mismo
genotipo (IS1311–C y SnaBI-SpeI PFGE–1-1), la cepa de referencia (ATCC 19698)
está muy adaptada al laboratorio como consecuencia de sucesivos pases en 7H9, medio
en el que ha crecido notablemente mejor, mientras que el crecimiento de la cepa de
campo (recién aislada) ha sido similar en ambos medios. No obstante, el 7H9 parece
haber sido más eficaz a la hora de recuperar células dañadas (por el tratamiento) de las
dos cepas, tal y como se desprende de los recuentos de la leche tratada a 400 o 500
MPa. También cabe destacar la aparente mayor resistencia de la cepa de campo en
comparación con la de referencia. Todos estos hallazgos demuestran la importancia de
las condiciones y procedimientos que se utilizan, pues pueden influir claramente en los
resultados de este tipo de estudios, cuyas conclusiones pueden tener gran alcance. Los

DISCUSIÓN GENERAL
178
resultados de este ensayo son prometedores, y garantizan la ejecución de más estudios
sobre la utilización de tratamientos HHP para inactivar este patógeno en leche y otros
productos.
Como resultado general de todo el trabajo realizado en la presente Tesis Doctoral, se
han podido esclarecer varios aspectos sobre el diagnóstico, la patogénesis, la
epidemiología y el control de la paratuberculosis, y lo que es más importante, se han
generado nuevos conocimientos, cuestiones e hipótesis que abren nuevos caminos para
la investigación de esta enfermedad.

CONCLUSIONES GENERALES


CONCLUSIONES GENERALES
181
1. La PCR específica de IS900 en muestras de sangre permite detectar animales
con infección paratuberculosa que no detectan otras técnicas como el ELISA, lo
que demuestra la existencia de una bacteriemia (“DNA-hemia”) en distintas
fases de la infección paratuberculosa y sugiere una estrategia de combinación de
técnicas para aumentar la sensibilidad del diagnóstico epidemiológico.
2. La técnica denominada IS1311 PCR-REA junto con la IS900 PCR, forman una
herramienta muy práctica para realizar la identificación y el genotipado general
de los aislamientos de Mycobacterium avium subsp. paratuberculosis de manera
sistemática por su fácil y económica ejecución. La IS1311 PCR-REA es capaz
de diferenciar entre cepas de tipo bovino, ovino y de bisonte de Mycobacterium
avium subsp. paratuberculosis, además de distinguir éstas del resto de
subespecies del complejo Mycobacterium avium.
3. La técnica denominada SnaBI-SpeI PFGE es un método de genotipado de cepas
de Mycobacterium avium subsp. paratuberculosis con bajo poder de
discriminación cuando se trata de aislamientos procedentes de bovino lechero,
principalmente de vacas de raza frisona/holstein. Sin embargo, en el caso de
aislados procedentes de especies y/o razas con carácter más regional, su poder de
discriminación se ve sensiblemente incrementado. Debido a su complejidad, no
resulta recomendable como primer método de genotipado de aislamientos de
esta micobacteria.
4. El análisis de similitudes entre los perfiles SnaBI-SpeI PFGE descritos agrupa
las cepas de Mycobacterium avium subsp. paratuberculosis en dos grandes
grupos generales. Uno de ellos comprende tipos de cepas aisladas únicamente de
pequeños rumiantes (tipos ovinos), mientras que en el otro se incluyen tipos de
cepas aisladas de todas las especies estudiadas (tipos bovinos).
5. La colección de aislamientos de Mycobacterium avium subsp. paratuberculosis
estudiada muestra que la especie bovina se infecta exclusivamente con cepas de
tipo bovino, mientras que la especie ovina se ve principalmente infectada por
cepas de tipo ovino. La especie caprina, está parasitada por ambos tipos de
cepas, pero la incidencia de cepas ovinas es mucho mayor de lo que la anterior
literatura reflejaba. Las cepas de tipo IS1311–B se encuentran infectando ovejas
y cabras procedentes de India y bisontes de EEUU, pero según la PFGE, no son
el mismo tipo de cepa.

CONCLUSIONES GENERALES
182
6. Existe un alto grado de homogeneidad genotípica entre los aislados bovinos de
Mycobacterium avium subsp. paratuberculosis en general. Los aislados de
origen caprino son más heterogéneos, pero menos que los de origen ovino, entre
los que se ha detectado una diversidad muy superior a la de los bovinos.
7. Se ha confirmado la infección policlonal con tres cepas diferentes de
Mycobacterium avium subsp. paratuberculosis en un toro semental, dos de las
cuales también se hallaban infectando otros animales de la misma explotación.
8. El genotipo (IS1311 PCR-REA, SnaBI-SpeI PFGE) de las cepas de
Mycobacterium avium subsp. paratuberculosis se correlaciona bien con las
características fenotípicas de crecimiento. Así, las cepas de tipo bovino crecen
mejor en HEY con micobactina y piruvato y las de tipo ovino en LJ sin piruvato
con micobactina, o en 7H11 con OADC con o sin micobactina. El medio 7H9
con OADC es idóneo para la propagación de todos los tipos de cepas analizados.
La utilización de varios medios para el aislamiento primario es necesaria para
evitar posibles sesgos en el aislamiento de cepas diferentes a las esperadas en
cada especie hospedadora.
9. Se ha detectado la presencia de IS900 en cinco aislamientos de Mycobacterium
avium obtenidos de tres pacientes de SIDA procedentes de Colombia. Estos
resultados obligan a interpretar con cautela los resultados de estudios previos
sobre la relación entre Mycobacterium avium subsp. paratuberculosis y la
enfermedad de Crohn basados en la detección de IS900. Esto implica que es
necesario el uso de una técnica alternativa en la identificación de los aislados de
Mycobacterium avium subsp. paratuberculosis que no procedan de las especies
de rumiantes sensibles a la paratuberculosis.
10. El tratamiento experimental de la leche con altas presiones hidrostáticas (HHP)
de 500 MPa a 5 o 20ºC de temperatura, produce reducciones medias de 4,07 y
5,20 log CFU/ml en el recuento de células viables de la cepa de campo 3644/02
y de la cepa de referencia ATCC 19698 de Mycobacterium avium subsp.
paratuberculosis, respectivamente. Estas reducciones son similares a las
producidas por pasterización, con la ventaja de que el tratamiento de HHP puede
aplicarse en temperaturas de refrigeración que no alteran las propiedades
organolépticas de la leche.

RESUMEN; LABURPENA; SUMMARY


RESUMEN; LABURPENA; SUMMARY
185
RESUMEN
La paratuberculosis es una afección crónica del intestino causada por
Mycobacterium avium subsp. paratuberculosis (Map), que afecta principalmente a
rumiantes. Es de difusión mundial y provoca importantes pérdidas económicas en el
sector ganadero. A pesar de no haber sido demostrado de manera concluyente, Map
también parece estar involucrado en la enfermedad de Crohn, y la detección de la
bacteria en posibles vehículos de transmisión para el ser humano ha aumentado la
preocupación en salud pública.
Este trabajo consta de cinco estudios cuyos objetivos están relacionados con
aspectos bacteriológicos de Map: confirmar la circulación de Map (o de su DNA) en la
sangre y estudiar su potencial diagnóstico, adaptar a Neiker métodos de genotipado
molecular y utilizarlos para caracterizar una colección importante de aislamientos de
Map, y evaluar tratamientos alternativos a los térmicos para inactivar esta bacteria en
leche destinada al consumo humano.
En el primer estudio se comparó el ELISA y la PCR de sangre en muestras de
bovino y ovino. El conjunto de muestras bovinas (94 de animales menores de dos años
y 184 de animales de entre 2 y 5 años) procedía de una explotación afectada, y se
componía de 80 positivas y 198 negativas a ELISA. La PCR detectó DNA de Map
(IS900) en 66 de estas muestras. El análisis general de los resultados mostró una clara
asociación pero una baja correlación negativa entre las dos técnicas (Fisher exact test
[FET] p = 0,0049; 45,3%, kappa = -0,1665 ± 0.0994). Se observó una ligera
concordancia entre las novillas (87,7%, kappa = 0,4471 ± 0,2435), que resultó negativa
entre vacas adultas (62,7%, kappa = -0,3670 ± 0,1057). En el caso de la especie ovina,
se utilizaron 496 muestras de sangre procedentes de 53 explotaciones, muestras entre las
que había 180 de parejas madre/cría. Se confirmó la presencia de DNA de Map en 61
muestras ovinas. La asociación entre los resultados de ELISA y PCR en estas muestras
era fuerte, pero con una baja concordancia (FET, p < 0,0005; kappa = 0,1622 ± 0,1188)
ligeramente mejor entre ovejas adultas (kappa = 0,2135 ± 0,1992) que entre corderos
(kappa = 0,1193 ± 0,1301). También se detectó una alta proporción de parejas
madre/cría positivas (FET p < 0,0001, kappa = 0.6269 ± 0.1854). Los resultados de la
PCR IS900, además de demostrar la circulación del DNA de Map en la sangre, indican
una potencial utilidad diagnóstica de esta técnica, especialmente en animales jóvenes.
Estas apreciaciones deberán ser confirmadas en estudios más extensos con animales
cuyo estatus de paratuberculosis sea determinado con métodos de referencia.
El segundo, tercero y cuarto estudios se centran en la verificación de métodos de
identificación y caracterización molecular de Map. En el segundo, se estudió la validez
de la PCR IS900 y del análisis de restricción con endonucleasas de amplificados de la
secuencia IS1311 (IS1311 PCR-REA) como estrategia de identificación y genotipado de

RESUMEN; LABURPENA; SUMMARY
186
cepas de Map, aplicándola a 38 aislamientos procedentes de vacas, ovejas, cabras y
bisontes de distintas regiones de España, India y EEUU, y a una colección de otras
micobacterias distintas de Map. Todos los aislados de Map eran positivos a IS900 e
IS1311, a diferencia de las otras micobacterias que eran negativas, excepto un aislado
de M. intracellulare con el que se obtuvo un amplificado de IS1311 perfectamente
distinguible del específico. De los aislamientos obtenidos en España, todos los que
procedían de bovino, tres de seis de los caprinos y uno de los diez ovinos, pertenecían al
tipo C (cattle type). Mientras, los nueve ovinos y tres caprinos restantes pertenecían al
tipo S (sheep type). Todos los aislados de India eran del tipo B (bison type), tipo que
sólo se había detectado en bisontes de EEUU, al que igualmente pertenecían los tres
analizados en este estudio. Los resultados de este estudio preliminar muestran
diferencias genotípicas entre aislamientos de Map relacionados con factores geográficos
y de hospedador que pueden ser explotados para el trazado epidemiológico de
aislamientos de Map con el método IS1311 PCR-REA.
En el tercer estudio se empleó la IS1311 PCR-REA para analizar 520 aislamientos
de Map cultivados de varias especies hospedadoras (bovino, ovino, caprino, bisonte,
ciervo y jabalí) procedentes de diversas localizaciones geográficas, con el fin de
conocer mejor la epidemiología de Map. Cuatro de los cultivos ovinos eran
pigmentados. Todos los aislamientos bovinos, un 4% de los ovinos y un 26% de los
caprinos eran del tipo C, así como los de ciervo y jabalí. En cambio, el 96% de las
ovejas y el 74% de las cabras procedentes de España se hallaban infectadas con cepas
de tipo S, igual que las tres ovejas de Portugal. De esta colección de aislamientos, 269
fueron además tipados con la técnica de electroforesis en gel de campo pulsado (PFGE)
utilizando las endonucleasas SnaBI y SpeI. El empleo de este método reveló 37 perfiles
SnaBI-SpeI diferentes, de los cuales 32 no habían sido descritos antes. El 85% de los
cultivos bovinos eran de los tipos llamados 2-1 y 1-1, previamente descritos en la
bibliografía. Sin contar con los aislamientos de pequeños rumiantes de India (todos ellos
con perfil 29-1), entre los cultivos ovinos se detectaron 10 genotipos distintos, y su
prevalencia no superó en ningún caso el 25%. El 43% de los aislamientos caprinos eran
del tipo 16-11 previamente descrito, aislamientos entre los que se detectaron otros tres
tipos más. Gracias a esta técnica de tipado, se pusieron de manifiesto algunas
diferencias interesantes entre las distintas razas estudiadas. El dendrograma que se
construyó a partir de las similitudes entre los patrones de PFGE de todas las cepas,
dividió los aislamientos en dos grandes grupos compatibles con los descritos en gran
parte de la bibliografía. En uno (cepas de tipo bovino o C) se incluían todas las cepas
procedentes de bovino, de ciervo, de jabalí, y algunas de pequeños rumiantes, mientras
que el otro (cepas de tipo ovino o S) lo componían exclusivamente cepas ovinas y
caprinas, que representaban 11 perfiles SnaBI-SpeI diferentes. Entre las cepas
pigmentadas ovinas se detectaron 2 tipos diferentes, y aunque ambos fueron incluidos
en el grupo de cepas ovinas, no formaban un subgrupo común. En cultivos sucesivos de

RESUMEN; LABURPENA; SUMMARY
187
varias muestras de un toro semental se descubrió la presencia de tres cepas diferentes de
Map, sugiriendo una infección mixta. Los tipos de las cepas de bisonte de EEUU y los
de pequeños rumiantes de India eran diferentes, lo que indica que no se trata de la
misma cepa. Se observaron diferencias en los requerimientos de cultivo asociadas con el
genotipo de los aislamientos. A pesar de que las distintas modalidades del medio de
cultivo 7H9 parecen cumplir con los requerimientos de todos los tipos de cepa, resulta
interesante utilizar otros medios para minimizar cualquier tipo de sesgo en el cultivo de
Map, así como subcultivar en medio sólido los cultivos líquidos para obtener colonias
puras. En España, la especie bovina se encuentra parasitada por cepas de tipo bovino,
mientras que las ovejas y cabras se ven principalmente afectadas por cepas de tipo
ovino. Se ha detectado una gran homogeneidad genotípica entre los aislados bovinos, a
diferencia de la heterogeneidad detectada entre los de origen ovino y caprino.
En el cuarto estudio se contó con varios aislamientos de M. avium procedentes de
pacientes de SIDA colombianos que habían sido exhaustivamente caracterizados por
métodos moleculares (Accuprobe, hsp65-PRA, ITS-1, mig, IS1245 PCR y RFLP,
IS1311 PCR y RFLP, e PCR IS900). Según la PCR, cinco de ellos procedentes de tres
pacientes poseían la secuencia específica de Map IS900 (17,4% de los pacientes). Como
resultado de la secuenciación de los amplificados se comprobó que ésta era idéntica a la
de la cepa K10 recientemente propuesta como de referencia. Esos cinco aislados y otros
dos IS900 negativos de uno de los pacientes fueron analizados con la IS1311 PCR-
REA, y el patrón obtenido correspondía en todos los casos al de M. avium. Para
descartar la presencia de DNA de Map, también se les aplicó la PCR para detectar la
secuencia exclusiva de Map Locus 251, siempre con resultado negativo. La presencia de
IS900 en aislados de M. avium de pacientes infectados con el HIV ya se había
observado, pero no se le ha atribuido la suficiente importancia. Los resultados de
estudios de detección de Map en pacientes de Crohn basados en la IS900, pueden verse
afectados, pues parece que su detección no implica necesariamente la presencia de Map
al menos en estos grupos de personas, aunque sigue estando asociada a enfermedad
clínica. El empleo de otras dianas genéticas puede confirmar la identidad de estas
bacterias portadoras de IS900.
En el quinto y último trabajo, en vista de la posibilidad de que células viables de
Map se encuentren en leche destinada al consumo humano, se planteó la posibilidad de
usar un método alternativo a los tratamientos térmicos para conseguir la inactivación de
Map en este potencial vehículo de transmisión para el ser humano, las altas presiones
hidrostáticas (HHP). Para ello, se inocularon de manera artificial varias muestras de
leche con distintas concentraciones de dos cepas diferentes de Map, que fueron
sometidas a tratamientos de 300, 400 y 500 MPa en temperaturas de 5 y 20ºC. Las
reducciones en las células viables de ambas cepas se incrementaron con el aumento de
la presión. Se pudieron observar diferencias significativas entre las cepas, y entre los
medios de cultivo. En cambio, el resultado de los tratamientos fue muy similar a ambas

RESUMEN; LABURPENA; SUMMARY
188
temperaturas. Con el tratamiento de 500 MPa se consiguieron reducciones medias de 4
log CFU/ml que son similares a las obtenidas con la pasterización, con la ventaja de que
el HHP se puede aplicar en condiciones de refrigeración que mantienen las cualidades y
características organolépticas de la leche.
LABURPENA
Paratuberkulosia Mycobacterium avium subsp. paratuberculosis (Map) bakterioak
heste mehean eraginiko gaixotasun kronikoa da, eta batez ere animalia hausnarkarien
artean ematen da. Mundu osoan zehar hedatuta dago eta galera ekonomiko
garrantzitsuak eragiten ditu abere-industrian. Nahiz eta oraindik inork ez duen froga
definitiborik aurkitu, badirudi Mapek Crohn gaixotasunaren eragilea dela. Bakterioa
gizakiaren elika katean topatu denez, osasun publikoaren inguruko ardura nabarmenki
areagotu egin da.
Lan hau bost estudio ezberdinez osatuta dago, eta bere helburuak Mapen alderdi
bakteriologikoekin erlazionatuta daude: Map (edo bere DNA) odolean zehar ibiltzen
dela baieztatzea eta horrek diagnostikoan erabiltzeko daukan ahalmena ikertzea,
karakterizazio molekularreko teknikak Neikerren erabiltzeko egokitzea eta hoiekin
Mapen isolamendu bilduma bat karakterizatzea, eta azkenik, gizakiaren kontsumorako
zuzendutako esnean tratamendu termikoen metodo alternatiboak saiatzea Map
inaktibatzeko.
Lehenengo estudioan ELISA eta odol PCRa konparatu ziren behi eta ardi laginekin.
Behien odol laginak esplotazio erasatu batetik hartu ziren (94 behi bi urte baino
gazteagoak eta 184 bi eta bost urte bitartekoak), hauetatik 80 ELISA positibo eta 198
ELISA negatibo izanik. PCRak Mapen DNAa (IS900) lagin hauetako 66tan detektatu
zuen. Emaitzen analisi orokorraren arabera teknika bien arteko asoziazioa argia zen
baina korrelazio negatiboa zen (Fisher exact test [FET] p = 0,0049; 45,3%, kappa = -
0,1665 ± 0.0994). Nahiz eta ahula izan, gazteen artean tekniken konkordantzia ikusi zen
(87,7%, kappa = 0,4471 ± 0,2435), behi helduen artekoa negatiboa izanik (62,7%,
kappa = -0,3670 ± 0,1057). Ardien kasuan, 53 esplotazio ezberdinetan hartutako 496
odol lagin bildu ziren, hauen artean 180 ama/kume bikoteak izanik. Mapen DNAa ardi
laginen 61 laginetan aurkitu zen. Konkordantzia baxua izan arren, ELISA eta PCRaren
emaitzen arteko asoziazioa handia izan zen (FET, p < 0,0005; kappa = 0,1622 ±
0,1188). Ardi helduen artean lortutako konkordantzia altuagoa zen (kappa = 0,2135 ±
0,1992) gazteetan baino (kappa = 0,1193 ± 0,1301). Ama/kume bikote positiboen
proportzio altua detektatu zen (FET p < 0,0001, kappa = 0.6269 ± 0.1854). IS900
PCRaren emaitzek erakusten duten bezala, Mapen DNAa odolean zehar ibiltzen da, eta
teknika honek diagnostikorako erabilera potentziala duela, batez ere animalia gazteen
artean. Etorkizunean, balorazio hauek paratuberkulosi estatus ezaguna (erreferentziazko

RESUMEN; LABURPENA; SUMMARY
189
teknikek jarritakoa) duten animaliekin egindako estudio handiagoetan egiaztatu behar
izango dira.
Hurrengo hiru estudioak (bigarrena, hirugarrena eta laugarrena) Mapen identifikazio
eta karakterizazio molekularrerako metodoetan oinarrituta daude. Bigarrenean, Mapen
identifikazio eta anduien genotipo azterketa orokorrerako, IS900 PCRaren eta
IS1311aren amplifikatuen endonukleasen bidezko errestrikzioaren (IS1311 PCR-REA)
baliotasuna aztertu zen. Horretarako, Espainia, India eta EEBBetako eskualde
ezberdinetako behi, ardi, ahuntz eta bisontetatik eratorritako 38 isolamendu erabili
ziren. Kontrol modura Mapen ezberdinak ziren beste mikobakterio bilduma bat erabili
zen. Mapen isolamendu guztiek IS900 eta IS1311 PCReetan positiboak ziren, eta
kontrol guztiak negatiboak, M. intracellulareren kontrolan emandako banda
inespezifikoa izan ezik (dena den tamaina ezberdina zuelaz aparte, REAren bidez
IS1311ren ezberdina zela froga zitekeen). Espainiar estatuko behi isolamendu guztiak,
ahuntzen 6 isolamenduetatik hiru, eta ardien 10etatik beste bat C (cattle type) motakoak
ziren. Aldiz, ardien beste 10ak eta ahuntzen beste hirurak S (sheep type) motakoak
ziren. Lehenago EEBBetako bisonte batzuetan besterik aurkitu ez zen B (bison type)
motakoak ziren Indiako isolamendu guztiak, gure estudioko bisonte amerikarrak bezala.
Atariko estudio honen emaitzek Mapen isolamenduen artean ostalari eta geografia
faktorekin erlazionatutako ezberdintasunak daudela eta IS1311 PCR-REAa Mapen
epidemiologia molekularra aztertzeko erabilgarria dela erakusten dute.
Hirugarren estudioan, Mapen epidemiologia hobeto ulertzeko asmoz puntu
geografiko ezberdinetako ostalari ezberdinetatik (behia, ardia, ahuntza, bisontea, oreina
eta basurdea) hazitako 520 isolamendu IS1311 PCR-REArekin aztertu ziren. Ardien
isolamenduetatik lau pigmentatuak ziren. Behien isolamendu guztiak, ardien
isolamenduen %4a, eta ahuntzen isolamenduen %26a, C motakoak ziren, orein eta
basurde isolamenduak bezala. Aldiz, Espainiako ardien %96a eta ahuntzen %74a S
motako anduiekin zeuden infektatuta, Portugaleko hiru ardiak bezala. Isolamendu
bilduma honetatik, 269 hartu ziren eremu pultsatupeko gel elektroforesiaren (PFGE)
bidez genotipatzeko, SnaBI eta SpeI entzimak erabiliz. Teknika honen bidez 37 SnaBI-
SpeI profil ezberdin aurkitu ziren, hauetatik 32 berriak izanik. Behien isolamenduen
%85a bibliografian lehenagotik deskribatuta zeuden 2-1 edo 1-1 genotipoa erakutsi
zuten. Indiako hausnarkari txikien isolamenduak kontutan hartu gabe (guztiak 29-1
izanik), ardien kultiboen artean 10 genotipo ezberdin aurkitu ziren, nahiz eta horietako
batek ere ez zeukan %25eko prebalentzia baino handiagoa. Ahuntzen isolamenduen
%43a bibliografian deskribatuta zegoen 16-11 genotipoa erakutsi zuten, eta espezie
honen gainerako isolamenduen artean beste hiru genotipo ezberdin aurkitu ziren espezie
ostalari hau infektatzen. Teknika honi ezker aztertutako arraza ezberdinen arteko
diferentzia interesgarriak somatu ziren. PFGE patroien antzekotasunetan oinarritutako
dendrogramak isolamenduak bi talde ezberdinetan banandu zituen, bibliografiaren parte
batean bezala. Talde batean (behi motako anduiak edo C anduiak) behi, orein eta

RESUMEN; LABURPENA; SUMMARY
190
basurdetik isolatutako kultibo guztiak, eta hausnarkari txikietatik isolatutako gutxi
batzuk biltzen ziren bitartean, bestean (ardi motako anduiak edo S anduiak), ardi edo
ahuntzen isolamenduen artean bakarrik aurkitu ziren 11 SnaBI-SpeI profil biltzen ziren.
Andui pigmentatuen artean bi genotipo aurkitu ziren, eta biak S motakoen artean
sailkatu ziren arren, hauek ez zuten azpitalde bakarra eratu elkarrekin. Hazitarako zezen
baten lagin ezberdinetako kultibo ezberdinetan (ezberdinak denboran zehar ere bai)
Mapen andui ezberdinak azaldu ziren, infekzio mixtoaren indikatzaile izanik. Indiako
hausnarkari txikien anduien eta EEBBetako bisonteen anduien genotipoak ezberdinak
ziren, beraz ez ziren andui berbera. Isolamenduaren genotipoarekin erlazionatuta
agertzen ziren hazkuntza ezberdintasunak somatu ziren. Nahiz eta 7H9 medioaren
modalitate ezberdinek Mapen andui guztien hazkuntzarako errekerimendu guztiak
barnean hartzen dituztela dirudien, zehartasun posibleak murrizteko, beste medio solido
batzuk erabiltzea interesgarria da, baita kultibo likidoak solido batean azpikultibatzea
ere, horrela kolonia puruak hazteko. Espainian behiak C motako anduiek infektazen
dituzte, ardiak eta ahuntzak batez ere S motako anduiek infektatzen dituzten bitartean.
Behi isolamenduen artean homogeneotasun genetiko oso handia ikusi izan da, ardi eta
ahuntz isolamenduen artean aldiz, heterogeneotasun handia da ikusi izan dena.
Laugarren estudioan, sakonki karakterizatuta (Accuprobe, hsp65-PRA, ITS-1, mig,
IS1245 PCR y RFLP, IS1311 PCR y RFLP, e PCR IS900) zeuden Kolonbiako HIES
gaixo batzuetatik isolatutako kultibo batzuk erabili dira. PCRaren arabera, hiru gaixo
ezberdinetatik (aztertutako gaixoen %17,4) hazitako bost isolamenduk Mapen IS900
sekuentzia espezifikoa zeukaten. Sekuentziazioaren arabera, anplifikatu hoien
sekuentzia erreferentzia gisa proposatu berri duten K10 Mapen anduiaren berdinak
ziren. Bost isolamendu horiek eta gaixo baten beste bi (isolamendu IS900 positibo bi
zituen eta beste bi negatibo) IS1311 PCR-REAren bidez aztertu ziren, eta emaitza M.
aviumen patroi berdina izan zen kasu guztietan. Mapen DNArik ez zegoela baieztatzeko
Locus 251 Mapen sekuentzia espezifikoa bilatu zen PCRaren bidez, eta isolamendu
guztietan negatiboa izan zen. Aurretiko beste lan batzuk IS900a HIESdun gaixotatik
hazitako M. aviumen isolamendu batzutan topatu zutela diote, baina aurkikuntza hauei
orain arte ez zaie merezi duten garrantzia eman. Gure emaitzak eta aurrekoak
(bibliografia), IS900an oinarritutako Mapen detekzioa erabiltzen duten Crohni buruzko
estudioen emaitzetan eragina eduki dezakete. Hau da, IS900a aurkitzeak ez du Mapen
beharrezko presentzia inplikatzen, pertsona talde mota honetan behintzat. Hala ere,
IS900a gaixotasun klinikoarekin erlazionatuta jarraitzen du. Beste itu genetikoen
erabilerari esker bakterio IS900-eramaileen identitatea konfirmatu daiteke.
Mapen zelula bideragarriak giza-kontsumorako zuzendutako esnean topa daitezkela
kontutan hartuz, azkeneko estudioan (bostgarrena) Map inaktibatzeko tratamendu
termikoen metodo alternatiboak aztertzea zen helburua, presio hidrostatiko altuak
(HHP) erabiltzea alegia. Horretarako esne lagin ezberdinak Mapen andui biren
kontzentrazio ezberdinez inokulatu ondoren, 300, 400 eta 500 MPaeko presioz tratatu

RESUMEN; LABURPENA; SUMMARY
191
ziren 5 edota 20ºCtan. Zelula bideragarrien kontaketetan lortutako murrizketak presioa
igotzearekin batera handitzen ziren. Erabilitako kultibo medioen artean eta anduien
artean ezberdintasun signifikatiboak aurkitu ziren. Ez zen temperaturaren (5 edo 20ºC)
eraginik nabaritu. 500 MPaeko tratamenduari esker zelula bideragarrien 4 log
CFU/mlko batazbesteko murrizketak lortu ziren, pasterizazioaren bidez lortzen direnen
antzekoak izanik. Hala ere, HHP tratamenduak duen abantaila hozte-temperaturatan
aplika daitezkela da, esnearen kualitate eta ezaugarri organoleptikoak mantenduz.
SUMMARY
Mycobacterium avium subsp. paratuberculosis (Map) causes a chronic
inflammation (paratuberculosis) of the intestines mainly in ruminant species. Its
prevalence in food-producing animals is increasing globally, causing significant
financial losses. Although there are no conclusive studies, it is suspected to be
responsible for Crohn’s disease. The detection of Map in potential transmission vehicles
for humans has increased public health concerns on Map.
The present work consists of five studies focused on some bacteriological aspects of
Map: to confirm the presence of Map (or its DNA) on cattle and sheep blood and to
study its potential use in diagnosis, to adjust molecular typing methods of Map in
Neiker and to use them for a big Map collection characterization, and finally, to
evaluate alternatives for the use of thermal inactivation treatments of micro-organisms
on milk destined for human consumption.
The first study was carried out to compare the performance of enzyme-linked
immunosorbent assay (ELISA) and blood polymerase chain reaction (PCR) for
diagnosis of paratuberculosis in cattle and sheep. For cattle, a set of 278 samples from
one paratuberculosis affected Friesian farm was used; it included 80 ELISA-positive
samples and 198 ELISA-negative samples from an age-matched group. Ninety-four
samples were from heifers and 184 were from 2-5-year-old cows. The IS900 PCR
detected 66 positive samples. The overall analysis showed a clear association (Fisher
exact test [FET] p = 0.0049) but a weak negative agreement (45.3%, kappa = -0.1665 ±
0.0994) between the two tests. It reflected a moderate agreement among heifers (87.7%,
kappa = 0.4471 ± 0.2435) and a moderate disagreement among cows (62.7%, kappa = -
0.3670 ± 0.1057). For sheep, 496 single blood samples from 53 Latxa dairy flocks; 180
of the blood samples were from dam/offspring pairs. In 61 ovine blood samples the
PCR detected the presence of Map’s DNA. The overall association between the 2 tests
on ovine samples was strong (FET, p < 0.0005), whereas the agreement was low (kappa
= 0.1622 ± 0.1188). There was slightly better agreement for ewes (kappa = 0.2135 ±
0.1992) than for lambs (kappa = 0.1193 ± 0.1301). There was also a highly unlikely
proportion of dam/offspring positive results (FET p < 0.0001, kappa = 0.6269 ±
0.1854). These results confirm the presence of Map’s DNA in cattle and sheep blood,

RESUMEN; LABURPENA; SUMMARY
192
and suggest that blood PCR testing might be a potentially useful new approach in
paratuberculosis diagnosis, especially in young animals. Further intensive studies on
animals with known paratuberculosis status (with reference techniques) are needed for
confirmation of these considerations.
The second, third and fourth studies are focused on identification and molecular
characterization methods of Map isolates. In the second one, the validity of IS900 PCR
and IS1311 restriction-endonuclease-analysis (IS1311 PCR-REA) as identification and
preliminary characterization methods are studied. These were applied to 38
Mycobacterium avium subsp. paratuberculosis (Map) isolates from cattle, sheep, goats
and bison from distinct regions of Spain, India and the United States of America (USA).
A control non-Map mycobacterial collection was used. All Map isolates were positive
in both IS900 and IS1311 PCR methods. None of the other isolates was positive in any
of the PCR assays performed, except M. intracellulare, which yielded a band easily
distinguishable from the specific one in the IS1311 PCR. In Spain, all eight bovine
isolates, three out of six caprine isolates and one of ten ovine isolates were of the C type
(cattle type), while the other nine ovine isolates and three caprine isolates were of the S
type (sheep type). In India, all five ovine isolates and six caprine isolates were of the B
type, and so were all three isolates from bison (Bison bison) from the USA. These
results show that there are genetic differences between Map isolates related to
geographic and host factors that have a potential use in the epidemiological tracing of
new paratuberculosis isolates by using the IS1311 PCR-REA.
In order to better understand the epidemiology of paratuberculosis, 520 isolates from
different hosts (cattle, sheep, goats, bison, deer and wild boar) and origins were cultured
and typed by IS1311 PCR-REA, in the third study. Four of the ovine isolates were
pigmented. All bovine isolates, 4 and 26% of Spanish ovine and caprine cultures,
respectively, and the deer and wild boar isolates, were IS1311-cattle type strains.
IS1311-sheep type encompassed 96% and 74% of isolates from Spanish sheep and
goats, and all three Portuguese sheep cultures. Two-hundred and sixty-nine of these
isolates were further characterized by pulsed-field gel electrophoresis (PFGE) using
SnaBI and SpeI endonucleases. Thirty-seven distinct multiplex PFGE profiles were
found, giving 32 novel profiles. Previously defined profiles 2-1 and 1-1 accounted for
the 85% of cattle isolates. All Indian isolates belonged to the novel 29-1 profile. Ten
distinct profiles were detected in Spanish sheep, none of them with an incidence higher
than 25%. 16-11 (43%) and another three profiles were identified in Spanish caprine
cultures. The use of PFGE showed some interesting differences between different
breeds. The hierarchical analysis, clustered all profiles found in cattle, “wild” hosts and
some small ruminants within the same group (C type strains). The other group included
11 profiles only found in Spanish sheep and goats, including Spanish pigmented profiles
(S type strains). Although the two profiles found in these pigmented isolates belonged to
the same S type group, they were not included in a separate subgroup. Consecutive

RESUMEN; LABURPENA; SUMMARY
193
cultures from different samples of one bull yielded three isolates with a different PFGE
genotype, suggesting mixed infection of the animal. The differences found between
profiles of Indian small ruminant isolates and those from American bisons indicate they
are not the same strain. Differences in growth requirements associated with isolate
genotype were observed. Although 7H9 broth based culture media seem to broadly
cover the growth requirements of most Map strains, the use of various solid media is
recommended to reduce any recovery biases, and subculture on solid media to obtain
pure colonies. Cattle in Spain are infected with C type strains, while small ruminants are
mainly infected with S type strains. High genetic homogeneity of isolates from cattle,
and heterogeneity of those from sheep and goats have been detected.
In the fourth study, various thoroughly characterized (Accuprobe, hsp65-PRA, ITS-
1, mig, IS1245 PCR y RFLP, IS1311 PCR y RFLP, e PCR IS900) M. avium isolates
from Colombian HIV positive human patients were further characterized. Up to five of
the isolates cultured from three patients were positive to IS900 by PCR (17.4% of
patients studied). These amplified products showed a nucleotide sequence identical to
that found in the genome of Map K10 bovine strain. That sequence has been recently
proposed as the reference sequence of IS900. The IS900 positive isolates, as well as two
other isolates obtained from a same patient were further studied with IS1311 PCR-REA
to confirm those results. All isolates gave the same M. avium pattern. To rule out the
presence of Map DNA, the isolates were tested for the presence of the Map specific
locus 251, and all isolates were negative. This is not the first report on the presence of
the IS900 element in M. avium isolates from HIV positive patients. This finding has not
been suitably considered in further IS900 based studies on Crohn’s Disease, since
detection of IS900 does not necessarily implicate presence of Map, in this group of
patients at least. However, positive IS900 amplification still appears to be associated to
clinical disease. Analysis of additional specific targets could help to clarify the identity
of the IS900-carrying micro-organisms detected.
Finally, in the fifth study, the objective was to evaluate the ability of high
hydrostatic pressures (HHP) to be used as an alternative to thermal treatments for
inactivation of Map in milk destined for human consumption. Several milk samples
were artificially inoculated with different dilutions of two Map strains for this purpose.
Samples were then treated with 300, 400 y 500 MPa pressures at 5 and 20ºC.
Reductions of M. avium subsp. paratuberculosis increased with pressure level.
Significant differences were also found between M. avium subsp. paratuberculosis
strains and between the media used, but the assayed temperature levels had no
significant effects on the counts. Average reductions of 4 log CFU/ml after treatment
with 500 MPa are comparable to those caused by thermal treatments. Since mild HHP
treatments could be applied to milk at refrigeration temperatures, minimizing
organoleptic and nutritional alteration, HHP treatment seem to be advantageous.


BIBLIOGRAFÍA CITADA


BIBLIOGRAFÍA CITADA
197
Ablordey A, Swings J, Hubans C, Chemlal K, Locht C, Portaels F y Supply P. 2005. Multilocus variable-number tandem repeat typing of Mycobacterium ulcerans. J.Clin.Microbiol., 43(4):1546-51.
Adaska JM y Anderson RJ. 2003. Seroprevalence of Johne's-disease infection in dairy cattle in California, USA. Prev.Vet.Med., 60(3):255-61.
Aduriz G, Juste RA, Garrido JM y Geijo MV. 2000. Epidemiología y control de la paratuberculosis bovina. En: Juste editor. Paratuberculosis. Bovis, Aula Veterinaria, vol.nº93, pp. 63-93. Luzan 5, Madrid, España.
Aduriz JJ. 1993. Epidemiología, diagnóstico y control de la paratuberculosis ovina en la Comunidad Autónoma del País Vasco. Tesis Doctoral. Universidad de Zaragoza, Aragón, España.
Aduriz JJ, Juste RA y Cortabarría N. 1995a. Lack of mycobactin dependence of mycobacteria isolated on Middlebrook 7H11 from clinical cases of ovine paratuberculosis. Vet.Microbiol., 45:211-7.
Aduriz JJ, Juste RA, Marco JC, Sáez de Ocáriz C, García AL y Cuervo LA. 1991. Diagnósticos de paratuberculosis en el SIMA: Análisis epidemiológico. ITEA, 11:653-5.
Aduriz JJ, Juste RA y Sáez de Ocáriz C. 1995b. An epidemiologic study of sheep paratuberculosis in the Basque Country of Spain: serology and productive data. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 19-26. IAP, Rehoboth, MA, USA.
Agasino CB, Ponce dL, Jasmer RM y Small PM. 1998. Epidemiology of Mycobacterium tuberculosis strains in San Francisco that do not contain IS6110. Int.J.Tuberc.Lung Dis., 2(6):518-20.
Aho AD, McNulty AM y Coussens PM. 2003. Enhanced expression of interleukin-1alpha and tumor necrosis factor receptor-associated protein 1 in ileal tissues of cattle infected with Mycobacterium avium subsp. paratuberculosis. Infect.Immun., 71(11):6479-86.
Al Shamali M, Khan I, Al Nakib B, Al Hassan F y Mustafa AS. 1997. A multiplex polymerase chain reaction assay for the detection of Mycobacterium paratuberculosis DNA in Crohn's disease tissue. Scand.J.Gastroenterol., 32(8):819-23.
Alexejeff-Goloff NA. 1929. Zur frage der pathogenese und bazillenausscheidung bei Rinderparatuberkulose. Zeitschr.Infekt.Haus., 36(4):313-7.
Allen WM, Berret S y Patterson DSP. 1974a. A biochemical study of experimental Johne's disease. I. Plasma leakage into the intestine of sheep. J.Comp.Pathol., 84:381-4.
Allen WM, Patterson DS y Slater TF. 1974b. A biochemical study of experimental Johne's disease. I. Protein metabolism in sheep and mice. J.Comp.Pathol., 84:391-8.
Aller B, Fernández-Díez M y Escudero-Díez A. 1973. Paratuberculosis ovina. Sup.Cient.Bol.Inf.Cons.Gen.Col.Vet.Esp., 196:11-8. Allix C, Walravens K, Saegerman C, Godfroid J, Supply P y Fauville-Dufaux M. 2006. Evaluation of the epidemiological relevance
of variable-number tandem-repeat genotyping of Mycobacterium bovis and comparison of the method with IS6110 restriction fragment length polymorphism analysis and spoligotyping. J.Clin.Microbiol., 44(6):1951-62.
Alpas H, Kalchayanand N, Bozoglu F, Sikes A, Dunne CP y Ray B. 1999. Variation in resistance to hydrostatic pressure among strains of food-borne pathogens. Appl.Environ.Microbiol., 65(9):4248-51.
Alvarez J, de Juan L, Briones V, Romero B, Aranaz A, Fernandez-Garayzabal JF y Mateos A. 2005. Mycobacterium avium subspecies paratuberculosis in fallow deer and wild boar in Spain. Vet.Rec., 156(7):212-3.
Aly SS y Thurmond MC. 2005. Evaluation of Mycobacterium avium subsp. paratuberculosis infection of dairy cows attributable to infection status of the dam. J.Am.Vet.Med.Assoc., 227(3):450-4.
Alzuherri HM, Little D y Clarke CJ. 1997. Altered intestinal macrophage phenotype in ovine paratuberculosis. Res.Vet.Sci., 63:139-43.
Amonsin A, Li LL, Zhang Q, Bannantine JP, Motiwala AS, Sreevatsan S y Kapur V. 2004. Multilocus short sequence repeat sequencing approach for differentiating among Mycobacterium avium subsp. paratuberculosis strains. J.Clin.Microbiol., 42(4):1694-702.
Anderson JR. 1980. Alimentary tract. Muir's textbook of pathology. ed. 11. Edward Arnold. London, UK. Angus KW. 1990. Intestinal lesions resembling paratuberculosis in a wild rabbit (Oryctolagus cuniculus). J.Comp.Pathol., 103:101-
5. Angus KW y Gilmour NJL. 1971. Effect of the rimino phenazine B663 (G30320) on Mycobacterium johnei infection and
reinfection in sheep. II Pathology. J.Comp.Pathol., 81:227-36. Arbeit RD, Slutsky A, Barber TW, Maslow JN, Niemczyk S, Falkinham III JO, O'Connor GT y Von Reyn CF. 1993. Genetic
diversity among strains of Mycobacterium avium causing monoclonal and polyclonal bacteremia in patients with AIDS. J.Infect.Dis., 167(6):1384-90.
Argueta C, Yoder S, Holtzman AE, Aronson TW, Glover N, Berlin OG, Stelma GN, Jr., Froman S y Tomasek P. 2000. Isolation and identification of nontuberculous mycobacteria from foods as possible exposure sources. J.Food Prot., 63(7):930-3.
Arques JL, Rodriguez E, Gaya P, Medina M, Guamis B y Nunez M. 2005. Inactivation of Staphylococcus aureus in raw milk cheese by combinations of high-pressure treatments and bacteriocin-producing lactic acid bacteria. J.Appl.Microbiol., 98(2):254-60.
Arsenault J, Girard C, Dubreuil P, Daignault D, Galarneau JR, Boisclair J, Simard C y Belanger D. 2003. Prevalence of and carcass condemnation from maedi-visna, paratuberculosis and caseous lymphadenitis in culled sheep from Quebec, Canada. Prev.Vet.Med., 59(1-2):67-81.
Askling J, Grahnquist L, Ekbom A y Finkel Y. 1999. Incidence of paediatric Crohn's disease in Stockholm, Sweden. Lancet, 354(9185):1179.
Aurtenetxe O, Sevilla I, Fernández de Mera I, Vicente J, Garrido JM, Gortazar C, Juste RA y Barral M. 2005. Micobacteriosis en mamiferos silvestres. VII Jorn.SECEM, 3-6 dic. 2005. Valencia, España.
Autschbach F, Eisold S, Hinz U, Zinser S, Linnebacher M, Giese T, Loffler T, Buchler MW y Schmidt J. 2005. High prevalence of Mycobacterium avium subspecies paratuberculosis IS900 DNA in gut tissues from individuals with Crohn's disease. Gut, 54(7):944-9.
Ayele WY, Bartos M, Svastova P y Pavlik I. 2004. Distribution of Mycobacterium avium subsp. paratuberculosis in organs of naturally infected bull-calves and breeding bulls. Vet.Microbiol., 103(3-4):209-17.
Ayele WY, Svastova P, Roubal P, Bartos M y Pavlik I. 2005. Mycobacterium avium subspecies paratuberculosis cultured from locally and commercially pasteurized cow's milk in the Czech Republic. Appl.Environ.Microbiol., 71(3):1210-4.
Badiola JJ, Balaguer L, Arnal C y García Marín JF. 1986. La paratuberculosis ovina en Aragón: trabajos realizados por el Departamento de Histología y Anatomía Patológica de la Facultad de Veterinaria de Zaragoza. Reu PTBC España, (ed. I), pp. 9-10.
Badiola JJ, García de Jalón JA y Cuervo LA. 1980. Paratuberculosis ovina. An.Fac.Vet.Zaragoza, XIV-XV:14-5. Bakker D y Willemsen PTJ. 2006. Evaluation of paratuberculosis vaccination studies in the Netherlands. VENoMYC, Use of
vaccines in the control of tuberculosis and paratuberculosis, 10-11 de Noviembre. Laguardia, Euskadi, Spain.

BIBLIOGRAFÍA CITADA
198
Baksh FK, Finkelstein SD, Ariyanayagam-Baksh SM, Swalsky PA, Klein EC y Dunn JC. 2004. Absence of Mycobacterium avium subsp. paratuberculosis in the microdissected granulomas of Crohn's disease. Mod.Pathol., 17(10):1289-94.
Balseiro A, Prieto JM, Espi A, Perez V y Garcia Marin JF. 2003. Presence of focal and multifocal paratuberculosis lesions in mesenteric lymph nodes and the ileocaecal valve of cattle positive to the tuberculin skin test. Vet.J., 166(2):210-2.
Balseiro AM. 2004. Paratuberculosis Bovina: valoración de las reacciones cruzadas con la prueba de la tuberculina, evaluación de técnicas diagnósticas y prevalencia en Asturias. Tesis Doctoral. Universidad de León, Castilla y León, España.
Bannantine JP, Baechler E, Zhang Q, Li L y Kapur V. 2002a. Genome scale comparison of Mycobacterium avium subsp. paratuberculosis with Mycobacterium avium subsp. avium reveals potential diagnostic sequences. J.Clin.Microbiol., 40(4):1303-10.
Bannantine JP, Barletta RG, Stabel JR, Paustian ML y Kapur V. 2004a. Application of the genome sequence to address concerns that Mycobacterium avium subspecies paratuberculosis might be a foodborne pathogen. Foodborne Pathog.Dis., 1(1):3-15.
Bannantine JP, Hansen JK, Paustian ML, Amonsin A, Li LL, Stabel JR y Kapur V. 2004b. Expression and immunogenicity of proteins encoded by sequences specific to Mycobacterium avium subsp. paratuberculosis. J.Clin.Microbiol., 42(1):106-14.
Bannantine JP, Huntley JF, Miltner E, Stabel JR y Bermudez LE. 2003a. The Mycobacterium avium subsp. paratuberculosis 35 kDa protein plays a role in invasion of bovine epithelial cells. Microbiology, 149(Pt 8):2061-9.
Bannantine JP, Palmer MV, Kapur V, Stabel JR y Paustian ML. 2006a. Development and use of a M. avium subsp. paratuberculosis partial protein array for discovery of novel antigens. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 146. IAP, Madison, WI, USA.
Bannantine JP y Paustian ML. 2006b. Identification of diagnostic proteins in Mycobacterium avium subspecies paratuberculosis by a whole genome analysis approach. Methods Mol.Biol., 345:185-96.
Bannantine JP y Stabel JR. 2001. Identification of two Mycobacterium avium subspecies paratuberculosis gene products differentially recognised by sera from rabbits immunised with live mycobacteria but not heat-killed mycobacteria. J.Med.Microbiol., 50(9):795-804.
Bannantine JP y Stabel JR. 2002b. Killing of Mycobacterium avium subspecies paratuberculosis within macrophages. BMC Microbiol., 2:2.
Bannantine JP, Zhang Q, Li LL y Kapur V. 2003b. Genomic homogeneity between Mycobacterium avium subsp. avium and Mycobacterium avium subsp. paratuberculosis belies their divergent growth rates. BMC Microbiol., 3(1):10.
Barclay R, Ewing DF y Ratledge C. 1985. Isolation, identification, and structural analysis of the mycobactins of Mycobacterium avium, Mycobacterium intracellulare, Mycobacterium scrofulaceum, and Mycobacterium paratuberculosis. J.Bacteriol., 164(2):896-903.
Barclay R y Ratledge C. 1988. Mycobactins and exochelins of Mycobacterium tuberculosis, M. bovis, M. africanum and other related species. J.Gen.Microbiol., 134(3):771-6.
Barrington GM, Gay JM, Eriks IS, Davis WC, Evermann JF, Emerson C, O'Rourke JL, Hamilton MJ y Bradway DS. 2003. Temporal patterns of diagnostic results in serial samples from cattle with advanced paratuberculosis infections. J.Vet.Diagn.Invest., 15(2):195-200.
Barta Z, Csipo I, Mekkel G, Zeher M y Majoros L. 2004. Seroprevalence of Mycobacterium paratuberculosis in patients with Crohn's Disease. J.Clin.Microbiol., 42(11):5432-3.
Bartos M, Hlozek P, Svastova P, Dvorska L, Bull T, Matlova L, Parmova I, Kuhn I, Stubbs J, Moravkova M, Kintr J, Beran V, Melicharek I, Ocepek M y Pavlik I. 2006. Identification of members of Mycobacterium avium species by Accu-Probes, serotyping, and single IS900, IS901, IS1245 and IS901-flanking region PCR with internal standards. J.Microbiol.Methods, 64(3):333-45.
Bascuñana CR y Belak K. 1996. Detection and identification of mycobacteria in formalin-fixed, paraffin-embedded tissues by nested PCR and restriction enzyme analisis. J.Clin.Microbiol., 34(10):2351-5.
Bauerfeind R, Benazzi S, Weiss R, Schliesser T, Willems H y Baljer G. 1996. Molecular characterization of Mycobacterium paratuberculosis isolates from sheep, goats and cattle by hybridization with a DNA probe to insertion element IS900. J.Clin.Microbiol., 34(7):1617-21.
Beard PM, Daniels MJ, Henderson D, Pirie A, Rudge K, Buxton D, Rhind S, Greig A, Hutchings MR, McKendrick I, Stevenson K y Sharp JM. 2001a. Paratuberculosis infection of nonruminant wildlife in Scotland. J.Clin.Microbiol., 39(4):1517-21.
Beard PM, Henderson D, Daniels MJ, Pirie A, Buxton D, Greig A, Hutchings MR, McKendrick I, Rhind S, Stevenson K y Sharp JM. 1999. Evidence of paratuberculosis in fox (Vulpes vulpes) and stoat (Mustela erminea). Vet.Rec., 145:612-3.
Beard PM, Rhind SM, Buxton D, Daniels MJ, Henderson D, Pirie A, Rudge K, Greig A, Hutchings MR, Stevenson K y Sharp JM. 2001b. Natural paratuberculosis infection in rabbits in Scotland. J.Comp.Pathol., 124(4):290-9.
Begara-McGorum I, Wildblood LA, Clarke CJ, Connor KM, Stevenson K, McInnes CJ, Sharp JM y Jones DG. 1998. Early immunopathological events in experimental ovine paratuberculosis. Vet.Immunol.Immunopathol., 63:265-87.
Beggs ML, Stevanova R y Eisenach KD. 2000. Species identification of Mycobacterium avium complex isolates by a variety of molecular techniques. J.Clin.Microbiol., 38(2):508-12.
Behrens H. 1955. Die Paratuberkulöse des Schafes. Dtsch.Tierarztl.Wochenschr., 13-14:133-4. Behymer DE, Riemann HP, Utterback W, Elmi C y Franti CE. 1991. Mass screening of cattle sera against 14 infectious disease
agents, using an ELISA system for monitoring health in livestock. Am.J.Vet.Res., 52(10):1699-705. Belknap EB, Getzy DM, Johnson LW y Ellis RP. 1994. Mycobacterium paratuberculosis infection in two llamas.
J.Am.Vet.Med.Assoc., 204(11):1805-8. Belloli A, Arrigoni N, Belletti GL, Proverbio D, Greppi G y Vacirca G. 1995. First result of paratuberculosis therapy in calves
intravenously infected with Mycobacterium paratuberculosis. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 144-6. IAP, Rehoboth, MA, USA.
Benazzi S, el Hamidi M y Schliesser T. 1996. Paratuberculosis in sheep flocks in Morocco: a serological, microscopical and cultural survey. Zentralbl.Veterinarmed.B, 43(4):213-9.
Bendixen PH. 1978. Immunological reactions caused by infection with Mycobacterium paratuberculosis. Nord.Vet.Med., 30:163-8. Bendixen PH, Bloch B y Jorgensen JB. 1981. Lack of intracellular degradation of Mycobacterium paratuberculosis by bovine
macrophages infected in vitro and in vivo: light microscopic and electron microscopic observations. Am.J.Vet.Res., 42(1):109-13.
Benedictus G. 1984. Evaluation of the organized control of paratuberculosis in the province of Friesland. Tijdschr.Diergeneeskd., 109(22):905-16.
Benedictus G y Bosma J. 1985a. Paratuberculosis: a surgical method of diagnosis in practice. Vet.Q., 7:217-21. Benedictus G, Dijkhuizen AA y Stelwagen J. 1985b. Economic losses to farms due to paratuberculosis in cattle.
Tijdschr.Diergeneeskd., 110(8):310-9. Benedictus G, Dijkhuizen AA y Stelwagen J. 1987. Economic losses due to paratuberculosis in dairy cattle. Vet.Rec., 121:142-6.

BIBLIOGRAFÍA CITADA
199
Benedictus G, Dinkla ETB y Wentink GH. 1988. Preliminary results of vaccination against paratuberculosis in adult dairy cattle. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 136-40. LCRV, Maisons-Alfort, France.
Benedictus G y Haagsma J. 1986. The efficacy of mesenteric lymph node biopsy in the eradication of paratuberculosis from an infected dairy farm. Vet.Q., 8:5-11.
Benedictus G y Kalis CHJ. 2003. Paratuberculosis: Eradication, Control and Diagnostic Methods. Acta Vet.Scand., 2003(44):231-41.
Bercovier H, Kafri O y Sela S. 1986. Mycobacteria possess a surprisingly small number of ribosomal RNA genes in relation to the size of their genome. Biochem.Biophys.Res.Commun., 136(3):1136-41.
Bergey GH, Harrison FC, Breed RS, Hammer BW y Huntoon FM. 1923, Bergey's Manual of Determinative Bacteriology. 1 Williams & Wilkins.Baltimore, MD, USA.
Berghaus RD, Farver TB, Anderson RJ, Adaska JM y Gardner IA. 2006a. Use of age and milk production data to improve the ability of enzyme-linked immunosorbent assay test results to predict Mycobacterium avium ssp. paratuberculosis fecal culture status. J.Vet.Diagn.Invest., 18(3):233-42.
Berghaus RD, Farver TB, Anderson RJ, Jaravata CC y Gardner IA. 2006b. Environmental sampling for detection of Mycobacterium avium ssp. paratuberculosis on large California dairies. J.Dairy Sci., 89(3):963-70.
Bernstein CN, Blanchard JF, Rawsthorne P y Collins MT. 2004. Population-based case control study of seroprevalence of Mycobacterium paratuberculosis in patients with Crohn's disease and ulcerative colitis. J.Clin.Microbiol., 42(3):1129-35.
Bernstein CN, Blanchard JF, Rawsthorne P y Wajda A. 1999. Epidemiology of Crohn's disease and ulcerative colitis in a central Canadian province: a population-based study. Am.J.Epidemiol., 149(10):916-24.
Beyerbach M, Ortmann G, Gerlach GF, Homuth M, Strutzberg K y Kreienbrock L. 2004. Considerations concerning diagnostic certainties and cut-off values for a bulk milk ELISA for Mycobacterium avium ssp. paratuberculosis. Dtsch.Tierarztl.Wochenschr., 111(5):220-5.
Beyerbach M, Rehm T, Kreienbrock L y Gerlach GF. 2001. Eradication of paratuberculosis in dairy herds: determination of the initial herd prevalence and modelling of prevalence development. Dtsch.Tierarztl.Wochenschr., 108(7):291-6.
Bhide M, Chakurkar E, Tkacikova L, Barbuddhe S, Novak M y Mikula I. 2006. IS900-PCR-based detection and characterization of Mycobacterium avium subsp. paratuberculosis from buffy coat of cattle and sheep. Vet.Microbiol., 112(1):33-41.
Billman-Jacobe H, Carrigan MJ, Cockram F, Corner LA, Gill IJ, Hill JF, Jessep T, Milner AR y Wood PR. 1992. A comparison of the interferon gamma assay with the absorbed ELISA for the diagnosis of Johne's disease in cattle. Aust.Vet.J., 69(2):25-8.
Blanchard B, Versmisse Y, Chatonnet S, Rebeyroles L, Fremont A y Couquet C. 2005. Detection of Mycobacterium avium subsp. paratubverculosis in bovine feces and milk using Adiapure extraction kit and Real Time Adiavet PCR kit. En: L Mewes and S Kenklies editores. 5. Internationales Symposium zur BHV 1-, BVD- und Paratuberkulose-Bekämpfung, Landesamt für Verbraucherschutz Sachsen-Anhalt. Stendal Bundesland Sachsen-Anhalt, Deutschland.
Blood DC y Radostits OM. 1989. Paratuberculosis (Johne's disease). Veterinary Medicine. ed. 7. pp. 722-9. Baillière & Tindall. London, UK.
Blood DC, Radostits OM y Henderson JA. 1983. Johne's disease. Veterinary Medicine. ed. 6. pp. 643-9. Baillière & Tindall. London, UK.
Boddinghaus B, Rogall T, Flohr T, Blocker H y Bottger EC. 1990. Detection and identification of mycobacteria by amplification of rRNA. J.Clin.Microbiol., 28(8):1751-9.
Boelaert F, Walravens K, Biront P, Vermeersch JP, Berkvens D y Godfroid J. 2000. Prevalence of paratuberculosis (Johne's disease) in the Belgian cattle population. Vet.Microbiol., 77(3-4):269-81.
Boever WJ y Peters D. 1974. Paratuberculosis in two herds of exotic sheep. J.Am.Vet.Med.Assoc., 165(9):822. Bogli-Stuber K, Kohler C, Seitert G, Glanemann B, Antognoli MC, Salman MD, Wittenbrink MM, Wittwer M, Wassenaar T,
Jemmi T y Bissig-Choisat B. 2005. Detection of Mycobacterium avium subspecies paratuberculosis in Swiss dairy cattle by real-time PCR and culture: a comparison of the two assays. J.Appl.Microbiol, 99(3):587-97.
Böhn R y Ley T. 1994. Desinfection of Mycobacterium paratuberculosis in cattle slurry. En: Diesch editor. Proc.8th Int.Congr.Anim.Hyg., p. DD10-DD15. ISAH, St. Paul, MN, USA.
Bolske G, Sternberg S, Viske D y Larsson B. 1999. Use of culture and serology in tracing and screening for paratuberculosis in Sweden. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 349-52. IAP, Providence, RI, USA.
Borody TJ, Leis S, Warren EF y Surace R. 2002. Treatment of severe Crohn's disease using antimycobacterial triple therapy: approaching a cure? Dig.Liver Dis., 34(1):29-38.
Bosshard C, Stephan R y Tasara T. 2006. Application of an F57 sequence-based real-time PCR assay for Mycobacterium paratuberculosis detection in bulk tank raw milk and slaughtered healthy dairy cows. J.Food Prot., 69(7):1662-7.
Braun RK y Buergelt CD. 1990. Use of an enzyme-linked immunosorbent assay to estimate prevalence of paratuberculosis in cattle of Florida. J.Am.Vet.Med.Assoc., 196:1251-4.
Brees DJ, Reimer SB, Cheville NF, Florance A y Thoen CO. 2000. Immunohistochemical detection of Mycobacterium paratuberculosis in formalin-fixed, paraffin-embedded bovine tissue sections. J.Vet.Diagn.Invest., 12(1):60-3.
Brett E. 1998, Johne's disease: an economic evaluation of control options for the New Zealand livestock industries. Agriculture New Zealand.Wellington, NZ.
Brey BJ, Radcliff RP, Clark DL, Jr. y Ellingson JL. 2006. Design and development of an internal control plasmid for the detection of Mycobacterium avium subsp. paratuberculosis using real-time PCR. Mol.Cell.Probes, 20(1):51-9.
Brotherston JG y Gilmour NJL. 1961a. Quantitative studies of Mycobacterium johnei in the tissues of sheep. I Routes of infection and assay of viable M. johnei. J.Comp.Pathol., 71:286-99.
Brotherston JG, Gilmour NJL y Samuel JM. 1961b. Quantitative studies of Mycobacterium johnei in the tissues of sheep. II Protection afforded by dead vaccines. J.Comp.Pathol., 71:300-11.
Brugère-Picoux J. 1987. Le diagnostic de la paratuberculare chez les ruminants. Rec.Med.Vet.Ec.Alfort, 163(5):539-46. Brumbaugh GW, Edwards JF, Roussel AJ y Thomson TD. 2000. Effect of monensin sodium on histological lesions of naturally
occurring bovine paratuberculosis. J.Comp.Pathol., 123:22-8. Brunello F, Pera A, Martini S, Marino L, Astegiano M, Barletti C, Gastaldi P, Verme G y Emmanuelli G. 1991. Antibodies to
Mycobacterium paratuberculosis in patients with Crohn's disease. Dig.Dis.Sci., 36:1741-5. Buergelt CD y Duncan JR. 1978a. Age and milk production data of cattle culled from a dairy herd with paratuberculosis.
J.Am.Vet.Med.Assoc., 173(5):478-80. Buergelt CD y Ginn PE. 2000. The histopathologic diagnosis of subclinical Johne's disease in North American bison (Bison bison).
Vet.Microbiol., 77(3-4):325-31. Buergelt CD, Hall C, McEntee K y Duncan JR. 1978b. Pathological evaluation of paratuberculosis in naturally infected cattle.
Vet.Pathol., 15:196-206.

BIBLIOGRAFÍA CITADA
200
Buergelt CD y Williams JE. 2004. Nested PCR on blood and milk for the detection of Mycobacterium avium subsp. paratuberculosis DNA in clinical and subclinical bovine paratuberculosis. Aust.Vet.J., 82(8):497-503.
Buffa M, Guamis B, Royo C y Trujillo AJ. 2001. Microbiological changes throughout ripening of goat cheese made from raw, pasteurized and high-pressure-treated milk. Food Microbiology, 18(1):45-51.
Bulaga LL y Collins MT. 1999. U.S. voluntary Johne's disease herd status program for cattle. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 39-47. IAP, Madison, WI, USA.
Bull TJ, Hermon-Taylor J, Pavlik I, El Zaatari F y Tizard M. 2000. Characterization of IS900 loci in Mycobacterium avium subsp. paratuberculosis and development of multiplex PCR typing. Microbiology, 146:2185-97.
Bull TJ, McMinn EJ, Sidi-Boumedine K, Skull A, Durkin D, Neild P, Rhodes G, Pickup R y Hermon-Taylor J. 2003a. Detection and verification of Mycobacterium avium subsp. paratuberculosis in fresh ileocolonic mucosal biopsy specimens from individuals with and without Crohn's disease. J.Clin.Microbiol., 41(7):2915-23.
Bull TJ, Sidi-Boumedine K, McMinn EJ, Stevenson K, Pickup R y Hermon-Taylor J. 2003b. Mycobacterial interspersed repetitive units (MIRU) differentiate Mycobacterium avium subspecies paratuberculosis from other species of the Mycobacterium avium complex. Mol.Cell.Probes, 17(4):157-64.
Burns DN, Wallace RJ, Jr., Schultz ME, Zhang YS, Zubairi SQ, Pang YJ, Gibert CL, Brown BA, Noel ES y Gordin FM. 1991. Nosocomial outbreak of respiratory tract colonization with Mycobacterium fortuitum: demonstration of the usefulness of pulsed-field gel electrophoresis in an epidemiologic investigation. Am.Rev.Respir.Dis., 144(5):1153-9.
Burrells C, Clarke CJ, Colston A, Kay JM, Porter J, Little D y Sharp JM. 1998. A study of immunological responses of sheep clinically-affected with paratuberculosis (Johne's disease). The relationship of blood, mesenteric lymph node and intestinal lymphocyte responses to gross and microscopic pathology. Vet.Immunol.Immunopathol., 66:343-58.
Burrells C, Clarke CJ, Colston A, Kay JM, Porter J, Little D y Sharp JM. 1999. Interferon-gamma and interleukin-2 release by lymphocytes derived from the blood, mesenteric lymph nodes and intestines of normal sheep and those affected with paratuberculosis (Johne's disease). Vet.Immunol.Immunopathol., 68:139-48.
Burton MS, Olsen JH, Ball RL y Dumonceaux GA. 2001. Mycobacterium avium subsp. paratuberculosis infection in an addax (Addax nasomaculatus). J.Zoo Wildl.Med., 32(2):242-4.
Butz P y Ludwig H. 1986. Pressure Inactivation of Microorganisms at Moderate Temperatures. Physica B + C, 139(1-3):875-7. Buza JJ, Hikono H, Mori Y, Nagata R, Hirayama S, Aodon g, Bari AM, Shu Y, Tsuji NM y Momotani E. 2004. Neutralization of
interleukin-10 significantly enhances gamma interferon expression in peripheral blood by stimulation with Johnin purified protein derivative and by infection with Mycobacterium avium subsp. paratuberculosis in experimentally infected cattle with paratuberculosis. Infect.Immun., 72(4):2425-8.
Caldow GL. 2006. The prospects for herd level control of paratuberculosis in cattle: a consultant's view. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 165-73. IAP, Madison, WI, USA.
Carpenter T, Gardner IA, Collins MT y Whitlock RH. 2004. Effects of prevalence and testing by enzyme-linked immunosorbent assay and fecal culture on the risk of introduction of Mycobacterium avium subsp. paratuberculosis-infected cows into dairy herds. J.Vet.Diagn.Invest., 16(1):31-8.
Carrera J y Bener AM. 1989. La paratuberculosis en la Comunidad Autónoma de Cataluña. En: Juste editor. 2ª Reu.PTBC España (1987), p. 45. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España.
Carrigan MJ y Seaman JT. 1990. The pathology of Johne's disease in sheep. Aust.Vet.J., 67(2):47-50. Cave MD, Eisenach KD, McDermott PF, Bates JH y Crawford JT. 1991. IS6110: Conservation of sequence in the Mycobacterium
tuberculosis complex and its utilization in DNA fingerprinting. Mol.Cell.Probes, 5(1):73-80. Cavirani S, Taddei S, Ossiprandi MC, Iotti A y Bottarelli E. 2006. Farm factors associated with Mycobacterium avium subespecies
paratuberculosis infections in Northern Italian dairy herds. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 182-6. IAP, Madison, WI, USA.
Cavusoglu C, Turhan A, Yaygin YE, Karaca Derici Y y Bilgic A. 2005. Evaluation of INNO-LiPA mycobacteria, INNO-LiPA Mycobacteria v2 and hsp65 sequencing for identification of the clinical nontuberculous mycobacterial isolates. Mikrobiyol.Bul., 39(4):437-45.
Cellier C, De Beenhouwer H, Berger A, Penna C, Carbonnel F, Parc R, Cugnenc PH, Le Quintrec Y, Gendre JP, Barbier JP y Portaels F. 1998. Mycobacterium paratuberculosis and Mycobacterium avium subsp. silvaticum DNA cannot be detected by PCR in Crohn's disease tissue. Gastroenterol.Clin.Biol., 22(8-9):675-8.
Çetinkaya B, Egan K, Harbour DA y Morgan KL. 1996. An abattoir-based study of the prevalence of subclinical Johne's disease in adult cattle in south west England. Epidemiol.Infect., 116:373-9.
Çetinkaya B, Erdogan HM y Morgan KL. 1998. Prevalence, incidence and geographical distribution of Johne's disease in cattle in England and the Welsh borders. Vet.Rec., 143(10):265-9.
Chacon O, Bermudez LEM y Barletta RG. 2004. Johne's Disease, Inflammatory Bowel Disease, and Mycobacterium paratuberculosis. Annu.Rev.Microbiol., 58:329-63.
Challans JA, Stevenson K, Reid HW y Sharp JM. 1994. A rapid method for detection and extraction of Mycobacterium avium subspecies paratuberculosis from clinical specimens. Vet.Rec., 134:95-6.
Chamberlin W, Graham DY, Hulten K, El-Zimaity HM, Schwartz MR, Naser S, Shafran I y El-Zaatari FA. 2001. Review article: Mycobacterium avium subsp. paratuberculosis as one cause of Crohn's disease. Aliment.Pharmacol.Ther., 15(3):337-46.
Chamberlin WM y Naser SA. 2006. Integrating theories of the etiology of Crohn's disease. On the etiology of Crohn's disease: questioning the hypotheses. Med.Sci.Monit., 12(2):RA27-33.
Chávez G. 1993. Estudio comparativo de las lesiones y de la respuesta inmunológica observadas en corderos infectados experimentalmente con Mycobacterium paratuberculosis y Mycobacterium avium subsp. silvaticum. Tesis Doctoral. Universidad de Zaragoza. Zaragoza, España.
Chevallier B, Versmisse Y y Blanchard B. 2003. Development of a PCR test to detect Mycobacterium avium subsp. paratuberculosis in bovine faeces. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 247-50. IAP, Madison, WI, USA.
Cheville NF, Hostetter J, Thomsen BV, Simutis F, Vanloubbeeck Y y Steadham E. 2001. Intracellular trafficking of Mycobacterium avium ss. paratuberculosis in macrophages. Dtsch.Tierarztl.Wochenschr., 108(6):236-43.
Chi J, VanLeeuwen J, Weersink A y Keefe G. 2002. Direct production losses and treatment costs from bovine viral diarrhoea virus, bovine leukosis virus, Mycobacterium avium subspecies paratuberculosis, and Neospora caninum. Prev.Vet.Med., 55(2):137.
Chiba M, Fukushima T, Horie Y, Iizuka M y Masamune O. 1998. No Mycobacterium paratuberculosis detected in intestinal tissue, including Peyer's patches and lymph follicles, of Crohn's disease. J.Gastroenterol., 33(4):482-7.
Chiodini RJ. 1983. Crohn's disease. En: Merkal editor. Proc.1st Int.Coll.Paratub., pp. 198-9. NADC, USDA, Ames, IA, USA. Chiodini RJ. 1993. Abolish Mycobacterium paratuberculosis Strain 18. J.Clin.Microbiol., 31:1956-8. Chiodini RJ. 1996. Immunology: resistance to paratuberculosis. Vet.Clin.North Am.Food Anim.Pract., 12(2):313-43.

BIBLIOGRAFÍA CITADA
201
Chiodini RJ. 1997. Debate: M. paratuberculosis in foods and the public health implications (Pro associacion between Johne's disease and Crohn's disease). En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., pp. 353-65. IAP, Rehoboth, MA, USA.
Chiodini RJ y Davis W. 1992. The cellular immunology of bovine paratuberculosis: the predominant response is mediated by cytotoxic gamma/delta T lymphocytes which prevent CD4+ activity. Microb.Pathog., 13:447-63.
Chiodini RJ, Kreeger JM y Thayer WR. 1993. Use of rifabutin in treatment of systemic Mycobacterium paratuberculosis infection in mice. Antimicrob.Agents Chemother., 37(8):1645-8.
Chiodini RJ y Van Kruiningen HJ. 1983. Eastern white-tailed deer as a reservoir of ruminant paratuberculosis. J.Am.Vet.Med.Assoc., 182(2):168-9.
Chiodini RJ y Van Kruiningen HJ. 1986a. The prevalence of paratuberculosis in culled New England cattle. Cornell Vet., 76:91-104.
Chiodini RJ, Van Kruiningen HJ y Merkal RS. 1984a. Ruminant paratuberculosis (Johne's disease): The current status and future prospects. Cornell Vet., 74:218-62.
Chiodini RJ, Van Kruiningen HJ, Merkal RS, Thayer WR y Coutu JA. 1984b. Characteristics of an unclassified Mycobacterium species isolated from patients with Crohn's disease. J.Clin.Microbiol., 20(5):966-71.
Chiodini RJ, Van Kruiningen HJ, Thayer WR y Coutu JA. 1986b. Spheroplastic phase of mycobacteria isolated from patients with Crohn's disease. J.Clin.Microbiol., 24(3):357-63.
Chiodini RJ, Van Kruiningen HJ, Thayer WR, Merkal RS y Coutu JA. 1984c. Possible role of mycobacteria in inflammatory bowel disease. I. An unclassified Mycobacterium species isolated from patients with Crohn's disease. Dig.Dis.Sci., 29(12):1073-9.
Cho D y Collins MT. 2006a. Comparison of the proteosomes and antigenicities of secreted and cellular proteins produced by Mycobacterium paratuberculosis. Clin.Vaccine Immunol., 13(10):1155-61.
Cho D, Sung N y Collins MT. 2006b. Identification of proteins of potential diagnostic value for bovine paratuberculosis. Proteomics, 6(21):5785-94.
Choy E, Whittington RJ, Marsh I, Marshall J y Campbell MT. 1998. A method for purification and characterization of Mycobacterium avium subsp. paratuberculosis from the intestinal mucosa of sheep with Johne's daisease. Vet.Microbiol., 64(1):51-60.
Christopher-Hennings J, Dammen MA, Weeks SR, Epperson WB, Singh SN, Steinlicht GL, Fang Y, Skaare JL, Larsen JL, Payeur JB y Nelson EA. 2003. Comparison of two DNA extractions and nested PCR, real-time PCR, a new commercial PCR assay, and bacterial culture for detection of Mycobacterium avium subsp. paratuberculosis in bovine feces. J.Vet.Diagn.Invest., 15(2):87-93.
Chui LW, King R, Lu P, Manninen K y Sim J. 2004. Evaluation of four DNA extraction methods for the detection of Mycobacterium avium subsp. paratuberculosis by polymerase chain reaction. Diagn.Microbiol.Infect.Dis., 48: 39-45.
Clark DL, Jr., Anderson JL, Koziczkowski JJ y Ellingson JL. 2006. Detection of Mycobacterium avium subspecies paratuberculosis genetic components in retail cheese curds purchased in Wisconsin and Minnesota by PCR. Mol.Cell.Probes, 20(3-4):197-202.
Clarke CJ. 1997a. Paratuberculosis and molecular biology. Vet.J., 153:245-7. Clarke CJ. 1997b. The pathology and pathogenesis of paratuberculosis in ruminants and other species. J.Comp.Pathol., 116:217-61. Clarke CJ y Little D. 1996a. The pathology of ovine paratuberculosis: gross and histological changes in the intestine and other
tissues. J.Comp.Pathol., 114(4):419-37. Clarke CJ, Patterson DSP, Armstrong KE y Low JC. 1996b. Comparison of the absorbed ELISA and agar gel immunodiffusion test
with clinicopathological findings in ovine clinical paratuberculosis. Vet.Rec., 139:618-21. Clarkston WK, Presti ME, Petersen PF, Zachary PE, Fan WX, Leonardi CL, Vernava AM, Longo WE y Kreeger JM. 1998. Role of
Mycobacterium paratuberculosis in Crohn's disease: a prospective, controlled study using polymerase chain reaction. Dis.Colon Rectum, 41(2):195-9.
Cocito C, Gilot P, Coene M, De Kesel M, Poupart P y Vannuffel P. 1994. Paratuberculosis. Clin.Microbiol.Rev., 7(3):328-45. Coelho AC, Pinto ML, Silva S, Coelho AM, Rodrigues J y Juste RA. 2006. Seroprevalence of ovine paratuberculosis infection in
the Northeast of Portugal. Small Rumin.Res., In press. Coetsier C, Havaux X, Mattelard F, Sadatte S, Cormont F, Buergelt K, Limbourg B, Latinne D, Bazin H, Denef JF y Cocito C.
1998. Detection of Mycobacterium avium subsp. paratuberculosis in infected tissues by new species-specific immunohistological procedures. Clin.Diagn.Lab.Immunol., 5(4):446-51.
Coetsier C, Vannuffel P, Blondeel N, Denef JF, Cocito C y Gala JL. 2000. Duplex PCR for differential identification of Mycobacterium bovis, M. avium and M. avium subsp. paratuberculosis in formalin-fixed paraffin-embedded tissues from cattle. J.Clin.Microbiol., 38(8):3048-54.
Coffin JW, Condon C, Compston CA, Potter KN, Lamontagne LR, Shafiq J y Kunimoto DY. 1992. Use of restriction fragment length polymorphisms resolved by pulsed-field gel electrophoresis for subspecies identification of mycobacteria in the Mycobacterium avium complex and for isolation of DNA probes. J.Clin.Microbiol., 30(7):1829-36.
Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, Gordon SV, Eiglmeier K, Gas S, Barry CE, III, Tekaia F, Badcock K, Basham D, Brown D, Chillingworth T, Connor R, Davies R, Devlin K, Feltwell T, Gentles S, Hamlin N, Holroyd S, Hornsby T, Jagels K, Krogh A, Mclean J, Moule S, Murphy L, Oliver K, Osborne J, Quail MA, Rajandream MA, Rogers J, Rutter S, Seeger K, Skelton J, Squares R, Squares S, Sulston JE, Taylor K, Whitehead S y Barrell BG. 1998. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature, 393(6685):537-44.
Cole ST, Eiglmeier K, Parkhill J, James KD, Thomson NR, Wheeler PR, Honore N, Garnier T, Churcher C, Harris D, Mungall K, Basham D, Brown D, Chillingworth T, Connor R, Davies RM, Devlin K, Duthoy S, Feltwell T, Fraser A, Hamlin N, Holroyd S, Hornsby T, Jagels K, Lacroix C, Maclean J, Moule S, Murphy L, Oliver K, Quail MA, Rajandream MA, Rutherford KM, Rutter S, Seeger K, Simon S, Simmonds M, Skelton J, Squares R, Squares S, Stevens K, Taylor K, Whitehead S, Woodward JR y Barrell BG. 2001. Massive gene decay in the leprosy bacillus. Nature, 409(6823):1007-11.
Colgrove GS, Thoen CO, Blackburn BO y Murphy CD. 1989. Paratuberculosis in cattle: a comparison of three serologic tests with results of fecal culture. Vet.Microbiol., 19:183-7.
Collins DM, Gabric DM y de Lisle GW. 1989. Identification of a repetitive DNA sequence specific to Mycobacterium paratuberculosis. FEMS Microbiol.Lett., 60(2):175-8.
Collins DM, Gabric DM y de Lisle GW. 1990a. Identification of two groups of Mycobacterium paratuberculosis strains by restriction analysis and DNA hybridization. J.Clin.Microbiol., 28(9):1591-6.
Collins DM, Hilbink F, West DM, Hosie BD, Cooke MM y de Lisle GW. 1993a. Investigation of Mycobacterium paratuberculosis in sheep by fecal culture, DNA characterisation and the polymerase chain reaction. Vet.Rec., 133:599-600.
Collins DM y Stephens DM. 1991. Identification of an insertion sequence, IS1081, in Mycobacterium bovis. FEMS Microbiol.Lett., 67(1):11-5.

BIBLIOGRAFÍA CITADA
202
Collins DM, Stephens DM y de Lisle GW. 1993b. Comparison of polymerase chain reaction tests and faecal culture for detecting Mycobacterium paratuberculosis in bovine faeces. Vet.Microbiol., 36:289-99.
Collins MT. 1995. Johne's disease diagnosis and control. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 325-44. IAP, Rehoboth, MA, USA.
Collins MT. 1996. Diagnosis of paratuberculosis. Vet.Clin.North Am.Food Anim.Pract., 12(2):357-71. Collins MT. 1997. Mycobacterium paratuberculosis: a potential food-borne pathogen? J.Dairy.Sci., 80(12):3445-8. Collins MT. 2002. Interpretation of a commercial bovine paratuberculosis enzime-linked immunosorbent assay by using likelihood
ratios. Clin.Diagn.Lab.Immunol., 9(6):1367-71. Collins MT. 2006. Paratuberculosis diagnostics: the cost versus sensitivity trade-off. En: Manning y Nielsen editores. Proc.8th
Int.Coll.Paratub., pp. 459-62. IAP, Madison, WI, USA. Collins MT, Angulo A, Buergelt CD, Hennager SG, Hietala SK, Jacobson RH y Whipple DF. 1993c. Reproducibility of a comercial
enzyme-linked immunosorbent assay for bovine paratuberculosis among eight laboratories. J.Vet.Diagn.Invest., 5:52-5. Collins MT, Keneflick KB, Sockett DC, Lambrecht RS, McDonald J y Jorgensen J. 1990b. Enhanced radiometric detection of
Mycobacterium paratuberculosis by using filter-concentrated bovine fecal specimens. J.Clin.Microbiol., 28:2514-9. Collins MT, Lambrecht RS y McDonald J. 1988. Radiometric detection of M. paratuberculosis from clinical specimens. En: Thorel
y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 179-98. LCRV, Maisons-Alfort, France. Collins MT, Lisby G, Moser C, Chicks D, Christensen S, Reichelderfer M, Hoiby N, Harms BA, Thomsen OO, Skibsted U y Binder
V. 2000. Results of multiple diagnostic tests for Mycobacterium avium subsp. paratuberculosis in patients with inflammatory bowel disease and in controls. J.Clin.Microbiol., 38(12):4373-81.
Collins MT, Sockett DC, Goodger WJ, Conrad TA, Thomas CB y Carr DJ. 1994. Herd prevalence and geographic distribution of, and risk factors for, bovine paratuberculosis in Wisconsin. J.Am.Vet.Med.Assoc., 204(4):636-41.
Collins MT, Wells SJ, Petrini KR, Collins JE, Schultz RD y Whitlock RH. 2005. Evaluation of five antibody detection tests for diagnosis of bovine paratuberculosis. Clin.Diagn.Lab.Immunol., 12(6):685-92.
Colston A, McConnell I y Bujdoso R. 1994. Cloning and expression in Escherichia coli of DNA encoding a 60 kDa stress protein of Mycobacterium paratuberculosis, the causative agent of Johne's disease. Microbiology, 140:3329-36.
Condron RJ, Schroen CJ, Black CA, Ridge SE y Hope A. 1995. Histological confirmation of subclinical infection with M. paratuberculosis in cattle. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 37-40. IAP, Providence, RI, USA.
Cook WE, Cornish TE, Shideler S, Lasley B y Collins MT. 1997. Radiometric culture of Mycobacterium avium paratuberculosis from the feces of tule elk. J.Wildl.Dis., 33(3):635-7.
Corn JL, Manning EJ, Sreevatsan S y Fischer JR. 2005. Isolation of Mycobacterium avium subsp. paratuberculosis from free-ranging birds and mammals on livestock premises. Appl.Environ.Microbiol., 71(11):6963-7.
Corner LA, Pfeiffer DU y Abbott KA. 2004. The respiratory tract as a hypothetical route of infection of cattle with Mycobacterium avium subspecies paratuberculosis. Aust.Vet.J., 82(3):170-3.
Corpa JM. 1999. Aspectos inmunopatológicos de la paratuberculosis de los pequeños rumiantes. Respuesta inmune asociada a la vacunación. Tesis Doctoral. Universidad de León. León, España.
Corpa JM, García Marín JF y Pérez V. 1997. Diagnóstico de paratuberculosis en ganado vacuno. Med.Vet., 15(11):618-24. Corpa JM, García Marín JF y Pérez V. 1999. Evolución de la respuesta inmune frente a paratuberculosis en ganado ovino y caprino
según la edad de vacunación. Med.Vet., 16(3):162-6. Corpa JM, Garrido JM, García Marín JF y Pérez V. 2000a. Classification of lesions observed in natural cases of paratuberculosis in
goats. J.Comp.Pathol., 122(4):255-65. Corpa JM, Pérez V y García Marin JF. 2000b. Control of paratuberculosis (Johne's disease) in goats by vaccination of adult animals.
Vet.Rec., 146:195-6. Corti S y Stephan R. 2002. Detection of Mycobacterium avium subspecies paratuberculosis specific IS900 insertion sequences in
bulk-tank milk samples obtained from different regions throughout Switzerland. BMC Microbiol., 2(1):15. Cousins DV, Evans RJ y Francis BR. 1995. Use of BACTEC radiometric culture method and polymerase chain reaction for the
rapid screening of faeces and tissues for Mycobacterium paratuberculosis. Aust.Vet.J., 72(12):458-62. Cousins DV, Whittington R, Marsh I, Masters A, Evans RJ y Kluver P. 1999. Mycobacteria distinct from Mycobacterium avium
subsp. paratuberculosis isolated from the faeces of ruminants posses IS900-like sequences detectable by IS900 polymerase chain reaction: implications for the diagnosis. Mol.Cell.Probes, 13(6):431-42.
Cousins DV, Williams SN, Hope A y Eamens GJ. 2000. DNA fingerprinting of Australian isolates of Mycobacterium avium subsp. paratuberculosis using IS900 RFLP. Aust.Vet.J., 78(3):184-90.
Coussens PM. 2004. Model for immune responses to Mycobacterium avium subspecies paratuberculosis in cattle. Infect.Immun., 72(6):3089-96.
Coussens PM, Chiang SK, Aho A y Kuipel M. 2006. Relationship between Mycobacterium avium subspecies paratuberculosis, IL-1a, and TRAF1 in primary bovine macrophages. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 146. IAP, Madison, WI, USA.
Coussens PM, Colvin CJ, Rosa GJ, Perez LJ y Elftman MD. 2003. Evidence for a novel gene expression program in peripheral blood mononuclear cells from Mycobacterium avium subsp. paratuberculosis infected cattle. Infect.Immun., 71(11):6487-98.
Coussens PM, Colvin CJ, Wiersma K, Abouzied A y Sipkovsky S. 2002. Gene expression profiling of peripheral blood mononuclear cells from cattle infected with Mycobacterium paratuberculosis. Infect.Immun., 70(10):5494-502.
Coussens PM, Pudrith CB, Skovgaard K, Ren X, Suchyta SP, Stabel JR y Heegaard PM. 2005. Johne's disease in cattle is associated with enhanced expression of genes encoding IL-5, GATA-3, tissue inhibitors of matrix metalloproteinases 1 and 2, and factors promoting apoptosis in peripheral blood mononuclear cells. Vet.Immunol.Immunopathol., 105(3-4):221-34.
Coussens PM, Verman N, Coussens MA, Elftman MD y McNulty AM. 2004. Cytokine gene expression in peripheral blood mononuclear cells and tissues of cattle infected with Mycobacterium avium subsp. paratuberculosis: evidence for an inherent proinflammatory gene expression pattern. Infect.Immun., 72(3):1409-22.
Cox JC, Drane DP, Jones SL, Ridge SE y Milner AR. 1991. Development and evaluation of a rapid absorbed enzyme immuno assay test for the diagnosis of Johne's disease in cattle. Aust.Vet.J., 68(5):157-60.
Cranwell MP. 1988. Johne's disease in a flock of sheep: presenting clinical and pathological signs, losses sustained and control methods adopted. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 71-4. LCRV, Maisons-Alfort, France.
Crawford JT, Cave MD y Bates JH. 1981. Characterization of plasmids from strains of Mycobacterium avium-intracellulare. Rev.Infect.Dis., 3(5):949-52.
Crohn BB, Ginzburg L y Oppenheimer G. 1932. Regional ileitis: a pathologic and clinical entity. J.Am.Med.Assoc., 99:1323-9. Crossley BM, Zagmutt-Vergara FJ, Fyock TL, Whitlock RH y Gardner IA. 2005. Fecal shedding of Mycobacterium avium subsp.
paratuberculosis by dairy cows. Vet.Microbiol., 107(3-4):257-63.

BIBLIOGRAFÍA CITADA
203
Crowther RW, Polydorou K, Nitti S y Phyrilla A. 1976. Johne's disease in sheep in Cyprus. Vet.Rec., 98:463. Cummins RE, Mullins D, Smith LJ y Ford MJ. 1996. Metastatic Crohn's disease: a case report. Pediatr.Dermatol., 13(1):25-8. Cuoghi F. 1993. Use of high-pressure in the food-industry: cold sterilization of milk and influence on its properties. Ind.Aliment.,
32(319):956-64. Cvetnic Z. 2000. Paratuberculosis in cattle in the Republic of Croatia. Vet.Stanika, 31(1):21-5. Daffe M y Draper P. 1998. The envelope layers of mycobacteria with reference to their pathogenicity. Adv.Microb.Physiol., 39:131-
203. Dale JW. 1995. Mobile genetic elements in mycobacteria. Eur.Respir.J.Suppl., 20:633s-48s. Dalziel TK. 1913. Chronic interstitial enteritis. Br.Med.J.(ii):1068-70. Damato JJ y Collins MT. 1984. Radiometric studies with gas-liquid and thin layer chromatography for rapid demonstratation of
hemin dependence and characterization of Mycobacterium haemophilum. J.Clin.Microbiol., 20(3):515-8. Damato JJ y Collins MT. 1990. Growth of Mycobacterium paratuberculosis in radiometric, Middlebrook and egg-based media.
Vet.Microbiol., 22:31-42. Damato JJ, Collins MT y McClatchy JK. 1982. Urease testing of Mycobacteria with BACTEC radiometric instrumentation.
J.Clin.Microbiol., 15(3):478-80. Damato JJ, Knisley C y Collins MT. 1987. Characterization of Mycobacterium paratuberculosis by gas-liquid and thin-layer
chromatography and rapid demonstration of mycobactin dependence using radiometric methods. J.Clin.Microbiol., 25(12):2380-3.
Daniels MJ, Ball N, Hutchings MR y Greig A. 2001. The grazing response of cattle to pasture contaminated with rabbit faeces and the implications for the transmission of paratuberculosis. Vet.J., 161(3):306-13.
Daniels MJ, Henderson D, Greig A, Stevenson K, Sharp JM y Hutchings MR. 2003a. The potential role of wild rabbits Oryctolagus cuniculus in the epidemiology of paratuberculosis in domestic ruminants. Epidemiol.Infect., 130(3):553-9.
Daniels MJ, Hutchings MR, Beard PM, Henderson D, Greig A, Stevenson K y Sharp JM. 2003b. Do non-ruminant wildlife pose a risk of paratuberculosis to domestic livestock and vice versa in Scotland? J.Wildl.Dis., 39(1):10-5.
Daniels MJ, Hutchings MR y Greig A. 2003c. The risk of disease transmission to livestock posed by contamination of farm stored feed by wildlife excreta. Epidemiol.Infect., 130(3):561-8.
Daniels MJ, Lees JD, Hutchings MR y Greig A. 2003d. The ranging behaviour and habitat use of rabbits on farmland and their potential role in the epidemiology of paratuberculosis. Vet.J., 165(3):248-57.
Dargatz DA, Byrum BA, Barber LK, Sweeney RW, Whitlock RH, Shulaw WP, Jacobson RH y Stabel JR. 2001a. Evaluation of a commercial ELISA for diagnosis of paratuberculosis in cattle. J.Am.Vet.Med.Assoc., 218(7):1163-6.
Dargatz DA, Byrum BA, Hennager SG, Barber LK, Kopral CA, Wagner BA y Wells SJ. 2001b. Prevalence of antibodies against Mycobacterium avium subsp. paratuberculosis among beef cow-calf herds. J.Am.Vet.Med.Assoc., 219(4):497-501.
Das SK, Sinha RP y Chauhan HVS. 1992. Chemoterapy of paratuberculosis in goats: streptimycin, rifampicin and levamisole versus streptomycin, rifampicin and dapsone. Ind.J.Anim.Sci., 62(1):8-13.
Das SK, Sinha RP y Soman JP. 1991. Isolation and characterization of Mycobacterium paratuberculosis from goats. Ind.J.Anim.Sci., 61(12):1289-91.
Davidson WR, Manning EJ y Nettles VF. 2004. Culture and serologic survey for Mycobacterium avium subsp. paratuberculosis infection among southeastern white-tailed deer (Odocoileus virginianus). J.Wildl.Dis., 40(2):301-6.
de Juan L, Alvarez J, Aranaz A, Rodriguez A, Romero B, Bezos J, Mateos A y Dominguez L. 2006a. Molecular epidemiology of Types I/III strains of Mycobacterium avium subspecies paratuberculosis isolated from goats and cattle. Vet.Microbiol., 115(1-3):102-10.
de Juan L, Alvarez J, Romero B, Bezos J, Castellanos E, Aranaz A, Mateos A y Dominguez L. 2006b. Comparison of four different culture media for isolation and growth of type II and type I/III Mycobacterium avium subsp. paratuberculosis strains isolated from cattle and goats. Appl.Environ.Microbiol., 72(9):5927-32.
de Juan L, Mateos A, Dominguez L, Sharp JM y Stevenson K. 2005. Genetic diversity of Mycobacterium avium subspecies paratuberculosis isolates from goats detected by pulsed-field gel electrophoresis. Vet.Microbiol., 106(3-4):249-57.
De Kesel M, Gilot P, Coene M y Cocito C. 1992. Composition and immunological properties of the protein fraction of A36, a major antigen complex of Mycobacterium paratuberculosis. Scand.J.Immunol., 36(2):201-12.
de Lisle GW, Cannon MC, Yates GF y Collins DM. 2006. Abattoir surveillance of paratuberculosis in farmed deer in New Zealand. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 601-4. IAP, Madison, WI, USA.
de Lisle GW, Collins DM y Huchzermeyer HFAK. 1992. Characterization of ovine strains of Mycobacterium paratuberculosis by restriction endonuclease analysis and DNA hybridization. Onderstepoort J.Vet.Res., 59:163-5.
de Lisle GW, Seguin P, Corner AH y Duncan JR. 1980. Bovine paratuberculosis I: A herd study using complement fixation and intradermal test. Can.J.Comp.Med., 44:177-82.
de Lisle GW, Yates GF y Collins DM. 1993. Paratuberculosis in farmed deer: case reports and DNA characterization of isolates of Mycobacterium paratuberculosis. J.Vet.Diagn.Invest., 5:567-71.
de Lisle GW, Yates GF y Montgomery H. 2003. The emergence of Mycobacterium paratuberculosis in farmed deer in New Zealand - a review of 619 cases. N.Z.Vet.J., 51(2):58-62.
De Smet KA, Brown IN, Yates M y Ivanyi J. 1995. Ribosomal internal transcribed spacer sequences are identical among Mycobacterium avium-intracellulare complex isolates from AIDS patients, but vary among isolates from elderly pulmonary disease patients. Microbiology, 141(Pt 10):2739-47.
Dell'Isola B, Poyart C, Goulet O, Mougenot JF, Sadoun-Journo E, Brousse N, Schmitz J, Ricour C y Berche P. 1994. Detection of Mycobacterium paratuberculosis by polymerase chain reaction in children with Crohn's disease. J.Infect.Dis., 169(2):449-51.
Denholm LJ, Ottaway SJ y Marshall DJ. 1997. Control of ovine Johne's disease in New South Wales (Australia). En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., pp. 151-8. IAP, Rehoboth, MA, USA.
Deutz A, Spergser J, Wagner P, Rosengarten R y Kofer J. 2005. Mycobacterium avium subsp. paratuberculosis in wild animal species and cattle in Styria/Austria. Berl.Munch.Tierartzl.Wochenschr., 118(7-8):314-20.
Devallois A, Picardeau M, Paramasivan CN, Vincent V y Rastogi N. 1997. Molecular characterization of Mycobacterium avium complex isolates giving discordant results in AccuProbe tests by PCR-restriction enzyme analysis, 16S rRNA gene sequencing, and DT1-DT6 PCR. J.Clin.Microbiol., 35(11):2767-72.
Díaz EJ. 1989. Paratuberculosis en Andalucía. En: Juste editor. 2ª Reu.PTBC España (1987), pp. 17-24. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España.
Dierckins MS, Sherman DM y Gendron F. 1990. Probable paratuberculosis in a Sicilian ass. J.Am.Vet.Med.Assoc.:459-61. Dimareli-Malli Z y Sarris K. 2001. Comparison of DNA probe test and cultivation methods for detection of Mycobacterium avium
subsp. paratuberculosis in caprine and ovine faeces. Aust.Vet.J., 79(1):47-50. Djonne B. 2003. Paratuberculosis in goats: a special focus on the Nordic countries. Acta Vet.Scand., 2003(44):257-9.

BIBLIOGRAFÍA CITADA
204
Djonne B, Jensen MR, Grant IR y Holstad G. 2003. Detection by immunomagnetic PCR of Mycobacterium avium subsp. paratuberculosis in milk from dairy goats in Norway. Vet.Microbiol., 92(1-2):135-43.
Djonne B, Pavlik I, Svastova P, Bartos M y Holstad G. 2005. IS900 restriction fragment length polymorphism (RFLP) analysis of Mycobacterium avium subsp. paratuberculosis isolates from goats and cattle in Norway. Acta Vet.Scand., 46(1-2):13-8.
Dohoo IR, Martin W y Stryhn H. 2003, Veterinary epidemiologic research. AVC Inc.Charlottetown, Canada. Dolz G, Araya LN, Suarez J y Jimenez C. 1999. Prevalence of antibodies to bovine paratuberculosis detected by a LAM-ELISA in
Costa Rica. Vet.Rec., 144(12):322-3. Donaghy JA, Totton NL y Rowe MT. 2003a. Evaluation of culture media for the recovery of Mycobacterium avium subsp.
paratuberculosis from Cheddar cheese. Lett.Appl.Microbiol., 37(4):285-91. Donaghy JA, Totton NL y Rowe MT. 2003b. Iodixanol development of a laboratory-scale technique to monitor the persistence of
Mycobacterium avium subsp. paratuberculosis in Cheddar cheese. ScientificWorldJournal, 3:1241-8. Donaghy JA, Totton NL y Rowe MT. 2004. Persistence of Mycobacterium paratuberculosis during manufacture and ripening of
cheddar cheese. Appl.Environ.Microbiol., 70(8):4899-905. Dow CT. 2006a. A proposed link between MAP and Type I Diabetes - Mapping the TRIGR. En: Manning y Nielsen editores.
Proc.8th Int.Coll.Paratub., pp. 284-8. IAP, Madison, WI, USA. Dow CT. 2006b. PCR evidence of MAP in Blau syndrome patient tissues. En: Manning y Nielsen editores. Proc.8th
Int.Coll.Paratub., pp. 278-83. IAP, Madison, WI, USA. Doyle TM. 1954. Isolation of Johne's bacilli from the udders of clinically affected cows. Br.Vet.J., 110:215-8. Doyle TM. 1956. Johne's disease. Vet.Rec., 68:869-86. Doyle TM. 1958. Foetal infection in Johne's Disease. Vet.Rec., 70:328. Dukes TW, Glover GJ, Brooks BW y Duncan JR. 1992. Paratuberculosis in Saiga Antelope (Saiga tatarica) and experimental
transmission to domestic sheep. J.Wildl.Dis., 28(2):161-70. Dumonceau JM, Van Gossum A, Adler M, Fonteyne PA, Van Vooren JP, Deviere J y Portaels F. 1996. No Mycobacterium
paratuberculosis found in Crohn's disease using the polymerase chain reaction. Dig.Dis.Sci., 41:421-6. Dundee L, Grant IR, Ball HJ y Rowe MT. 2001. Comparative evaluation of four decontamination protocols for the isolation of
Mycobacterium avium subsp. paratuberculosis from milk. Lett.Appl.Microbiol., 33(3):173-7. Dunkin GW. 1934. Johne's disease. Its economic significance and early deteccion. Vet.Rec., 14(52):1551-9. Dunkin GW y Balfour-Jones SEB. 1935. Preliminary investigation of a disease of sheep possessing certain characteristics
simulating Johne's disease. J.Comp.Pathol.Ther., 48:236-9. Dupont C, Thompson K, Heuer C, Gicquel B y Murray A. 2005. Identification and characterization of an immunogenic 22 kDa
exported protein of Mycobacterium avium subspecies paratuberculosis. J.Med.Microbiol., 54(Pt 11):1083-92. Dvorska L, Bartos M, Martin G, Erler W y Pavlik I. 2001. Strategies for differentiation, identification and typing of medically
important species of mycobacteria by molecular methods. Vet.Med.(Praha), 46(11-12):309-28. Dvorska L, Bull TJ, Bartos M, Matlova L, Svastova P, Weston RT, Kintr J, Parmova I, van Soolingen D y Pavlik I. 2003. A
standardised restriction fragment length polymorphism (RFLP) method for typing Mycobacterium avium isolates links IS901 with virulence for birds. J.Microbiol.Methods, 55(1):11-27.
Eamens G, Spence S y Turner M. 2001. Survival of Mycobacterium avium subsp. paratuberculosis in amitraz cattle dip fluid. Aust.Vet.J., 79(10):703-6.
Eamens GJ, Whittington RJ, Marsh IB, Turner MJ, Saunders V, Kemsley PD y Rayward D. 2000. Comparative sensitivity of various faecal culture methods and ELISA in dairy cattle herds with endemic Johne's disease. Vet.Microbiol., 77(3-4):357-67.
Eda S, Bannantine JP, Waters WR, Mori Y, Whitlock RH, Scott MC y Speer CA. 2006. A highly sensitive and subspecies-specific surface antigen enzyme-linked immunosorbent assay for diagnosis of Johne's disease. Clin.Vaccine Immunol., 13(8):837-44.
Eda S, Elliott B, Scott MC, Waters WR, Bannantine JP, Whitlock RH y Speer CA. 2005. New method of serological testing for Mycobacterium avium subsp. paratuberculosis (Johne's disease) by flow cytometry. Foodborne Pathog.Dis., 2(3):250-62.
Eiglmeier K, Simon S, Garnier T y Cole ST. 2001. The integrated genome map of Mycobacterium leprae. Lepr.Rev., 72(4):462-9. El Zaatari FA, Naser SA, Markesich DC, Kalter DC, Engstand L y Graham DY. 1996. Identification of Mycobacterium avium
complex in sarcoidosis. J.Clin.Microbiol., 34(9):2240-5. El-Zaatari F, Naser SA, Hulten K, Burch P y Graham DY. 1999. Characterization of Mycobacterium paratuberculosis p36 antigen
and its seroreactivities in Crohn's disease. Curr.Microbiol., 39(2):115-9. El-Zaatari FAK, Naser SA, Engstrand L, Burch PE, Hachem CY, Whipple DL y Graham DY. 1995. Nucleotide sequence analysis
and seroreactivities of the 65 K heat shock protein from Mycobacterium paratuberculosis. Clin.Diagn.Lab.Immunol., 2:657-64.
Elguezabal N, Pavón A, Garrido JM, Geijo MV, Sevilla I, Tejada A, Izeta A, Casado R, Ochotorena I y Juste RA. 2006. High frequency of Mycobacterium avium subsp. paratuberculosis DNA in inflammatory bowel disease patient blood and controls detected by nested PCR. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 289-96. IAP, Madison, WI, USA.
Ellingson JEL, Stabel JR y Whitlock RH. 1999. Detection of Mycobacterium avium subspecies paratuberculosis in free-ranging bison by PCR. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 420-3. IAP, Madison, WI, USA.
Ellingson JL, Anderson JL, Koziczkowski JJ, Radcliff RP, Sloan SJ, Allen SE y Sullivan NM. 2005a. Detection of viable Mycobacterium avium subsp. paratuberculosis in retail pasteurized whole milk by two culture methods and PCR. J.Food Prot., 68(5):966-72.
Ellingson JL, Bolin CA y Stabel JR. 1998. Identification of a gene unique to Mycobacterium avium subespecies paratuberculosis and application to diagnosis of paratuberculosis. Mol.Cell.Probes, 12(3):133-42.
Ellingson JL, Cheville JC, Brees D, Miller JM y Cheville NF. 2003. Absence of Mycobacterium avium subspecies paratuberculosis components from Crohn's disease intestinal biopsy tissues. Clin.Med.Res., 1(3):217-26.
Ellingson JL, Koziczkowski JJ y Anderson JL. 2004. Comparison of PCR prescreening to two cultivation procedures with PCR confirmation for detection of Mycobacterium avium subsp. paratuberculosis in U.S. Department of Agriculture fecal check test samples. J.Food Prot., 67(10):2310-4.
Ellingson JL, Stabel JR, Bishai WR, Frothingham R y Miller JM. 2000. Evaluation of the accuracy and reproducibility of a practical PCR panel assay for rapid detection and differentiation of Mycobactrium avium subspecies. Mol.Cell.Probes, 14(3):153-61.
Ellingson JL, Stabel JR, Radcliff RP, Whitlock RH y Miller JM. 2005b. Detection of Mycobacterium avium subspecies paratuberculosis in free-ranging bison (Bison bison) by PCR. Mol.Cell.Probes, 19(3):219-25.
Elsaghier A, Prantera C, Moreno C y Ivanyi J. 1992. Antibodies to Mycobacterium paratuberculosis-specific protein antigens in Crohn's disease. Clin.Exp.Immunol., 90(3):503-8.
Emery DL y Whittington RJ. 2004. An evaluation of mycophage therapy, chemotherapy and vaccination for control of Mycobacterium avium subsp. paratuberculosis infection. Vet.Microbiol., 104(3-4):143-55.

BIBLIOGRAFÍA CITADA
205
Englund S, Ballagi-Pordány A, Bölske G y Johansson KE. 1999. Single PCR and nested PCR with a mimic molecule for detection of Mycobacterium avium subsp. paratuberculosis. Diagn.Microbiol.Infect.Dis., 33:163-71.
Englund S, Bolske G y Johansson KE. 2002. An IS900-like sequence found in a Mycobacterium sp. other than Mycobacterium avium subsp. paratuberculosis. FEMS Microbiol.Lett., 209(2):267-71.
Eppleston J. 2006. The potential impact of vaccination site lesions on the value of sheep previously vaccinated against ovine paratuberculosis in Australia. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 195-9. IAP, Madison, WI, USA.
Eppleston J y Whittington RJ. 2001. Isolation of Mycobacterium avium subsp. paratuberculosis from the semen of rams with clinical Johne's disease. Aust.Vet.J., 79(11):776-7.
Erardi FX, Failla ML y Falkinham III JO. 1987. Plasmid-encoded copper resistance and precipitation by Mycobacterium scrofulaceum. Appl.Environ.Microbiol., 53(8):1951-4.
Estonba A, Iriondo M, Oyanguren O, Garrido JM, Geijo MV, Sevilla I y Juste RA. 2006. Bovine 3'-UTR (GT11) variant of the NRAMP1 gene under-represented among paratuberculosis seropositive cattle. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 150. IAP, Madison, WI, USA.
European Commission. 2000. Possible links between Crohn's disease and Paratuberculosis: report of the Scientific Committee on Animal Health and Animal Welfare. http://ec.europa.eu/food/fs/sc/scah/out38_en.pdf,
Falkinham III JO. 1996. Epidemiology of infection by nontuberculous mycobacteria. Clin.Microbiol.Rev., 9(2):177-215. Fang Y, Wu WH, Pepper JL, Larsen JL, Marras SA, Nelson EA, Epperson WB y Christopher-Hennings J. 2002. Comparison of
real-time, quantitative PCR with molecular beacons to nested PCR and culture methods for detection of Mycobacterium avium subsp. paratuberculosis in bovine fecal samples. J.Clin.Microbiol., 40(1):287-91.
Fang Z, Doig C, Morrison N, Watt B y Forbes KJ. 1999. Characterization of IS1547, a new member of the IS900 family in the Mycobacterium tuberculosis complex, and its association with IS6110. J.Bacteriol., 181(3):1021-4.
Farreras P. 1915. Sobre la enfermedad de Johne. Revista Veterinaria de España, IX:277-83. Fawcett AR, Goddard PJ, McKelvey WAC, Buxton D, Reid HW, Greig A y Macdonald AJ. 1995. Johne's disease in a herd of
farmed red deer. Vet.Rec., 136:165-9. Feizabadi MM, Robertson ID, Cousins DV, Dawson DJ y Hampson DJ. 1997a. Use of multilocus enzyme electrophoresis to
examine genetic relationships amongst isolates of Mycobacterium intracellulare and related species. Microbiology, 143(Pt 4):1461-9.
Feizabadi MM, Robertson ID, Cousins DV y Hampson DJ. 1996. Genomic analysis of Mycobacterium bovis and other members of the Mycobacterium tuberculosis complex by isoenzyme analysis and pulsed-field gel electrophoresis. J.Clin.Microbiol., 34(5):1136-42.
Feizabadi MM, Robertson ID, Hope A, Cousins DV y Hampson DJ. 1997b. Differentiation of Australian isolates of Mycobacterium paratuberculosis using pulsed-field gel electrophoresis. Aust.Vet.J., 75(12):887-9.
Fernández de Luco D, Calvete C, Gortázar C, Bolea R y García Marín JF. 1995. Paratuberculosis en el conejo silvestre (Oryctolagus cuniculus). VII Reu.Soc.Esp.Anat.Pat.Vet., p. 19. SEAP, León, España.
Fernández V, Sánchez-García R y Velasco J. 2001. Estimación serológica e intradermoreacción doble (Tuberculosis-Paratuberculosis) en 30 rebaños caprinos de la comarca de Béjar (Salamanca). XXVI Jornadas Científicas y V Internacionales de la SEOC, pp. 714-21. Junta de Andalucía, Sevilla, España.
Ferreira R, Fonseca L y Lilenbaum W. 2002. Comparison between a commercial and an in-house ELISA or anti-M. avium paratuberculosis antibodies detection in dairy herds in Rio de Janeiro, Brazil. Rev.Latinoam.Microbiol., 44(3-4):129-32.
Ferreira R, Fonseca LS y Lilenbaum W. 2003. Detection of anti-Mycobacterium avium subsp paratuberculosis antibodies in Brazilian herds. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 463-4. IAP, Madison, WI, USA.
Ferroglio E, Nebbia P, Robino P, Rossi L y Rosati S. 2000. Mycobacterium paratuberculosis infection in two free-ranging Alpine ibex. Rev.- Off.Int.Epizoot., 19(3):859-62.
Fidler HM, Thurrell W, McJohnson M, Rook GAW y McFadden JJ. 1994. Specific detection of Mycobacterium paratuberculosis DNA associated with granulomatous tissue in Crohn's disease. Gut, 35(4):506-10.
Fischer O, Matlova L, Dvorska L, Svastova P, Bartl J, Melicharek I, Weston RT y Pavlik I. 2001. Diptera as vectors of mycobacterial infections in cattle and pigs. Med.Vet.Entomol., 15(2):208-11.
Fischer OA, Matlova L, Bartl J, Dvorska L, Svastova P, du MR, Melicharek I, Bartos M y Pavlik I. 2003a. Earthworms (Oligochaeta, Lumbricidae) and mycobacteria. Vet.Microbiol., 91(4):325-38.
Fischer OA, Matlova L, Dvorska L, Svastova P, Bartl J, Weston RT y Pavlik I. 2004a. Blowflies Calliphora vicina and Lucilia sericata as passive vectors of Mycobacterium avium subsp. avium, M. a. paratuberculosis and M. a. hominissuis. Med.Vet.Entomol., 18(2):116-22.
Fischer OA, Matlova L, Dvorska L, Svastova P y Pavlik I. 2003b. Nymphs of the Oriental cockroach (Blatta orientalis) as passive vectors of causal agents of avian tuberculosis and paratuberculosis. Med.Vet.Entomol., 17(2):145-50.
Fischer OA, Matlova L, Dvorska L, Svastova P, Peral DL, Weston RT, Bartos M y Pavlik I. 2004b. Beetles as possible vectors of infections caused by Mycobacterium avium species. Vet.Microbiol., 102(3-4):247-55.
Fleischmann RD, Alland D, Eisen JA, Carpenter L, White O, Peterson J, DeBoy R, Dodson R, Gwinn M, Haft D, Hickey E, Kolonay JF, Nelson WC, Umayam LA, Ermolaeva M, Salzberg SL, Delcher A, Utterback T, Weidman J, Khouri H, Gill J, Mikula A, Bishai W, Jacobs JW, Jr., Venter JC y Fraser CM. 2002. Whole-genome comparison of Mycobacterium tuberculosis clinical and laboratory strains. J.Bacteriol., 184(19):5479-90.
Flesch IE y Kaufmann SH. 1999. Effect of fetal calf serum on cytokine release by bone marrow-derived macrophages during infection with intracellular bacteria. Immunobiology, 200(1):120-7.
Francis J. 1943. Infection of laboratory animals with Mycobacterium johnei. J.Comp.Pathol.Ther., 53:140-50. Francis J, Madinaveitia J, Macturk HM y Snow GA. 1949. Isolation from acid-fast bacteria of a growth-factor for Mycobacterium
johnei and a precursor of phthiocol. Nature, 163:365. Frank TS y Cook SM. 1996. Analysis of paraffin sections of Crohn's disease for Mycobacterium paratuberculosis using polymerase
chain reaction. Mod.Pathol., 9(1):32-5. Fraqueli C, Carpi G, Bregoli M, Ostanello F, Pasolli C, Pozzato N y Rosati S. 2006. Epidemiology of paratuberculosis in two red
deer (Cervus elaphus) populations of Trentino (Northern Italy). En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 605-12. IAP, Madison, WI, USA.
Freudenberger RS y Simafranca SM. 2006. Cutaneous infection with rapidly-growing mycobacterial infection following heart transplant: a case report and review of the literature. Transplant.Proc., 38(5):1526-9.
Fridriksdottir V, Gunnarsson E, Sigurdarson S y Gudmundsdottir KB. 2000. Paratuberculosis in Iceland: epidemiology and control measures, past and present. Vet.Microbiol., 77(3-4):263-7.
Fridriksdottir V, Gunnarsson E, Sigurdarson S y Gudmundsdottir KB. 2003a. Paratuberculosis in Iceland: ongoing research. Acta Vet.Scand., 2003(44):273.

BIBLIOGRAFÍA CITADA
206
Fridriksdottir V, Hjartardottir S, Poulsen S, Sigurdarson S y Gunnarsson E. 2003b. Paratuberculosis in sheep in Iceland: Is eradication by vaccination possible? En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., p. 517. IAP, Madison, WI, USA.
Frothingham R y Meeker-O'Connell WA. 1998. Genetic diversity in the Mycobacterium tuberculosis complex based on variable numbers of tandem DNA repeats. Microbiology, 144(Pt 5):1189-96.
Frothingham R y Wilson KH. 1993. Sequence-based differentiation of strains in the Mycobacterium avium complex. J.Bacteriol., 175(10):2818-25.
Frothingham R y Wilson KH. 1994. Molecular phylogeny of the Mycobacterium avium complex demonstrates clinically meaningful divisions. J.Infect.Dis., 169(2):305-12.
Fuentes O y Cebrian J. 1988. El conejo de monte ¿reservorio de la paratuberculosis ovina y caprina? Symposium de Patologia Ovina y Caprina, vol.I, p. 32. AIDA, Zaragoza.
Fujimura Y y Owen RL. 1996. M cells as portals of infection: clinical and pathophysiological aspects. Infect.Agents Dis., 5(3):144-56.
Fujita H, Eishi Y, Ishige I, Saitoh K, Takizawa T, Arima T y Koike M. 2002. Quantitative analysis of bacterial DNA from Mycobacteria spp., Bacteroides vulgatus, and Escherichia coli in tissue samples from patients with inflammatory bowel diseases. J.Gastroenterol., 37(7):509-16.
Gao A, Mutharia L, Chen S, Rahn K y Odumeru J. 2002. Effect of pasteurization on survival of Mycobacterium paratuberculosis in milk. J.Dairy Sci., 85(12):3198-205.
Gao A, Odumeru J, Raymond M y Mutharia L. 2005. Development of improved method for isolation of Mycobacterium avium subsp. paratuberculosis from bulk tank milk: effect of age of milk, centrifugation, and decontamination. Can.J.Vet.Res., 69(2):81-7.
García Marín JF, Benazzi S, Pérez V y Badiola JJ. 1992a. Study of the entrance of M. paratuberculosis in the lamb intestinal mucosa using immunohistochemical methods for antigen detection. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 371-7. IAP, Providence, RI, USA.
García Marín JF, Chávez G, Aduriz JJ, Pérez V, Juste RA y Badiola JJ. 1992b. Prevalence of paratuberculosis in infected goats flocks and comparison of different methods of diagnosis. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 157-63. IAP, Providence, RI, USA.
García Marín JF, Chávez G, Pérez V y Badiola JJ. 1995a. Pathological findings in lambs experimentally infected with Mycobacterium avium subsp silvaticum and Mycobacterium avium subsp paratuberculosis. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., p. 208. IAP, Rehoboth, MA, USA.
García Marín JF, Juste RA, Peris B, Badiola JJ y Sáez de Ocáriz C. 1988a. Study of a trial of vaccinated and non vaccinated experimental infected lambs with M.paratuberculosis. CEC Workshop, CEC. Crete, Greece.
García Marín JF, Juste RA, Peris B, Marco JC, Saez de Ocariz C y Badiola JJ. 1988b. Study of the initial phases of the experimental infection with Mycobacterium paratuberculosis in vaccinated and non-vaccinated sheep. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 354-60. LCRV, Maisons-Alfort, France.
García Marín JF, Pérez V y Badiola JJ. 1992c. Prevalence and type of paratuberculosis lesions in sheep and their relation with the diagnosis by AGID test. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 172-80. IAP, Providence, RI, USA.
García Marín JF, Pérez V, García de Jalón JA, De las Heras M, Barberá M, Fernández de Luco D y Badiola JJ. 1994. Diagnóstico de casos clínicos de paratuberculosis ovina y caprina. Med.Vet., 11(9):491-502.
García Marín JF, Pérez V y González J. 2000. La interacción bovino-bacteria y los mecanismos de la enfermedad. En: Juste editor. Paratuberculosis. Bovis, Aula Veterinaria, vol.nº98, pp. 29-38. Luzán 5, Madrid, España.
García Marín JF, Tellechea J, Corpa JM, Gutiérrez M y Pérez V. 1999a. Relation between ovine paratuberculois lesions and cellular and humoral immune responses in diagnostic tests. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 593-8. IAP, Madison, WI, USA.
García Marín JF, Tellechea J, Gutiérrez M, Corpa JM y Pérez V. 1999b. Evaluation of two vaccines (killed and attenuated) against small ruminant paratuberculosis. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., IAP, Madison, WI, USA.
García Marín JF, Tellechea J, Gutiérrez M y Pérez V. 1995b. Histopathological evaluation of a killed vaccine against paratuberculosis in sheep. Proc.13th Eur.Congr.Vet.Pathol., Edinburgh, UK.
García Marín JF, Tellechea J, Gutiérrez M, Pérez V y Juste RA. 1997. Paratuberculosis vaccination in sheep modifies and limits the development of lesions. En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., p. 223. IAP, Rehoboth, MA, USA.
García-Graells C, Masschalck B y Michiels CW. 1999. Inactivation of Escherichia coli in milk by high-hydrostatic-pressure treatment in combination with antimicrobial peptides. J.Food Prot., 62(11):1248-54.
García-Pariente C, González J, Ferreras MC, Fuertes M, Benavides J, Reyes LE, Moreno O, García Marín JF y Pérez V. 2003. Paratuberculosis vaccination of adult animals in two flocks of dairy sheep. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 507-10. IAP, Madison, WI, USA.
García-Pariente C, Pérez V, Geijo MV, Moreno O, Muñoz M, Fuertes M, Puentes E, Doce J, Ferreras MC y García Marín JF. 2006. The efficacy of a killed vaccine against paratuberculosis (SILIRUM®) in cattle. A field study. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 260. IAP, Madison, WI, USA.
Gardner IA, Tavornpanich S, Carpenter TE, Johnson WO y Anderson RJ. 2006. Targeted sampling of subpopulations can improve detection of Mycobacterium avium subsp. paratuberculosis (Map) in dairy herds. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 260. IAP, Madison, WI, USA.
Garnier T, Eiglmeier K, Camus JC, Medina N, Mansoor H, Pryor M, Duthoy S, Grondin S, Lacroix C, Monsempe C, Simon S, Harris B, Atkin R, Doggett J, Mayes R, Keating L, Wheeler PR, Parkhill J, Barrell BG, Cole ST, Gordon SV y Hewinson RG. 2003. The complete genome sequence of Mycobacterium bovis. Proc.Natl.Acad.Sci.U.S.A., 100(13):7877-82.
Garrido F, Hooghuis H y León L. 1989. Apuntes sobre paratuberculosis. En: Juste editor. 2ª Reu.PTBC España (1987), pp. 47-54. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España.
Garrido F y León Vizcaino L. 1979. Diagnóstico de paratuberculosis caprina en Andalucía. VII Congr.Nac.Microbiol., SEM. Cádiz, España.
Garrido JM. 2001. Puesta a punto de las técnicas de PCR en heces y de ELISA para el diagnóstico de la paratuberculosis. Estudio de prevalencia en ganado bovino. Tesis Doctoral. Universidad de Zaragoza, Aragón, España.
Garrido JM, Aduriz G, Garcia-Marin JF, Perez V, Corpa JM y Juste RA. 1999a. A new assessment of the absorbed ELISA for diagnosis of ovine paratuberculosis. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 395-9. IAP, Madison, WI, USA.
Garrido JM, Aduriz G, Geijo MV, Sevilla I y Juste RA. 2003. Comparison of different indirect ELISA methods on reference cattle. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., IAP, Madison, WI, USA.

BIBLIOGRAFÍA CITADA
207
Garrido JM, Aduriz G, Juste RA y Geijo MV. 2000a. Los métodos de diagnóstico de la paratuberculosis en el ganado vacuno. En: Juste editor. Paratuberculosis. Bovis, Aula Veterinaria, vol.nº93, pp. 49-61. Luzan 5, Madrid, España.
Garrido JM, Cortabarria N, Aduriz G y Juste RA. 1999b. Use of a PCR method on Faecal Samples for Diagnosis of Sheep Paratuberculosis. En: Manning y Collins editores. Proc Int Coll PTBC, (ed. VI), vol.VI, pp. 407-12. IAP, Madison, WI, USA.
Garrido JM, Cortabarria N, Oguiza JA, Aduriz G y Juste RA. 2000b. Use of a PCR method on fecal samples for diagnosis of sheep paratuberculosis. Vet.Microbiol., 77:379-86.
Garrido JM, Plazaola JM, Sevilla I, Geijo MV, Elguezabal N, Aduriz G, Doce J, Puentes E y Juste RA. 2006. Control of paratuberculosis in dairy farms in the Basque Country. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 261. IAP, Madison, WI, USA.
Gasteiner J, Wenzl H, Fuchs K, Jark U y Baumgartner W. 1999. Serological cross-sectional study of paratuberculosis in cattle in Austria. Zentralbl.Veterinarmed.B, 46(7):457-66.
Gerlach GF. 2002. Paratuberculosis: the pathogen and routes of infection. Dtsch.Tierarztl.Wochenschr., 109(12):504-6. Gervilla R, Sendra E, Ferragut V y Guamis B. 1999. Sensitivity of Staphylococcus aureus and Lactobacillus helveticus in ovine
milk subjected to high hydrostatic pressure. J.Dairy Sci., 82(6):1099-107. Gezon HM, Bither HD, Gibbs HC, Acker EJ, Hanson LA, Thompson JK y Jorgenson RD. 1988. Identification and control of
paratuberculosis in a large goat herd. Am.J.Vet.Res., 49(11):1817-23. Ghadiali AH, Strother M, Naser SA, Manning EJ y Sreevatsan S. 2004. Mycobacterium avium subsp. paratuberculosis strains
isolated from Crohn's disease patients and animal species exhibit similar polymorphic locus patterns. J.Clin.Microbiol., 42(11):5345-8.
Giacometti M, Tolari F, Mannelli A y Lanfranchi P. 1995. Seroepidemiologic investigations in the Alpine ibex (Capra i. ibex) of Piz Albris in the canton of Grigioni (Switzerland). Schweiz.Arch.Tierheilkd., 137(12):537-42.
Giese SB y Ahrens P. 2000. Detection of Mycobacterium avium subsp. paratuberculosis in milk from clinically affected cows by PCR and culture. Vet.Microbiol., 77(3-4):291-7.
Gilmour NJL y Angus KW. 1971. Effect of the riminophenazine B663 (G 30320) on Mycobacterium johnei infection and reinfection in sheep. I. Bacteriology and hypersensitivity. J.Comp.Pathol., 81:221-26.
Gilmour NJL y Angus KW. 1973. Effect of revaccination on Mycobacterium johnei infection in sheep. J.Comp.Pathol., 83:437-45. Gilmour NJL y Angus KW. 1991. Johne's disease. En: Martin, W. B. y Aitken, I. D. editores. Diseases of sheep. ed. 2. pp. 95-9.
Blackwell Scientific Publications. Oxford, UK. Gilmour NJL y Brotherston JG. 1966. Further studies on immunity to Mycobacterium johnei in sheep. J.Comp.Pathol., 76:341-9. Gilot P, De Kesel M, Coene M y Cocito C. 1992. Induction of cellular immune reactions by A36, an antigen complex of
Mycobacterium paratuberculosis: comparison of A36 and johnin components. Scand.J.Immunol., 36(6):811-21. Gitnick G, Collins J, Beaman B, Brooks D, Arthur M, Imaeda T y Palieschesky M. 1989. Preliminary report on isolation of
mycobacteria from patients with Crohn's disease. Dig.Dis.Sci., 34(6):925-32. Glawischnig W, Awad-Masalmeh M, Khaschabi D y Schonbauer M. 2004. Detection of Mycobacterium avium subsp.
paratuberculosis from the testicles of a clinically infected breeding animal. Berl.Munch.Tierartzl.Wochenschr., 117(3-4):136-9.
Gobin J y Horwitz MA. 1996. Exochelins of Mycobacterium tuberculosis remove iron from human iron-binding proteins and donate iron to mycobactins in the M. tuberculosis cell wall. J.Exp.Med., 183(4):1527-32.
Godfroid J, Boelaert F, Heier A, Clavareau C, Wellemans V, Desmecht M, Roels S y Walravens K. 2000. First evidence of Johne's disease in farmed red deer (Cervus elaphus) in Belgium. Vet.Microbiol., 77(3-4):283-90.
Gomes MJP, Asanome W y Ribeiro VR. 2006. Prevalence of infection caused by Mycobacterium avium subsp. paratuberculosis (Map) in dairy herds in Rio Grande do Sul-South Brazil. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., IAP, Madison, WI, USA.
Gonda MG, Chang YM, Shook GE, Collins MT y Kirkpatrick BW. 2006. Genetic variation of Mycobacterium avium ssp. paratuberculosis infection in US Holsteins. J.Dairy.Sci., 89(5):1804-12.
González J. 1998. Empleo de un método histológico para el estudio de la prevalencia de la paratuberculosis ovina. Tesina de Licenciatura. Universidad de León, Castilla y León, España.
González J. 2003. Caracterización lesional y evaluación de técnicas diagnósticas de la paratuberculosis bovina. Tesis Doctoral. Universidad de León, Castilla y León, España.
González J, Geijo MV, García-Pariente C, Verna A, Corpa JM, Reyes LE, Ferreras MC, Juste RA, Garcia-Marin JF y Pérez V. 2005. Histopathological classification of lesions associated with natural paratuberculosis infection in cattle. J.Comp.Pathol., 133(2-3):184-96.
Goodfellow M y Magee JG. 1998. Taxonomy of mycobacteria. En: Gangadharam, P. R. y Jenkin, P. A. editores. Mycobacteria I: basic aspects. pp. 1-71. International Thomson Publishing. New York.
Goodger WJ, Collins MT, Nordlund KV, Eisele C, Pelletier J y Thomas CB. 1996. Epidemiologic study of on-farm management practices associated with prevalence of Mycobacterium paratuberculosis infections in dairy cattle. J.Am.Vet.Med.Assoc., 208(11):1877-80.
Graham DY, Markesich DC y Yoshimura HH. 1987. Mycobacteria and inflammatory bowel disease. Results of culture. Gastroenterology, 92(2):436-42.
Grant IR. 2005. Zoonotic potential of Mycobacterium avium ssp. paratuberculosis: the current position. J.Appl.Microbiol., 98(6):1282-93.
Grant IR, Ball HJ, Neill SD y Rowe MT. 1996. Inactivation of Mycobacterium paratuberculosis in cows' milk at pasteurization temperatures. Appl.Environ.Microbiol., 62(2):631-6.
Grant IR, Ball HJ y Rowe MT. 1998a. Effect of high-temperature, short-time (HTST) pasteurization on milk containing low numbers of Mycobacterium paratuberculosis. Lett.Appl.Microbiol., 26:166-70.
Grant IR, Ball HJ y Rowe MT. 1998b. Isolation of Mycobacterium paratuberculosis from milk by immunomagnetic separation. Appl.Environ.Microbiol., 64(9):3153-8.
Grant IR, Ball HJ y Rowe MT. 1999. Effect of higher pasteurization temperatures, and longer holding times at 72 degrees C, on the inactivation of Mycobacterium paratuberculosis in milk. Lett.Appl.Microbiol., 28:461-5.
Grant IR, Ball HJ y Rowe MT. 2002a. Incidence of Mycobacterium paratuberculosis in bulk raw and commercially pasteurized cows' milk from approved dairy processing establishments in the United Kingdom. Appl.Environ.Microbiol., 68(5):2428-35.
Grant IR, Hitchings EI, McCartney A, Ferguson F y Rowe MT. 2002b. Effect of commercial-scale high-temperature, short-time pasteurization on the viability of Mycobacterium paratuberculosis in naturally infected cows' milk. Appl.Environ.Microbiol., 68(2):602-7.
Grant IR, Kirk RB, Hitchings EI y Rowe MT. 2003. Comparative evaluation of the MGIT and BACTEC culture systems for the recovery of Mycobacterium avium subsp. paratuberculosis from milk. J.Appl.Bacteriol., 95(1):196-201.

BIBLIOGRAFÍA CITADA
208
Grant IR, O'Riordan LM, Ball HJ y Rowe MT. 2001a. Incidence of Mycobacterium paratuberculosis in raw sheep and goats' milk in England, Wales and Northern Ireland. Vet.Microbiol., 79(2):123-31.
Grant IR, Pope CM, O'Riordan LM, Ball HJ y Rowe MT. 2000. Improved detection of Mycobacterium avium subsp. paratuberculosis in milk by immunomagnetic PCR. Vet.Microbiol., 77(3-4):369-78.
Grant IR y Rowe MT. 2004. Effect of chemical decontamination and refrigerated storage on the isolation of Mycobacterium avium subsp. paratuberculosis from heat-treated milk. Lett.Appl.Microbiol., 38(4):283-8.
Grant IR, Rowe MT, Dundee L y Hitchings E. 2001b. Mycobacterium avium ssp. paratuberculosis: its incidence, heat resistance and detection in milk and dairy products. Int.J.Dairy Technol., 54:2-14.
Grant IR, Williams AG, Rowe MT y Muir DD. 2005a. Efficacy of various pasteurization time-temperature conditions in combination with homogenization on inactivation of Mycobacterium avium subsp. paratuberculosis in milk. Appl.Environ.Microbiol., 71(6):2853-61.
Grant IR, Williams AG, Rowe MT y Muir DD. 2005b. Investigation of the impact of simulated commercial centrifugation and microfiltration conditions on levels of Mycobacterium avium ssp. paratuberculosis in milk. Int.J.Dairy Technol., 58(3):138-49.
Green EP, Tizard MLV, Moss MT, Thompson JK, Winterbourne DJ, McFadden JJ y Hermon-Taylor J. 1989. Sequence and characteristics of IS900, an insertion element identified in a human Crohn's disease isolate of Mycobacterium paratuberculosis. Nucleic Acids Res., 17(22):9063-73.
Green SL, Lifland BD, Bouley DM, Brown BA, Wallace RJ, Jr. y Ferrell JE, Jr. 2000. Disease attributed to Mycobacterium chelonae in South African clawed frogs (Xenopus laevis). Comp.Med., 50(6):675-9.
Greenstein RJ. 2003. Is Crohn's disease caused by a mycobacterium? Comparisons with leprosy, tuberculosis, and Johne's disease. Lancet Infect.Dis., 3(8):507-14.
Greig A, Stevenson K, Henderson D, Perez V, Hughes V, Pavlik I, Hines ME, McKendrick I y Sharp JM. 1999. Epidemiological study of paratuberculosis in wild rabbits in Scotland. J.Clin.Microbiol., 37(6):1746-51.
Greig A, Stevenson K, Pérez V, Pirie AA, Grant JM y Sharp JM. 1997. Paratuberculosis in wild rabbits (Oryctolagus cuniculus). Vet.Rec., 140:141-3.
Griffin JF, Spittle E, Rodgers CR, Liggett S, Cooper M, Bakker D y Bannantine JP. 2005. Immunoglobulin G1 enzyme-linked immunosorbent assay for diagnosis of Johne's Disease in red deer (Cervus elaphus). Clin.Diagn.Lab.Immunol., 12(12):1401-9.
Griffith DE, Girard WM y Wallace RJ, Jr. 1993. Clinical features of pulmonary disease caused by rapidly growing mycobacteria. An analysis of 154 patients. Am.Rev.Respir.Dis., 147(5):1271-8.
Groenen PMA, Bunschoten A, van Soolingen D y Van Embden JDA. 1993. Nature of DNA polymorphism in the direct repeat cluster of Mycobacterium tuberculosis; application for strain differentiation by a novel typing method. Mol.Microbiol., 10(5):1057-65.
Groenendaal H y Galligan DT. 2003a. Economic consequences of control programs for paratuberculosis in midsize dairy farms in the United States. J.Am.Vet.Med.Assoc., 223(12):1757-63.
Groenendaal H, Nielen M y Hesselink JW. 2003b. Development of the Dutch Johne's disease control program supported by a simulation model. Prev.Vet.Med., 60(1):69-90.
Guerrero C, Bernasconi C, Burki D, Bodmer T y Telenti A. 1995. A novel insertion element from Mycobacterium avium, IS1245, is a specific target for analysis of strains relatedness. J.Clin.Microbiol., 33(2):304-7.
Guerrero C, Bernasconi C, Burki D, y Telenti A. 1994. IS1311, an insertion sequence in Mycobacterium avium. GeneBank Accession number U16276. http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nucleotide&val=564031.
Guijarro E. 1990. Paratuberculosis en la Comunidad Autónoma de Castilla y León. En: Juste editor. 3ª Reu.PTBC España (1989), pp. 42-3. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España.
Gumber S y Whittington RJ. 2006. Comparison of BACTEC 460 and MGIT 960 systems for the culture of Mycobacterium avium subsp. paratuberculosis S strain and observations on the effect of inclusion of ampicillin in culture media to reduce contamination. Vet.Microbiol.(In press).
Gunnarsson E y Fodstad FH. 1979. Cultural and biochemical characteristics of Mycobacterium paratuberculosis isolated from goats in Norway. Acta Vet.Scand., 20:122-34.
Gurtler V y Stanisich VA. 1996. New approaches to typing and identification of bacteria using the 16S-23S rDNA spacer region. Microbiology, 142(Pt 1):3-16.
Gwozdz JM, Reichel MP, Murray C, Manktelow W, West DM y Thompson KG. 1997. Detection of Mycobacterium avium subsp. paratuberculosis in ovine tissues and blood by the polymerase chain reaction. Vet.Microbiol., 51:233-44.
Gwozdz JM y Thompson KG. 2002. Antigen-induced production of interferon-gamma in samples of peripheral lymph nodes from sheep experimentally inoculated with Mycobacterium avium subsp. paratuberculosis. Vet.Microbiol., 84(3):243-52.
Gwozdz JM, Thompson KG, Manktelow BW, Murray A y West DM. 2000a. Vaccination against paratuberculosis of lambs already infected experimentally with Mycobacterium avium subespecies paratuberculosis. Aust.Vet.J., 78(8):560-6.
Gwozdz JM, Thompson KG, Murray A, Reichel MP, Manktelow BW y West DM. 2000b. Comparison of three serological tests and an interferon-gamma assay for the diagnosis of paratuberculosis in experimentally infected sheep. Aust.Vet.J., 78(11):779-83.
Gwozdz JM, Thompson KG, Murray A, West DM y Manktelow BW. 2000c. Use of the polymerase chain reacton assay detection of Mycobacterium avium subspecies paratuberculosis in blood and liver biopsies from experimentally infected sheep. Aust.Vet.J., 78:622-4.
Hacker U, Huttner K y Konow M. 2004. [Investigation of serological prevalence and risk factors of paratuberculosis in dairy farms in the state of Mecklenburg-Westpommerania, Germany]. Berl.Munch.Tierartzl.Wochenschr., 117(3-4):140-4.
Hagan WA. 1935. Vaccination against Johne's disease. Cornell Vet., 25:344-53. Hall G, Katanik M, Tuohy M y Shoultis M. 2004. Use of comercial amplification tests in the clinical microbiology laboratory: test
selection and Quality Assurance. En: Persing, D. H., Tenover, F. C., Versalovic, J., Tang, Y. W., Unger, E. R., Relman, D. A. y White, T. J. editores. Molecular microbiology: diagnostic principles and practice. pp. 27-36. ASM. Washington, DC, USA.
Halldorsdottir S, Englund S, Nilsen SF y Olsaker I. 2002. Detection of Mycobacterium avium subsp. paratuberculosis by buoyant density centrifugation, sequence capture PCR and dot blot hybridisation. Vet.Microbiol., 87(4):327-40.
Hamilton HL, Cooley AJ, Adams JL y Czuprynsky CJ. 1991. Mycobacterium paratuberculosis monoassociated nude mice as a paratuberculosis model. Vet.Pathol., 28:146-55.
Hammer P, Kiesner C, Walte HG, Knappstein K y Teufel P. 2002. Heat resistance of Mycobacterium avium ssp paratuberculosis in raw milk tested in a pilot plant pasteurizer. Kiel.Milchwirt.Forschungsber., 54(275):275-303.
Hammer P, Walte HG, Kiesner C y Teufel P. 2006. Heat resistance of Mycobacterium avium subsp. paratuberculosis in skim milk and cream tested in a pilot plant pasteurizer. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 297-303. IAP, Madison, WI, USA.

BIBLIOGRAFÍA CITADA
209
Hanna BA, Walters SB, Bonk SJ y Tick LJ. 1995. Recovery of mycobacteria from blood in mycobacteria growth indicator tube and Lowenstein-Jensen slant after lysis-centrifugation. J.Clin.Microbiol., 33(12):3315-6.
Hanzlikova M y Vilimek L. 1989. Occurrence of paratuberculosis in cattle in Slovakia in the period 1982-1987. Veterinarstvi, 39(9):462-3.
Hardin LE y Thorne JG. 1996. Comparison of milk with serum ELISA for the detection of paratuberculosis in dairy cows. J.Am.Vet.Med.Assoc., 209(1):120-2.
Harris NB y Barletta RG. 2001. Mycobacterium avium subsp. paratuberculosis in veterinary medicine. Clin.Microbiol.Rev., 14(3):489-512.
Hasting APM, Blackburn CDW y Crowther JS. 2001. Mycobacterium paratuberculosis and the commercial pasteurization of milk. Food Bioprod.Process., 79(C2):83-8.
Hendrick S, Duffield T, Leslie K, Lissemore K, Archambault M y Kelton D. 2005a. The prevalence of milk and serum antibodies to Mycobacterium avium subspecies paratuberculosis in dairy herds in Ontario. Can.Vet.J., 46(12):1126-9.
Hendrick SH, Duffield TE, Kelton DE, Leslie KE, Lissemore KD y Archambault M. 2005b. Evaluation of enzyme-linked immunosorbent assays performed on milk and serum samples for detection of paratuberculosis in lactating dairy cows. J.Am.Vet.Med.Assoc., 226(3):424-8.
Hendrick SH, Kelton DF, Leslie KE, Lissemore KD, Archambault M, Bagg R, Dick P y Duffield TF. 2006. Efficacy of monensin sodium for the reduction of fecal shedding of Mycobacterium avium subsp. paratuberculosis in infected dairy cattle. Prev.Vet.Med., 75(3-4):206-20.
Hendrick SH, Kelton DF, Leslie KE, Lissemore KD, Archambault M y Duffield TF. 2005c. Effect of paratuberculosis on culling, milk production, and milk quality in dairy herds. J.Am.Vet.Med.Assoc., 227(8):1302-8.
Herman L, De Jonghe V, Dumon I, Grijspeerdt K, Naydenski H y D'Haese E. 2006. Clumping of Mycobacterium avium subsp. paratuberculosis in milk and feces and growth activation after milk heating. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 304-10. IAP, Madison, WI, USA.
Hermon-Taylor J. 1998. The causation of Crohn's disease and treatment with antimicrobial drugs. Ital.J.Gastroenterol.Hepatol., 30(6):607-10.
Hermon-Taylor J. 2002. Treatment with drugs active against Mycobacterium avium subespecies paratuberculosis can heal Crohn's disease: more evidence for a neglected public health tragedy. Dig.Liver Dis., 34:9-12.
Hermon-Taylor J, Bull T, Sheridan, J.M., Cheng JSML y Sumar N. 2000. Causation of Crohn's disease by Mycobacterium avium subspecies paratuberculosis. Can.J.Gastroenterol., 14(6):521-39.
Hilbink F, West DM, de Lisle GW, Kittelberger R, Hoise BD, Hutton J, Cooke MM y Penrose M. 1994. Comparison of a complement fixation test, a gel diffusion test and two absorbed and unabsorbed ELISAs for the diagnosis of paratuberculosis in sheep. Vet.Microbiol., 41:107-16.
Hill BB, West M y Brock KV. 2003. An estimated prevalence of Johne's disease in a subpopulation of Alabama beef cattle. J.Vet.Diagn.Invest., 15(1):21-5.
Hillerma K. 1966. A disease resembling paratuberculosis (Johne's disease) in roe deer (Capreolus capreolus L.). An aetiological and patho-anatomical study. Acta Vet.Scand., 7(4):330-63.
Hines ME, Barker SA y Snider TG. 1995. Isolation of DNA from Mycobacterium paratuberculosis by matrix solid phase dispersion (MSPD). En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 250-6. IAP, Rehoboth, MA, USA.
Hines ME2, Stiver S, Giri D, Whittington L, Watson C, Johnson J, Pence M, Baldwin C, Sangster L y Aly S. 2006. Efficacy of spheroplastic and cell wall competent vaccines for Johne's disease in experimentally challenged baby goats. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 262. IAP, Madison, WI, USA.
Hines ME2 y Styer EL. 2003. Preliminary characterization of chemically generated Mycobacterium avium subsp. paratuberculosis cell wall deficient forms (spheroplasts). Vet.Microbiol., 95(4):247-58.
Hirst HL, Garry FB, Morley PS, Salman MD, Dinsmore RP, Wagner BA, McSweeney KD y Goodell GM. 2004. Seroprevalence of Mycobacterium avium subsp paratuberculosis infection among dairy cows in Colorado and herd-level risk factors for seropositivity. J.Am.Vet.Med.Assoc., 225(1):97-101.
Hole NH. 1953. The diagnosis of Johne's disease. Proc.15th Int.Vet.Congr., vol.Pt.1, pp. 173-7. Stockholm, Sweden. Holzmann CB, Jorge MC, Traversa MJ, Schettino DM, Medina L y Bernardelli A. 2004. [A study of the epidemiological behaviour
of bovine paratuberculosis using time series in Tandil in the province of Buenos Aires, Argentina]. Rev.- Off.Int.Epizoot., 23(3):791-9.
Homuth M, Valentin-Weigand P, Rohde M y Gerlach GF. 1998. Identification and characterization of a novel extracellular ferric reductase from Mycobacterium paratuberculosis. Infect.Immun., 66(2):710-6.
Hoover DG, Metrick C, Papineau AM, Farkas DF y Knorr D. 1989. Biological effects of high hydrostatic-pressure on food microorganisms. Food Technol., 43(3):99-107.
Hope AF, Kluver PF, Jones SL y Condron RJ. 2000. Sensitivity and specificity of two serological tests for the detection of ovine paratuberculosis. Aust.Vet.J., 78(12):850-6.
Hostetter J, Steadham E, Haynes J, Bailey T y Cheville N. 2003. Phagosomal maturation and intracellular survival of Mycobacterium avium subspecies paratuberculosis in J774 cells. Comp.Immunol.Microbiol.Infect.Dis., 26(4):269-83.
Hostetter JM, Steadham EM, Haynes JS, Bailey TB y Cheville NF. 2002. Cytokine effects on maturation of the phagosomes containing Mycobacteria avium subspecies paratuberculosis in J774 cells. FEMS Immunol.Med.Microbiol., 34(2):127-34.
Hruska K, Bartos M, Kralik P y Pavlik I. 2005. Mycobacterium avium subsp. paratuberculosis in powdered infant milk: paratuberculosis in cattle: the public health problem to be solved. Vet.Med.(Praha), 50(8):327-35.
Huchzermeyer HFAK y Bastianello SS. 1992. Serological, microscopic, cultural and pathological findings from 135 sheep originating from a flock in South Africa. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 140-6. IAP, Providence, RI, USA.
Huda A y Jensen HE. 2003a. Comparison of histopathology, cultivation of tissues and rectal contents, and interferon-gamma and serum antibody responses for the diagnosis of bovine paratuberculosis. J.Comp.Pathol., 129(4):259-67.
Huda A, Jungersen G, Christoffersen AB y Lind P. 2003b. Diagnosis of bovine paratuberculosis by interferon-gamma (IFN-gamma) test. Acta Vet.Scand., 2003(44):281.
Huda A, Jungersen G y Lind P. 2004. Longitudinal study of interferon-gamma, serum antibody and milk antibody responses in cattle infected with Mycobacterium avium subsp. paratuberculosis. Vet.Microbiol., 104(1-2):43-53.
Hughes VM, Smith S, García-Sánchez A y Stevenson K. 2006. Proteomic comparison of Mycobacterium avium subspecies paratuberculosis grown in vitro and isolated from clinical cases of ovine paratuberculosis. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 158. IAP, Madison, WI, USA.
Huitema H. 1967. Johne's disease in cattle and vaccination. Bull.Off.Int.Epizoot., 68(1):743-8.

BIBLIOGRAFÍA CITADA
210
Hulten K, Almashhrawi A, El Zaatari FA y Graham DY. 2000a. Antibacterial therapy for Crohn's disease: a review emphasizing therapy directed against mycobacteria. Dig.Dis.Sci., 45(3):445-56.
Hulten K, El-Zimaity HM, Karttunen TJ, Almashhrawi A, Schwartz MR, Graham DY y El-Zaatari FA. 2001. Detection of Mycobacterium avium subspecies paratuberculosis in Crohn's diseased tissues by in situ hybridization. Am.J.Gastroenterol., 96(5):1529-35.
Hulten K, Karttunen TJ, El Zimaity HM, Naser SA, Almashhrawi A, Graham DY y El Zaatari FA. 2000b. In situ hybridization method for studies of cell wall deficient M. paratuberculosis in tissue samples. Vet.Microbiol., 77(3-4):513-8.
Hulten K, Karttunen TJ, El Zimaity HM, Naser SA, Collins MT, Graham DY y El Zaatari FA. 2000c. Identification of cell wall deficient forms of M. avium subsp. paratuberculosis in paraffin embedded tissues from animals with Johne's disease by in situ hybridization. J.Microbiol.Methods, 42(2):185-95.
Huntley JF, Stabel JR, Paustian ML, Reinhardt TA y Bannantine JP. 2005. Expression library immunization confers protection against Mycobacterium avium subsp. paratuberculosis infection. Infect.Immun., 73(10):6877-84.
Hurley SS y Ewing E. 1983. Results of a field evaluation of a whole cell bacterin. En: Merkal editor. Proc.1st Int.Coll.Paratub., pp. 244-8. NADC, USDA, Ames, IA, USA.
Hutchinson LJ. 1996. Economic impact of paratuberculosis. Vet.Clin.North Am.Food Anim.Pract., 12(2):373-81. Ibbotson JP, Lowes JR, Chahal H, Gaston JS, Life P, Kumararatne DS, Sharif H, Alexander-Williams J y Allan RN. 1992. Mucosal
cell-mediated immunity to mycobacterial, enterobacterial and other microbial antigens in inflammatory bowel disease. Clin.Exp.Immunol., 87(2):224-30.
Iinuma Y, Ichiyama S, Hasegawa Y, Shimokata K, Kawahara S y Matsushima T. 1997. Large-restriction-fragment analysis of Mycobacterium kansasii genomic DNA and its application in molecular typing. J.Clin.Microbiol., 35(3):596-9.
Ikonomopoulos J, Pavlik I, Bartos M, Svastova P, Ayele WY, Roubal P, Lukas J, Cook N y Gazouli M. 2005. Detection of Mycobacterium avium subsp. paratuberculosis in retail cheeses from Greece and the Czech Republic. Appl.Environ.Microbiol., 71(12):8934-6.
Ikonomopoulos JA, Gorgoulis VG, Kastrinakis NG, Zacharatos PV, Kokotas SN, Evangelou K, Kotsinas AG, Tsakris AG, Manolis EN y Kittas CN. 2000. Sensitive differential detection of genetically related mycobacterial pathogens in archival material. Am.J.Clin.Pathol., 114(6):940-50.
Imaeda T. 1984. Cell wall deficient forms. En: Kubica, G. P. y Wayne, L. G. editores. The mycobacteria. ed. 15. pp. 671-80. Marcel Dekker, Inc. New York, USA.
Inderlied CB, Kemper CA y Bermudez LEM. 1993. The Mycobacterium avium complex. Clin.Microbiol.Rev., 6(3):266-310. Jakob HJ y Schiefer B. 1969. Presencia de paratuberculosis entre las ovejas. Panorama Veterinario. pp. 87-95. Jakobsen MB, Alban L y Nielsen SS. 2000. A cross sectional study of paratuberculosis in 1155 Danish dairy cows. Prev.Vet.Med.,
46:15-27. Jaravata CV, Smith WL, Rensen GJ, Ruzante JM y Cullor JS. 2006. Detection of Mycobacterium avium subsp. paratuberculosis in
bovine manure using Whatman FTA card technology and Lightcycler real-time PCR. Foodborne Pathog.Dis., 3(2):212-5. Jayarao BM, Pillai SR, Wolfgang DR, Griswold DR, Rossiter CA, Tewari D, Burns CM y Hutchinson LJ. 2004. Evaluation of
IS900-PCR assay for detection of Mycobacterium avium subspecies paratuberculosis infection in cattle using quarter milk and bulk tank milk samples. Foodborne.Pathog.Dis, 1(1):17-26.
Jensen AG, Bennedsen J y Rosdahl VT. 1989. Plasmid profiles of Mycobacterium avium/intracellulare isolated from patients with AIDS or cervical lymphadenitis and from environmental samples. Scand.J.Infect.Dis., 21(6):645-9.
Jessup DA, Abbas B y Behymer D. 1981. Paratuberculosis in tule elk in California. J.Am.Vet.Med.Assoc., 179(11):1252-4. Johansen TB, Djonne B, Jensen MR y Olsen I. 2005. Distribution of IS1311 and IS1245 in Mycobacterium avium subspecies
revisited. J.Clin.Microbiol., 43(5):2500-2. Johne HA y Frothingham L. 1895. Ein eigenthümlicher Fall von Tuberculose beim Rind. Deut.Zeits.Tiermed.Vergl.Pathol., 21:438-
54. Johnson HW, Larsen AB, Henley AB y Groth AH. 1949. Studies on Johnin: VI.- The relationship of the allergens of
Mycobacterium paratuberculosis, Mycobacterium tuberculosis var. avium, bovis, and hominis, and Mycobacterium phlei. Am.J.Vet.Res., 35:138-41.
Johnson YJ, Kaneene JB, Gardiner JC, LLoyd JW, Sprecher DJ y Coe PH. 2001. The effect of subclinical Mycobacterium paratuberculosis infection on milk production in Michigan dairy cows. J.Dairy Sci., 84(10):2188-94.
Johnson-Ifearulundu Y y Kaneene JB. 1999a. Distribution and environmental risk factors for paratuberculosis in dairy cattle herds in Michigan. Am.J.Vet.Res., 60(5):489-596.
Johnson-Ifearulundu Y, Kaneene JB y LLoyd JW. 1999b. Herd -level economic analysis of the impact of paratuberculosis on dairy herds. J.Am.Vet.Med.Assoc., 214(6):822-6.
Jones PH, Farver TB, Beaman B, Cetinkaya B y Morgan KL. 2006. Crohn's disease in people exposed to clinical cases of bovine paratuberculosis. Epidemiol.Infect., 134(1):49-56.
Jones RL. 1989. Review of the economic impact of Johne's disease in the United States. En: Milner, A. R. y Wood, P. R. editores. Johne's disease: current trends in research, diagnosis and management. pp. 46-50. CSIRO. Melbourne, Australia.
Jorgensen JB. 1969. Paratuberculosis in pigs. Experimental infection by oral administration of Mycobacterium paratuberculosis. Acta Vet.Scand., 10:275-87.
Jorgensen JB. 1977. Survival of Mycobacterium paratuberculosis in slurry. Nord.Vet.Med., 29:267-70. Jorgensen JB. 1982. An improved medium for culture of Mycobacterium paratuberculosis from bovine faeces. Acta Vet.Scand.,
23:325-35. Jorgensen JB. 1983. The effect of vaccination on the excretion of Mycobacterium paratuberculosis. En: Merkal editor. Proc.1st
Int.Coll.Paratub., pp. 249-54. NADC, USDA, Ames, IA, USA. Jorgensen JB y Jensen PT. 1978. Enzyme-linked immunosorbent assay (ELISA) for detection of antibodies to Mycobacterium
paratuberculosis in cattle. Acta Vet.Scand., 19:310-2. Jovanovic Z. 1999. Epidemiology of Crohn's disease in the Rijeka-Istra region. Lijec.Vjesn., 121(1-2):8-13. Jubb KVF, Kennedy PC y Palmer N. 1985. Mycobacterial enteritis. En: Jubb, K. V. F., Kennedy, P. C. y Palmer, N. editores.
Pathology of Domestic Animals. pp. 155-9. Academic Press Inc. London, UK. Jucker MT y Falkinham III JO. 1990. Epidemiology of infection by nontuberculous mycobacteria IX. Evidence for two DNA
homology groups among small plasmids in Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium scrofulaceum. Am.Rev.Respir.Dis., 142(4):858-62.
Judge J, Kyriazakis I, Greig A, Allcroft DJ y Hutchings MR. 2005. Clustering of Mycobacterium avium subsp. paratuberculosis in rabbits and the environment: how hot is a hot spot? Appl.Environ.Microbiol., 71(10):6033-8.
Judge J, Kyriazakis I, Greig A, Davidson RS y Hutchings MR. 2006. Routes of Intraspecies Transmission of Mycobacterium avium subsp. paratuberculosis in Rabbits (Oryctolagus cuniculus): a Field Study. Appl.Environ.Microbiol., 72(1):398-403.

BIBLIOGRAFÍA CITADA
211
Jungersen G, Grell SN, Clemensen A, Roust T y Howard CJ. 2006. Interleukin-12 potentiation of the interferon-gamma test rescues day-old blood samples for the diagnosis of paratuberculosis. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 501-6. IAP, Madison, WI, USA.
Jungersen G, Huda A, Hasen JJ y Lind A. 2002. Interpretation of the gamma interferon test for diagnosis of subclinical paratuberculosis in cattle. Clin.Diagn.Lab.Immunol., 9(2):453-60.
Juste RA. 1990. Estudio experimental de las fases iniciales de la paratuberculosis ovina. Tesis Doctoral. Universidad de Zaragoza, Aragón, España.
Juste RA. 1997a. Debate/Discussion. Nomenclature: M. paratuberculosis vs M. avium subsp. paratuberculosis and strain definitions and differentiation. En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., pp. 102-13. IAP, Rehoboth, MA, USA.
Juste RA. 1997b. Johne's disease: a review of current knowledge. En: Allworth editor. Proc.4th Int.Congr.Sheep Vet., pp. 140-50. ASVS, Armidale, NSW, Australia.
Juste RA, Aduriz G, Bascones M, Foley E, Bargar TW y Barletta RG. 1993. Effect of iron on mycobactin production and dependence in Mycobacteria. Proc.74th Annu.Meet.CRWAD, p. 64. Chicago, IL, USA.
Juste RA y Aduriz JJ. 1990a. Diagnóstico. En: Juste editor. Paratuberculosis. Ovis, Aula Veterinaria, pp. 49-62. Luzán 5, Madrid, España.
Juste RA, Badiola JJ, Arnal MC, Balaguer L, García Marín JF, Sáez de Ocáriz C y Marco JC. 1991a. A survey of ovine paratuberculosis in Aragón (Spain) by different methods. The Paratuberculosis Newsletter, 3:3-4.
Juste RA, Cuervo LA, Gelabert JL, Sáez de Ocáriz C, Marco JC y Camon J. 1983. Paratuberculosis bovina en Vizcaya. Hyg.Pec., 5(1):57-66.
Juste RA, García Marín JF, Peris B, Sáez de Ocáriz C y Badiola JJ. 1994. Experimental infection of vaccinated and non-vaccinated lambs with Mycobacterium paratuberculosis. J.Comp.Pathol., 110(2):185-94.
Juste RA, Garrido JM y Aduriz G. 2000. El agente causal de la paratuberculosis y su situación taxonómica. En: Juste editor. Paratuberculosis. Bovis, Aula Veterinaria, pp. 13-28. Luzan 5, Madrid, España.
Juste RA, Marco JC, Saez de Ocariz C y Aduriz JJ. 1991b. Comparison of different media for the isolation of small ruminant strains of Mycobacterium paratuberculosis. Vet.Microbiol., 28:385-90.
Juste RA y Sáez de Ocáriz C. 1990b. Control. En: Juste editor. Paratuberculosis. Ovis, Aula Veterinaria, pp. 77-85. Luzán 5, Madrid.
Juste RA, Sáez de Ocáriz C y Aduriz JJ. 1990c. Investigación serológica de la paratuberculosis ovina: técnicas y resultados. En: Juste editor. 3ª Reu.PTBC España (1989), pp. 17-20. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España.
Juste RA, Sáez de Ocáriz C y Marco JC. 1988. Diagnosis of clinical paratuberculosis in sheep: A comparison of pathological, serological, and bacteriological methods. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 220-6. LCRV, Maisons-Alfort, France.
Juste RA, Sáez de Ocáriz C, Portu J y Aldamiz M. 1995. Seroreactivity of Crohn's patients against paratuberculosis antigens. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., p. 321. IAP, Rehoboth, MA, USA.
Kalis CH, Barkema HW, Hesselink JW, van Maanen C y Collins MT. 2002. Evaluation of two absorbed enzyme-linked immunosorbent assays and a complement fixation test as replacements for fecal culture in the detection of cows shedding Mycobacterium avium subspecies paratuberculosis. J.Vet.Diagn.Invest., 14(3):219-24.
Kalis CH, Collins MT, Hesselink JW y Barkema HW. 2003a. Specificity of two tests for the early diagnosis of bovine paratuberculosis based on cell-mediated immunity: the Johnin skin test and the gamma interferon assay. Vet.Microbiol., 97(1-2):73-86.
Kalis CH, Hesselink JW, Barkema HW y Collins MT. 2000. Culture of strategically pooled bovine fecal samples as a method to screen herds for paratuberculosis. J.Vet.Diagn.Invest., 12(6):547-51.
Kalis CH, Hesselink JW, Barkema HW y Collins MT. 2001. Use of long-term vaccination with a killed vaccine to prevent fecal shedding of Mycobacterium avium subsp. paratuberculosis in dairy herds. Am.J.Vet.Res., 62(2):270-4.
Kalis CHJ, Barkema HW y Hesselink JW. 1999a. Certification of dairy herds as free of paratuberculosis using cultures of strategically pooled fecal samples. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 55-8. IAP, Madison, WI, USA.
Kalis CHJ, Benedictus G, van Weering HJ, Flamand F y Haagsma J. 1992. Experiences with the use of an experimental vaccine in the control of paratuberculosis in The Netherlands. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 484-94. Proc.8th Int.Coll.Paratub., Providence, RI, USA.
Kalis CHJ, Collins MT, Hesselink JW y Barkema HW. 2003b. Rapid eradication of Paratuberculosis. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 413-6. IAP, Madison, WI, USA.
Kalis CHJ, Hesselink JW y Barkema HW. 1999b. Vaccination does not prevent faecal shedding of Mycobacterium avium subsp. paratuberculosis. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 153-6. IAP, Madison, WI, USA.
Kallinowski F, Wassmer A, Hofmann MA, Harmsen D, Heesemann J, Karch H, Herfarth C y Buhr HJ. 1998. Prevalence of enteropathogenic bacteria in surgically treated chronic inflammatory bowel disease. Hepatogastroenterology, 45(23):1552-8.
Kanazawa K, Haga Y, Funakoshi O, Nakajima H, Munakata A y Yoshida Y. 1999. Absence of Mycobacterium paratuberculosis DNA in intestinal tissues from Crohn's disease by nested polymerase chain reaction. J.Gastroenterol., 34(2):200-6.
Karpinski T. 1972. Isolation and cultivation of Mycobacterium johnei. Bull.Vet.Inst.Pulawy, 16:1-2. Katayama N, Suzuki T, Ootake M, Totsuka T, Kamata S y Yokomizo Y. 2003. Effects of ammonia treatment under various
conditions on viability of Mycobacterium avium subsp paratuberculosis inoculated in low moisture roughage. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 16-20. IAP, Madison, WI, USA.
Kaufmann SH. 1995. Immunity to intracellular microbial pathogens. Immunol.Today, 16(7):338-42. Kaufmann SH. 2001. How can immunology contribute to the control of tuberculosis? Nat.Rev.Immunol., 1(1):20-30. Kauppinen J, Mantyjarvi R y Katila ML. 1994. Random amplified polymorphic DNA genotyping of Mycobacterium malmoense.
J.Clin.Microbiol., 32(7):1827-9. Kennedy DJ y Benedictus G. 2001. Control of Mycobacterium avium subsp. paratuberculosis infection in agricultural species. Rev.-
Off.Int.Epizoot., 20(1):151-79. Kennedy DJ, Citer L y Sergeant ESG. 2006. Increasing improvement of herd owners in controlling paratuberculosis through
assurance based trading. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 20-5. IAP, Madison, WI, USA. Keswani J y Frank JF. 1998. Thermal inactivation of Mycobacterium paratuberculosis in milk. J.Food Prot., 61(8):974-8. Khare S, Ficht TA, Santos RL, Romano J, Ficht AR, Zhang S, Grant IR, Libal M, Hunter D y Adams LG. 2004. Rapid and sensitive
detection of Mycobacterium avium subsp. paratuberculosis in bovine milk and feces by a combination of immunomagnetic bead separation-conventional PCR and real-time PCR. J.Clin.Microbiol., 42(3):1075-81.

BIBLIOGRAFÍA CITADA
212
Khol JL, Dreier S, Stein B, Fuchs K, Gütler S y Baumgartner W. 2006. Prevalence of Mycobacterium avium ssp. paratuberculosis in Austrian cattle 2002/03 in comparison to 1997/98. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. Abstract. IAP, Madison, WI, USA.
Kim JR y Kim CH. 1997. Genomic polymorphism in clinical mycobacterial strains analyzed by pulsed-field gel electrophoresis. J.Microbiol., 35(3):172-6.
Kim SG, Shin SJ, Jacobson RH, Miller LJ, Harpending PR, Stehman SM, Rossiter CA y Lein DA. 2002. Development and application of quantitative polymerase chain reaction assay based on the ABI 7700 system (TaqMan) for detection and quantification of Mycobacterium avium subsp. paratuberculosis. J.Vet.Diagn.Invest., 14(2):126-31.
Kim YG, Bech-Nielsen S, Gordon JC, Slemons RD y Spangler E. 1988. Fecal culture of Mycobacterium paratuberculosis in cattle using a centrifugation specimen processing step. Acta Vet.Scand., 84:203-5.
Kim YG, Bech-Nielsen S, Gordon JC, Slemons RD y Spangler E. 1989. Comparison of two methods for isolation of Mycobacterium paratuberculosis from bovine fecal samples. Am.J.Vet.Res., 50(7):1110-3.
Kirschner P y Bottger EC. 1998. Species identification of mycobacteria using rDNA sequencing. Methods Mol.Biol., 101:349-61. Kirschner P, Springer B, Vogel U, Meier A, Wrede A, Kiekenbeck M, Bange FC y Bottger EC. 1993. Genotypic identification of
mycobacteria by nucleic acid sequence determination: report of a 2-year experience in a clinical laboratory. J.Clin.Microbiol., 31(11):2882-9.
Klausen J, Huda A, Ekeroth L y Ahrens P. 2003. Evaluation of serum and milk ELISAs for paratuberculosis in Danish dairy cattle. Prev.Vet.Med., 58(3-4):171-8.
Klijn N, Herrewegh AA y de Jong P. 2001. Heat inactivation data for Mycobacterium avium subsp. paratuberculosis: implications for interpretation. J.Appl.Bacteriol., 91(4):697-704.
Kobayashi K, Blaser MJ y Brown WR. 1989. Immunohistochemical examination for mycobacteria in intestinal tissues from patients with Crohn's disease. Gastroenterology, 96:1009-15.
Koenig GJ, Hoffsis GF, Shulaw WP, Bech-Nielsen S, Rings DM y St Jean G. 1993. Isolation of Mycobacterium paratuberculosis from mononuclear cells in tissues, blood, and mammary glands of cows with advanced paratuberculosis. Am.J.Vet.Res., 54(9):1441-5.
Koets A, Hoek A, Langelaar M, Overdijk M, Santema W, Franken P, Eden W y Rutten V. 2006a. Mycobacterial 70 kD heat-shock protein is an effective subunit vaccine against bovine paratuberculosis. Vaccine, 24(14):2550-9.
Koets A, Rutten V, Hoek A, van Mil F, Muller K, Bakker D, Gruys E y van Eden W. 2002. Progressive bovine paratuberculosis is associated with local loss of CD4(+) T cells, increased frequency of gamma delta T cells, and related changes in T-cell function. Infect.Immun., 70(7):3856-64.
Koets AP. 2006. Immunology of bovine paratuberculosis: the hosts' perspective. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 77-87. IAP, Madison, WI, USA.
Koets AP, Adugna G, Janss LL, van Weering HJ, Kalis CH, Wentink GH, Rutten VP y Schukken YH. 2000. Genetic variation of susceptibility to Mycobacterium avium subsp. paratuberculosis infection in dairy cattle. J.Dairy Sci., 83(11):2702-8.
Koets AP, Langelaar M, Hoek A, Bakker D, Willemsen PTJ, van Eden W y Rutten V. 2006b. Evidence for distinct host response patterns in cows experimentally infected with M. avium subspecies paratuberculosis. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 154. IAP, Madison, WI, USA.
Koets AP, Rutten VP, de Boer M, Bakker D, Valentin-Weigand P y van Eden W. 2001. Differential changes in heat shock protein-, lipoarabinomannan-, and purified protein derivative-specific immunoglobulin G1 and G2 isotype responses during bovine Mycobacterium avium subsp. paratuberculosis infection. Infect.Immun., 69(3):1492-8.
Koets AP, Rutten VPMG, Hoek A, Bakker D, van Zijderveld F, Müller KE y van Eden W. 1999. Heat-shock protein-specific T-cell responses in various stages of bovine paratuberculosis. Vet.Immunol.Immunopathol., 70:105-15.
Kohler H, Gyra H, Zimmer K, Drager KG, Burkert B, Lemser B, Hausleithner D, Cubler K, Klawonn W y Hess RG. 2001. Immune reactions in cattle after immunization with a Mycobacterium paratuberculosis vaccine and implications for the diagnosis of M. paratuberculosis and M. bovis infections. J.Vet.Med.B Infect.Dis.Vet.Public Health, 48(3):185-95.
Koo HC, Park YH, Ahn J, Waters WR, Hamilton MJ, Barrington G, Mosaad AA, Palmer MV, Shin S y Davis WC. 2004. New latex bead agglutination assay for differential diagnosis of cattle infected with Mycobacterium bovis and Mycobacterium avium subsp. paratuberculosis. Clin.Diagn.Lab.Immunol., 11(6):1070-4.
Kopecky KE, Booth GD, Merkal RS y Larsen AB. 1971. Hematologic changes associated with intravenous johnin testing in cattle. Am.J.Vet.Res., 32(9):1343-7.
Kopecky KE, Larsen AB y Merkal RS. 1967. Uterine infection in bovine paratuberculosis. Am.J.Vet.Res., 28(125):1043-5. Koren O, Elad D, Schafer M, Garazi S, Meltser R y Klement E. 2006. Johne´s disease control programs in the Israeli dairy herds.
En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 26-9. IAP, Madison, WI, USA. Kormendy B. 1990. Paratuberculosis in a cattle herd: comparison of allergic, serologic and faecal microscopic tests. Acta
Microbiol.Hung., 37(2):219-22. Kormendy B. 1994. The effect of vaccination on the prevalence of paratuberculosis in large dairy herds. Vet.Microbiol., 41(1-
2):117-25. Kormendy B, Nagy GY y Illes J. 1984. Cross-reactions between Mycobacterium avium and Mycobacterium paratuberculosis strains
in complement fixation and gel precipitation tests. Acta Vet.Hung., 32(1/2):3-7. Kremer L, Gurcha SS, Bifani P, Hitchen PG, Baulard A, Morris HR, Dell A, Brennan PJ y Besra GS. 2002. Characterization of a
putative alpha-mannosyltransferase involved in phosphatidylinositol trimannoside biosynthesis in Mycobacterium tuberculosis. Biochem.J., 363(Pt 3):437-47.
Kreuzpaintner G, Kirschner P, Wallner A, Kolble R, Hesterberg R, Thomas L y Borchard F. 1995. Mycobacteria of Runyon groups I, II and IV do not play an aetiological role in Crohn's disease. Eur.J.Gastroenterol.Hepatol., 7(12):1177-82.
Kruip TA, Muskens J, van Roermund HJ, Bakker D y Stockhofe-Zurwieden N. 2003. Lack of association of Mycobacterium avium subsp. paratuberculosis with oocytes and embryos from moderate shedders of the pathogen. Theriogenology, 59(7):1651-60.
Kudahl A, Nielsen SS y Sorensen JT. 2004. Relationship between antibodies against Mycobacterium avium subsp. paratuberculosis in milk and shape of lactation curves. Prev.Vet.Med., 62(2):119-34.
Kuehnel MP, Goethe R, Habermann A, Mueller E, Rohde M, Griffiths G y Valentin-Weigand P. 2001. Characterization of the intracellular survival of Mycobacterium avium ssp. paratuberculosis: phagosomal pH and fusogenicity in J774 macrophages compared with other mycobacteria. Cell.Microbiol., 3(8):551-66.
Kunze ZM, Wall S, Appelberg R, Silva MT, Portaels F y McFadden JJ. 1991. IS901, a new member of a widespread class of atypical insertion sequences, is associated with pathogenicity in Mycobacterium avium. Mol.Microbiol., 5(9):2265-72.
Lagadic MA, Menec LM y Argenté G. 1983. Techniques de cultures de Mycobacterium paratuberculosis: Leur utilisation en routine dans laboratoire de diagnostic. Rec.Med.Vet.Ec.Alfort, 159(10):801-7.

BIBLIOGRAFÍA CITADA
213
Lambeth C, Reddacliff LA, Windsor P, Abbott KA, McGregor H y Whittington RJ. 2004. Intrauterine and transmammary transmission of Mycobacterium avium subsp. paratuberculosis in sheep. Aust.Vet.J., 82(8):504-8.
Lambrecht RS y Collins MT. 1992. Mycobacterium paratuberculosis: Factors that influence mycobactin dependence. Diagn.Microbiol.Infect.Dis., 15:239-46.
Lamo-Castellvi S, Capellas M, Lopez-Pedemonte T, Hernandez-Herrero MM, Guamis B y Roig-Sagues AX. 2005. Behavior of Yersinia enterocolitica strains inoculated in model cheese treated with high hydrostatic pressure. J.Food Prot., 68(3):528-33.
Langelaar M, Koets A, Muller K, van Eden W, Noordhuizen J, Howard C, Hope J y Rutten V. 2002. Mycobacterium paratuberculosis heat shock protein 70 as a tool in control of paratuberculosis. Vet.Immunol.Immunopathol., 87(3-4):239-44.
Langelaar MF, Hope JC, Rutten VP, Noordhuizen JP, van Eden W y Koets AP. 2005a. Mycobacterium avium ssp. paratuberculosis recombinant heat shock protein 70 interaction with different bovine antigen-presenting cells. Scand.J.Immunol., 61(3):242-50.
Langelaar MF, Weber CN, Overdijk MB, Muller KE, Koets AP y Rutten VP. 2005b. Cytokine gene expression profiles of bovine dendritic cells after interaction with Mycobacterium avium ssp. paratuberculosis (M.a.p.), Escherichia coli (E. coli) or recombinant M.a.p. heat shock protein 70. Vet.Immunol.Immunopathol., 107(1-2):153-61.
Larsen AB. 1972. Paratuberculosis: The status of our knowledge. J.Am.Vet.Med.Assoc., 161:1539-41. Larsen AB, Hawkins WW y Merkal RS. 1964. Experimental vaccination of sheep against Johne's disease. Am.J.Vet.Res.,
25(107):974-6. Larsen AB y Kopecky KE. 1970. Mycobacterium paratuberculosis in reproductive organs and semen of bulls. Am.J.Vet.Res.,
31:255-8. Larsen AB, Merkal RS y Cutlip RC. 1975. Age of cattle as related to resistance to infection with Mycobacterium paratuberculosis.
Am.J.Vet.Res., 36(3):255-7. Larsen AB, Merkal RS y Vardaman TH. 1956. Survival time of Mycobacterium paratuberculosis. Am.J.Vet.Res., 17(64):549-51. Larsen AB y Miller JM. 1978a. Effect of dexamethasone on Mycobacterium paratuberculosis infections in hamsters.
Am.J.Vet.Res., 39(11):1866-7. Larsen AB y Moon HW. 1972a. Experimental Mycobacterium paratuberculosis infection in chickens. Am.J.Vet.Res., 33:1231-5. Larsen AB, Moon HW y Merkal RS. 1971. Susceptibility of swine to Mycobacterium paratuberculosis. Am.J.Vet.Res., 32(4):589-
95. Larsen AB, Moon HW y Merkal RS. 1972b. Susceptibility of horses to Mycobacterium paratuberculosis. Am.J.Vet.Res.,
33(11):2185-9. Larsen AB, Moyle AI y Himes EM. 1978b. Experimental vaccination of cattle against paratuberculosis (Johne`s disease) with killed
bacterial vaccines: A controlled field study. Am.J.Vet.Res., 39:65-9. Larsen AB, Stalheim OHV y Hughes DE. 1981. Mycobacterium paratuberculosis in the semen and genital organs of a semen-donor
bull. J.Am.Vet.Med.Assoc., 179:169-71. Larsen AB y Vardaman TH. 1953. The effect of isonicotinic acid hidrazide on Mycobacterium paratuberculosis.
J.Am.Vet.Med.Assoc., 122(913):309-10. Larsen AB, Vardaman TH, Kopecky KE y Merkal RS. 1965. An extended study of a herd of cattle naturally infected with Johne`s
disease: the significance of the hemagglutination test. Am.J.Vet.Res., 26(111):254-7. Lawrence WE. 1956. Congenital infection with M. paratuberculosis in cattle. Vet.Rec., 68:312-4. Leao SC, Briones MR, Sircili MP, Balian SC, Mores N y Ferreira-Neto JS. 1999. Identification of two novel Mycobacterium avium
allelic variants in pig and human isolates from Brazil by PCR-restriction enzyme analysis. J.Clin.Microbiol., 37(8):2592-7. Lebrun L, Weill FX, Lafendi L, Houriez F, Casanova F, Gutierrez MC, Ingrand D, Lagrange P, Vincent V y Herrmann JL. 2005.
Use of the INNO-LiPA-MYCOBACTERIA assay (version 2) for identification of Mycobacterium avium-Mycobacterium intracellulare-Mycobacterium scrofulaceum complex isolates. J.Clin.Microbiol., 43(6):2567-74.
Leclerc MC, Haddad N, Moreau R y Thorel MF. 2000. Molecular characterization of environmental mycobacterium strains by PCR-restriction fragment length polymorphism of hsp65 and by sequencing of hsp65, and of 16S and ITS1 rDNA. Res.Microbiol., 151(8):629-38.
Lee H, Stabel JR y Kehrli ME, Jr. 2001. Cytokine gene expression in ileal tissues of cattle infected with Mycobacterium paratuberculosis. Vet.Immunol.Immunopathol., 82(1-2):73-85.
Leid JG, Hunter D y Speer CA. 2002. Early diagnosis of Johne's disease in the American bison by monoclonal antibodies directed against antigen 85. Ann.N.Y.Acad.Sci., 969:66-72.
León L, Garrido F, Pardo A, Hermoso M y Carranza J. 1984. Observaciones sobre la paratuberculosis caprina en España meridional. Arch.Zootec., 33(127):205-17.
Lepper AWD. 1989. The aetiology and pathogenesis of Johne's disease. En: Milner, A. R. y Wood, P. R. editores. Johne's disease: current trends in research, diagnosis and management. pp. 74-86. CSIRO. Melbourne, Australia.
Leslie P, Riemann HP, West G y Moe A. 1988. Vaccination field trial for Mycobacterium paratuberculosis (Johne's disease) in the caprine species. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 110-29. LCRV, Maisons-Alfort, France.
Levy-Frebault VV y Portaels F. 1992. Proposed minimal standards for the genus Mycobacterium and for description of new slowly growing Mycobacterium species. Int.J.Syst.Bacteriol., 42(2):315-23.
Levy-Frebault VV, Thorel MF, Varnerot A y Gicquel B. 1989. DNA polymorphism in Mycobacterium paratuberculosis, "wood pigeon mycobacteria", and related mycobacteria analyzed by field inversion gel electrophoresis. J.Clin.Microbiol., 27(12):2823-6.
Li L, Bannantine JP, Zhang Q, Amonsin A, May BJ, Alt D, Banerji N, Kanjilal S y Kapur V. 2005. The complete genome sequence of Mycobacterium avium subspecies paratuberculosis. Proc.Natl.Acad.Sci.U.S.A., 102(35):12344-9.
Libke KG y Walton AM. 1975. Presumptive paratuberculosis in a Virginia white-tailed deer. J.Wildl.Dis., 11(4):552-3. Liebler EM, Pohlenz JF y Cheville NF. 1988a. Gut associated lymphoid tissue in the large intestine of calves. II. Electron
microscopy. Vet.Pathol., 25:509-15. Liebler EM, Pohlenz JF y Woode GN. 1988b. Gut associated lymphoid tissue in the large intestine of calves. I. Distribution and
histology. Vet.Pathol., 25:503-8. Lillini E, Bitonti G, Gamberale F y Cersini A. 2006. Prevalence of bovine paratuberculosis in the Latium region (Italy). En:
Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 638-44. IAP, Madison, WI, USA. Lillini E, Gamberale F, De Grossi L, Cersini A, Scherm B y Fagiolo A. 2003. Comparative study on detection of Mycobacterium
avium subsp. paratuberculosis by PCR diagnosis and conventional culture on feaces of water-buffalo herds in the Latium region (Italy). En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 283-6. IAP, Madison.
Linton CJ, Jalal H, Leeming JP y Millar MR. 1994. Rapid discrimination of Mycobacterium tuberculosis strains by random amplified polymorphic DNA analysis. J.Clin.Microbiol., 32(9):2169-74.
Lisby G, Andersen J, Engbaek K y Binder V. 1994. Mycobacterium paratuberculosis in intestinal tissue from patients with Crohn's disease demonstrated by a nested primer polymerase chain reaction. Scand.J.Gastroenterol., 29(10):923-9.

BIBLIOGRAFÍA CITADA
214
Liu X, Feng Z, Harris NB, Cirillo JD, Bercovier H y Barletta RG. 2001. Identification of a secreted superoxide dismutase in Mycobacterium avium ssp. paratuberculosis. FEMS Microbiol.Lett., 202(2):233-8.
Lloyd JB, Whittington RJ, Fitzgibbon C y Dobson R. 2001. Presence of Mycobacterium avium subspecies paratuberculosis in suspensions of ovine trichostrongylid larvae produced in faecal cultures artificially contaminated with the bacterium. Vet.Rec., 148(9):261-3.
Loftus EV, Jr., Silverstein MD, Sandborn WJ, Tremaine WJ, Harmsen WS y Zinsmeister AR. 1998. Crohn's disease in Olmsted County, Minnesota, 1940-1993: incidence, prevalence, and survival. Gastroenterology, 114(6):1161-8.
Lombard JE, Byrem TM, Wagner BA y McCluskey BJ. 2006. Comparison of milk and serum enzyme-linked immunosorbent assays for diagnosis of Mycobacterium avium subspecies paratuberculosis infection in dairy cattle. J.Vet.Diagn.Invest., 18(5):448-58.
Lombard JE, Garry FB, McCluskey BJ y Wagner BA. 2005. Risk of removal and effects on milk production associated with paratuberculosis status in dairy cows. J.Am.Vet.Med.Assoc., 227(12):1975-81.
Lopez-Pedemonte TJ, Roig-Sagues AX, Trujillo AJ, Capellas M y Guamis B. 2003. Inactivation of spores of Bacillus cereus in cheese by high hydrostatic pressure with the addition of nisin or lysozyme. J.Dairy Sci., 86(10):3075-81.
Losinger WC. 2005. Economic impact of reduced milk production associated with Johne's disease on dairy operations in the USA. J.Dairy Res., 72(4):425-32.
Lovell R, Levi ML y Francis J. 1944. Studies on the survival of Johne's bacilli. J.Comp.Pathol., 34:120-9. Lucas JN, Cousins DV, Mills AJ y Van Wijk JG. 2003. Identification of Mycobacterium avium subsp. avium in an alpaca with
lesions resembling paratuberculosis. Aust.Vet.J., 81(9):567-9. Lugton I. 1999. Mucosa-associated lymphoid tissues as sites for uptake, carriage and excretion of tubercle bacilli and other
pathogenic mycobacteria. Immunol.Cell Biol., 77(4):364-72. Lund BM, Gould GW y Rampling AM. 2002. Pasteurization of milk and the heat resistance of Mycobacterium avium subsp.
paratuberculosis: a critical review of the data. Int.J.Food Microbiol., 77(1-2):135-45. Luyven G, Vom SA y Sasserath M. 2002. Paratuberculosis eradication programs in Northrhine-Westfalia.
Dtsch.Tierarztl.Wochenschr., 109(12):524-7. Lyle PAS y Merkal RS. 1983. Comparison of ELISA and gel diffusion precipitin tests for paratuberculosis in cattle, sheep and
goats. En: Merkal editor. Proc.1st Int.Coll.Paratub., pp. 109-12. NADC, USDA, Ames, IA, USA. Machackova M, Lamka J, Yayo Ayele W, Parmova I, Svastova P, Amemori T y Pavlik I. 2003a. Infection of ruminants by
uncultivable strains of Mycobacterium avium subsp. paratuberculosis in the Czech Republic. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 191-6. IAP, Madison, WI, USA.
Machackova M, Matlova L, Lamka J, Smolik J, Melicharek I, Hanzlikova M, Docekal J, Cvetnic Z, Nagy G, Lipiec M, Ocepek M y Pavlik I. 2003b. Wild boar (Sus scrofa) as a possible vector of mycobacterial infections: review of literature and critical analysis of data from Central Europe between 1983 to 2001. Vet.Med.(Praha), 48(3):51-65.
Machackova M, Svastova P, Lamka J, Parmova I, Liska V, Smolik J, Fischer OA y Pavlik I. 2004. Paratuberculosis in farmed and free-living wild ruminants in the Czech Republic (1999-2001). Vet.Microbiol., 101(4):225-34.
Macham LP, Ratledge C y Nocton JC. 1975. Extracellular iron acquisition by mycobacteria: role of the exochelins and evidence against the participation of mycobactin. Infect.Immun., 12(6):1242-51.
Mackintosh C, Haigh JC y Griffin F. 2002. Bacterial diseases of farmed deer and bison. Rev.- Off.Int.Epizoot., 21(2):249-63. Mackintosh CG, de Lisle GW, Collins DM y Griffin JF. 2004. Mycobacterial diseases of deer. N.Z.Vet.J., 52(4):163-74. Maher M, Rowe M, Egan J, Hill C y Murphy P. 2004. Detection and survival of Mycobacterium avium subsp. paratuberculosis.
Project code FS005, http://www.relayresearch.ie/public/p_research_project_details.asp?project_id=6&RTID=6. Mainar-Jaime RC y Vázquez-Boland JA. 1998. Factors associated with seroprevalence to Mycobacterium paratuberculosis in small-
ruminants farms in the Madrid region (Spain). Prev.Vet.Med., 34:317-27. Malik R, Shaw SE, Griffin C, Stanley B, Burrows AK, Bryden SL, Titmarsh J, Stutsel MJ, Carter SA, Warner A, Martin P, Wigney
DI y Gilpin C. 2004. Infections of the subcutis and skin of dogs caused by rapidly growing mycobacteria. J.Small Anim.Pract., 45(10):485-94.
Malik R, Wigney DI, Dawson D, Martin P, Hunt GB y Love DN. 2000. Infection of the subcutis and skin of cats with rapidly growing mycobacteria: a review of microbiological and clinical findings. J.Feline Med.Surg., 2(1):35-48.
Manning EJ, Steinberg H, Rossow K, Ruth GR y Collins MT. 1998. Epizootic of paratuberculosis in farmed elk. J.Am.Vet.Med.Assoc., 213(9):1320-1.
Marco I, Ruiz M, Juste R, Garrido JM y Lavin S. 2002. Paratuberculosis in free-ranging fallow deer in Spain. J.Wildl.Dis., 38(3):629-32.
Markesich DC, Graham DY y Yoshimura HH. 2004. Progress in culture and subculture of spheroplasts and fastidious acid-fast bacilli isolated from intestinal tissues. J.Clin.Microbiol., 26(8):1600-3.
Marly J, Thorel MF, Perrin GG, Pardon P y Guerrault PJ. 1988. Suivi de vaccination de chevrettes contre la paratuberculose: Consequences cliniques, serologiques et allergiques et epreuve virulente. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 99-109. LCRV, Maisons-Alfort, France.
Marri PR, Bannantine JP, Paustian ML y Golding GB. 2006. Lateral gene transfer in Mycobacterium avium subspecies paratuberculosis. Can.J.Microbiol., 52(6):560-9.
Marsh I, Whittington R y Cousins D. 1999. PCR-restriction endonuclease analysis for identification and strain typing of Mycobacterium avium subsp. paratuberculosis and Mycobacterium avium subsp. avium based on polymorphisms in IS1311. Mol.Cell.Probes, 13(2):115-26.
Marsh I, Whittington R y Millar D. 2000. Quality control and optimized procedure of hybridization capture-PCR for the identification of Mycobacterium avium subsp. paratuberculosis in faeces. Mol.Cell.Probes, 14(4):219-32.
Marsh IB, Bannantine JP, Paustian ML, Tizard ML, Kapur V y Whittington RJ. 2006. Genomic comparison of Mycobacterium avium subsp. paratuberculosis sheep and cattle strains by microarray hybridization. J.Bacteriol., 188(6):2290-3.
Marsh IB y Whittington RJ. 2001. Progress towards a rapid polymerase chain reaction diagnostic test for the identification of Mycobacterium avium subsp. paratuberculosis in faeces. Mol.Cell.Probes, 15(2):105-18.
Marshall DJ, Ottaway SJ, Eamens GJ y Manchester PE. 1997. Validation of a diagnostic strategy for detection of ovine Johne's disease in New South Wales sheep flocks. En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., vol.1996, pp. 286-9. IAP, Madison, WI, USA.
Mason O, Marsh IB y Whittington RJ. 2001. Comparison of immunomagnetic bead separation-polymerase chain reaction and faecal culture for the detection of Mycobacterium avium subsp paratuberculosis in sheep faeces. Aust.Vet.J., 79(7):497-500.
Massone AR, Martin AA, Ibargoyen GS y Gimeno EJ. 1990. Immunohistochemical method for the visualization of Mycobacterium paratuberculosis in bovine tissues. J.Vet.Med.B Infect.Dis.Vet.Public Health, 37:251-3.
Matsiota-Bernard P, Waser S, Tassios PT, Kyriakopoulos A y Legakis NJ. 1997. Rapid discrimination of Mycobacterium avium strains from AIDS patients by randomly amplified polymorphic DNA analysis. J.Clin.Microbiol., 35(6):1585-8.

BIBLIOGRAFÍA CITADA
215
Matsiota-Bernard P, Zinzendorf N, Onody C y Guenounou M. 2000. Comparison of clarithromycin-sensitive and clarithromycin-resistant Mycobacterium avium strains isolated from AIDS patients during therapy regimens including clarithromycin. J.Infect., 40(1):49-54.
Matzanke BF, Bohnke R, Mollmann U, Reissbrodt R, Schunemann V y Trautwein AX. 1997. Iron uptake and intracellular metal transfer in mycobacteria mediated by xenosiderophores. Biometals, 10(3):193-203.
Mayer G. 2005. Bacteriology. Exchange of genetic information. En: Hunt, R. C. editor. Microbiology and Immunology On-line. http://pathmicro.med.sc.edu/2003-bacpdf/geneexchange2003.pdf. University of South Carolina School of Medicine. Columbia, SC, USA.
Mazars E, Lesjean S, Banuls AL, Gilbert M, Vincent V, Gicquel B, Tibayrenc M, Locht C y Supply P. 2001. High-resolution minisatellite-based typing as a portable approach to global analysis of Mycobacterium tuberculosis molecular epidemiology. Proc.Natl.Acad.Sci.U.S.A., 98(4):1901-6.
Mazurek GH, Hartman S, Zhang Y, Brown BA, Hector JS, Murphy D y Wallace RJ, Jr. 1993. Large DNA restriction fragment polymorphism in the Mycobacterium avium-M. intracellulare complex: a potential epidemiologic tool. J.Clin.Microbiol., 31(2):390-4.
Mazurier S, van de GA, Heuvelman K y Wernars K. 1992. RAPD analysis of Campylobacter isolates: DNA fingerprinting without the need to purify DNA. Lett.Appl.Microbiol., 14(6):260-2.
McCausland IP. 1980. Apparent Johne's disease in a sheep. Aust.Vet.J., 56(11):564. McClure HM, Chiodini RJ, Anderson DC, Swenson RB, Thayer WR y Coutu JA. 1987. Mycobacterium paratuberculosis infection
in a colony of stumptail Maccaques (Maccaca arctoides). J.Infect.Dis., 155:1011-9. McDonald WL, O'Riley KJ, Schroen CJ y Condron RJ. 2005. Heat inactivation of Mycobacterium avium subsp. paratuberculosis in
milk. Appl.Environ.Microbiol., 71(4):1785-9. McDonald WL, Ridge SE, Hope AF y Condron RJ. 1999. Evaluation of diagnostic test for Johne's disease in young cattle.
Aust.Vet.J., 77(2):113-9. McEwen AD. 1939. Investigations on Johne`s disease of sheep. J.Comp.Pathol., 52:69-87. McEwen AD y Samuel JM. 1958. The intravenous inoculation of sheep with graded dosses of Mycobacterium johnei.
J.Comp.Pathol., 68:45. McFadden JJ, Butcher PD, Chiodini RJ y Hermon-Taylor J. 1987. Crohn's disease-isolated mycobacteria are identical to
Mycobacterium paratuberculosis, as determined by probes that distinguish between mycobacterial species. J.Clin.Microbiol., 25(5):796-801.
McFadden JJ y Houdayer C. 1988. No evidence for antibodies to mycobacterial A60 antigen in Crohn's disease sera by enzime-linked immunoabsorbent assay (ELISA). J.Med.Bacteriol., 25:295-8.
McGee ZA, Wittler RG, Gooder H y Charache P. 1971. Wall-defective microbial variants: terminology and experimental design. J.Infect.Dis., 123(4):433-8.
McKenna SL, Keefe GP, Barkema HW, McClure J, VanLeeuwen JA, Hanna P y Sockett DC. 2004. Cow-level prevalence of paratuberculosis in culled dairy cows in Atlantic Canada and Maine. J.Dairy Sci., 87(11):3770-7.
McKenna SL, Keefe GP, Barkema HW y Sockett DC. 2005a. Evaluation of three ELISAs for Mycobacterium avium subsp. paratuberculosis using tissue and fecal culture as comparison standards. Vet.Microbiol., 110(1-2):105-11.
McKenna SL, Sockett DC, Keefe GP, McClure J, VanLeeuwen JA y Barkema HW. 2005b. Comparison of two enzyme-linked immunosorbent assays for diagnosis of Mycobacterium avium subsp. paratuberculosis. J.Vet.Diagn.Invest., 17(5):463-6.
McNab WB, Meek AH, Duncan JR, Martin S.W. y Van Dreumel A.A. 1991. An epidemiological study of paratuberculosis in dairy cattle in Ontario: study desing and prevalence estimates. Can.J.Vet.Res., 55:246-51.
McNabb A, Eisler D, Adie K, Amos M, Rodrigues M, Stephens G, Black WA y Isaac-Renton J. 2004. Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources. J.Clin.Microbiol., 42(7):3000-11.
Menchén V. 1995. Estudio inmunocitoquímico de la tuberculosis y paratuberculosis caprina. Tesis Doctoral. Universidad de Murcia, Murcia, España.
Mendes S, Albuquerque T, Afonso A, Boinas F y Amado A. 2006. Comparison of the methods used for the diagnosis of paratuberculosis infection in herds of small ruminants in Portugal. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., IAP, Madison, WI, USA.
Menendez MC, Garcia MJ, Navarro MC, Merchand JA, Rivera-Gutierrez S, Garcia-Sanchez L y Cox RA. 2002. Characterization of an rRNA operon (rrnB) of Mycobacterium fortuitum and other mycobacterial species: implications for the classification of mycobacteria. J.Bacteriol., 184(4):1078-88.
Merkal RS. 1973. Laboratory diagnosis of bovine paratuberculosis. J.Am.Vet.Med.Assoc., 163(9):1100-2. Merkal RS. 1983. Report on the International Colloquium on research in paratuberculosis. National Animal Disease Center, Ames,
Iowa. Merkal RS. 1984. Paratuberculosis: Advances in cultural, serologic and vaccination methods. J.Am.Vet.Med.Assoc., 184(8):939-43. Merkal RS y Cullogh Mc. WG. 1982a. A new mycobactin, mycobactin J, from Mycobaterium paratuberculosis. Curr.Microbiol.,
7:333-5. Merkal RS y Curran BJ. 1974. Growth and metabolic characteristics of Mycobacterium paratuberculosis. Appl.Microbiol., 28:276-
9. Merkal RS, Kopecky KE, Larsen AB y Ness RN. 1970. Immunologic mechanisms in bovine paratuberculosis. Am.J.Vet.Res.,
31(3):475-85. Merkal RS, Kopecky KE, Larsen AB y Thurston JR. 1964. Improvements in the techniques for primary cultivation of
Mycobacterium paratuberculosis. Am.J.Vet.Res., 25(107):1290-3. Merkal RS y Larsen AB. 1973. Clofazimine treatment of cows naturally infected with Mycobacterium paratuberculosis.
Am.J.Vet.Res., 34:27-8. Merkal RS, Larsen AB y Booth GD. 1975. Analysis of the effects of inapparent bovine paratuberculosis. Am.J.Vet.Res., 36(6):837-
8. Merkal RS, Miller JM, Hintz AM y Bryner JH. 1982b. Intrauterine inoculation of Mycobacterium paratuberculosis into guinea pigs
and cattle. Am.J.Vet.Res., 43(4):676-8. Merkal RS y Richards WD. 1972. Inhibition of fungal growth in the cultural isolation of mycobacteria. Appl.Microbiol., 24:205-7. Merkal RS y Whipple DL. 1982c. Effectiveness of disinfectants on Mycobacterium paratuberculosis. Proc.86th
Annu.Meet.USAHA, pp. 514-8. Merkal RS, Whipple DL, Sacks JM y Snyder GR. 1987. Prevalence of Mycobacterium paratuberculosis in ileocecal lymph nodes of
cattle culled in the United States. J.Am.Vet.Med.Assoc., 190:676-80. Mertens B y Knorr D. 1992. Developments of nonthermal processes for food preservation. Food Technol., 46(5):124-33.

BIBLIOGRAFÍA CITADA
216
Metzger-Boddien C, Khaschabi D, Schonbauer M, Boddien S, Schlederer T y Kehle J. 2006. Automated high-throughput immunomagnetic separation-PCR for detection of Mycobacterium avium subsp. paratuberculosis in bovine milk. Int.J.Food Microbiol., 110(3):201-8.
Meylan M, Nicolet J, Busato A, Burnens A y Martig J. 1995. [The prevalence of paratuberculosis in the Plateau de Diesse region]. Schweiz.Arch.Tierheilkd., 137(1):22-5.
Meylan M, Nicolet J, Oppliger A, Burnens A y Martig J. 1994. Evaluation de deux techniques d'ELISA pour le diagnostic de la paratuberculose bovine. Schweiz.Arch.Tierheilkd., 136:377-81.
Meylan M, Rings M, Shulaw WP, Kowalski JJ, Bech-Nielsen S y Hoffis GF. 1996. Survival of Mycobacterium paratuberculosis and preservation of inmunoglobulin G in bovine calostrum under experimental conditions simulating pasteurization. Am.J.Vet.Res., 57(11):1580-5.
Michel AL y Bastianello SS. 2000. Paratuberculosis in sheep: an emerging disease in South Africa. Vet.Microbiol., 77(3-4):299-307.
Mijs W, de Haas P, Rossau R, Van der LT, Rigouts L, Portaels F y van Soolingen D. 2002. Molecular evidence to support a proposal to reserve the designation Mycobacterium avium subsp. avium for bird-type isolates and 'M. avium subsp. hominissuis' for the human/porcine type of M. avium. Int.J.Syst.Evol.Microbiol., 52(Pt 5):1505-18.
Millar D, Sanderson J, Withey S, Tizard M, Doran T y Hermon-Taylor J. 1996. IS900 PCR to detect Mycobacterium paratuberculosis in retail supplies of whole pasteurized cows' milk in England and Wales. Appl.Environ.Microbiol., 62(9):3446-52.
Millar DS, Withey S.J., Tizard MLV, Ford J y Hermon-Taylor J. 1995. Solid-phase hybridization capture of low-abundance target DNA sequences: application to the polymerase chain reaction detection of Mycobacterium paratuberculosis and Mycobacterium avium subsp. silvaticum. Anal.Biochem., 226:325-30.
Miller DS, Collins MT, Smith BB, Anderson PR, Kramsky J, Wilder J y Hope A. 2000. Specificity of four serologic assays for Mycobacterium avium ss paratuberculosis in llamas and alpacas: a single herd study. J.Vet.Diagn.Invest., 12:345-53.
Miller J, Jenny A, Rhyan J y Saari DSD. 1997. Detection of Mycobacterium bovis in formalin-fixed, paraffin-embedded tissues of cattle and elk by amplification of an IS6110 sequence specific for Mycobacterium tuberculosis complex in formalin-fixed. J.Vet.Diagn.Invest., 9:244-9.
Miller JM, Jenny A y Ellingson JL. 1999. Polymerase chain reaction identification of Mycobacterium avium in formalin-fixed, paraffin-embedded animal tissues. J.Vet.Diagn.Invest., 11:436-40.
Miller JM, Jenny AL y Payeur JB. 2002. Polymerase chain reaction detection of Mycobacterium tuberculosis complex and Mycobacterium avium organisms in formalin-fixed tissues from culture-negative ruminants. Vet.Microbiol., 87(1):15-23.
Milner AR, Lepper AWD, Symonds WN y Gruner E. 1987. Analysis by ELISA and Western blotting of antibody reactivities in cattle infected with Mycobacterium paratuberculosis after absorption of serum with M. phlei. Res.Vet.Sci., 42:140-4.
Milner AR, Mack WN, Coates KJ, Hill J, Gill IJ y Sheldrick P. 1990. The sensitivity and specificity of a modified ELISA for the diagnosis of Johne's disease from a field trial in cattle. Vet.Microbiol., 25:193-8.
Minnikin DE. 1982. Lipids: Complex Lipids, Their Chemistry, Biosynthesis and Roles. En: Ratledge, C y Stanford, J. editores. Physiology, Identification and Classification. The Biology of the Mycobacteria. pp. 95-184. Academic Press. London, UK.
Mishina D, Katsel P, Brown ST, Gilberts EC y Greenstein RJ. 1996. On the etiology of Crohn disease. Proc.Natl.Acad.Sci.U.S.A., 93(18):9816-20.
Mobius P, Lentzsch P, Moser I, Naumann L, Martin G y Kohler H. 2006. Comparative macrorestriction and RFLP analysis of Mycobacterium avium subsp. avium and Mycobacterium avium subsp. hominissuis isolates from man, pig, and cattle. Vet.Microbiol., 117(2-4):284-91.
Mokresh AH y Butler DG. 1990. Granulomatous enteritis following oral inoculation of newborn rabbits with Mycobacterium paratuberculosis of bovine origin. Can.J.Vet.Res., 54:313-9.
Molina Caballero JM, Anguiano A, Serrano E y Uceda A. 1993. Use of an ELISA for serodiagnosis of clinical paratuberculosis in goats. Study by western-blotting of false-positive reactions. Rev.- Off.Int.Epizoot., 12(2):629-38.
Molina A, Morera L y Llanes D. 1991. Enzyme-linked immunosorbent assay for detection of antibodies against Mycobacterium paratuberculosis in goats. Am.J.Vet.Res., 52(6):863-8.
Momotani E, Whipple DL, Thiermann AB y Cheville NF. 1988. Role of M cells and macrophagos in the entrance of Mycobacterium paratuberculosis into domes of ileal Peyer's patches in calves. Vet.Pathol., 25:131-7.
Moreira AR, Paolicchi F, Morsella C, Zumarraga M, Cataldi A, Fabiana B, Alicia A, Piet O, van Soolingen D y Isabel RM. 1999. Distribution of IS900 restriction fragment polymorphism types among animal Mycobacterium avium subsp. pratuberculosis isolates from Argentina and Europe. Vet.Microbiol., 70(3-4):251-9.
Morgan SM, Ross RP, Beresford T y Hill C. 2000. Combination of hydrostatic pressure and lacticin 3147 causes increased killing of Staphylococcus and Listeria. J.Appl.Microbiol., 88(3):414-20.
Morrison NE. 1965. Circumvention of the mycobactin requirement of Mycobacterium paratuberculosis. J.Bacteriol., 89:762-9. Mortensen H, Nielsen SS y Berg P. 2004. Genetic variation and heritability of the antibody response to Mycobacterium avium
subspecies paratuberculosis in Danish Holstein cows. J.Dairy Sci., 87(7):2108-13. Mosmann TR, Cherwinski H, Bond MW, Giedlin MA y Coffman RL. 1986. Two types of murine helper T cell clone. I. Definition
according to profiles of lymphokine activities and secreted proteins. J.Immunol., 136(7):2348-57. Moss MT, Green EP, Tizard MLV, Malik ZP y Hermon-Taylor J. 1991. Specific detection of Mycobacterium paratuberculosis by
DNA hybridisation with a fragment of the insertion element IS900. Gut, 32(4):395-8. Moss MT, Malik ZP, Tizard MLV, Green EP, Sanderson JD y Hermon-Taylor J. 1992a. IS902, an insertion element of the chronic-
enteritis-causing Mycobacterium avium subsp.silvaticum. J.Gen.Microbiol., 138:139-45. Moss MT, Sanderson JD, Tizard MLV, Hermon-Taylor J, El-Zaatari FAK, Markesich DC y Graham DY. 1992b. Polymerase chain
reaction detection of Mycobacterium paratuberculosis and Mycobacterium avium subsp. silvaticum in long term cultures from Crohn's disease and control tissues. Gut, 33(9):1209-13.
Motiwala AS, Amonsin A, Strother M, Manning EJ, Kapur V y Sreevatsan S. 2004. Molecular epidemiology of Mycobacterium avium subsp. paratuberculosis isolates recovered from wild animal species. J.Clin.Microbiol., 42(4):1703-12.
Motiwala AS, Strother M, Amonsin A, Byrum B, Naser SA, Stabel JR, Shulaw WP, Bannantine JP, Kapur V y Sreevatsan S. 2003. Molecular epidemiology of Mycobacterium avium subsp. paratuberculosis: evidence for limited strain diversity, strain sharing, and identification of unique targets for diagnosis. J.Clin.Microbiol., 41(5):2015-26.
Motiwala AS, Strother M, Theus NE, Stich RW, Byrum B, Shulaw WP, Kapur V y Sreevatsan S. 2005. Rapid detection and typing of strains of Mycobacterium avium subsp. paratuberculosis from broth cultures. J.Clin.Microbiol., 43(5):2111-7.
Moyle AI. 1975. Culture and cull procedure for control of paratuberculosis. J.Am.Vet.Med.Assoc., 166(7):689-90. Muehlherr JE, Zweifel C, Corti S, Blanco JE y Stephan R. 2003. Microbiological quality of raw goat's and ewe's bulk-tank milk in
Switzerland. J.Dairy Sci., 86(12):3849-56.

BIBLIOGRAFÍA CITADA
217
Mullerad J, Hovav AH, Nahary R, Fishman Y y Bercovier H. 2003. Immunogenicity of a 16.7 kDa Mycobacterium paratuberculosis antigen. Microb.Pathog., 34(2):81-90.
Mullerad J, Michal I, Fishman Y, Hovav AH, Barletta RG y Bercovier H. 2002. The immunogenicity of Mycobacterium paratuberculosis 85B antigen. Med.Microbiol.Immunol.(Berl.), 190(4):179-87.
Mullis K, Faloona F, Scharf S, Saiki R, Horn G y Erlich H. 1986. Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harb.Symp.Quant.Biol., 51(Pt 1):263-73.
Munjal SK, Boehmer J, Beyerbach M, Strutzberg-Minder K y Homuth M. 2004. Evaluation of a LAM ELISA for diagnosis of paratuberculosis in sheep and goats. Vet.Microbiol., 103(1-2):107-14.
Muñoz M, García Marín JF, García-Pariente C, Reyes LE, Verna A, Moreno O, Fuertes M, Doce J, Puentes E, Garrido JM y Pérez V. 2006. Efficacy of a killed vaccine (SILIRUM®) in calves challenged with MAP. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 208-17. IAP, Madison, WI, USA.
Mura M, Bull TJ, Evans H, Sidi-Boumedine K, McMinn L, Rhodes G, Pickup R y Hermon-Taylor J. 2006. Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp. paratuberculosis within Acanthamoeba polyphaga. Appl.Environ.Microbiol., 72(1):854-9.
Murcia MI. 2004. Caracterización molecular de aislados clínicos pertenecientes al complejo Mycobacterium avium. Tesis Doctoral. Departamento de Medicina Preventiva, Salud Pública y Microbiología. Facultad de Medicina de la Universidad Autónoma de Madrid, Madrid, España.
Murcia MI, Leao SC, Ritacco V, Palenque E, de Oliveira RS, Reniero A, Menendez MC, Telles MA, Hadad DJ, Barrera L y Garcia MJ. 2004. Distribution of PRA patterns of clinical isolates of the Mycobacterium avium complex from Spain and South America. Biomedica, 24(Supp 1):60-4.
Murcia MI, Tortoli E, Menendez MC, Palenque E y Garcia MJ. 2006. Mycobacterium colombiense sp. nov., a novel member of the Mycobacterium avium complex and description of MAC-X as a new ITS genetic variant. Int.J.Syst.Evol.Microbiol., 56(Pt 9):2049-54.
Murray A, Oliaro J, Schlup MMT y Chadwick VS. 1995. Mycobacterium paratuberculosis and inflamatory bowel disease: frequency distribution in serial colonoscopic biopsies using the polymerase chain reaction. Microbios, 83:217-28.
Muskens J, Barkema HW, Russchen E, van Maanen K, Schukken YH y Bakker D. 2000. Prevalence and regional distribution of paratuberculosis in dairy herds in The Netherlands. Vet.Microbiol., 77(3-4):253-61.
Muskens J, Elbers AR, van Weering HJ y Noordhuizen JP. 2003. Herd management practices associated with paratuberculosis seroprevalence in Dutch dairy herds. J.Vet.Med.B Infect.Dis.Vet.Public Health, 50(8):372-7.
Mussa DM y Ramaswamy HS. 1997. Ultra high pressure pasteurization of milk: Kinetics of microbial destruction and changes in physico-chemical characteristics. Lebensm.Wiss.Technol., 30(6):551-7.
Mutwiri GK, Kosecka U, Benjamin M, Rosendal S, Perdue M y Butler DG. 2001. Mycobacterium avium subspecies paratuberculosis triggers intestinal pathophysiologic changes in beige/scid mice. Comp.Med., 51(6):538-44.
Mutwiri GK, Rosendal S, Kosecka U, Yager JA, Perdue M, Snider D y Butler DG. 2002. Adoptive transfer of BALb/c mouse splenocytes reduces lesion severity and induces intestinal pathophysiologic changes in the Mycobacterium avium subspecies paratuberculosis beige/scid mouse model. Comp.Med., 52(4):332-41.
Nagata R, Muneta Y y Mori Y. 2006. PPE proteins of Mycobacterium avium subspecies paratuberculosis induce interferon-gamma production by peripheral blood mononuclear cells from infected cattle. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 158. IAP, Madison, WI, USA.
Nakase H, Nishio A, Tamaki H, Matsuura M, Asada M, Chiba T y Okazaki K. 2006. Specific antibodies against recombinant protein of insertion element 900 of Mycobacterium avium subspecies paratuberculosis in Japanese patients with Crohn's disease. Inflamm.Bowel Dis., 12(1):62-9.
Naser SA, Felix J, Liping H, Romero C, Naser N, Walsh A y Safranek W. 1999. Occurrence of the IS900 gene in Mycobacterium avium complex derived from HIV patients. Mol.Cell.Probes, 13(5):367-72.
Naser SA, Ghobrial G, Romero C y Valentine JF. 2004. Culture of Mycobacterium avium subspecies paratuberculosis from the blood of patients with Crohn's disease. Lancet, 364(9439):1039-44.
Naser SA, Hulten K, Shafran I, Graham DY y El Zaatari FA. 2000a. Specific seroreactivity of Crohn's disease patients against p35 and p36 antigens of M. avium subsp. paratuberculosis. Vet.Microbiol., 77(3-4):497-504.
Naser SA, Schwartz D y Shafran I. 2000b. Isolation of Mycobacterium avium subsp. paratuberculosis from breast milk of Crohn's disease patients. Am.J.Gastroenterol., 95(4):1094-5.
Naser SA, Shafran I, Schwartz D, El Zaatari F y Biggerstaff J. 2002. In situ identification of mycobacteria in Crohn's disease patient tissue using confocal scanning laser microscopy. Mol.Cell.Probes, 16(1):41-8.
Navarro JA, Bernabé A, Gómez MA, Sánchez J y Gómez S. 1991. Mycobacterial antigen detection by immunohistochemistry in goat paratuberculosis. J.Vet.Med.B Infect.Dis.Vet.Public Health, 38:231-7.
Nebbia P, Robino P, Ferroglio E, Rossi L, Meneguz G y Rosati S. 2000. Paratuberculosis in red deer (Cervus elaphus hippelaphus) in the western Alps. Vet.Res.Commun., 24(7):435-43.
Newsholme SJ y Pletcher JM. 1981. A mycobateriosis in a sheep resembling paratuberculosis (Johne's disease). J.S.Afr.Vet.Assoc., June:143-4.
Nielsen SS y Agger JF. 2000a. Prevalence of paratuberculosis in Danish dairy herds. Proceedings of the 9th Symposium of the International Society of Veterinary Epidemiology and Economics, pp. 267-9.
Nielsen SS, Enevoldsen C y Grohn YT. 2002a. The Mycobacterium avium subsp. paratuberculosis ELISA response by parity and stage of lactation. Prev.Vet.Med., 54(1):1-10.
Nielsen SS, Grohn YT y Enevoldsen C. 2002b. Variation of the milk antibody response to paratuberculosis in naturally infected dairy cows. J.Dairy Sci., 85(11):2795-802.
Nielsen SS, Gronbaek C, Agger JF y Houe H. 2002c. Maximum-likelihood estimation of sensitivity and specificity of ELISAs and faecal culture for diagnosis of paratuberculosis. Prev.Vet.Med., 53(3):191-204.
Nielsen SS, Houe H, Thamsborg SM y Bitsch V. 2001. Comparison of two enzyme-linked immunosorbent assays for serologic diagnosis of paratuberculosis (Johne's disease) in cattle using different subspecies strains of Mycobacterium avium. J.Vet.Diagn.Invest., 13(2):164-6.
Nielsen SS, Kolmos B y Christoffersen AB. 2004. Comparison of contamination and growth of Mycobacterium avium subsp. paratuberculosis on two different media. J.Appl.Microbiol., 96(1):149-53.
Nielsen SS, Thamsborg SM, Houe H y Bitsch V. 2000b. Bulk-tank milk ELISA antibodies for estimating the prevalence of paratuberculosis in Danish dairy herds. Prev.Vet.Med., 44:1-7.
Nielsen SS y Toft N. 2003. Optimisation of the validity of ELISA and fecal culture tests for paratuberculosis: Selection of population or correction by population characteristics? En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 400-5. IAP, Madison, WI, USA.

BIBLIOGRAFÍA CITADA
218
Nielsen SS y Toft N. 2006. Age-specific characteristics of ELISA and fecal culture for purpose-specific testing for paratuberculosis. J.Dairy Sci., 89(2):569-79.
Nisbet DI, Gilmour NJL y Brotherston JG. 1962. Quantitative studies on Mycobacterium johnei in tissue of sheep. III intestinal histopathology. J.Comp.Pathol., 72:80-91.
Niv Y, Abuksis G y Fraser GM. 1999. Epidemiology of Crohn's disease in Israel: a survey of Israeli kibbutz settlements. Am.J.Gastroenterol., 94(10):2961-5.
Nordlund KV, Goodger WJ, Pelletier J y Collins MT. 1996. Associations between subclinical paratuberculosis and milk production, milk components, and somatic cell counts in dairy herds. J.Am.Vet.Med.Assoc., 208(11):1872-6.
Novi C, Rindi L, Lari N y Garzelli C. 2000. Molecular typing of Mycobacterium avium isolates by sequencing of the 16S-23S rDNA internal transcribed spacer and comparison with IS1245-based fingerprinting. J.Med.Microbiol., 49(12):1091-5.
O'Doherty A, O'Grady D, O'Farrell K, Smith T y Egan J. 2002a. Survey of Johne's disease in imported animals in the Republic of Ireland. Vet.Rec., 150(20):634-6.
O'Doherty A, O'Grady D, Smith T y Egan J. 2002b. Mycobacterium avium subsp. paratuberculosis in pasteurised and unpasteurised milk in the Republic of Ireland. Irish J.Agr.Food Res., 41(1):117-21.
O'Mahony J y Hill C. 2004. Rapid real-time PCR assay for detection and quantitation of Mycobacterium avium subsp. paratuberculosis DNA in artificially contaminated milk. Appl.Environ.Microbiol., 70(8):4561-8.
O'Reilly CE, Kelly AL, Murphy PM y Beresford TP. 2001. High Pressure treatment: application in cheese manufacture and ripening. Trends Food Sci.Technol., 12:51-9.
O'Reilly CE, O'Connor L, Anderson W, Harvey P, Grant IR, Donaghy J, Rowe M y O'Mahony P. 2004. Surveillance of bulk raw and commercially pasteurized cows' milk from approved Irish liquid-milk pasteurization plants to determine the incidence of Mycobacterium paratuberculosis. Appl.Environ.Microbiol., 70(9):5138-44.
Obasanjo IO, Gröhn YT y Hussini MO. 1997. Farm factors associated with the presence of Mycobacterium paratuberculosis infection in dairy herds on the New York State Paratuberculosis Control Program. Prev.Vet.Med., 32:243-51.
Ocepek M, Krt B y Pogacnik M. 2003. Seroprevalence of paratuberculosis in bulls' mothers herds in Slovenia. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 337-9. IAP, Madison, WI, USA.
Ocepek M, Posedi J y Pislak M. 1998. Prevalence of bovine paratuberculosis in Slovenia in 1997 and 1998. Zbornik Veterinarske Fakultete Univerza Ljubljana, 36(1):111-9.
Odumeru J, Gao A, Chen S, Raymond M y Mutharia L. 2001. Use of the bead beater for preparation of Mycobacterium paratuberculosis template DNA in milk. Can.J.Vet.Res., 65:201-5.
Olaison G, Sjodahl R y Tagesson C. 1990. Glucocorticoid treatment in ileal Crohn's disease: relief of symptoms but not of endoscopically viewed inflammation. Gut, 31(3):325-8.
Olcott BM, Snider TG, Turnquist SE, Kreeger JM y Miller JE. 1992. Paratuberculosis in the state of Louisiana: Prevalence and distribution. En: Chiodini y Kreeger editores. Proc Int Coll PTBC, (ed. III), vol.III, pp. 390-401. IAP, Providence, RI, USA.
Oliveira RS, Sircili MP, Oliveira EM, Balian SC, Ferreira-Neto JS y Leao SC. 2003. Identification of Mycobacterium avium genotypes with distinctive traits by combination of IS1245-based restriction fragment length polymorphism and restriction analysis of hsp65. J.Clin.Microbiol., 41(1):44-9.
Oliveira RS, Sircili MP, Ueki SY, Telles MA, Schnabel B, Briones MR y Leao SC. 2000. PCR-restriction enzyme analysis of a bone marrow isolate from a human immunodeficiency virus-positive patient discloses polyclonal infection with two Mycobacterium avium strains. J.Clin.Microbiol., 38(12):4643-5.
Olsen I, Boysen P, Kulberg S, Hope JC, Jungersen G y Storset AK. 2005. Bovine NK cells can produce gamma interferon in response to the secreted mycobacterial proteins ESAT-6 and MPP14 but not in response to MPB70. Infect.Immun., 73(9):5628-35.
Olsen I, Johansen TB, Billman-Jacobe H, Nilsen SF y Djonne B. 2004. A novel IS element, ISMpa1, in Mycobacterium avium subsp. paratuberculosis. Vet.Microbiol., 98(3-4):297-306.
Olsen I, Reitan LJ, Holstad G y Wiker HG. 2000. Alkyl hydroperoxide reductases C and D are major antigens constitutively expressed by Mycobacterium avium subsp. paratuberculosis. Infect.Immun., 68(2):801-8.
Olsen I y Storset AK. 2001a. Innate IFN-gamma production in cattle in response to MPP14, a secreted protein from Mycobacterium avium subsp. paratuberculosis. Scand.J.Immunol., 54(3):306-13.
Olsen I, Tryland M, Wiker HG y Reitan LJ. 2001b. AhpC, AhpD, and a secreted 14-kilodalton antigen from Mycobacterium avium subsp. paratuberculosis distinguish between paratuberculosis and bovine tuberculosis in an enzyme-linked immunosorbent assay. Clin.Diagn.Lab.Immunol., 8(4):797-801.
Olsen I, Wiker HG, Johnson E, Langeggen H y Reitan LJ. 2001c. Elevated antibody responses in patients with Crohn's disease against a 14-kDa secreted protein purified from Mycobacterium avium subsp. paratuberculosis. Scand.J.Immunol., 53(2):198-203.
Olsen I, Wiker HG, Johnson E, Langeggen H y Reitan LJ. 2003. Elevated antibody responses in patients with Crohn's disease against MPP14, a 14 kDa secreted protein purified from Mycobacterium avium subsp. paratuberculosis. Acta Vet.Scand., 44(3-4):287.
Olsen JE, Jorgensen JB y Nansen P. 1985. On the reduction of Mycobacterium paratuberculosis in bovine slurry subjected to batch mesophilic or thermophilic anaerobic-digestion. Agricultural Wastes, 13(4):273-80.
Ostrowski M, Mundo SL, Harris NB, Barletta RG y Lopez OJ. 2003. B-cell epitopes in the immunodominant p34 antigen of Mycobacterium avium ssp. paratuberculosis recognized by antibodies from infected cattle. Scand.J.Immunol., 58(5):511-21.
Ott SL, Wells SJ y Wagner BA. 1999. Herd-level economic losses associated with Johne's disease on US dairy operations. Prev.Vet.Med., 40:179-92.
Overduin P, Schouls L, Roholl P, van der ZA, Mahmmod N, Herrewegh A y van Soolingen D. 2004. Use of multilocus variable-number tandem-repeat analysis for typing Mycobacterium avium subsp. paratuberculosis. J.Clin.Microbiol., 42(11):5022-8.
Ozbek A, Michel FC, Strother M, Motiwala AS, Byrum BR, Shulaw WP, Thornton CG y Sreevatsan S. 2003. Evaluation of two recovery methods for detection of Mycobacterium avium subsp. paratuberculosis by PCR: direct-dilution--centrifugation and C(18)-carboxypropylbetaine processing. FEMS Microbiol.Lett., 229(2):145-51.
Pak SI, Kim D y Salman M. 2003. Estimation of paratuberculosis prevalence in dairy cattle in a province of Korea using an enzyme-linked immunosorbent assay: application of Bayesian approach. J.Vet.Sci., 4(1):51-6.
Paling RW, Waghela S, Macowan KJ y Heath BR. 1988. The occurrence of infectious diseases in mixed farming of domesticated wild herbivores and livestock in Kenya. II. Bacterial diseases. J.Wildl.Dis., 24(2):308-16.
Paliwal OP, Kumar R y Somvanshi R. 1985. The inmune spectrum of Mycobacterium johnei infections in goats. Indian Vet.J., 62:743-7.
Paliwal OP, Rajya BS y Krishna SG. 1984. Comparative evaluation of diagnostic tests for paratuberculosis in goats. Ind.J.Anim.Sci., 54(7):657-9.

BIBLIOGRAFÍA CITADA
219
Palmer MV, Stoffregen WC, Carpenter JG y Stabel JR. 2005. Isolation of Mycobacterium avium subsp paratuberculosis (Map) from feral cats on a dairy farm with Map-infected cattle. J.Wildl.Dis., 41(3):629-35.
Paolicchi F, Cirone K, Marsella C, Gioffré A, Cataldi A y Romano M. 2006. Isolation of Mycobacterium avium subsp. paratuberculosis (MAP) from commercial pasteurised milk. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., IAP, Madison, WI, USA.
Paolicchi FA, Zumarraga MJ, Gioffre A, Zamorano P, Morsella C, Verna A, Cataldi A, Alito A y Romano M. 2003. Application of different methods for the diagnosis of paratuberculosis in a dairy cattle herd in Argentina. J.Vet.Med.B Infect.Dis.Vet.Public Health, 50(1):20-6.
Park YH, Yoo HS, Yoon JW, Yang SJ, An JS y Davis WC. 2000. Phenotypic and functional analysis of bovine gammadelta lymphocytes. J.Vet.Sci., 1(1):39-48.
Patterson DS y Allen WM. 1972. Chronic mycobacterial enteritis in ruminants as a model of Crohn's disease. Proc.R.Soc.Med., 65(11):998-1001.
Patterson JP, LaVenture M, Hurley SS y Davis JP. 1988. Accidental self-inoculation with Mycobacterium paratuberculosis bacterin (Johne's bacterin) by veterinarians in Wisconsin. J.Am.Vet.Med.Assoc., 192(9):1197-9.
Paustian ML, Amonsin A, Kapur V y Bannantine JP. 2004. Characterization of novel coding sequences specific to Mycobacterium avium subsp. paratuberculosis: implications for diagnosis of Johne's Disease. J.Clin.Microbiol., 42(6):2675-81.
Paustian ML, Kapur V y Bannantine JP. 2005. Comparative genomic hybridizations reveal genetic regions within the Mycobacterium avium complex that are divergent from Mycobacterium avium subsp. paratuberculosis isolates. J.Bacteriol., 187(7):2406-15.
Pavlas M. 1990. Biologic characteristics of mycobacterium strains isolated from cattle from herds with clinical paratuberculosis. Vet.Med.(Praha), 35(9):523-8.
Pavlik I, Bartl J, Dvorska L, Svastova P, du MR, Machackova M, Yayo AW y Horvathova A. 2000a. Epidemiology of paratuberculosis in wild ruminants studied by restriction fragment length polymorphism in the Czech Republic during the period 1995-1998. Vet.Microbiol., 77(3-4):231-51.
Pavlik I, Bazant J, Vitasek J, Machackova M, Matlova L, Rozsypalova Z y Parmova I. 2000b. Paratuberculosis in imported cattle to the Czech Republic. Veterinarstvi, 51:159-63.
Pavlik I, Horvathova A, Dvorska L, Bartl J, Svastova P, du Maine R y Rychlik I. 1999a. Standardisation of restriction fragment length polymorphism analysis for Mycobacterium avium subspecies paratuberculosis. J.Microbiol.Methods, 38(1-2):155-67.
Pavlik I, Horvathova A, Dvorska L, Svastova P, du Maine R, Fixa B y Rychlik I. 1999b. Homogeneity/Heterogeneity of Mycobacterium avium subsp. paratuberculosis strains: Correlation between RFLP-type and source (Animal, Environment, Human). En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 321-30. IAP, Madison, WI, USA.
Pavlik I, Horvathova A, Dvorska L, Svastova P, du Maine R, Fixa B y Rychlik I. 1999c. Reliability of diagnostic methods (clinical examination, faecal culture, skin and serological test) for paratuberculosis of cattle and sheep during the 1988-1998 eradication and control programme. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 371-83. IAP, Madison, WI, USA.
Pavlik I, Matlova L, Bartl J, Svastova P, Dvorska L y Whitlock R. 2000c. Parallel faecal and organ Mycobacterium avium subsp. paratuberculosis culture of different productivity types of cattle. Vet.Microbiol., 77(3-4):309-24.
Pavlik I, Matlova L, Dvorska L, Shitaye JE y Parmova I. 2005. Mycobacterial infections in cattle and pigs caused by Mycobacterium avium complex members and atypical mycobacteria in the Czech Republic during 2000-2004. Vet.Med.(Praha), 50(7):281-90.
Pavlik I, Parmova I, Yayo Ayele W, Machackova M, Lamka J, Svoboda J, Pokorny J, Bazant J y Vitasek J. 2003. Paratuberculosis control programme in cattle in 1992-2001 by faecal culture and associated economic losses. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., p. 514. IAP, Madison, WI, USA.
Pavlik I, Pavlas BM y Kosková RS. 1995. Characterization by restriction endonuclease analysis and DNA hybridization using IS900 of bovine, ovine, caprine and human dependent strains of Mycobacterium paratuberculosis isolated in various localities. Vet.Microbiol., 45(4):311-8.
Pavlik I, Rozsypalova Z, Vesely T, Bartl J, Matlova L, Vrbas V, Valent L, Rajsky D, Mracko I, Hirko M y Miscovic P. 2000d. Control of paratuberculosis in five cattle farms by serological test and faecal culture during the period 1990-1999. Vet.Med.(Praha), 45:61-70.
Pavlik I, Svastova P, Bartl J, Dvorska L y Rychlik I. 2000e. Relationship between IS901 in the Mycobacterium avium Complex strains isolated from birds, animals, humans, and the environment and virulence for poultry. Clin.Diagn.Lab.Immunol., 7(2):212-7.
Payne JM y Rankin JD. 1961. The pathogenesis of experimental Johne's disease in calves. Res.Vet.Sci., 2:167-74. Pearce LE, Truong HT, Crawford RA, Yates GF, Cavaignac S y de Lisle GW. 2001. Effect of turbulent-flow pasteurization on
survival of Mycobacterium avium subsp. paratuberculosis added to raw milk. Appl.Environ.Microbiol., 67(9):3964-9. Pearson JK y McClelland TG. 1955. Uterine infection and congenital Johne's disease in cattle. Vet.Rec., 67:615-6. Pence M, Baldwin C y Black CC, III. 2003. The seroprevalence of Johne's disease in Georgia beef and dairy cull cattle.
J.Vet.Diagn.Invest., 15(5):475-7. Pérez V. 1992. Estudio de la paratuberculosis en la especie ovina. Tesis Doctoral. Universidad de Zaragoza, Aragón, España. Pérez V. 2006. Assessment of the efficacy of vaccination against paratuberculosis: the use of pathological methods. VENoMYC,
Use of vaccines in the control of tuberculosis and paratuberculosis, 10-11 de Noviembre. Laguardia, Euskadi, Spain. Pérez V, Bolea R, Chávez G, Cortabarría N, Juste RA, Badiola JJ y García Marín JF. 1995a. Efficiency of PCR and culture in the
detection of Mycobacterium avium subsp. silvaticum and Mycobacterium avium subsp. paratuberculosis in tissue samples of sheep. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 97-101. IAP, Rehoboth, MA, USA.
Pérez V, Chávez G, Gutiérrez M, Tellechea J, Badiola JJ y García Marín JF. 1995b. Evaluation of the response to AGID and Gamma-Interferon tests in lambs infected with Mycobacterium avium subsp. silvaticum and Mycobacterium avium subsp. paratuberculosis and their relation with the diagnosis of ovine paratuberculosis. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 91-6. IAP, Rehoboth, MA, USA.
Pérez V, Corpa JM y García Marín JF. 2000. El cuadro clínico y lesional de la paratuberculosis bovina. En: Juste editor. Paratuberculosis. Bovis, Aula Veterinaria, vol.nº93, pp. 39-47. Luzan 5, Madrid, España.
Pérez V, García Marín JF y Badiola JJ. 1996. Description and classification of different types of lesion associated with natural paratuberculosis infection in sheep. J.Comp.Pathol., 114(2):107-22.
Pérez V, García Marín JF, Bru R, Moreno B y Badiola JJ. 1995c. Resultados obtenidos en la vacunación de ovinos adultos frente a paratuberculosis. Med.Vet., 12(3):196-201.
Pérez V, Tellechea J, Badiola JJ, Gutiérrez J y García Marín JF. 1997. Relation between serologic response and pathologic findings in sheep with naturally acquired paratuberculosis. Am.J.Vet.Res., 58:799-803.

BIBLIOGRAFÍA CITADA
220
Pérez V, Tellechea J, Corpa JM, Gutiérrez M y García Marín JF. 1999. Relation between pathologic findings and cellular immune responses in sheep with naturally acquired paratuberculosis. Am.J.Vet.Res., 60(1):123-7.
Pérez V, Verna A, Muñoz M, García-Pariente C, Benavides J, Ferreras MC y García Marín JF. 2006. Immunohistochemical detection of TGF-b in paratuberculosis granulomatous lesions. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 122-6. IAP, Madison, WI, USA.
Peris B. 1992. Contribución al conocimiento de los aspectos inmunopatológicos de la paratuberculosis ovina mediante el estudio de la respuesta humoral y celular local. Tesis Doctoral. Universidad de Zaragoza, Aragón, España.
Petit E. 2001. Serological study of bovine paratuberculosis in Yonne in 1998/1999. REVUE EPIDEMIOLOGIE ET SANTE ANIMALE. EPIDEMIOLOGY AND ANIMAL HEALTH, vol.40, pp. 23-39. France.
Piaggio J, Nunez A y Gil A. 2003. Johne's disease serological prevalence in Uruguayan dairy cows. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 455-6. IAP, Madison, WI, USA.
Picardeau M y Vincent V. 1997. Characterization of large linear plasmids in mycobacteria. J.Bacteriol., 179(8):2753-6. Pickup RW, Rhodes G, Arnott S, Sidi-Boumedine K, Bull TJ, Weightman A, Hurley M y Hermon-Taylor J. 2005. Mycobacterium
avium subsp. paratuberculosis in the catchment area and water of the River Taff in South Wales, United Kingdom, and its potential relationship to clustering of Crohn's disease cases in the city of Cardiff. Appl.Environ.Microbiol., 71(4):2130-9.
Pickup RW, Rhodes G, Bull TJ, Arnott S, Sidi-Boumedine K, Hurley M y Hermon-Taylor J. 2006. Mycobacterium avium subsp. paratuberculosis in lake catchments, in river water abstracted for domestic use, and in effluent from domestic sewage treatment works: diverse opportunities for environmental cycling and human exposure. Appl.Environ.Microbiol., 72(6):4067-77.
Pieters J. 2001. Entry and survival of pathogenic mycobacteria in macrophages. Microbes.Infect, 3(3):249-55. Pillai SR y Jayarao BM. 2002. Application of IS900 PCR for detection of Mycobacterium avium subsp. paratuberculosis directly
from raw milk. J.Dairy Sci., 85(5):1052-7. Pillai SR, Jayarao BM, Gummo JD, Hue EC, Tiwari D, Stabel JR y Whitlock RH. 2001. Identification and sub-typing of
Mycobacterium avium subsp. paratuberculosis and Mycobacterium avium subsp. avium by randomly amplified polymorphic DNA. Vet.Microbiol., 79(3):275-84.
Pitcock JA y Gisler DB. 1961. Paratuberculosis (Johne's disease) in the monkey (Macaca mulatta). Tech.Doc.Rep.SAMTDR USAF Sch.Aerosp.Med., 61-86:1-4.
Plante Y, Remenda BW, Chelack BJ y Haines DM. 1996. Detection of Mycobacterium paratuberculosis in formalin-fixed paraffin-embedded tissues by the polymerase chain reaction. J.Vet.Diagn.Invest., 4:23-7.
Podolsky DK. 2002. Inflammatory bowel disease. N.Engl.J.Med., 347(6):417-29. Poupart P, Coene M, Van Heuverswyn H y Cocito C. 1993. Preparation of a specific RNA probe for detection of Mycobacterium
paratuberculosis and diagnosis of Johne's disease. J.Clin.Microbiol., 31(6):1601-5. Power SB, Haagsma J y Smyth Dp. 1993. Paratuberculosis in farmed red deer (Cervus elaphus) in Ireland. Vet.Rec., 132:213-6. Prudvi Reddy K, Sriraman PK, Gopal Naidu NR y Rama Rao P. 1984. Pathology of Johne's disease in sheep. Indian Vet.Med.J.,
61:179-84. Quadri LE, Sello J, Keating TA, Weinreb PH y Walsh CT. 1998. Identification of a Mycobacterium tuberculosis gene cluster
encoding the biosynthetic enzymes for assembly of the virulence-conferring siderophore mycobactin. Chem.Biol., 5(11):631-45.
Quist CF, Nettles VF, Manning EJ, Hall DG, Gaydos JK, Wilmers TJ y Lopez RR. 2002. Paratuberculosis in key deer (Odocoilecus virginianus clavium). J.Wildl.Dis., 38(4):729-37.
Raizman EA, Wells SJ, Godden SM, Bey RF, Oakes MJ, Bentley DC y Olsen KE. 2004. The distribution of Mycobacterium avium ssp. paratuberculosis in the environment surrounding Minnesota dairy farms. J.Dairy Sci., 87:2959-66.
Raizman EA, Wells SJ, Jordan PA, DelGiudice GD y Bey RR. 2005. Mycobacterium avium subsp. paratuberculosis from free-ranging deer and rabbits surrounding Minnesota dairy herds. Can.J.Vet.Res., 69(1):32-8.
Rajeev S, Zhang Y, Sreevatsan S, Motiwala AS y Byrum B. 2005. Evaluation of multiple genomic targets for identification and confirmation of Mycobacterium avium subsp. paratuberculosis isolates using real-time PCR. Vet.Microbiol., 105(3-4):215-21.
Rajya BS y Singh CM. 1961. Studies on the pathology of Johne's disease in sheep: III. Pathologic changes in sheep with naturally occurring infections. Am.J.Vet.Res., 22:189-203.
Rastogi N, Legrand E y Sola C. 2001. The mycobacteria: an introduction to nomenclature and pathogenesis. Rev.- Off.Int.Epizoot., 20(1):21-54.
Ratledge C. 1982. Nutrition, Growth and Metabolism. En: Ratledge, C. y Stanford, J. editores. Physiology, Identification and Classification. The Biology of the Mycobacteria. pp. 185-271. Academic Press. London.
Ratledge C. 2004. Iron, mycobacteria and tuberculosis. Tuberculosis, 84(1-2):110-30. Ratledge C y Ewing M. 1996. The occurrence of carboxymycobactin, the siderophore of pathogenic mycobacteria, as a second
extracellular siderophore in Mycobacterium smegmatis. Microbiology, 142(Pt 8):2207-12. Reddacliff L, Eppleston J, Windsor P, Whittington R y Jones S. 2006. Efficacy of a killed vaccine for the control of paratuberculosis
in Australian sheep flocks. Vet.Microbiol., 115(1-3):77-90. Reddacliff LA, Vadali A y Whittington RJ. 2003a. The effect of decontamination protocols on the numbers of sheep strain
Mycobacterium avium subsp. paratuberculosis isolated from tissues and faeces. Vet.Microbiol., 95(4):271-82. Reddacliff LA y Whittington RJ. 2003b. Experimental infection of weaner sheep with S strain Mycobacterium avium subsp.
paratuberculosis. Vet.Microbiol., 96(3):247-58. Reichel MP, Kittelberger R, Penrose M, Meynell RM, Cousins D, Ellis T, Mutharia LM, Sugden EA, Johns AH y de Lisle GW.
1999. Comparison of serological tests and faecal culture for the detection of Mycobacterium avium subsp. paratuberculosis infection in cattle and analysis of the antigens involved. Vet.Microbiol., 66:135-50.
Reisner BS, Gatson AM y Woods GL. 1994. Use of Gen-Probe AccuProbes to identify Mycobacterium avium complex, Mycobacterium tuberculosis complex, Mycobacterium kansaii, and Mycobacterium gordonae directly from BACTEC TB broth cultures. J.Clin.Microbiol., 33(12):2995-8.
Reviriego FJ, Moreno MA y Domínguez L. 2000. Soil type as a putative risk factor of ovine and caprine paratuberculosis seropositivity in Spain. Prev.Vet.Med., 43:43-51.
Reyes LE, González J, Ferreras MC, García-Pariente C, Benavides J, Fuertes M, García Marín JF y Pérez V. 2003. Evaluation of different adjuvants in the vaccination against paratuberculosis in sheep. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., p. 526. IAP, Madison, WI, USA.
Richards WD, Harris SK y Jarnagin JL. 1983. The isolation of Mycobacterium paratuberculosis from animals in the United States: A 16-year summary. En: Merkal editor. Proc.1st Int.Coll.Paratub., pp. 30-41. NADC, USDA, Ames, IA, USA.
Richards WD y Thoen CO. 1977. Effect of freezing on the viability of Mycobacterium paratuberculosis in bovine feces. J.Clin.Microbiol., 68:253-7.

BIBLIOGRAFÍA CITADA
221
Ridge SE, Harkin JT, Badman RT, Mellor AM y Larsen JW. 1995a. Johne's disease in alpacas (Lama pacos) in Australia. Aust.Vet.J., 72(4):150-3.
Ridge SE, Hope A y Condron RJ. 1995b. Johne's disease control in 25 victorian dairy herds 1990-1993. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 111-24. IAP, Rehoboth, MA, USA.
Ridge SE, Morgan IR, Condron RJ, Sockett DC y Collins MT. 1992. Comparison of the Johne's absorbed EIA and the complement-fixation test for the diagnosis of Johne's disease in cattle. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 22-8. IAP, Providence, RI, USA.
Ridley DS. 1974. Histological classification and the immunological spectrum of leprosy. Bull.World Health Organ., 51:451-65. Riemann H, Zaman MR, Ruppanner R, Aalund O, Jorgensen JB, Worsaae H y Behymer D. 1979. Paratuberculosis in cattle and
free-living exotic deer. J.Am.Vet.Med.Assoc., 174(8):841-3. Riemann HP y Abbas B. 1983. Diagnosis and control of bovine paratuberculosis (Johne's disease). Adv.Vet.Sci.Comp.Med.,
27:481-506. Ringdal G. 1963. Johne's disease in pigs. Nord.Vet.Med., 15:217-38. Ringuet H, Akoua-Koffi C, Honore S, Varnerot A, Vincent V, Berche P, Gaillard JL y Pierre-Audigier C. 1999. hsp65 sequencing
for identification of rapidly growing mycobacteria. J.Clin.Microbiol., 37(3):852-7. Ritz M, Jugiau F, Rama F, Corcoux P, Semenou M y Federighi M. 2000. Inactivation of Listeria monocytogenes by high hydrostatic
pressure: effects and interaction of treatment variables studied by analysis of variance. Food Microbiology, 17:375-82. Robbe-Austerman S, Gardner IA, Thomsen BV, Morrical DG, Martin BM, Palmer MV, Thoen CO y Ewing C. 2006a. Sensitivity
and specificity of the agar-gel-immunodiffusion test, ELISA and the skin test for detection of paratuberculosis in United States Midwest sheep populations. Vet.Res., 37(4):553-64.
Robbe-Austerman S, Krull AC y Stabel JR. 2006b. Time delay, temperature effects and assessment of positive controls on whole blood for the gamma interferon ELISA to detect paratuberculosis. J.Vet.Med.B Infect.Dis.Vet.Public Health, 53(5):213-7.
Robbe-Austerman S, Stabel JR y Palmer MV. 2006c. Evaluation of the gamma interferon ELISA in sheep subclinically infected with Mycobacterium avium subspecies paratuberculosis using a whole-cell sonicate or a johnin purified-protein derivative. J.Vet.Diagn.Invest., 18(2):189-94.
Robbi C, Rossi I, Nardelli S, Marangon S, Vincenzi G y Vicenzoni G. 2003. Prevalence of paratuberculosis (Johne's disease) in dairy farms in northeastern Italy. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 457-60. IAP, Madison, WI, USA.
Roca Soler R. 1935. Paratuberculosis en una vaca. Rev.Hig.San.Pecu., 25:839. Rodriguez E, Arques JL, Nunez M, Gaya P y Medina M. 2005. Combined effect of high-pressure treatments and bacteriocin-
producing lactic acid bacteria on inactivation of Escherichia coli O157:H7 in raw-milk cheese. Appl.Environ.Microbiol., 71(7):3399-404.
Rodríguez S. 2000. Desarrollo y evaluación de una prueba ELISA indirecta para el serodiagnóstico de paratuberculosis ovina. Tesina de Licenciatura. Universidad de Córdoba, Andalucía, España.
Rodriguez-Lazaro D, D'Agostino M, Herrewegh A, Pla M, Cook N y Ikonomopoulos J. 2005. Real-time PCR-based methods for detection of Mycobacterium avium subsp. paratuberculosis in water and milk. Int.J.Food Microbiol., 101(1):93-104.
Rodriguez-Lazaro D, Lloyd J, Herrewegh A, Ikonomopoulos J, D'Agostino M, Pla M y Cook N. 2004. A molecular beacon-based real-time NASBA assay for detection of Mycobacterium avium subsp. paratuberculosis in water and milk. FEMS Microbiol.Lett., 237(1):119-26.
Rogall T, Flohr T y Bottger EC. 1990. Differentiation of Mycobacterium species by direct sequencing of amplified DNA. J.Gen.Microbiol., 136(9):1915-20.
Rohde RF y Shulaw WP. 1990. Isolation of Mycobacterium paratuberculosis from the uterine flush fluids of cows with clinical paratuberculosis. J.Am.Vet.Med.Assoc., 197(11):1482-3.
Rohonczy EB, Balachandran AV, Dukes TW, Payeur JB, Rhyan JC, Saari DA, Whiting TL, Wilson SH y Jarnagin JL. 1996. A comparison of gross pathology, histopathology, and mycobacterial culture for the diagnosis of tuberculosis in elk (Cervus elaphus). Can.J.Vet.Res., 60(2):108-14.
Roiz MP, Palenque E, Guerrero C y García MJ. 1995. Use of restriction fragment length polymorphism as a genetic marker for typing Mycobacterium avium strains. J.Clin.Microbiol., 33(5):74-6.
Romero C, Hamdi A, Valentine JF y Naser SA. 2005. Evaluation of surgical tissue from patients with Crohn's disease for the presence of Mycobacterium avium subspecies paratuberculosis DNA by in situ hybridization and nested polymerase chain reaction. Inflamm.Bowel Dis., 11(2):116-25.
Rosseels V, Marche S, Roupie V, Govaerts M, Godfroid J, Walravens K y Huygen K. 2006. Members of the 30- to 32-kilodalton mycolyl transferase family (Ag85) from culture filtrate of Mycobacterium avium subsp. paratuberculosis are immunodominant Th1-type antigens recognized early upon infection in mice and cattle. Infect.Immun., 74(1):202-12.
Rossiter CA. 1992. A philosophy for successful Johne's disease control: experience from the New York State Paratuberculosis program. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 428-39. IAP, Providence, RI, USA.
Rossiter CA. 1996. Farm-specific approach to paratuberculosis (Johne's disease) control. Vet.Clin.North Am.Food Anim.Pract., 12(2):383-415.
Rossiter CA, Lein DH y Shin S. 1992. Objectives and current status of the NYS Paratuberculosis Erradication and Certification Program. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 448-54. IAP, Providence, RI, USA.
Roth A, Fischer M, Hamid ME, Michalke S, Ludwig W y Mauch H. 1998. Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences. J.Clin.Microbiol., 36(1):139-47.
Rothel JS, Jones SL, Corner LA, Cox JC y Wood PR. 1990. A sandwich enzyme immunoassay for bovine interferon-y and its use for the detection of tuberculosis in cattle. Aust.Vet.J., 67(4):134-7.
Roussel AJ, Libal MC, Whitlock RL, Hairgrove TB, Barling KS y Thompson JA. 2005. Prevalence of and risk factors for paratuberculosis in purebred beef cattle. J.Am.Vet.Med.Assoc., 226(5):773-8.
Rowan NJ, MacGregor SJ, Anderson JG, Cameron D y Farish O. 2001. Inactivation of Mycobacterium paratuberculosis by pulsed electric fields. Appl.Environ.Microbiol., 67(6):2833-6.
Rowbotham DS, Mapstone NP, Trejdosiewicz LK, Howdle PD y Quirke P. 1995. Mycobacterium paratuberculosis DNA not detected in Crohn's disease tissue by fluorescent polymerase chain reaction. Gut, 37:660-7.
Rubery E. 2001. A review of the evidence for a link between exposure to Mycobacterium paratuberculosis (Map) and Crohn's disease (CD) in humans. A report for the Food Standards Agency. pp. 1-55. Food Standards Agency, London, UK.
Runyon EH. 1959. Anonymous mycobacteria in pulmonary disease. Med.Clin.N.Am., 43:273-90. Ryan P, Bennett MW, Aarons S, Lee G, Collins JK, O'Sullivan GC, O'Connell J y Shanahan F. 2002. PCR detection of
Mycobacterium paratuberculosis in Crohn's disease granulomas isolated by laser capture microdissection. Gut, 51(5):665-70.

BIBLIOGRAFÍA CITADA
222
Ryan P, Kelly RG, Lee G, Collins JK, O'Sullivan GC, O'Connell J y Shanahan F. 2004. Bacterial DNA within granulomas of patients with Crohn's disease: detection by laser capture microdissection and PCR. Am.J.Gastroenterol., 99(8):1539-43.
Saint-Marc B, Argenté G, Guillemin F, Milward F, Lacoste F, Reynaud G, Brun A, Vandeputte J y B. 1992a. Efficiency of a live 316 F strain vaccine against paratuberculosis in ruminants. 17th World Buiatrics Congress, 1, pp. 96-101. St. Paul, MN, USA.
Saint-Marc B, Guillemin F, Milward F, Reynauld G, Lacoste F y Brun A. 1992b. Vaccination against paratuberculosis: new perspectives. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 469-74. IAP, Providence, RI, USA.
Salgado M, Manning EJ y Collins MT. 2005. Performance of a Johne's disease enzyme-linked immunosorbent assay adapted for milk samples from goats. J.Vet.Diagn.Invest., 17(4):350-4.
Sanderson JD, Moss MT, Tizard MLV y Hermon-Taylor J. 1992a. Mycobacterium paratuberculosis DNA in Crohn's disease tissue. Gut, 33(7):890-6.
Sanderson JD, Moss MT, Tizard MLV y Hermon-Taylor J. 1992b. PCR detection of Mycobacterium paratuberculosis in Crohn's disease tissue DNA extracts. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 201-7. IAP, Providence, RI, USA.
Sanna E, Woodall CJ, Watt NJ, Clarke CJ, Pittau M, Leoni A y Nieddu AM. 2000. In situ-PCR for the detection of Mycobacterium paratuberculosis DNA in paraffin-embedded tissues. Eur.J.Histochem., 44(2):179-84.
Santamaría C. 1989. Incidencia de paratuberculosis en el ovino de Navarra. En: Juste editor. 2ª Reu.PTBC España (1987), p. 46. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España.
Sasahara KC, Gray MJ, Shin SJ y Boor KJ. 2004. Detection of viable Mycobacterium avium subsp. paratuberculosis using luciferase reporter systems. Foodborne.Pathog.Dis, 1(4):258-66.
Saxegaard F. 1985. Isolation of Mycobacterium paratuberculosis from intestinal mucosa and mesenteric lymph nodes of goats by use of selective Dubos medium. J.Clin.Microbiol., 22(2):312-3.
Saxegaard F. 1990. Experimental infection of calves with an apparently specific goat-pathogenic strain of Mycobacterium paratuberculosis. J.Comp.Pathol., 102:149-56.
Saxegaard F y Baess I. 1988. Relationship between Mycobacterium avium, Mycobacterium paratuberculosis and "wood pigeon mycobacteria". APMIS, 96:37-42.
Schleig PM, Buergelt CD, Davis JK, Williams E, Monif GR y Davidson MK. 2005. Attachment of Mycobacterium avium subspecies paratuberculosis to bovine intestinal organ cultures: method development and strain differences. Vet.Microbiol., 108(3-4):271-9.
Schroen CJ, Kluver PF, Butler KL, McDonald WL, Hope AF y Condron RJ. 2003. Factors affecting the survival of Mycobacterium avium subsp. paratuberculosis in soil. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 10-5. IAP, Madison, WI, USA.
Schwartz D, Shafran I, Romero C, Piromalli C, Biggerstaff J, Naser N, Chamberlin W y Naser SA. 2000. Use of short-term culture for identification of Mycobacterium avium subsp. paratuberculosis in tissue from Crohn's disease patients. Clin.Microbiol.Infect., 6(6):303-7.
Scott PR, Clarke CJ y King TJ. 1995. Serum protein concentrations in clinical cases of ovine paratuberculosis (Johne's disease). Vet.Rec., 12:173.
Seaman JT, Gardner IA y Dent CHR. 1981. Johne's disease in sheep. Aust.Vet.J., 57(2):102-3. Seaman JT y Thomson DR. 1984. Johne's disease in sheep. Aust.Vet.J., 61:227-9. Sechi LA, Manuela M, Francesco T, Amelia L, Antonello S, Giovanni F y Stefania Z. 2001. Identification of Mycobacterium avium
subsp. paratuberculosis in biopsy specimens from patients with Crohn's disease identified by in situ hybridization. J.Clin.Microbiol., 39(12):4514-7.
Sechi LA, Mara L, Cappai P, Frothingam R, Ortu S, Leoni A, Ahmed N y Zanetti S. 2006. Immunization with DNA vaccines encoding different mycobacterial antigens elicits a Th1 type immune response in lambs and protects against Mycobacterium avium subspecies paratuberculosis infection. Vaccine, 24(3):229-35.
Sechi LA, Mura M, Tanda E, Lissia A, Fadda G y Zanetti S. 2004. Mycobacterium avium sub. paratuberculosis in tissue samples of Crohn's disease patients. New Microbiol., 27(1):75-7.
Sechi LA, Scanu AM, Molicotti P, Cannas S, Mura M, Dettori G, Fadda G y Zanetti S. 2005. Detection and isolation of Mycobacterium avium subspecies paratuberculosis from intestinal mucosal biopsies of patients with and without Crohn's disease in Sardinia. Am.J.Gastroenterol., 100(7):1529-36.
Secott TE, Lin TL y Wu CC. 2002. Fibronectin attachment protein is necessary for efficient attachment and invasion of epithelial cells by Mycobacterium avium subsp. paratuberculosis. Infect.Immun., 70(5):2670-5.
Secott TE, Lin TL y Wu CC. 2004. Mycobacterium avium subsp. paratuberculosis fibronectin attachment protein facilitates M-cell targeting and invasion through a fibronectin bridge with host integrins. Infect.Immun., 72(7):3724-32.
Secott TE, Ohme AM, Barton KS, Wu CC y Rommel FA. 1999. Mycobacterium paratuberculosis detection in bovine feces is improved by coupling agar culture enrichment to an IS900-specific polymerase chain reaction assay. J.Vet.Diagn.Invest., 11:441-7.
Seitz SE, Heider LE, Hueston WD, Bech-Nielsen S, Rings DM y Spangler L. 1989. Bovine fetal infection with Mycobacterium paratuberculosis. J.Am.Vet.Med.Assoc., 194:1423-6.
Seixas C, Esteves F, Albuquerque T, Afonso A, Botelho A, Santos C, Vala H y Amado A. 2006. Sheep paratuberculosis study in Portugal, Serra da Estrela Region (preliminary results). En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., IAP, Madison, WI, USA.
Selby WS. 2004. Mycobacterium avium subspecies paratuberculosis bacteraemia in patients with inflammatory bowel disease. Lancet, 364: 1013-4.
Semret M, Turenne CY y Behr MA. 2006. Insertion sequence IS900 revisited. J.Clin.Microbiol., 44(3):1081-3. Sergeant ES. 2001. Ovine Johne's disease in Australia: the first 20 years. Aust.Vet.J., 79(7):484-91. Sergeant ES. 2003. Estimated flock-prevalence and distribution of ovine Johne's disease in Australia at December 2001. Aust.Vet.J.,
81(12):768-9. Sergeant ES y Baldock FC. 2002a. The estimated prevalence of Johne's disease infected sheep flocks in Australia. Aust.Vet.J.,
80(12):762-8. Sergeant ES, Marshall DJ, Eamens GJ, Kearns C y Whittington RJ. 2003. Evaluation of an absorbed ELISA and an agar-gel
immuno-diffusion test for ovine paratuberculosis in sheep in Australia. Prev.Vet.Med., 61(4):235-48. Sergeant ES, Whittington RJ y More SJ. 2002b. Sensitivity and specificity of pooled faecal culture and serology as flock-screening
tests for detection of ovine paratuberculosis in Australia. Prev.Vet.Med., 52(3-4):199-211. Serraino A, Marchetti G, Sanguinetti V, Rossi MC, Zanoni RG, Catozzi L, Bandera A, Dini W, Mignone W, Franzetti F y Gori A.
1999. Monitoring of transmission of tuberculosis between wild boars and cattle: genotypical analysis of strains by molecular epidemiology techniques. J.Clin.Microbiol., 37(9):2766-71.

BIBLIOGRAFÍA CITADA
223
Seuna E y Seppanen J. 2003. National control strategies and ongoing research in Finland. Acta Vet.Scand., 2003(44):251-3. Sevilla I, Aduriz G, Garrido JM, Geijo MV y Juste RA. 2003. A preliminary survey on the prevalence of paratuberculosis in dairy
cattle in Spain by bulk milk PCR. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., pp. 332-6. IAP, Madison, WI, USA.
Sevilla I, Singh SV, Garrido JM, Aduriz G, Rodríguez S, Geijo MV, Whittington RJ, Saunders V, Whitlock RH y Juste RA. 2005. PCR-REA genotype of paratuberculosis strains isolated from different host species and geographic locations. Rev.- Off.Int.Epizoot., 24(3):1061-6.
Shafran I, Kugler L, El Zaatari FA, Naser SA y Sandoval J. 2002a. Open clinical trial of rifabutin and clarithromycin therapy in Crohn's disease. Dig.Liver Dis., 34(1):22-8.
Shafran I, Piromalli C, Decker JW, Sandoval J, Naser SA y El Zaatari FA. 2002b. Seroreactivities against Saccharomyces cerevisiae and Mycobacterium avium subsp. paratuberculosis p35 and p36 antigens in Crohn's disease patients. Dig.Dis.Sci., 47(9):2079-81.
Shanahan F. 2002. Crohn's disease. Lancet, 359(9300):62-9. Sherman DM. 1985. Current concepts in Johne's disease. Vet.Med., 80:77-84. Sherman DM, Markham RJF y Bates F. 1984. Agar-gel immunodiffusion test for diagnosis of clinical paratuberculosis in cattle.
J.Am.Vet.Med.Assoc., 185:179-82. Shin SJ, Wu CW, Steinberg H y Talaat AM. 2006. Identification of novel virulence determinants in Mycobacterium
paratuberculosis by screening a library of insertional mutants. Infect.Immun., 74(7):3825-33. Shin SJ, Yoo HS, McDonough SP y Chang YF. 2004. Comparative antibody response of five recombinant antigens in related to
bacterial shedding levels and development of serological diagnosis based on 35 kDa antigen for Mycobacterium avium subsp. paratuberculosis. J.Vet.Sci., 5(2):111-7.
Shinnick TM y Good RC. 1994. Mycobacterial taxonomy. Eur.J.Clin.Microbiol.Infect.Dis., 13(11):884-901. Shulaw WP, Bech-Nielsen S, Rings DM, Getzy DM y Woodruff TS. 1993. Serodiagnosis of paratuberculosis in sheep by use of
agar gel immunodiffusion. Am.J.Vet.Res., 54(1):1319. Shulaw WP, Gordon JC, Bech-Nielsen S, Pretzman CI y Hoffsis GF. 1986. Evidence of paratuberculosis in Ohio's white-tailed deer
as determined by an enzyme-linked immunosorbent assay. Am.J.Vet.Res., 47:2539-42. Sigur-Dardottir OG, Press CM y Evensen O. 2001. Uptake of Mycobacterium avium subsp. paratuberculosis through the distal
small intestinal mucosa in goats: an ultrastructural study. Vet.Pathol., 38(2):184-9. Sigurdardottir OG, Bakke-McKellep AM, Djonne B y Evensen O. 2005. Mycobacterium avium subsp. paratuberculosis enters the
small intestinal mucosa of goat kids in areas with and without Peyer's patches as demonstrated with the everted sleeve method. Comp.Immunol.Microbiol.Infect.Dis., 28(3):223-30.
Sigurdardottir OG, Press CM, Saxegaard F y Evensen O. 1999. Bacterial isolation, inmunological response, and histopathological lesions during the early subclinical phase of experimental infection of goat kids with Mycobacterium avium subsp. paratuberculosis. Vet.Pathol., 36:542-50.
Sigurdsson B. 1960. A killed vaccine against paratuberculosis (Johne's disease) in sheep. Am.J.Vet.Res., 21:54-67. Sigurdsson B y Tryggvadottir AG. 1949. Immunization with heat-killed Mycobacterium paratuberculosis in mineral oil.
J.Bacteriol., 58:271-8. Singh N, Singh SV, Gupta VK, Sharma VD, Sharma RK y Katoch VM. 1996. Isolation and identification of Mycobacterium
paratuberculosis from naturally infected goat herds in India. Ind J Vet Path, 20(2):104-8. Singh SP, Salamon H, Lahti CJ, Farid-Moyer M y Small PM. 1999. Use of pulsed-field gel electrophoresis for molecular
epidemiologic and population genetic studies of Mycobacterium tuberculosis. J.Clin.Microbiol., 37(6):1927-31. Singh SV, Shing AV, Singh R, Sandhu KS, Gupta VK y Misra S. 2006. Survey of ruminant population of Northern India for the
incidence of Mycobacterium avium subsp. paratuberculosis infection. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., IAP, Madison, WI, USA.
Sivakumar P, Tripathi BN y Singh N. 2005. Detection of Mycobacterium avium subsp. paratuberculosis in intestinal and lymph node tissues of water buffaloes (Bubalus bubalis) by PCR and bacterial culture. Vet.Microbiol., 108(3-4):263-70.
Slutsky AM, Arbeit RD, Barber TW, Rich J, Fordham C, Pieciak W, Barlow MA y Maslow JN. 1994. Polyclonal infections due to Mycobacterium avium complex in patients with AIDS detected by pulsed-field gel electrophoresis of sequential clinical isolates. J.Clin.Microbiol., 32(7):1773-8.
Smelt JPPM. 1998. Recent advances in the microbiology of high pressure processing. Trends Food Sci.Technol., 9(4):152-8. Smith HW. 1960. The examination of milk for the presence of Mycobacterium johnei. J.Pathol.Bacteriol., 80(2):440-2. Smith WL, Lagunas-Solar MC y Cullor JS. 2002. Use of pulsed ultraviolet laser light for the cold pasteurization of bovine milk.
J.Food Prot., 65(9):1480-2. Smith WL, McGarvey KL y Cullor JS. 2003. The use of spiral plating and microscopic colony counting for the rapid quantitation of
Mycobacterium paratuberculosis. Lett.Appl.Microbiol., 36(5):293-6. Smole SC, McAleese F, Ngampasutadol J, Von Reyn CF y Arbeit RD. 2002. Clinical and epidemiological correlates of genotypes
within the Mycobacterium avium complex defined by restriction and sequence analysis of hsp65. J.Clin.Microbiol., 40(9):3374-80.
Snow GA. 1965. The structure of Mycobactin P, a growth factors for Mycobacterium johnei and the significance of its iron complex. Biochem.J., 94:160-5.
Snow GA. 1970. Mycobactins: iron chelating growth factors from mycobacteria. Bacteriol.Rev., 34:99-125. Snow GA y Waite AJ. 1969. Chemical and biological properties of mycobactins isolated from various mycobacteria. Biochem.J.,
115:1031-45. Sockett DC, Conrad TA, Thomas CB y Collins MT. 1992. Evaluation of four serological tests for bovine paratuberculosis.
J.Clin.Microbiol., 30(2):1134-9. Soini H y Musser JM. 2001. Molecular diagnosis of mycobacteria. Clin.Chem., 47(5):809-14. Sorensen O, Rawluk S, Wu J, Manninen K y Ollis G. 2003. Mycobacterium paratuberculosis in dairy herds in Alberta. Can.Vet.J.,
44(3):221-6. Souza CD, Evanson OA y Weiss DJ. 2006a. Mitogen activated protein kinase p38 pathway is an important component of the anti-
inflammatory response in Mycobacterium avium subsp. paratuberculosis-infected bovine monocytes. Microb.Pathog., 41(2-3):59-66.
Souza CD, Evanson OA y Weiss DJ. 2006b. Regulation by Jun N-terminal kinase/stress activated protein kinase of cytokine expression in Mycobacterium avium subsp. paratuberculosis-infected bovine monocytes. Am.J.Vet.Res., 67(10):1760-5.
Spahr U y Schafroth K. 2001. Fate of Mycobacterium avium subsp. paratuberculosis in Swiss hard and semihard cheese manufactured from raw milk. Appl.Environ.Microbiol., 67(9):4199-205.

BIBLIOGRAFÍA CITADA
224
Spangler E, Bech-Nielsen S y Heider LE. 1992a. Diagnostic performance of two serologic tests and fecal culture for subclinical paratuberculosis, and associations with production. Prev.Vet.Med., 13:185-95.
Spangler E, Bech-Nielsen S, Heider LE y Dorn CR. 1992b. Interpretation of an enzyme immunosorbent test using different cut-offs between positive and negative samples for diagnosis of paratuberculosis. Prev.Vet.Med., 13:197-204.
Spears HN. 1959. Vaccination against Johne's disease: the results of a field trial experiment. Vet.Rec., 71(45):1154-6. Speer CA, Scott MC, Bannantine JP, Waters WR, Mori Y, Whitlock RH y Eda S. 2006. A novel enzyme-linked immunosorbent
assay for diagnosis of Mycobacterium avium subsp. paratuberculosis infections (Johne's Disease) in cattle. Clin.Vaccine Immunol., 13(5):535-40.
St Jean G. 1996. Treatmnet of clinical paratuberculosis in cattle. Vet.Clin.North Am.Food Anim.Pract., 12(2):417-30. St Jean G y Jernigan AD. 1991. Treatment of Mycobacterium paratuberculosis infection in ruminants. Vet.Clin.North Am.Food
Anim.Pract., 7(3):793-804. Stabel JR. 1995. Temporal effects of tumor necrosis factor-alpha on intracellular survival of Mycobacterium paratuberculosis.
Vet.Immunol.Immunopathol., 45(3-4):321-32. Stabel JR. 1996. Production of g-interferon by peripheral blood mononuclear cells: an important diagnostic tool for detection of
subclinical paratuberculosis. J.Vet.Diagn.Invest., 8:345-50. Stabel JR. 1997. An improved method for cultivation of Mycobacterium paratuberculosis from bovine fecal samples and
comparison to three other methods. J.Vet.Diagn.Invest., 9(4):375-80. Stabel JR. 2000a. Cytokine secretion by peripheral blood mononuclear cells from cows infected with Mycobacterium
paratuberculosis. Am.J.Vet.Res., 61(7):754-60. Stabel JR. 2000b. Johne's disease and milk: do consumers need to worry? J.Dairy Sci., 83(7):1659-63. Stabel JR. 2000c. Transitions in immune responses to Mycobacterium paratuberculosis. Vet.Microbiol., 77(3-4):465-73. Stabel JR. 2001. On-farm batch pasteurization destroys Mycobacterium paratuberculosis in waste milk. J.Dairy Sci., 84(2):524-7. Stabel JR y Ackermann MR. 2002a. Temporal Mycobacterium paratuberculosis infection in T-cell receptor (TCR)-alpha and TCR-
delta-deficient mice. Vet.Immunol.Immunopathol., 89(3-4):127. Stabel JR, Ackermann MR y Goff JP. 1996. Comparison of polyclonal antibodies to three different preparations of Mycobacterium
paratuberculosis in immunohistochemical diagnosis of Johne's disease in cattle. J.Vet.Diagn.Invest., 8:469-73. Stabel JR y Bannantine JP. 2005. Development of a nested PCR method targeting a unique multicopy element, ISMap02, for
detection of Mycobacterium avium subsp. paratuberculosis in fecal samples. J.Clin.Microbiol., 43(9):4744-50. Stabel JR, Bosworth TL, Kirkbride TA, Forde RL y Whitlock RH. 2004a. A simple, rapid, and effective method for the extraction
of Mycobacterium paratuberculosis DNA from fecal samples for polymerase chain reaction. J.Vet.Diagn.Invest., 16(1):22-30. Stabel JR y Lambertz A. 2004b. Efficacy of pasteurization conditions for the inactivation of Mycobacterium avium subsp.
paratuberculosis in milk. J.Food Prot., 67(12):2719-26. Stabel JR, Steadham EM y Bolin CA. 1997. Heat inactivation of Mycobacterium paratuberculosis in raw milk: are current
pasteurization conditions effective? Appl.Environ.Microbiol., 63(12):4975-7. Stabel JR, Waldren CA y Garry F. 2001a. Gamma radiation effectively destroys Mycobacterium paratuberculosis in milk.
J.Dairy Sci., 84(Suppl.1):27. Stabel JR, Wells SJ y Wagner BA. 2002b. Relationships between fecal culture, ELISA, and bulk tank milk test results for Johne's
disease in US dairy herds. J.Dairy Sci., 85(3):525-31. Stabel JR y Whitlock RH. 2001b. An evaluation of a modified interferon-gamma assay for the detection of paratuberculosis in dairy
herds. Vet.Immunol.Immunopathol., 79(1-2):69-81. Stackebrandt E, Rainey FA y Ward-Rainey NL. 1997. Proposal for a new hierarchic classification system, Actinobacteria Classis
nov. Int.J.Syst.Bacteriol., 47(2):479-91. Stahl DA y Urbance JW. 1990. The division between fast- and slow-growing species corresponds to natural relationships among the
mycobacteria. J.Bacteriol., 172(1):116-24. Stainsby KJ, Lowes JR, Allan RN y Ibbotson JP. 1993. Antibodies to Mycobacterium paratuberculosis and nine species of
environmental mycobacteria in Crohn's disease and control subjects. Gut, 34:371-4. Stamp JT y Watt JA. 1954. Johne's disease in sheep. J.Comp.Pathol., 64(1):26-38. Stark KD, Frei-Staheli C, Frei PP, Pfeiffer DU, Danuser J, Audige L, Nicolet J, Strasser M, Gottstein B y Kihm U. 1997.
[Frequency and cost of health problems in Swiss dairy cows and their calves (1993-1994)]. Schweiz.Arch.Tierheilkd., 139(8):343-53.
Stehman SM. 1996. Paratuberculosis in small ruminants, deer, and South American camelids. Vet.Clin.North Am.Food Anim.Pract., 12(2):441-55.
Steinberg H. 1988. Johne's disease (Mycobacterium paratuberculosis) in a Jimela Topi (Damaliscus lunatus jimela). J.Zoo Anim.Med., 19(1-2):33-41.
Stephan R, Buhler K y Corti S. 2002. Incidence of Mycobacterium avium subspecies paratuberculosis in bulk-tank milk samples from different regions in Switzerland. Vet.Rec., 150(7):214-5.
Stevenson K. 1997. Debate/Discussion. Nomenclature: M. paratuberculosis vs M. avium subsp paratuberculosis and strain definitions and differentiation. En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., pp. 96-101. IAP, Rehoboth, MA, USA.
Stevenson K. 2006. Studying the pathogenesis of paratuberculosis: the enduring challenge. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 88-97. IAP, Madison, WI, USA.
Stevenson K, Hughes VM, de Juan L, Inglis NF, Wright F y Sharp JM. 2002. Molecular characterization of pigmented and nonpigmented isolates of Mycobacterium avium subsp. paratuberculosis. J.Clin.Microbiol., 40(5):1798-804.
Stevenson K, Inglis NF, Rae B, Burrells C, Donachie W y Sharp JM. 1992. Identification and characterization of immunogenic proteins of Mycobacterium paratuberculosis. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 238-40. IAP, Providence, RI, USA.
Stevenson K, Inglis NF, Rae B, Donachie WD y Sharp JM. 1991. Complete nucleotide sequence of a gene encoding the 70 kDa heat shock protein of Mycobacterium paratuberculosis. Nucleic Acids Res., 19(16):4552.
Stevenson K y Sharp JM. 1997. The contribution of molecular biology to Mycobacterium avium subspecies paratuberculosis research. Vet.J., 153(3):269-86.
Stich RW, Byrum B, Love B, Theus N, Barber L y Shulaw WP. 2004. Evaluation of an automated system for non-radiometric detection of Mycobacterium avium paratuberculosis in bovine feces. J.Microbiol.Methods, 56(2):267-75.
Storset AK, Slettedal IO, Williams JL, Law A y Dissen E. 2003. Natural killer cell receptors in cattle: a bovine killer cell immunoglobulin-like receptor multigene family contains members with divergent signaling motifs. Eur.J.Immunol., 33(4):980-90.

BIBLIOGRAFÍA CITADA
225
Stratmann J, Dohmann K, Heinzmann J y Gerlach GF. 2006. Peptide aMptD-mediated capture PCR for detection of Mycobacterium avium subsp. paratuberculosis in bulk milk samples. Appl.Environ.Microbiol., 72(8):5150-8.
Stratmann J, Strommenger B, Stevenson K y Gerlach GF. 2002. Development of a peptide-mediated capture PCR for detection of Mycobacterium avium subsp. paratuberculosis in milk. J.Clin.Microbiol., 40(11):4244-50.
Streeter RN, Hoffis GF, Bech-Nielsen S, Shulaw WP y Rings M. 1995. Isolation of Mycobacterium paratuberculosis from calostrum and milk of subclinically infected cows. Am.J.Vet.Res., 56:1322-4.
Strommenger B, Stevenson K y Gerlach GF. 2001. Isolation and diagnostic potential of ISMav2, a novel insertion sequence-like element from Mycobacterium avium subspecies paratuberculosis. FEMS Microbiol.Lett., 196(1):31-7.
Stuart P. 1962. The diagnosis of Johne's disease in cattle and the effect of vaccination on tuberculin and johnin tests. Bull.Off.Int.Epizoot., 58:33-50.
Stuart P. 1965. Vaccination against Johne's disease in cattle exposed to experimental infection. Br.Vet.J., 121:289-318. Suenaga K, Yokoyama E, Nishimori I, Sano S, Morita M, Okazaki K y Onishi S. 1999. Serum antibodies to Mycobacterium
paratuberculosis in patients with Crohn's disease. Dig.Dis.Sci., 44(6):1202-7. Suenaga K, Yokoyama Y, Okazaki K y Yamamoto Y. 1995. Mycobacteria in the intestine of Japanese patients with inflammatory
bowel disease. Am.J.Gastroenterol., 90(1):76-80. Sugden EA, Brooks BW, Corner AH, Nielsen KH, Turcotte C, Shapiro J y Duncan JR. 1988. A study of indirect enzyme
immunoassay for ovine paratuberculosis using A and D protein antigens and lipoarabinomannan polysaccharide antigen. En: Thorel y Merkal editores. Proc.2nd Int.Coll.Paratub., pp. 231-5. LCRV, Maisons-Alfort, France.
Sugden EA, Brooks BW, Young NM, Watson DC, Nielsen KH y Corner AH. 1991. Chromatographic purification and characterization of antigens A and D from Mycobacterium paratuberculosis and their use in enzyme-linked immunosorbent assays for diagnosis of paratuberculosis in sheep. J.Clin.Microbiol., 29(8):1659-64.
Sugden EA, Corner AH, Samagh DS, Duncan JR, Brooks BW, Turcotte C, Nielsen KH y Stewart RD. 1989. Serodiagnosis of ovine paratuberculosis using lipoarabinomannan in an enzime-linked immunosorbent assay. Am.J.Vet.Res., 50:850-4.
Sung N y Collins MT. 1998. Thermal tolerance of Mycobacterium paratuberculosis. Appl.Environ.Microbiol., 64(3):999-1005. Sung N y Collins MT. 2000. Effect of three factors in cheese production (pH, salt, and heat) on Mycobacterium avium subsp.
paratuberculosis viability. Appl.Environ.Microbiol., 66(4):1334-9. Supply P, Allix C, Lesjean S, Cardoso-Oelemann M, Rusch-Gerdes S, Willery E, Savine E, de Haas P, van Deutekom H, Roring S,
Bifani P, Kurepina N, Kreiswirth B, Sola C, Rastogi N, Vatin V, Gutierrez MC, Fauville M, Niemann S, Skuce R, Kremer K, Locht C y van Soolingen D. 2006. Proposal for standardization of optimized Mycobacterial Interspersed Repetitive Unit-Variable Number Tandem Repeat typing of Mycobacterium tuberculosis. J.Clin.Microbiol., (in press).
Supply P, Magdalena J, Himpens S y Locht C. 1997. Identification of novel intergenic repetitive units in a mycobacterial two-component system operon. Mol.Microbiol., 26(5):991-1003.
Supply P, Mazars E, Lesjean S, Vincent V, Gicquel B y Locht C. 2000. Variable human minisatellite-like regions in the Mycobacterium tuberculosis genome. Mol.Microbiol., 36(3):762-71.
Surujballi OP, Romanowska A, Sugden EA, Turcotte C y Jolley ME. 2002. A fluorescence polarization assay for the detection of antibodies to Mycobacterium bovis in cattle sera. Vet.Microbiol., 87(2):149-57.
Swanson DS, Kapur V, Stockbauer K, Pan X, Frothingham R y Musser JM. 1997. Subspecific differentiation of Mycobacterium avium complex strains by automated sequencing of a region of the gene (hsp65) encoding a 65-kilodalton heat shock protein. Int.J.Syst.Bacteriol., 47(2):414-9.
Sweeney RW. 1996. Transmission of paratuberculosis. Vet.Clin.North Am.Food Anim.Pract., 12(2):305-12. Sweeney RW, Jones DE, Habecker P y Scott P. 1998. Interferon-gamma and interleukin 4 gene expression in cows infected with
Mycobacterium paratuberculosis . Am.J.Vet.Res., 59(7):842-7. Sweeney RW, Whitlock RH, Buckley CL y Spencer PA. 1995. Evaluation of a commercial enzyme-linked immunosorbent assay for
the diagnosis of paratuberculosis in dairy cattle. J.Vet.Diagn.Invest., 7(4):488-93. Sweeney RW, Whitlock RH, Buckley CL, Spencer PA, Rosenberger AE y Hutchinson LJ. 1994. Diagnosis of paratuberculosis in
dairy cattle, using enzyme linked immunosorbent assay for detection of antibodies against Mycobacterium paratuberculosis in milk. Am.J.Vet.Res., 55(7):905-9.
Sweeney RW, Whitlock RH y Rosenberger AA. 1992a. Mycobacterium paratuberculosis cultured from milk and supramammary lymph nodes of infected asymptomatic cows. J.Clin.Microbiol., 30(1):166-71.
Sweeney RW, Whitlock RH y Rosenberger AN. 1992b. Mycobacterium paratuberculosis isolated from fetuses of infected cows not manifesting sings of the disease. Am.J.Vet.Res., 53(4):477-80.
Tafti AK y Rashidi K. 2000. The pathology of goat paratuberculosis: gross and histopathological lesions in the intestines and mesenteric lymph nodes. J.Vet.Med.B Infect.Dis.Vet.Public Health, 47(7):487-95.
Tasara T y Stephan R. 2005. Development of an F57 sequence-based real-time PCR assay for detection of Mycobacterium avium subsp. paratuberculosis in milk. Appl.Environ.Microbiol., 71(10):5957-68.
Taylor AW. 1945. Ovine paratuberculosis. J.Comp.Pathol., 55:41-4. Taylor AW. 1951. Varieties of Mycobacterium johnei isolated from sheep. J.Pathol.Bacteriol., 63:333-7. Taylor AW. 1953. Experimental Johne's disease in cattle. J.Comp.Pathol.Ther., 63:355-73. Taylor TK, Wilks CR y McQueen DS. 1981. Isolation of Mycobacterium paratuberculosis from the milk of a cow with Johne's
disease. Vet.Rec., 109:532-3. Telenti A, Marchesi F, Balz M, Bally F, Böttger EC y Bodmer T. 1993. Rapid identification of mycobacteria to the species level by
polymerase chain reaction enzyme analysis. J.Clin.Microbiol., 31(2):175-8. Tellechea J, Gutiérrez M, Pérez V y García Marín JF. 1994. Valoración preliminar de las respuestas inmune humoral y celular en
ovinos y caprinos inmunizados frente a paratuberculosis con vacunas inactivadas y atenuadas. XVIII Jorn.Científ.SEOC, SEOC. Albacete, España.
Temple RM, Muscoplat CC, Thoen CO, Himes EM y Johnson DW. 1979. Observations on diagnostic tests for paratuberculosis in a deer herd. J.Am.Vet.Med.Assoc., 175(9):914-5.
Tessema MZ, Koets AP, Rutten VP y Gruys E. 2001. How does Mycobacterium avium subsp. paratuberculosis resist intracellular degradation? Vet.Q., 23(4):153-62.
Tharaldsen J, Djonne B, Fredriksen B, Nyberg O y Siguroardottir O. 2003. The National Paratuberculosis Program in Norway. Acta Vet.Scand., 2003(44):243-6.
Thierry D, Cave MD, Eisenach KD, Crawford JT, Bates JH, Gicquel B y Guesdon JL. 1990. IS6110, an IS-like element of Mycobacterium tuberculosis complex. Nucleic Acids Res., 18(1):188.
Thoen CO y Baum KH. 1988. Current knowledge on paratuberculosis. J.Am.Vet.Med.Assoc., 192(11):1609-11. Thoen CO y Haagsma J. 1996a. Molecular techniques in the diagnosis and control of paratuberculosis in cattle.
J.Am.Vet.Med.Assoc., 209(4):734-7.

BIBLIOGRAFÍA CITADA
226
Thoen CO, Himes EM y Campbell JH. 1996b. Isolation of Mycobacterium avium serotype 3 from a white-headed tree duck (Dendrocigna viduata). Av.Dis., 20(3):587-92.
Thoen CO y Muscoplat CC. 1979. Recent developments in diagnosis of paratuberculosis. J.Am.Vet.Med.Assoc., 174(8):838-40. Thoen CO, Owen WJ, Himes EM y Hall MR. 1983. Experimental exposure of cattle to Mycobacterium paratuberculosis. En:
Merkal editor. Proc.1st Int.Coll.Paratub., pp. 120-6. NADC, USDA, Ames, IA, USA. Thomas GA, Swift GL, Green JT, Newcombe RG, Braniff-Mathews C, Rhodes J, Wilkinson S, Strohmeyer G y Kreuzpainter G.
1998. Controlled trial of antituberculous chemotherapy in Crohn's disease: a five year follow up study. Gut, 42(4):497-500. Thomas GW. 1983. Paratuberculosis in a large goat herd. Vet.Rec., 113:464-6. Thorel MF, Huchzermeyer HF y Michel AL. 2001. Mycobacterium avium and Mycobacterium intracellulare infection in mammals.
Rev.- Off.Int.Epizoot., 20(1):204-18. Thorel MF, Krichevsky MI y Levy-Frebault VV. 1990. Numerical taxonomy of mycobactin-dependent mycobacteria, emended
description of Mycobacterium avium, and description of Mycobacterium avium subsp. avium subsp. nov., Mycobacterium avium subsp. paratuberculosis subsp. nov., and Mycobacterium avium subsp. silvaticum subsp. nov. Int.J.Syst.Bacteriol., 40(3):254-60.
Thoresen OF, Falk K y Evensen O. 1994a. Comparison of immunohistochemistry, acid-fast staining, and cultivation for detection of Mycobacterium paratuberculosis in goats. J.Vet.Diagn.Invest., 6(2):195-9.
Thoresen OF y Olsaker I. 1994b. Distribution and hybridization patterns of the insertion element IS900 in clinical isolates of Mycobacterium paratuberculosis. Vet.Microbiol., 40(3-4):293-303.
Thorne JG y Hardin LE. 2000. Estimated pevalence of paratuberculosis in Missouri, USA cattle. Prev.Vet.Med., 31:51-7. Thornton CG, MacLellan KM, Stabel JR, Carothers C, Whitlock RH y Passen S. 2002. Application of the C(18)-
carboxypropylbetaine specimen processing method to recovery of Mycobacterium avium subsp. paratuberculosis from ruminant tissue specimens. J.Clin.Microbiol., 40(5):1783-90.
Timpe A y Runyon EH. 1954. The relationship of "atypical" acid-fast bacteria to human disease. J.Lab.Clin.Med., 44:202-9. Timson WJ y Short AJ. 1965. Resistance of microorganisms to hydrostatic pressure. Biotechnol.Bioeng., 7(1):139-59. Tiveljung A, Soderholm JD, Olaison G, Jonasson J y Monstein HJ. 1999. Presence of eubacteria in biopsies from Crohn's disease
inflammatory lesions as determined by 16S rRNA gene-based PCR. J.Med.Microbiol., 48(3):263-8. Toft N, Nielsen SS y Jorgensen E. 2005. Continuous-data diagnostic tests for paratuberculosis as a multistage disease. J.Dairy Sci.,
88(11):3923-31. Torres-Anjel MJ y Hedrick TI. 1971. Spore removal by centrifugation and its effect on ultra-high temperature commercial
sterilization of milk. J.Dairy Sci., 54(3):326-30. Tortoli E. 2003. Impact of genotypic studies on mycobacterial taxonomy: the new mycobacteria of the 1990s. Clin.Microbiol.Rev.,
16(2):319-54. Tortoli E, Mariottini A y Mazzarelli G. 2003. Evaluation of INNO-LiPA MYCOBACTERIA v2: improved reverse hybridization
multiple DNA probe assay for mycobacterial identification. J.Clin.Microbiol., 41(9):4418-20. Tortoli E, Rindi L, Garcia MJ, Chiaradonna P, Dei R, Garzelli C, Kroppenstedt RM, Lari N, Mattei R, Mariottini A, Mazzarelli G,
Murcia MI, Nanetti A, Piccoli P y Scarparo C. 2004. Proposal to elevate the genetic variant MAC-A, included in the Mycobacterium avium complex, to species rank as Mycobacterium chimaera sp. nov. Int.J.Syst.Evol.Microbiol., 54(Pt 4):1277-85.
Trujillo AJ. 2002. Applications of high-hydrostatic pressure on milk and dairy products. High Pressure Res., 22(3-4):619-26. Tryland M, Olsen I, Vikoren T, Handeland K, Arnemo JM, Tharaldsen J, Djonne B, Josefsen TD y Reitan LJ. 2004. Serologic
survey for antibodies against Mycobacerium avium subsp. paratuberculosis in free-ranging cervids from Norway. J.Wildl.Dis., 40(1):32-41.
Tunkl B y Aleraj Z. 1965. The recovery of M. johnei in the semen of bull from one station fro artificial insemination. Vet.Glasnik, 19:845-9.
Turenne CY, Semret M, Cousins DV, Collins DM y Behr MA. 2006. Sequencing of hsp65 distinguishes among subsets of the Mycobacterium avium complex. J.Clin.Microbiol., 44(2):433-40.
Turenne CY, Tschetter L, Wolfe J y Kabani A. 2001. Necessity of quality-controlled 16S rRNA gene sequence databases: identifying nontuberculous Mycobacterium species. J.Clin.Microbiol., 39(10):3637-48.
Twort FW y Ingram GLY. 1911. Further experiments with the Mycobacterium enteriditis chronicae pseudotuberculosae bovis, Johne, and with vaccines prepared from this micro-organism. Zentralbl.Bakteriol.[Orig A], 67(3):126-35.
Twort FW y Ingram GLY. 1912. A method for isolating and cultivating the Mycobacterium enteriditis chronicae pseudotuberculosae bovis, Johne, and some experiments on the preparation of a diagnostic vaccine for pseudo-tuberculous enteritis of bovines. Proc.R.Soc.Lond., B, Biol.Sci., 84:517-42.
Twort FW y Ingram GLY. 1913, A monograph on Johne's disease. Baltiere,Tindall and Cox.London, UK. Tzen CY, Wu TY y Tzen CY. 2006. Detection of mycobacteria in Crohn's disease by a broad spectrum polymerase chain reaction.
J.Formos.Med.Assoc., 105(4):290-8. Valentin-Weigand P y Goethe R. 1999. Pathogenesis of Mycobacterium avium subspecies paratuberculosis infections in ruminants:
still more questions than answers. Microbes.Infect., 1(13):1121-7. Vallée H y Rinjard P. 1926. Etudes sur l'entérite paratuberculeuse des bovidés. Revue Générale de Médecine Vétérinaire, 409:1-9. Vallée H, Rinjard P y Vallée P. 1934. Sur la prémunition de l'entérite paratuberculeuse des bovidés. Revue Générale de Médecine
Vétérinaire, 43:777-9. van der Giessen J, van Dijk L, Bleumink-Pluym N, Eger T, Haagsma J y van der Zeijst B. 1995. The spatial distribution of
Mycobacterium paratuberculosis in infected cattle. Implications for pathogenesis and diagnosis. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 61-6. IAP, Rehoboth, MA, USA.
van der Giessen JWB, Eger A, Haagsma J, Haring RM, Gaastra W y van der Zeijst BAM. 1992a. Amplification of 16S rRNA sequences to detect Mycobacterium paratuberculosis. J.Med.Microbiol., 36:255-63.
van der Giessen JWB, Haring RM, Vauclare E, Haagsma J y van der Zeijst BAM. 1992b. Evaluation of the abilities of three diagnostic tests based on the polymerase chain reaction to detect Mycobacterium paratuberculosis in cattle: Application in a control program. J.Clin.Microbiol., 30(5):1216-9.
Van Embden JD, Cave MD, Crawford JT, Dale JW, Eisenach KD, Gicquel B, Hermans P, Martin C, McAdam RA, Shinnick TM y Small PM. 1993. Strain identification of Mycobacterium tuberculosis by DNA fingerprinting: recommendations for a standardized methodology. J.Clin.Microbiol., 31(2):406-9.
van Kooten HC, Mackintosh CG y Koets AP. 2006. Intra-uterine transmission of paratuberculosis (Johne's disease) in farmed red deer. N.Z.Vet.J., 54(1):16-20.
Van Kruiningen HJ. 1999. Lack of support for a common etiology in Johne's disease of animals and Crohn's disease in humans. Inflamm.Bowel Dis., 5(3):183-91.

BIBLIOGRAFÍA CITADA
227
Van Kruiningen HJ, Chiodini RJ, Thayer WR, Coutu JA, Merkal RS y Runnels PL. 1986. Experimental disease in infant goats induced by a Mycobacterium isolated from a patient with Crohn's disease. A preliminary report. Dig.Dis.Sci., 31(12):1351-60.
Van Kruiningen HJ, Ruiz B y Gumprecht L. 1991. Experimental disease in young chickens induced by a Mycobacterium paratuberculosis isolate from a patient with Crohn's disease. Can.J.Vet.Res., 55:199-202.
Van Schaik G, Kalis CHJ, Benedictus G, Dijkhuizen AA y Huirne RBM. 1996. Cost-benefit analysis of vaccination against paratuberculosis in dairy cattle. Vet.Rec., 139(25):624-7.
Van Schaik G, Schukken YH, Crainiceanu C, Muskens J y VanLeeuwen JA. 2003a. Prevalence estimates for paratuberculosis adjusted for test variability using Bayesian analysis. Prev.Vet.Med., 60(4):281-95.
Van Schaik G, Stehman SM, Schukken YH, Rossiter CR y Shin SJ. 2003b. Pooled fecal culture sampling for Mycobacterium avium subsp. paratuberculosis at different herd sizes and prevalence. J.Vet.Diagn.Invest., 15(3):233-41.
van Soolingen D, Bauer K, Ritacco V, Carpenter JL, Pavlík I, Vincent V, Rastogi N, Gori A, Bodmer T, Garcelli C y Garcia MJ. 1998. IS1245 restriction fragment length polymorphism typing of Mycobacterium avium isolates: proposal for standardization. J.Clin.Microbiol., 36(10):3051-4.
Vandegraaff R, Barton MD, Barry GR y Vanwijk JGA. 1995. Prevalence of Mycobacterium paratuberculosis in Dairy Cattle in South-Australia. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 9-18. IAP, Rehoboth, MA, USA.
VanLeeuwen JA, Forsythe L, Tiwari A y Chartier R. 2005. Seroprevalence of antibodies against bovine leukemia virus, bovine viral diarrhea virus, Mycobacterium avium subspecies paratuberculosis, and Neospora caninum in dairy cattle in Saskatchewan. Can.Vet.J., 46(1):56-8.
VanLeeuwen JA, Keefe GP, Tremblay R, Power C y Wichtel JJ. 2001. Seroprevalence of infection with Mycobacterium avium subspecies paratuberculosis, bovine leukemia virus, and bovine viral diarrhea virus in maritime Canada dairy cattle. Can.Vet.J., 42(3):193-8.
Vannuffel P, Gilot P, Limbourg B, Naerhunzen B, Dieterich C, Coene M, Machtelinckx L y Cocito C. 1994. Development of species-specific enzyme-linked immunosorbent assay for diagnosis of Johne's disease in cattle. J.Clin.Microbiol., 32(5):1211-6.
Vanuffel P, Dieterich C, Naerhuyzen B, Gilot P, Coene M, Fiasse R y Cocito C. 1994. Occurrence, in Crohn's disease, of antibodies directed against a species-specific recombinant polypeptide of Mycobacterium paratuberculosis. Clin.Diagn.Lab.Immunol., 1:241-3.
Vary PH, Andersen PR, Green E, Hermon-Taylor J y McFadden JJ. 1990. Use of highly specific DNA probes and the polymerase chain reaction to detect Mycobacterium paratuberculosis in Johne's disease. J.Clin.Microbiol., 28(5):933-7.
Vaughan JA, Lenghaus C, Stewart DJ, Tizard ML y Michalski WP. 2005. Development of a Johne's disease infection model in laboratory rabbits following oral administration of Mycobacterium avium subspecies paratuberculosis. Vet.Microbiol., 105(3-4):207-13.
Vesosky B, Turner OC, Turner J y Orme IM. 2004. Gamma interferon production by bovine gamma delta T cells following stimulation with mycobacterial mycolylarabinogalactan peptidoglycan. Infect.Immun., 72(8):4612-8.
Vialard J, Fleury C y Lacheretz A. 1994. Pathological aspects of caprine paratuberculosis. Rec.Med.Vet.Ec.Alfort, 170(8-9):553-8. Vialard J, Lacheretz A, Thiercy A y Prave M. 1993. Detection of bovine paratuberculosis by culture of pooled faeces.
Rev.Med.Vet., 144(6):527-33. Villanueva M. 1989. Primer diagnóstico de paratuberculosis en la región de Cantabria. En: Juste editor. 2ª Reu.PTBC España
(1987), p. 32. Informes Técnicos, Gobierno Vasco, Gasteiz, Euskadi, España. Vogel O. 1977. Paratuberculosis in a dog. Berl.Munch.Tierartzl.Wochenschr., 90(21):419-21. Waldner CL, Cunningham GL, Janzen ED y Campbell JR. 2002. Survey of Mycobacterium avium subspecies paratuberculosis
serological status in beef herds on community pastures in Saskatchewan. Can.Vet.J., 43(7):542-6. Wall S, Kunze ZM, Saboor S, Soufleri I, Seechurn P, Chiodini RJ y McFadden JJ. 1993. Identification of spheroplast-like agents
isolated from tissues of patients with Crohn's disease and control tissues by polymerase chain reaction. J.Clin.Microbiol., 31(5):1241-5.
Wallace RJ, Jr., Zhang Y, Brown BA, Fraser V, Mazurek GH y Maloney S. 1993. DNA large restriction fragment patterns of sporadic and epidemic nosocomial strains of Mycobacterium chelonae and Mycobacterium abscessus. J.Clin.Microbiol., 31(10):2697-701.
Walmsley RS, Ibbotson JP, Chahal H y Allan RN. 1996. Antibodies against Mycobacterium paratuberculosis in Crohn's disease. Q.J.Med., 89:217-21.
Wang C, Turnbull BW, Grohn YT y Nielsen SS. 2006. Estimating receiver operating characteristic curves with covariates when there is no perfect reference test for diagnosis of Johne's disease. J.Dairy Sci., 89(8):3038-46.
Warren JB, Rees HC y Cox TM. 1986. Remission of Crohn's disease with tuberculosis chemotherapy. N.Engl.J.Med.(Jan.16):182. Waters WR, Miller JM, Palmer MV, Stabel JR, Jones DE, Koistinen KA, Steadham EM, Hamilton MJ, Davis WC y Bannantine JP.
2003. Early induction of humoral and cellular immune responses during experimental Mycobacterium avium subsp. paratuberculosis infection of calves. Infect.Immun., 71(9):5130-8.
Wayne LG, Good RC, Böttger EC, Butler R, Dorsch M, Ezaki T, Gross W, Jonas V, Kilburn JO, Kirschner P, Krichevsky MI, Ridell M, Shinnick TM, Springer B, Stackebrandt E, Tarnok Z, Tasaka H, Vincent V, Warren NG, Knott CA y Johnson R. 1996. Semantide- and chemotaxonomy-based analyses of some problematic phenotypic clusters of slowly growing mycobacteria, a cooperative study of the internetional working group on mycobacterial taxonomy. Int.J.Syst.Bacteriol., 46(1):280-97.
Wayne LG, Good RC, Tsang AY, Butler R, Dawson DJ, Groothuis D, Gross W, Hawkins J, Kilburn JO, Kubin M, Schroder KH, Silkox VA, Smith C, Thorel MF, Woodley CL y Yakrus MA. 1993. Serovar determination and molecular taxonomic correlation in Mycobacterium avium,Mycobacterium intracellulare,and Mycobacterium scrofulaceum:a ccoperative study of the international working group on mycobacteria taxonomy. Int.J.Syst.Bacteriol., 43(3):482-9.
Wayne LG y Kubica GP. 1986. Mycobacteria. Family Mycobacteriaceae Chester 1897. En: Sneath, P. H. A., Mair, N. S., Sharpe, M. E. y Holt, J. G. editores. Bergey's Manual of Systematic Bacteriology. pp. 1436-57. Williams & Wilkins. Baltimore, MD, USA.
Weber A, Gurke R, Bauer K y Schreyer K. 1992a. Bacterial studies on the occurrence of Mycobacterium paratuberculosis in fecal samples of zoo ruminants. Berl.Munch.Tierartzl.Wochenschr., 105(5):161-4.
Weber MF, van Roermund HJW, Velthuis AGJ, Koeijer AA, de Jong MCM y Nielen M. 2006. Milk quality assurance programmes for paratuberculosis: stochastic simulation of within-herd infection dynamics and economics. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 60-73. IAP, Madison, WI, USA.
Weber VA y Gurke R. 1992b. Bakteriologiske untersuchungen zum vorkommen von Mycobacterium paratuberculosis in kotproben von damwild (Dama dama L.). Z.Jagdwiss., 38:55-9.

BIBLIOGRAFÍA CITADA
228
Weiss DJ, Evanson OA, Deng M y Abrahamsen MS. 2004a. Gene expression and antimicrobial activity of bovine macrophages in response to Mycobacterium avium subsp. paratuberculosis. Vet.Pathol., 41(4):326-37.
Weiss DJ, Evanson OA, Deng M y Abrahamsen MS. 2004b. Sequential patterns of gene expression by bovine monocyte-derived macrophages associated with ingestion of mycobacterial organisms. Microb.Pathog., 37(4):215-24.
Weiss DJ, Evanson OA, McClenahan DJ, Abrahamsen MS y Walcheck BK. 2001. Regulation of expression of major histocompatibility antigens by bovine macrophages infected with Mycobacterium avium subsp. paratuberculosis or Mycobacterium avium subsp. avium. Infect.Immun., 69(2):1002-8.
Weiss DJ, Evanson OA, Moritz A, Deng MQ y Abrahamsen MS. 2002. Differential responses of bovine macrophages to Mycobacterium avium subsp. paratuberculosis and Mycobacterium avium subsp. avium. Infect.Immun., 70(10):5556-61.
Weiss DJ, Evanson OA y Souza CD. 2006. Mucosal immune response in cattle with subclinical Johne's disease. Vet.Pathol., 43(2):127-35.
Wells SJ. 2006. The prospects for herd level control of paratuberculosis in cattle: A scientist's view. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., pp. 174-81. IAP, Madison, WI, USA.
Wells SJ y Wagner BA. 2000. Herd-level risk factors for infection with Mycobacterium paratuberculosis in US dairies and association between familiarity of the herd manager with the disease in that herd and use of preventive measures. J.Am.Vet.Med.Assoc., 216(9):1450-7.
Wells SJ, Whitlock RH, Lindeman CJ y Fyock T. 2002. Evaluation of bacteriologic culture of pooled fecal samples for detection of Mycobacterium paratuberculosis. Am.J.Vet.Res., 63(8):1207-11.
Wentink GH, Bongers JH, Zeeuwen AAPA, Jaartsveld FHJ y Van de Broek J. 1992. Efficiency of vaccination against paratuberculosis using an inactivated whole cell vaccine. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 475-83. IAP, Providence, RI, USA.
West DM. 1997. Johne's disease in New Zealand: history from first cases to current situation. Int Cong Sheep Vet, IV, pp. 151-4. University of New England. Armidale,Australia.
Whan L, Ball HJ, Grant IR y Rowe MT. 2005a. Development of an IMS-PCR assay for the detection of Mycobacterium avium ssp. paratuberculosis in water. Lett.Appl.Microbiol., 40(4):269-73.
Whan L, Ball HJ, Grant IR y Rowe MT. 2005b. Occurrence of Mycobacterium avium subsp. paratuberculosis in untreated water in Northern Ireland. Appl.Environ.Microbiol., 71(11):7107-12.
Whan L, Grant IR y Rowe MT. 2006. Interaction between Mycobacterium avium subsp. paratuberculosis and environmental protozoa. BMC Microbiol., 6:63.
Whan LB, Grant IR, Ball HJ, Scott R y Rowe MT. 2001. Bactericidal effect of chlorine on Mycobacterium paratuberculosis in drinking water. Lett.Appl.Microbiol., 33(3):227-31.
Whipple D, Kapke P y Vary C. 1990. Identification of restriction fragment length polymorphisms in DNA from Mycobacterium paratuberculosis. J.Clin.Microbiol., 28(11):2561-4.
Whipple DL, Callihan DR y Jarnagin JL. 1991. Cultivation of Mycobacterium paratuberculosis from bovine fecal specimens and a suggested standardized procedure. J.Vet.Diagn.Invest., 3(4):368-73.
Whipple DL y Merkal RS. 1983. Modifications in the techniques for cultivation of Mycobaterium paratuberculosis. En: Merkal editor. Proc.1st Int.Coll.Paratub., pp. 82-92. NADC, USDA, Ames, IA, USA.
Whist SK, Storset AK y Larsen HJ. 2000. The use of interleukin-2 receptor expression as a marker of cell-mediated immunity in goats experimentally infected with Mycobacterium avium ssp. paratuberculosis. Vet.Immunol.Immunopathol., 73(3-4):207-18.
Whitlock RH. 1992. Laboratory diagnosis of Johne's disease. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 1-21. IAP, Providence, RI, USA.
Whitlock RH. 1998. Johne's disease (Paratuberculosis): diagnostic tests for individual animal and the herd. En: Junta ANEMBE editor. V Congr.Int.Med.Bov., pp. 69-72. Sitges, Cataluña, España.
Whitlock RH, McAdams SC, Fyock TL y Sweeney RW. 2003. An International Proficiency study for culture or detection of Mycobacterium avium sp. paratuberculosis in bovine fecal samples. En: Juste, Geijo, y Garrido editores. Proc.7th Int.Coll.Paratub., p. 226. IAP, Madison, WI, USA.
Whitlock RH, Rosenberger AE, Sweeney RW y Hutchinson LJ. 1992a. Culture techniques and media constituents for the isolation of Mycobacterium paratuberculosis from bovine fecal samples. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 94-111. IAP, Providence, RI, USA.
Whitlock RH, Rosenberger AE, Sweeney RW y Spencer PA. 1997. Distribution of M. paratuberculosis in tissues of cattle from herds infected with Johne's disease. En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., pp. 168-74. IAP, Rehoboth, MA, USA.
Whitlock RH, Sweeney RW, Fyock TL, McAdams S, Gardner IA y McClary DG. 2006a. Johne's disease: The effect of feeding Monensin to reduce the bio-burden of Mycobacterium avium subspecies paratuberculosis in neo-natal calves. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 270. IAP, Madison, WI, USA.
Whitlock RH, Sweeney RW, Fyock TL y Smith J. 2006b. MAP Super-Shedders: Another factor in the control of Johne's disease. En: Manning y Nielsen editores. Proc.8th Int.Coll.Paratub., p. 164. IAP, Madison, WI, USA.
Whitlock RH, Sweeney RW, Hutchinson LT y Van Buskirk M. 1992b. Pennsylvania Johne's disease control program. En: Chiodini y Kreeger editores. Proc.3rd Int.Coll.Paratub., pp. 455-74. IAP, Providence, RI, USA.
Whitlock RH, Sweeney RW, Hutchinson LT y Van Buskirk M. 1995. Pennsylvania Johne's disease program (1973 to 1993): a review of the twenty year program. En: Chiodini, Collins, y Bassey editores. Proc.4th Int.Coll.Paratub., pp. 102-10. IAP, Rehoboth, MA, USA.
Whitlock RH, Wells SJ, Sweeney RW y Van Tiem J. 2000. ELISA and fecal culture for paratuberculosis (Johne's disease): sensitivity and specificity of each method. Vet.Microbiol., 77(3-4):387-98.
Whitlock RH, West S, Layton B, Ellingson JL, Stabel J, Rossiter CA, Buergelt CD, Pavlik I, Ginn P, Collins MT, Juste RA y Habecker P. 1999. Paratuberculosis in bison: a comparison of PCR, culture and histopathology. En: Manning y Collins editores. Proc.6th Int.Coll.Paratub., pp. 424-38. IAP, Madison, WI, USA.
Whittington R, Marsh I, Choy E y Cousins D. 1999a. Polymorphism in IS1311, an insertion sequence common to Mycobacterium avium and Mycobacterium avium subsp. paratuberculosis, can be used to distinguish between and within these species. Mol.Cell.Probes, 12(6):349-58.
Whittington R, Marsh I, McAllister S, Turner MJ, Marshall DJ y Fraser CA. 1999b. Evaluation of a modified BACTEC12B radiometric medium and solid media for culture of Mycobacterium avium subsp. paratuberculosis from sheep. J.Clin.Microbiol., 37(4):1077-83.
Whittington RJ, Eamens GJ y Cousins DV. 2003. Specificity of absorbed ELISA and agar gel immuno-diffusion tests for paratuberculosis in goats with observations about use of these tests in infected goats. Aust.Vet.J., 81(1-2):71-5.

BIBLIOGRAFÍA CITADA
229
Whittington RJ, Fell S, Walker D, McAllister S, Marsh I, Sergeant E, Taragel CA, Marshall DJ y Links IJ. 2000a. Use of pooled fecal culture for sensitivite and economic detection of Mycobacterium avium subsp. paratuberculosis. J.Clin.Microbiol., 38(7):2550-6.
Whittington RJ, Hope AF, Marshall DJ, Taragel CA y Marsh I. 2000b. Molecular epidemiology of Mycobacterium avium subsp. paratuberculosis: IS900 restriction fragment length polymorphism and IS1311 polymorphism analyses of isolates from animals and a human in Australia. J.Clin.Microbiol., 38(9):3240-8.
Whittington RJ, Lloyd JB y Reddacliff LA. 2001a. Recovery of Mycobacterium avium subsp. paratuberculosis from nematode larvae cultured from the faeces of sheep with Johne's disease. Vet.Microbiol., 81(3):273-9.
Whittington RJ, Marsh I, Turner MJ, McAllister S, Choy E, Eamens GJ, Marshall DJ y Ottaway SJ. 1998. Rapid detection of Mycobacterium paratuberculosis in clinical samples from ruminants and in spiked environmental samples by modified BACTEC 12B radiometric culture and direct confirmation by IS900 PCR. J.Clin.Microbiol., 36(3):701-7.
Whittington RJ, Marsh I, McAllister S, Turner MJ, Marshall DJ y Fraser CA. 1999c. Evaluation of modified BACTEC12B radiometric medium and solid media for culture of Mycobacterium avium subsp. paratuberculosis from sheep. J.Clin.Microbiol., 37(4):1077-83.
Whittington RJ, Marsh IB y Reddacliff LA. 2005. Survival of Mycobacterium avium subsp. paratuberculosis in dam water and sediment. Appl.Environ.Microbiol., 71(9):5304-8.
Whittington RJ, Marsh IB y Whitlock RH. 2001b. Typing of IS1311 polymorphisms confirms that bison (Bison bison) with paratuberculosis in Montana are infected with a strain of Mycobacterium avium subsp. paratuberculosis distinct from that occurring in cattle and other domesticated livestock. Mol.Cell.Probes, 15(3):139-45.
Whittington RJ, Marshall DJ, Nicholls PJ, Marsh IB y Reddacliff LA. 2004. Survival and dormancy of Mycobacterium avium subsp. paratuberculosis in the environment. Appl.Environ.Microbiol., 70(5):2989-3004.
Whittington RJ, Reddacliff L, Marsh I, McAllister S y Saunders V. 2000c. Temporal patterns and quantification of excretion of Mycobacterium avium subsp. paratuberculosis in sheep with Johne's disease. Aust.Vet.J., 78(1):34-7.
Whittington RJ, Reddacliff L, Marsh I y Saunders V. 1999d. Detection of Mycobacterium avium subsp. paratuberculosis in formalin-fixed paraffin-embedded intestinal tissue by IS900 polymerase chain reaction. Aust.Vet.J., 77(6):392-7.
Whittington RJ y Sergeant ES. 2001c. Progress towards understanding the spread, detection and control of Mycobacterium avium subsp. paratuberculosis in animal populations. Aust.Vet.J., 79(4):267-78.
Whittington RJ, Taragel CA, Ottaway S, Marsh I, Seaman J y Fridriksdottir V. 2001d. Molecular epidemiological confirmation and circumstances of occurrence of sheep (S) strains of Mycobacterium avium subsp. paratuberculosis in cases of paratuberculosis in cattle in Australia and sheep and cattle in Iceland. Vet.Microbiol., 79(4):311-22.
Wilesmith JW. 1982. Johne's disease: A retrospective study of vaccinated herds in Great Britain. Br.Vet.J., 138(4):321-31. Williams ES, Snyder SP y Martin KL. 1983. Experimental infection of some north american ruminants and domestic sheep with
Mycobacterium paratuberculosis: clinical and bacteriological findings. J.Wildl.Dis., 19(3):185-91. Williams ES, Spraker TR y Scoonveld GG. 1979. Paratuberculosis (Johne's disease) in bighorn sheep and a Rocky Mountain goat in
Colorado. J.Wildl.Dis., 15(2):221-7. Williams SL, Harris NB y Barletta RG. 1999. Development of a firefly luciferase-based assay for determining antimicrobial
susceptibility of Mycobacterium avium subsp. paratuberculosis. J.Clin.Microbiol., 37(2):304-9. Wilson DJ, Rossiter CA, Han HR y Sears PM. 1997. Financial effects of Mycobacterium paratuberculosis on mastitis, culling and
milk production in clinically normal dairy cattle. En: Chiodini, Hines, y Collins editores. Proc.5th Int.Coll.Paratub., pp. 151-8. IAP, Rehoboth, MA, USA.
Wilton S y Cousins D. 1992. Detection and identification of multiple mycobacterial pathogens by DNA amplification in a single tube. PCR Methods Appl., 1(4):269-73.
Windsor PA, Bush R, Links I y Eppleston J. 2005. Injury caused by self-inoculation with a vaccine of a Freund's complete adjuvant nature (Gudair) used for control of ovine paratuberculosis. Aust.Vet.J., 83(4):216-20.
Windsor PA y Eppleston J. 2006. Lesions in sheep following administration of a vaccine of a Freund's complete adjuvant nature used in the control of ovine paratuberculosis. N.Z.Vet.J., 54(5):237-41.
Winterhoff C, Beyerbach M, Homuth M, Strutzberg K y Gerlach GF. 2002. Establishment and evaluation of an ELISA for the detection of antibodies in milk against Mycobacterium avium subspecies paratuberculosis. Dtsch.Tierarztl.Wochenschr., 109(5):230-4.
Wiszniewska A, Szteyn J, Fus MM y Gomolka-Pawlicka M. 2004. Optimisation of the time of milk sample decontamination with 0.75% cethylpirydinium chloride upon identification of Mycobacterium avium subsp. paratuberculosis. Pol.J.Vet.Sci., 7(2):113-6.
Wood PR, Corner LA y Plackett P. 1990a. Development of a simple, rapid in vitro cellular assay for bovine tuberculosis based on the production of gamma interferon. Res.Vet.Sci., 49(1):46-9.
Wood PR, Rothel JS, McWarters PGD y Jones SL. 1990b. Production and characterization of monoclonal antibodies specific for bovine gamma-interferon. Vet.Immunol.Immunopathol., 25(1):37-46.
Yakrus MA y Straus WL. 1994. DNA polymorphisms detected in Mycobacterium haemophilum by pulsed-field gel electrophoresis. J.Clin.Microbiol., 32(4):1083-4.
Yamamoto T. 2005. Factors affecting recurrence after surgery for Crohn's disease. World J.Gastroenterol., 11(26):3971-9. Yamori S y Tsukamura M. 1992. Comparison of prognosis of pulmonary diseases caused by Mycobacterium avium and by
Mycobacterium intracellulare. Chest, 102(1):89-90. Yao J, Burton JL, Saama P, Sipkovsky S y Coussens PM. 2001. Generation of EST and cDNA microarray resources for the study of
bovine immunobiology. Acta Vet.Scand., 42(3):391-405. Yokomizo Y, Kishima M, Mori Y y Nishimori K. 1991. Evaluation of enzyme-linked immunosorbent assay in comparison with
complement fixation test for the diagnosis of subclinical paratuberculosis in cattle. J.Vet.Med.Sci., 53(4):577-84. Yokomizo Y, Merkal RS y Lyle PAS. 1983. Enzyme-linked immunosorbent assay (ELISA) for detection of bovine immunoglobulin
G1 antibody to a protoplasmatic antigen of Mycobacterium paratuberculosis . Am.J.Vet.Res., 44(11):2205-7. Yokomizo Y, Yugi H y Merkal RS. 1985. A method for avoiding false-positive reaction in an enzyme-linked immunoassay
(ELISA) for the diagnosis of bovine paratuberculosis. Nippon Juigaku Zasshi, 47(1):111-9. Yoshimura HH y Graham D. 1988. Nucleic acid hybridization studies of mycobactin-dependent mycobacteria. J.Clin.Microbiol.,
26(7):1309-12. Zhang Q, Kennon R, Koza MA, Hulten K y Clarridge JE, III. 2002. Pseudoepidemic due to a unique strain of Mycobacterium
szulgai: genotypic, phenotypic, and epidemiological analysis. J.Clin.Microbiol., 40(4):1134-9. Zhang Y, Mazurek GH, Cave MD, Eisenach KD, Pang Y, Murphy DT y Wallace RJ, Jr. 1992. DNA polymorphisms in strains of
Mycobacterium tuberculosis analyzed by pulsed-field gel electrophoresis: a tool for epidemiology. J.Clin.Microbiol., 30(6):1551-6.

BIBLIOGRAFÍA CITADA
230
Zhang Y, Rajagopalan M, Brown BA y Wallace RJ, Jr. 1997. Randomly amplified polymorphic DNA PCR for comparison of Mycobacterium abscessus strains from nosocomial outbreaks. J.Clin.Microbiol., 35(12):3132-9.
Zhao BY, Collins MT y Czuprynsky CJ. 1997. Effects of gamma interferon and nitric oxid on the interaction of Mycobacterium avium subsp. paratuberculosis with bovine monocytes. Infect.Immun., 65(5):1761-6.
Zur LS, Goethe R, Darji A, Valentin-Weigand P y Weiss S. 2003. Activation of macrophages and interference with CD4+ T-cell stimulation by Mycobacterium avium subspecies paratuberculosis and Mycobacterium avium subspecies avium. Immunology, 108(1):62-9.

TESIS DOCTORALES PUBLICADAS
Nº 1. La raza Latxa: Sistemas de producción y características reproductivas. EDUARDO URIARTE
EGURCEGUI
Nº 2. Estudio y puesta a punto de un método simplificado de control lechero cualitativo en la raza
ovina Latxa y su inclusión en el plan de selección. GUSTAVO ADOLFO MARIA LEVRINO
Nº 3. Implicaciones tecnológicas de la composición química del pescado con especial referencia a los
lípidos. ROGELIO POZO CARRO
Nº 4. Estudio de suelos de Vizkaia. MARGARITA DOMINGO URARTE
Nº 5. El Maedi o neumonía progresiva en el conjunto de las enfermedades respiratorias crónicas del
ganado ovino en la Comunidad Autónoma Vasca. LORENZO GONZÁLEZ ANGULO
Nº 6. Estudio experimental de las fases iniciales de la paratuberculosis ovina. RAMÓN A. JUSTE JORDAN
Nº 7. Identificación, origen y factores fisicoquímicos que condicionan la contaminación por elementos
metálicos de sedimentos de ríos. ESTILITA RUIZ ROMERA
Nº 8. Análisis financiero de proyectos de inversión en repoblaciones forestales. ÁLAVARO AUNOS GÓMEZ
Nº 9. Desarrollo y evaluación del sistema integrado de diagnóstico y recomendación (DRIS) para la
fertilización de las praderas permanentes. Marta Rodríguez Julia
Nº 10. Estudio de las mieles producidas en la Comunidad Autónoma del País Vasco. MARÍA TERESA
SANCHO ORTIZ
Nº 11. La biomasa microbiana como agente de las transformaciones de nitrógeno en el suelo tras el
enterrado de la paja de cereal. JESÚS ÁNGEL OCIO ARMENTIA
Nº 12. Análisis jurídico y económico de la implementación de la política agraria comunitaria en la
Comunidad Autónoma del País Vasco. BEATRIZ PÉREZ DE LAS HERAS
Nº 13. Nemátodos formadores de quistes (Globodera spp.) en patata (Solanum tuberosum L.): caracteri-
zación taxonómica, reproducción y actividad de las formas juveniles. AZUCENA SALAZAR BAYONA
Nº 14. Ensayo comparativo de tres métodos de tratamiento antihelmítico estratégico en rebaños de ove-
jas latxas. ANA LUISA GRACIA PÉREZ
Nº 15. Estudio sobre una encefalitis vírica similar al Louping-ill en el ganado ovino de la Comunidad
Autónoma Vasca. DANIEL FERNÁNDEZ DE LUCO MARTINÉZ
Nº 16. Análisis de caracteres involucrados en la selección y mejora de Lupinus hispanicus Boiss. et
Reuter. VERÓNICA ARRIETA PICO
Nº 17. Contribución al estudio de fermentaciones artesanales e industriales de Rioja Alavesa. MILAGROS
VIÑEGRA GARCÍA
Nº 18. Estudio del manejo de la alimentación en los rebaños ovinos de raza Latxa y su influencia sobre
los resultados reproductivos y de producción de leche. LUIS Mª. OREGUI LIZARRALDE
Nº 19. El sector pesquero vizcaíno, 1800-1960. Análisis de la interacción de los elementos ambiental,
extractivo y comercial en la pesquería. JOSÉ AGUSTÍN MAIZ ALCORTA
Nº 20. Epidemiología, diagnóstico y control de la paratuberculosis ovina en la Comunidad Autónoma
del País Vasco. J. J. ADURIZ RECALDE
Nº 21. Agrupación de poblaciones locales de maíz (Zea mays L.) mediante caracteres morfológicos y
parámetros ambientales. JOSÉ IGNACIO RUIZ DE GALARRETA GÓMEZ
Nº 22. Estudio del potencial melífero de Bizkaia. AMELIA CERVELLO MARTÍNEZ

Nº 23. Influencia de los procesos de salado y ahumado sobre las características fisicoquímicas del
queso Idiazabal (compuestos nitrogenados). FRANCISCO C. IBAÑEZ MOYA
Nº 24. El Euskal Artzain Txakurra (el perro pastor vasco) descripción y tipificación racial. MARIANO
GÓMEZ FERNÁNDEZ
Nº 25. Evaluación de diferentes ciclos de selección recurrente en dos poblaciones sintéticas de maíz.
GOTZONE GARAY SOLACHI
Nº 26. Valoración agronómica de la gallinaza: Compostaje. ADOLFO MENOYO PUELLES
Nº 27. Relación clima-vegetación en la Comunidad Autónoma del País Vasco. AMELIA ORTUBAY FUENTES
Nº 28. Influencia de los procesos de salado y ahumado tradicional sobre las características microbioló-
gicas y organolépticas del queso Idiazabal. FRANCISCO J. PÉREZ ELORTONDO
Nº 29. Mastitis en la oveja Latxa: epidemiología, diagnóstico y control. JUAN C. MARCO MELERO
Nº 30. Contribución al conocimiento anatomopatológico y diagnóstico de la tuberculosis caprina y
ovina por Mycobacterium bovis. M.ª MONTSERRAT GUTIÉRREZ CANCELA
Nº 31. Estudio de factores que pueden influir en la calidad de la pluma de gallos Eusko-oiloa (Variedad
Marradune) para la fabricación de moscas artificiales utilizadas en la pesca de la trucha. ROSA
M.ª ECHARRI TOMÉ
Nº 32. Estudio de la fracción lipídica durante la maduración del queso Idiazabal. Influencia de los pro-
cesos tecnológicos del tiempo de permanencia en salmuera y ahumado. ANA ISABEL NÁJERA
ORTIGOSA
Nº 33.- Influencia del tipo de cuajo y adición de cultivo iniciador sobre los compuestos nitrogenados
durante la maduración del queso Idiazabal. M.ª SOLEDAD VICENTE MARTÍN
Nº 34. Estudio de la infección por Borrelia burgdorferi, grupo Ehrlichia phagocytophila y virus de la
encefalitis ovina en las poblaciones de ixódidos de la Comunidad Autónoma Vasca. MARTA
BARRAL LAHIDALGA
Nº 35. Lipolisis en el queso Idiazabal: efecto de la época de elaboración, del cultivo iniciador, de la pas-
teurización y del tipo de cuajo. FELISA CHAVARRI DÍAZ DE CERIO
Nº 36. Aspectos inmunopalógicos de la paratuberculosis de los pequeños rumiantes. Respuesta inmune
asociada a la vacunación. JUAN MANUEL CORPA ARENAS
Nº 37. Desarrollo y evaluación de nuevas técnicas de diagnóstico del Maedi-Visna. ANA BELÉN
EXTRAMIANA ALONSO
Nº 38. Estudios sobre Patogenia y Diagnóstico de la Adenomatosis Pulmonar Ovina. MARÍA MERCEDES
GARCÍA GOTI
Nº 39. Análisis de los factores de explotación que afectan a la producción lechera en los rebaños de
raza Latxa de la CAPV. ROBERTO J. RUIZ SANTOS
Nº 40. Crecimiento y producción de repoblaciones de Pinus radiata D. Don en el Territorio Histórico
de Gipuzkoa (País Vasco). LUIS MARIO CHAUCHARD BADANO
Nº 41. Puesta a punto de técnicas PCR en heces y de Elisa para el diagnóstico de la Paratuberculosis.
Estudio de prevalencia en ganado bovino. JOSEBA M. GARRIDO URKULLU
Nº 42. Epidemiología y diagnóstico de la leptospirosis y la neosporosis en explotaciones de bovino
lechero de la CAPV. RAQUEL ACHAERANDIO GALDOS
Nº 43. Relaciones aire-agua en sustratos de cultivo como base para el control del riego. Metodología de
laboratorio y modelización. VALENTÍN TERÉS TERÉS
Nº 44. Zonas endémicas de enfermedad de Lyme en la CAPV: estudio del papel de los micromamíferos
en el mantenimiento de Borrelia burgdorferi sensu lato en el medio natural. HORACIO GIL GIL

Nº 45. Optimización del esquema de mejora de la raza Latxa: análisis del modelo de valoración e intro-ducción de nuevos caractéres en el objetivo de selección. ANDRÉS LEGARZA ALBIZU
Nº 46. Influencia de las condiciones de almacenamiento, reimplantación y lluvia ácida en la viabilidadde Pinus radiata D. Don. MIREN AMAIA MENA PETITE
Nº 47. Estudio sobre encefalopatías en peces: patogenicidad del nodavirus causante de la enfermedad yretinopatía vírica (ERV) y transmisión experimental del prión scrapie a peces. RAQUEL ARANGUREN
RUIZ
Nº 48. Enfermedades transmitidas por semilla en judía-grano (Phaseolus vulgaris L.): detección, controlsanitario y mejora genética. ANA MARÍA DÍEZ NAVAJAS
Nº 49. Pastoreo del ganado vacuno en zonas de montaña y su integración en los sistemas de producciónde la CAPV. NEREA MANDALUNIZ ASTIGARRAGA
Nº 50. Aspectos básicos de la mejora genética de patata (Solanum tuberosum L.) a nivel diploide. LEIRE
BARANDALLA URTIAGA
Nº 51. El cuajo de cordero en pasta: preparación y efecto en los procesos proteolíticos y lipolíticos de lamaduración del queso de Idiazabal. Mª. ÁNGELES BUSTAMANTE GALLEGO
Nº 52. Dinámica de la población de atún blanco (Thunnus alalunga Bonnaterre 1788) del AtlánticoNorte. JOSU SANTIAGO BURRUTXAGA
Nº 53. El pino radiata (Pinus radiata D.Don) en la historia forestal de la Comunidad Autónoma de eus-kadi. Análisis de un proceso de forestalismo intensivo. MARIO MICHEL RODRÍGUEZ
Nº 54. Balance hídrico y mineral del pimiento de Gernika (Capsicum annuum L., cv Derio) en cultivohidropónico. Relaciones con la producción. HUGO MACÍA OLIVER
Nº 55. Desarrollo de métodos moleculares y su aplicación al estudio de la resistencia genética y patoge-nia molecular del Scrapie. DAVID GARCÍA CRESPO
Nº 56. Estudio epidemiológico y experimental de la transmisión y control del virus Maedi-Visna enovino lechero de raza Latxa del País Vasco. VEGA ÁLVAREZ MAIZTEGUI
Nº 57. Desarrollo y aplicación de técnicas de diagnóstico serológico para el estudio de la transmisióncalostral y horizontal del virus Maedi-Visna (VMV) en ovino. MARA ELISA DALTABUIT TEST
Nº 58. Integral Study of Calving Ease in Spanish Holstein Population. EVANGELINA LÓPEZ DE MATURANA
LÓPEZ DE LACALLE